L’INFORMATION GENETIQUE - SVT NC€™EXPRESSION DE L’INFORMATION GENETIQUE ET SON CONTROLE Flux...
111
L’INFORMATION GENETIQUE
Transcript of L’INFORMATION GENETIQUE - SVT NC€™EXPRESSION DE L’INFORMATION GENETIQUE ET SON CONTROLE Flux...

L’INFORMATION GENETIQUE

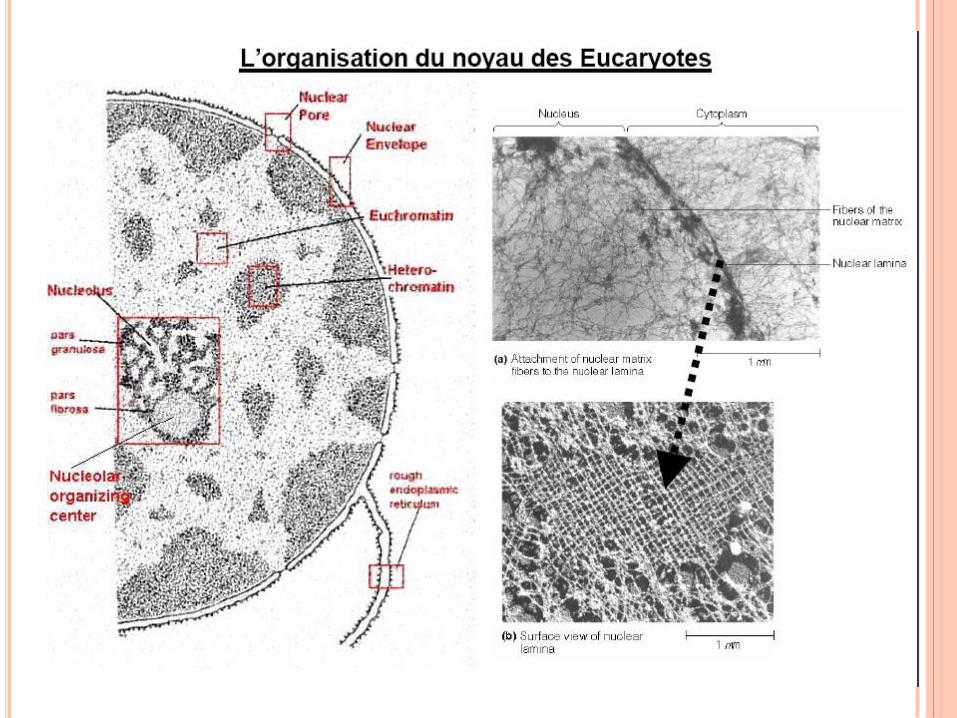
LE SUPPORT STRUCTURALE
DE L’INFORMATION GENETIQUE

TRAVAUX DE BOVERI
Travail sur la
fécondation (1888) =>
rôle du spermatozoïde
et de l’ovule
(fécondation œuf
oursin)
Étude de cas de
polyspermie
Développement normal
dépend d’une
combinaison
particulière de
chromosomes
Chaque chromosomes
individuels doivent
posséder des qualités
différentes
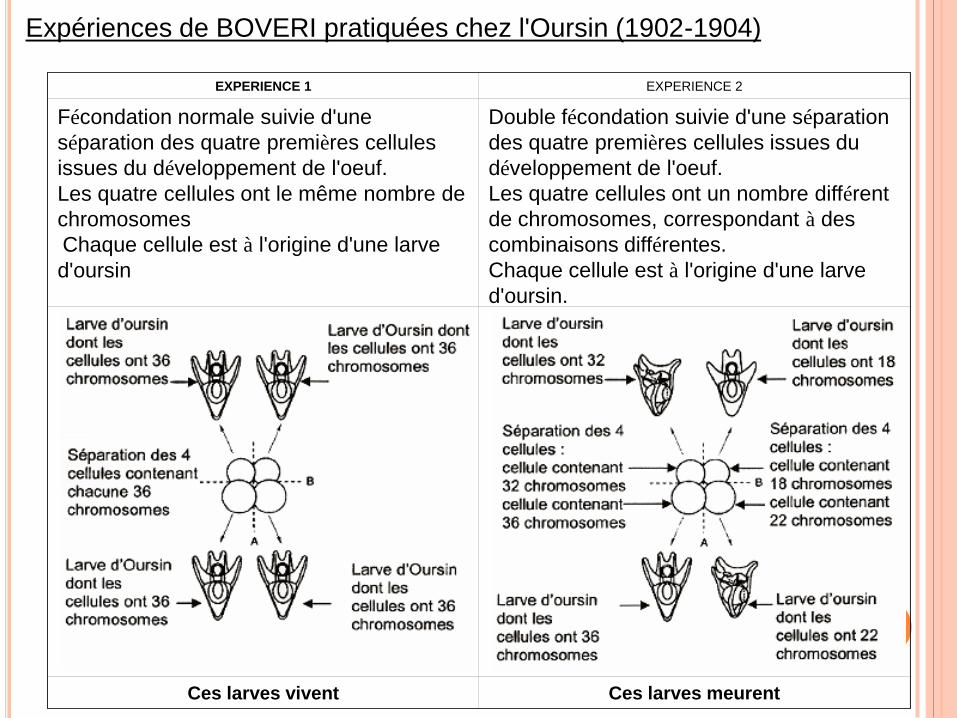
Expériences de BOVERI pratiquées chez l'Oursin (1902-1904)
EXPERIENCE 1 EXPERIENCE 2
Fécondation normale suivie d'une
séparation des quatre premières cellules
issues du développement de l'oeuf.
Les quatre cellules ont le même nombre de
chromosomes
Chaque cellule est à l'origine d'une larve
d'oursin
Double fécondation suivie d'une séparation
des quatre premières cellules issues du
développement de l'oeuf.
Les quatre cellules ont un nombre différent
de chromosomes, correspondant à des
combinaisons différentes.
Chaque cellule est à l'origine d'une larve
d'oursin.
Ces larves vivent Ces larves meurent

TRAVAUX DE SUTTON
o Formation des spermatozoides des sauterelles pas méiose
o Paires de chromosomes : chromosomes homologues => paires de facteurs héréditaires (cf Mendel)
o Paire de chromosome qui se séparent lors de la formation des gamètes (idem particules de Mendel)
o Réunion de ces chromosomes durant la fécondation (idem mendel)
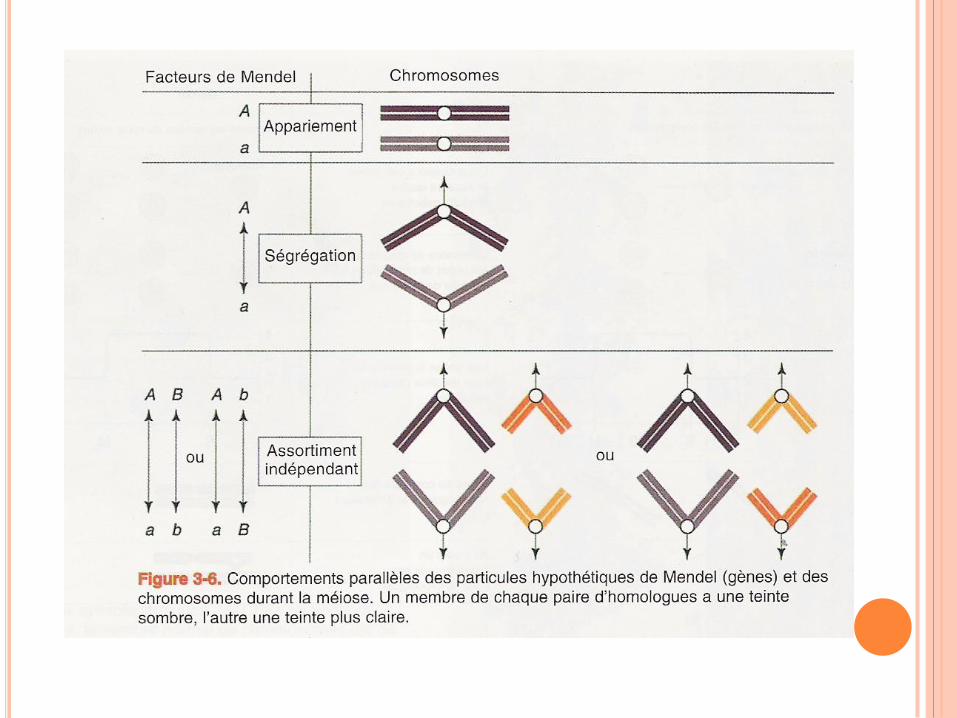

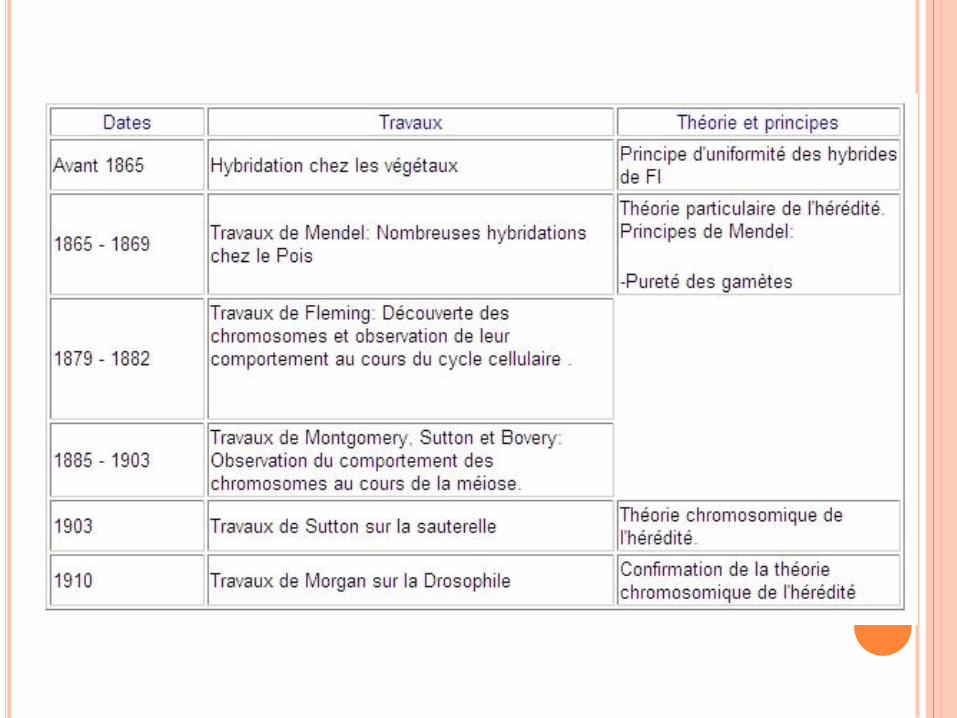
THÉORIE CHROMOSOMIQUE DE L’HEREDITÉ
Parallélisme entre le comportement des gènes et
des chromosomes suggère que les gènes sont
localisé sur les chromosomes
Sutton et Boveri proposent indépendamment la théorie
chromosomique de l’hérédité : les « facteurs
mendéliens » responsables de la transmission héréditaire
des caractères sont situés sur les chromosomes.
Critique sévère de cette théorie pendant des
années faute de preuves expérimentales…
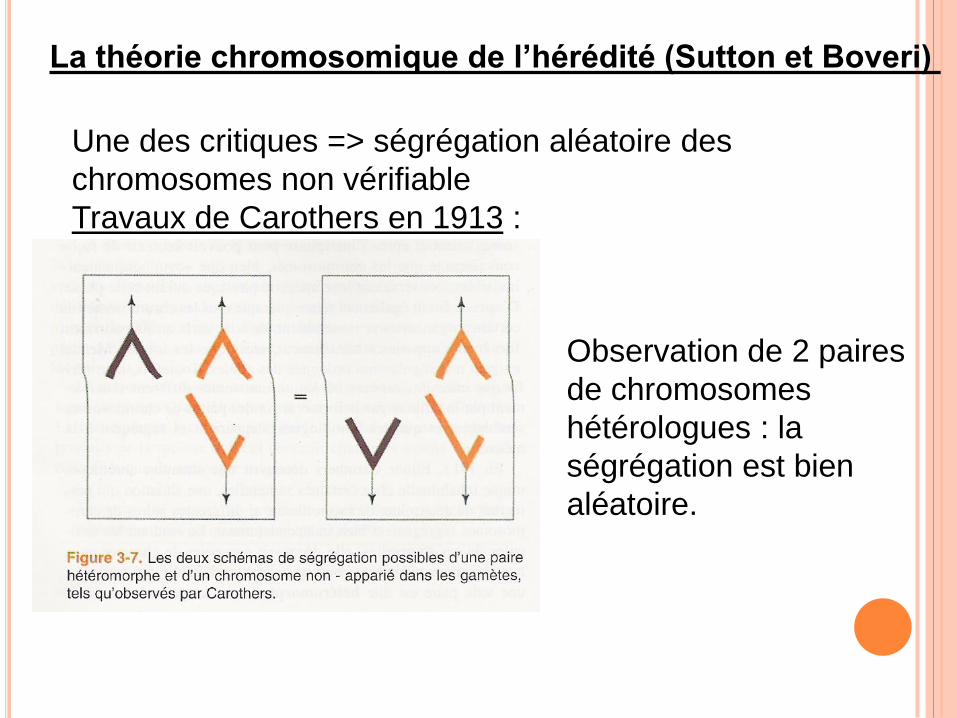
La théorie chromosomique de l’hérédité (Sutton et Boveri)
Une des critiques => ségrégation aléatoire des
chromosomes non vérifiable
Travaux de Carothers en 1913 :
Observation de 2 paires
de chromosomes
hétérologues : la
ségrégation est bien
aléatoire.

TRAVAUX DE MORGAN
Observation des chromosomes sexuels chez la drosophile
Etude de croisement qui ne correspondent pas aux prévisions de Mendel
Henking en 1891 découvrent un corps non apparié : le corps X => plus tard nommé chromosome X par Wilson.
Chez d’autres insectes : chromosome hétéromorphe : chromosome X et Y
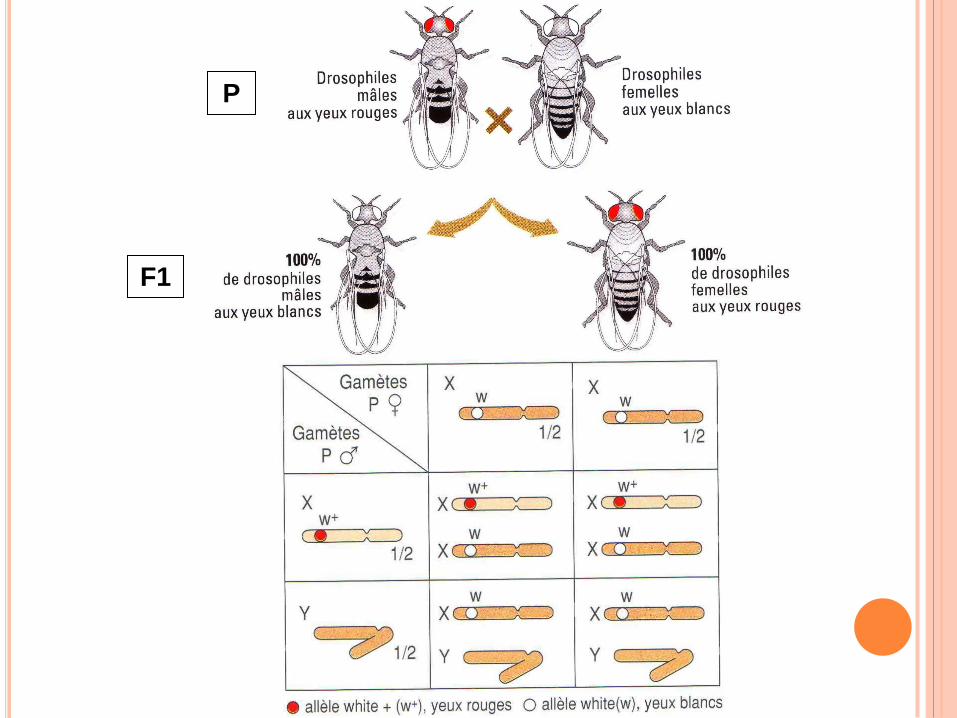
P
F1
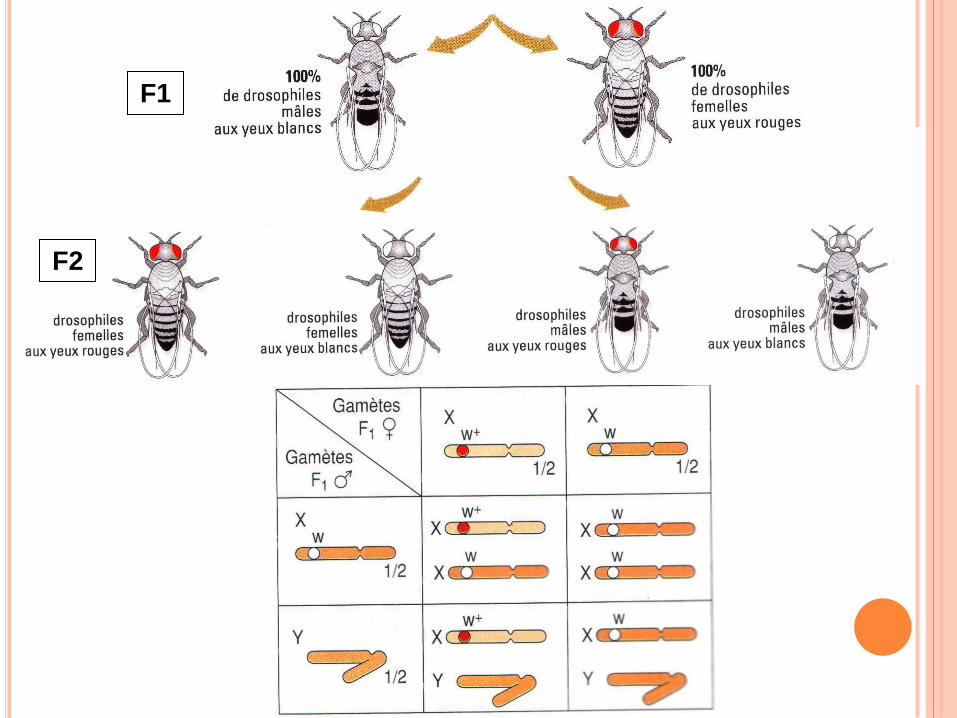
F2
F1
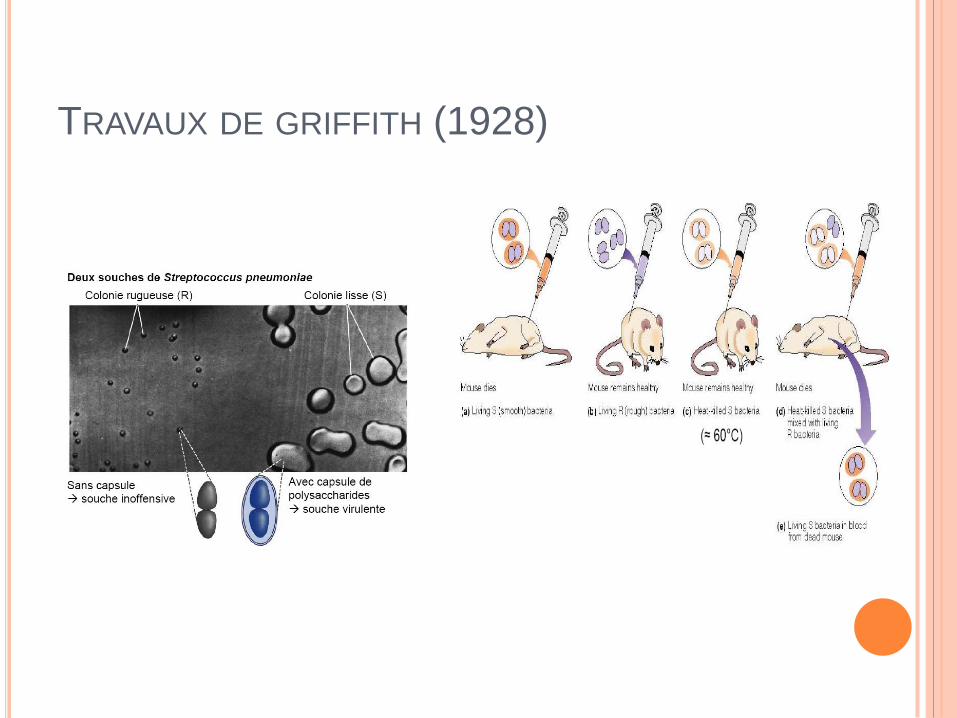
TRAVAUX DE GRIFFITH (1928)
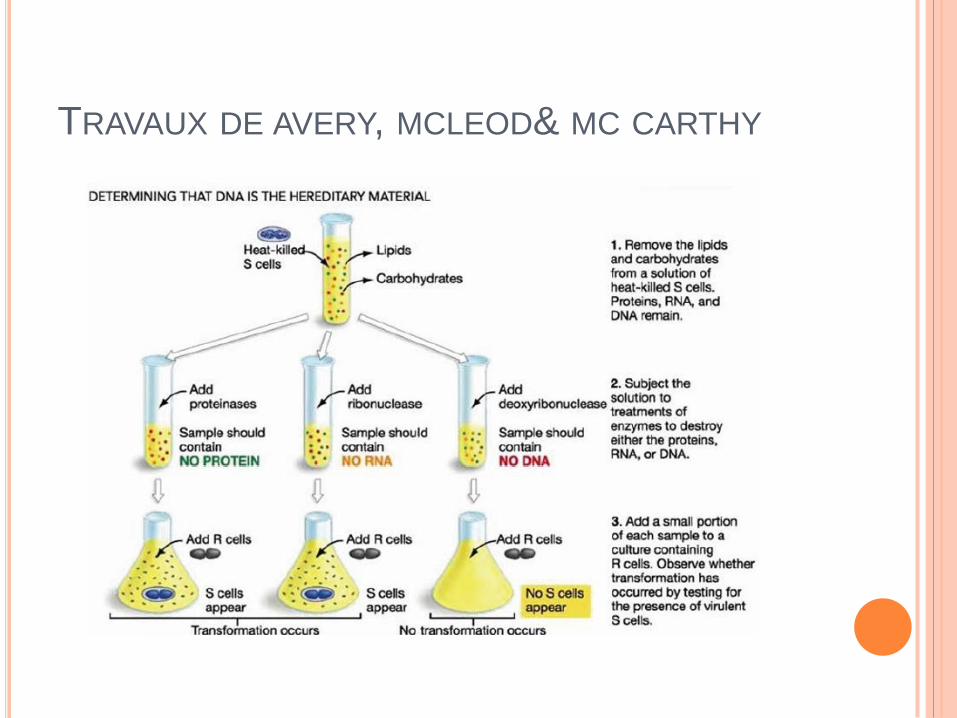
TRAVAUX DE AVERY, MCLEOD& MC CARTHY
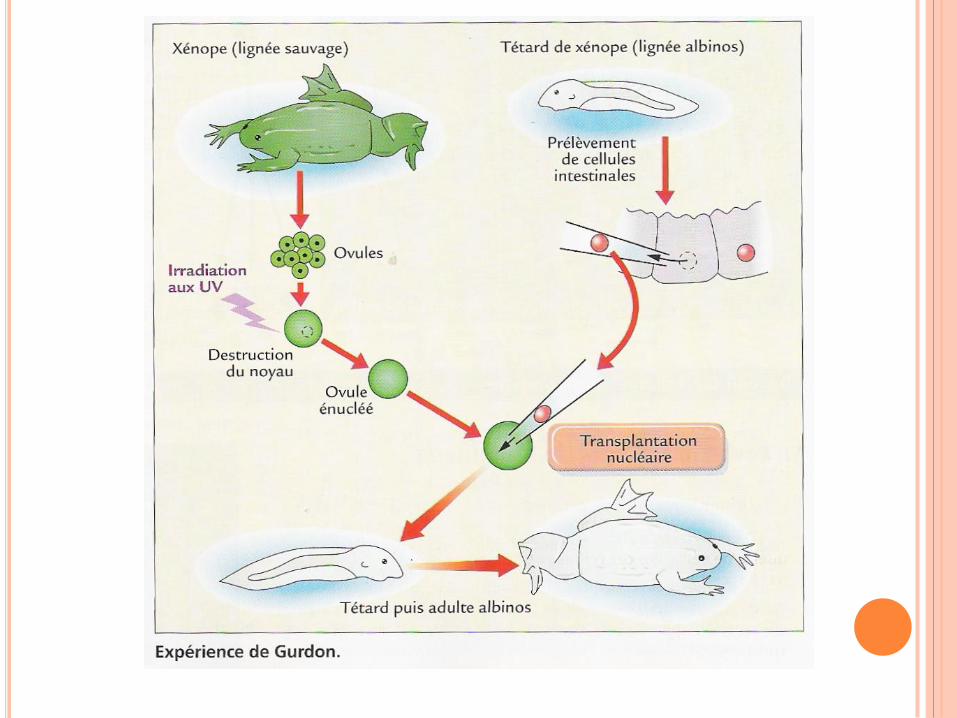
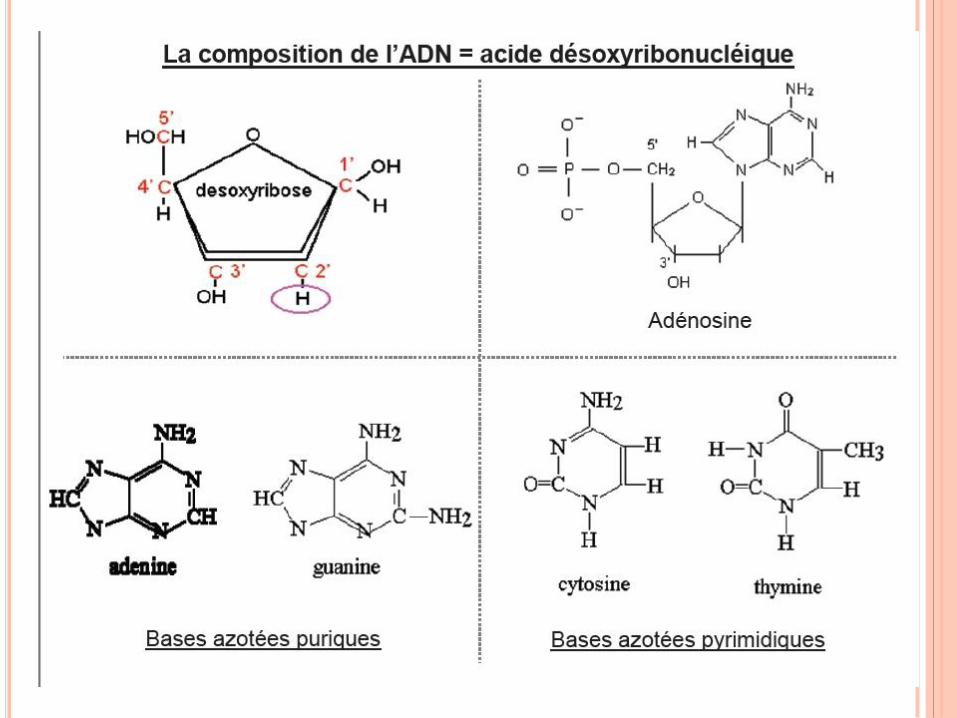
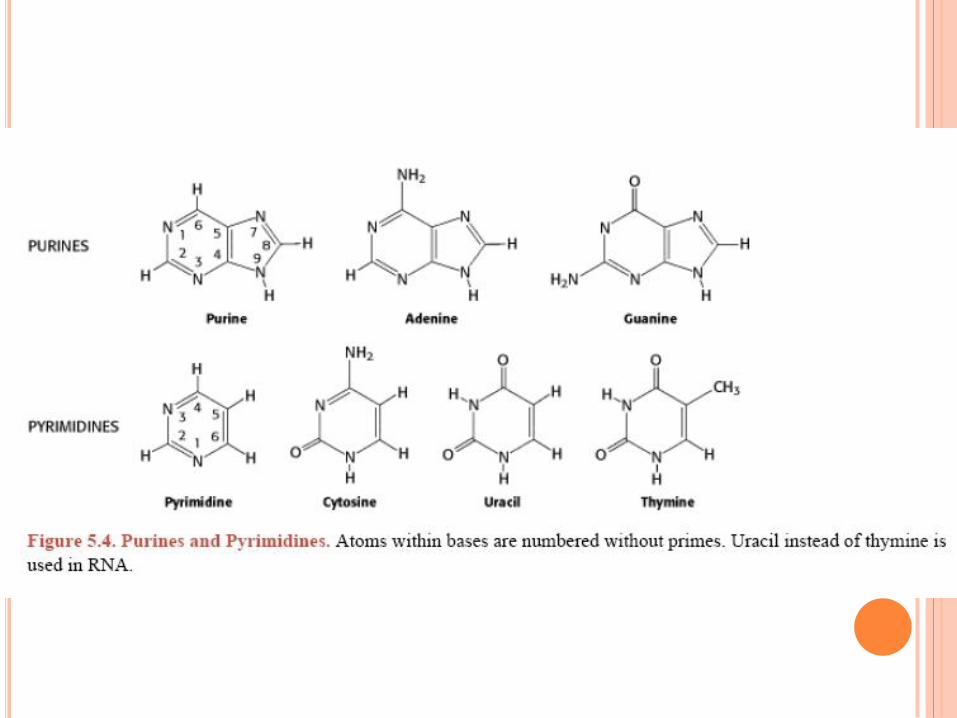

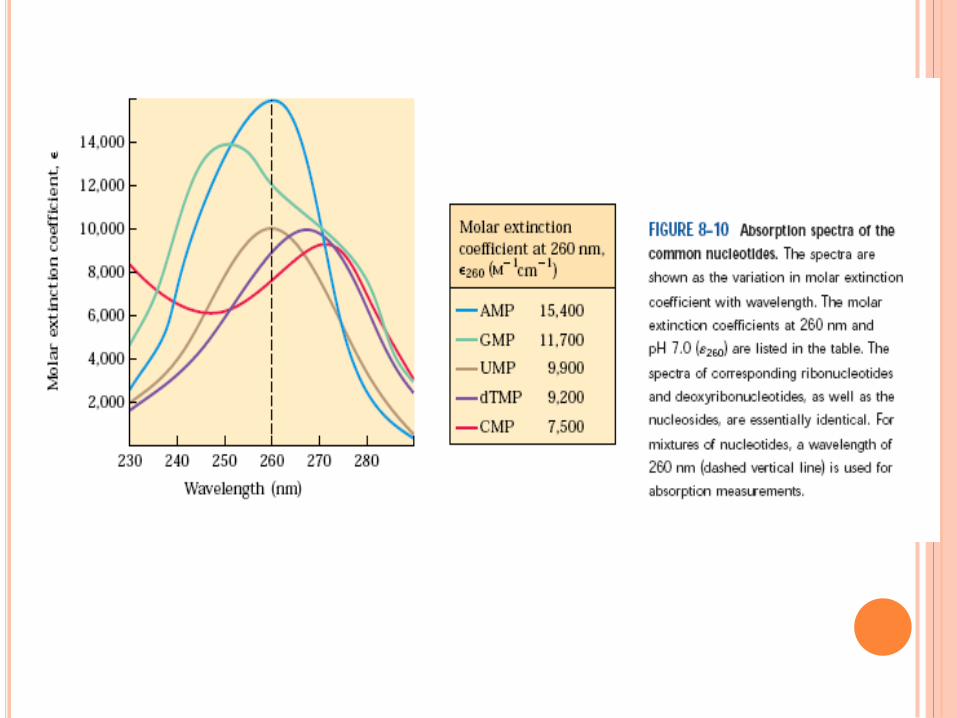
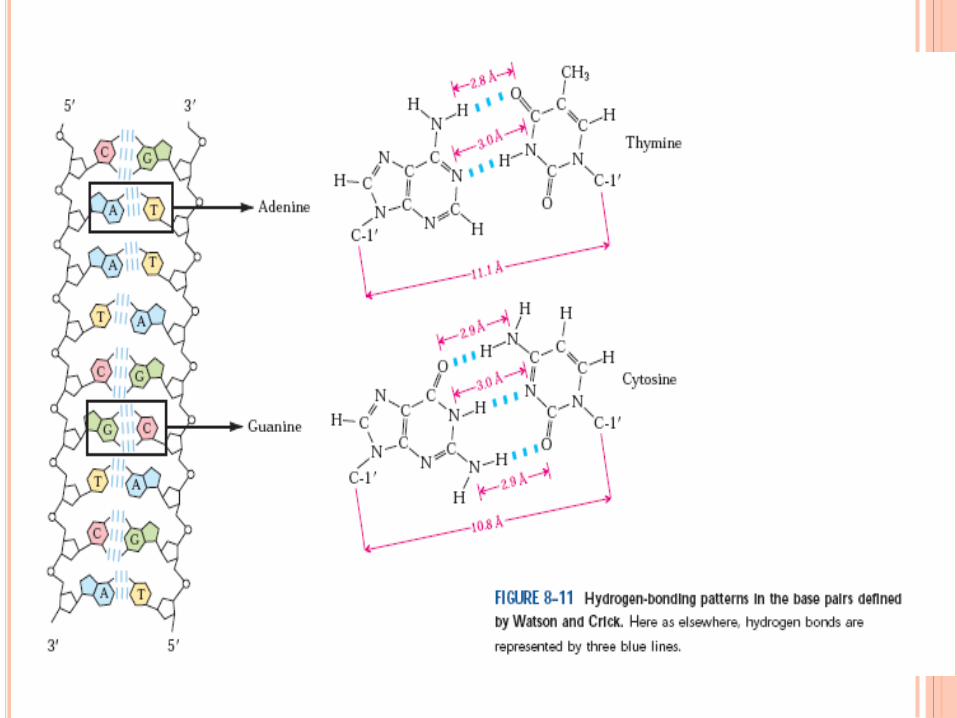
WATSON ET CRICK,1952
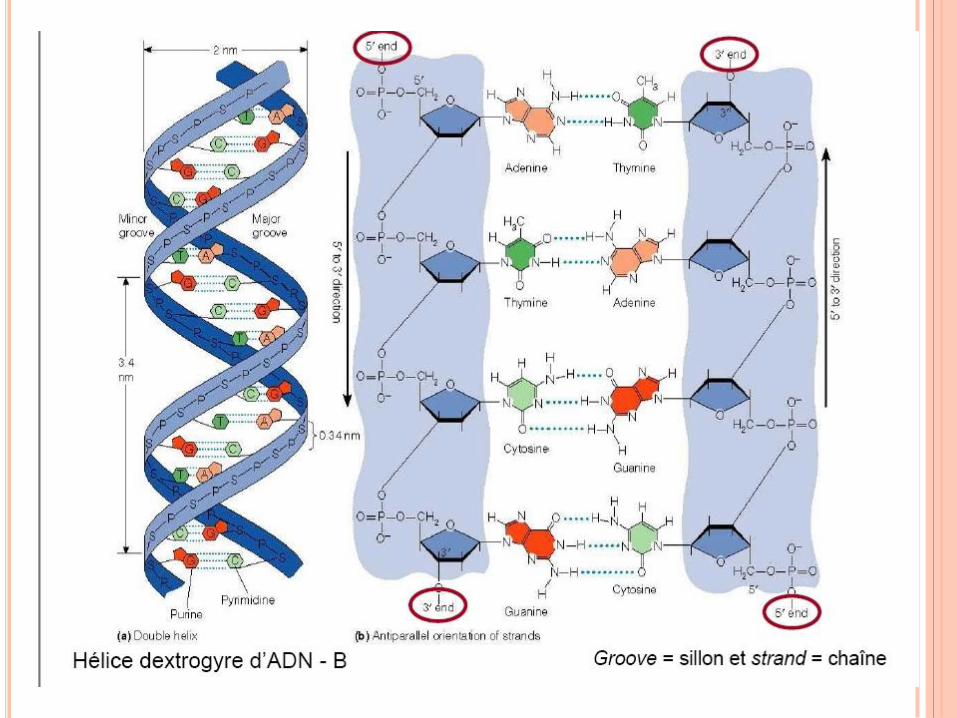
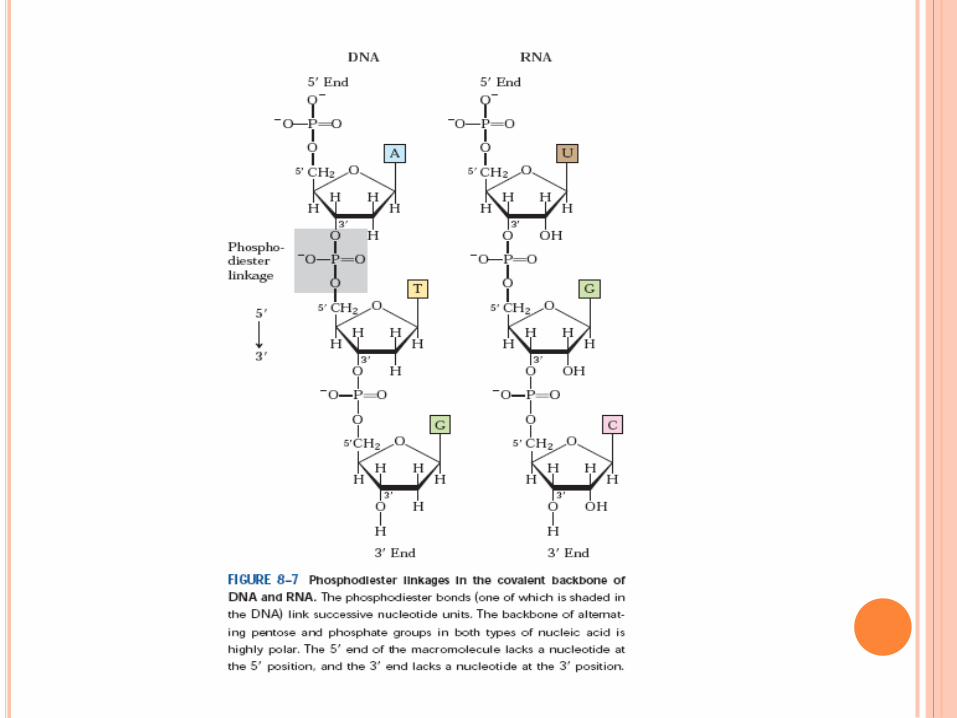

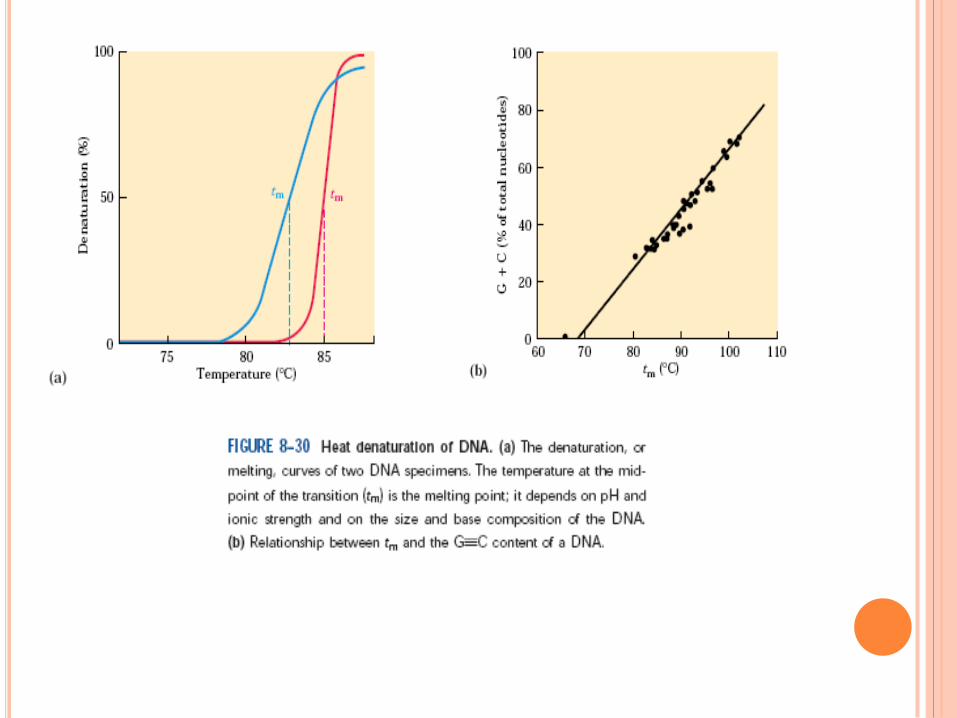
Règle de Chargaff
A = T , C = G , A + G = T + C

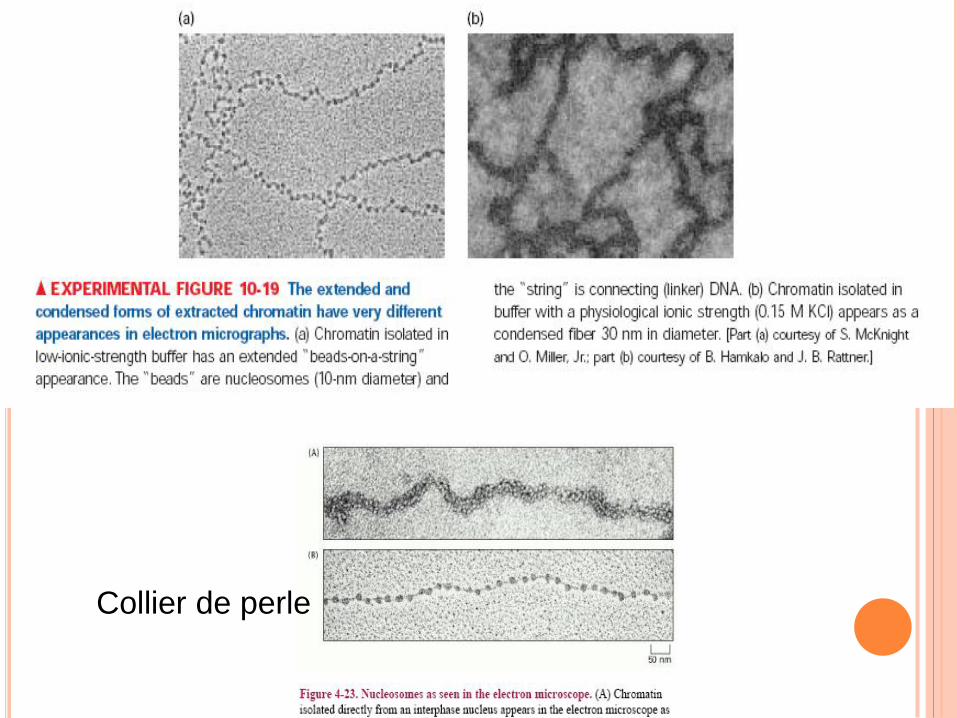
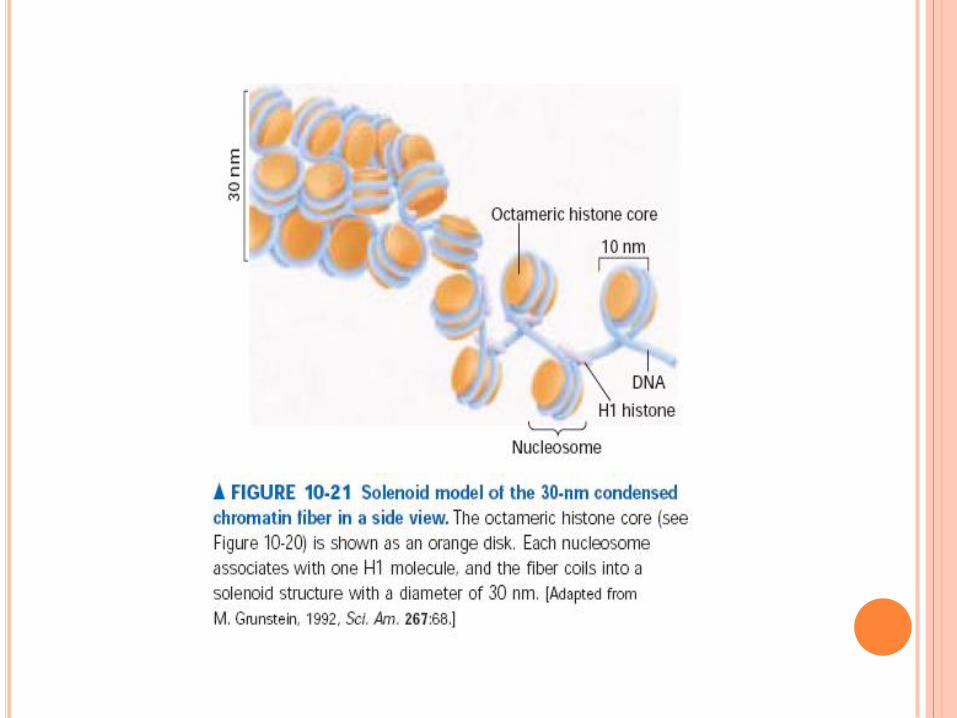
Collier de perle
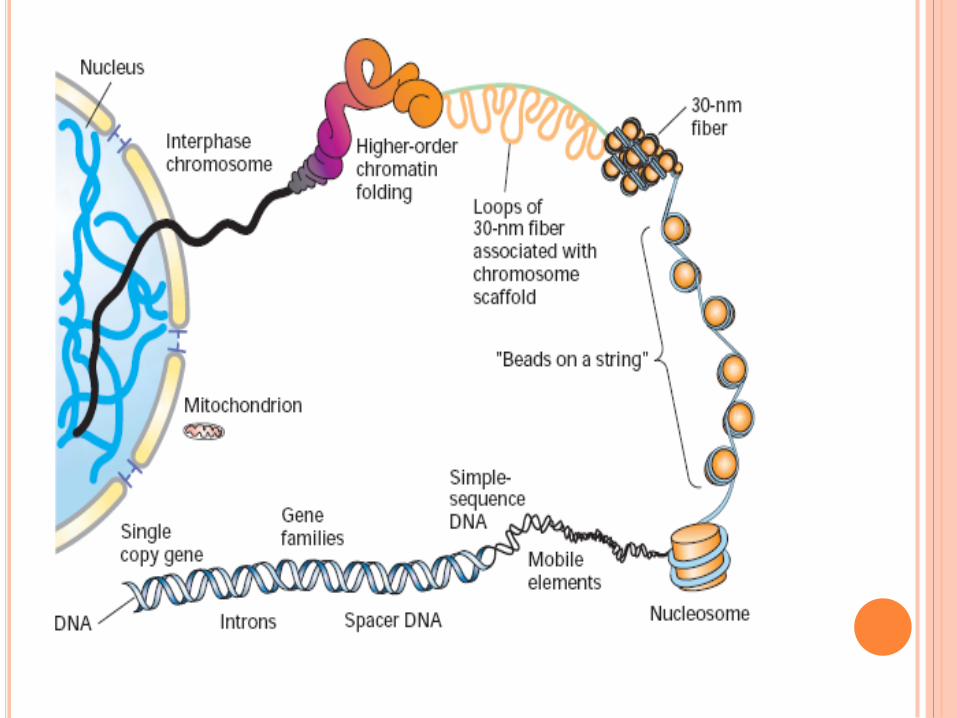
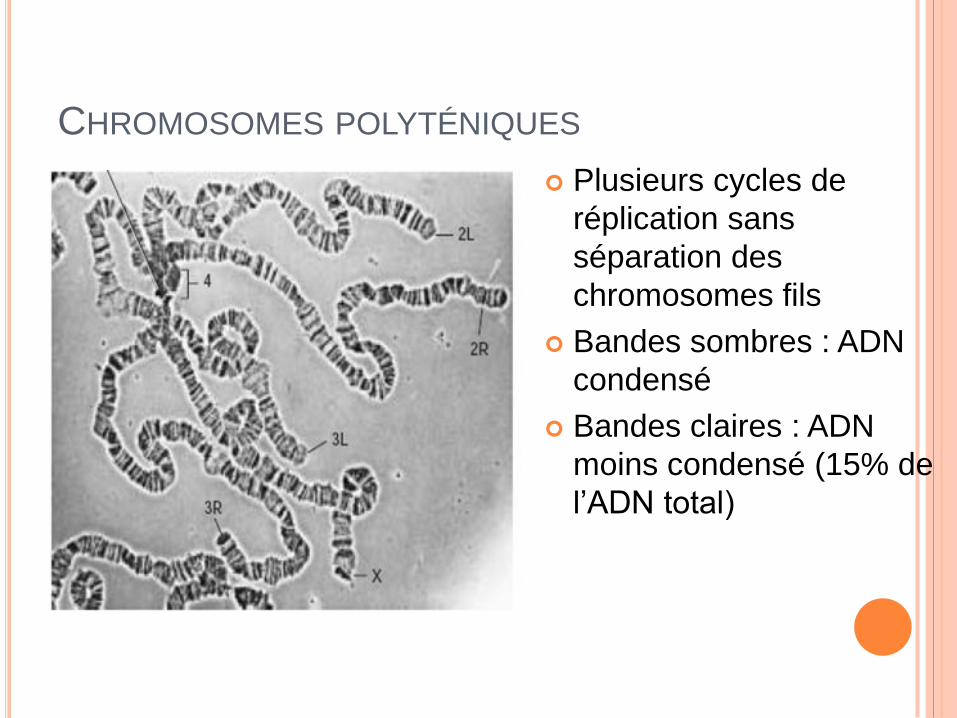
CHROMOSOMES POLYTÉNIQUES
Plusieurs cycles de
réplication sans
séparation des
chromosomes fils
Bandes sombres : ADN
condensé
Bandes claires : ADN
moins condensé (15% de
l’ADN total)


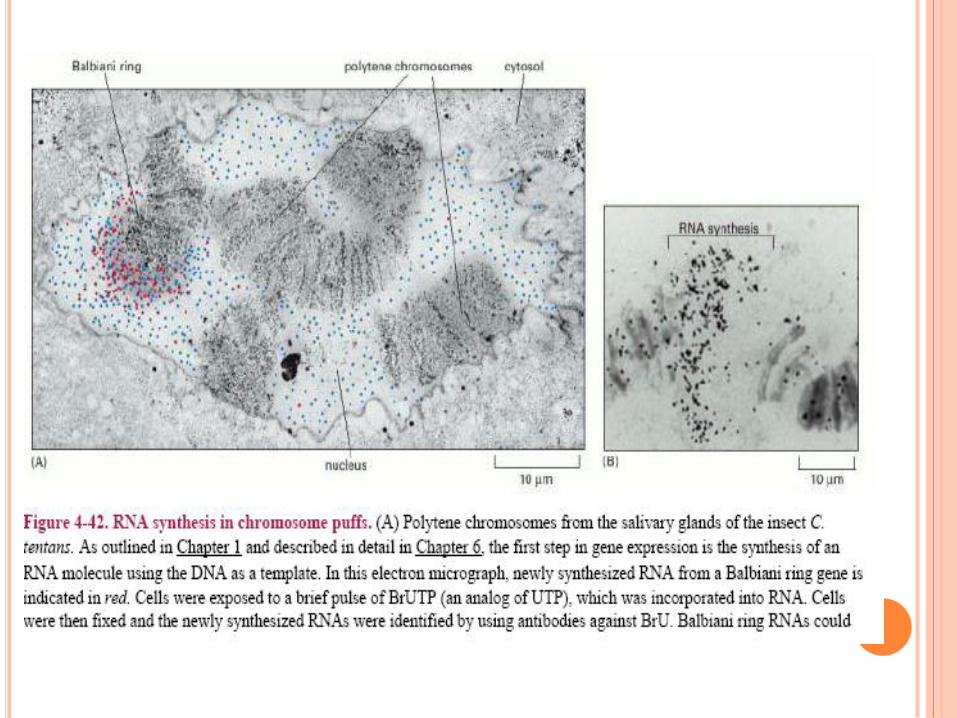
ANNEAUX DE BALBIANI OU PUFFS
Zone ou ADN est
décondensé
Lieu de synthèse ARN
intense (diapo suivante)
Puffs : gènes fonctionnels
qui varient en fonction du
temps
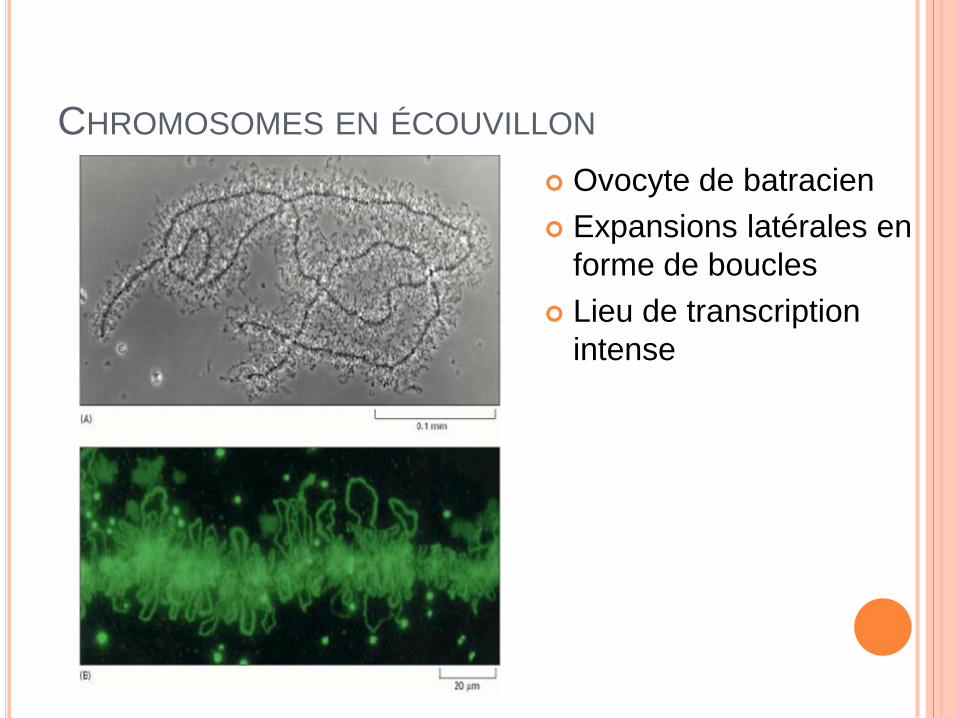
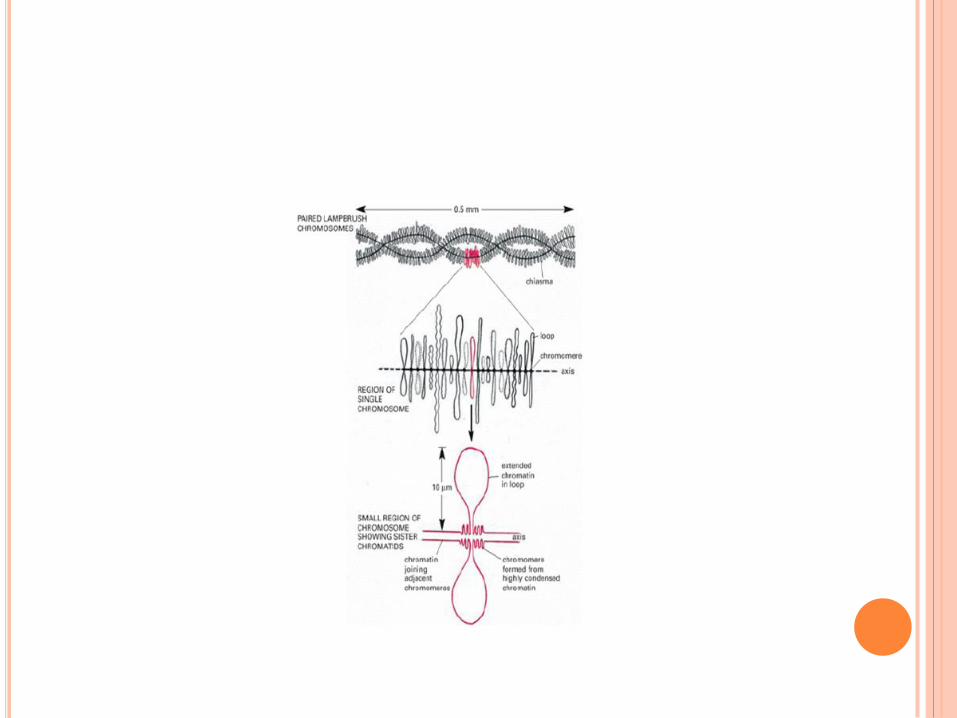
CHROMOSOMES EN ÉCOUVILLON
Ovocyte de batracien
Expansions latérales en
forme de boucles
Lieu de transcription
intense

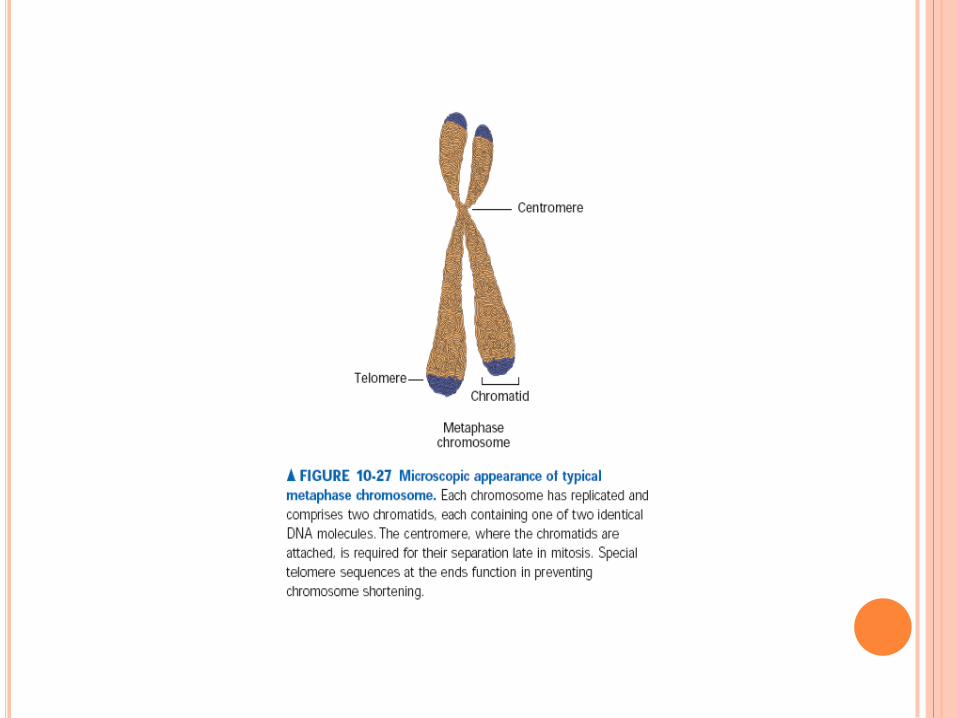
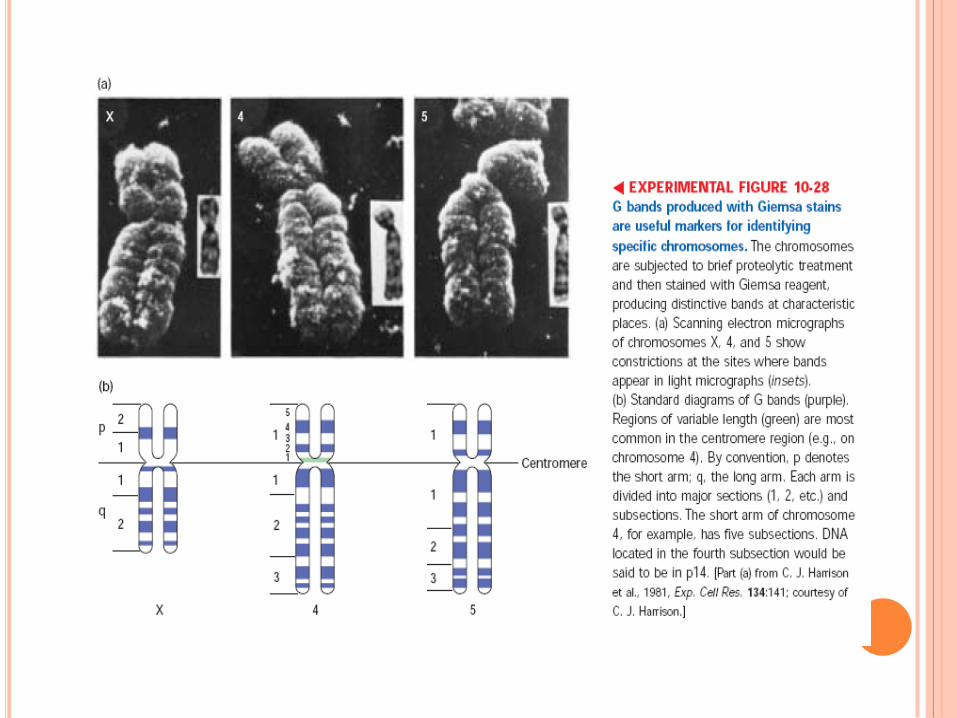
CARYOTYPE
Traitement à la colchicine qui bloque la mitose en
métaphase
Classement par ordre taille et forme
Autosome/gonosome
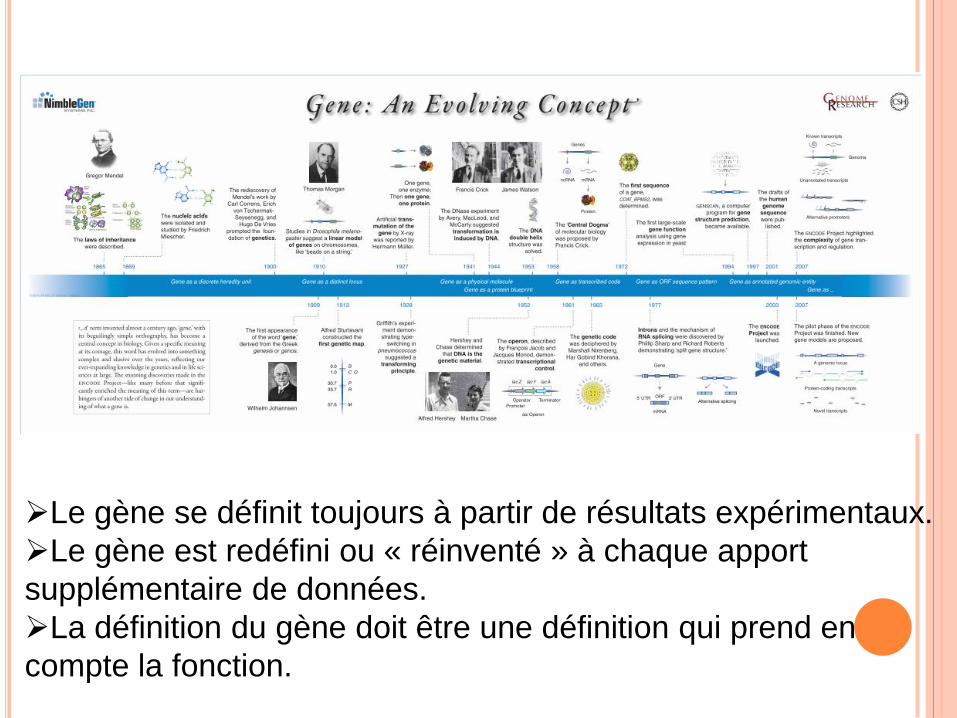
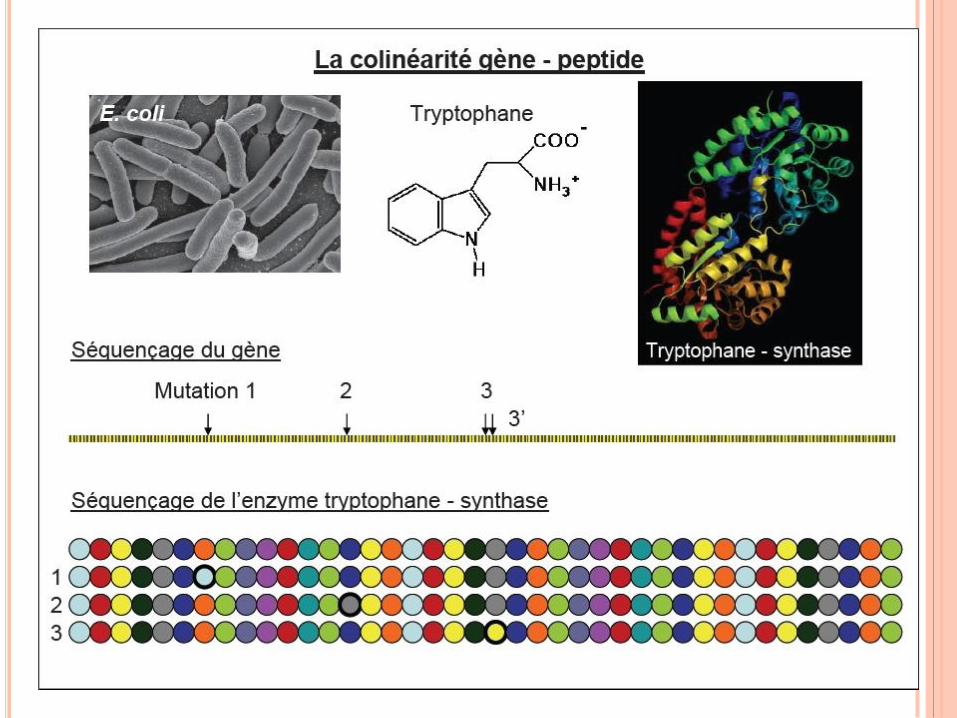
Le gène se définit toujours à partir de résultats expérimentaux.
Le gène est redéfini ou « réinventé » à chaque apport
supplémentaire de données.
La définition du gène doit être une définition qui prend en
compte la fonction.
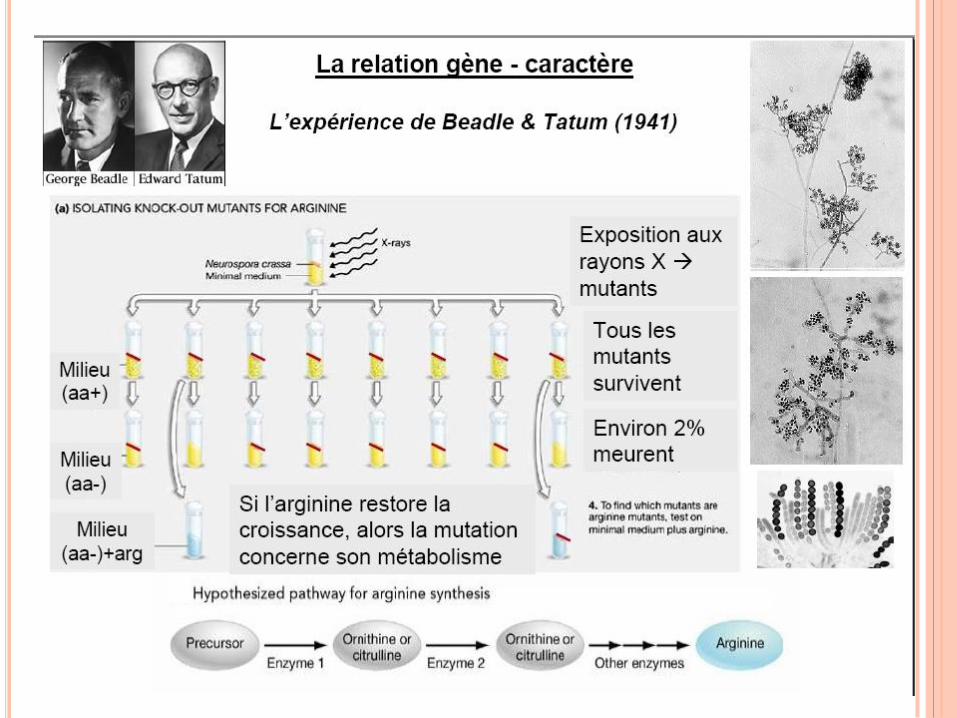
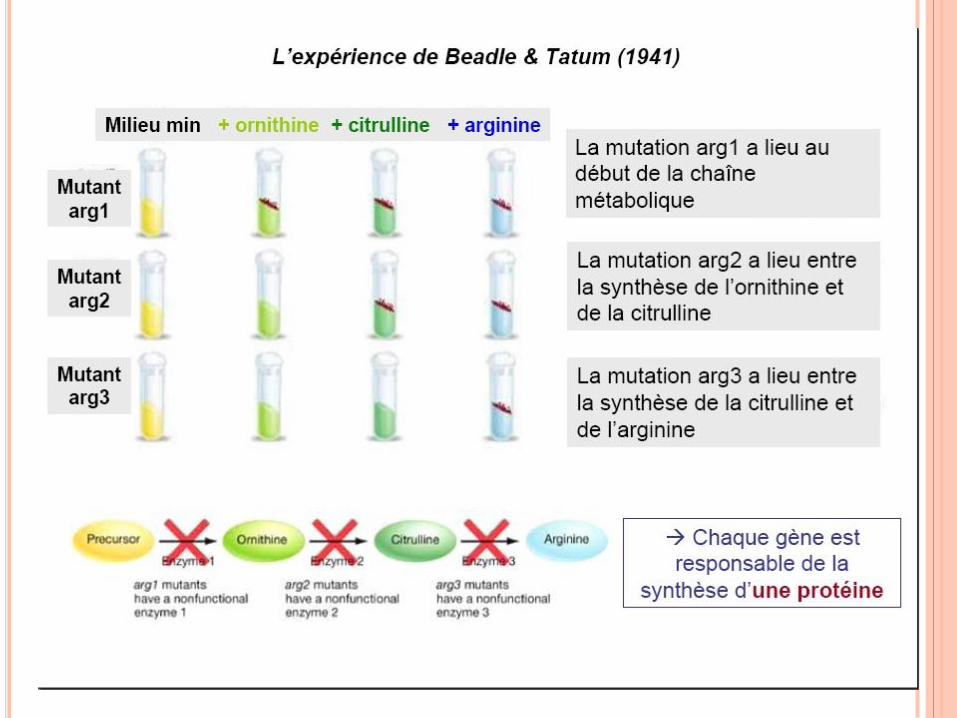
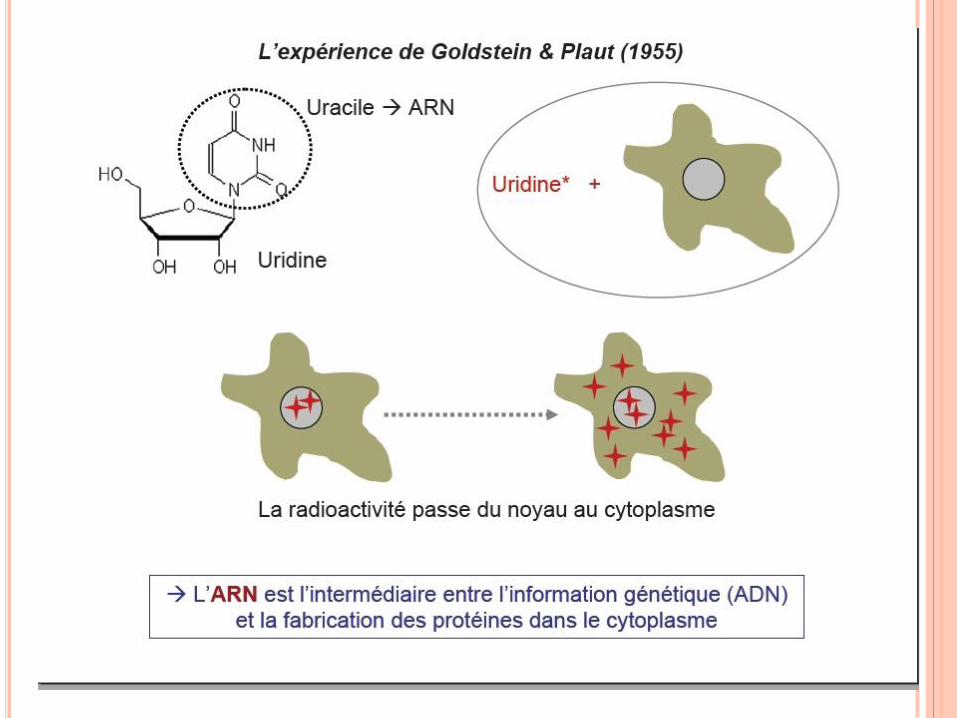
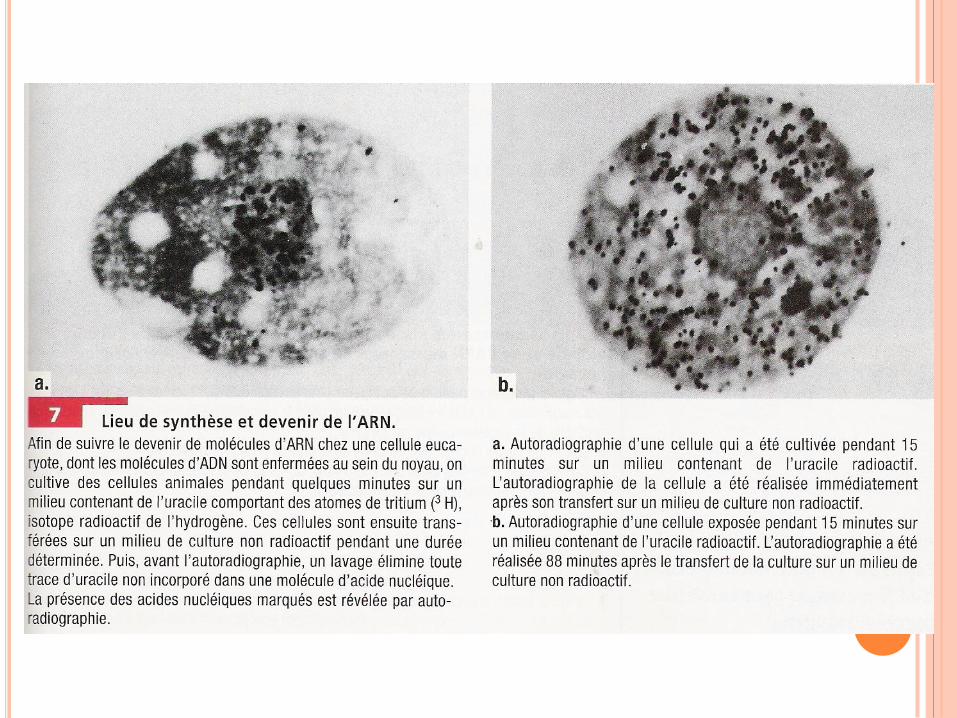

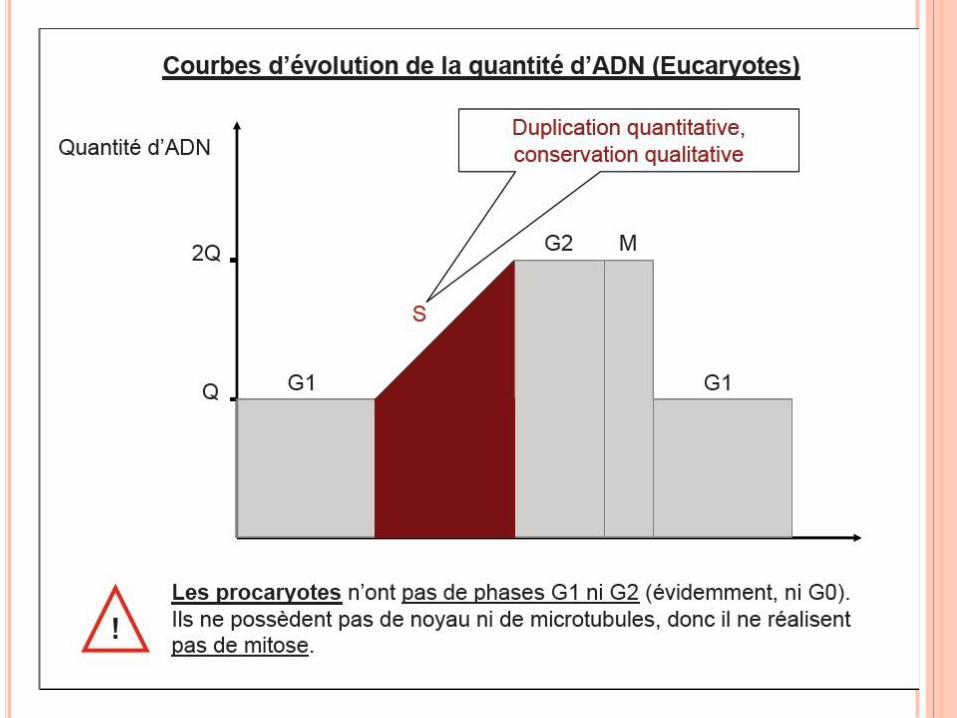
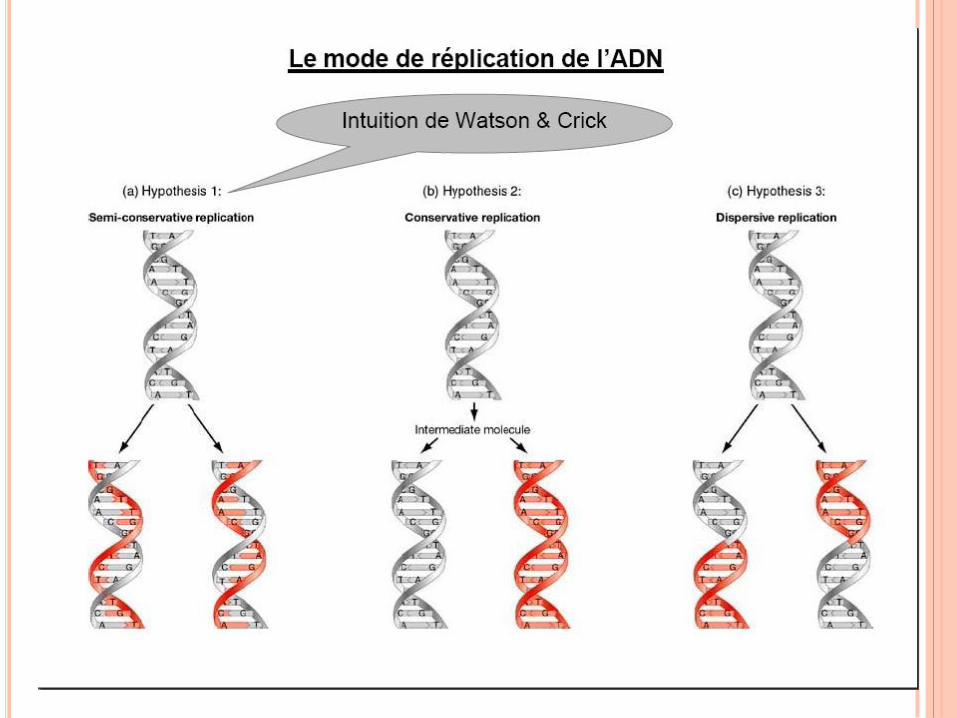
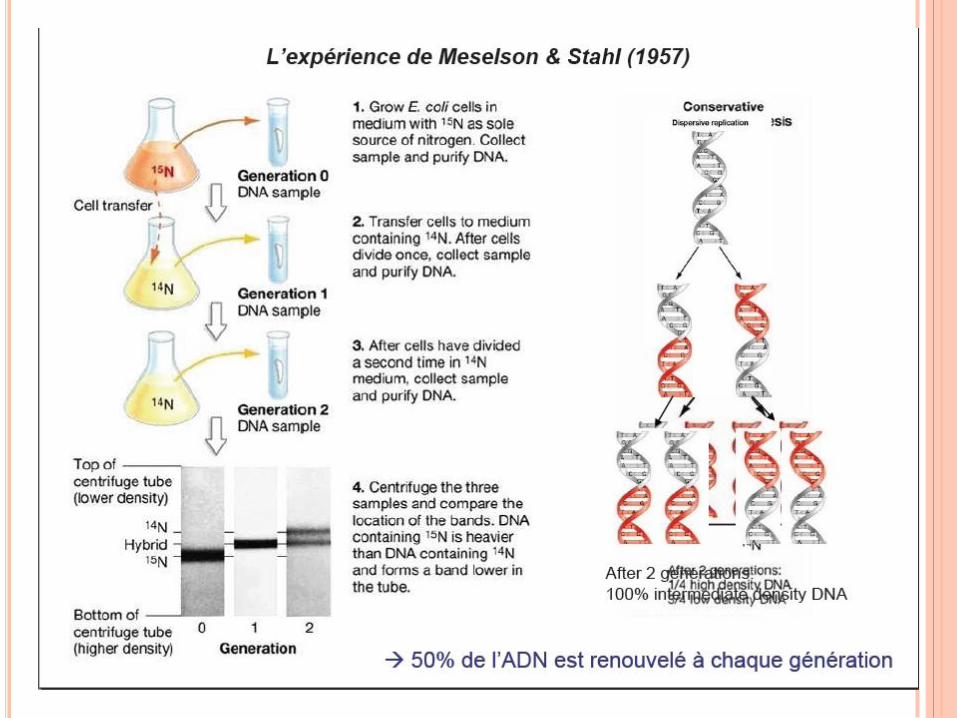
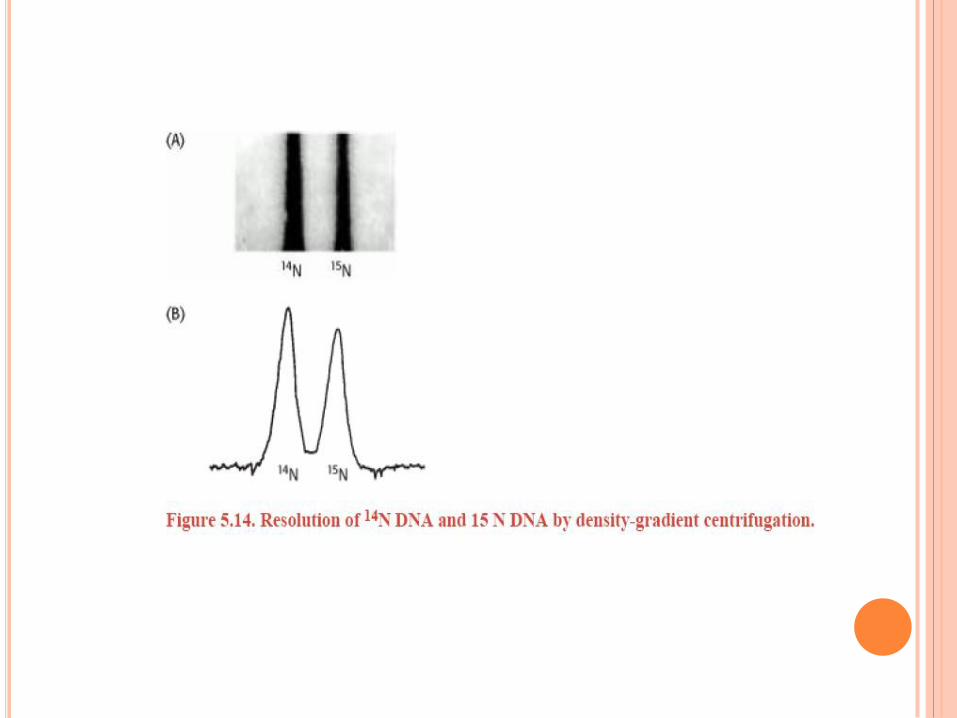
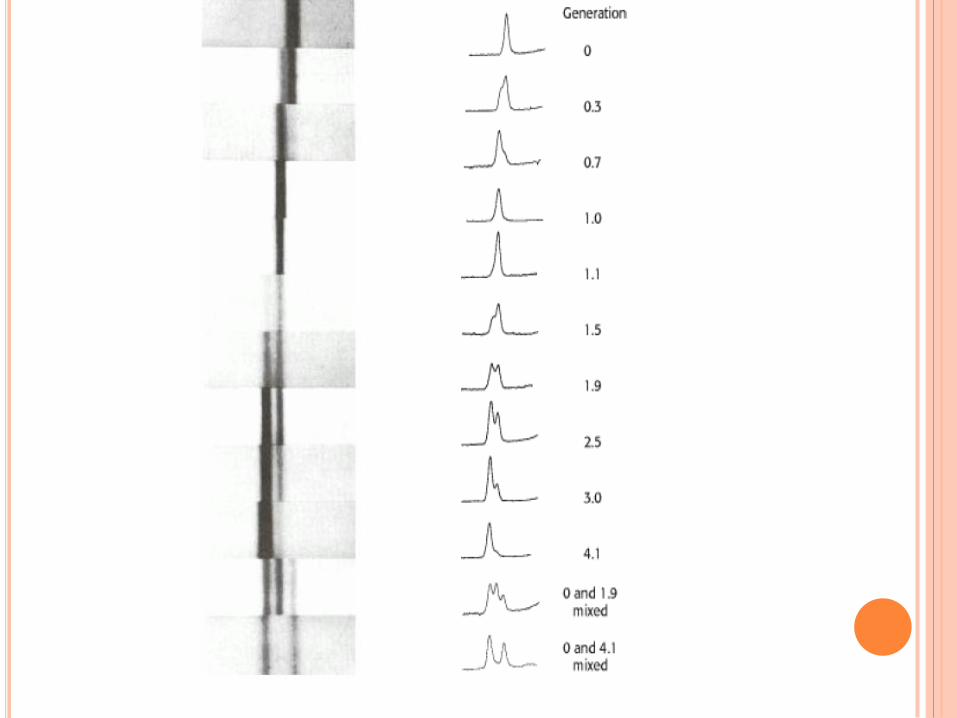
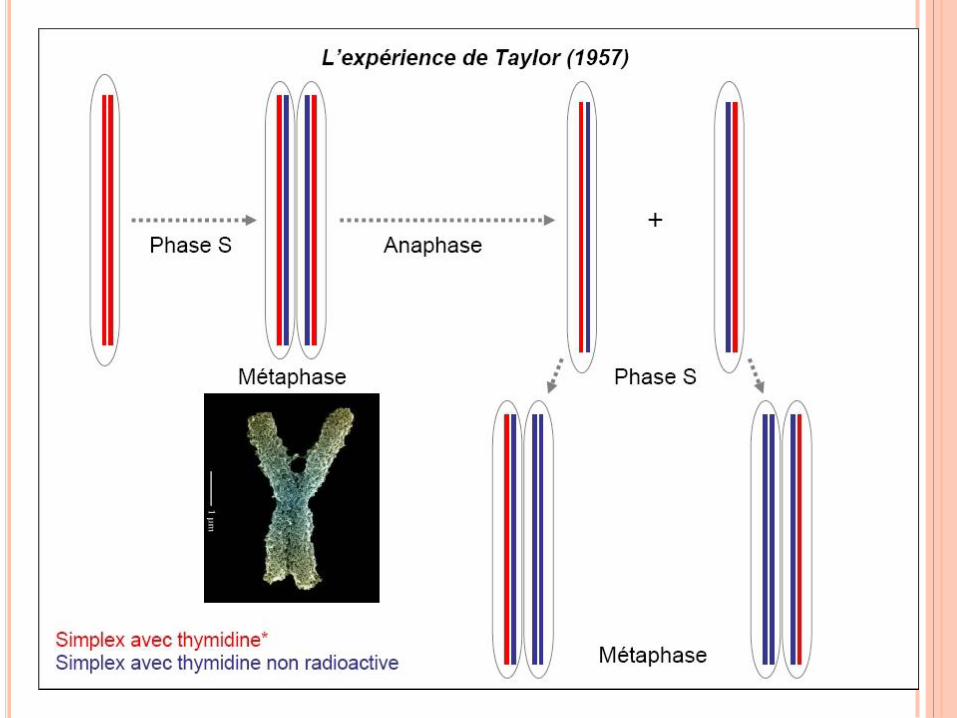
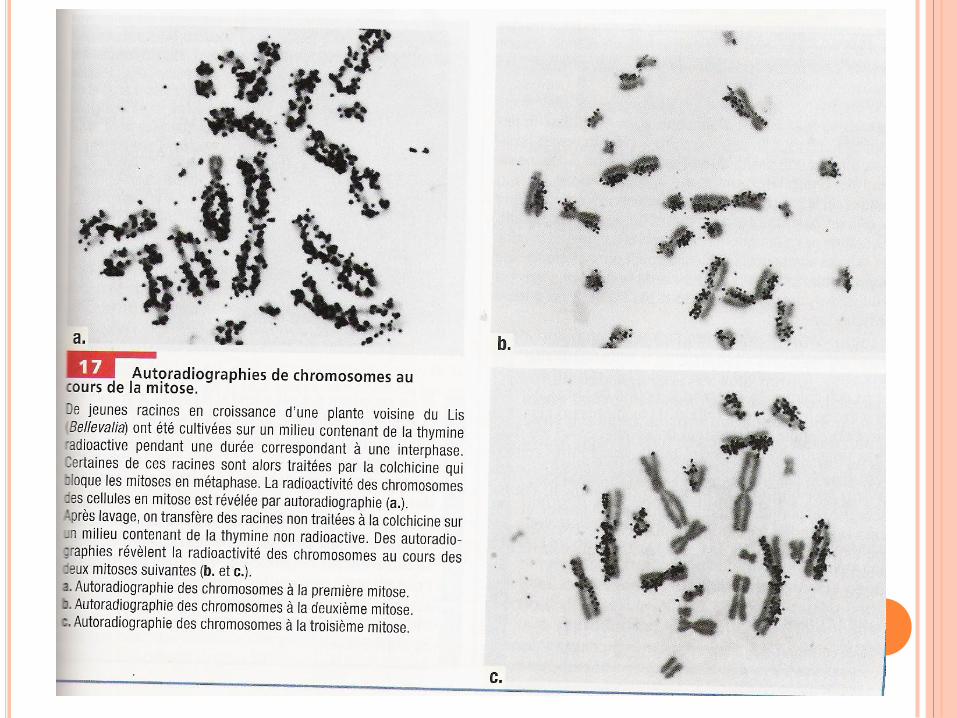
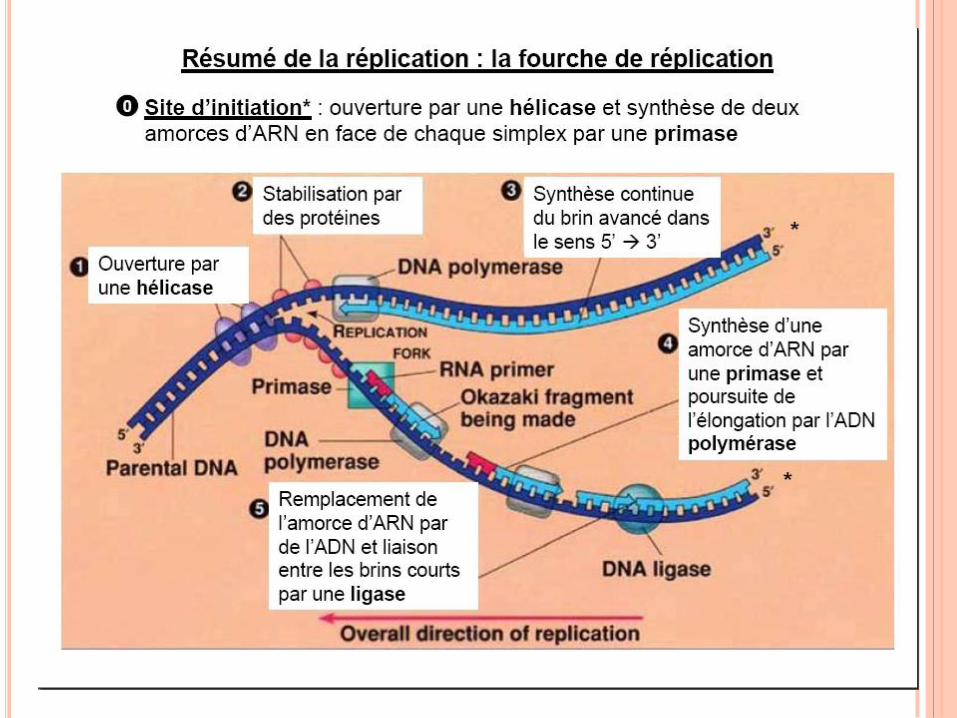
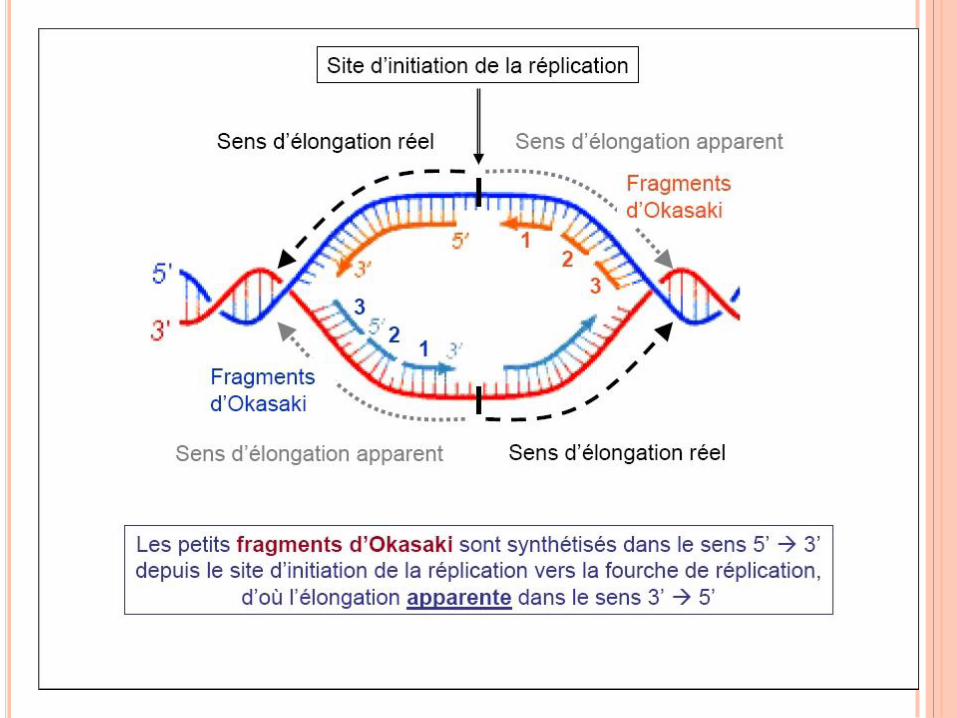
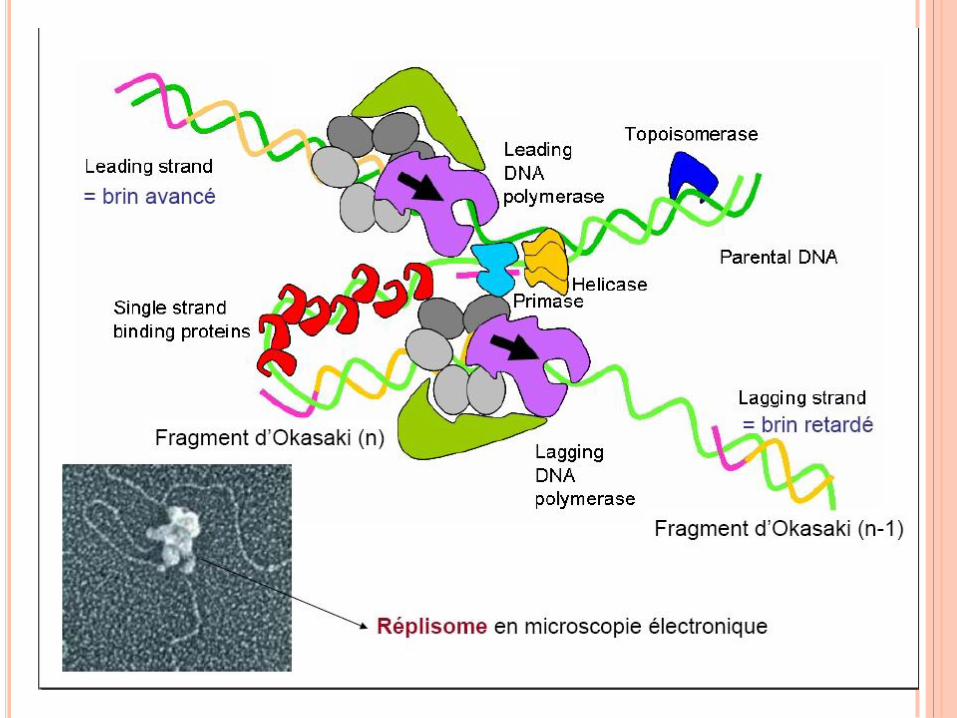
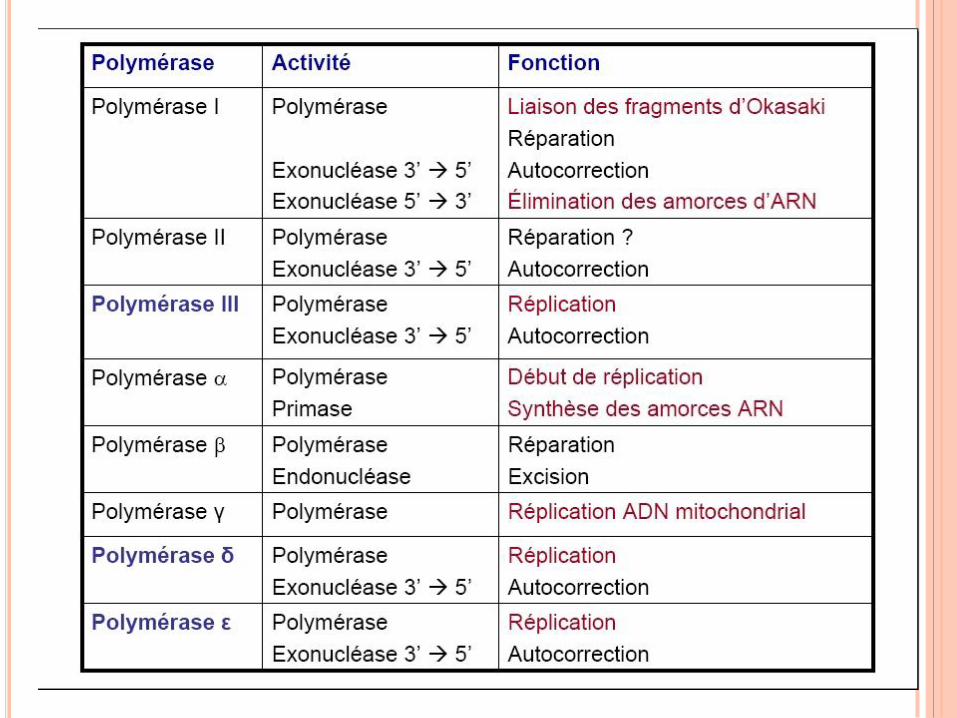
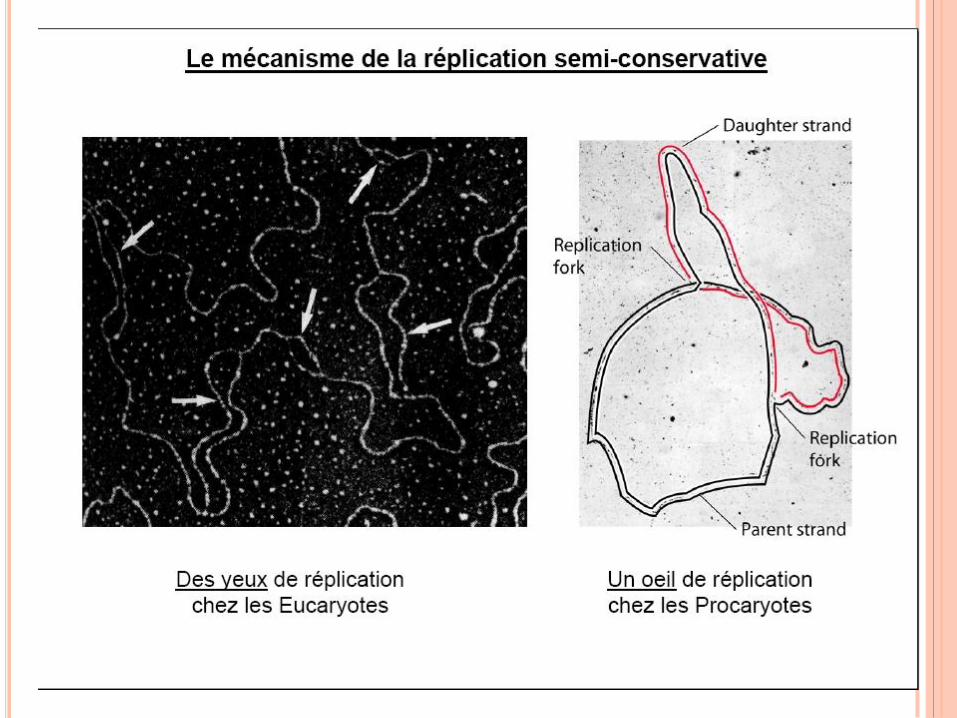
RÉPLICATION DE L’ADN

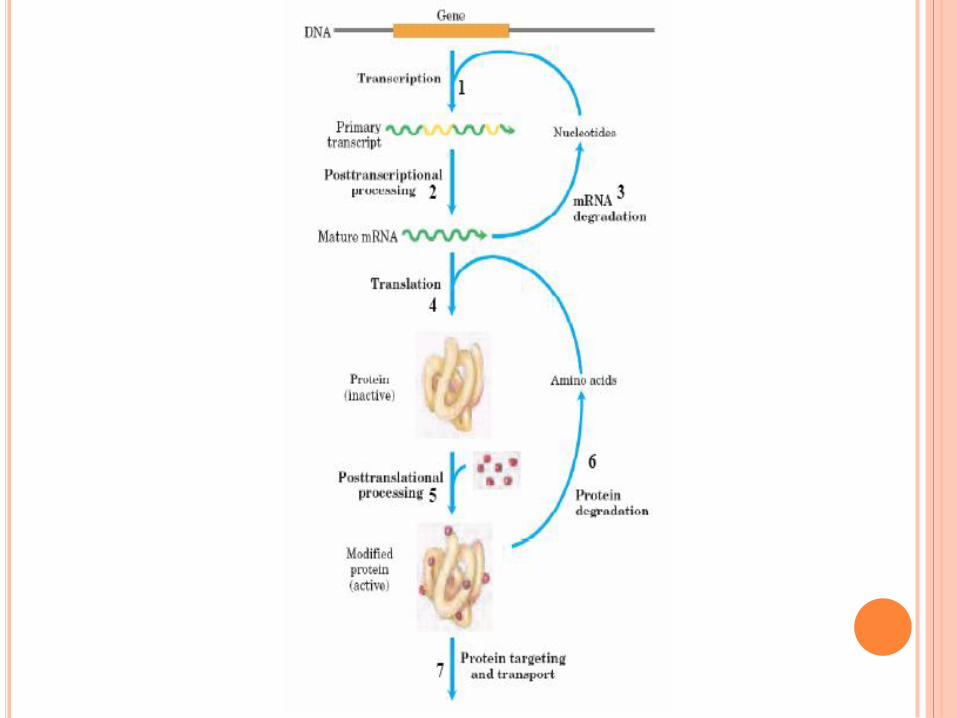
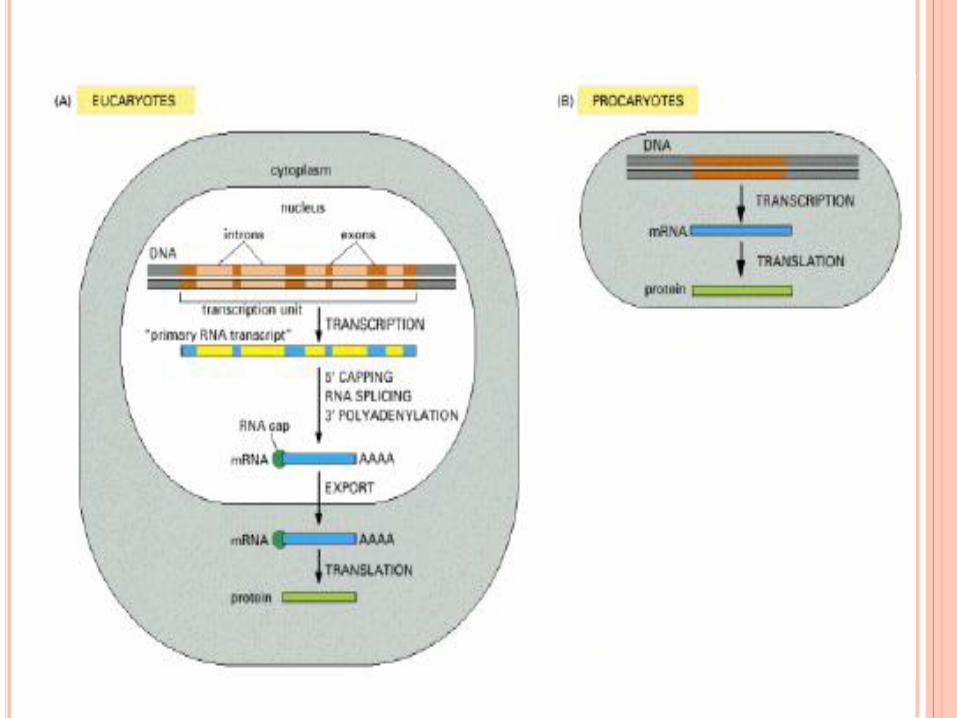
L’EXPRESSION DE
L’INFORMATION GENETIQUE ET
SON CONTROLE
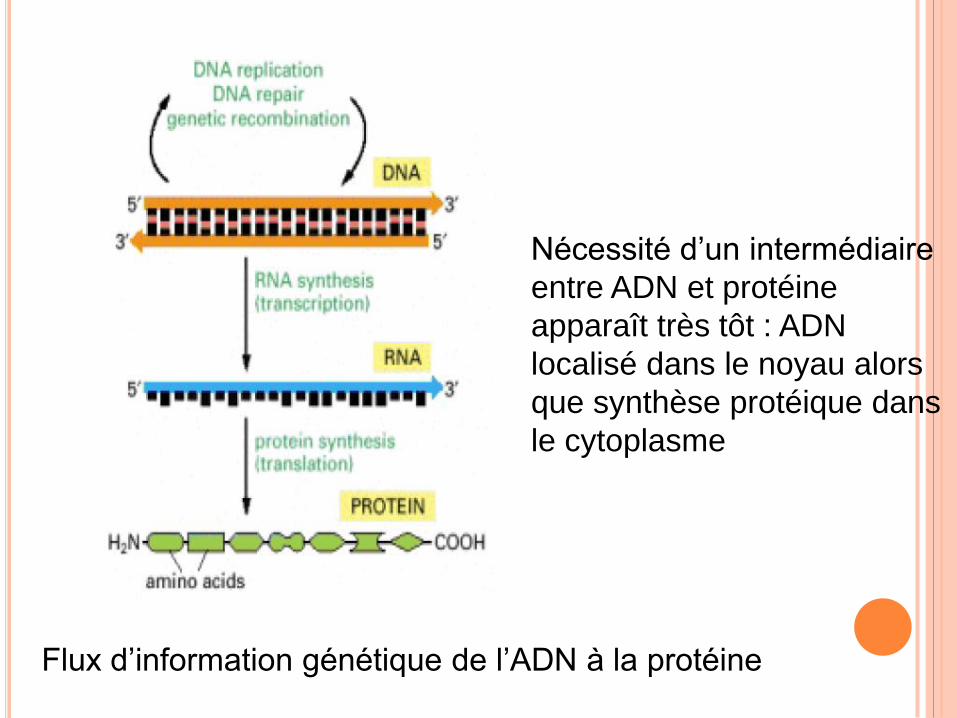
Flux d’information génétique de l’ADN à la protéine
Nécessité d’un intermédiaire
entre ADN et protéine
apparaît très tôt : ADN
localisé dans le noyau alors
que synthèse protéique dans
le cytoplasme
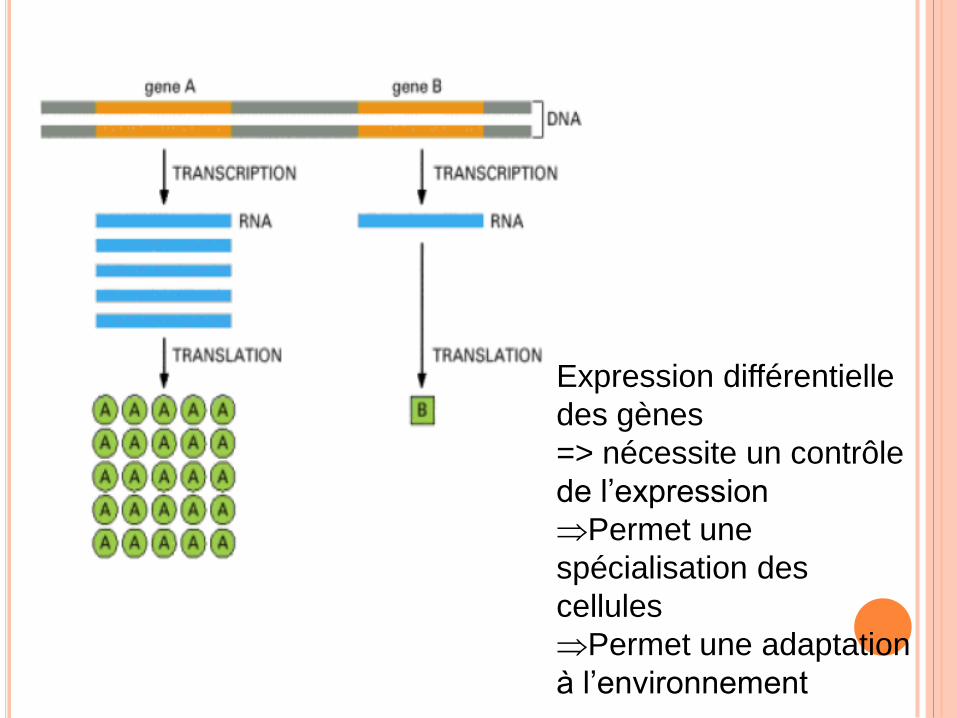
Expression différentielle
des gènes
=> nécessite un contrôle
de l’expression
Permet une
spécialisation des
cellules
Permet une adaptation
à l’environnement
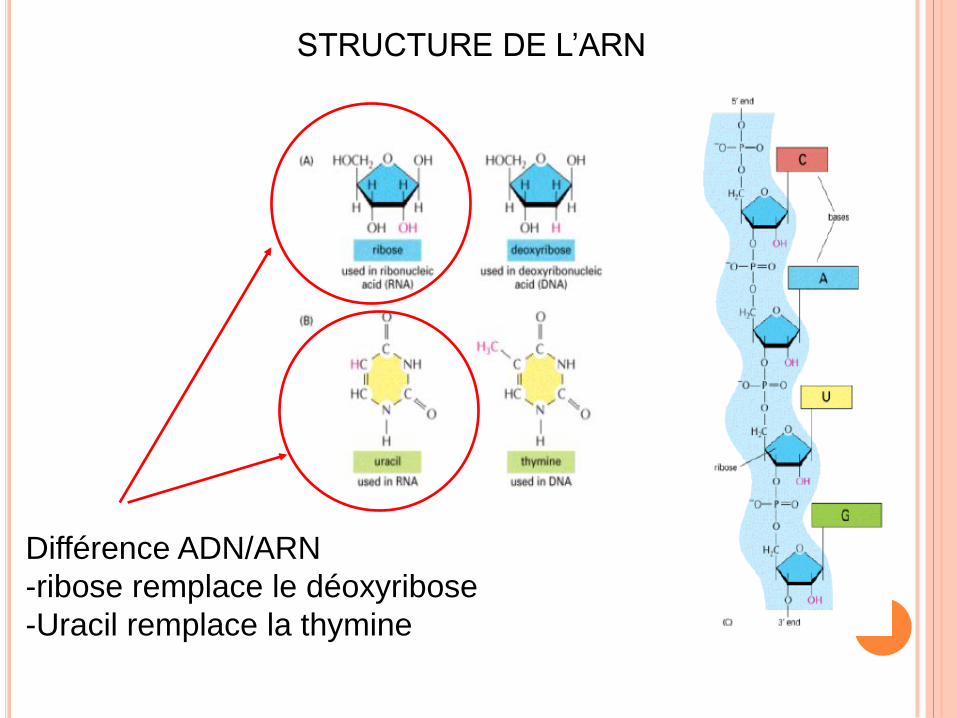
STRUCTURE DE L’ARN
Différence ADN/ARN
-ribose remplace le déoxyribose
-Uracil remplace la thymine
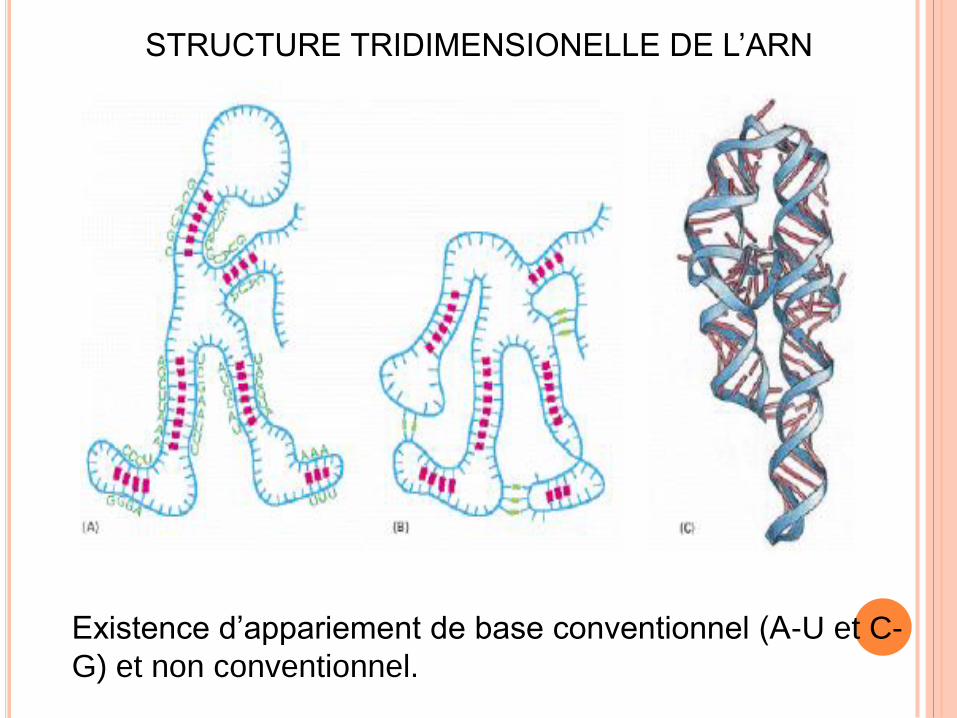
STRUCTURE TRIDIMENSIONELLE DE L’ARN
Existence d’appariement de base conventionnel (A-U et C-
G) et non conventionnel.

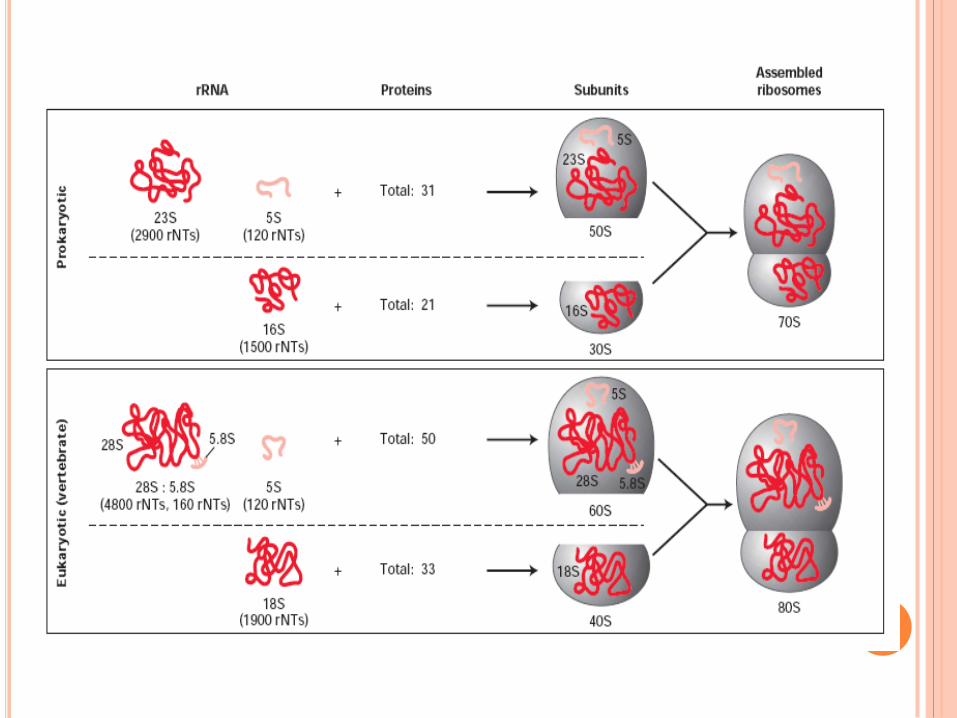
Les ARNr et les ARNt représentent 95% des ARN chez E Coli

EXPRESSION DE L’INFORMATION
GÉNÉTIQUE CHEZ LES PROCARYOTES
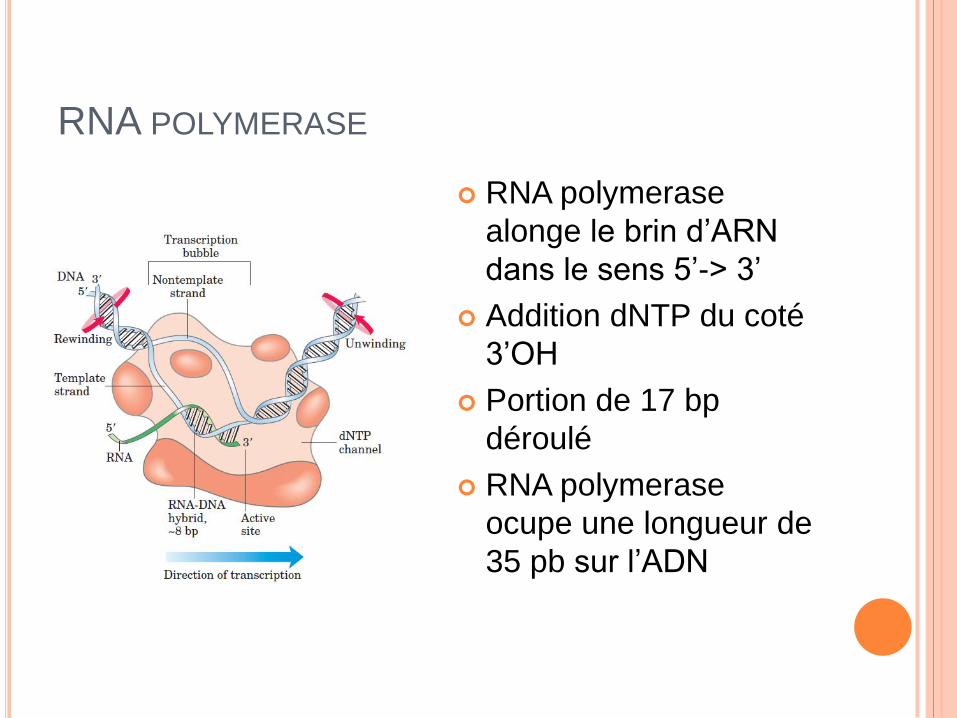
RNA POLYMERASE
RNA polymerase
alonge le brin d’ARN
dans le sens 5’-> 3’
Addition dNTP du coté
3’OH
Portion de 17 bp
déroulé
RNA polymerase
ocupe une longueur de
35 pb sur l’ADN

Les ARN polymérases catalysent la synthèse de l’ARN à
partir d’une matrice d’ADN
Une seule ARN
polymerase chez les
procaryotes mais
existence de plusieurs
facteurs nécessaires à
l’initiation de la
transcription : les facteurs
sigma
Facteur sigma 70 permet
la synthèse des ARNm par
ex
Facteur sigma reconnait
séquence signal et recrute
cœur polymérase

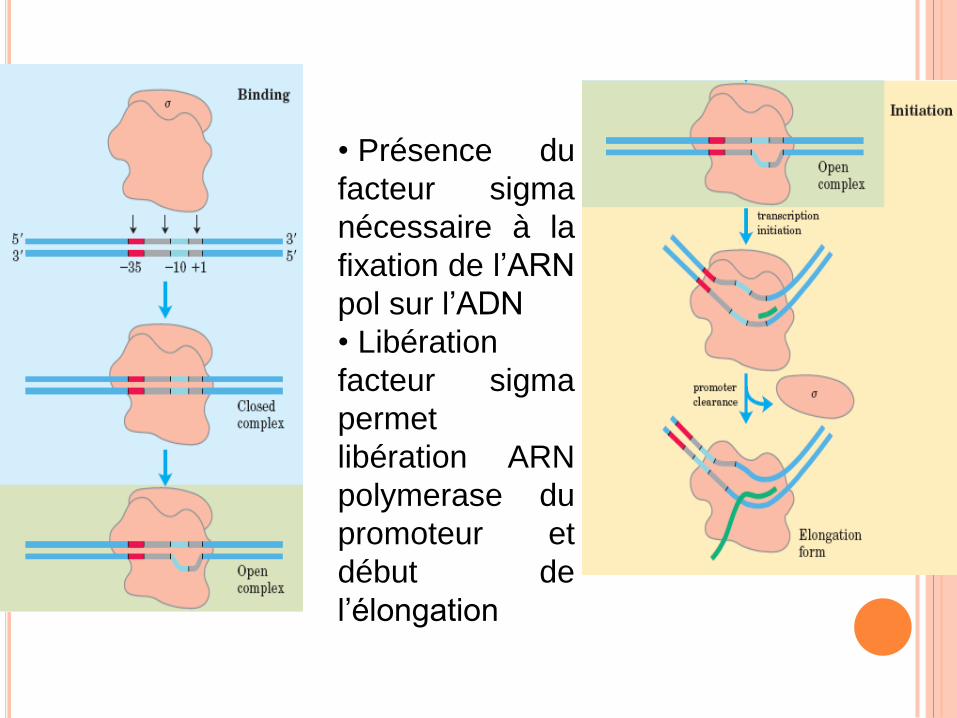
• Présence du
facteur sigma
nécessaire à la
fixation de l’ARN
pol sur l’ADN
• Libération
facteur sigma
permet
libération ARN
polymerase du
promoteur et
début de
l’élongation

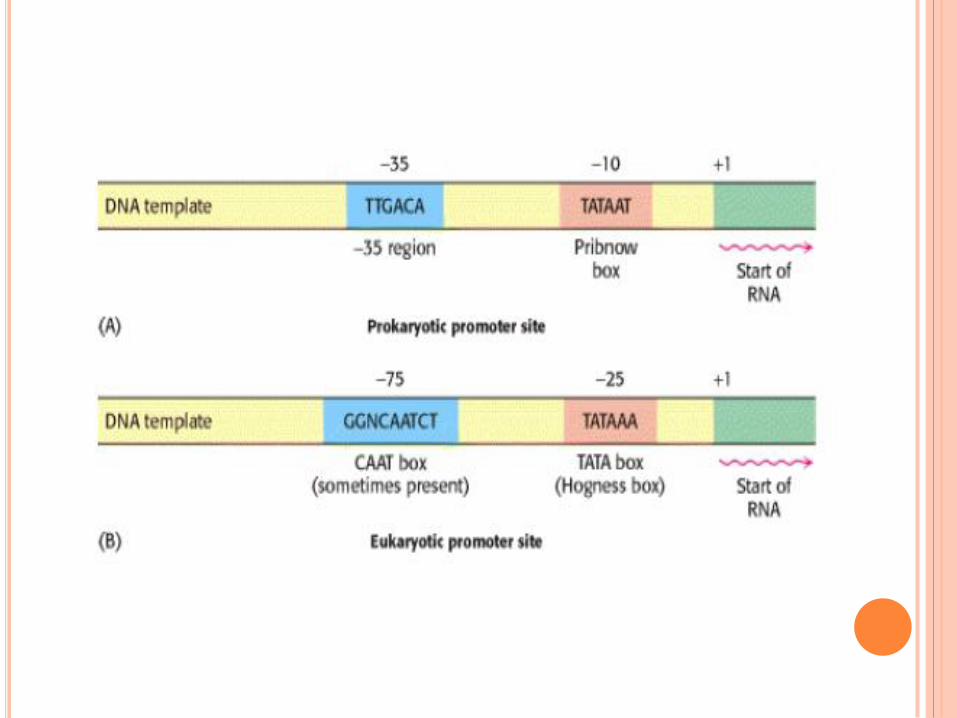
PROMOTEURS
Région –10 (boite TATA ou boite Pribnow) et –35 retrouvées dans
tous les gènes bactériens : site de fixation des facteurs sigma.
Séquence UP (upstream promoter) : seulement dans certains gènes
hautement exprimés => fixation SU alpha
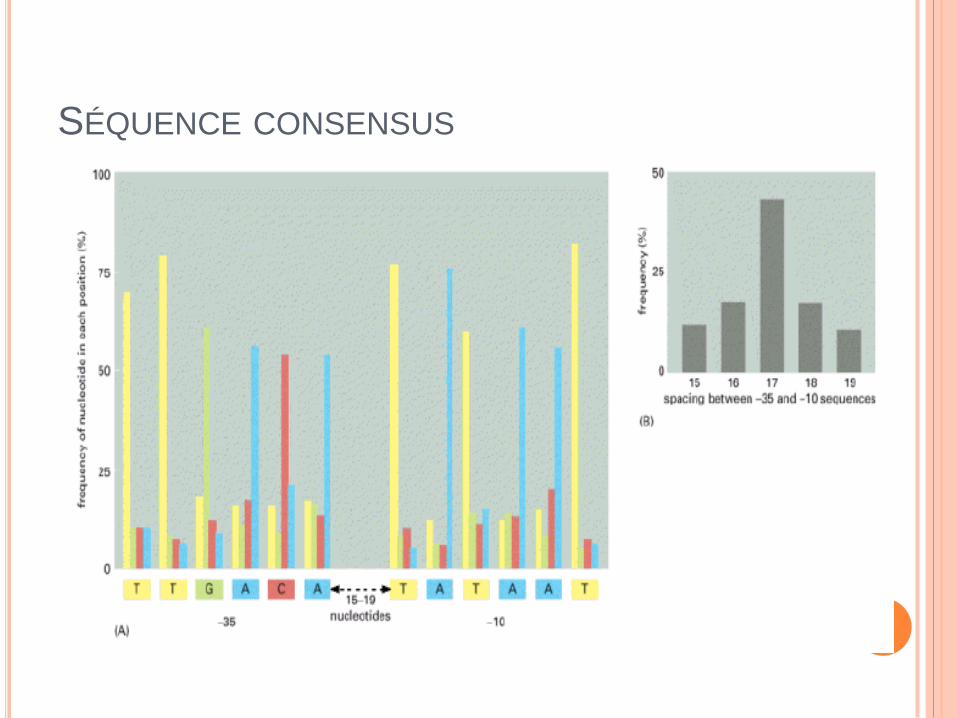
SÉQUENCE CONSENSUS
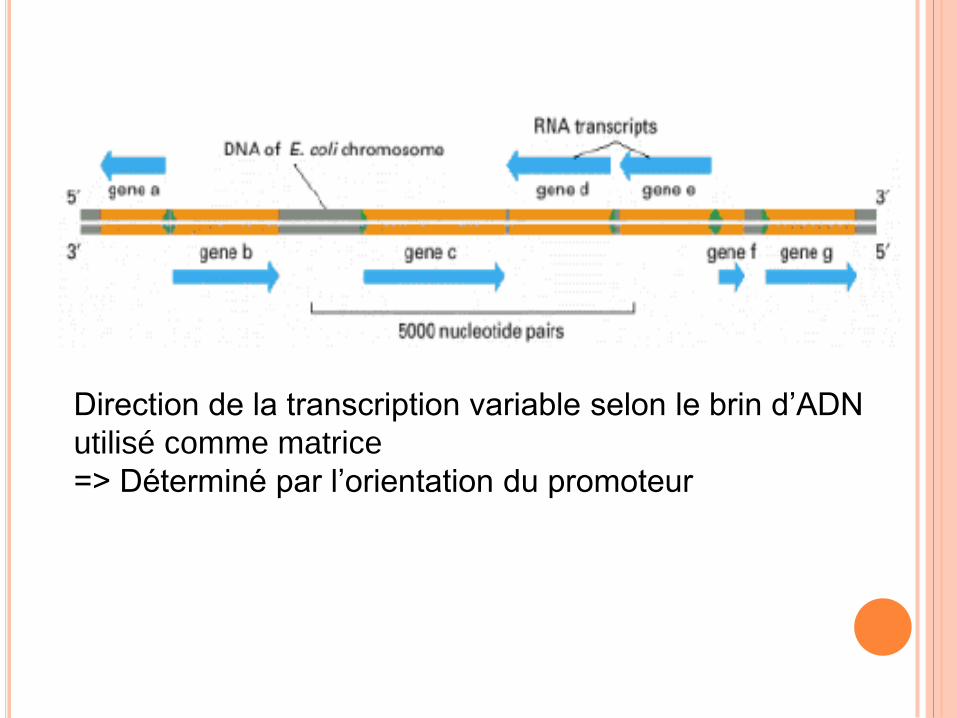
Direction de la transcription variable selon le brin d’ADN
utilisé comme matrice
=> Déterminé par l’orientation du promoteur
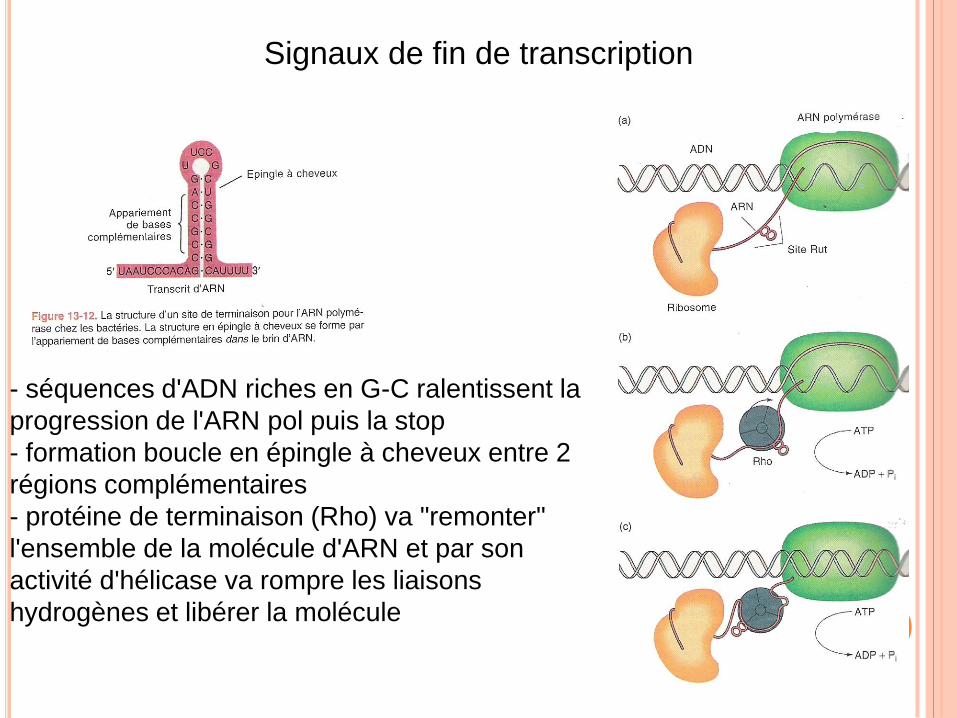
Signaux de fin de transcription
- séquences d'ADN riches en G-C ralentissent la
progression de l'ARN pol puis la stop
- formation boucle en épingle à cheveux entre 2
régions complémentaires
- protéine de terminaison (Rho) va "remonter"
l'ensemble de la molécule d'ARN et par son
activité d'hélicase va rompre les liaisons
hydrogènes et libérer la molécule
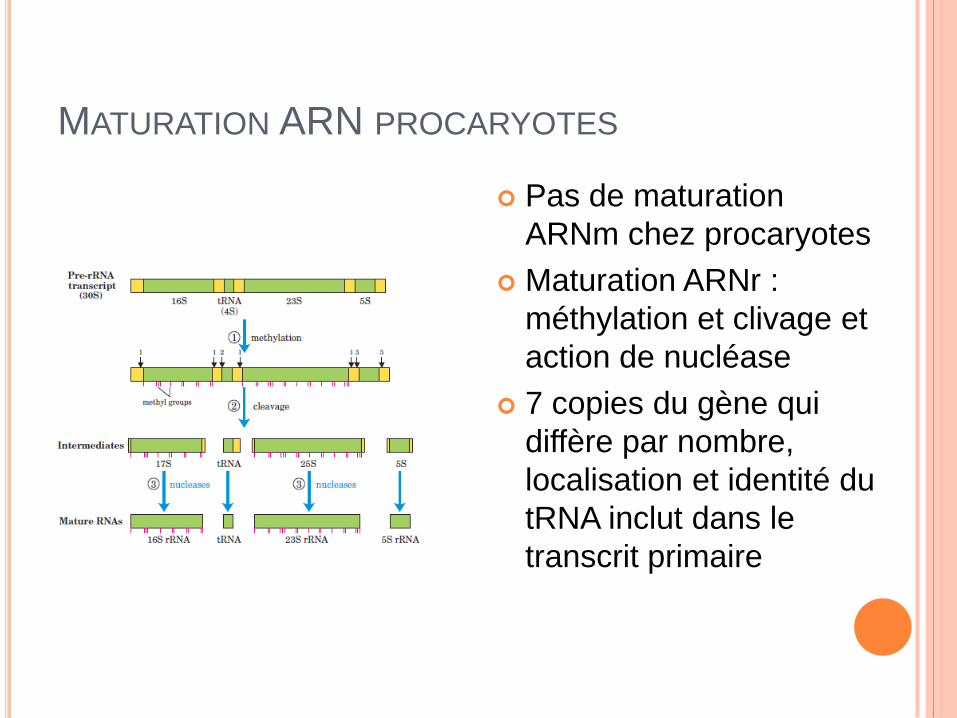
MATURATION ARN PROCARYOTES
Pas de maturation
ARNm chez procaryotes
Maturation ARNr :
méthylation et clivage et
action de nucléase
7 copies du gène qui
diffère par nombre,
localisation et identité du
tRNA inclut dans le
transcrit primaire
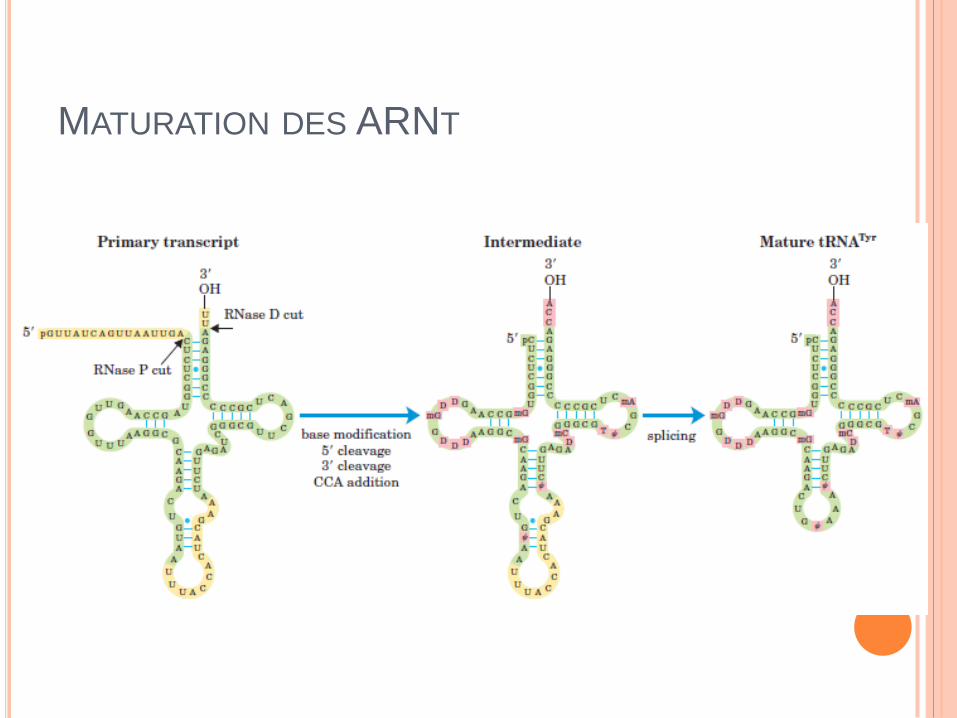
MATURATION DES ARNT

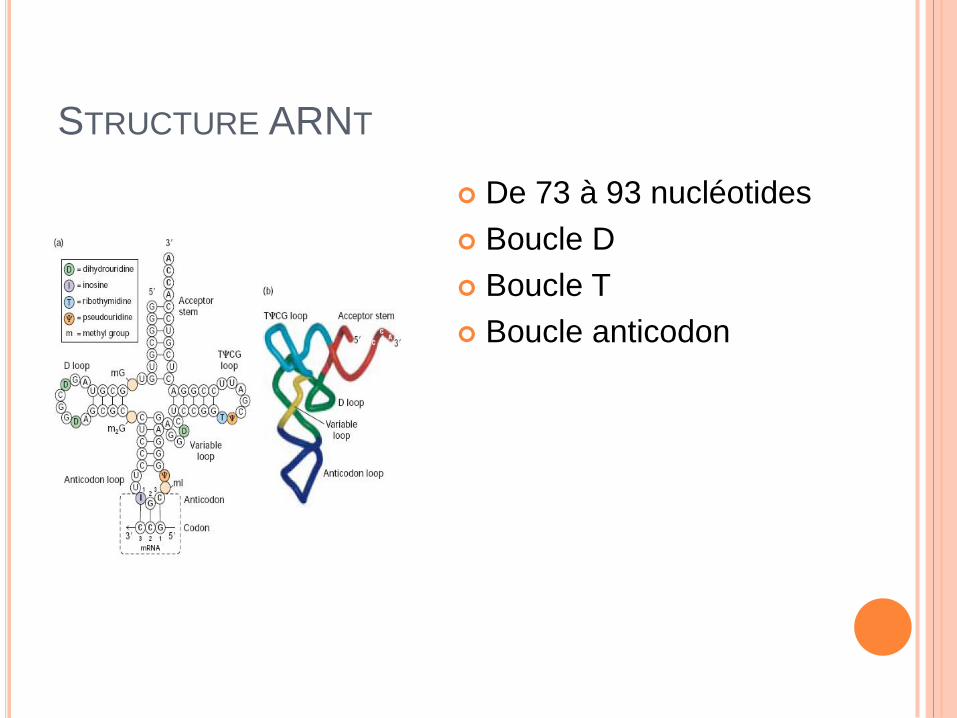
STRUCTURE ARNT
De 73 à 93 nucléotides
Boucle D
Boucle T
Boucle anticodon
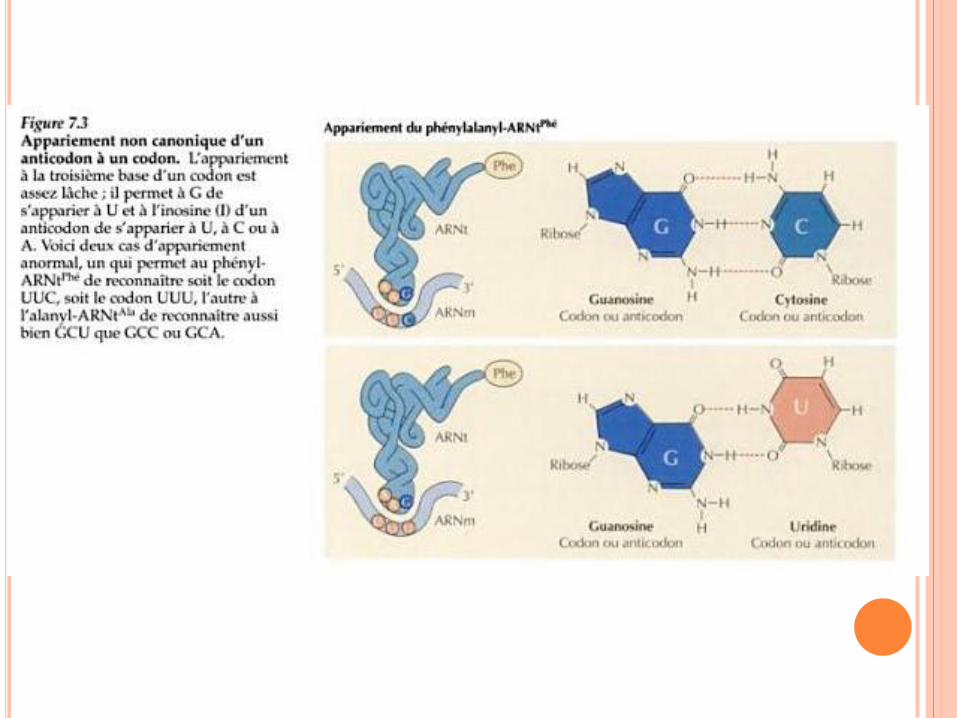
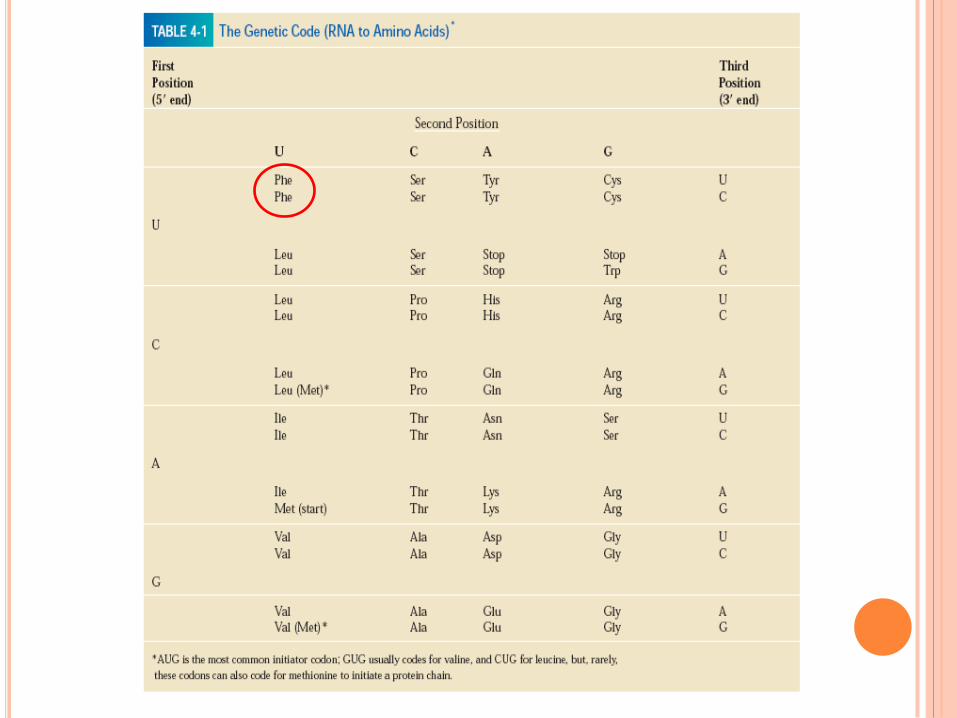
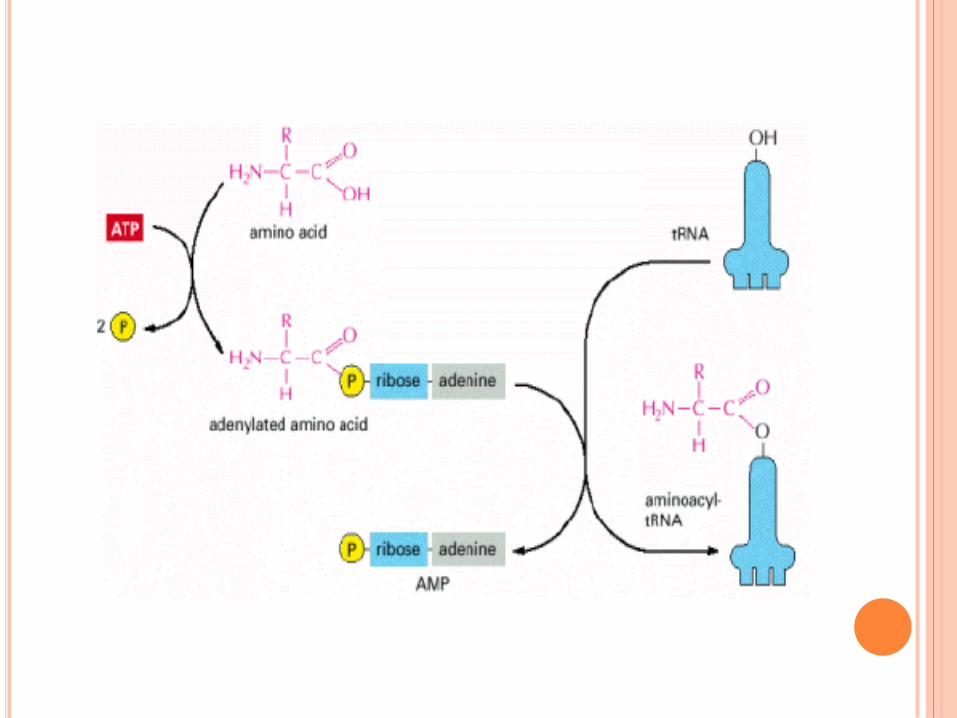
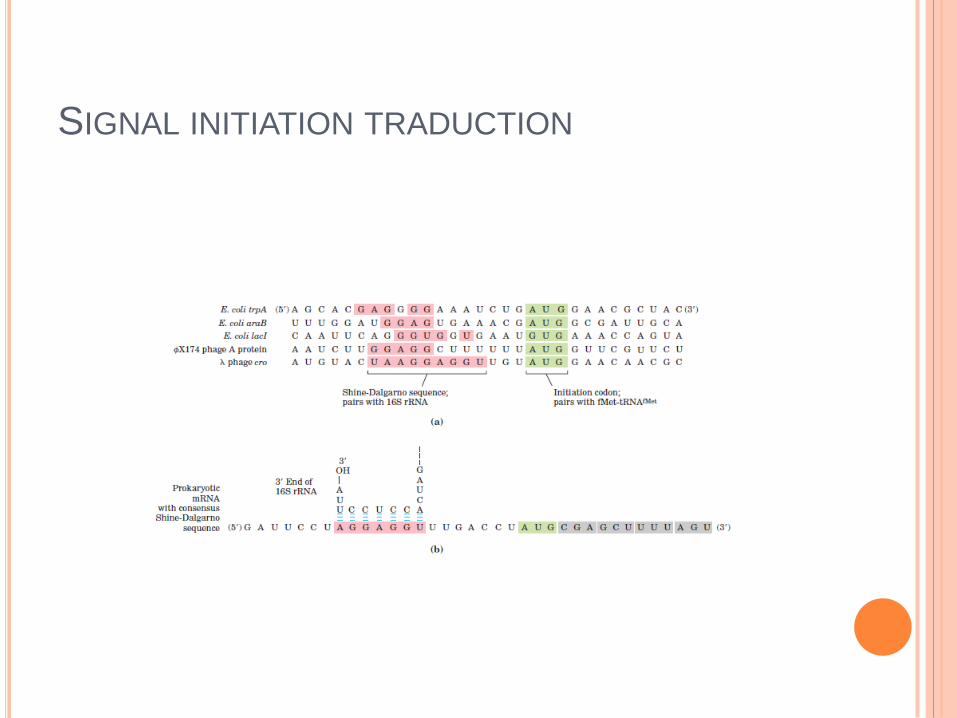
SIGNAL INITIATION TRADUCTION
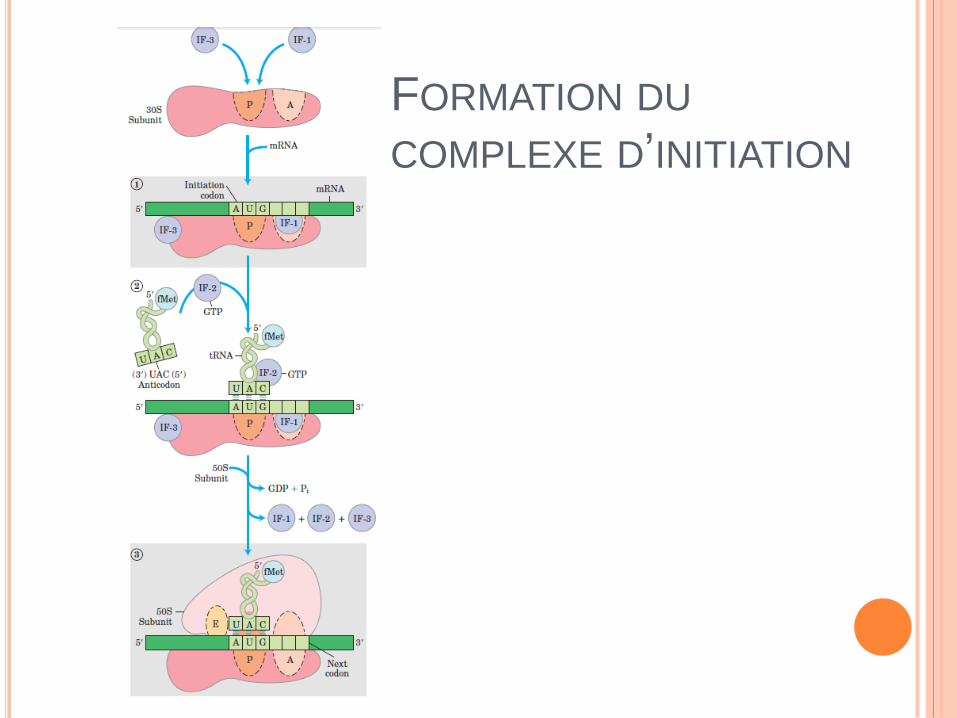
FORMATION DU
COMPLEXE D’INITIATION
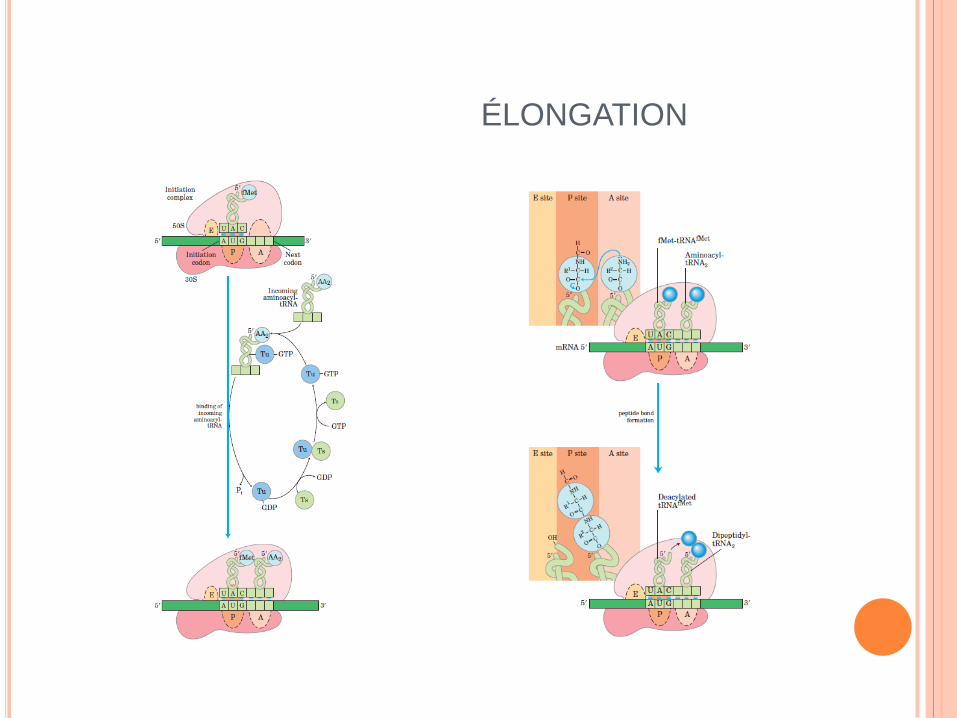
ÉLONGATION
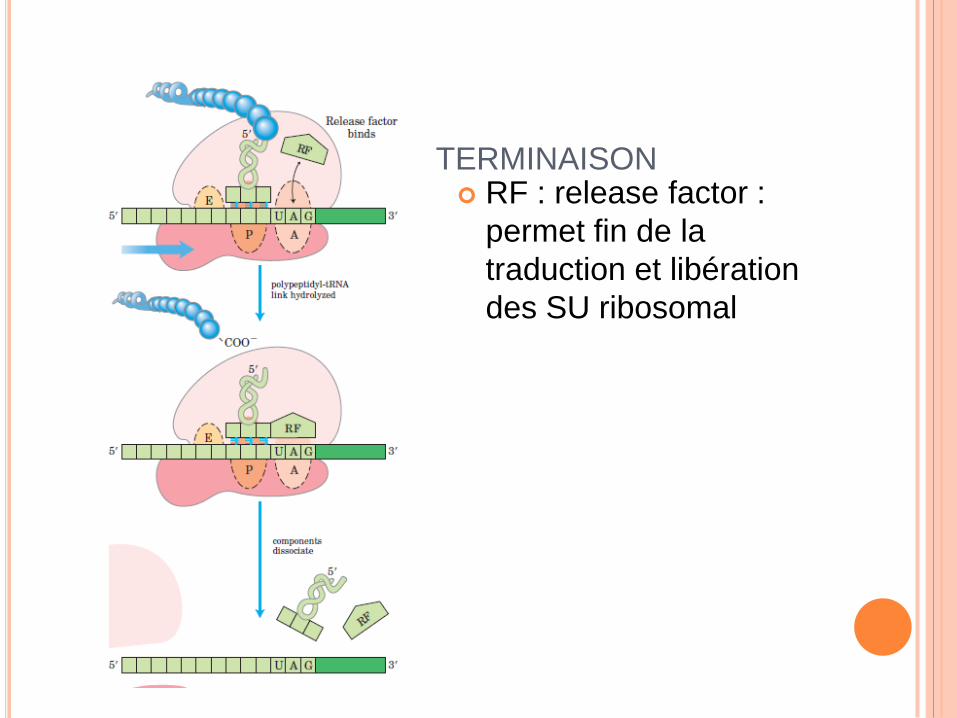
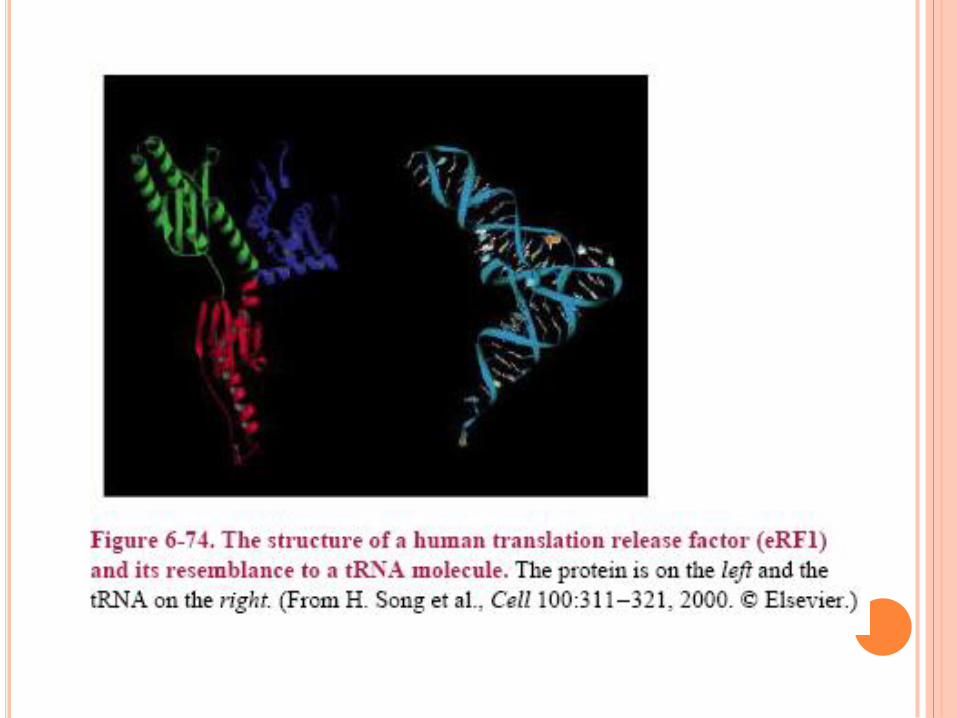
TERMINAISON RF : release factor :
permet fin de la
traduction et libération
des SU ribosomal
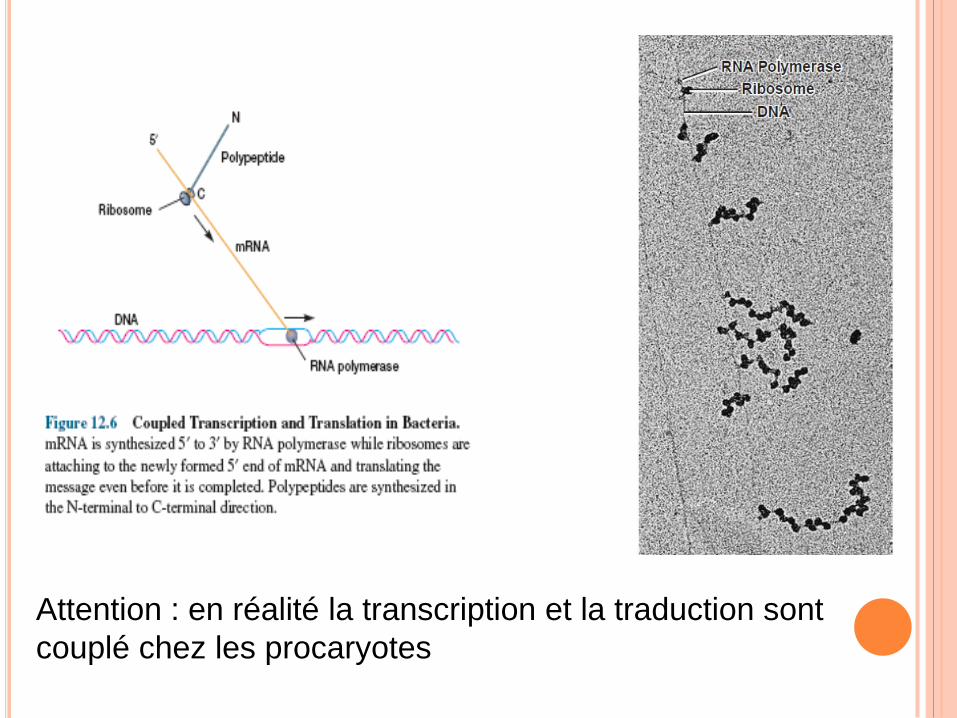
Attention : en réalité la transcription et la traduction sont
couplé chez les procaryotes

EXPRESSION GÉNÉTIQUE EUCARYOTE
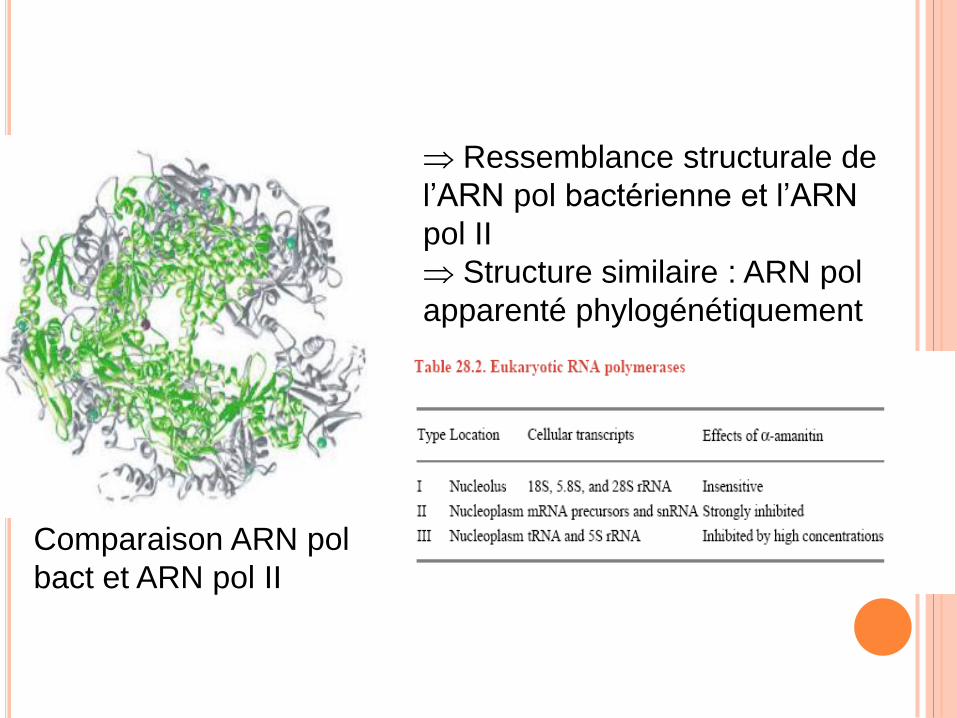
Ressemblance structurale de
l’ARN pol bactérienne et l’ARN
pol II
Structure similaire : ARN pol
apparenté phylogénétiquement
Comparaison ARN pol
bact et ARN pol II
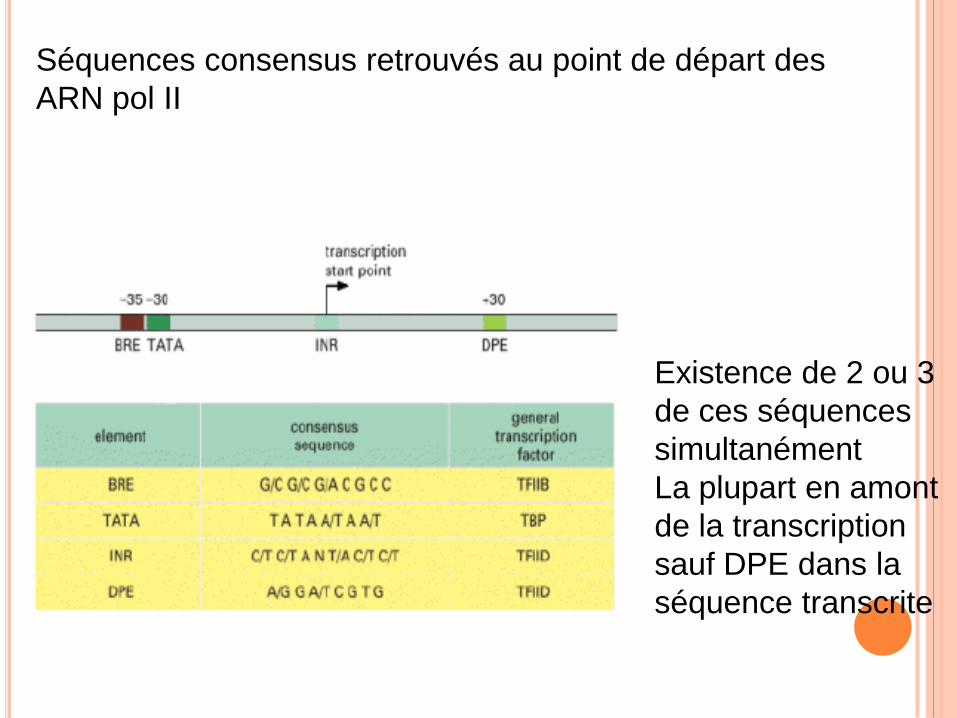
Séquences consensus retrouvés au point de départ des
ARN pol II
Existence de 2 ou 3
de ces séquences
simultanément
La plupart en amont
de la transcription
sauf DPE dans la
séquence transcrite
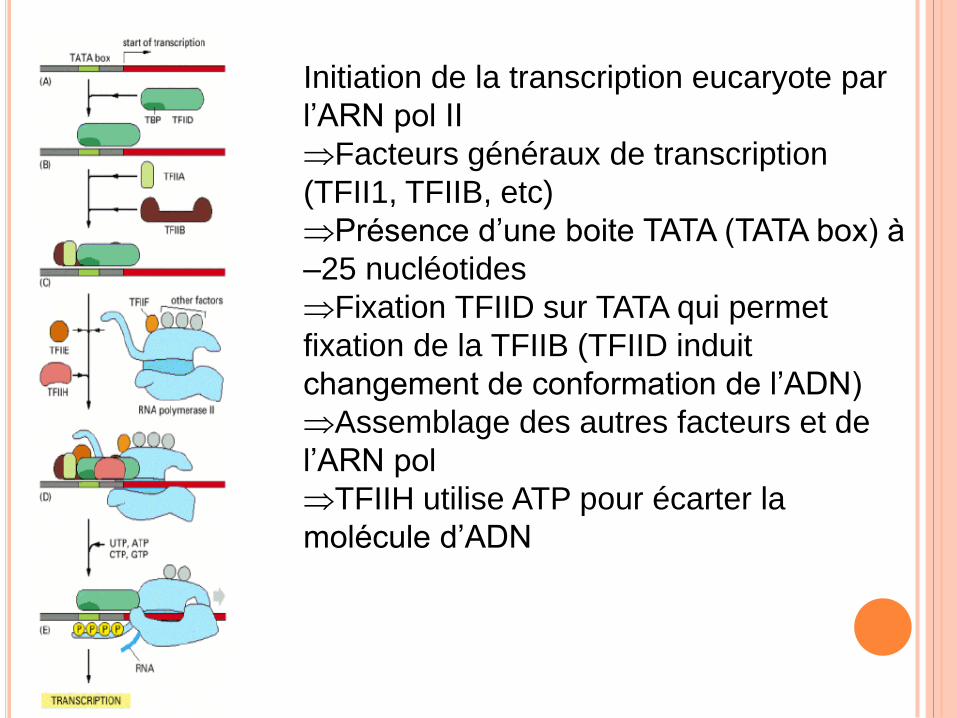
Initiation de la transcription eucaryote par
l’ARN pol II
Facteurs généraux de transcription
(TFII1, TFIIB, etc)
Présence d’une boite TATA (TATA box) à
–25 nucléotides
Fixation TFIID sur TATA qui permet
fixation de la TFIIB (TFIID induit
changement de conformation de l’ADN)
Assemblage des autres facteurs et de
l’ARN pol
TFIIH utilise ATP pour écarter la
molécule d’ADN
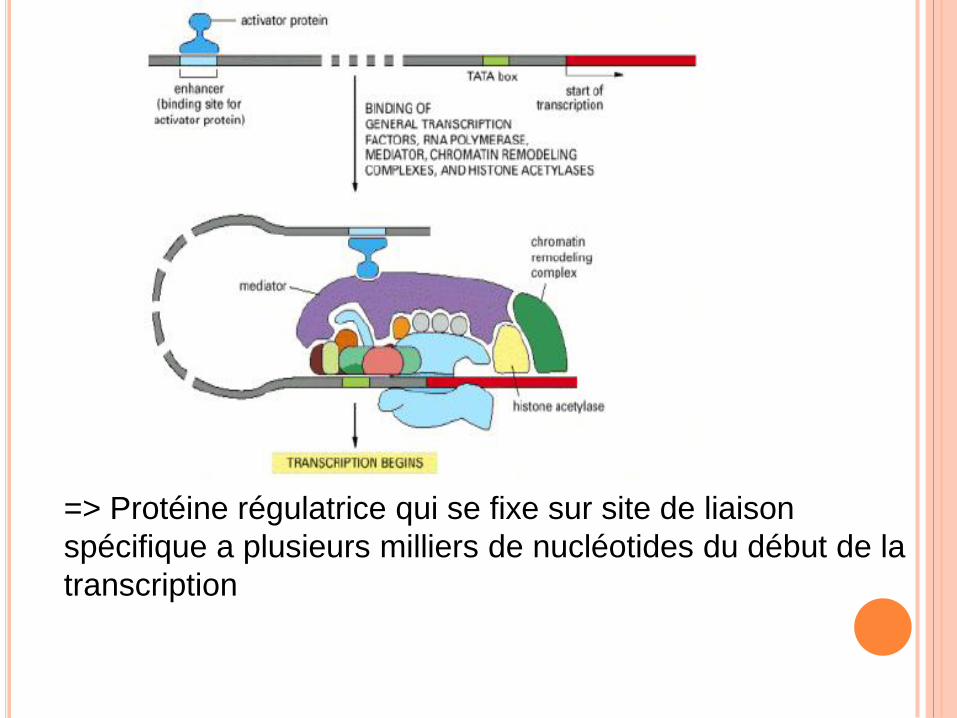
=> Protéine régulatrice qui se fixe sur site de liaison
spécifique a plusieurs milliers de nucléotides du début de la
transcription
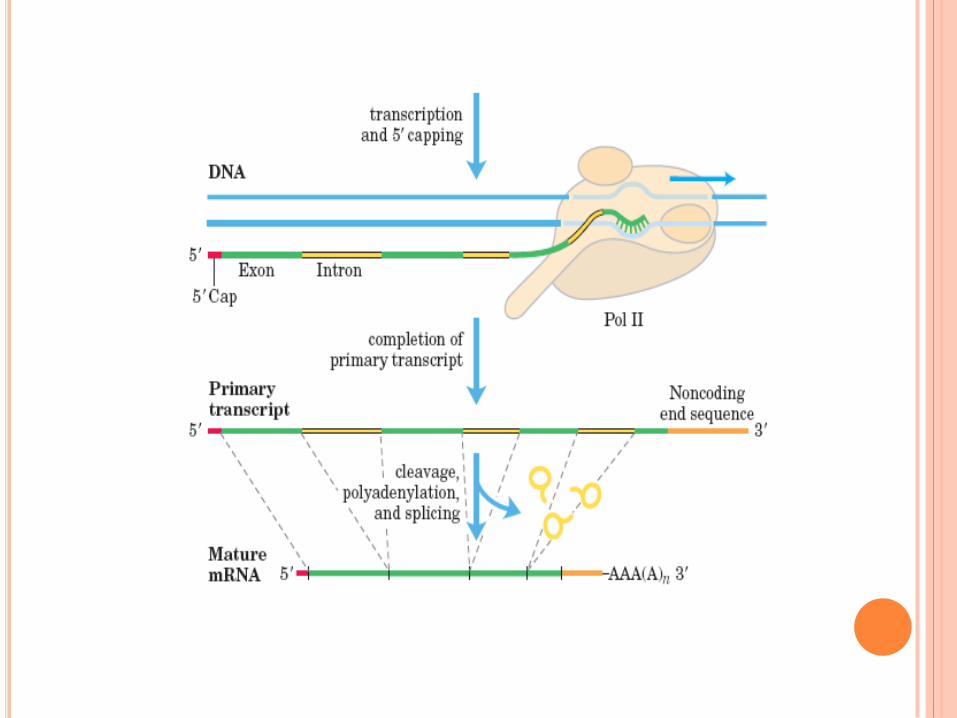
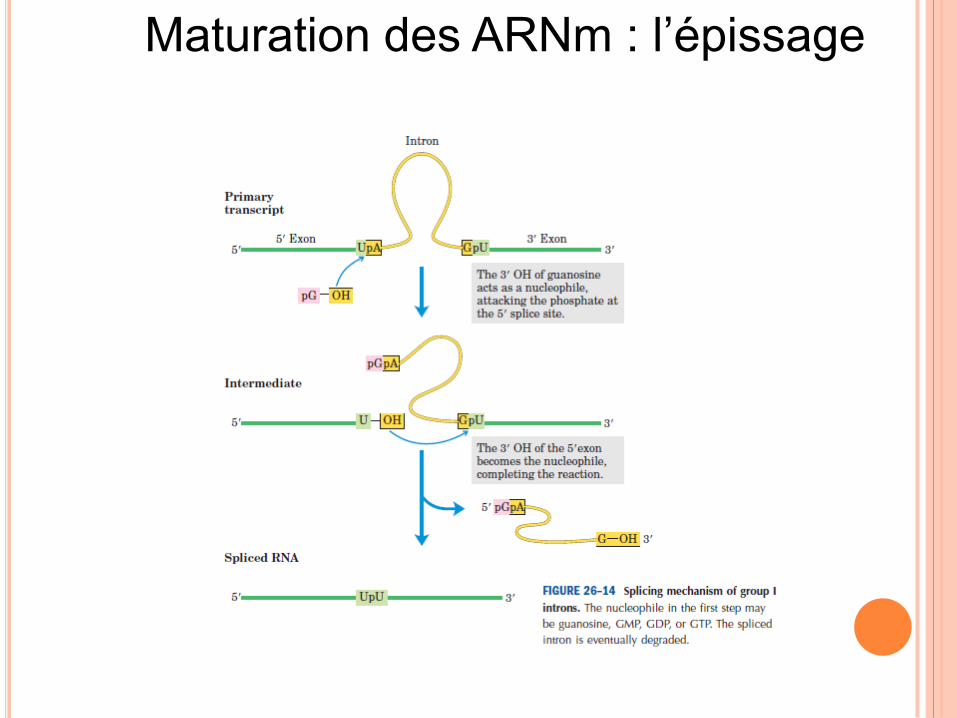
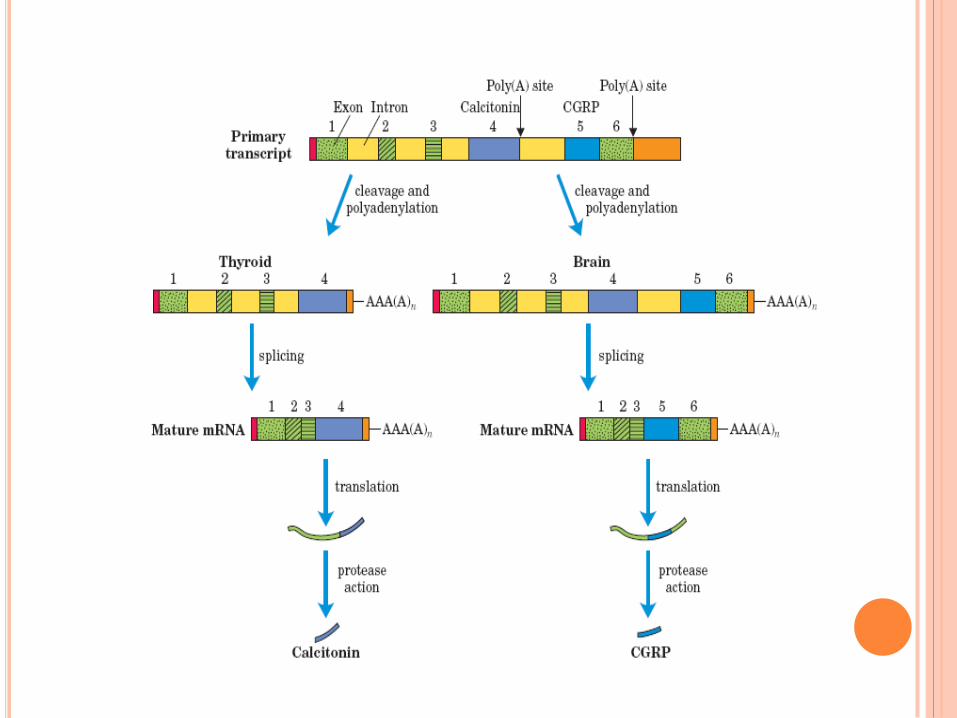
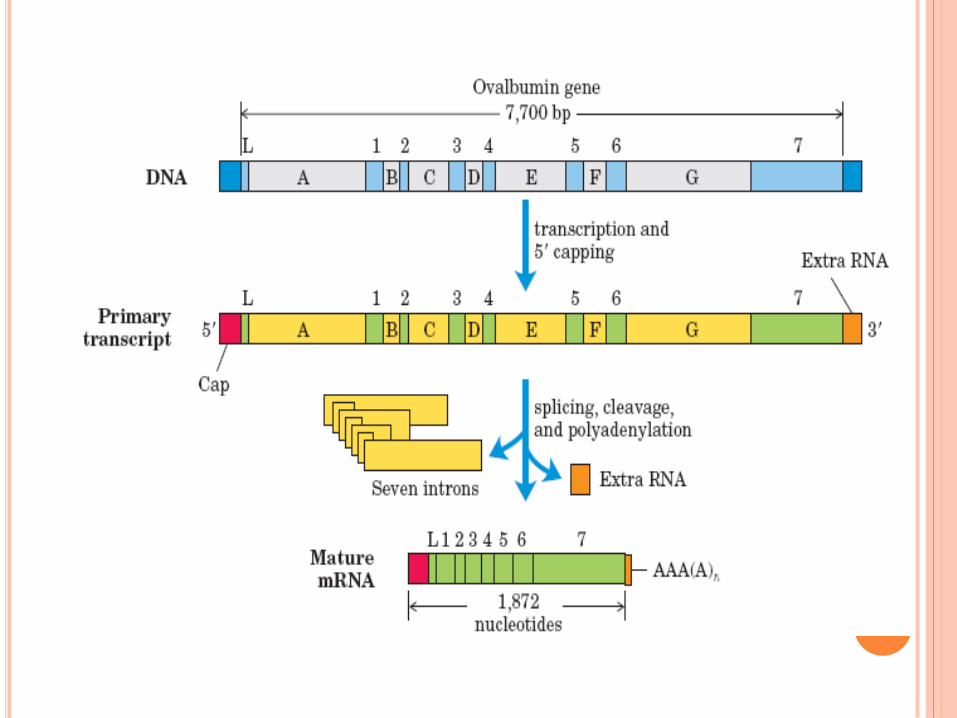
Maturation des ARNm : l’épissage
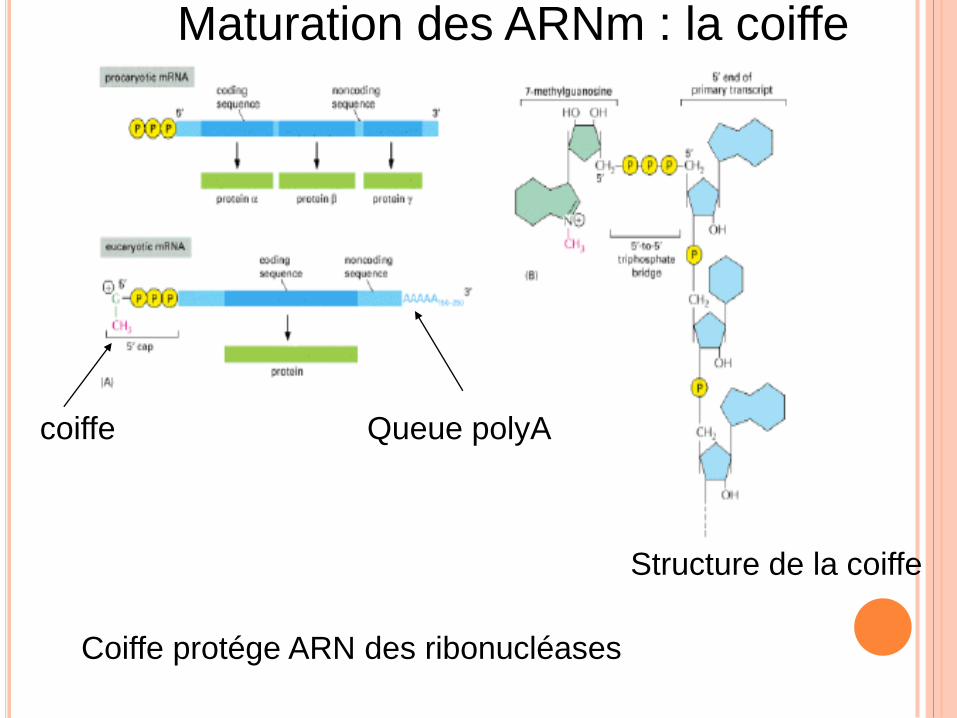
coiffe Queue polyA
Structure de la coiffe
Maturation des ARNm : la coiffe
Coiffe protége ARN des ribonucléases
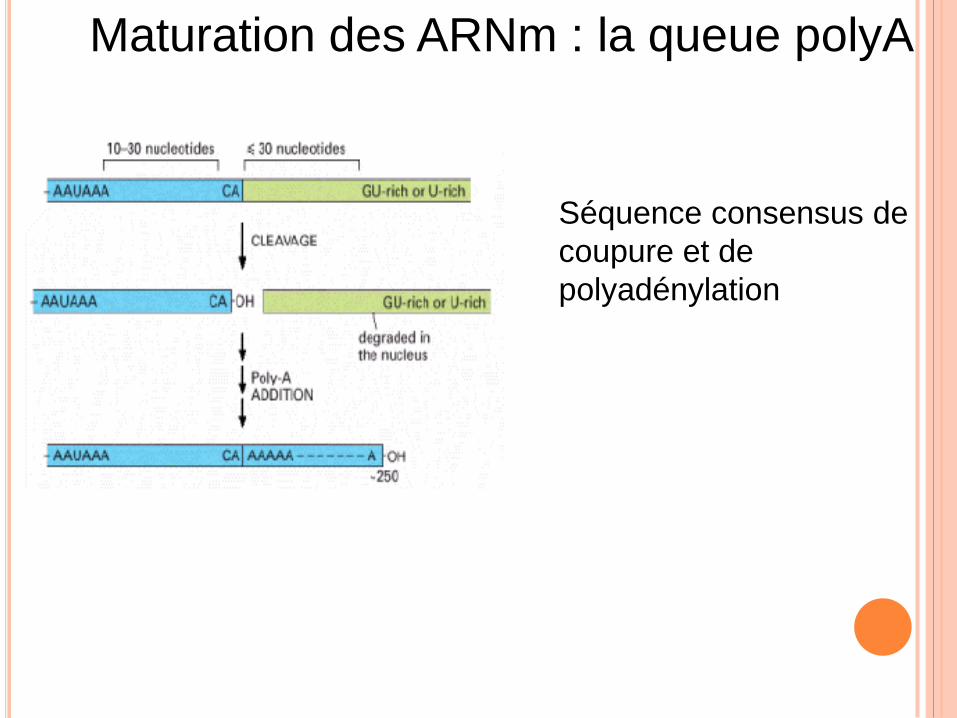
Maturation des ARNm : la queue polyA
Séquence consensus de
coupure et de
polyadénylation
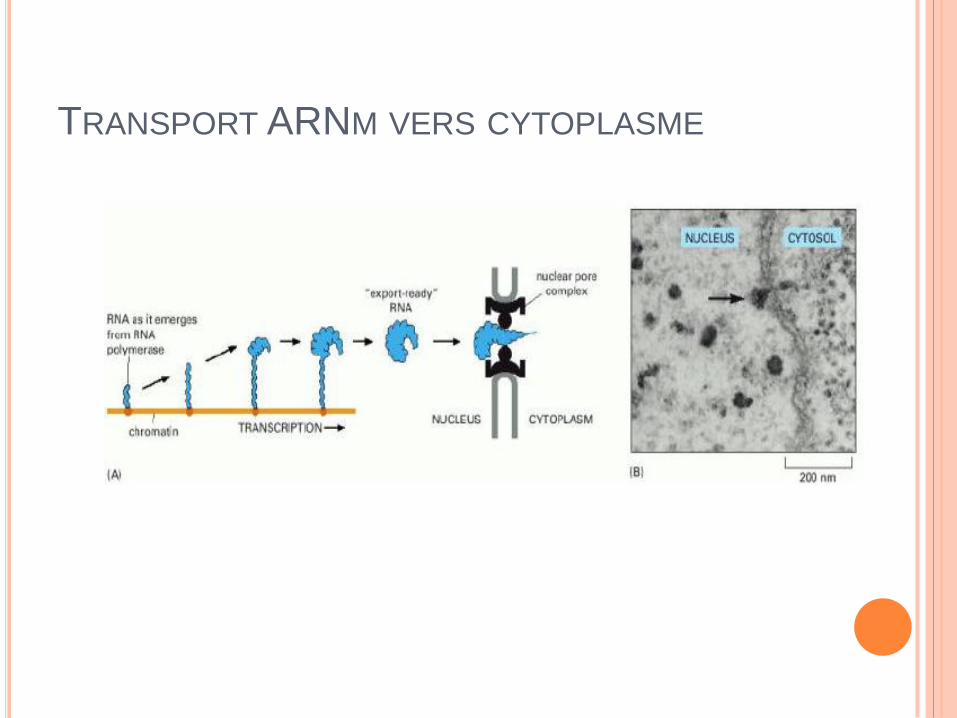
TRANSPORT ARNM VERS CYTOPLASME
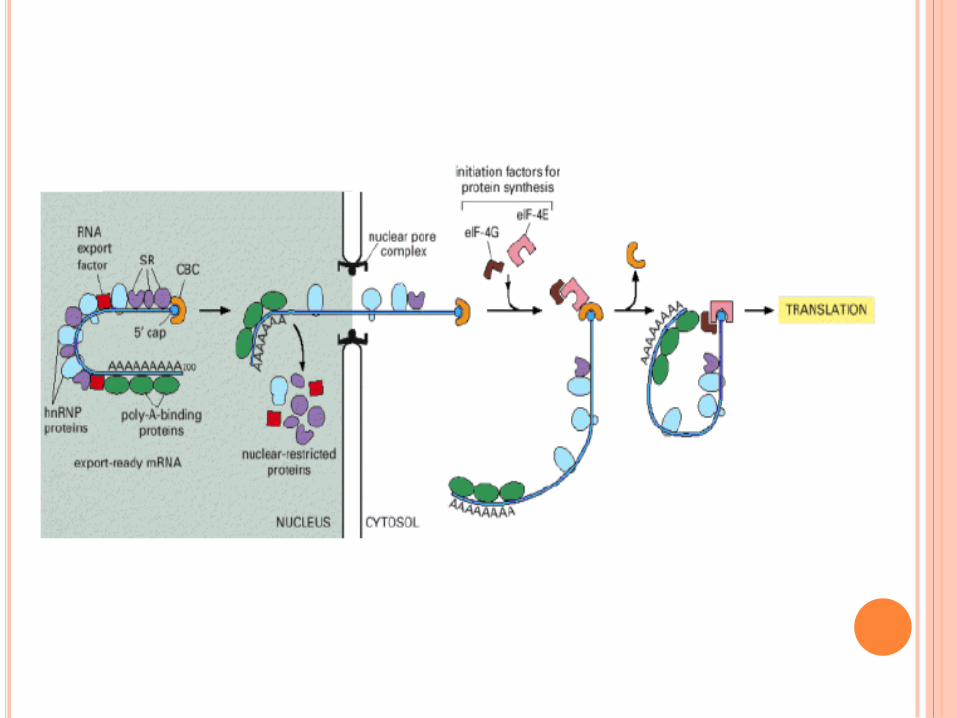

SÉQUENCE SIGNAL DE TRANSLOCATION DANS
LE RER
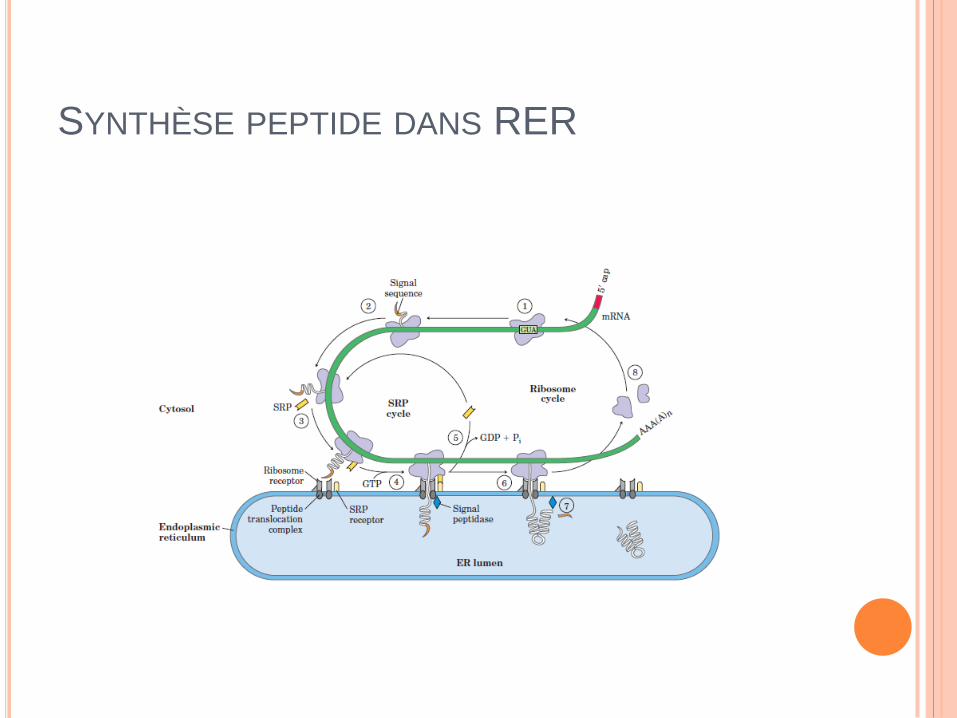
SYNTHÈSE PEPTIDE DANS RER
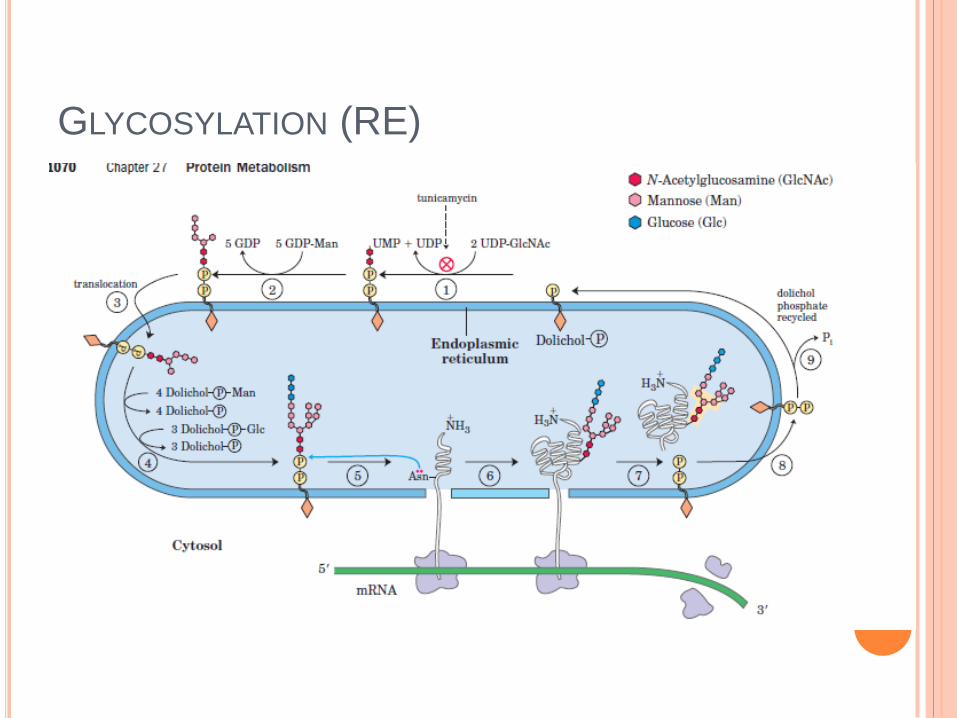
GLYCOSYLATION (RE)
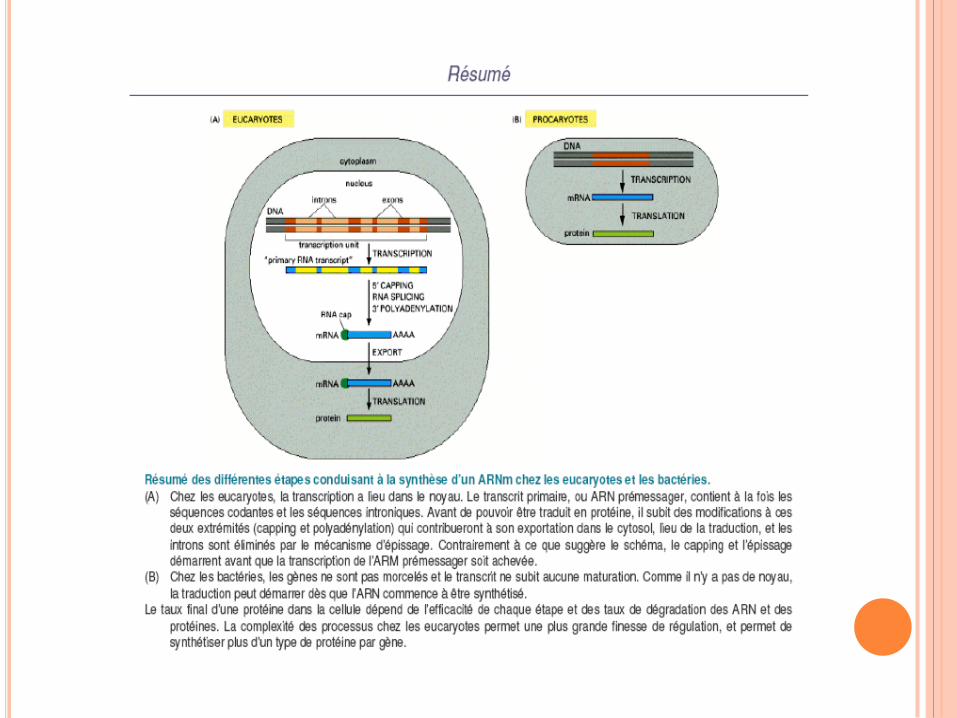

Modifications post-traductionnelles
et adressage protéique
• Elles permettent des modifications d’activité (inhibition, activation…)
• Se sont des réactions chimiques catalysées par des enzymes la plupart du temps.
• Trois sortes de modif post traductionnelles:
- fixation de protéines à des lipides
- glycosylation de protéines.
- phosphorylation de protéines.

CONTRÔLE DE L’EXPRESSION
GÉNÉTIQUE