
Etude des anisoptères des cours d’eau des Pays de la...
24
0 Clémence Monvoisin Soutenu à Rennes le 18.06.13 Jury: J.Pétillon S.Dugravot Rapport de stage Master 1 Patrimoine Naturel et Biodiversité UFR SVE Etude des cortèges d’Anisoptères des cours d’eau et de l’implication des microhabitats ligneux dans leur répartition Stage du 02.04.13 au 06.06.13 Correspondant universitaire : Julien Pétillon Maître de stage : Franck Herbrecht GRETIA Pays de la Loire
Transcript of Etude des anisoptères des cours d’eau des Pays de la...

0
Clémence Monvoisin
Soutenu à Rennes le 18.06.13
Jury:
J.Pétillon
S.Dugravot
Rapport de stage
Master 1 Patrimoine Naturel et Biodiversité
UFR SVE
Etude des cortèges d’Anisoptères des cours d’eau
et de l’implication des microhabitats ligneux
dans leur répartition
Stage du 02.04.13 au 06.06.13
Correspondant universitaire : Julien Pétillon
Maître de stage : Franck Herbrecht
GRETIA Pays de la Loire

1
Remerciements
Je tiens à remercier l’association du GRETIA pour m’avoir accueillie chaleureusement au
sein de l’équipe. Ce fut un plaisir de travailler avec des personnes passionnées par leur métier
et avec l’envie de le partager.
Cette étude est le fruit d’un travail inter-associatif, je remercie donc les bénévoles et salariés
de Mayenne Nature Environnement, du Groupe Naturaliste de Loire-Atlantique, de Bretagne
vivante Nantes, du CPIE Vallées de la Sarthe et du Loir, d’Entomologie Tourangelle et
Ligérienne et de la LPO, sans qui je n’aurai pu réaliser ce rapport.
Les précieuses explications de Thomas Cherpitel sur la réalisation du protocole en 2012
m’ont aidé dans la compréhension et la mise en place de cette étude.
Merci aux relecteurs pour avoir consacré du temps à ce travail et m’avoir aiguillée dans sa
rédaction.
Enfin, un grand merci à Franck Herbrecht pour avoir partagé ses connaissances, ses
expériences et pour nos discussions militantistes.
*Photographie de couverture : A gauche : Mâle de Gomphus vulgatissimus et son exuvie, Mathieu Lagarde. A droite : Emergence de Gomphus pulchellus, Clémence Monvoisin.

2
Sommaire
Introduction ............................................................................................................................. 3
Matériel et Méthodes ............................................................................................................ 5
1) Présentation des sites ....................................................................................................... 5
2) Modèle biologique ........................................................................................................... 5
3) Protocole expérimental .................................................................................................... 6
4) Analyses statistiques ....................................................................................................... 7
Tableau des abréviations…………………………………………………………………….
Résultats ................................................................................................................................... 8
1) Diversité et cortèges d’espèces présents sur la Sèvre, la Mayenne, l’Huisne et l’Erdre . 8
2) Effet de la présence de ligneux sur la densité et la richesse spécifique des espèces ..... 10
3) Lien entre la richesse en microhabitats et les cortèges d’espèces ................................. 10
Discussion ............................................................................................................................. 14
1) Cortèges d’espèces présents sur la Sèvre, l’Erdre, la Mayenne et l’Huisne ................. 14
2) Effet de la diversité des microhabitats ligneux sur les cortèges d’espèces ................... 16
Conclusion ............................................................................................................................. 17
Bibliographie ........................................................................................................................ 19

3
Introduction
Dans le cadre du Schéma d’Aménagement et de Gestion des Eaux, de la Trame Bleue
et de la Directive Cadre sur l’Eau, la détermination de nouveaux indicateurs biologiques est
attendue afin d’améliorer l’évaluation de l’état général des cours d’eau et d’en assurer le suivi.
Pour cela, l’approfondissement des connaissances sur les espèces et leurs relations avec cet
environnement est indispensable (Chessel et Mercier, 1993). L’ordre des Odonates constitue
l’un des groupes d’insectes caractéristiques des milieux aquatiques.
Présents depuis le Permien inférieur (Wootton, 1981), les Odonates ont développé une
plasticité leur permettant de coloniser divers milieux aux conditions environnementales
changeantes. Ils ont ainsi marqué l’évolution du paysage (De Ricqles, 1988) et sont présents
dans différents types de milieux aquatiques (lentiques, lotiques). En France métropolitaine,
près de cent taxons sont répartis en une dizaine de familles (Grand et Boudot, 2006).
Cependant, la régression progressive des zones humides, la réaffectation des étangs et lacs, la
pollution des cours d’eau et leurs recalibrage provoquent le déclin des populations d’Odonates
dans de nombreux pays. Par conséquent, ils sont à même de refléter les modifications globales
subies par l’environnement (Grand et Boudot, 2006).
Toutefois, l’utilisation de ce groupe comme indicateurs de l’état général des milieux suscite
encore des interrogations.
Jarry et Vidal (1960) estiment qu’il constitue « un mauvais matériel car leurs espèces sont
trop peu nombreuses pour que l’on puisse disposer d’un faisceau suffisant pour que chaque
habitat conditionne une population différente ». De même, la tolérance de certaines libellules
face aux polluants en ferait des outils indiciels de bio-surveillance inappropriés (Masselot et
Nel 2003).
A l’inverse, de nombreux spécialistes considèrent les Odonates comme un outil phare dans le
suivi des milieux aquatiques. Leur écologie est relativement bien connue, leur détermination
aisée et ils peuvent faire l’objet de méthodes d’échantillonnage simples et efficaces ce qui
permet de les utiliser facilement (Oertli, 2008). Les gestionnaires les choisissent d’ailleurs
bien souvent comme éléments clés pour l’analyse des habitats (Houard et al., 2012). Les
peuplements d’Odonates sont également reconnus comme de bons descripteurs biologiques.
Ils mettraient en évidence la structuration des milieux aquatiques, leur diversité en

4
microhabitats, leurs modifications générales, et traduiraient ainsi leur état de santé (Houard et
al., 2012). D’après Dommanget (1989), ils sont particulièrement sensibles à la réduction de la
végétation riveraine et celle aux alentours de leurs habitats.
Au Brésil, par exemple, il a été montré que certaines espèces d’Odonates sont dépendantes
des forêts rivulaires : la composition et la structuration de cette végétation influent sur la
diversité de ces espèces (Ferreira-Peruquetti et De Marco, 2002 ; De paiva Silva et al., 2010).
Enfin, d’autres auteurs avancent que le potentiel bio-indicateur des Odonates est une affaire
d’échelle : l’utilisation de ce groupe à un niveau spécifique serait effectivement peu
pertinente, mais les cortèges d’espèces peuvent réellement traduire les caractéristiques
générales du milieu (Dommanget 1989, Corbet 1999). Schmidt (1985) emploie ainsi le terme
de RSO « Representative Spectrum of Odonates », comme outil de diagnostic d’un habitat.
Dans le but d’alimenter la réflexion sur la gestion des cours d’eau et de leur berge, il est donc
primordial d’approfondir les connaissances sur les liens entre les cortèges d’espèces et leurs
habitats, en particulier avec la composition et la structuration végétale des berges.
Pour cela, cette étude a été basée sur la récolte des exuvies d’Odonates. La collecte de cette
dernière mue larvaire présente effectivement un intérêt particulier car elle n’engendre pas ou
peu de perturbations sur le milieu (Baïlo Ndiaye, 2010), contrairement à une approche centrée
sur les larves. De plus, la présence d’exuvies garantit l’autochtonie des espèces concernées
(Grand et Boudot, 2006), ce que ne peut assurer l’observation des imagos, compte tenu de
leur capacité dispersion.
Le sous-ordre des Anisoptères a été choisi comme modèle biologique. En effet, la petite taille
des exuvies de Zygoptères les rend plus difficiles à découvrir et influerait donc fortement sur
l’exhaustivité de la prospection.
En France, on dénombre actuellement 62 espèces et sous-espèces d’Anisoptères, regroupées
en six familles: les Aeshnidés, les Gomphidés, les Cordulégastridés, les Macromiidés, les
Corduliidés et les Libellulidés.
Dans le cadre de ce travail, diverses hypothèses peuvent être émises : les cortèges d’espèces
sont variables sur les cours d’eau ; la présence ou l’absence de végétation rivulaire ligneuse
influent sur la richesse spécifique des Anisoptères et leur abondance ; la composition et la

5
structuration de la végétation rivulaire ligneuse prédisent la présence de cortèges particuliers
d’espèces.
La prospection des exuvies a été réalisée sur quatre cours d’eau des Pays de la Loire. Les
caractéristiques majeures du milieu, ainsi que la composition et la structuration végétale
ligneuse des berges ont été relevées, simultanément aux collectes.
Matériel et Méthodes
1) Présentation des sites
Le choix des sites et des tronçons de cours d’eau s’est fait sur différents critères. L’élément
déterminant fut le calibre du cours d’eau, le but étant d’étudier la répartition des exuvies
d’Anisoptères des rivières de moyennes envergure, mais suffisamment important pour que
l’institution soit intéressée par leur gestion (domaine public). L’eau devait y être courante et la
prospection en canoë réalisable. De plus, les sites devaient être potentiellement intéressants en
termes de richesse spécifique et peu prospectés antérieurement afin d’enrichir les inventaires.
L’étude a ainsi été réalisée sur des tronçons de quatre cours d’eau des Pays de la Loire :
La Sèvre nantaise (entre le pont de la Grossière et Terbin, à Boussay), l’Erdre dans sa partie
amont (entre Nort-sur-Erdre et Joué-sur-Erdre), l’Huisne (entre Saint-Mars-La-Brière et
Monfort-le-Gesnois et entre Brêteau et Vouvray-sur-Huisne) et la Mayenne amont (entre Gué
de Loré et la Frételière).
2) Modèle biologique
Les Odonates ont un cycle de vie marqué par différentes étapes successives. Les œufs pondus
par les femelles dans l’eau, sur la vase ou dans la végétation rivulaire, se développent pour
donner naissance à des larves aquatiques. Celles-ci subissent de nombreuses mues, suivies, en
fin de développement, par de profondes modifications internes (métamorphose incomplète).
L’imago se forme bel et bien à l’intérieur du tégument du dernier stade larvaire. Lors de la
dernière mue, dite imaginale, l’individu devient définitivement aérien ; en se fixant sur un
support, il va s’extraire de son exuvie. Suivra une période de maturation sexuelle au bout de
laquelle intervient la reproduction.

6
Cette étude fut basée sur la collecte systématique des exuvies.
3) Protocole expérimental
Les exuvies d’Anisoptères ont été récoltées le plus exhaustivement possible à vue, le long des
berges. Ces dernières, ont été parcourues à partir de la rivière, en canoë, et pour chaque cours
d’eau, différents tronçons ont été étudiés. Ils ont varié en fonction des conditions
expérimentales et des capacités de prospection.
Cet échantillonnage continu irrégulier a nécessité la présence de quatre équipages (deux
naturalistes par canoë) qui ont opéré simultanément sur des sections différentes d’une même
rivière. Les deux rives ont été prospectées et des sous-sections ont été identifiées. Elles
correspondent à des longueurs de berge sur lesquelles le milieu apparaissait homogène en
termes de conformation de berge et de ripisylve. Un lot d’exuvie fut récolté pour chaque sous-
section.
Pour déterminer la longueur de ces sous-sections, des seuils d’échelle ont été mis en place : un
seuil minimal a été fixé à 15 m, sauf cas particulier (ouvrages, ampoule lentiques,
hétérogénéité soudaine). De plus, si la ripisylve ou la conformation de la berge changeait à
l’évidence sur une longueur de 50 m ou plus, il a fallu impérativement établir une nouvelle
sous-section.
Deux visites successives sur chaque tronçon de rivière ont été réalisées afin de récolter les
espèces de phénologies différentes. Pour cela deux dates de prospection des berges ont été
fixées pour les différents sites :
- L’Huisne : 06.06.2012 et 11.07.2012 - La Mayenne : 29.05.2012 et 26.06.2012
- La Sèvre : 25.05.2012 et 01.07.2012 - L’Erdre 31.05.2012 et 26.06.2012
Lors de la deuxième prospection, les sous-sections identifiées au premier passage ont été
conservées.
Ajoutées à la récolte des exuvies, des variables environnementales ont été prises en compte
pour décrire les sous-sections. Le choix des paramètres à relever s’est fait à partir d’études
bibliographiques (Dommanget 1989, Meurgey et al 2000, Masselot et Nel 2003), afin
d’inclure les facteurs pouvant jouer un rôle dans la constitution des cortèges d’espèces. Par

7
ailleurs, ces critères ont été choisis en permettant une prise de note efficace sur le terrain.
Des variables qualitatives ont donc été relevées. Celles concernées par notre problématique
sont : l’ombrage (absent, faible, moyen, fort) ; la présence, la nature et la dominance
d’essences végétales ligneuses.
Des microhabitats ont ainsi pu être définis pour chaque sous-section par la diversité et la
structure particulière des essences ligneuses. L’ombrage a traduit indirectement l’impact de
cette végétation sur le cours d’eau en termes d’ensoleillement.
Matériel requis
Des piluliers ont été nécessaires pour la récolte des exuvies ainsi que des fiches de terrain
pour relever les modalités environnementales. Des photographies aériennes à échelle
1/2500ème
ont permis de se localiser sur le cours d’eau et identifier les sous-sections.
Travail au laboratoire
La détermination des exuvies a été réalisée sous loupe binoculaire, à l’aide de la clé de
détermination simplifiée des exuvies d’Anisoptères réalisée pour l’occasion (Cherpitel et
Herbrecht, 2012).
4) Analyses statistiques
Les longueurs de berges prospectées ayant varié pour chaque cours d’eau, les abondances
d’espèces ont donc été abordées sur la base des densités d’exuvies sur 100 mètres linéaires
(La Sèvre nantaise : 12km de berge, l’Erdre dans sa partie amont : 13.6km de berge,
l’Huisne : 13.2km de berge, la Mayenne amont : 7,4 km de berge).
La diversité des cortèges et la répartition des effectifs des espèces ont été analysées à l’aide du
logiciel Past. La richesse spécifique, l’indice de Shannon-Weaver et d’équitabilité de Piélou
ont ainsi pu être calculés. Par ailleurs, une analyse en composante principale (ACP) a été
réalisée sur les densités d’espèces sous R afin de définir les cortèges.
Ce même logiciel a ensuite été utilisé pour tester les différentes hypothèses (Test de Chi2 et
test de Wilcoxon).
De la même façon, une analyse de co-inertie a été réalisée entre les densités d’espèces et les
variables environnementales (Dolédec and Chessel, 1994). Pour cela, l’ACP déjà effectuée
8
ainsi qu’une analyse en composante multiples (ACM) sur le tableau des modalités
environnementales, ont été utilisés.
Résultats
1) Diversité et cortèges d’espèces présents sur la Sèvre, la Mayenne, l’Huisne et
l’Erdre
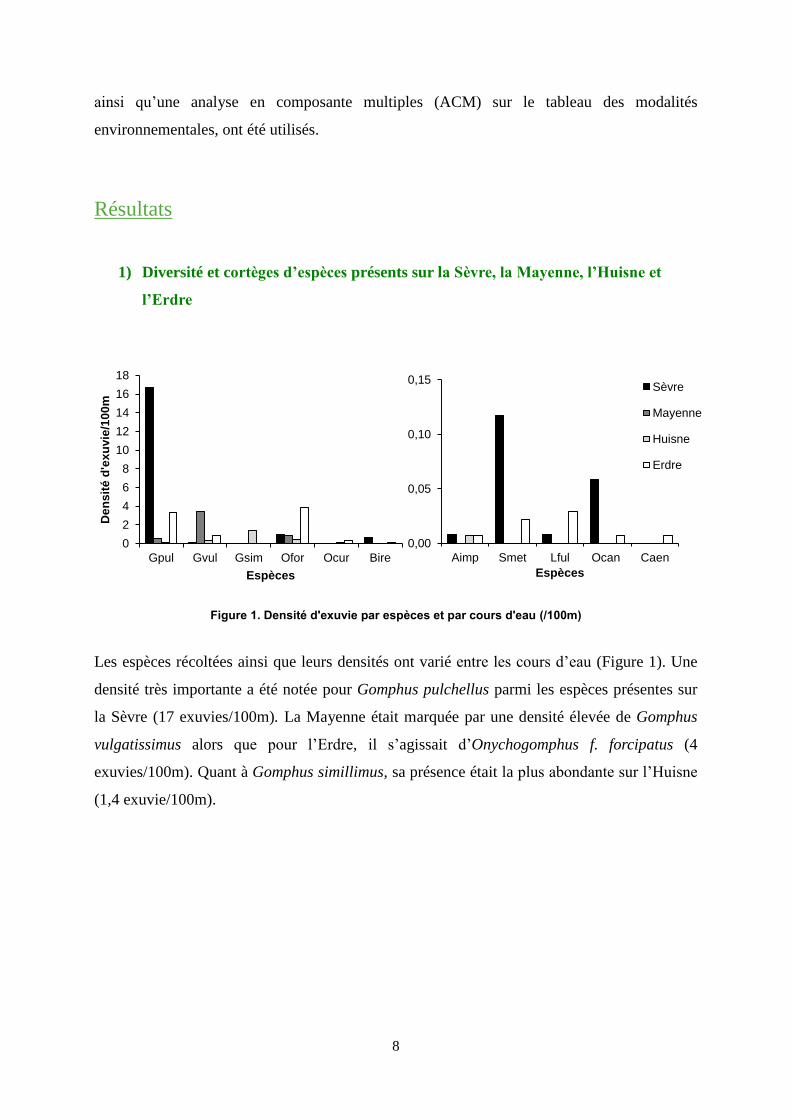
Figure 1. Densité d'exuvie par espèces et par cours d'eau (/100m)
Les espèces récoltées ainsi que leurs densités ont varié entre les cours d’eau (Figure 1). Une
densité très importante a été notée pour Gomphus pulchellus parmi les espèces présentes sur
la Sèvre (17 exuvies/100m). La Mayenne était marquée par une densité élevée de Gomphus
vulgatissimus alors que pour l’Erdre, il s’agissait d’Onychogomphus f. forcipatus (4
exuvies/100m). Quant à Gomphus simillimus, sa présence était la plus abondante sur l’Huisne
(1,4 exuvie/100m).
0
2
4
6
8
10
12
14
16
18
Gpul Gvul Gsim Ofor Ocur Bire
Den
sit
é d
'ex
uvie
/10
0m
Espèces
0,00
0,05
0,10
0,15
Aimp Smet Lful Ocan Caen
Espèces
Sèvre
Mayenne
Huisne
Erdre
9
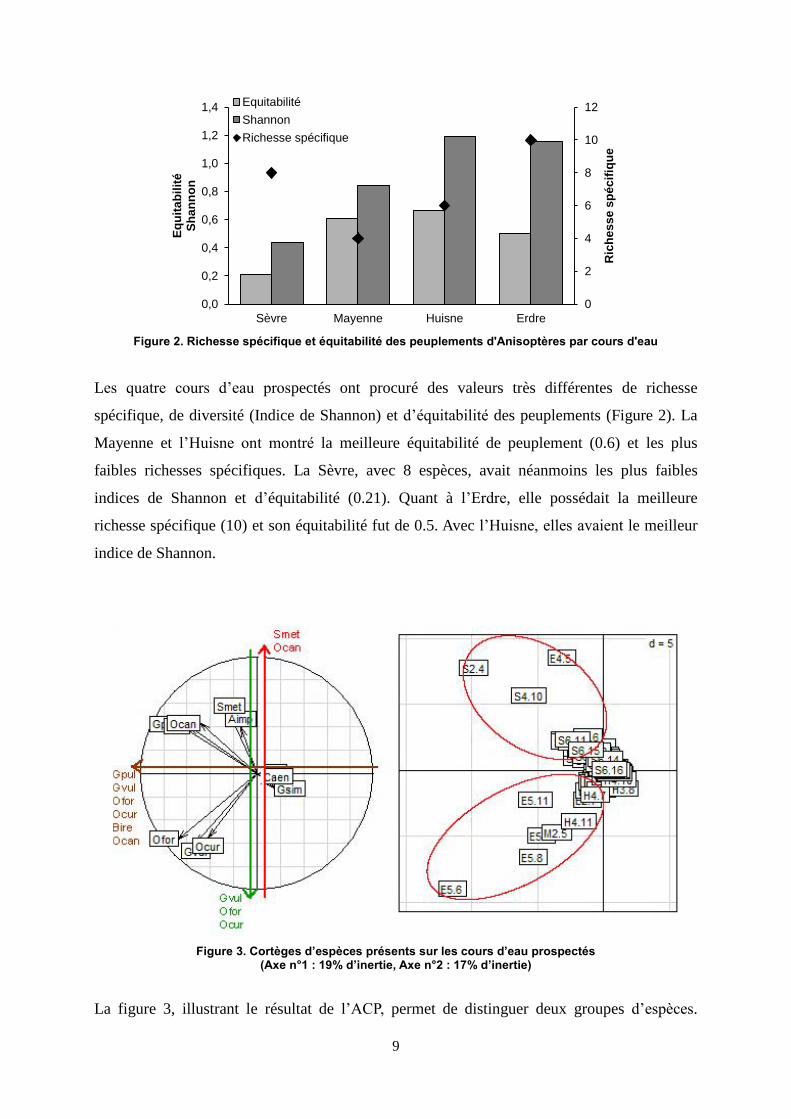
Figure 2. Richesse spécifique et équitabilité des peuplements d'Anisoptères par cours d'eau
Les quatre cours d’eau prospectés ont procuré des valeurs très différentes de richesse
spécifique, de diversité (Indice de Shannon) et d’équitabilité des peuplements (Figure 2). La
Mayenne et l’Huisne ont montré la meilleure équitabilité de peuplement (0.6) et les plus
faibles richesses spécifiques. La Sèvre, avec 8 espèces, avait néanmoins les plus faibles
indices de Shannon et d’équitabilité (0.21). Quant à l’Erdre, elle possédait la meilleure
richesse spécifique (10) et son équitabilité fut de 0.5. Avec l’Huisne, elles avaient le meilleur
indice de Shannon.
Figure 3. Cortèges d’espèces présents sur les cours d’eau prospectés (Axe n°1 : 19% d’inertie, Axe n°2 : 17% d’inertie)
La figure 3, illustrant le résultat de l’ACP, permet de distinguer deux groupes d’espèces.
0
2
4
6
8
10
12
0,0
0,2
0,4
0,6
0,8
1,0
1,2
1,4
Sèvre Mayenne Huisne Erdre
Ric
he
ss
e s
pé
cif
iqu
e
Eq
uit
ab
ilit
é
Sh
an
no
n
Equitabilité
Shannon
Richesse spécifique

10
Gomphus pulchellus, Orthetrum cancellatum, Boyeria irene et Somatochlora metallica
forment un premier groupe. Le deuxième est constitué de Gomphus vulgatissimus, Oxygastra
curtisii et Onychogomphus f.forcipatus. Le premier groupe est lié à des sous-sections de la
Sèvre (Figure 3 et Figure 1), le deuxième davantage à des sous-sections de l’Erdre, de la
Mayenne et de l’Huisne.
Les sous-sections S2.4, S4.10 et E4.5, qui se distinguent particulièrement au travers de l’ACP,
ne montraient cependant pas de modalités environnementales communes particulières. De
même pour les sous-sections E5.6, E5.8, E5.11, M2.5 et H4.11.
2) Effet de la présence de ligneux sur la densité et la richesse spécifique des espèces
Pour comprendre l’effet de la présence de ligneux sur la composition des cortèges, des tests
statistiques ont été réalisés sur les données de l’Erdre, de la Sèvre et de l’Huisne réunis. Celles
de la Mayenne n’ont pas été prises en compte car elles ne comptaient qu’une sous-section
sans arbre.
Deux hypothèses ont été émises:
- la présence de ligneux influe sur la richesse spécifique des exuvies ;
- la présence de ligneux influe sur la densité des exuvies.
L’hypothèse nulle - la présence de ligneux n’a pas d’influence sur la richesse spécifique des
exuvies récoltées - ne peut être rejetée (Test de Chi2, X² = 28.39, p-value = 0.29, 25 ddl).
De même, la présence de ligneux ne semble pas influer significativement sur la densité
d’exuvies récoltés (Test de Wilcoxon, W = 752.5, p-value = 0.24).
Les cours d’eau étant constitués de cortèges d’espèces très différents, les mêmes tests ont été
effectués sur chaque rivière séparément. Mais à nouveau, les hypothèses nulles n’ont pu être
rejetées.
3) Lien entre la richesse en microhabitats et les cortèges d’espèces
La nature, la diversité et la dominance des ligneux ayant permis de définir des microhabitats,
ces derniers peuvent être mis en cause dans la richesse spécifique des espèces d’Anisoptères.
11
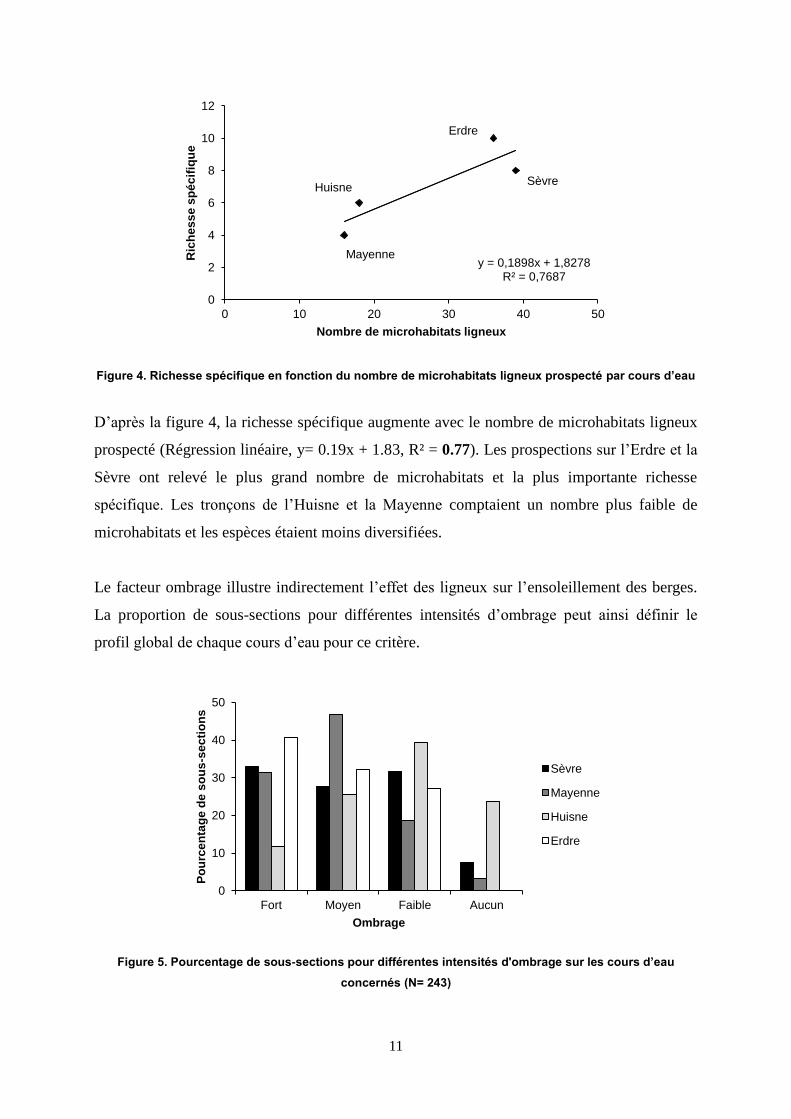
Figure 4. Richesse spécifique en fonction du nombre de microhabitats ligneux prospecté par cours d’eau
D’après la figure 4, la richesse spécifique augmente avec le nombre de microhabitats ligneux
prospecté (Régression linéaire, y= 0.19x + 1.83, R² = 0.77). Les prospections sur l’Erdre et la
Sèvre ont relevé le plus grand nombre de microhabitats et la plus importante richesse
spécifique. Les tronçons de l’Huisne et la Mayenne comptaient un nombre plus faible de
microhabitats et les espèces étaient moins diversifiées.
Le facteur ombrage illustre indirectement l’effet des ligneux sur l’ensoleillement des berges.
La proportion de sous-sections pour différentes intensités d’ombrage peut ainsi définir le
profil global de chaque cours d’eau pour ce critère.
Figure 5. Pourcentage de sous-sections pour différentes intensités d'ombrage sur les cours d’eau
concernés (N= 243)
y = 0,1898x + 1,8278 R² = 0,7687
0
2
4
6
8
10
12
0 10 20 30 40 50
Ric
he
ss
e s
pé
cif
iqu
e
Nombre de microhabitats ligneux
Mayenne
Huisne
Erdre
Sèvre
0
10
20
30
40
50
Fort Moyen Faible Aucun
Po
urc
en
tag
e d
e s
ou
s-s
ec
tio
ns
Ombrage
Sèvre
Mayenne
Huisne
Erdre
12
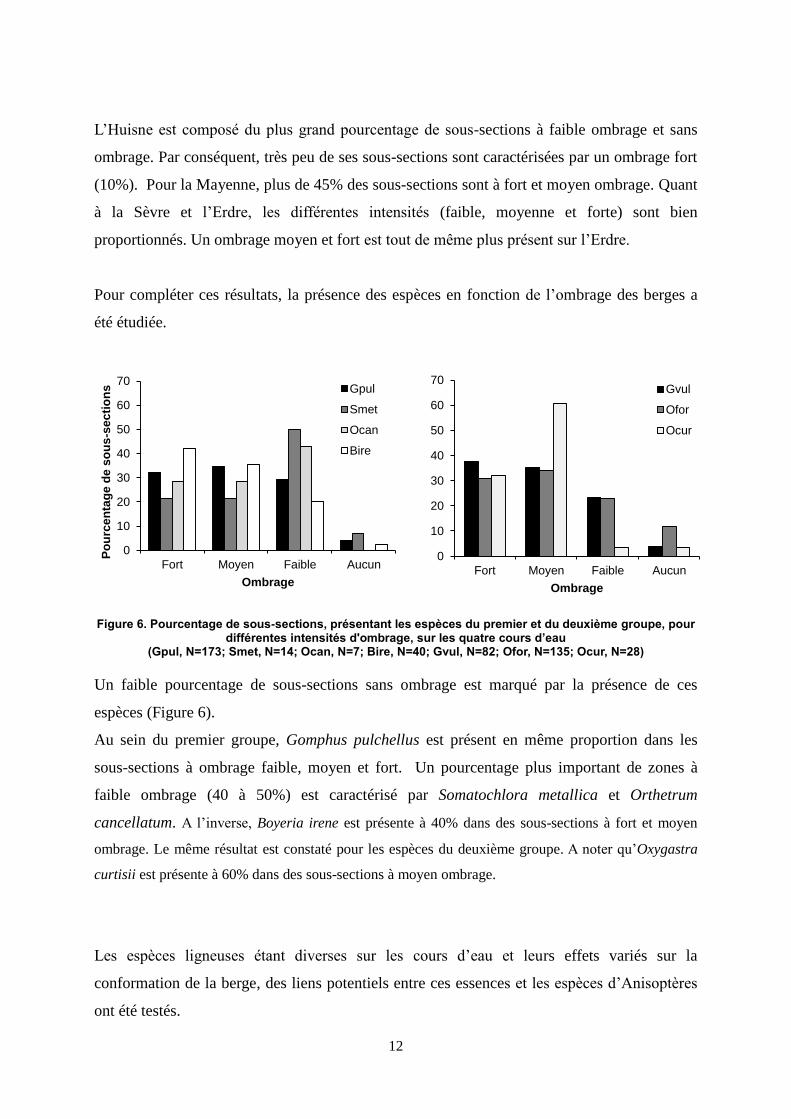
L’Huisne est composé du plus grand pourcentage de sous-sections à faible ombrage et sans
ombrage. Par conséquent, très peu de ses sous-sections sont caractérisées par un ombrage fort
(10%). Pour la Mayenne, plus de 45% des sous-sections sont à fort et moyen ombrage. Quant
à la Sèvre et l’Erdre, les différentes intensités (faible, moyenne et forte) sont bien
proportionnés. Un ombrage moyen et fort est tout de même plus présent sur l’Erdre.
Pour compléter ces résultats, la présence des espèces en fonction de l’ombrage des berges a
été étudiée.
Figure 6. Pourcentage de sous-sections, présentant les espèces du premier et du deuxième groupe, pour différentes intensités d'ombrage, sur les quatre cours d’eau
(Gpul, N=173; Smet, N=14; Ocan, N=7; Bire, N=40; Gvul, N=82; Ofor, N=135; Ocur, N=28)
Un faible pourcentage de sous-sections sans ombrage est marqué par la présence de ces
espèces (Figure 6).
Au sein du premier groupe, Gomphus pulchellus est présent en même proportion dans les
sous-sections à ombrage faible, moyen et fort. Un pourcentage plus important de zones à
faible ombrage (40 à 50%) est caractérisé par Somatochlora metallica et Orthetrum
cancellatum. A l’inverse, Boyeria irene est présente à 40% dans des sous-sections à fort et moyen
ombrage. Le même résultat est constaté pour les espèces du deuxième groupe. A noter qu’Oxygastra
curtisii est présente à 60% dans des sous-sections à moyen ombrage.
Les espèces ligneuses étant diverses sur les cours d’eau et leurs effets variés sur la
conformation de la berge, des liens potentiels entre ces essences et les espèces d’Anisoptères
ont été testés.
0
10
20
30
40
50
60
70
Fort Moyen Faible Aucun
Ombrage
Gvul
Ofor
Ocur
0
10
20
30
40
50
60
70
Fort Moyen Faible Aucun
Po
urc
en
tag
e d
e s
ou
s-s
ec
tio
ns
Ombrage
Gpul
Smet
Ocan
Bire
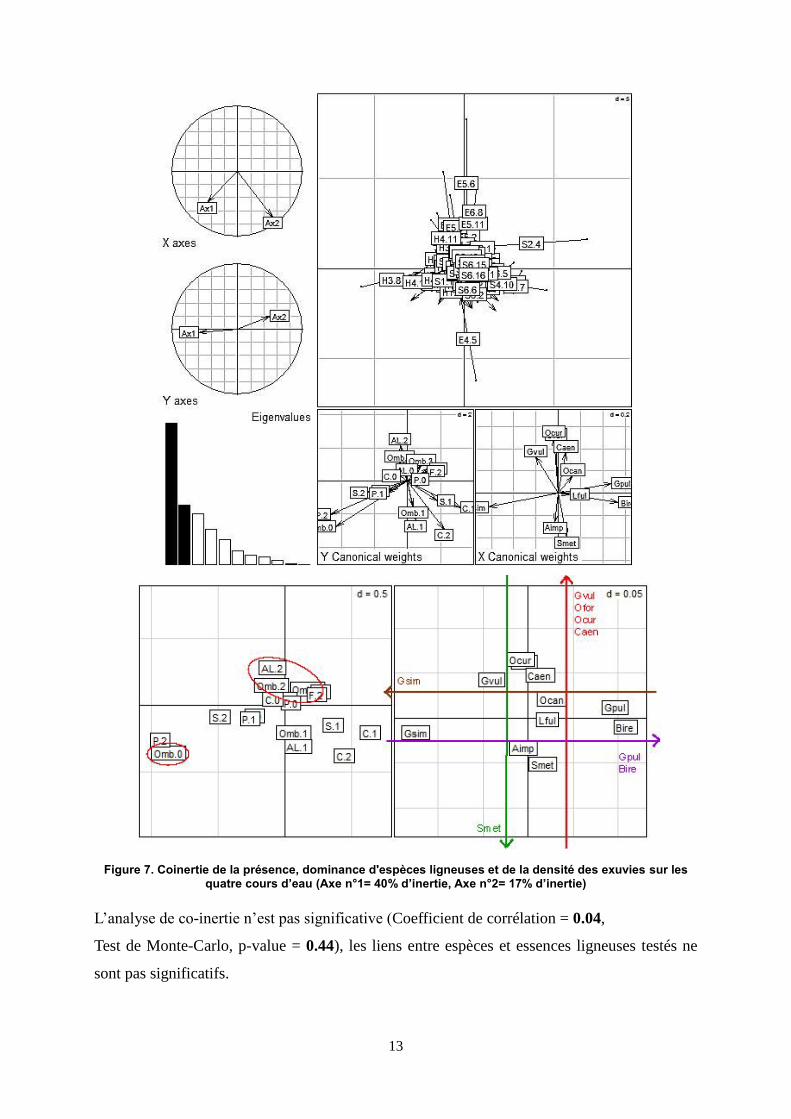
13
Figure 7. Coinertie de la présence, dominance d'espèces ligneuses et de la densité des exuvies sur les quatre cours d’eau (Axe n°1= 40% d’inertie, Axe n°2= 17% d’inertie)
L’analyse de co-inertie n’est pas significative (Coefficient de corrélation = 0.04,
Test de Monte-Carlo, p-value = 0.44), les liens entre espèces et essences ligneuses testés ne
sont pas significatifs.

14
Discussion
1) Cortèges d’espèces présents sur la Sèvre, l’Erdre, la Mayenne et l’Huisne
La valence écologique de certains Odonates des eaux courantes permet leur développement
dans différents habitats (Grand et Boudot, 2006). Cependant, un ensemble d’espèces
affectionnent plus particulièrement ces milieux d’eau courante (Dommanget, 1989).
Les caractéristiques générales des cours d’eau étudiés peuvent donc donner une première
explication sur la nature des cortèges d’espèces des quatre cours d’eau.
En effet, Gomphus vulgatissimus, Onychogomphus f.forcipatus et Oxygastra curtisii
préféreraient les eaux faiblement à moyennement courante (Meurgey et al 2000, Djikstra
2007). Ce cortège est présent sur l’Erdre et l’Huisne. Or l’Erdre, dans sa partie amont,
possède différentes physionomies, des portions à courant rapide et d’autres plus lents (SAGE
Estuaire Loire). Quant à l’Huisne, elle est alimentée par une nappe souterraine qui assure un
bon débit d’étiage, même durant les périodes les plus sèches (SAGE Huisne). Ces critères
globaux conviendraient à ces espèces.
La Mayenne, semble accueillir un cortège similaire où l’on constate cependant l’absence
d’Oxygastra curtisii. Cette rivière, très dégradée en termes de qualité d’eau et de
morphologie, a été perturbée par de nombreux ouvrages (e.g. microcentrales). Par ailleurs, des
prélèvements d’eau trop importants (e.g. eau potable, irrigation) engendrent une forte
réduction du débit ainsi que des étiages marqués, sur une grande partie du territoire (SAGE
Mayenne). Ces éléments peuvent éventuellement expliquer la richesse spécifique très faible
de cette rivière.
Le deuxième cortège présent est composé de : Gomphus pulchellus, Boyeria irene,
Somatochlora metallica et Orthetrum cancellatum. Parmi ces espèces, trois sont
caractéristiques des milieux stagnants ou très lents : Somatochlora metallica, Orthetrum
cancellatum, Gomphus pulchellus (Meurgey et al, 2000 ; Djikstra, 2007), bien que cette
dernière soit une espèce de grande valence, susceptible de se développer dans une grande
variété de milieux.
La formation de ce cortège sur la Sèvre pourrait s’expliquer par la présence de nombreux
ouvrages, notamment de chaussées de moulins, de seuils, de vannes et déversoirs, ce qui

15
ralentit considérablement le courant le long de cette rivière (SAGE Sèvre nantaise). De plus,
la dynamique d’écoulement a été perturbée par des recalibrages du lit. Ces caractéristiques en
font une rivière d’eau très calme, voire quasiment stagnante sur certaines zones. De même, la
présence de ce cortège sur l’Erdre est sans doute possible par l’existence de portions d’eaux
plus lentes.
Quant à Boyeria irene, ses larves semblent se développer aussi bien dans les eaux faiblement
courantes que vives (Votat, 1993 ; Meurgey et al, 2000).
Par ailleurs, la répartition des effectifs de ces espèces varient entre ces quatre cours d’eau. Les
peuplements d’Anisoptères de la Sèvre apparaissent très déséquilibrés. En effet, l’espèce
Gomphus pulchellus domine largement les autres. Ce phénomène intervient généralement en
réponse à des perturbations subies par le milieu, ce qui fragilisent certaines populations alors
qu’elles profitent à quelques autres (Baïlo Ndiaye, 2010). Une hypothèse peut être émise : les
nombreux aménagements et travaux réalisés sur la Sèvre ont influé sur les peuplements
d’Anisoptères. A ce stade, il est difficile d’affirmer si cela est due aux impacts directs
d’aménagements anciens, à la persistante encore aujourd’hui d’impacts induits ou à d’autres
types de perturbations, plus contemporaines, les trois pouvant agir de concert.
Les peuplements des autres cours d’eau semblent mieux équilibrés, aucune espèce ne
dominant largement les autres.
Les effectifs de Libellula fulva, Cordulia aenea et Anax imperator sont trop limité pour
permettre leur rattachement à un cortège. Cependant, ces espèces se développent
préférentiellement dans des milieux stagnants (Meurgey et al, 2000 ; Djikstra, 2007), ce qui
pourrait expliquer leur présence sur quelques sections de la Sèvre et de l’Erdre.
De même, la faible densité de Gomphus simillimus sur l’Huisne et la Mayenne, ne paraît pas
caractériser un autre cortège. Cette espèce de cours d’eau (GRETIA, 2012), se développerait
indépendamment des autres. Il est intéressant de constater que la présence de Gomphus
simillimus semble aller de pair avec l’absence de Gomphus pulchellus. Des hypothèses
peuvent être émises : les conditions écologiques préférentielles de ces espèces s’opposent, ou
bien une compétition particulière a lieu entre elles. Cependant, une autre information est à
prendre en compte : Gomphus simillimus est en marge de son aire de répartition, en Pays de la
Loire (quasiment absente en Loire-Atlantique et Vendée) (GRETIA, 2012), ce qui pourrait
expliquer son absence sur l’Erdre et sur la Sèvre.

16
2) Effet de la diversité des microhabitats ligneux sur les cortèges d’espèces
La présence ou l’absence d’arbres, sur une portion de cours d’eau, ne semble pas être un
facteur clé dans la répartition des effectifs des espèces. Cependant, la diversité des
microhabitats ligneux serait bien un critère déterminant. Plusieurs auteurs l’ont aussi
constaté : les milieux aquatiques riches en microbiotopes sont essentiels pour l’odonatofaune,
selon Dommanget, 1989 ; et plus la végétation rivulaire est variée, plus le nombre d’espèces
présentes sera grand, d’après Djikstra, 2007. A l’opposé, quand la diversité des microhabitats
est faible, certaines espèces euryèces peuvent dominer le peuplement (Ferreira-Peruquetti et
De Marco, 2002).
Or, un phénomène différent a lieu sur la Sèvre, sa richesse en microhabitats ligneux constaté
est relativement grande, et pourtant il y a dominance de l’espèce Gomphus pulchellus.
L’hypothèse de perturbations en cours sur cette rivière, déjà émise précédemment, peut être à
nouveau suggérée. Selon Moore (1991), des changements importants dans la végétation
rivulaire, par la mise en place de nouveaux aménagements, peuvent modifier l’équilibre de
ces peuplements. Baïlo Ndiaye (2010) mentionne également que l’abondance et la diversité
des Odonates peuvent être bouleversées par des changements de masses d’eau et de la
végétation associée.
La diversité des microhabitats ligneux influerait bien sur la richesse spécifique des espèces
présentes. L’hypothèse selon laquelle des cortèges particuliers seraient liés à la nature et la
dominance des essences rivulaires n’a pas été confirmée. Cependant, d’après plusieurs études
(Votat 1993, Doucet, 2009), certaines interactions semblaient possible. Par exemple, entre
Oxygastra curtisii et la dominance de l’aulne, ou bien avec un ombrage moyen de la berge.
En effet, il a déjà été prouvé qu’Oxygastra curtisii montrait une préférence pour les cours
d’eau ombragés (Dommanget, 1987 ; Heidemann et Seidenbusch, 2002). Son développement
larvaire aurait lieu dans, ou à proximité, des lacis racinaires de certains arbres (Leipelt &
Suhling, 2001). Or l’Erdre, rivière où cette espèce est la plus abondante parmi les cours d’eau
étudiés, est bien celle qui compte le plus grand ombrage globale, ce qu’avait déjà constaté à
juste titre Gurliat (2004).
De plus, des femelles d’Oxygastra curtisii ont été observées pondant autour du système
racinaire des aulnes (Alnus sp.) (Votat 1993, Herbrecht et Dommanget, 2006). Ses larves
étant très sédentaires (com. pers. Herbrecht), elles restent fixées dans les racines ou le

17
sédiment jusqu’à leur émergence. Ainsi, les exuvies sont majoritairement ramassées sur cette
essence (Doucet, 2009 ; Douillard et al., 2004). Elle jouerait donc un rôle primordial dans le
cycle de vie d’Oxygastra curtisii.
Ce choix porté sur l’aulne, ou parfois sur le saule (Herbrecht et Dommanget, 2006),
s’expliquerait par la faculté de ces essences à émettre un important lacis racinaire immergé.
Ce microhabitat forme une niche particulière et permet aux larves de se cacher.
Les espèces supposées associées à Oxygastra curtisii : Gomphus vulgatissimus et
Onychogomphus f.forcipatus, seraient donc, elles aussi, potentiellement liées à des milieux
ombragés. La Mayenne compte bien une grande partie de zones prospectées avec un ombrage
moyen à fort, ce qui pourrait expliquer le développement plus abondant de Gomphus
vulgatissimus. Quant à Onychogomphus f.forcipatus, elle est présente sur les quatre cours
d’eau, il est donc difficile de définir un lien particulier. Il est à noter que sa densité est plus
importante sur l’Erdre, ce qui pourrait être lié à la majorité de sous-sections à fort ombrage.
Cependant, et contrairement à Oxygastra curtisii, elles semblent aussi se développer dans les
zones à plus faible ombrage.
Enfin, l’Huisne s’illustre par la présence de Gomphus simillimus. Cette espèce est quasi-
absente des autres cours d’eau et, par conséquent, elle semble affectionner les conditions de
vie particulières de cette rivière. La particularité de l’Huisne est qu’elle possède de
nombreuses zones dépourvues d’arbres, l’ombrage y est donc plus faible que sur les autres
cours d’eau. Ce critère pourrait être en lien avec le développement de cette espèce.
Conclusion
Les espèces d’Anisoptères et leur environnement sont liés par des relations complexes
difficiles à définir. De multiples facteurs sont impliqués tout au long de leur cycle de vie.
Les effets de certaines variables, telles que la nature des sédiments, la nature et la présence
d’hydrophytes, d’hélophytes, d’herbacées ainsi que la qualité physico-chimiques de l’eau,
restent encore à étudier sur ces rivières. De plus, l’échantillonnage réalisé n’a pu être
standardisé en termes de longueur totale de berge prospectée par cours d’eau, les
comparaisons entre sites semblent donc moins pertinentes.

18
Les recherches doivent être poursuivies sur plusieurs années et plusieurs cours d’eau pour
tenter de définir les facteurs impliqués dans la présence de cortèges particuliers d’espèces
d’Anisoptères.

19
Bibliographie
Baïlo Ndiaye A., 2010, Intégration de la biodiversité d’eau douce dans le processus de
développement en Afrique, Module de formation des formateurs sur le suivi des Odonates,
47pp, Document non publié.
Cherpitel T., Herbrecht F., 2012, Clé de détermination des exuvies d’Anisoptères, Groupe
d’étude des invertébrés armoricains, 11pp, Document non publié.
Chessel D. et Mercier P., 1993, Couplage de triplets statistiques et liaisons espèces-
environnement, in J. D. Lebreton and B. Asselain, Biométrie et Environnement, Masson,
Paris, pages : 15-44.
Corbet P.S., 1999, Dragonflies: Behaviour and Ecology of Odonata, Editions Harley Books,
Colchester: 862 pp.
De paiva Silva D., De Marco P., Chaves Resende D., 2010, Adult odonate abundance and
community assemblage measures as indicators of stream ecological integrity: A case of study,
Elsevier, Ecological Indicators 10: 744-752.
De Ricqles A., 1988, Les odonates de Dordogne et leur intérêt comme indicateurs de
l’évolution des milieux à moyen terme, Revue d’Ecologie Terre Vie, 43 : 177-194.
Djikstra K.-D.B, 2007, Guide des libellules de France et d’Europe, Delachaux et Niestlé,
320pp.
Dolédec, S., and D. Chessel, 1994, Co-inertia analysis: an alternative method for studying
species-environment relationships, Freshwater Biology 31:277-294.
Dommanget J-L., 1987, Etude faunistique et bibliographique des odonates de
France. Inventaire de faune et de flore (MNHN), Secrétariat de la Faune et de la Flore, Paris,
36 : 283 pp.

20
Dommanget J-L., 1989, Utilisation des Odonates dans le cadre de la gestion des zones
humides, Beaufort F.et Maurin H. (Editeurs), Utilisation des inventaires d’invertébrés pour
l’identification et la surveillance d’espaces de grand intérêt faunistiques, Inventaires de Faune
et de Flore (MNHN), Secrétariat de la Faune et de la Flore, Paris, 53 : 93-110.
Doucet G., 2009, Suivi de l’émergence d’Oxygastra curtisii et de Gomphus Graslinii sur un
étang du centre de la Dordogne, Martinia, 25(4).
Douillard E., Durand O., Gabory O., Samson N., 2004, Du nouveau sur le cycle biologique et
l’état des populations de La Cordulie à corps fin (Oxygastra curtisii, Dale, 1834) dans les
Mauges, Mauges Nature bulletin de synthèse n°6 – 2004, 63-67.
Ferreira-Peruquetti, P.S., De Marco Jr., P., 2002, Efeito da alteracão ambiental sobre
comunidades de Odonata em riachos de Mata Atlântica de Minas Gerais, Brasil, Revue
Zoologique brésilienne, 19: 317–327.
Grand D., Boudot J-P., 2006, Les libellules de France, Belgique et Luxembourg, Biotope,
Mèze (Collection Parthénope), 480 pp.
GRETIA, 2012, Plan national d’actions en faveur des odonates : Déclinaison Pays de la Loire
(2012-2015), Rapport pour la DREAL Pays de la Loire, 203pp.
Gurliat P., 2004, Contribution à la connaissance des Odonates de l'Erdre et de ses affluents.
Martinia, 20(3) : 125-130.
Heidemann H. et Seidenbusch R., 2002, Larves et exuvies des libellules de France et
l’Allemagne (sauf Corse), Société française d’Odonatologie, Bois-d’Arcy, 416pp.
Herbrecht F. et Dommanget J-L., 2006, Sur le développement larvaire d’Oxygastra curtisii
(Dale, 1834) dans les eaux stagnantes (Odonata, Anisoptera, Corduliidae), Martinia, vol. 22
(2) : 89-94.
Houard X., Jaulin S., Dupont P. et Merlet F., 2012, Définition des listes d’insectes pour la
cohérence nationale de la TVB -Odonates, Orthoptères et Rhopalocères, OPIE, 29 pp + 71 pp

21
d’annexes.
Jarry D. et Vidal H., 1960, Introduction à l’étude écologique des Odonates de la région
montpelliéraine, Vie et Milieu, 11 : 261-283.
Leipelt K.G., Suhling F., 2001, Habitat selection of larval Gomphus graslinii and Oxygastra
curtisii (Odonata: Gomphidae, Corduliidae), International Journal of Odonatology, 4 (1) : 23-
34.
Masselot G. et Nel A., 2003, Odonates sont-ils des taxons bio-indicateurs ?, Martinia, 19(1):5-
38.
Meurgey F., Herbrecht F., Gurliat P., Dortel F., Boureau A., Dusoulier F. et Williamson T.,
2000, Atlas préliminaire des Odonates de Loire-Atlantique, Martinia 16(1) : 28.
Moore N.W., 1991, The development of dragonfly communities and the consequences of
territorial behavior: a 27 years study on small ponds at Woodwalton Fen, Cambridgeshire,
United Kingdom, Odonatologica, 25: 49-58.
Oertli B., 2008, The use of dragonflies in the assessment and monitoring of aquatic habitats,
In Cordoba-Aguilar A. (Ed.), Model Organisms for Ecological and Evolutionary Research,
Oxford University Press, Oxford, 79-95.
Schmidt E., 1985, Habitat inventarization, characterization, and bioindication by a
“representative spectrum of odonata species (RSO)”, Odonatologica, 14: 127-133.
Votat P., 1993, les Odonates du Nord-Est de la Mayenne, du Sud-Ouest de l’Orne et du Nord-
Ouest de la Sarthe (suite). Notes sur quelques espèces remarquables ou rares. Martinia 9(2) :
35-41.
Wootton R.J., 1981, Palaeozoic insects, Annual Review of Entomology, 26: 319-344.

22
Sites internet
http://www.sagemayenne.org
http://www.sagehuisne.org
http://www.sevre-nantaise.com
http://www.sage-estuaire-loire.org

23
Résumé
L’ordre des Odonates constitue l’un des groupes d’insectes caractéristiques des écosystèmes
aquatiques. La présence de cortèges particuliers d’espèces serait influencée par l’état général des cours
d’eau. Il est donc primordial d’approfondir les connaissances sur les liens entre ces cortèges et leur
environnement.
Basée sur la collecte systématique des exuvies du sous-ordre des Anisoptères, cette étude a été réalisée
sur des tronçons de quatre cours d’eau des Pays de la Loire. La composition et la structuration végétale
ligneuse des berges prospectées ont été relevées.
Deux cortèges d’espèces ont été identifiés sur les rivières. Les espèces Gomphus vulgatissimus,
Onychogomphus f.forcipatus et Oxygastra curtisii, composent le premier groupe. Elles affectionnent
particulièrement les eaux faiblement à moyennement courantes, présentes sur l’Erdre et l’Huisne. Pour
le deuxième, composé de Gomphus pulchellus, Somatochlora metallica et Orthetrum cancellatum, ce
sont les milieux d’eaux lentes à stagnantes qui sont privilégiés. Sur plusieurs des rivières, de
nombreux ouvrages et aménagements ont considérablement ralenti le courant de l’eau, ce qui explique
la présence de ce cortège.
Par ailleurs, la diversité des berges en microhabitats est de grand intérêt pour l’odonatofaune. Plus la
végétation rivulaire ligneuse est variée, plus la richesse spécifique des Anisoptères sera grande. Des
liens directs entre la présence de certaines essences et celle de libellules n’ont cependant pas été
identifiés.
Néanmoins, des interactions déjà mise en évidence ont à nouveau été constatées, telles que la
préférence d’Oxygastra curtisii pour les berges ombragées. D’autres restent inexpliquées, comme la
présence discrète de Gomphus simillimus sur certains cours d’eau.
Mots clés: Odonates, Indicateurs, Berge, Arbre, Ombrage


















