
L’impact des nuisances acoustiques sur les cétacés du ...
182
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 1 L’impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale Dr Alexandre Gannier Klymene Recherche Marine Contrat n°13031 – 83400PC du 30 octobre 2013 Rapport Final 16 avril 2014
Transcript of L’impact des nuisances acoustiques sur les cétacés du ...

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 1
L’impact des nuisances acoustiques sur les cétacés du
Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier
Klymene Recherche Marine
Contrat n°13031 – 83400PC du 30 octobre 2013
Rapport Final
16 avril 2014

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 2
Résumé :
La contribution des pollutions acoustiques à la dégradation des habitats des cétacés avait
été considérée comme secondaire jusqu’à une époque assez récente, en Méditerranée et ailleurs.
Cette étude propose des moyens de l’évaluer, permettant ainsi la prévention des impacts des
fortes intensités sonores sur les cétacés. Dans une première partie, nous donnons les clefs de la
compréhension de cette problématique complexe : le monde acoustique sous-marin, les facultés
acoustiques des cétacés. Nous nous focalisons sur les huit espèces communes en Méditerranée
occidentale, en fournissant un catalogue des émissions sonores de ces espèces. Dans une
seconde partie, nous exposons l’état de l’art : dans ce domaine plus que dans d’autres, les
avancées scientifiques majeures sont très récentes. L’examen des résultats récents nous permet
de proposer une typologie des impacts provoqués par les pollutions acoustiques sur les cétacés :
de la simple perception jusqu’à la lésion traumatique grave, constatée dans certains cas d’intensité
sonore très forte. Dans une troisième partie, nous élaborons un modèle de prévision des impacts
largement basé, mais pas uniquement, sur la notion de « dose sonore perçue » : pour des sons
impulsifs (type sismique ou battage de pieux) et des sons non impulsifs (type sonar), ce modèle
permet d’estimer des impacts allant du dérangement léger (sans interruption d’activité) jusqu’à la
lésion létale, en passant par la désertion d’un habitat. Nous appliquons ce modèle à un cas
générique de pollution sonore de type « transmission de sonar à moyenne fréquence », simulation
effectuée grâce à un logiciel spécialisé (et développé antérieurement à l’étude). La quatrième
partie est une discussion des points faibles et des points forts de l’approche, au regard de ce qui
se pratique ailleurs dans le monde. Cette partie est importante, car la problématique de la
prévision des impacts sonores sur les cétacés est en rapide évolution, et il faut avoir conscience
des zones sombres qui subsistent. Du reste, on constate malheureusement chaque année que
des accidents graves se produisent en raison de l’imperfection des méthodes de mitigation, ou de
leur mauvaise mise en pratique. L’imperfection des méthodes est largement due à l’imprécision
inhérente à chaque étape de la démarche de prévision, qui est complexe. Mais cette étude permet
de connaître les phénomènes qui se produisent lorsque des cétacés sont soumis à de fortes
intensités sonores, et fournit des moyens permettant de prévenir les effets les plus graves.
Citation :
GANNIER A., 2014. - L’impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la
Méditerranée nord-occidentale. Klymene Recherche Marine et Sanctuaire Pelagos (Partie
française) : 182 p.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 3
Introduction _______________________________________ ________________5
A\ Eléments fondamentaux ____________________________ _____________8
1) Eléments d’acoustique ____________________________ __________________ 8
a) Les ondes sonores ________________________________________________________ 8b) Différentes catégories de sons _______________________________________________ 9c) Ambiance sonore sous-marine______________________________________________ 10
2) Le monde acoustique des cétacés __________________ _________________ 13
a) Audition des cétacés______________________________________________________ 13b) Production des sons par les cétacés _________________________________________ 18c) Sensibilité des cétacés aux nuisances sonores _________________________________ 22
3) Répertoires des huit espèces de cétacés dans le S anctuaire _____________ 24
a) Répertoire du Dauphin bleu et blanc _________________________________________ 25b) Répertoire du Dauphin commun_____________________________________________ 27c) Répertoire du Grand dauphin _______________________________________________ 28d) Répertoire du Dauphin de Risso_____________________________________________ 29e) Répertoire du Globicéphale noir_____________________________________________ 30f) Répertoire du Ziphius de Cuvier_____________________________________________ 32g) Répertoire du Cachalot____________________________________________________ 33h) Répertoire du Rorqual commun _____________________________________________ 35
B\ Synthèse bibliographique _________________________ ______________36
1) Principales sources anthropiques pouvant impacter les cétacés __________ 36
a) Les explosions __________________________________________________________ 38b) Le battage de pieux ______________________________________________________ 42c) Le forage_______________________________________________________________ 45d) Les sismiques pneumatiques _______________________________________________ 48e) Les sonars _____________________________________________________________ 62f) Le trafic maritime ________________________________________________________ 78
2) Synthèse des effets des nuisances sonores sur les cétacés ______________ 84
a) Lésions traumatiques graves ou létales _______________________________________ 84b) Effets auditifs mesurés lors d’expériences en bassin_____________________________ 89c) Effets observés lors d’expériences d’exposition contrôlée________________________ 108d) Autres effets comportementaux observés ____________________________________ 114e) Synthèse des effets des nuisances sonores __________________________________ 117
3) Typologie retenue pour la gradation des effets de s nuisances sonores____ 119
a) Rationalité fondamentale de l’approche ______________________________________ 119b) Les différents niveaux de perturbation _______________________________________ 121
C\ Résultats ________________________________________ ____________125
1) Proposition d'un modèle de prévision des effets __ ____________________ 127
a) Variante pour les sons non-impulsifs ________________________________________ 128b) Variante du modèle pour les sons impulsifs (type sismique ou explosion) ___________ 132c) Audiogrammes de référence ______________________________________________ 135
2) Prévision des distances significatives pour les e ffets __________________ 141
a) Utilisation d’un modèle de propagation sphérique ______________________________ 142b) Limites du modèle de propagation sphérique__________________________________ 144

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 4
3) Application du modèle à un cas de forte nuisance sonore_______________ 147
a) Proposition d’un scénario de transmission de sonar ____________________________ 148b) Peuplement de cétacés considéré __________________________________________ 149c) Impact de la transmission sonar sur le peuplement _____________________________ 150
D\ Discussion _______________________________________ ___________157
1) Points forts et limites du modèle proposé ________ ____________________ 159
a) Sur les métriques employées ______________________________________________ 159b) Sensibilités des résultats aux seuils adoptés pour les effets comportementaux _______ 160c) Cas spécifique du Ziphius_________________________________________________ 161
2) Comparaison avec d'autres approches disponibles __ __________________ 163
a) Audiométrie par espèce ou regroupement d’espèces ?__________________________ 163b) Type de pondération fréquentielle __________________________________________ 164
Conclusion _________________________________________ _____________167
Références bibliographiques ________________________ _______________168
Index des figures __________________________________ _______________177
Index des tableaux _________________________________ _______________180
Annexes ____________________________________________ ____________182

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 5
Introduction
Depuis une trentaine d’années, on s’est progressivement rendu compte que les cétacés
pouvaient non seulement être dérangés dans leur cycle biologique par des pollutions sonores
d’origine humaine, mais aussi être mortellement affectés par des intensités sonores élevées.
Longtemps délaissée par les autorités chargées de réglementer les impacts des activités
humaines sur le milieu marin, la menace que fait peser la pollution acoustique sur la viabilité des
habitats de cétacés est maintenant prise au sérieux1. Cette problématique fait d’ailleurs partie des
aspects devant être pris en compte pour que le Sanctuaire Pelagos demeure un espace favorable
aux huit espèces de cétacés que l’on y rencontre régulièrement.
Au sein du milieu marin, les cétacés sont des prédateurs qui doivent se déplacer sur des
dizaines ou des centaines de kilomètres pour localiser leur proies, que ce soit une concentration
d’euphausiacés, des bancs de poissons, ou une agrégation de calmars océaniques. Une fois à
quelques kilomètres d’une ressource, les cétacés doivent approcher et capturer les proies en
évoluant au sein d’un milieu à trois dimensions où la visibilité s’étend au mieux à quelques
dizaines de mètres, et souvent bien moins. En Méditerranée nord-occidentale, la prédation des
cétacés se déroule en grande partie entre le crépuscule et l’aube, souvent en profondeur, dans
des conditions d’obscurité. Dans ce contexte, c’est le sens auditif qui est utilisé par les cétacés
pour localiser les proies, que ce soit le mysticète de Méditerranée, ou les odontocètes qui
approchent et capturent leurs proies par écholocalisation.
La plupart des espèces a une sociabilité élevée : le groupe a une importance primordiale
dans tous les aspects du cycle biologique des cétacés, y compris bien sûr la reproduction, la
protection contre les prédateurs, et les stratégies pour localiser et capturer les proies. Mis à part
pour la communication très rapprochée, c’est encore le sens acoustique qui est utilisé pour
organiser les individus dans l’ensemble de ces fonctions vitales. Les baleines, cachalots, dauphins
et autres odontocètes échangent des signaux sonores durant leur activité, pour se coordonner,
mais aussi pour reformer les groupes après qu’au cours d’une phase d’activité les individus se
soient dispersés. Dans ce cas également, des distances de plusieurs centaines, plusieurs milliers
de mètres, voire plus, séparent les conspécifiques, et il est vital que les signaux de communication
portent suffisamment loin.
Pour ces raisons, il est justifié d’affirmer que les caractéristiques acoustiques du milieu marin
ont une importance primordiale pour les cétacés. C’est pour cette raison que les facultés auditives
des cétacés sont très développées, englobant un grand domaine de fréquence (de 10 Herz à
200 kHz), variable selon les espèces, et une gamme de sensibilité très élevée : un dauphin est
capable d’entendre des pressions sonores aussi faibles que 10-4 Pa et de supporter une pression
1 Voir la Directive européenne EU 2008-56 du 25 juin 2008.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 6
sonore dépassant 104 Pa. Par conséquent, les cétacés sont extrêmement sensibles aux
modifications de leur environnement sonore.
Comme l’anatomie de leur oreille interne est analogue à celle des autres mammifères, y
compris l’Homme, face à une pollution sonore, les cétacés développent une gamme de réactions
qui est similaire à celle qui est observée chez les mammifères terrestres, allant de la simple gêne à
une surdité définitive, en passant par la douleur et l’altération temporaire de l’audition (ou TTS en
anglais2). Il arrive aussi que dans certaines configurations, une réaction comportementale
d’évasion en réponse à une nuisance sonore élevée entraîne l’échouage massif de cétacés, et
donc très souvent la mort de nombreux individus. Un échouage aux conséquences fatales peut
également se produire alors que des individus ont subi des altérations de leur audition ou des
lésions internes non fatales à la suite d’une insonification accidentelle. Enfin, en présence
d’intensités sonores extrêmement fortes, les cétacés peuvent subir des lésions mortelles
immédiates. Face à une pollution sonore, la gamme des effets possibles est donc très étendue. La
région du Sanctuaire Pelagos n’est pas épargnée par l’ensemble des nuisances sonores
anthropiques que l’on rencontre sur la planète.
Le Sanctuaire Pelagos et les régions adjacentes sont situés dans une zone maritime très
fréquentée, où l’on trouve l’ensemble des activités humaines, la plupart étant malheureusement
bruyantes. Le trafic maritime commercial y est intense en raison de nombreux ports de commerce,
et de l’attrait touristique de la région, le trafic de plaisance à moteur y est extrêmement dense
pendant la saison chaude (et en expansion rapide depuis deux décennies), périodiquement des
campagnes de prospection sismique ont lieu, ainsi que des travaux lourds d’agrandissement de
port ou d’infrastructure, enfin il y a plusieurs ports militaires en France et en Italie. Les flottes
militaires comprennent des frégates anti-sous-marines (ASM), des navires de guerre des mines,
etc., qui sont munis de sonars de forte puissance ; les avions et les hélicoptères de lutte ASM sont
munis également des sonars aéroportés (bouées acoustiques, sonars trempés). L’ensemble de
ces moyens militaires présente un grand potentiel de nuisance acoustique pour les cétacés, même
si ce trafic militaire n’est pas le plus important, numériquement. L’utilisation de sonars ASM est à
l’origine de plusieurs échouages massifs de Ziphius, depuis 1963, le dernier en date s’étant produit
au début du mois d’avril 2014, en Grèce.
Si le traité du Sanctuaire Pelagos mentionne explicitement les pollutions acoustiques, il ne
les proscrit pas. Déjà bruyante il y a une vingtaine d’années3, l’ambiance sonore dans les eaux de
Méditerranée nord-occidentale s’est fortement dégradée, notamment dans les eaux du Sanctuaire.
Certains indices suggèrent que des portions d’habitats normalement favorables seraient en passe
d’être délaissées, ce qui pourrait conduire à la disparition de certains groupes résidents de cétacés
2 TTS pour Temporary Threshold Shift, soit une hausse temporaire du seuil d’audition (la hausse du seuilsignifiant une moins bonne audition).
3 Selon les enregistrements que nous avons réalisés à cette époque.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 7
de plusieurs espèces. Il est donc indispensable de porter la problématique de l’effet des nuisances
sonores sur les cétacés à la connaissance d’un plus grand nombre de personnes, d’attirer
l’attention sur les effets les plus nocifs des fortes intensités sonores, de disposer d’outils
permettant de déterminer l’ampleur des impacts produits. C’est l’objet de cette étude.
Dans une première partie, on présentera les éléments fondamentaux de l’acoustique, étape
nécessaire pour évoquer ensuite le monde acoustique des cétacés, ce qui nous amènera à une
description du répertoire acoustique des cétacés de Méditerranée nord-occidentale, associée à
des documents sonores.
Ensuite, on passera en revue les principales sources de nuisance sonore qui ont des effets
sur les cétacés, avant de présenter une synthèse des éléments scientifiques qui permettent
d’élaborer une échelle des effets des pollutions sonores sur les différentes espèces, en me
focalisant sur les espèces du Sanctuaire, ou sur des espèces très voisines. Cet examen permettra
de présenter une typologie de ces effets, en partant du plus bénin jusqu’au plus grave, et en les
mettant en rapport avec les niveaux d’insonification auxquels sont soumis les cétacés.
Une échelle de détermination des effets des insonifications sera proposée, elle exprimera les
différents effets produits en fonction des niveaux sonores perçus ou reçus, selon le cas. Des
courbes audiométriques permettront de transformer, pour chaque espèce, les niveaux sonores
reçus en niveaux perçus. En couplant ces niveaux sonores à une loi de propagation du son, on
pourra déterminer les distances correspondant aux différentes classes d’impact pour une source
sonore de caractéristiques données. Le tout constituera mon modèle de prévision des impacts,
modèle que j’appliquerai ensuite à un cas typique de nuisance sonore, celui d’une transmission de
sonar anti-sous-marin4. Cet exemple permettra de matérialiser l’impact provoqué par une source
sonore de forte puissance sur un peuplement méditerranéen de cétacés5.
Enfin, on comparera l’approche proposée à d’autres méthodes disponibles dans la littérature,
en soulignant les points forts du modèle, tout en expliquant pourquoi il demeure néanmoins d’une
précision limitée.
L’ensemble de l’étude donnera des bases solides pour comprendre l’impact des fortes
intensités sonores sur les cétacés, et fournira le moyen d’évaluer l’ordre de grandeur des impacts
dans des cas de figures typiques.
4 Les caractéristiques de ce sonar seront cependant « génériques », sans rapport précis avec un engin réel,de même que les conditions de transmission des « pings » du sonar.5 Un second cas d’application, celui d’une prospection sismique par canon pneumatique, avait été envisagéau départ de l’étude ; mais finalement, il n’a pas été retenu.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 8
A\ Eléments fondamentaux
Définir les bases de l’acoustique, décrire les différents types élémentaires de sons, et les
composantes de bases de l’ambiance sonore sous-marine. Décrire les spécificités des cétacés en
terme de production et de réception des sons, fournir une base de données du répertoire des
espèces de Méditerranée nord-occidentale, aborder les conséquences des nuisances sonores
d’origine anthropique.
1) Eléments d’acoustique
a) Les ondes sonores
Le son est une onde de pression caractérisée par sa célérité de propagation et sa fréquence,
lesquels permettent de calculer la longueur d’onde. La célérité du son n’est pas fixe, dans l’eau
elle varie en fonction de la densité (donc de la température, de la pression et de la salinité), mais
elle est moins variable que dans l’air. La célérité du son dans l’eau de mer vaut de l’ordre de
1530 m/s (contre 341 m/s dans l’air à 15°C). La fré quence se définit en Herz (ou cycle/s), par
exemple une fréquence de 440 Hz correspond au « la » des musiciens. Pour chaque espèce
animale dotée d’une ouïe, il y a une fréquence en deçà de laquelle les sons ne sont pas entendus
(fréquence min pour l'homme ≅≅≅≅ 20 Hz), et une fréquence au-delà de laquelle les sons ne sont pas
audibles (fréquence max pour l'homme ≅≅≅≅ 18 kHz). Entre ces deux bornes, qui limitent le domaine
audible d’une espèce, les sons ne sont pas entendus avec la même sensibilité.
La longueur de l’onde de pression sonore est définie par la relation λ = c / f , où λ est en m
quand c est en m/s et f est en Herz. Ce qui par exemple donne pour un « la », une longueur
d’onde de 341/440 = 0,77 m dans l’air, mais de 1530/440 = 3,47 m dans l’eau de mer. Cette
notion est importante car la longueur d’onde est en rapport avec les organes qui produisent et
reçoivent les sons. De plus, la détection d’un objet par écholocalisation n’est pas possible quand
cet objet est significativement plus petit que la longueur d’onde6.
Toutes espèces confondues, les cétacés sont concernées par des fréquences allant de
10 Hz (appels des grands balénoptères) à 150-200 kHz (écholocalisation de certains delphinidés),
soit des longueurs d’onde allant de 150 m à 1 mm, environ.
La puissance sonore d’une source est établie en watt, comme toutes les puissances, et
l’intensité sonore se définit comme un flux de puissance à travers une unité de surface, en W/m2.
On démontre que cette intensité I est reliée à la pression sonore p par la relation simplifiée
suivante : I = p2 / ρ.c , avec ρ la masse volumique du fluide. En raison de la différence entre les
masses volumiques de l’air et de l’eau, ainsi que la différence entre leurs célérités, une puissance
6 En fait, c’est une variable appelée « nombre d’onde » qui intervient.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 9
acoustique donnée (par ex. 1 W) génère une pression sonore 60 fois plus élevée dans l’eau que
dans l’air. C’est pour cette raison que l’eau est un milieu très favorable à la transmission des sons.
• Les décibels
En acoustique, la quantification des niveaux sonores et un grand nombre de calculs
s’effectuent en utilisant les décibels. Cette notation permet d’exprimer les niveaux sonores en
fonction de leur perception par l’ouïe humaine (et celle des mammifères), comme le niveau de
pression sonore :
Lp = 20 log10 (p/p0) , p0 = 1 µPa dans l’eau (20 µPa dans l’air)
L’échelle des pressions sonores audibles (dans l’air, pour l’homme, de 20 µPa à 100 Pa) est
ainsi réduite à une échelle allant de 0 à 133 dB. On dit conventionnellement que 120 dB est le
seuil de douleur pour la fréquence de 1000 Hz. Notons que la quantité de 1 décibel (dB) est en fait
la variation minimale à laquelle est sensible une oreille avertie. Dans cette étude, l’expression
« niveau sonore » sera équivalente à l’expression « niveau de pression sonore ».
Les décibels s’appliquent aussi aux autres variables utilisées en acoustique ; une autre
grandeur que nous utiliserons est le niveau de source, Ls, qui égal au niveau de pression sonore
mesuré à une distance de 1 m de la source. Le niveau de source est la manière habituelle
d’exprimer la puissance acoustique d’une source sonore, il est noté ainsi : dB re 1µPa@1m. Des
niveaux de source supérieurs à 200 dB indiquent des sources sonores de très forte puissance.
On observe que la notation « décibel » s’applique à des grandeurs de nature diverse.
b) Différentes catégories de sons
L’évolution de la pression sonore dans le temps, et son contenu en fréquence7 sont les deux
caractéristiques qui permettent de classer les sons en trois catégories principales :
• les sons impulsifs
Les sons extrêmement brefs produits par des chocs, des chutes, des claquements, des
explosions appartiennent à la catégorie « impulsions ». Leur durée est de l’ordre de quelques µs à
quelques ms. Quelques exemples : un claquement de main, un claquement de cavitation, un clic
de cachalot, une explosion chimique ou pneumatique.
Les clics d’écholocalisation des odontocètes sont des impulsions, ils peuvent dans certains
cas être à bande étroite et modulation de fréquence (marsouins, ziphiidés, kogiidés, quelques
dauphins), mais ils sont le plus souvent à bande large (cachalot, la plupart des dauphins).
o Exemple sonore 1: airgun
7 Pour les besoins de cette étude, on définira la basse fréquence pour des sons à moins de 1 kHz, lamoyenne fréquence pour des sons compris entre 1 et 5 kHz, la haute fréquence pour des sons comprisentre 5 et 20 kHz, et ultra-haute fréquence pour ce qui est au delà.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 10
• les sons purs et composés
Il s’agit de sons de durée importante (en tout cas supérieure à quelques 1/100è de s), dont la
caractéristique en fréquence est une sinusoïde (son pur), ou plusieurs sinusoïdes dont les
fréquences « harmoniques » sont multiples d’une fondamentale (cas du son composé).
Les sifflements, y compris ceux de cétacés, sont des exemples de sons purs ou composés,
selon le cas, ainsi que certains cris ou appels de baleine. Les sonars scientifiques ou militaires
émettent également ce type de son.
Dans la plupart des cas, les sifflements de dauphins ont une fréquence modulée, qui varie
régulièrement au cours de la durée du sifflement. Cette durée est en général comprise entre moins
d’un dixième de seconde et quelques secondes.
Exemple sonore 2: sonar
• les cris pulsés
Les moyens et grands delphinidés produisent aussi des sons qui ressemblent à des sons
composés mais qui sont en fait des trains d’impulsions très serrées. A l’oreille, ils rendent comme
des miaulements, ou des couinements, etc.
Exemple sonore 3: cri pulsé d’orque
• les bruits
En acoustique, le terme « bruit » a une signification précise : il s’agit d’un son dont le
contenu en fréquence n’est pas « discret », c’est-à-dire qui n’est pas composé d’une ou plusieurs
fréquences particulières, mais d’un nombre infini de fréquences élémentaires adjacentes qui
forment un spectre continu.
Le bruit du vent, les bruits d’hélice, les frottements divers, sont des exemples de bruits. Un
cas courant de bruit utilisé en laboratoire est le « bruit blanc », pour lequel la densité spectrale est
constante entre deux bornes. En milieu sous-marin, les bruits masquent les signaux de
communication, en particulier ceux des cétacés.
Exemple sonore 4: bruit de NGV
c) Ambiance sonore sous-marine
Les bruits produits au-dessus de la surface ne pénètrent pratiquement pas le milieu
aquatique ; de même, les sons produits au sein de la colonne d’eau s’en échappent très peu à
travers la surface, en raison des très grandes différences d’impédance acoustique. Une proportion
mineure des bruits sous-marins sont transmis par le sol, en fonction de la nature du fond. Au total,
le volume sous-marin se comporterait comme un réservoir acoustique si l’énergie sonore n’y était
pas progressivement absorbée par des effets mécaniques et électrochimiques. L’absorption du
son dans l’eau au fur et à mesure de son parcours dépend énormément de la fréquence : à
100 Hz, l’absorption n’est que de 10-3 dB par km, alors qu’à 100 kHz elle vaut 20 dB par km. Donc,
le milieu sous-marin se comporte un peu comme un réservoir de bruits à basse fréquence.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 11
La conséquence de ces phénomènes est que le monde sous-marin est naturellement
bruyant pour les basses fréquences, alors que pour les hautes fréquences le bruit naturel ambiant
est plutôt faible. Un grande nombre de phénomènes naturels contribuent au bruit ambiant, y
compris la composante biologique, et les aspects climatiques : ainsi, le vent, les vagues, la pluie,
ont une grande influence sur le niveau de bruit ambiant. Pour prévoir le bruit ambiant sous-marin,
on utilise des abaques issus des courbes de Wenz, qui indiquent la densité spectrale de bruit en
fonction de la fréquence et de nombreux paramètres (Fig.1).
Exemple sonore 5: Cachalot en ambiance sous-marine peu anthropisée
Le bruit ambiant a tendance à masquer les sons des cétacés, en fonction des fréquences
utilisées par les différentes espèces. Plus le bruit ambiant augmente, plus les cétacés doivent
augmenter le niveau de leurs vocalisations pour maintenir une même distance de communication
ou d’écholocalisation.
Toutes les composantes anthropiques du bruit sous-marin augmentent le bruit ambiant et ont
tendance à entraver les communications des cétacés. En raison de l’effet « réservoir » du milieu
sous-marin et de la faible absorption des sons à basse fréquence, le bruit basse fréquence a
fortement augmenté dans les 50 dernières années du fait de la circulation de milliers de navires
marchands dans presque toutes les zones géographiques du monde. McDonald et al. (2006) ont
ainsi mesuré une hausse de 15 dB du bruit pour des fréquences de 30 à 50 Hz, au large de la
Californie. En Méditerranée nord-occidentale, il n’y a pas de résultat publié sur l’évolution du bruit
ambiant, mais depuis deux décennies l’augmentation est très forte8.
Exemple sonore 6: Cachalot en ambiance sous-marine anthropisée
8 Tendance démontrée par nos enregistrements, notamment dans la bande des 0-20 milles.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 12
Figure 1 : Abaque de bruit ambiant sous-marin (d’après Richardson et al., 1995).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 13
2) Le monde acoustique des cétacés
Dans ce milieu sous-marin extrêmement acoustique, les cétacés dans leur ensemble ont
développé des caractéristiques uniques d’audition et de production des sons. Leur appareil auditif,
bien que gardant une organisation semblable à celle des autres mammifères, a énormément
évolué, surtout au niveau de l’oreille externe et moyenne, et n’a évidemment rien à voir avec celui
des autres vertébrés marins (en premier lieu les poissons). Leurs organes de vocalisation gardent
un point commun avec celui des mammifères terrestres, celui de fonctionner grâce aux voies
aériennes supérieures. Par ailleurs, aussi bien au niveau de l’audition que de la vocalisation, les
mysticètes et les odontocètes ont subi des évolutions distinctes9. Pourtant, il n’est pas exact
d’affirmer que les répertoires des mysticètes soient moins diversifiés que ceux des odontocètes,
comme on le lit parfois : une espèce comme le Mégaptère (Megaptera novaeangliae) produit des
sons beaucoup plus variés que le Cachalot, les Ziphiidés ou les marsouins.
Les huit espèces communes en Méditerranée nord-occidentale ne représentent pas un
échantillon complet de toutes les familles de cétacés marins : on ne trouve qu’un mysticète, le
Rorqual commun (Balaenoptera physalus), et il n’y a aucun phocoenidé, par exemple. Mais parmi
les odontocètes, on compte des représentants de familles essentielles (petits et grands
delphinidés, ziphiidés, physéteridés), ce qui fait que les sons de cétacés audibles dans le
Sanctuaire sont tout de même assez diversifiés et couvrent une bonne partie du spectre, entre
15 Hz et 100 kHz. Grâce aux enregistrements du GREC, je pourrai donc présenter une grande
partie du monde acoustique des cétacés dans un catalogue sonore10.
a) Audition des cétacés
Aspects anatomiques
Les trois parties fonctionnelles de l’appareil auditif des mammifères sont présentes chez les
cétacés : oreille externe, oreille moyenne et oreille interne. L’oreille externe des cétacés conserve
un orifice auditif, mais dont la fonctionnalité est limitée : chez les odontocètes, la réception des
sons à moyenne et haute fréquence s’effectue par la mandibule (Fig.2) et chez les mysticètes, le
circuit de l’oreille externe semble passer par l’orifice auriculaire et une structure souple spécifique
appelée le « doigt de gant ». La partie osseuse de la mandibule des odontocètes est constituée
dans sa partie postérieure d’un voile concave, à l’intérieur duquel se trouve une masse fibreuse et
graisseuse très bonne conductrice des sons qui joint l’oreille moyenne à la membrane tympanique.
9 La production des sons par les mysticètes n’est pas encore précisément élucidée.10 Pour les enregistrements de Rorqual commun, j’aurai recours à une source extérieure car leséquipements embarqués du GREC ne permettent pas d’enregistrer les sons à très très basse fréquence.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 14
Figure 2 : Réception du son chez les odontocètes (d’après Ketten in Au et al., 2000).
Celle-ci est très particulière car elle fait partie avec l’oreille interne du complexe de la bulle
tympano-périotique : entre le tympan et la fenêtre ovale de l’oreille interne, les osselets de l’oreille
moyenne sont très modifiés. La bulle tympanique osseuse a pour propriété de ne pas être liée
rigidement à la boîte cranienne, chez les odontocètes. La cavité de l’oreille moyenne contient de
l’air et un plexus artério-veineux. Une partie de la bulle tympanique renferme l’appareil cochléal de
l’oreille interne, qui n’est pas différent par essence de celui des autres mammifères. La cochlée en
forme de spirale renferme la membrane basilaire qui recueille les vibrations et les transforme en
impulsions micro-électriques grâce à des cellules ciliées, lesquelles sont présentes en quantité 6 à
17 fois supérieure chez les dauphins, par rapport aux humains. Les proportions de l’appareil
cochléal sont très variables et strictement liées au domaine de fréquence de chaque espèce de
cétacé, elles sont très adaptées aux hautes fréquences chez les odontocètes (Fig.3).
Figure 3 : Oreille moyenne et interne de dauphin (tiré de Thewissen in Perrin et al., 2002)

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 15
L’oreille interne est reliée à la partie auditive du cerveau (très volumineuse) par un faisceau
nerveux de diamètre très important, confirmant ainsi l’importance du sens auditif chez les cétacés.
Ainsi, malgré sa proximité fonctionnelle avec celle des mammifères terrestres, l’anatomie du
système auditif des cétacés a des traits spécifiques, plus accentués chez les odontocètes que
chez les baleines à fanons. Ces caractères lui confèrent une grande sensibilité, y compris sans
doute aux agressions sonores.
Audiogrammes
La sensibilité de l’oreille dépend de la fréquence ; l’audiogramme est la courbe définissant le
seuil d’audition (donc le plus faible niveau sonore audible) en fonction de la fréquence. Chaque
espèce a son audiogramme11, que l’on détermine par expérimentation. Parmi 85 espèces de
cétacés, seules 19 ont été sujettes à des expériences audiométriques, c’est peu mais cela
représente un grand progrès par rapport à 2002 (Fig.4). Mais on ne dispose pas à l’heure actuelle
d’audiogramme de mysticète ou de cachalot, par exemple.
Figure 4 : Audiogrammes d'odontocètes existant en 2002 (extrait de NRC, 2003).
En effet, les seuils auditifs sont habituellement déterminés par des expériences "psycho-
acoustiques" réalisées en milieu captif et pendant lesquelles on soumet des cétacés à des tests
auditifs, souvent avec des sons purs. Ces tests sont analogues à ceux auxquels sont soumis les
humains quand on établit leur audiogramme. Les individus doivent au préalable avoir été dressés,
afin de leur apprendre à donner la bonne réponse à la question posée: "entends-tu le son que l'on
11 Et même chaque individu a son audiogramme, qui de plus varie avec l’âge.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 16
vient de t'envoyer ?". Cela prend des mois. Il faut ensuite plusieurs semaines pour établir
l'audiogramme d'un cétacé avec cette méthode. Pour cette raison, jusqu'à présent, seuls des
odontocètes de taille moyenne ou petite pouvant être maintenus en captivité ont été étudiés sur le
plan de l'audiométrie.
Une méthode d'audiométrie électro-physiologique "AEP" a été mise au point pour pallier la
quasi-impossibilité expérimentale de mesurer l'audition des cétacés qui ne peuvent être maintenus
en captivité. Elle consiste à mesurer la sensibilité auditive grâce à des électrodes implantées sur la
peau au-dessus de la tête pour capter les micro-potentiels électriques qui émanent du cortex
auditif: c'est l’ABR, "auditory brain stem" (Nachtigall et al., 2007). Les niveaux sonores sont alors
projetés soit par des transducteurs disposés à faible distance du cétacé, soit par des transducteurs
appliqués contre le mandibule inférieure des cétacés (cas du "jawphone"). L'avantage de cette
méthode est sa rapidité (pas besoin de dressage) et son aptitude à être mise en oeuvre à
l'extérieur, comme par exemple dans un bassin temporaire sur un cétacé malade et en cours de
réhabilitation (Nachtigall et al., 2005), ou sur le rivage après l'échouage d'un cétacé vivant (Cook et
al., 2006), ou bien encore sur un bateau après la capture temporaire d'un dauphin (Nachtigall et
al., 2008).
Un des grands avantages de la méthode AEP est qu'elle permet l'obtention rapide
d'audiogrammes, ce qui permet d'étudier des groupes entiers (Houser et al., 2008), ou de mesurer
les pertes d'audition (TTS) consécutives à l’exposition préalable à des niveaux sonores élevés.
Toutefois, les résultats des tests par AEP ne concordent pas exactement avec les expériences
psycho-acoustiques, on obtient fréquemment des seuils d’audition un peu plus forts (de l’ordre de
5 à 10 dB). D'autre part, les seuils auditifs obtenus sur des individus échoués, et parfois mourants,
ne sont pas forcément représentatifs de ceux d'individus en bonne santé. Malgré ces
imperfections, la méthode AEP a permis de connaître grossièrement le domaine de sensibilité
auditive pour des odontocètes comme le cachalot (Ridgway & Carder, 2001), ou des mésoplodons
(Cook et al., 2006; Finneran et al., 2009).
Les courbes d'audiométrie établies jusqu'à présent sur les delphinidés, marsouins et
bélouga, ont plusieurs points communs (Fig.4) :
- elles montrent la forme classique en cuvette de celles des autres mammifères,
- la meilleure audition pour les cétacés testés est obtenue entre 10 et 80 kHz,
- à basse fréquence, la diminution de la sensibilité suit une pente approximative de 12 dB/octave12,
- pour ces odontocètes, la sensibilité optimale vaut entre 30 et 60 dB (re 1µPa) environ.
Il faut remarquer que des écarts de 10 dB ou plus peuvent exister pour des expériences
menés sur des individus différents d'une même espèce, écarts dus aux sujets eux-mêmes (Houser
& Finneran, 2006), à leur âge, ou aux conditions d'expérience, notamment le bruit de fond dans le
12 Multiplication de la fréquence par un facteur 2 ou 1/2

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 17
bassin, qui a un effet masquant. Les résultats de certaines expériences semblent non
représentatifs (Schlundt et al., 2011 sur le globicéphale tropical, par ex). La précision d’une courbe
audiométrique est donc de cet ordre. Parmi les huit espèces du Sanctuaire, la totalité des
delphinidés a subi des expériences d’audiométrie, liste à laquelle on peut ajouter le cachalot qui a
été étudié dans des conditions plus basiques (Tab.1). Pour les mysticètes, des travaux de
modélisation mécanique de l’oreille externe et moyenne (Tubelli et al., 2012) ont permis de
déterminer une bande audible pour le Rorqual de Minke (B.acutorostrata), d’environ 30 Hz à 7,5
kHz (modèle « doigt de gant »). La meilleure sensibilité est estimée grossièrement à 65 dB.
Espèce Bande passante @-10 dB etseuil optimal
méthode Référence
grand dauphin [8 à 100 kHz] ; 42 dB à 50 kHz comportementale Johnson, 1967
orque [10 à 30 kHz] ; 34 dB à 20 kHz comportementale Szymanski et al., 1999
pseudorque [15 à 70 kHz] ; 40 dB à 64 kHz comportementale Thomas et al., 1988
dauphin bleu et blanc [29 à 123 kHz] ; 42 dB à 64 kHz comportementale Kastelein et al., 2003
dauphin de Risso [20 à 100 kHz] ; 50 dB à 90 kHz électrophysiologique Nachtigall et al., 2005
dauphin commun [44 à 90 kHz] ; 53 dB à 64 kHz électrophysiologique Popov et al., 1998
globicéphale noir [23 à 45 kHz] ; 53 dB à 40 kHz électrophysiologique Pacini et al., 2010a
globicéphale tropical [20 à 70 kHz] ; 78 dB à 40 kHz électrophysiologique Schlundt et al., 2011
orque pygmée [25 à 50 kHz] ; 51 dB à 30 kHz électrophysiologique Pacini et al., 2010b
lagénorhynque Pacifique [16 à 100 kHz] ; 64 dB à 64 kHz comportementale Tremel et al., 1998
lagénorhynque à bec blc [29 à 123 kHz] ; 40 dB à 130 kHz électrophysiologique Nachtigall et al., 2008
mésoplodon de Blainville [35 à 60 kHz] ; 49 dB à 50 kHz comportementale Pacini et al., 2011
bélouga [20 à 100 kHz] ; 37 dB à 32 kHz comportementale in Au et al., 2000
sotalie fluviatile [64 à 100 kHz] ; 50 dB à 85 kHz comportementale Sauerland et al., 1998
sousa [30 à 50 kHz] ; 47 dB à 45 kHz électrophysiologique Li et al., 2012
marsouin commun [15 à 120 kHz] ; 37 dB à 40 kHz comportementale Kastelein et al., 2002
marsouin du Yang-Tse [40 à 110 kHz] ; 48 dB à 54 kHz électrophysiologique Popov et al., 2005
dauphin de l’Amazone [60 à 100 kHz] ; 50 dB à 80 kHz ? in Au et al., 2000
dauphin du Yang-Tsé [16 à 80 kHz] ; 59 dB à 32 kHz ? in Au et al., 2000
cachalot [5 à 40 kHz] ?; < 60 dB ? AEP sur nouveau-né Ridgway & Carder, 2001
rorqual de Minke [0.2 à 3 kHz] ?; 65 dB à 1 kHz ? modélisation Tubelli et al., 2012
Tableau 1 : Liste des audiogrammes de cétacés publiés (janv. 2014)

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 18
b) Production des sons par les cétacés
Tous les cétacés produisent des sons essentiellement grâce à des circulations d’air dans
leurs voies aériennes supérieures, et la plupart du temps par circulation interne. Un autre mode
d’émission acoustique pratiqué par les cétacés est le choc d’une partie du corps (ou de la totalité)
avec la surface : sauts et battements de caudale ou de pectorale assurent cette fonction.
Aspects anatomiques
Les parties anatomiques qui assurent les émissions sonores sont mieux connues pour les
odontocètes que pour les mysticètes ; dans les deux cas, des progrès scientifiques viennent
constamment mettre à jour cette problématique.
Pour ce qui est des baleines à fanons, il y a évidence que les vocalisations sont émises dans
une zone du bas larynx dans lequel on a observé des caractères anatomiques spécifiques, tout au
moins chez le Mégaptère (Mercado et al., 2010). Cependant, la variété des émissions acoustiques
des mysticètes est forte selon les espèces : extrêmement réduite chez les plus grands
balénoptères, elle est grande chez les balénidés et maximale chez le Mégaptère. Il y a donc des
chances pour que les spécificités anatomiques liées à la production des sons soient moindres
chez le Rorqual bleu (B. musculus) et commun.
Pour ce qui est des odontocètes, on n’observe pas d’unicité anatomique de l’appareil
vocalisateur ; il semble cependant qu’il y ait un cas extrême d’évolution chez le Cachalot, alors que
pour les autres familles, les caractères anatomiques dérivent d’un même schéma basé sur une
complexification des voies aériennes supérieures.
Chez le Cachalot, le conduit respiratoire se divise sous l’évent en un conduit gauche
assurant la fonction respiratoire, et un conduit droit assurant la fonction production des sons
(Fig.5). Le conduit droit comprend un sac « vestibulaire » producteur de clics, associé au museau
de singe, et un sac « naso-frontal » réflecteur des clics, situé sur la paroi crânienne.
Figure 5 : Anatomie de l’appareil émetteur sonore du Cachalot (d’après K.Das, 2003)

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 19
Le reste de la tête a pour fonction de conduire et focaliser les clics (spermaceti et « junk »13).
Cette singularité anatomique est associée à une spécialisation absolue du répertoire, puisque le
Cachalot ne produit que des impulsions de type clic, qui sont organisées de manières différentes
selon leur fonction élémentaire, soit écholocalisation, soit communication (Madsen et al. 2002).
Chez les delphinidés, les modifications des voies aériennes supérieures consistent en deux
paires de sacs aériens/museau de singe, une droite et une gauche, pas exactement identiques
(Fig.6). La circulation interne de l’air dans la paire droite ou la paire gauche assure la production
sonore. Ces deux ensembles permettent la production simultanée de deux types de sons, par
exemple un train de clics dans une paire de sacs et un sifflement dans l’autre. Les toutes dernières
informations suggèrent que la paire droite servirait à produire les clics, et la paire gauche les
sifflements (Madsen et al., 2013).
Figure 6 : Anatomie de l’appareil émetteur sonore chez le Grand dauphin (d’après K.Das, 2003)
Les appareils producteurs de sons n’ont pas été étudiés pour tous les odontocètes ;
cependant la littérature suggère que le système de « complexe de sacs aériens » a été observé
pour plusieurs delphinidés, et qu’il serait également valable pour les ziphiidés et les phocoenidés
(Huggenberger, 2009). Pour le Marsouin commun, Huggenberger (2008) observe que la région
hyoïde-larynx joue également un rôle pour la production des sons. Etant donné, que plusieurs
familles d’odontocètes présentent des répertoires sonores très spécifiques, il n’est pas exclu que
les adaptations anatomiques soient également substantiellement différentes. On voit donc que ce
domaine de la recherche est encore ouvert.
Domaines de fréquence utilisés
Les fréquences sonores émises par les cétacés obéissent en général aux grandes lois de
l’acoustique : plus les animaux sont gros, plus les sons émis sont à basse fréquence. Cependant,
ce principe s’applique de manière distincte aux sons purs ou composés (émissions tonales) et aux
13 Poche de tissu huileux et fibreux située sous le spermaceti et conduisant les sons

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 20
impulsions (Fig.7). A une extrémité du diagramme des fréquences, on a les appels du Rorqual
bleu, et à l’autre extrémité les clics du Marsouin ; lesquels sont plus hauts en fréquence que ceux
du Cachalot, d’une magnitude environ.
Figure 7 : Emissions sonores de cétacés (extrait de NRC, 2003).(Cuvier's B W = ziphius, Pilot W = globicéphale, Striped D = dauphin bleu et blanc, Sperm W = cachalot)
Pour ce qui est des delphinidés, qui ont souvent un répertoire composé au moins de
sifflements et de clics d’écholocalisation, on a remarqué que les sifflements comprenaient en
général une fondamentale entre 2 et 15 kHz (audible par les humains) et s'étendaient dans la
bande 20-35 kHz (May-Collado et al., 2007 ; 2009), bien que cela ne soit pas systématique
(Oswald et al., 2004). Il est admis (Supin et al., 1995, in Au & Hastings, 2008) qu’il y a une
corrélation nette entre la fréquence maximale des sifflements émis par les différentes espèces de
dauphins, et leur taille respective (c’est une relation inverse).
Espèce Fréquences(kHz)
Duréemoyenne (s)
référence (s)
Dauphin bleu et blanc 7,8 à 15,1 0,73 Gannier et al., 2009
Dauphin commun 8,5 à 13,1 0,47 Gannier et al., 2009
Grand dauphin 6,4 à 12,7 0,71 Gannier et al., 2009
Dauphin de Risso 8,2 à 14,5 0,65 Rendell et al., 1999 - Gannier et al., 2009
Globicéphale noir 3,2 à 6,4 0,89 Gannier et al., 2009
Orque 4,2 à 9,0 1,11 Riesch et al., 2005
Tableau 2 : Domaines de fréquence et durée des cris et sifflements de quelques espèces.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 21
Les clics d'écholocalisation des delphinidés ont une énergie spectrale maximale à ultra-haute
fréquence, comme pour l'orque pygmée (70-85 kHz ; Madsen et al., 2004a), le faux orque (30-
70 kHz ; Madsen et al., 2004b), le dauphin de Risso (50-90 kHz ; Phillips et al., 2003 ; Madsen et
al., 2004b). Pour ce dernier et l'orque, delphinidé de très grande taille, les spectres des clics sont
bimodaux, avec un premier pic entre 20 et 30 kHz, et un second entre 40 et 60 kHz (Au et al.,
2004). Comme indiqué plus haut, la fréquence des clics d’écholocalisation est en relation avec la
taille des proies de chaque espèce, par exemple, une fréquence de 30 kHz indique que la
dimension de la cible est de l’ordre de 5 cm14. Pour les ziphiidés, le tableau est similaire, avec des
fréquences d'énergie maximale de 42 kHz pour le ziphius (Zimmer et al., 2005) et une centroïde
de 38 kHz pour le mésoplodon de Blainville (Johnson et al., 2006) ou de 30 à 50 kHz pour le
mésoplodon de Gervais (Gillespie et al., 2009). Pour le cachalot, l'énergie maximale du clic se
situerait à 15 kHz (Madsen et al., 2002), donc en rapport avec des proies plus grosses.
Il apparaît donc clairement que l'énergie spectrale des clics se trouve à plus haute fréquence
pour les espèces les plus petites, ce qu'ont déjà observé Au et al. (2004), et quelle que soit la
famille d'odontocète (delphinidés, ziphiidés, physéteridés).
Espèce Fréquencecentroïde
kHz (*)
Fréquenced'énergie
max kHz (*)
Niveau desource pic-pic dB (**)
référence (s)
Dauphin de Risso 75 49 220 Madsen et al. 2004
Ziphius de Cuvier 42 40 214 Zimmer et al. 2005
Mésoplodon de Blainville 38 Johnson et al. 2006
Cachalot 15 15 220-236 Madsen et al. 2002
Faux orque 49 40 220 Madsen et al. 2004
Orque pygmée 70-85 40 197-223 Madsen et al. 2004
Orque 38 195 - 224 Au et al. 2004
Tableau 3 : Domaines de fréquence et niveau de source des clics d'écholocalisation de quelquesodontocètes.(*) uniquement quand le résultat provient d'un système d'enregistrement à bande large. (**) quand lesconditions expérimentales ont permis de déterminer l'orientation dans l'axe
En juxtaposant ces résultats sur les clics d'écholocalisation et ceux concernant l'audition, il
apparaît une correspondance entre le domaine de meilleure sensibilité auditive (Tab.3) et la bande
de fréquence d'énergie maximale des clics d'écholocalisation, quels que soient les genres
d'odontocète. On peut utiliser ce paradigme de la similarité des bandes fréquentielles d'émission et
de réception, pour tenter de définir le domaine de meilleure audition des espèces pour lesquelles
on ne dispose pas d’audiogramme, à condition de disposer d’une bonne description spectrale des
clics d'écholocalisation.
14 Ce qui ne veut pas dire que les poissons ou les calmars recherchés mesurent 5 cm, c’est simplement la
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 22
c) Sensibilité des cétacés aux nuisances sonores
Même si cela peut sembler évident, il faut observer que les cétacés sont sensibles aux
nuisances sonores dans la mesure où ils perçoivent les bruits en question avec une forte intensité.
En effet, mis à part les très fortes pressions sonores qui peuvent entraîner des lésions aux
organes même si elles ne sont pas perçues par le système auditif, le reste des insonifications
passe par le filtre de l’appareil auditif de chaque espèce avant de produire ses effets. Par
conséquent, les éléments dont nous disposons sur l’audition des espèces et leurs vocalisations
nous permettent de distinguer les bandes de fréquences auxquelles seront sensibles les cétacés
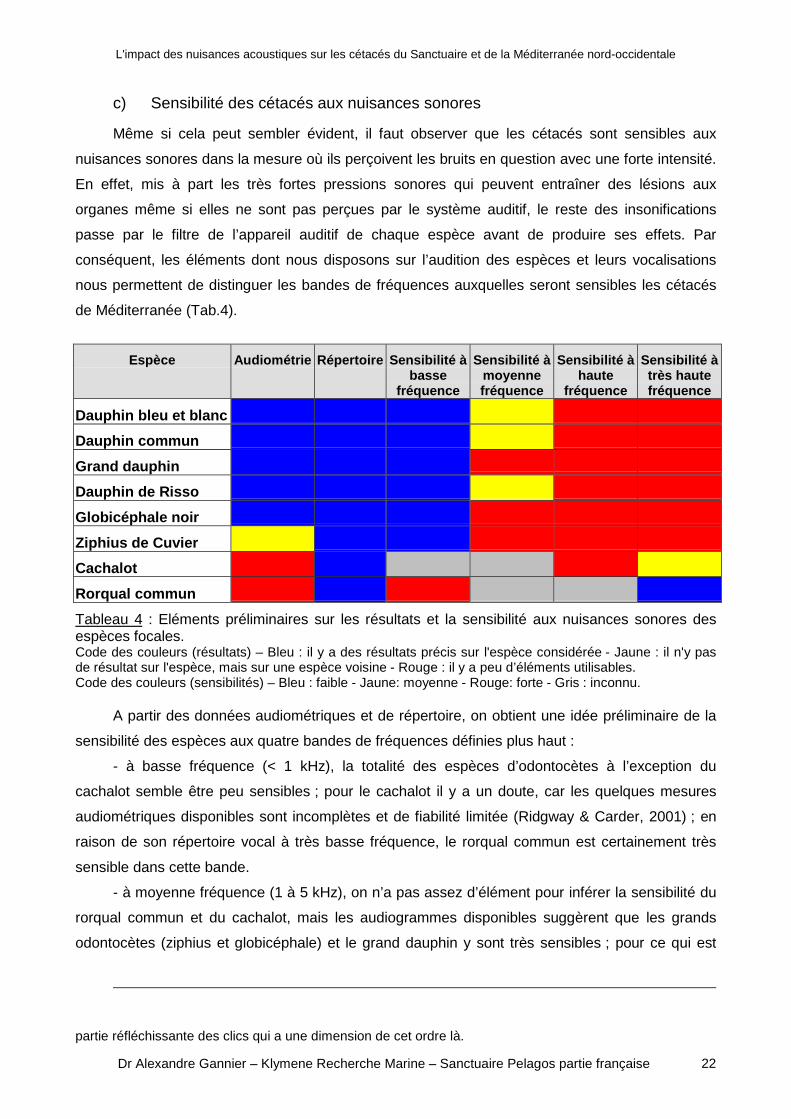
de Méditerranée (Tab.4).
Espèce Audiométrie Répertoire Sensibilité àbasse
fréquence
Sensibilité àmoyennefréquence
Sensibilité àhaute
fréquence
Sensibilité àtrès hautefréquence
Dauphin bleu et blanc
Dauphin commun
Grand dauphin
Dauphin de Risso
Globicéphale noir
Ziphius de Cuvier
Cachalot
Rorqual commun
Tableau 4 : Eléments préliminaires sur les résultats et la sensibilité aux nuisances sonores desespèces focales.Code des couleurs (résultats) – Bleu : il y a des résultats précis sur l'espèce considérée - Jaune : il n'y pasde résultat sur l'espèce, mais sur une espèce voisine - Rouge : il y a peu d’éléments utilisables.Code des couleurs (sensibilités) – Bleu : faible - Jaune: moyenne - Rouge: forte - Gris : inconnu.
A partir des données audiométriques et de répertoire, on obtient une idée préliminaire de la
sensibilité des espèces aux quatre bandes de fréquences définies plus haut :
- à basse fréquence (< 1 kHz), la totalité des espèces d’odontocètes à l’exception du
cachalot semble être peu sensibles ; pour le cachalot il y a un doute, car les quelques mesures
audiométriques disponibles sont incomplètes et de fiabilité limitée (Ridgway & Carder, 2001) ; en
raison de son répertoire vocal à très basse fréquence, le rorqual commun est certainement très
sensible dans cette bande.
- à moyenne fréquence (1 à 5 kHz), on n’a pas assez d’élément pour inférer la sensibilité du
rorqual commun et du cachalot, mais les audiogrammes disponibles suggèrent que les grands
odontocètes (ziphius et globicéphale) et le grand dauphin y sont très sensibles ; pour ce qui est
partie réfléchissante des clics qui a une dimension de cet ordre là.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 23
des petits delphinidés et du dauphin de Risso, les éléments audiométriques et de répertoire
suggèrent une sensibilité moyenne.
- à haute fréquence (5 à 20 kHz), les éléments audiométriques et de répertoire suggèrent
que tous les odontocètes de Méditerranée pourraient être très sensibles ; pour le rorqual commun,
l’absence totale d’éléments audiométriques ne permet pas de se prononcer.
- à très haute fréquence (> 20 kHz), même si on n’a pas d’éléments audiométriques, on peut
avancer que le rorqual commun est peu sensible, car il n’y a pas d’exemple d’audiogramme de
cétacé dont la bande passante soit aussi étendue des très basses fréquences aux très hautes
fréquences ; pour tous les odontocètes, même le cachalot, il est à ce stade vraisemblable que la
sensibilité aux très hautes fréquences soit élevée.
Un autre aspect important dans l’évaluation de la conséquence des insonifications est la
prise en compte de la durée d’exposition. Dans les phénomènes de fatigue auditive en particulier,
les conséquences d’une onde de niveau sonore et de fréquence donnée dépendent de la durée
d’exposition. Aussi bien dans le cas d’un son non-impulsif (type sonar) que dans le cas
d’impulsions (prospection sismique, battage de pieu), le risque de pertes de sensibilité auditive doit
tenir compte du temps d’insonification, par exemple le nombre de secondes du signal de sonar, ou
le nombre d’impulsions du canon pneumatique. Pour résoudre ce problème, on est amené à
exprimer certains critères en « dose d’exposition sonore » (SEL pour Sound Exposure Level), une
quantité qui intègre l’intensité sonore sur le temps d’exposition.
Cette première impression basée sur des données acoustiques objectives va devoir être
confirmée par des résultats expérimentaux portant sur les effets réellement observés sur les huit
espèces, lors d’études opportunistes (pendant l’exposition à des fortes intensités sonores) ou
dédiées (exposition contrôlée à des intensités sonores connues). Pour les basses fréquences, on
doit s’attendre à une forte sensibilité du rorqual, et peut-être du cachalot. Pour les hautes
fréquences, tous les odontocètes semblent potentiellement sensibles aux nuisances. Pour les
moyennes fréquences, le tableau semble plus contrasté, avec aussi davantage d’incertitude.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 24
3) Répertoires des huit espèces de cétacés dans le Sanctuaire
La presque totalité des enregistrements acoustiques présentés ici a été obtenue lors des
prospections du GREC depuis 1990. En particulier, depuis 1994 un hydrophone est remorqué en
permanence à l’arrière du voilier utilisé pour les prospections, ce qui a permis d’enregistrer les sept
espèces d’odontocètes de Méditerranée nord-occidentale à de multiples occasions. Par contre, le
matériel utilisé ne permet pas d’enregistrer le Rorqual commun : en effet, ce mysticète est
caractérisé par ses émissions à très basse fréquence (< 30 Hz) et cette bande n’est pas
accessible à un système remorqué, puisque en dessous de 100 Hz, le bruit lié à l’écoulement de
l’eau autour de l’antenne de l’hydrophone est très important. Pour les enregistrements de
balénoptère, nous aurons donc recours à des sources extérieures.
L’hydrophone en lui-même n’a pratiquement pas varié au cours de la période 1990-2010,
ainsi que le pré-amplificateur qui se trouve dans l’antenne, qui filtre par construction les sons de
fréquence inférieure à 200 Hz. Par contre les moyens d’enregistrement ont fortement évolué, avec
les techniques de numérisation : dans une première période, c’est un enregistreur analogique de
type « walkman » (Sony TCD-30 ou WMD-6, bande passante 0,2-16 kHz), puis à partir de 1995,
un enregistreur digital à bande a été utilisé (types TCD-7 et TCD-8, bande passante 0,2-22 kHz),
puis à partir de 2005, un enregistreur numérique de type Marantz (bande passante 0,2-24 kHz).
Enfin, depuis 2010, l’enregistrement des sons se fait directement sur le système informatique de
monitoring, avec une bande passante réglable, mais en général fixée à 0,2-48 kHz.
La plupart du temps, à bord, un filtre analogique est interposé entre l’hydrophone et
l’enregistreur, et ce filtre a souvent été réglé pour atténuer les sons de fréquence inférieure à
1 kHz.
Ma présentation du répertoire est organisée par espèce, en commençant par le Dauphin
bleu et blanc et en terminant par le Rorqual commun. Pour chaque espèce, on trouvera un court
commentaire de présentation du répertoire, suivi par un tableau de référence dans lequel on aura
une liste de noms de fichiers, avec les méta-données de l’enregistrement, et un commentaire sur
chaque enregistrement. Le nom du fichier (ex : Sc108075_4) est relié par un lien hypertexte au
fichier *.wav et permet donc l’écoute du son considéré15.
15 A condition que les fichiers sonores soient placés dans le répertoire convenable.
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 25
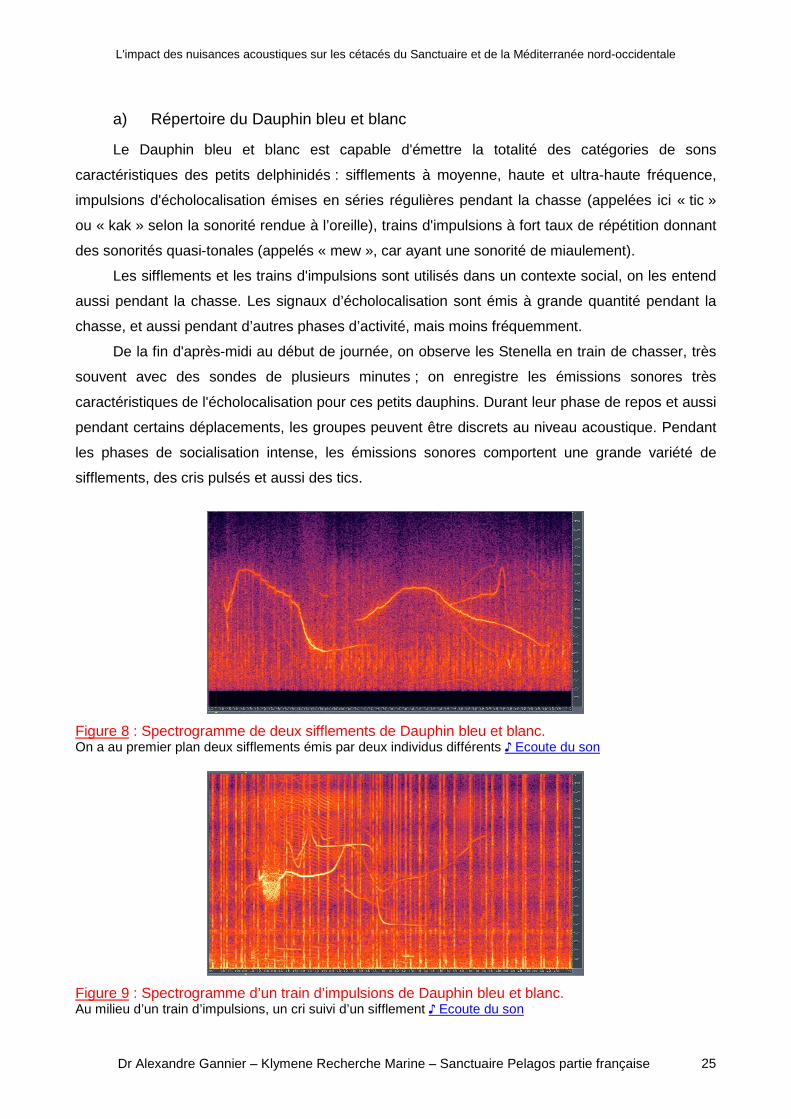
a) Répertoire du Dauphin bleu et blanc
Le Dauphin bleu et blanc est capable d'émettre la totalité des catégories de sons
caractéristiques des petits delphinidés : sifflements à moyenne, haute et ultra-haute fréquence,
impulsions d'écholocalisation émises en séries régulières pendant la chasse (appelées ici « tic »
ou « kak » selon la sonorité rendue à l’oreille), trains d'impulsions à fort taux de répétition donnant
des sonorités quasi-tonales (appelés « mew », car ayant une sonorité de miaulement).
Les sifflements et les trains d'impulsions sont utilisés dans un contexte social, on les entend
aussi pendant la chasse. Les signaux d’écholocalisation sont émis à grande quantité pendant la
chasse, et aussi pendant d’autres phases d’activité, mais moins fréquemment.
De la fin d'après-midi au début de journée, on observe les Stenella en train de chasser, très
souvent avec des sondes de plusieurs minutes ; on enregistre les émissions sonores très
caractéristiques de l'écholocalisation pour ces petits dauphins. Durant leur phase de repos et aussi
pendant certains déplacements, les groupes peuvent être discrets au niveau acoustique. Pendant
les phases de socialisation intense, les émissions sonores comportent une grande variété de
sifflements, des cris pulsés et aussi des tics.
Figure 8 : Spectrogramme de deux sifflements de Dauphin bleu et blanc.On a au premier plan deux sifflements émis par deux individus différents ♪ Ecoute du son
Figure 9 : Spectrogramme d’un train d’impulsions de Dauphin bleu et blanc.Au milieu d’un train d’impulsions, un cri suivi d’un sifflement ♪ Ecoute du son

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 26
Tous les enregistrements ont été réalisés avec l'hydrophone remorqué HP30-ST,
comprenant deux capteurs Benthos AQ-4 d'une bande passante supérieure à 30 kHz (Tab.5).
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat-Lon)
Origine
Sc99018_03-sifs Sifflements complexes dans ungroupe de 20 (Pm en fond)
6,0 25/06/9913h50
41N1005E51
Données GREC
Sc99080_03-kaks-sifs Kaks et sifflements d’un petitgrou^pe en chasse
8,0 10/07/9920h52
35N3503W57
Données GREC
Sc99080_04-kak « Forêt » de kak, typique de lachasse pélagique.
7,9 idem idem Données GREC
Sc99096_01-sif-coin Sifflements et bourdon dans ungroupe en prédation
11,4 14/07/9900h50
37N2000W25
Données GREC
Sc101006_01-sif-HF Sifflement haute fréquence avecharmonique.
6,5 04/07/0119h32
42N4506E34
Données GREC
Sc101007_01-sif-HF Sifflement très modulés 2,6 05/07/0106h02
42N0105E57
Données GREC
Sc101016_15-sif-mew Clics directs et nombreux sifflenchevêtrés. Miaulements
6,5 11/07/0117h09
40N0304E21
Données GREC
Sc2001-C36_1-kak Forêt de kaks, et miaulement decapture. Prédation de nuit
11,0 14/08/0120h50
43N0006E55
Données GREC
Sc102029_04-siffs Sifflements forts de plusieursindividus dans un grand groupe
7,5 09/07/0208h59
40N1504E17
Données GREC
Sc102029_07-sif-pulses Cris pulsés et sifflements,miaulements.
10,8 idem idem Données GREC
Sc102034_04-sifs-couin Sifflements très variés et autressons sociaux, grand groupe
14,4 10/07/0211h56
40N0904E14
Données GREC
Sc102034_17-sifs-couin Cris pulsés, sifflements et clicsdans un groupe en socialisation
6,9 idem idem Données GREC
Sc2003-C82_6-kak-mew Prédation d’un seul individu,chronométrage possible
8,3 04/08/0320h35
43N0106E49
Données GREC
Sc2004-18.07_3h-kak-mew Prédation nocturne dans ungroupe pélagique
16,7 18/07/0403h28
42N1004E35
Données GREC
Sc2006_19.11-13h-sif-bruit Sifflements en milieu côtierbruyant
6,3 19/11/0613h53
43N3507E15
Données GREC
Sc2007_07.07-23h-kak-bruit Prédation nocturne au large enambiance bruyante
7,6 07/07/0723h18
43N1008E26
Données GREC
Sc2008_5.08-6h-kak Prédation à l’aube au large enambiance normale
12 05/08/0806h01
39N0210E49
Données GREC
Tableau 5 : Enregistrements de dauphins bleus et blancs (activables par lien hypertexte).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 27
b) Répertoire du Dauphin commun
Comme le Dauphin bleu et blanc, le Dauphin commun est capable d'émettre toutes les
catégories caractéristiques de sons des petits delphinidés : sifflements de moyenne, à ultra-haute
fréquence, impulsions d'écholocalisation émises en séries régulières pendant la chasse. Il y a
aussi des sons plus exotiques, comme des aboiements, liés à des contextes sociaux agités.
Dans les groupes importants, l’activité sociale est souvent forte en journée, ce qui se traduit
par des groupes très bruyants : sifflements nombreux et d’intensité élevée. Il arrive aussi de
rencontrer des groupes silencieux. Durant les phases de chasse méso-pélagique, on entend le
même type de caquètement que celui des Stenella. Il est difficile de distinguer les deux espèces.
Figure 10 : Spectrogramme de vocalisations de Dauphin commun.On est près d’un groupe où il y a beaucoup de sifflements enchevêtrés avec des impulsions ♪ Ecoute du son
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat-Lon)
Origine
Dd94101_06-reponse Sons et série de couinements 6,0 07/08/9413h25
41N3209E32
Données GREC
Dd94101_05-sifs Sifflements HF 8,2 idem idem Données GREC
Dd94101_06-sifs Sifflements HF 2,2 idem idem Données GREC
Dd97080_05-kak-mew Kaks de chasse, mew capture 7,7 28/07/9718h20
38E3508E47
Données GREC
Dd97080_03-sifs Deux sifflements simples HF 5,8 idem idem Données GREC
Dd99045_01-sif Une série de sifflements HF 4 03/07/9909h52
36N5402W39
Données GREC
Dd99045_01-sifs-simples Deux sifflements simples 3 idem idem Données GREC
Dd107097_17h00-tics-kaks Une rafale de tics puis deskaks du groupe en chasse
13,5 22/07/0716h52
42N1010E01
Données GREC
Dd107097_17h00-tics Des clics directs forts 5,8 idem idem Données GREC
Dd107097_17h00-sifs Sifflements mélangés avecdes clics, groupe nombreux
6,7 idem idem Données GREC
Tableau 6 : Enregistrements de dauphins communs (activables par lien hypertexte).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 28
c) Répertoire du Grand dauphin
La variété des sons émis par le Grand dauphin est très grande, notamment pour les
sifflements et les trains d’impulsions à contexte social, qui rendent des sonorités très diverses. En
ce qui concerne les sifflements, on en distingue deux catégories fonctionnelles : les sifflements
« signature » propres à chaque individu, et les sifflements destinés aux autres communications.
Les sifflements peuvent être aussi bien des sons simples (sans harmonique), que des sons
composés (avec harmoniques).
Le timbre des sifflements est sensiblement moins aigu (Tab.1) et comme typiquement les
groupes de Grand dauphin sont moins importants que ceux des petits delphinidés, on a souvent
une activité acoustique moins continue, ou moins confuse. Egalement, il est fréquent de rencontrer
des groupes de Tursiops peu vocaux, voire silencieux, dans des activités de repos, de voyage, ou
même en chasse.
Figure 11 : Spectrogramme d’un sifflement de Grand Dauphin.Un sifflement avec harmoniques précède une série de clics proches ♪ Ecoute du son
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat-Lon)
Origine
Tt99082_15-sif-et-tics Deux sifflements et rafales 3,1 11/07/9911h51
35N592W52
Données GREC
Tt99082_15-sif-bizarre Sifflements bizarre et normal 5 idem idem Données GREC
Tt99082_15-sifs-rafales Sifflements forts et rafales 4,9 idem idem Données GREC
Tt102033_05-sifs Des sifflements répétés 11,6 09/07/0218h20
40N084E14
Données GREC
Tt106069-sifs-tics Sifflements de plusieurs et tics 7,7 30/06/0617h51
40N114E04
Données GREC
Tt106069-sifs Sifflements de 2 ou 3individus, quelques tics
10,8 idem idem Données GREC
Tt106074-sifs-couine Sifflements de plusieursindividus, couinements
11 02/07/0609h54
40N0804E06
Données GREC
Tt106074-sifs Sifflements haute fréquence 4,7 idem idem Données GREC
Tableau 7 : Enregistrements de grands dauphins (activables par lien hypertexte).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 29
d) Répertoire du Dauphin de Risso
Le Dauphin de Risso peut émettre les différentes catégories de sons des petits delphinidés :
sifflements à moyenne, et haute fréquence, impulsions d'écholocalisation émises en séries
régulières pendant la chasse et la socialisation. D’après notre expérience, le Grampus est un peu
plus discret que d’autres en ce qui concerne les sifflements. Par contre, l’espèce peut souvent être
repérée grâce aux bourdonnements fréquents qu’elle émet lors de phases de socialisation, entre
autres, qui sont des trains d’impulsions, avec aussi des miaulements (taux de répétition rapide).
Le Grampus est aussi identifiable à distance grâce à ses clics d'écholocalisation, plus
espacés que ceux de petits delphinidés, qui peuvent sembler similaires à ceux du globicéphale.
Mais on n'entend pas de cris pulsés proprement dits, tels que ceux émis par le globicéphale.
En milieu de journée et en été, les Dauphins de Risso peuvent être inactifs ou se déplacent
assez silencieusement. En fin d'après-midi, on les observe en train de chasser. Leurs émissions
sonores sont alors assez distinctes car elles comportent une grande proportion de trains
d'impulsions très serrées (Fig.12). En général, il peut arriver que les groupes soient discrets au
niveau acoustique.
Figure 12 : Spectrogramme de bourdonnement de Dauphin de Risso.On a un train d’impulsion proche avec des bourdonnements ♪ Ecoute du son
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat-Lon)
Origine
Gg99090_03-tics-sif Clics et sifflements. 7,3 13/07/9911h46
37N1101W33
Données GREC
Gg99113_03-mew-tics Sifflements, bourdon, clicsdirects et indirects.
8,3 21/07/9915h39
40N1204E10
Données GREC
Gg99113_11-tics-sif-mew Clics, bourdons variés,couac, siffl.
10 idem idem Données GREC
Gg102042_11-tics-mew Bourdon et miaulementsavec clics de cach en fond.
6 12/07/0217h10
40N1404E08
Données GREC
Gg107069_10h49-clic-sif Tics dans un groupe enphase de chasse.
10,3 15/07/0710h43
41N3710E36
Données GREC
Tableau 8 : Enregistrements de Dauphins de Risso (activables par lien hypertexte).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 30
e) Répertoire du Globicéphale noir
Le répertoire du Globicéphale noir comprend les sifflements, les cris pulsés et les clics
d’écholocalisation. Les sifflements de globicéphale se déclinent avec de nombreuses variantes,
mais ont généralement un domaine de fréquence entre 1 et 8 kHz. Le globicéphale se distingue
des autres delphinidés communs par les cris pulsés, qui sont assez proches de ceux de l'orque
par leur structure et leur sonorité. La fonction des cris pulsés est avant tout la communication, y
compris la reconnaissance de l'appartenance au groupe et celle de l'individu.
En Méditerranée, l'activité diurne est souvent le repos ou la socialisation, la nuit étant
consacrée à la chasse. Un groupe est souvent silencieux pendant le repos, mais très audible
lorsque les cris pulsés sont émis en socialisation. Les clics d’écholocalisation émis pendant la
prédation sont souvent audibles à plusieurs kilomètres. Les clics d'écholocalisation des
globicéphales sont généralement facilement distincts parmi les grands delphinidés, par leur timbre,
qui signale une fréquence plus basse (différence audible), et leur rythme assez lent.
Figure 13 : Spectrogramme de cri complexe pulsé de Globicéphale noir.Au sein d’un groupe, on entend des cris complexes ♪ Ecoute du son
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 31
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat-Lon)
Origine
Gm97026_19-cris Des cris complexes d’ungroupe au repos
12,7 08/07/9711h06
42N5105E25
Données GREC
Gm97044_17-clics Un train de clics 6,5 10/07/9708h39
42N5305E45
Données GREC
Gm97044_18-cris-clics Deux cris entrecoupés parun train de clics
6,3 idem idem Données GREC
Gm98118_16-2cris Deux cris avec un clic decachalot en fond
5,5 16/08/9809h03
43N2207E09
Données GREC
Gm98118_16-cris Sifflements, rafales declics, couinements
8,3 idem idem Données GREC
Gm98118_16-clics-cris Sifflements, rafales declics, couinements
6,8 idem idem Données GREC
Gm99038_07-cris Cris et quelques clics d’ungroupe en chasse
25,4 01/07/9902h03
37N1900W21
Données GREC
Gm99038_07-clics-cris Clics en rafale et quelquescris simples
14 idem idem Données GREC
Gm99047_06-cris Cris variés dans un groupesemblant en voyage
22 03/07/9910h07
36N2502W40
Données GREC
Gm99047_05-cris-mew Bourdon, couic, et miaule-ment dans un groupe
11,8 idem idem Données GREC
Gm103132_07-cris Cris enchevêtrés dans ungroupe en repos social
19,4 12/08/0315h42
43N2207E30
Données GREC
Gm107116_18h00-cris Cris répétés forts, avec unbruit de navire filtré
31,6 25/080716h33
42N4906E16
Données GREC
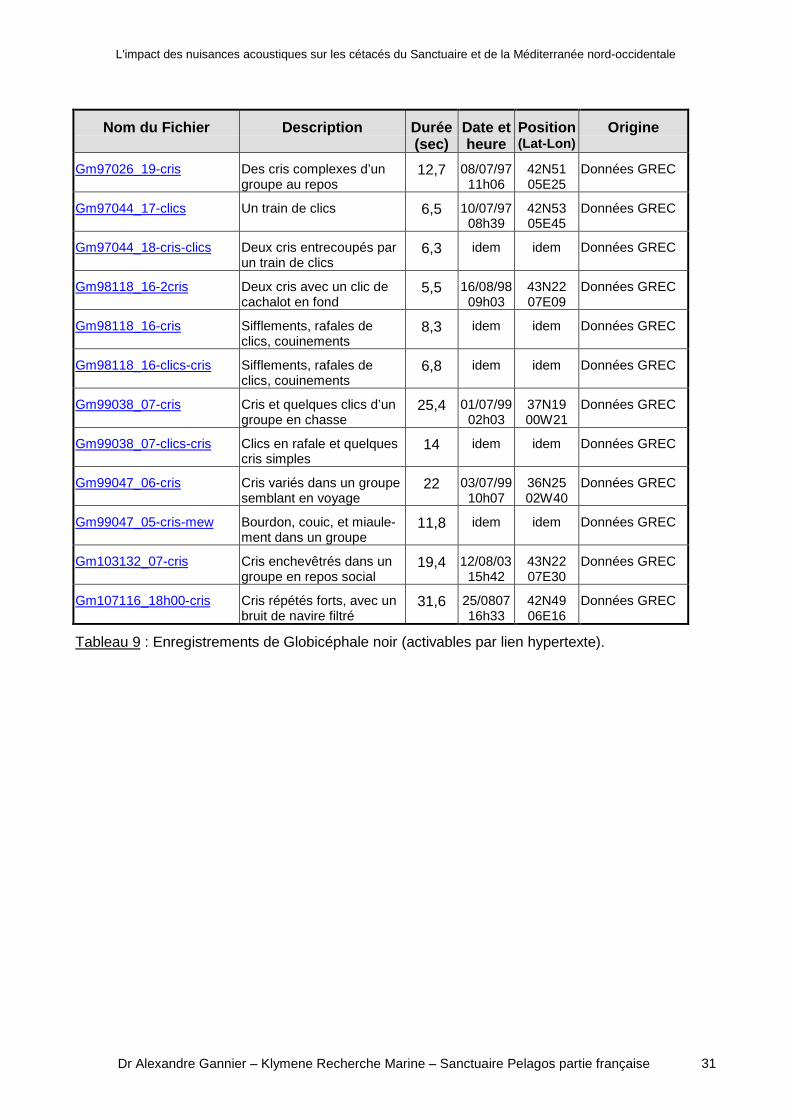
Tableau 9 : Enregistrements de Globicéphale noir (activables par lien hypertexte).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 32
f) Répertoire du Ziphius de Cuvier
Le répertoire du Ziphius est beaucoup moins varié que ceux des delphinidés : il est composé
essentiellement de clics et de bourdonnements (impulsions serrées) émis dans les très hautes
fréquences, avec une énergie maximale entre 30 et 40 kHz. L’énergie présente entre 10 et 20 kHz
fait que ces clics sont audibles par l’homme à faible distance. La caractéristique de ces clics est
d’être à bande étroite et modulation de fréquence, donc une structure différente de celle des
dauphins ou du Cachalot. Le Ziphius émet aussi des sifflements, ceux-ci étant peu puissants et
peu modulés16.
Les conditions de nos enregistrements de Ziphius sont particulières : le bateau est tenu
quasi-immobile, l’hydrophone se positionne quasiment verticalement avec l’antenne située à
plusieurs dizaines de mètres de profondeur. Le filtrage analogique à bord est réglé sur 3 à 5 kHz,
et le gain analogique est augmenté. Ces conditions permettent des enregistrements jusqu’à une
distance estimée d’au moins 500 m, lorsque les Ziphius sont en sonde profonde.
Les Ziphius sont actifs en prédation à n’importe quelle heure de la journée ou de la nuit ,
alternant sondes de chasse et sondes de récupération. On les observe parfois en repos, ou bien
en socialisation.
Figure 14 : Spectrogramme de clics de Ziphius.Une série de clics d’un individu (avec des échos de la surface) ♪ Ecoute du son
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat-Lon)
Origine
Zc110031_16h19_tic Clics direct et indirectsprès de 4 indiv. en sonde
7,8 09/08/1017h47
41N4210E16
Données GREC
Zc110031_17h45_tic-sif Clics indirects avec un siffl. 10,4 idem idem Données GREC
Zc111069_18h00_tic Clics de niveau faible d’unseul individu à 800 m
10 18/08/1117h07
44N008E28
Données GREC
Tableau 10 : Enregistrements de Ziphius en Méditerranée.
16 Certains scientifiques semblent contester ce point.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 33
g) Répertoire du Cachalot
Le Cachalot n'émet que des vocalisations de type "impulsions". La bande spectrale de ces
impulsions est assez large, environ de 2 à 40 kHz, tandis que les autres caractéristiques, comme
le niveau de source, le taux de répétition, la directivité ou l'organisation dans le temps, sont
variables selon leur fonction. Celle-ci est variable, soit strictement la prédation (clic, creak), soit
strictement la communication (coda, clang), soit encore une combinaison comme les codas émis
par les cachalots dispersés durant la prédation. D'autres catégories de sons, comme les
« chirrups » (crépitements), les « squeals » (crissement), ne sont pas encore associés
précisément à un contexte d'activité. Par contre, les sons de trompette émis juste après le début
de la sonde ont été associés à la purge des conduits aériens avant le début d'émission des clics.
Les clics normaux dits "réguliers" sont utilisés pour l'écholocalisation et sont émis pendant la
majeure partie de la sonde, à partir du moment où le cachalot a débuté sa descente, puis à un
rythme presque régulier quand l'animal est en phase profonde, et jusqu'au moment où il débute sa
remontée. Durant la phase de chasse, les séries de clics réguliers sont interrompues par des
creaks correspondant à des phase de poursuite-capture des proies, on en compte souvent entre
20 et 30 par sonde, en Méditerranée. Ce nombre varie selon les bassins océaniques (Watwood et
al., 2006). Les codas sont des trains d'impulsions dont la structure temporelle (taux de répétition)
est fortement organisée et stéréotypée, la durée de la séquence étant souvent inférieure à 1
seconde ; leur fonction est sociale, ils peuvent être émis en plongée ou en surface. Les clics lents
ou « clang » ne sont entendus qu'en présence de mâles adultes, dans des situations variables. Ils
semblent avoir un rôle dans la communication à longue distance. Le chirrup ou crépitement est un
train d'impulsions rapides dont la structure temporelle (taux de répétition) ne paraît pas
stéréotypée. Le squeal ou miaulement ressemble à un cri, de nature impulsionnelle à taux très
rapide. Il est souvent émis en présence de nourrissons ou juvéniles, ce qui suggère qu’il serait
produit par ces derniers. Le Cachalot chasse potentiellement jour et nuit. Des groupes peuvent se
réunir pour se reposer (ils sont alors souvent silencieux) ou socialiser.
Figure 15 : Spectrogramme de clic de Cachalot.Une série de clics de chasse (avec des échos de surface, bruit de cargo) ♪ Ecoute du son

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 34
Figure 16 : Spectrogramme de creak de Cachalot.Un creak de capture, avec amplitude et intervalle inter-clic décroissants ♪ Ecoute du son
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat-Lon)
Origine
Pm99_clang Un individu émet 2 clangs 8,1 1999 Bassinouest
Données GREC
Pm100_coda8 Un individu émet 1 coda à8 impulsions
1,2 2000 Bassinest
Données GREC
Pm101021_coda Un indiv. émet des codas(d’abord lent puis rapide)
10 13/07/0112h54
39N574E29
Données GREC
Pm103090_chir Émission de chirrups parun individu
9,2 03/08/0321h32
42N586E51
Données GREC
Pm103103_tromp Trompette par un individuqui vient de sonder
6 06/08/0311h23
42N476E09
Données GREC
Pm104007_04clic Clic et silence de chasse(bruit de navire)
23 04/07/0412h43
43N237E10
Données GREC
Pm104020_03clic 2 à 3 individus cliquent enchasse
44 18/07/0406h50
42N134E33
Données GREC
Pm104020_05clic-silence Un individu clique avecdeux phases de silence
39 idem idem Données GREC
Pm104036_03clic Un individu clique avecvariation de rythme
34 21/07/0412h37
42N576E28
Données GREC
Pm104067_02clic-creak Un individu clique puisémet un creak de capture
27 02/08/04 42N596E44
Données GREC
Pm105010-3_1clic-coda Un individu clique et unautre émet des codas
30 04/07/0512h01
43N2407E09
Données GREC
Pm105010-3_2clic Un individu clique, etindividu clique plus proche
21 idem idem Données GREC
Pm105039-5_1m Au moins 4 individus detaille différente en chasse
23,5 13/07/0511h49
42N535E43
Données GREC
Pm105092_codaclic Un individu émet 4 codaspuis des clics de descente
41,4 27/07/0518h14
43N0405E29
Données GREC
Tableau 11 : Enregistrements de Cachalot (activables par lien hypertexte).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 35
h) Répertoire du Rorqual commun
Les vocalisations du Rorqual commun sont parmi les plus simples chez les mysticètes : elles
se composent d’appels dans la bande des 15-20 Hz, d’une durée d’environ 1 seconde et à bande
plutôt étroite, et qui sont généralement produits en longues séquences avec un rythme spécifique.
Il est généralement admis que ces appels sont émis par les mâles. La période des appels
(intervalle inter notes) est autour de 7 à 26 secondes (14-15 s en nord Méditerranée), avec des
séquences stéréotypées durant typiquement 3 à 20 min et entrecoupées de silences de plusieurs
minutes. La période peut varier avec la saison. Ces vocalisations s’apparentent donc à un
« chant » mais dont les notes ne seraient pas variées (deux types de notes). En Atlantique et dans
le sud-ouest de la Méditerranée, des sons modulés dans la bande 135-140 Hz peuvent également
être entendus, mais plus rarement.
Les sons de Rorqual commun que nous présentons ici ont été aimablement mis à disposition
par le Dr Manuel Castellote, que nous remercions particulièrement.
Figure 17 : Spectrogramme d’un appel de Rorqual commun.Un train d’appels très basse fréquence qui se répète ♪ Ecoute du son (fréquence x 10)
Nom du Fichier Description Durée(sec)
Date etheure
Position(Lat/Long)
Origine
Bp classic backbeat ext Pelagos Appel classiqueméditerranéen
4,4 Manuel Castellote(fréquence x 10)
Bp downsweep Appel modulédescendant
4,5 Manuel Castellote
Bp 134-140 Hz upsweeps Appel moduléascendant
9,8 Manuel Castellote
Tableau 12 : Enregistrements de Rorqual commun (activables par lien hypertexte).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 36
B\ Synthèse bibliographique
Dans le milieu marin, le bruit ambiant naturel (tel qu’il était avant l’invention de la machine à
vapeur) comportait des événements naturels très bruyants tels que glissements de terrain,
tremblements de terre, éruptions volcaniques sous-marines. Cependant, ceux-ci étaient rares, de
portée locale, et avaient un caractère basse ou très basse fréquence. Ainsi, leur influence sur la
plupart des espèces de cétacés était-elle limitée. Seules les fortes précipitations avaient, et ont
toujours, la faculté d’impacter le milieu par un bruit ambiant significatif à moyenne fréquence
(Fig.1). Mais, hormis en zone équatoriale, elles n’ont pas le caractère de fréquence qui pourraient
les rendre gênantes pour le sens auditif des cétacés, et de toutes façons elles n’atteignent pas un
niveau sonore dangereux pour eux. Avec l’amplification de l’industrialisation, des bruits nocifs de
toutes sortes ont envahi de vastes régions des océans, qui offrent par ailleurs des habitats très
favorables à de nombreuses espèces de cétacés. Les eaux du Sanctuaire Pelagos ne sont
exemptes d’aucune de ces menaces acoustiques17 ; nous allons donc passer en revue les plus
fortes des nuisances sonores que l’on peut rencontrer en Méditerranée occidentale.
1) Principales sources anthropiques pouvant impacte r les cétacés
Les cétacés sont des animaux très mobiles : la plupart des espèces peuvent se déplacer à
des vitesses supérieures à 5 noeuds pendant de nombreuses heures sans risquer l’épuisement.
Par conséquent, dans une majorité des cas, lorsqu’une nuisance sonore dangereuse et prolongée
se produit sans prendre les cétacés par surprise, ceux-ci peuvent échapper aux niveaux sonores
véritablement dangereux en s’éloignant de la zone critique, et donc ne pas subir d’insonification
fatale. Par conséquent, les cas de nuisance sonore entraînant un risque fatal pour les cétacés sont
les suivants :
o la forte intensité sonore survient brutalement avec un niveau d’emblée excessif,
o la vitesse de la source sonore est supérieure à la vitesse d’évasion des cétacés,
o le mouvement de la source est imprévisible et entraîne le piégeage des cétacés,
o le mouvement d’évasion des cétacés est entravé par un obstacle,
o les cétacés n’ont pas leur faculté habituelle d’évasion, en raison par exemple de la
présence de nouveau-nés.
Cependant, même si les cétacés ne sont pas soumis à ce genre de risque fatal, ils peuvent
encourir d’autres risques sévères du fait d’une exposition à des niveaux sonores élevés, comme :
o une érosion temporaire de leurs facultés auditives (TTS),
o une nutrition insuffisante ou une exposition aux prédateurs, suite à une évasion
prolongée,
17 Le terme « menace » semble trop fort, mais il ne l’est pas dans la mesure où les plus fortes intensitéssonores peuvent faire fuire durablement des cétacés d’une région qu’ils affectionnent, voire même les tuer.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 37
o un risque d’échouage, dans le cas où leur trajectoire d’évasion les amène dans une zone
inhabituelle par rapport à leur habitat normal.
Selon leur nature, les sources de nuisance sonore que nous allons passer en revue
soumettent les cétacés à ces différents risques. Les facteurs qui influent sur la gravité des
conséquences possibles sont : le niveau de source, le type de bruit (impulsif ou non), la fréquence,
la durée, la localisation de la source et son mouvement. Pour bien documenter la problématique,
nous présenterons les caractéristiques essentielles de chacune d’entre elles, et nous donnerons
un ou plusieurs exemples parmi les conséquences observées chez les cétacés.
Nous commencerons par des sources de nuisance sonore qui sont fixes (explosions, battage
de pieux, forage), ensuite nous aborderons les sources sonores mobiles (à bord de bateaux), avec
des nuisances sonores biens connues, comme les sismiques de prospection géophysique, et les
sonars. Nous terminerons par l’influence des bruits de trafic maritime, un sujet dont l’importance a
longtemps été négligée, mais qui est l’objet d’un intérêt grandissant depuis une décennie. Notre
examen ne sera pas exhaustif : il existe d’autres sources sonores anthropiques qui peuvent
impacter les cétacés. Mais, dans le contexte de la Méditerranée nord-occidentale et du Sanctuaire
Pelagos, les sources que nous avons sélectionnées sont celles qui présentent le risque le plus
grave, ou qui sont les plus fréquentes.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 38
a) Les explosions
Si l'utilisation des explosifs en mer est beaucoup moins répandue en temps de paix qu’en
temps de guerre, elle n'en est pas moins habituelle dans plusieurs cas précis. En premier lieu, on
a les utilisations militaires des explosifs, comme dans le cas d'entraînement avec des munitions
réelles. Il semble que ces exercices militaires aient souvent lieu dans des polygones bien
identifiés, mais qui peuvent être situés dans des aires marines riches en mammifères marins, voire
même protégées. D'autres opérations militaires incluent le déminage, le pétardage de cadavres de
baleines (en Méditerranée), ou le pétardement : ce dernier cas est assez fréquent dans le
périmètre de Pelagos (plus d’une dizaine d’occurrences par année). Le pétardement consiste le
plus souvent à faire exploser des munitions historiques avec une charge de contreminage. Les
charges sont variables, entre quelques dizaines et plus de 200 kg (archives personnelles).
L’opération est menée par un équipage de la Marine Nationale, généralement en dehors de la
saison estivale.
Les utilisations civiles des explosifs incluent le dérochement effectué dans le cas de travaux
portuaires ou d'élargissement de passes, par conséquent en milieu côtier. Richardson et al. (1995)
signalent le cas d'une explosion au Canada, avec 5 tonnes d'explosifs, qui avait semble-t-il abouti
à la mort de Mégaptères : lors de la séquence d'explosions, ceux-ci n'avaient pas été observés
réagissant au bruit, mais deux individus avaient été retrouvés morts le lendemain avec des
fractures (Ketten et al., 1993).
Cette catégorie de nuisance sonore est caractérisée par une faible fréquence dans le temps,
une localisation assez précise dans l'espace, mais des conséquences potentiellement létales pour
les cétacés. En dehors de cas très rares, les explosions ont lieu dans des endroits fixes, souvent
déterminés longtemps à l’avance, et sont souvent uniques.
• Nature des ondes sonores
Les explosions génèrent des ondes de choc et des ondes sonores de très forte intensité.
L’explosion produit d’abord une onde de choc très rapide, dépendant de la nature de l’explosif,
suivie par des pics de pression positive et négative liés aux oscillations de la bulle des gaz issus
de l’explosion. A une certaine distance, l’ensemble des oscillations de pression finit par ressembler
à une onde acoustique, notamment parce que l’onde de choc est rattrapée par les ondes de
pression. C’est une onde impulsionnelle d’une durée inférieure à deux centièmes de seconde. Le
maximum d’énergie est souvent autour de 50-100 Hz, mais l’onde comporte une énergie
importante jusqu'à 1000 Hz et au-delà. Le niveau du pic sonore résultant dépend de la profondeur
à laquelle se trouve la charge explosive et de la profondeur où l’on se trouve, en plus bien sûr
d’être une fonction croissante de la charge d’explosif en question. Vu la complexité des
phénomènes, il est difficile de donner une formulation simple pour le niveau sonore produit par une
explosion. Cependant, une équation approximative donne le niveau de source en bande large de

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 39
l’onde de choc initiale, qui est le plus élevé du train d’ondes produit (NRC, 2003, p72 - converti ici
pour une charge w en kg) :
SL0-p (en dB 1µPa@1m) = 274 + 7,5.log(w)
Le niveau en bande large (toute la bande audible) est ainsi de 274 dB0-p re 1µPa@1m pour
une charge de 1 kg, ou encore 289 dB pour une charge de 100 kg.
Cette onde de choc extrêmement brutale et dangereuse se convertit en quelques dizaines
de mètres en une onde de pression un peu moins intense, pour laquelle Richardson et al. (1995,
p151) proposent une formulation. Ces auteurs présentent un abaque de détermination du niveau
pic de pression maximale en fonction de la charge w et de la distance d, pour une charge
explosant en pleine eau, c’est-à-dire hors de la proximité du fond et de la surface (Fig.18). Leur
abaque peut être transcrit avec la règle empirique suivante :
o Lpic = 202 dB0-p pour une charge w de 0,1 kg et une distance d de 1 km,
o on enlève 20 dB par magnitude de distance,
o on ajoute 7 dB par magnitude de charge,
o la relation donne un niveau de pression de 223 dB pour 100 kg à 1 km, ou un niveau
de pression de 203 dB pour 100 kg à 10 km
Dans le cadre de notre étude, je propose que cette règle serve de référence. On remarque
que la détermination ci-dessus s’applique à l’élévation brutale de pression, sans référence à une
fréquence particulière. Selon Richardson et al. (1995), les expériences menées ont montré que les
lésions constatées sur les organes des animaux dépendaient au premier degré de ce niveau de
pression. On observe que l’énergie maximale des explosions est située entre 10 et 500 Hz ; pour
les hautes fréquences, la densité d’énergie baisse d’environ 20 dB par magnitude de fréquence.
• Effets observés sur des cétacés
Il y a quelques exemples d’effets observés lors d’explosions en milieu naturel, en dehors du
cas relaté plus haut. Van Bree & Kristensen (1974) avaient relevé l'échouage atypique de 4
Ziphius sur l'île de Bonaire (Antilles), événement qu'ils ont rapproché d'exercices navals avec des
explosions qui avaient eu lieu deux jours avant l'échouage. Bien que les paramètres des
explosions ne soient pas connus, il paraît probable que les charges faisaient plusieurs dizaines à
plusieurs centaines de kg, donc capables de générer des niveaux pic-pic de plus de 210 dB à
plusieurs kilomètres de distance. Les auteurs ont d'ailleurs fait le rapprochement avec un
échouage de plusieurs dauphins à bec blanc (Lagenorhynchus albirostris) qui avait eu lieu en
Hollande en 1964, peu après qu'un navire militaire eut procédé à des essais de charges de
profondeur.
D’autre part, Madsen & Mohl (2000) ont conduit une expérience avec des explosions de très
faible charge, 1 g de TNT, dans le nord de la Norvège, non loin de plusieurs cachalots. Le niveau

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 40
de source de ces charges a été évalué à 231 dBRMSep18 en valeur efficace équivalente au pic. En
tenant compte de la position d'un cachalot, il avait été estimé que l'animal situé à 840 m avait reçu
un niveau de 173 dBRMSep. Les auteurs n'ont détecté aucun changement dans le rythme des clics
émis par l'individu, donc pas de dérangement apparent. Pour quatre autres cachalots en plongée,
les effets ont été également indétectables sur le rythme des clics, de même que pour un cachalot
qui respirait en surface à 450 m des charges et qui avait reçu un niveau d'environ 179 dBRMSep. On
peut observer que la signature spectrale de la détonation était assez voisine de celle d'un clic de
cachalot, avec plus d'énergie à basse fréquence. Pour cette raison, ces détonations pourraient ne
pas déranger les cachalots, bien que le niveau soit assez fort et clairement dans la bande audible
du cachalot.
Des expérimentations ont également été menées en captivité, sur des dauphins Tursiops
truncatus et un bélouga Delphinapterus leuca (Finneran et al., 2000). Le but était de soumettre les
cétacés à des intensités sonores équivalentes à celles d'explosions de 5 kg ou 500 kg d'explosifs
de type HBX-1 à des distances allant de 1,7 à 55,6 km, les ondes étant générées par des
transducteurs piézo-électriques pouvant générer des signaux équivalents à ceux des explosifs. On
mesurait les effets des explosions en pratiquant des tests d'acuité auditive avant et après
l'exposition aux bruits. Les tests d'audition ont été réalisés selon la méthode comportementale,
uniquement à des fréquences de 1,2 à 2,4 kHz, correspondant à des fréquences situées 1/2 et 1
octave au-dessus du pic d'énergie spectrale des explosions. Pour les insonifications simulant une
charge de 500 kg à 1,9 km, une baisse d’acuité auditive non significative (3 à 6 dB) a été observée
chez les dauphins mais pas chez le bélouga. Il faut noter qu’un fort bruit blanc masquant était créé
pendant les expériences (dans le but de créer un bruit ambiant constant) ; ce bruit masquant a
probablement influé sur les résultats en augmentant la résilience des sujets face aux détonations.
Autre phénomène intéressant, des dérangements comportementaux croissants ont été notés avec
les expositions les plus fortes : refus de quitter le poste d'exposition après une détonation,
mouvements de nage autour de l'enclos avant de se rendre au poste d'audiométrie, refus de
rejoindre le poste de mesure de l'audiométrie, émissions de vocalisations après l'exposition. Ces
effets comportementaux ont moins touché le bélouga que les dauphins (Finneran et al., 2000).
Les effets auditifs des explosions sur les cétacés sont encore largement évalués sur la base
d’expériences réalisées sur les humains ou d’autres mammifères (Richardson et al., 1995). Pour
les explosions chimiques, le risque est aussi une lésion sévère au niveau des autres organes, en
rapport avec l'onde de pression générée. Des expériences sur des mammifères terrestres
submergés ont permis de déterminer les charges dangereuses : la charge létale augmenterait de
manière régulière avec la masse des animaux (Richardson et al., 1995).
18 Unité signifiant « valeur efficace équivalente au pic »

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 41
Figure 18 : Prévision du niveau de pression (dB) en fonction de la charge (kg) d’explosifs(Richardson et al., 1995, p151).
• Principes de mitigation applicables pour le risque « explosion »
Du fait de l’immobilité de la source sonore, la mitigation du risque peut s’appuyer sur une
surveillance soignée et préalable de la zone critique, dont les dimensions peuvent être
déterminées en fonction de la charge d’explosif, et des autres paramètres de l’explosion. Dans le
cas courant où ces dimensions critiques dépassent les capacités normales de la détection visuelle,
une surveillance acoustique semble appropriée. Des procédés de « mise en alerte » permettent
éventuellement de provoquer un éloignement ou une augmentation de la détectabilité des cétacés.
De toutes façons, la protection préventive des cétacés doit impérativement reposer sur une
adéquation entre la capacité de surveillance et la dimension critique de la zone englobant la
source.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 42
b) Le battage de pieux
Le battage de pieux au marteau pilon est une technique usuelle pour réaliser les fondations
des éoliennes marines19, il s'agit d'une source de bruit très puissante qui perturbe l'habitat des
cétacés côtiers. L’impact environnemental du battage de pieu a été intensivement étudié en mer
du Nord et en Baltique, en raison de l’essor sans précédent de l’industrie éolienne dans ces
régions (Carstensen et al., 2006 ; Tougaard et al., 2009). Par conséquent, l’impact a été surtout
évalué sur le Marsouin commun (Phocoena phocoena), une espèce non répertoriée en
Méditerranée occidentale, ces dernières décennies.
En Méditerranée nord-occidentale, les usines éoliennes marines ne sont pas encore
répandues, ce qui suggère sans doute un fort développement dans le futur. Le battage de pieux
est aussi couramment employé lors de l’agrandissement de grandes infrastructures portuaires.
Comme pour les explosions, ce qui caractérise le battage de pieux, c’est l’immobilité de la
source sonore, et la nature impulsive. De manière évidente, le battage de pieux est pratiqué en
eaux peu profondes : la nuisance sonore s’applique donc au premier chef aux espèces côtières.
Ce qui différencie cette nuisance sonore des marteaux-pilons par rapport au cas précédent, c’est
le fait qu’elle se produit en longues séries, avec une impulsion toutes les quelques secondes.
Selon le cas, cette nuisance sonore peut se prolonger pendant des jours ou des mois,
éventuellement sans interruption nocturne.
• Nature des ondes sonores
Ce sont le plus souvent des mesures in situ qui permettent d’évaluer correctement la nature
et l’intensité des ondes sonores produites : en effet, la faible profondeur et la topographie sous-
marine très variable rendent les modèles de propagation peu performants. David (2006) et Nedwell
et al. (2007) ont publié des rapports détaillés sur les ondes sonores produites par le battage de
pieux : ils observent des ondes à basse fréquence, dont l’énergie est maximale entre 100 et
1000 Hz (Fig.19).
Dans l'ensemble des sites, les niveaux de sources déterminés d'après les mesures valent
environ 250 dB: le premier facteur qui influe sur ce niveau est le diamètre des piles (autour de 4m
dans ces cas). En second lieu, le niveau de source dépend également de la nature du sol et de la
force du martèlement. Dans les cas évoqués par Nedwell et al. (2007), les niveaux de source ont
été évalués entre 243 et 257 dB re1µPa@1m20.
19 Ce n’est pas la seule, il existe également la technique de la fondation gravitaire, ou le forage.20 C’est une mesure globale, sur l’ensemble du spectre de fréquence.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 43
Figure 19 : Spectre d'énergie de l'impulsion d’un marteau-pilon à 100 m et à 10 km de la source(Nedwell et al., 2007).Cas d'un marteau de 450 kJ environ et de piles de plus de 4 m de diamètre.
La forme de l'onde sonore varie beaucoup entre la proximité de sa source (100 m) et la
grande distance (10 km), en raison des multiples réflexions, réfractions et des interactions avec le
sol. D'une durée de 0,5 s près de la source, les impulsions finissent par être presque continues à
grande distance (Nedwell et al., 2007). Les auteurs mentionnent un niveau de pression sonore pic-
pic en bande large de 206 dB à 100 m de la source (re 1µPa) et de 146 dB à 10 km de la source.
Les niveaux de sources extrêmement élevés, alliés à la répétition dans le temps de milliers
d’impulsions, impliquent que le battage de pieux est l'un des phénomènes les plus nuisibles pour
l'environnement acoustique des cétacés, au même titre que les canons pneumatiques, avec la
propriété d'être plus intenses que ces derniers dans les "hautes" basses fréquences voisines de
1000 Hz. Il n’est donc pas étonnant que des réactions très significatives aient été mises en
évidence chez les cétacés lors des études d’impact.
• Effets observés sur des cétacés
A l’exception des explosions, les marteaux-pilons génèrent les intensités sonores les plus
élevées, à égalité avec celles provoquées par les canons pneumatiques des prospections
sismiques. En raison du caractère fixe de la source, la conséquence la plus fréquemment
observée lors du battage de pieux est la désertion de l’habitat par les cétacés, phénomène d’une
ampleur variable, et potentiellement lié à l’espèce concernée, en l’occurrence à son audiométrie.
Si la battage de pieux est potentiellement létal à courte distance, les bruits précurseurs à
l’opération à pleine puissance du marteau-pilon suffisent probablement à éloigner les cétacés de la
zone critique.
En mer du Nord, la réaction du Marsouin commun aux constructions d'usines éoliennes a été
étudiée par Carstensen et al. (2006). Le marsouin est un odontocète dont la sensibilité auditive est

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 44
très bonne de 3 à 100 kHz, mais pas à basse fréquence. La vérification de la présence des
marsouins, cétacés discrets, a été réalisée grâce à la mise en place de balises T-Pod, des
compteurs de clics d'écholocalisation. Le temps d'attente entre deux séries successives de clics a
servi de mesure indirecte de la présence des marsouins. Lors des travaux d'enfoncement d'un
pieu par battage et vibration, ce temps d'attente est passé de 6 heures à 3-5 jours, selon les
emplacements des balises autour du site du marteau-pilon, suggérant que les marsouins avaient
quasiment déserté le site et ses environs pendant les travaux. Dans une étude plus récente,
Tougaard et al. (2009) ont pu estimer que pour une source de niveau égal à 235 dB re1µPa@1m,
le rayon de la zone effectivement concernée par la baisse de fréquentation des marsouins était
compris entre 1 et 20 km. Ces résultats ont confirmé l’étude antérieure de Würsig et al. (2000) à
Hong-Kong, sur une population résidente de dauphins à bosse Sousa chinensis.
Le Marsouin commun possède une audition dont la sensibilité maximale se trouve dans le
domaine des hautes et ultra-hautes fréquences ; si ses facultés auditives sont tout de même
bonnes entre 500 et 1000 Hz (environ 80-90 dB de seuil, Kastelein et al., 2002), elles décroissent
rapidement pour les fréquences plus basses. Les résultats obtenus pour cette espèce donnent une
idée approximative de ce que pourrait être la réaction des dauphins de Méditerranée à des
opérations de battage de pieux, en particulier les espèces qui fréquentent la zone côtière (Grand
dauphin, Dauphin commun, etc.).
• Principes de mitigation applicables pour le risque « battage de pieux »
Le battage de pieux étant une activité fixe, il est possible d’éviter les conséquences les plus
graves en effectuant le démarrage de chaque session avec une puissance réduite, de manière à
permettre un éloignement des cétacés. Puisque les opérations de battage sur un site donné sont
prolongées, il est souhaitable de diminuer la puissance sonore rayonnée par la source, par
exemple en installant autour du site un rideau de bulles. Schultz et al. (2006) ont testé une
enveloppe de mousse placée autour des pieux, qui a aboutit à des réductions 10 à 20 dB des
niveaux de pression sonore entre 1 et 10 kHz, des fréquences importantes pour les odontocètes.
Si elle n’est pas spectaculaire, la désertion des sites du fait des travaux peut néanmoins avoir des
conséquences néfastes significatives au niveau d’un groupe de cétacés, puisqu’elle éloigne les
cétacés d’un habitat favorable à leur nutrition (en général).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 45
c) Le forage
La technique de forage est employée dans le domaine pétrolier, et pour construire des
infrastructures côtières et sous-marines. Elle est intensément employée pour les hydrocarbures
offshore ; ces dernières ont été exploitées dans un premier temps sur le plateau continental (dès
les années 70), puis sur le talus continental, et maintenant en milieu profond. Les conséquences
du forage sur les cétacés ont été étudiées en Arctique, au large des côtes américaines, puis
progressivement dans le monde entier. Les espèces concernées par les études d’impact sous
surtout les mysticètes fréquentant ces habitats (Richardson et al., 1995) : Baleines franches
(Balaena mysticetus, Eubalaena sp), grises (Eschrichtius robustus), plus récemment les autres
espèces (Megaptera novaeangliae), et plus rarement les petits et moyens odontocètes.
En Méditerranée nord-occidentale, cette nuisance sonore n’est pas encore très répandue, en
raison de la rareté des forages pétroliers, les autres applications du forage étant plus
épisodiques21.
Cette source sonore est immobile, comme les deux précédentes, par contre elle est de type
continu, à bande large. Les niveaux d’intensité sonore transmis au milieu marin sont moindres,
mais l’énergie sonore est élevée en raison de la continuité des émissions.
• Nature des ondes sonores
Les bruits de forage sont caractéristiques de ceux d’une machine rotative assortie à des
frottements de matériaux durs. Nedwell & Brooker (2008) ont rapporté des mesures réalisées en
zone de faible profondeur (< 30 m) sur un fond rocheux, avec des trous forés jusqu'à 7,4 m dans le
substrat, avec un diamètre de 1,15 m et un régime de 37 tours par minute. Les niveaux ont été
mesurés en bande large à une distance de 54 m, à 10 m d'immersion : les auteurs ont trouvé de
127-133 dB en valeur efficace (re1µPa), diminuant à 115-116 dB à 830 m de distance. Au-delà, le
niveau de bruit du forage devient inférieur au niveau ambiant moyen. Le niveau spectral est
relativement important jusqu'à une fréquence de 500 Hz (Fig.20).
Le relevé des niveaux sonores en fonction de la distance suggère un niveau de source de
l'ordre de 162 dB re1µPa@1m, avec des pertes par propagation en 16.log(R) (Nedwell & Brooker,
2008). Ces résultats sont en accord avec les éléments exposés par Richardson et al. (1995). On
observe sur ces mesures que le maximum d’énergie spectrale des ondes sonores du forage se
situe dans la bande 20 – 200 Hz, avec plus de 90 dB par Hz jusqu’à environ 1 kHz.
21 Les forages pétroliers et gaziers sont par contre répandus en Méditerranée centrale et orientale.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 46
Figure 20 : Bruits enregistrés le long d'une radiale lors du forage d'un substrat rocheux (d'aprèsNedwell & Brooker, 2008)
• Effets observés sur des cétacés
Les réactions aux plates-formes ou aux bateaux de forage ont été notées ou étudiées dans
le golfe du Mexique ou en Alaska, avec un plus grand nombre d'expériences visant les mysticètes,
particulièrement les baleines du Groenland, B. mysticetus (Richardson et al., 1995). En
Méditerranée occidentale, le Rorqual commun est le seul mysticète résident ; bien que ses seuils
de réaction puissent être différents de ceux de la Baleine du Groenland, les expériences réalisées
dans l’Arctique sont précieuses. Richardson et al. (1995) citent plusieurs résultats cohérents qui
montrent des réactions d’évitement pour des niveaux de pression sonore reçus supérieurs à
115 dB (en bande large), ou encore 105 dB mesurés dans la bande 1/3 d’octave22 la plus forte. Il
est très intéressant de noter que des expériences similaires réalisées sur des Baleines grises par
Malme (in Richardson et al., 1995) suggèrent à peu près le même niveau de 115-120 dB (bande
large) comme seuil de réaction significative. Ces résultats indiquent une bonne convergence des
seuils de réactions pour deux mysticètes appartenant à des genres différents, ce qui est
encourageant pour une possibilité d’extrapolation à B. physalus. Par ailleurs, selon Richardson et
22 Bande de 1/3 d’octave : bande de fréquence située autour d’une fréquence centrale Fc, dont la limiteinférieure est Fc x 2 -1/6 (soit Fc :1,12) et la limite supérieure est Fc x 2 1/6 (soit 1,12.Fc). La largeur de labande est de 26% de la fréquence inférieure. Il y a trois bandes 1/3 d’octave dans une bande d’octave.Dans le cas de sons à bande large, l’oreille humaine restitue le niveau sonore contenu dans une bande de1/3 d’octave. Un son de fréquence F est peu masqué par un bruit dont la fréquence se situe en dehors de labande de 1/3 d’octave qui l’encadre.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 47
al. (1995), les baleines peuvent montrer une certaine habituation lorsque les forages sont
prolongés.
Pour les odontocètes, les expériences ont visé particulièrement le bélouga, lequel a montré
une certaine tolérance au bruit des forages, s'approchant ou ne modifiant pas son comportement
selon le cas jusqu'à des distances de 200-400 m d'une source simulant le bruit d'un forage, avec
des évolutions jusqu'à une distance de 50 à 200 m (Richardson et al., 1995). En bassin, le bélouga
s'est approché de sources de bruit de forage jusqu'à être soumis à des niveaux sonores de l'ordre
de 153 dB. Mais les résultats semblent assez équivoques, et suggèrent que les réactions des
bélougas sont très variables en fonction des conditions environnementales générales, ou en
fonction du contexte socio-comportemental des groupes exposés. Au total, cependant, les
odontocètes semblent montrer une grande tolérance aux bruits de forage en n'évitant pas la
proximité des sites (Richardson et al., 1995). Cela concorde avec les mesures de Nedwell &
Brooker (2008), qui montrent que le bruit sous-marin des forages n'est pas très significatif au-delà
d'une distance de l'ordre d'un km de la source.
• Principes de mitigation applicables pour le risque « forage »
Cette source basse fréquence a démontré son potentiel de nuisance jusqu’à une distance de
10 km, pour des mysticètes. Pour les odontocètes, apparemment les réactions ne sont observées
qu’à courte distance (quelques centaines de mètres). Mais les cas cités dans la littérature ont été
étudiés en zone néritique. En ce qui concerne le Rorqual commun et le milieu méditerranéen, la
première mesure de mitigation à appliquer dans un cas concret serait l’étude précise de la
propagation du son, afin de déterminer à quelle distance on trouve le niveau sonore de 115 dB.
Dans le cas d’une propagation difficile à modéliser, une campagne de mesures des niveaux
sonores permettrait de préciser les mesures de mitigation.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 48
d) Les sismiques pneumatiques
Les prospections géophysiques par explosion d’air comprimé, que l’on peut appeler
« sismique pneumatique »23, sont utilisées intensivement pour étudier la structure du sous-sol
dans le cas de prospections pétrolières (hydrocarbures en général) ou de recherches scientifiques.
Le son produit s’apparente à des explosions (autrefois, on employait des explosifs), elles sont
émises en séries d’au moins plusieurs heures par un navire important qui se déplace à vitesse
faible (souvent de l’ordre de 4-7 noeuds).
Les sismiques à canons pneumatiques sont de loin les sources sonores puissantes les plus
fréquemment utilisées. Il y a en permanence autour du globe plusieurs dizaines de prospections
sismiques en activité. Leur impact sur les cétacés a été étudié depuis plus de trois décennies
(Richardson et al., 1995). D’abord, les recherches se sont surtout focalisées sur les baleines à
fanons, soit parce que les recherches pétrolières avaient lieu dans des régions fréquentées par les
mysticètes (Arctique), soit parce que la nature surtout basse fréquence de l'énergie sonore
semblait menacer en priorité ces espèces. Evans & Nice (1997) examinent l’ensemble de la
bibliographie disponible, comprenant la caractérisation des sources dans différents cas, les
réactions des cétacés constatées de facto, en particulier sur les mysticètes (Baleine du Groenland
et Baleine grise), sur les odontocètes (particulièrement le Marsouin commun et le Dauphin
commun), et aussi sur des poissons. La sensibilité des petits odontocètes aux impulsions de
sismique est moindre que celle des mysticètes en raison de leurs courbes audiométriques qui sont
décalées vers les hautes fréquences, à cause de l’écholocalisation dans la bande 10-100 kHz.
Mais les odontocètes peuvent cependant être perturbés au niveau de leur facultés de
communication dans la bande 0,1-10 kHz, car l’énergie transmise par les impulsions sismiques est
significative dans cette bande jusqu’à plus de 10 km de la source.
Richardson et al. (1995) indiquent que la puissance des systèmes sismiques est fonction
d'une part du volume des canons, de moins de 1 litre pour une simple unité, à 130 litres pour des
grands dispositifs 3D24, et d'autre part de la pression, souvent entre 14 et 31 bars. Selon les
auteurs, le niveau de source atteint par les sismiques peut dépasser 250 dB (en pic-pic
re1µPa@1m). Ainsi, une campagne de sismique est facilement audible à plusieurs centaines de
kilomètres à la ronde (observations personnelles). Par nature, les sismiques pneumatiques sont de
nature plutôt basse fréquence, les émissions sonores sont espacées de plusieurs secondes et
émises en séries durant généralement entre quelques heures et plusieurs jours, mais une
campagne dans une région donnée peut se prolonger plusieurs mois. De très grande puissance,
les ondes atteignent des niveaux forts sur des milliers de km2. La source sonore se déplace
23 Adaptation libre de l’expression « airgun seismic survey »24 Dans la cas d’un dispositif 3D, les canons pneumatiques sont déployés dans une configuration de rateau,avec plusieurs lignes d’émetteurs, de manière à balayer une surface de grande largeur derrière le (ou les)bateau. Pour un système 2D, les canons sont l’un derrière l’autre sur une seule ligne remorquée.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 49
lentement le long de segments de droites organisés pour couvrir de manière systématique toute
une région. Les émissions sonores ont lieu jour et nuit, et potentiellement dans des conditions de
mer variables (pas seulement par beau temps). Tous paramètres confondus, le potentiel de
nuisance des sismiques vis-à-vis des populations de cétacés est certainement sans égal.
• Nature des ondes sonores
Richardson et al. (1995) caractérisent les ondes reçues des canons pneumatiques à plus
d'un kilomètre comme des impulsions durant environ 50 ms (pour un canon unique) avec une
prédominance basse fréquence, mais avec une directivité nette : dans un plan vertical, le niveau
propagé à plus de 40° sur les côtés du canon est in férieur d’au moins 10 dB à celui qui est
rayonné vers le dessous (Fig.21). L'onde sonore produite à la source dure environ 20 ms, elle est
constituée d'une oscillation positive-négative complète qui s'amortit très rapidement (NRC, 2003 ;
Fig.22). Plus on s'éloigne de la source, plus la durée de l'onde reçue s'allonge du fait des
réflexions sur le fond et la surface.
Figure 21 : Directivité de canons pneumatiques multiples (Richardson et al., 1995 p138)
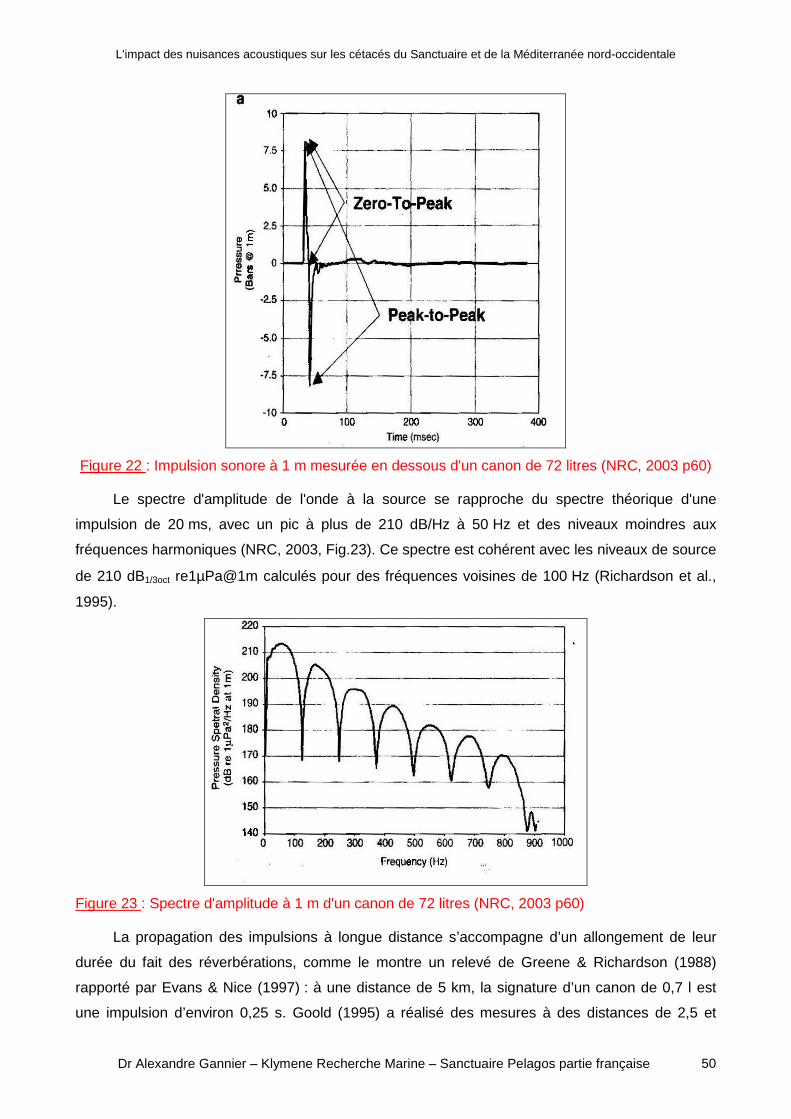
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 50
Figure 22 : Impulsion sonore à 1 m mesurée en dessous d'un canon de 72 litres (NRC, 2003 p60)
Le spectre d'amplitude de l'onde à la source se rapproche du spectre théorique d'une
impulsion de 20 ms, avec un pic à plus de 210 dB/Hz à 50 Hz et des niveaux moindres aux
fréquences harmoniques (NRC, 2003, Fig.23). Ce spectre est cohérent avec les niveaux de source
de 210 dB1/3oct re1µPa@1m calculés pour des fréquences voisines de 100 Hz (Richardson et al.,
1995).
Figure 23 : Spectre d'amplitude à 1 m d'un canon de 72 litres (NRC, 2003 p60)
La propagation des impulsions à longue distance s’accompagne d’un allongement de leur
durée du fait des réverbérations, comme le montre un relevé de Greene & Richardson (1988)
rapporté par Evans & Nice (1997) : à une distance de 5 km, la signature d’un canon de 0,7 l est
une impulsion d’environ 0,25 s. Goold (1995) a réalisé des mesures à des distances de 2,5 et

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 51
10 km, pour des profondeurs de 50-80 m. Le spectre d’amplitude montre une décroissance du
niveau spectral pour les fréquences moyennes et hautes (> 5 kHz), environ 20 dB/Hz, plus
importante que pour les fréquences basses à moyennes (0 à 5 kHz), avec environ 10 dB/Hz. Cet
effet peut être dû à l’absorption des hautes fréquences et aux interactions entre la surface et le
fond (Fig.24). Tout en restant centré sur les basses fréquences, 50-100 Hz, la propagation des
impulsions se fait en produisant une modulation « chirp » vers le bas dans les milieux peu
profonds, alors qu’en milieux profonds la modulation semble se faire vers le haut.
Figure 24 : Spectres d'amplitude à 2,5 et 10 km d'un canon (Goold, 1995 in Evans & Nice, 1997).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 52
Selon Richardson et al. (1995, p137), les ondes de pression atteignent des valeurs de
niveau de source variant entre 222 dBp-p re1µPa@1m (canon de 0,16 l et 14 bars) et 261 dBp-p
(canon de 66 l et 13 bars). Même si les énergies sonores sont prédominantes à moins de 200 Hz
(>200 dB1/3oct. re1µPa@1m), il y a des énergies importantes entre 200 Hz et 2000 Hz (>180 dB1/3oct.
re1µPa@1m), ce qui explique que les odontocètes soient susceptibles d'être perturbés par ces
ondes à plusieurs kilomètres. Le niveau de pression sonore maximal qui peut être infligé à un
animal localisé sous un grand dispositif de canons sismiques est de l'ordre de 235-240 dBRMS en
bande large (NRC, 2003). D’autre part, Richardson et al. (1995) et McCauley et al. (2000)
indiquent que les niveaux propagés sont souvent sensiblement plus faibles près de la surface (de
3 à 10 m) que dans la couche plus profonde (au-delà de 10 m), en raison des interférences de
type miroir de Lloyd.
Des évaluations des ondes sonores produites par les sismiques ont récemment été réalisées
pour des canons pneumatiques employés en géophysique fondamentale, tels ceux d’IFREMER
(Lurton & Antoine, 2007). Lurton & Antoine (2007) donnent les caractéristiques de plusieurs
sources sismiques employées, dont la sismique haute-résolution (HR) de 236 dB @1m et la
sismique multi-traces (SMT) de 252 dB @1m. La première émet une impulsion de 4 ms, durée du
pic principal, toutes les 5 sec, et la seconde une impulsion de 10 ms toutes les 20 sec. La
modélisation de la propagation prend en compte l'effet de surface et les réflexions sur le fond les
plus énergétiques. Pour la SMT, on constate que l'isoplèthe des 180 dB reçus est en gros une
sphère de 2000 m de rayon centrée sous la source à une immersion de 2000 m. L'isoplèthe des
200 dB reçus est une ovoïde d'à peu près 1000 m de diamètre. En surface, le niveau sonore de
plus de 180 dB s'étend à moins d'un km autour de la source. La totalité du volume étudié soit un
cylindre de 5000 m de profondeur et de 15 000 m de diamètre, est insonifiée par un niveau basse
fréquence de plus de 160 dB (Fig.25). Pour la sismique à haute résolution de 234 dB, l'isoplèthe
des 180 dB reçus est une ovoïde de 500 m de diamètre à peu près, sous la source. L'isoplèthe
des 160 dB a une empreinte de 2500 m de rayon à 2000 m d'immersion, mais en surface il s'étend
à moins de 500 m autour de la source.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 53
Figure 25 : Champ acoustique propagé pour la sismique multi-traces de 252 dB de l'IFREMER,pour sa fréquence d'énergie maximale (< 100Hz), d'après Lurton et Antoine, 2007.
Il y a de plus en plus de campagnes sismiques qui sont assorties de programmes de
surveillance et de mesures acoustiques, nécessaires pour évaluer et atténuer les nuisances
apportées à la faune de cétacés. Dans ce cadre, Tolstoy et al. (2004) ont indiqué les niveaux
transmis par un dispositif de 20 canons jusqu'à des distances de 10 km (niveau de source de 252
dB0-p @1m, Madsen et al., 2006), avec des expériences de calibration faites en zone de plateau
(z=30 m) et en zone océanique (z=3200 m). Pour le site peu profond, les niveaux de pression
sonore en bande large de 160 et 180 dBRMS ont été mesurés à environ 12 km et 3,5 km,
respectivement. Pour le site profond, le niveau en bande large de 160 dBRMS a été mesuré à
environ 2,5 km. On observe donc une atténuation beaucoup plus rapide du bruit en milieu
océanique qu'en milieu néritique.
Lors d'une campagne sismique en Sibérie sur des zones de profondeur faible (20 m),
Rutenko et al. (2007) ont voulu déterminer le rayon de sécurité nécessaire pour éviter de déranger
les baleines grises (Eschrichtius robustus) : avec un dispositif de canons assez voisin du
précédent, ils avaient calculé un rayon de sécurité de 4-5 km pour un niveau de 163 dBRMS, alors
que les mesures ont montré que ce niveau était atteint en fait à 7 km. Cet écart important en terme
de distance ne traduit en fait que quelques dB en terme de niveaux sonores propagés.
En résumé, les sismiques industrielles exposent les cétacés à des niveaux de pression
sonore supérieurs à 180 dBp-p jusqu’à des distances supérieures à 2 km, avec une prédominance
des fréquences inférieures à 500 Hz. Les énergies sonores sont maximales en profondeur sous la
source, et peuvent être plus faibles de l’ordre de 10 à 30 dB en surface. Bien que les impulsions

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 54
soient courtes (moins d’1/10è de s), le fait qu’elles soient émises en longues séries implique que
des énergies sonores très importantes sont transférées au milieu marin et aux cétacés des
environs.
• Métrique à utiliser pour les impulsions sonores
Par rapport aux autres ondes sonores, la mesure des niveaux sonores des impulsions doit
intégrer le fait qu’une pression très importante est propagée, mais pendant un temps très court.
Par conséquent la métrique utilisée doit intégrer la notion de temps, pour être comparable à la
métrique utilisée pour les énergies sonores non-impulsives. La plupart des auteurs reprennent la
proposition de Richardson et al. (1995) d'adopter une quantification des impulsions sonores en
équivalent-énergie exprimé en dB re1µPa2.s, intégrant le temps, qui semble propice à la
quantification des énergies sonores reçues et des effets.
Ainsi, McCauley et al. (2000) ont établi pour deux types de canon pneumatique et des
distances de 1,5 à 64 km que le signal sonore est bien caractérisé par une expression
proportionnelle à son énergie en dB re1µPa2.s, plus significative que la valeur efficace (dBrms)25 ou
la valeur pic-pic. Les auteurs ont effectué leurs mesures essentiellement dans des eaux peu
profondes (< 50m). Ils proposent des valeurs de correction empiriques pour passer du niveau
équivalent-énergie au niveau efficace:
niveau dBRMS = niveau dB re1µPa2.s - 10.log(τ)
Cela se traduit en pratique par une correction positive variant de 11,4 à 14,6 dB (car τ < 1s).
Les mêmes auteurs indiquent la correction pour passer du niveau équivalent-énergie au niveau
pic-pic, selon les nombreux cas qu'ils ont traités :
niveau dBp-p = niveau dB re1µPa2.s + 27,3 à 30,5 dB
• Effets observés sur des cétacés
L’impact des sismiques pneumatiques est potentiellement létal du fait des niveaux sonores
très élevés qui sont transmis dans l’eau : récemment, dans le golfe de Guinée on a observé la
mort de dauphins à peu de distance d’une sismique en activité (Gray & Van Waerebeek, 2011).
Mais en général, on observe des effets moins violents, quoique très significatifs, sur les
populations de cétacés, du fait de la faible vitesse de la source. En effet, les cétacés ont souvent la
possibilité de s’évader de la zone fortement insonifiée à l’approche du bateau. Nous résumons ci-
dessous les nombreux travaux portant sur les effets des prospections sismiques sur les cétacés,
dans un ordre de gravité décroissante.
25 Les valeurs efficaces souffrent souvent d'imprécision en raison de la difficulté pour définir un critère uniquepour les temps de début et de fin de l'impulsion (ou la durée τ de l'onde).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 55
o Mort ou échouage mortel
Les impulsions des canons pneumatiques sont moins brutales que celles des explosions
chimiques et ont par conséquent moins de chance de provoquer des dommages physiques par
effet de choc (Gordon et al., 2004). Crum & Yao (1996) ont calculé que la croissance des micro-
bulles d'azote présentes dans les tissus pouvait intervenir en-deçà de 210 dB de niveau de
pression, pour une fréquence de 500 Hz, mais cet effet n’a jamais été mis en évidence pour des
sismiques. Par contre, il semble que les systèmes sismiques puissent être liés à des échouages
accidentels de ziphiidés : une corrélation a été observée en 2002, dans le golfe de Californie, et en
2000 aux Galapagos (Gentry, 2002 ; Malakoff, 2002 in Gordon et al., 2004). De plus, deux
échouages atypiques de mésoplodons de Sowerby ont été constatés aux Açores durant l'été 2009,
et il semble que des campagnes sismiques puissent être mises en cause (M. Fernandez,
comm. pers.). Ces échouages pourraient être dus à des effets comportementaux provoqués par
les tirs sismiques sur les ziphiidés, comme une fuite, ou une perte de facultés induite par
l’interruption du cycle de plongée complexe des cétacés.
o Lésion définitive des organes auditifs (PTS ou permanent threshold shift)
Des lésions aux organes auditifs ont été mis en évidence chez des mammifères marins à la
suite d'explosions chimiques, mais il n'y a pas de cas avéré faisant suite à des tirs sismiques
(Gordon et al., 2004). Par contre, des sons intenses provoquent des TTS (perte temporaire
d'audition), et des sons très intenses ou prolongés entraînent des dommages irréversibles aux
facultés auditives des cétacés (PTS). Les premiers sont induits par une sollicitation excessive au
niveau de l'oreille interne (fatigue auditive), alors que les seconds peuvent découler de la
destruction de cellules ciliées au niveau de la cochlée, ou même d'une lésion sévère de cet organe
de l'oreille interne (Gordon et al., 2004).
En raison du manque de cas documentés chez les cétacés, le risque de PTS doit être
abordé grâce aux études ayant porté sur les mammifères terrestres, en particulier sur le
développement de forts TTS chez les humains. Ward (1968 in Richardson et al., 1995) a étudié
l’effet de la durée de l’impulsion sur la perte auditive : il suggère que le seuil de risque de PTS
diminuerait de 2 dB pour chaque doublement de la durée de l'impulsion, jusqu'à la valeur limite de
200 ms au-delà de laquelle il serait constant. Le seuil de risque diminuerait aussi de 5 dB pour
chaque augmentation d'un facteur 10 du nombre d'impulsions sonores reçues par l'oreille
humaine. Ainsi, le seuil de risque pour une oreille humaine serait de 148 dB pour 1 impulsion de
200 ms, de 138 dB pour 100 impulsions de 200 ms et de 164 dB pour 100 impulsions de 25 µs,
(exprimé en dB re20µPa).
Richardson et al. (1995) effectuent une transposition des résultats pour des humains aux
cétacés sur la base d'une équivalence des dynamiques des deux appareils auditifs (Tab.13).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 56
Niveau pic de risque dePTS pour :
être humaindB re 20 µµµµPa
cétacé trèssensible seuil
auditif de 40 dB
cétacé moinssensible seuil
auditif de 60 dB
1 impulsion de 25 µs 174 214 234
100 impulsions de 25 µs 164 204 224
1 impulsion de 25 ms 154 194 214
10 impulsions de 25 ms 149 189 209
100 impulsions de 25 ms 144 184 204
1000 impulsions de 25 ms 139 179 199
1 impulsion de 200 ms 148 188 208
10 impulsions de 200 ms 143 183 203
Tableau 13 : Critère de risque de dommage auditif définitif pour l'être humain, extrapolé pour desmammifères marins (adapté de Richardson et al., 1995).
Pour les cétacés, les seuils sont en dB re1µPa.
Dans le cas d’un cétacé doué d’un seuil d’audition de 40 dB, il suffirait d’une exposition de
moins d’une heure à des impulsions de niveau pic-pic de 179 dB pour aboutir à une surdité
définitive. Ce niveau correspond à une distance à la source de l’ordre de quelques km.
Mais en réalité, pour l'oreille humaine soumise à des impulsions sonores, le seuil d'apparition
d'une PTS serait beaucoup plus rapide que ne le suggère l’extrapolation précédente (Laroche et
al., 1989). Et, sur la base de l'anatomie comparée de l'oreille moyenne et interne des odontocètes
et des humains, il n'y a pas de raison de supposer que les cétacés soient mieux protégés quand ils
sont soumis à des sollicitations sonores intenses et brusques (Gordon et al., 2004)26. Les seuils
exposés plus haut (Tab.13) sont donc à interpréter avec une grande prudence.
o Diminution temporaire de la sensibilité auditive (TTS pour temporary threshold shift)
L’apparition de TTS, phénomène réversible de fatigue auditive, a été étudiée sur deux
espèces de cétacés, en bassin : Finneran et al. (2002) ont procédé à l'insonification de Grand
dauphin et de Bélouga par des impulsions de canon hydraulique, d’une durée de 10 à 20 ms. Ces
expériences consistaient en l'émission d'une seule impulsion d'intensité croissante au cours de la
série de tests. Pour le Bélouga, un TTS est apparu à des fréquences de 400 Hz et 30 kHz, pour
des niveaux sonores reçus de 226 dBp-p, alors que pour le Grand dauphin, aucune perte
significative n'a été constatée pour un niveau de 228 dBp-p. La récupération du seuil auditif initial
est intervenue après quelques minutes, confirmant que le TTS du Bélouga avait été léger. Le
niveau de 226 dBp-p est à comparer au niveau de 214 dB indiqué plus haut (Tab.13) pour une
impulsion de 25 ms : l’apparition d’un TTS léger se produit avec un niveau pic-pic supérieur de 10
26 Les osselets de l'oreille moyenne sont plus massifs et plus raidis, et la contraction des muscles stapédiauxpourrait avoir moins d'effet atténuateur que chez les mammifères terrestres, en cas de son intense. Parcontre, l'oreille interne est fortement irriguée, ce qui rendrait plus difficile une fatigue auditive due à dessollicitations intenses prolongées.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 57
dB à celui qui est indiqué comme pouvant provoquer un PTS ; cela suggère que la sensibilité
auditive du Bélouga était moins bonne que 60 dB pour la fréquence du canon hydraulique, ou que
la transposition « risque de PTS humain – risque cétacé » n’est pas bonne, ou encore que les
conditions expérimentales sont trop spécifiques pour que les résultats soient généralisables. Nous
reviendrons sur cette expérience en bassin dans le chapitre consacré à ce type d’étude ($B.2b).
o Réactions comportementales
Les niveaux sonores élevés perçus par les cétacés entraînent aussi des réactions
comportementales, dont l’impact est variable : de la simple altération d’activité jusqu’à l’évasion
par fuite, il y a toute une variété d’effets qui ont été constatés en mer, en particulier par des
observateurs indépendants présents lors de prospections sismiques27.
Stone (2003) et Stone & Tasker (2009) ont étudié l'effet des tirs sismiques sur différentes
variables de l'observation des cétacés pendant les campagnes sismiques ; cette étude porte sur
1652 observations de cétacés obtenues de 1997 à 2000. Le premier effet observé est une
diminution du taux d'observation de la plupart des espèces de cétacés lorsque les sismiques sont
en opération (Fig.26).
Figure 26 : Effet des sismiques sur le taux de détection des cétacés (d'ap. Stone & Tasker, 2006).
Ceci traduit l'éloignement des cétacés de la route du navire, au fur et à mesure de sa
progression ; il est spécialement sensible pour les cachalots, les globicéphales et les delphinidés.
27 En Europe, depuis plus d’une décennie, le Joint Nature Conservation Comitee (JNCC) britannique obligeles compagnies à embarquer des observateurs indépendants.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 58
Le second effet mesuré est celui de l’augmentation de la distance minimale des cétacés au
bateau, qui confirme une évasion à l’approche de la sismique.
L'activité sismique influence également significativement de nombreux comportements
visibles en surface, d'une manière générale ou pour certaines espèces (Stone, 2003 ; Stone &
Tasker, 2006). L'activité de nourrissage a ainsi été négativement affectée par les tirs à grande
échelle, puisqu'elle a été observée dans 3,6% des cas pendant les tirs, contre 8,3 % en l’absence
de tirs. Les interactions positives vers le bateau (approche, étrave, etc.) ont fortement décru
pendant les tirs (4,8 % des cas contre 12,7 %), alors que les interactions négatives (éloignement,
etc.) ont augmenté pendant les tirs (3,6 % contre 0,8 %). Les changements de direction par
rapport au bateau sont également assez éloquents, puisque 6,8 % des cétacés se rapprochent du
bateau pendant les tirs, contre 20,4 % qui s'éloignent, alors que 19,2 % des cétacés se
rapprochent du bateau en dehors des séances de tirs, et que 10,3 % s'en éloignent. Cet effet est
statistiquement significatif pour le groupe des mysticètes, les globicéphales, et le groupe des petits
odontocètes (Stone & Tasker, 2006).
En résumé, Stone (2003) et Stone & Tasker (2006) montrent que les tirs sismiques ont pour
effet d'éloigner les cétacés de la route du bateau, de diminuer leur activité de chasse (y compris
pour les cachalots que l'on retrouve davantage en surface), et de générer une activité de nage
rapide, pour la plupart des espèces.
Gordon et al. (2004) rappellent que des effets comportementaux très significatifs, tels que
l'évitement de la zone des tirs jusqu'à plus de 30 km, des sondes plus courtes et des respirations
moins nombreuses, ont affecté plusieurs espèces de mysticètes (Baleine du Groenland, Baleine
grise, Baleine à bosse, Baleine bleue). Les niveaux occasionnant ces dérangements légers
peuvent descendre jusqu’à 125-133 dB, dans le cas de la baleine du Groenland. Différents auteurs
repris dans Richardson et al. (1995) indiquent que des effet comportementaux clairs (évasion) sont
constatés en général pour des niveaux sonores pic-pic de l’ordre de 160 dB, pour les espèces
citées précédemment. Dans une étude récente portant sur le Rorqual commun en Méditerranée,
Castellote et al. (2012) ont constaté une variation dans le rythme et la fréquence des appels des
individus quand une sismique était en activité. Mais l’étude ne donne pas les niveaux sonores
reçus par les baleines.
Pour la plupart des petits et moyens odontocètes (globicéphales, dauphins, marsouins), qui
ont une audition à basse fréquence probablement moins sensible que les mysticètes, les réactions
comportementales aux sismiques peuvent éventuellement avoir lieu pour des niveaux plus forts
(donc à des distances plus faibles). Weir (2008) a mis en évidence la réaction d'un groupe de
globicéphales tropicaux (Globicephala macrorhynchus) pendant la procédure de ramp-up28 d'un
gros système sismique au large du Gabon. Selon Richardson et al. (1995), un tel dispositif pouvait
28 Le ramp-up ou « soft start » est une procédure de montée progressive de la puissance d’une source, quien théorie doit permettre l’éloignement de la faune sensible.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 59
avoir un niveau de pression à la source de 260 dBp-p. Après plusieurs minutes de mouvements
erratiques et d'immobilité, les cétacés ont pris une route les éloignant du navire, ils se sont trouvés
à environ 1500 m dans le travers du bateau lorsque la source a atteint sa puissance nominale.
Weir (2008) a observé que les cétacés n'ont pas choisi de s'éloigner à vive allure de la source au
cours des 30 minutes qu'a duré l'insonification. La pratique d'un ramp-up n'a pas entraîné la
réaction supposée, un éloignement. Dans une étude récente, Thompson et al. (2013) ont quantifié
les effets produits par une sismique 2D de 7,7 l, sur des Marsouins communs en mer du Nord. Ils
ont constaté une évasion de l’habitat jusqu’à 10 km de part et d’autre de la source, une distance
correspondant à des niveaux pic-pic de 165-172 dB. Les mêmes auteurs indiquent que les
marsouins revenaient sur la zone insonifiée en moyenne plusieurs heures après l’activité de la
sismique.
Pour le Cachalot dans le golfe du Mexique, Jochens et al. (2008) et Miller et al. (2009) ont
étudié l’impact de sismiques industrielles sur des individus préalablement taggés. Ils rapportent
que des cachalots ont continué à chasser alors qu'ils étaient exposés à des niveaux sonores de
135-160 dBp-p (119-147 dBRMS). Mais un individu est resté inactif après le taggage et pendant
l'utilisation des canons, la chasse n'ayant repris qu'après l'arrêt des impulsions sonores. Miller et
al. (2009) ont aussi observé que le taux de prédation des cachalots diminuait significativement de
19 % pendant l'exposition aux canons pneumatiques, et que les ondulations motrices des
cachalots diminuaient de 6% environ. Par contre, le comportement élémentaire de sonde restait
inchangé pour 7 des 8 cachalots taggés. Ces expériences ont donc montré un certain
dérangement se traduisant par une tendance à chasser moins efficacement pendant l'exposition
sonore à des niveaux de 138-160 dBp-p.
Figure 27 : Impulsion de canon airgun reçue par un cachalot à 3 km et 15 m de profondeur.
Durant cette même campagne, Madsen et al. (2006) ont montré que les impulsions sonores
reçues par les cachalots à quelques kilomètres comprennent au moins deux composantes, une
directe et une réverbérée, et que leur énergie s'étendaient jusqu'aux moyennes et hautes

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 60
fréquences (Fig.27). L'impulsion réverbérée avait une durée voisine de 0,5 s. Ce facteur n’est pas
du tout anodin pour la prise en compte des réactions de quasiment tous les odontocètes.
En résumé, les effets comportementaux sont constatés pour des niveaux différents chez les
mysticètes et les odontocètes, avec une évasion provoquée par des niveaux voisins de 160 dB
pour les baleines, contre 170 dB environ pour des marsouins. Des effets comportementaux
significatifs mais portant moins à conséquence sont observés pour des niveaux pic-pic reçus de
l’ordre de 130 dB chez les baleines, et 140-160 dB pour les cachalots, et à partir de 165 dB pour
des marsouins.
o Effets sur les proies des cétacés
Un effet marquant des tirs sismiques a été mis en évidence au large de la Norvège : il s’agit
de l’éloignement de certaines espèces de poissons (Evans & Nice, 1997). Des chercheurs ont
montré expérimentalement qu’en présence d’une source de 249 dB de niveau de source, les
morues et les haddocks évacuaient significativement le secteur situé jusqu’à 30 km de la source,
avec une baisse d’abondance de plus de 45 % de ces poissons, allant jusqu’à 70 % autour de la
source. En effet, les poissons ont une audition qui favorise souvent les basses fréquences (Au &
Hastings, 2008). Il semble évident qu’une baisse d’abondance des proies conduit à un
changement de distribution des cétacés prédateurs. Evans & Nice (1997) citent des résultats
montrant que les poissons pouvaient être blessés pour des niveaux reçus de plus de 220 dB, et
également que les oeufs et larves de poissons subissaient une mortalité élevée pour des niveaux
de 236 dB.
Cet effet sur les proies n’est pas encore assez documenté, mais dans le cas de campagnes
sismiques prolongées dans une même région, il peut bien sûr influer sur la fréquentation d’un
habitat par les cétacés.
• Principes de mitigation applicables pour le risque « sismique »
Les principes de mitigation applicables pour minimiser les impacts aux populations de
cétacés découlent de la nature de la source sonore « sismique ». Deux catégories d’impact
doivent guider toute action de mitigation : d’abord les impacts sévères (mort, lésion auditive),
ensuite les impacts sur les populations découlant de l’évasion d’habitat. Dans les deux cas, les
propriétés de la source sont : très forte intensité et audibilité par toutes les espèces, mouvement
assez lent, longue (quelques heures) ou très longue (plusieurs mois) durée de transmission dans
une même région.
Pour faire face au premier type de risque, les compagnies de prospection s’inspirent
généralement du code de bonne conduite de la JNCC (JNCC, 2004), qui comporte l’obligation de
démarrage progressif des tirs (ramp-up ou soft-start), et l’embarquement d’observateurs
indépendants (éventuellement doublés par une surveillance acoustique). Le démarrage progressif
doit en théorie permettre l’évasion des cétacés présents à proximité de la source, et les
observateurs embarqués sont censés détecter les cétacés encore présents dans la zone critique

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 61
et, en principe, obtenir un arrêt des sismiques si un risque d’accident se présente. En général, les
précautions prises lors de campagnes sismiques se bornent à cette catégorie de risque, tout en
ignorant certains aspects d’une mitigation efficace, à savoir, l’impossibilité de détecter certains
groupes de cétacés la nuit et par mauvais temps, et l’extrême difficulté de détecter les ziphiidés.
La seconde catégorie de risques, celle qui est liée à la désertion de l’habitat pendant la
prospection sismique, est souvent peu abordée, voire totalement ignorée. Pourtant, c’est dans
certains cas celle qui peut avoir le plus d’impact sur une population, notamment lorsque des
prospections sismiques se prolongent pendant des mois. La première étape de la mitigation de ce
risque « désertion d’habitat » consiste à évaluer l’impact de la prospection, ce qui nécessite déjà
une évaluation du peuplement de cétacés présents à la période prévue. Les étapes suivantes sont
naturellement les mesures éventuelles pour minimiser l’impact de la « désertion d’habitat ».

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 62
e) Les sonars
Les sonars en général29 sont les sources sonores intenses les plus fréquemment utilisées
par l'homme en milieu marin, car leur diversité s'étend des dispositifs utilisés par les plaisanciers
(écho-sondeurs), en passant par ceux employés par les pêcheurs professionnels, par les navires
océanographiques, par les bateaux militaires, ainsi que par différents engins pilotés et non pilotés,
dont certains sont mis en oeuvre par des aéronefs. Mais en fait, si l’homme ou la femme de la rue
pense immédiatement aux engins militaires quand on lui parle de « sonar », c’est que ces derniers
comptent parmi les engins sonores les plus puissants, et que leur nocivité vis-à-vis des cétacés a
été mise en évidence à l'occasion de plusieurs accidents médiatisés.
Les caractéristiques affectant l'impact d'un sonar sur des cétacés sont les suivantes :
- sa puissance, avec son niveau de source et sa cadence d'émission (ou cycle de charge),
- le type de signal, avec sa fréquence, son mode de modulation, sa durée d'impulsion,
- sa directivité, c’est-à-dire l’angle couvert par le faisceau dans l’espace,
- ses conditions spatiales et temporelle d'emploi, y compris son immersion, le mouvement,
de la source (vitesse et direction), la durée et la concentration des transmissions,
- les précautions prises lors de sa mise en oeuvre.
Si certains sonars militaires ont été mis en cause lors d’accidents survenus à des cétacés,
c'est a posteriori qu'ils réunissent les caractéristiques qui les rendent plus nocifs que les autres : ils
sont de forte puissance, à basse-moyenne fréquence, peu directifs, les impulsions sont assez
longues (une ou plusieurs secondes), la durée d’utilisation est longue également (une ou plusieurs
dizaines d’heures), et dans un secteur géographique qui peut être restreint. Mais notre examen ne
doit pas ce restreindre à ces sonars-là, car d’une part, d’autres sonars peuvent réunir des
caractéristiques qui les rendent potentiellement dangereux, et d’autre part, le contraste avec
d’autres engins réputés inoffensifs peut faciliter la compréhension des accidents et de leurs
causes.
• Nature des onde sonores de quelques sonars
Pour des raisons de confidentialité industrielle ou militaire, il est difficile de connaître les
caractéristiques techniques fines des différents types de sonar. Les sonars utilisés en pêche
professionnelle ou en pêche-plaisance peuvent être mono-faisceaux et bi-fréquence, par exemple
50 kHz/200 kHz avec des directivités prononcées et des niveaux de source parfois supérieurs à
210 dB (pêche-plaisance) et supérieurs à 225 dB (pêche-professionnel). Il y a aussi des sondeurs
de pêche qui sont panoramiques (Lurton & Antoine, 2007). Les sondeurs de recherche
appartiennent à des catégories très diverses en terme de fréquence, de niveau de source et de
29 La terminologie anglo-saxone parle de « active sonars » (LFAS, MFAS) lorsque la terminologie françaisedéfinit simplement par sonar un engin envoyant des sons et qui analyse les échos reçus en retour.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 63
directivité. Lurton & Antoine (2007) dressent une liste des principaux systèmes sonars employés
par l'IFREMER à bord de ses navires océanographiques.
- Les sondeurs de sédiments sont destinés à l'exploration des strates sédimentaires
superficielles, ils émettent dans une gamme de fréquences moyennes (typiquement 1 à 8 kHz) et à
des niveaux de source assez importants (typiquement 200 à 215 dB re1µPa@1m). Les signaux
sont le plus souvent modulés en fréquence durant plusieurs dizaines de millisecondes, et avec des
cadences assez rapides de l'ordre de la seconde. Ils sont directifs (faisceau unique de quelques
dizaines de degrés) et émettent verticalement.
- Les sondeurs multifaisceaux sont des systèmes à large couverture transversale, destinés à
effectuer la cartographie des fonds marins. Ils peuvent être divisés en trois catégories selon leur
fréquence de travail : les sondeurs « grands fonds », (12 à 30 kHz), les sondeurs « plateau » (70 à
100 kHz) et les systèmes haute fréquence (au delà de 200 kHz). Les niveaux de source maximaux
des sondeurs multifaisceaux sont assez élevés (225 à 240 dB re1µPa@1m). Les signaux utilisés
sont très majoritairement non modulés, et durent de 0,1 à 20 ms selon les systèmes, les
fréquences utilisées et les conditions d'utilisation. Les cadences de transmission sont comprises
entre 0,5 s et 20 s, en fonction de la hauteur d'eau ; les plus rapides correspondent aux hauteurs
d'eau les plus faibles, et aux impulsions les plus courtes. La forte puissance des sondeurs
multifaisceaux est contrebalancée par leur directivité très sélective: typiquement le faisceau est un
éventail aplati d'ouverture 120°-150° dans le sens transversal et 1°- 3° dans le sens longitudinal.
- Les sondeurs monofaisceaux couvrent les mêmes fréquences que les sondeurs
multifaisceaux : de 12 kHz (grands fonds) à plusieurs centaines de kHz. Les cadences d'émission
peuvent être comparables à celles des sondeurs multifaisceaux, à hauteur d'eau égale. Les
sondeurs monofaisceaux émettent de manière très directive vers le fond : le lobe est étroit
(inférieur à 16°). Pour la configuration la plus pé nalisante chez IFREMER (38 kHz, 231 dB
re1µPa@1m), l’isoplèthe 180 dB re1µPa est à 260 m : à cette distance, la largeur de la zone
éclairée par le faisceau (angulairement 7°) est de 32 m.
- Les sonars panoramiques de pêche émettent de manière omnidirectionnelle autour du
navire porteur, en favorisant des directions proches de l'horizontale. Leurs niveaux émis sont
comparables ou inférieurs (du fait de leur caractère panoramique) à ceux des sondeurs
monofaisceaux, mais les signaux émis peuvent être plus longs : en effet, de grandes portées sont
obtenues grâce à l'emploi de signaux modulés en fréquence. Ces sonars sont par nature peu
sélectifs spatialement ; ils sont susceptibles d'être pointés dans toutes les directions.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 64
Catégorie Type FréquencekHz
Impulsion etcadence
Niveau desource (dB)
Directivité Emploi
Recherche: sondeurde sédiment
3-6 50 ms, 1 sec 203 30° l'Atalante
Recherche: sondeurde sédiment
1,8 - 5,3 50 ms, 1 sec 214 45° - 20° Le Suroît LePourquoi Pas
Recherche: sondeurmultifaisceaux
KongsbergEM 300
30 1 à 10 ms, 1 à20 sec
234 1° x 150° L'Atalante
Recherche: sondeurmultifaisceaux
KongsbergEM 1000
95 0,2 à 2 ms,0,1 à 1 sec
226 3° x 150° Le Suroît
Recherche: sondeurmonofaisceau
EA-600 12-38-200 0,06 à 16 ms,>0,05 sec
223-231-228 16°-7°-7° Pourquoi pas
Sonar latéral Edge TechDF-1000
100-400 0,1 à 1 ms,0,1 à 1 sec
220 1° ou2°x170°
L'Europe
Panoramique depêche
Furuno CH-32 60 32 ms, 0,25 à2 sec
222 9°x 8° Gwen-Drez
Sondeur de pêche Furuno 620 50/200 variable 600W rms 28°/5,5° plaisance
Militaire ASM Exp. TVDS LF 0,45 -0,7 2+2 sec,chaque 60 sec
214-228 faisceau23°x360°
SaclantCen
Militaire ASM Exp. TVDS MF 2,8 -3,3 2+ 2 sec,chaque 60 sec
223-226 faisceau20°x360°
SaclantCen
Militaire tactique SQS-53C 2,6 -3,3 0,5 à 2 sec,chaque 26 sec
≥ 235 faisceau 40°x120°/360°
US Navy etOTAN
Militaire tactique SQS-56 6,8-7,5-8,2 chaque 26 sec 223 faisceau30°x360°
marines US etOTAN
Militaire ASM SURTASSLFA
0,1-0,5 6-100 s chaq.360-900 s
240(18*215)
faisceau5,5°x360°
US Navy -4 systèmes
Militaire ASM remorquéSLASM
< 3 ? 220 < ? < 230 2 frégatesMarine Nation.
Militaire tactique remorqué2087
0,5 - 2 ? 220 < ? < 230 Frégates T.23Royal Navy
Militaire tactique remorqué2149
< 3 ? 220 < ? < 230 FrégatesAquitaine
Militaire anti-mines remorqué2093
> 50 ? multi-fréquences
? 210 < ? < 230 ? Fabriqué parThalès
Militaire anti-mines remorquéDUBM 44
> 50 ? ? 210 < ? < 230 à ouverturesynthétique
MarineNationale
Militaire anti-mines de coque2022 et 2193
> 50 ? bandelarge
? 210 < ? < 230 ? Fabriqué parThalès
Militaire aéroporté bouéelargable
ASM
Militaire aéroporté trempé soushélicoptère
ASM
Tableau 14: Caractéristiques de sonars actifs utilisés dans différentes activités humaines.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 65
Les sonars militaires tactiques
On peut les appeler tactiques car ils ont vocation à être utilisé en combat, contre des sous-
marins, par des frégates, navires puissants et manoeuvrants. Ils appartiennent à différentes
catégories spécifiques à leur emploi : les sonars à basse fréquence anti-sous-marin à longue
portée, les sonars à moyenne fréquence (MFAS pour Medium Frequency Active Sonar) anti-sous-
marin. Selon les technologies, ils peuvent être montés sur la coque des navires (SQS-53C), ou
remorqués à des immersions variables (2087 britannique, 2149 français). On connaît rarement
leurs caractéristiques techniques générales, mais les sonars TVDS (Towed Vertically Directive
Source), les deux sonars tactiques SQC-53 et SQS-56 (et le système LFA américain) font
exception, en raison des études d'impact qu'ils ont nécessitées, ou des rapports d'accidents
publiés (Zimmer, 2004).
Les sonars tactiques à moyenne fréquence (1 à 5 kHz) émettent des signaux variés et
adaptés à différentes fonctions (Evans & England, 2001) : les signaux à bande étroite et fréquence
constante sont adaptés à la détermination des mouvements (effets Doppler), et les signaux à
fréquence modulée (FM) sont adaptés à la détection des objets en milieu réverbérant. Le sonar
SQS-53C émet une série d’impulsions courtes (fréquence env. 3 kHz), avec différentes
composantes qui peuvent être variées, chaque série étant séparée de la suivante par un délai
dépendant de l’horizon qui est scruté. Aux Bahamas en 2000, un des navires émettait ainsi une
série de trois impulsions de 0,5 sec pendant 2 sec, suivie par un silence de 24 sec avant l’émission
suivante, soit un cycle de 26 sec. Les TVDS employés en Grèce en 1996 avaient par contre une
durée de cycle plus longue, avec deux impulsions de 2 sec toutes les minutes, et deux bandes de
fréquence actives. Les sonars remorqués de type 2087, 2149 et SLASM sont des « poissons » à
immersion variable (quelques centaines de mètres) qui émettent dans le domaine de la basse-
moyenne fréquence, mais pour lesquels les caractéristiques de signal et de cycle ne sont pas
publiées. On doit avoir des impulsions de durée supérieure à 1 sec, une ou plusieurs fois par
minute.
D'après la comparaison entre les engins civils les plus puissants et les militaires (Tab.14), on
remarque que ce qui distingue davantage les deux catégories est d'abord la durée des impulsions,
inférieure à 10 ms pour le multifaisceaux de recherche EM300, mais supérieure à une seconde
pour les engins militaires. En second lieu, les engins civils à forte puissance se situent dans le
domaine des très hautes fréquences (plusieurs dizaines de kHz), mais on est dans le domaine des
moyennes-basses fréquences pour les militaires. Enfin, les sondeurs de recherche puissants
émettent un faisceau de grande ouverture (env. 150°) dans un plan vertical perpendiculaire à l'axe
du bateau, mais de très faible ouverture dans le plan longitudinal (1 à 3°). Au contraire, les sonars
tactiques puissants émettent un faisceau de grande extension sur un disque horizontal atteignant
360° (couvrant tout le tour du navire) et dont l'ou verture verticale est importante (30°) pour les
moyennes fréquences et plus faible (5,5° ?) pour le s basses fréquences. Les volumes insonifiés et

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 66
la durée d'insonification sont donc beaucoup plus importants pour les sonars militaires que pour
les engins civils de niveau de source équivalent. Certains sonars militaires (en général européens)
ont également la propriété d’émettre à des immersions variables.
Le sonar SurTASS LFA
Le Surveillance Towed Array Sensor System - Low Frequency Active, est un système très
basse fréquence de surveillance à très longue portée, différent dans son utilisation et dans sa
conception des sonars actifs tactiques. Il n'est pas destiné à être remorqué à grande vitesse, se
déplaçant à environ 3 noeuds au plus (Fig.28).
Figure 28 : Configuration générale d'un sonar SURTASS-LFA (d'après FSEIS, 2007).Vertical sous le bateau, l’émetteur, horizontal derrière le bateau, le récepteur.
Voici les caractéristiques de la composante active (LFA) du système (FSEIS, 2007):
- la source est une antenne verticale de 18 projecteurs qui transmettent un faisceau omnidirectionnel dans leplan horizontal, avec une épaisseur réduite dans le plan vertical et qui peut être dirigé légèrement vers lehaut ou vers le bas ; l'immersion est de l'ordre de 122 m,
- chaque projecteur du LFA a un niveau de source égal ou inférieur à 215 dB @1m, mais le champ sonorede l'antenne ne peut pas être supérieur à celui d'un seul projecteur (215 dB),
- la fréquence est entre 100 et 500 Hz, et les ondes transmises peuvent être variables, y compris dessignaux continus ou à fréquence modulée,
- le signal transmis est typiquement une combinaison de différentes ondes sonores qui varient en fréquenceet en durée; le ping est formé de ce train d'ondes et dure entre 6 et 100 s, avec une moyenne de 60 s,chaque composante du train ne durant pas plus de 10 s,
- le cycle a un taux de charge30 de moins de 20%, entre 2003 et 2007 il a été en moyenne de 7,5 à 10 %:l'intervalle entre 2 trains d'ondes est typiquement de 6 à 15 minutes.
30 Le taux de charge d’un sonar est la proportion de temps durant laquelle il transmet des impulsions,lorsqu’il est en fonctionnement. Typiquement moins de 10%, par ex. 2 sec toutes les 20 s.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 67
C’est un système utilisé à quatre exemplaires par l’US Navy, il est sans doute présent en
Méditerranée.
Autres sonars militaires
A l’autre extrémité du spectre des sonars militaires, on trouve des engins à très haute
fréquence destinés à la localisation et à l’identification des mines et des petits objets sous-marins.
Les sonars français DUBM-21 (coque) et DUBM-44 (remorqué, profond) sont employés sur les
chasseurs de mines ou autres bâtiments de surveillance anti-mines. Les caractéristiques de
puissance de ces engins ne nous sont pas connues, la puissance est moindre que les sonars
tactiques ; le caractère très haute fréquence (50 à 100 kHz ? ou plus ?) de ces systèmes les rend
potentiellement agressifs vis-à-vis des cétacés. De plus, nombre de ces sonars émettent à
plusieurs fréquences, en bande large, permettant aux utilisateurs d’obtenir une image acoustique
du fond et des objets immergés.
Autres dispositifs acoustiques dont les caractéristiques sont peu connues, les sonars
aéroportés : soit les bouées acoustiques actives larguées par des avions de patrouille, soit des
sonars trempés déployés par des hélicoptères volant en stationnaire. Dans les deux cas, la finalité
est la lutte ASM, donc les fréquences ne sont ni très hautes (question de longueur d’onde et
absorption) ni très basses (taille réduite des engins). Ces sonars sont très couramment employés
lors d’exercices à grande échelle, ou d’entraînements. Les dernières générations de sonars
héliportés sont sans doute d’une puissance accrue (type sonar « Flash »). Pour les sonars
aéroportés, la nocivité vis-à-vis des cétacés peut éventuellement découler de la soudaineté du
déclenchement des transmissions, dans des sites ou quelques minutes auparavant il y avait peu
d’activité humaine, donc pas de « préavis » permettant éventuellement une évasion des cétacés.
• Effets des sonars sur les cétacés
L’effet des sonars sur les cétacés est de mieux en mieux connu, car les accidents multiples
de ces deux dernières décennies ont incité les organismes militaires à s’intéresser au problème :
les travaux scientifiques sont maintenant nombreux. Ils portent sur les échouages eux-mêmes,
mais aussi sur les effets comportementaux et auditifs des sonars.
o Echouages atypiques en relation avec des transmissions de sonar
Par rapport justement aux cas d’accidents les plus spectaculaires, ceux qui se traduisent par
des échouages multiples atypiques, il faut relativiser le petit nombre qui est officiellement
répertorié, depuis 1996 : il y a évidemment de nombreux cas où des accidents liés aux sonars
n'ont pas été connus car les cétacés ne se sont pas échoués, ou bien les échouages n'ont pas été
signalés. Notons que très souvent les manoeuvres navales n’entraînent pas d'échouage atypique.
Au regard du nombre d'utilisations de sonars de tous types chaque année dans le monde
(Hildebrandt, 2004 ; Lurton & Antoine, 2007), si les accidents liés aux sonars sont assez peu
fréquents, c'est qu'ils sont causés par certains types de matériel seulement, ou dans des
conditions particulières. Plusieurs publications ou rapports présentent un tableau initialement

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 68
compilé par Hildebrandt (2004, mis à jour dans OSPAR, 2009) et présentant une liste des
échouages de deux Ziphius de Cuvier au moins remontant à 1963, avec un cas recensé en 1914
(ICES, 2005 ; OSPAR, 2009). Ce type de tableau n’est pas dépourvu d’intérêt, mais il réunit des
éléments disparates qui en majorité ne sont pas explicitement reliés à des utilisations de sonars
actifs. De plus, le critère le plus significatif pour définir un échouage atypique de ziphiidés serait de
ne considérer que les effectifs supérieurs ou égaux à 3, puisque un effectif de 2 comprend l’unité
élémentaire « mère + jeune », dont l’échouage conjoint ne constitue pas un événement atypique.
Le récapitulatif que je propose réunit un nombre plus restreint d’accidents, portant sur une
période plus récente et pendant lesquels la présence de plusieurs navires militaires a été
constatée et rapportée dans un document rapportant l’échouage (critère minimum), ou bien la
présence navale a été documentée dans un rapport d’accident (critère idéal). Nous ne nous
restreignons pas aux ziphiidés. Par rapport au rapport OSPAR (2009), la mention d’un accident en
Alaska en 2004 est ajoutée, car elle est documentée dans Lurton & Antoine (2007), ainsi bien sûr
que plusieurs accidents récents en Méditerranée.
Lieu et date Cétacéséchoués
Exercice Sonars employés Référence
Grèce, mai 1996 12 Zc essai SaclantCen TVDS BF et MF Frantzis et al., 2004
Grèce, sept 1997 9 Zc oui inconnus Frantzis et al., 2004
Madère, mai 2000 3 Zc OTAN modèles desmarines de l’OTAN
Freitas et al., 2004
Bahamas, mars 2000 9 Zc+3 Md+ 2Ba+1 Sf
manoeuvre US sonars tactiques MF Evans & England,2001
Canaries, septembre 2002 7 Zc+ 3 Md+ 1Me+ 3 ziph. sp
Neo Tapon 2002 sonars tactiques MF,marines OTAN
Cox et al., 2006
Alaska, juin 2004 6 ziphiidés Northern Edge 04 US ... Lurton & Antoine,2007
Canaries, juillet 2004 4 Zc morts Majestic Eagle 04 modèles desmarines de l’OTAN
Jaber et al., 2007 inOSPAR, 2009
Caroline du nord, janvier2005
Gma, Pe, Ks émissions de sonar pardes navires US
sonars tactiques MF Hohn et al., 2006
Hawaii, juillet 2004 1 + (175) Pe RIMPAC 2004 sonars tactiques MF Southall et al., 2006
Alboran, janvier 2006 > 4 Zc présence de naviresUK et US
sonars tactiques MF Arbelo et al., 2007
Cornouaille UK, juin 2008 > 26 Dd exercice ASM RoyalNavy et OTAN >30 u.
sonars tactiques MFHF et héliportés
Jepson et al., 2009Jepson et al., 2013
Sicile, février 2011 2 Zc Proud Manta 2011 sonars tactiques MF Emails Podesta etal., 20 fév.2011
Ionienne, nov./déc. 2011 > 9 Zc Mare Aperto 2011navires italiens OTAN
sonars tactiques MF Emails Frantzis etal., 30 nov.2011
Tyrrhénienne, octobre2012
>= 1 Zc Corsican Lion 2012marine française et UK
sonars tactiques MF Corse Matin25/10/12, 14/03/13
Tableau 15 : Echouages accidentels de cétacés liés à l'emploi de sonars militaires.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 69
Comme on peut s’en rendre compte, malgré les progrès scientifiques, les échouages liés
aux sonars continuent à se produire, en particulier en Méditerranée. C’est pourquoi on ne peut
considérer le problème comme résolu, malgré les dizaines de millions qui ont été consacrés aux
recherches autour de cette problématique.
Les échouages atypiques sont l’aspect le plus remarquable des conséquences possibles des
transmissions de sonar militaire, et il est possible que certains d’entre eux soient le résultat
tragique d’un effet comportemental (évasion ou fuite, comme dans le cas de la Cornouailles en
2008 – Tab.15), alors que d’autres sont la conséquence ultime de lésions peut-être non létales
causées aux cétacés par les fortes intensités sonores. Il est certain que les conséquences
comportementales de l’utilisation des sonars sont extrêmement fréquentes, mais sont rarement
relatées, car observées uniquement par des personnels militaires qui n’ont pas pour mission soit
de relever ce genre d’observation, soit de les diffuser31.
Les effets comportementaux ont pourtant été parfois rapportés, ou bien intensivement
étudiés lors de Controled Exposure Experiments (CEE).
o Effets comportementaux observés en mer – hors CEE
Des observations opportunistes ont tout d’abord suggéré que les sonars provoquaient des
réactions d’évasion chez diverses espèces de cétacés. Ainsi, Watkins et al. (1993) rapportent que
des cachalots taggés en octobre 1991 avec des balises-transpondeurs émettant des signaux à 32
et 36 kHz n'ont pas semblé avoir de réaction à ces sons artificiels, ainsi qu'aux impulsions
d'interrogation (30 kHz) et de recherche (36 kHz) produites par le bateau. Par contre les auteurs
signalent que les cachalots ont cessé leur activité et disparu de la zone d'expérience lorsqu'ils ont
reçu un signal fort de sonar de sous-marin (fréquence inconnue). De par ses propres émissions
sonores, le cachalot est supposé avoir une meilleure audition dans la bande 2-20 kHz qu'à la
fréquence de 30 kHz et au-delà. Des réactions d'évitement avaient également été constatées par
Maybaum (1993) lors de l'expérience de transmission d'un sonar de fréquence 3-3,6 kHz sur des
Mégaptères : les vitesses d'éloignement étaient d'autant plus rectilignes que le niveau de la source
était fort (celui-ci n'est pas donné).
En Méditerranée, dans la région du Sanctuaire Pelagos, Rendell & Gordon (1999) ont
observé en août 1994 la réaction acoustique d'un groupe de globicéphales noirs à un signal de
sonar dans la bande de fréquence de 4-5 kHz et qui était composé de plusieurs impulsions courtes
et d'une impulsion longue de type "cri" de 2 s. de durée. Le cycle complet était de 42 s et la source
a été identifiée plus tard comme un sonar remorqué derrière un navire militaire italien. La réaction
des globicéphales était l'émission d'un nombre beaucoup plus élevé de sifflements pendant et
après chaque transmission du sonar.
31 Observons que le Marine Mammals Risk Mitigation Project du NATO Undersea Reseach Center préconiseque des observateurs dédiés rapportent tout événement liés à des groupes de cétacés, pendant lestransmissions sonar.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 70
Plus récemment, lors du grand exercice naval RIMPAC de 2004, Southall et al. (2006)
mentionnent qu’un groupe de 150-200 dauphins d'Electre ont été confiné dans la baie d'Hanalei
(juillet 2004) à Kaua'i (îles Hawaii). Le groupe a pu recevoir des niveaux sonores de 138-149 dB
de sonars dont l'un semblait être un SQS-53C de fréquence centrale 3 kHz. Six sonars de types
différents étaient utilisés au moment de ces événements, à une trentaine de milles de la baie où
les dauphins étaient confinés. On peut en déduire que des petits delphinidés océaniques
réagissent à un niveau de 140-150 dB en se rapprochant dangereusement du rivage.
Cet événement se rapproche d’un autre cas plus récent, qui s’est produit en Manche en
2008 (Tab.15) et durant lequel un grand groupe de Dauphins communs (Delphinus delphis) s’est
trouvé piégé dans un estuaire de Cornouailles à cause de l’activité acoustique anti-sous-marine
d’un grand exercice naval. Un total de 26 dauphins a fini par s’échouer et leur autopsie exhaustive
n’a révélé aucune cause particulière d’échouage, hormis une réaction à la présence des sonars
anti-sous-marins (Jepson et al., 2013).
Les effets comportementaux des transmissions de sonar apparaissant peu à peu comme
significatifs voire sévères, plusieurs gros programmes de CEE ont été lancés de part et d’autre de
l’Atlantique afin de mieux mesurer ces effets. Les espèces visées ont tout d’abord été les ziphiidés,
espèces les plus critiques, mais les expériences se sont progressivement étendues à beaucoup
d’autres espèces, delphinidés, ou mysticètes.
o Effets comportementaux observés en mer lors de CEE
Les programmes principaux de CEE sont au nombre de trois, ils ont tous pour objectif
d’étudier les réactions des cétacés aux sonars militaires :
- Bahamas Behavioral Response Study : il a eu pour espèces focales principales le Mésoplodon
de Blainville (M. densirostris), mais aussi plusieurs espèces de delphinidés tropicaux (Tab.16),
- 3S experiments : ce programme a lieu principalement dans les mers nordiques européennes et a
pour espèces focales principales l’Orque, le Cachalot, le Globicéphale noir, et plus récemment
l’Hyperoodon arctique, et des mysticètes (Tab.16),
- Biological and Behavioral Studies of Marine Mammals in Southern California – SoCal : ce
programme traite des réactions comportementales des espèces du Pacifique tempéré, y compris
les ziphiidés, delphinidés et mysticètes que l’on rencontre au large de la Californie (Tab.16),
Les expériences menées en Méditerranée par le NURC (NATO Undersea Research Center)
et le WHOI (Woods Hole Oceanographic Institute) en collaboration avec certains scientifiques
européens, visent en priorité le Ziphius, mais aussi le Cachalot et, de manière opportuniste,
d’autres espèces. Ce programme n’est pas l’objet d’un suivi aussi visible que les trois précédents,
il ne semble pas que des Ziphius ou d’autres odontocètes de Méditerranée aient été l’objet
d’expériences de CEE (D’Amico, 2009), bien que des taggages aient été réussis. Les points
communs de ces programmes sont l’aspect pluri-annuel, l’emploi de balises digitales

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 71
enregistreuses Dtag ou leur équivalent, et des moyens d’investigation très lourds mis à disposition
par les organismes liés à la défense.
Ces moyens comprennent au moins un navire source de sonar, un bateau d’observation des
cétacés, un semi-rigide avec l’équipe de « taggeurs ». Sans entrer dans les détails des résultats
des études, d’ailleurs pas forcément disponibles, nous proposons de résumer ci-dessous les
expériences effectuées durant ces trois programmes, entre 2007 et 2013.
� Le programme (Bahamas) Behavioral Response Study
Chronologiquement, c’est le second des grands programmes, il se déroule sur la zone
d’exercice AUTEC de l’US Navy aux Bahamas. Même si ce n’était pas officiellement affiché, il
visait à comprendre l’accident survenu à des cétacés en 2000 aux Bahamas lors d’un exercice
naval (Tab.15). Ce sont d’ailleurs des sons proches de ceux du sonar tactique SQS-53 qui sont
projetés sur les cétacés. Le principe pratique de l'expérience est de poser une balise numérique
DTag sur un cétacé, et d'enregistrer les réactions de l'animal à différents sons qui sont transmis
depuis un navire-source. Ces sons comprennent des sons artificiels (sonar, fréquences modulées,
bruits blancs, fréquence de 3 kHz environ) et naturels (cris d'orques). Les expériences ont été
réalisées sur 4 espèces (Boyd et al., 2008, 2009) : le Mésoplodon de Blainville, le Globicéphale
tropical (G. macrorhynchus), le Dauphin d’Electre (Peponocephala electra) et le Pseudorque
(Pseudorca crassidens). Même si le nombre d’expériences a été réduit par rapport aux prévisions,
certains résultats sont très intéressants (Tab.16) :
Espèce taggée Année Type d’Expérience Effet observé
M. densirostris 2007 : 5 comportement -
M. densirostris 2007 : 1 playback sonar MF et orque interruption d’activité et fuite
M. densirostris 2008 : 1 playback bruit blanc MF interruption d’activité
G. macrorhynchus 2007 : 2 comportement -
G. macrorhynchus 2007 : 2 playback sonar MF et orque changement d’activité vocale
G. macrorhynchus 2008 : 2 playback bruit blanc et sonar MF peu d’effet
P . crassidens 2008 : 2 playback bruit blanc et sonar MF évasion accélérée activité vocale
P. electra 2008 : 1 playback bruit blanc et sonar MF peu d’effet
Tableau 16 : Expériences de CEE durant le programme Bahamas BRS 2007 et 2008.
Les expériences Bahamas BRS se sont achevées après 2008. Si leur nombre est assez
faible, quelques résultats inattendus ont été obtenus : en premier lieu, les réactions fortes des
Mésoplodons, en second lieu, des changements d’activité plus légers chez les trois autres
espèces. Il sera intéressant de reporter les effets observés en fonction des niveaux sonores reçus
(voir §B.2). Les expériences Bahamas BRS servent plus ou moins de modèles aux autres
programmes pluri-annuels.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 72
� Le programme 3S
Au contraire du précédent, le programme « Sea Mammals Sonar Safety » est animé
principalement par des pays européens (Norvège, Hollande), avec la participation d’organismes de
recherche gouvernementaux US (WHOI) et britanniques (SMRU). Son objectif est principalement
d’étudier les réactions des cétacés à des sonars utilisés par ces pays d’Europe, engins remorqués
à des fréquences de 1-2 kHz (appelés LFAS) et 6-7 kHz (MFAS), et aussi à des sons d’orques ;
les espèces étudiées ont été d’abord l’Orque, le Cachalot et le Globicéphale noir (G.melas).
Actuellement, une nouvelle phase du programme s’intéresse à de nouvelles espèces (phase 3S2).
Les expériences sont du même type que celles de Bahamas BRS et sont réalisées en
Norvège (Kvadsheim et al., 2007 ; 2009) ou en Atlantique nord. Les espèces sont taggées puis
suivies par un bateau d’observation, un autre navire muni de sonar opérant des approches avec
les deux types de sonar (LF et BF). La récupération des données des tags, couplées avec les
observations des deux bateaux permet de décrire les effets des sonars sur les cétacés, et de les
interpréter. La première grande phase du programme 3S1 s’est déroulée de 2006 à 2009.
Espèce Année MFAS LFAS Silence LF-DS Orque Type de réaction observée
Orcinus orca 2006 1 augmentation de vitesse
Orcinus orca x2 2006 1 interruption d’activité et évasion
Orcinus orca 2008 1 1 1 1 activité vocale – fuite panique
Orcinus orca x2 2009 1 1 1 1 accélération et évasion
Globicephala melas 2008 1 1 activité vocale et changt de dir.
G. melas x2 2008 1 1 activité vocale et fuite
Globicephala melas 2008 1 1 1 évasion lente
Globicephala melas 2008 1 1 1 1 interruption d’activité
G. melas x2 2009 1 1 1 1 1 interruption d’activité - évasion
Globicephala melas 2009 1 1 1 1 1 interruption d’activité - évasion
P. macrocephalus 2008 1 1 interruption d’activité
P. macrocephalus 2008 1 1 1 1 interruption d’activité - évasion
P. macrocephalus 2009 1 1 1 1 1 interruption d’activité
P. macrocephalus 2009 1 1 1 1 1 interruption d’activité - réaction
Tableau 17 : Expériences de transmission réalisées durant le programme 3S de 2006 à 2009
Quatorze expériences ont été conduites sur trois espèces (Oo, Gm, Pm), avec durant
chaque expérience des expositions multiples successives. Les expositions sont les suivantes:
- LFAS 1-2 kHz (13), MFAS 6-7 kHz (13) en mode « upsweep »32,
- LFAS en mode « downsweep » (7),
32 Downsweep : le son du sonar est modulé avec une fréquence qui diminue ; Upsweep, c’est l’inverse.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 73
- son naturel d'Orque (8),
- approche simple en mode silencieux (9),
Les résultats du programme 3S1 ne sont pas encore exploités finement mais Kvadsheim et
al. (2009) et Miller et al. (2009) ont relevé que les deux grands delphinidés peuvent avoir des
réactions très diverses ou très fortes selon les niveaux reçus et les fréquences : réponses vocales,
stratégies d'évasion complexes, fuite parfois avec panique. Les transmissions semblent aussi avoir
un effet sur les sondes de prédation des trois espèces.
Le programme se continue avec la phase 3S2, prévue pour les étés 2011-2012-2013, et a
priori axée sur des mysticètes (M. novaeangliae, B. acutorostrata) et sur un ziphiidé, l’Hyperoodon
(Hyperoodon ampulatus). Dans cette phase, les transmissions sonar sont uniquement à basse
fréquence, et il y aussi des play-back de cris d’Orque. Cette phase est toujours en cours, mais
deux rapports d’étape permettre de connaître la teneur des expériences réalisées (Kvadsheim et
al., 2011 ; 2012). Les expériences CEE qui ont réussi en 2011 et 2012 ont porté sur le Mégaptère,
et à une seule reprise sur un Rorqual de Minke (Tab.18).
Espèce Année LFAS Orque Bruit Silence Type de réaction observée
Megaptera novaeangliae 2011 6 3 3 2
Megaptera novaeangliae 2012 7 5 5
peu accentuée vis à vis dusonar, forte pour les orques
B. acutorostrata 2011 1 1 1
B. acutorostrata 2012
accélération et évasion pour lesonar
Tableau 18 : Expériences de transmission réalisées durant le programme 3S en 2011 et 2012
L’effet des transmissions sonar à basse fréquence est faible pour les Mégaptères et semble
forte pour le Rorqual de Minke. Cette différence reste à confirmer, car elle repose sur une seule
expérience pour le petit rorqual, néanmoins elle suggère que pour des espèces de mysticètes pas
très éloignées en terme phylogénique, la différence de réaction comportementale à un sonar peut
être très grande.
� Le programme SoCal
Le programme “Biological and Behavioral Studies of Marine Mammals in Southern
California” se déroule dans la zone d’exercice militaire au large de San Diego, dans laquelle se
rencontrent de nombreuses espèces de cétacés de tous genres. Le programme SoCal est prévu
pour se dérouler de 2010 à 2015, mettant en oeuvre des expériences de CEE avec des
transmissions de signaux de sonar et un réalisme croissant allant jusqu’à l’emploi de véritables
sonars avec des navires de l’US Navy. Les espèces visées sont en premier lieu le Ziphius, mais
aussi les delphinidés, et les mysticètes. A la différence du programme 3S, les transmissions de
sonar sont dans au milieu de la gamme des moyennes fréquences, vers 3500 Hz, ce qui
correspond aux sonars les plus puissants utilisés par l’US Navy.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 74
De nombreuses expériences ont été réalisées par SoCal depuis 2010, sur au moins 7
espèces dont trois mysticètes (Rorqual bleu B. musculus, Rorqual commun, Mégaptère), deux
ziphiidés (Ziphius et Bérardie Berardius bairdi), et un delphinidé, le Dauphin de Risso. D’autres
espèces ont été soumises aux bruits de sonar, mais sans avoir été taggées. Les sons qui sont
projetés sur les animaux sont de deux types : signal de sonar type SQS-53C, bruit blanc dans
l’octave centrée sur 3,5 kHz. Je résume ci-dessous les résultats obtenus, tels qu’ils sont exposés
dans Southall et al. (2011 ; 2012 ; 2013). D’autres résultats sur ces espèces ont été obtenus
durant SoCal13, mais il ne sont pas encore exposés33.
Espèce Année MFAS Noise Base Type de réaction observée
B. musculus 2010 max 160 pas de chgt d’activité ni d’évasion
B. musculus 2011 max 145 pas de chgt d’activité ni d’évasion
B. musculus 2011 max 135 modification d’activité avec évasion
B. musculus 2012 max 135 continuation de chasse profonde
B. musculus 2012 continuation de sonde de sub-surface
B. physalus 2010 max 160 pas de chgt d’activité ni d’évasion
B. physalus 2011 max 145 continuation de chasse profonde
B. physalus 2011 max 150 continuation de sonde de sub-surface
M. novaeangliae 2012 max 160 modification de l’activité sans évasion
P. macrocephalus 2010 max 150 pas de chgt d’activité ni d’évasion
P. macrocephalus 2010 max 145 pas de chgt d’activité ni d’évasion
Ziphius cavirostris 2010 max 135 interruption d’activité avec fuite
Ziphius cavirostris 2011 max 140 interruption d’activité
Berardius bairdii 2012 max 140 dérangement sans chgt d’activité
Grampus griseus 2010 max 135 pas de chgt d’activité ni d’évasion
Grampus griseus 2011 max 145 pas de chgt d’activité ni d’évasion
Grampus griseus 2012 max 135 pas de chgt d’activité ni d’évasion
Tableau 19 : Expériences de transmission réalisées durant le programme SOCAL de 2010 à 2012.
Les informations partielles données dans les rapports d’étude permettent d’avoir une idée
sur les effets des sonars à moyenne fréquence sur plusieurs espèces, y compris certains des
cétacés habituels du Sanctuaire Pelagos. Les expériences SoCal confirment que le Ziphius est
beaucoup plus sensible que les autres espèces aux transmissions à 3,5 kHz. Nous exploiterons
ces résultats plus en détail au §B.2.
Les trois principaux programmes de CEE ont donné des résultats récents (entre 2008 et
2013) très utiles pour notre problématique.
33 Une demande a été faite auprès de B. Southall en vue d’obtenir des éléments sur ces résultats (12/2013).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 75
o Effets des transmissions sonar sur l’audition des cétacés
Comme les autres sources sonores puissantes, les sonars peuvent provoquer des atteintes
à l’appareil auditif des cétacés, temporaires (TTS) ou définitives (PTS). Seules les pertes auditives
temporaires ont été étudiées, en bassin, les expériences portant jusqu’à présent sur trois
espèces : le Bélouga, le Grand dauphin et le Marsouin commun.
Dans un premier temps, les expériences ont consisté à rechercher le seuil de niveau sonore
reçu conduisant à une perte auditive mesurable, pour des sons de durée « unité » (1 seconde) et
de différentes fréquences (Schlundt et al., 2000). Dans un second temps, les recherches se sont
concentrées sur des fréquences usuelles de sonar tactique (comme par exemple 3 kHz), en
recherchant l’effet de la durée d’exposition sur la parte auditive temporaire obtenue (Finneran et
al., 2005 ; Kastelein et al., 2012 ; 2013). Très récemment, les recherches visent à obtenir des
modèles permettant de prévoir l’occurrence, l’amplitude et la durée des pertes auditives chez le
Grand dauphin, en fonction de la dose sonore efficace reçue (donc notion de durée d’exposition, et
de niveau sonore relatif). Il apparaît que l’espèce Tursiops truncatus est prise comme étalon pour
l’établissement d’une loi permettant de prédire les effets auditifs des transmissions de sonar
(Finneran et al., 2010 ; 2011 ; 2013). Nous indiquons ci-dessous les expériences réalisées en
bassin sur les pertes auditives provoquées sur les cétacés par des sons simples, analogues aux
sons de sonar, à différentes fréquences et sur trois espèces (Tab.20).
D’autres expériences similaires ont été conduites avec des signaux de type « bruit blanc par
octave » sur le Grand dauphin et le Marsouin commun, elles seront également prises en compte
dans notre analyse approfondie (§B.2).
Les signaux qui ont été utilisés pour ces expériences ont bien souvent un rapport direct avec
ceux des sonars (fréquence, durée), notamment pour les travaux les plus récents, après 2005.
Mais la finalité des expériences menées, notamment aux USA, n’est pas seulement de mesurer
directement les effets de ces signaux, elle est aussi de développer un modèle général de prévision
des effets auditifs des intensités sonores sur les cétacés, en utilisant le Grand dauphin comme
modèle de laboratoire (Finneran et al., 2010).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 76
Espèce Insonification seuil et fréquencedu TTS
Référence
grand dauphin Son pur 0.4 kHz 1 sec pas de TTS Schlundt et al., 2000
grand dauphin Son pur 3 kHz 1 sec 194 dB à 4,5 kHz Schlundt et al., 2000
grand dauphin Son pur 10 kHz 1 sec 192 dB à 15 kHz Schlundt et al., 2000
grand dauphin Son pur 20 kHz 1 sec 196 dB à 30 kHz Schlundt et al., 2000
grand dauphin Son pur 75 kHz 1 sec 182 dB à 100 kHz Schlundt et al., 2000
grand dauphin Son pur 3 kHz 1-8 sec 190 dB à 4,5 kHz Finneran et al., 2005
grand dauphin Son pur 20 kHz – 185 dB - 64 s TTS de 35 dB à 30 kHz Finneran et al., 2007
grand dauphin Séries de 3 chirps vers 3 kHz TTS à 203 dB à 5,6 kHz Mooney et al., 2009
grand dauphin Séries de 4 sons de 16 s à 3 kHz TTS à dB à kHz Finneran et al., 2010
grand dauphin idem pour 3 et 20 kHz TTS à dB à kHz Finneran & Schlundt, 2010
grand dauphin Son de 16 s à 3 et 80 kHz TTS à dB à kHz Finneran & Schlundt, 2013
bélouga Son pur 0.4 kHz 1 sec pas de TTS Schlundt et al., 2000
bélouga Son pur 3 kHz 1 sec 195 dB à 4,5 kHz Schlundt et al., 2000
bélouga Son pur 10kHz 1 sec 192 dB à 15 kHz Schlundt et al., 2000
bélouga Son pur 20kHz 1 sec 200 dB à 40 kHz Schlundt et al., 2000
marsouincommun
Son pur à 1.5 kHz de 60 min 154 dB à 1,5 et 2 kHz Kastelein et al., 2013
Tableau 20 : Synthèse des insonifications et TTS des dauphins et des bélougas en sons purs.Les pertes de sensibilité obtenues (TTS) sont dites « masquées » car les expériences se sont dérouléesavec un bruit blanc artificiel (masquant).
• Principes de mitigation applicables pour le risque « sonar »
Comme pour les prospections sismiques, la mitigation doit permettre d’éliminer les impacts
sévères (mort, lésion auditive) des sonars, et de minimiser ceux découlant de l’évasion d’habitat.
Les sources sonar sont de très forte intensité et audibles par différentes espèces en fonction de
leurs fréquences (LF, MF, THF). Il y a des différences importantes entre les sources sonar et les
sismiques au niveau de leur fréquence, de leur directivité, de leur mouvement, de la durée des
transmissions. En ce qui concerne la directivité , les sonars sont en général optimisés pour obtenir
une grande portée horizontale, ce qui les rend d’autant plus nuisibles pour les cétacés à des
distances de plusieurs kilomètres. En ce qui concerne le mouvement, la vitesse de la source est
variable mais peut souvent dépasser les 10 noeuds, c’est-à-dire se situer certainement au-delà
des capacités d’évasion soutenue de la plupart des espèces de cétacés. En ce qui concerne la
durée, une série de signaux peut excéder une demi-journée (cas documentés notamment par l’US
Navy), ou parfois ne comprendre que quelques minutes de transmission. Les transmissions les
plus longues sont certainement celles du sonar de surveillance à longue portée SurTaSS-LFA,
avec des séries de plusieurs semaines, mais cet engin demeure un cas particulier. Typiquement,

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 77
une série de transmissions de sonar est beaucoup plus courte qu’une campagne sismique. En
revanche, la soudaineté du début des transmissions constitue un problème aggravant les effets
des sonar, surtout lorsque la doctrine d’emploi ne prévoit pas de « ramp-up ».
Pour faire face au premier type de risque, il est nécessaire de disposer de moyens de
détection visuelle et acoustique garantissant qu’aucun cétacé ne se trouve dans la zone critique où
les niveaux sonores peuvent provoquer des lésions ou des incapacités auditives. Cette condition
doit être remplie aussi bien au démarrage de la source que lors de son mouvement. A l’instar des
campagnes sismiques, l’impossibilité de détecter certains cétacés la nuit ou par mauvais temps,
ainsi que l’extrême difficulté de détecter les ziphiidés, constituent un obstacle sévère à la mitigation
du premier type de risque. Le contrôle de ce risque grave peut être facilité par la mitigation
« stratégique » : le choix d’une zone où les espèces les plus vulnérables sont absentes.
Le risque lié à la désertion de l’habitat est moins accentué que pour les sismiques, en raison
des durées typiquement assez courtes des transmissions sonar.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 78
f) Le trafic maritime
Le bruit ambiant en milieu sous-marin subit une augmentation générale en proportion du
développement du trafic maritime, en premier lieu du trafic marchand. Les bruits issus de la
circulation maritime ont subi une évolution récente, avec le développement de la navigation de
loisir à moteur, expression qui désigne toutes sortes d’engins, depuis le jet-ski jusqu’au yacht de
150 m de long. L’intensité de cette activité de loisir a largement dépassé celle de la navigation
commerciale dans beaucoup de régions où se trouvent des cétacés, comme par exemple certains
sites du Sanctuaire Pelagos. Or les caractéristiques acoustiques de la navigation de loisir diffèrent
totalement de celles de la navigation commerciale classique (Erbe 2002, 2013). Si les niveaux
sonores sont en général moindres en raison de la taille réduite des unités, les régimes rapides des
hélices et leur faible immersion ont tendance à créer des phénomènes de cavitation et de nuages
de bulles qui provoquent du bruit à moyenne fréquence, donc avec un dérangement potentiel de
l’activité acoustique des odontocètes.
• Nature des ondes sonores du trafic maritime
Trafic commercial classique
Malgré tout, le bruit des grands navires est demeuré une composante dominante du bruit
ambiant à basse fréquence, et les croissances de cette catégorie de bruit ont été documentées
dans plusieurs régions de la planète où des augmentations importantes de trafic maritime ont eu
lieu (McDonald et al., 2006 ; Southall & Scholik-Schlomer, 2008 ; Erbe et al., 2012). Le nombre
total de navires commerciaux a doublé entre 1965 et 2003 et les projections prévoient que le
tonnage transporté par ces grands navires de fret va encore doubler ou tripler d'ici à 2025. Par
exemple, de 2001 à 2005, le nombre de tankers et de porte-containers dans le monde a augmenté
respectivement de 37 % et de 30 % (Southall & Scholik-Schlomer, 2008). Il y a donc une
augmentation prévisible de la contribution de ce type de bruit au niveau sonore ambiant en milieu
marin. Le bruit rayonné par les navires commerciaux a augmenté d'environ 3 dB par décade
depuis 1950 (McDonald et al., 2006 ; Southall & Scholik-Schlomer, 2008).
Hatch et al. (2008) ont analysé les niveaux sonores de ce type de trafic maritime dans le
sanctuaire marin de Stellwagen Bank (Atlantique NW, USA), avec une moyenne d'environ 10
passages de grands navires chaque jour. Avec un ensemble de 9 enregistreurs autonomes, ils ont
mesuré un niveau sonore basse fréquence de 119,5 dB en moyenne (en valeur efficace, de 10 à
1000Hz – Fig.29). Ils ont mesuré le niveau de source des navires, qui atteint 186 dB pour un
pétrolier dans la bande de 71 à 141 Hz (TBF). Ce niveau de bruit impacte la communication de
Baleine franche de l'Atlantique nord, une espèce menacée d'extinction. Hatch et al. (2008)
observent que l'empreinte acoustique du trafic de grands navires dépasse sur une très grande
surface le seuil de dérangement fixé par le Marine Mammals Protection Act.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 79
La contribution du trafic océanique au bruit ambiant est parfois plus importante dans une
zone de talus que dans une zone néritique, comme au large de la Californie, sur les sites de Eel
Point et San Nicolas, où l'on observe une différence de plus de 15 dB (Southall & Scholik-
Schlomer, 2008). Le bruit ambiant dans les zones peu profondes peut varier dans des proportions
très importantes, dans une fourchette de 40 dB, selon le site considéré, même dans l'hémisphère
nord, beaucoup plus fréquenté que l'hémisphère sud.
Figure 29 : Niveaux sonores moyens en tiers d'octave reçus sur 9 enregistreurs autonomes enavril et mai 2006 sur le sanctuaire marin de Stellwagen Bank (d'après Hatch et al., 2008).
Malgré l'accroissement du tonnage commercial, il y a eu peu d'évolutions sur le spectre des
bruits rayonnés par les grands navires de fret, ceux-ci provenant en majorité des machines pour
les très basses fréquences, et des hélices pour les fréquences plus hautes jusqu'à 1000 Hz. Par
contre, pour les navires rapides de moindre tonnage dont les caractéristiques spectrales sont
différentes : le bruit rayonné par ces engins est très important en moyenne fréquence. Nous avons
pu observer et enregistrer ces navires rapides en Méditerranée. Ces caractéristiques « moyenne
fréquence » propres aux "fast-ferries" ou aux grands navires rapides impliquent que leurs
nuisances acoustiques s'étendent au domaine de communication des odontocètes, voire au
domaine de l'écholocalisation de certains d'entre eux (cachalots, ziphius).
Trafic de loisir
Contrairement au trafic marchand habituel, l’énergie sonore produite par les embarcations
motorisées de loisir empiète largement sur les moyennes fréquences, un domaine acoustique
important pour beaucoup d’odontocètes (communication et écholocalisation). La propulsion de ces
engins diffère de celle de la plupart des navires marchands, en ce qui concerne le régime des
moteurs, leur type (hélices carénées, hélices rapides, « turbines »). Elle est souvent génératrice de
cavitation généralisée, synonyme de bruit à bande large et moyenne fréquence. Si toutes les
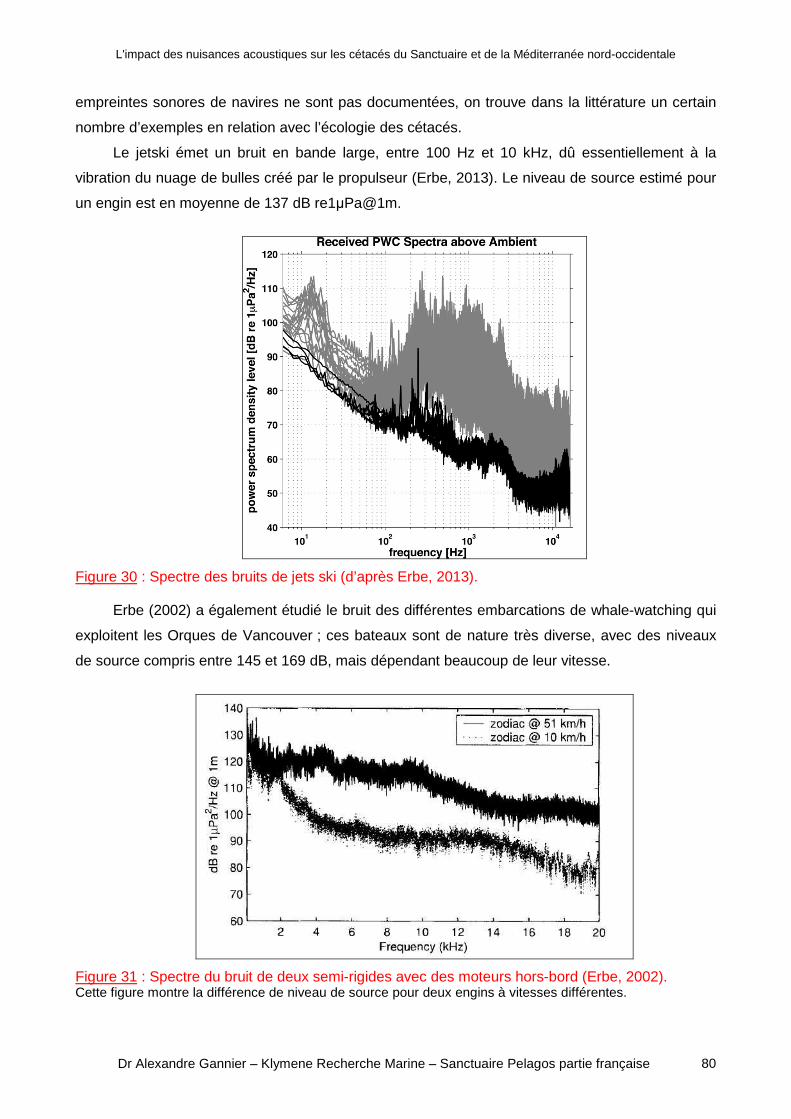
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 80
empreintes sonores de navires ne sont pas documentées, on trouve dans la littérature un certain
nombre d’exemples en relation avec l’écologie des cétacés.
Le jetski émet un bruit en bande large, entre 100 Hz et 10 kHz, dû essentiellement à la
vibration du nuage de bulles créé par le propulseur (Erbe, 2013). Le niveau de source estimé pour
un engin est en moyenne de 137 dB re1µPa@1m.
Figure 30 : Spectre des bruits de jets ski (d’après Erbe, 2013).
Erbe (2002) a également étudié le bruit des différentes embarcations de whale-watching qui
exploitent les Orques de Vancouver ; ces bateaux sont de nature très diverse, avec des niveaux
de source compris entre 145 et 169 dB, mais dépendant beaucoup de leur vitesse.
Figure 31 : Spectre du bruit de deux semi-rigides avec des moteurs hors-bord (Erbe, 2002).Cette figure montre la différence de niveau de source pour deux engins à vitesses différentes.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 81
Il n’y a pas de résultats équivalents pour tous les types de bateaux à moteur rencontrés en
Méditerranée. Les enregistrements que j’ai réalisés de manière opportuniste confirment cette
augmentation du bruit à moyenne fréquence en zone péri-côtière, liée au trafic intense
d’embarcations rapides de taille comprise entre 5 et 30 mètres. L’ambiance acoustique qui règne
par exemple dans la zone des 5 milles au large d’Antibes semble être défavorable à la présence
de dauphins ou de cachalots.
• Effet des bruits du trafic maritime sur les cétacés
L’augmentation du bruit du trafic maritime se produit en premier lieu autour des grands
centres de population, où se trouvent les noeuds portuaires vers lesquels convergent les lignes de
navigation, et où se développent les activités de loisirs motorisés (voir par exemple Erbe et al.,
2012). Les bruits de bateaux ont été reconnus récemment comme des nuisances importantes pour
les cétacés, entraînant des réactions significatives dans les cas où ils dépassent un certain seuil,
comme l'augmentation du taux de vocalisations chez les Grands dauphins (Buckstaff, 2004), ou de
leur niveau de source chez les Orques à Vancouver (Erbe, 2002; Holt et al., 2009), ou une
augmentation de leur stress physiologique (Baleine franche, Rolland et al., 2012). De plus, dans
nombre de cas étudiés, l’augmentation du niveau de bruit sous-marin est due à l’exploitation
commerciale des cétacés (whale-watching).
En cas de masquage de leur communication, la réponse des odontocètes est d'augmenter le
niveau sonore de leurs propres émissions. Holt et al. (2009) l’ont montré pour les orques de
Vancouver qui sont soumises quotidiennement sur leur habitat préférentiel au bruit des
embarcations de whale-watching : en présence d'un bruit de fond dû aux bateaux dans la bande
des 1-40 kHz, les orques ont émis des cris d'un niveau de source valant entre 133 et 174 dB, en
corrélation très nette avec le niveau de bruit ambiant. Ce type de réaction au masquage, appelé
effet Lombard, a aussi été démontré pour les bélougas du Saint-Laurent (Scheifele et al., 2005),
une population menacée soumise à de nombreuses nuisances acoustiques (McQuinn et al., 2011).
Il paraît évident que ce type de réponse au masquage est limité par la capacité des cétacés
à élever le niveau sonore de leurs communications. Ainsi, malgré le fort niveau de bruit, les orques
continuent à fréquenter cet habitat critique en ce qui concerne leur alimentation à base de saumon,
tant que leur capacité physique leur permet de communiquer. Jensen et al. (2009) ont déterminé
que les rayons de communication du Grand dauphin (Australie) et du Globicéphale tropical
(Canaries) pouvaient être diminués de 26 % et 58 %, respectivement, à cause du trafic même pour
des bateaux naviguant lentement.
Il existe aussi des cas bien documentés de variation du comportement de chasse du Ziphius
en mer Ligure lors du passage d'un navire bruyant: (Aguilar Soto et al., 2006), et de Mésoplodons
de Blainville (M. densirostris) aux Bahamas (Pirotta et al., 2012). Dans le premier cas, un individu
taggé a interrompu son activité acoustique et sa chasse lors du passage d’un navire très bruyant
(niveau reçu à bande large atteignant 136 dB). Le bruit par tiers d’octave excédait alors de 20 dB

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 82
re1µPa le bruit ambiant enregistré auparavant (valeur de 125 dB entre 1 et 10 kHz) . La sonde en
question (la n°4) a duré 42 minutes, soit 15 minute s de moins que la moyenne des sept autres
sondes, bien qu'elle se soit déroulée à une profondeur égale (Aguilar Soto et al., 2006). Le nombre
de "buzzes" de capture de proies a été très inférieur à celui des autres sondes (Fig.32), car le
temps de chasse a fortement diminué pendant le passage du navire bruyant, avec 17 min contre
34 min pour les autres sondes. Les auteurs expliquent que la portée du biosonar du ziphius
pourrait avoir été réduite de 42 %. Ce phénomène s’est produit alors que l’insonification du ziphius
ne concernait pas, pour l’essentiel, la bande de fréquence utilisée par l’animal pour son
écholocalisation, au-delà de 20 kHz, même s’il y avait aussi une hausse significative du bruit dans
cette bande. On remarque que la sonde suivante (n°5 ) a été également de durée assez courte,
avec un nombre apparent de captures plus faible.
Figure 32 : Effet du bruit d’un bateau sur la sonde d’un Ziphius (Aguilar Soto et al., 2006).
Niveaux de bruit en moyenne et très haute fréquence et quantité de sons de captures enregistrés lors de 8sondes successives, comprenant le passage d'un bateau très bruyant.
Il apparaît donc clairement que les bruits du trafic maritime affectent significativement les
capacités de communication des cétacés (mysticètes, delphinidés, ziphiidés), et que de plus ils
peuvent diminuer les facultés d’écholocalisation des odontocètes. Par conséquent, lorsque ces
bruits sont forts et pérennes ils peuvent dégrader la qualité environnementale d’un milieu marin, et
aboutir à la désertion d’un habitat qui était auparavant favorable à une espèce.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 83
En raison de l’extraordinaire croissance du trafic motorisé (commercial et de loisir) dans la
bande la plus côtière (celle des 10 milles) du Sanctuaire Pelagos, et de l’augmentation des
nuisances sonores correspondantes, on doit se poser la question de la disparition éventuelle de
certains groupes côtiers de dauphins.
La directive européenne de 2008 et un document de travail34 fixent des limites de niveau
sonore ambiant pour les bandes de basse fréquence, et détermine un cadre méthodologique pour
évaluer la quantité d’énergie sonore introduite dans le milieu marin35.
• Principes de mitigation applicables pour le risque « bruit du trafic maritime »
Les autres sources de bruit prises en compte dans cette étude émanent d’activités
industrielles ou militaires, dont les caractéristiques générales sont : un nombre réduit de sources,
localisées dans l’espace, une forte puissance sonore unitaire des sources, une signature réduite
dans le domaine de fréquence, une répartition dans le temps irrégulière. En ce qui concerne le
bruit du trafic maritime, on a au contraire : une multiplicité de sources de faible (130 dB
re1µPa@1m) à moyenne puissance (190 dB), avec des fréquences recouvrant tout le spectre de
la basse à la moyenne fréquence (0.1-10 kHz), réparties sur un très grand domaine spatial, avec
plusieurs régions de concentration, un grand étalement dans le temps chaque année de manière
pérenne (au moins 6 mois de saison chaude pour le trafic de loisir, de nos jours), et un cycle
jour/nuit accentué, pour le trafic de loisir.
La mitigation pourrait s’articuler autour de deux principes : identification des sources les plus
bruyantes, et action visant à diminuer leur impact, d’une part, identification des habitats critiques
les plus exposés au bruit, et action visant à diminuer l’intensité sonore localement.
34 Directive 2008/56/EC du 17 juin 2008 – Document de travail SEC(2011) 1255 du 14 octobre 2011.35 Ces limites sont un niveau de pression sonore de 100 dBrms pour les bandes 1/3 d’octave de 63 et 125Hz, en moyenne annuelle (Erbe et al., 2012).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 84
2) Synthèse des effets des nuisances sonores sur le s cétacés
Dans cette partie, on reprend les résultats publiés sur les effets provoqués par les fortes
intensités sonores sur les cétacés, en les organisant dans le sens d’une gravité décroissante à
l’échelle de l’individu36. Nous débutons l’examen par les cas où des fortes intensités sonores ont
entraîné la mort, directement, ou à la suite de lésions irrémédiables mais non immédiatement
létales à des organes vitaux. Ensuite, nous abordons les pertes de sensibilité auditive provoquées
par les fortes insonifications, ces effets étant mesurés lors d’expériences en bassin. Puis, on
examinera les effets comportementaux plus ou moins sévères provoqués par les sons
anthropiques, et mis en évidence lors d’expériences d’exposition contrôlée, en milieu naturel. Dans
une partie distincte, on résumera les effets comportementaux observés également en milieu
naturel, mais pour lesquels on n’a pas d’indication aussi précise des intensités sonores reçues.
Enfin, la synthèse de tous ces résultats sera exposée autour d’un tableau permettant de saisir la
cohérence d’une relation entre « sévérité des effets » et « intensité (ou dose) reçue ou perçue ».
a) Lésions traumatiques graves ou létales
C’est le cas de cétacés ayant subi des traumatismes graves ou létaux en liaison très
probable ou certaine avec des émissions sonores anthropiques. On observe parfois en mer,
pendant l’insonification, l’accident lui même, mais en général les traumatismes sont révélés par
l’autopsie de cadavres échoués à proximité d’un site de pollution sonore aigüe, et peu après que
les nuisances sonores se soient produites. Les lésions traumatiques létales comprennent aussi les
baisses de sensibilité auditive permanentes (PTS) : dans un monde aquatique où la faculté
auditive est d’importance primordiale, la surdité partielle (c’est-à-dire concernant un domaine de
fréquence particulier) est une infirmité qui entame le pronostic vital d’un individu.
• Lésion traumatique immédiatement ou rapidement léta le
Ce type d’accident est rarement constaté en pleine mer, pendant une transmission sonore.
Un accident s’est produit en mars 2009 lors d’une prospection sismique 3D (donc de forte
puissance), durant laquelle un Dauphin tacheté pantropical (S. attenuata) s’est immobilisé à
l’approche des canons sismiques (distance de 600 m), tentant de garder sa tête hors de l’eau, puis
coulant, avant que les canons pneumatiques (toujours actifs) ne passent à proximité de l’endroit où
il avait disparu (Gray & Van Waerebeek, 2011). Je conclus que cet événement illustre le cas d’un
dauphin qui ne s’est pas évadé à l’approche de la forte intensité sonore, a tenté d’y échapper en
sortant sa tête, puis a été tué par l’énergie sonore alors qu’il était sous l’eau. J’observe que
36 En effet, c’est au niveau de l’individu qu’il est plus facile d’organiser cet examen, car il faudrait de trèslongs développements pour établir si la surdité partielle d’un individu est un dommage plus sévère que lestress physiologique d’une population qui a été déplacée hors d’une zone propice à son alimentation.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 85
l’opération de mitigation du risque avec observateurs dédiés à bord n’a pas évité cet accident. On
retiendra qu’un dauphin à environ 500 m d’un réseau d’airgun a été conduit à la mort.
Dans de nombreux cas, la lésion traumatique est constatée après coup, lors de l’échouage
du cétacé vivant ou mort. Ainsi, Richardson et al. (1995) signalent le cas d'une explosion au
Canada, avec 5 tonnes d'explosifs, qui a semble-t-il abouti à la mort de mégaptères : lors de la
séquence d'explosions, ceux-ci n'ont pas été observés réagissant au bruit (par exemple en
s'éloignant), mais deux individus ont été retrouvés morts le lendemain avec des fractures (Ketten
et al., 1993). Ketten et al. (1993) ont mis en évidence les fractures et lésions de l'oreille moyenne
et interne, donnant substance à des cas moins bien documentés auparavant comme celui de
plusieurs ziphius aux Antilles, échoués après des explosions d’origine militaire (Van Bree &
Kristensen, 1974). De même, Wang & Yang (2006) rapportent l'échouage au sud de Taïwan d'un
mésoplodon (M. gingkodens) pendant des manoeuvres navales au large, et cet individu, mort,
souffrait de fractures des os les plus fins du crâne, y compris au niveau de la bulle tympanique.
Des lésions hémorragiques, ont été constatées en différents endroits de la tête et de la cage
thoracique. Dans la même période, plusieurs groupes de cétacés se sont échoués, y compris un
groupe de globicéphales vivants. A Taïwan, Yang et al. (2008) ont signalé deux autres séries
d'échouages suspects survenus en 2005, y compris sur des mésoplodons et d'autres ziphiidés,
des cachalots nains, des globicéphales et des orques pygmées, d'autres dauphins. Dans les cas
où des examens post-mortem ont été effectués, on a constaté des hémorragies sévères,
notamment au niveau du melon. A l'époque de ces échouages massifs et atypiques, il y avait une
forte activité militaire dans les eaux voisines de Taïwan, mais d’autres causes possibles n'ont pas
été écartées.
Les conséquences traumatiques de ces explosions ne sont pas une surprise, puisqu’elles
ont été bien étudiées pour protéger les humains. Pour les explosions chimiques, le risque pris en
compte pour les mammifères terrestres est aussi une lésion sévère au niveau des organes autres
qu’auditifs, en rapport avec l'intensité de l'onde de pression générée par l'explosion. Des
expériences ont été réalisées avec des mammifères terrestres submergés, pour lesquels les
charges provoquant des lésions ont été déterminées (Richardson et al., 1995) : la charge
nécessaire pour provoquer la mortalité augmenterait de manière régulière avec la masse des
animaux. Goertner (1982 in Richardson et al., 1995) a extrapolé ces données et les a couplées à
des modèles de propagation pour fournir des modèles simplifiés de distance de sécurité à
appliquer aux mammifères marins pour éviter les lésions légères aux poumons ou aux intestins.
Par exemple, pour une charge de 545 kg explosant à 38 m d'immersion, cela donne
approximativement une distance de sécurité de 1200 m pour un bébé dauphin de 10 kg, de 800 m
pour un dauphin de 100 kg, de 600 m pour un globicéphale de 1 tonne, de 300 m pour une baleine
de 10 tonnes.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 86
Young (1991, in Richardson et al., 1995) donne des valeurs-guide simplifiées à partir des
résultats précédents de rayons de sécurité (Rsécu), pour des charges de masse w avec une
profondeur d’explosion de 30 m. Ces valeurs sont à interpréter en fonction des conditions locales
de topographie et de bathymétrie (R en m, w en kg) :
- jeune dauphin : Rsécu = 220 w 0.28 800 m pour 100 kg d’explosifs- dauphin adulte : Rsécu = 165 w 0.28 600 m pour 100 kg d’explosifs- cétacé de 6 mètres : Rsécu = 124 w 0.28 450 m pour 100 kg d’explosifs
Par ailleurs, Gaspin (1983, in Richardson et al., 1995) a déduit d’autres niveaux de sécurité
d'après des expériences sur des humains, et les distances trouvées sont plus importantes que
celles extrapolées pour les dauphins d’après les expériences sur les animaux terrestres.
- humain au large : Rsécu = 1335 w 0.18 3050 m pour 100 kg d’explosifs- humain faible fond, 9m : Rsécu = 608 w 0.18 1400 m pour 100 kg d’explosifs
En fonction de l’état des connaissances scientifiques exposées ci-dessus, il n’est pas
étonnant que les distances préconisées pratiquement pour les sources impulsives explosives
soient majorées du côté de la sécurité. Le NATO Undersea Research Center donne l'exemple de
la source SUS 61/82 dont la charge explosive vaut 0,8 kg : des distances de sécurité de 3000 m
sont préconisées pour les plongeurs, et de 2000 m pour les cétacés (NURC, 2006). Les distances
préconisées par les règles empiriques ci-dessus sont ainsi fortement majorées par le NURC pour
les grands cétacés et pour les dauphins juvéniles, si on se rapporte à la faible charge d’explosifs
(moins d’un kg) prise en compte dans le document.
Si l’on se réfère aux divers règlements, aux événements publiés, aux tables, il semble
qu’une explosion unique, de nature chimique ou pneumatique, puisse provoquer une lésion sévère
si la valeur de pression reçue atteint 235 dB en valeur pic.
Un autre type d’accident a été mis en évidence, mais cette fois sur des échouages de
cétacés encore vivants, ou des cadavres très frais : il s’agit des embolies gazeuse et graisseuse
d’abord diagnostiquées par Jepson et al. (2003). Leur réalité n’est plus contestée : elles ont été
constatées à différentes reprises, avec des gravités diverses. Fernandez et al. (2005) ont examiné
8 ziphius, un mésoplodon de Blainville et un mésoplodon de Gervais, sur les 14 ziphiidés échoués
vivants ou trouvés morts en mer le 24 septembre 2002, aux Canaries (Tab.15). Ils notent qu'aucun
pathogène n'a été identifié et que tous les individus examinés souffraient d'hémorragies et de
congestions sévères et diffuses au niveau des tissus graisseux mandibulaires, de l'oreille, du
cerveau et des reins. Des lésions associées à des bulles de gaz et des embolies graisseuses37
sont observées au niveau des vaisseaux sanguins et des organes vitaux. Les embolies
graisseuses sont des processus secondaires associés aux syndromes de décompression sévères,
les graisses des moelles osseuses étant des sites privilégiés de stockage de l'azote sursaturé. Les
embolies graisseuses ne sont pas réversibles, au contraire des bulles d'azote apparues au cours
37 Largage de graisses dans la circulation sanguine

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 87
d'un accident de décompression qui peuvent être résorbées par une recompression. L'accident de
décompression pourrait survenir de deux manières : soit directement par la stimulation acoustique
de la diffusion gazeuse, selon ce qui a été montré pour des niveaux sonores entre 190 et 210 dB
(Crum & Mao, 1996 ; Crum et al., 2005) en présence de sursaturation des tissus en azote, soit
sous l'effet d'un dérangement comportemental affectant le cycle de plongée des ziphiidés.
Ce type de lésion a été constaté également lors de l’échouage multiple de Ziphius de janvier
2006 en mer d’Alboran (Arbelo et al., 2007) et peut-être lors de l’échouage multiple de Ziphius à
Corfou, en 2011. Aux Bahamas (accident de 2000), les examens post mortem ont été moins
poussés qu'aux Canaries, mais ont montré des traumatismes au niveau de la tête, peut-être
insuffisants pour provoquer directement la mort (Ketten, 2003). Les travaux de modélisation
acoustique et mécanique de la tête d'un ziphius montrent que les caractères anatomiques (tissus
graisseux, sinus) favorisent une forte transmission des intensités sonores du mandibule à l'oreille
interne, et dans une certaine mesure au niveau du melon (Soldevilla et al. 2005 ; Krysl et al.,
2006 ; Cranford et al., 2008). La conjugaison des graisses "stockeuses" d'azote et de forts niveaux
sonores font de la tête des ziphiidés, et des autres odontocètes, un lieu privilégié pour la formation
de bulles de gaz, et d'embolies.
Les modélisations les plus récentes de ce phénomène suggèrent que le risque d'une létalité
très rapide sous l'effet d’une insonification non-impulsive existe pour des niveaux sonores reçus
élevés (plus de 190 dB) et des tissus sursaturés en azote, correspondant à des distances de
l’ordre de la centaine de mètres pour les plus puissantes sources continues connues.
Pour les plongeurs extrêmes, c'est-à-dire les ziphiidés, les ondes sonores perturbent le cycle
de plongée, ce qui pourrait déclencher un accident lié au métabolisme de la plongée ou de sa
récupération. Ce cas d’accident « secondaire » appartient plutôt à la classe des dérangements
comportementaux, et sera traité dans le paragraphe correspondant.
• Lésions sévères létales à terme
Dans ce cas, la lésion entraîne une incapacité permanente du cétacé, qui conduit à sa mort
à moyen ou long terme, comme par exemple une lésion affectant ses capacités à percevoir son
environnement, ses proies ou ses prédateurs. Nous plaçons dans cette catégorie la perte de
sensibilité auditive permanente (PTS), qui survient lorsqu'une exposition sonore à un niveau
suffisant pour produire un TTS est prolongée, ou avec un niveau sonore encore plus élevé. Le
PTS peut être provoquée aussi bien par des sons impulsifs que par des sons stationnaires : après
un long temps d'exposition, ou soumis à un niveau extrêmement élevé, l'oreille perd définitivement
de la sensibilité pour une bande de fréquence un peu supérieure à celle de l’insonification, car les
cellules de la cochlée, dans l'oreille interne, ont subi des dommages irrémédiables. Pour les
humains, de nombreux résultats sur les pertes définitives, et règlements, proviennent de mesures
d’audiométrie réalisées a posteriori chez des travaiIleurs ayant été soumis à des bruits importants

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 88
au cours de leur carrière. Les PTS ont été également étudiées en laboratoire sur des petits
mammifères.
Les PTS sont associés à la destruction ou à la forte altération des cellules ciliées, et de
l'organe de Corti, situés tout au long de la cochlée. Le seuil sonore provoquant un PTS est un
paramètre peu accessible dans la littérature38, bien qu'il corresponde à une réalité physiologique
identifiée par certains auteurs, notamment sur des petits mammifères (Price, 1981 ; Roberto et al.,
1985 ; Saunders et al., 1985). L'apparition de pertes auditives est liée à deux phénomènes bien
distincts (Laroche et al., 1989) : d'une part, les fortes intensités sonores provoquent un épuisement
métabolique au niveau du processus bio-chimique qui permet la génération des signaux
électriques au sein de la cochlée (liquides lymphatiques), et d'autre part elles causent un
endommagement mécanique des cellules ciliées qui transmettent les vibrations dans l'oreille
interne, voire la destruction de ces cellules pour des sons impulsifs extrêmement forts (> 143 dBpic
chez l'humain - Price, 1981). Le second processus ne s'amortit pas rapidement car il nécessite une
réparation cellulaire des cellules ciliées, et peut-être un processus de remplacement de ces
cellules. Quand le dommage a été trop important, les zones les plus affectées de l'organe de Corti
restent dépourvues de cellules ciliées, c'est une perte définitive (PTS) pour les fréquences
concernées (Roberto et al., 1985). Les conditions d'insonification causant des PTS rapides sont
fréquemment des trains d'impulsions, car le réflexe d'amortissement par l'oreille moyenne, observé
chez les mammifères terrestres, n'a alors pas le temps d'être activé.
Figure 33 : Organe de Corti (section radiale de la cochlée) avec les cellules ciliées "hair cells"(Saunders et al., 1985)
La littérature scientifique suggère que les niveaux provoquant des forts TTS donnent une
base pour connaître les niveaux seuils de PTS (Price, 1981 ; Laroche et al. 1989 ; Melnick, 1991).
Pour trois espèces de mammifères (homme, chimpanzé, chinchilla) Melnick (1991) a mis en
38 L’éthique scientifique interdit de réaliser des expérimentations de ce type, ou du moins de les publier.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 89
évidence l'existence d'un seuil de niveau sonore correspondant à une perte auditive de 8-10 dB,
niveau au-delà duquel la progression des TTS est très rapide. Ces résultats rejoignent ceux de
Finneran et al. (2007) pour les dauphins, avec des TTS supérieurs à 20 dB obtenus lors
d'expositions de 64 s à 20 kHz et 186 dB reçu, ou ceux de Lucke et al. (2009) pour de très forts
TTS dans le cas d'impulsions de canons pneumatiques. André et al. (2003) ont réalisé des
expériences audiométriques sur un Dauphin bleu et blanc, qui démontraient que ce dauphin était
atteint de surdité permanente, bien que la cause de cette infirmité n’ait pas été identifiée.
En fonction de tout ceci, je propose qu’un niveau sonore égal au seuil d'apparition d’un TTS
significatif (6 dB de perte auditive) augmenté de 15 dB, peut provoquer une perte auditive
persistante de 20 dB, laquelle constitue un bon critère pour définir le risque de PTS :
seuil PTS = seuil TTS + 15 dB
Nous allons voir comment le seuil de TTS est obtenu, et en fonction de quelle métrique on
peut l’exprimer (§B.2.b).
D’autres types de lésions dues à des insonifications excessives peuvent entraîner la mort à
moyen terme : on pense à des hémorragies importantes, ou à des embolies gazeuses ayant des
séquelles, les traumatismes pouvant affecter la capacité des cétacés à plonger, et donc à se
nourrir.
b) Effets auditifs mesurés lors d’expériences en bassin
Les effets auditifs chez les mammifères, y compris les humains, se mesurent lors
d’expériences au cours desquelles on soumet les sujets à des insonifications fortes et/ou
prolongées, et on mesure leur audiométrie avant et après ces épreuves. Les expériences
consistent à rechercher les conditions d’apparition d’une perte auditive temporaire, ou bien même
une perte définitive, mais dans ce cas pas pour les humains et, semble-t-il, pas pour des
mammifères supérieurs. Grâce à des expériences, il est établi que l’apparition des phénomènes de
fatigue auditive (TTS), au sein de l’oreille interne, est régie par des lois similaires pour les
mammifères placentaires ; les expériences menées sur les cétacés depuis 2000 environ, le
confirment.
Dans un premier temps, ces expériences menées à San Diego39, ont eu pour objectif
d’établir les bases du phénomène TTS chez les cétacés, avec des expériences sur le Grand
dauphin et le Bélouga (par ex. Finneran & Schlundt, 2000) et des insonifications impulsives ou en
son pur. Dans un second temps, des expériences analogues ont été conduites sur le Grand
dauphin, mais aussi en Europe sur le Marsouin commun, en utilisant des sons en rapport direct
avec des nuisances anthropiques, comme des signaux aux fréquences de sonars US ou
européens (Finneran et al., 2005 ; Kastelein et al., 2012 ; 2013), ou des sismiques pneumatiques
(Lucke et al., 2009). Dans un troisième temps, actuellement , les expériences s’orientent vers
39 Au centre U.S. Navy Marine Mammal Program du Space and Naval Warfare Systems Center.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 90
l’obtention de points expérimentaux pour valider des modèles de prévisions des TTS (Finneran et
al., 2010).
La synthèse de l’ensemble de ces résultats pour la proposition d’un critère de prévision des
impacts est au coeur de notre étude. Pour déterminer les seuils de risque d’apparition des pertes
auditives temporaires (TTS), notre approche est la suivante :
(1) considérer l’ensemble des résultats expérimentaux disponibles sur le Grand dauphin pour
déterminer les contours d’un critère de prévision applicable à tous les cétacés
(2) examiner les résultats disponibles sur le Bélouga, le Marsouin commun et le Marsouin du
Yangtsé, pour confirmer et préciser ce critère
(3) formaliser le critère de risque de TTS pour le rendre facilement applicable aux différentes
espèces de cétacés.
Au cours de cette synthèse, on examinera les résultats traitant des sources impulsives (type
airgun) et des sources non-impulsives avant de décider s’il vaut mieux proposer un critère
« universel » de prévision des TTS, applicable aux deux catégories de bruit, ou bien deux versions
du critère.
Les critères de prévision des TTS sont d’autant plus importants qu’ils servent de base à la
détermination des seuils de sécurité pour éviter l’apparition de pertes auditives définitives, ou PTS.
� Prise en compte de la durée d’exposition dans la métrique des insonifications
La durée d’exposition intervient fortement dans les phénomènes de fatigue auditive (TTS) ou
de surdité permanente (PTS). Pour la prendre en compte, on utilise une métrique qui intègre
l’énergie sonore au cours du temps d’exposition : la dose d’exposition sonore SEL. Cette métrique
permet de déterminer la dose aussi bien dans le cas d’un son non-impulsif intermittent, tel que lors
d’un cycle d’émissions de sonar, que lors d’un train d’impulsions (sismique, battage de pieu).
L’exposition sonore, en Pa2.s est calculée en intégrant la pression sonore au carré au cours
du temps d’exposition (Southall et al., 2007):
E (t) = ∫0,T P2(t).dt
Le niveau d’exposition sonore (SEL) en est l’expression relative en décibels (dB re1 µPa2.s):
SEL = 10.log10 ∫0,T (P2(t).dt / p2ref) ]
L’expression permet d’exprimer les niveaux d’énergie reçus dans les cas de sons non-
impulsifs de durée variable et de trains d’impulsions, et de déterminer les niveaux de risque. A la
base, on utilise l’expression suivante :
SEL = 10.log10 [ Σ1,N ∫0,T (Pn2(t).dt / p2
ref) ]
où P est la pression sonore, N le nombre d’expositions au son, et pref la pression de
référence (1 µPa). La sommation sur n indique que pour n expositions identiques, la dose
d’exposition augmente en fonction du log10 (n) :

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 91
SEL = 10.log10 (n) + 10.log10 [ ∫0,T (P2(t).dt / p2ref) ]
SELn = 10.log10 (n) + SEL1
où SEL1 est la dose (ou niveau) d’exposition pour un événement de transmission
donc SEL2 = SEL1 + 3 dB, SEL3 = SEL1 + 5 dB, SEL10 = SEL1 + 10 dB, SEL100 = SEL1 + 20 dB
La sommation est identique pour les deux catégories de sons (type « sonar » ou type
« airgun / battage de pieux »). La dose d’exposition totale est ensuite à comparer aux seuils de
TTS/PTS obtenus pour un événement de type « chirp » sonar de durée une seconde, ou bien pour
un événement de type « impulsion », exprimé en dBSEL (dB re re1 µPa2.s). Pour un son de type
sonar, les seuils sont exprimés pour une durée de 1 seconde (dBSEL 1 sec = dBrms), correspondant à
beaucoup d’expériences en bassin. Pour un son impulsif, le seuil de TTS/PTS est toujours exprimé
pour une seule impulsion : dans les expériences, on trouve la valeur seuil en dBpic-pic et en dBSEL.
Les résultats existants suggèrent que des impulsions de même SEL ont le même effet.
SELn < SELseuil(TTS) → SEL1 < SELseuil(TTS) - 10.log 10 (n)
La comparaison du seuil de TTS/PTS à la dose cumulée exprimée en SELn suppose qu’il n’y
a pas eu de relaxation de fatigue auditive entre deux expositions successives. Selon les résultats
récemment publiés, une relaxation apparaît surtout si les expositions sont séparées par de longs
intervalles (Finneran et al., 2010). Donc l’application d’un modèle simplifié basé sur les SEL se
situe du côté de la sécurité, pour les cétacés.
• Pertes de sensibilité auditive (TTS) observées chez le Grand dauphin
o Sons et bruits non impulsifs : bruit blanc
Un Grand dauphin T. truncatus a été soumis à des insonifications de type "bruit blanc" par
octave, dans les installations de Kanehoe Bay (Hawaii), où le bruit ambiant est fort (Au et al.,
1999 ; Nachtigall et al., 2003). Les seuils d'audition à la fréquence de 7,5 kHz ont été mesurés
avant et après une exposition de 30 minutes à un bruit blanc autour de l'octave 5-10 kHz, par
méthode comportementale. Des TTS de l'ordre de 12–18 dB ont été obtenus chez le dauphin pour
des niveaux sonores de 179 dB. Ce niveau était de 96 dB au dessus du seuil d'audition du
dauphin à 7,5 kHz. Nachtigall et al. (2003) ont rapporté plus en détails les tests d'insonification
réalisés par Au et al. (1999). Compte tenu des différentes positions occupées par le dauphin dans
l'enclos, les insonifications ont duré jusqu'à 55 min, les doses sonores ont été recalculées en
fonction de l'exposition réelle du dauphin. Mais ces expériences souffrent de plusieurs points
faibles méthodologiques : le TTS a été déterminé à la fréquence de 7,5 kHz, centrale par rapport à
l'insonification, alors que les TTS les plus forts ont lieu à une fréquence supérieure d'une demi-
octave à celle de l'insonification. De plus, les seuils d’audition post-exposition ont été déterminés
longtemps après l'insonification, donc les TTS ont été sous-estimés en valeur.
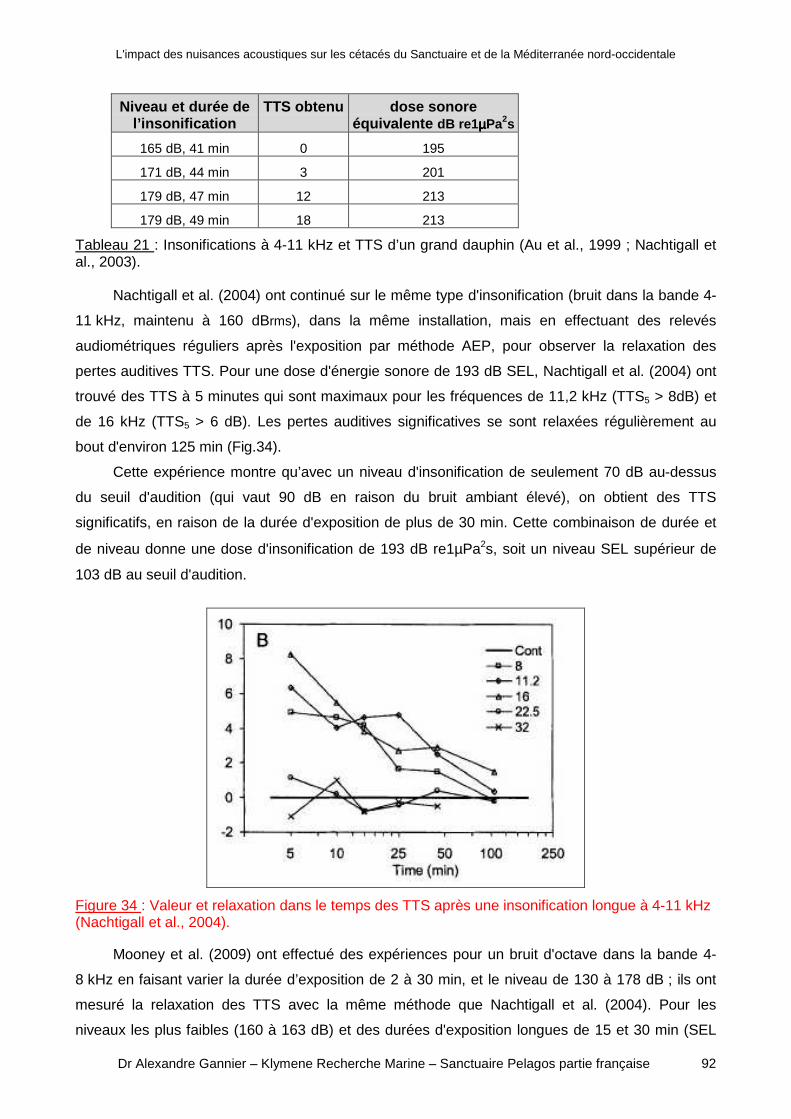
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 92
Niveau et durée del’insonification
TTS obtenu dose sonoreéquivalente dB re1 µµµµPa2s
165 dB, 41 min 0 195
171 dB, 44 min 3 201
179 dB, 47 min 12 213
179 dB, 49 min 18 213
Tableau 21 : Insonifications à 4-11 kHz et TTS d’un grand dauphin (Au et al., 1999 ; Nachtigall etal., 2003).
Nachtigall et al. (2004) ont continué sur le même type d'insonification (bruit dans la bande 4-
11 kHz, maintenu à 160 dBrms), dans la même installation, mais en effectuant des relevés
audiométriques réguliers après l'exposition par méthode AEP, pour observer la relaxation des
pertes auditives TTS. Pour une dose d'énergie sonore de 193 dB SEL, Nachtigall et al. (2004) ont
trouvé des TTS à 5 minutes qui sont maximaux pour les fréquences de 11,2 kHz (TTS5 > 8dB) et
de 16 kHz (TTS5 > 6 dB). Les pertes auditives significatives se sont relaxées régulièrement au
bout d'environ 125 min (Fig.34).
Cette expérience montre qu’avec un niveau d'insonification de seulement 70 dB au-dessus
du seuil d'audition (qui vaut 90 dB en raison du bruit ambiant élevé), on obtient des TTS
significatifs, en raison de la durée d'exposition de plus de 30 min. Cette combinaison de durée et
de niveau donne une dose d'insonification de 193 dB re1µPa2s, soit un niveau SEL supérieur de
103 dB au seuil d'audition.
Figure 34 : Valeur et relaxation dans le temps des TTS après une insonification longue à 4-11 kHz(Nachtigall et al., 2004).
Mooney et al. (2009) ont effectué des expériences pour un bruit d'octave dans la bande 4-
8 kHz en faisant varier la durée d’exposition de 2 à 30 min, et le niveau de 130 à 178 dB ; ils ont
mesuré la relaxation des TTS avec la même méthode que Nachtigall et al. (2004). Pour les
niveaux les plus faibles (160 à 163 dB) et des durées d'exposition longues de 15 et 30 min (SEL
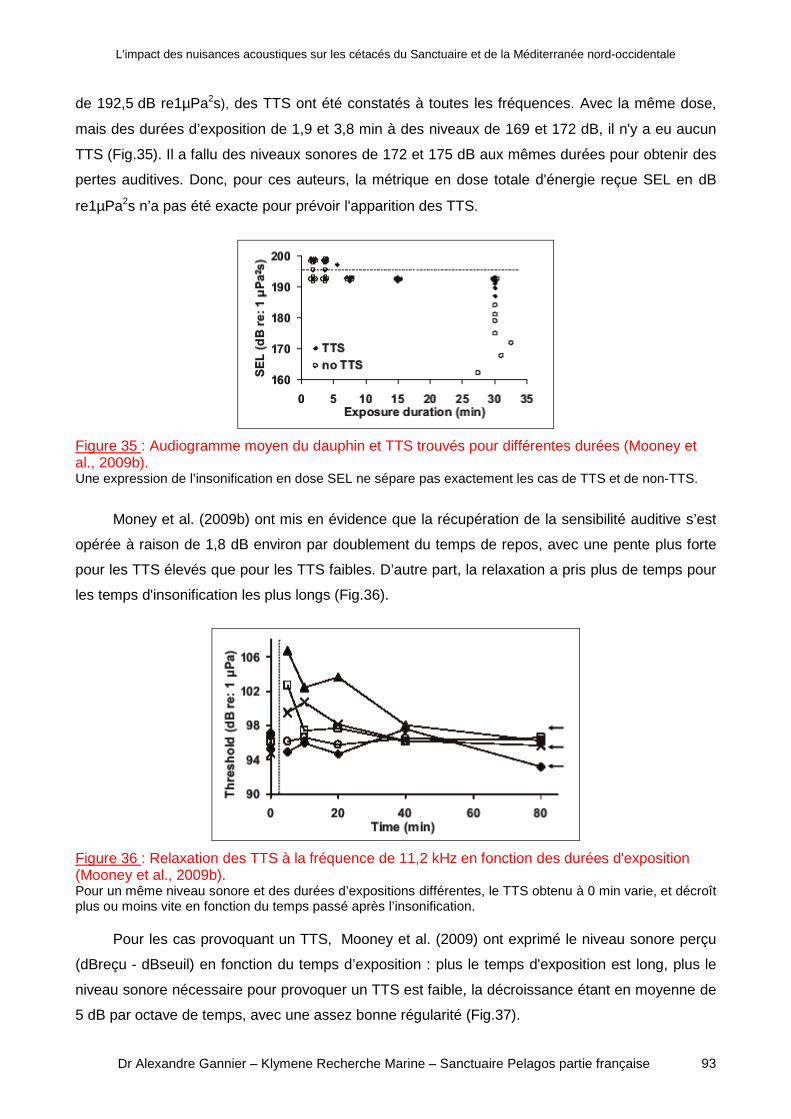
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 93
de 192,5 dB re1µPa2s), des TTS ont été constatés à toutes les fréquences. Avec la même dose,
mais des durées d’exposition de 1,9 et 3,8 min à des niveaux de 169 et 172 dB, il n'y a eu aucun
TTS (Fig.35). Il a fallu des niveaux sonores de 172 et 175 dB aux mêmes durées pour obtenir des
pertes auditives. Donc, pour ces auteurs, la métrique en dose totale d'énergie reçue SEL en dB
re1µPa2s n’a pas été exacte pour prévoir l'apparition des TTS.
Figure 35 : Audiogramme moyen du dauphin et TTS trouvés pour différentes durées (Mooney etal., 2009b).Une expression de l’insonification en dose SEL ne sépare pas exactement les cas de TTS et de non-TTS.
Money et al. (2009b) ont mis en évidence que la récupération de la sensibilité auditive s’est
opérée à raison de 1,8 dB environ par doublement du temps de repos, avec une pente plus forte
pour les TTS élevés que pour les TTS faibles. D’autre part, la relaxation a pris plus de temps pour
les temps d'insonification les plus longs (Fig.36).
Figure 36 : Relaxation des TTS à la fréquence de 11,2 kHz en fonction des durées d'exposition(Mooney et al., 2009b).Pour un même niveau sonore et des durées d’expositions différentes, le TTS obtenu à 0 min varie, et décroîtplus ou moins vite en fonction du temps passé après l’insonification.
Pour les cas provoquant un TTS, Mooney et al. (2009) ont exprimé le niveau sonore perçu
(dBreçu - dBseuil) en fonction du temps d’exposition : plus le temps d'exposition est long, plus le
niveau sonore nécessaire pour provoquer un TTS est faible, la décroissance étant en moyenne de
5 dB par octave de temps, avec une assez bonne régularité (Fig.37).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 94
Figure 37 : Niveau d'exposition net causant un TTS en fonction des durées d'expositions (Mooneyet al., 2009b).
Ces résultats sur les pertes auditives provoquées par des insonifications en bruit blanc
montrent que des niveaux sonores relativement faibles provoquent des TTS lorsque les durées
d’exposition deviennent très longues. Pour exprimer la valeur de l’insonification qui provoque un
TTS, la métrique en SEL (niveau d’exposition en dB re1µPa2s) fournit une indication utile, même si
elle n’est pas exacte. Dans le cas de TTS d’une valeur de 10 dB ou plus, les temps nécessaires
pour la relaxation de la perte auditive sont de l’ordre d’une heure.
o Sons et bruits non impulsifs : sons simples
Les expériences visant à déterminer les seuils d'apparition de TTS avec des sons
sinusoïdaux ou légèrement modulés ont été fréquemment réalisées sur le Grand dauphin
(Schlundt et al., 2000 ; Finneran et al., 2005 ; 2007 ; 2010a,b,c ; Finneran & Schlundt, 2013). Pour
certaines de ces expériences, les tests auditifs ont été comportementaux, alors que pour d'autres,
la méthode électro-physiologique a été employée ; certains écarts entre les résultats peuvent
découler des méthodes choisies pour mesurer l'audition, avant et après les insonifications.
Schlundt et al. (2000) ont exposé 5 Grands dauphins (et 2 Bélougas) à des sons simples
d'une durée de 1 sec et à des fréquences de 400 Hz, 3, 10, 20, et 75 kHz. La mesure de seuils
auditifs s'est faite en présence de bruit de fond artificiel (afin de stabiliser les conditions de bruit
ambiant pendant toute la durée des expériences), avec une méthode comportementale et pour des
fréquences égales ou supérieures à celles des insonifications. Schlundt et al. (2000) ont ainsi
montré l'apparition de TTS chez ces odontocètes (Tab.22). En raison du bruit de masquage
artificiel, la perte auditive est appelée MTTS (voir p75).
Les niveaux nécessaires pour provoquer un MTTS significatif (défini comme une hausse de
seuil auditif ≥ 6 dB), ont été généralement compris entre 192 and 201 dB re1µPa, avec une
variabilité entre les dauphins. A 400 Hz, aucun des sujets n'a eu de perte auditive temporaire
jusqu'à la valeur maximale expérimentée, 193 dB re1µPa, à 3 kHz, les MTTS ont été très forts. Le
plus souvent, les MTTS ont eu lieu à des fréquences supérieures à celle de l’insonification.
Observons que les TTS ont été mesurés entre 2 et 18 min après les insonifications, ce qui induit
une imprécision et une sous-estimation, puisque pour des faibles MTTS, des durées de quelques
minutes suffisent pour qu'un dauphin ait récupéré ses facultés auditives.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 95
Fréquence Niveau reçu(dB rms)
Bruit ( dBre1µµµµPa2/Hz)
ntest
seuil d’apparition etvaleur du MTTS
400 Hz 179 à 193 95 2 non
à 194dB, 7dB @ 3 kHz
à 194dB, 16 dB @4,5 kHz3 kHz 160 à 202 67 2
à 194dB, 16 dB @4,5 kHz
3 kHz 180 à 201 90 2 non
10 kHz 180 à 197 63 2 à 192dB, 7 dB @5 kHz
20 kHz 160 à 197 75 2 à 193dB, 8 dB @40 kHz
20 kHz 180 à 201 75 2 non
20 kHz 178 à 202 63 2 à 196dB, 6 dB @30 kHz
75 kHz 160 à 194 70 2 à 182dB, 8 dB @100 kHz
Tableau 22 : Insonifications et MTTS pour des Grands dauphins exposés à des sons purs de 1 sde durée et de fréquence variable (d'après Schlundt et al., 2000).
Finneran et al. (2005) ont effectué le même type d'expérience sur deux Grands dauphins, en
gardant une unique fréquence d'insonification de 3 kHz, mais en faisant varier les durées
d’exposition de 1 à 8 s (les durées supérieures à 1 sec ont été testées avec un seul dauphin). Les
auteurs ont conduit ces expériences en ambiance non bruitée. Pour prendre en compte la durée
dans l'insonification, les auteurs ont utilisé la mesure en dose équivalente en SEL, exprimée en dB
re1µPa2s, un doublement de durée conduisant à un SEL augmenté de 3 dB. Le dauphin sujet de
ces expériences avait un seuil auditif d'environ 90 dB à 3 kHz (Finneran et al., 2005).
Les expériences 1 et 2 ont consisté en des insonifications de 1 s, et des vérifications de TTS
respectivement à 4,5 et 3 kHz. Les niveaux imposés ont varié de 100 à 200 dB. L'expérience 3 a
consisté en des insonifications de durée variable (2 à 8 s) et des niveaux imposés de 140 à 195
dB. Après chaque insonification, les tests auditifs comportementaux ont été effectués au bout de 4
min (TTS4) et de 10 min (TTS10).
Les niveaux sonores provoquant des TTS ont été de 197 dB pour 1 s, de plus de 190 dB
pour 2 s, de plus de 184 dB pour 4 s, et de plus 190 dB pour 8 s. Finneran et al. (2005) déduisent
que les doses sonores entraînant un TTS significatif sont de 195 dBSEL, mais on s’aperçoit sur les
graphes que le TTS commence à apparaître à 190 dBSEL (Fig.38).
Niveau et duréed’exposition
Fréquence dutest auditif
seuil de TTSobtenu en dB
seuil de TTSobtenu en SEL
seuil de TTS enSEL effectif
100 à 200 dB, 1 sec 4,5 kHz 197 dB 197 dB env. 111 dB SEL*
180 à 200 dB, 1 sec 3 kHz - - -
144 à 195 dB, 2 sec 4,5 kHz 190 - 195 dB > 190 dB env. 100 dB SEL°
144 à 195 dB, 4 sec 4,5 kHz 184 - 195 dB > 190 dB env. 100 dB SEL°
144 à 195 dB, 8 sec 4,5 kHz 190 - 195 dB > 190 dB env. 100 dB SEL°
Tableau 23 : Insonifications et TTS des dauphins insonifiés à 3 kHz avec des durées variables(d'après Finneran et al., 2005).(*) pour une durée de 1 sec, le SEL est égal au niveau sonore rms. (°) seuil auditif du dauphin évalué à 90dB pour 3 kHz
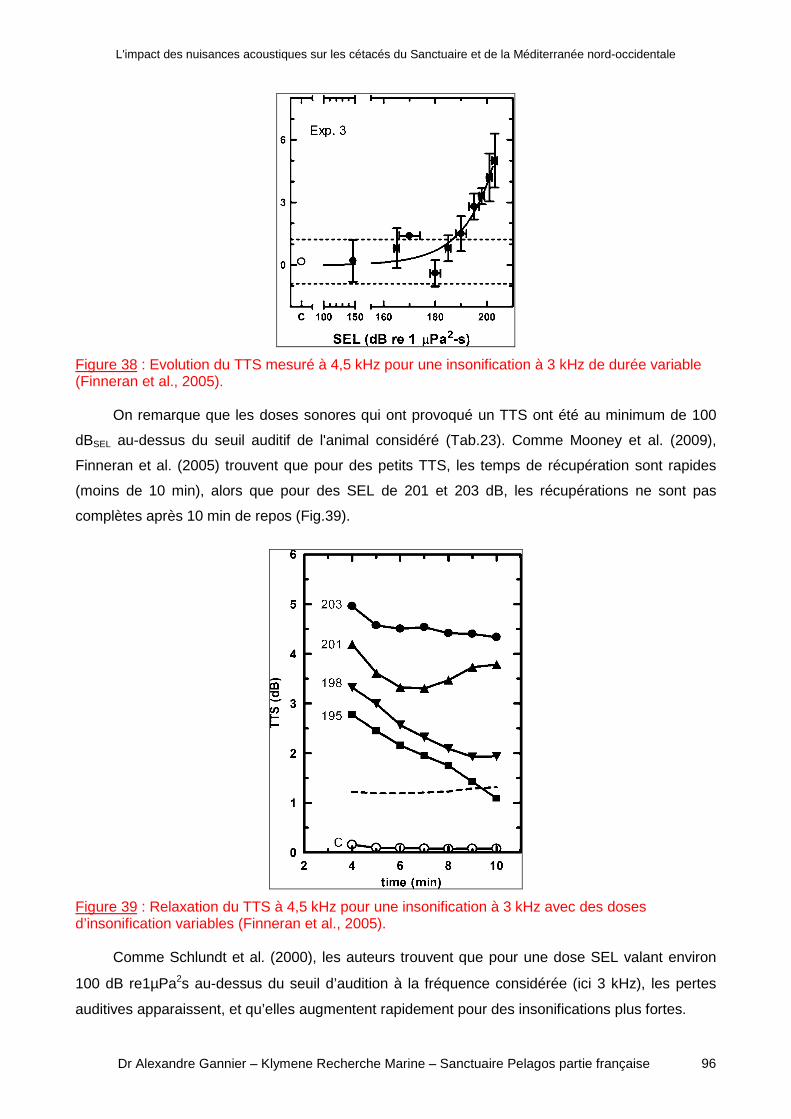
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 96
Figure 38 : Evolution du TTS mesuré à 4,5 kHz pour une insonification à 3 kHz de durée variable(Finneran et al., 2005).
On remarque que les doses sonores qui ont provoqué un TTS ont été au minimum de 100
dBSEL au-dessus du seuil auditif de l'animal considéré (Tab.23). Comme Mooney et al. (2009),
Finneran et al. (2005) trouvent que pour des petits TTS, les temps de récupération sont rapides
(moins de 10 min), alors que pour des SEL de 201 et 203 dB, les récupérations ne sont pas
complètes après 10 min de repos (Fig.39).
Figure 39 : Relaxation du TTS à 4,5 kHz pour une insonification à 3 kHz avec des dosesd’insonification variables (Finneran et al., 2005).
Comme Schlundt et al. (2000), les auteurs trouvent que pour une dose SEL valant environ
100 dB re1µPa2s au-dessus du seuil d’audition à la fréquence considérée (ici 3 kHz), les pertes
auditives apparaissent, et qu’elles augmentent rapidement pour des insonifications plus fortes.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 97
Mooney et al. (2009b) ont soumis un Grand dauphin à des séries de 3 chirps de 0,5 s
modulés à 3 kHz, répétés toutes les 26 s40, avec un niveau reçu de 203 dBrms au maximum. Les
tests auditifs étaient réalisés pour la fréquence de 5,6 kHz uniquement, et par la méthode
électrophysiologique. Le bruit de fond était fort au-dessus de 3 kHz (Mooney et al., 2009a).
Les auteurs ont constaté que des TTS apparaissaient seulement pour des niveaux sonores
de 203 dB reçus pendant 5 séries consécutives de chirps, correspondant à des doses SEL totales
de 214 dB re1µPa2s. Ces pertes auditives s'amortissaient 10 à 20 minutes après l'insonification.
Compte-tenu des données sur ce dauphin, les niveaux d'exposition ayant entraîné le début de TTS
sont environ de 114 dBSEL au-dessus du seuil d'audition. Mais, le cycle de transmission permettait
une certaine relaxation de la fatigue auditive entre deux séries de chirps, ce qui explique que la
dose seuil de TTS ait été aussi élevée.
Pour explorer le domaine des expositions très longues, Finneran et al. (2007) ont réalisé des
expériences sur une femelle Grand dauphin, avec des insonifications à 20 kHz et des durées de
64 s (tests 1 et 2) ou de 16 s répétées 3 fois (test 3). Cette étude a été exécutée dans un bassin
avec un faible bruit de fond. Les tests audiométriques comportementaux étaient réalisés de
manière accélérée après les insonifications, pour mesurer les TTS peu de temps après les
expositions aux sons intenses. Les seuils d'audition étaient d'environ 62 dB pour le domaine 6-
30 kHz (méthode comportementale) et 75-80 dB pour le domaine 10-40 kHz avec la méthode
AEP.
Le niveau de l’exposition a été maintenu en moyenne à 185-186 dBrms, ce qui équivaut à
des SEL de 203-204 dB re1µPa2s (exposition de 64 s) ; pour le test n°3, le nivea u maintenu a été
de 193 dB, soit un SEL de 210 dB re1µPa2s (3 fois 16 s d'insonification, séparées par 11 et 13
min). Les pertes auditives TTS engendrées par les trois types d’exposition ont été très importantes
aux fréquences de 30 (35 dB) et 40 kHz (20 dB), et nulles ou négligeables pour les autres
fréquences (10, 20, 50, 70 kHz). La perte auditive à 30 kHz a eu tendance à persister plus de
quatre jours après l'insonification (Fig.40), ce qui indique que le TTS a été très sévère. Il s'agit
d'une perte auditive persistante. La perte auditive a été très importante à la fois pour l’insonification
unique de 64 s et la série de 3 insonifications de 16 s (au moins 26 dB de TTS mesuré 17 min
après la fin de l’exposition, dans ce dernier cas), ce qui suggère que la relaxation du TTS n’a pas
été importante pendant les 11-13 min de repos entre deux insonifications de 16 s.
40 Il s’agit d’un cycle de transmission se rapprochant de celui d’un sonar de type SQS-53.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 98
Figure 40 : Pertes auditives TTS engendrées à 30 et 40 kHz par une insonification de 64 sec et186 dB à 20 kHz (méthode comportementale) (Finneran et al., 2007).
Les insonifications ont donc engendré des surdités partielles très importantes pour une
durée de plusieurs heures à plusieurs jours, principalement à 30 et 40 kHz (Fig.41).
Figure 41 : Variations de l'audiogramme engendrées par une insonification de 64 s et 185 dB à20 kHz (audiométrie par méthode AEP) (Finneran et al., 2007).
Donc, Finneran et al. (2007) ont généré chez le Grand dauphin de très forts TTS par des
expositions de plus d’une minute à un niveau de 123-124 dB au-dessus du seuil d’audition (niveau
perçu). Ces très fortes pertes auditives proviennent de la dose d'exposition qui est supérieure de
142 dBSEL au seuil auditif à 62 dB. Les pertes auditives subsistent plus d’une journée, pour
30 kHz.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 99
D’autres expériences du même type ont été menées avec des insonifications aux fréquences
de 3 et 20 kHz (Finneran et al., 2010a ; b ; c), dans l’optique de déterminer les phénomènes
d’augmentation des TTS pour de longues durées d’insonification et à des fréquences distinctes.
Finneran et al. (2010a) ont soumis un Grand dauphin à des insonifications d’une durée de 16 s
pour 3 et 20 kHz, le dauphin ayant des seuils d’auditions (S) de 81 et 66 dB, respectivement, à ces
fréquences. Ils ont exprimé les doses sonores nécessaires pour obtenir une perte auditive au
moins égale à 6 dB, 4 minutes après l’exposition (TTS4). A 3 kHz, le niveau SEL provoquant cette
perte significative valait 191 dB re1µPa2s, soit S + 110 dB, et à 20 kHz, 181 dB re1µPa2s, soit S +
115 dB. Passé ce stade, la croissance des TTS est très rapide, surtout pour la fréquence de
20 kHz, pour laquelle les auteurs ont obtenu des TTS supérieurs à 25 dB (Fig.42).
Figure 42 : Evolution de la perte auditive (TTS4) en fonction du niveau sonore pour une duréed’exposition de 16 s et les fréquences de 3 et 20 kHz (Finneran et al., 2010a).
Finneran et al. (2010b) ont aussi réalisé des expériences à la fréquence de 3 kHz, en variant
les niveaux sonores et avec des durées d’exposition croissantes, jusqu’à 128 s. Ils ont observé
l’augmentation des pertes auditives avec le temps d’exposition : des TTS4 de plus de 20 dB ont été
obtenus avec des expositions de 100 s à un niveau sonore de 197 dB re1µPa (soit S + 116 dB).
Exprimé en terme de dose SEL (dB re1µPa2s), un TTS4 de 1 dB est obtenu pour une dose de
S + 99 dB, 2 dB pour S + 112 dB, 3 dB pour une dose de S + 121 dB, 15 dB pour une dose de
S + 137 dB. Les auteurs montrent aussi que la croissance des pertes auditives est non-linéaire
aussi bien lorsqu’on augmente la durée d’exposition que lorsqu’on fait croître le niveau sonore
transmis (Fig.43). Nous observons qu’il n’y a pas stricte équivalence entre les résultats
expérimentaux, à 3 kHz, exposés par Finneran et al. (2010a) et ceux exposés dans Finneran et al.
(2010b) : nous voyons ici les limites de la précision de l’approche expérimentale, et sans doute de
la métrique employée pour exprimer les insonifications (doses SEL perçues).
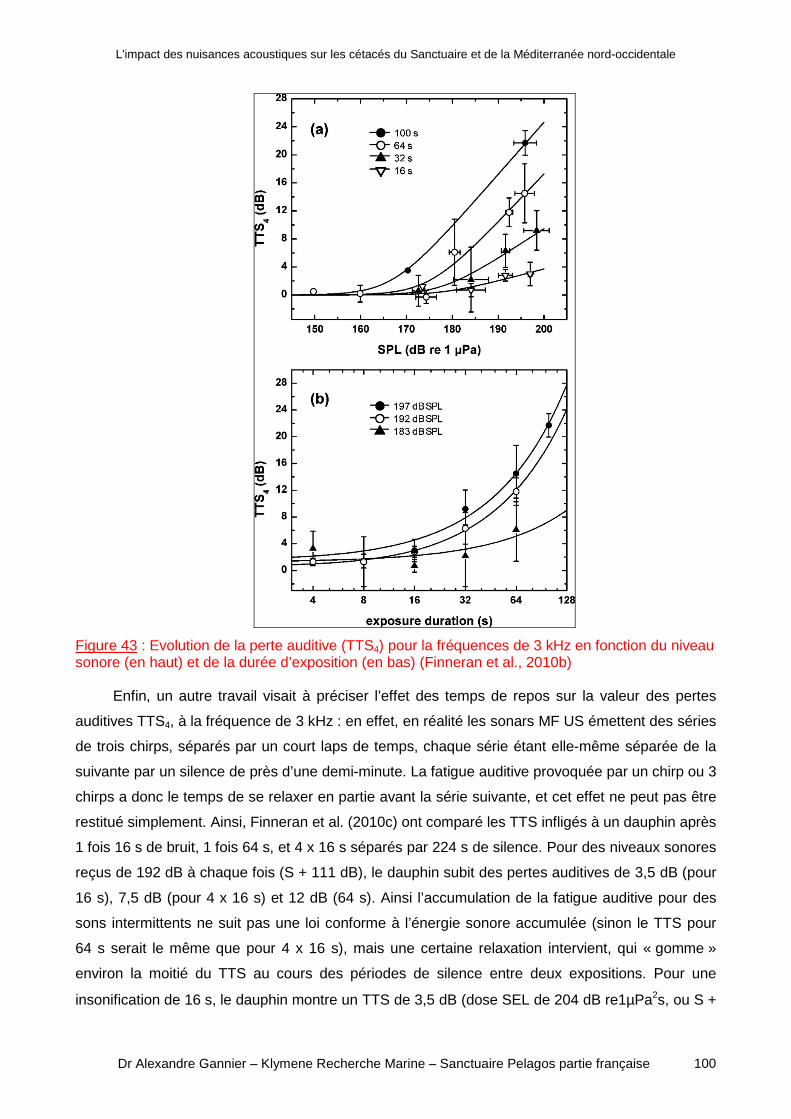
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 100
Figure 43 : Evolution de la perte auditive (TTS4) pour la fréquences de 3 kHz en fonction du niveausonore (en haut) et de la durée d’exposition (en bas) (Finneran et al., 2010b)
Enfin, un autre travail visait à préciser l’effet des temps de repos sur la valeur des pertes
auditives TTS4, à la fréquence de 3 kHz : en effet, en réalité les sonars MF US émettent des séries
de trois chirps, séparés par un court laps de temps, chaque série étant elle-même séparée de la
suivante par un silence de près d’une demi-minute. La fatigue auditive provoquée par un chirp ou 3
chirps a donc le temps de se relaxer en partie avant la série suivante, et cet effet ne peut pas être
restitué simplement. Ainsi, Finneran et al. (2010c) ont comparé les TTS infligés à un dauphin après
1 fois 16 s de bruit, 1 fois 64 s, et 4 x 16 s séparés par 224 s de silence. Pour des niveaux sonores
reçus de 192 dB à chaque fois (S + 111 dB), le dauphin subit des pertes auditives de 3,5 dB (pour
16 s), 7,5 dB (pour 4 x 16 s) et 12 dB (64 s). Ainsi l’accumulation de la fatigue auditive pour des
sons intermittents ne suit pas une loi conforme à l’énergie sonore accumulée (sinon le TTS pour
64 s serait le même que pour 4 x 16 s), mais une certaine relaxation intervient, qui « gomme »
environ la moitié du TTS au cours des périodes de silence entre deux expositions. Pour une
insonification de 16 s, le dauphin montre un TTS de 3,5 dB (dose SEL de 204 dB re1µPa2s, ou S +

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 101
123 dB) et une insonification de 64 s (SEL de 210 dB re1µPa2s, ou S + 129 dB) provoque un TTS
de 12 dB.
En résumé (Tab.24), le seuil de TTS pour des insonifications en son pur de durée variable se
situe dans la plage de 100-115 dB au dessus du seuil auditif à la fréquence considérée. On
observe que la fréquence du TTS maximal est localisée autour de 1,5 fois la fréquence de
l'insonification, la fatigue auditive ne se situant pas sur tout le domaine audible.
Insonification TTS à la fréquence1,5 f
Seuil de TTS rapportéau seuil auditif
Référence
Son pur 3 kHz - 194 dB - 1 s MTTS 16 dB à 4,5 kHz ? Schlundt et al., 2000
Son pur 20 kHz - 196 dB - 1 s MTTS 6 dB à 30 kHz < 109 dB Schlundt et al., 2000
Son pur 3 kHz - 194 dB - 1 s seuil 197 dB à 4,5 kHz 111 dB Finneran et al., 2005
Son pur 3 kHz - 2 à 8 s 190-195 dB à 4,5 kHz env. 100 dB SEL Finneran et al., 2005
Son pur 20 kHz - 186 dB - 64 s TTS de 35 dB à 30 kHz <<< 142 dB SEL Finneran et al., 2007
Série de 3 sons 16 s à 20 kHz TTS > 26 dB à 30 kHz <<< 146 dB SEL Finneran et al., 2007
Série de 3 sons 0,5 s à 3 kHz seuil 203 dB à 5,6 kHz env.114 dB SEL Mooney et al., 2009
Série de 3 sons 16 s à 3 kHz seuil de 191 dB SEL env. 110 dB SEL Finneran et al., 2010a
Série de 3 sons 16 s à 20 kHz seuil de 181 dB SEL env. 115 dB SEL Finneran et al., 2010a
Son pur à 3 kHz durée 4 à 128 s seuil de 177 dB SEL env. 99 dB SEL Finneran et al., 2010b
Son pur à 3 kHz durée 16 s seuil < 204 dB SEL < 123 dB SEL Finneran et al., 2010c
Son pur à 3 kHz durée 64 s TTS 12 dB à 211 SEL << 130 dB SEL Finneran et al., 2010c
Son pur à 3 kHz durée 4 x 16 s TTS 7,5 dB à 210 SEL << 129 dB SEL Finneran et al., 2010c
Tableau 24: Synthèse des insonifications en sons purs et des TTS observés chez le Granddauphin.
Une insonification d’un niveau SEL inférieur à 100 dB au-dessus du seuil d’audition à la
fréquence considérée ne semble pas produire de perte auditive de type TTS.
o Cas des impulsions
Le Grand dauphin a été soumis à des impulsions sonores d’origine variée, canons
hydrauliques ou explosifs simulés. La pression sonore d’une explosion évolue en quelques
millisecondes : l’expression d’une métrique pertinente pour quantifier les effets auditifs de ce
phénomène sonore est un problème ardu. L’utilisation d’une valeur efficace (dBrms) se heurte au
problème de la définition de l’intervalle de temps sur lequel calculer ce niveau. L’utilisation d’un
niveau sonore pic-pic (dBp-p) est envisageable, mais ce niveau ne restitue pas la totalité de
l’énergie sonore reçue. C’est une expression en niveau d’exposition SEL (dB re1µPa2.s) qui
convient le mieux, car elle donne une métrique équivalente pour les impulsions de durées et de
valeurs maximales différentes, en tenant compte du nombre de répétitions. Nous exprimons les
différents résultats d’expériences avec leurs niveaux SEL et leurs niveaux pic-pic.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 102
Finneran et al. (2000) ont soumis deux dauphins et un Bélouga à des explosions simulées
en présence d’un bruit masquant artificiel, les sons étant générés par des transducteurs piézo-
électriques. Pour une charge simulée de 500 kg explosant à 1,7 km, l'onde sonore avait une durée
de 9,5 ms et un niveau pic-pic de 221 dB a été mesuré près de la tête des cétacés, avec un niveau
d’exposition SEL de 179 dB re1µPa2.s. Le dauphin a subi un TTS de l'ordre de 3 à 6 dB mesuré à
la fréquence de 2400 Hz (pas optimale pour ce cas précis). Avec un seuil d’audition (masqué) à
118 dB, le niveau SEL reçu était de 61 dB SEL au-dessus du seuil (et de 103 dBp-p).
Finneran et al. (2002) ont procédé à l'insonification des mêmes deux espèces, le Grand
dauphin et le Bélouga, par des impulsions de canon hydraulique, toujours en présence d’un bruit
masquant artificiel. Le canon produisait une impulsion dure de 10 à 20 ms environ, proche de celle
d’un canon pneumatique. Pour le Grand dauphin, aucune perte auditive n'a été constatée pour un
niveau de reçu de 228 dBp-p. Les niveaux d’exposition sonore reçus pendant les impulsions les
plus fortes étaient de 185-188 dB re1µPa2.s . Le seuil d'audition (masqué) se situait à 117-126 dB
pour le Grand dauphin, les niveaux SEL reçus ont donc été de 60-70 dB au-dessus du seuil (ou
91-98 dBp-p).
En résumé, pour des impulsions de 220 à 229 dBp-p appartenant aux deux catégories ci-
dessus, le grand dauphin a présenté une faible perte auditive dans un cas (explosifs simulés) mais
pas dans l’autre. Ces niveaux sont supérieurs de plus de 100 dB à son seuil auditif, et une fois
exprimés en SEL, ils sont de 60 à 70 dB supérieurs au même seuil. Dans les deux cas, le
masquage par un bruit de fond artificiel a induit une augmentation du seuil auditif et la
généralisation de ces résultats n’est pas possible. Les deux ensembles de résultats sur les effets
auditifs d’impulsions portant sur le Grand dauphin (Finneran et al., 2000 ; 2002) indiquent que des
valeurs pic-pic supérieures à 220 dB, ou encore supérieures de plus de 100 dBp-p et 60-70 dBSEL
au seuil auditif masqué n’ont pas provoqué de TTS significatif, mais semblent approcher ou
légèrement dépasser le seuil admissible.
• Pertes de sensibilité auditive (TTS) observées chez d’autres espèces
o Cas des impulsions
Finneran et al. (2000) donne des résultats de TTS après des explosions simulées pour un
Bélouga (Delphinapterus leucas), qui avait des seuils d’audition de 119-123 dB en moyenne. Mes
deux expositions les plus fortes (niveaux reçus de 220 et 221 dBp-p) ont provoqué une variation de
seuil d’audition de l’ordre de 2-4 dB pour le bélouga, jugée non significative selon le critère adopté
par les auteurs. Comme pour le Grand dauphin, les auteurs suggèrent que le masquage par un
bruit artificiel, en élevant le seuil auditif, a eu pour conséquence d’augmenter la résistance des
cétacés aux pertes auditives.
Pour le canon hydraulique, Finneran et al. (2002) ont constaté des MTTS pour le bélouga,
pour niveaux reçus supérieurs ou égal à 202 dBp-p, c’est-à-dire inférieurs de 20 dB au seuil

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 103
observé pour le Grand dauphin. Ce niveau est de seulement 85 dBp-p au dessus du seuil
d’audition (masqué) de l’animal.
Lucke et al. (2009) ont étudié les pertes TTS induites par des impulsions simples de type
« airgun » sur un Marsouin commun. La mesure des seuils auditifs a été réalisée par la méthode
électrophysiologique AEP. Les installations étaient localisées en milieu semi-ouvert à proximité
d’un port et étaient donc soumises à un bruit ambiant variable correspondant au trafic portuaire. Le
seul auditif était beaucoup plus élevé que celui estimé dans des cas antérieurs (Fig.44).
Figure 44 : Seuils d'audition du marsouin mesuré par AEP (Lucke et al., 2009).Triangles : seuils mesurés lors de cette étude.Autres symboles : audiométries issues d’étude antérieures.
A une fréquence d’audition de 4 kHz, les auteurs ont constaté un TTS de 8 dB pour un
niveau sonore reçu de 199,7 dBp-p ou une dose d’exposition (SEL) de 164,3 dB re1µPa2s. Avec
un seuil d'audition de 100-117dB, la valeur du niveau sonore provoquant ce TTS était de 83-100
dBp-p (ou 47-64 dB SEL au-dessus du même seuil). Des TTS de 21 dB ont été obtenus pour des
expositions légèrement plus fortes (Fig.45) : la croissance du TTS en fonction du niveau reçu a
donc été très forte dès que l'on a dépassé le niveau de 200 dBp-p.
La récupération de l'audition normale après l'insonification (relaxation du TTS) a été très
lente puisque 29 heures après, le marsouin n'avait pas encore retrouvé sa sensibilité initiale. Le
seuil d’audition élevé au départ suggère que le marsouin était soumis à un fort bruit de fond. Les
valeurs d’insonification entraînant des MTTS sont très faibles par comparaison avec des résultats
sur d’autres odontocètes : 83 dBp-p ou 45 dBSEL au dessus du seuil.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 104
Figure 45 : Pertes auditives TTS à 4 kHz pour le Marsouin commun (Lucke et al., 2009).Seuils d’auditions (ordonnées) obtenus après expériences en fonction d’insonification exprimés soit en dBp-p (ovales) soit en niveau SEL (rectangles)
En résumé, pour des impulsions de 220 à 229 dBp-p appartenant aux deux catégories ci-
dessus, le Grand dauphin présente peu de perte auditive. Ces niveaux sont supérieurs de plus de
100dB à son seuil auditif. Le Bélouga, au contraire est plus sensible, puisqu’il montre une
tendance à un MTTS pour un niveau de 220 dBp-p (explosion) et un MTTS significatif pour un
niveau de 226 dBp-p (watergun). Avec un bruit de fond non contrôlé, le marsouin subit une perte
auditive pour un niveau de 200 dBp-p, valeur qui correspond à un niveau de moins de 100 dBp-p
au dessus de son seuil auditif.
Espèce Impulsion Seuil de TTS en niveau reçu Seuil de TTS rapporté auseuil auditif
marsouin commun Airgun < 200 dBp-p ou 164 dB SEL 83-100 dBp-p ou 47-64 dB SEL
grand dauphin Explosion > 220 dBp-p ou 177 dB SEL >102 dBp-p ou 59 dB SEL
grand dauphin Watergun > 229 dBp-p ou 187 dB SEL >112 dBp-p ou 70 dB SEL
bélouga Explosion ≥ 220 dBp-p ou 179 dB SEL ≥ 97 dBp-p ou 54 dB SEL
bélouga Watergun 220 dBp-p ou SEL 103 dBp-p ou 63 dB SEL
Tableau 25: Synthèse des insonifications et MTTS en sons impulsifs.
Le nombre d’expériences est réduit, et de plus toutes ont été réalisées dans des conditions
de bruit de fond élevé, et des mesures de TTS en quantité insuffisante. Les données issues de ces
tests semblent peu fiables. Les valeurs de 200 dBp-p ou 100 dBp-p effectif semblent pouvoir être
prises comme seuil indicatif pour éviter les TTS chez ces cétacés, avec précaution.
o Cas des sons non-impulsifs
Selon Schlundt et al. (2000), pour des sons simples de durée 1 s, un Bélouga a montré des
TTS pour des niveaux reçus variant entre 195 dB à 3 kHz et 200 dB à 20 kHz (Tab.24). Pour les

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 105
hautes fréquences, et aussi pour les très hautes fréquences (75 kHz), le Bélouga est moins
sensible, il n’a pas subi de TTS. A 20 kHz, les niveaux de 110 à 114 dB au-dessus du seuil auditif
ont provoqué des TTS significatifs aux fréquences de 20 à 40 kHz. Ces résultats justifient qu’un
niveau perçu de 100 dB soit retenu comme valeur seuil pour l’apparition des TTS. Cependant, les
conditions de ces expériences doivent inciter à de la prudence dans l’interprétation de ces
résultats : le seuil d’audition du cétacés était déjà très élevé en raison du masquage employé pour
stabiliser le niveau de bruit de fond lors des expériences (87 dB à 20 kHz, par ex.).
Fréquence Niveau reçu(dB rms)
TTS bélouga à lafréquence de
TTS bélouga rapportéau seuil auditif
3 kHz 160 à 202 pour 195 dB, 12 dB @ 4,5 kHz
10 kHz 180 à 197 pour 192 dB, 7 dB @ 20 kHz
20 kHz 180 à 201 pour 197 dB, 8 dB @ 40 kHz pour 110 dB, 8 dB @ 40k
Tableau 26 : Insonifications et MTTS des bélougas (d'après Schlundt et al., 2000).
Kastelein et al. (2013) ont effectué des expériences similaires sur un Marsouin commun,
dans un bassin avec peu de bruit de fond : l’animal a été soumis a un son modulé à bande étroite
de 154 dB de niveau sonore à la fréquence de 1,5 kHz, soit une dose sonore SEL de 190 dB
re1µPa2s. Les TTS mesurés 1 à 4 min après les insonifications étaient de 14 dB à 1,5 kHz et de 11
dB à 2 kHz. Comme la sensibilité audiométrique à 1,5 kHz était de 78 dB, le niveau reçu a été de
76 dB au-dessus du seuil, ou encore 112 dB SEL. Comme cette dose reçue a entraîné un TTS de
plus de 10 dB, on peut estimer que l’emploi d’une valeur de 100 dB SEL en valeur effective (au-
dessus du seuil d’audition) comme critère d’apparition des TTS est valable. Par ailleurs, Kastelein
et al. (2013) montrent que lors de ces expériences, les pertes auditives partielles ont duré près
d’une heure.
Dans une autre série d’expériences, les auteurs ont recherché les TTS provoqués par des
bruits blancs par octave à 4 kHz, fréquence pour laquelle le Marsouin avait une sensibilité auditive
de 60 dB, et pour des temps d’exposition variant de 7,5 à 240 min. Des seuils de TTS ont été
trouvés pour des doses sonores de 150 et 160 dB re1µPa2s (avec des niveaux reçus de 124 et
136 dB, respectivement, pour des expositions de 7,5 min). On a dans ces deux cas des doses
effectives de 90 et 100 dBSEL. Un TTS très important de 15 dB a été provoqué par une dose de
190 dB, correspondant à un niveau reçu de 148 dB pendant 240 min. On constate donc que la
valeur de 100 dBSEL au-dessus du seuil d’audition constitue ici aussi une référence utile pour
déterminer un risque de TTS, même si pour un niveau reçu plus faible, elle correspond déjà à un
TTS de 5 dB.
Popov et al. (2011) ont réalisé des expériences de fatigue auditive sur deux Marsouins du
Yangtsé (Neophocoena phocaenoides asiaeorientalis) avec des bruits par demi-octave entre 22,5
et 128 kHz, pour des durées d’exposition de 1 à 30 minutes et un niveau reçu de 150 dB re 1µPa.
La combinaison du niveau de pression sonore et des durées a conduit à des doses d’exposition de

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 106
168 dBSEL (1 min) à 183 dBSEL (30 min). En fonction des seuils d’audition aux fréquences du bruit
(75 dB à 22,5 kHz, 62,5 dB à 45 kHz, 62,5 dB à 90 kHz), les niveaux d’exposition ont été de 93 à
130 dBSEL au-dessus des seuils ; mais ces derniers ont été obtenus par méthode AEP et sont un
peu supérieurs aux valeurs réelles de sensibilité auditive. Les TTS mesurés ont été très importants
dans quasiment tous les cas, au moins 17 dB et jusqu’à 45 dB, avec des durées de récupération
pouvant excéder 20 heures. Popov et al. (2011) n’avaient pas comme objectif de rechercher les
seuils de TTS.
Le même type d’expérience a été réalisé sur deux Bélougas (Popov et al., 2013), mais en
adoptant un niveau sonore de 165 dB re 1µPa et des fréquences de bruit 11,2 à 90 kHz. L’objectif
était toujours d’observer les très forts TTS et leurs temps de récupération, donc les seuils
d’apparition des TTS n’ont pas été observés. En effet, pour le temps minimal d’insonification
(1 min) la dose d’exposition était de 183 dBSEL, soit entre 120 et 135 dB au-dessus du seuil
d’audition à la fréquence considérée (65 dB à 11,2 kHz, 60 dB à 22,5 kHz, 50 dB à 45 et 90 kHz).
Insonification Espèce Seuil de TTSrapporté au seuil
Référence
Son pur 20 kHz - 197 dB - 1 s Bélouga < 110 dB Schlundt et al., 2000
Bruit 11,2 kHz – 1 à 30 min – 165 dB Bélouga <<< 120 dB SEL* Popov et al., 2013
Bruit 22.5 kHz – 1 à 30 min – 165 dB Bélouga <<< 125 dB SEL* Popov et al., 2013
Bruit 45 kHz – 1 à 30 min – 165 dB Bélouga <<< 135 dB SEL* Popov et al., 2013
Son modulé 1,5 kHz - 154 dB Marsouin commun << 112 dB SEL Kastelein et al., 2013
Bruit blanc 4 kHz – 7,5 min – 124 dB Marsouin commun env. 90 dB SEL Kastelein et al., 2012
Bruit blanc 4 kHz – 7,5 min – 136 dB Marsouin commun env 100 dB SEL Kastelein et al., 2012
Bruit blanc 4 kHz – 240 min – 148 dB Marsouin commun <<< 130 dB SEL Kastelein et al., 2012
Bruit 22,5 kHz – 1 à 30 min – 150 dB Marsouin Yang-Tsé << 93 dB SEL* Popov et al., 2011
Bruit 45 kHz – 1 à 30 min – 150 dB Marsouin Yang-Tsé << 105 dB SEL* Popov et al., 2011
Bruit 90 kHz – 1 à 30 min – 150 dB Marsouin Yang-Tsé << 105 dB SEL* Popov et al., 2011
Tableau 27 : Insonifications et TTS pour 4 espèces en sons non-impulsifs.(*) mesuré par méthode AEP, donc certainement sous-estimé.
Au travers de ces exemples et des études réalisées sur le Grand dauphin (Tab.27), il
apparaît que le critère d’une dose de 100 dBSEL au-dessus du seuil d’audition, donc 100 dB effectif,
permet de séparer avec justesse la plupart des cas d’insonification : en-deçà de ce seuil, les
pertes auditives sont rares, et au-dessus, elles sont quasi-systématiques.
L’avantage évident de ce critère, même s’il est approximatif, est qu’il s’exprime en tenant
compte de la sensibilité auditive, d’une part, et en tenant compte de la durée des expositions,
d’autre part. On peut attribuer au critère des « 100 dBSEL » une précision de +/- 10 dB : à l’intérieur
de cette fourchette se trouve tous les cas de pertes auditives disponibles dans la littérature.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 107
Au terme de notre examen bibliographique exhaustif, on sait que les pertes auditives
temporaires (TTS) surviennent quand les cétacés sont soumis à des intensités sonores fortes,
aussi bien en présence d’impulsions de type airgun ou explosion, qu’en présence de sons non-
impulsifs comme ceux des sonars.
Dans le cas des sons non-impulsifs, les seuils critiques au delà desquels se produisent les
TTS peuvent être exprimés en terme de dose d’exposition sonore SEL (en dB re1µPa2s), c’est-à-
dire la valeur équivalente à un niveau de pression sonore appliqué pendant une seconde : en deçà
d’un seuil de 100 dBSEL effectif (mesuré par rapport au seuil d’audition), il semble que les
odontocètes ne souffrent pas de TTS significatif. Pour chaque doublement de la durée
d’exposition, le niveau de pression sonore tolérable diminue de 3 dB, par exemple 97 dB effectif
pour 2 s, 94 dB pour 4 s, etc. Si l’exposition dure moins qu’une seconde, le niveau sonore tolérable
passe au dessus de 100 dB (par ex., 103 dB pour 0,5 s, etc.).
Dans le cas des impulsions, le critère de niveau tolérable est plus difficile à exprimer, à
cause de résultats expérimentaux insuffisants. Il semble qu’au delà d’un niveau de pression
sonore de 200 dBp-p ou encore de 100 dBp-p au dessus du seuil d’audition, il y ait un risque
d’apparition de TTS. En présence d’impulsions répétées, comme celles d’une prospection
sismique ou du battage de pieu, les expériences sur les cétacés ne fournissent pas d’indication sur
la démarche à adopter. La notion de dose d’exposition SEL a sans doute une utilité, mais elle n’a
pas été testée.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 108
c) Effets observés lors d’expériences d’exposition contrôlée
Les expériences d’exposition contrôlée, ou CEE comme « controled exposure experiments »
ou encore « behavioral response studies », sont pratiquées depuis plus d’une décennie sur des
cétacés sauvages. Elles servent à mesurer les effets comportementaux provoqués par les
insonifications : variation de mouvement, changement d’activité ou de comportement acoustique,
modification du cycle de sonde.
Les effets comportementaux, ainsi que les sons reçus, sont mesurés grâce à une balise de
type DTag (Johnson & Tyack, 2003) qui est fixée sur l’animal avant l’exposition au son, grâce à
une ventouse. En général, on enregistre le comportement basal du cétacé pendant au moins un
cycle de sonde complet, avant de procéder à l’insonification. Puis on procède à la transmission
d’une ou plusieurs séquences du son en question. Ensuite, la balise est récupérée et les données
des capteurs sont déchargées sur un ordinateur. Dans l’idéal, la transmission de sons identiques
sur différents individus de la même espèce et sur différentes espèces permet de dresser un
tableau des effets comportementaux provoqués par un type de signal donné. La classification des
effets selon leur sévérité, en fonction du niveau sonore transmis aux cétacés, permet d’évaluer
l’impact comportemental de chaque catégorie de pollution sonore sur les différentes espèces.
Les effets comportementaux des insonifications concernent généralement un nombre
beaucoup plus grand de cétacés que les effets auditifs, car ils sont provoqués par des niveaux
sonores sensiblement plus faibles. Donc, en partant d’une source sonore de puissance donnée,
les effets comportementaux concernent des individus situés à des distances pouvant être 10 fois
ou 100 fois plus grandes que les effets de fatigue auditive (ou TTS). Selon leur sévérité, les effets
comportementaux ont donc un impact potentiellement plus fort que les effets auditifs, car ils
concernent un très grand nombre d’individus.
Par ailleurs, il ne fait aucun doute que les effets comportementaux découlent de la
perception du niveau sonore par un individu d’une espèce donnée : il faut convertir un « niveau
reçu » en un « niveau perçu » : dB perçu = dB reçu – seuil auditif (f) , ce qui nécessite de
connaître le seuil auditif de l’animal à la fréquence donnée. Il est donc très important de connaître
l’audiogramme de l’espèce en question pour évaluer la conséquence comportementale d’une
insonification donnée. Dans le cas où l’audiogramme du cétacé n’est pas connu, la prévision des
effets comportementaux est plus difficile, mais elle est encore possible de façon empirique si l’on
dispose de suffisamment d’exemples de perturbations observées.
C’est pour cette raison que les expériences de CEE ont connu un essor considérable ces
dernières années (programmes Bahamas BRS, 3S, SoCal) : parmi les espèces qui ont subi ces
expériences, il y en a un grand nombre pour lesquelles on n’a pas d’audiogramme. Ces
expériences sont nombreuses et les résultats ne sont pas tous publiés sous une forme facilement
utilisable. Néanmoins, les rapports d’étude permettent en général de recueillir des informations
précieuses sur les réactions des espèces aux insonifications (Tab.28).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 109
Programme Espèce Référence
Bahamas BRS Globicéphale tropical – Mésoplodon deBlainville
BRS-07 Cruise Report (January 2008)
Bahamas BRS Globicéphale tropical – Mésoplodon deBlainville – Pseudorque - Péponocéphale
BRS 2008 Preliminary Report (October 2008)
B & BRS SoCal Rorqual bleu – Rorqual commun –Cachalot – Dauphin de Risso - Ziphius
SOCAL-10 Project Report (February 2011)
B & BRS SoCal Rorqual bleu – Dauphin de Risso -Ziphius
SOCAL-11 Project Report (March 2012)
B & BRS SoCal Cachalot – Dauphin de Risso – Rorqualbleu – Rorqual commun – Ziphius –Grand dauphin – Dauphin commun
SOCAL-12 Project Report (April 2013)
B & BRS SoCal SOCAL-13 Project Report
BRS 3S Norway Orque – Globicéphale noir - Cachalot SOI – Technical Report (August 2011)
BRS 3S-2011 Mégaptère – Petit rorqual 3S-2011 Cruise report – FFI-2011- 01289
BRS 3S-2012 Mégaptère – Rorqual commun 3S-2012 Cruise report – FFI-2012 - 02058
Tableau 28 : Différents programmes d’expériences d’exposition contrôlée.
Lors de ces expériences, des sons proches des signaux de sonar sont transmis, et les
réactions des cétacés varient depuis l’indifférence jusqu’à la fuite avec panique, dans quelques
cas. La gradation de ces effets comportementaux peut être organisée selon deux catégories
génériques, de sévérité croissante : (1) effet comportemental sans évasion de l’habitat, (2) réaction
d’évasion de l’habitat. Dans le premier cas, l’intensité sonore perçue provoque éventuellement des
variations de comportement ou d’activité, alors que dans le second le niveau sonore perçu est
intense au point d’obliger les cétacés à quitter la zone où ils se trouvaient.
Pour ce qui est des effets sans évasion, on propose deux niveaux de dérangement : (1)
l’activité est altérée mais se poursuit, (2) l’activité est interrompue. Pour ce qui est de l’évasion, on
propose trois modalités : (1) éloignement, vitesse lente et cohésion du groupe maintenue, (2) fuite,
vitesse rapide avec ou sans maintien de la cohésion, (3) panique, mouvement rapide et
incohérent, avec désordre dans le groupe.
Notre méthode de synthèse des résultats de CEE consistera à placer sur des graphes
(fréquence x niveau sonore) les points expérimentaux extraits des rapports d’étude. Sur ces
graphes, on aura porté également les audiogrammes des espèces, quand ils sont connus, ou des
groupes d’espèces (représentés par au moins une espèce). Pour ce qui concerne la Méditerranée,
deux espèces posent des problèmes : le Rorqual commun (pas d’audiogramme connu, ni pour
l’espèce, ni pour aucun mysticète), et le Cachalot (pas d’audiogramme, mais quelques points
AEP). Pour ces espèces, on aura recours à des audiogrammes synthétiques, forcément très
approximatifs. La comparaison des points expérimentaux de CEE et des audiogrammes permet
d’exprimer les différentes sévérités de réaction en fonction des niveaux sonores perçus.
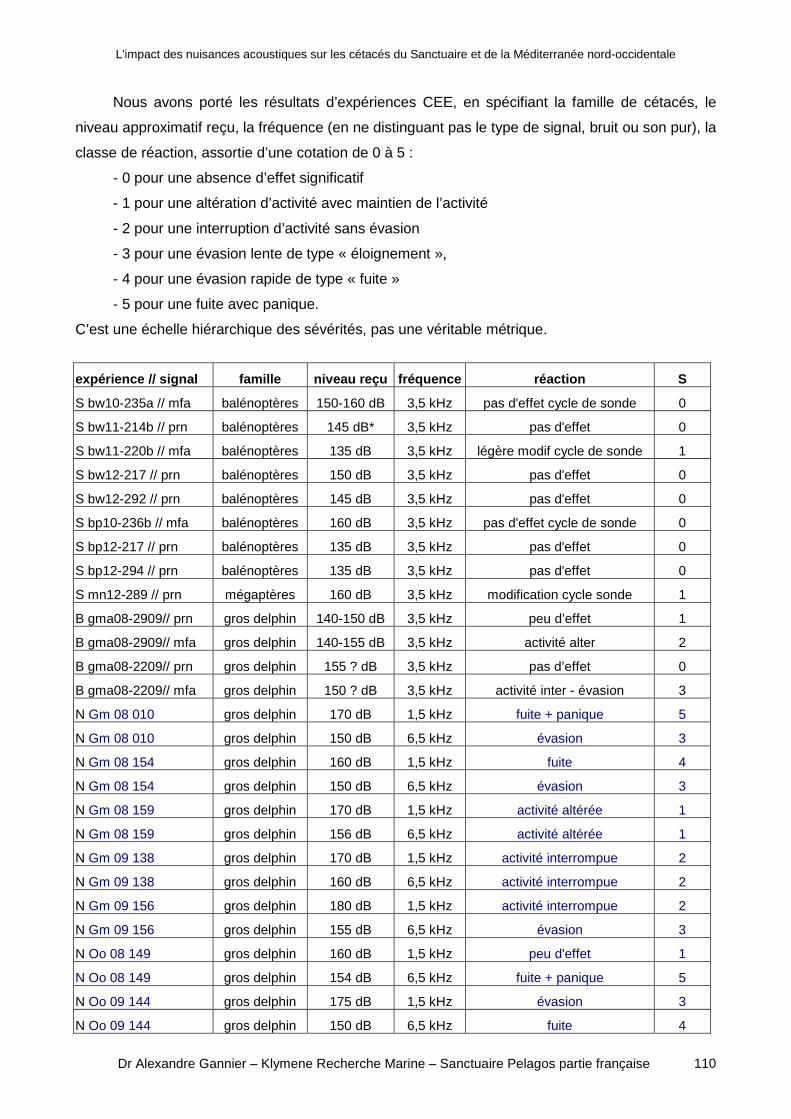
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 110
Nous avons porté les résultats d’expériences CEE, en spécifiant la famille de cétacés, le
niveau approximatif reçu, la fréquence (en ne distinguant pas le type de signal, bruit ou son pur), la
classe de réaction, assortie d’une cotation de 0 à 5 :
- 0 pour une absence d’effet significatif
- 1 pour une altération d’activité avec maintien de l’activité
- 2 pour une interruption d’activité sans évasion
- 3 pour une évasion lente de type « éloignement »,
- 4 pour une évasion rapide de type « fuite »
- 5 pour une fuite avec panique.
C’est une échelle hiérarchique des sévérités, pas une véritable métrique.
expérience // signal famille niveau reçu fréquence réac tion S
S bw10-235a // mfa balénoptères 150-160 dB 3,5 kHz pas d'effet cycle de sonde 0
S bw11-214b // prn balénoptères 145 dB* 3,5 kHz pas d'effet 0
S bw11-220b // mfa balénoptères 135 dB 3,5 kHz légère modif cycle de sonde 1
S bw12-217 // prn balénoptères 150 dB 3,5 kHz pas d'effet 0
S bw12-292 // prn balénoptères 145 dB 3,5 kHz pas d'effet 0
S bp10-236b // mfa balénoptères 160 dB 3,5 kHz pas d'effet cycle de sonde 0
S bp12-217 // prn balénoptères 135 dB 3,5 kHz pas d'effet 0
S bp12-294 // prn balénoptères 135 dB 3,5 kHz pas d'effet 0
S mn12-289 // prn mégaptères 160 dB 3,5 kHz modification cycle sonde 1
B gma08-2909// prn gros delphin 140-150 dB 3,5 kHz peu d’effet 1
B gma08-2909// mfa gros delphin 140-155 dB 3,5 kHz activité alter 2
B gma08-2209// prn gros delphin 155 ? dB 3,5 kHz pas d’effet 0
B gma08-2209// mfa gros delphin 150 ? dB 3,5 kHz activité inter - évasion 3
N Gm 08 010 gros delphin 170 dB 1,5 kHz fuite + panique 5
N Gm 08 010 gros delphin 150 dB 6,5 kHz évasion 3
N Gm 08 154 gros delphin 160 dB 1,5 kHz fuite 4
N Gm 08 154 gros delphin 150 dB 6,5 kHz évasion 3
N Gm 08 159 gros delphin 170 dB 1,5 kHz activité altérée 1
N Gm 08 159 gros delphin 156 dB 6,5 kHz activité altérée 1
N Gm 09 138 gros delphin 170 dB 1,5 kHz activité interrompue 2
N Gm 09 138 gros delphin 160 dB 6,5 kHz activité interrompue 2
N Gm 09 156 gros delphin 180 dB 1,5 kHz activité interrompue 2
N Gm 09 156 gros delphin 155 dB 6,5 kHz évasion 3
N Oo 08 149 gros delphin 160 dB 1,5 kHz peu d'effet 1
N Oo 08 149 gros delphin 154 dB 6,5 kHz fuite + panique 5
N Oo 09 144 gros delphin 175 dB 1,5 kHz évasion 3
N Oo 09 144 gros delphin 150 dB 6,5 kHz fuite 4

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 111
B pc08-2609// prn gros delphin 150-160 dB 3,5 kHz activité inter - évasion 3
B pc08-2609// mfa gros delphin 150-155 dB 3,5 kHz activité inter - fuite 4
B pc08-2809// prn gros delphin 155-160 dB 3,5 kHz évasion 3
B pc08-2809// mfa gros delphin 145-155 dB 3,5 kHz évasion 3
S gg11-164a // mfa d.océaniques 140-145 dB 3,5 kHz pas d'effet (Gg en surface) 0
S gg12-209 // prn d.océaniques 135 dB 3,5 kHz pas d'effet 0
B pe08-2909 // mfa d.océaniques (140 dB) 3,5 kHz regroupemt - immobilisation 2
B md07-0209 // mfa ziphiidés 145 dB 3,5 kHz activité inter - évasion 3
B md08-2709 // prn ziphiidés 140-150 dB 3,5 kHz altération d'activité 1
S zc10-272a // mfa ziphiidés 110 dB 3,5 kHz dérangement prédation 1
S zc10-272a // mfa ziphiidés 135 dB 3,5 kHz interruption prédation 2
S zc11-267a // mfa ziphiidés 140 dB 3,5 kHz modif de la descente sonde 1
S bb12-267a // mfa ziphiidés 140 dB 3,5 kHz pas d’effet significatif 0
S pm10-242a // mfa cachalot 150-160 dB 3,5 kHz pas d'effet sur sonde 0
N Pm 08 152 cachalot 155 dB 1,5 kHz activité interrompue 2
N Pm 08 152 cachalot 145 dB 6,5 kHz pas de réaction 0
N Pm 09 141 cachalot 170 dB 1,5 kHz activité interrompue 2
N Pm 09 141 cachalot 150 dB 6,5 kHz activité altérée 1
N Pm 09 142 cachalot 165 dB 1,5 kHz activité interrompue 2
N Pm 09 142 cachalot 145 dB 6,5 kHz pas de réaction 0
N Pm 09 160 cachalot 170 dB 1,5 kHz activité interrompue 2
N Pm 09 160 cachalot 150 dB 6,5 kHz pas de réaction 0
Tableau 29 : Différents points expérimentaux obtenus lors d’expériences de CEE.
(N= programme 3S Norway, B = programme BRS Bahamas, S= programme SoCal)
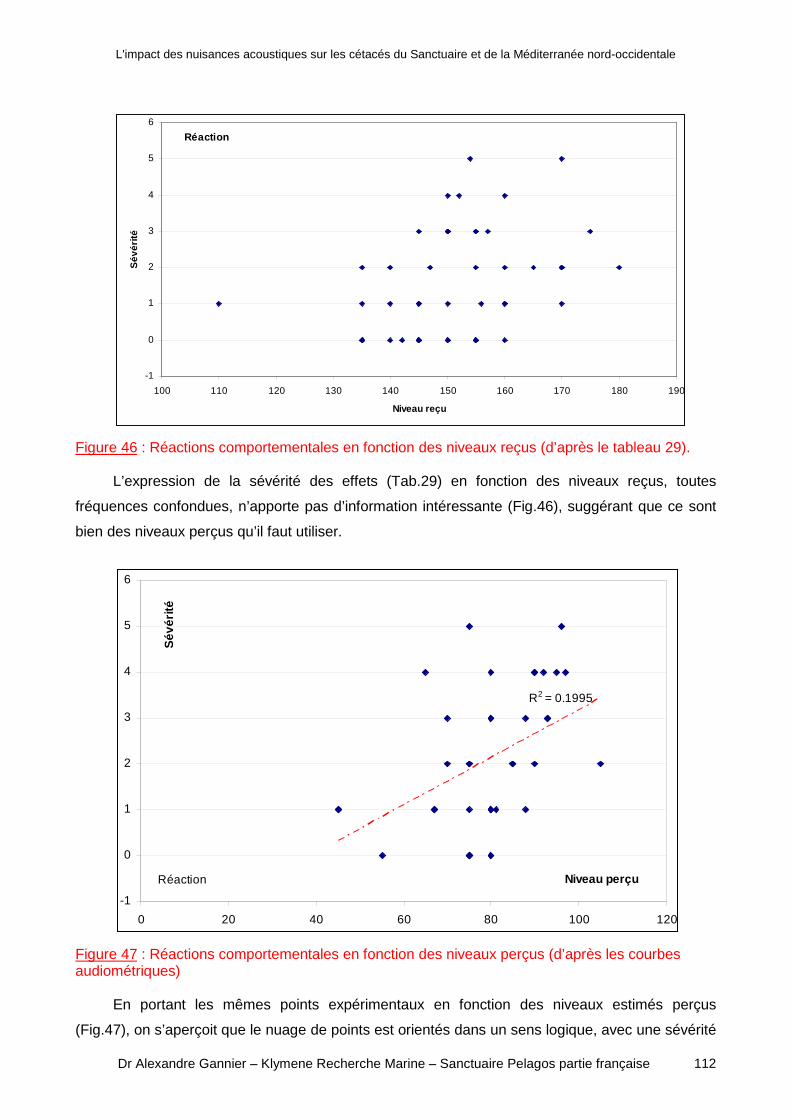
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 112
Réaction
-1
0
1
2
3
4
5
6
100 110 120 130 140 150 160 170 180 190
Niveau reçu
Sév
érité
Figure 46 : Réactions comportementales en fonction des niveaux reçus (d’après le tableau 29).
L’expression de la sévérité des effets (Tab.29) en fonction des niveaux reçus, toutes
fréquences confondues, n’apporte pas d’information intéressante (Fig.46), suggérant que ce sont
bien des niveaux perçus qu’il faut utiliser.
Réaction
R2 = 0.1995
-1
0
1
2
3
4
5
6
0 20 40 60 80 100 120
Niveau perçu
Sév
érité
Figure 47 : Réactions comportementales en fonction des niveaux perçus (d’après les courbesaudiométriques)
En portant les mêmes points expérimentaux en fonction des niveaux estimés perçus
(Fig.47), on s’aperçoit que le nuage de points est orientés dans un sens logique, avec une sévérité

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 113
de réaction accrue pour des niveaux perçus plus forts. On ne peut pas attendre une allure plus
précise compte-tenu d’une part de la nature hiérarchique de l’expression de la sévérité, et d’autre
part de l’imprécision des courbes audiométriques utilisées, en particulier celle qui concerne le
cachalot.
En ce qui concerne les impulsions de canons sismiques, des expériences d'exposition
contrôlée ont eu lieu dans le golfe du Mexique (Jochens et al., 2008), avec le taggage de 8
cachalots et l'insonification avec des canons pneumatiques, avec une pression de 14 bars et un
volume de 27 à 50 litres ; cette capacité correspond à des niveaux de sources calculés de 260
dBp-p @1m pour la bande 3-800 Hz (Miller et al., 2009). Les cachalots ont continué à chasser alors
qu'ils étaient exposés à des niveaux sonores de 135-160 dBp-p, par contre Miller et al. (2009) ont
observé que le taux de prédation des cachalots diminuait de 19% pendant l'exposition aux canons
pneumatiques. On peut donc affirmer que l’activité avait été altérée (sévérité 1) pour ce niveau
reçu.
Les expériences réalisées sur les mysticètes (Rorqual bleu, commun et Mégaptère) n’ont
pas été reportées sur le graphe sévérité de réaction = f(niveau perçu), car notre audiogramme
synthétique de mysticètes est trop peu fiable pour effectuer la transformation niveau reçu/niveau
perçu. Pour les fréquences de 3,5 kHz, des signaux de sonar reçus à 140-150 dB ne provoquent
en général pas de réaction chez ces mysticètes. On peut estimer que les niveaux perçus sont très
certainement inférieurs à 80 dB pour cette fréquence, chez les deux balénoptères.
Les expériences d’exposition contrôlée sont encore assez restreintes : elles ne traitent
pratiquement que des signaux de sonar à basse-moyenne fréquence, et peu d’espèces de petits
delphinidés. Néanmoins, ces résultats sont les seuls utilisables pour exprimer la sévérité des
dérangements en fonction des niveaux sonores. On peut ainsi proposer avec le graphe ci-dessus
(Fig.47) des seuils correspondants à chaque classe principale de réaction comportementale,
même si ces limites restent imprécises :
- pour des niveaux inférieurs à 60 dB, il y aurait peu de dérangements d’activité,
- pour des niveaux entre 60 et 75 dB, il y a peu d’évasions et beaucoup d’interruption d’activité,
- pour des niveaux entre 75 et 90 dB, il y a des interruptions d’activité, des évasions, des fuites,
- pour des niveaux supérieurs à 90 dB, il y a toujours évasion, fuite ou panique.
Il y a bien sûr des points expérimentaux qui ne collent pas à cette échelle ; même si l’on
augmentait le nombre d’expériences, on aurait encore une grande variabilité des réactions
observées en fonction des niveaux perçus. En effet, ces phénomènes comportementaux
dépendent de beaucoup de facteurs (contexte social, facteurs environnementaux, historique de la
transmission sonore). Donc, il ne faut pas attendre d’un seuil de niveau sonore qu’il définisse
précisément un seul type de réaction. Le seuil donne plutôt un ordre d’idée de ce qui peut se
passer lorsqu’une intensité sonore de niveau donné est perçue par un groupe de cétacés.
Les points expérimentaux disponibles pour les petits delphinidés sont encore trop peu nombreux.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 114
d) Autres effets comportementaux observés
En dehors des effets comportementaux observés lors des programmes d’expériences CEE,
d’autres réactions ont été documentées lors d’émissions de sons intenses : une grosse base de
résultats existe pour ce qui concerne les prospections sismiques (Stone & Tasker, 2006),
également il y a plusieurs résultats intéressants sur les réactions provoquées par les plates-formes
pétrolières sur les mysticètes, en plus il y a plusieurs cas d’échouages multiples en lien avec des
sonars pour lesquels aucune lésion traumatique n’a été mise en évidence, donc qui relèveraient
simplement de phénomènes comportementaux. Contrairement aux expériences CEE, ces résultats
ne sont généralement pas assortis d’estimation des niveaux sonores reçus.
• Réactions constatées lors de prospections sismiques
Les prospections sismiques industrielles autour des îles Britanniques ont permis d’identifier
des réactions comportementales sur beaucoup d’espèces (Stone & Tasker, 2006), dont une
diminution du taux de détection due à l’évasion des cétacés à l’approche de la source sonore
(§.B.1d). Cette évasion est également mise en évidence par des graphes de comparaison des
distances d’observation des cétacés (Fig.48) : il y a une très forte augmentation des distances
minimales d’observation des petits odontocètes (marsouins et dauphins). Par exemple, à peine
20 % des petits odontocètes détectés sont vus à moins de 500 m du navire en cas de tir sismique,
contre environ 50 % en période d’inactivité des tirs. Les différences s’amenuisent à partir de 3 km
de distance. Ainsi une forte proportion des petits cétacés évacuent la zone qui se trouve à moins
de 500 m de la source41. La proportion de baisse de détection à courte distance est un peu
moindre pour les globicéphales, cachalots et mysticètes (Stone & Tasker, 2006).
Figure 48 : Augmentation des distances d’observation pour les petits odontocètes, entre unesituation de tir sismique (carré noir) et de non-activité sismique (triangle blanc).
41 En considérant un niveau de source de 255 dB, et un « abattement » de 20 dB dû à la directivité vers lebas des impulsions pneumatiques, on a des niveaux sonores de l’ordre de 195 dBp-p à la distance de 100 m

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 115
Une autre statistique qui permet de percevoir le phénomène d’évasion est celle portant sur
l’orientation des cétacés par rapport au bateau (ou de la source) : il y a une diminution d’un facteur
de 2,5 de la proportion des cétacés se rapprochant du navire, et multiplication par deux de la
proportion de cétacés s’éloignant du navire. Cette statistique est valable pour toutes les espèces à
l’exception du cachalot (Stone & Tasker, 2006).
Pour les tirs sismiques, McCauley et al. (2000) indiquent que pour des Mégaptères le niveau
suscitant une évasion est de l’ordre de 169-176 dBp-p (baleines en migration) ou de l’ordre de 155
dBp-p (baleines avec nourrisson au repos). Ces valeurs sont en accord avec des résultats plus
anciens (Richardson et al., 1995). Ces niveaux sonores provoquant l’évasion expliquent pourquoi
des mysticètes s’éloignent à plus d’un km d’un navire de prospection sismique.
Ces éléments, en accord avec notre §B.1d, suggèrent que le niveau sonore reçu de 160
dBp-p est intéressant pour définir le seuil d’évasion des mysticètes, y compris le Rorqual commun.
Avec Thompson et al. (2013), nous avons une indication pour un seuil de 170 dBp-p valable pour
les marsouins, et par extension aux petits odontocètes.
Rappelons que les seuils d’évitement/évasion cités pour des bruits à large bande et basse
fréquence, tels que les forages pétroliers, sont d’environ 115-120 dB pour des mysticètes comme
la baleine grise ou la baleine du Groenland (Richardson et al., 1995).
• Evasion ou fuite de dauphins face à des bruits de s onar
Les évasions de dauphins lors de transmission de sonars MF ou HF sont régulièrement
observés, mais pas souvent rapportées avec le formalisme scientifique, sauf pour quelques
événements qui ont conduit à un échouage massif de dauphins.
En premier lieu, on a le cas des dauphins d'Electre des îles Hawaii en juillet 2004 (Southall
et al., 2006) : un groupe important s’est rapproché de la côte au point de presque s’échouer.
L’intervention humaine a permis d’éviter l’échouage massif. D’après des calculs approximatifs
effectués après coup, des niveaux sonores de 138-149 dB d’un sonar à moyenne fréquence ont pu
conduire les dauphins à se réfugier dans cette baie peu profonde.
Un cas d’échouage atypique qui s’est produit en janvier 2005 en Caroline du Nord et qui
concernait en majorité 33 globicéphales semble être relié de près à la transmission d’un sonar à
moyenne fréquence de l’US Navy (Hohn et al., 2006). Certaines lésions dans les graisses
mandibulaires suggèrent que quelques individus auraient pu être blessés par les intensités
sonores, mais pas les autres. La grégarité est toujours un facteur aggravant les échouages.
Le cas des Dauphins communs de Cornouailles en 2008 se rapproche de l’événement de
Hawaii : un grand groupe s’est trouvé piégé dans un estuaire à cause d’un grand exercice naval ;
26 dauphins ont fini par s’échouer (Jepson et al., 2013). Il n’y a pas eu de calcul des niveaux
sonores, plusieurs sonars ayant été utilisés dans les 24 heures précédant le piégeage des
dauphins.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 116
Le dernier échouage de ce type a été répertorié à Madagascar en mai 2008 et concerne à
nouveau des dauphins d’Electre qui se sont réfugiés dans une baie peu profonde et ont fini par
s’échouer peu à peu. D’après l’enquête menée, la cause la plus probable du piégeage du groupe
de dauphins a été une prospection océanographique utilisant un sonar multi-faisceaux de
fréquence 12 kHz.
Enfin, cette fois sur la côte Pacifique des USA, en 2003, plusieurs marsouins se sont
échoués morts ou vivants peu de temps après qu’un destroyer de l’US Navy ait parcouru le détroit
de Puget Sound en utilisant son sonar moyenne fréquence. Des orques ont également été
observées avec un comportement très inhabituel, pendant les transmissions sonar.
• Expériences en bassin sur un Marsouin commun
Kastelein et al. (2006 ; 2008) ont montré en bassin des zones d’évitement correspondant à
des niveaux perçus de 75-85 dB, pour des sons purs de différentes fréquences.
Ces quelques exemples démontrent qu’une réaction comportementale suffit à provoquer des
échouages massifs accidentels lors de l’emploi de sonars à moyenne et haute fréquence. Il y a de
nombreux cas d’échouages de cétacés vivants qui sont demeurés inexpliqués, mais qui pourraient
être dus à l’emploi sans précaution d’engins acoustiques puissants. La combinaison d’une forte
intensité sonore perçue par les cétacés et d’une configuration topographique favorable à un
piégeage du groupe sont les deux facteurs cruciaux. Une autre condition porte sur la durée
nécessaire de la transmission sonore : en effet, un groupe de delphinidés, se trouvant souvent à
plusieurs kilomètres d’un littoral, ne va pas s’échouer avant d’avoir parcouru la distance qui le
sépare de la côte. Par conséquent, une émission sonore puissante, mais brève, pourra provoquer
une fuite panique qui ne durera que quelques minutes, et donc qui ne sera pas suffisante pour
entraîner un échouage. La grégarité et des facteurs « psychologiques » propres aux espèces sont
également des facteurs à considérer : notons que les globicéphales, les péponocéphales, et
certains petits delphinidés comme D. delphis sont des candidats préférentiels aux échouages
accidentels dus aux sources sonores. Par contre, des espèces comme Tursiops truncatus ou
Orcinus orca ne semblent pas particulièrement vulnérables à ce type d’événement.
On observe que des petits delphinidés océaniques de l’espèce P. electra réagissent à un
niveau de 140-150 dB en se rapprochant dangereusement du rivage, alors que les expériences de
CEE (aux Bahamas) avaient montré qu’un niveau d’environ 135 dB à moyenne fréquence
provoquait une réaction de regroupement et d’immobilisation. Ces niveaux reçus correspondraient
à des niveaux perçus de l’ordre de 40 à 50 dB pour cette espèce. Mais des espèces telles que le
Dauphin de Risso semblent ne pas réagir à des niveaux reçus de 145 dB (environ 55 dB perçus).
Ainsi, en complément des résultats obtenus par CEE qui suggéraient peu d’interruption
d’activité en dessous d’un niveau perçu de 60 dB (mais avec une insuffisance de points

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 117
expérimentaux pour les petits delphinidés), nous prenons en compte un niveau seuil de 45 dB
perçus pour l’apparition de réactions comportementales chez certaines espèces.
e) Synthèse des effets des nuisances sonores
On peut classer les effets des pollutions sonores dans l’ordre d’une gravité croissante
(gravité considérée à l’échelle d’un individu et non à celle d’une population), en mettant en regard
les niveaux sonores reçus, les niveaux perçus ou des doses sonores accumulées, selon la
métrique qui est la plus adaptée au type d’effet considéré.
Notre synthèse de la littérature scientifique sur les effets des intensités sonores met en
évidence d’une part, les atteintes aux facultés auditives mesurés par des expériences en captivité
(principalement sur le Grand dauphin), et d'autre part, les effets comportementaux observés lors
de travaux en milieu naturel, avec ou sans CEE. Par ailleurs, la prévision des effets traumatiques
aigus et létaux observés indirectement sur des cétacés s’appuie sur des études expérimentales
non spécifiques menées dans le passé pour évaluer la résistance des mammifères aux explosions.
Si les trois catégories d’effets se produisent pour des ondes sonores impulsives et non-
impulsives, elles ne peuvent pas être prédites en utilisant les mêmes métriques.
- D’abord, les effets traumatiques aigus affectent différents organes surtout à la suite de
transitoires provenant d’explosions chimiques ou d’impulsions pneumatiques. C’est donc une
métrique basée sur des niveaux pic-pic reçus qui convient en premier lieu (dBp-p re1µPa), avec
une expression complémentaire basée sur un niveau d’énergie sonore reçue, qui permet de
prendre en compte la durée de l’impulsion (dBSEL re1µPa2.s).
- Pour les effets auditifs, pertes permanentes PTS, ou pertes temporaires TTS, les seuils
peuvent être exprimés en niveau d’exposition sonore perçue SEL pour tenir compte de la durée de
l’insonification (dBSEL re1µPa2.s). Pour une durée de 1 seconde, le niveau d’exposition (en dBSEL)
est égal au niveau de pression sonore, et pour chaque doublement de la durée d’exposition, à
niveau d’exposition égal, le niveau de pression diminue de 3 dB. Pour les sons impulsifs uniques,
une métrique complémentaire est basée sur les niveaux pic-pic perçus (dBp-p re1µPa).
- Pour les effets comportementaux, ce sont des niveaux sonores perçus qui doivent servir de
métrique à la prévision de la sévérité des réactions, en valeur pic-pic pour les impulsions, et en
valeur efficace pour les sons stationnaires. Une égalité de niveaux perçus ne signifie pas des
réactions de sévérité égale pour des sons de différentes fréquences, car une perception auditive
égale ne signifie pas que la gêne (notion psychologique) ou la peur, et par conséquent la réaction,
soit la même. Le niveau perçu est une métrique plus adéquate que le niveau sonore reçu.
Avant d’exprimer une gradation des effets (Tab.30), il faut rappeler que la précision d’une
telle prévision ne peut pas être bonne, en raison de la variabilité inter-individuelle dans la
perception des sons, et des éléments contextuels qui échappent aux descriptions expérimentales
qui ont été réalisées. Ainsi, la précision des seuils exprimés ici est en général moins bonne que +/-
10 dB.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 118
Sons non-impulsifs ImpulsionsPerturbation seuil niveau
perçuseuil niveau
reçuseuil niveau
perçuseuil niveau
reçu Dérangement léger sans évasion 40 à 60 dB so ? so
Interruption d’activité à évasion 60 à 75 dB so ? so
Evasion lente à fuite 75 à 90 dB so 70-80 dBp-p ? mysticètes160-170 dBp-p
Fuite avec ou sans panique > 90 dB so ? so
Perte temporaire d’audition 100 dB SEL so 100 dB p-p
70 dB SEL
220 dB p-p
Perte définitive d’audition 115 dB SEL so 115 dB p-p 235 dB p-p
Croissance des micro-bulles d’azote so > 190 dB so ?
Risque de traumatisme aigü létal so > 220 dB so > 230 dBp-p
Tableau 30 : Gradation des effets induits par les pollutions sonores (à partir de résultats relevés enbassin et en liberté sur plusieurs espèces).
(so = sans objet ; SEL en re1µPa2.s ; autres niveaux en dB re1µPa)
Commentaires du tableau
° des perturbations peu sévères (altérations d’activité, regroupement) peuvent apparaître pour
des niveaux perçus de 40 à 60 dB ;
° des dérangements significatifs (interruption d’activité, voire évasion) interviennent pour des
niveaux perçus de 60 à 75 dB , voire moins pour les petits delphinidés et petits ziphiidés ;
° des dérangements sévères de type interruption d’activité, évasion ou fuite interviennent pour
des niveaux de 75 à 90 dB perçus, variabilité selon les espèces et le contexte ; pour les séries
d’impulsions, 160 dBp-p reçu provoquent l’évasion de mysticètes et 70 à 80 dBp-p perçu
provoquent l’évasion de marsouins et petits delphinidés ;
° des évacuations de type fuite interviennent des niveaux perçus supérieurs à 90 dB avec un fort
risque de panique ;
° pour les sons non-impulsifs, très peu de TTS ont été observés avec des doses inférieures à
100 dBSEL ; le critère de TTS pour les impulsions est de 100 dBp-p pour une impulsion unique
ou de 70 dBSEL, mais il y a trop peu d’expériences réalisées pour que le critère soit utilisable,
surtout dans le cas de séries d’impulsions multiples ;
° le critère de PTS retenu ici est le seuil provoquant un TTS significatif (supérieur à 6 dB),
augmenté de 15 dB, valeur aboutissant à des TTS persistants ; il y a donc une marge de
sécurité de plusieurs décibels par rapport aux véritables pertes d’audition définitives ;
° d’après les expériences, une croissance des bulles peut intervenir dès 190 dB dans un tissu
sursaturé en azote, et 210 dB dans le cas de tissus non sursaturés ;

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 119
3) Typologie retenue pour la gradation des effets d es nuisances sonores
L’impact des nuisances sonores sur les cétacés se décline selon une échelle qui est
généralement une fonction de l’intensité des nuisances sonores auxquelles les individus sont
exposés. Les effets ne sont pas fondamentalement différents de ceux qui sont observés chez les
humains et chez les mammifères terrestres : allant de la gêne légère jusqu’à la surdité
permanente, ou pour des sons extrêmement forts, à des lésions aigües aux cavités aériennes de
l’organisme. Certains effets comportementaux sont par contre spécifiques aux cétacés, et ont des
conséquences parfois mortelles : pour des niveaux sonores perçus modérés ou assez forts, et en
présence d’une topographie défavorable (proximité d’une côte ou d’une baie piègeante), les
cétacés peuvent s’échouer vivants, puis mourir. Par ailleurs, certains cétacés plongeurs profonds
comme les ziphiidés sont vulnérables à des accidents de décompression lorsqu’ils sont obligés
d’interrompre leur cycle de sonde complexe, à la suite de niveaux sonores excessifs. Des
embolies gazeuses et graisseuses ont été diagnostiquées à plusieurs reprises à la suite
d’échouages atypiques liés à des exercices de sonars militaires.
La gradation des effets des nuisances sonores sur les cétacés doit donc prendre en compte
les deux aspects des phénomènes cités ci-dessus : d’une part, une sévérité croissante des effets
pour des niveaux sonores reçus ou perçus de plus en plus forts ; d’autre part, des effets
potentiellement mortels découlant de conséquences comportementales à des niveaux sonores
modérés à assez forts.
a) Rationalité fondamentale de l’approche
Notre approche vise à prévoir l’impact d’une pollution sonore sur un peuplement de cétacés
en déterminant les effets subis par les individus des différentes espèces, en fonction de leur
distance par rapport à la source au moment où celle-ci émet et pendant le temps où elle émet.
L’approche est possible à partir de la connaissance des paramètres de la source (puissance,
fréquence, cycle d’émission) et de sa localisation dans l’espace et dans le temps. L’aspect
temporel est très important car dans la plupart des régions et notamment en Méditerranée nord-
occidentale, le peuplement de cétacés varie en fonction de la saison, voire même en fonction du
moment de la journée.
L’approche est proposée dans un cadre quasi-statique : elle établit les effets provoqués par
les intensités sonores au moment de l’émission de la source et dans les secondes qui suivent. Elle
prévoit les mouvements d’évasion provoqués par les intensités sonores, mais ne prend pas en
compte ces mouvements pour la détermination des niveaux sonores suivants, et donc pour
l’intégration des effets produits à l’échelle de plusieurs minutes ou plusieurs heures.
La rationalité de cette étude reprend celle qui avait été exposée par Gannier & Mifsud
(2011), en intégrant dans la synthèse de l’état de l’art issue des nombreuses connaissances
nouvelles publiées depuis 2010 :

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 120
- nouveaux audiogrammes de cétacés,
- nouveaux travaux sur les pertes auditives provoquées par les sons,
- nouveaux résultats d’expériences d’exposition controlée,
- nouvelles publications sur la problématique de l’effet des pollutions sonores sur les cétacés
Notre analyse a mis en évidence l’importance de l’expression des niveaux sonores en terme
de perception du son par les cétacés, aussi bien pour les principaux effets auditifs, que pour la
plupart sinon la totalité des effets comportementaux. La connaissance des audiogrammes est donc
un pré-requis pour toute évaluation de l’impact des nuisances sonores. En Méditerranée, les
audiogrammes sont connus grâce à des expériences pour 5 des 8 espèces communes du
peuplement, expériences réalisées à au moins une reprise dans des conditions satisfaisantes sur
un individu de l’espèce en question. Pour le Ziphius, on peut utiliser un audiogramme de
Mésoplodon de Blainville (Pacini et al., 2011) : du fait de l’écologie et de la taille voisines des deux
espèces, ainsi que de la similarité des répertoires vocaux, l’approximation est valable. Pour le
Cachalot, le problème est plus difficile, mais Ridgway & Carder (2001) ont obtenu une allure de
sensibilité auditive grâce à une expérience électro-physiologique sur un nourrisson. Un
raisonnement et quelques calculs nous permettront de proposer un audiogramme synthétique
approximatif pour cette espèce. Enfin, pour le Rorqual commun, la juxtaposition d’un raisonnement
fondé sur le bruit de fond en milieu marin et des travaux de modélisation mécanique de l’oreille
moyenne (Tubelli et al., 2012) permettent de proposer également un audiogramme synthétique.
Dans les deux derniers cas, évidemment, la précision n’est pas bonne.
Parmi les effets les plus dangereux des pollutions sonores sur les cétacés, il y les lésions
aigües sur des organes aériens ou auditifs causées par les contraintes mécaniques provoquées
dans les tissus par les ondes de choc ou les impulsions sonores de niveau extrêmement fort. Cette
catégorie d’effets souvent létaux découle d’une métrique des niveaux reçus, contrairement aux
précédentes. Avec les sources sonores connues, de tels effets ne peuvent se produire que si les
cétacés se trouvent à courte distance de la source au moment où elle commence à émettre.
D’autres réactions létales peuvent être provoquées par des niveaux sonores moindres, sur la
base d’effets comportementaux (obéissant à la métrique des effets perçus) ayant des
conséquences graves, comme un échouage vivant ou un accident de décompression.
La prévision des effets des pollutions sonores doit donc intégrer un ensemble de
phénomènes complexes, lésions aigües, fatigue auditive, réactions comportementales, et qui se
combinent parfois, pour aboutir à la détermination des conséquences variées sur les individus
exposés, allant de la mort à une légère altération de l’activité des cétacés. Il est important de
hiérarchiser d’abord les différents niveaux de perturbation.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 121
b) Les différents niveaux de perturbation
On établit ici les différents niveaux de perturbation en les hiérarchisant en fonction de leurs
conséquences vitales sur un individu ou un groupe d’individus. Cette hiérarchie n’est pas
forcément celle que l’on établirait en prenant pour base une « population » : par exemple, vue à
l’échelle d’une population, la mort brutale d’un individu est peut-être moins grave que le
déplacement de plusieurs centaines de cétacés à l’écart d’une aire favorable à la nutrition, pendant
des semaines.
On propose une hiérarchie des perturbations comprenant 5 niveaux :
- effet critique : pollution sonore conduisant à la mort d’un ou plusieurs individus,
- effet sévère : pollution sonore diminuant les facultés vitales d’un ou plusieurs individus,
- effet répulsif : pollution sonore conduisant des cétacés à quitter leur habitat favorable,
- effet significatif : pollution sonore diminuant l’efficacité d’une activité vitale des cétacés,
- effet non significatif : pollution sonore ressentie mais ne provoquant pas d’effet significatif.
Dans l’idéal, cette hiérarchie des effets serait liée de manière directe à une échelle
croissante des niveaux perçus et reçus. En réalité, c’est un peu plus compliqué, en raison de
couplage entre effets comportementaux et physiques, et des conséquences possibles des
évasions, comme les échouages.
• Pollution sonore conduisant à la mort d’un ou plusi eurs individus
La mort de cétacés à la suite de niveaux sonores élevés peut survenir selon différents
scénarios (Fig.49), directs ou indirects :
(1) le cétacé meurt sous l’effet d’une intensité extrêmement forte, immédiatement (sous
l’effet d’une lésion aigüe), ou plus tard (sous l’effet d’une incapacité permanente, comme une
surdité définitive). Ces effets graves découlent de niveaux reçus excessifs (lésions variées,
embolie provoquée par une diffusion gazeuse), ou de doses perçues excessives (PTS)
(2) le cétacé subit une lésion grave, comme une embolie gazeuse ou graisseuse, en raison
d’une réaction comportementale provoquée par un niveau perçu trop élevé (cas de l’accident de
décompression chez les ziphiidés)
(3) le cétacé subit une diminution prolongée de certaines de ses facultés (comme un TTS) ,
sous l’effet de doses sonores perçues excessives, qui le conduit à s’échouer vivant ; juste avant
l’échouage, son pronostic vital n’était pas engagé
(4) le cétacé, ou le groupe, est conduit à une évasion sous l’effet de niveaux perçus
excessifs ; dans un contexte géographique particulier, cette évasion aboutit à un échouage vivant,
avant lequel, le pronostic vital n’était pas engagé
Ces conséquences critiques peuvent être évitées en s’assurant que les cétacés sont absents
d’une zone critique (Z1) entourant la source et dont les limites sont déterminées par les niveaux
perçus ou reçus correspondant aux cas (1) et (2) ci-dessus. Ce risque critique est équivalent au
« level-A catch » des réglementations US.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 122
Figure 49 : Scénarios pouvant conduire à la mort de cétacés en raison d’une insonification.
• Pollution sonore diminuant les facultés vitales des individus
Dans cette catégorie, on a principalement les diminutions temporaires de facultés auditives
ou TTS, qui sont provoquées par des doses sonores perçues supérieures à des seuils qui ont été
déterminés par expérimentation (§B.2). Ces pertes de sensibilité auditive affectent surtout la bande
de fréquence immédiatement supérieure au bruit de la source sonore (par ex, entre 3,5 et 7 kHz si
c’est un sonar à 3,5 kHz qui a provoqué le problème). Si les insonifications sont voisines des seuils
d’apparition du TTS, les pertes durent quelques minutes, mais si la dose sonore est plus forte, les
pertes d’audition peuvent se prolonger plusieurs heures.
Même si le TTS n’affecte pas toute la bande audible du cétacé, il détériore la faculté
essentielle de l’individu à entendre ses conspécifiques, des prédateurs éventuels, ou des proies,
ou encore une menace quelconque. Il y a des cas également où le TTS peut diminuer la capacité
d’écholocalisation d’une espèce, lorsque la source est à une fréquence un peu inférieure à la
bande de fréquence utilisée par l’odontocète pour la détection.
Ces conséquences sévères peuvent être évitées en s’assurant que les cétacés sont absents
d’une zone d’insécurité (Z2) entourant la source et dont les limites sont déterminées par les doses
perçues correspondant au risque de TTS. Ce risque est équivalent au « level-B catch » des
réglementations US.
• Pollution sonore conduisant les cétacés à quitter u n habitat favorable
Mis à part lorsqu’ils sont en voyage entre deux habitats favorables, circonstances dans
lesquelles ils empruntent des routes quand même proches de leur habitat fondamental, les

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 123
cétacés sont localisés dans des sites où leur activité de prédation est optimisée, ou bien leur
activité de reproduction (accouplement, mise-bas, élevage des jeunes), ou bien encore où ils sont
le plus en sécurité possible vis-à-vis des prédateurs.
Par conséquent, si les forts niveaux sonores provoquent une évasion des cétacés en dehors
du site qu’ils ont choisi, cela implique toujours un coût écologique. Ce coût peut être
éventuellement estimé, assez facilement si les cétacés ont dû interrompre leur activité de
nourrissage. En effet, le déplacement forcé des cétacés peut dans certains cas empêcher la
poursuite de leur prédation, les proies étant souvent distribuées de façon hétérogène.
Enfin, il y a d’autres raisons pour lesquelles une évasion peut avoir des conséquences
fortes : s’il s’agit d’une fuite prolongée, qui peut aboutir à l’éclatement d’un groupe au détriment de
nourrissons, ou de couples mère-nourrisson, exposant ces individus à des dangers
supplémentaires, voire à la mort, s’il s’agit de nouveau-nés incapables de rejoindre leur groupe.
Dans un contexte géographique défavorable (baie fermée), une fuite prolongée peut aussi
provoquer un échouage massif, avec mort probable des individus ; on se retrouve dans le
scénario 4 ci-dessus (Fig.49).
La zone correspondant à ces risques de gravité variable est appelée zone d’évasion (Z3).
Dans le cas de sources très puissantes, elle s’étend à des distances importantes car elle
correspond à des niveaux sonores perçus relativement modérés.
• Pollution sonore diminuant l’efficacité d’une activ ité vitale des cétacés
On est ici dans le cas où les cétacés subissent les niveaux sonores sans chercher à
s’évader de la zone : soit parce que le niveau perçu n’est pas excessif, soif parce qu’il à un
caractère intermittent et prévisible, les cétacés préfèrent demeurer dans un site favorable et
continuer leur activité. Cependant, on mesure des variations de comportement qui traduisent une
gêne, comme une séquence de respiration modifiée, ou un cycle de sonde raccourci, ou un
nombre de proies capturées plus faible. Il y a donc un impact de la pollution sonore sur les
cétacés, significatif, mais il est assez faible si la pollution sonore ne se prolonge pas pendant des
semaines ou des mois.
La zone correspondant à ces altérations de comportement est appelée zone de gêne (Z4).
Elle s’étend souvent à des distances de plusieurs dizaines de kilomètres car elle correspond à des
niveaux sonores perçus modérés.
• Pollution sonore ressentie ne provoquant pas d’effe t significatif
En deçà d’un certain niveau, la source sonore devient un élément du bruit de fond dans la
région. S’il n’y a pas d’effet significatif auditif ou comportemental, la source sonore contribue à
l’élévation du niveau de bruit ambiant ; elle peut donc entraîner un masquage supplémentaire des
communications des cétacés. Certains effets peuvent être mesurés, comme une élévation du
niveau des appels des cétacés, ou une modification de leur rythme ou de leur fréquence. L’impact

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 124
de la pollution sonore peut être qualifié de léger ou de non significatif. Les niveaux sonores
émanant de la source se propagent à des distances d’autant plus grandes qu’ils sont à basse
fréquence.
La zone correspondant à ces effets légers est appelée zone de perception (Z5). Elle s’étend
souvent à des centaines de kilomètres autour de la source.
Figure 50 : Zonage des perturbations encourues par les cétacés au cours d’une insonification.
Une bonne partie de la démarche de prévision des impacts consiste à dimensionner ces
différentes zones.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 125
C\ Résultats
La prévision des impacts des nuisances sonores sur les cétacés (et aussi les autres
mammifères) s’appuie nécessairement sur quatre ensembles d’éléments, outre bien sûr la
connaissance des caractéristiques de la source sonore :
o la nature du peuplement de cétacés insonifié (diversité et densité du peuplement), ainsi
que les éléments comportementaux attachés à chaque espèce, en particulier ceux
touchant aux cycles de plongée, et aux vitesses de déplacement,
o les énergies sonores reçues par les cétacés, telles qu’elles sont déduites par l’application
d’un modèle de propagation sonore aux sons issus de la source,
o les caractéristiques de sensibilité acoustiques des cétacés, ou audiogrammes, qui
permettent de déterminer quels sont les niveaux sonores perçus par chaque espèce,
o un modèle de prévision des effets produits par les énergies sonores perçues et reçues par
les cétacés.
Il s’agit donc d’une affaire complexe. Au départ, il est parfois extrêmement difficile d’obtenir
les caractéristiques acoustiques précises d’une source sonore puissante, pour les engins
militaires, mais pas seulement. Même si les éléments sur la nature du peuplement ne sont pas le
sujet précis de cette étude, on sait qu’ils ne sont pas forcément aussi précis qu’il serait
nécessaire42.
Pour ce qui concerne la propagation, l’application d’un modèle est un domaine en soi : des
ouvrages et des logiciels permettant de résoudre plus ou moins précisément le problème
complexe de la propagation existent, les logiciels étant le plus souvent utilisables par des
spécialistes. La propagation du son en milieu marin est un phénomène très sensible aux
conditions physiques régnant dans le milieu au moment de la transmission des sons par la source,
en particulier dépendant de la nature du fond, de l’état de la surface de la mer, et surtout de la
structure verticale de densité de la colonne d’eau (Urick, 1975). La prise en compte correcte de la
bathycélérimétrie43 dans l’eau conditionne la bonne détermination de la propagation et donc des
niveaux reçus ; celle-ci peut-être obtenue grâce à des relevés climatologiques disponibles dans
certaines bases de données, ou même par des mesures in situ. A l’aide de la bathycélérimétrie, un
logiciel spécialisé permet de déterminer la propagation du son depuis la source jusqu’au récepteur
(dans notre cas, les cétacés). A défaut d’approche sophistiquée de ce type, il est possible d’avoir
une approximation de la propagation sonore en utilisant simplement la loi sphérique : pour cette
étude, c’est cette solution simplifiée qui est adoptée, car le travail sur la propagation du son dans
l’eau n’est pas au coeur de notre recherche (cet aspect est développé par les industriels du
secteur).
42 Ils peuvent être obtenus par Pelagos à partir des multiples études réalisées depuis une vingtaine d’années

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 126
La prévision des effets en fonction des énergies sonores reçues ou perçues est au coeur de
notre problématique : la mise en oeuvre de l’approche présentée au chapitre précédent demande
la transformation des niveaux reçus en niveaux perçus, à l’aide d’audiogrammes qui sont
disponibles ou qui seront développés. Ensuite, la prévision des impacts est une application de la
synthèse bibliographique que nous venons de réaliser, chaque catégorie d’impact étant reliée à
des seuils ou à des fourchettes de niveaux sonores.
Notre modèle de prévision des effets découle de l’examen des résultats expérimentaux
(§B.2). Il reprend les principes exposés lors de la publication d’un travail antérieur (Gannier &
Mifsud, 2011), mais avec des évolutions rendues possibles par les nombreuses publications
récentes.
Figure 51 : Problématique de la prévision de l’impact d’une insonification sur les cétacés.
43 Profil d’évolution verticale de la célérité du son dans la colonne d’eau

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 127
1) Proposition d'un modèle de prévision des effets
Le modèle de prévision des effets découle directement de la typologie définie plus haut
(§B.3), qui organise les effets de sévérité croissante (de la perception sans dérangement de la
nuisance sonore à la lésion immédiatement létale) en fonction des niveaux sonores perçus, pour
ce qui est des effets comportementaux, des doses sonores perçues, pour ce qui est des effets
auditifs, et des niveaux sonores reçus, pour ce qui est des conséquences physiologiques non
auditives.
Le modèle proposé définit les effets de manière qualitative, en fonction de seuils sonores
atteints ou dépassés, et non quantitative : par exemple, le dépassement d’un seuil de dose sonore
donnée entraîne un risque de perte auditive temporaire (TTS)44, mais le modèle ne détermine pas
l’étendue et la durée de celle-ci. Ou bien, le modèle prévoit un risque d’évasion, mais ne détermine
pas la vitesse de l’évasion.
Dans bien des cas, les effets des sons non impulsifs et impulsifs obéissent à des métriques
très similaires, les unes étant exprimées en niveau sonore efficace (dBrms), et les autres en
niveau sonore pic-pic (dBp-p). Dans le cas des pertes de sensibilité auditive, les deux métriques
sont exprimées en niveau d’exposition sonore, mais qui ne se calculent pas de la même façon.
Cependant, nous préférons séparer la présentation du modèle de prévision pour les deux
catégories de bruit : les niveaux de connaissance ne sont pas les mêmes (plus d’expériences ont
été réalisées pour les sons non impulsifs), et on utilise un modèle de prévision dans un cas
particulier, qui appartient à l’une ou l’autre des deux catégories de sons.
De plus, la compréhension du problème est parfois ardue, sans qu’il soit besoin de la
complexifier en présentant les deux cas de figure à la fois.
44 Un modèle quantitatif donnerait la valeur du TTS en dB (par ex 20 dB) et sa persistance dans le temps.
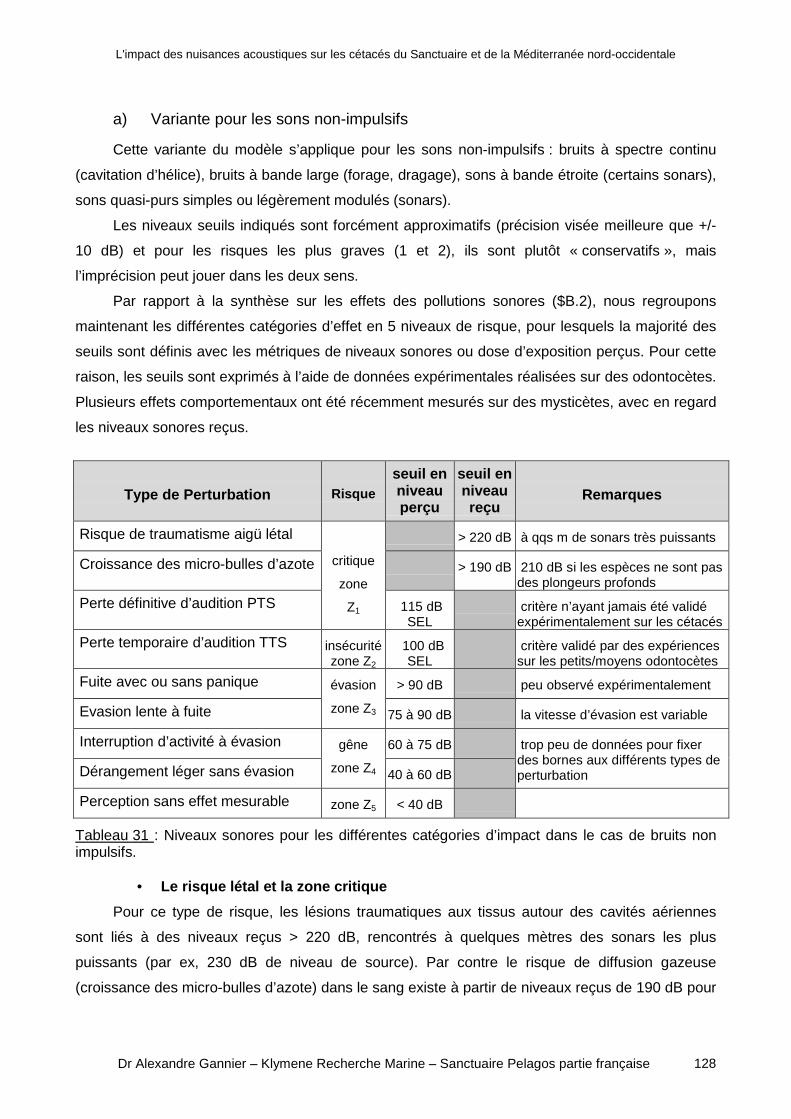
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 128
a) Variante pour les sons non-impulsifs
Cette variante du modèle s’applique pour les sons non-impulsifs : bruits à spectre continu
(cavitation d’hélice), bruits à bande large (forage, dragage), sons à bande étroite (certains sonars),
sons quasi-purs simples ou légèrement modulés (sonars).
Les niveaux seuils indiqués sont forcément approximatifs (précision visée meilleure que +/-
10 dB) et pour les risques les plus graves (1 et 2), ils sont plutôt « conservatifs », mais
l’imprécision peut jouer dans les deux sens.
Par rapport à la synthèse sur les effets des pollutions sonores ($B.2), nous regroupons
maintenant les différentes catégories d’effet en 5 niveaux de risque, pour lesquels la majorité des
seuils sont définis avec les métriques de niveaux sonores ou dose d’exposition perçus. Pour cette
raison, les seuils sont exprimés à l’aide de données expérimentales réalisées sur des odontocètes.
Plusieurs effets comportementaux ont été récemment mesurés sur des mysticètes, avec en regard
les niveaux sonores reçus.
Type de Perturbation Risqueseuil enniveauperçu
seuil enniveaureçu
Remarques
Risque de traumatisme aigü létal > 220 dB à qqs m de sonars très puissants
Croissance des micro-bulles d’azote > 190 dB 210 dB si les espèces ne sont pasdes plongeurs profonds
Perte définitive d’audition PTS
critique
zone
Z1 115 dBSEL
critère n’ayant jamais été validéexpérimentalement sur les cétacés
Perte temporaire d’audition TTS insécuritézone Z2
100 dBSEL
critère validé par des expériencessur les petits/moyens odontocètes
Fuite avec ou sans panique > 90 dB peu observé expérimentalement
Evasion lente à fuite
évasion
zone Z3 75 à 90 dB la vitesse d’évasion est variable
Interruption d’activité à évasion 60 à 75 dB
Dérangement léger sans évasion
gêne
zone Z4 40 à 60 dB
trop peu de données pour fixerdes bornes aux différents types deperturbation
Perception sans effet mesurable zone Z5 < 40 dB
Tableau 31 : Niveaux sonores pour les différentes catégories d’impact dans le cas de bruits nonimpulsifs.
• Le risque létal et la zone critique
Pour ce type de risque, les lésions traumatiques aux tissus autour des cavités aériennes
sont liés à des niveaux reçus > 220 dB, rencontrés à quelques mètres des sonars les plus
puissants (par ex, 230 dB de niveau de source). Par contre le risque de diffusion gazeuse
(croissance des micro-bulles d’azote) dans le sang existe à partir de niveaux reçus de 190 dB pour

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 129
des cétacés sursaturés en azote, et de 210 dB pour les autres. Le niveau de 190 dB correspond à
une distance critique d’une centaine de mètres d’un sonar puissant.
Le risque de perte auditive définitive correspond à des doses d’exposition de 115 dBSEL (re
1µPa2.s) perçues. Par exemple, pour un cétacé ayant une sensibilité auditive de 65 dB à la
fréquence de la pollution sonore, cela donne une dose reçue de 180 dBSEL. Cette dose est
l’énergie reçue à une distance d’environ 300 m suite à une transmission d’une seconde (niveau
sonore de 180 dB) d’un sonar très puissant. Nous aurions dans ce cas virtuel une zone critique
circulaire de 300 m de rayon. Si le son durait 4 sec, le niveau sonore de 174 dB reçu
correspondrait à un cercle de 630 m de rayon.
• Le risque de TTS et la zone d’insécurité
Ce type de risque est documenté pour les petits et moyens odontocètes et correspond à une
dose perçue supérieure à 100 dBSEL (re 1µPa2.s), ce seuil a un caractère approximatif. Dans le cas
de sons transmis selon un cycle comportant des temps de repos (cas de tous les sonars), le seuil
est « pessimiste » en considérant les temps d’émission mis bout à bout sur l’ensemble du cycle, et
il est « optimiste » si l’on ne considère que le temps de transmission d’un « chirp ».
Par exemple, pour un cétacé ayant une sensibilité auditive de 65 dB à la fréquence de la
pollution sonore, cela donne une dose reçue de 165 dBSEL. Cette dose est l’énergie reçue à une
distance d’environ 1800 m suite à une transmission d’une seconde (niveau sonore de 165 dB) d’un
sonar très puissant. Nous aurions dans ce cas virtuel une zone d’insécurité circulaire de 3,5 km de
diamètre. Si le son durait 4 sec, le niveau sonore de 159 dB reçu correspond à un cercle de 7 km
de diamètre. La zone d’insécurité définie par ce risque est vaste, dans le cas d’engins puissants.
• Le risque et la zone d’évasion
Le risque d’évasion dans ses différentes modalités (éloignement, évasion rapide, fuite avec
panique) est présent pour une large fourchette de niveaux perçus, entre 75 et 100 dB de niveau
sonore. Le point commun est que les cétacés quittent la zone insonifiée à ces niveaux. Dans le
haut de la fourchette on a les comportements de fuite rapide, avec panique, avec les risques
inhérents pour les individus vulnérables d’un groupe. Dans le bas de la fourchette, on a les
évasions avec des vitesses plus ou moins soutenues, allant d’un rythme normal de voyage, à une
allure rapide, non soutenable sur le long terme. Dans la zone des forts niveaux perçus, on peut
éventuellement observer des comportements différents « d’évasion », comme des cétacés tentant
de maintenir la tête hors de l’eau pour échapper au bruit intense. En général, ces niveaux forts ont
pour conséquence des comportements aériens visibles.
L’évasion est également possible pour des niveaux perçus inférieurs à 75 dB, selon le
contexte social, écologique, et la psychologie de l’espèce en question. On ne doit pas imaginer les
réactions comme homogènes, même pour une espèce donnée ; pour un niveau perçu donné, la
réaction comportementale d’un groupe de cétacés se situe sur une palette.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 130
Pour un cétacé ayant une sensibilité auditive de 65 dB à la fréquence de la pollution sonore,
le niveau perçu de 90 dB correspond à 155 dB reçu, soit une distance d’environ 5500 m dans le
cas d’un sonar très puissant (230 dB de niveau de source). Un niveau perçu de 75 dB correspond
lui à un niveau reçu de 140 dB, soit une distance de 30 km environ. Nous aurions dans ce cas
virtuel des zones d’évasion très étendues, correspondant à de nombreux groupes de cétacés.
Figure 52 : Rappel du zonage autour d’une source sonore.
• Altération d’activité et zone de gêne
On trouve à nouveau dans cette catégorie des effets comportementaux variables
correspondant à la large fourchette de niveaux perçus, entre 40 et 75 dB, et à la psychologie de
l’espèce. Les modifications de comportement altérant l’activité, et allant jusqu’à son interruption,
occupent quand même plutôt le haut de la fourchette, entre 60 et 75 dB perçus.
Pour ce qui est des balénoptères, comme le Rorqual commun, on observe que des niveaux
reçus de 150-160 dB à 3,5 kHz ne provoquent pas de changement significatif d’activité.
Pour un cétacé ayant une sensibilité auditive de 65 dB à la fréquence de la pollution sonore,
le niveau perçu de 60 dB correspond à 125 dB reçu, soit une distance d’environ 170 km dans le
cas d’un sonar très puissant (230 dB de niveau de source). Un niveau perçu de 75 dB correspond
lui à un niveau reçu de 140 dB, soit une distance de 30 km environ. Entre ces deux cercles
concentriques, sur une surface de plus de 80 000 km2, le modèle prévoit que de nombreux
groupes de cétacés auraient leur activité impactée par l’émission de ce sonar. La zone de gêne
concernerait potentiellement des milliers de cétacés.
Quand à la zone correspondant au niveau perçu de 40 dB, que nous avons définie comme
limite de la zone de gêne légère, il faudrait faire un calcul de propagation incluant l’absorption pour

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 131
en évaluer les frontières, mais elle s’étendrait à plusieurs centaines de km d’un sonar très
puissant.
• Zone de perception
Les niveaux perçus inférieurs à 40 dB correspondent à des distances supérieures à 500 km,
ce qui signifie qu’un engin de forte puissance transmettant des sons dans une bande de fréquence
où les cétacés ont une sensibilité auditive correcte (65 dB) est audible par les cétacés à l’échelle
du bassin nord-occidental.
On remarque que les niveaux perçus correspondant aux différentes zones de risque ont
potentiellement des étendues qui changent d’ordre de grandeur, ce qu’illustre l’exemple virtuel
d’un sonar de 230 dB de niveau de source, avec des cétacés ayant une sensibilité auditive de
65 dB à la fréquence de la source.
On est dans des distances inférieures au kilomètre pour la zone critique, de quelques
kilomètres pour la zone d’insécurité, puis on passe à des dizaines et des centaines de km pour les
zones d’évasion et de gêne. Ceci est naturellement dû à la nature de la loi de propagation
sphérique, qui donne une décroissance linéaire du niveau sonore en fonction du logarithme de la
distance, donc une décroissance lente à grande distance.
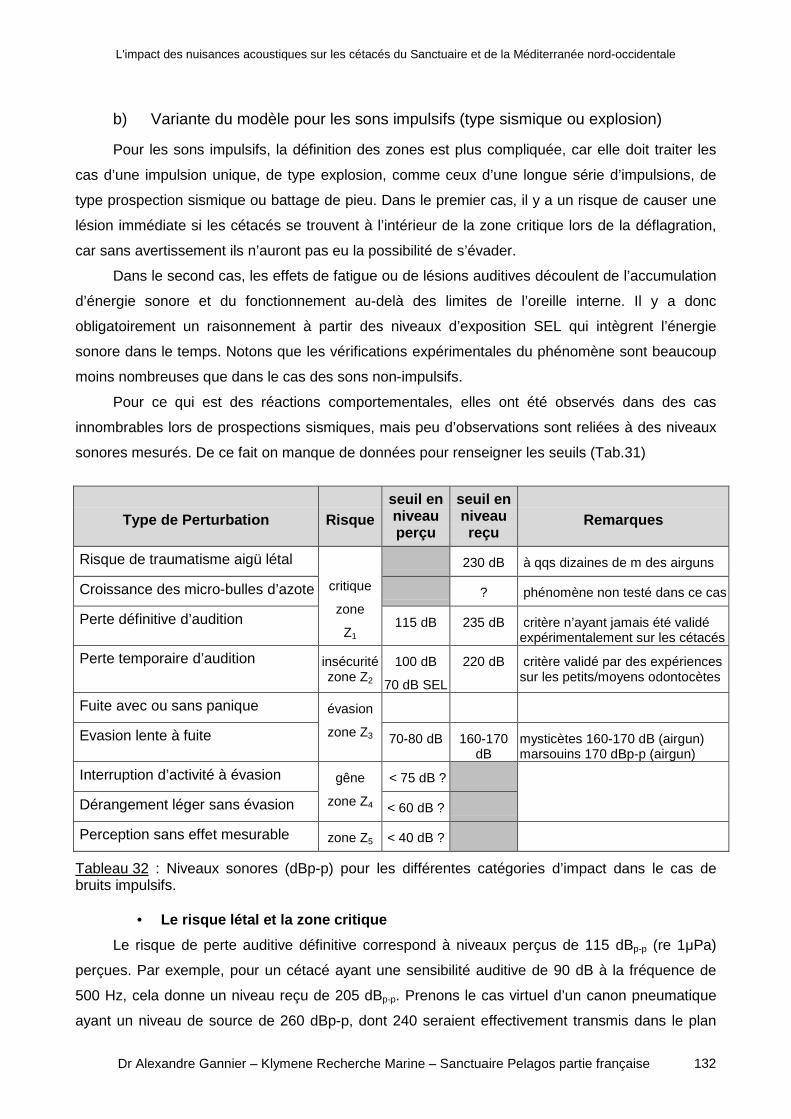
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 132
b) Variante du modèle pour les sons impulsifs (type sismique ou explosion)
Pour les sons impulsifs, la définition des zones est plus compliquée, car elle doit traiter les
cas d’une impulsion unique, de type explosion, comme ceux d’une longue série d’impulsions, de
type prospection sismique ou battage de pieu. Dans le premier cas, il y a un risque de causer une
lésion immédiate si les cétacés se trouvent à l’intérieur de la zone critique lors de la déflagration,
car sans avertissement ils n’auront pas eu la possibilité de s’évader.
Dans le second cas, les effets de fatigue ou de lésions auditives découlent de l’accumulation
d’énergie sonore et du fonctionnement au-delà des limites de l’oreille interne. Il y a donc
obligatoirement un raisonnement à partir des niveaux d’exposition SEL qui intègrent l’énergie
sonore dans le temps. Notons que les vérifications expérimentales du phénomène sont beaucoup
moins nombreuses que dans le cas des sons non-impulsifs.
Pour ce qui est des réactions comportementales, elles ont été observés dans des cas
innombrables lors de prospections sismiques, mais peu d’observations sont reliées à des niveaux
sonores mesurés. De ce fait on manque de données pour renseigner les seuils (Tab.31)
Type de Perturbation Risqueseuil enniveauperçu
seuil enniveaureçu
Remarques
Risque de traumatisme aigü létal 230 dB à qqs dizaines de m des airguns
Croissance des micro-bulles d’azote ? phénomène non testé dans ce cas
Perte définitive d’audition
critique
zone
Z1115 dB 235 dB critère n’ayant jamais été validé
expérimentalement sur les cétacés
Perte temporaire d’audition insécuritézone Z2
100 dB
70 dB SEL
220 dB critère validé par des expériencessur les petits/moyens odontocètes
Fuite avec ou sans panique
Evasion lente à fuite
évasion
zone Z3 70-80 dB 160-170dB
mysticètes 160-170 dB (airgun)marsouins 170 dBp-p (airgun)
Interruption d’activité à évasion < 75 dB ?
Dérangement léger sans évasion
gêne
zone Z4 < 60 dB ?
Perception sans effet mesurable zone Z5 < 40 dB ?
Tableau 32 : Niveaux sonores (dBp-p) pour les différentes catégories d’impact dans le cas debruits impulsifs.
• Le risque létal et la zone critique
Le risque de perte auditive définitive correspond à niveaux perçus de 115 dBp-p (re 1µPa)
perçues. Par exemple, pour un cétacé ayant une sensibilité auditive de 90 dB à la fréquence de
500 Hz, cela donne un niveau reçu de 205 dBp-p. Prenons le cas virtuel d’un canon pneumatique
ayant un niveau de source de 260 dBp-p, dont 240 seraient effectivement transmis dans le plan

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 133
horizontal : nous aurions dans ce cas une zone critique circulaire de 55 m de rayon, à l’intérieur de
laquelle le cétacé serait rendu partiellement sourd en une seule impulsion. Dans la cas d’une série
d’impulsions, ce disque augmente de diamètre à chaque doublement du nombre d’impulsions.
• Le risque de TTS et la zone d’insécurité
Ce type de risque correspond à un niveau perçu supérieur à 100 dBp-p. Par exemple, pour un
cétacé ayant une sensibilité auditive de 90 dB à la fréquence 500 Hz, cela donne un niveau perçu
de 190 dBp-p. Nous aurions dans ce cas virtuel une zone d’insécurité circulaire de 700 m de
diamètre, pour une impulsion unique. La zone d’insécurité définie par ce risque est en réalité plus
vaste dans le cas d’une prospection sismique, en raison du nombre d’impulsions transmises à
chaque minute. On calcule alors en utilisant le seuil de 70 dB re 1µPa2.s : connaissant le niveau
SEL d’une impulsion à la source, on en déduit à quelle distance le niveau SEL vaut 70 dB. A
chaque doublement du nombre d’impulsions, le niveau d’exposition à une distance donnée
augmente de 3 dB.
• Le risque et la zone d’évasion
Le risque d’évasion dans ses différentes modalités (éloignement, évasion rapide, fuite avec
panique) est présent pour une large fourchette de niveaux pic-pic perçus, entre 75 et 100 dB. Pour
des niveaux proches de 100 dBp-p, on observe probablement des comportements de fuite rapide,
ou de panique, peut-être avec des réactions d’immobilisation pour certaines espèces, ou des
cétacés tentant de maintenir la tête hors de l’eau pour échapper au bruit intense.
Pour un cétacé ayant une sensibilité auditive de 90 dB à la fréquence de la pollution sonore,
le niveau perçu de 90 dB correspond à 180 dB reçu, soit une distance d’environ 1000 m dans le
cas d’un airgun industriel (240 dB de niveau de source dans le plan horizontal). Cette distance
d’évasion rapide est cohérente avec les réactions observées sur les cétacés lors des prospections
sismiques industrielles.
Il y a un manque de données expérimentales pour documenter cette zone dans le cas des
sons impulsifs.
• Altération d’activité et zone de gêne
On trouve à nouveau dans cette catégorie des effets comportementaux variables
correspondant à la large fourchette de niveaux perçus, entre 40 et 75 dB, et à la psychologie de
l’espèce.
Un résultat intéressant a montré que des Cachalots soumis à des impulsions d’airgun, avec
des niveaux reçus de 140-160 dBp-p, avaient une altération d’activité mais sans interruption, sauf
peut-être pour un individu. Avec ces niveaux reçus, on se trouve vraisemblablement dans le haut
de la fourchette 40-75 dB, sans doute entre 60 et 75 dB perçus : en prenant 150 dBp-p pour

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 134
l’altération d’activité, cela correspond à une sensibilité auditive de 75 – 90 dB à 500 Hz, ce qui est
plausible.
La distance correspondant à 150 dBp-p reçus, pour un airgun de 240 dBp-p, est au delà de
10 km, en tenant compte de l’absorption, ce qui est considérable.
• Zone de perception
Pour un cétacé ayant une sensibilité auditive de 90 dB à 500 Hz, les niveaux perçus à 40
dBp-p correspondent à des distances supérieures à 300 km, ce qui signifie qu’un canon
pneumatique de puissance industrielle est audible par les cétacés à l’échelle du bassin nord-
occidental, au minimum.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 135
c) Audiogrammes de référence
L’utilisation du modèle de prévision nécessite un audiogramme de référence pour chaque
espèce : une courbe de sensibilité auditive qui soit représentative de la moyenne des individus de
l’espèce. Nous donnons ci-dessous les audiogrammes utilisés pour les 8 espèces du peuplement
de Méditerranée nord-occidentale. Cinq de ces espèces ont fait l’objet d’expériences d’audiométrie
en bassin (les delphinidés), les audiogrammes utilisés sont des versions lissées de l’audiogramme
initialement publié, à l’aide du programme excel, extrapolés jusqu’à la fréquence de 100 Hz, et
éventuellement modifiés si cela est justifié. Une correction systématique sera de baisser de 5 dB le
seuil d’audition lorsqu’il est obtenu par méthode électrophysiologique AEP. L’audiogramme du
Ziphius sera remplacé par un audiogramme de Mésoplodon de Blainville obtenu par AEP (Pacini
et al., 2011). L’audiogramme de Cachalot sera basé sur la courbe de sensibilité électro-
physiologique obtenue par Ridgway & Carder (2001) sur un nourrisson, et calé en valeur absolue
sur le bruit ambiant par temps calme. L’audiogramme de Rorqual commun sera basé sur la courbe
de sensibilité modélisée par Tubelli et al. (2012).
• Audiogramme de Dauphin bleu et blanc
Le seul audiogramme connu pour le Dauphin bleu et blanc a été obtenu par Kastelein et al.
(2003), par méthode comportementale sur un individu juvénile réhabilité, dans un bassin isolé avec
peu de bruit de fond. Les signaux expérimentaux ont été des sons sinusoïdaux de 2 s dans le
domaine 0,1-160 kHz. Aucune modification n’est nécessaire pour que cet audiogramme serve de
référence.
Figure 53 : Audiogramme de Dauphin bleu et blanc (d’après Kastelein et al., 2003).
Le domaine de meilleure audition du Dauphin bleu et blanc est entre 29 et 123 kHz, la
décroissance de sensibilité entre 16 et 0,5 kHz est de 11 dB/octave.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 136
• Audiogramme de Dauphin commun
Le seul audiogramme publié de Dauphin commun (D. delphis) est celui de Popov & Klishin
(1998), obtenu par méthode électro-physiologique ABR, avec des signaux sinusoïdaux très courts.
Le dauphin était un specimen de mer Noire, échoué vivant et malade, et qui a été maintenu dans
un bassin 4 jours avant que sa mort intervienne. Les fréquences de 10 à 152 kHz ont été testées.
Nous apportons les modifications suivantes : baisse de 5 dB des seuils trouvés, continuation de
l’audiogramme vers les basses fréquences avec une pente de 11/dB par octave.
Figure 54 : Audiogramme de Dauphin commun (d’après Popov & Klishin, 1998, modifié).
Le domaine de meilleure audition du Dauphin commun se situe entre 45 et 90 kHz.
• Audiogramme de Grand dauphin
Parmi les nombreux audiogrammes de Grand dauphin (T. truncatus) existants, c’est celui de
Johnson (1966, in Au et al., 2000) qui continue à faire référence. Il a été obtenu par méthode
comportementale, avec des signaux sinusoïdaux de 3 sec. Le dauphin était un jeune adulte. Les
fréquences de 75 Hz à 150 kHz ont été testées. Aucune modification n’est nécessaire pour que cet
audiogramme serve de référence.
Le domaine de meilleure audition du Grand dauphin est très large, entre 10 et 100 kHz, la
décroissance de sensibilité entre 10 et 0,5 kHz est de 9.7 dB/octave.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 137
Figure 55 : Audiogramme de Grand dauphin (d’après Johnson, 1966 in Au et al., 2000).
• Audiogramme de Dauphin de Risso
Un des deux audiogrammes publiés de Dauphin de Risso (G. griseus) est celui de Nachtigall
et al. (2005), obtenu par méthode électro-physiologique AEP et suivi EFR, avec des signaux
sinusoïdaux très courts. Le dauphin était un nouveau-né d’Atlantique, échoué vivant, qui a été
maintenu en bassin durant le temps de l’expérience, mais qui est mort après. Il y avait un bruit de
fond très faible dans le bassin, pour les fréquences testées : de 4 à 110 kHz. Nous apportons les
modifications suivantes : baisse de 5 dB des seuils trouvés, continuation de l’audiogramme à partir
de 8 kHz vers les basses fréquences avec une pente de 11/dB par octave.
Figure 56 : Audiogramme de Dauphin de Risso (d’après Nachtigall et al., 2005, modifié).
Le domaine de meilleure audition du dauphin de Risso est entre 20 et 100 kHz.
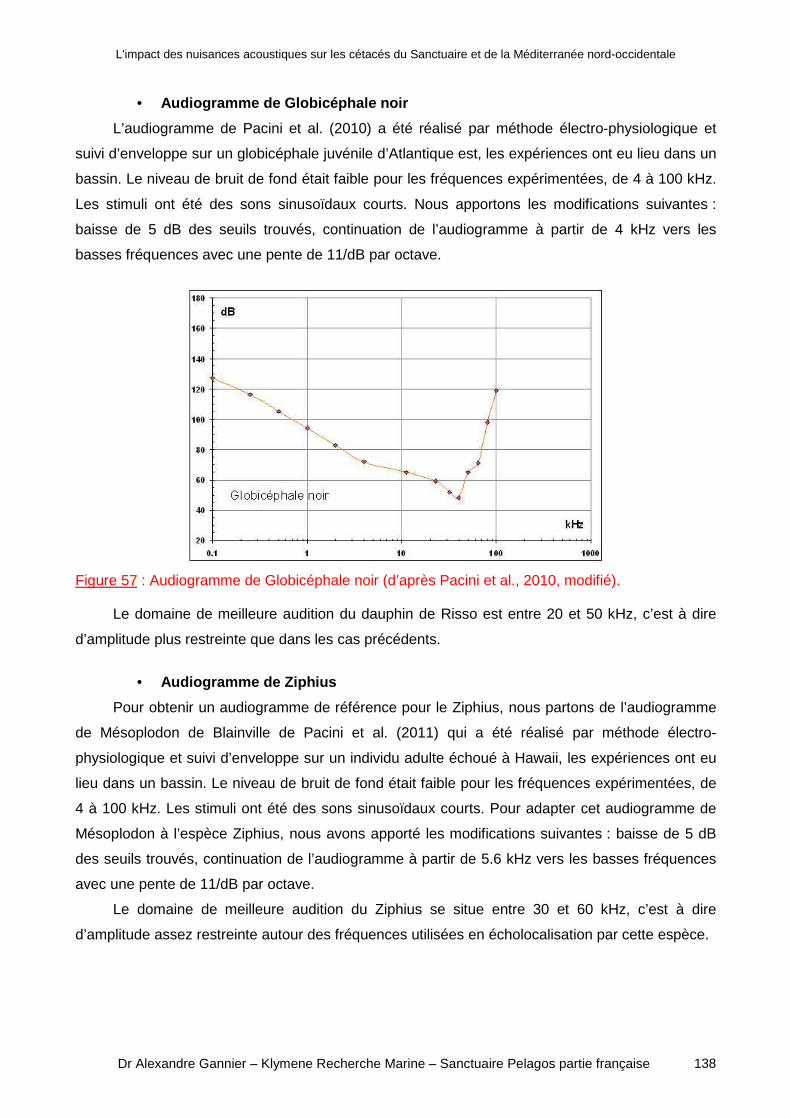
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 138
• Audiogramme de Globicéphale noir
L’audiogramme de Pacini et al. (2010) a été réalisé par méthode électro-physiologique et
suivi d’enveloppe sur un globicéphale juvénile d’Atlantique est, les expériences ont eu lieu dans un
bassin. Le niveau de bruit de fond était faible pour les fréquences expérimentées, de 4 à 100 kHz.
Les stimuli ont été des sons sinusoïdaux courts. Nous apportons les modifications suivantes :
baisse de 5 dB des seuils trouvés, continuation de l’audiogramme à partir de 4 kHz vers les
basses fréquences avec une pente de 11/dB par octave.
Figure 57 : Audiogramme de Globicéphale noir (d’après Pacini et al., 2010, modifié).
Le domaine de meilleure audition du dauphin de Risso est entre 20 et 50 kHz, c’est à dire
d’amplitude plus restreinte que dans les cas précédents.
• Audiogramme de Ziphius
Pour obtenir un audiogramme de référence pour le Ziphius, nous partons de l’audiogramme
de Mésoplodon de Blainville de Pacini et al. (2011) qui a été réalisé par méthode électro-
physiologique et suivi d’enveloppe sur un individu adulte échoué à Hawaii, les expériences ont eu
lieu dans un bassin. Le niveau de bruit de fond était faible pour les fréquences expérimentées, de
4 à 100 kHz. Les stimuli ont été des sons sinusoïdaux courts. Pour adapter cet audiogramme de
Mésoplodon à l’espèce Ziphius, nous avons apporté les modifications suivantes : baisse de 5 dB
des seuils trouvés, continuation de l’audiogramme à partir de 5.6 kHz vers les basses fréquences
avec une pente de 11/dB par octave.
Le domaine de meilleure audition du Ziphius se situe entre 30 et 60 kHz, c’est à dire
d’amplitude assez restreinte autour des fréquences utilisées en écholocalisation par cette espèce.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 139
Figure 58 : Audiogramme de Ziphius (d’après l’audiogramme de Mésoplodon de Blainville dePacini et al., 2011, modifié).
• Audiogramme synthétique de Cachalot
Pour obtenir un audiogramme de référence pour le Cachalot, nous partons des courbes de
sensibilités de Ridgway & Carder (2001) qui ont été établies sur un nourrisson en bassin, par
méthode électro-physiologique. Le domaine de meilleure réponse est de 8 à 20 kHz. Nous avons
relevé le spectre de bruit de fond par temps calme (Richardson et al., 1995), auquel nous avons
rajouté 25 dB de rapport critique (Au & Hastings, 2008). Nous avons ensuite calé le domaine de
meilleure audition sur ce bruit de fond, et prolongé l’audiogramme vers les basses fréquences par
une pente de 11 dB/octave. Pour les hautes fréquences, nous avons tenu compte d’une sensibilité
assez bonne jusqu’à 40 kHz, et prolongé la courbe jusqu’au point (100 kHz, 140 dB).
Figure 59 : Audiogramme synthétique de Cachalot (élaboré d’après des données variées).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 140
Le domaine de meilleure audition du Cachalot se situerait ainsi entre 4 et 40 kHz, une plage
assez large des moyennes et hautes fréquences, conformes aux fréquences d’écholocalisation de
cette espèce.
Naturellement, cet audiogramme synthétique est une référence approximative temporaire.
• Audiogramme synthétique de Rorqual commun
Pour le Rorqual commun, nous avons également dû opérer par raisonnement d’après des
données expérimentales diverses, la principale étant la modélisation de l’oreille moyenne de
Rorqual de Minke par Tubelli et al. (2012), qui montre que la meilleure audition de cette espèce se
trouverait entre 1 et 2,5 kHz, avec un domaine de 30 Hz à 8 kHz (domaine à – 40 dB), assez bien
couplé au domaine des vocalisations de cette espèce. Nous reprenons cette courbe de sensibilité
en la translatant vers les basses fréquences de 2 octaves, et en la calant en sensibilité sur la
valeur de 65 dB, entre 200 Hz et 1 kHz, ce qui correspond à 50 dB de plus que les sensibilités les
meilleures constatées chez les odontocètes à ces fréquence. Mais le Rorqual commun utilise les
basses fréquences pour sa communication. De plus, nous avons continué la courbe de sensibilité
vers les basses fréquences en la plaçant parallèle au niveau de bruit de fond.
Figure 60 : Audiogramme synthétique de Rorqual commun (d’après Tubelli, et données variées).
Le domaine de meilleure audition du Rorqual commun se situerait ainsi entre 20 et 1000 Hz,
avec une audition déjà dégradée à partir de 2 kHz.
Cet audiogramme synthétique ne peut pas être tenu pour fiable. Il sera utilisé avec
précaution en attendant qu’une modélisation de l’oreille du Rorqual commun soit proposée par des
scientifiques spécialistes.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 141
2) Prévision des distances significatives pour les effets
Dans notre problématique, nous continuons notre cheminement inverse qui part des effets
provoqués sur les cétacés pour remonter jusqu’à la source sonore. Nous venons de déterminer le
moyen de passer des niveaux sonores reçus aux niveaux sonores perçus (les audiogrammes de
référence). Il nous reste maintenant à trouver le moyen de déterminer les niveaux sonores reçus à
partir des paramètres de la source : il s’agit d’utiliser une loi de propagation. Cette étape est
(comme les autres) cruciale, et en pratique elle peut être la cause de très grandes imprécisions sur
l’estimation de l’impact d’une pollution sonore au peuplement de cétacés, un facteur 10 sur le
nombre d’individus impactés, par exemple.
En effet, une erreur de 10 dB sur l’estimation du niveau reçu par un cétacé (donc une erreur
identique sur le niveau perçu) entraîne une mauvaise détermination des effets : on peut par
exemple se situer 5 dB au-dessus du seuil de TTS, ou 5 dB en-dessous. A l’inverse, si l’on prend
la détermination de la distance de sécurité correspondant à un niveau seuil, par exemple, celui
correspondant à l’apparition de TTS, une erreur de 10 dB sur détermination du niveau reçu
entraîne une évaluation complètement fausse du nombre de cétacés impactés.
Prenons le cas d’un sonar de 230 dB de niveau de source émettant 4 « pings » de 1
seconde de durée, à la fréquence de 3,5 kHz, sensiblement dans le plan horizontal, et d’une
population de dauphins avec une densité de 1 individu/km2 :
- le seuil de TTS de 100 dBSEL vaut 94 dB perçu pendant la durée de 4 secondes,
- 94 dB perçu correspond à un niveau sonore reçu de 189 dB (audiogramme Fig.53),
- les pertes par propagation de la source au niveau seuil sont de (230 – 189 =) 41 dB,
- avec une loi sphérique, ces pertes se produisent sur une distance de 112 m 45,
- la surface du disque où le niveau est supérieur au seuil vaut donc: 39 388 m2,
- le nombre de dauphins présents dans cette zone est : 0,04 soit 0 dauphin,
- en revanche, si les pertes par propagation ont été sous-estimées de 10 dB,
- le calcul donne : (idem avec 51 dB, donc une distance de 355 m) soit 0.4 dauphins,
- 10 dB d’erreur entraîne un facteur 10 sur la surface et le nombre de cétacés impactés.
Dans une problématique d’étude d’impact réelle, l’opérateur industriel ou étatique de la
source, ou le bureau d’étude en charge de l’étude, doit fournir un diagramme des niveaux
propagés autour de la source, donnant les niveaux sonores en fonction de la distance radiale et de
la profondeur. Cette propagation doit tenir compte des propriétés précises du milieu lors de la
pollution sonore : topographie et nature du fond, profil vertical de la célérité du son dans l’eau (ou
bathy-célérimétrie). Il existe des moyens simples et relativement peu coûteux de connaître la
45 41 = 20.log (distance) soit : distance = 10 (41/20) = 112 m, en négligeant l’absorption.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 142
bathy-célérimétrie in situ. Ces diagrammes de propagation sont élaborés grâce à des logiciels
spécifiques qui sont en possession de tous les opérateurs ou bureau d’études sérieux.
Dans une problématique d’étude générique (comme celle-ci), ou d’examen d’une étude
d’impact, l’emploi de logiciels de propagation n’est pas forcément nécessaire ou possible. Dans
ces cas, on est amené à utiliser un modèle idéal, et simple, qui est la loi de propagation sphérique.
En réalité, le son ne se propage pas au sein du milieu marin de façon aussi simple que ce qui est
décrit par la loi sphérique. Cependant, dans beaucoup de cas la loi sphérique fournit une
approximation utile, et en tout cas facile, de la réalité.
a) Utilisation d’un modèle de propagation sphérique
La loi de propagation sphérique suppose que l’énergie sonore d’une source se propage de
manière égale dans toutes les directions, au sein d’un milieu fluide homogène, de densité égale,
où il n’y a pas de frontière. La propagation sphérique s’écrit :
i = W / 4π.r2
où i est l’intensité sonore en watt/m2, W la puissance sonore en watt, et r le rayon de la sphère où
est mesurée i . Une fois convertie en pression sonore et exprimée en dB, cette relation devient :
Lr = Ls – 20.log (r)
où Lr est le niveau sonore reçu (dB 1µPa@1m) à la distance radiale r, Ls est le niveau de source
(dB re 1µPa@1m). En réalité, il faut ajouter un terme pour tenir compte de l’absorption sans l’eau,
avec le coefficient α qui dépend de la fréquence :
Lr = Ls – 20.log (r) – α.r
L’influence de l’absorption est faible pour les fréquences basses/moyennes et à des
distances de quelques kilomètres, car le coefficient α est de l’ordre de 0,2 – 0,3 dB/km à 5 kHz, ce
qui ne contribue que des quelques décibels aux pertes totales jusqu’à 10 km de distance de la
source. Par conséquent, pour les risques graves comme les atteintes à l’audition, on peut négliger
ce terme. Pour les très hautes fréquences et à longue distance, l’absorption contribue
substantiellement à la baisse du niveau sonore à longue distance, par ex. 16 dB/km à 50 kHz pour
une température de 15°C, et ne peut pas être négli gée.
Le terme [– 20.log (r) – α.r] est appelé « pertes par propagation ». On l’obtient facilement à
l’aide d’un tableur46. Cette loi exprime bien que la décroissance du niveau sonore est maximale
dans les premiers 500 mètres, là où les dangers seront les plus forts pour les cétacés (Fig.61).
Plus on s’éloigne des hypothèses de la loi sphérique, plus les résultats donnés par ce
modèle s’éloignent de la réalité, comme par exemple :
- au voisinage immédiat de la surface de l’eau,
- en milieu peu profond (plateau continental), et près du rivage,
- dans une colonne d’eau stratifiée, comme celle des mers tempérées en saison chaude.
46 Une feuille excel avec ce genre de calcul est fournie en annexe de cette étude.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 143
Figure 61 : Valeurs approximatives du coefficient d’absorption en fonction de la fréquence.
Figure 62 : Pertes par propagation sphérique en fonction de la distance.
En milieu peu profond, on observe souvent qu’une loi quasi-sphérique fonctionne assez bien :
Lr = Ls – 15.log (r) – α.r

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 144
b) Limites du modèle de propagation sphérique
La propagation du son dans l’eau s’écarte plus ou moins du modèle sphérique, mais le
champ d’applicabilité de cette loi simple n’est pas négligeable. Les principales causes
d’inexactitude du modèle sont : l’existence de frontières à l’espace aquatique (surface, sol), et la
stratification de la colonne d’eau, plus ou moins forte, avec des phénomènes associés au trajet
des ondes sonores, dans bien des cas. Ces propriétés peuvent causer de fortes erreurs dans les
niveaux sonores prévus à une certaine distance par le modèle sphérique. Il est donc nécessaire de
délimiter le champ de validité du modèle sphérique, en comparant ses prévisions à celle d’un
logiciel prenant en compte les conditions réelles de la propagation : par exemple un logiciel de
tracé de rayons.
Dans cette optique, nous avons modélisé la propagation avec un logiciel de tracé de rayons,
afin de comparer les résultats d’un modèle réaliste avec le modèle sphérique pour les différentes
périodes de l’année. Nous avons choisi les mois de janvier, mars, mai, juillet, septembre et
novembre et nous avons traité deux cas de propagation : le cas d’une source immergée à 10 m,
correspondant à un sonar de coque, et le cas d’une source immergée à 200 m, correspondant à
un sonar à immersion variable de type « poisson », comme il en existe pas mal en Europe. Dans
les deux cas nous avons déterminé les pertes par propagation pour un récepteur (dauphin,
baleine, etc.) immergé à 20 m.
La comparaison nous permet donc de délimiter les cas où la loi sphérique est valable à +/- 5
dB près, en fonction des saisons, et aussi d’indiquer dans quels cas les pertes par propagation
prédites par cette loi sont plus fortes que la réalité (du modèle), ou moins fortes.
• Source immergée à 10 m et récepteur à 20 m
Dans l’ensemble des cas, la détermination des pertes par propagation n’est pas correcte
au-delà d’une distance de 5 km (Fig.63).
Figure 63 : Pertes par propagation pour une source immergée à 10 m.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 145
Pour la saison froide (janvier à mars), les pertes sont un peu surestimées par la loi
sphérique, entre 5 et 20 km. Pour la saison chaude, seule la zone très proche de la source est
correctement décrite par la loi sphérique.
Figure 64: Pertes par propagation pour une source immergée à 10 m - zone proche.
On observe que pour les distances inférieures à 750 m, la description donnée par le modèle
sphérique est correcte, pour toutes les saisons. Au-delà de cette distance, pour la saison chaude,
les pertes par propagation réelles - modélisées - sont beaucoup plus élevées que celles
déterminées par une loi sphérique : en réalité, la propagation subit une très forte atténuation.
• Source immergée à 200 m et récepteur à 20 m
Pour une source immergée à 200 m, la détermination des pertes par propagation n’est
correcte pour aucune saison, dans la fourchette de 0 à 20 km (Fig.65). Mais en saison froide, la loi
sphérique surestime un peu la réalité modélisée, jusqu’à 10 dB environ.
Figure 65 : Pertes par propagation pour une source immergée à 200 m.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 146
En regardant de plus près, les pertes par propagation sont très fortes à très courte distance
pour toutes les saisons, engendrant une zone « sourde » jusqu’à 300 m, en surface. Puis,
l’ensemble des courbes rejoint la loi sphérique jusqu’à une distance de 2 km environ. Ensuite,
seules celles de la saison froide restent assez proches de la loi sphérique.
Figure 66 : Pertes par propagation pour une source immergée à 200 m (source proche).
En conclusion la loi de propagation sphérique donne :
- une approximation correcte de la réalité pour une source en sub-surface, jusqu’à la distance
de 750 m environ ; ensuite elle n’est convenable que pour la saison froide,
- une approximation correcte de la réalité pour une source semi-profonde, entre 300 et 2000 m
de distance , et au-delà pour la saison froide. Mais entre 0 et 300 m, les résultats de la loi
sphérique se situent du côté de la sécurité.
Il y a donc bien un domaine d’applicabilité pour la loi sphérique, dans le cadre de notre
problématique, il se situe dans le domaine des courtes distances47.
47 Attention, ces résultats sont valables pour la zone profonde nord-méditerranéenne.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 147
3) Application du modèle à un cas de forte nuisance sonore
Nous traitons ici de manière générique de l’impact d'un scénario de transmission de sonar
militaire en saison froide. Le choix de la saison froide, le mois de mars en l’occurrence, s’explique
par l’application d’une loi de propagation sphérique : nous venons de voir que les pertes par
propagation étaient assez bien déterminées par cette loi, lorsqu’il n’y a pas de stratification de la
colonne d’eau.
Nous employons le modèle d’impact que nous avons mis au point plus haut. Le réalisme de
l’étude impose une détermination en conditions dynamiques : l’impact sur les cétacés sera évalué
en intégrant un mouvement de réponse de leur part. C’est-à-dire qu’au cours de la transmission
des « pings » du sonar, les cétacés recevront les intensités successives en changeant de position
relative. La détermination de l’impact dans un cas dynamique nécessite l’utilisation d’un logiciel,
qui a été développé préalablement à l’exécution de cette étude.
En fonction d’une information largement diffusée (voir Prézelin, 2008), nous choisissons un
sonar de type « poisson » : les sonars à immersion variable sont en plein développement pour
toutes les marines européennes, et présents sur toutes les frégates anti-sous-marines de la
Marine Nationale (cf article Mer et Marine). Selon cet article, nous pourrions fixer la profondeur
d’immersion du sonar à 150 m ; cependant l’utilisation du modèle de propagation sphérique ne
demande pas explicitement de fixer une immersion.
Le choix des caractéristiques de fréquence du sonar fictif dont l’impact sera évalué n’est pas
simple : seuls quelques sonars US ont vu leurs caractéristiques publiées, et pour le reste nous
devons utiliser des informations non confidentielles. En fonction d’enregistrements réalisés près de
l’île de Port-Cros en 2008, nous choisissons un sonar de 4500 Hz fréquence fondamentale, qui
émet un « chirp » de 0,5 s de durée toutes les 12 secondes, pendant une durée totale de 2
minutes, puis qui se déplace pendant 5 minutes avant de recommencer.
La question du niveau de source du sonar est délicate, car ce type d’information n’est pas du
domaine public. Le sonar connu le plus puissant est le type SQS-53C (US), dont le niveau de
source est de 235 dB environ (pour une fréquence de 3,5 kHz) ; c’est un sonar de coque. Lors de
réunions publiques pour la partie française du Sanctuaire Pelagos, nous avons entendu des
militaires dire que les sonars français étaient « beaucoup moins puissants » que ce sonar
américain48. Les sonars « miniatures » inspirés des modèles militaires qui sont utilisés pour les
expériences 3S font 214 et 199 dB de niveau de source, respectivement pour les fréquences de 1-
2 kHz et 6-7 kHz (Miller et al., 2011). En fonction de ces informations, nous choisissons pour notre
sonar fictif un niveau de source de 220 dB.
Il reste ensuite à choisir un scénario de transmission pour notre exercice fictif.
48 Ce qui s’explique bien car des sonars de type « poisson » se doivent d’être beaucoup plus légers que dessonars de coque, car ils doivent être mis à la mer, remorqués, puis récupérés à bord du navire.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 148
a) Proposition d’un scénario de transmission de sonar
Il est vraiment très rare de trouver des informations publiques sur les scénarios de
transmission lors des exercices sonar, car ce sont des informations d’ordre tactique qui justement
ne doivent pas être divulguées. Les seuls cas bien documentés sont, à ma connaissance, les cas
qui sont liés à des échouages de ziphiidés. Nous avons par exemple le cas du scénario de
transmission d’un navire US, lors de l’échouage de Ziphius en janvier 2006, en mer d’Alboran
(Fig.67). Dans cet exemple, on voit que le sonar ratisse la zone au large d’Almeria, pendant plus
de 24 heures.
Figure 67 : Exemple de transmission sonar par un navire US en janvier 2006.(carte réalisée d’après un document diffusé avec autorisation).
Il est évident pour l’intérêt de l’étude que l’exemple de détermination d’impact doit se situer
dans un contexte intéressant pour le Sanctuaire Pelagos, donc mettre en jeu la zone Nord du
Sanctuaire et intégrer la présence de la base navale de Toulon. Les exercices sonar dans la
région nord-méditerranéenne sont en premier lieu effectués par la Marine Nationale, très
probablement. Des militaires français ont indiqué lors de réunions du Sanctuaire Pelagos que les
navires français n’opéraient pas avec des tactiques identiques à celles mis en oeuvre par les
navires US. N’ayant aucune connaissance des tactiques d’emploi des sonars par des frégates de
la Marine, nous allons établir un scénario imaginaire. Nous basons ce scénario sur un e-article de
Mer et Marine, un site d’information bien connu (www.meretmarine.com), qui contient quelques
vagues indications.
Notre scénario met en jeu une seule frégate, au mois de mars, avec le type de sonar fictif
décrit plus haut. On va supposer que le bateau effectue un entraînement à la recherche ASM, à
partir d’un point situé au large de Porquerolles, sur 2600 m de profondeur, pendant à peu près 5
heures. Le scénario reprend le cycle de transmission indiqué ci-dessus : le sonar émet un « chirp »
de 0,5 sec (niveau de source 220 dB à 4,5 kHz, faisceau aplati omnidirectionnel) toutes les 12
secondes, pendant une durée d’une minute (soit 2,5 sec par min). La frégate se déplace dans une
direction pendant 30 minutes à 10 noeuds avant de changer de direction, pour tenter de localiser

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 149
un hypothétique sous-marin. Ce scénario ne correspond à aucune réalité, mais pour notre
problématique, il n’est pas insensé.
b) Peuplement de cétacés considéré
Pour établir un peuplement de cétacés approximatif, on part des densités absolues ou
relatives issues de l’étude de population de 2001 (Gannier, 2006) et on enlève les espèces
migratrices qui sont absentes en fin d’hiver, c’est-à-dire le Globicéphale noir, ainsi que le Grand
dauphin (car la simulation s’effectue au large). On re-calcule des densités estivales à l’aide des
indices d’abondance relative, pour les espèces suivantes : cas du Cachalot, Dauphin de Risso.
Pour les espèces migratrices partielles, on pondère les densités issues de cette étude par un
coefficient d’abondance relative issu de l’étude saisonnière de Laran & Dulau (2007). Pour les
espèces nomades ou supposées sédentaires (Dauphin de Risso), on garde la densité estivale. On
suppose par souci de simplification que ce peuplement estimé s’applique pour la zone allant du
large jusqu’à l’isobathe 200 m.
Espèce Densité estivale(Gannier, 2006)
Densitécorrigée
Coefficienthiver / été
Densitéhivernale
Taille degroupe
Dauphinbleu et blanc
0,75 ind./km2
2,22 .10-2 g./km2
0,752,22 .10-2
0,50/1,1 = 0,45(Laran et, 2007)
0,32 ind./km2
2,0 .10-2 gr./km2
16
Dauphin deRisso
relative1,96 / 82,5
0,0170.0017 *
1 0,017 ind./km2
1,70 .10-3 gr./km2
10
Cachalot relative0,64 / 3,12
0,004 **0,004
0,32/1,89 = 0,17(Laran, 2006)
0,7 .10-3 ind./km2
0,7 .10-3 gr./km21
Rorqualcommun
0,018 ind./km2
1,13 .10-2 g./km2
0,0181,13 .10-2
0,5/5 = 0,1(Laran et, 2007)
0,0018 ind./km2
1,8 .10-3 gr./km2
1
Tableau 33 : Définition du peuplement local de cétacés
(*) Calculé comme ceci : 0,75 * 1,96/82,5 – arrondi à 0,017, taille de groupe prise à 10 ind.(**) Calculé comme ceci : 0,018 * 0,64/3,12 - arrondi à 0,004, taille de groupe prise à 1 ind.
Les densités hivernales estimées correspondent à la présence de :
- 2 groupes de 16 individus de Dauphin bleu et blanc pour 100 km2,
- 1,7 groupes de 10 individus de Dauphin de Risso pour 1000 km2,
- 0,7 individu Cachalot pour 1000 km2,
- 1,7 individu Rorqual commun pour 1000 km2.
En posant que la simulation de l’exercice sonar se déroule au sein d’une zone carrée de 60
kilomètres de côté (32,5 milles), on a une surface de 3600 kilomètres carrés. On trouvera ainsi
dans cette aire : 72 groupes de Dauphin bleu et blanc, 6 groupes de Dauphin de Risso, 2
Cachalots et 6 Rorquals.
C’est l’impact de la série de transmission du sonar sur ce peuplement que nous allons
déterminer maintenant, grâce au logiciel Sauvkipeu 1.6.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 150
c) Impact de la transmission sonar sur le peuplement
On évalue l’impact de la transmission sonar qui a été définie plus haut en plaçant au hasard
le peuplement de cétacés dans la zone de 60 km x 60 km que nous avons choisie, ce qui est fait
par le logiciel49.
Ensuite, on fait parcourir la zone d’étude par la source sonore, avec le scénario de
transmission défini plus haut (chirp de 0,5 sec, toutes les 12 secondes, niveau de source 220 dB à
4,5 kHz, pendant une durée de 1 minute ; déplacement de la source en ligne droite pendant 30
minutes à 10 noeuds, puis changement de direction).
Le logiciel calcule les niveaux sonores reçus et perçus par chacun des groupes de cétacés
dans la zone, les doses d’exposition, et en déduit les effets provoqués. Ce calcul s’effectue pas
par pas au fur et à mesure du parcours du sonar dans la zone. La sommation des effets est
tabulée à la fin du parcours simulé, une carte des déplacements des différents groupes est
disponible pour illustration des effets d’évasion.
Bien que le logiciel fonctionne avec une répartition au hasard des cétacés, nous le faisons
tourner à trois reprises pour chacune des 4 espèces de cétacé du peuplement. Pour chaque
espèce, le logiciel donne accès aux informations suivantes :
- une carte montrant le parcours de la source sonore, avec les positions occupées par les groupes
de cétacés au cours de la transmission,
- les doses sonores reçues et perçues par chacun des groupes à l’issue du scénario,
- l’évolution des niveaux sonores perçus et reçus au cours du scénario, pour chacun des groupes.
• Impact sur le Rorqual commun
La densité de population aboutit à la présence de 6 individus dans la zone au moment du
scénario (Fig.68). Les vitesses d’évasion choisies sont : (i) éloignement 2 m/s (+/- 0,5), (ii) fuite
5 m/s (+/- 1). L’audiogramme de synthèse évalue que la sensibilité du Rorqual commun à la
fréquence de 4,5 kHz est très mauvaise (seuil > 140 dB), aussi les baleines ne perçoivent
quasiment pas le son du sonar, même lorsque la bateau passe à quelques kilomètres. Donc, les
doses perçues sont très faibles (20 dB), même si les doses reçues sont assez élevées (plus de
170 dB pour les deux derniers individus.
Selon notre modèle, les réactions comportementales des Rorquals communs à la
transmission de ce sonar sont nulles.
Notons que nos résultats de simulation sont confortés par les résultats expérimentaux
obtenus lors des CEE (SoCal 2010 et 2012), pendant lesquels des Rorquals communs n’ont
montré aucune réaction à des niveaux reçus de 150 à 160 dB pour la fréquence de 3,5 kHz.
49 Le logiciel Sauvkipeu a été développé sur fonds propres par Klymene Recherche Marine, depuis 2010.
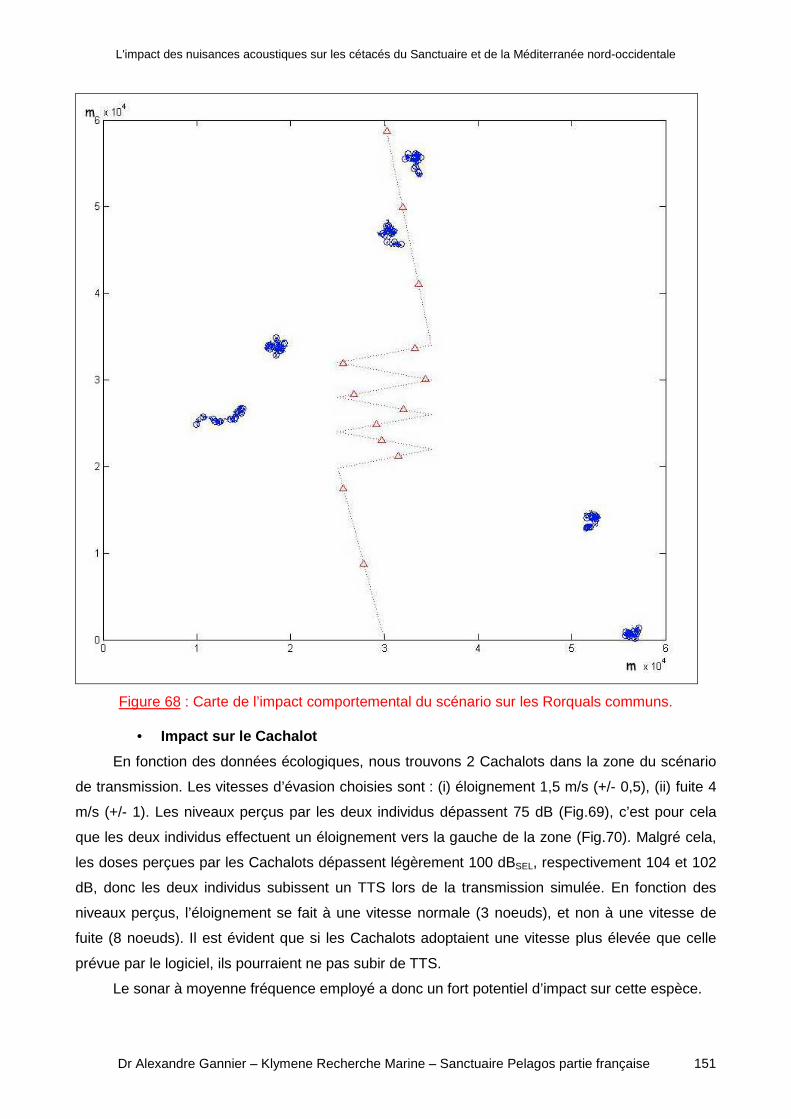
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 151
Figure 68 : Carte de l’impact comportemental du scénario sur les Rorquals communs.
• Impact sur le Cachalot
En fonction des données écologiques, nous trouvons 2 Cachalots dans la zone du scénario
de transmission. Les vitesses d’évasion choisies sont : (i) éloignement 1,5 m/s (+/- 0,5), (ii) fuite 4
m/s (+/- 1). Les niveaux perçus par les deux individus dépassent 75 dB (Fig.69), c’est pour cela
que les deux individus effectuent un éloignement vers la gauche de la zone (Fig.70). Malgré cela,
les doses perçues par les Cachalots dépassent légèrement 100 dBSEL, respectivement 104 et 102
dB, donc les deux individus subissent un TTS lors de la transmission simulée. En fonction des
niveaux perçus, l’éloignement se fait à une vitesse normale (3 noeuds), et non à une vitesse de
fuite (8 noeuds). Il est évident que si les Cachalots adoptaient une vitesse plus élevée que celle
prévue par le logiciel, ils pourraient ne pas subir de TTS.
Le sonar à moyenne fréquence employé a donc un fort potentiel d’impact sur cette espèce.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 152
Figure 69 : Niveaux perçus par les Cachalots au cours du scénario de transmission.
Figure 70 : Carte de l’impact comportemental du scénario sur les Cachalots.
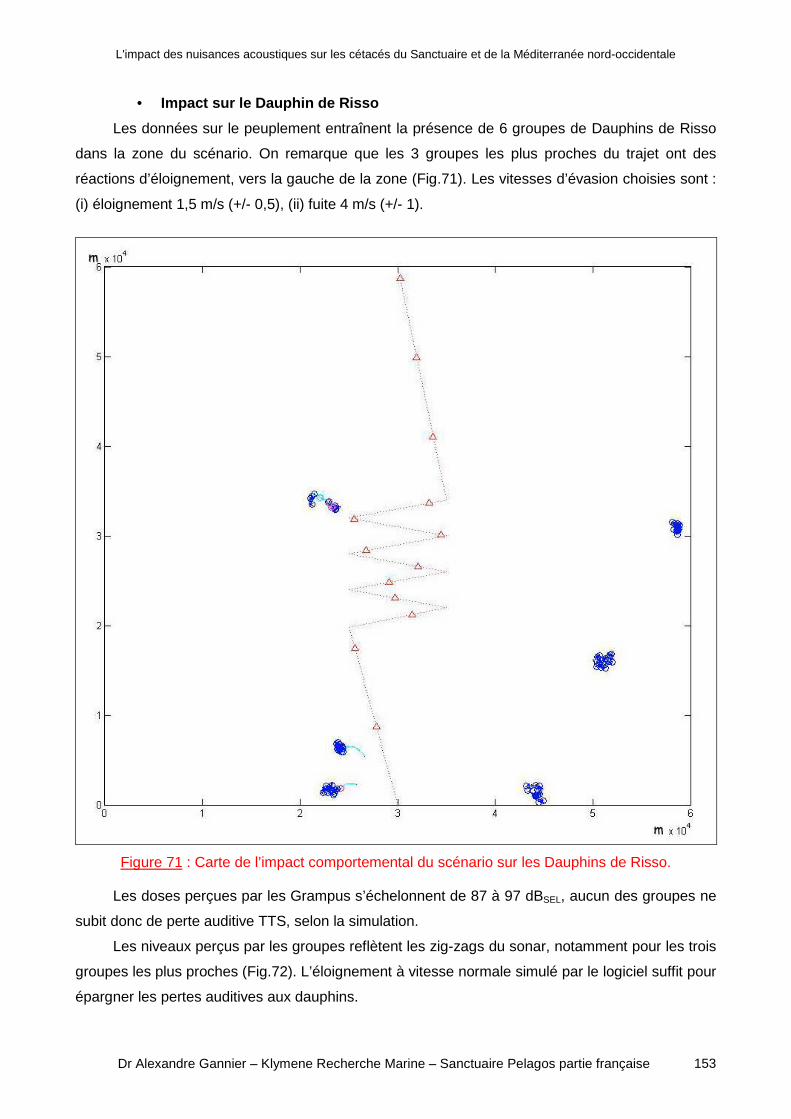
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 153
• Impact sur le Dauphin de Risso
Les données sur le peuplement entraînent la présence de 6 groupes de Dauphins de Risso
dans la zone du scénario. On remarque que les 3 groupes les plus proches du trajet ont des
réactions d’éloignement, vers la gauche de la zone (Fig.71). Les vitesses d’évasion choisies sont :
(i) éloignement 1,5 m/s (+/- 0,5), (ii) fuite 4 m/s (+/- 1).
Figure 71 : Carte de l’impact comportemental du scénario sur les Dauphins de Risso.
Les doses perçues par les Grampus s’échelonnent de 87 à 97 dBSEL, aucun des groupes ne
subit donc de perte auditive TTS, selon la simulation.
Les niveaux perçus par les groupes reflètent les zig-zags du sonar, notamment pour les trois
groupes les plus proches (Fig.72). L’éloignement à vitesse normale simulé par le logiciel suffit pour
épargner les pertes auditives aux dauphins.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 154
Figure 72 : Niveaux perçus par les Dauphins de Risso au cours du scénario de transmission.
• Impact sur les Dauphins bleus et blancs
Les données écologiques génèrent la présence de 72 groupes de Stenella dans la zone du
scénario. Les vitesses d’évasion choisies sont : (i) éloignement 2 m/s (+/- 0,5), (ii) fuite 4 m/s
(+/- 1). Les niveaux perçus sont très variables selon la position du groupe par rapport au trajet du
bateau (Fig.73). Malgré une sensibilité moyenne du Dauphin bleu et blanc à la fréquence de 4,5
kHz, environ 90 dB, certains niveaux perçus dépassent 70 dB.
Figure 73 : Niveaux perçus par les Dauphins de Risso au cours du scénario de transmission.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 155
On observe des légers mouvements d’évasion pour les groupes les plus proches du trajets,
mais la plupart des dauphins ne sont pas perturbés.
Figure 74 : Carte de l’impact comportemental du scénario sur les Dauphins bleus et blancs.
Figure 75 : Doses perçues par les Dauphins bleus et blancs au cours du scénario de transmission.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 156
La longue durée des transmissions (plus de 6 heures au total) entraîne une accumulation
importante de l’exposition sonore pour une douzaine de groupes de dauphins, bien que les
niveaux perçus soient rarement très élevés (Fig.73).
En résumé , au travers des quatre espèces présentes dans la zone de transmission, nous
pouvons prévoir plusieurs types d’impact de pollution sonore, grâce à la simulation.
Nous avons le cas d’une espèce très peu sensible à la fréquence de la pollution sonore, le
Rorqual commun, qui par conséquent ne perçoit quasiment pas le sonar, et n’a aucune réaction
comportementale, ni bien sûr aucune perte auditive.
Nous avons le cas d’une espèce assez sensible à la fréquence du sonar, le Cachalot, qui
effectue une évasion dès le début du scénario, et qui s’éloigne de manière constante du trajet du
sonar. Mais la longue durée de l’insonification et la faible vitesse d’éloignement sont la cause
d’une dose sonore perçue supérieure au seuil de TTS, et donc le logiciel prévoit des TTS.
Nous avons ensuite le cas d’une espèce également sensible à cette fréquence, le Dauphin
de Risso, (environ 72 dB de seuil d’audition), certains des groupes effectuent également des
évasions, mais pour laquelle les doses perçues restent inférieures au seuil et donc on ne prévoit
pas de pertes auditives.
Enfin nous avons le cas d’une population dense de Dauphin bleu et blanc, assez peu
sensible à la fréquence du sonar (90 dB de seuil) : seuls les groupes proches du trajet effectuent
des manoeuvres d’évasion, mais de faible amplitude. Du fait de la durée de l’exposition, les doses
perçues dépassent 90 dBSEL pour certains groupes, mais n’atteignent pas le seuil de TTS.
Il va de soi que ces résultats découlent d’une simulation, dans lesquels nombre de
paramètres sont fixés en fonction de l’état des connaissances, encore insuffisantes notamment les
seuils comportementaux ; par conséquent les effets simulés sont approximatifs. Néanmoins, les
résultats délivrés par cette étude sont conformes à certaines observations réalisées en mer, soit
lors d’expériences sonores, soit de manière opportuniste.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 157
D\ Discussion
La prévision des effets des fortes intensités sonores sur les mammifères marins fait
intervenir des champs de connaissance divers, dont les résultats se combinent pour aboutir au
final à une détermination des impacts sur un peuplement de cétacés. En raison des imprécisions
rencontrées à chaque étape du processus, il est impossible d’obtenir une bonne précision sur cette
prévision. En raison de sa complexité, ce processus ne peut être mené à bien qu’à l’aide d’un
logiciel informatique qui simule les différentes étapes. En réalité, un tel logiciel de simulation ne
peut être réaliste que depuis très peu d’années.
La première source d’approximation réside dans la détermination des niveaux sonores
propagés autour de la source à différentes distances. Même si cet aspect fait appel à de la
physique, il reste soumis à l’influence des facteurs naturels qu’une simulation réalisée à l’avance
ne peut pas forcément restituer. Et il faut déjà que les caractéristiques techniques de la source
sonore soient bien connus, ce qui n’est pas évident. Dans notre cas, l’option a été claire : se placer
en conditions hivernales pour être en mesure d’utiliser une loi propagation sphérique. Dans le cas
d’une prévision d’impact en conditions estivales, la difficulté est plus grande.
Lorsque l’on veut chiffrer les impacts, le second facteur d’imprécision est d’ordre écologique :
il s’agit tout simplement des incertitudes sur le peuplement. Si la composition est souvent connue,
les densités de populations sont souvent estimées grossièrement. Sur ce point, la population du
Sanctuaire Pelagos est mieux placée que la moyenne, grâce aux travaux menés durant la dernière
décennie. Mais l’incertitude sur les densités se chiffre quand même en dizaines de pour cent.
Mis à part les gravissimes lésions traumatiques provoquées par des niveaux extrêmes
rencontrés à très courte distance des sources, la quantification des impacts repose sur une bonne
détermination des effets physiologiques générant des pertes auditives. Les progrès récents pour
ce qui est des sons non-impulsifs ont permis de préciser les critères d’apparition des TTS. Les
seuils d’apparition des pertes auditives permanentes restent quant à eux en partie fondés sur la
prise en compte d’un facteur de sécurité. Le principal facteur d’incertitude pour la prévision des
TTS demeure l’effet d’accumulation de fatigue auditive lors d’une succession d’expositions
intermittentes, ce qui est précisément notre cas. La simple prévision par addition des énergies
sonores, ou accumulation des effets sans relaxation lors de périodes de repos, est certainement
pessimiste. Mais la mise en oeuvre d’une approche alternative reste délicate, car une erreur sur la
quantification de la fatigue auditive pourrait avoir une conséquence grave sur la prévision d’impact.
Une grande partie de la justesse de la prévision des effets dépend de la prise en compte des
facteurs comportementaux, en particulier les seuils de niveau perçu qui provoquent des évasions
plus ou moins rapides. En effet, dans le cas d’une pollution sonore de durée assez courte (comme
dans pour notre scénario, même si l’exposition dure plusieurs heures), une interruption d’activité
peut avoir de faibles conséquences pour la vie des cétacés. En revanche, les seuils fixés pour le

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 158
déclenchement de l’évasion à vitesse normale, et celui qui déclenche la fuite (à vitesse rapide) ont
une grande importance sur les conséquences auditives, ainsi d’ailleurs que les vitesses d’évasion
adoptées dans le processus de modélisation. Il est évident que le seuil comme la vitesse sont des
facteurs qui interviennent directement sur la distance entre le sonar et les cétacés et donc les
niveaux reçus. Les seuils ont été déterminés par examen des résultats d’expériences CEE, mais
ceux-ci sont peu nombreux pour certaines espèces. Quant aux vitesses d’évasion, lente ou rapide,
leurs valeurs découlent de ce que nous avons observé en mer, sur les différentes espèces du
peuplement. L’influence de cet aspect de la modélisation de l’impact peut être mis en évidence
grâce à quelques tests.
Enfin, la philosophie de notre approche, dont le trait principal consiste à se fonder sur une
métrique des niveaux perçus pour évaluer les effets, est originale. Une comparaison avec la
principale approche proposée par ailleurs, celle qui est exposée Southall et al. (2008), est de
toutes façons constructive. En effet, notre approche, comme celle de Southall et al. (2008), n’est
pas figée puisqu’elle évolue avec la publication incessante de nouveaux résultats expérimentaux. Il
est important de distinguer les points où les approches convergent, et ceux où elles diffèrent.
La discussion sur la rationalité de l’approche va donc s’articuler sur ces deux aspects :
certains points critiques de la mise en oeuvre de notre modèle de prévision d’impact, et la
comparaison la philosophie du modèle avec celle d’une autre approche existante.
Pour ce qui est de la prévision des seuils comportementaux, seuils dont le dépassement
provoque l’éloignement, la fuite, ou l’immobilisation-panique des cétacés, il est vrai que la
sensibilité d’un modèle de prévision d’impact peut être importante. On va illustrer ce point en
testant la modélisation des impacts avec différentes vitesses et différents seuils pour une même
espèce, et en comparant les résultats obtenus.
Enfin, un point spécifique portera sur le cas d’une espèce non traitée dans notre exemple,
pour conserver le réalisme de notre peuplement de cétacés, mais très importante pour la
problématique : il s’agit du Ziphius. Nous effectuerons donc une simulation spécifique, comme si le
scénario de transmission de sonar se déroulait dans une zone de peuplement dense de Ziphius.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 159
1) Points forts et limites du modèle proposé
Nous avons évoqué plus haut (§C.1) les limites de notre approche sur la partie
« modélisation des effets reçus », processus physique qui nécessite normalement une
connaissance précise de la bathycélérimétrie. Notre scénario de transmission de sonar étant
localisé au mois de mars et au large, les incertitudes sur la détermination de la propagation des
niveaux sonores ne sont pas de première grandeur. Mais pour tout scénario de pollution sonre se
produisant entre le mois de mai et le mois de novembre, la prise en considération d’une
propagation proche de la réalité (par exemple grâce à un logiciel de tracé de rayons) est
impérative pour prévoir les effets du son au-delà d’une distance de 700-800 m, donc pour tout ce
qui est du domaine du comportemental.
a) Sur les métriques employées
Un des points forts du modèle utilisé réside dans la prise en compte des niveaux perçus pour
la prévision des effets comportementaux. En effet, dans la plupart des approches récentes, on
considérait les niveaux reçus pour prévoir l’apparition des TTS, ou alors des pondérations
fréquentielles par grands groupes d’espèces, mais qui en fait n’apportent aucune différence de
prise en compte de l’audition sur l’essentiel du domaine de fréquence (200 à 50 000 Hz), entre un
marsouin et un cachalot par exemple (Southall et al., 2007). Or depuis l’étude de Finneran &
Schlundt (2010), il est devenu évident que le seuil d’apparition des TTS, et leur amplitude, dépend
fortement de l’audition de l’espèce, et de la fréquence de l’insonification. Finneran & Schlundt
(2010) expriment en effet la valeur du TTS mesuré lors d’expériences à l’aide de la relation
suivante :
TTS = a. log10 [1+10 (Lr-b)/10 ]
où a est un paramètre de pente de croissance du TTS et b un niveau sonore seuil dépendant de la
fréquence. Or, les niveaux seuils exprimés pour les fréquences de 3 et 20 kHz (resp. 156 et
165 dB) reflètent à peu de choses près la différence de seuil d’audition pour ces deux fréquences,
pour le dauphin considéré. A une constante près, la valeur (Lr – b) de la relation de Finneran &
Schlundt (2010) est donc égale à un niveau perçu Lp : Lr – b(f) = (Lr –seuil (f) – k) = (Lp – k).
Notre méthode de niveaux perçus pour prévoir l’apparition d’un TTS en fonction de la
fréquence de l’insonification est donc confortée par ces travaux expérimentaux.
Un des autres points forts de notre approche, qui est conforté par d’autres méthodes
actuelles (comme Southall et al., 2008), est la prise en compte de la variable « durée
d’insonification » pour la prévision des TTS, sous la forme d’une expression du seuil en terme de
dose d’exposition sonore SEL (en dB re 1µPa2.s). Les expériences sur le Grand dauphin avec des
sons intermittents de sonar menées par Finneran et al. (2010a et b) montrent que la prévision du
seuil d’apparition de TTS et de l’amplitude de TTS est mieux décrite par un modèle complexe, qui
intègre les temps morts entre deux impulsions de sonar, que par le simple modèle d’accumulation

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 160
d’énergie basé sur le SEL. Cependant, la différence n’est pas importante, d’une part, et d’autre
part une métrique SEL simple se situe du côté de la sécurité des cétacés, en ce sens qu’elle a
tendance à légèrement sous-estimer le temps d’exposition nécessaire pour provoquer un TTS.
Dernier point, les résultats expérimentaux sont encore trop peu nombreux pour justifier l’emploi
d’un modèle complexe et qui reste fragile.
Par conséquent, l’utilisation dans notre approche d’une métrique en SEL pour tenir compte
de la durée d’insonification dans la prévision des TTS et des PTS reste aujourd’hui encore la
meilleure option.
b) Sensibilités des résultats aux seuils adoptés pour les effets comportementaux
La prévision des effets comportementaux et acoustiques repose en partie sur les seuils
adoptés pour définir les mouvements d’évasion, éloignement et fuite, principalement, ainsi que les
vitesses moyennes adoptées pour ces modalités d’évasion. Pour évaluer la sensibilité de nos
prévisions à ces paramètres, on reprend la prévision d’impact réalisée pour le Dauphin bleu et
blanc en faisant varier deux couples de valeurs, (seuil d’éloignement-seuil de fuite) et (vitesse
d’éloignement-vitesse de fuite) dans un sens, puis dans l’autre. Cela nous donne quatre cas de
modélisation, pour lesquels on peut comparer les prévisions, au travers des doses d’expositions
perçues par l’ensemble des groupes de dauphins.
Cas Seuild’éloignement
Seuil de fuite Vitessed’éloignement
Vitesse defuite
référence 65 dB perçus 80 dB perçus 2 m/s (0,5) 4 m/s (1)
seuils plus forts 70 dB perçus 85 dB perçus 2 m/s (0,5) 4 m/s (1)
seuils plus faibles 60 dB perçus 75 dB perçus 2 m/s (0,5) 4 m/s (1)
vitesses + élevées 65 dB perçus 80 dB perçus 2.5 m/s (0,3) 5 m/s (1)
vitesse + faibles 65 dB perçus 80 dB perçus 1.5 m/s (0,5) 3 m/s (1)
Tableau 34 : Paramètres utilisés pour évaluer la sensibilité de la prévision (Dauphin bleu et blanc)
Pour ces évaluations, nous conservons le même scénario de transmission du sonar que
dans la prévision d’impact effectuée plus haut. On constate que les variations sur les doses
sonores perçues par les dauphins ne sont pas drastiques (Fig.76). L’influence des variations de
vitesses d’évasion n’est pas univoque. Par contre, une augmentation des seuils d’évasion se
traduit par un plus grand nombre de groupes de dauphins exposés à des doses supérieures à 90
dBSEL, et vice versa.
Cela confirme que les études CEE doivent être analysées avec soin pour en déduire les
niveaux sonores qui provoquent différents types de réaction, si possible sur des espèces très
proches de celles qui sont analysées dans une étude d’impact.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 161
Figure 76 : Influence des seuils et vitesses sur les doses d’exposition sonore des dauphins.
Malgré tout, la variation des seuils et des vitesses ne fait pas apparaître de cas de TTS ou
de PTS, ce qui indique que notre prévision d’impact est quand même assez robuste vis-à-vis de la
conséquence sévère que constitue une perte auditive partielle.
c) Cas spécifique du Ziphius
Il serait incongru dans une telle étude de ne pas aborder spécifiquement le cas du Ziphius,
espèce méditerranéenne qui est une victime signalée des exercices navals avec sonar. Le Ziphius
ne faisant pas partie du peuplement habituel au large des îles du Levant50, nous avons abordé son
cas en faisant rejouer le scénario sonar dans une zone où il y aurait une forte densité de Ziphius.
Les paramètres de population sont les suivants : densité de 0,04 individu par km2, 2 individus
par groupe. Les paramètres de la source sonore sont les mêmes que précédemment : niveau de
source de 220 dB re1µPa@1m, 4,5 kHz, 5 pings de 0,5 sec par minute, vitesse du bateau 5 m/s,
parcours en zig-zags avec 8 points de passage (Fig.77). Les paramètres de comportement sont
les suivants : vitesse normale d’évasion 2,5 m/s (+/-0,5) – vitesse de fuite 3,5 m/s (+/-1) – seuil
d’évasion 60 dB perçus – seuils de fuite 75 dB perçus – seuils de panique 90 dB perçus.
Comme dans les cas de l’étude principale, nous avons fait tourner le logiciel à trois reprises
et nous donnons la moyenne des doses perçues : sur 72 groupes de Ziphius, 67 subissent une
perte auditive TTS, et 5 sont indemnes. Les doses totales perçues sont supérieures à 110 dB
re1µPa2.s (un groupe), comprises entre 100 et 110 dB (66 groupes) et inférieures à 100 dB (5
groupes).
On constate que l’impact prévu du scénario sonar est énorme, dans un tel cas.
Les doses perçues dépassent le seuil de TTS dans la majorité des cas, en raison de la
bonne sensibilité auditive du Ziphius à la fréquence de 4,5 kHz (Fig.58) qui est de 72 dB environ.
50 Bien qu’il soit signalé de temps à autre.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 162
La majorité des groupes de Ziphius réagissent par une évasion rapide (couleur verte) à l’approche
du sonar, mais cette évasion ne suffit pas à leur épargner une forte exposition aux doses sonores.
Figure 77 : Application de la simulation du scénario sonar à une population de Ziphius.
Même si par essence, les prévisions ne peuvent pas être exactes, la différence de la
simulation pour le Ziphius par rapport aux autres cas est impressionnante. Il est évident que
l’utilisation du sonar à moyenne fréquence dans une zone densément peuplée de Ziphius est à
éviter, en Méditerranée comme ailleurs. La simulation rejoint ici l’expérience : des échouages de
Ziphius ont été observés à de nombreuses reprises lors d’exercices de sonar51.
51 Comme cela vient à nouveau de se produire en Grèce (1er avril 2014).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 163
2) Comparaison avec d'autres approches disponibles
Le développement et l’emploi de systèmes de prévision et de mitigation des impacts des
nuisances sonores date d’une dizaine d’années, avec l’apparition de logiciels complets depuis un
peu moins de temps. Un logiciel complet permet de prévoir les impacts d’une source dans un
contexte géo-référencé donné, avec la connaissance du peuplement local basé sur des bases de
données intégrées dont la précision est variable52.
Le « moteur » qui permet à un logiciel de prévoir les pertes auditives a généralement été
développé par ailleurs, sous forme d’un critère qui s’inspire des résultats expérimentaux
disponibles. Dans les cas où les principes actifs du logiciel sont connus, on s’aperçoit qu’il y a un
point commun : la prévision des pertes auditives est basée sur l’accumulation de l’énergie sonore
reçue au cours du scénario de transmission, c’est-à-dire la métrique en SEL, mesure du niveau
d’exposition en dB re 1µPa2.s. C’est le cas du critère de Southall et al. (2007), et du critère en
cours de révision pour le National Oceanic and Atmosphere Administration53, qui s’en inspire
directement.
La partie active d’un logiciel qui permet de prévoir les réactions comportementales est en
général moins explicite. Pour le logiciel Sakamata, il apparaît une référence aux niveaux sonores
qui entraînent des réactions d’aversion ou d’inconfort chez les humains (Benders et al., 2004 ;
Verboom & Kastelein, 2005). Pour d’autres logiciels, les expériences récentes de CEE ont
certainement été utilisées pour exprimer des critères de réaction, ce que nous avons également
fait dans cette étude avec le matériel scientifique disponible. Il est clair que la détermination des
effets comportementaux est un enjeu extrêmement important pour la prévision des impacts des
pollutions sonores, ce qui justifie d’ailleurs les efforts scientifiques actuels (plusieurs gros
programmes de CEE).
a) Audiométrie par espèce ou regroupement d’espèces ?
Pour revenir à la prévision des impacts auditifs, mais cette fois sur la prise en compte de
l’audition des différentes espèces, plusieurs approches existent : en général, le principe est de
comparer les doses sonores reçues à des courbes de sensibilités qui sont déterminées pour des
groupes d’espèces de cétacés. Ces groupes sont basés sur la classification zoologique, en
distinguant les mysticètes des odontocètes. Ces derniers sont eux-mêmes subdivisés en sous-
groupes. Dans l’approche de Southall et al. (2007), il y a deux sous-groupes d’odontocètes : celui
des cétacés « moyenne fréquence » et celui des cétacés « haute fréquence ». Le premier
52 Certains logiciels sont développés et commercialisés par des industriels du sonar, comme Sakamata(TNO) ou ERMC (BaeD).53 Draft Guidance for Assessing the Effects of Anthropogenic Sound on Marine Mammals - AcousticThreshold Levels for Onset of Permanent and Temporary Threshold Shifts (23 décembre 2013)

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 164
regroupe les cachalots, la plupart des delphinidés, les ziphiidés, les monodontidés, et le second,
les marsouins, les dauphins de rivière et les dauphins du genre Cephalorhynchus54.
Les impacts auditifs générés sur toutes les espèces d’un même groupement reposent sur
une pondération dérivant d’une courbe unique de sensibilité. Or, les espèces incluses dans un
même groupement ont souvent un audiogramme très distinct, comme par exemple pour nos
espèces méditerranéennes, le Globicéphale noir et le Dauphin bleu et blanc (Fig.78).
Figure 78 : Comparaison des audiogrammes du Dauphin bleu et blanc et du Globicéphale noir.
Pour le domaine basse et moyenne fréquence, très important pour notre problématique, la
différence de sensibilité entre des deux espèces avoisine 20 dB, ce qui est très important en terme
de conséquence sur la vulnérabilité aux énergies sonores.
Pour cette raison, une approche basée sur les audiogrammes des espèces, telle que la
nôtre, est meilleure qu’une approche basée sur des grands regroupements d’espèces, même si
nous avons été conduits à utiliser un audiogramme semi-synthétique pour le Cachalot et Le
Rorqual commun.
b) Type de pondération fréquentielle
Dans toutes les approches récentes, ce ne sont pas les niveaux reçus qui servent de
métrique pour prévoir les pertes auditives, mais des niveaux qui sont pondérés en fonction de la
sensibilité auditive. La méthode de pondération est variable : dans le cas de notre approche, la
pondération se déduit directement de l’audiogramme, puisque le niveau perçu est égal au niveau
reçu diminué du seuil d’audition à la fréquence considérée. Prenons l’exemple du Dauphin bleu et
blanc : avec l’approche de Southall et al. (2008), on a une pondération extrêmement plate, pour
laquelle le niveau sonore « actif » pour la détermination des pertes auditives est égal au niveau
reçu sur la plus grande partie du domaine (Fig.79).
54 Les regroupements ont été légèrement modifiés dans la proposition de mise à jour du NOAA.
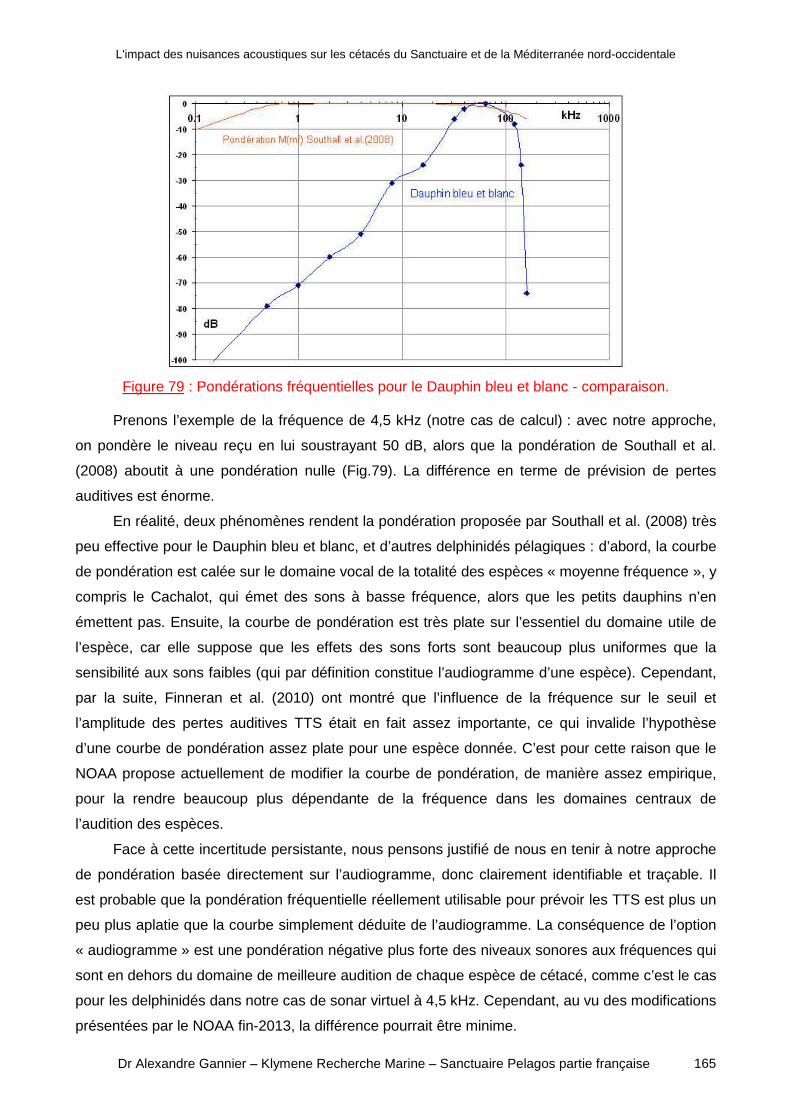
L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 165
Figure 79 : Pondérations fréquentielles pour le Dauphin bleu et blanc - comparaison.
Prenons l’exemple de la fréquence de 4,5 kHz (notre cas de calcul) : avec notre approche,
on pondère le niveau reçu en lui soustrayant 50 dB, alors que la pondération de Southall et al.
(2008) aboutit à une pondération nulle (Fig.79). La différence en terme de prévision de pertes
auditives est énorme.
En réalité, deux phénomènes rendent la pondération proposée par Southall et al. (2008) très
peu effective pour le Dauphin bleu et blanc, et d’autres delphinidés pélagiques : d’abord, la courbe
de pondération est calée sur le domaine vocal de la totalité des espèces « moyenne fréquence », y
compris le Cachalot, qui émet des sons à basse fréquence, alors que les petits dauphins n’en
émettent pas. Ensuite, la courbe de pondération est très plate sur l’essentiel du domaine utile de
l’espèce, car elle suppose que les effets des sons forts sont beaucoup plus uniformes que la
sensibilité aux sons faibles (qui par définition constitue l’audiogramme d’une espèce). Cependant,
par la suite, Finneran et al. (2010) ont montré que l’influence de la fréquence sur le seuil et
l’amplitude des pertes auditives TTS était en fait assez importante, ce qui invalide l’hypothèse
d’une courbe de pondération assez plate pour une espèce donnée. C’est pour cette raison que le
NOAA propose actuellement de modifier la courbe de pondération, de manière assez empirique,
pour la rendre beaucoup plus dépendante de la fréquence dans les domaines centraux de
l’audition des espèces.
Face à cette incertitude persistante, nous pensons justifié de nous en tenir à notre approche
de pondération basée directement sur l’audiogramme, donc clairement identifiable et traçable. Il
est probable que la pondération fréquentielle réellement utilisable pour prévoir les TTS est plus un
peu plus aplatie que la courbe simplement déduite de l’audiogramme. La conséquence de l’option
« audiogramme » est une pondération négative plus forte des niveaux sonores aux fréquences qui
sont en dehors du domaine de meilleure audition de chaque espèce de cétacé, comme c’est le cas
pour les delphinidés dans notre cas de sonar virtuel à 4,5 kHz. Cependant, au vu des modifications
présentées par le NOAA fin-2013, la différence pourrait être minime.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 166
De plus, l’approche Southall/NOAA conserve son inconvénient fondamental, qui est de
regrouper un trop grand nombre d’espèces différentes, au détriment de la justesse de la
pondération à apporter pour chaque espèce. Enfin, la révision en cours (document NOAA de
décembre 2013), consiste à proposer une courbe de pondération composite, réunion de deux
courbes. Outre que cette construction est empirique, ce type de solution est de toute évidence
destiné à pallier temporairement les incertitudes réelles sur cette problématique. Elle n’est pas
destinée à durer longtemps : d’ici quelques années au plus, la recherche éclaircira davantage ce
point encore nébuleux, mais très important, des démarches de prévision d’impact sonore.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 167
Conclusion
La problématique des effets des pollutions sonores sur les cétacés voit les connaissances
progresser très vite depuis que l’on a constaté à quel point ces mammifères pouvaient être
touchés par les activités bruyantes. Cette étude donne un état des lieux en intégrant les progrès
scientifiques de ces dernières années, notamment dans les deux domaines clefs que sont d’une
part, la compréhension des phénomènes de fatigue auditive, et, d’autre part, l’importance des
réactions comportementales. Pour ce qui est de la prévision des pertes de sensibilité auditive, il
est devenu évident depuis quelques années que les effets d’un niveau sonore élevé sur une
espèce donnée dépendent fortement de la fréquence de l’insonification, et de la durée de
l’exposition au son. Toute approche de prévision des impacts qui ne prendrait pas en compte cet
aspect serait vouée à l’échec. En ce qui concerne les réactions comportementales, les résultats
récents des expériences d’exposition contrôlée, sur de nombreuses espèces, démontrent qu’une
large gamme de réactions se produit lorsque les cétacés perçoivent les sources sonores, même à
des niveaux beaucoup plus faibles que ceux qui provoquent des atteintes auditives. Ces réactions
comportementales concernent un nombre beaucoup plus grand d’individus que les effets auditifs,
aussi leur prise en compte est obligatoire dans toute étude d’impact sérieuse. De plus, les
réactions comportementales peuvent provoquer des échouages massifs, et leur prévision est
nécessaire pour déterminer le risque d’impact auditif sur les individus proches de la source.
Après avoir justifié les bases théoriques et expérimentales de notre approche de la prévision
des impacts sonores, dans laquelle les niveaux perçus ont une grande importance, nous avons
effectué une prévision d’impact dans le cas du scénario choisi : celui d’un exercice de transmission
de sonar à moyenne fréquence. Le sonar a des caractéristiques « génériques » arbitraires, et le
scénario lui-même est imaginaire. Par contre, nous choisissons d’exercer la simulation en eaux
profondes et en conditions hivernales, pour des raisons de faisabilité. L’outil de simulation est un
logiciel de prévision mis au point par Klymene Recherche Marine, plusieurs années en amont de
l’étude, et utilisé auparavant dans des cas réels. Cette simulation nous conduit à déterminer les
impacts de la transmission sonar sur un peuplement de cétacés comprenant le Dauphin bleu et
blanc, le Dauphin de Risso, le Cachalot et le Rorqual commun. Les résultats de la prévision
comprennent une gamme d’effets, allant de l’indifférence quasi-totale (Rorqual commun) à
l’évasion rapide (Dauphin de Risso) et à des atteintes auditives (Cachalot). Nous discutons ces
résultats, en même temps que les tenants et les aboutissants de notre approche. L’approche par
simulation est aujourd’hui indispensable si l’on veut prévoir les effets biologiques et
comportementaux de manière réaliste ; elle peut évoluer en fonction des progrès de la science.
L’effet des pollutions sonores a été largement ignoré dans la mise en oeuvre de la protection
des cétacés en Méditerranée, et ailleurs. La détérioration acoustique du milieu marin a pu, et
pourra, provoquer la désertion d’habitats locaux par certaines espèces vulnérables.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 168
Références bibliographiques
Une bibliothèque pdf des références utilisées est fournie, en fonction de la disponibilité des
fichiers. Les références fournies sont signalées par un nom de premier auteur écrit en BLEU.
Aguilar Soto N., M. Johnson, P.T. Madsen, P.L. Tyack, A. Bocconcelli, J.F. Borsani, 2006. Doesintense ship noise disrupt foraging in deep diving Cuvier's beaked whales (Ziphiuscavirostris)? Mar.Mam.Sci. 22(3): 690–699.
Arbelo M., Sierra E., Mendez M., Godinho A., Ramirez G., Caballero M.J, Fernandez A., 2007.Atypical beaked whale mass stranding in Almeria’s coasts: pathological study. 21st AnnualConference of the E.C.S. (San Sebastian). Abstracts : 75.
Au W.W. L., P.E. Nachtigall, J L. Pawloski 1999. Temporary threshold shift in hearing induced byan octave band of continuous noise in the bottlenose dolphin. J. Acoust. Soc. Am.106(4):abstract.
Au W.W.L., 2000. Hearing in whales and dolphins: an overview. Pp1-43 in Au W.W.L., PopperA.N., Fay R.R. (Eds). Hearing by whales and dolphins, Springer Handbook of AuditoryResearch: 485pp.
Au W.W.L.,& Hastings M.C., 2008. Principles of Marine Bioacoustics. Springer: 679pp.
Au W.W.L., Ford J.K.B., Horne J.K., Newman Allman K.A., 2004. Echolocation signals of free-ranging killer whales (Orcinus orca) and modeling of foraging for chinook salmon(Oncorhynchus tshawytscha). J.Acoust.Soc.Am. 115(2): 901-909
André M., A. Supin, E. Delory, C. Kamminga, E. Degollada, J. M. Alonso, 2003. Evidence ofdeafness in a striped dolphin, Stenella coeruleoalba. Aquatic Mammals 29(1): 3–8.
Benders F.P.A., Beerens S.P., Verboom W.C., 2004. SAKAMATA - A tool to avoid whalestrandings. Proceedings of UDT Conference (Nice, June 2004) : 6pp.
Boyd Y., Claridge D., Clarck C., Southall B., Tyack P., 2007. Behavioral Research Study 2007Cruise Report. executive summary: 11pp.
Boyd Y., 2009. Analysis and application of BRS data (Latte and other approaches). Présentationorale à ESOMM3, Lerici (7-9 sept 09).
Buckstaff K.C., 2004. Effects of watercraft noise on the acoustic behavior of bottlenose dolphinsT.truncatus in Sarasota Bay, Florida. Mar.Mam.Sci. 20(4): 709-725.
Carstensen J., O.D. Henriksen, J. Teilmann, 2006. Impacts of offshore wind farm construction onharbour porpoises: acoustic monitoring of echolocation activity using porpoise detectors (T-PODs). Mar.Ecol. Prog.Ser. (321): 295–308.
Cook M.L.H., R.A. Varela, J.D. Goldstein, S.D. McCulloch, G.D. Bossart, J.J. Finneran, D. Houser,D.A. Mann, 2006. Beaked whale auditory evoked potential hearing measurements. J CompPhysiol A 192: 489–495.
Cox T.M., T.J. Ragrn, A.J. Read, E. Vos, R.W. Baird, K. Balcomb et al., 2006. Understanding theimpacts of anthropogenic sound on beaked whales. J. Cet. Res. Manage. 7(3): 177–187.
Cranford T.W., M.F. McKenna, M.S. Soldevilla, S.M. Wiggins, J.A. Goldbogen, R.E. Shadwick, P.Krysl, J.A. St. Leger, J.A. Hildebrand, 2008. Anatomic geometry of sound transmission andreception in Cuvier’s beaked whale (Ziphius cavirostris). The Anatomical Record 291: 353–378
Crum L.A., Yi Mao, 1996. Acoustically enhanced bubble growth at low frequencies and itsimplications for human diver and marine mammal safety. J.Acoust.Soc.Am. 99 (5), 2898-2907.
Crum L.A., M.R. Bailey, J. Guan, P.R. Hilmo, S.G. Kargl, T.J. Matula, O.A. Sapozhnikov, 2005.Monitoring bubble growth in supersaturated blood and tissue ex vivo and the relevance tomarine mammal bioeffects. Acoustical Research Letter Online 6(3).
D'Amico A., 2010. Latest Behavioral Response Study Builds Upon Years of Marine MammalResearch.Currents Summer 2010: 48-55.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 169
David J.A., 2006. Likely sensitivity of bottlenose dolphins to pile-driving noise. Water andEnvironment Journal 20: 48-54.
Erbe C., 2002. Underwater noise of whale-watching boats and potential effects on killer whales(Orcinus orca), based on an acoustic impact model. Mar.Mam.Sci. 18(2): 394-418.
Erbe C., A. MacGillivray, R. Williams, 2012. Mapping cumulative noise from shipping to informmarine spatial planning. J.Acoust.Soc.Am. 132(5): EL423-428
Erbe C., 2013. Underwater noise of small personal watercraft (jet skis). J. Acoust. Soc. Am. 133(4),EL326: 5pp
Evans P.G.H., Nice H, 1997. Review of the effects of underwater sound generated by seismicsurveys on cetaceans. Report of the Sea Watch Foundation: 50pp.
Evans D.L., England G.R., 2001. Joint Interim Report Bahamas Marine Mammal Stranding Eventof 14-16 March 2000. NOAA/U.S. Navy: 66pp.
Fernandez A., J.F. Edwards, F. Rodriguez, A. Espinosa de los Monteros, P. Herraez, P. Castro,J.R. Jaber, V. Martin, M. Arbelo, 2005. ‘‘Gas and Fat Embolic Syndrome’’ Involving a massstranding of beaked whales (Family Ziphiidae) exposed to anthropogenic sonar signals. Vet.Pathol. 42: 446-457.
Finneran J.J., C.E. Schlundt, D.A. Carder, J.A. Clark, J.A. Young, J.B. Gaspin, S.H. Ridgway,2000. Auditory and behavioral responses of bottlenose dolphins (Tursiops truncatus) and abeluga whale (Delphinapterus leucas) to impulsive sounds resembling distant signatures ofunderwater explosions. J.Acoust.Soc.Am. 108(1): 417-431.
Finneran J J., C E. Schlundt, R Dear, D A. Carder, S H. Ridgway, 2002. Temporary shift in maskedhearing thresholds in odontocetes after exposure to single underwater impulses from aseismic watergun. J.Acoust.Soc.Am. 111(6): 2929-2940.
Finneran J.J., D.A. Carder, C.E. Schlundt, S.H. Ridgway, 2005. Temporary threshold shift inbottlenose dolphins (Tursiops truncatus) exposed to mid-frequency tones. J.Acoust.Soc.Am.118(4): 2696-2705.
Finneran J.J., C.E. Schlundt, B. Branstetter, R.L. Dear, 2007. Assessing temporary threshold shiftin a bottlenose dolphin (Tursiops truncatus) using multiple simultaneous auditory evokedpotentials. J.Acoust.Soc.Am. 122(2): 1249-1264.
Finneran J.J., D.S. Houser,B. Mase-Guthrie, R.Y. Ewing, R.G. Lingenfelser, 2009. Auditory evokedpotentials in a stranded Gervais’ beaked whale (Mesoplodon europaeus). J.Acoust.Soc.Am.126(1): 484-490.
Finneran J.J., C.E. Schlundt, 2010. Frequency-dependent and longitudinal changes in noise-induced hearing loss in a bottlenose dolphin (Tursiops truncatus) (L). J. Acoust. Soc. Am.128(2): 567-570
Finneran J.J., D.A. Carder, C.E. Schlundt, R.L. Dear, 2010. Growth and recovery of temporarythreshold shift at 3 kHz in bottlenose dolphins: Experimental data and mathematical models. J.Acoust. Soc. Am. 127(5): 3256-3266
Finneran J.J., D.A. Carder, C.E. Schlundt, R.L. Dear, 2010. Temporary threshold shift in abottlenose dolphin (Tursiops truncatus) exposed to intermittent tones. J. Acoust. Soc. Am.127(5): 3267-3272
Finneran J.J., C.E. Schlundt, 2013. Effects of fatiguing tone frequency on temporary threshold shiftin bottlenose dolphins (Tursiops truncatus). J. Acoust. Soc. Am. 133(3): 1819-1826.
Frantzis A., 2004. The first mass stranding that was associated with the use of active sonar(Kyparissiakos gulf, Greece 1996). Proceedings of the workshop on active sonars andcetaceans (La Palmas, 8 March 2003): 14-20.
Freitas L., 2004. The stranding of three Cuvier's beaked whales Ziphius cavirostris in Madeiraarchipelago- May 2000. Proceedings of the workshop on active sonars and cetaceans (LaPalmas, 8 March 2003): 28-32.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 170
Gannier A., 2006. Le peuplement estival de cétacés dans le Sanctuaire Marin Pelagos(Méditerranée nord-occidentale): distribution et abondance. Mammalia 70(1): 17-27.
Gannier A., Fuchs S., Quèbre P., Oswald J.N., 2010. Performance of a contour-basedclassification method for whistles of Mediterranean delphinids. Applied Acoustics 71: 1063-1069.
Gannier A., Mifsud L., 2011. From experimental data to a scale of sound-induced effectsexpressed in terms of perceived levels. Fourth International Conference on the Effects ofSound in the Ocean on Marine Mammals (Amsterdam, 5-9 September 2011). Abstract
Gentry R.L., 2002. Mass Stranding of Beaked Whales in the Galapagos Islands, April 2000. Unp.note, November 4, 2002.
Gillespie D., C. Dunn, J. Gordon, D. Claridge, C. Embling, I. Boyd, 2009. Field recordings ofGervais’ beaked whales Mesoplodon europaeus from the Bahamas. J. Acoust. Soc. Am.125(5): pp.
Gordon J., D. Gillespie, J. Potter, A. Frantzis, M. P. Simmonds, R. Swift, D. Thompson, 2004. AReview of the effects of seismic surveys on marine mammals. Marine Technology SocietyJournal 37(4): 16-45.
Gray H., K. Van Waerebeek, 2011. Postural instability and akinesia in a pantropical spotteddolphin, Stenella attenuata, in proximity to operating airguns of a geophysical seismic vessel.Journal for Nature Conservation 19: 363– 367.
Hatch L., C. Clark, R. Merrick, S. Van Parijs, D. Ponirakis, K Schwehr, M. Thompson, D. Wiley,2008. Characterizing the relative contributions of large vessels to total ocean noise fields: acase study using the Gerry E. Studds Stellwagen Bank National Marine Sanctuary.Environmental Management 42: 735–752.
Hildebrand J. 2004. Source of Anthropogenic Sound in the Marine Environment. Marine MammalCommission. Source : www.mmc.gov/sound/internationalwrkshp/backgroundpapers.html
Hohn A A., D S. Rotstein, C.A. Harms, B.L. Southall, 2006. Report on marine mammal unusualmortality event UMESE0501Sp: multispecies mass stranding of pilot whales (Globicephalamacrorhynchus), Minke whale (Balaenoptera acutorostrata), and dwarf sperm whales (Kogiasima) in North Carolina on 15-16 January 2005. NOAA Technical Memorandum NMFS-SEFSC-537: 230pp.
Holt M.M., D.P. Noren, V. Veirs, C.K. Emmons, S. Veirs, 2009. Speaking up: Killer whales (Orcinusorca) increase their call amplitude in response to vessel noise. J.Acoust.Soc.Am. 125(1):ET27.
Houser D.S., J.J. Finneran, 2006. A comparison of underwater hearing sensitivity in bottlenosedolphins (Tursiops truncatus) determined by electrophysiological and behavioral methods. J.Acoust. Soc. Am. 120(3): 1713-1722.
Houser D.S., A Gomez-Rubio, J.J. Finneran, 2008. Evoked potential audiometry of 13 Pacificbottlenose dolphins (Tursiops truncatus g.). Marine Mammal Science 24(1): 28–41
Huggenberger S., M.A. Rauschmann, H.H.A. Oelschlagger, 2008. Functional Morphology of theHyolaryngeal Complex of the Harbor Porpoise (Phocoena phocoena): Implications for its Rolein Sound Production and Respiration. The Anatomical Record 291:1262–1270
Huggenberger S., M.A. Rauschmann, T J. Vogl, H.H.A. Oelschlager, 2009. Functional morphologyof the nasal complex in the harbor porpoise (Phocoena phocoena L.). The Anatomical Record292: 902–920
ICES 2005. Report of the Ad-hoc Group on Impacts of Sonar on Cetaceans and Fish (AGISC). CM2006/ACE06: 25pp.
Jensen F.H., L. Bejder, M. Wahlberg, N. Aguilar Soto, M. Johnson, P.T. Madsen, 2009. Vesselnoise effects on delphinid communication. M.E.P.S. 395: 161–175
Jepson P.D., Arbelo M., Deaville R., Patterson I.A.P., Castro P., Baker J.R., Degollada E., RossH.M., Herraez P., Pocknell A.M., Rodriguez F., Howie F.E., Espinosa A., Reid R.J., Jaber J.R.,

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 171
Martin V., Cunninghan A.A., Fernandez A., 2003. Gas-bubble lesions in stranded cetaceans.Nature 435: 575–576.
Jepson P.D., R. Deaville, K. Acevedo-Whitehouse, J. Barnett, A. Brownlow, R.L. Brownell Jr., F.C.Clare, N. Davison, R.J. Law, J. Loveridge, S.K. Macgregor, S. Morris, S. Murphy, R. Penrose,M.W. Perkins, E. Pinn, H. Seibel, U. Siebert, E. Sierra, V. Simpson, M.L. Tasker, N. Tregenza,A.A. Cunningham, A. Fernandez, 2013. What caused the UK’s largest common dolphin(Delphinus delphis) mass stranding event?. PLOS One 8(4) e60953: 18pp
Jochens A., D. Biggs, K. Benoit-Bird, D. Engelhardt, J. Gordon, C. Hu, N. Jaquet, M. Johnson, R.Leben, B. Mate, P. Miller, J. Ortega-Ortiz, A. Thode, P. Tyack, B. Würsig, 2008. Sperm whaleseismic study in the Gulf of Mexico. Synthesis report. OCS Study-MMS 2008-006: 341pp.
Johnson M.P., Madsen P.T., Zimmer W.M.Z., Aguilar de Soto N., & Tyack P.T. 2004. Beakedwhales echolocate on prey. Proceedings of the Royal Society Biological Letters.DOI10.1098/rsbl.2004.0208 .
Johnson M., P.T. Madsen, W.M.X. Zimmer, N. Aguilar de Soto, P.L. Tyack, 2006. ForagingBlainville’s beaked whales (Mesoplodon densirostris) produce distinct click types matched todifferent phases of echolocation. The Journal of Experimental Biology 209: 5038-5050
Joint Nature Concervation Council, 2004. Guidelines for minimising the disturbance to marinemammals from seismic surveys. JNCC report: 9pp.
Kastelein R.A., P. Bunskoek, M. Hagedoorn, W.W.L. Au, D. de Haan, 2002. Audiogram of a harborporpoise (Phocoena phocoena) measured with narrow-band frequency-modulated signals. J.Acoust. Soc. Am. 112(1): 334-344.
Kastelein R.A., M. Hagedoorn, W.W. L. Au, D. de Haan, 2003. Audiogram of a striped dolphin(Stenella coeruleoalba). JASA 113(2) : 1130-1337.
Kastelein R.A., N. Jennings, W.C. Verboom, D. de Haan, N.M. Schooneman, 2006. Differences inthe response of a striped dolphin (Stenella coeruleoalba) and a harbour porpoise (Phocoenaphocoena) to an acoustic alarm. Marine Environmental Research 61: 363-378
Kastelein R.A., W.C. Verboom, N. Jennings, D. de Haan, S. van der Heul, 2008. The influence of70 and 120 kHz tonal signals on the behavior of harbor porpoises (Phocoena phocoena) in afloating pen. Marine Environmental Research 66: 319–326
Kastelein R.A., W C. Verboom, N Jennings, D de Haan, 2008. Behavioral avoidance thresholdlevel of a harbor porpoise (Phocoena phocoena) for a continuous 50 kHz pure tone (L).J.Acoust.Soc.Am. 123(4): 1858-1861.
Kastelein R.A., N. Steen, R. Gransier, P.J. Wensveen, C.A. F. de Jong, 2012. Threshold receivedsound pressure levels of single 1–2 kHz and 6–7 kHz up-sweeps and down-sweeps causingstartle responses in a harbor porpoise (Phocoena phocoena). J. Acoust. Soc. Am. 131(2):2325-2333.
Kastelein R.A., R. Gransier, L. Hoek, M. Rambags, 2013. Hearing frequency thresholds of a harborporpoise (Phocoena phocoena) temporarily affected by a continuous 1.5kHz tone. J. Acoust.Soc. Am. 134(3): 2286-2292.
Ketten D.R., J. Lien, S. Todd, 1993. Blast injury in humpback whale ears: Evidence andimplications (A). J. Acoust. Soc. Am. 94(3): 1849-1850.
Ketten D.R., 2005. Beaked whale necropsy findings for strandings in the Bahamas, Puerto Rico,and Madeira, 1999-2002. WHOI Technical report 2005-09: 38pp
Krysl P., T. W. Cranford, S. M. Wiggins, J. A. Hildebrand, 2006. Simulating the effect of high-intensity sound on cetaceans: Modeling approach and a case study for Cuvier’s beaked whale(Ziphius cavirostris). J.Acoust.Soc.Am. 120 (4): 2328–2339.
Kvadsheim P., F. Benders, P. Miller, L. Doksæter, F. Knudsen, P. Tyack, N. Nordlund, F-P Lam, F.Samarra, L. Kleivane
, O.R. Godo, 2007. Herring (sild), killer whales (spekkhogger) and sonar
– the 3S-2006 cruise report with preliminary results. FFI-rapport 2007/01189: 79pp.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 172
Kvadsheim P., F.-P. Lam, P. Miller, A.C. Alves, R. Antunes, A. Bocconcelli, S. van Ijsselmuide,Lars Kleivane, M. Olivierse, F. Visser, 2009. Cetaceans and naval sonar – the 3S-2009 cruisereport. FFI-rapport 2009/01140: 133pp.
Laran S., Dulau-Drouot V., 2007. Seasonal variation of striped dolphins, fin- and sperm whales’abundance in the Ligurian Sea (Mediterranean Sea). J. Mar. Biol. Ass. U.K. 87: 345–352
Laroche C., R. Hétu, S. Poirier, 1989. The growth of and recovery from TTS in human subjectsexposed to impact noise. J. Acoust. Soc. Am. 88(4): 1681-1690.
Li S., D. Wang, K. Wang, E.A. Taylor, E. Cros, W. Shi, Z. Wang, L. Fang, Y. Chen, F. Kong, 2012.Evoked-potential audiogram of an Indo-Pacific humpback dolphin (Sousa chinensis). TheJournal of Experimental Biology 215: 3055-3063
Lucke K., U. Siebert, P.A. Lepper, M.-A. Blanchet, 2009. Temporary shift in masked hearingthresholds in a harbour porpoise (Phocoena phocoena) after exposure to seismic airgunstimuli. J. Acoust. Soc. Am. 125(6): 4060-4070.
Lurton X., L. Antoine, 2007. Analyse des risques pour les mammifères marins liés à l’emploi desméthodes acoustiques en océanographie. Rapport final DOP/CB/NSE/AS/07-07 : 88p
Madsen P.T., B. Møhl, 2000. Sperm whales (Physeter catodon L. 1758) do not react to soundsfrom detonators. J.Acoust.Soc.Am. 107(1): 668-671.
Madsen P.T., M. Wahlberg, B. Møhl, 2002. Male sperm whale (Physeter macrocephalus) acousticsin a high-latitude habitat: implications for echolocation and communication. Behav. Ecol.Sociobiol. 53: 31-41.
Madsen P.T., I. Kerr, R. Payne, 2004. Source parameter estimates of echolocation clicks from wildpygmy killer whales (Feresa attenuata) (L). J.Acoust.Soc.Am. 116(4) Pt. 1: 1909-1912.
Madsen P.T., I. Kerr, R. Payne, 2004. Echolocation clicks of two free-ranging, oceanic delphinidswith different food preferences: false killer whales Pseudorca crassidens and Risso’s dolphinsGrampus griseus. The Journal of Experimental Biology 207: 1811-1823.
Madsen P.T., M. Johnson, P.J.O. Miller, N. Aguilar Soto, J. Lynch, P. Tyack, 2006. Quantitativemeasures of air-gun pulses recorded on sperm whales (Physeter macrocephalus) usingacoustic tags during controlled exposure experiments. J.Acoust.Soc.Am. 120(4): 2366–2379.
Madsen P. T., N. Aguilar de Soto, P. Arranz, M. Johnson, 2013. Echolocation in Blainville’s beakedwhales (Mesoplodon densirostris). J Comp Physiol A 199: 451-469.
May-Collado L J., D. Wartzok, 2007. The freshwater dolphin Inia geoffrensis geoffrensis produceshigh frequency whistles. J. Acoust. Soc. Am. 121(2): 1203-1212.
May-Collado L.J., D. Wartzok, 2009. A characterization of Guyana dolphin (Sotalia guianensis)whistles from Costa Rica: the importance of broadband recording systems. J. Acoust. Soc.Am. 125(3): 1202–1213
MacCauley R.D., Fewtrell J., Duncan a.J., et al., 2000. Marine seismic surveys: a study ofenvironmental implications. APPEA Journal: 692-708
McDonald M.A., J.A. Hildebrand, S.M. Wiggins, 2006. Increases in deep ocean ambient noise inthe Northeast Pacific west of San Nicolas Island, California. J.Acoust.Soc.Am. 120(2): 711-718.
Maybaum H. L., 1993. Response of humpback whales to sonar sounds. J.Acoust.Soc.Am. 94(3) pt2; 126 th Meeting of the AAA. Abstract.
Melnick W., 1991. Human temporary threshold shift (TTS) and damage risk. J.Acoust. Soc.Am.90(1): 147-154.
Mercado E. III, J.N. Schneider, A.A. Pack, L.M. Herman, 2010. Sound production by singinghumpback whales. J. Acoust. Soc. Am. 127(4): 2698-2691.
Miller P., Kvadsheim P., Lam F.-P., Tyack P.T., 2009. Behaviour of killer, sperm and long-finnedpilot whales during CEE with sonar signals. Présentation orale à ESOMM3, Lerici (7-9 sept09).

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 173
Miller P.J.O., M.P. Johnson, P.T. Madsen, N. Biassoni, M. Quero, P.L. Tyack, 2009. Using at-seaexperiments to study the effects of airguns on the foraging behavior of sperm whales in theGulf of Mexico. Deep-Sea Research I 56:1168–1181.
Miller P., Antunes R., A.C. Alves, P. Wensveen, P. Kvadsheim, L. Kleivane, N. Nordlund, F.-P.Lam, S. van IJsselmuide, F. Visser, P. Tyack, 2011. The 3S experiments: studying thebehavioural effects of naval sonar on killer whales (Orcinus orca), sperm whales (Physetermacrocephalus), and long-finned pilot whales (Globicephala melas) in Norwegian waters.Scottish Oceans Inst. Tech. Rept., SOI-2011-001: 290pp.
Mooney T.A., P.E. Nachtigall, M. Breese, S. Vlachos, W.W.L. Au, 2009. Predicting temporarythreshold shifts in a bottlenose dolphin (Tursiops truncatus): the effects of noise level andduration. J.Acoust. Soc.Am. 125(3): 1816–1826
Mooney T.A., P.E. Nachtigall, S. Vlachos, 2009. Sonar-induced temporary hearing loss in dolphins.Biol. Lett. (on line): 4pp.
Nachtigall P.E., J.L. Pawloski, W.W.L. Au, 2003. Temporary threshold shifts and recovery followingnoise exposure in the Atlantic bottlenosed dolphin (Tursiops truncatus). J.Acoust.Soc.Am.113(6): 3425-3429.
Nachtigall P.E., A.Y. Supin, J. Pawloski, W.W.L. Au, 2004. Temporary threshold shift after noiseexposure in the bottlenose dolphin (T. truncatus) measured using evoked auditory potentials.Mar.Mam.Sci. 20(4): 673-687.
Nachtigall P.E., M M. L. Yuen, T.A. Mooney, K.A. Taylor, 2005. Hearing measurements from astranded infant Risso’s dolphin, Grampus griseus. Journal of Experimental Biology 208 : 4181-4188.
Nachtigall P.E., T.A. Mooney, K.A. Taylor, M.M.L. Yuen, 2007. Hearing and auditory evokedpotential methods applied to odontocete cetaceans. Aquatic Mammals 33(1): 6-13,
National Research Council, 2003. Ocean Noise and Marine Mammals. Report of the NationalAcademy of Sciences: 192 pp.
Nedwell J.R., Parvin S.J, Edwards B., Workman R., Brooker A.G., Kynoch J.E., 2007.Measurement and interpretation of underwater noise during construction and operation ofoffshore windfarms in UK waters. Subacoustech Report No. 544R0738 to COWRIE Ltd. ISBN:978-0-9554279-5-4.
Nedwell J.R., Brooker A G, 2008. Measurement and assessment of background underwater noiseand its comparison with noise from pin pile drilling operations during installation of the SeaGentidal turbine device, Strangford Lough. Subacoustech Report No. 724R0120 to COWRIE Ltd.ISBN: 978-0-9557501-9-9.
NURC, 2006. NATO Undersea Research Centre Human Diver and Marine Mammal Risk MitigationRules and Procedures - Marine Mammal Risk Mitigation Project. NURC Special PublicationNURC-SP-2006-008: 30pp.
OSPAR Commission, 2009. Overview of the impacts of anthropogenic underwater sound in themarine environment. Biodiversity Series. 134pp.
Oswald J.N., S. Rankin, J. Barlow, 2004. The effect of recording and analysis bandwidth onacoustic identification of delphinid species. J. Acoust. Soc. Am. 116(5): 3178-3185.
Pacini A.F., P.E. Nachtigall, L.N. Kloepper, M. Linnenschmidt, A. Sogorb, S. Matias, 2010.Audiogram of a formerly stranded long-finned pilot whale (Globicephala melas) measuredusing auditory evoked potentials. The Journal of Experimental Biology 213: 3138-3143
Pacini A.F., P.E. Nachtigall, L.N. Kloepper, M. Linnenschmidt, 2010. Hearing measurement of astranded pygmy killer whale. Conference of the ECS (Stralsund), poster.
Pacini A.F., P.E. Nachtigall, C.T. Quintos, T.D. Schofield, D.A. Look, G.A. Levine, J.P. Turner,2011. Audiogram of a stranded Blainville’s beaked whale (Mesoplodon densirostris) measuredusing auditory evoked potentials. The Journal of Experimental Biology 214: 2409-2415.
Philips J.D., P.E. Nachtigall, W.W.L. Au, J.L. Pawloski, H.L. Roitblat, 2003. Echolocation in theRisso’s dolphin, Grampus griseus. JASA 113(1): 605-616.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 174
Pirotta E., R. Milor, N. Quick, D. Moretti, N. Di Marzio, P. Tyack, I. Boyd, G. Hastie, 2012. Vesselnoise affects beaked whale behavior: results of a dedicated acoustic response study. PLosOne 7(8): 8pp.
Popov V.V., Klishin V.O., 1998. EEG study of hearing in the common dolphin, Delphinus delphis.Aquatic Mammals 24(1): 13-20.
Popov V.V., A.Ya. Supin, D. Wang, K. Wang, J Xiao, S. Li, 2005. Evoked-potential audiogram of theYangtze finless porpoise Neophocaena phocaenoides asiaeorientalis (L). JASA 117(5): 2728-2731.
Popov V.V., A.Ya. Supin, D. Wang, K. Wang, L. Dong, S. Wang, 2011.Noise-induced temporarythreshold shift and recovery in Yangtze finless porpoises Neophocaena phocaenoidesasiaeorientalis. J. Acoust. Soc. Am. 130(1): 574-584.
Popov V.V., A.Ya. Supin, V.V. Rozhnov, D.I. Nechaev, E.V. Sysuyeva, V.O. Klishin, M.G.Pletenko, M.B. Tarakanov, 2013. Hearing threshold shifts and recovery after noise exposure inbeluga whales, Delphinapterus leucas. The Journal of Experimental Biology 216, 1587-1596.
Prézelin B., 2008. Flottes de combat. Editions maritimes et d’Outremer : 1312pp.
Price G.R., 1981. Implications of a critical level in the ear for assessment of noise hazard at highintensities. JASA 69(1): 171-177.
Rendell L., Gordon J., 1999. Vocal Response of long-finned pilot whales to military sonar in theLigurian Sea. Mar.Mam.Sci. 15(1): 198-204
Rendell L.E., J.N. Matthews, A. Gill, J.C.D. Gordon, D.W. Macdonald, 1999. Quantitative analysisof tonal calls from five odontocete species, examining interspecific and intraspecific variation.J. Zool. Lond. 249: 403-410.
Richardson W.J., Greene C.R., Malme C.I., Thomson D.H., 1995. Marine Mammals and Noise.Academic Press (San Diego, London): 576pp.
Ridgway S.H., Carder D.A., 2001. Assessing hearing and sound production in cetaceans notavailable for behavioral audiograms: experiences with sperm, pygmy sperm and gray whales.Aquatic Mammals 27(3): 267-276.
Riesch R.D., Ford J.K.B., Thomsen F., 2005. Stability and group specificity of stereotyped whistlesin resident killer whales, Orcinus orca, off British Columbia. Animal Behaviour: 13pp
Roberto M., Hamernick R.P., Salvi R.J., Henderson D., Milone R., 1985. Impact noise and theequal energy hypothesis. JASA 77(4): 1514-1520.
Rolland R.M., S.E. Parks, K.E. Hunt, M. Castellote, P.J. Corkeron, D.P. Nowacek, S.K. Wasser,S.D. Kraus, 2012. Evidence that ship noise increases stress in right whales. Proc. R. Soc. B279: 2363-2368.
Rutenko A.N., S.V. Borisov, A.V. Gritsenko, M.R. Jenkerson, 2007. Calibrating and monitoring thewestern gray whale mitigation zone and estimating acoustic transmission during a 3D seismicsurvey, Sakhalin Island, Russia. Environ. Monit. Assess. 134: 21–44
Sauerland M., G. Dehnhardt, 1998. Underwater audiogram of a tucuxi (Sotalia fluviatilisguianensis). J. Acoust. Soc. Am. 103(2): 1199-1204.
Saunders J.C., Dear S.P., Schneider M.E.., 1985. The anatomical consequences of acoustic injury:a review and tutorial. JASA 78(3): 833-860.
Schlundt C.E., J.J. Finneran, D.A. Carder, S.H. Ridgway J., 2000. Temporary shift in maskedhearing thresholds of bottlenose dolphins, Tursiops truncatus, and white whales,Delphinapterus leucas, after exposure to intense tones. Acoust. Soc. Am. 107(6): 3496-3508.
Schlundt C.E., R.L. Dear, D.S. Houser, A.E. Bowles, T. Reidarson, J.J. Finneran, 2011. Auditoryevoked potentials in two short-finned pilot whales (Globicephala macrorhynchus). J. Acoust.Soc. Am. 129(2): 1111-1116.
Schultz M., Betke K., Nehls G., 2006. Underwater noise reduction of pile driving for offshore windturbines – Evaluation of several techniques under offshore conditions. Report summary: 5pp.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 175
Soldevilla M.S., M.F. McKenna, S.M. Wiggins, R.E. Shadwick, T.W. Cranford, J.A. Hildebrand,2007. Cuvier’s beaked whale (Ziphius cavirostris) head tissues: physical properties and CTimaging. J Exp Biology 208: 2319-2332
Southall B.L., Scholik-Schlomer A., 2008. Final report of the NOAA International Conference:Potential Application of Vessel-Quieting Technology on Large Commercial Vessels (1-2 May,2007, Silver Spring, MD, U.S.A.). 47pp
Southall B.L., R Braun, F M.D. Gulland, A D., Heard, R W. Baird, S M. Wilkin, T K. Rowles, 2006.Hawaiian Melon-headed Whale (Peponocephala electra) mass stranding event of July 3-4,2004. NOAA Technical Memorandum NMFS-OPR-31- April: 78pp.
Southall B.L., A.E. Bowles, W.T. Ellison, J.J. Finneran, R.L. Gentry, C.R. Greene, D. Kastak, D.R.Ketten, J.H. Miller, P.E. Nachtigall, W.J. Richardson, J.A. Thomas, P.L. Tyack, 2007. Marinemammal noise exposure criteria: initial scientific recommendations. Aquatic Mammals 33(4):411-521.
Southall B., J. Calambokidis, P. Tyack, D. Moretti, J, Hildebrand, C., Kyburg, R. Carlson, A.Friedlaender, E. Falcone, G. Schorr, A. Douglas, S. DeRuiter, J. Goldbogen, J. Barlow, 2011.Biological and Behavioral Response Studies of Marine Mammals in Southern California, 2010(“SOCAL‐10”). PROJECT REPORT, 26 February 2011 : 29pp.
Southall B., J. Calambokidis, P. Tyack, D. Moretti, A. Friedlaender, S. DeRuiter, J. Goldbogen, E.Falcone, G. Schorr, A. Douglas, A. Stimpert, J. Hildebrand, C. Kyburg, R. Carlson, T. Yack, J.Barlow, 2012. Biological and Behavioral Response Studies of Marine Mammals in SouthernCalifornia, 2011 (“SOCAL‐11”). FINAL PROJECT REPORT, 8 March 2012 : 55pp.
Southall B., J. Calambokidis, Barlow, D. Moretti, A. Friedlaender, A. Stimpert, A. Douglas, K.Southall, P. Arranz, S. DeRuiter, E. Hazen, J. Goldbogen, E. Falcone, G. Schorr, 2013.Biological and Behavioral Response Studies of Marine Mammals in Southern California, 2012(“SOCAL‐12”). FINAL PROJECT REPORT, 4 April 2013 : 40pp.
Stone C.J., 2003. The effects of seismic activity on marine mammals in UK waters, 1998-2000.JNCC Report No. 323: 78pp.
Stone C.R. & Tasker M., 2006. The effects of seismic airguns on cetaceans in UK waters. J.C.R.M.8(3): 255-263.
Szymanski M.D., D.E. Bain, K. Kiehl, S. Pennington, S. Wong, K.R. Henry, 1999. Killer whale(Orcinus orca) hearing: Auditory brainstem response and behavioral audiograms. J. Acoust.Soc. Am. 106(2): 1134-1141
Thomas J., Chun N., Au W., Pugh K., 1988. Underwater audiogram of a false killer whale(Pseudorca crassidens). J. Acoust. Soc. Am. 84(3): 936-940.
Thompson P.M., Brookes K.L., Graham I.M., Barton T.M., Needham K., Bradbury G., MerchantN.D., 2013. Short-term disturbance by a commercial two-dimensionnal seismic survey doesnot lead to long-term displacement of harbour porpoises. Proc.T.Soc. B 280: 9pp.
Tougaard J., Damsgaard Henriksen O., L.A. Miller, 2009. Underwater noise from three types ofoffshore wind turbines: estimation of impact zones for harbor porpoises and harbor seals. J.Acoust. Soc. Am. 125(6): 3755-3773.
Tremel D.P., Thomas J.A., Ramirez K.T., Dye G.S., Bachman W.A., Orban A.N., Grimm K.K.,1998. Underwater hearing sensitivity of a Pacific white-sided dolphin, Lagenorhynchusobliquidens. Aquatic Mammals 24(2): 63-69
Tubelli A.A., A. Zosuls, D.R. Ketten, M. Yamato, D.C. Mountain, 2012. A prediction of the minkewhale (Balaenoptera acutorostrata) middle-ear transfer function. J. Acoust. Soc. Am. 132(5):3263-3272.
Urick R.J., 1975. Principles of Underwater Acoustics (2nd Edition). McGraw-Hill Book Company:384 pp.
Van Bree P.J.H., Kristensen I., 1974. In the intriguig stranding of four Cuvier's beaked whales, Zcavirostris, on the lesser antillean island of Bonaire. Bijdragen Tod De Tierkunde 44(2): 235-238.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 176
Verboom W.C., R.A. Kastelein, 2005. Some examples of marine mammal ‘discomfort thresholds’ inrelation to man-made noise. Proceedings of the European conference on Undersea DefenceTechnology, UDT Europe 2005. June 21-23, Amsterdam, The Netherlands.
Wang J Y., S. Yang, 2006. Unusual cetacean stranding events of Taiwan in 2004 and 2005. J.Cetacean Res. Manag. 8(3): 283–292.
Watkins W.A., M.A Daher, K.M. Fristrup, T.J. Howald, G. N Di Sciara, 1993. Sperm whale taggedwith transponders and tracked underwater by sonar. Mar.Mamm.Sci. 9(1): 55-67.
Watwood S.L., P.J.O. Miller, M. Johnson, P.T. Madsen & P.L. Tyack, 2006. Deep-diving foragingbehaviour of sperm whales (Physeter macrocephalus). Journal of Animal Ecology 75: 814–825.
Weir C.R., 2008. Short-finned pilot whales (Globicephala macrorhynchus) respond to an airgunramp-up procedure off Gabon. Aquatic Mammals 34: 349–354.
Würsig B., Greene C.R., Jefferson T.A, 2000. Development of an air bubble curtain to reduceunderwater noise of percussive piling. Mar. Environ. Res. 49: 79–93.
Yang W-C., L-S. Chou, P. D. Jepson, R. L. Brownell, Jr, D. Cowan, P-H. Chang, H-I. Chiou, C-J.Yao, T. K. Yamada, J-T. Chiu, P-J. Wang, A. Fernández, 2008. Unusual cetacean mortalityevent in Taiwan, possibly linked to naval activities. Veterinary Record 162: 184-186
Zimmer W.M., 2004. Sonar systems and stranding of beaked whales. Proceedings of the workshopon active sonars and cetaceans (La Palmas, 8 March 2003): 8-13.
Zimmer W.M. X., M.P. Johnson, P.T. Madsen, P.L. Tyack, 2005. Echolocation clicks of free-ranging Cuvier’s beaked whales (Ziphius cavirostris). JASA 117(6): 3919-3927.

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 177
Index des figures
Figure 1 : Abaque de bruit ambiant sous-marin (d’après Richardson et al., 1995)......................... 12
Figure 2 : Réception du son chez les odontocètes (d’après Ketten in Au et al., 2000). ................. 14
Figure 3 : Oreille moyenne et interne de dauphin (tiré de Thewissen in Perrin et al., 2002) .......... 14
Figure 4 : Audiogrammes d'odontocètes existant en 2002 (extrait de NRC, 2003)........................ 15
Figure 5 : Anatomie de l’appareil émetteur sonore du Cachalot (d’après K.Das, 2003)................. 18
Figure 6 : Anatomie de l’appareil émetteur sonore chez le Grand dauphin (d’après K.Das, 2003) 19
Figure 7 : Emissions sonores de cétacés (extrait de NRC, 2003). (Cuvier's B W = ziphius, Pilot W =
globicéphale, Striped D = dauphin bleu et blanc, Sperm W = cachalot) ........................................ 20
Figure 8 : Spectrogramme d’un sifflement de Dauphin bleu et blanc............................................. 25
Figure 9 : Spectrogramme d’un train d’impulsions de Dauphin bleu et blanc................................. 25
Figure 10 : Spectrogramme de vocalisations de Dauphin commun............................................... 27
Figure 11 : Spectrogramme d’un sifflement de Grand Dauphin. .................................................... 28
Figure 12 : Spectrogramme de bourdonnement de Dauphin de Risso. ......................................... 29
Figure 13 : Spectrogramme de cri complexe pulsé de Globicéphale noir. ..................................... 30
Figure 14 :Spectrogramme de clics de Ziphius. ............................................................................ 32
Figure 15 : Spectrogramme de clic de Cachalot............................................................................ 33
Figure 16 : Spectrogramme de creak de Cachalot. ....................................................................... 34
Figure 17 : Spectrogramme d’un appel de Rorqual commun. ....................................................... 35
Figure 18 : Prévision du niveau de pression en fonction de la charge d’explosifs (Richardson et al.,
1995, p151)................................................................................................................................... 41
Figure 19 : Spectre d'énergie de l'impulsion d’un marteau-pilon à 100m et à 10 km de la source
(Nedwell et al., 2007). ................................................................................................................... 43
Figure 20 : Bruits enregistrés le long d'une radiale lors du forage d'un substrat rocheux (d'après
Nedwell & Brooker, 2008) ............................................................................................................. 46
Figure 21 : Directivité de canons pneumatiques multiples (Richardson et al., 1995 p138) ............ 49
Figure 22 : Impulsion sonore à 1m mesurée en dessous d'un canon de 72 litres (NRC, 2003 p60)
..................................................................................................................................................... 50
Figure 23 : Spectre d'amplitude à 1m d'un canon de 72 litres (NRC, 2003 p60)............................ 50
Figure 24 : Spectres d'amplitude à 2,5 et 10 km d'un canon (Goold, 1995 in Evans & Nice, 1997).
..................................................................................................................................................... 51
Figure 25 : Champ acoustique propagé pour la sismique multi-traces de 252 dB de l'IFREMER,
pour sa fréquence d'énergie maximale (< 100Hz), d'après Lurton et Antoine, 2007...................... 53
Figure 26 : Effet des sismiques sur le taux de détection des cétacés (d'ap. Stone & Tasker, 2006).
..................................................................................................................................................... 57
Figure 27 : Impulsion de canon airgun reçue par un cachalot à 3 km et 15 m de profondeur. ....... 59

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 178
Figure 28 : Configuration générale d'un sonar SURTASS-LFA (d'après FSEIS, 2007). ................ 66
Figure 29 : Niveaux sonores moyens en tiers d'octave reçus sur 9 enregistreurs autonomes en
avril et main 2006 sur le sanctuaire marin de Stellwagen Bank (d'après Hatch et al., 2008). ........ 79
Figure 30 : Spectre des bruits de jets ski (d’après Erbe, 2013). .................................................... 80
Figure 31 : Spectre du bruit de deux semi-rigides avec des moteurs hors-bord, montrant la
différence de niveau de source pour deux engins à vitesses différente (d’après Erbe, 2002)........ 80
Figure 32 : Effet du bruit d’un bateau sur la sonde d’un Ziphius (Aguilar Soto et al., 2006) ........... 82
Figure 33 : Organe de Corti (section radiale de la cochlée) avec les cellules ciliées "hair cells"
(Saunders et al., 1985) ................................................................................................................. 88
Figure 34 : Valeur et relaxation dans le temps des TTS après une insonification longue à 4-11 kHz
(Nachtigall et al., 2004). ................................................................................................................ 92
Figure 35 : Audiogramme moyen du dauphin et TTS trouvés aux différentes fréquences (Mooney
et al., 2009b)................................................................................................................................. 93
Figure 36 : Relaxation des TTS à la fréquence de 11,2 kHz en fonction des durées d'exposition,
(Mooney et al., 2009b). ................................................................................................................. 93
Figure 37 : Niveau d'exposition net causant un TTS en fonction des durées d'expositions (Mooney
et al., 2009b)................................................................................................................................. 94
Figure 38 : Evolution du TTS mesuré à 4,5 kHz pour une insonification à 3 kHz de durée variable
(Finneran et al., 2005)................................................................................................................... 96
Figure 39 : Relaxation du TTS à 4,5 kHz pour une insonification à 3 kHz avec des doses
d’insonification variables (Finneran et al., 2005). .......................................................................... 96
Figure 40 : Pertes auditives TTS engendrées à 30 et 40 kHz par une insonification de 64 sec et
186 dB à 20 kHz (méthode comportementale) (Finneran et al., 2007). ......................................... 98
Figure 41 : Variations de l'audiogramme engendrées par une insonification de 64 sec et 185 dB à
20 kHz (audimétrie par méthode AEP) (Finneran et al., 2007). ..................................................... 98
Figure 42 : Evolution de la perte auditive (TTS4) en fonction du niveau sonore pour une durée
d’exposition de 16 s et les fréquences de 3 et 20 kHz (Finneran et al., 2010a). ............................ 99
Figure 43 : Evolution de la perte auditive (TTS4) pour la fréquences de 3 kHz en fonction du
niveau sonore et de la durée d’exposition de 4 à 100 s et (Finneran et al., 2010b) ..................... 100
Figure 44 : Seuils d'audition du marsouin mesuré par AEP (Lucke et al., 2009). Triangles ......... 103
Figure 45 : Pertes auditives TTS à 4 kHz pour le Marsouin commun (Lucke et al., 2009). .......... 104
Figure 46 : Réactions comportementales en fonction des niveaux reçus (d’après le tableau) ..... 112
Figure 47 : Réactions comportementales en fonction des niveaux perçus (d’après les courbes
audiométriques) .......................................................................................................................... 112
Figure 48 : Augmentation des distances d’observation pour les petits odontocètes, entre une
situation de tir sismique (carré noir) et de non-activité sismique (triangle blanc). ........................ 114
Figure 49 : Scénarios pouvant conduire à la mort de cétacés en raison d’une insonification....... 122

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 179
Figure 50 : Zonage des perturbations encourues par les cétacés au cours d’une insonification.. 124
Figure 51 : Problématique de la prévision de l’impact d’une insonification sur les cétacés.......... 126
Figure 52 : Rappel du zonage autour d’une source sonore. ........................................................ 130
Figure 53 : Audiogramme de Dauphin bleu et blanc (d’après Kastelein et al., 2003)................... 135
Figure 54 : Audiogramme de Dauphin commun (d’après Popov & Klishin, 1998, modifié). ......... 136
Figure 55 : Audiogramme de Grand dauphin (d’après Johnson, 1966 in Au et al., 2000). ........... 137
Figure 56 : Audiogramme de Dauphin de Risso (d’après Nachtigall et al., 2005, modifié)........... 137
Figure 57 : Audiogramme de Globicéphale noir (d’après Pacini et al., 2010, modifié). ................ 138
Figure 58 : Audiogramme de Ziphius (d’après l’audiogramme de Mésoplodon de Blainville de
Pacini et al., 2011, modifié). ........................................................................................................ 139
Figure 59 : Audiogramme synthétique de Cachalot (élaboré d’après des données variées)........ 139
Figure 60 : Audiogramme synthétique de Rorqual commun (élaboré d’après des données variées).
................................................................................................................................................... 140
Figure 61 : Valeurs approximatives du coefficient d’absorption en fonction de la fréquence. ...... 143
Figure 62: Pertes par propagation sphérique en fonction de la distance. .................................... 143
Figure 63 : Pertes par propagation pour une source immergée à 10 m....................................... 144
Figure 64: Pertes par propagation pour une source immergée à 10 m - zone proche. ................ 145
Figure 65 : Pertes par propagation pour une source immergée à 200 m..................................... 145
Figure 66 : Pertes par propagation pour une source immergée à 200 m (source proche). .......... 146
Figure 67 : Exemple de transmission sonar par un navire US en janvier 2006............................ 148
Figure 68 : Carte de l’impact comportemental du scénario sur les Rorquals communs. .............. 151
Figure 69 : Niveaux perçus par les Cachalots au cours du scénario de transmission.................. 152
Figure 70: Carte de l’impact comportemental du scénario sur les Cachalots............................... 152
Figure 71 : Carte de l’impact comportemental du scénario sur les Dauphins de Risso................ 153
Figure 72 : Niveaux perçus par les Dauphins de Risso au cours du scénario de transmission.... 154
Figure 73 : Niveaux perçus par les Dauphins de Risso au cours du scénario de transmission.... 154
Figure 74 : Carte de l’impact comportemental du scénario sur les Dauphins bleus et blancs. ..... 155
Figure 75 : Doses perçues par les Dauphins bleus et blancs au cours du scénario de transmission.
................................................................................................................................................... 155
Figure 76 : Influence des seuils et vitesses sur les doses d’exposition sonores des dauphins. ... 161
Figure 77 : Application de la simulation du scénario sonar à une population de Ziphius. ............ 162
Figure 78 : Comparaison des audiogrammes du Dauphin bleu et blanc et du Globicéphale noir.164
Figure 79 : Pondérations fréquentielles pour le Dauphin bleu et blanc - comparaison................. 165

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 180
Index des tableaux
Tableau 1 : Liste des audiogrammes de cétacés publiés (janv. 2014) _____________________ 17
Tableau 2 : Domaines de fréquence et durée des cris et sifflements de quelques espèces. ____ 20
Tableau 3 : Domaines de fréquence et niveau de source des clics d'écholocalisation de quelques
odontocètes. _________________________________________________________________ 21
Tableau 4 : Eléments préliminaires sur les résultats et la sensibilité aux nuisances sonores des
espèces focales. ______________________________________________________________ 22
Tableau 5 : Enregistrements de dauphins bleus et blancs (activables par lien hypertexte). ____ 26
Tableau 6 : Enregistrements de dauphins communs (activables par lien hypertexte)._________ 27
Tableau 7 : Enregistrements de grands dauphins (activables par lien hypertexte). ___________ 28
Tableau 8 : Enregistrements de Dauphins de Risso (activables par lien hypertexte).(provisoire) 29
Tableau 9 : Enregistrements de Globicéphale noir (activables par lien hypertexte). __________ 31
Tableau 10 : Enregistrements de Ziphius en Méditerranée. _____________________________ 32
Tableau 11 : Enregistrements de Cachalot (activables par lien hypertexte). ________________ 34
Tableau 12 : Enregistrements de Rorqual commun (activables par lien hypertexte).__________ 35
Tableau 13 : Critère de risque de dommage auditif définitif pour l'être humain, extrapolé pour des
mammifères marins (adapté de Richardson et al., 1995). ______________________________ 56
Tableau 14: Caractéristiques de sonars actifs utilisés dans différentes activités humaines. ____ 64
Tableau 15 : Echouages accidentels de cétacés liés à l'emploi de sonars militaires. _________ 68
Tableau 16 : Expériences de CEE durant le programme Bahamas BRS 2007 et 2008. _______ 71
Tableau 17 : Expériences de transmission réalisées durant le programme 3S de 2006 à 2009 _ 72
Tableau 18 : Expériences de transmission réalisées durant le programme 3S en 2011 et 2012_ 73
Tableau 19 : Expériences de transmission réalisées durant le programme SOCAL de 2010 à 2012.
___________________________________________________________________________ 74
Tableau 20 : Synthèse des insonifications et TTS des dauphins et des bélougas en sons purs. 76
Tableau 21 : Insonifications à 4-11 kHz et TTS d’un grand dauphin (Au et al., 1999 ; Nachtigall et
al., 2003). ___________________________________________________________________ 92
Tableau 22 : Insonifications et MTTS pour des Grands dauphins exposés à des sons purs de 1 s
de durée et de fréquence variable (d'après Schlundt et al., 2000). _______________________ 95
Tableau 23 : Insonifications et TTS des dauphins insonifiés à 3 kHz avec des durées variables
(d'après Finneran et al., 2005).___________________________________________________ 95
Tableau 24: Synthèse des insonifications en sons purs et des TTS observés chez le Grand
dauphin. ___________________________________________________________________ 101
Tableau 25: Synthèse des insonifications et MTTS en sons impulsifs. ___________________ 104
Tableau 26 : Insonifications et MTTS des bélougas (d'après Schlundt et al., 2000). _________ 105
Tableau 27 : Insonifications et TTS pour 4 espèces en sons non-impulsifs. _______________ 106

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 181
Tableau 28 : Différents programmes d’expériences d’exposition contrôlée. _______________ 109
Tableau 29 : Différents points expérimentaux obtenus lors d’expériences de CEE. _________ 111
Tableau 30 : Gradation des effets induits par les pollutions sonores (à partir de résultats relevés en
bassin et en liberté sur plusieurs espèces). ________________________________________ 118
Tableau 31 : Niveaux sonores pour les différentes catégories d’impact dans le cas de bruits non
impulsifs. ___________________________________________________________________ 128
Tableau 32 : Niveaux sonores (dBp-p) pour les différentes catégories d’impact dans le cas de
bruits impulsifs. ______________________________________________________________ 132
Tableau 33 : Définition du peuplement local de cétacés ______________________________ 149
Tableau 34 : Paramètres utilisés pour évaluer la sensibilité de la prévision (Dauphin bleu et blanc)
__________________________________________________________________________ 160

L'impact des nuisances acoustiques sur les cétacés du Sanctuaire et de la Méditerranée nord-occidentale
Dr Alexandre Gannier – Klymene Recherche Marine – Sanctuaire Pelagos partie française 182
Annexes
Annexe 1 (électronique) : Bibliothèque des sons inclus dans le rapport.
note 1 : pour que les liens hypertextes fonctionnent, il faut impérativement que le rapport soit situé
dans un répertoire « Documents/AcouPelagos » et que les fichiers sons (.wav) soient dans
« Documents/AcouPelagos/SonsPelagos ».
note 2 : les fichiers sont font partie intégrante de l’étude et ne doivent pas être distribués
séparément.
Annexe 2 (électronique) : Références bibliographiques existant en fichiers pdf.
Les fichiers de la bibliothèque peuvent être placés dans un répertoire
« Documents/AcouPelagos/BiblioPelagos ».
Annexe 3 (électronique) : Feuille de calcul de la propagation et du niveau reçu.
Cette feuille de calcul ne peut pas être séparée du rapport d’étude et ne doit pas être distribuée
sans autorisation des auteurs.


















