ÉTUDE CLINIQUE ET GÉNÉTIQUE DE L’ATAXIE RÉCESSIVE DE LA … · périphériques normales et...
45
NICOLAS DUPRÉ ÉTUDE CLINIQUE ET GÉNÉTIQUE DE L’ATAXIE RÉCESSIVE DE LA BEAUCE Mémoire présenté à la Faculté des études supérieures de l’Université Laval dans le cadre du programme de Maîtrise en Épidémiologie pour l’obtention du grade de Maître ès Sciences (M.Sc.) DÉPARTEMENT DE MÉDECINE SOCIALE ET PRÉVENTIVE FACULTÉ DE MÉDECINE UNIVERSITÉ LAVAL QUÉBEC 2008 ©Nicolas Dupré, 2008
Transcript of ÉTUDE CLINIQUE ET GÉNÉTIQUE DE L’ATAXIE RÉCESSIVE DE LA … · périphériques normales et...

NICOLAS DUPRÉ
ÉTUDE CLINIQUE ET GÉNÉTIQUE DE L’ATAXIE
RÉCESSIVE DE LA BEAUCE
Mémoire présenté
à la Faculté des études supérieures de l’Université Laval
dans le cadre du programme de Maîtrise en Épidémiologie
pour l’obtention du grade de Maître ès Sciences (M.Sc.)
DÉPARTEMENT DE MÉDECINE SOCIALE ET PRÉVENTIVE
FACULTÉ DE MÉDECINE
UNIVERSITÉ LAVAL
QUÉBEC
2008
©Nicolas Dupré, 2008

II
Résumé
Un échantillon de 64 patients répertoriés au sein de 30 familles d’origine Canadienne-
Française montrant des caractéristiques cliniques très semblables et provenant de la même
région du Québec a été évalué de façon détaillée. Une histoire clinique standardisée, un
examen neurologique ciblé, une imagerie cérébrale, des conductions nerveuses
périphériques, et une analyse du gène SYNE1 ont été réalisés sur tous les patients
disponibles. En se basant sur l’échantillon répertorié, l’Ataxie Récessive de la Beauce est
un syndrome cérébelleux dégénératif caractérisé par un âge de début dans la trentaine, une
évolution lente, un degré de handicap modéré, une dysarthrie significative, de légers signes
oculomoteurs, parfois des réflexes vifs aux membres inférieurs, des conductions nerveuses
périphériques normales et une atrophie cérébelleuse diffuse à l’imagerie. Dans notre
échantillon, nous avons identifié 7 mutations, démontrant ainsi l’hétérogénéité génétique.
Les patients présentant des mutations différentes ne présentaient pas d’atteintes cliniques
permettent de les différencier. Il est probable que cette nouvelle maladie devienne une
cause fréquente, à travers le monde, d’ataxie héréditaire débutant à un âge moyen.

III
Summary
We ascertained 64 probands and affected members of 30 French-Canadian families all
showing similar clinical features and originating from the same region of Quebec. After
informed consent, we performed detailed clinical history, neurological examination, brain
imaging, nerve conduction studies, and SYNE1 mutation detection of all available subjects.
Based on the cases examined, Autosomal Recessive Cerebellar Ataxia Type 1 is a
cerebellar syndrome characterized by: recessive transmission; middle-age onset (mean
31.60 years, range 17-46); slow progression and moderate disability; significant dysarthria;
mild oculomotor abnormalities; occasional brisk reflexes in the lower extremities; normal
nerve conduction studies and diffuse cerebellar atrophy on imaging. We identified a total
of 7 mutations in our population, thereby providing evidence of genotypic heterogeneity.
Patients with different mutations did not show significant phenotypic heterogeneity. We
expect that this disease will be a common cause of middle-age onset recessive ataxia
worldwide.

IV
Avant-Propos
Le présent mémoire est le fruit d’une collaboration étroite entre des neurologues,
généticiens et épidémiologistes. L’auteur de ce mémoire est également le premier auteur de
l’article inséré dans le mémoire et publié dans la revue Annals of Neurology (2006; 62: 93-
98), dont la version n’a été que peu modifiée par l’insertion d’une figure supplémentaire.
Le Docteur Nicolas Dupré a mené le projet sous la supervision des Docteurs Jean-Pierre
Bouchard et René Verreault, avec un financement accordé au Docteur Guy A. Rouleau.
Les analyses génétiques ont été réalisées par le Docteur François Gros-Louis en majeure
partie, avec la contribution de Danielle de Verteuil. Le projet a nécessité l’implication de
cliniciens neurologues ayant contribué à l’évaluation des patients, soit les Docteurs Steve
Verreault, Denis Brunet, et Bernard Brais.

V
Table des matières
Résumé……………………………………………………………………………………...II Summary……………………………………………………………………………..…….III Avant propos………………………………………………………...……………………..IV Liste des figures et tableaux…………………...…………………………………..……….VI Introduction………………………………………………………………...………………..7 Article………………………………………………………………………………………20 Conclusion………………………………………………………………………………….35 Bibliographie……………………………………………………………………………….37

VI
Liste des figures et tableaux
Figure 1 Âge de début par décade……………………………………………………26 Figure 2 Imagerie par résonance magnétique cérébrale d’un patient
atteint de l’Ataxie Récessive de la Beauce…………………………………27 Figure 3 Mutations identifiées dans le gène SYNE1………………………………...30 Figure 4 Répartition des génotypes dans la population étudiée……………………...31 Tableau 1 Évaluation clinique des patients atteints de l’ARB……………………...…25 Tableau 2 Mutations connues causant l’ARB…………………………………………29

7
Introduction
Les dernières décennies ont été marquées par l'arrivée de meilleurs outils cliniques et
moléculaires qui ont révolutionné la classification de nombreuses maladies. La population
de l’est du Canada, compte tenu de son héritage génétique particulier, a contribué à
certaines de ces avancées scientifiques en permettant l’identification de maladies
considérées comme rares dans d'autres régions du monde. Sa plus grande homogénéité
génétique a permis la description fine de présentations cliniques ainsi que de mettre en
lumière des variations phénotypiques. Dans l’introduction, nous présentons une synthèse
des diverses ataxies, neuropathies et paraparésies spastiques héréditaires qui ont été décrites
dans l’est du Canada (1).
Les ataxies héréditaires
Ataxie avec apraxie oculomotrice de type 2 (AAO-2, AOA-2, MIM 606002)
L’ AAO-2 est une maladie autosomique récessive avec une pénétrance dépendante de l’âge,
et débutant entre 10 et 25 ans. Elle est caractérisée principalement par une atrophie
cérébelleuse, une neuropathie axonale sensitivomotrice, une apraxie oculomotrice, et une
alpha-foetoprotéine sérique élevée (2). Le symptôme initial est une démarche ataxique
chez la majorité des patients, qui peut être précédée chez quelques patients par un
strabisme, une dystonie ou un tremblement d’attitude. La maladie progresse lentement :
une minorité de patients se retrouvent en fauteuil roulant après 10 ans d’évolution, alors
que d'autres restent ambulatoires plus de 20 ans après le début des symptômes. La
démarche ataxique est présente chez tous les patients au cours de la maladie, alors que
d'autres caractéristiques peuvent également se développer comme des postures dystoniques
des mains, des mouvements choréiques, et d’un tremblement d’attitude, des membres ou de
la tête (3). Quelques patients peuvent avoir un signe de Babinski ou des perturbations
sphinctériennes. Des signes cliniques de neuropathie (abolition ou diminution des réflexes
tendineux, déficits sensitif ou moteur) sont retrouvés chez la majorité des patients. Les
fonctions cognitives peuvent être modérément altérées. L'apraxie oculomotrice,

8
caractérisée par une dissociation des mouvements conjugués de la tête et des yeux, est
présente chez la plupart des patients. En outre, une poursuite saccadée, un nystagmus dans
le regard fixe, et une abduction limitée sont habituellement présents (3). Les résultats
d’imagerie par résonance magnétique (IRM) montrent invariablement une atrophie
cérébelleuse. Les études de conduction nerveuse périphérique montrent des amplitudes
diminuées compatibles avec une neuropathie axonale sensitivomotrice (3).
Chez les Canadiens français, la description initiale de l’AAO-2 repose sur une famille de 20
enfants dont 7 étaient atteints, la mère provenant de la région de Québec et le père étant
d’origine acadienne (4). Plus tard, cette famille a été étudiée avec 10 autres familles
d'ascendance canadienne-française, pour un total de 17 patients atteints (5). Tous les
patients présentaient un phénotype assez homogène, avec une ataxie commençant entre 8 et
16 ans, accompagnée d'une polyneuropathie sensitivomotrice sévère. L'apraxie
oculomotrice n’était jamais présente, excepté chez quelques patients plus âgés. L'alpha-
foetoprotéine était invariablement élevée, et l’IRM cérébrale montrait toujours une atrophie
ponto-cérébélleuse significative. Chez 13 des 18 porteurs de ces familles canadiennes-
françaises, un haplotype commun a été trouvé en utilisant des marqueurs sur le
chromosome 9q34. L’AAO-2 a également été décrite dans d'autres régions du globe,
comme l'Algérie, le Vietnam, la France, le Portugal, l’Espagne, le Cap Vert, la Turquie, le
Japon, le Pakistan, la Tunisie et le Royaume-Uni (6), (2). Des mutations dans le gène qui
code pour la protéine senataxine causent l’ AAO-2 (2). La fonction de la senataxine est
peu connue, mais on pense que celle-ci aurait une activité ARN et ADN hélicase, et
pourrait ainsi agir dans la voie de réparation de l'ADN. Des mutations dans le gène causant
l’AAO-2 sont également impliquées dans la sclérose latérale amyotrophique de type 4
(MIM 602433) (7), mais cette variante n’a jamais été décrite chez les Canadiens français.
Ataxie de Friedreich (AF ; MIM 229300)
L'ataxie progressive est le symptôme principal de cette maladie, qui commence
habituellement autour de la puberté par un trouble de la marche. Selon Harding (8), les
caractéristiques essentielles sont (1) une transmission autosomique récessive, (2) un début
avant 25 ans, (3) une ataxie progressive des membres et de la démarche, (4) une aréflexie

9
des membres inférieures, (5) des signes électrophysiologiques de neuropathie axonale
sensitive suivie (dans un délai de 5 ans après le début), (6) une dysarthrie, (7) une aréflexie
aux quatre membres, (8) une perte de proprioception et du sens de la vibration, (9) une
réponse cutanée plantaire en extension, et (10) une faiblesse de type pyramidale aux
membres inférieurs. Les autres caractéristiques cliniques incluent une cardiomyopathie, une
scoliose, un diabète, et une surdité. L’IRM révèle une atrophie de la moelle épinière, avec
un tronc cérébral et un cervelet intacts (9).
L’AF est l'ataxie héréditaire la plus fréquente internationalement, avec une prévalence
estimée chez les Caucasiens de 1/29000-1/50000 (9). Bouchard et Barbeau ont décrit un
groupe de 8 familles étroitement liées, avec la forme typique d’AF, dans la région de
Rimouski (Québec) (10). Plus tard, ils ont étudié la généalogie complète de 14 familles
dispersées à travers le Québec, y compris les 8 de la région de Rimouski (11). Le couple
fondateur, qui a vraisemblablement introduit la copie anormale du gène de l’AF au Québec
est arrivé en Nouvelle-France vers 1634 du Perche (France) (11). L’homme possédait une
concession de terre à Beauport, près de la ville de Québec, et n'était ainsi pas d’ascendance
acadienne. L’AF a été également étudiée chez les Acadiens des provinces maritimes et les «
Cajuns » de la Louisiane (12). On avait pensé à l’origine que ces cas pouvaient représenter
une entité différente de l’AF, puisqu'ils différaient des cas classiques principalement quant
à la vitesse de progression de la maladie et à l'incidence limitée de certains des signes
cardinaux. Un total de 14 patients avec l’AF acadiens du Nouveau Brunswick et 28
patients « Cajuns » de Louisiane d’origine acadienne ont été examinés par Barbeau (12).
L’AF chez les patients d’origine acadienne ou « Cajun » était assez uniforme cliniquement,
et présentait une espérance de vie plus élevée et une cardiomyopathie moins fréquente que
chez les patients originaires du Québec. Les études d’haplotypes ont confirmé que les
variantes acadienne et « Cajun » étaient bien liées au locus de l’AF sur le chromosome 9
(13),(14). Les patients acadiens et « Cajuns » partagent le même haplotype principal, bien
que cet haplotype soit rare tant au sein de la population caucasienne atteinte de l’AF que
chez les patients du Québec qui ne sont pas d’ascendance acadienne (15),(16).

10
Le locus de l’AF a été situé (17) au début sur le chromosome 9q13-q21.1, et le screening de
patients pour une mutation dans le gène X25 a mené à l'identification d'une répétition trop
élevée de trinucléotides (GAA) au sein de son premier intron (18). Le gène X25 comprend
sept exons et code pour une protéine appelée frataxine. Cette protéine est exprimée à un
niveau élevé dans la moelle épinière, et à un niveau plus bas dans le cervelet et le cortex
cérébral (18). Dans de grandes études de patients caucasiens atteints de l’AF, il y a une
forte corrélation entre la taille de l'allèle (GAA), l'âge du début ainsi que la progression de
la maladie (19). Dans l’AF acadienne, cependant, il semble y avoir peu de corrélation entre
la taille de l’expansion et la sévérité clinique (20). La pathophysiologie de l’AF a été
intensivement passée en revue ailleurs (9), (21).
Ataxie récessive spastique avec leucodystrophie (ARSAL, Ataxie récessive de
Portneuf).
L’ARSAL se caractérise principalement par la présence d’une ataxie spastique progressive
dont l’âge de diagnostic et le phénotype sont extrêmement variables. L’âge de début moyen
est de 15 années (intervalle de 2-59 ans) (22). La majorité des cas présente des urgences
mictionnelles (57%), une dysarthrie (74%), une dystonie (57%), un nystagmus horizontal
léger (44%), une scoliose (35%), et un retard mental léger (44%). Par ailleurs, on ne
retrouve pas de polyneuropathie. L’imagerie par résonance magnétique révèle une atrophie
cérébelleuse diffuse chez tous les sujets atteints (100%), une atrophie cérébrale légère à
modérée dans 42.9% des cas et la présence de leucodystrophie chez plus de 52% des
patients. La localisation de ces changements est majoritairement périventriculaire ou juxta-
corticale. Aucune corrélation n’a pu être établie entre la présence de leucodystrophie et
l’âge du diagnostic ou le déclin cognitif (22).
Un total de 23 patients qui présentaient des signes cliniques de l’ARSAL provenant de 17
familles canadiennes-françaises ont été recrutés. Plus de 59% avaient une généalogie
connue pour la région de Portneuf (Québec). Une étude génomique complète a été réalisée
avec 500 marqueurs microsatellites à des intervalles approximatifs de 7cM (22). L’analyse
en multipoint LOD a permis de cibler le locus de l’ARSAL avec un LOD de 2.08 pour le
marqueur microsatellite D2S2321. Le génotypage fin de la cohorte a permis de réduire la

11
région candidate de l’ARSAL à environ 0.9 cM (1.25Mb) sur le chromosome 2q33-34 entre
les marqueurs D2S2321 etD2S2274 (LOD score de 5.95). L’analyse des haplotypes chez
tous les patients ARSAL suggère qu’une mutation fondatrice au sein de la population
étudiée soit partagée par plus de 97% des chromosomes porteurs. L’intervalle candidat de
l’ARSAL selon la base de donnée UCSC (version de mars 2006) contient plus de 34 gènes
prédits (22). À ce jour, plus de la moitié des gènes candidats ont été séquencés et aucune
mutation n’a été découverte. Des analyses sont actuellement effectuées afin d’identifier les
mutations causales dans les autres gènes candidats intéressant cet intervalle.
Ataxie sensitive autosomique dominante (ASAD, ADSA)
L'âge typique du début des symptômes survient en général dans la troisième ou quatrième
décennie (entre 28 et 55 ans), et la maladie ne semble pas affecter l’espérance de vie (23).
Il n'y a aucun signe d'anticipation génétique. Le mode de transmission est autosomique
dominant avec une pénétrance dépendante de l’âge. La plupart des patients éprouvent au
commencement des difficultés à la marche, et certains rapportent également une l'instabilité
qui tend à augmenter à l'obscurité. Les réflexes ostéotendineux sont habituellement
diminués dans les membres supérieurs et sont abolis dans les membres inférieurs. Quelques
sujets atteints ont des réponses plantaires en extension et une faiblesse au niveau des
membres inférieurs. La sensibilité dans toutes ses modalités (tact, douleur, température,
proprioception, et vibration) est invariablement diminuée. Il n'y a jamais d’ulcérations
significatives aux pieds. Les dysfonctionnements du système nerveux autonome, tels que
des troubles sphinctériens ou une hypotension orthostatique, ne sont pas présents. Il n'y a
aucun signe d’atteinte cérébelleuse. L’imagerie par résonance magnétique cérébrale et
spinale est normale. Les études de conduction nerveuse périphérique demeurent dans les
limites de la normale, et ce même à un stade avancé de la maladie. Les potentiels
somatosensitifs, cependant, sont ralentis ou absents dans les membres.
La première famille décrite est d'origine Anglo-Saxonne et comprend 10 membres vivants
atteints (23). Une deuxième famille comprenant 9 membres atteints a été publiée
récemment (24). Ces deux familles sont originaires de Moncton (Nouveau Brunswick,
Canada) et portent le même haplotype, ce qui confirme leur ascendance commune. Le gène

12
responsable de cette condition n'a pas été encore caractérisé, mais le locus SNAX1 a été
défini sur un intervalle chromosomique de 9.1 cM sur le chromosome 8p12-8q12.1. Un
certain nombre de gènes candidats intéressant cet intervalle sont actuellement examinés.
Cette maladie neurodégénérative est caractérisée par une ataxie proprioceptive avec
également une atteinte des voies pyramidales. Aucune autopsie n'a encore été effectuée.
L’ASAD partage certaines caractéristiques cliniques avec les neuropathies héréditaires.
Cependant, à la différence de ces dernières, les études de conduction nerveuse sont
normales dans l’ASAD. Cette caractéristique oriente plutôt vers une atteinte prédominante
des fibres nerveuse sensitives pré-ganglionnaires (cordons postérieurs).
Ataxie spastique héréditaire autosomique dominante de type 1 (ASHAD-1 ; SAX 1 ;
MIM 108600)
L’âge typique du début est dans les deux premières décennies, et la maladie ne semble pas
affecter l’espérance de vie de manière significative. Le mode de transmission est
autosomique dominant avec une pénétrance dépendante de l’âge. La plupart des patients se
présentent au début avec une spasticité des membres inférieurs, une maladresse des mains,
et des secousses involontaires de la tête (25). Dans la trentaine, la spasticité progresse aux
membres inférieurs, et de nouveaux symptômes apparaissent tels qu’une dysarthrie, une
dysphagie, des anomalies oculomotrices (saccades lentes, regard vertical altéré, rétraction
palpébrale), et une dystonie. Aucun des patients ne montre d’amyotrophie. L’imagerie
cérébrale ou les données électrophysiologiques sont normales.
Trois familles ont été rapportées à ce jour, toutes dans une région rurale isolée de la
province de Terre-Neuve. Dans cette région, l’ASHAD-1 contribue de façon
disproportionnée au fardeau des maladies neurodégénératives de l'adulte. Au sein de ces
trois familles anglo-saxonnes, 38 individus atteints ont été identifiés et évalués.
L’ASHAD-1 a été décrite également en Iran (26) et aux États-Unis (la Virginie) (27), sans
confirmation génétique toutefois. Le gène causant l’ASHAD-1 a été localisé au
chromosome 12p13 (25). La recherche des haplotypes au moyen de 13 marqueurs
polymorphiques a indiqué que chacune des trois familles de Terre-Neuve partage un
haplotype commun avec des régions recombinantes réduisant l’ensemble à un intervalle de

13
5 cM. Les données neuropathologiques disponibles montrent de la dégénérescence des
régions cortico-spinales et des cordons postérieurs (25).
Ataxie récessive spastique du Charlevoix-Saguenay (ARSCS ; ARSACS ; MIM
270550)
Tôt lorsqu’ils commencent à marcher (12-18 mois), les patients atteints de l’ ARSCS
montrent des signes de spasticité des membres inférieurs (28). Les parents mentionnent
que leurs enfants présentent un trouble de l’équilibre et une tendance à faire des chutes
(28). Le parole peut être légèrement dysarthrique dans l'enfance, et devient explosive vers
l'âge adulte. Les signes neurologiques précoces incluent un signe de Babinski, une
perturbation de la poursuite oculaire, et la présence de fibres myélinisées rayonnant à partir
du disque optique au fond d’œil. Bien qu'ils aient un développement moteur perturbé, les
enfants atteints de l’ ARSCS ne présentent aucun déficit intellectuel (29). La maladie
s’aggrave à l'adolescence et vers 20 ans, avec une augmentation progressive du tonus
musculaire et des réflexes ostéotendineux. Une amyotrophie distale est alors
habituellement observée (28). Le confinement au fauteuil apparaît à un âge moyen de
41ans (17-57 ans). L’âge moyen de décès est de 51 ans (30). Les études de conduction
nerveuse périphériques démontrent des signes de démyélinisation précoce et de neuropathie
axonale progressive, ces derniers confirmés par la biopsie de nerf (31). L'atrophie du
vermis cérébelleux supérieur est toujours présente à l’imagerie par résonance magnétique
(IRM), même chez les plus jeunes patients. La moelle épinière est atrophiée de manière
diffuse.
Avec plus de 300 individus atteints, l’ ARSCS est la plus fréquents de toutes les ataxies
héréditaires du Québec. La plupart des familles proviennent des régions de Charlevoix et
du Saguenay-Lac-Saint-Jean. Pour la période 1941-1985, la fréquence des porteurs fut
estimée à 1/22 dans la région de Saguenay-Lac-Saint-Jean (32). L’ ARSCS a été également
décrite dans d'autres pays, tels le Japon (33), la Turquie (34), la Tunisie (35), l’Espagne
(36), (37), et l’Italie (38). Le gène de l’ARSACS a été localisé à un intervalle de 0.42 cM
sur le chromosome 13q12 (39) avec un haplotype prépondérant chez la plupart des cas
canadien-français, confirmant de ce fait l'existence d'un effet fondateur. Des mutations

14
dans le gène SACS (40), qui codent pour la sacsine, sont responsables de l’ ARSCS. On
connaît peu de choses sur la fonction de SACS, qui code pour une gigantesque protéine de
4579 acides aminés et est exprimé dans une grande variété de tissus incluant le système
nerveux central. La présence d’un domaine DNAJ (heat-shock) dans la sacsine suggère
une fonction chaperonne à cette protéine (40). Le domaine de liaison HEPN suggère un
rôle dans la liaison à l’ATP et à l’ADN. Deux mutations dans le gène ARSCS ont été
découvertes chez les Canadiens-Français : la 6594delT explique environ 92% et la
5254C_T environ 4% des cas. Il est intéressant de noter que depuis la découverte des
deux premières mutations responsables de cette ataxie au SLSJ, plusieurs autres mutations
ont été identifiées à l’étranger.
Les paraparésies spastiques héréditaires et les ataxies spastiques
Paraparésie spastique autosomique dominante de type 4 (SPG-4, FSP2 ; MIM
182601)
L'âge de début de paraparésie spastique autosomique dominante de type 4 est très variable,
s'étendant de l'enfance jusqu’à l’âge adulte avancé, et la maladie ne semble pas affecter
l’espérance de vie (41). Le mode de transmission est autosomique dominant. Les patients
peuvent se présenter soit avec le phénotype non compliqué ou avec le phénotype
compliqué. Dans le phénotype non compliqué, les symptômes seront limités à une
paraparésie spastique progressive avec des degrés variables de sévérité, quelques patients
pouvant être en mesure de marcher de manière autonome jusqu'à un stade avancé dans la
maladie, tandis que d’autres se retrouveront tôt en fauteuil roulant (41). Dans la forme
compliquée, la paraparésie spastique peut être associée à une vessie neurogène, à de
l'incontinence fécale, à des anomalies cognitives, à des crises d’épilepsie, ou encore à une
neuropathie périphérique (41). La résonance magnétique cérébrale et spinale est
invariablement normale. Les conductions nerveuses périphériques peuvent être anormales
dans la forme compliquée.
Un total de 8 familles canadienne-françaises avec cette maladie ont été rapportées au
Québec (41). La maladie a été décrite également dans d'autres régions du monde, dont en

15
France (42), en Hollande (42), en Angleterre (origine Galloise) (43), en Amérique du Nord
(caucasiens) (44), en Tunisie (44), et au Danemark (45). La paraparésie spastique
autosomique dominante de type 4 a été localisée au chromosome 2p22-21. Toutes les
familles canadiennes-françaises décrites présentent des mutations différentes du gène SPG-
4, sauf pour deux d'entre elles, qui partagent une mutation apparemment plus commune
chez les Caucasiens (G1801A) (41). La paraparésie spastique autosomique dominante de
type 4, comme les autres formes de paraparésies spastiques, est liée à l’atteinte des voies
cortico-spinales. On sait désormais que la protéine spastine interagit avec les microtubules
par leurs régions N-terminale, le tout médié par une activité ATPase. Les mutations de la
spastine semblent ainsi altérer la régulation des microtubules au sein des longs axones (46).
Leucodystrophie autosomique dominante (ADLD, PMD-AD-Late-onset type, MS-like
disorder ; MIM 169500)
L'âge typique du début de la maladie est dans la troisième ou la quatrième décennie, et la
maladie affecte l’espérance de vie de façon significative. Le mode de transmission est
autosomique dominant avec une pénétrance dépendante de l’âge. Onze patients canadien-
français avec la leucodystrophie autosomique dominante ont été examinés et des études
généalogiques ont été réalisées (47). Le phénotype des familles canadienne-françaises s'est
avéré semblable à celui des autres familles précédemment décrites : les 6 individus
examinés ont montré une instabilité progressive à la marche, avec un syndrome du
motoneurone supérieur (spasticité; hyperréflexie; réponses plantaires en extension), et des
anomalies du système nerveux autonome (dysfonctionnement intestinal/vésical;
hypotension orthostatique). L’imagerie par résonance magnétique cérébrale montre une
légère atrophie corticale avec des changements symétriques de la substance blanche, les
plus prononcés étant dans les régions fronto-pariétales, en association avec une atrophie
diffuse de la moelle épinière.
Quatre familles dans le monde ont été rapportées avec leucodystrophie autosomique
dominante, une d’origine canadienne-française (Charlevoix, Québec) (47), (48) ; une
d’origine écossaise/irlandaise (49); une d’origine américaine/irlandaise (50); une d’origine
suédoise (51). Des études de localisation ont montré que le locus de la leucodystrophie

16
autosomique dominante se situait sur le chromosome 5q31, avec un LOD score de 5.72 sur
le marqueur D5S804 (52). Par la suite, le gène Lamin B1 a été identifié comme étant
responsable de leucodystrophie autosomique dominante (53). En effet, les sujets atteints de
leucodystrophie autosomique dominante présentent une duplication génomique qui a pour
conséquence de placer une copie supplémentaire du gène Lamin B1, et ainsi créer une
surexpression de ce gène. Ceci aurait pour conséquence de nuire à l’intégrité de la
membrane nucléaire cellulaire, tout en affectant de façon prédominante les astrocytes et en
épargnant les oligodendrocytes (53).
Les Neuropathies héréditaires
Neuropathie sensitivomotrice héréditaire avec agénésie du corps calleux
(NSMH/ACC, HMSN/ACC, ACCPN, syndrome d'Andermann ; MIM 218000)
La NSMH/ACC est une maladie autosomique récessive d’apparition précoce. Elle est
caractérisée par un retard du développement psychomoteur, une neuropathie
sensitivomotrice sévère avec aréflexie, un degré variable d'agénésie du corps calleux, une
amyotrophie, et une hypotonie (54). D'autres caractéristiques associées incluent une ptose
palpébrale, une paralysie du regard vertical, une asymétrie faciale, un tremblement, une
rétraction des tendons d’Achille, et une scoliose (54). La maladie se manifeste au départ
par une hypotonie et un retard dans le développement psychomoteur. L'âge moyen du
début de la marche assistée est de 3.8 ans. La maladie progresse implacablement, et tous
les patients deviennent par la suite confinés au fauteuil roulant dès l'adolescence.
L’espérance de vie est raccourcie (33 ans, en moyenne), bien qu'un certain nombre de
patients aient survécu jusqu’à la cinquantaine. Une majorité de patients présente un retard
intellectuel léger à modéré, et près de 40% des patients développent des caractéristiques «
psychotiques » en fin d’adolescence (idées paranoïdes, hallucinations visuelles et
auditives). Les conductions nerveuses sensitives sont invariablement absentes dès la
naissance (54). La résonance magnétique cérébrale montre une dysgénésie partielle ou
totale de corps calleux (54).

17
La NSMH/ACC est retrouvée principalement dans la population canadienne française du
Québec (région du Saguenay-Lac-St-Jean et de Charlevoix). Le taux de porteur est estimé
à 1/23 habitants dans le Saguenay-Lac-St-Jean, et les reconstructions généalogiques ont
prouvé que tous les porteurs obligatoires ont des ancêtres communs, soutenant de ce fait un
effet fondateur (55). La NSMH/ACC a également a été rapportée dans d'autres régions du
globe comme l’Italie (56), (57), l’Autriche (58), la Tanzanie (59), la Turquie (57), la
Hollande, et l'Afrique du Sud (communication personnelle). La preuve d’un effet
fondateur dans la population canadienne-française a été confirmée par l'analyse des
haplotypes, qui a prouvé que le marqueur D15S1232 était conservé et se retrouvait dans
plus de 97% des chromosomes atteints (57). Des mutations dans le gène SLC12A6, qui
code pour le co-transporteur chlore-potassium (KCC3), se sont avérées responsables de la
NSMH/ACC. Le KCC3 est exprimé à la fois dans le système nerveux central et
périphérique (57). Le KCC3 serait impliqué dans la régulation de la volémie cellulaire ou
dans la régulation des concentrations intracellulaire de K+ et de Cl- (60). Des autopsies de
patients avec NSMH/ACC ont été effectuées, montrant des gonflements axonaux dans la
substance blanche et un oedème axonal sévère dans l’ensemble du système nerveux
périphérique (54).
Neuropathie sensitive héréditaire de type 2 (NSH-2, HSAN-2, HSN-2, Neurogenic
Acroosteolysis, MIM 201300)
La NSH-2 est une maladie autosomique récessive débutant typiquement dans les deux
premières décennies. La maladie peut avoir une variété de manifestations phénotypiques,
mais la caractéristique principale consiste en une perte de sensibilité dans les extrémités.
Les cas décrits au sein des familles canadienne-françaises (61), (62) ont les caractéristiques
suivantes : les enfants sont habituellement normaux à la naissance, avec un développement
moteur et des capacités d’apprentissage normales durant l’enfance; pendant la première
décennie, ils développent habituellement des troubles sensitifs dans les extrémités
(difficultés à identifier des objets avec leurs mains, blessures cutanées fréquentes,
ulcérations) ; par la suite, les patients développent une acroostéolyse, évidente à l'examen
clinique et aux rayons X des membres. Les cas décrits en Nouvelle-Écosse et à Terre-
Neuve (63) ont les caractéristiques suivantes : les patients se présentent tôt dans l'enfance

18
avec un engourdissement des mains et des pieds, aggravé par le froid, ainsi qu’une
diminution de la sensibilité au toucher, à la douleur, et à la température. La perte de
sensibilité dans les membres est principalement distale, mais peut également impliquer le
tronc chez quelques patients. Dans certains cas, d'autres éléments sont présents, tels une
atrophie musculaire, des réflexes ostéotendineux diminués, des ulcérations, des infections,
et des amputations des membres inférieurs. Le dysfonctionnement du système nerveux
autonome (l'hypotension orthostatique et une diminution de la transpiration) n'est pas
retrouvé chez ces patients. Dans la NSH-2, les conductions nerveuses sensitives sont
toujours anormales (absence de potentiel ou amplitudes sévèrement diminuées) (62).
Un total de 13 familles d'origine canadienne-française de diverses régions du Québec ont
été décrites avec la NSH-2 (61), (62), (64). Des 16 cas examinés pour des mutations dans
le gène causant la NSH-2, 9 étaient homozygotes pour la mutation de c.943C→T, 1 était
homozygote pour la mutation c.918-919insA, et 6 étaient des hétérozygotes (64). Par
ailleurs, la NSH-2 a été la première fois signalée à Terre-Neuve au début des années 1900.
Les membres de la famille principale sont venus du Dorset (Royaume-Uni) en tant que
migrants issus du déplacement massif des colons protestants du sud-ouest de l’Angleterre et
des colons catholiques d'Irlande méridionale (65). Une grande famille consanguine
multigénérationelle a été décrite, ainsi qu’une plus petite famille. Tous les cas affectés
étaient porteurs de la mutation homozygote c.594delA dans le gène causant la NSH-2. Une
autre petite famille de Nouvelle-Écosse, d’origine ethnique incertaine, a été décrite avec
une mutation homozygote c.918-919insA (63). Selon la biopsie et les données
électrophysiologiques obtenues, le dysfonctionnement du gène causant la NSH-2 entraîne
des anomalies autant pour les fibres nerveuses sensitives que motrices, bien que les
changements soient plus sévères dans les fibres sensitives. Les biopsies de nerfs sensitifs
montrent une perte sévère ou complète des fibres myélinisées (62). On ne sait pas à ce
stade dans quelle mesure le système nerveux autonome est impliqué. Le gène causant la
NSH-2 est particulier parce qu'il se compose d'un seul exon qui se retrouve dans la région
intronique d'un autre gène, le PRPKWNK1. Sa fonction est inconnue, mais il pourrait
entraîner l’expression d’une protéine jouant un rôle de facteur neurotrophique (63).

19
L’Ataxie cérébelleuse récessive de la Beauce
Au cours des dernières années, une nouvelle forme d’ataxie héréditaire a été identifiée dans
la région de la Beauce (Québec). Afin de déterminer le gène responsable de cette nouvelle
forme d’ataxie nommée ataxie récessive de la Beauce (ARB), une étude génomique
complète a été réalisée (66). Cette étude a localisé le gène causant l’ARB sur le
chromosome 6q. La séquence complète chez tous les patients atteints d’ARB du seul gène
inclu dans cette intervalle (SYNE-1) a permis d’identifier plusieurs mutations au sein la
population étudiée. Le travail d’identification du gène complété, l’objectif du présent
mémoire a été centré sur la caractérisation génotypique et phénotypique de cette maladie
qui n’avait jamais été décrite ailleurs dans le monde.

20
CHAPITRE I: Clinical and Genetic Study of Autosomal
Recessive Cerebellar Ataxia Type 1
Nicolas Dupré MD MSc (1), François Gros-Louis PhD (2), Nicolas Chrestian MD (1),
Steve Verreault MD MSc (1), Denis Brunet MD (1), Danielle de Verteuil (2), Bernard
Brais MD PhD (2), Jean-Pierre Bouchard MD (1) and Guy A. Rouleau MD PhD (2)
(1)Faculty of Medicine, Laval University, Department of Neurological Sciences, CHAUQ -
Enfant-Jésus, 1401, 18th street, Quebec City, QC, Canada, G1J 1Z4
(2) Center for the Study of Brain Diseases, Université de Montréal, CHUM Research
Center - Notre-Dame Hospital, J.A. de Sève Pavilion, 1560, Sherbrooke Street East,
Montreal, QC, Canada, H2L 4M1

21
ABSTRACT
Objective: Define the phenotype and genotype of a cluster of families with a relatively pure
cerebellar ataxia referred to as Autosomal Recessive Cerebellar Ataxia type 1 (ARCA-1).
Methods: We ascertained 64 probands and affected members of 30 French-Canadian
families all showing similar clinical features and originating from the same region of
Quebec. After informed consent, we performed detailed clinical history, neurological
examination, brain imaging, nerve conduction studies, and SYNE1 mutation detection of all
available subjects. Results: Based on the cases examined, ARCA-1 is a cerebellar
syndrome characterized by: recessive transmission; middle-age onset (mean 31.60 years,
range 17-46); slow progression and moderate disability; significant dysarthria; mild
oculomotor abnormalities; occasional brisk reflexes in the lower extremities; normal nerve
conduction studies and diffuse cerebellar atrophy on imaging. We identified a total of 7
mutations in our population, thereby providing evidence of genotypic heterogeneity.
Patients with different mutations did not show significant phenotypic heterogeneity.
Interpretation: We identified a cluster of French-Canadian families with a new recessive
ataxia of relatively pure cerebellar type caused by mutations in SYNE1. The function of
SYNE-1 is thus critical in the maintenance of cerebellar structure in humans. We expect
that this disease will be a common cause of middle-age onset recessive ataxia worldwide.

22
Introduction
The hereditary ataxias can be divided based on their mode of inheritance into autosomal
dominant, autosomal recessive, X-linked and the mitochondrial ataxias. These disease
categories share the prototypic feature of impaired walking, although they usually present a
variety of other neurological symptoms such as pyramidal signs, peripheral neuropathy,
extrapyramidal signs, cognitive loss, or retinopathy. The autosomal recessive ataxias are
also a heterogeneous group of disorders comprised mainly of Friedreich’s Ataxia, Ataxia
Telangiectasia, Ataxia with Vitamin E Deficiency, Autosomal Recessive Spastic Ataxia of
Charlevoix-Saguenay (ARSACS), Abetalipoproteinemia, Ataxia with Oculomotor Apraxia
type 1 (AOA-1) and type 2 (AOA-2) (67).
Over the last decade, we have identified a large cluster of French-Canadian families whose
ancestors originate mostly from the same region of the Province of Quebec (Canada). This
region, located in South-Eastern Quebec near the US border, is called Beauce. The affected
individuals in these families all share similar clinical characteristics that define this new
disease entity we named ARCA-1, also known as Recessive Ataxia of Beauce (RAB) (68),
(69). Genome-wide linkage and fine-mapping analysis on selected families with ARCA-1
established a minimum candidate interval of about 0.5-Mb on chromosome 6q containing a
single causal gene (SYNE1) (66). Spanning over 0.5-Mb of genomic DNA, SYNE1 is one
of the biggest gene in the human genome formed of 147 exons that encodes a 27,652-kb
mRNA and an 8797 amino acid long protein. To date, 5 different truncating mutations
within Syne1 have been described (66). In the present paper, we report 2 novel mutations,
also leading to premature termination of the protein, and define the full clinical and
molecular spectrum of ARCA-1.
Subjects and methods
Subjects were referred to the study protocol by their treating physician based on a
preliminary assessment consistent with the core features of ARCA-1 (ataxia and dysarthria
of middle-age onset with cerebellar atrophy) and a recessive family history (parents

23
unaffected). They were informed of the procedures entailed in the protocol and signed,
prior to their participation, a consent form approved by the local ethics review boards
(Centre Hospitalier de l’Université de Montréal and Centre Hospitalier Affilié Universitaire
de Québec). All 64 available affected and unaffected members of the 30 recruited families
underwent a thorough neurological examination, and were examined independently by at
least two neurologists. We also used an assessment scale (70) to grade severity of
symptoms: dysarthria (0 = no impairment; 1 = mild dysarthria but comprehensible; 2 =
moderate dysarthria with interruption in flow; 3 = severe dysarthria and incomprehensible,
very difficult to understand; 4 = completely unintelligible); dysmetria (0 = no impairment; 1
= mild dysmetria but reaches the target; 2 = moderate dysmetria, reaches target after several
attempts; 3 = severe dysmetria, short of target after many attempts; 4 = cannot use hands);
gait (0 = normal; 1 = stance width increased, mildly unstable gait but can walk without
support; 2 = moderately unstable gait and needs support for walking; 3 = unable to walk,
needs the assistance of two persons; 4 = wheelchair bound). Twenty-two subjects
underwent electrophysiological studies (D.B) including compound motor action potentials
of median, ulnar, tibial and peroneal nerves, as well as sensory nerve action potentials of
median, ulnar, radial and sural nerves, with standard values for filters, stimulus duration
and electrode positioning. Brain imaging with magnetic resonance imaging was performed
on 50 affected subjects.
Upon receipt of informed consent, blood samples were obtained from affected individuals
in 30 families. DNA was extracted from peripheral blood by standard methods (71). For
mutation screening, a set of 154 PCR primer pairs were designed from genomic DNA to
amplify each exon of the SYNE1 gene, including the flanking splice sites and the
untranslated regions (66). Products were PCR-amplified, checked on agarose gels, and
then sequenced using the forward primers for all of the amplicons. Each fragments
containing mutations were PCR-amplified a second time and sequenced with the reverse
primer in order to confirm that the identified mutations are not due to PCR artefact.

24
Results
Sample case: This 42 year-old woman originates from the Beauce region in Quebec. On
her initial visit, she mentioned that around the age of 30 she started noticing that her speech
was slurred. During the same period, she also noticed some mild walking impairment. As
she advanced in her thirties, the slurred speech and walking impairment progressed.
Strangers would sometimes wonder if she had been drinking alcohol, and her gait was
becoming more wide-based with a tendency to fall if she did not pay more attention than
usual during her movements. Additionally, her hands would feel clumsier. On
neurological examination at age 42, she had normal fundoscopy, normal ocular saccades
and pursuit, and no nystagmus. She had significant cerebellar dysarthria. Strength was
normal throughout, with no spasticity. Reflexes were normal in the upper and lower limbs,
with down-going toes. Sensory examination was normal to pain, temperature, vibration,
proprioception, and light touch. Gait was broad-based. There was significant dysmetria on
finger-to-nose and heel-to-shin, as well as cerebellar hypotonia (Holmes’ sign) in the upper
limbs. Magnetic resonance imaging revealed diffuse cerebellar atrophy and nerve
conduction studies were normal. Genetic testing confirmed that she was homozygous for
the g.306434A>G mutation.
Overall, the ARCA1 phenotype consists of a middle-age onset disease that presents with
either dysarthria, cerebellar ataxia or both coincidentally (table 1 & figure 1). Over time,
all patients develop significant dysarthria and ataxia, with other associated features such as
dysmetria, brisk lower extremity tendon reflexes, and minor abnormalities in saccade and
smooth pursuit. None of the subjects evaluated showed extrapyramidal signs, cognitive
loss, retinopathy, cardiomyopathy, sensory abnormalities, or autonomic disturbances. The
disease progresses slowly and evolves into a moderate degree of disability. There appears
to be no effect on life expectancy. Nerve conduction studies performed on 22 affected
individuals were always normal, showing therefore no sign of a peripheral sensory or motor
neuropathy in this disease. Single-fiber electromyography was also normal when
performed on one subject, suggesting preserved function at the neuromuscular junction.

25
Table 1: Clinical results of patients with ARCA-1 (n=64). ____________________________________________________________________ Sex Male 37 (58%) Female 27 (42%) First complaint Dysarthria 8 (12.5%) Ataxia 40 (62.5%) Both 16 (25%) Age of onset (dysarthria) Mean 34.79 SD 7.62 range 17-50 Age of onset (ataxia) Mean 31.60 SD 7.81 range 17-45 Age at evaluation Mean 45.36 SD 10.71 range 24-69 Duration at evaluation Mean 14.33 SD 9.77 range 3-40 ____________________________________________________________________ Assessment scale Dysarthria Mean 1.78 range 0-3 Dysdiadochokinesis Mean 1.26 range 0-2 Dysmetria Mean 1.44 range 0-3 Gait Mean 1.48 range 0-3 ____________________________________________________________________ Neurological examination Dysarthria 64 (100%) Nystagmus 6 (9.4%) Slow saccades 20 (31.2%) Abnormal pursuit 28 (43.8%) Brisk reflexes in the lower limbs 21 (32.8%) Ankle clonus and/or Babinski sign 4 (6.2%) Dysmetria on finger-to-nose 58 (90.6%) Dysmetria on heal-to-shin 58 (90.6%) Ataxia 63 (98.4%) _____________________________________________________________________

26
Figure 1: Age of onset by decade.

27
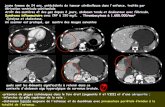
Imaging findings by CT or MRI on 50 affected individuals invariably showed marked
diffuse cerebellar atrophy (figure 2). On detailed review of imaging in 38 affected subjects,
there was no cerebral cortical atrophy; no midbrain, pontine or bulbar atrophy; no atrophy
of inferior olives; no white matter changes.
Figure 2: Magnetic resonance imaging of a 43 years old ARCA-1 patient after 5 years of
disease evolution. Sagittal T1 shows marked diffuse cerebellar atrophy with no cerebral
cortical atrophy; no midbrain, pontine or bulbar atrophy.

28
Genetic analysis revealed 7 mutations (table 2, figure 3 & 4), including 5 previously
described mutations (66) and 2 novel truncating mutations (g.409218C>T and g.281100-
281101delTG). The most frequent mutation, g.306434A>G, was present homozygously in
20/64 subjects (31.2%). This same mutation was present heterozygously with the
g.310067A>G mutation in 11/64 (17.2%), with the g.247012A>T mutation in 6/64 (9.4%),
with the g.426494C>T mutation in 1/64 (1.6%), and with the g.334338-334342delATTTG
mutation in 4/64 (6.2%). There was 1/64 (1.6%) g.310067A>G homozygote, 3/64 (4.7%)
g.247012A>T homozygotes, 1/64 (1.6%) g.247012A>T / g.409218C>T heterozygote.
Finally, 6/64 (9.4%) carried a known mutation on one chromosome and an unknown
mutation on the other chromosome, while in 9/64 (14.0%) the mutation was not found
using the technique described above and in 2/64 (3.1%) genetic testing could not be
performed. The g.306434A>G mutation was present on 50.8% of chromosomes tested,
while mutations remain to be identified in close to one-fifth of carrier chromosomes. We
performed genotype-phenotype correlation studies by segregating based on the most
common genotypes (g.306434A>G homozygotes and g.306434A>G / g.310067A>G
heterozygotes) using the following parameters: age of onset (dysarthria, ataxia, overall);
disease duration; eye movement abnormalities. This analysis gave no statistically
significant differences. Additionally, when we analysed the clinical data of patients bearing
the less common genotypes, we were unable to demonstrate that they showed any atypical
clinical features.

29
Table 2: Known mutations causing ARCA-1. Variants* Exons/Introns Protein changes Total carrier
chromosomes tested in patient population (n=124)
_________________________________________________________________________ g.306434A>G Intron 81 Premature stop 63 (50.8%) at position 5244 g.310067A>G Intron 84 Premature stop at 13 (10.5%) position 5402 g.247012A>T Exon 56 R2906X 16 (12.9%) g.426494C>T Exon 126 Q7640X 2 (1.6%) g.334338-334342delATTTG Exon 93 Premature stop at 4 (3.2%)
position 5880 g.409218C>T Exon 118 Q7386X 1 (0.8%) g.281100-281101delTG Exon 71 4077X 1 (0.8%) Unknown 24 (19.3%) _________________________________________________________________________ * Variants were named according to the genomic DNA sequence NM_033071; nucleotide
"A" from the ATG initiation codon is referred as 1. Allele frequencies of variants were
0/380 French-Canadian controls.

30
Figure 3: SYNE1 identified truncating mutations.
(A) Sequence traces of normal individual (top) and ARCA-1 patients (lower) showing the
novel detected mutations within exons 71 and 118 of SYNE1.
(B) Protein structure of SYNE-1. Known ARCA-1 disease causing mutations are in black
and newly identified mutations lie within boxes. Light grey region defines areas rich in
predicted spectrin repeats; grey boxes at the N-terminal part of the protein correspond to
calponin-homology domains involved in actin binding; dark-grey box indicates the C-
terminal nuclear envelope binding domains containing sequences homologous to the
Drosophila protein Klarsicht.

31
Figure 4 : Segregation of the various genotypes in the population studies.
51% (Intron 81)
10% (Intron 84)
13% (Exon 56)
19% (Unknown)

32
Discussion
ARCA-1 is thus a new recessive relatively pure cerebellar ataxia that is caused by various
mutations in SYNE1. ARCA-1 shows relative homogeneity of the phenotype, despite being
caused by more than seven different mutations. The age of onset does not vary
significantly in function of given mutations like in trinucleotide repeat disorders, whereas
the disease shows little associated features accompanying the core symptoms of dysarthria
and dysmetria. SYNE1 encodes a protein of about 8,797 amino acid residues (>1,000 kDa)
(66). The protein contains two N-terminal actin-binding regions that comprise tandem
paired calponin-homology-domains, a transmembrane domain, multiple spectrin repeats
and a C-terminal KASH domain. Although SYNE1 is expressed in multiple tissues, its
greatest level in the central nervous system of mice is in the cell bodies of the Purkinje cells
and in neurons of the olivary region of the brainstem, while in humans it is also expressed
predominantly in the cerebellum; it is not expressed in glial cells. In the peripheral nervous
system, SYNE-1 is involved in anchoring specialized myonuclei underneath the
neuromuscular junctions (66). It was found in a muscle biopsy of an ARCA-1 patient that
fewer myonuclei come to lie beneath the neuromuscular junction, although this has no
consequences clinically, electrophysiologically, or ultrastructurally. SYNE-1 is part of the
spectrin family of structural proteins that share a common function of linking the plasma
membrane to the actin cytoskeleton. This family also includes dystrophin (Duchenne and
Becker muscular dystrophies)(72), SPTBN2 (Spinocerebellar ataxia type 5 ) (73),
PLEKHG4 (16q-Autosomal dominant cerebellar ataxia) (74) and Spnb4 (75).
The closest description of ARCA-1 phenotype in the literature is Holmes hereditary ataxia.
Holmes’ type of hereditary ataxia (76) was described a century ago (1907) in a family of
eight siblings, with four of them presenting with middle-age onset dysarthria, ataxia and
hypogonadism (not present in ARCA-1). The inheritance pattern was most likely
autosomal recessive, since the parents were not affected. Autopsy of one affected case
showed diffuse cerebellar atrophy with no pontine or olivary involvement. To our
knowledge, no linkage or gene defect responsible for this type of ataxia has been reported
as yet.

33
(ARSACS) is the most common of all autosomal recessive ataxias in Quebec with more
than 300 affected individuals. ARSACS patients exhibit early onset signs of spasticity in
the lower limbs usually observed at gait initiation (12-18 months) (28). The clinical picture
noticed by parents from early childhood is always that of a gait ataxia with a tendency to
fall. Nerve conduction studies demonstrate signs of progressive axonal sensory-motor
neuropathy. AOA-2 is also present in Quebec, where more than 10 families have been
described. It is characterized mainly by cerebellar atrophy, axonal sensory-motor
neuropathy and elevated serum α-fetoprotein (77). The main differences clinically between
these other recessive ataxias common in Quebec and ARCA-1 is the earlier age of onset,
the higher degree of disability, and the associated peripheral neuropathy.
Of interest, 16q-Autosomal Dominant Cerebellar Ataxia is characterized by an age of onset
>55 years and sensorineuroral hearing impairment (74), which is different from what we
found in ARCA-1. However, there are similarities in that it is also a relatively pure
cerebellar syndrome caused by mutations in PLEKHG4, also part of the spectrin family of
structural proteins. On pathology, 16q-Autosomal dominant cerebellar ataxia shows
peculiar degeneration of Purkinje cells that undergo shrinkage and are surrounded by
amorphous material. Spinocerebellar Ataxia type 5 is characterized by a slowly
progressive cerebellar syndrome beginning mostly in the third decade (78), (79). The most
consistent clinical feature is downbeat nystagmus, while other common features included
gait, stance, and limb ataxia; dysarthria; intention tremor and resting tremor; impaired
smooth pursuit; gaze-evoked nystagmus. Symptom progression is slow, and all patients
remain ambulatory despite disease duration of up to 30 years. MRI shows atrophy of the
cerebellar vermis and hemispheres. Again, this other ataxia caused by mutations in a
spectrin family protein shows striking similarities with ARCA-1 in terms of the
predominant cerebellar involvement, middle-age onset, relatively slow progression, and
moderate degree of disability.
In conclusion, ARCA-1, Spinocerebellar Ataxia type 5 and 16q-Autosomal Dominant
Cerebellar Ataxia taken together allow us to define a new category of hereditary ataxias

34
related to the spectrin family of structural proteins. Despite significant genetic
heterogeneity, this category of ataxia shares many common clinical features. We expect
that this new category of inherited ataxias may be more frequent than previously thought,
mainly through the contribution of ARCA-1, since we have encountered important genetic
heterogeneity even within a homogeneous founder population. We speculate that a
significant proportion of yet undiagnosed recessive or “sporadic” ataxias may be due to
SYNE1 mutations, which would have great repercussions on our ability to diagnose more
precisely these ataxia types in specialized clinics worldwide.
ACKNOWLEDGEMENTS
This work was supported by the National Ataxia Foundation (Funding), the Canadian
Genetic Disease Network (Funding), the Canadian Institute of Health Research
(Scholarships of FGL and ND) and the Association des Ataxies Familiales (Scholarship of
DV). France Gosselin and Madeleine Plante performed blood collection of patients and
obtained their consent. Finally, we would also like to thank the family members who
participated in this study.

35
Conclusion
Nous avons récapitulé dans l’introduction les diverses ataxies héréditaires, paraparésies
spastiques héréditaires, et enfin neuropathies héréditaires de l’est du Canada. Le travail
préliminaire a été effectué en grande partie par des cliniciens astucieux comme André
Barbeau et Jean-Pierre Bouchard, qui ont contribué considérablement à la description des
sous-types phénotypiques de ces maladies. En tant que tel, les maladies suivantes ont été
identifiés la première fois chez des patients de l’est du Canada: l’Ataxie récessive spastique
du Charlevoix-Saguenay, la Neuropathie sensitive héréditaire de type 2, l’ARB, et la
Neuropathie sensitivomotrice héréditaire avec agénésie du corpus calleux. Le contexte
historique particulier de cette région du Canada a provoqué les effets fondateurs et
l'isolement génétique qui ont eu comme conséquence le développement de certaines
maladies rares au sein de cette population. Le travail de base résidait dans la
démonstration, par des études généalogiques, d’ancêtres communs aux individus se
présentant avec des phénotypes semblables. Ceci a été suivi, dans l'ère génomique, de
l'identification des haplotypes de fondateur et de la relative homogénéité génétique de ces
individus.
L’ARB se caractérise principalement par une atteinte lentement progressive sous forme de
dysarthrie ou d’ataxie cérébelleuse débutant généralement dans la deuxième ou la troisième
décennie (âge de début moyen 30.4 années ; intervalle de 17-46 ans). Il y a peu d’autres
signes cliniques associés si ce n’est des anomalies mineures des mouvements extra-
oculaires (diminution de la vitesse des saccades), une incoordination des membres ou
encore des réflexes ostéotendineux parfois vifs aux membres inférieurs. Par ailleurs, on ne
retrouve pas de signes extra-pyramidaux, d’atteinte du système nerveux autonome, ou de
polyneuropathie. Sur le plan des examens para-cliniques, l’imagerie par résonance
magnétique révèle une atrophie cérébelleuse diffuse sans atteinte du tronc cérébral chez les
sujets atteints, et ce, même après seulement quelques années d’évolution de la maladie; les
conductions nerveuses périphériques sont toujours normales, donc sans signe d’atteinte
neuropathique.

36
Au cours des dix dernières années, des familles canadiennes-françaises originaires de la
Beauce et du Bas-St-Laurent (Québec) ont été identifiées, dont plusieurs membres
présentaient des signes cliniques de l’ARB. Afin de déterminer le gène responsable de l’
ARB, une étude génomique complète a été réalisée avec 388 marqueurs microsatellites à
des intervalles approximatifs de 10cM (66). Cette étude a localisé le gène causant l’ARB
sur le chromosome 6q entre les marqueurs D6S420 et GATA186B06. La séquence
complète chez tous les patients atteints d’ARB du seul gène inclu dans cette intervalle
(SYNE-1) a permis d’identifier 7 mutations au sein la population étudiée. Chaque mutation
sied sur son propre haplotype, dont 2 sont nettement plus fréquents, ce qui confirme l’effet
fondateur. La protéine encodée par SYNE-1 étant constituée de nombreuses répétitions
interagissant avec les spectrines, on peut émettre l’hypothèse que les mutations retrouvées
chez les patients atteints de l’ARB affectent la capacité de cette dernière à s’attacher à la
spectrine, ce qui pourrait avoir un impact significatif sur la structure nucléaire des cellules
de Purkinje. L’ARB peut ainsi être classifiée dans une nouvelle catégorie d’ataxies
héréditaires - celles reliées à des défauts d’attachement à la spectrine - qui comprend
désormais l’ARB, l’Ataxie spinocérébelleuse de type 5, et l’Ataxie cérébelleuse
autosomique dominant du chromosome 15q, toutes trois caractérisées pas une
dégénérescence cérébelleuse tardive et lentement progressive.

37
Bibliographie
1. Dupre N, Bouchard JP, Brais B, Rouleau GA. Hereditary ataxia, spastic paraparesis
and neuropathy in the French-Canadian population. Can J Neurol Sci 2006;33:149-157.
2. Moreira MC, Klur S, Watanabe M, et al. Senataxin, the ortholog of a yeast RNA
helicase, is mutant in ataxia-ocular apraxia 2. Nat Genet 2004;36:225-227.
3. Le Ber I, Bouslam N, Rivaud-Pechoux S, et al. Frequency and phenotypic spectrum
of ataxia with oculomotor apraxia 2: a clinical and genetic study in 18 patients. Brain
2004;127:759-767.
4. Bouchard J, Bedard P, Bouchard R. Study of a family with progressive ataxia,
tremor and severe distal amyotrophy. Can J Neurol Sci 1980;7:345-349.
5. Duquette A, Bouchard JP, Marchand L, et al. French Canadian Cluster of
Autosomal Recessive Ataxia Associated with a CMT-Like Polyneuropathy. Neurology
2003;60 (Suppl 1):A461.
6. Nemeth AH, Bochukova E, Dunne E, et al. Autosomal recessive cerebellar ataxia
with oculomotor apraxia (ataxia-telangiectasia-like syndrome) is linked to chromosome
9q34. Am J Hum Genet 2000;67:1320-1326.
7. Chen YZ, Bennett CL, Huynh HM, et al. DNA/RNA helicase gene mutations in a
form of juvenile amyotrophic lateral sclerosis (ALS4). Am J Hum Genet 2004;74:1128-
1135.
8. Harding AE. Friedreich's ataxia: a clinical and genetic study of 90 families with an
analysis of early diagnostic criteria and intrafamilial clustering of clinical features. Brain
1981;104:589-620.
9. Alper G, Narayanan V. Friedreich's ataxia. Pediatr Neurol 2003;28:335-341.

38
10. Bouchard JP, Barbeau A, Bouchard R, Paquet M, Bouchard RW. A cluster of
Friedreich's ataxia in Rimouski, Quebec. Can J Neurol Sci 1979;6:205-208.
11. Barbeau A, Sadibelouiz M, Roy M, Lemieux B, Bouchard JP, Geoffroy G. Origin of
Friedreich's disease in Quebec. Can J Neurol Sci 1984;11:506-509.
12. Barbeau A, Roy M, Sadibelouiz M, Wilensky MA. Recessive ataxia in Acadians
and "Cajuns". Can J Neurol Sci 1984;11:526-533.
13. Keats BJ, Ward LJ, Shaw J, Wickremasinghe A, Chamberlain S. "Acadian" and
"classical" forms of Friedreich ataxia are most probably caused by mutations at the same
locus. AM J Med Genet 1989;33:266-268.
14. Sirugo G, Duclos F, Fujita R, et al. Mapping the Friedreich ataxia locus (FRDA) by
linkage disequilibrium analysis with highly polymorphic microsatellites. Biomed
Pharmacother 1994;48:219-224.
15. Richter A, Poirier J, Mercier J, et al. Friedriech ataxia in Acadian families from
Easter Canada: Clinical diversity with conserverd haplotypes. AM J Med Genet
1996;64:594-601.
16. Sirugo G, Keats B, Fujita R, et al. Friedreich ataxia in Louisiana Acadians:
demonstration of a founder effect by analysis of microsatellite-generated extended
haplotypes. Am J Hum Genet 1992;50:559-566.
17. Chamberlain S, Shaw J, Wallis J, et al. Genetic homogeneity at the Friedreich ataxia
locus on chromosome 9. Am J Hum Genet 1989;44:518-521.
18. Campuzano V, Montermini L, Molto M, et al. Friedriech ataxia: autosomal
recessive disease caused by an intronic GAA triplet repeat expansion. Science
1996;271:1423-1427.

39
19. Durr A, Cossee M, Agid Y, et al. Clinical and genetic abnormalities in patients with
Friedreich's ataxia. N Engl J Med 1996;335:1169-1175.
20. Montermini L, Richter A, Morgan K, et al. Phenotypic variability in Friedreich
ataxia: role of the associated GAA triplet repeat expansion. Ann Neurol 1997;41:675-682.
21. Voncken M, Ioannou P, Delatycki MB. Friedreich ataxia-update on pathogenesis
and possible therapies. Neurogenetics 2004;5:1-8.
22. Thiffault I, Rioux MF, Tetreault M, et al. A new autosomal recessive spastic ataxia
associated with frequent white matter changes maps to 2q33-34. Brain 2006;129:2332-
2340.
23. Valdmanis PN, Simoes Lopes AA, Gros-Louis F, Stewart JD, Rouleau GA, Dupre
N. A novel neurodegenerative disease characterised by posterior column ataxia and
pyramidal tract involvement maps to chromosome 8p12-8q12.1. J Med Genet 2004;41:634-
639.
24. Valdmanis PN, Brunet D, St-Onge J, Weston L, Rouleau GA, Dupre N. A founder
haplotype for autosomal dominant sensory ataxia in Eastern Canada. Neurology
2006;67:2239-2242.
25. Meijer IA, Hand CK, Grewal KK, Stefanelli MG, Ives EJ, Rouleau GA. A locus for
autosomal dominant hereditary spastic ataxia, SAX1, maps to chromosome 12p13. Am J
Hum Genet 2002;70:763-769.
26. Mahloudji M. Hereditary spastic ataxia simulating disseminated sclerosis. J Neurol
Neurosurg Psychiatry 1963;26:511-513.
27. Gayle RF, Williams JP. A familial disease of the central nervous system resembling
multiple sclerosis. Sth. Med. J. 1933;26:242-246.

40
28. Bouchard JP, Richter A, Mathieu J, et al. Autosomal recessive spastic ataxia of
Charlevoix-Saguenay. Neuromuscul Disord 1998;8:474-479.
29. Bouchard JP, Barbeau A, Bouchard R, Bouchard RW. Autosomal recessive spastic
ataxia of Charlevoix-Saguenay. Can J Neurol Sci 1978;5:61-69.
30. Bouchard JP. Recessive spastic ataxia of Charlevoix-Saguenay. In: de Jonghe
JMBV, ed. Hereditary Neuropathies and Spinocerebellar Atrophies. Amsterdam: Elsevier
Science Publisher, 1991: 451-459.
31. Peyronnard JM, Charron L, Barbeau A. The neuropathy of Charlevoix-Saguenay
ataxia: an electrophysiological and pathological study. Can J Neurol Sci 1979;6:199-203.
32. De Braekeleer M, Giasson F, Mathieu J, Roy M, Bouchard JP, Morgan K. Genetic
epidemiology of autosomal recessive spastic ataxia of Charlevoix-Saguenay in northeastern
Quebec. Genet Epidemiol 1993;10:17-25.
33. Ogawa T, Takiyama Y, Sakoe K, et al. Identification of a SACS gene missense
mutation in ARSACS. Neurology 2004;62:107-109.
34. Gucuyener K, Ozgul K, Paternotte C, et al. Autosomal recessive spastic ataxia of
Charlevoix-Saguenay in two unrelated Turkish families. Neuropediatrics 2001;32:142-146.
35. Mrissa N, Belal S, Hamida CB, et al. Linkage to chromosome 13q11-12 of an
autosomal recessive cerebellar ataxia in a Tunisian family. Neurology 2000;54:1408-1414.
36. Pascual-Castroviejo I, Pascual-Pascual SI, Viano J, Martinez V. [Charlevoix-
Saguenay type recessive spastic ataxia. A report of a Spanish case]. Rev Neurol
2000;31:36-38.
37. Criscuolo C, Sacca F, De Michele G, et al. Novel mutation of SACS gene in a
Spanish family with autosomal recessive spastic ataxia. Mov Disord 2005;20:1358-1361.

41
38. Criscuolo C, Banfi S, Orio M, et al. A novel mutation in SACS gene in a family
from southern Italy. Neurology 2004;62:100-102.
39. Richter A, Rioux JD, Bouchard JP, et al. Location score and haplotype analyses of
the locus for autosomal recessive spastic ataxia of Charlevoix-Saguenay, in chromosome
region 13q11. Am J Hum Genet 1999;64:768-775.
40. Engert JC, Berube P, Mercier J, et al. ARSACS, a spastic ataxia common in
northeastern Quebec, is caused by mutations in a new gene encoding an 11.5-kb ORF. Nat
Genet 2000;24:120-125.
41. Meijer IA, Hand CK, Cossette P, Figlewicz DA, Rouleau GA. Spectrum of SPG4
mutations in a large collection of North American families with hereditary spastic
paraplegia. Arch neurol 2002;59:281-286.
42. Hazan J, Lamy C, Melki J, Munnich A, de Recondo J, Weissenbach J. Autosomal
dominant familial spastic paraplegia is genetically heterogeneous and one locus maps to
chromosome 14q. Nat Genet 1993;5:163-167.
43. Reid E, Grayson C, Rubinsztein DC, Rogers MT, Rubinsztein JS. Subclinical
cognitive impairment in autosomal dominant "pure" hereditary spastic paraplegia. J Med
Genet 1999;36:797-798.
44. Hentati A, Pericak-Vance MA, Lennon F, et al. Linkage of a locus for autosomal
dominant familial spastic paraplegia to chromosome 2p markers. Hum Mol Genet
1994;3:1867-1871.
45. Nielsen JE, Koefoed P, Abell K, et al. CAG repeat expansion in autosomal
dominant pure spastic paraplegia linked to chromosome 2p21-p24. Hum Mol Genet
1997;6:1811-1816.

42
46. Errico A, Ballabio A, Rugarli EI. Spastin, the protein mutated in autosomal
dominant hereditary spastic paraplegia, is involved in microtubule dynamics. Hum Mol
Genet 2002;11:153-163.
47. Simoes Lopes A, Verreault S, Bouchard JP, Rouleau GA. Further evidence of
linkage between adult-onset autosomal dominant leukodystrophy and chromosome 5q23.
Am J Hum Genet 2002;73:471.
48. Verreault S, Langlois M, Thibault M, Bouchard JP, Simoes Lopes A, Rouleau GA.
A kindred of hereditary adult-onset autosomal dominant leukodystrophy in Charlevoix,
Quebec. Can J Neurol Sci 2002;29:S57.
49. Brown RT, Polinsky RJ, Schwankhaus J, et al. Adrenergic dysfunction in hereditary
adult-onset leukodystrophy. Neurology 1987;37:1421-1424.
50. Eldridge R, Anayiotos CP, Schlesinger S, et al. Hereditary adult-onset
leukodystrophy simulating chronic progressive multiple sclerosis. N Engl J Med
1984;311:948-953.
51. Marklund L, Melin M, Melberg A, Giedraitis V, Dahl N. Adult-onset autosomal
dominant leukodystrophy with autonomic symptoms restricted to 1.5 Mbp on chromosome
5q23. Am J Med Genet B Neuropsychiatr Genet 2006;141:608-614.
52. Coffeen CM, McKenna CE, Koeppen AH, et al. Genetic localization of an
autosomal dominant leukodystrophy mimicking chronic progressive multiple sclerosis to
chromosome 5q31. Hum Mol Genet 2000;9:787-793.
53. Padiath QS, Saigoh K, Schiffmann R, et al. Lamin B1 duplications cause autosomal
dominant leukodystrophy. Nat Genet 2006;38:1114-1123.
54. Dupre N, Howard HC, Mathieu J, et al. Hereditary motor and sensory neuropathy
with agenesis of the corpus callosum. Ann Neurol 2003;54:9-18.

43
55. De Braekeleer M, Dallaire A, Mathieu J. Genetic epidemiology of sensorimotor
polyneuropathy with or without agenesis of the corpus callosum in northeastern Quebec.
Hum Genet 1993;91:223-227.
56. Battistella P, Drigo P, Laverda A, Casara G, De Martin P, Condini A. The
Andermann syndrome. Progressive neuropathy, mental retardation with agenesis of the
corpus callosum. Ital J Pediatrics 1987;13:200-202.
57. Howard H, Mount D, Rochefort D, et al. Mutations in the K-Cl cotransporter KCC3
cause a severe peripheral neuropathy associated with agenesis of the corpus callosum. Nat
Genet 2002;32:384-392.
58. Hauser E, Bittner P, Liegl C, Bernert G, Zeitlhofer J. Occurrence of Andermann
Syndrome out of French Canada - Agenesis of the Corpus Callosum with Neuronopathy.
Neuropediatrics 1993;24:107-110.
59. Deleu D, Bamanikar S, Muirhead D, Louon A. Familial Progressive Sensorimotor
Neuropathy with Agenesis of the Corpus callosum (Andermann Syndrome): A Clinical,
Neuroradiological and Histopathological Study. Eur Neurol 1997;37:104-109.
60. Race J, Makhlouf F, Logue P, Wilson F, Dunham P, Holtzman E. Molecular cloning
and functional characterization of KCC3, a new K-Cl cotransporter. Am J Physiol
1999;277:C1210-C1219.
61. Hould F, Verret S. [Hereditary radicular neuropathy with sensory loss: study of a
French-Canadian family]. Laval Med 1967;38:454-459.
62. Ohta M, Ellefson RD, Lambert EH, Dyck PJ. Hereditary sensory neuropathy, type
II. Clinical, electrophysiologic, histologic, and biochemical studies of a Quebec kinship.
Arch neurol 1973;29:23-37.

44
63. Lafreniere RG, MacDonald ML, Dube MP, et al. Identification of a novel gene
(HSN2) causing hereditary sensory and autonomic neuropathy type II through the Study of
Canadian Genetic Isolates. Am J Hum Genet 2004;74:1064-1073.
64. Roddier K, Thomas T, Marleau G, et al. Two mutations in the HSN2 gene explain
the high prevalence of HSAN2 in French Canadians. Neurology 2005;64:1762-1767.
65. Rahman P, Jones A, Curtis J, et al. The Newfoundland population: a unique
resource for genetic investigation of complex diseases. Hum Mol Genet 2003;12:167-172.
66. Gros-Louis F, Dupre N, Dion P, et al. Mutations in SYNE1 lead to a newly
discovered form of autosomal recessive cerebellar ataxia. Nat Genet 2007;39:80-85.
67. Di Donato S, Gellera C, Mariotti C. The complex clinical and genetic classification
of inherited ataxias. II. Autosomal recessive ataxias. Neurol Sci 2001;22:219-228.
68. Dupré N, Bouchard JP, Verreault S, Rivest D, Puymirat J, Rouleau G. Recessive
ataxia of the Beauce, a new form of hereditary ataxia of pure cerebellar type. Neurology
2002;58:A35.
69. Dupre N, Gros-Louis F, Verreault S, Bouchard J-P, Rouleau G. Recessive Ataxia of
the Beauce, a New Form of Hereditary Ataxia, Maps to Chromosome 6. Neurology
2006;66:A274.
70. Alusi SH, Worthington J, Glickman S, Bain PG. A study of tremor in multiple
sclerosis. Brain 2001;124:720-730.
71. Miller SA, Dykes DD, Polesky HF. A simple salting out procedure for extracting
DNA from human nucleated cells. Nucleic Acids Res 1988;16:1215.
72. Koenig M, Hoffman EP, Bertelson CJ, Monaco AP, Feener C, Kunkel LM.
Complete cloning of the Duchenne muscular dystrophy (DMD) cDNA and preliminary

45
genomic organization of the DMD gene in normal and affected individuals. Cell
1987;50:509-517.
73. Ikeda Y, Dick KA, Weatherspoon MR, et al. Spectrin mutations cause
spinocerebellar ataxia type 5. Nat Genet 2006;38:184-190.
74. Ishikawa K, Toru S, Tsunemi T, et al. An autosomal dominant cerebellar ataxia
linked to chromosome 16q22.1 is associated with a single-nucleotide substitution in the 5'
untranslated region of the gene encoding a protein with spectrin repeat and Rho guanine-
nucleotide exchange-factor domains. Am J Hum Genet 2005;77:280-296.
75. Parkinson NJ, Olsson CL, Hallows JL, et al. Mutant beta-spectrin 4 causes auditory
and motor neuropathies in quivering mice. Nat Genet 2001;29:61-65.
76. Holmes G. A form of cerebellar degeneration of the cerebellum. Brain 1907;30:466-
489.
77. Duquette A, Roddier K, McNabb-Baltar J, et al. Mutations in senataxin responsible
for Quebec cluster of ataxia with neuropathy. Ann Neurol 2005;57:408-414.
78. Stevanin G, Herman A, Brice A, Durr A. Clinical and MRI findings in
spinocerebellar ataxia type 5. Neurology 1999;53:1355-1357.
79. Burk K, Zuhlke C, Konig IR, et al. Spinocerebellar ataxia type 5: clinical and
molecular genetic features of a German kindred. Neurology 2004;62:327-329.