
ROLE DU SYSTEME RACINAIRE DANS LA RESISTANCE DU MIL A …
51
ROLE DU SYSTEME RACINAIRE DANS LA RESISTANCE DU MIL A LA SECHERESSE. MEMOIRE DE FIN DE PREMIERE ANNEE DE FORMATION A LA RECHERCHE Sani DAOUDA OUSMANE Institut des Radio-Isotopes. Université de Niamey. BP 10727Niamey Niger. Octobre 1991 1
Transcript of ROLE DU SYSTEME RACINAIRE DANS LA RESISTANCE DU MIL A …

ROLE DU SYSTEME RACINAIRE DANS LA RESISTANCE DU MIL A LA SECHERESSE.
MEMOIRE DE FIN DE PREMIERE ANNEE DE FORMATION A LA RECHERCHE
Sani DAOUDA OUSMANE Institut des Radio-Isotopes. Université de Niamey.
BP 10727 Niamey Niger.
Octobre 1991
1

Ce mémoire termine la première année d’Üne formation à la recherche qui s’insere dans le cadre d’un contrat “formation-insertion” de I’ORSTOM et fait l’objet d’une convention entre I’ORSTOM et l’Université de Niamey (République du Niger). Le but final de cette formation est la présentation d’une these de doctorat dont le thème est: “Etude de l’alimentation hydrique et minérale du mil en - milieu contraignant”.
Durant cette première année d’investigation deux opérations ont été conduites: - l’étude de la réaction du système racinaire du mil face à un déficit hydrique post- floral, réalisee en plein champ pendant la contre-saison froide 1988-1989 et la contre-saison chaude 1990; w l’étude de la réaction du système racinaire du mil face à un déficit hydrique de début de cycle, menée en rhizotron au CERAAS de Bambey au SCnégal durant l’hivernage 1990.
Les principaux résultats obtenus sont présentés dans ce document.

DEFINITION DU PROBLEME
Le déficit hydrique apparaît dans une plante quand ses pertes en eau (transpiration) excèdent ses possibilités d’absorption racinaire. Cette contrainte hydrique se répercute rapidement sur les activités métaboliques de la plante et, à plus long terme, sur sa morphologie. Le déficit hydrique peut intervenir de mani&re transitoire, pendant les heur& chaudes de la journée ou bien être plus durable s’il résulte de conditions climatiques défavorables.
Dans la zone tropicale semi-aride, deux périodes de fortes probabilités de sécheresse menacent plus particulièrement les cultures annuelles: la première au moment de l’installation de la culture, la seconde à partir de la floraison (Sivakumar,l986). L.a seconde situation (appelée indifféremment sécheresse terminale, ou de fin de cycle, ou post-florale) est assurCment la plus dangereuse. Elle survient en effet alors que les resemis ne sont plus possibles et affecte la fécondation, ce qui peut remettre en cause le rendement en grain escompté.
Face au dCficit hydrique la plante rCagit par des mécanismes morpho- physiologiques d’évitement ou de tolérance à la sécheresse (Levitt 1980, Turner 1986). L’un des plus importants de ces mécanismes est l’extension du syst&me racinaire (Passioura 1983, Clarke et al. 1984, Jones et al. 1984, Muchow et al. 1986). Il est généralement admis que l’utilisation de la réserve en eau du sol est étroitement liée à la morphologie (profondeur et densité) et à l’activité (ConductivitC hydraulique notamment) du système racinaire (Gregory et al. 1984, Ludlow et aL 1988, Gregory 1989). Il est Cvident què le système racinaire le plus profond met à la disposition du vCgCta1 le plus grand volume de sol à exploiter. Jordan (1983) montre chez le sorgho, qu’un enracinement profond peut augmenter jusqu’à 20% le rendement au cours des années sCches. L’amélioration, en même temps que la sCcurisation de la production agricole, pourraient donc passer par l’identification de cultivars à enracinement profond. Un effet génotypique sur la profondeur et la forme de l’enracinement a en effet été démontré chez différentes espèces cultivées (Shriniwas et al. (1974), Batcho et al. (1990) sur le mil; Hurd (1974) sur le blé; Taylor et a!. (1978) sur le niébé; Mathews et al. (1988) sur l’arachide; Jordan et al. (1979) sur le sorgho). Mais la question se pose des relations entre un enracinement profond et le développement des parties aériennes d’une part (concurrence ou synergie entre les différents organes?) et d’autre part la dynamique d’utilisation de l’eau du sol chez le mil. Turner et Nicholas (1987) obtiennent une plus grande biomasse à la floraison et un rendement en grain supérieur chez le génotype qui a montré la plus grande vitesse de croissance racinaire en phase d’installation. Mais dans le cas d’un stress hydrique terminal, qui intervient à la floraison, au moment même où on semble admettre que la croissance du syst&me racinaire prend fin, quel sera le type d’enracinement à la floraison le

plus favorable, pour une meilleure utilisation de l’eau du sol et le maintien du rendement ? Un enracinement étendu et dense, à grande capacité et rapidite d’exploitation des réserves du sol, ou bien un système racinaire plus ou moins suspendu, laissant généralement de l’eau disponible résiduelle à maturité ?.
.
Quel est le role de cette structure du système racinaire à la floraison pour le comportement ultérieur du mil concernant l’exploitation de la réserve en eau du sol, le maintien de la morphologie et des capacites photosynthetiques, le remplissage des grains, etc. pendant une sécheresse terminale ?
Qu’il s’agisse d’un système racinaire Ctendu et dense ou d’un système racinaire suspendu, quelle serait la réaction face à un déficit hydrique terminal ? Une extension (néogenèse) en profondeur ou latérale ? Comment alors différencier ce phénomène morphologique du système racinaire, d’une simple entrée en activité de racines déjà présentes dans le sol ? Cette distinction est importante car l’incidence des deux processus sur l’utilisation des ressources carbonées de la plante et donc sur le rendement est différente.
La repense à ces questions permettra de préciser, pour les conditions de cultures étudiées (sol, regime hydrique), les strategies d’utilisation de l’eau du sol développées par les différents cultivars de mil ainsi que les réactions morpho- physiologiques qui leur sont associées. Pour ce faire il faut:
l/ caractériser la structure initiale de la plante à la floraison a travers la répartition de la biomasse entre les différents organes (racines, tiges, feuilles, épis);
2/ déterminer les liens existant entre cette structure à la floraison et la croissance racinaire en phase d’installation de la culture; son r81e dans l’exploitation post-florale de la réserve en eau du sol, et le maintien du rendement, en période de déficit hydrique;
3/ déterminer l’évolution morphologique du système racinaire au cours d’une période de stress hydrique post-floral;
4/ parallèlement, caractériser l’activité du système racinaire et sa dynamique par le suivi de l’extraction de l’eau du sol et l’étude de sa répartition dans le profil durant la période de déficit hydrique;
5/ relier ce fonctionnement racinaire à la croissance et à l’état hydrique de la partie aérienne pour en préciser les conséquences sur l’élaboration du rendement.
La méthodologie adoptée repose sur une approche comparative du fonctionnement racinaire de cultivars réputés plus ou moins résistants à. la sécheresse, soumis à des régimes hydriques naturels (pluvial) ou contrôlés (contre- saison, rhizotron), avec témoins irrigués.

. 1. ESSAI EN RHIZOTRON
Résumé:
La vitesse de colonisation verticale du système racinaire est étudiée en rhizotron pour six cultivars de mil (Pennzktum glaucum) et pour trois traitements hydriques en début de cycle, du semis jusqu’à la montaison (40 jours). Il est mis en évidence une relation entre la vitesse d’enracinement et l’état hydrique du sol d’une part, et une dispersion variétale de la morphologie du système racinaire en condition d’alimentation hydrique normale d’autre part.

INTRODUCTION
Cette étude fait suite à des essais de criblage de variétés de mil pour la résistance à la sécheresse (Bidinger et al. 1988) et d’analyse des mécanismes morphophysiologiques de cette résistance (Do et aL. 1989, Anonyme 1990, Da Silva et al. 1989), sous l’effet d’un stress hydrique en fin de cycle notamment. Elle a pour but de déterminer le comportement de six cultivars de mil face à un stress hydrique appliqué en début de cycle (40 premiers jours après le semis). Dans ces conditions, la survie du jeune plant dépend de son aptitude à coloniser rapidement le sol et à disposer d’une réserve hydrique potentielle importante. *
L’essai a été réalisé durant l’hivernage 1990 à Bambey au Sénégal collaboration avec plusieurs chercheurs: E. Batcho, D. Laffray, P. Louguet laboratoire de physiologie végétale et d’écophysiologie végétale appliquée l’université Paris XII, DJ. Annerose, J.L Khalfaoui et A. Fofana CERAAS/CNRA de Bambey, F. Do et O.S. Daouda de I’IRI.
en du de du
On ne traitera dans ce rapport, que de la vitesse de colonisation du sol par les racines des différents cultivars, en raison de l’indisponibilité des données relatives aux autres paramètres racinaires (biomasse et longueur totales).
1. MATERIEL ET METHODES
1.1. Le climat
L’essai est réalisé en fin d’hivernage, d’août à septembre 1990. De façon générale, le climat est comparable à celui des environs de Niamey: vents et rayonnements forts, températures élevées aboutissant à une demande evaporative intense, supérieure à Smm/jour (Dancette, 1976). Le degré d’hygrométrie est faible sauf justement en hivernage où il peut atteindre des valeurs supérieures à 90%.
1.2. Le sol
Les rhizotrons utilisés pour l’essai sont remplis avec de la terre fine récoltée à partir d’horizons de surface du sol décrit ci-aprés. II s’agit d’un sol ferrugineux tropical profond, comparable à celui de Niamey mais plus riche en argile. La densité apparente en sol non cultivé est de l’ordre de 1.6 donnant une porosité totale faible (40%). L’eau utile correspondant à la différence entre l’humidité à la capacité au champ et le point de flétrissement varie entre 6 et 12 mm par tranche de 10 cm de sol. La capacité d’échange cationique est faible et le complexe absorbant faiblement saturé. L’azote total est de 0.02 à 0.1% en surface avec une nette diminution en profondeur. Le C/N (entre 8 et 15) indique une matière organique labile.

1.2. Dispositif expérimental
Il comporte 6 variétés, 3 traitements et 6 répétitions. Soit en tout un ensemble de 108 rhizotrons entièrement randomisés.
1.2.1. Le rhizotron
Il a été décrit par le CERAAS (1990). Il est composé de tubes en PVC de 100 cm de hauteur et 16 cm de diamètre ayant une face plane transparente en plexiglas. Le tube est disposé en position inclinée de 45” sur un chassis métallique du cote de la face transparente. Sous l’effet du geotropisme, les racines viennent s’appliquer en partie contre le plexiglas permettant ainsi leur observation.
Le remplissage du rhizotron se fait avec de la terre fine qui, dans notre cas a été mélangée à 4 g d’engrais NPK 10 21 21 correspondant à 40 kg pour 10000 poquets, dose plutôt modérée. La terre est tassee de manière à reconstituer au mieux la situation réelle au champ (densité de 1.45 à 1.65). Une couche horizontale de gravillons de 5 cm d’épaisseur est creée au milieu du rhizotron de façon a rompre les flux capillaires et à tester la capacite des racines à franchir un “obstacle”.
Le rhizotron est recouvert d’un film plastique noir qui crée les conditions d’obscurite habituelles au milieu racinaire. Un autre film dans la partie supérieure empêche l’entrée de la pluie, mais limite aussi l’évaporation directe du sol.
1.2.2. Le matdriel v4gCtal
Il comprend 5 varietés cultivées de cycle moyen (90 jours) et une forme sauvage : - deux cultivars améliorés par I’ICRISAT: ICMVIS85327 (IC27) et ICMVIS86330
(IC30). Ils ont montre des comportements opposes face à un stress hydrique terminal;
- une variété sélectionnée par I’IRAT et vulgarisée par I’INRAN, HKP. Elle est considéree comme résistante;
- une forme sauvage (sp violweum), fournie par les généticiens de I’ORSTOM; - deux cultivars fournis par le service semencier du CNRA/ISRA de Bambey:
SOUNA et IBV8004.
Les quatre premières variétés sont celles sur lesquelles travaillent en condition de stress hydrique post-floral, les équipes de l’IRI/ORSTOM à Niamey, des laboratoires de physiologie végétale des Universités Paris VII et Paris XII et l’equipe associée CEA/CIRAD/ORSTOM au Centre d’Etudes Nucléaires de Cadarache.

1.2.3. L’zkimentation hydrique
Trois conditions hydriques sont testées. Elles sont analogues à des situations reelles qui se présentent dans le sol en début d’hivernage:
- le traitement hydrique 1, appelé Cl. II s’agit d’une alimentation en eau sans limitation, qui maintien le profil hydrique du sol à la capacité au champ pendant toute la durée de l’essai. L’arrosage est quotidien a la dose 200 cm3 par paquet (rhizotron);
- le traitement hydrique 2, appelé C2. C’est une alimentation hydrique déficitaire qui consiste à partir d’un sol porté à la capacité au champ et ne plus arroser. C’est le cas assez fréquent d’une première pluie très abondante suivie d’une longue période de sécheresse;
- le traitement hydrique 3, appelé C3. C’est un autre scénario où le profil initial est à la capacité au champ dans sa partie inférieure (50 à 100 cm), avec une humidité plus faible dans la partie superieure. Ce cas peut se rencontrer soit lors d’un semis tardif ou d’un resemis, soit en prtsence de nappe perchee ou peu profonde.
1.3. Mbthodes de mesure
1.3.1. Le suivi de l’avancement du front racinaire
On mesure la racine la plus longue, observée à travers le plexiglas, deux fois bar semaine sur chaque rhizotron. La vitesse d’avancement du front racinaire est calculée par simple différence.
1.3.2. La colonisation du sol
Une note entre 1 et 5 permet d’apprécier visuellement le niveau de colonisation du sol Dar les racines à travers la face transoarente du rhizotron.

X.3.3. Les autres param&es mesur&
Plusieurs parametres de caractérisation du systeme racinaire ont également Cte mesurés à la fin de l’essai, quarante jours après le semis (Batcho er aL 1991). Il s’agit de:
- la distribution des racines. A la fin de l’essai le rhizotron est découpé en tranches de 20 cm, puis la longueur totale et la masse des racines sont déterminées pour chaque tranche. Le nombre de racines est aussi déterminé pour l’horizon O-20 cm;
- la racine la plus longue. Sur la moitié des répétitions, le systéme racinaire est dégagé intact et la racine la plus longue est mesurée;
- le volume racinaire. Il est déterminé en plongeant l’ensemble des racines dans une eprouvette graduee contenant de l’eau. Le volume d’eau déplacée donne le volume des racines.
2. RESULTATS
Le tableau 1.1 donne pour chacun des 4 génotypes dans les trois conditions hydriques, les valeurs moyennes d’avancement du front racinaire (cm/jour) entre la germination et la 5è semaine (montaison);
Tsblcau 1.1 : Vitesses moyernes d'avancement du front racineire entre La germination et le 56 semaine (cm/jour, moycmc f kart-type).
IC30 IC27 HKP SAUVAGE
Cl 1.6 f 0.2 2.1 f 0.5 1.9 f 0.5 1.3 t 0.8 c2 3.3 t 0.3 3.5 f 0.3 3.3 f 0.3 2.6 î 1.6 c3 3.0 f 0.3 2.9 î 0.3 2.9 f 0.4 2.7 t 1.5
En phase d’installation. le déficit hydrique accélère l’extension verticale du système racinaire chez le mil. En moyenne, sur l’ensemble de la période, il n’apparaît pas de différence significative entre les 2 traitements où l’alimentation en eau est restreinte (C2 et C3). En revanche, si on distingue les 3 premières semaines de croissance de la période qui suit on constate que jusqu’à la 3è semaine, en C2 (contrainte hydrique moyenne) la vitesse d’enracinement est comparable à celle du témoin irrigué (Cl) et nettement inférieure à celle du traitement C3 (contrainte hydrique sevère).
La situation s’inverse au cours de la période suivante (semaines 4 et 5), avec des vitesses d’enracinement deux fois plus fortes en CL? qu’en C3. Dans ce même

temps, l’avancement du front racinaire est ralenti chez le témoin irrigué face à un déficit hydrique en phase d’installation, la capacité! d’extension en profondeur du systeme racinaire reste donc la même que le deficit soit précoce (traitement C3) ou tardif (traitement C2). Elle ne diffère pas non plus entre les 3 varietés cultivées. Pour la forme sauvage, l’extension verticale du front racinaire sous l’effet du déficit hydrique est moins forte que chez les cultivars et également moins sensible au type de déficit (precoce ou tardif).
Le tableau 1.2 distingue les trois premières semaines de début de croissance de la période qui suit.
Tableau 1.2 : Vitesses moyennes dlevancement du front racineire (cm/jour) pour Les semaines 1 B 3 et 4 et 5.
Après 4 semaines, en absence de déficit hydrique, la variété IC27 montre une colonisation du rhizotron à la fois plus dense sur les 60 premiers centimétres et plus profonde que les variétés HKP et IC30. Sauf pour la forme sauvage dont l’enracinement est faible et uniforme sur 60 cm de profondeur, les densités racinaires apparaissent plus fortes en surface qu’en profondeur.
Tableau 1.3 : Profils de densit6 racinaire en condition Cl: irrigation journaliere.
Prof. 1 c30 1 C27 (cm
0
20
40
60
80
100
4 4
1’ 3
2
5
5
I 5
1
1
HKP SAUVAGE 4 2
4 2
Ill 3 2
En revanche, en situation de déficit hydrique (tableaux 1.4 et 1.5) la densité de racines est plus faible en surface qu’en profondeur. Les rhizotrons yont colonisés par les racines jusque dans leur partie inférieure, sans différence sensible entre les quatre génotypes testés.
- 10 -

Tableau 1.4: Profils de demit racinairc en condition CZ: rkerve maximale eu semis et sans irrigation ultCrieure.
Prof IC30 IC27 HKP SAUVAGE (cm)
0
20
40
60
80
100
2
3
1 4
2
1
3
2
11 4
3
2
2
3
1 4
2
2
2
2
l 3
3
1
Tableau 1.5 : Profils de densit6 racinaire en condition C3: rherve maximale seulement en profondeur, au semis et sans irrigation ulterieure.
Prof. IC30 fC27 HKP SAUVAGE (cm)
O---
-l 3 2 3 2 20
2 2 2 2 40
100 LEElBE 3 3 3 2
60 3 3 3 3
80 2 3 3 2
3. CONCLUSION
L’essai a permis de mettre en évidence deux points essentiels :
- l’effet du stress hydrique sur la croissance et la forme du système racinaire du mil: en phase d’installation des plantes, l’avancement du front, racinaire est stimulé par un déficit hydrique, que celui-ci soit précoce ou tardif. Cette extension verticale du système racinaire s’accompagne d’un colonisation plus importante des horizons humides. Hormis pour la forme sauvage dont l’enracinement est gCnCralement moins rapide et moins dense que celui des formes cultivees, la reaction des plantes est la même entre les cultivars.
- deux types d’enracinement semblent se dégager: un enracinement profond qui, même en condition d’irrigations fréquentes, colonise le sol en profondeur (IC27) et un enracinement plus superficiel susceptible de s’étendre en profondeur en cas de déficit hvdriaue tIC30. HKP et sauvage).
- ll-

II. ESSAI DE CONTRE SAISON FROIDE 1988-1989
1. CADRE EXPERIMENTAL
1.1. Le climat
Le tableau 11.1 (Cf. fin du chapitre) dorme les moyennes des principaux param&res climatiques pour les environs de Niamey (Sivakumar, 1986) et le tableau 11.2 (Cf. fin du chapitre) les valeurs pour les années d’exphrimentation 1988-1989 et 1990. Sur toute l’étendue du Niger méridional (zone à vocation agricole) on observe :
- une saison de pluie estivale courte, à vents dominants humides du sud-ouest. La pluviométrie annuelle moyenne est de l’ordre de 500 mm, mais variable en quantité et aléatoire dans sa distribution. 75% des pluies tombent durant les mois de juillet, août et septembre;
ih une saison s&che de 7 à 10 mois à vents continentaux de secteur nord-est armattan);
- une insolation de l’ordre de 3000 heures par an. Des températures Clevées, jusqu’à 40°C en mars, avril et mai. Une hygrométrie moyenne faible, rarement plus de 50% sauf en saison des pluies.
La période de contre-saison froide se situe de mi-novembre A mi-février. Elle est caractCrisée par des températures moins ClevCes (maximum 32”C), des humidités relatives faibles (moins de 15% à 12H) et une pluviométrie nulle. La contre-saison chaude s’Ctend sur les mois de mars à mai. Les températures plus ClevCes (ph~s de 4O”C), la demande évaporative va jusqu’à 8 mm/j et les humidités relatives sont faibles jusqu’à l’arrivée des premières pluies, fin mai début juin.
Dans l’ensemble, les années d’essai 88, 89 et 90 se rapprochent d’une annCe normale, en dépit des l’écarts pluviométriques.
1.2. Le sol
L’essai a été conduit sur un sol ferrugineux tropical peu lessivé, développé sur un matériau sableux. Les principales caractéristiques physiques et chimiques sont rCsumées dans le tableau 11.3. La fraction fine constituée essentiellement de kaolinite et de sesquioxydes, est faible et varie d’un site à l’autre (entre 1 et 15%). Sur la base des taux d’argile et de sable fin, on peut distinguer le site 4 des sites 1, 9
- 12 -

et 12. D’une manière génerale, la matière organique est caractérisee par un C/N inferieur à 10, le SOI est profond et perméable, la capacité de rétention en eau variant de 70 à 80 mm dans le premier mètre de soi (Daouda et a1.,1991).
. 2. MATERIEL ET METHODES
Le dispositif statistique est un essai factoriel à deux facteurs (un facteur variétal à 4 niveaux, un facteur hydrique a 2 niveaux) à 4 répétitions non randomisées (3 pour le mil sauvage) regroupées en deux parties Cgales correspondant chacune à un des traitements hydriques (fig. 11.1).
2.1. Le matkiel vég#tal
II s’agit de quatre génotypes de mil (Penniwtum gfaucum) a cycle moyen de 90 jours, présentant des comportements contrastés face au stress hydrique post- floral :
s HKP, variété selectionnée par I’IRAT, vulgarisée par I’INRAN, considtrte comme résistante à la sécheresse;
e ICMVIS85327 et ICMVIS86330, cultivars améliorés par I’ICRISAT, ayant montré des comportements opposés face à un stress hydrique terminal (Bidinger et a~!, 1988), désignés respectivement IC27 et IC30, par la suite;
- un mil sauvage (Subspecies viohceum), supposé être résistant.
Ils constituent le facteur variétal d’un essai comparatif de la croissance et du developpement du mil sous alimentation hydrique differenciée.
2.2. L’alimentation hydrique
L’alimentation hydrique représente le second facteur étudié. A partir de la floraison, l’irrigation jusqu’alors uniforme fait place à deux traitements hydriques:
- un traitement irrigué où l’arrosage se poursuit tout au long du cycle cultural,
- un traitement stressé où l’irrigation est interrompue à la floraison pour provoquer ‘un stress hydrique post-floral.
L’irrigation au goutte à goutte est conduite au moyen de goutteurs de 41/h de débit, à raison de deux goutteurs par mètre sur la ligne de semis. L’apport d’eau est auotidien en début de cycle, puis il est ‘effectué tous les 2 jours. La dose est
- 13 -

calculée sur la base de I’ETP PENMAN (moyenne 1949-1984) et des coefficients culturaiut Ctablis par Dancette (1983).
2.3. Conduite de l’essai
Les quatre vari&& sont semCes en poquets (dCmariés à trois brins), avec un espacement de lm x lm (densité de 10000 poquets par hectare). Le semis a été fait le 27 octobre 1988 et la récolte le 16 février 1989. Un apport modé,rk d’engrais en solution (45 unités d’azote et 22.5 uni& de phosphore) a été effectué en deux fois: au dépariage et à la moptaison. Les sarcla-binages et les traitements phytosanitaires ont été effectuks dès que nécessaire.
2.4. M&hode d’estimation de la biomasse racinaire
2.4.1. Le carottage racinaire
Cette étape consiste à prélever des Cc~antillons de terre ou carottes d’un volume connu a différentes profondeurs, et à extraire les racines par lavage à l’eau. Le carottier que nous avons utilisé a un diamètre de 70 mm et 200 mm de hauteur, soit un volume utile de 770 cm3.
Sur chacune des rCpétitkms (un poquet par parcelle élémentaire), les prClévements sont faits dans 2 plans privilégiés: sur la ligne de semis et perpendiculairement à la ligne de semis (Fgure 11.2). Trois sites de carottage caractérisent chaque plan de mesure: sous le poquet, à 25 cm et SO cm du poquet. Les carottes sont prélevées tous les 20 cm jusqu’à 140 cm de profondeur.
2.4.2, La m&hode de calcul de la masse de racine
Cette méthode a été mise au point en 1990 par T’hiéry et al.(Cf. annexe). Par carottage on mesure la densité volumique de racines, exprimée en mg de racine/cm3 de sol, en 7 profondeurs et sur 5 sites par poquet, ce qui donne 35 mesures par poquct. Le principe de calcul de la masse de racines est l’intégration de la densité volumique sur !‘ensemble du poquet. Le voisinage du poquet est divise en 8 octants, tenant compte de la dissymétrie due à l’irrigation en ligne de goutteurs:
1 deux octants sont axés sur la ligne de l’irrigatio:,, - deux octants sur la ligne perpendiculaire; - quatre octants sur la diagonale.
Au niveau de chaque octant on passe de la densité volumiqKe à la densité surfacique par une projection sur l’un des plans de base de l’octant. Pour obtenir la
- 14 -

masse il suffit d’intégrer sur le plan dé projection, puis de faire la somme des 8 octants. L’octant diagonal est considért’ comme la moyenne des deux autres octants.
2.5. Mesure de l’eau du sol
LE suivi de I’humiditC du sol est assure par la méthode neutronique, sur des tubes d’ac& de 2 mètres de profondeur utile. Les relevés des profils hydriques sont effectuCs une fois tous les deux jours pendant la phase de stress hydrique. Une des 4 placettes-&Pétitions est affectee au bilan hydrique et comporte un tube de mesure neutronique, situé en interligne de semis entre deux poquets.
3. RESULTATS
3.1. Caracttirisation de la parcelle avant l’application du stress Lydrique
3.1.1. Les stocks hydriques avant l’application du stress
Le tableau II.4 donne les stocks hydriques à l’arrêt de l’irrigation (JAE 60), pour chacun des génotypes au niveau de la parcelle de bilan.
Tableau II.4 : stocks hydriques (mm) à l’arrêt de l’irrigation (JAE 60) ; CC=Capacité au champ.
Prof(cm) IC30 lC27 HKP SAUVAGE
O-50 66 47 38 18 50-100 53 67 76 72
100-150 91 102 126 105 150-200 100 106 137 110
O-200 291 322 377 335 Stock Cc 27s 357 357 357
On constate, d’une part, que les .réserves sont pleines au moment de l’arrêt de l’irrigation. et d’au:re par qu’elles sont comparables entre .les @notypes, SUT le
Premier mètre de sol.
3.1.2. La biomasse adrienne des cultivars
s#. . I-e tableau 11.5 donne les valeurs de quelques paramètres morphologiques aerlens, 59 jours après emergence, 3 jours avant l’arrêt de l’irrigation.

Tableau 11.5 : gianesse &ieme des trois cultivero A 59 JAE (UV: surface folirirt verte, cm*; nb 1: tire & tiges; HSt: matit!re s&chc l Crieme totale, g/m*).
Il ressort de ce tableau, que le cultivar 00 a une biomasse aérienne plus importante que les autres gCnotypes au départ du stress hydrique. Il a une surface d’echange (surface foliaire verte) et un nombre de talles plus Clevés que les deux autres cultivars.
3.2, LYvolution au cours du stress hydrique
3.2.1. LVvolution des stocks hydriques
Le tableau II.6 donne les variations de stock au cours de la période du déficit hydrique. En nCgligeant le drainage en profondeur (stocks hydrique voisins des réserves à la capacité au champ : Cf tableau 11.1) on peut les assimiler aux consommations en eau des cultures.
Tableau 11.6 : Variations de stock (mn> au cours de la tiriode du déficit hydrique. (VS%: pourcentage de variation par rapport au stock initial).
7- Prof(cm) lC30 1c27 HKP SAUVAGE
O-50 36 34 24 35 50-100 26 33 37 35
100-150 46 28 40 31 150-200 33 32 24 24
O-200 142 127 125 125 VS% 49 39 33 37
On constate un comportement identique de IC27, HKP et du sauvage, comportement différent pour IC30 : sur 50 jours, IC30 a évaporé 142 mm contre 12s en moyenne pour les trois autres génotypes.
Le tableau 11.7 donne pour chaque génotype, la répartition entre les couches O-100 et 100-200 cm de l’eau consommée au cours de deux périodes consécutives (O-10 et 10-50 jours après le début de stress).
- 16 -

i‘
Tableau 11.7 : Repartition dans Lt sol de l'eau consonrnk pw its differtnts gktotypes au cours 6 déficit hydriqw (X de La consomnstion totale de chaux p&riode, JADS: jours aprk le d&ut de stress)
JADS
lC30 IC27 HKP SAUVAGE
O-10 10-50 O-10 10-50 O-10 10-50 O-10 10-50
Prof(cm) D-100 72 27 80 52 42 85 44
100-200 28 73 20 48 27 58 15 56
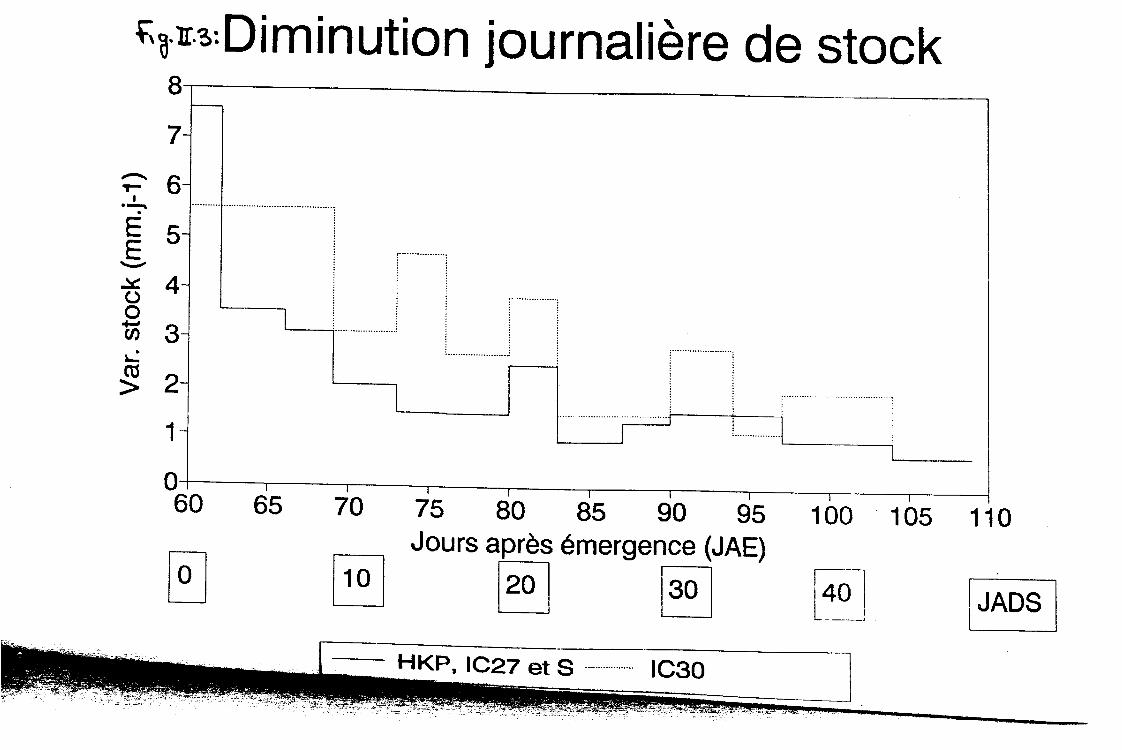
Au cours des 10 premiers jours, les trois quarts environ de l’eau consommée par les cultures proviennent du premier mètre de sol, sans différence notable entre les quatre génotypes. En revanche au cours des 40 jours suivants, le cultivar IC30 extrait plus d’eau en profondeur (73%) que les trois autres qui restent comparables entre eux. Ceci se voit clairement à travers les figures II.3, II.4 et 11.5, présentant les variations journalières de stock au cours du stress hydrique. La figure II.3 compare IC30 à la moyenne des autres génotypes. On constate que 15 jours après l’application de la contrainte hydrique, la diminution de stock est encore élevée chez le premier alors qu’elle tombe à moins de 2mm/jour chez les autres. Les figures II.4 et II.5 montrent que cette différence provient des couches profondes (100 - 150 cm) que le cultivar IC30 semble mieux exploiter.
3.2.2. L’bolution de la partie akienne
Elle est représentee sur les figures II.6 à II.8 et concerne la surface foliaire verte, le nombre de tiges et le poids des Cpis des trois cultivars (moyenne par poquet).
La surface foliaire verte (figure 11.6) diminue au cours du stress hydrique avec des pentes comparables entre les trois cultivars, Mais le cultiva-r IC30 qui avait au départ une surface foliaire verte superieure de 40% à celle des deux autres variétés, conserve cette supériorité tout le long de l’essai. Il a de même un nombre de tiges supérieur à celui des autres variétés pendant toute la durée du stress hydrique (Fig. 11.7).
L’évolution du poids des epis est identique pour les trois cultivars, jusqu’à 78 JAE (Fig.JI.8). A partir de cette date, la masse des épis augmente fortement chez IC27 et suriout IC30, alors qu’elle n’évolue pratiquement plus chez HKP.
Lc tableau II.8 donne la situation à 85 JAE. Lc cultivar IC30 a une biomasse aérienne supérieure, .en particulier au niveau du poids des épis : 62.7 g/ooouet contre 43 et 27 nour IC27 et HKP respectivement. L’r1 1 1 .
- 17 -

Tableau 11.8: gianasse abrierwe des trois cultiiars b 85 JAE (SFv: surface foliaire verte, cm?; Epis et Kit: poids d’6pis et de metiere sèche totale, g/m’)
3.3. Caract6risation de la parcelle B la récolte
3.3.1. Les stocks hydriques à la r&olte
Ils sont donnés dans le tableau 11.9. Les zones les plus humides au moment de l’application du stresse hydrique restent les plus humides à la fin. Les horizons de surface (0 - 100 cm) sont bien asséchés de façon uniforme par tous les cultivars. Ceci est également illustré par la figure 2.4.
Tableau 11.9: Stocks hydriques 0110 b la r6colte.
O-200 149 1% 253 210
3.3.2. La biomasse aMenne & la r&olte
Les rendements en matière ‘sèche totale et en grain sont donnés dans tableau 11.10. Les rendements obtenus pour les parcelles de bilan hydrique sont comparables aux moyennes générales des 4 répétitions, pour chaque cultivar. Ce qui confirme la bonne représentativité de ces parcelles réservées au bilan hydrique.
Tableau 11.10: Rendements en grains et matière séche totale B la récolte (g/m'). Xi: moyenne pour Le cultivar i; EH: valeur obtenue pour !a parcelle du bilan hydrique.
grains mst
IC30 IC27 Xi ElH Xi BM
31 27 62 77 114 103 181 196
HKP Xi EH
65 64 200 184
- 18 -

Le cultivar IC30, qui jusqu’au JAE 85 avait la plus forte biomasse aérienne, se retrouve à maturité avec une production de matière seche totale et un rendement en grains inférieurs à ceux des deux autres cultivars.
3.3.3. La biomasse racinaire A la rtkolte
La distribution des racines dans le sol à la récolte, exprimée en mg de racines par cm3 de sol, est donnée dans les tableaux II.11 pour l’aplomb du poquet et II.12 pour la zone latérale autour du poquet.
Tableau 11.11 : Densité de racines B l~splomb du poquet (mg/cm3)
Proftcm) IC30 IC27
O-40 7.22 4.50 40-140 0.80 0.09
HKP SAUVAGE
3.79 4.93 0.07 0.18
Tableay 11.12 : bensite de racines lat&alernent autour du poquet 0fWcm 1
Prof(cm) IC30 IC27
O-40 1.65 2.12 40-140 0.20 0.10
HKP SAUVAGE
0.98 1.60 0.05 0.22
Sous le poquet, le cultivars IC30 se distingue des autres génotypes par une densité racinaire sensiblement plus forte, notamment en profondeur.
Latéralement aussi IC30 est celui des trois cultivars qui développe la plus forte biomasse racinaire en profondeur, a égalité avec le mil sauvage. En surface, c’est IC27 qui colonise le mieux le sol.
- 190

t ! Au total, IC30 et le mil sauvage parviennent B une meilleure occupation des
horizons de profondeur à la récolte, tandis que HKP montre le plus faible dkveloppement racinaire des trois cultivars.
4. DISCUSSION-CONCLUSION
L’évolution de la biomasse aérienne et la consommation en eau au cours du stress hydrique, ainsi que la structure de la plante à la récolte sont résumées dans le tableau 11.13.
Le cultivar IC30 ayant, au moment de l’application du stress, la biomasse aérienne la plus importante, montre entre 78 et 85 JAE la plus forte augmentation du poids d’épis (Fig. 11.8). Cette croissance rapide des épis est liée à une surface foliaire verte (source d’assimilats carbonés) supérieure à celle des deux autres cultivars et à une consommation en eau importante soutenue notamment par l’exploitation des horizons profonds entre 72 et 82 JAE (Fig. II.5).
La consommation en eau, de même que les surfaces foliaires initiales des deux autres cultivars sont pratiquement Cgales, mais IC27 se distingue de HKP en maintenant sa surface foliaire verte plus longtemps: entre 77 et 85 JAE, sa surface assimilatrice est en moyenne supérieure de 50% à celle de HKP (Fig. 11.6). Dans le même temps, le poids de ses épis augmente encore fortement, tandis que HKP ne varie plus.
A partir de 85 JAE, la consommation en eau devient inférieure à 2mm/jour pour tous les génotypes, et la croissance des Cpis de 100 et HKP se trouve bloquée. Malgré un poids d’Cpis supérieur, IC30 produit à la récolte moins de grains que les deux autres cultivars.
Ainsi, le cultivar ayant la biomasse la plus importante avant l’application du stress s’est comporté le mieux face à la sécheresse pendant les 25 jours de déficit hydrique. On peut penser qu’il aurait produit le plus de grain (ayant le poids d’épis le plus élevé) à maturité si le déficit hydrique, au lieu de se prolonger encore deux semaines, avait é.té interrompu à 85 JAE.
Cependant, une baisse de matière sèche totale est enregistrée B la récolte. Où est passée cette biomasse ? Nous avons observé en rhizotron et en phase d’installation (Cf. lère partie) un effet de rhizogénèse lié au stress hydrique. Le phénom&ne est-il le même, et la biomasse aérienne perdue au cours du stress aurait-elle été affectée à l’extension du système racinaire ? Ceci expliquerait en partie le développement important du système racinaire et le faible rendement en grains observés chez le cultivar IC30. Pour le préciser, il faudra suivre en paralkle l’évolution des biomasses souterraine et aérienne au cours du stress.
- 20 -

Tableau 1Z.13 faible).
: Rkapitulatif des rkultats (+++: fort; ++: moyen; +
IC30 Ic27 HKP SAUVAGE1
MSt initiale(59JAE) +++ MSt (85 JAE) +*+ HSt rkolte (109 JAE) + racine rkolte (109 JAE)++* eau consde 142 59 II 109 JAE
++ + ++ + +++ +++ ++ 127 1;5 1;5
Il est à noter que les résultats présentés, reposant sur 2 et 4 répétitions additionnées respectivement pour les biomasses aériennes et racinaires et sur un sondage neutronique par parcelle pour le bilan hydrique, méritent d’être vérifies. En revanche, la similitude de comportement entre d’une part IC30 et le mil sauvage et d’autre part IC27 et HKP, déjà constatée en phase d’installation (essai rhizotron), n’est pas le fait de hasard et mérite plus d’investigations.
-211
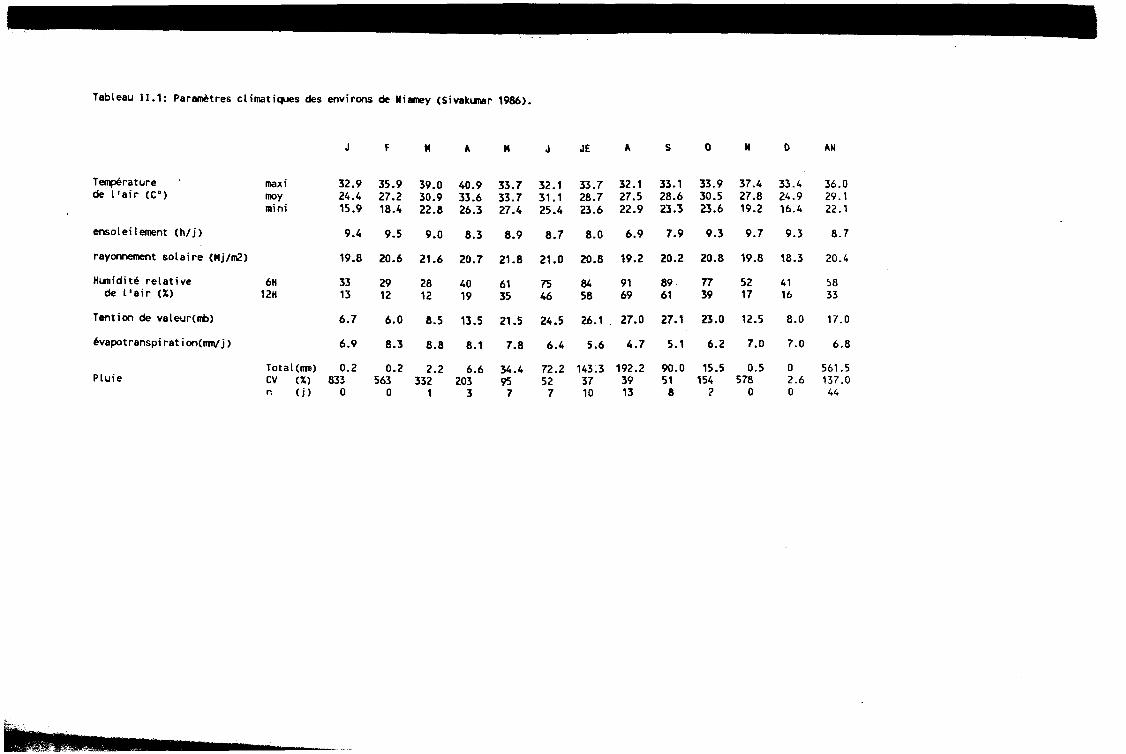
Tableau 11.1: Partitres climatiques des environs de Nicuney (Sivakuaar 1986).
J f n A II J JE A s 0 N D AN
35.9 39.0 40.9 33.7 32.1 33.7 32.1 33.1 33.9 37.4 33.4 36.0 27.2 30.9 33.6 33.7 31.1 28.7 27.5 28.6 30.5 27.8 24.9 29.1 18.4 22.8 26.3 27.4 25.4 23.6 22.9 23.3 23.6 19.2 16.4 22.1
9.5 9.0 8.3
20.7
40 19
13.5
8.1
8.9 8.7
21.0
75 46
24.5
6.4
8.0 6.9 7.9 9.3 9.7 9.3
19.8 20.6 21.6 21.8 20.8 19.8 18.3
6H 33 29 28 12H 13 12 12
61 35
20.8 19.2 20.2
84 91 89 58 69 61
77 52 41 39 17 16
6.7 6.0 8.5 21.5 26.1 . 27.0 27.1
5.6 4.7 5.1
143.3 192.2 90.0 37 39 51
23.0 12.5 8.0
6.9 8.3 8.8 7.8 6.2 7.0 7.0
Total(m) 0.2 0.2 2.2 6.6 34.4 R.2 cv C?i) 833 563 332 203 95 52
15.5 0.5 D 154 578 2.6
8.7
20.4
58 33
17.0
6.8
561.5 137.0
Température * de L'air (CD)
maxi 32.9 MY 24.4 mini 15.9
9.4 ensoleilement (h/j)
raycxvwnent solaire (Mj/m2)
Hunidite relative de l'air (X)
Tention de valeur(&)
6vapotranspiration(mn/j)
Pluie n (j) 0 0 1 3 7 7 10 13 8 ? 0 0 44

Tableau 11.2: parametres climatiques pour les annkes d'essai 1988-1989 et 1990.
1909 1988
J F M A M J JE AT s 0 N D AN
Teapérature maxi 30.0 32.0 32.0 42.0 41.0 39.4 35.6 32.0 35.0 36.8 36.0 31.2 mini 13.0 17.0 20.5 22.0 27.6 27.5 25.0 23.5 24.0 19.8 16.5 19.4
Ensoleillement (heures/jour) 9.7 8.5 9.3 10.3 10.8 8.0 6.6 7.6 8.3 9.8 9.2 9.4 Rayonnement (J/cm2) Nunidite relative 6h 38.0 30.9 40.8 30.0 51.3 59.9 74.7 87.4 03.6 59.8 66.6 42.9
12h 13.0 12.0 13.0 13.6 26.0 38.0 47.0 66.0 55.0 26.0 17.5 14.4 ETP 4.0 4.8 6.1 6.3 8.5 6.9 6.0 5.0 5.0 4.2 4.0 Pluie 0.0 0.0 0.0 1.2 1.2 25.4 119.5
31::: 152.6 0.0 0.0 0.0
I
1990
J F M A M J JE AT S 0 N D
Température msxi 32.6 33.1 36.4 41.3 40.6 38.4 32.6 31.4 35.4 38.4 38.0 35.2 mini 16.6 15.9 18.8 25.8 28.8 27.4 24.2 27.2 25.0 21.9 17.4 17.2
Ensoleillement (heures/jour) 9.5 8.7 Rayonnement (J/cm2) 17.0 18.9
21-g 8.4 9.9 9.5 7.40 9.7 8.3 9.9 10.0 8.9 19.8 16.2
Hunidite relative 6h 49.5 42.1 31:4 47.7 59.5 65.3 62.4 80.8 78.1 68.9 56.2 52.6 l2h 15.3 12.8 8.3 21.5 35.6 40.8 62.5 56.1 52.1 27.6 15.6 15.6
ETP 4.3 4.6 6.6 J.3 7.4 6.7 5.0 5.6 8;::
5.1 4.3 4.1 Pluie 0 0 0 0 75.7 89.9 113.2 113 0 0 0
I AN
-- 36.1 22.2
9.2
57.9 30.3
5.4 474.2

Tableau 11.3: caracteristiques physico-chimiques du sol- Parcelle de Llessai de contre-saison. Les sites 1, 4, 9 et 12 sont distants d'environ 10 a 30 m (voir plan parcellaire fig.1).
graru.il&trie (X) hunidite pH C Y PlOOO PI000
C/M
horizon argile kquiv.
Limon limon sable fin gros. fin
ékaents échangeables CEC me/lOOg lae/1009
Ca Na Mg K
O-40 0.7 1.1 1.6 20.3 40-150 1.7 1.5 0.9 29.4
140-250 5.8 0.4 1.1 24.0 240-260 3.6 0.9 1.3 21.0
sable matiere gros. org.
76.3 0.21 66.5 0.05 68.7 0.02 73.2 a.02
P
1.7 6.2 1.2 0.10 12 2.1 2-o 5.4 0.3 0.06 5 2.9 4.3 5.8 0.1 0.06 1.7 2.8 3.2 6.0 0.1 0.05 2.0 0.9
0.6 0.013 0.17 0.13 1.2 O-6 0.015 0.16 0.074 1.3 1.2 0.02 0.58 0.185 2.1 1.0 0.027 0.45 0.20 1.6
O-50 2.5 0.7 1.3 28.5 67.0 0.07 50-100 8.2 0.9 1.3 30.6 59.0 0.10
100-180 8.7 1.8 2.4 7.2 39.9 0.07 WI-240 13.4 2.4 3.1 53.0 28.1 0.10 240-260 7.1 1.4 2.6 62.3 26.6 0.02
7:: 5.9
10.0 6.0
6.1 0.7 0.08 8.7 2.4 0.9 0.047 0.32 0.157 1.6 5.7 0.6 0.09 6.7 9.2 2.4 0.033 0.68 0.168 3.0 6.1 0.4 0.07 5.7 4.7 2.4 0.033 0.61 0.089 3.0 6.4 0.6 0.09 6.7 1.5 3.5 0.046 1.82 0.159 4.8 6.9 0.1 0.06 1.7 0.5 2.1 0.031 1.08 0.119 2.9
O-50 50-120
120-200 200-250
1.7 5.1 4.9 2.7
0.5 1.2 21.6 75.0 0.12 0.8 1.0 16.4 76.7 0.10 0.2 0.5 14.5 79.9 0.07 0.5 0.0 11.9 84.9 0.02
::"7 3.8 2.3
6.1 0.7 0.08 8.7 3.3 5.8 0.6 0.09 6.7 2.7 5.4 0.4 0.08 5.0 3.3 5.3 0.1 0.03 3.3 1.7
0.4 0.8
0.011 0.12 0.076 1.0 0.012 0.39 0.149 1.7 0.017 o-39 0.110 1.8 0.015 0.21 0.096 1.3 0.8
O-50 1.8 1.6 0.0 21.7 74.9 0.17 2.1 5.6 1.0 0.11 9.1 4.1 50-150 4.1 0.7 0.8 21.5 72.9 0.07 3.4 6.3 0.4 0.07 5.7 9.4
150-200 5.8 0.6 1.8 24.8 67.0 0.17 4.7 6.6 1.0 0.08 12.5 10.9 200-250 3.1 0.3 1.1 20.9 74.6 0.07 2.9 7.0 0.4 0.03 13.3 4.6
0.9 0.018 0.26 0.151 1.5 1.4 0.018 0.44 0.15 l-4 1.5 0.023 0.72 0.297 2.3 0.9 0.017 0.22 0.454 1.3
site 1
site 4
site 9
site 12
i NB: carbon organique méthode ANNE;
azote methode KJELDAHL; capacité d'échange méthode de IIETSCN

-l--.-
-r--
----
1.--
----
----
----
--.
: l--
--
-.--

c 0 0
0 c z 9,
u 5 v)
2 a C
Un
c D mm
w
u1 0 m
Var
. sto
ck (
mm
.j-1)
)
+ N
cdP
m6,
~ I
r - 1
i...
......
......
......
......
. i
-:
......
......
......
.....
_:
j- -
......
......
......
......
i
r !A .‘T

Fa- n++Diminution journalière de stock O-50 cm
l *- .
E
g
5 0
5 I
‘m >
2.5
2
1.5
1 !
-.- .
60 65 70 ?5 SO 85 90 95 100 105 110
Jours après émergence (JAE)
[7 10 q 20 il 40 ) JADS /

Var
. sto
ck (
mm
&1)
‘PP
P
d-LA
&
. .
. .
.i
;ii
i .
. .
. . .
. . .
. .
!, 1 5 :. . .
. . . . . .:
i
r..
. . .
. .
. . . .
I-T- . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ..:
: . .
j.

~;px: figure : biomasse aérienne 88-89 7000-
6000-
5000-
4OOD-
3000-
zooo-
L ooo-
O- 55 60 65 70 75 80 85 90 !
jours après émergence

cl 0
~3U&J~Ul~ sqde sinol . Ç6 06 Ç8 08 C;L OL 99 09 çç
0
2
.v
.9
.8
-0 1
31
471
-9 1

gp figure : biomasse aérienne 88-89 70
60 - . . .
50 - . .
40 -
30 -
20 l- -.
10 l- ..
o-. I--
65 70 75 85 90 95
jours après émergence

III. ESSAI DE CONTRE SAISON CHAUDE (FEVRIER-MAI 1990)
Résumé :
Le comportement du système racinaire face à un stress hydrique post-floral est étudié pour 3 cultivars de mil (Pennketum glaucum) et un mil sauvage en plein champ en saison seche et chaude. II est mis en évidence une évolution du systeme racinaire apres la floraison qui, dans le cas du traitement stress6 peut aller jusqu’à un accroissement de biomasse de 53% par rapport à la floraison. Les variétés ayant montré des capacités opposées à maintenir le rendement face aux conditions hydriques déficitaires présentent des différences de structure racinaire des la floraison en conditions normales, différences qui s’accentuent avec le déficit hydrique.
- 22 -

INTRODUCTION
Pour les varietés cultivées (IC30, IC27 et HKP), nous avons analysé la structure du système racinaire à la floraison (biomasse totale et distribution), sa dynamique en conditions d’alimentation hydrique déficitaire, ainsi que la quantité d’eau totale extraite par chacun des trois cultivars au cours du stress hydrique.
1. MATERIEL ET METHODES
L’essai est réalisé sur la même parcelle, qu’en contre-saison froide 88-89. Pour améliorer la précision statistique le nombre de répétitions est porté à 6 pour les paramètres relatifs à la biomasse aérienne, à 4 pour les racines et à 8 pour ceux du bilan hydrique. Plus particulièrement pour les racines, un troisiéme plan de carottage (plan diagonal) a étC introduit pour mieux cerner la distribution autour du poquet. Pour caractériser l’extraction de l’eau et l’avancement des racines deux positions par rapport au poquet ont été choisies (20 et 50 cm du poquet).
2. RESULTATS
2.1. Caractbrisation de la parcelle au moment de l’application du stress hydrique
2.1.1. L& stocks hydriques de depart
11 s’agit des stocks hydriques calculés entre 20 et 200 cm de profondeur. Les mesures de surface. entachées d’erreur n’ont pas été prises en considération. Cependant, les horizons de surface (0 - 30 cm) sont tr& homogènes sur tout le champ et a priori ne doivent pas gêner une Ctude comparative. Deux remarques essentielles se dégagent du tableau 111.1:
- il y a une bonne uniformite entre les tubes à 20 et à SO cm du poquet au niveau d’une même parcelle de bilan, malgré le mode d’irrigation utilisé (goutte-à-goutte). En effet, les bulbes d’humectation se sont rejoints à la floraison, avant l’arrêt de l’irrigation.
- il y a une hétérogéneité considérable des stocks hydriques entre les parcelles élémentaires allant de 114 à 187 mm, ce qui justifie la nécessité de plusieurs répétitions. Mais les moyennes générales de ces stocks initiaux ne diffèrent pas significativement entre les cultivars.
- 23 -

Tableau 111.1. Stocks hydriques (IIBTI) entre 20 et 200 cm avant ('srrët de l'irrigation.
IC30 1C27 HKP r+?tition -
1 178 135 186 II 147 151
20 cm du poquet III 151 147 127 IV 139 157 187
moyenne f 6cart-type 139 f 12 146 f 9 163 f 29
1 114 127 155 Il 137 144 139
50 cm du poquet III 158 146 125 IV 144 149 187
moyenne f &xrt-type 138 f 18 144 f 13 151 f 27
moyenne g6nérale f 6cart-type 139 f 15 145 f 10 157 î 25
2.1.2. La biomasse aMenne B la floraison
Statistiquement il n’apparaît aucune différence significative entre variétés. Cependant, on note un développement végétatif (limbes verts, tiges et graines, matière stche totale) en moyenne plus important pour IC27, tandis qu’JC30 montre un poids d’épis légèrement supérieur à celui des 2 autres cultivars. Il faut noter que, comparé aux autres cultivars, IC30 a une avance de 3 jours dans le développement végétatif de ses brins-maîtres.
Tableau 111.2: biomssse drienne B La floraison (moyenne et Ccsrt-type)
1c30 IC27 HKP
Limbes Verts 39 t 12 47 f 10 38 t 13 liges * Gaines 82 i 16 115 f 41 93 f 25 Epis 28 î 10 22 t 10 15î 6 Mati6re Sche totale 158 t 30 201 f 64 159 f 42
2.1.3. La biomasse racinaire a la floraison
Le tableau des biomasses racinaires à la floraison (tableau III 3) montre également une nette supériorité du cultivar IC27 (significative au seuil de 1% entre O-40 et 0 - 200 cm de profondeur).
- 24 -

Tableau 111.3 :' Biomasse racirwire B la floraison (g/m2) Onoyenne î écart-type)
-l
Prof (cm) IC30 IC27 HKP
O-40 40 î 11 70 f 14 48 î 8 40-200 61 2 18î 6 15 f 5
O-200 46 t 12 88 i 13 63 f 10 I l
Tableau III.4 : Répartition de la biomasse racinaire dans le profil (% de la biomasse racinaire totale)
Prof (cm) If.30 IC27 HKP
O-40 87 79 76 , 40-200 13 21 24
En terme de distribution verticale, les cultivars IC27 et HKP colonisent le sol plus en profondeur que IC30 qui reste superficiel. Une observation similaire a déjà été faite en rhizotron, en a phase d’instahation des plantes.
Ainsi, avant l’application du stress hydrique (60 JAE), le cultivar IC27 montre un développement végétatif plus important que les 2 autres cultivars, tant au niveau aérien qu’au niveau racinaire. Cependant, en ce qui concerne le poids des épis, le cultivar IC30 montre une supériorité, probablement à mettre en relation avec les 3 jours d’avance qu’il avait dans son développement vegétatif, et qui correspondent à 3 jours de remplissage des épis.
2.2. Evolution des stocks en eau du sol et des param&res tiorphologiques des plantes au cours de la contrainte hydrique.
2.2.1. LVvolution des stocks hydriques
Les variations de stock hydrique ont été mesurées après 12 jours de contrainte. Le tableau III5 donne les valeurs cumulées entre 20 et 200 cm de profondeur mesurées à 20 et 50 cm de distance du poquet.
On constate que les variations de stock sont comparables lu 20 et 50 cm du poquet pour une méme cultivar. Les différences entre cultivars ne sont pas significatives mais IC27 a la plus forte valeur absolue.
- 25 -

Tableau 111.5 : Variation de stock (mn) au cours de ta contrainte hydrique, sur 20 - 200 cm de profondeur. W: moyenne des mesures B 20 et 50 cm de distance)
En figure III.1 est présentée la distribution verticale de la variation moyenne de stock pour chacun des trois cultivars. Dans les horizons intermédiaires (80 - 140 cm) elle est plus importante sous le cultivar IC27.
2.2.2. LVvolution de la partie akienne au cours de la contrainte hydrique (62 A 77 JAE)
L’évolution de la biomasse aérienne entre 62 et 77 JAE se caractérise par une réduction de la matière sèche aérienne totale, et notamment une forte diminution du poids de limbes verts, correspondant à une sénescence foliaire. Seules les épis maintiennent leur croissance, déjà bien amorcée pour IC30 avant la contrainte hydrique.
2.2.3. Evolution de la partie akienne apres la réirrigation (77 & 92 JAE)
La reirrigation intervenue à 77 JAE (après 15 jours de contrainte) se traduit globalement par une augmentation de la biomasse aerienne totale (tableau 111.6). Cette augmentation est le fait, principalement du maintien de la croissance des épis et dans le cas d’IC30, de la reprise de croissance des tiges et des gaines.
2.3. Caractkisation de la parcelle à la récolte
2.3.1 La biomasse akienne A la rkolte
La structure de la biomas$e aérienne à la récolte est résumée dans le tableau 11.7. On remarque que IC30 a pu rattraper IC27 sur le plan du développenient végétatif, grâce à la reprise de croissance après la réirrigation. Le cultivar IC27, à la structure plus développée (limbes verts, épis et tiges + gaines) a produit plus de grains que HKP.
- 26 -

Tableau II i.7 : Biomasse aerienne à ta récolte (moyennes et karts types en g/ni2)
1 IC30 IC27 HKP
Li*s Verts 13ï 2 16 t 11 12* 5 Epis 64 f 22 51 f 13 40 f 11 Tiges + Gaines 91 t 14 85 f 28 77i8 Matiere Sche totale 213 f 26 195 f 55 177 î 25 Rendement Grains 2Oi 6 17i 5 13î 3
2.3.2. La biomasse racinaire A la dcolte
La situation de la biomasse racinaire en fin de cycle est résumée dans le tableau 111.8. IC30 a la masse de racines la plus importante; 56 contre 40 g/m2 pour ,les deux autres cultivars. Ceci correspond à une accroissement de 21% par rapport à la floraison pour IC30 et une baisse de -53 et -36% respectivement pour IC27 et HKP.
Tableau III.8 : Biomsse racinaire iJ fa récolte et variation par rapport & Le floraison (moyenne et kart type en g/m2)
!iElii-...,....,,
3. DISCUSSION - CONCLUSION
Les résultats de l’essai 1990 sont résumés dans le tableau III.9. Le cultivar IC30 qui avait une avance de 3 jours sur le cycle de développement a montre la plus forte production de grain à maturité, malgré un système racinaire initialement moins développé que ceux de IC27 et HKP. Cette situation illustre comment, pour le mil aussi, l’interaction entre le stade phenologique et la structure végétative peut modifier l’impact de la sécheresse sur le rendement final (Robelin, 1963; Robins et al., 1953).
Par ailleurs, au moment de l’application du stress hydrique, les cultivars IC27 et HKP étaient au même stade phénologique mais avec des développements vegétatifs différents (tableau 111.9) (matière sèche aérienne, matière sèche racinaire). Grâce à un système racinaire initialement plus développé, exploitant mieux les réserves en profondeur, IC27 a pu maintenir un poids de limbes verts plus important durant toute la période de déficit hydrique. Conséquemment, il fini avec une masse d’épis et un rendement final superieurs.
-27-

Tableau III.9 : Rkepitulstif des dsuitats (!!Se: Mstiere Sèche s&ienne; + : faibLe'++ moyen +++ fort).
pilzz&r2j ;;- 7
1 Rerxkmnt Grein +++ ++ + Racines p1 la r&olte +++ ++ l
Ainsi, la structure Initiale la plus développée, tant de point de vue de la biomasse aérienne que de la biomasse racinaire, semble pouvoir résister mieux à un déficit hydrique post-floral notamment par le maintien d’une surface photosynthétique plus importante, soutenue par une meilleure exploitation des reserves hydriques de profondeur.
Ces résultats, statistiquement peu significatifs dans la plupart des cas, méritent être confirmés à travers une amélioration de la precision dans les essais futurs. En revanche, la logique cohérente de classement des cultivars trouvée à tous les niveaux (système racinaire. e,xploitation de réserve de sol, maintien de la surface foliaire verte, croissance des épis, etc.) n’est pas le fait du hasard et mérite plus d’investigation. D’autre part, une diminution globale de biomasse aérienne a été enregistrée au cours du stress. Une hypothése envisageable est celle d’un détournement de matiere séche vers les racines, hypothèse dejà émise en contre- saison froide 1988. Cependant, l’essai de 1990 a montré chez IC30 un maintien et chez IC27 et HKP une perte significative de biomasse racinaire au cours des 30 jours séparant floraison et récolte. Ici, il faut préciser que la contrainte hydrique a duré 15 jours, après quoi l’irrigation a et& rétablie sur les 15 jours suivants. Ainsi la structure du systeme racinaire à la recolte intégre-t-elle les effets cumulés d’une sécheresse et d’une réhumectation. Si les effets dr l’alternance dessication- réhumectation sur les racines, ne sont pas encore bien connus, l’action de rhizogenèse suivant une réhumectation a été plusieurs fois soulignée (Vartanian 1981; Picard 1973). Ainsi, dans notre cas, nous mesurons l’influence d’un stress hydrique post-floral de durée limitée, sur la structure finale de la plante, ce qui est d’ailleurs plus conforme à la réalité. Mais pour la réaction même du système racinaire à la contrainte hydrique, elle doit être mesurée de maniére dynamique au cours du stress hydrique, en relation avec la partie aérienne. Ceci doit être pris en compte pour les essais futurs.
.

CONCLUSION GENERALE
k trois essais (rhizotron, contre-saison froide et contre-saison chaude) . nous ont permis de dégager un certain nombre de points importants concernant la
réaction du mil face aux stress hydriques de début et de fin de cycle et de formuler des hypothèses.
En phase d’installation (germination-montaison), le stress hydrique stimule une extension verticale du systéme racinaire avec une colonisation plus importante en profondeur. Aucune différence variétale n’apparaît dans cette réponse au déficit hydrique. En revanche, en situation d’alimentation hydrique satisfaisante, certains cultivars (IC27) colonisent le sol plus profondément que d’autres (Hw, imo) durant la phase d’installation des cultures. Ceci pourrait conditionner leur comportement face à un déficit hydrique plus tardif.
En phase de remplissage des grains, l’essai de contre-saison 88-89 montre que dans un premier temps le cultivar qui supporte le mieux le stress hydrique est celui qui a la structure initiale la plus développée. En raison d’une consommation en eau importante, soutenue par l’exploitation des horisons profonds, et d’une surface foliaire verte (sources d’assimilats carbonés) supérieure, il maintient mieux la croissance des épis dans les 25 premiers jours. Ensuite, quand le déficit hydrique se prolonge, la faiblesse de la réserve en eau disponible pénalise fortement le rendement final. Cependant, l’essai de contre-saison 1990 révèle aussi un effet déterminant du stade phénologique atteint par la plante lors du déclenchement du déficit hydrique. Indépendamment de leur biomasse aérienne, les plantes ayant le développement phénologique le plus avancé au début du stress sont celles qui maintiennent la plus forte production de grains.
Il existe donc une relation entre la capacité de la plante à maintenir le rendement et celle de son système racinaire à exploiter les horizons profonds lors de la sécheresse. L’extension du systéme racinaire semble être liée à la structure de la plante avant le stress et notamment à l’importance de la surface foliaire verte. Ceci implique des transferts de matiere sèche entre parties aériennes et racines. Or, même en situation de déficit hydrique prolongé (contre-saison 88-89), on constate que l’extraction de l’eau par les racines est loin d’être complète en deçà de 150 cm de profondeur. L.a croissance des racines en réponse à la sécheresse a donc une limite dans l’espace (capacité de colonisation du sol en profondeur) et dans le temps. Ce dernier point reste particulièrement obscur: il pose le problème de la coordination du développement des différents organes de la plante et de l’interaction entre phénologie et réponse au déficit hydrique.
- 29 -

REFERENCES BIBLIOGRAPHIQUES
ANONYME, 1990. Etude agropbysiologique des mkanismes de rCsistance du mil à la sécheresse. Rapport Annuel 1990, Jnstitut des Radio-Jsotopes, Université de Niamey, 14 p.
BATCHO, E., DAOUDA, O.S., DO, F., ANNEROSE, DJ.M., KHALFAOUI, J.L., FOFANA, A., LAFFRAY, D., LOUGUET, P. 1990. Etude de la croissance racinaire de six cultivars de mil (Pennisetum americanum L.). Revue du Réseau pour I’Amkhoration de la Produch’vité Agricole en Milieu Aride, 2: 51-65.
BIDINGER, FR., MAHALAKSHMJ, V. and RAO, G.D.P., 1987. Assessment of drought resistance in Pearl millet. IJ. Estimation of genotype response to stress. Aust. J. Agric. Res., 38: 49-59.
CHOPART, J.L., 1980. Etude au champ des systémes racinaires des principales cultures pluviales au Stnégal. These de Doctorat, Institut National Polytechnique de Toulouse, 148 p.
CIARKE, J.M., and TOWNLEY-SMITH, T.F. 1984. Screening and selection techniques for improving drought resistance. In: Crop Breeding, a Contemporary Basis. Vose, P.B. and Blixt, S.G. (eds). Pergamon Press, Oxford, UK pp 137-162.
DANCETTE, C., 1983. Estimation des besoins en eau des principales cultures pluviales en zones soudano-sahtliennes. Agron. Trop. 38,4: 281-293.
DIAS DA SILVA, J.C., NUNES DE PINHO, J.L., LAFFRAY, D. et LOUGUET, P., 1990. Effet d’une contrainte hydrique controk sur divers paramètres hydriques et physiologiques de cultivars de mil et sorgho sensibles et résistants a la sécheresse. Rev. Rh. Amklior. Product. Agri. Milieu Atide, 2: 79-92.
GREGORY, PJ., 1989. The role of root characteristics in moderating the effects of drought. In: Drougth Resistance in Cereals. BAKER, F.WJ. (ed), Wallingford. pp 141-150.
4 GREGORY, PJ., SHEPHERD, K-D. and COOPER, PJ.M., 1984. Effects of fertilizer on root growth and water use of barley in Northern Syria. J: A@. SC., 103: 429-438.
ij
1 HURD, Eh., 1974. Phenotype and drougth tolerance in wheat. Agit. Meteor., 14: 39-55.
f JONES, J.W. and ZUR, B., 1984. Similation of possible adaptative mechanisms in crops subjected to water stress. hfgation Science, 5: 251-264.
1 I
JORDAN, W.R., MILLER, F.R. and MORRIS, DE., 1979. Genetic variation in root and shoot growth of sorghum in hydroponics. Crop Sciertce, 19: 468-472.
JORDAN, W.R., DOUGAS, WA. and SHOUSE, PJ., 1983. Strategies and trop improvement for drougth-prone regions. 111: Plant Production and Management Under Drougth Conditions. Stone, J.F. and Willis, W. (eds). Elseviers, Amsterdam, Netherlands. pp 251-264
LEVJTT, J., 1980. Drought avoidance. //r: Responses of Plants to Environmental Stress. Lelitt, J. (ed). Physiological Ecology. II: 93-128.
LUDLOW, M.M. and MUCHOW, R.C., 1988. Criticai evaluation of the possibilities for modifying crops for hight production per unit of prccipitation. bt: Drought Researcb Priorities for the Dryland Trop&. Bidinger, F.R. and Johansen, C (eds). ICRISAT, Patancheru, India. pp 179-211.

MATHEWS, R.B., HARRIS, D., NAGESWARA RAO, R.C., WILIAMS, J.S. and WADIA, K.D.R. 1988. Tbe physiological basis for yield differences between four genotypes of groundnut (Amchis
hypogea) in response to drought. 1. Dry matter production and water use. Exp. Agri., 24: 191-202.
MORGAN, J.M. 1988. The use of coleoptile response to water stress to differentiate wheat genotypes for osmoregulation, growth and yield.Ann. Bof., 62: 193-198.
MUCHOW, R.C. and ‘SINCLAIR, T.R. 1986. Water and nitrogen limitations in soybean grain production. II. Field and mode1 analyses. Field Crops Res., 15: 143-156.
PASSIOURA, J.P. 1982. The role of root system characteristics in drought resistance of trop plants. In: Drought Resistance of Crops With Special Emphasis in Rice. International Rice Research Institute (ed), Los Banos, Laguna, Phillipines. pp 71-82.
PASSIOURA, J.P. 1983. Roots and drought resistance. In: Plant Production and Management Under Drought Conditions. Stone, J.F. and Willis, W. (eds). Elsevier, Amsterdam, Netherlands. pp 265-280.
SANI, D.O. , SICOT, M., et MARINI, P. , 1990. Caractérisation de la diversité hydrodynamique d’un sol ferrugineux tropical, differencié sur sable dunaire en milieu soudano-sahélien. Sivakumar, S.V.K., Wallace, J.S., Renard, C. et Giroux, C. (eds), 1991. Soi1 Water Balance in the Soudano-sahélien Zone (Proc. Int. Workshop, Niamey, Niger, february 1991) n”l99: 209-216.
SHRINIWAS and SUBBIAH, B.V. 1976. Root studies of some Bajra hybrids (Pennisetum fyphoides
1
Stapj). J. Nue. Agi. Bio., 5: 15-16
t SIBAND, P., 1983. Essai d’analyse du fonctionnement du mil en zone sahélienne. Thèse de Doctorat,
! Université des Sciences et Techniques du Languedoc, 302 p.
SIVAKUMAR, S.V.K. 1986. Programme Ouest Africain de I’ICRISAT. Rapport Annuel, Centre SahClien de I’ICRISAT, 1986, Niamey, Niger. pp 47-53.
TAYLOR, H.M., BURNE’IT, E. and BOOTH, G.D. 1978. Toproot elongation rate of soybeans. Zeitschnj? fit Acker- und Pfanzenbau, 146: 33-39.
THIERY, J.M., 1985. “VOYONS”, Programme de Simulation Conversationnelle en Physico-chimie et Agronomie, 156-157.
TURNER, N.C. 1986. Crop water del%: a decade of progress. Adv. Agron., 39: 1-51.
TURNER, N.C. and NICHOLAS, M.E. 1987. Drought resistance of wheat for light textural soils in a Mediterranean climate. In: Drought Tolerance in Winter Cereals. Srivastava, J.P., Porceddu, E., Acevedo, E. and Varma, S. (eds), Jones Wiley, New York. pp 203-216.
-31-

Contribution I l'étude des profils racinaires du Mil et méthode de calcul do ln masse racinaire sous le poquet. DECEMBRE 1990
P. SANTUCCI, J, THIERY*, S. DAOUDA OUSMANE, F. DG, P. MARINI.
D&partement de Radio-Agronomie, Institut des Radio-Isotopes, Université de B.P. 10727, NIAMEY, NIGER.
Département de Physiologie Végétale et des EcosystBmes, Centre d'Etudes Nucléaires de Cadarache, 13108, SAINT-PAUL-LEZ-DURANCE, FRANCE.
INTRODUC!J!ION
L'approche du eystème racinaire des végétaux suflrieurs n’est pae ais(5e: sa localisation souterraine dans un milieu dense et opaque, son intimité avec
font que toute dtude de l'architecture racinaire, forcément destructrice, ne peut aboutir qu'a l'elaboration d'un modèle simplifi6 de cette structure spatiale.
L'objet de cet écrit est alors de mettre au point un outil de représentation de l'enracinement d'un végétal, applique ici dans le cas du Mil, et de le prolonger par une méthode de calcul de la masse racinaire dans un volume de sol donné.
MATERIELS BT METHODE
Protocole expérimental et prise de l'information
L'enracinement du Mil, variété ICMVIS 86330, semé à raison d'un paquet par m2 a éte étudie en contre-saison froide sous le controle du facteur alimentation hydrique a deux niveaux: stressé ou irrigué (deeau de goutteure
parallele aux lignes de eemfs).
Pour un poquet (fig.1). exprimée en mg/dm3,
on a détermine la densité volumique de racines a, en sept profondeurs et sur cinq points de carottage:
- points de carottage: . Ocm, sur la plante; . 25 et 50cm sur le pl;~n parallèle a la ligne de
semie, ainsi que sur le plan orthogonal B la ligne de semis;
- profondeurs: . O-20cm donnant une valeur a 1Ocm (le collet n'est pae compté);
. 20-40cm donnant une valeur B 3Ocm;
. 40-60cm donnant une valeur B 5Ocm;
. 60-80cm donnant une valeur a 70~x1;
. 80-100cm donnant une valeur a 9Ocm;
. lOO-120cm donnant une valeur 8 1lOcm;
. 120-140cm donnant une valeur a 130cm;
ce gui donne en tout 35 mesures par poguet qui déterminent deux plans de représentation: chaque plan (parallale ou orthogonal h la ligne de semis) est

defini par la profondeur, la distance au poquet, et la valeur Q. Il est donc possible de donner une représentation de l'enracinement par la cartographie en lignes d'ieodensite a=cte dans le plan considéré du profil (utilisation du module de topographie du logiciel SURFER4* (1990) [l] (méthode du Kriging).
Détermination de la masse racinaire BOUE le poquet
Le principe de calcul. de la masse de racines est lgfnt6gration de la densité volumique sur l'ensemble du poguet.
Soit p la profondeur du profil, i.e. la profondeur maximum d'enracinement, pour laquelle a =O (condition aux limites), inter-poquete:
et eoit dl/2 la demi-dietance
longueur p, En conaidke donc le cylindre de base carree 2dl/2*2dl/2 de
meeure oit en remarquant que la limitation h dl/2 n'eet pae g&nante dans la
il y a, en moyenne, autant de racinee du poquet considerd au-del& de dl/2 que de racines du paquet voisin qui penètrent dans le profil étudi6.
La base carree est subdivi&e en huit octante (fig..2) afin de tenir compte de la dieymétrie due au rdseau d'irrigation (2 plans d'étude), et dventuellement de mesures faites eur la biesectrice; dans le cae où il n'y aurait pas de mesure sur cette bissectrice, i.e. celui gui noue intéreese, on prendra comme valeurs les moyennes dee mesures sur les deux plans orthogonaux.
L'idée est d'eneuite étudier chague octant, de le "replier comme un Bventail" pour projeter une densite eurfacfgue dans un des trois plans de base. Pour obtenir la masse, il suffira d'integrer eur le plan de projection, puie de sommer lee masses de chaque octant.
Octante centres sur la ligne d'irrigation. Soit r le rayon des carottes et soit h leur épaisseur.
Soient 0, A et B les points de carottage, il faut consid6rer 3 tonee concentriques (fig.3) 1, II et III. On poee A=d(O,A) et B=d(O,B). On ee propose de calculer le volume asaoci6 aux secteurs hachurés.
1: le secteur de la carotte 0 est de volume vl=1/8 x r2 h;
II: cachant que l'aire d'une couronne de rayon moyen R, d'épaisseur e est ut [(R+e/2)2-(R-e/2)2)=2r R e, la surface du secteur centrd sur A et d'dpaiseeur 2r est:
s2=l/8 2n A 2r=1/2 r A r,
et le volume v2 du secteur II est:
v2=1/2 TI A r h;
III: de même, v3=l/2 VT B r h par symétrie algèbrique de A et B.
-2-

Si mi est la masse de racines dans chaque volume vi, mi=vi ui (ui densite volmique mmurde), la densite nurfacique u' projetée sur le plan mddian de l'octant est alors égale a la masse du secteur sur sa eection verticale:
1: a1 '=a1 1/6 lr r2 h /(r h)=ol (n r/8);
II! u2'=a2 1/2 R A r h/(2r h)=o2 (n Aj4);
III: a3 *=a3 (n B/4);
On retrouve en fait le résultat gue la densité surfacigue est 6gale à la densité volumique multipliee par la longueur de l'arc moyen, et les coefficient correctifs ti de ai*=di ci sont:
cl: arc moyen de rayon r/2: 1/8 2n r/2=n r/8;
c2: arc moyen de rayon AI 1/8 2n A=n A/4?
c3: arc moyen de rayon B: u B/4;
Pour trouver .la masse totale de racines dans l'octant, $1 faut intégrer sur la surface S du plan vertical conwfderér
moctant" JJ
u'(x,Y) dx dy; s
ie plan étant défini par sa profondeur p et par son rayon R' tel que la eurface de l'octant soit identique a celle du triangle ieoc&le de base définie par la mediatrice e.kre deux poquets voisins:
ff W2/8=d@ tg(2 n/16),
R'=dl,; [8/n tg (~/8))~/~.
Octante centrés eut la ligne orthogonale h la ligne d'irrigation. Puisque la géométrie est la même, lès calculs sont identiques au cas précbdent.
Octante diagonaux. Les calculs sont eimilairee jusqu'au rayon d'intégration R" qu'il faut calculer:
surface(octant diagonal)+surf.(oct.horizontal)Lautf.(guadrant),
eurf.(oct,diag.)=dl/22-dAi22 tg(tr/8)=n Rw2/8,
donc R"=dl/2 [8/n (1-tg(n/8))1/2.
R'/* p Et dam toue les cas %ctant" J J o'(x,Y) ~JC dy obtenue par l'uti-
0 0 lieation d*: module d'intbgration du logiciel SURFER (1990) [2] Sur la
"cartographie" de a' calculée à partir des valeurs a-i, déduites des meeuree ui et des coefficients ci.
-3-

Alors,
Rtotale" 2 nbctant parallele à la ligne de semis +2 'kctant orthogonal h la ligne de semis +4 rrbctant diagonale
RESULTATS BT DISCUSSION
Repiésentation de la densité volumigue
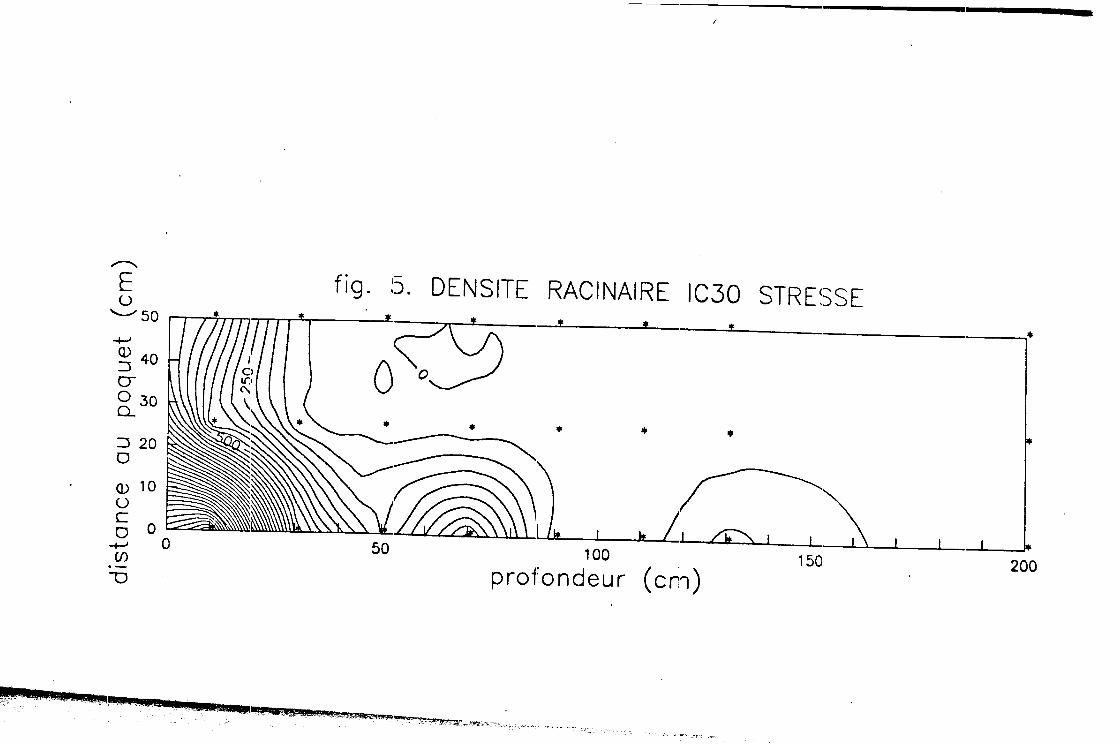
En considérant que o-0 dès que la profondeur dbpasse 2m, on a pu représenter les cartes moyennes d'isodensite pour le Mil irrigué et pour le etressé (fig.4 et 5), ce gui nous permet de constater qu'en conditione de stress le Mil recherche l'eau en profondeur,. alors que eoua irrigation le eyet&me racinaire reste en surface.
Calcul den masses racinaires -----
En prenant p=200cm, dl/2=50cm, r=3.75cm, A=25cm, et B=SOcm, on trouve les coefficients de correction cl=1.47cm, c2=19.64cm et c3=39.27cm, ainsi que les rayons d'intégration R'=51.35cm et R"=61.07cm.
Le tab.1 donne les principaux résultats concernant les masses; ceci permet de constater que le Mil stress& développe, a terme, plus de masse racinaire que le Mil irrigu6. On s'aperçoit auesi qu'il existe une tr&s forte dieytnétrie induite par le réseau d'irrigation: la masse racinaire eet regroupée dans la zone où la disponibilité en eau est la plus forte.'
CONCLUSION
L'approche du eystème racinaire du Mil au travere de ma densité volumique eet possible: cette variable permet d'appréhender non seulement la distribution spatiale (aspect qualitatif), maie aussi de calculer des maeees racinaires aussi bien dans le profil tout entier que dans certains horizons (aspect quantitatif).
REFERENCES BIBL.IOGRAPHIQVES
[l] SURFER4O, 1990. Pages 4-l a 4-102 in SURFERO Version 4, Reference Manual, Golden Software,' Inc., 809 14th zreet, P.O. Box 281, Golden, Colorado 80402-0281, U.S.A..
[2) SURFER4O, 1990. Pages 8-5 à 8-29 in SURFER Version 4, Reference Manual, Golden Software, Inc., 809 14th Street, P.G. Box 281, Golden, Colorado 80402-0281, U.S.A..
-4-

fig. 1. Poquet d’étude i i I
... . . . . . ... . . . . . . . .................................... .................. .... .............. .............. ............. .............. ........... ........... . ........... ........... ........... . . . . . ... ...
point de carottage
r6seau d’irrigation
fig. 2. Diuision de a base carr6e du poquet en huit octants

fig. 3. Etude des trois secteurs sur un octant
carotte de rayon r

dist
ance
ou
po
quet
(c
m)
0 0
8 g
0 ts
; F
4
.4 4 0
dist
ance
au
po
quet
(c
m)
%o
. CO
l %
o-
. CO
7 7 nL 0 3.
n T
4 4

r niveau
du facteur
etresse
irrigué
masse octant masse octant masse octant Maeee parallsle h orthogonal a diagonal Totale
la 1. de eemia la ligne
14.7 g 13.2 g 19.5 g 134 g
14.1 7.6 15.2 104
Tableau 1. Massee racinaires aou8 le poquet.


















