
Rapport de stage Master 2_2014/2015
48
IBEA Institut de Biologie et d’Écologie Appliquée U.C.O. Établissement d’enseignement supérieur privé 3, place André Leroy 49008 Angers Chambre d’Agriculture du Maine et Loire POLE ENVIRONNEMENT 14, avenue Jean Joxé 49100 Angers Etude de la biodiversité fonctionnelle dans des agroécosystèmes du Maine-et-Loire Soutenu par THEBAULT David Rapport de stage Master 2 Ecologie des Ressources Naturelles et Développement Durable (ERNDD) Mars/Août 2014 Nom du maître de stage : Ambroise Bécot Nom du tuteur : Didier Georges
-
Upload
david-thebault -
Category
Environment
-
view
317 -
download
0
Transcript of Rapport de stage Master 2_2014/2015

IBEA Institut de Biologie et d’Écologie Appliquée
U.C.O.
Établissement d’enseignement supérieur privé
3, place André Leroy
49008 Angers
Chambre d’Agriculture du Maine et Loire
POLE ENVIRONNEMENT
14, avenue Jean Joxé 49100 Angers
Etude de la biodiversité fonctionnelle dans des agroécosystèmes du Maine-et-Loire
Soutenu par THEBAULT David
Rapport de stage Master 2
Ecologie des Ressources Naturelles et Développement Durable (ERNDD)
Mars/Août 2014
Nom du maître de stage : Ambroise Bécot
Nom du tuteur : Didier Georges

Ce rapport a été rédigé en Century Gothic, taille 10, qui économise 30% d’encre à
l’impression par rapport à une police classique


CHARTE DE NON PLAGIAT
Protection de la propriété intellectuelle
Tout travail universitaire doit être réalisé dans le respect intégral de la propriété intellectuelle
d’autrui. Pour tout travail personnel, ou collectif, pour lequel le candidat est autorisé à utiliser
des documents (textes, images, musiques, films etc.), celui-ci devra très précisément signaler
le crédit (référence complète du texte cité, de l’image ou de la bande-son utilisés, sources
internet incluses) à la fois dans le corps du texte et dans la bibliographie. Il est précisé que
l’UCO dispose d’un logiciel anti-plagiat dans dokeos.uco.fr, aussi est-il demandé à tout
étudiant de remettre à ses enseignants un double de ses travaux lourds sur support
informatique.
Cf. « Prévention des fraudes à l’attention des étudiants »
Je soussigné(e), ……………………………………………………., étudiant(e) en
…………………………………………………
m’engage à respecter cette charte.
Fait à ……………………………………..……………, le……………………………………..
Signature :

Remerciements
Tous mes remerciements à Monsieur Ambroise Bécot, chargé de mission biodiversité
au pôle territoire et développement durable de la chambre d’agriculture du Maine-et-Loire
et à Madame Virginie Guichard, responsable de ce pôle pour l’intérêt et la confiance portés
à mon égard durant ces six mois de formation.
Je remercie tout particulièrement Monsieur Julien Pétillon, enseignant-chercheur à
Rennes 1, pour ses précieux conseils et m’avoir accueilli dans ses locaux lors de
l’identification et l’analyse statistique des araignées.
Je remercie également, tous les agriculteurs du réseau ARBRE qui m’ont accueilli,
accompagné et donné de leurs temps pour me transmettre leurs conseils, expériences et
connaissances, mais aussi l’ensemble du personnel de la chambre d’agriculture pour leur
accueil chaleureux et sympathique.
Je remercie mon tuteur, Monsieur Didier Georges, pour ses conseils quant à
l’organisation de ce rapport.
Merci enfin aux étudiants en Master 2 de Rennes, pour leur bonne humeur sans faille,
leur soutien et leur sympathie.
À Ambroise Bécot, Marion et Angélique, merci pour vos multiples relectures et conseils
pour la rédaction de ce rapport.

Table des matières
Introduction ................................................................................................ 1
Présentation de la structure ..................................................................... 4
1. Organisation ......................................................................................... 4
2. Une répartition territoriale des missions ........................................... 4
Chapitre 1 : Observatoire Agricole de la Biodiversité (OAB) ............ 5
1. Matériel et méthode .......................................................................... 5
2. Résultats ................................................................................................ 6
3. Discussion et Conclusion .................................................................... 7
Chapitre 2 : Etude des communautés d’araignées dans des
agroécosystèmes ...................................................................................... 9
1. Matériel et méthode .......................................................................... 9
1.1 Sites d’études et choix des parcelles ............................................................. 9
1.2 Dispositif expérimental .................................................................................... 10 1.2.1 Période ........................................................................................................................................... 10 1.2.2 Pièges barbers .............................................................................................................................. 11
1.3 Données locales et paysagères ................................................................... 12
1.4 Méthode d’identification .............................................................................. 13
1.5 Base de données ............................................................................................ 13
1.6 Analyses des données .................................................................................... 13 1.6.1 Présentation générale des prélèvements ............................................................................... 13 1.6.2 Comparaison des communautés d’araignées en fonction du type de pratique
agricole (conventionnelle / biologique) .............................................................................................. 14 1.6.3 Influence des variables paysagères sur les communautés d’araignées .......................... 16

2
THEBAULT David
2. Résultats : ............................................................................................ 16
2.1 Présentation générale des prélèvements ................................................... 16
2.2 Comparaison des communautés d’araignées en fonction du type de
pratique agricole ....................................................................................................... 17 2.2.1 Comparaison des assemblages de communautés d’araignées ...................................... 17 2.2.2 Comparaison de l’abondance et de la richesse spécifique des communautés
d’araignées ................................................................................................................................................ 21
2.3 Influence des variables paysagères sur les communautés d’araignées 25
3. Discussion ............................................................................................ 28
3.1 Présentation générale des prélèvements ................................................... 28
3.2 Comparaison des communautés d’araignées en fonction du type de
pratique agricole (conventionnelle / biologique) ................................................ 29 3.2.1 Comparaison des assemblages de communautés d’araignées ...................................... 29 3.2.2 Comparaison de l’abondance et de la richesse spécifique des communautés
d’araignées ................................................................................................................................................ 29
3.3 Influence des variables paysagères sur les communautés d’araignées 31
4. Conclusion .......................................................................................... 33
Bibliographie ............................................................................................. 34
Sites web consultés .................................................................................................... 35
Annexes ....................................................................................................... A
Annexe 1 : Descriptions des quatre protocoles de l’OAB .................................... A
Annexe 2 : Liste des 86 espèces identifiées dans l’étude ...................................... E

1
THEBAULT David
Introduction
Il existe de nombreuses définitions de la diversité biologique ou biodiversité. La
définition la plus communément admise, est : « la variabilité des organismes vivants de toute
origine y compris, entre autres, les écosystèmes terrestres, marins et autres écosystèmes
aquatiques et les complexes écologiques dont ils font partie : cela comprend la diversité au
sein des espèces et entre espèces ainsi que celle des écosystèmes ». Elle s’exprime par la
diversité génétique, la diversité des espèces et la diversité des écosystèmes. C'est aussi la
position choisie à l’issue de la Convention sur la diversité biologique (CDB), établie dans le
cadre du sommet planétaire sur l’environnement et le développement, à Rio de Janeiro en
1992. La Convention sur la diversité biologique (CDB) est le premier texte de droit
international définissant la diversité biologique et la reconnaissant comme une valeur que les
États doivent protéger. La France a ratifié cette dernière le 7 juillet 1994. Depuis le sommet
planétaire de Rio de Janeiro (1992), la biodiversité est devenue un objectif politique majeur.
De nombreux autres engagements internationaux ont suivi ce sommet. En Europe, on peut
citer la directive Oiseaux, la directive Habitats, le réseau Natura 2000, et la Directive cadre sur
l’eau.
De fait, si la "biodiversité" a d’abord paru privilégier les grands écosystèmes naturels et
la nature "cathédrale", sous l’effet d’une médiatisation, mais aussi d’une mobilisation réussie
notamment de grandes ONG telles que l’UICN, le World Wide Fund (WWF) ou « Conservation
International », l’intérêt des écosystèmes semi-naturels et de leurs biodiversités ordinaires, n’en
est pas moins primordial. La biodiversité dite ordinaire, domestique et sauvage, joue un rôle
essentiel dans les écosystèmes. Nous la côtoyons constamment en milieu urbain dans les
jardins, les bosquets, dans les espaces laissés en friche, ainsi qu’en milieu agricole.
En milieu agricole, trois grandes catégories de biodiversité ordinaire peuvent être
définies en fonction de leurs rôles au sein de l’agroécosystème (Peeters A., 2004). Il y a la
biodiversité domestique choisie par l’agriculteur, la biodiversité para-agricole (ou biodiversité
sauvage fonctionnelle) ayant un grand rôle dans le fonctionnement de l’agroécosystème, et
la biodiversité extra-agricole (ou biodiversité sauvage spontanée) ayant un faible rôle – ou
supposé comme tel - dans le fonctionnement de l’agroécosystème. La biodiversité sauvage
fonctionnelle joue un rôle particulièrement important dans le fonctionnement de
l’agroécosystème. Cette biodiversité peut-être parfois problématique avec notamment les
espèces ravageuses de cultures et « les mauvaises herbes », mais est aussi et surtout source
d’un grand nombre de services écosystémiques (auxiliaire de culture, pollinisation, fertilisation
du sol, etc.).
Les rapports entre agriculture et biodiversité sont complexes. En effet, l’agriculture
participe à la fois à favoriser la biodiversité (sélection de races et de variétés, création de
structures paysagères constituant des habitats particuliers, etc.), mais participe aussi à sa
réduction.
L'influence de l'homme sur les écosystèmes est omniprésente (Foley et al., 2005 in
Prieto-Benitez & Méndez, 2010). Les humains ont transformé ou dégradé 39 à 50 % de la
surface de la Terre (Vitousek et al., 1997 in Prieto-Benitez & Méndez, 2010). La destruction des
forêts, la gestion et le remplacement par des plantations ont été particulièrement intenses
ces 300 dernières années (Foley et al., 2005 in Prieto-Benitez & Méndez, 2010), d'abord dans
les zones tempérées et plus récemment dans les tropiques (Lewis, 2006 in Prieto-Benitez &
Méndez, 2010). Les terres cultivées et les pâturages occupent maintenant 40 % de la surface
de la Terre (Foley et al., 2005 in Prieto-Benitez & Méndez, 2010). La transformation des terres
en agroécosystèmes a souvent eu pour conséquence une simplification du paysage et la
difficulté d’y maintenir un grand nombre d'espèces (Tscharntke et al., 2005).

2
THEBAULT David
Initialement en France le nombre et la diversité des petites régions agricoles ont
contribué à renforcer la biodiversité du territoire national. Mais après la seconde guerre
mondiale, l’agriculture française s’est réorganisée et le paysage agricole a été fortement
modifié (Agriculture et Biodiversité – ESCo, 2008). Les conséquences de ces aménagements
dans les pratiques agricoles et dans le paysage agricole, tels que : la concentration des
productions à l’échelle des exploitations et des régions, l’agrandissement de la taille des
parcelles, la suppression de nombreuses structures paysagères, la forte augmentation de
l’utilisation d’engrais chimiques et de produits phytosanitaires ; ont eu des conséquences
environnementales et paysagères négatives majeures ayant pour résultat la simplification du
paysage et une faible proportion d’habitats non agricoles (Schmidt et al., 2005). Tout cela a
abouti à une baisse importante de la répartition et de l’abondance de nombreuses espèces,
faunistique et floristique, associées aux terres agricoles (Hole et al., 2005). Les araignées font
partie de ces espèces ayant été particulièrement affectées par ces modifications récentes
des pratiques et du paysage agricole (Prieto-Benitez & Méndez, 2010). Depuis les années
2000, les préoccupations générales quant à l’impact négatif de l’agriculture sur les espèces
a conduit certains pays et exploitants agricoles à chercher à remettre en avant des pratiques
agricoles moins intensifs et plus respectueuses de l’environnement. L’Agriculture Biologique
(AB) est une de ces pratiques et se caractérise notamment par l’absence d’utilisation de
produits phytosanitaires. De nos jours, il est reconnu que les facteurs locaux, comme le type
de pratique agricole, et le contexte paysager influence les communautés d’araignées
(Schmidt et al., 2005).
Les tendances récentes de l’utilisation d’un mode d’agriculture alternatif sans
utilisation de produits phytosanitaire ont conduits à un intérêt accru des araignées comme
agents potentiels de lutte biologique. Actuellement, les araignées sont considérées comme
des agents de lutte biologique efficaces contre les ravageurs dans les agroécosystèmes
(Lang et al., 1999). De plus, des recherches récentes ont mis en évidence le rôle important
des prédateurs et en particulier des araignées dans la structure des communautés et
indirectement sur la production primaire (Bruno & Cardinale, 2008).
Les araignées sont des prédateurs présents dans tous les écosystèmes terrestres
(Sunderland, 1999) et font partie des groupes les plus diversifiés de la planète (Wise, 1993 in
Prieto-Benitez & Méndez, 2010). Elles se trouvent au niveau trophique le plus élevé parmi les
invertébrés (Main, 1987 in Prieto-Benitez & Méndez, 2010). La plupart des araignées sont des
prédateurs généralistes (Riechert et Luczak, 1982). Ils chassent principalement des insectes
(Turnbull, 1973 in Prieto-Benitez & Méndez, 2010) à tous leurs stades de développement (œufs,
larves, adultes) (Riechert et Lockley, 1984 in Prieto-Benitez & Méndez, 2010). Selon les familles,
les araignées chassent à l'affût, chassent en courant (dans la végétation ou au sol), ou tissent
des toiles (Cardoso et al., 2011, Uetz et al., 1999). Les araignées dans les agroécosystèmes ont
été jusqu’à maintenant relativement négligées malgré leur importance (Bruno et Cardinale,
2008). Les agroécosystèmes semblent être dominés par quelques espèces d'araignées ou «
agrobiontes » qui se développent dans des conditions perturbées (Luczak, 1979).
Suite au sommet planétaire de Rio (2002) et à cette dynamique autour de la
biodiversité de ces dernières années, de nombreux pays se sont dotés de stratégies
nationales sur la biodiversité et le développement durable. La France s’est dotée d’une
stratégie nationale pour la biodiversité en février 2004. La stratégie française reprend les
grands objectifs de la convention, à savoir la conservation et l’utilisation durable de la
diversité biologique et le partage équitable des bénéfices issus de cette utilisation. La
stratégie nationale propose également la mise en œuvre de plans d’action sectoriels en
faveur de la biodiversité.
Le Ministère de l’Alimentation, de l’Agriculture et de la Pêche (MAAP) pilote un de ces plans
d’action, le plan d’action « agriculture et biodiversité ». Ce dernier a commencé à être mis
en œuvre en 2004. L’une des actions phares est « de promouvoir les démarches coordonnées

3
THEBAULT David
des agriculteurs et de leurs partenaires visant à stopper, à l’échelle de leur territoire, la perte
de biodiversité (Agriculture et Biodiversité – ESCo, 2008)». Les priorités du plan d’action
« agriculture et biodiversité » ont reçu l’appui du plan de Développement Rural National
2007-2013, dont la préservation de la biodiversité constitue également l’un des enjeux
majeurs.
Ce stage est issu d’une commande de la chambre d’agriculture du Maine-et-Loire
dans le cadre du Projet Agricole Départemental (PAD), débuté en 2008. Le projet agricole
départemental vise à maintenir et valoriser l’agriculture de Maine-et-Loire, en conservant un
grand nombre d’actifs et des productions animales et végétales compétitives, et en prenant
en compte les différentes composantes du développement durable, particulièrement en
matière d’environnement et de biodiversité. Pour la biodiversité, les objectifs et les pistes
d’actions proposés par le PAD sont : 1) l’amélioration de la connaissance et l’acquisition de
références sur les interactions entre agriculture et biodiversité ; 2) définir des priorités par
milieu et/ou par espèces de manière concertée entre les acteurs ; 3) encourager le partage
d’expérience et la sensibilisation des agriculteurs; et 4) renforcer les actions de promotion des
pratiques déjà identifiées comme favorables à la biodiversité.
Pour l’encadrement de ce stage, nous avons une cellule technique et scientifique :
- La chambre d’agriculture est la structure d’accueil.
- Le CPIE joue le rôle de relai scientifique et naturaliste local.
- L’université de Rennes 1, avec Mr. PETILLON Julien, est le référent scientifique.
Cette étude s'inscrit dans le contexte de l’observation de la biodiversité fonctionnelle
dans des agroécosystèmes du Maine-et-Loire, à différentes échelles taxonomiques. Plus
précisément, nous désirons participer à l’amélioration des connaissances sur l’état de la
biodiversité en milieu agricole, car actuellement nous manquons encore de données. En
cela, nous voulons mettre en évidence les interactions entre agriculture et biodiversité d’une
manière générale.
Mon travail a porté sur deux grands volets de l’observation de la biodiversité en milieu
agricole.
Le premier volet a été le déploiement de l’Observatoire Agricole de la Biodiversité (OAB).
L’OAB est un projet scientifique national, avec des protocoles simple, basée sur le principe
des sciences participatives. L’objectif principal de ce projet est pédagogique. En effet, il a
pour but de fournir des outils de sensibilisation et d’éducation à la biodiversité aux acteurs du
monde agricole. Dans une moindre mesure, un autre des objectifs de l’OAB est la mise en
place d’indicateurs de biodiversité en milieu agricole via des observations ex-situ sur le terrain
en présence des agriculteurs. Cela afin d’avoir une meilleure compréhension de la
biodiversité ordinaire sauvage en milieu agricole, de son évolution et des liens de ces derniers
avec les pratiques agricoles.
Le second grand volet, de l’observation de la biodiversité en milieu agricole, a été l’étude de
l’impact des pratiques agricoles, et de la structuration du paysage sur les communautés
d’araignées dans des agroécosystèmes. Nos hypothèses sont d’une part que la richesse
spécifique et l’abondance des communautés d’araignées sont plus importantes en
agriculture biologique. D’autre part, que les variables paysagères non cultivées auront un
effet bénéfique sur les communautés d’araignées et ceci d’autant plus sur les individus
présents en bord de parcelle.

4
THEBAULT David
Pour des raisons de compréhension, ce rapport a été décomposé en deux chapitres:
Chapitre 1 : Observatoire Agricole de la Biodiversité (OAB)
Chapitre 2 : Etude des communautés d’araignées dans des agroécosystèmes
Présentation de la structure
1. Organisation
Le réseau des Chambres d'agriculture a été créé dans les années 1920 pour être un
interlocuteur privilégié des instances publiques et pour représenter les intérêts du monde
agricole. Il contribue activement depuis les années 60 au développement agricole en
reprenant la mission publique assumée jusque là par l'Etat. Aujourd'hui, cette double mission
se déploie dans le champ économique, social et environnemental ainsi qu'à l'échelle locale,
nationale et européenne.
Présentes dans chaque département et chaque région, les Chambres d'agriculture sont des
établissements publics dirigés par 4 200 élus professionnels, tous représentants des diverses
activités du secteur agricole. Animées, en lien avec les élus, par 7 800 collaborateurs, les
activités des Chambres contribuent au dynamisme de chaque département et région dans
une logique de développement durable.
L’Assemblée permanente des Chambres d’agriculture (ou APCA) coordonne à l’échelle
nationale les Chambres d’agriculture. Dans son rôle de tête de réseau, l’APCA assure la
formation des collaborateurs et des élus, anime le dialogue social, met à disposition des
dirigeants des références techniques et financières issues des données des Chambres et un
centre de ressources et d’expertises national.
2. Une répartition territoriale des missions
Les principales missions menées par les Chambres d'agriculture concernent : l'appui aux
entreprises, les ressources et la gestion des bases de données, l'économie et la politique
agricole, les territoires et le développement local, les démarches qualité des produits et
l'appui aux filières territorialisées, les analyses et les comptabilités, la promotion de l'agriculture
et de ses métiers, ainsi que l'agronomie et l'environnement. Sur le plan de l’environnement, à
l’échelle locale, ces missions sont inscrites dans le Projet Agricole Départemental (PAD).

5
THEBAULT David
Chapitre 1 : Observatoire Agricole de la Biodiversité (OAB)
1. Matériel et méthode
L’Observatoire Agricole de la Biodiversité (OAB) est un dispositif mis en place depuis 4 ans
(2010). L’OAB est une initiative du Ministère en charge de l’Agriculture. La coordination
scientifique est gérée par le muséum national d’histoire naturelle en partenariat avec
l’université de Rennes 1 pour le protocole « vers de terre » et un laboratoire du CNRS (LADYSS)
pour l’approche sociologique. L’Assemblée national permanente des Chambres
d’agricultures (APCA) s’occupe de l’animation nationale du projet.
Le réseau de l’OAB regroupe de nombreuses personnes.
- Les coordinateurs nationaux sont les structures décrites précédemment et forment les
animateurs locaux.
- Les animateurs locaux participent à l’OAB au sein de leurs structures. Ils animent un groupe
sur leur territoire en mobilisant et en formant les agriculteurs intéressés. La chambre
d’agriculture est l’animateur locale en Maine-et-Loire en charge de déployer l’OAB. Il existe
une grande diversité de structures locales, telles que : les chambres d’agricultures, les lycées
d’enseignement agricole, les Fédérations des chasseurs, coopératives, associations
naturaliste, etc.
- Les observateurs réalisent les observations de terrain, transmettent leurs données sur le site
internet de l’OAB et échangent autour de leur résultats. Durant ce stage, j’ai rempli le rôle
d’animateur et d’observateur de l’OAB en Maine-et-Loire.
Les protocoles de l’OAB ont été adaptés aux besoins et contraintes des agriculteurs grâce à :
une étude de faisabilité menée en 2010, une phase de test sur le terrain et une étude
sociologique. L’OAB comprend quatre protocoles d’étude : abeilles sauvages solitaires,
invertébrés à la surface du sol, vers de terre, et papillons. Le détail des protocoles sont visibles
en annexe 1. Ces protocoles sont volontairement ciblés sur les taxons les plus « représentatifs »
et pouvant être facilement observé par les professionnels du monde agricole. Tous les
agriculteurs intéressés peuvent participer. Le choix d’un ou plusieurs protocoles est libre. En
2013, 417 exploitations sont inscrites à l’OAB et 672 parcelles participent aux protocoles.
Nous avons commencé par prendre contact avec les agriculteurs du réseau A.R.B.R.E
susceptibles d’être intéressés par la démarche de l’OAB. Le réseau A.R.B.R.E ou « Agriculteurs
Respectueux de la Biodiversité et des Richesses de l’Environnement » est un réseau
d'agriculteurs volontaires pour intégrer la question de la biodiversité dans leur outil de
production, et pour répondre à des enjeux agricoles ainsi que de territoires1. Le premier suivi
mensuel de l’OAB a eu lieu fin avril. Les autres suivis ont eu lieu fin mai, mi-juillet et mi-août.
Une partie des suivis a été réalisée par les agriculteurs eux-mêmes. Cependant, j’ai été
présent auprès de l’ensemble des exploitants au moins lors du premier suivi. Le suivi du mois
de juin n’a pas eu lieu, car nous avons préféré focalisé notre temps sur la collecte des
araignées (chapitre 2 de ce rapport). Les relevés mensuels, réalisés sur le terrain, sont ensuite
saisis en ligne sur le site de l’OAB. Le traitement et l’exploitation de nos données sont réalisés
à l’échelle nationale par les coordinateurs nationaux. De courtes vidéos pédagogiques ont
été réalisées, sur le but ainsi que sur les protocoles de l’OAB, afin d’aider les agriculteurs
désirant intégrer l’OAB à mieux appréhender la démarche de cette dernière.
1 Source : CA 49 (Chambre d’Agriculture du Maine-et-Loire)
6
THEBAULT David
Les participants à l’OAB 2014 en Maine-et-Loire, sont : 24 exploitants agricoles du réseau
A.R.B.R.E, suivi par moi-même, ainsi que des viticulteurs non affiliés au réseau A.R.B.R.E et suivis
par le syndicat du Saumur-Champigny. La mise en place des protocoles a eu lieu en
présence des agriculteurs. Les protocoles que j’ai réalisés ont fini d’être mis en place début
avril. Au total, cela représente 72 protocoles, qui ont été répartis et mis en place sur la
période du mois de mars. Plus précisément, on a : 34 protocoles abeilles solitaires ; 19
protocoles planches à invertébrés ; 9 protocoles vers de terre et 10 protocoles papillons.
Parmi ces 72 protocoles, 10 protocoles abeilles solitaires et 10 protocoles papillon sont à la
charge du syndicat du Saumur-Champigny.
2. Résultats
Nous avons choisi pour la suite de ce chapitre de nous focaliser uniquement sur les résultats
des protocoles vers de terre. En effet, les données des autres protocoles ne sont pas encore
complétées à cette époque de l’année. Bien que le nombre de réplicats spatiaux des
protocoles vers de terre soit ici limités, ils ne nécessitent pas de réplicats temporels pour être
valide scientifiquement, à l’inverse des 3 autres protocoles de l’OAB (suivis chaque mois
jusqu’en octobre). Il est donc ainsi possible d’extrapoler ces résultats et de conserver une
validité scientifique tout en gardant un certain recul sur l’exploitation de ces résultats.
Les protocoles vers de terre étudiés ici sont issus à la fois de l’Observatoire Agricole de la
Biodiversité (OAB) et des Effets Non Intentionnelles (ENI) de 2014. Les protocoles ENI ont été
réalisés par le pôle agronomique de la chambre d’agriculture. Les protocoles OAB et ENI
étant identiques, il nous a paru plus pertinent de regrouper les protocoles. Au total, cela
représente 14 exploitations agricoles du Maine-et-Loire, échantillonnées entre mi-mars et mi-
avril.
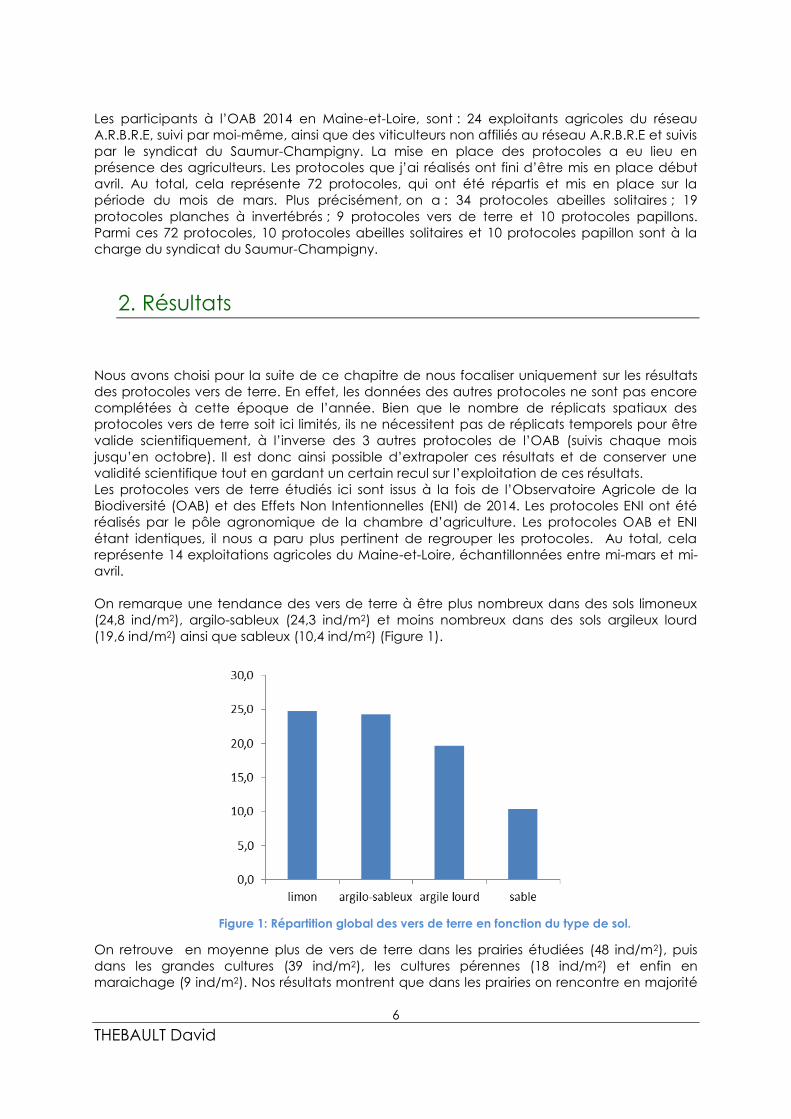
On remarque une tendance des vers de terre à être plus nombreux dans des sols limoneux
(24,8 ind/m2), argilo-sableux (24,3 ind/m2) et moins nombreux dans des sols argileux lourd
(19,6 ind/m2) ainsi que sableux (10,4 ind/m2) (Figure 1).
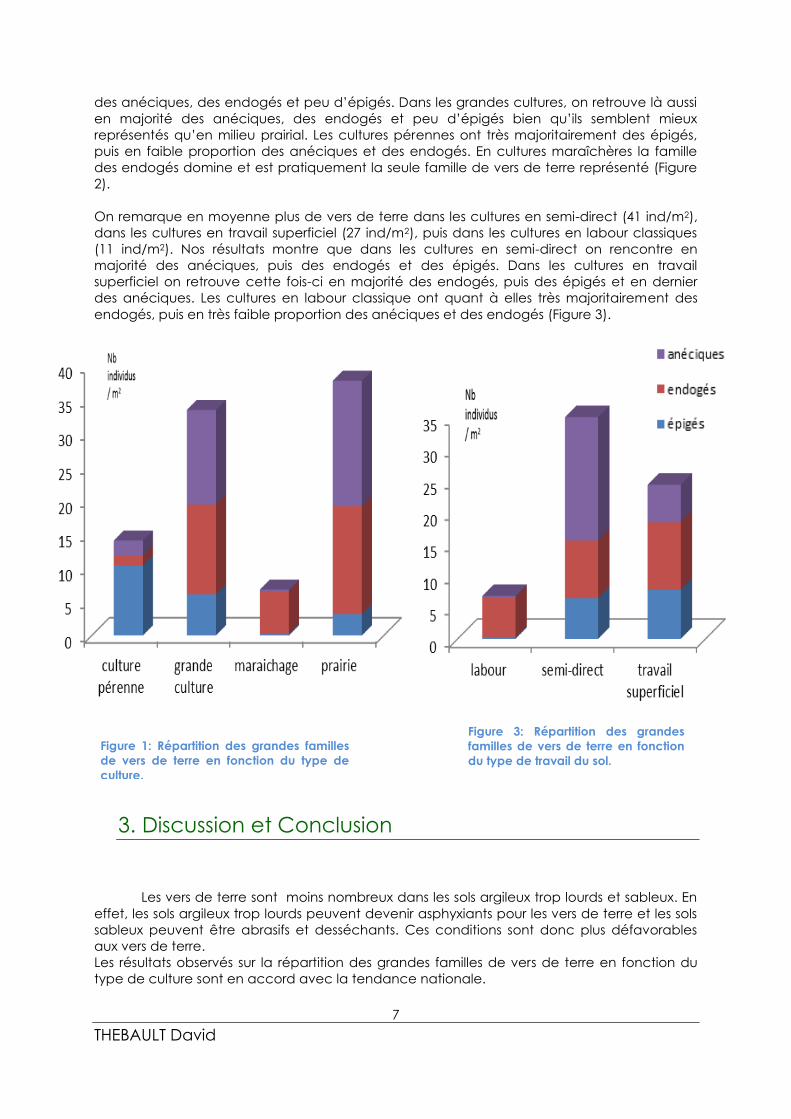
On retrouve en moyenne plus de vers de terre dans les prairies étudiées (48 ind/m2), puis
dans les grandes cultures (39 ind/m2), les cultures pérennes (18 ind/m2) et enfin en
maraichage (9 ind/m2). Nos résultats montrent que dans les prairies on rencontre en majorité
Figure 1: Répartition global des vers de terre en fonction du type de sol.
7
THEBAULT David
des anéciques, des endogés et peu d’épigés. Dans les grandes cultures, on retrouve là aussi
en majorité des anéciques, des endogés et peu d’épigés bien qu’ils semblent mieux
représentés qu’en milieu prairial. Les cultures pérennes ont très majoritairement des épigés,
puis en faible proportion des anéciques et des endogés. En cultures maraîchères la famille
des endogés domine et est pratiquement la seule famille de vers de terre représenté (Figure
2).
On remarque en moyenne plus de vers de terre dans les cultures en semi-direct (41 ind/m2),
dans les cultures en travail superficiel (27 ind/m2), puis dans les cultures en labour classiques
(11 ind/m2). Nos résultats montre que dans les cultures en semi-direct on rencontre en
majorité des anéciques, puis des endogés et des épigés. Dans les cultures en travail
superficiel on retrouve cette fois-ci en majorité des endogés, puis des épigés et en dernier
des anéciques. Les cultures en labour classique ont quant à elles très majoritairement des
endogés, puis en très faible proportion des anéciques et des endogés (Figure 3).
3. Discussion et Conclusion
Les vers de terre sont moins nombreux dans les sols argileux trop lourds et sableux. En
effet, les sols argileux trop lourds peuvent devenir asphyxiants pour les vers de terre et les sols
sableux peuvent être abrasifs et desséchants. Ces conditions sont donc plus défavorables
aux vers de terre.
Les résultats observés sur la répartition des grandes familles de vers de terre en fonction du
type de culture sont en accord avec la tendance nationale.
Figure 1: Répartition des grandes familles
de vers de terre en fonction du type de
culture.
Figure 3: Répartition des grandes
familles de vers de terre en fonction
du type de travail du sol.

8
THEBAULT David
Les prairies sont plus favorables aux lombriciens, car le milieu est globalement peu perturbé
(travail du sol et traitements chimiques) et offre une alimentation en quantité importante. Les
quantités moyennes observées sur les prairies de notre échantillon (48 ind/m2) sont
supérieures à la moyenne nationale (environ 32 ind/m²). Cette tendance peut-être due à un
facteur local et doit être confirmée par un plus grand échantillonnage. Dans le cas présent,
les quantités observées en grande culture (39 ind/m2) sont particulièrement abondantes
(moyenne nationale de l’OAB 2013 autour de 21 ind/m²). Cela peut s’expliquer en partie par
une plus grande proportion de grande culture en semi-directs ou en travail du sol simplifié,
dans nos parcelles étudiées, et donc une plus grande proportion de sols favorable au
développement des populations de vers de terre. La quantité importante d’épigés
particulièrement sensibles au travail du sol, semble confirmer cette hypothèse.
Les données concernant le maraîchage ne portent que sur une parcelle et ne doivent donc
pas être extrapolées. On peut toutefois préciser qu’il s’agit d’une parcelle au sol sableux ce
qui peut en partie expliquer la faible quantité globale de vers de terre et la présence très
majoritaire des endogés.
Les données concernant la culture pérenne (ici vigne) ne portent que sur une parcelle et ne
doivent donc pas être extrapolées. L’abondance globale est à peu près équivalente à la
moyenne nationale 2013 de l’OAB. En revanche, les proportions des endogés et des épigés
sont inversement proportionnelles à la moyenne.
L’OAB a globalement bien fonctionné chez les agriculteurs. Seuls 4 protocoles
planches à invertébrés et 1 protocole abeille ont été arrêtés pour cause de dégradation. De
nombreux agriculteurs (35% des agriculteurs) se sont approprié les protocoles en participant
par eux-mêmes aux suivis, et plus largement intéressés à la biodiversité observée au sein de
leurs parcelles. D’autres agriculteurs (35%) n’ont pas participé aux suivis faute de temps, mais
se sont tout de même vivement intéressés aux résultats obtenus chez eux. Ceci est en partie
dû au fait que les protocoles sont réalisés au sein d’un réseau d’agriculteurs particulièrement
sensibles à la biodiversité. Cependant, une proportion non négligeable d’exploitants
agricoles (30%) ne s’est que très peu voir pas intéressés aux protocoles expérimentaux mis en
place chez eux.
Le choix des parcelles s’est en partie porté sur celles venant ou allant être exploitées avec
des techniques culturales simplifiées. L’objectif était ici de réaliser un constat présent (à t0) de
l’état de la biodiversité sur ces parcelles et de voir l’évolution de cette dernière au cours du
temps. Dans d’autres cas, les parcelles étaient en pratiques culturales simplifiées depuis un
certain temps, et les résultats de l’OAB venaient ou pas les conforter dans leur choix de
pratique.
Il est important de préciser qu’un mauvais résultat aux protocoles de l’OAB, ne signifie pas
forcément qu’il y ait une mauvaise biodiversité globale sur les parcelles étudiées, mais que la
biodiversité n’a pas été mise en évidence via les protocoles de l’OAB.
Fort de cette expérience le réseau A.R.B.R.E pourrait concentrer ses efforts de déploiement
de l’OAB sur les agriculteurs particulièrement sensibles aux questions de biodiversité et de
chercher à compléter leurs attentes par des protocoles faunistiques et floristiques
complémentaires au sein de leur exploitation, quitte à réduire le nombre des protocoles.
Suite à cela, il serait approprié de faire participer éventuellement ces agriculteurs à des
réunions thématiques sur la biodiversité et l’OAB en présence d’autres agriculteurs. De cette
façon, il serait possible d’arriver progressivement à la mise en place d’un réseau
d’agriculteurs particulièrement impliqués sur cette thématique et demandeur de
connaissances, plutôt que de chercher à faire participer un grand nombre d’agriculteurs à
l’OAB pas toujours prêt à s’investir sur cette thématique.

9
THEBAULT David
Chapitre 2 : Etude des communautés d’araignées dans
des agroécosystèmes
1. Matériel et méthode
1.1 Sites d’études et choix des parcelles
Le Maine-et-Loire (49) est un territoire propice à l’agriculture notamment du fait d’un relief
très peu marqué. C’est le troisième département français (derrière la Marne et la Saône-et-
Loire) en surface agricole utilisée. L’activité agricole s’exerce sur 64 % de la superficie, soit 457
000 hectares. L’agriculture biologique représente 3,3 % des 64 %, soit 15 700 ha.2
Le choix des sites d’études a commencé par une sélection d’une zone homogène sur le
plan géologique (Figure 4) et climatique (Figure 5), afin de limiter l’effet de ces variables sur
notre étude (Lafage & Petillon, « en préparation »). Pour notre étude, les parcelles choisies
sont donc exclusivement sur des sols schisteux et dans des secteurs où l’amplitude des
précipitations se situe entre 600 et 700mm. Cela correspond à l’Ouest du département.
On a fait ce choix de parcelle compte tenu du caractère hétérogène des caractéristiques
pédoclimatique du Maine-et-Loire. En effet, L’ouest du département (Figure 4) est dominé
par le Massif armoricain avec des sols, plus anciens, essentiellement constitués de schiste, de
gneiss et de granite. À l’est, il y a le bassin Parisien, plus jeune, avec des sols calcaires,
constitués de grès, de tuffeau et de falun coquillier. Enfin la vallée de la Loire elle-même
constitue d’un territoire géologique à part entière, traversant d’est en ouest l’Anjou, et
constitué de sables et de graviers.3 L’ouest du département est également une zone ayant
des écarts de températures et de précipitations faibles (Figure 5).
Le choix des parcelles a été affiné en fonction de trois autres critères : 1) occupation
du sol par des cultures de blé d’hiver ou de triticale (culture très proche dans la nature dont
on considère la différence avec le blé comme négligeable), 2) cohérence avec les
2 Source : CA 49 (Chambre d’Agriculture du Maine-et-Loire) 3 Source : http://www.meteobell.com/__anjou_geomorphologie.php
Figure 4: Carte géologique du Maine-et-Loire Figure 5: Carte climatique du Maine-et-Loire
Ü Ü

10
THEBAULT David
modalités de l’étude (culture biologique, cultures conventionnelles en travail du sol
traditionnel (ou « labour normal ») ou en semi-direct) et 3) de l’hétérogénéité des variables
paysagères (milieu ouvert ou fermé).
Les 24 parcelles échantillonnées sont représentées, par des points bleus, sur la carte
ci-dessous (Figure 6). Parmi ces 24 parcelles, il y a : huit cultures conventionnelles en travail
classique du sol, huit cultures conventionnelles en semi-direct, et huit cultures biologiques en
travail classique du sol. Toutes étaient semées avec du blé d’hiver, à l’exception de trois
parcelles semées avec du triticale. La modalité « culture biologique en semi-direct » aurait
été pertinente pour cette étude cependant il n’a pas été possible de trouver un nombre de
réplicats spatiales suffisants.
Les points en rouge représentent les 12 parcelles ayant pu être analysées lors de cette étude
(six en cultures conventionnelles et six en biologiques) sur les 24 parcelles échantillonnées à
l’origine.
1.2 Dispositif expérimental
1.2.1 Période
L’étude a eu lieu entre mi-mai et fin juin. Cette période correspond au pic démographique
ainsi que de maturité sexuelle des populations d’araignées et est donc la plus pertinente
pour notre étude. Le protocole expérimental a été mis en place du 17 mai au 31 mai. Deux
campagnes de relevés (ou réplicats temporels) ont eu lieu, à 15 jours d’intervalle après la
mise en place des protocoles. La première campagne a eu lieu du 1er juin au 15 juin et la
deuxième campagne a eu lieu du 16 juin au 30 juin.
Par souci de temps, les données traitées dans ce rapport correspondent uniquement aux
données de la campagne 2. En effet, il a été observé durant la récolte de cette campagne
Figure 6: Répartition des parcelles de l’étude. A= Colineau P2, B= Coutard P, C= Cloarec P, D= Bidault P1, E=
Poupin P3, F= Poupin P1, G= Poupin P4, H= Pauvert P1, I= Aligon P2, J= Aligon P3, K= Perdrieau P, L= Cogné P1

11
THEBAULT David
un nombre d’individus plus abondant que dans la campagne 1. Cela s’explique
notamment par une pluviométrie moins importante, en phase avec la moyenne saisonnière.
Il nous a paru plus judicieux de traiter en priorité ce jeu de données.
1.2.2 Pièges barbers
L’échantillonnage des araignées a été effectué à l’aide de pièges barbers (H=12 cm, Ø=8,5
cm) (Knapp & Ruzicka, 2012). Ces derniers étaient protégés des intempéries par des assiettes
en plastique (Ø=20 cm) positionnées à 15 cm au-dessus du sol. Le liquide collecteur était
constitué d’un mélange de propylène glycol (40 %), d’eau (59 %) et de liquide vaisselle (1 %).
Les pièges barbers étaient remplis au 2/3. Le bord de ces derniers était disposé précisément
au niveau du sol (<1mm) afin d’éviter tout obstacle pouvant biaiser l’échantillonnage.
Pour chacune des 24 parcelles, neuf pièges barbers ont été disposés sur trois stations, elles-
mêmes disposées selon un pseudo-gradient de distance à la lisière de la parcelle : bord de
parcelle (Station 1), mi-distance entre le bord et le milieu de la parcelle (Station 2), et milieu
de parcelle (Station3).
Sur chaque station a été positionné un piquet rigide surplombé d’un fanion rouge, afin de
faciliter la visibilité de ces dernières lors des relevés de pièges.
Au total, 436 pièges barbers ont été relevés. Chaque piège d’une station était distant de 10m
les uns des autres, cette distance permettant de considérer les trois pots d’une même station
comme des réplicats spatiaux indépendants. Les barbers situés sur la station 1 en bord de
parcelle étaient distants de moins de 10m des bordures des parcelles afin de prendre en
compte un effet de lisière.
Photographies 1 et 2: Photos illustrant la mise en place d’un piège barber sur une des parcelles

12
THEBAULT David
Figure 7: Schéma d’une parcelle échantillonnée
Sur ces 436 pièges Barber, 72 ont pu être étudiés. Pour notre étude, seules les stations en
bord et en milieu de parcelle ont été conservées (Figure 7 - Stations colorées en rouge épais),
c’est-à-dire les stations les plus opposées sur le pseudo-gradient de distance à la lisière de la
parcelle. Le manque de temps ayant contraint à une sélection des stations et une limitation
du nombre de pièges à étudier.
1.3 Données locales et paysagères
En parallèle, des pièges barbers, des variables environnementales ont été relevées, à
l’échelle locale et à l’échelle paysagère. Ainsi pour l’échelle locale, des relevés de
pluviométrie, de type de pratique agricole (conventionnelle, biologique), et de type de
travail du sol (conventionnelle avec labour normal ou en semi-direct) ont été notés sur les 24
parcelles. La pluviométrie a été relevée quotidiennement par les agriculteurs sur la parcelle
concernée. Pour notre étude, seule l’incidence du travail du sol n’a pas été prise en compte.
Le manque de temps ayant contraint à une sélection des parcelles.
À l’échelle paysagère, un certain nombre de variables ont été relevées pour chacune des 24
parcelles. Afin de déterminer l’échelle la plus pertinente pour la prise en compte du paysage
en rapport avec le taxon étudié, nous avons testé différents rayons (100, 200 et 500 mètres) et
ceci pour chaque station (S1, S2, S3). La structure du paysage a été caractérisée en
considérant comme unité élémentaire (patch) la parcelle agricole.
Dans ces rayons ont été calculées, à l’aide du logiciel ArcGIS (version 10.2) :
- les longueurs de haie, et de cours d’eau (en mètres linéaires)
- les cultures, les prairies, le boisement, le bâtiment, les mares, et les bandes enherbées
(en pourcentages (%) de surface)
Centre de la parcelle
10 m
Gradient de distance à la lisière de parcelle
Piège
barber
Lisière (bord)
de la parcelle
Station 1 : en bord de
parcelle (bp)
Station 2 : mi-distance
entre Station 1 et 3
Station 3 : milieu
de parcelle (mp)

13
THEBAULT David
Ces informations ont pu être acquises par l’utilisation de photos aériennes, de couches SIG et
complétées par des enquêtes de terrain. Étant donné que seules les deux stations en bord et
milieu de parcelle ont été conservées pour notre étude, alors seuls les rayons (100, 200 et 500
mètres) de ces stations ont été conservés pour l’analyse.
1.4 Méthode d’identification
Les araignées (Araneae) collectées ont été stockées (dans de l’alcool à 70 degrés), triées,
dénombrées, puis identifiées jusqu’à l’espèce sous loupe binoculaire par l’observation des
pièces génitales des individus adultes et matures et à l’aide de l’utilisation d’ouvrages de
référence (« Spiders of Great Britain and Ireland », Michael J. Roberts Vol. 1, 2 et 3).
1.5 Base de données
Le jeu de données final, réalisé sur les 12 parcelles (72 pots), se compose de : 86 espèces
(3750 individus au total) réparties au sein de 42 genres et 12 familles. La liste complète des
espèces est fournie dans l’annexe 2.
Les données étudiées - issues de la 2éme campagne - sont séparées par pièges, et regroupent
les stations bord et milieu de chaque parcelle. À chaque piège est calculé l’abondance
totale, l’abondance par type de chasse, la richesse spécifique totale et la richesse
spécifique par type de chasse.
Les catégories de type de chasse déterminées pour cette étude sont : Chasseuses à l’Affût
(CA), Chasseuses Coureuses (CC) (dans la végétation et au sol), ainsi que les Tisseuses de
Toiles (TT) (en nappe, géométrique et en réseau) (Cardoso et al., 2011, Uetz et al., 1999).
1.6 Analyses des données
Toutes les données ont été analysées à l’aide des logiciels R (version 3.1.1, 2014), et PAST
(version 3.02a, 2014).
1.6.1 Présentation générale des prélèvements
Le nombre d’individus (Nbr ind), la richesse spécifique totale observée (Sobs), la richesse
spécifique totale estimée (Sest), la complétude (Sobs/Sest), l’indice de diversité de Simpson
(1-D), et l’équitabilité (J) pour chaque parcelle ont été calculés avec le logiciel PAST.
La richesse spécifique estimée a été calculée en faisant la moyenne des estimateurs de
biodiversité : Chao2, Bootstrap, ainsi que Jackknife 1 et 2. La richesse spécifique estimée nous
as permis de calculer la complétude de chaque parcelle.
La complétude permet d’estimer si l’effort d’échantillonnage réalisé sur une parcelle est
suffisant et que la richesse spécifique observée est suffisamment proche de la richesse
spécifique estimée. Elle se calcule en divisant la richesse spécifique observée par la richesse
spécifique estimée. Elle peut être considérée comme satisfaisante lorsque l’indice est
supérieur à 0,75.

14
THEBAULT David
L’indice de diversité de Simpson (1-D) est issu de l’indice de Simpson (D). Le maximum de
diversité d’une population étant représenté par la valeur 1, et le minimum de diversité par la
valeur 0 (Krebs, 1989). Cet indice de diversité donne plus de poids aux espèces abondantes
qu’aux espèces rares. C’est pour cela que nous avons également étudié l’indice
d’équitabilité J de Piélou (1966). Il permet d’exprimer la dominance d’une espèce. Il peut
varier de 0 à 1, il est maximal quand les espèces ont des abondances identiques dans le
peuplement et il est minimal quand une seule espèce domine tout le peuplement.
1.6.2 Comparaison des communautés d’araignées en fonction du type de pratique
agricole (conventionnelle / biologique)
Au-delà de la simple présentation des données, notre objectif premier a été de comparer les
communautés d’araignées en fonction du type de pratique agricole.
1.6.2.1 Comparaison des assemblages de communautés d’araignées
Ainsi, nous avons commencé par étudier la répartition spatiale du peuplement
d’araignées de notre étude. Cela a été réalisé en trois temps.
Tout d’abord nous avons analysé le peuplement d’araignées selon une méthode
statistique d’analyse de similarité des espèces, entre les parcelles, dans le but d’identifier des
assemblages de peuplement. Cette technique de classification produit des schémas
d’arborescence (aussi appelés dendrogrammes) qui relie les données selon leur niveau de
ressemblance (ou similarité). Le dendrogramme réalisé a été calculé à partir de l’occurrence
des espèces d’araignées.
Puis nous avons affiné notre étude, par une analyse multivariée, afin de connaitre plus
précisément les espèces présentes dans les assemblages. Pour cette seconde analyse, les
données étant en effectifs et organisées dans un tableau de contingence, une analyse
factorielle des correspondances a été réalisée.
Cependant les analyses de type AFC accordent un poids important aux espèces atypiques,
c’est-à-dire rares ou particulières. Nous avons donc choisi de restreindre le jeu de données,
en retirant les espèces rares, et en ne conservant que les 20 espèces les plus communes.
Cela contribue également à une meilleure lisibilité du graphique.
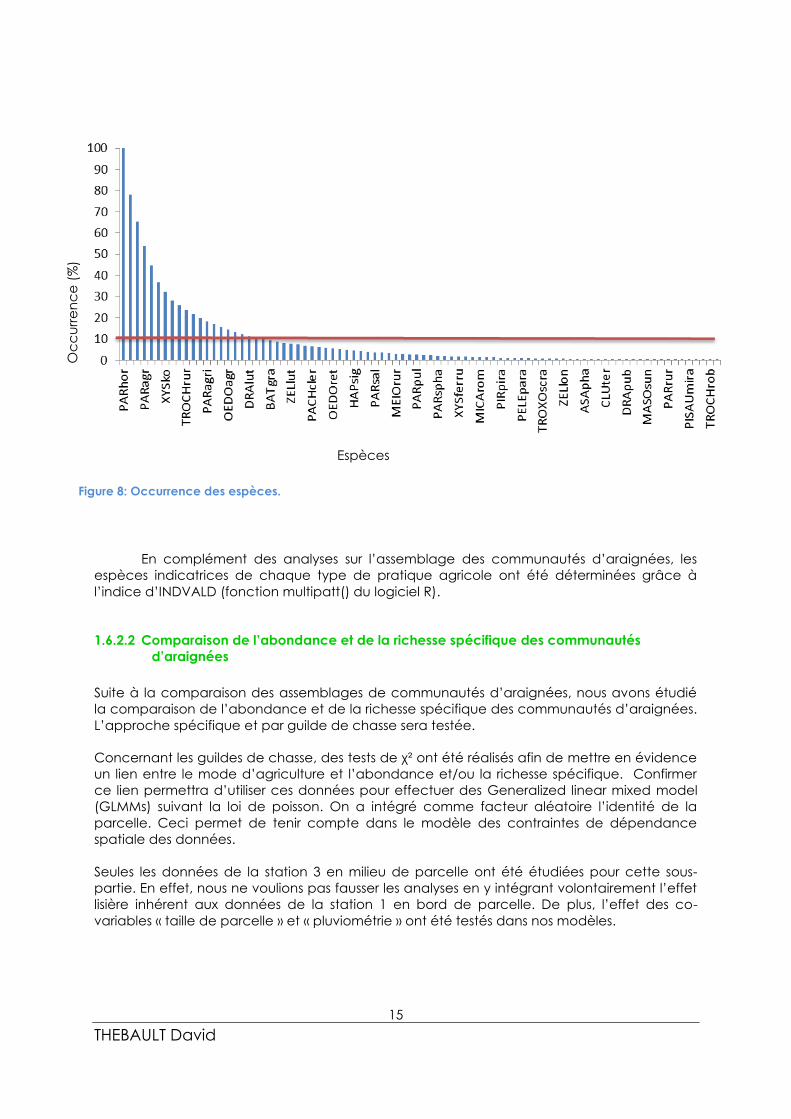
Ces 20 espèces dominantes représentent 90 % de l’occurrence totale (figure 8). Ces espèces
sont :
Pardosa hortensis, Pardosa proxima, Oedothorax apicatus, Pardosa agrestis, Ozyptila
simplex, Ozyptila fuscus, Xysticus kochi, Walckenaeria vigilax, Pardosa nigriceps, Trochosa
ruricola, Erigone dentipalpis, Tenuiphantes tenuis, Pardosa agricola, Zelotes civicus, Erigone
atra, Oedothorax agrestis, Zelotes tenuis, Zelotes pusillus, Drassylus lutetianus, et Pardosa
palustris.
15
THEBAULT David
En complément des analyses sur l’assemblage des communautés d’araignées, les
espèces indicatrices de chaque type de pratique agricole ont été déterminées grâce à
l’indice d’INDVALD (fonction multipatt() du logiciel R).
1.6.2.2 Comparaison de l’abondance et de la richesse spécifique des communautés
d’araignées
Suite à la comparaison des assemblages de communautés d’araignées, nous avons étudié
la comparaison de l’abondance et de la richesse spécifique des communautés d’araignées.
L’approche spécifique et par guilde de chasse sera testée.
Concernant les guildes de chasse, des tests de χ² ont été réalisés afin de mettre en évidence
un lien entre le mode d’agriculture et l’abondance et/ou la richesse spécifique. Confirmer
ce lien permettra d’utiliser ces données pour effectuer des Generalized linear mixed model
(GLMMs) suivant la loi de poisson. On a intégré comme facteur aléatoire l’identité de la
parcelle. Ceci permet de tenir compte dans le modèle des contraintes de dépendance
spatiale des données.
Seules les données de la station 3 en milieu de parcelle ont été étudiées pour cette sous-
partie. En effet, nous ne voulions pas fausser les analyses en y intégrant volontairement l’effet
lisière inhérent aux données de la station 1 en bord de parcelle. De plus, l’effet des co-
variables « taille de parcelle » et « pluviométrie » ont été testés dans nos modèles.
Oc
cu
rre
nc
e (
%)
Espèces
Figure 8: Occurrence des espèces.
16
THEBAULT David
1.6.3 Influence des variables paysagères sur les communautés d’araignées
Notre second objectif a été d’étudier l’influence des variables paysagères sur les
communautés d’araignées.
L’influence des variables paysagères avec ou sans interaction avec la position dans la
parcelle a été testée en réalisant des Generalised Linear Models (GLMs), et en utilisant les
richesses spécifiques et abondances totales ou les richesses spécifiques et abondances par
guilde de chasse comme facteur à expliquer. Sur chacune des 12 parcelles, deux stations
ont été étudiées. La station en bord de parcelle a pour objectif de représenter l’effet lisière
facilement colonisable par les communautés d’araignées, car le plus proche du contexte
paysager environnant. La station en milieu de culture, représente la zone de la parcelle la
moins facilement colonisable, car la plus éloignée du contexte paysager entourant la
parcelle.
Pour tous les tests réalisés, le taux d’erreur accepté est de 5 %. Tous les résultats présentent
une probabilité P-value (ou P) inférieure à ce pourcentage seront jugés significativement
différents.
8 C1 0
8 C2 6 C2
8 C1 0
8 C2 6 C2
8 C1 0
8 C2 0
Pour résumer :
Nbr de
parcellesPratiques Agricole
Stations
par
436 pièges barbers ; Estimation : 22 710 individus 72 pièges barbers ; 3750 individus
travail classique du sol
semi-direct
Nbr de
parcellesPratiques Agricole
bio
conv
Camp. Camp.
Données mobilisées pour l'analyse
S1, S2, S3
Données échantillonnées sur le terrain
Stations
par
S1, S3
bio travail classique du sol
conv travail classique du sol
conv semi-directconv
travail classique du sol
2. Résultats :
2.1 Présentation générale des prélèvements
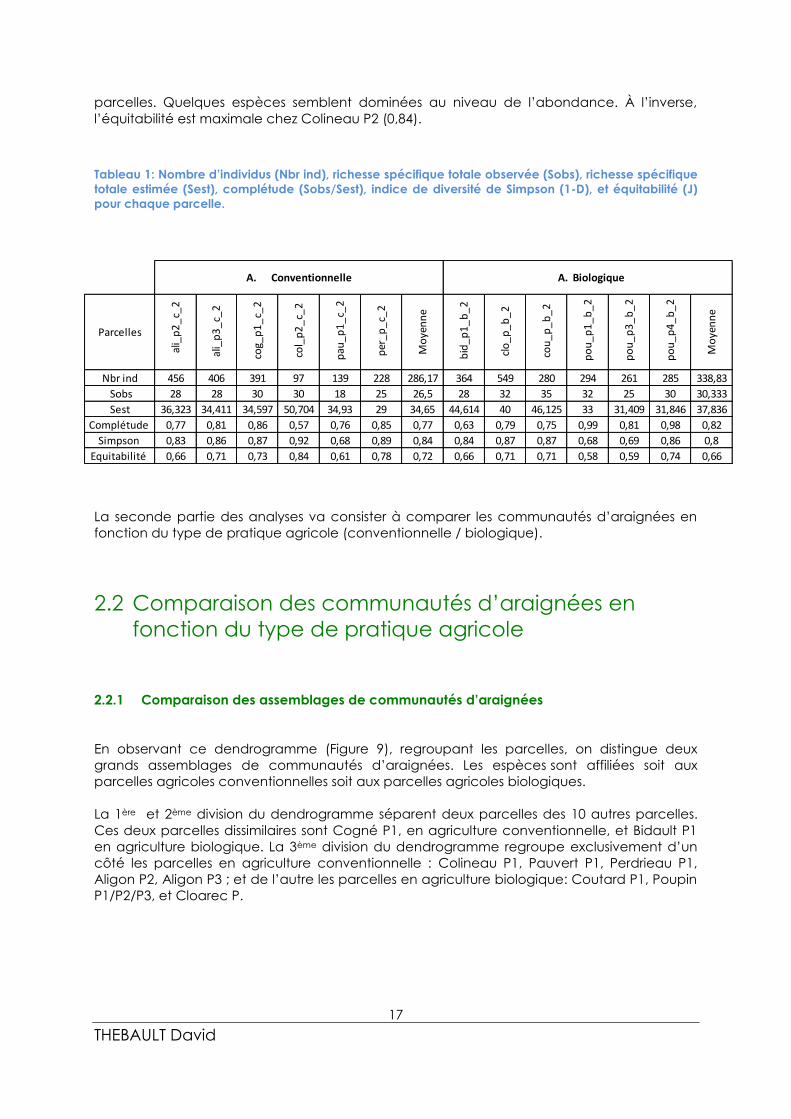
Les 12 parcelles étudiées ont une abondance variant de 97 à 549 individus (Tableau 1).
La richesse spécifique des parcelles varie de 18 à 35. L’abondance et la richesse spécifique
observées sont légèrement supérieures en agriculture biologique qu’en conventionnelle.
La complétude peut être considérée comme satisfaisante lorsque l’indice de
complétude est supérieur à 0,75. C’est le cas ici pour 10 des 12 parcelles. Seules les parcelles
de Colineau P2 et de Bidault P1 semblent avoir eu un effort d’échantillonnage (ou
complétude) insuffisant. Avec des indices de complétude moyens de 0,77 pour les parcelles
conventionnelles et de 0,82 pour les parcelles biologiques, les complétudes moyennes sont
suffisantes.
L’indice de diversité de Simpson est minimal dans les parcelles de Poupin P1 et Pauvert
P1 (0,68), et maximale chez Colineau P2 (0,92). L’ensemble des parcelles tend globalement à
avoir une grande diversité. L’équitabilité est minimale dans les parcelles Poupin P1 et P3 (0,58
et 0,59). La répartition du nombre d’individus par espèce est donc moins bonne sur ces
17
THEBAULT David
parcelles. Quelques espèces semblent dominées au niveau de l’abondance. À l’inverse,
l’équitabilité est maximale chez Colineau P2 (0,84).
Tableau 1: Nombre d’individus (Nbr ind), richesse spécifique totale observée (Sobs), richesse spécifique
totale estimée (Sest), complétude (Sobs/Sest), indice de diversité de Simpson (1-D), et équitabilité (J)
pour chaque parcelle.
La seconde partie des analyses va consister à comparer les communautés d’araignées en
fonction du type de pratique agricole (conventionnelle / biologique).
2.2 Comparaison des communautés d’araignées en
fonction du type de pratique agricole
2.2.1 Comparaison des assemblages de communautés d’araignées
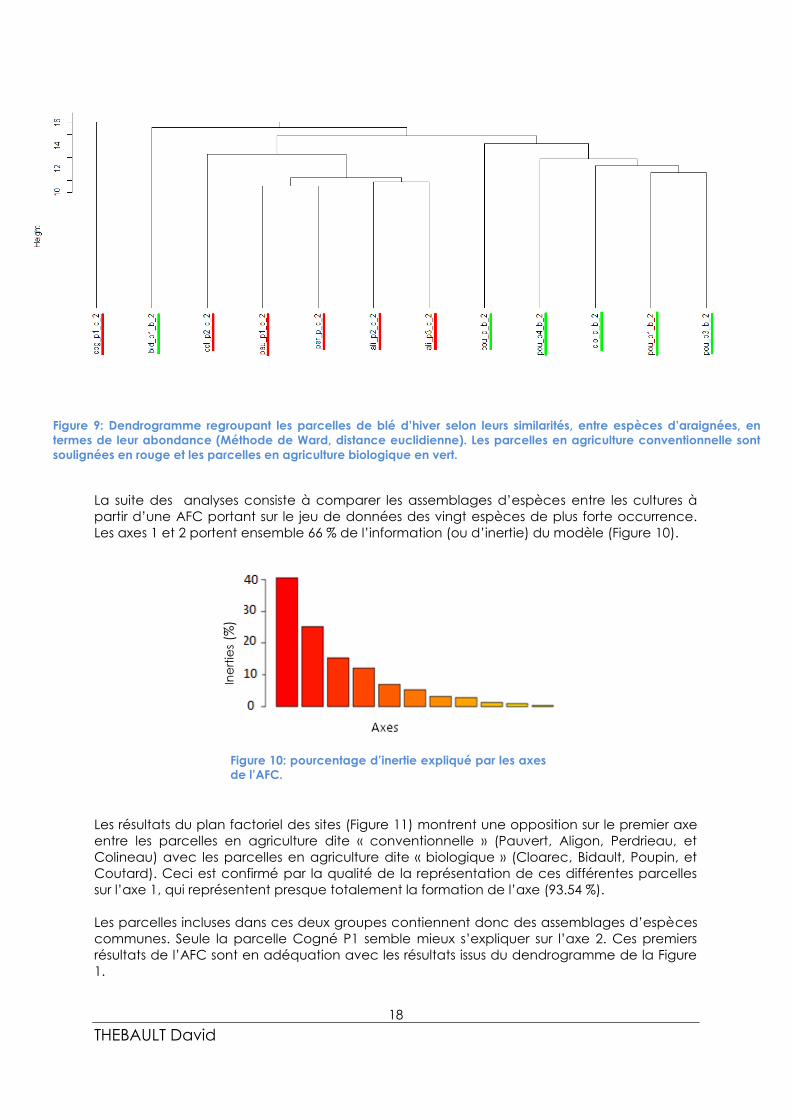
En observant ce dendrogramme (Figure 9), regroupant les parcelles, on distingue deux
grands assemblages de communautés d’araignées. Les espèces sont affiliées soit aux
parcelles agricoles conventionnelles soit aux parcelles agricoles biologiques.
La 1ère et 2ème division du dendrogramme séparent deux parcelles des 10 autres parcelles.
Ces deux parcelles dissimilaires sont Cogné P1, en agriculture conventionnelle, et Bidault P1
en agriculture biologique. La 3ème division du dendrogramme regroupe exclusivement d’un
côté les parcelles en agriculture conventionnelle : Colineau P1, Pauvert P1, Perdrieau P1,
Aligon P2, Aligon P3 ; et de l’autre les parcelles en agriculture biologique: Coutard P1, Poupin
P1/P2/P3, et Cloarec P.
Parcelles
al
i_p
2_
c_2
ali_
p3
_c_
2
cog_
p1
_c_
2
col_
p2
_c_
2
pau
_p
1_
c_2
per
_p
_c_
2
Mo
yen
ne
bid
_p
1_
b_
2
clo
_p
_b
_2
cou
_p
_b
_2
po
u_
p1
_b
_2
po
u_
p3
_b
_2
po
u_
p4
_b
_2
Mo
yen
ne
Nbr ind 456 406 391 97 139 228 286,17 364 549 280 294 261 285 338,83
Sobs 28 28 30 30 18 25 26,5 28 32 35 32 25 30 30,333
Sest 36,323 34,411 34,597 50,704 34,93 29 34,65 44,614 40 46,125 33 31,409 31,846 37,836
Complétude 0,77 0,81 0,86 0,57 0,76 0,85 0,77 0,63 0,79 0,75 0,99 0,81 0,98 0,82
Simpson 0,83 0,86 0,87 0,92 0,68 0,89 0,84 0,84 0,87 0,87 0,68 0,69 0,86 0,8
Equitabilité 0,66 0,71 0,73 0,84 0,61 0,78 0,72 0,66 0,71 0,71 0,58 0,59 0,74 0,66
A. BiologiqueA. Conventionnelle
18
THEBAULT David
Figure 9: Dendrogramme regroupant les parcelles de blé d’hiver selon leurs similarités, entre espèces d’araignées, en
termes de leur abondance (Méthode de Ward, distance euclidienne). Les parcelles en agriculture conventionnelle sont
soulignées en rouge et les parcelles en agriculture biologique en vert.
Figure 10: pourcentage d’inertie expliqué par les axes
de l’AFC.
Ine
rtie
s (%
)
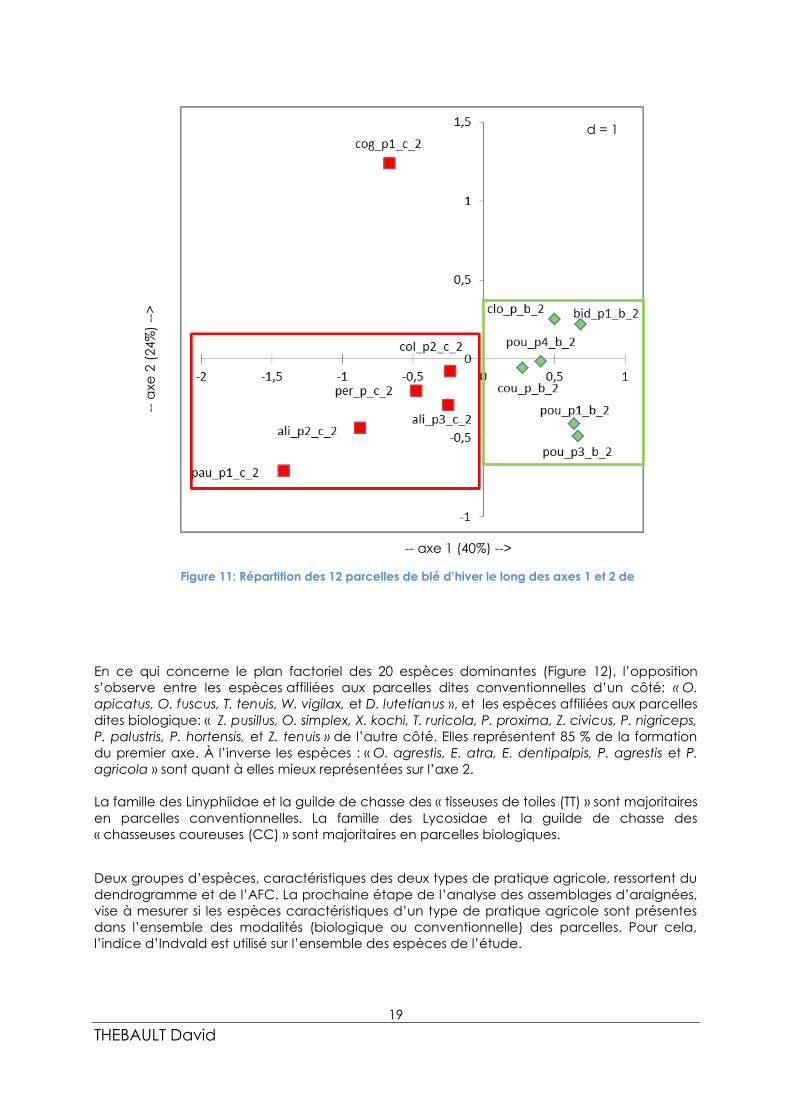
La suite des analyses consiste à comparer les assemblages d’espèces entre les cultures à
partir d’une AFC portant sur le jeu de données des vingt espèces de plus forte occurrence.
Les axes 1 et 2 portent ensemble 66 % de l’information (ou d’inertie) du modèle (Figure 10).
Les résultats du plan factoriel des sites (Figure 11) montrent une opposition sur le premier axe
entre les parcelles en agriculture dite « conventionnelle » (Pauvert, Aligon, Perdrieau, et
Colineau) avec les parcelles en agriculture dite « biologique » (Cloarec, Bidault, Poupin, et
Coutard). Ceci est confirmé par la qualité de la représentation de ces différentes parcelles
sur l’axe 1, qui représentent presque totalement la formation de l’axe (93.54 %).
Les parcelles incluses dans ces deux groupes contiennent donc des assemblages d’espèces
communes. Seule la parcelle Cogné P1 semble mieux s’expliquer sur l’axe 2. Ces premiers
résultats de l’AFC sont en adéquation avec les résultats issus du dendrogramme de la Figure
1.
19
THEBAULT David
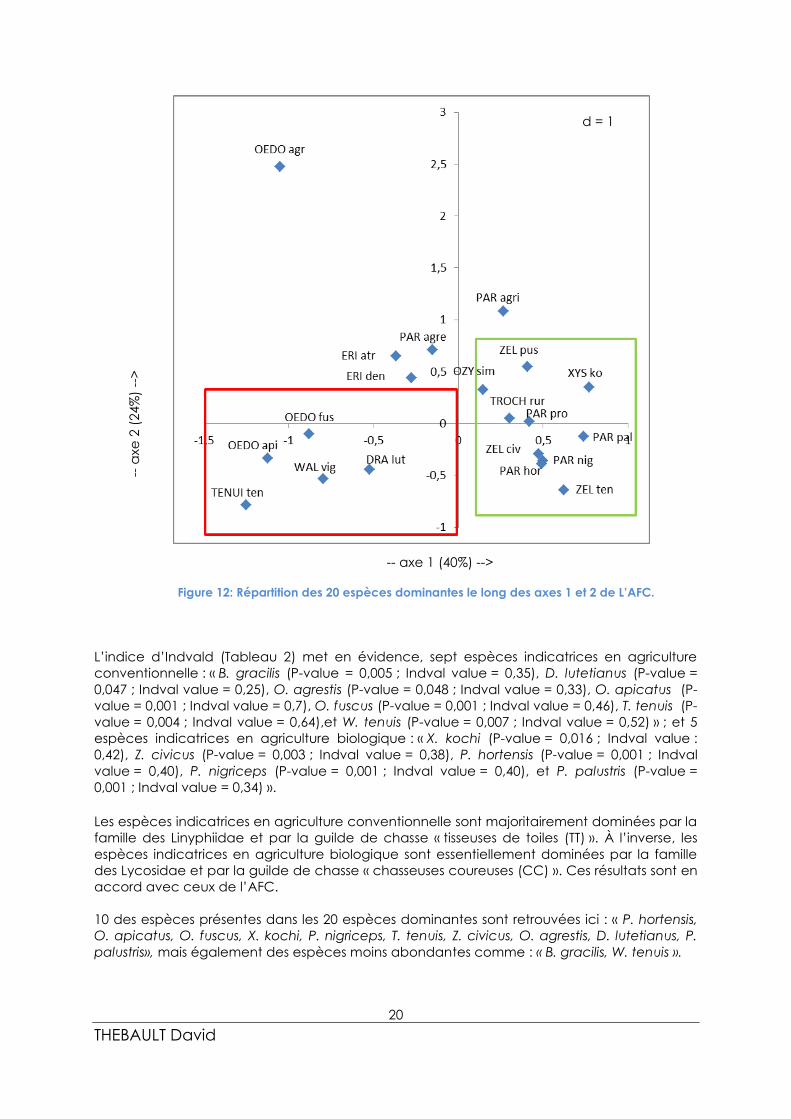
En ce qui concerne le plan factoriel des 20 espèces dominantes (Figure 12), l’opposition
s’observe entre les espèces affiliées aux parcelles dites conventionnelles d’un côté: « O.
apicatus, O. fuscus, T. tenuis, W. vigilax, et D. lutetianus », et les espèces affiliées aux parcelles
dites biologique: « Z. pusillus, O. simplex, X. kochi, T. ruricola, P. proxima, Z. civicus, P. nigriceps,
P. palustris, P. hortensis, et Z. tenuis » de l’autre côté. Elles représentent 85 % de la formation
du premier axe. À l’inverse les espèces : « O. agrestis, E. atra, E. dentipalpis, P. agrestis et P.
agricola » sont quant à elles mieux représentées sur l’axe 2.
La famille des Linyphiidae et la guilde de chasse des « tisseuses de toiles (TT) » sont majoritaires
en parcelles conventionnelles. La famille des Lycosidae et la guilde de chasse des
« chasseuses coureuses (CC) » sont majoritaires en parcelles biologiques.
Deux groupes d’espèces, caractéristiques des deux types de pratique agricole, ressortent du
dendrogramme et de l’AFC. La prochaine étape de l’analyse des assemblages d’araignées,
vise à mesurer si les espèces caractéristiques d’un type de pratique agricole sont présentes
dans l’ensemble des modalités (biologique ou conventionnelle) des parcelles. Pour cela,
l’indice d’Indvald est utilisé sur l’ensemble des espèces de l’étude.
-- axe 1 (40%) -->
-- a
xe
2 (
24
%)
-->
d = 1
Figure 11: Répartition des 12 parcelles de blé d’hiver le long des axes 1 et 2 de
L’AFC.
20
THEBAULT David
L’indice d’Indvald (Tableau 2) met en évidence, sept espèces indicatrices en agriculture
conventionnelle : « B. gracilis (P-value = 0,005 ; Indval value = 0,35), D. lutetianus (P-value =
0,047 ; Indval value = 0,25), O. agrestis (P-value = 0,048 ; Indval value = 0,33), O. apicatus (P-
value = 0,001 ; Indval value = 0,7), O. fuscus (P-value = 0,001 ; Indval value = 0,46), T. tenuis (P-
value = 0,004 ; Indval value = 0,64),et W. tenuis (P-value = 0,007 ; Indval value = 0,52) » ; et 5
espèces indicatrices en agriculture biologique : « X. kochi (P-value = 0,016 ; Indval value :
0,42), Z. civicus (P-value = 0,003 ; Indval value = 0,38), P. hortensis (P-value = 0,001 ; Indval
value = 0,40), P. nigriceps (P-value = 0,001 ; Indval value = 0,40), et P. palustris (P-value =
0,001 ; Indval value = 0,34) ».
Les espèces indicatrices en agriculture conventionnelle sont majoritairement dominées par la
famille des Linyphiidae et par la guilde de chasse « tisseuses de toiles (TT) ». À l’inverse, les
espèces indicatrices en agriculture biologique sont essentiellement dominées par la famille
des Lycosidae et par la guilde de chasse « chasseuses coureuses (CC) ». Ces résultats sont en
accord avec ceux de l’AFC.
10 des espèces présentes dans les 20 espèces dominantes sont retrouvées ici : « P. hortensis,
O. apicatus, O. fuscus, X. kochi, P. nigriceps, T. tenuis, Z. civicus, O. agrestis, D. lutetianus, P.
palustris», mais également des espèces moins abondantes comme : « B. gracilis, W. tenuis ».
Figure 2:
-- a
xe
2 (
24
%)
-->
-- axe 1 (40%) -->
d = 1
Figure 12: Répartition des 20 espèces dominantes le long des axes 1 et 2 de L’AFC.
21
THEBAULT David
Tableau 2: Espèces indicatrices pour un type de pratique agricole. L’ensemble des espèces sont
significativement indicatrices avec une P-value < 0,05 et un Indval value≥ 0,25. L’analyse a été réalisée
en abondance totale.
Famille Espèce Pratique A P-value Indval value Type de chasse
Linyphiidae Bathyphantes gracilis conv 0,005 0,35 Tisseuses de toiles
Gnaphosidae Drassyllus lutetianus conv 0,047 0,25 Chasseuses coureuses
Linyphiidae Oedothorax agrestis conv 0,048 0,33 Tisseuses de toiles
Linyphiidae Oedothorax apicatus conv 0,001 0,7 Tisseuses de toiles
Linyphiidae Oedothorax fuscus conv 0,001 0,46 Tisseuses de toiles
Linyphiidae Tenuiphantes tenuis conv 0,004 0,64 Tisseuses de toiles
Linyphiidae Walckenaeria tenuis conv 0,007 0,52 Tisseuses de toiles
Thomisidae Xysticus kochi bio 0,016 0,42 Chasseuses à l’affût
Gnaphosidae Zelotes civicus bio 0,003 0,38 Chasseuses coureuses
Lycosidae Pardosa hortensis bio 0,001 0,4 Chasseuses coureuses
Lycosidae Pardosa nigriceps bio 0,001 0,4 Chasseuses coureuses
Lycosidae Pardosa palustris bio 0,001 0,34 Chasseuses coureuses
2.2.2 Comparaison de l’abondance et de la richesse spécifique des communautés
d’araignées
Pour cette sous-partie, seules les données issues de la station en milieu de parcelle ont été
conservées, cela afin de ne pas avoir d’effet lisière.
Nous avons commencé par étudier la différence de proportion des communautés
d’araignées entre agriculture conventionnelle et biologique. Dans un premier temps, nous
avons regardé la différence (de proportion) d’individus totaux en agriculture biologique (49,7
%) et en conventionnel (50,3 %). Les proportions étant quasiment identiques, entre les deux
types d’agriculture, nous avons choisi dans un second temps de catégoriser les individus par
guilde de chasse (Figures 13 et 14) : Chasseuses à l’affût (CA) ; Chasseuses Coureuses (CC) ;
et Tisseuses de Toiles (TT).
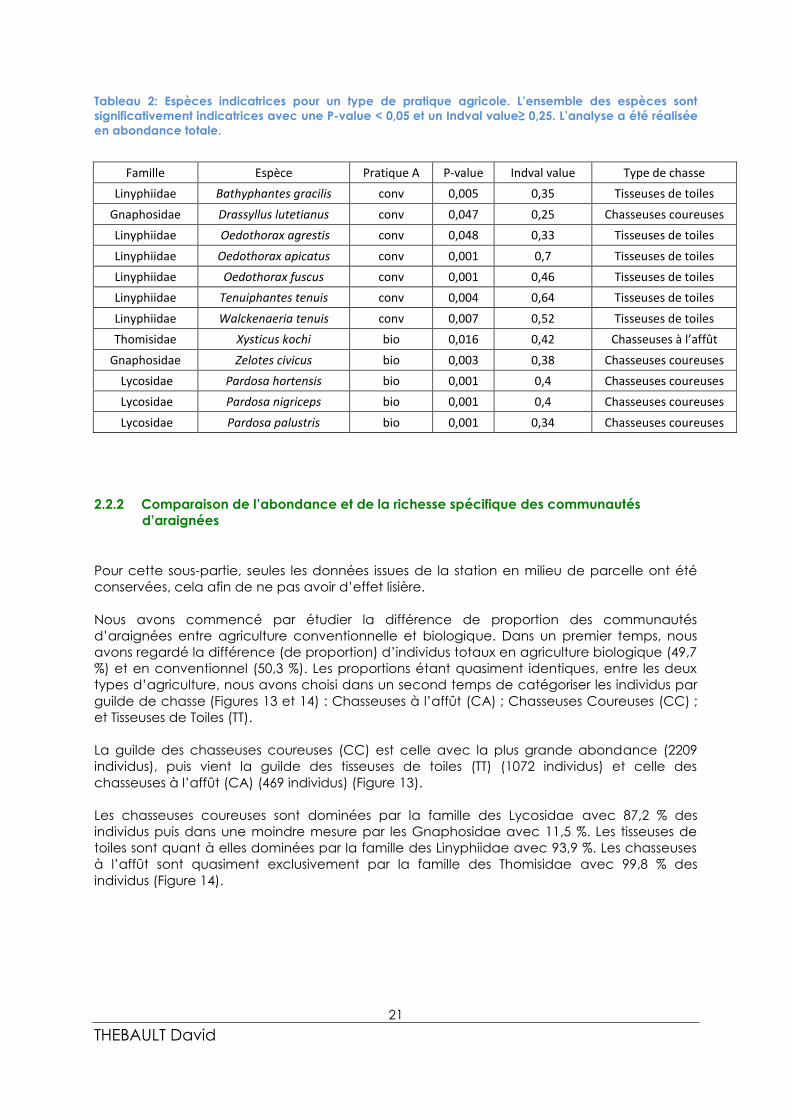
La guilde des chasseuses coureuses (CC) est celle avec la plus grande abondance (2209
individus), puis vient la guilde des tisseuses de toiles (TT) (1072 individus) et celle des
chasseuses à l’affût (CA) (469 individus) (Figure 13).
Les chasseuses coureuses sont dominées par la famille des Lycosidae avec 87,2 % des
individus puis dans une moindre mesure par les Gnaphosidae avec 11,5 %. Les tisseuses de
toiles sont quant à elles dominées par la famille des Linyphiidae avec 93,9 %. Les chasseuses
à l’affût sont quasiment exclusivement par la famille des Thomisidae avec 99,8 % des
individus (Figure 14).
22
THEBAULT David
Pro
po
rtio
n
d’in
div
idu
s p
ar
fam
ille
(%)
Type de chasse
Le test de chi-deux (Figure 15) de comparaison des proportions d’individus, en culture
biologique et conventionnelle, a mis en évidence des différences significatives pour les
chasseuses à l’affût (P-value = 0.003), pour les chasseuses coureuses (P-value < 0,001), et pour
les tisseuses de toiles (P-value < 0,001).
Figure 13: Nombre d’individus en
fonction du type de chasse.
No
mb
re d
’in
div
idu
s
Type de chasse
Figure 14: Proportion d’individus par
famille en fonction du type de
chasse.
Pro
po
rtio
n d
’in
div
idu
s
Figure 15: Résultats des tests χ2 de comparaison, des proportions d’individus, des différentes
guildes de chasse (CA, CC, TT) en fonction du type de pratique agricole. Des lettres différentes
ont été attribuées lorsque le test est significatif (P-value < 0,05).
Type de chasse

23
THEBAULT David
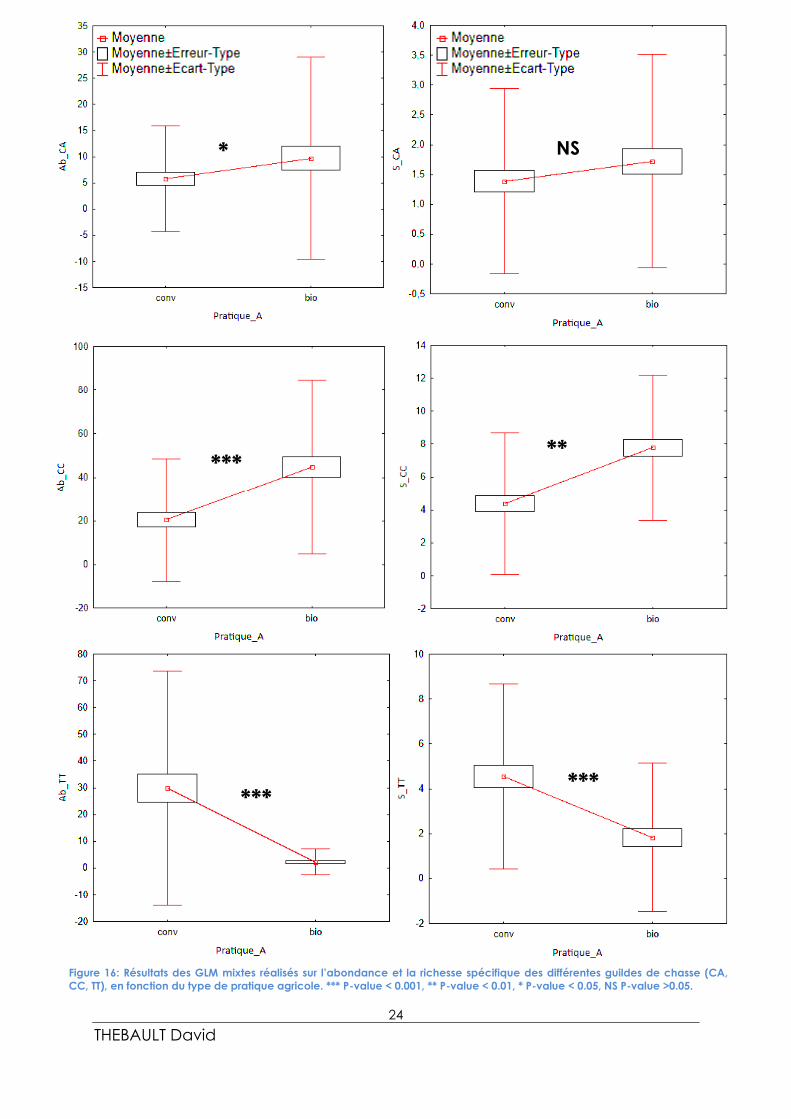
Suite à ces résultats, des GLMMs (Figure 16) ont été réalisées sur l’abondance et la richesse
spécifique afin d’étudier l’influence du type d’agriculture sur la composition du peuplement
d’araignées. Seules les données issues de la station en milieu de parcelle ont été conservées
pour les GLMMs. Les résultats issus des χ2 ci-dessus nous ont encouragés à continuer de
travailler pour cette sous-partie, et pour la partie III, par type de guilde de chasse et non pas
seulement sur l’ensemble de la population d’araignées.
Pour l’ensemble des GLMMs, les variables « taille de parcelle » et « pluviométrie » ont été
testées en co-variables explicatives, mais se sont révélées non significatives (P-value > 0,05).
Ainsi, seuls les modèles simples avec la variable explicative « type de pratique agricole » ont
été retenus.
Les GLMMs réalisés sur l’abondance et la richesse spécifique de l’ensemble de la population
d’araignées se sont révélés non significatifs (P-value > 0,05). Cependant, les GLMMs réalisées
sur les abondances et les richesses spécifiques des différentes guildes de chasse se sont
toutes révélées significatives (P-value < 0,05) excepté pour la richesse spécifique des
chasseuses à l’affût (P-value > 0,05).
L’abondance des chasseuses à l’affût est significativement supérieure en culture biologique
(P-value = 0.04264) qu’en culture conventionnelle. Il n’y a cependant pas de différence
significative au niveau de la richesse spécifique des chasseuses à l’affût (P-value > 0,05).
L’abondance des chasseuses coureuses est elle aussi significativement supérieure en culture
biologique (P-value = 2.2e-16), mais aussi leur richesse spécifique (P-value = 4.777e-05).
À l’inverse, l’abondance (P-value = 5.11e-10) et la richesse spécifique (P-value = 0.001323)
des tisseuses de toiles sont significativement inférieures en culture biologique.
24
THEBAULT David
Figure 16: Résultats des GLM mixtes réalisés sur l’abondance et la richesse spécifique des différentes guildes de chasse (CA,
CC, TT), en fonction du type de pratique agricole. *** P-value < 0.001, ** P-value < 0.01, * P-value < 0.05, NS P-value >0.05.
* NS
** ***
*** ***
G
G
G
GF
H
G
H
G
Hµ
***
**
µµ
25
THEBAULT David
2.3 Influence des variables paysagères sur les
communautés d’araignées
Pour finir, nous avons étudié l’effet du contexte paysager et de la position dans la parcelle sur
les communautés d’araignées. Nous avons testé individuellement l’effet de ces variables
paysagères sur : l’abondance totale, l’abondance par type de guilde de chasse (CA, CC,
TT), par richesse spécifique total, par richesse spécifique des différentes guildes de chasse, et
ceci dans un rayon de 500, 200, et 100m. L’ensemble des parcelles agricoles
(conventionnelles et biologiques) a été utilisé pour cette sous-partie.
On observe que la prise en compte des composantes du paysage permet d’expliquer une
partie de la déviance de la composition des communautés d’araignées (Tableau 3). L’effet
direct du paysage, sans interaction avec la position dans la parcelle, est plus souvent
significatif sur la population d’araignées dans le buffer le plus proche de la zone
d’échantillonnage, ici de 100m et avec 23 effets du paysage significatifs (Tableau3). À
l’inverse, l’effet direct du paysage, sans interaction avec la position dans la parcelle, tend à
être moins souvent significatif lorsqu’on augmente le rayon de l’étude : 200m (20) et 500m
(16) (Tableau 3).
Cependant, plus on tend à augmenter le rayon d’étude du contexte paysager autour des
zones d’échantillonnage et plus l’effet du contexte paysager, en interaction avec la position
dans la parcelle, est souvent significatif : 100m (21), 200m (29), et 500m (34) (Tableau 3).
Les effets du contexte paysager au sens global (avec ou sans interactions) ressortent plus
souvent significatifs pour un rayon de 500m, autour de la station échantillonnée, avec 50
effets significatifs (Tableau 3). Nous avons donc choisi de faire apparaitre prioritairement les
résultats de ce rayon (Tableau 4).
Tableau 3: Récapitulatif du nombre de variables paysagères significatives par buffer (500, 200,100m),
avec ou sans interactions, sur les variables Richesse spécifique et Abondances totales ou par guilde de
chasse.
Tout d’abord, on remarque que les variables paysagères ont nettement plus d’effets
significatifs sur les abondances (46) de la communauté d’araignées que sur leurs richesses
spécifiques (4), que ce soit pour l’ensemble des individus ou par guilde de chasse (Tableau
4).
Si l’on regarde l’abondance totale, on observe un effet négatif direct des variables
paysagères : bâtiment, mare, cours d’eau, haie, sur celle-ci et ceci quelle que soit la position
dans la parcelle. Les variables paysagères : culture et bande enherbée ont un effet positif
sur l’abondance totale en interaction avec la position dans la parcelle. En effet, ces effets
Buffer
Effets variables paysagères 500m 200m 100m
Paysage 16 20 23
Paysage : Position parcelle 34 29 21
Total 50 49 44

26
THEBAULT David
positifs sont plus élevés en bord de parcelle qu’en milieu de parcelle. Les variables
paysagères : prairie et boisement, ont elles un effet négatif sur l’abondance totale des
araignées en interaction avec la position dans la parcelle. En effet, ces effets négatifs sont
plus élevés en milieu de parcelle qu’en bord de parcelle.
L’abondance des chasseuses à l’affût, subit un effet direct du contexte paysager, et ce
quelle que soit la position dans la parcelle. Les variables paysagères boisement et haie
impactent positivement l’abondance des chasseuses à l’affût alors que les variables
bâtiment et cours d’eau l’impactent négativement. L’abondance des chasseuses à l’affût
subit également un effet des variables paysagères en interaction avec la position dans la
parcelle. Les variables paysagères : culture, mare et bande enherbée ont un effet positif en
bord de parcelle. Le milieu de parcelle de bande enherbée a également un effet positif. Les
effets de ces variables en bord de parcelles sont là aussi plus élevés. De plus, la variable
paysagère prairie impacte négativement l’abondance des chasseuses à l’affût et cela de
façon plus importante en milieu de parcelle.
L’abondance des chasseuses coureuses subit elle aussi un effet direct du contexte paysager,
que ce soit la position en bord ou milieu de parcelle. Les variables paysagères boisement et
haie impactent positivement l’abondance des chasseuses coureuses alors que les variables
bâtiment et cours d’eau l’impactent négativement. L’abondance des chasseuses coureuses
subit également un effet des variables paysagères en interaction avec la position dans la
parcelle. Les variables paysagères : culture, prairie et bande enherbée ont un effet positif en
bord de parcelle, et en milieu de parcelle, sauf le milieu de parcelle de prairie qui n’est pas
significatif. Les effets de ces variables en bord de parcelles sont là aussi plus élevés. De plus,
la variable paysagère mare impacte négativement l’abondance des chasseuses coureuses
et cela de façon plus importante en milieu de parcelle.
Pour finir avec les abondances d’araignées, nous nous attardons ici sur l’abondance des
tisseuses de toiles. On remarque que seule la variable paysagère bande enherbée a un effet
négatif direct, quelle que soit la position dans la parcelle, sur l’abondance des tisseuses de
toiles. L’abondance des tisseuses de toiles subit également un effet des variables paysagères
en interaction avec la position dans la parcelle. La variable paysagère culture a un effet
positif en bord de parcelle et en milieu de parcelle. L’effet de la culture est supérieur en bord
de parcelle. De plus, les variables paysagères : prairie, boisement, cours d’eau, haie, et
bâtiment impactent négativement l’abondance des tisseuses de toile et cela de façon plus
importante en milieu de parcelle, excepté pour la variable bâtiment qui est corrélée
positivement aux communautés d’araignées tisseuses de toiles en milieu de parcelle. De plus,
on remarque que la variable mare a bien un effet négatif sur les communautés d’araignées
tisseuses de toiles en milieu de parcelle, mais cet effet n’est pas significatif en bord de
parcelle.
Si l’on s’intéresse maintenant à la richesse spécifique totale des araignées, on remarque que
seule la variable paysagère cours d’eau en interaction avec la position milieu de la parcelle
a un effet significatif sur celle-ci. Cet effet de la variable paysagère cours d’eau est négatif.
La richesse spécifique des chasseuses à l’affût est impactée quant à elle négativement par
la variable mare et ceci quelle que soit la position dans la parcelle. La richesse spécifique des
chasseuses coureuses est impactée quant à elle négativement par la variable cours d’eau et
ceci quelle que soit la position dans la parcelle. Pour la richesse spécifique des tisseuses de
toiles, on remarque que seule la variable paysagère cours d’eau en interaction avec la
position milieu de la parcelle a un effet significatif sur celle-ci. Cet effet de la variable
paysagère cours d’eau est là aussi négatif.
27
THEBAULT David
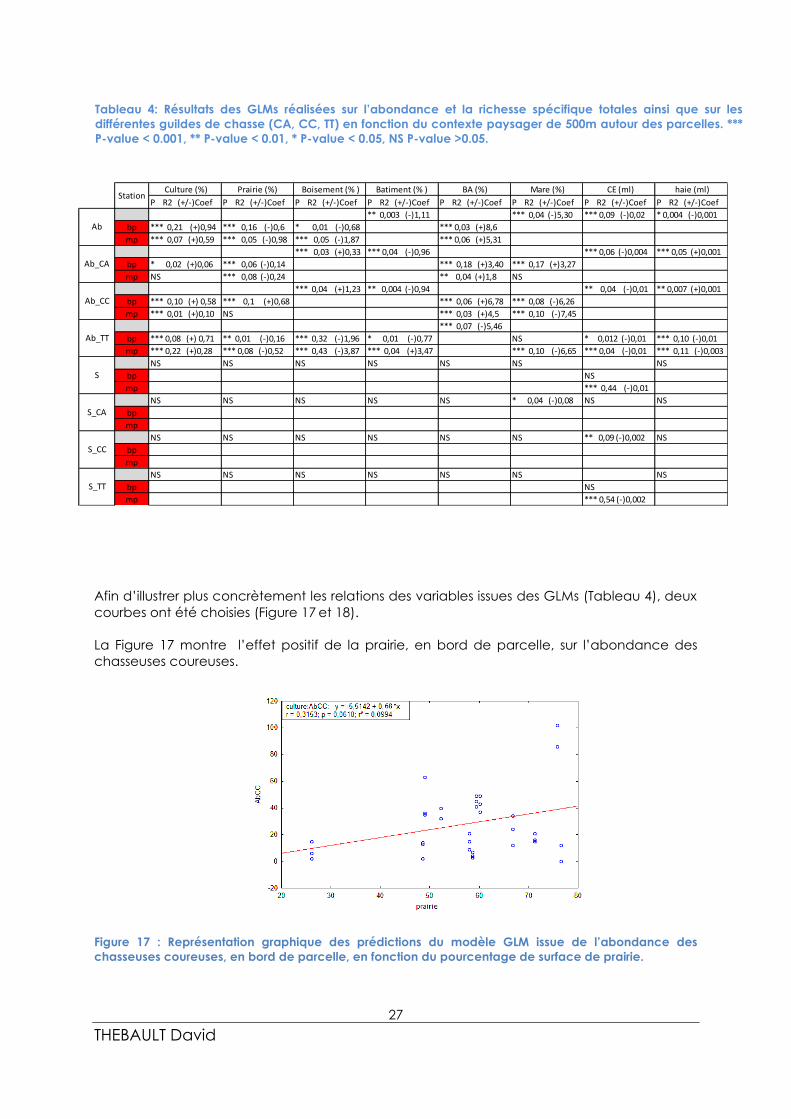
Afin d’illustrer plus concrètement les relations des variables issues des GLMs (Tableau 4), deux
courbes ont été choisies (Figure 17 et 18).
La Figure 17 montre l’effet positif de la prairie, en bord de parcelle, sur l’abondance des
chasseuses coureuses.
Figure 17 : Représentation graphique des prédictions du modèle GLM issue de l’abondance des
chasseuses coureuses, en bord de parcelle, en fonction du pourcentage de surface de prairie.
Tableau 4: Résultats des GLMs réalisées sur l’abondance et la richesse spécifique totales ainsi que sur les
différentes guildes de chasse (CA, CC, TT) en fonction du contexte paysager de 500m autour des parcelles. ***
P-value < 0.001, ** P-value < 0.01, * P-value < 0.05, NS P-value >0.05.
Culture (%) Prairie (%) Boisement (% ) Batiment (% ) BA (%) Mare (%) CE (ml) haie (ml)
P R2 (+/-)Coef P R2 (+/-)Coef P R2 (+/-)Coef P R2 (+/-)Coef P R2 (+/-)Coef P R2 (+/-)Coef P R2 (+/-)Coef P R2 (+/-)Coef
** 0,003 (-)1,11 *** 0,04 (-)5,30 *** 0,09 (-)0,02 * 0,004 (-)0,001
bp *** 0,21 (+)0,94 *** 0,16 (-)0,6 * 0,01 (-)0,68 *** 0,03 (+)8,6
mp *** 0,07 (+)0,59 *** 0,05 (-)0,98 *** 0,05 (-)1,87 *** 0,06 (+)5,31
*** 0,03 (+)0,33 *** 0,04 (-)0,96 *** 0,06 (-)0,004 *** 0,05 (+)0,001
bp * 0,02 (+)0,06 *** 0,06 (-)0,14 *** 0,18 (+)3,40 *** 0,17 (+)3,27
mp NS *** 0,08 (-)0,24 ** 0,04 (+)1,8 NS
*** 0,04 (+)1,23 ** 0,004 (-)0,94 ** 0,04 (-)0,01 ** 0,007 (+)0,001
bp *** 0,10 (+) 0,58 *** 0,1 (+)0,68 *** 0,06 (+)6,78 *** 0,08 (-)6,26
mp *** 0,01 (+)0,10 NS *** 0,03 (+)4,5 *** 0,10 (-)7,45
*** 0,07 (-)5,46
bp *** 0,08 (+) 0,71 ** 0,01 (-)0,16 *** 0,32 (-)1,96 * 0,01 (-)0,77 NS * 0,012 (-)0,01 *** 0,10 (-)0,01
mp *** 0,22 (+)0,28 *** 0,08 (-)0,52 *** 0,43 (-)3,87 *** 0,04 (+)3,47 *** 0,10 (-)6,65 *** 0,04 (-)0,01 *** 0,11 (-)0,003
NS NS NS NS NS NS NS
bp NS
mp *** 0,44 (-)0,01
NS NS NS NS NS * 0,04 (-)0,08 NS NS
bp
mp
NS NS NS NS NS NS ** 0,09 (-)0,002 NS
bp
mp
NS NS NS NS NS NS NS
bp NS
mp *** 0,54 (-)0,002
S_CA
S_CC
S
Ab_TT
Ab_CC
S_TT
Station
Ab_CA
Ab
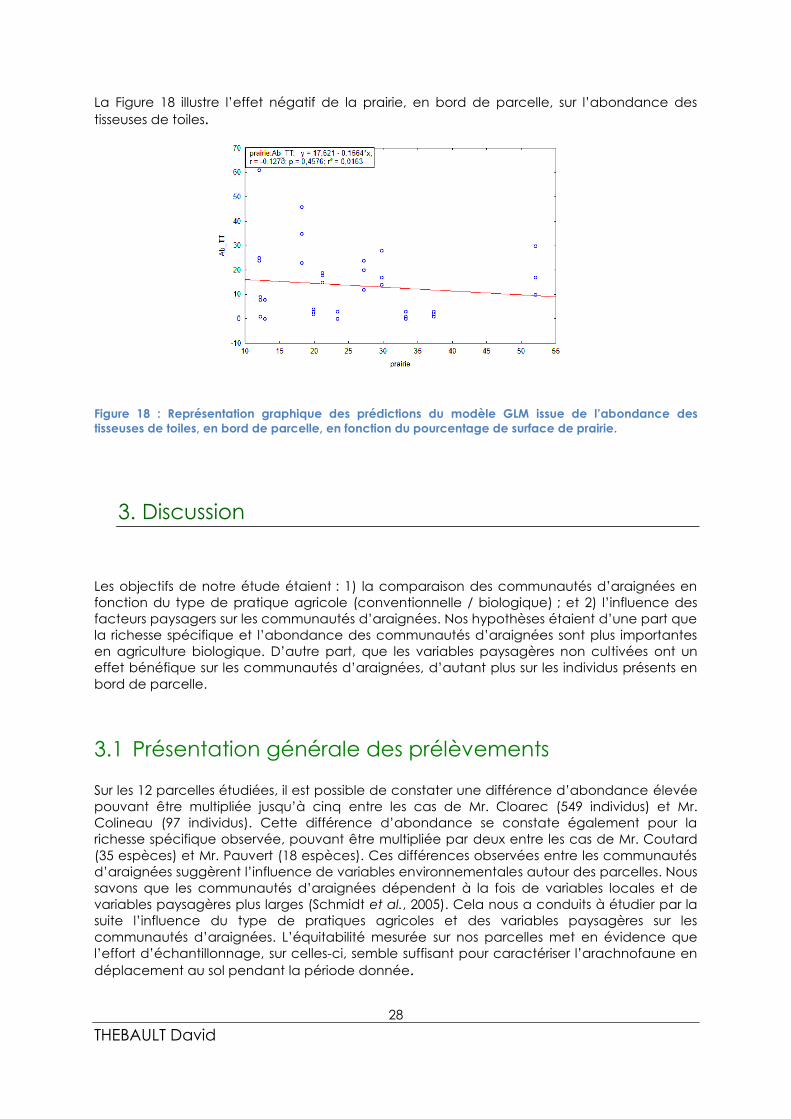
28
THEBAULT David
La Figure 18 illustre l’effet négatif de la prairie, en bord de parcelle, sur l’abondance des
tisseuses de toiles.
Figure 18 : Représentation graphique des prédictions du modèle GLM issue de l’abondance des
tisseuses de toiles, en bord de parcelle, en fonction du pourcentage de surface de prairie.
3. Discussion
Les objectifs de notre étude étaient : 1) la comparaison des communautés d’araignées en
fonction du type de pratique agricole (conventionnelle / biologique) ; et 2) l’influence des
facteurs paysagers sur les communautés d’araignées. Nos hypothèses étaient d’une part que
la richesse spécifique et l’abondance des communautés d’araignées sont plus importantes
en agriculture biologique. D’autre part, que les variables paysagères non cultivées ont un
effet bénéfique sur les communautés d’araignées, d’autant plus sur les individus présents en
bord de parcelle.
3.1 Présentation générale des prélèvements
Sur les 12 parcelles étudiées, il est possible de constater une différence d’abondance élevée
pouvant être multipliée jusqu’à cinq entre les cas de Mr. Cloarec (549 individus) et Mr.
Colineau (97 individus). Cette différence d’abondance se constate également pour la
richesse spécifique observée, pouvant être multipliée par deux entre les cas de Mr. Coutard
(35 espèces) et Mr. Pauvert (18 espèces). Ces différences observées entre les communautés
d’araignées suggèrent l’influence de variables environnementales autour des parcelles. Nous
savons que les communautés d’araignées dépendent à la fois de variables locales et de
variables paysagères plus larges (Schmidt et al., 2005). Cela nous a conduits à étudier par la
suite l’influence du type de pratiques agricoles et des variables paysagères sur les
communautés d’araignées. L’équitabilité mesurée sur nos parcelles met en évidence que
l’effort d’échantillonnage, sur celles-ci, semble suffisant pour caractériser l’arachnofaune en
déplacement au sol pendant la période donnée.

29
THEBAULT David
3.2 Comparaison des communautés d’araignées en
fonction du type de pratique agricole
(conventionnelle / biologique)
3.2.1 Comparaison des assemblages de communautés d’araignées
Premièrement, cette étude montre une grande cohérence des assemblages de
communautés d’araignées en fonction du type de pratique agricole. En effet, on constate
sur nos parcelles du Maine-et-Loire deux grands assemblages de communautés d’araignées
avec des espèces affiliées aux parcelles agricoles conventionnelles ou aux parcelles
agricoles biologiques. Cela suggère que les parcelles biologiques et conventionnelles sont
clairement différenciées les unes des autres sur le plan écologique et que ces différences
sont favorables à certaines espèces d’araignées et défavorables à d’autres. Nos premières
analyses (dendrogramme et AFC) ont tendu à montrer qu’en culture conventionnelle, on
retrouve essentiellement la famille des Linyphiidae et la guilde de chasse des tisseuses de
toiles ; et qu’en culture biologique, on retrouve en majorité la famille des Lycosidae et la
guilde de chasse des chasseuses coureuses.
Nous savons que la quasi-totalité des espèces indicatrices de la pratique agricole
conventionnelle sont des Linyphiidae tisseuses de toiles (6 sur 7). De plus, la majorité des
espèces indicatrices de la pratique agricole biologique sont des Lycosidae chasseuses
coureuses (3 sur 5). Ces espèces indicatrices viennent donc corroborer les premières
observations évoquées précédemment.
Seules les parcelles de Mr. Cogné et Mr. Bidault semblent avoir des assemblages de
communautés différentes aux autres parcelles.
Cette dissimilarité de la parcelle de Bidault P1 peut notamment s’expliquer par un
effort d’échantillonnage qui s’est révélé insuffisant sur cette dernière : l’assemblage
représenté sur cette parcelle étant ainsi considéré non représentatif de la réalité, car
incomplet.
En ce qui concerne la parcelle de Mr. Cogné, cela peut s’expliquer par le fait qu’il ait
répandu de l’insecticide sur la période de l’étude et donc l’assemblage d’araignées observé
ne correspond pas à ce que l’on aurait pu mettre en évidence (Prieto-Benitez & Méndez,
2010).
3.2.2 Comparaison de l’abondance et de la richesse spécifique des communautés
d’araignées
Nous avons établi dans notre étude qu’il n’y a pas de différence significative, au
niveau des proportions d’effectifs totaux des araignées, entre la culture conventionnelle et
biologique. Cependant, après avoir choisi de regrouper les individus par type de chasse,
nous avons mis en évidence des différences significatives, au niveau de leurs proportions
d’effectifs respectifs. Ceci pour les trois guildes de chasse : chasseuses à l’affût, chasseuses
coureuses et tisseuses de toiles. Ces résultats nous ont confortés dans notre idée de comparer
l’abondance et la richesse spécifique des communautés d’araignées par guilde de chasse
et non pas seulement sur la totalité des données. Contrairement à ce qui est observé dans la
littérature, la famille des Lycosidae domine nos échantillons suivis des Linyphiidae. En effet,
traditionnellement on observe dans les cultures d’Europe centrale une majorité de
Linyphiidae puis des Lycosidae (Sunderland, 1999). Cette sous-représentation des Linyphiidae

30
THEBAULT David
peut être liée à un sous-échantillonnage dû à un biais humain ou au fait que lors des
échantillonnages cette famille n’ait pas encore atteint son maximum démographique.
Aucune différence significative d’abondance et de richesse spécifique, sur
l’ensemble de la population d’araignées, en fonction du type d’agriculture n’a été mise en
évidence. Cela ne correspond pas avec ce qui est vu dans la littérature et qui tend à
montrer qu’il y ait plus d’abondance et de richesse spécifique dans les parcelles biologiques
(Schmidt et al., 2005 ; Clough et al., 2005). Cependant, actuellement de nombreuses études
semblent montrer qu’il n’y ait pas toujours de différences au niveau de l’abondance et de la
richesse spécifique totale entre les parcelles biologiques et conventionnelles (Prieto-Benitez &
Méndez, 2010). Cela peut s’expliquer par le fait d’une réplication insuffisante des unités
d’échantillonnage ou d’un manque de compatibilité entre les parcelles biologique et
conventionnelle. En effet, dans certains cas les parcelles biologiques étaient bordées pour la
plupart de terres gérées de manière conventionnelle (Schmidt et al., 2005a). De plus,
l’absence de différence significative de l’abondance et de la richesse spécifique totale,
entre les cultures biologiques et conventionnelles, peut s’expliquer par le fait que l’efficacité
d’un type d’agriculture dépend en grande partie de la complexité du paysage ou plus
précisément du pourcentage élevé de surface non cultivées et non urbanisées (Concepción
et al., 2008). Nos hypothèses sont donc : soit les parcelles biologiques étudiées ont un
paysage environnant avec une surface non cultivée insuffisante, soit les parcelles
conventionnelles étudiées ont un paysage environnant avec une surface non cultivée
suffisante pour atténuer la différence de richesse spécifique et d’abondance de l’ensemble
de la communauté d’araignées entre cultures biologiques et conventionnelles. C’est la
deuxième hypothèse qui est validée ici.
Des différences significatives, entre les cultures biologiques et conventionnelles, ont
été mises en évidence pour la richesse spécifique et l’abondance des guildes de chasse.
L’abondance des chasseuses à l’affût, ainsi que l’abondance et la richesse spécifique des
chasseuses coureuses sont significativement plus importantes en culture biologique. Ces
résultats sont en adéquation avec ceux trouvés dans l’étude de Feber et al. (1998). L’effectif
des chasseuses à l’affût étant plus limité dans notre étude par rapport aux autres guildes, ces
résultats sont à prendre avec précautions. Cette plus grande abondance et richesse
spécifique des chasseuses à l’affût et des coureuses s’explique par : premièrement le fait que
les insecticides sont absents en culture biologique, les populations d’araignées ne subissent
donc pas de mortalité liée à ce facteur (Pekar & Kocourek, 2004 in Prieto-Benitez & Méndez,
2010). Cependant, Laster et Brazzel (1968) ont montré que les araignées avaient une
tolérance aux pesticides et qu’elles étaient moins sensibles aux insecticides que certains
autres insectes prédateurs comme les coccinelles.
Deuxièmement, les cultures biologiques ont une plus grande couverture et diversité
d’adventices (« mauvaises herbes »). Cela a pour effet de fournir aux populations
d’araignées une plus grande complexité structurelle et de lieux où se cacher à la surface du
sol (Sunderland & Samu, 2000).
Troisièmement, les champs biologiques reçoivent de plus importants apports de fumier
et ont une rotation des cultures plus complexe permettant d’améliorer la qualité du sol. Cela
peut bénéficier aux araignées en augmentant la disponibilité des insectes tels que les
collemboles, ou les moucherons. Ces proies supplémentaires sont d’autant plus importantes
que dans les cultures de blé d’hiver, surtout au printemps, la quantité de proies est un facteur
limitant (Harwood et al., 2001).
Il a également été montré que l’abondance et la richesse spécifique des tisseuses de
toiles ne sont pas significativement plus importantes en culture biologique. C’est cohérent
avec les résultats trouvés par Feber et al. (1998). Cependant, il a également été montré dans
notre étude une abondance et une richesse spécifique significativement plus importante en
culture conventionnelle pour les tisseuses de toiles. Ce serait dû au fait que bien que le milieu
en culture biologique soit globalement plus favorable aux araignées tisseuses de toile, ces

31
THEBAULT David
dernières, par leur mode de déplacement aérien aléatoire (ou « ballooning ») sont
incapables de s’orienter en direction des cultures biologiques. De plus en Maine-et-Loire, il
n’y a que 8 % de cultures biologiques contre 92 % de cultures conventionnelles. Cette
tendance est également observée en France. Cette faible proportion de cultures
biologiques rend la tâche d’autant plus compliquée pour les araignées tisseuses de toiles, car
il y aura proportionnellement moins de chance pour ces araignées d’arriver jusqu’aux
cultures biologiques. Une autre raison pouvant expliquer la plus faible abondance et richesse
spécifique des araignées tisseuses de toiles est que les cultures biologiques se trouvent
majoritairement dans des contextes bocagers, et les haies sont un obstacle important au
mode de déplacement aérien passif des araignées tisseuses de toiles. Une dernière raison qui
expliquerait cette plus faible abondance et richesse spécifique des tisseuses de toiles en
culture biologique serait liée au travail du sol plus fréquent en culture biologique (Everts et al.,
1989 in Clough et al., 2007). En effet, en culture biologique, du fait de l’absence de
pesticides, les agriculteurs doivent réaliser un travail du sol plus fréquent afin du lutter contre
le « salissement » des parcelles les adventices. Les araignées tisseuses de toiles auront
beaucoup de difficultés à éviter ces perturbations répétées, car leur mode de déplacement
aléatoire n’est par définition pas prévisible. Elles sont donc soumises aux conditions
climatiques du moment.
3.3 Influence des variables paysagères sur les
communautés d’araignées
Nos résultats révèlent que les effets du paysage (ou des variables paysagères) sont
visibles sur les abondances mais pas sur les richesses spécifiques des araignées, que ce soit sur
l’ensemble des araignées ou par guildes de chasse. C’est cohérent avec ce que nous
pensions et cela s’explique par la nature même de la variable « richesse spécifique ». En
effet, il suffit d’avoir un individu pour être comptabilisé comme une espèce, ce qui tend à
réduire l’information et à limiter la variation dans le jeu de données. La conséquence est
d’avoir moins de probabilité d’avoir une corrélation significative de celle-ci avec les variables
paysagères, d’autant plus que le nombre d’espèces d’araignées en milieu agricole reste
modeste et en général ne dépasse que rarement les 120 espèces (Prieto-Benitez & Méndez,
2010).
La réponse des araignées face au paysage est comprise jusqu’à 500m autour de la
zone d’étude. Clough et al. (2007) ont montré que l’effet du contexte paysager sur les
communautés d’araignées pouvait aller jusqu’à 3 km selon les espèces. Notre étude a
également mis en évidence que l’effet du paysage est surtout marqué quand les guildes de
chasse sont considérées, d’où l’importance d’utiliser cette approche pour cette étude. De
plus, il a été prouvé qu’il y a de meilleures réponses des araignées au contexte paysager
dans un buffer de 500m. Dans ce buffer, la plupart des réponses des araignées au contexte
paysager diffèrent selon la position dans la parcelle. En effet, on remarque qu’une réponse
positive des communautés d’araignées, ici une augmentation de leur abondance, face à
l’augmentation d’une variable paysagère sera plus marquée en bord de parcelle et tendra
à s’atténuer en milieu de parcelle. Ce phénomène peut s’expliquer par le fait que toutes les
espèces ne peuvent se déplacer jusqu’au centre de la parcelle. Par opposition, une réponse
négative des communautés d’araignées, ici une baisse de leur abondance, face à
l’augmentation de la variable paysagère sera encore plus marquée en milieu de parcelle
qu’en bord de parcelle, car l’effet limitant de la capacité de dispersion peut accroitre cet
effet négatif déjà marqué en bord de parcelle. Cependant, il est important de signaler qu’il y
a peu de différences significatives observées entre les bords et milieux de parcelles. Au final,
contrairement aux hypothèses précédentes cela illustre donc une grande capacité de
dispersion des araignées et pour les trois types de chasses. Dans le buffer de 100m la réponse

32
THEBAULT David
des communautés d’araignées au contexte paysager est la même quelle que soit la position
dans la parcelle.
On a également remarqué un effet positif des cultures sur l’abondance des
chasseuses coureuses et des tisseuses de toiles. C’est inattendu et cela témoigne de
l’importance d’habitats similaires, bien que non pérennes (ex : prairies), à proximité de nos
cultures.
Nos résultats montrent que l’abondance des chasseuses coureuses, en grande
majorité représentées par les Lycosidae, est corrélée positivement à une majorité des
surfaces paysagères non cultivées (prairies, bois, bandes enherbées, et haies) et donc
corrélées positivement au pourcentage de surfaces non cultivées dans le paysage
environnant la zone d’étude. Ces résultats sont en accord avec d’autres publications
(Clough et al., 2007). De fait, les paysages moins dominés par les terres arables ont des
habitats plus pérennes. Ces habitats pérennes non cultivés semblent donc particulièrement
importants pour les espèces d’araignées de type chasseuses coureuses. Ceux-ci peuvent
non seulement servir de lieux pour passer l’hiver (abri contre les prédateurs, reproduction, ou
lieux de repos) en raison d’une perturbation réduite, mais sont également plus diversifiés en
espèces et ont des ressources alimentaires plus prévisibles (Sunderland, 1999). Il semble que
les Lycosidae utilisent donc les cultures de manière facultative pour chasser et chassent
prioritairement dans les habitats non cultivés et les écotones comme les bandes enherbées
ou les bords de parcelles (Sunderland & Samu, 2000).
À l’inverse, nous avons mis en évidence que l’abondance des tisseuses de toiles, en
grande majorité représentées par les Linyphiidae, est corrélée négativement à l’ensemble
des surfaces paysagères non cultivées (prairie, bois, bandes enherbées, et haies). Ces
résultats sont également en accord avec ce qui est vu dans la littérature (Clough et al.,
2007). Cela tend à montrer que pour les Linyphiidae les habitats plus pérennes sont non
seulement pas importants, mais défavorables. Ces résultats peuvent s’expliquer : d’une part,
par le fait que les habitats non cultivés sont, de par la nature passive du mode de dispersion
des Linyphiidae, un frein à leur circulation. Cela est d’autant plus impactant dans le cas de
bois et de haies. Il semble que cette famille utilise prioritairement les cultures pour chasser,
contrairement aux Lycosidae. D’autre part, l’effet négatif des prairies sur la population de
Linyphiidae peut s’expliquer par la compétition inter-guilde de chasse et même inter-
spécifique. En effet, nous savons que les prairies ont un effet positif sur la population de
Lycosidae et que ces dernières tendent à évoluer dans ce type de milieu. Une compétition
entre les Linyphiidae et les Lycosidae est donc envisageable, d’autant qu’il est établi que les
principaux ennemis naturels des araignées sont d’autres araignées (Wise, 1993 in Schmidt et
al., 2005b).
Il a également été montré que les variables paysagères semblent avoir globalement
un effet négatif sur l’abondance total des communautés d’araignées.
Maintenant qu’il a été démontré que le contexte paysager impacte les
communautés d’araignées nous pouvons supposer qu’il explique en partie la répartition des
Lycosidae chasseuses coureuses et/ou des Linyphiidae tisseuses de toiles en fonction des
cultures biologiques et conventionnelles. En effet, la présence importante des éléments
paysagers corrélés positivement pour les Lycosidae (ici les boisements, bandes enherbées,
haies) et/ou les éléments paysagers corrélés négativement pour les Linyphiidae (ici les
prairies, haies) autour des parcelles biologiques peuvent expliquer la répartition observée.
Après vérification sur les données cartographiques SIG, cette hypothèse est en partie validée.
En effet, il semble y avoir plus de prairies et de haies en culture biologique, favorisant ainsi les
Lycosidae et à l’inverse défavorisant les Linyphiidae.

33
THEBAULT David
4. Conclusion
Nous avons mis en évidence que la richesse spécifique et l’abondance des communautés
d’araignées sont plus importantes en agriculture biologique lorsque l’on compare par guilde
de chasse, excepté pour les tisseuses de toiles. L’hypothèse de notre premier objectif est
donc en partie validée. Concernant le second objectif, nous avons à la fois démontré un
effet bénéfique des variables paysagères non cultivées sur les araignées chasseuses
coureuses et ceci d’autant plus sur les individus présents en bord de parcelle, mais nous
avons également mis en évidence un effet négatif des variables paysagères non cultivées et
ceci est d’autant plus accentué en milieu de parcelle. La seconde hypothèse est donc là
aussi en partie validée. Cette étude a mis en évidence la grande disparité existant au sein
des araignées et l’importance de ne pas se contenter d’une approche uniquement
spécifique mais aussi de travailler à l’échelle des groupes fonctionnels, tels que les guildes de
chasse, qui permettent d’avoir des résultats plus probants.
Favoriser le maintien de l’abondance et la richesse en espèce des araignées dans les
agroécosystèmes a pour conséquence de favoriser le maintien de leur rôle d’auxiliaire de
cultures, donc de favoriser la régulation des ravageurs de cultures et ainsi indirectement de
contribuer à la productivité agricole. La gestion des communautés d’araignées doit se faire
à l’échelle locale, mais également à l’échelle paysagère. Cela implique que les mesures de
gestion devraient inclure la connaissance de leurs exigences spatiales.
Suite à ce travail, il serait intéressant de poursuivre l’étude et de traiter le reste des données
recueillies sur le terrain afin de valider ou non ces conclusions. L’étude de la variable travail
du sol sera particulièrement intéressante du fait que très peu d’études ont été réalisées sur ce
sujet. De plus, il serait intéressant d’intégrer à ces analyses des variables complémentaires,
pouvant également impacter les communautés d’araignées (Clough et al., 2007) :
l’historique cultural des parcelles, les taux de pesticides dans les cultures conventionnelles et
leur fréquence, ainsi que le pourcentage de connectivité entre les habitats.
Ce stage a été complet et formateur. J’ai eu à la fois énormément de terrain, de rencontres
et de discussions avec des acteurs du monde agricole, ainsi qu’un travail rédactionnel et
analytique important. Pour cette dernière partie, j’ai été amené à utiliser des outils statistiques
et cartographiques. Les acteurs du monde agricole que j’ai rencontrés étaient d’une grande
diversité : élus, directeurs de pôle et autres personnels de la chambre, exploitants agricoles :
et m’ont permis d’avoir une vision globale et cohérente de ce milieu. De plus, ce stage m’a
permis de compléter mes connaissances naturalistes, via mon travail réalisé sur le terrain
(OAB et étude sur les araignées) et mes échanges avec les associations naturalistes et
l’université de Rennes 1.

34
THEBAULT David
Bibliographie
Cardoso, P., Erwin, T.L., Borges, P.A.V., New, T.R., 2011. The seven impediments in invertebrate
conservation and how to overcome them. Biological Conservation, 144, 2647–2655.
Clough, Y., Kruess, A., Kleijn, D., Tscharntke, T., 2005. Spider diversity in cereal fields: comparing
factors at local, landscape and regional scales. Journal of Biology, 32, 2007–2014.
Clough, Y., Holzschuh, A., Gabriel, D., Purtauf, T., Kleijn, D., Kruess, A., Steffan-Dewenter, I.,
Tscharntke, T., 2007. Alpha and beta diversity of arthropods and plants in organically and
conventionally managed wheat fields. Journal of Applied Ecology 44, 804–812.
Concepcion, E., Dıaz, M., Baquero, R., 2008. Effects of landscape complexity on the
ecological effectiveness of agri-environment schemes. Landscape Ecology, 23, 135–148.
Feber, R.E., Bell, J., Johnson, P.J., Firbank, L.G., Macdonald, D.W., 1998. The effects of organic
farming on surface-active spider (Araneae) assemblages in wheat in southern England (UK).
Journal of Arachnology, 26, 190 –202.
Harwood, J.D., Sunderland, K.D., Symondson, W.O.C., 2001. Living where the food is: web
location by linyphiid spiders in relation to prey availability in winter wheat. Journal of Applied
Ecology, 38, 88–99.
Hole, D.G., Perkins, A.J., Wilson, J.D., Alexander, I.H., Grice, P.V., Evans, A.D., 2005. Does
organic farming benefit biodiversity? Biological Conservation, 122, 113–130.
Knapp, M., Ruzicka, J., 2012. The effect of pitfall trap construction and preservative on catch
size, species richness and species composition of ground beetles (Coleoptera: Carabidae).
European Journal of Entomology, 109, 419–426.
Krebs, C.J., 1989. Ecological methods. Harper & Row Publishers, New York., 654 pp.
Lafage, D., Petillon, J., « En préparation ». Disentangling the influence of local and landscape
factors on alpha and beta diversities : opposite response of plants and ground-dwelling
arthropods in wet meadows.
Lang, A., Filser, J., Henschel, J.R., 1999. Predation by ground beetles and wolf spiders on
herbivorous insects in a maize crop. Agriculture Ecosystems and Environment, 72, 189–199.
Laster, M., Brazzel, J.R., 1968. A comparison of predator populations in cotton under different
control programs in Mississippi. J. Economical of Entomology, 61, 714–719.
Le Roux, X., Barbault, R., Baudry, J., Burel, F., Doussan, I., Garnier, E., 2008. Agriculture et
Biodiversité. Valoriser les Synergies. Expertise scientifique collective, synthèse du rapport, INRA,
France.
Luczak, J., 1979. Spiders in agrocoenoses. Polish Ecological Studies, 5, 151–200.
Peeters, A., Malijean, J.F., Biala, K., Brouckaert, V., 2004. Les indicateurs de biodiversité pour
les prairies : un outil d’évaluation de la durabilité des systèmes d’élevage. Revue fourrages,
178, 217-232.
Prieto-Benítez, S., Méndez, M., 2010. Effects of land management on the abundance and
richness of spiders (Araneae): a meta-analysis. Biological Conservation, 144, 683–691.

35
THEBAULT David
Riechert, S.E., Luczak, J., 1982. Spider foraging: behavioural response to prey. Spider
Communication. Princeton University Press, 353–384.
Samu, F., Sunderland, K.D., Szinetár, C., 1999. Scale-dependent dispersal and distribution
patterns of spiders in agricultural systems. Journal of Arachnology, 27, 325–332.
Schmidt, M.H., Roschewitz, I., Thies, C., Tscharntke, T., 2005. Differential effects of
landscape and management on diversity and density of ground-dwelling farmland spiders.
Journal of Applied Ecology, 42, 281–287.
Schmidt, M.H., Tscharntke, T., 2005a. The role of perennial habitats for Central European
farmland spiders. Agriculture, Ecosystems and Environment, 105, 235–242.
Schmidt, M.H., Tscharntke, T., 2005b. Landscape context of sheetweb spider (Araneae:
Linyphiidae) abundance in cereal fields. Journal of Biogeography, 32, 467–473.
Sunderland, K., Samu, F., 2000. Effects of agricultural diversification on the abundance,
distribution, and pest control potential of spiders: a review. Entomologia Experimentalis and
Applicata, 95, 1–13.
Tscharntke, T., Klein, A.M., Kruess, A., Steffan-Dewenter, I., Thies, C., 2005. Landscape
perspectives on agricultural intensification and biodiversity ecosystem service management.
Ecology Letters, 8, 857–874.
Uetz, G. W., Halaj, J., Cady, A. B., 1999. Guild structure of spiders in major crops. Journal of
Arachnology, 27, 270–280.
Sites web consultés
Chambre d’agriculture du Maine-et-Loire. [En ligne]. « http://www.maine-et-
loire.chambagri.fr/ » (Page consultée le 2 juin 2004).
Météo Bell. [En ligne]. http://www.meteobell.com/__anjou_geomorphologie.php (Page
consultée le 20 Juin 2014)

A
THEBAULT David
Annexes
Annexe 1 : Descriptions des quatre protocoles de l’OAB

B
THEBAULT David

C
THEBAULT David

D
THEBAULT David
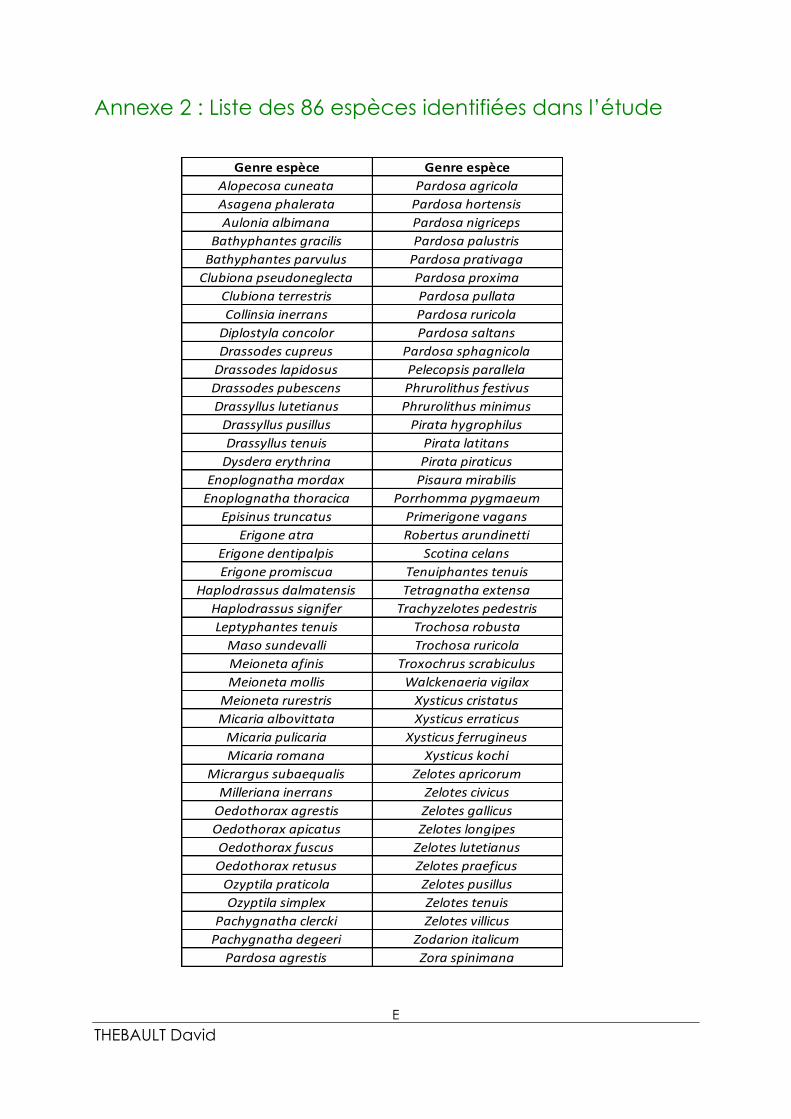
E
THEBAULT David
Annexe 2 : Liste des 86 espèces identifiées dans l’étude
Genre espèce Genre espèce
Alopecosa cuneata Pardosa agricola
Asagena phalerata Pardosa hortensis
Aulonia albimana Pardosa nigriceps
Bathyphantes gracilis Pardosa palustris
Bathyphantes parvulus Pardosa prativaga
Clubiona pseudoneglecta Pardosa proxima
Clubiona terrestris Pardosa pullata
Collinsia inerrans Pardosa ruricola
Diplostyla concolor Pardosa saltans
Drassodes cupreus Pardosa sphagnicola
Drassodes lapidosus Pelecopsis parallela
Drassodes pubescens Phrurolithus festivus
Drassyllus lutetianus Phrurolithus minimus
Drassyllus pusillus Pirata hygrophilus
Drassyllus tenuis Pirata latitans
Dysdera erythrina Pirata piraticus
Enoplognatha mordax Pisaura mirabilis
Enoplognatha thoracica Porrhomma pygmaeum
Episinus truncatus Primerigone vagans
Erigone atra Robertus arundinetti
Erigone dentipalpis Scotina celans
Erigone promiscua Tenuiphantes tenuis
Haplodrassus dalmatensis Tetragnatha extensa
Haplodrassus signifer Trachyzelotes pedestris
Leptyphantes tenuis Trochosa robusta
Maso sundevalli Trochosa ruricola
Meioneta afinis Troxochrus scrabiculus
Meioneta mollis Walckenaeria vigilax
Meioneta rurestris Xysticus cristatus
Micaria albovittata Xysticus erraticus
Micaria pulicaria Xysticus ferrugineus
Micaria romana Xysticus kochi
Micrargus subaequalis Zelotes apricorum
Milleriana inerrans Zelotes civicus
Oedothorax agrestis Zelotes gallicus
Oedothorax apicatus Zelotes longipes
Oedothorax fuscus Zelotes lutetianus
Oedothorax retusus Zelotes praeficus
Ozyptila praticola Zelotes pusillus
Ozyptila simplex Zelotes tenuis
Pachygnatha clercki Zelotes villicus
Pachygnatha degeeri Zodarion italicum
Pardosa agrestis Zora spinimana

Résumé
Etude de la biodiversité fonctionnelle dans des agroécosystèmes du Maine-et-Loire
La transformation des terres en agroécosystèmes a souvent eu pour conséquence une
simplification du paysage et une baisse de la biodiversité. La biodiversité fonctionnelle joue un rôle
particulièrement important dans le fonctionnement de l’agroécosystème. Cette étude s'inscrit
dans le contexte de l’observation de la biodiversité fonctionnelle dans des agroécosystèmes du
Maine-et-Loire, à différentes échelles taxonomiques. La première partie a été le déploiement de
l’Observatoire Agricole de la Biodiversité (OAB). La seconde partie a été l’étude de l’impact des
pratiques agricoles, et de la structuration du paysage sur les communautés d’araignées dans des
agroécosystèmes. Notre étude met en évidence deux grands assemblages de communautés
d’araignées avec des espèces affiliées aux parcelles agricoles conventionnelles (Linyphiidae) ou
aux parcelles agricoles biologiques (Lycosidae). Aucune différence significative d’abondance et
de richesse spécifique, sur l’ensemble de la population d’araignées, n’a été mise en évidence en
fonction du type d’agriculture mais des différences significatives ont été mises en évidence
lorsque l’on travail par guildes de chasse. En effet, il y a plus de Chasseuses Coureuses (CC) et à
l’affût (CA) en agriculture biologique mais à l’inverse moins de Tisseuses de Toiles (TT). Nous avons
également mis en évidence un impact du contexte paysager sur l’abondance des communautés
d’araignées mais pas sur leur richesse spécifique. De plus, il a été prouvé qu’il y a de meilleures
réponses des araignées au contexte paysager dans un buffer de 500m. L’augmentation de la
surface de variables paysagères non cultivés a un effet positif sur l’abondance des chasseuses
coureuses et des chasseuses à l’affût mais négatif sur l’abondance des tisseuses de toiles et
l’abondance totale. Cette étude a mis en évidence la grande disparité existant au sein des
araignées et l’importance de les étudier non pas seulement par une approche spécifique, mais
par groupes fonctionnelles, notamment par guildes de chasses. De plus, que la gestion des
communautés d’araignées doit se faire à l’échelle locale, mais également à l’échelle paysagère.
Mots-clés :
Agriculture, biodiversité, Araneae, blé d'hiver, gestion locale, paysage.
Abstract
Study of functional biodiversity in agroecosystems in Maine-et-Loire
Land transformation in agroecosystems has often led to a simplification of the landscape and a
decline of biodiversity. Functional biodiversity plays a particularly important role in the functioning
of the agro-ecosystem. The purpose of this study is the observation of functional biodiversity in
agroecosystems in Maine-et-Loire, at different taxonomic levels. The first part was the deployment
of the Agricultural Biodiversity Observatory (ABO). The second part was the study of the impact of
agricultural practices and the structuring of the landscape on spider communities in
agroecosystems. Our study highlights two major assemblies spider communities affiliated with
conventional agricultural plots (Linyphiidae) or organic agricultural plots (Lycosidae) species. No
significant difference in abundance and species richness, on the total population of spiders, has
been demonstrated in the type of agriculture but significant differences were highlighted when
working by hunting guilds. Indeed, there are more hunters runners (CC) and hunters on the lookout
(CA) in organic farming but conversely less sheetweb (TT). We have also demonstrated an impact
of landscape context on the abundance of spider communities but not on their species richness.
In addition, it has been proven that there are better answers of spiders to the landscape context in
a buffer of 500m around the study site. Increasing the surface of uncultivated landscape variables
has a positive effect on the abundance of hunters runners and hunters on the lookout but
negative on the abundance of sheetweb and total abundance. This study has highlighted the
great disparity in the spiders and the importance of studying not only by a specific approach, but
by functional groups, including hunts guilds. In addition, the management of spider communities
should be at the local level, but also in the wider landscape.
Keywords :
Agriculture, biodiversity, Araneae, winter wheat, local management, landscape.



![[Archive- Laval 2006] Rapport de stage de Master - Nautilus](https://static.fdocuments.fr/doc/165x107/55b4ed83bb61eb362d8b45d1/archive-laval-2006-rapport-de-stage-de-master-nautilus.jpg)
![Stéphanie LÉVÊQUE Master DEFI RAPPORT de STAGE · Stéphanie LÉVÊQUE Master DEFI RAPPORT de STAGE [stage effectué du 30 mars au 30 juin 2015] Centre International de Recherche](https://static.fdocuments.fr/doc/165x107/5b9bee7209d3f221608c3f21/stephanie-leveque-master-defi-rapport-de-stephanie-leveque-master-defi.jpg)













