
Laboratoire de Génétique COURS MAGISTAL ...
46
REPUBLIQUE DE COTE D’IVOIRE MINISTERE DE L’ENSEIGNEMENT SUPERIEUR UNION – DISCIPLINE – TRAVAIL ET DE LA RECHERCHERCHE SCIENTIFIQUE 01 BP V 34 Abidjan 01 - Tel./Fax : 22 44 35 31 22 BP 582 Abidjan 22 - Tél./Fax : 22 44 44 73 www.univ-cocody-ci [email protected] Laboratoire de Génétique COURS MAGISTAL -------------------- UE : GENETIQUE FORMELLE ----------------- ECUE 1 : GENETIQUE FORMELLE CHEZ LES ORGANISMES HAPLOIDES
Transcript of Laboratoire de Génétique COURS MAGISTAL ...

REPUBLIQUE DE COTE D’IVOIRE MINISTERE DE L’ENSEIGNEMENT SUPERIEUR UNION – DISCIPLINE – TRAVAIL ET DE LA RECHERCHERCHE SCIENTIFIQUE
01 BP V 34 Abidjan 01 - Tel./Fax : 22 44 35 31 22 BP 582 Abidjan 22 - Tél./Fax : 22 44 44 73 www.univ-cocody-ci [email protected]
Laboratoire de Génétique
COURS MAGISTAL --------------------
UE : GENETIQUE FORMELLE
-----------------
ECUE 1 : GENETIQUE FORMELLE CHEZ LES
ORGANISMES HAPLOIDES

2
TABLE DES MATIERES
PREMIERE PARTIE : BASES FONDAMENTALES POUR L’ANALYSE GENETIQUE CHEZ LES
ORGANISMES HAPLOIDES ................................................................................................................. 3
CHAPITRE I : CYCLE DE REPRODUCTION, CYCLES CHROMOSOMIQUES, MECANISME DE LA
MEIOSE ET CONSEQUENCES GENETIQUES DE LA MEIOSE ..................................................... 4
1- LE CYCLE DE REPRODUCTION ................................................................................................ 4 2- LES CYCLES CHROMOSOMIQUES .......................................................................................... 5 3- LA FECONDATION .................................................................................................................. 6 4- LA MEIOSE .............................................................................................................................. 6 5- LES CONSEQUENCES GENETIQUES DE LA MEIOSE .............................................................. 10 6- CONCLUSION ........................................................................................................................ 16
CHAPITRE II – EXEMPLES DE CYCLES DE REPRODUCTION CARCTERISTIQUES DES
ASCOMYCETES.................................................................................................................... 17
1- LE CYCLE de Neurospora crassa (la moisissure du pain) .................................................... 17 2- LE CYCLE de Saccharomyces cerevisiae (la levure de boulangerie) .................................. 18 3- ELEMENTS DE REFERENCES POUR l’ANALYSE GENETIQUE ................................................ 19
DEUXIEME PARTIE : TRANSMISSION DES CARACTERES HEREDITAIRES CHEZ LES ORGANISMES
HAPLOIDES ....................................................................................................................................... 20
CHAPITRE I : SEGREGATION D'UN COUPLE D'ALLELES .......................................................... 21
1- Cas d'un organisme à tétrades ordonnés : Neurospora crassa ........................................... 21 2- Cas d'un organisme à tétrades non ordonnés : Ascobolus immersus ................................. 25 3- Analyse de spores en vrac ................................................................................................... 25
CHAPITRE II : SEGREGATION DE DEUX COUPLES D'ALLELES LOCALISES SUR 2 PAIRES DE
CHROMOSOMES DIFFERENTS ............................................................................................. 26
1- Cas d'un organisme à tétrades non ordonnées : Ascobolus ................................................ 26 2- Cas d'un organisme à tétrades ordonnées : Neurospora crassa ......................................... 30 3- Analyse de spores en vrac ................................................................................................... 32
CHAPITRE III - SEGREGATION DE DEUX COUPLES D'ALLELES LOCALISES SUR LA MEME PAIRE
DE CHROMOSOMES ............................................................................................................ 34
1- Cas des organismes à tétrades non ordonnés ................................................................... 34 2- Cas des organismes à tétrades ordonnées .......................................................................... 39 3- Cas de spores en vrac .......................................................................................................... 39
CHAPITRE IV : LES CARTES FACTORIELLES .......................................................................... 40
1- Définition ............................................................................................................................. 40 2- Etablissement d'une carte factorielle .................................................................................. 40 3- Croisement tri-factoriel ou test à 3 points ........................................................................... 41 4- Notion d'interférence .......................................................................................................... 42

3
PREMIERE PARTIE :
BASES FONDAMENTALES POUR L’ANALYSE GENETIQUE CHEZ LES
ORGANISMES HAPLOIDES

4
CHAPITRE I : CYCLE DE REPRODUCTION, CYCLES CHROMOSOMIQUES,
MECANISME DE LA MEIOSE ET CONSEQUENCES GENETIQUES DE LA
MEIOSE
Tous les êtres vivants, des virus à l'homme, possèdent une propriété commune qui est le pouvoir
de reproduction c'est à dire la transmission de l'information héréditaire des ascendants aux
descendants.
Chez les eucaryotes, cette information est portée par l'ADN. La quasi totalité de l'ADN cellulaire
est localisée dans le noyau au niveau des chromosomes. Les chromosomes sont donc les
supports de l'hérédité.
Dans ce chapitre nous allons étudier successivement le cycle de reproduction, les cycles
chromosomiques,la fécondation, la méiose et les conséquences génétiques de la méiose pour
comprendre comment les chromosomes sont transmis d’une génération à la suivante à travers la
reproduction sexuée.
1- LE CYCLE DE REPRODUCTION
Chaque espèce est caractérisée par un nombre constant de chromosomes dans chacun des
noyaux de chacune de ses cellules somatiques et germinales.
Ce nombre est de 46 pour l'espèce humaine, 20 pour le maïs, 8 pour la drosophile, 14 pour le
mil, 24 pour l’aubergine, 176 pour le laurier etc… Cependant les cellules reproductrices ou
gamètes contiennent la moitié de ce nombre de chromosomes. Ces cellules reproductrices sont
les ovules et les spermatozoïdes chez l’homme et chez les animaux et les ovules et les grains de
pollen chez les végétaux.
La question qui se pose est de savoir par quels mécanismes le nombre de chromosomes se
maintient constant de génération en génération au cours de la reproduction sexuée.
Prenons l’exemple de l'espèce humaine.

5
Schéma 1 : Le cycle de reproduction de l’espèce humaine
Au cours de la fécondation il y a fusion entre 2 cellules reproductrices ou gamètes à 23
chromosomes chacun, l'ovule provenant de la mère et le spermatozoïde provenant du père.
La cellule œuf ou zygote qui dérive de cette fusion a 46 chromosomes.
La multiplication de ce zygote par mitoses successives va donner un individu dont toutes les
cellules somatiques et germinales seront à 46 chromosomes. Chez cet individu qui sera mâle ou
femelle, les testicules ou les ovaires renferment les cellules germinales à 46 chromosomes à
partir desquelles va se dérouler la gamétogenèse ou l’ovogenèse qui permettra de réduire le
nombre de chromosomes de moitié et de produire des spermatozoïdes ou des ovules à 23
chromosomes.
Ainsi des individus adultes d’une génération produiront des gamètes mâles et femelles à 23
chromosomes dont la fusion donnera, à la génération suivante, naissance à un nouvel individu à
46 chromosomes.
Les 23 chromosomes d'un gamète sont morphologiquement différents ; ils sont représentés en 1
seul exemplaire ; on dit que le gamète est à n chromosomes ou encore qu'il est haploïde.
Les 46 chromosomes du zygote et de toutes cellules somatiques et germinales qui en dérivent
par mitoses, sont représentés en 2 exemplaires ; on dit que le zygote et toutes les cellules
somatiques et germinales sont à 2 n chromosomes ou encore qu'elles sont diploïdes.
Dans la genèse d'un individu on peut ainsi définir deux phases : la PHASE DIPLOIDE où les cellules
contiennent 2 n chromosomes et la PHASE HAPLOÏDE où les cellules ne contiennent que n
chromosomes.
2- LES CYCLES CHROMOSOMIQUES
Au cours des générations sexuées successives il y a un cycle d'alternance des phases diploïde et
haploïde. Cette alternance est assurée par La MEIOSE qui fait passer de la phase diploïde à la
phase haploïde et par La FECONDATION qui fait passer de la phase haploïde à la phase diploïde.
Ces deux processus biologiques complémentaires que sont la Méiose et la Fécondation
permettent de maintenir constant le nombre de chromosomes de l'espèce humaine.

6
La méiose et la fécondation ne sont pas spécifiques à la seule espèce humaine, elles se déroulent
chez la quasi totalité des eucaryotes. Mais l'importance relative des phases haploïde et diploïde
permet cependant de définir 3 principaux cycles chromosomiques : le cycle diplobiontique, le
cycle haplobiontique et le cycle haplodiplobiontique
Schéma 2 : Les trois principaux cycles chromosomiques
Le cycle diplobiontique est caractérisé par une phase diploïde prépondérante au cours de
laquelle s'opère la multiplication cellulaire. La phase haploïde est réduite aux gamètes. C'est le
cas chez l'espèce humaine, le maîs, la drosophile, le pois, d'une façon générale chez les plantes et
les animaux supérieurs ;
Le cycle haplobiontique est caractérisé par une phase haploïde prépondérante au cours de
laquelle s'opère la multiplication cellulaire. La phase diploïde est réduite au seul zygote. C'est le
cas chez les champignons : Ascobolus, Sordaria, Neurospora ;
le cycle haplodiplobiontique est caractérisé par des multiplications cellulaires aussi bien en phase
haploïde que diploïde. Il y a ainsi alternance de deux formes, l'une haploïde et l'autre diploïde.
C'est le cas chez la levure de boulangerie (Saccharomyces cerevisiae).
3- LA FECONDATION
La fécondation est le processus qui, par la fusion de 2 gamètes haploïdes, conduit à la formation
d'une cellule diploïde ou zygote et permet ainsi le passage de la phase haploïde à la phase
diploïde. Chez la majorité des espèces, la fécondation a lieu entre des gamètes de tailles
différentes : on parle d'anisogamie. Le gamète femelle est immobile et possède un cytoplasme
riche en réserves. Le gamète mâle beaucoup plus petit est mobile et contient très peu de
cytoplasme. Il existe donc une différence au niveau de leur participation respective à la
constitution du cytoplasme.
Par contre gamètes mâle et femelle ont une contribution équivalente à la fabrication du noyau
du zygote à 2n chromosomes au cours de caryogamie. En effet les 2 gamètes qui contiennent
chacun n chromosomes apportent non seulement le même nombre de chromosomes, mais aussi
les mêmes types de chromosomes.
4- LA MEIOSE
La méiose est le processus qui fait passer de la phase diploïde à 2n chromosomes à la phase
haploïde à n chromosomes. Elle permet d'obtenir à partir d'une cellule diploïde 4 cellules
haploïdes. La méiose est un ensemble de 2 divisions nucléaires successives. La première est

7
précédée d'une duplication des chromosomes et procède à la séparation des chromosomes
homologues. La seconde procède à une division des chromosomes.
4-1- Description cytologique de la méiose
La méiose consiste en 2 divisions successives nommées DIVISION I et DIVISION II. Elles sont
subdivisées chacune en quatre phases d'après la morphologie et le mouvement des
chromosomes. La DIVISION I est précédée par l’interphase
4-1-1 L’Interphase
Le noyau interphasique est diffus. L’évènement majeur qu'il abrite n’est pas optiquement
visible. Il s’agit de la réplication de la molécule d'ADN de chaque chromosome. Chaque
chromosome se trouve ainsi composé de 2 molécules d'ADN filles avec pour conséquence la
duplication de chaque chromosome en 2 chromatides sœurs.
Schéma 3 : la réplication de la molécule d’ADN et la duplication de chaque chromosome
au cours de l’interphase

8
Schéma 4 : La description cytologique de la méiose
4-1-2 La DIVISION I ou division réductionnelle
Elle se compose de 4 phases : la prophase I, la métaphase I, l'anaphase I et la télophase I.
Prophase I
Elle est longue et complexe. Cette complexité a amené les cytologistes à la subdiviser en 5 stades
qui se succèdent de manière continue. Ces stades sont suivants :
Leptotène
Les chromosomes s'individualisent et apparaissent au microscope optique sous forme de
filaments fins, très longs et enchevêtrés. Chaque chromosome qui est déjà composé de 2
molécules d'ADN n'apparaît pas encore divisé en 2 chromatides sœurs.
Zygotène
Ce stade correspond au début de l'appariement des chromosomes homologues. Chaque
chromosome tout en se raccourcissant et en devenant épais va s'apparier à son homologue. Ce
phénomène d'appariement est le plus caractéristique et le plus important de la méiose. Le noyau
contient ainsi n paires de chromosomes ou n bivalents.
Pachytène
A ce stade, l'épaississement des chromosomes est important, de sorte que chaque chromosome
apparaît nettement formé de 2 chromatides sœurs étroitement accolés et d’un chromosome.
Chaque chromatide contient une molécule d'ADN : c'est la visualisation de la réplication de l'ADN
qui s’est opérée à l’interphase. Les chromosomes homologues sont entièrement appariés et cet
appariement est strictement homologue, il se fait point par point sur toute la longueur des
chromosomes. Les chromosomes homologues appariés forment des bivalents. Chaque bivalent
compte donc à ce stade 4 chromatides avec seulement 2 centromères et devient une tétrade.

9
Diplotène
L'attraction mutuelle entre les chromosomes homologues se relâche de telle sorte que les deux
paires de chromatides homologues de chaque tétrade ont tendance à se séparer comme si elles
se repoussaient mais restent associés en certains points appelés les chiasmas.
Diacinèse
A ce stade les chromosomes continuent à s'épaissir et atteignent leur maximum de
condensation. Les forces de répulsion éloignent progressivement les chromosomes homologues
l'un de l'autre et provoquent ainsi le glissement des chiasmas vers les extrémités des
chromosomes. C'est la terminalisation des chiasmas qui a comme effet la diminution du Le
nombre de chiasmas.
Métaphase I
Elle débute avec la disparition de l'enveloppe nucléaire et la formation du fuseau achromatique.
Les chromosomes homologues encore reliés par des chiasmas se disposent au niveau du plan
équatorial avec leurs centromères situés de part et d'autre de ce plan.
Anaphase I
La terminalisation prend fin. Les derniers chiasmas se séparent. Avec leurs centromères non
encore clivés, les chromosomes homologues formés chacun de 2 chromatides migrent vers
chaque pôle de la cellule, par l'Intermédiaire des fibres contractiles.
Télophase I
A chaque pôle de la cellule on a n chromosomes formés chacun de 2 chromatides. Selon les cas il
y a une reconstitution d'un noyau inter phasique avec l'apparition d'une membrane nucléaire
(cas de la sauterelle) ou une amorce immédiate de la seconde division (cas du Trillium).
Le bilan de la première division de la méiose ( DIVISION I) est la réduction par deux du nombre de
chromosomes par noyau
4-1-3 : La DIVISION II ou division équationnelle
Elle est comparable à une mitose normale. Les deux noyaux issus de la division subissent
chacun une seconde division, parallèlement et de façon synchrone.
Prophase II
En fin de prophase II, les chromosomes sont à nouveau contractés.
Métaphase II
Les chromosomes se disposent au niveau du plan équatorial avec leurs centromères situés sur ce
plan.
Anaphase II
L'anaphase II débute par le clivage du centromère de chaque chromosome et se poursuit par la
migration vers des pôles opposés des deux chromosomes fils
Télophase II
La membrane nucléaire réapparaît. Les quatre cellules correspondant à ces quatre noyaux
haploïdes sont appelées les quatre produits de méiose.

10
Le bilan de la deuxième division de la méiose (DIVISION II) est la division de chaque noyau issu
de la DIVISION I en 2 noyaux qui lui sont identiques.
Le schéma ci-après illustre les bilans des deux divisions de la méiose.
Schéma 5 : Bilans des deux divisions de la méiose
5- LES CONSEQUENCES GENETIQUES DE LA MEIOSE
II est nécessaire de revenir sur certains aspects de la méiose pour en voir les conséquences au
niveau génétique.
5-1- Le Crossing-over et la Recombinaison intra chromosomique
Considérons le devenir d'une seule paire de chromosomes homologues au cours de la méiose.
Au stade diplotène de la prophase I, les chromosomes homologues formés de 2 chromatides
sont en répulsion mais restent associés en un point appelé le chiasma. Le chiasma est une
figure cytologique qui montre 2 chromatides homologues entrecroisées en X.
Ce chiasma est la conséquence cytologique d'un phénomène appelé crossing-over.
Le crossing-over se déroule en deux étapes. La première consiste en une cassure en deux
points homologues de 2 chromatides homologues. La deuxième consiste en une soudure
entre des segments de chromatides homologues donc en un échange physique réciproque de
segments de chromatides homologues. Chacun des chromosomes homologues qui a subi le
crossing-over est formé d’une chromatide parentale et d’une chromatide remaniée.
Il s'est ainsi produit une recombinaison qui touche la structure physique des chromosomes, on
parle de recombinaison intra-chromosomique.

11
Schéma 6 : Le crossing-over et ses conséquences cytologique et génétique
A la métaphase I, les centromères des chromosomes homologues se placent de part et d'autre
du plan équatorial.
A télophase I, chaque noyau contient un chromosome formé d'une chromatide de type
parental et d'une chromatide remanié.
A la télophase II, on obtient 2 gamètes avec chacun un chromosome de type parental et 2
gamètes avec chacun un chromosome de type remanié.
En plus donc des gamètes de type parental on a des gamètes de type recombiné

12
Schéma 7 : Les gamètes parentaux et recombinés résultant d’un crossing-over
La distinction entre les gamètes de type parental et les gamètes de type recombiné peut se
faire en suivant le devenir au cours de la méiose de deux couples d’allèles A/a et B/b portés
par une paire de chromosomes homologues
Supposons que le chromosome d’origine paternelle porte les allèles a et B et que le
chromosome d’origine maternelle porte les allèles A et b.
Le noyau qui subi la méiose contient 1 paire de chromosomes homologues portant les
couples d’allèles A/a et B/b
Nous allons considérer les deux événements qui peuvent se produire dans l’intervalle qui
sépare les deux couples d’allèles A/a et B/b :
- si aucun crossing-over n'a lieu dans l'intervalle qui sépare les deux couples d’allèles A/a
et B/b , on aura 4 gamètes dont 2 gamètes Ab et 2 gamètes aB. Les gamètes Ab et
aB sont des gamètes de type parental.
Schéma 8 : Les gamètes parentaux résultant de l’absence de crossing-over
dans l’intervalle séparant les deux couples d’allèles A/a et B/b
- si un crossing-over a lieu dans l'intervalle qui sépare les deux couples d’allèles A/a et
B/b on aura 4 gamètes : Ab et aB sont des gamètes de type parental et AB et ab qui
sont des gamètes de type recombiné.

13
Schéma 9 : Les gamètes parentaux et recombinés résultant d’un crossing-over
dans l’intervalle séparant les deux couples d’allèles A/a et B/b
Ces deux événements n’ont pas la même probabilité car le crossing-over est un événement
rare, aussi :
- si on considère un grand nombre de méioses, la fréquence de méioses ayant subies un
crossing-over sera faible. Cela se traduira par le fait que la fréquence des gamètes de
type recombiné sera inférieure à la fréquence des gamètes de type parental.
- si Inversement on considère 1000 gamètes et que l'on trouve 450 Ab, 450 aB,
50 AB, 50 ab on pourra conclure que :
- les gamètes Ab et aB sont les gamètes de type parental ;
- AB et ab sont les gamètes de type recombiné ;
- les 2 couples A/a et B/b sont liés et donc qu'ils sont portés par des
chromosomes différents.
Le pourcentage de recombinaison est de : [(50 + 50)/ 1000] x100 = 10%
Ce pourcentage qui est lié au pourcentage de crossing-over permettra d'estimer la distance
génétique qui sépare les 2 couples d’allèles A/a et B/b.
Cette distance dans ce cas est de 10 UR.
5-2- La position aléatoire des centromères en métaphase I et recombinaison
inter-chromosomique
Considérons deux paires de chromosomes et suivons leur devenir au cours de la méiose en
faisant abstraction des remaniements dûs au crossing-over.
A la Métaphase I, les centromères peuvent se placer de manière aléatoire de part et d'autre
du plan équatorial selon 2 dispositions

14
Schéma 10 : Les gamètes parentaux et recombinés résultant de la recombinaison inter-chromosomique
impliquant 2 paires de chromosomes homologues
La première disposition (I) permettra d’obtenir à la fin de la méiose 4 gamètes qui
contiendront chacun 2 chromosomes de la même origine parentale ;
La seconde disposition (II) permettra d’obtenir 4 gamètes qui contiendront chacun 2
chromosomes d’origines parentales différentes
Ainsi on aura en plus des gamètes de type parental, des gamètes de type recombiné. Il s'est
produit une recombinaison entre les chromosomes non homologues, on parle de
recombinaison inter-chromosomique.
La distinction entre les gamètes de type parental et les gamètes de type recombiné peut se
faire en utilisant deux couples d’allèles Aa et B/b portés par deux paires de chromosomes
différents. Supposons que :
- la 1ère paire d'homologues porte le couple d’allèles A/a avec A sur le chromosome
d'origine paternel et a sur le chromosome d'origine maternelle
- la 2ème paire porte le couple d’allèles B/b avec B sur le chromosome d'origine
paternelle et b sur le chromosome d'origine maternelle.
Le noyau qui subi la méiose contient 2 paires de chromosomes homologues portant
respectivement les couples d’allèles A/a et B/b
En métaphase I les centromères de ces chromosomes peuvent se placer de manière aléatoire

15
de part et d’autre du plan équatorial selon 2 dispositions.
Schéma 11 : Les gamètes parentaux et recombinés résultant de la recombinaison inter-chromosomique
impliquant 2 paires de chromosomes homologues portant respectivement les couples d’allèles A/a
et B/b
La première disposition (I) permettra d’obtenir 4 gamètes dont 2 AB et 2 ab. Ces gamètes
sont des gamètes de type parental.
La seconde disposition (II) permettra d’obtenir 4 gamètes dont 2 Ab et 2 aB. Ces gamètes
sont des gamètes de type recombiné.
Ces 2 dispositions ont la même probabilité, aussi,
- si on considère un grand nombre de méiose, la moitié de ces méioses donnera des
gamètes de type parental et l'autre moitié donnera des gamètes de type recombiné.
On aura donc fr P= fr R ; on aura donc comme gamètes 1/4 AB ; 1/4 ab ; 1/4 Ab et 1/4
aB
- si, Inversement, son observe 1000 gamètes et que l'on trouve 250 AB ; 250 ab ; 250 Ab
et 250 aB on pourra conclure que les 2 couples d’allèles sont indépendants et donc
qu'ils sont portés par des chromosomes différents.
Ce que nous venons de voir avec 2 paires de chromosomes peut s'étendre à tous les
chromosomes d'un même organisme.
Dans le cas de 2 paires de chromosomes il y a 4 types de gamètes, soit 2².
Dans le cas de 3 paires de chromosomes il y a 8 types de gamètes, soit 23.
Dans le cas de n paires de chromosomes il y aura donc 2n types de gamètes.

16
Dans l'espèce humaine où on a 23 paires de chromosomes on aura donc 223 types de gamètes
Ainsi chaque Homme étant le résultat de la fusion de 2 gamètes un provenant du père
et l’autre provenant de la mère, la probabilité d'obtenir 2 fois le même individu est
mathématiquement infime.
Cette probabilité est encore plus infime si l'on tient compte du brassage intra
chromosomique. On comprend ainsi que chaque homme est de par son génotype unique sur
la terre.
6- CONCLUSION
D'un point de vue générale, il apparaît que la sexualité c'est à dire le jeu alterné de la
fécondation et de la méiose :
- assure la continuité biologique par le maintien de la stabilité du nombre de
chromosome et de la quantité d'ADN par cellule
- et crée la diversité biologique qui constitue un atout considérable pour la survie d'une
espèce.
Il est clair que plus divers sont les individus d'une population, plus grande est l'adaptabilité de
celle-ci aux changements des conditions d'existence.
Du point de vue de l'étude des mécanismes de l'hérédité, la connaissance du devenir des
chromosomes au cours de la méiose et des différents types de recombinaisons
chromosomiques constituera la base indispensable pour comprendre la transmission des
caractères héréditaires.
La recombinaison intra-chromosomique est associée au crossing-over qui se produit au stade
diplotène de la prophase I et à la liaison entre deux couples d’allèles localisés sur 1 paire de
chromosomes homologues
La recombinaison inter-chromosomique est associée à la position aléatoire des centromères
des chromosomes homologues en Métaphase I et à l’indépendance entre deux couples
d’allèles localisés sur 2 paires de chromosomes homologues

17
CHAPITRE II – EXEMPLES DE CYCLES DE REPRODUCTION CARCTERISTIQUES DES
ASCOMYCETES
1- LE CYCLE de Neurospora crassa (la moisissure du pain)
Schéma 12 : Le cycle de reproduction de Neurospora crassa
Neurospora crassa est un champignon ascomycète filamenteux haploide à n = 7. Ses
fructifications ou périthèces sont des sortes de petites bouteilles noires de 1 à 2 mm de haut
à maturité. Ces périthèces contiennent de petits sacs allongés appelés des asques qui
renferment chacun 8 spores disposées linéairement.
Chaque spore germe en produisant des filaments qui vont croître et se ramifier pour donner
un mycélium formé d’articles plurinucléés. Un mycélium constitue une souche qui peut être
multipliée végétativement par repiquage de fragments de ce mycélium.
Un mycélium différencie des organes reproducteurs :
- les conidies qui peuvent germer et produire un nouveau mycélium. Mais ils servent surtout
d'éléments mâles ;
- les ascogones qui servent d'éléments femelles.
Cependant un mycélium cultivé seul ne produit jamais de fructifications. Celles-ci
n'apparaissent que si les conidies d'un mycélium issu d'une spore de type conjugal A
fécondent les ascogones d'un mycélium issu d'une spore de type conjugal a : on dit qu'il y a
hétérothallisme.
A l'intérieur de l'ascogone fécondé, il y a multiplication synchrone du noyau de la conidie et du
noyau de l'ascogone. Il se différencie des hyphes ascogènes formés de 3 à 5 articles
contenant chacun un noyau A et un noyau a. Ces articles sont à n+n chromosomes, on dit
qu'ils sont dicaryotiques. Les derniers articles de ces hyphes se différencient en crochets
dangeardiens. La caryogamie a lieu dans la cellule apicale de chaque crochet. Le noyau
diploide obtenu subit une méiose suivie d’une mitose dite mitose post- méiotique à l'intérieur
de cette cellule qui s'allonge parallèlement pour former un asque qui contient 8 spores ou
ascospores
Cet asque disposés selon un ordre qui provient du fait que les fuseaux de division ne se

18
chevauchent pas quand les noyaux se divisent.
Par le jeu de mitoses synchrones au niveau de chaque hyphe ascogène, une série de crochets
donnera naissance à un bouquet d’asques provenant d’un seul couple de noyaux.
Pendant ce temps l'ascogone s'est développé, son enveloppe s'est épaissie et pigmentée, il est
devenu un périthèce qui peut contenir plus de cent asques
La phase diploïde est réduite au noyau diploïde. Le cycle dure environ dix jours.
2- LE CYCLE de Saccharomyces cerevisiae (la levure de boulangerie)
Schéma 13 : Le cycle de reproduction de Saccharomyces cerevisiae
Saccharomyces cerevisiae est un ascomycète unicellulaire dont le cycle est
haplodiplobiontique. Il est capable de se multiplier sous deux formes : une forme diploïde
(2n = 32 chromosomes) et une forme haploïde (n = 16 chromosomes).
Deux cellules haploïdes provenant à l’origine de deux spores qui différent par un gène que
l’on nomme le signe (+/- ou a/α) peuvent fusionner pour donner un zygote. Après la
caryogamie la cellule diploide obtenue se multiplie par bourgeonnement et les cellules
diploïdes qui en résultent peuvent être cultivées indéfiniment tant que l'environnement est
favorable. Si les nutriments viennent à manquer, ces cellules subissent la méiose et donnent
chacune quatre spores haploïdes enfermées dans un asque. Ces spores bourgeonnent en
donnant des cellules haploïdes qui peuvent proliférer indéfiniment en donnant à nouveau des
lignées de cellules haploïdes.
Les phases haploïdes et diploïdes peuvent avoir une importance égale dans la nature. La

19
phase diploïde est prépondérante quand les conditions de vie sont favorables. Les spores
apparaissent quand le milieu devient défavorable.
Saccharomyces cerevisiae peut se reproduire par voies asexuée ou sexuée.
3- ELEMENTS DE REFERENCES POUR l’ANALYSE GENETIQUE Les organismes haploïdes qui vont servir de matériel d'étude sont les champignons
ascomycètes.
Le caractère qui permet de regrouper tous les Ascomycètes tient aux faits suivants :
- la méiose se produit dans une cellule, qui en s'allongeant devient un asque ;
- les produits d’une méiose restent enfermés ensemble dans l'asque.
Chez certains ascomycètes (Neurospora, Sordaria) non seulement les 4 produits de la méiose
sont contenus dans l'asque, mais de plus, ils y sont ordonnés : Ces champignons sont à asques
ordonnés
Chez d'autres ascomycètes (Ascobolus) les fuseaux de division se chevauchent quand les
noyaux se divisent. Les 4 produits de méiose ne sont pas disposés selon un ordre :
ces champignons sont à asques non ordonnés.
Chez Neurospora, Sordaria et Ascobolus, la méiose est suivie d'une mitose dite post méiotique
de sorte que l'asque renferme 8 spores.
L'observation des caractères se fait en phase haploïde au niveau des produits de méiose
(spores ou ascospores) ou sur les individus qui en résultent par multiplication végétative
(mycélium).
Les caractères étudiés sont :
- des caractères morphologiques : taille, forme, pigmentation ;
- des caractères biochimiques : auxotrophie par rapport à un métabolite de croissance (Acide
Aminé, Vitamine etc..)
II existe deux niveaux d'analyse :
- analyse de tétrades ou d'asques
dans ce cas l'analyse porte sur un échantillon de tétrades ; les 4 produits d'une même
méiose sont donc identifiés. De plus au niveau de tétrades ordonnées, l'ordre permet de
de déceler avec précision les stades où se produit de la ségrégation des allèles.
- analyse en vrac
dans ce cas l'analyse porte sur un échantillon de produits de méiose. Les 4 produits d'une
même méiose ne sont pas identifiés.

20
DEUXIEME PARTIE :
TRANSMISSION DES CARACTERES HEREDITAIRES
CHEZ LES ORGANISMES HAPLOIDES

21
CHAPITRE I : SEGREGATION D'UN COUPLE D'ALLELES
1- Cas d'un organisme à tétrades ordonnés : Neurospora crassa
Le croisement entre une souche sauvage à spores noires et une souche mutante à spores
blanches a donné une descendance composée comme suit :
Schéma 14 : Descendance composée d’asques pré-réduits et post-réduits
L'observation de la descendance permet de constater les faits suivants :
- Chaque asque renferme uniquement des spores noires et des spores blanches, jamais de
spores de coloration intermédiaire.
Les spores obtenues sont rigoureusement identiques à l'un ou l'autre parent. Chaque spore ou
gamète emporte à l'état pur l'une ou l'autre des potentialités héréditaires des parents
provisoirement réunies dans le zygote. Ce fait illustre la notion de pureté des gamètes
- Chaque asque renferme 4 spores noires et 4 spores blanches. On dit que l'asque présente
une ségrégation 1-1.
Pour expliquer ce fait nous allons supposer que le caractère coloration des spores est sous la
dépendance d'un couple d'allèle b+/b qui occupe un emplacement caractéristique ou locus sur
l'un des chromosomes de Neurospora. Ce locus est à une certaine distance du centromère du
chromosome qui le porte. L'allèle b détermine le phénotype "spore blanche" et l'allèle b+
détermine le phénotype "spore noire".
Ces 2 allèles sont réunis au cours de la caryogamie au sein du zygote dont le noyau diploïde
possède 2 chromosomes homologues qui portent l'un l'allèle b+ et l'autre l'allèle b. Ce noyau
va subir la méiose pour donner 4 produits. Deux d’entre contiendront b, les 2 autres
contiendront b+. La mitose post méiotique donnera 4 spores b+: noires et 4 spores b :
blanches.
Les 2 allèles qui étaient réunies dans le noyau diploïde du zygote sont séparés dans les
produits de la méiose. La question qui se pose est de savoir à quel moment s'effectue la
séparation ou la ségrégation de ces 2 allèles.
C'est l'analyse des faits suivants qui va nous apporter la réponse.

22
- la descendance est composée de 6 types d'asques quant à l'ordonnancement des spores.
Si on n'observe que les demi-asques, ces 6 types peuvent être regroupés en 2 classes.
La Classe 1 : regroupe 2 types d’asques dont les demi-asques contiennent soit exclusivement
des spores noires soit exclusivement des spores blanches. Ces asques sont à demi-asques
sont homogènes.
Schéma 15 : Formation des asques pré-réduits
En suivant le devenir du couple d’allèles b+/b au cours de la méiose, il apparaît qu'à la fin de
la 1ère division un noyau fils contenait 1 chromosome avec les 2 chromatides portant le même
allèle b+, l'autre noyau fils contenait 1 chromosome avec les 2 chromatides portant le même
allèle b.
Les 2 allèles du couple b+/b se sont donc séparés à la 1ère division de méiose avant la
réduction du nombre de chromosomes. Les asques à demi-asques homogènes sont dits
asques pré-réduits.
L'apparition des 2 types d’asques pré-réduits dépend de la position aléatoire ces centromères
de part et d’autre du plan équatorial à la métaphase I. Les 2 dispositions possibles sont
équiprobables. Les 2 types d’asques qui en résultent auront donc des fréquences ou des
effectifs statistiquement équivalents.
La Classe II : regroupe 4 types d’asques dont les demi-asques contiennent à la fois des spores
noires et des spores blanches. Ces asques sont à demi-asques sont hétérogènes.

23
Schéma 16 : Formation des asques post-réduits
En suivant le devenir du couple d’allèles b+/b au cours de la méiose et en faisant intervenir
l'existence d'un crossing-over entre le centromère et le locus du couple d'allèle b+/b, il
apparaît qu'à la fin de la 1ère division, chacun des 2 noyaux fils contenait un chromosome avec
les 2 chromatides portant des allèles différents, l'une porte b+, l'autre porte b.
Les 2 allèles du couple b+/b se sont donc séparés à la 2ème division de méiose après la
réduction du nombre de chromosomes. Les asques à demi-asques hétérogènes sont dits
asques post-réduits.
L'apparition des 4 types d’asques post-réduits dépend de la migration des chromosomes à
l'anaphase II. Comme cette migration se fait de manière aléatoire, on aura autant de chance
de trouver dans chaque demi-asque la séquence b+b+ ou bb. Au total en combinant les 2
demi-asques dont l'histoire est indépendante au cours de la 2ème division de méiose on
obtient les 4 produits de méioses possibles avec la même probabilité.
Les 4 types asques qui en résultent auront donc des fréquences ou des effectifs
statistiquement équivalents.
Notion de distance
A l'origine d'un asque post-réduit, nous savons maintenant qu'il se produit un crossing-over
entre le locus du gène considéré et son centromère. Si l'on postule que les crossing-over sont
distribués de manière uniforme tout au long du chromosome, la fréquence des crossing-over
sera plus ou moins grande selon que le locus du gène considéré est plus ou moins loin de son
centromère.
Cela revient donc à dire que la fréquence de crossing-over est fonction de la distance qui
sépare le locus d'un gène de son centromère.
Cette fréquence nous donne donc un moyen de mesurer les distances qui séparent les gènes
de leurs centromères.
La distance génétique est exprimée en centimorgan CM (de Morgan qui a découvert le
crossing-over) ou en unité de recombinaison UR
Comment calcule-t-on cette distance entre le gène et le centromère ?
L’expression de la distance gène-centromère est la suivante :
dg-c = x 100 (UR)

24
Dans chaque asque, il y avait à l'origine, dans le noyau qui a subi la méiose, 2 chromosomes
homologues dupliqués, donc 4 chromatides.
Dans chaque asque pré-réduit il n'y a pas eu de crossing-over entre le locus du gène et le
centromère, il n'y a donc pas eu de remaniement chromatidique.
Dans chaque asque post réduit il y a eu 1 crossing-over entre le locus du gène et le
centromère . Le crossing-over se faisant entre 2 chromatides sur les quatre en présence, ces 2
chromatides sont remaniées ; il y a donc eu 2 remaniements chromatidiques. L'expression de
la distance devient donc :
dg-c = x 100 (UR)
dg-c = 1/2 % Post Réduction (UR)
Dans notre exemple on aura donc dg b+/b-c = x 100 = 9, 86 UR
Relation entre % de Post réduction et distance gène centromère
Double crossing over et remaniements chromatidiques
Jusqu'à présent nous avons postulé qu'à l'origine d'un asque post-réduit il ne se produit
qu'un seul crossing-over. En fait, au cours de la méiose il peut se produire plusieurs
crossing-over à la fois au niveau d'une paire de chromosomes homologues : on parle dans
ce cas de crossing-over multiples.
Considérons le cas où il se produit 2 crossing-over ou 1 double crossing-over dans l'intervalle
gène-centromère.
Ces 2 crossing-over pourront toucher :
- 2 chromatides dans ce cas on aboutit à un asque pré-réduit
- les 4 chromatides dans ce cas on aboutit à un asque pré-réduit ;
- 3 chromatides dans ces 2 cas on aboutit dans à un asque post réduit.
Comme ces 4 situations sont équiprobables, il apparaît que des 16 remaniements
chromatidiques qui se produisent (4 par situation) seules 4 sont décelables (2 par asque post
réduit). Ainsi donc les 12/16 ou 3/4 des remaniements chromatidiques ne sont pas décelables.
La relation dg-c = 1/2 % Post Réduction n'est donc correcte que si le segment gène-
centromère est suffisamment petit pour que les crossing-over multiples soient négligeables.
S'ils ne le sont plus, le calcul de la distance entre le gène et le centromère sur la base de la
relation dg-c = 1/2 % Post Réduction, donnera une sous évaluation de cette distance.
Limites du % de post réduction
La limite inférieure de la post-réduction est égale à zéro. Dans ce cas la distance gène-
centromère est nulle. Le locus du gène considéré est très près de son centromère, le gène est
donc toujours pré-réduit.
Si on considère un gène très éloigné de son centromère, de telle manière que de nombreux
crossing-over puissent avoir lieu dans le segment gène-centromère, on peut admettre que les
allèles du gène ne gardent aucun souvenir du centromère auquel ils étaient primitivement
rattachés. Leur ségrégation se fera indépendamment de celle du centromère de sorte
qu'aucune contrainte ne sera plus exercée sur le moment de leur disjonction.
Dans ce cas les 6 types de tétrades obtenus ont la même fréquence qui est égale à 1/6. Les
tétrades pré-réduites ont une fréquence de 2/6 et les tétrades post-réduites ont une 4/6. La
fréquence de post réduction est donc de 4/6 ou 2/3. La limite supérieure du % de post
réduction est donc égale à 66%.

25
2- Cas d'un organisme à tétrades non ordonnés : Ascobolus immersus
Le croisement entre une souche sauvage à spores noires et une souche mutante à spores
blanches a donné une descendance composée de 2486 asques contenant chacun 4 spores
noires et 4 spores blanches.
L’analyse de cette descendance permet de conclure que chaque asque présente une
ségrégation 1-1. Le caractère coloration des spores est donc sous la dépendance d'un couple
d'allèles b/b+.
Dans le cas des tétrades non ordonnées, on observe un seul type d'asque. Du fait de l'absence
d'ordonnancement des spores dû au chevauchement des fuseaux au cours des divisions de la
méiose, la pré-réduction et la post réduction ne sont pas décelées.
Il ne sera donc pas possible dans ce cas d'estimer la distance gène-centromère.
3- Analyse de spores en vrac
Qu'il utilise Neurospora ou Ascobolus comme matériel d'étude, l'expérimentateur peut
récolter seulement une population de spores en vrac.
Comme dans chaque asque, pour un couple d'allèles a+/a, on observe 4 spores a+ et 4 spores
a, au niveau d'une population de spores prises en vrac, on aboutit à 50% de spores a+ et 50 %
de spores a c'est à dire à une ségrégation 1-1,
Ainsi, quel que soit le niveau d'analyse (tétrades ordonnées, tétrades non ordonnées, spores
en vrac) la ségrégation d'un couple d'allèles est une ségrégation 1-1 et réciproquement.

26
CHAPITRE II : SEGREGATION DE DEUX COUPLES D'ALLELES LOCALISES SUR 2
PAIRES DE CHROMOSOMES DIFFERENTS
1- Cas d'un organisme à tétrades non ordonnées : Ascobolus
Chez Ascobolus, le croisement entre une souche mutante à spores blanches et une souche
mutante à spores rondes, a donné la descendance ci-après :
Schéma 17 : Descendance composée de DP, DR et TT dans le cas deux 2 couples d’allèles
localisés sur 2 pairs de chromosomes différents
Ce croisement fait intervenir 2 caractères la coloration de la spore et la forme de la spore
Pour le caractère coloration, dans chaque asque on a 4 spores noires et 4 spores blanches,
c'est à dire une ségrégation 1-1 qui permet de conclure que ce caractère est sous la
dépendance d'un couple d'allèles b+/b.
Pour le caractère forme de la spore dans chaque asque on a 4 spores ovales et 4 spores
rondes, c'est à dire une ségrégation 1-1 qui permet de conclure que ce caractère est sous la
dépendance d'un couple d'allèles r+/r.
Quelle relation existe-t-il entre ces 2 couples d'allèles ?
Pour le comprendre faisons abstraction des asques et intéressons nous uniquement aux
spores. En totalisant toutes les spores présentant le même phénotype sans tenir compte du
type d'asque d'origine, ce qui revient à procéder à une analyse en vrac, on obtient les résultats
suivants :
En considérant le croisement de départ : [blanche ovales] x [noire ronde] ; il apparait que la
Types de spores
Phénotypes
Génotypes
Effectifs
Pourcentages
Fréquences
Parental
[br+] br+
1032
25,6%
1/4
Parental
[b+r] b+r 1032
25,6 %
1/4
Recombiné
[b+r+] b+r+ 980
24,4 %
1/4
Recombiné
[br]
br
980
24,4%
1/4

27
descendance est composée des spores de type parental br+ et b+r et des spores de type
recombiné b+r+ et br
Compte tenu des fluctuations aléatoires, on peut dire que les fréquences observées ne sont
pas significativement différentes de 1/4. Chacun des 4 génotypes est également présent.
Cela revient à dire qu'un allèle donné d'un couple d'allèles a autant de chance de s'associer à
chacun des allèles de l'autre couple. Schématisons ces associations qui se font de manière
aléatoire
Les ségrégations des 2 couples d'allèles sont donc indépendantes. Cette indépendance se
traduit aussi par l'existence dans la descendance d'autant de produits parentaux que de
produits recombinés (P = R). Elle se traduit encore par le fait que le pourcentage de
recombinaison x 100 = 50 %
Considérons maintenant la descendance en asques:
On distingue 3 types d'asques :
- ceux qui ne renferment que les 2 associations parentales br+ et b+ r : ils sont appelés Ditype
Parental, DP
- ceux qui ne renferment que les 2 associations recombinées br et b+r+ : ils sont appelés Ditype
Recombiné, DR ;
- ceux qui renferment à la fois les 2 associations parentales br+ et b+ r et les 2 associations
recombinés br et b+r+ c'est à dire les 4 associations : ils sont appelés Tétratype TT.
Quels sont les origines de ces asques ?
Au cours de la méiose selon qu'il se produise ou non un crossing-over entre un couple d'allèles
et son centromère, différents types d'asques peuvent exister.
Considérons les différents événements et leurs conséquences
- aucun crossing-over ne se produit ni entre r+/r et son centromère ni entre b+/b et son
centromère : on a une pré-réduction des 2 couples d'allèles. On obtient des DP et des DR
avec autant de DP que de DR

28
Schéma 18 : Formation de DP et DR résultant de l’absence de crossing-over entre les 2
couples d’allèles et leurs centromères
- aucun crossing-over ne se produit entre b+/b et son centromère et un crossing-over se
produit entre r+/r et son centromère : on a une pré-réduction pour b+/b associée à une
post-réduction pour r+/r. On obtient uniquement des TT.
- Schéma 19 : Formation de TT résultant de l’absence de crossing-over entre l’un des 2 couples
d’allèles et son centromère associée l’existence d’ un crossing-over entre l’autre couple d’ allèles et
son centromère
- aucun crossing-over ne se produit entre r+/r et son centromère et un crossing-over se
produit entre b+/b et son centromère : on a une pré-réduction pour r+/r associée à une
post-réduction pour b+ b. On obtient uniquement des TT.
- Schéma 19 bis : Formation de TT résultant de l’absence de crossing-over entre l’un des 2 couples
d’allèles et son centromère associée l’existence d’ un crossing-over entre l’autre couple d’ allèles et
son centromère
- un crossing-over se produit aussi bien entre r+/r et son centromère que entre b+/b et son
centromère : on a une post-réduction des 2 couples d'allèles. Dans ce cas les 2 noyaux issus
de la 1ère division de méiose sont identiques du point de vue des associations d'allèles

29
qu'ils renferment. A l'anaphase II après le clivage des centromères du fait des ségrégations
indépendantes des chromosomes on aura 2 situations équiprobables pour chaque noyau
qui aboutissent à deux demi asques ayant la même fréquence. Comme les devenirs des 2
noyaux issus de la 1ère division de méiose sont indépendants on aboutit à 4 combinaisons
équiprobables qui donnent : 1/4 de DP, 1/4 de DR et 1/2de TT.
- Schéma 20 : Formation de DP, DR et TT résultant de l’existence d’ un crossing-over entre les deux
couples d’allèles et leurs centromères
Estimation des fréquences des asques DP, DR et TT
Il est possible de donner une formulation des fréquences des DP, DR et TT, en désignant par :
- x la fréquence de post réduction du couple r+/r et (1-x) sa fréquence de pré-réduction
- y la fréquence de post réduction du couple b+/b et p (1-y) sa fréquence de pré-réduction.nc
Considérons tous les différents évènements, leurs fréquences ainsi que les asques qui en
résultent :
- pré-réduction des 2 couples d'allèles : (1-x)(1-y) dans ce cas on a 1/2 de DP et 1/2 de DR
- pré-réduction d'un couple associée à la post réduction de l'autre. Il existe 2 éventualités et
on a donc : x (1-y) + y (1-x) dans ce cas on a que des TT
- post réduction des 2 couples d'allèles :x.y dans ce cas on a 1/4 de DP, 1/4 de DR et 1/2 de TT
Les fréquences des différents asques sont donc les suivantes :
- fr DP = 1/2 (1 - x) (1 - y) + 1/4 xy
- fr DR = 1/2 (1 - x) (1 - y) + 1/4 xy
- fr TT = x (1 - y) + y(1 - x) + 1/2 xy.
L'indépendance des ségrégations des 2 couples d'allèles se traduit donc par fr DP = fr DR.
Etude de quelques cas particuliers
Puisque nous savons que la fréquence de post-réduction varie de 0 à 2/3 nous allons

30
déterminer les fréquences des DP, DR et TT dans ces cas limites.
1er cas : x = 0 et y = 0
dans ce cas les 2 couples d'allèles sont très proches de leurs centromères respectifs, ils sont
donc toujours pré-réduits, fr DP = fr DR et fr TT = 0
2e cas : x = 0 ou y = 0
dans ce cas un des 2 couples d'allèles est très proche de son centromère, il est donc toujours
pré-réduit. On a fr DP = fr DR de plus si x=O, frTT=y et si y=O, frTT=x
Dans ce cas la fréquence de TT est équivalente à la fréquence de post-réduction du couple
d'allèles distant de son centromère.
La distance de ce couple d'allèles à son centromère sera donc : dg-c = 1/2 % TT
Chez les champignons à tétrades non ordonnées cette situation est intéressante car elle
permet de calculer la distance qui sépare un couple d'allèles de son centromère sans
détecter d'asques post réduits.
3e cas : x= 2/3 et / ou y =2/3 dans ce cas au moins un couple d'allèles est très éloigné de son
centromère. On a fr DP = fr DR avec fr DP = 1/6; fr DR = 1/6; fr TT = 2/3
D'une façon générale il apparaît donc que dans le cas de l'indépendance on a:
- fr DP = fr DR
- 1/6 < fr DP < 1/2 ;
- 1/6 < fr DP < 1/2 ;
- 0 < fr TT < 2/3
2- Cas d'un organisme à tétrades ordonnées : Neurospora crassa
Imaginons 2 couples d'allèles a+/a et b+/b localisés sur 2 paires de chromosomes différents et
à une certaine distance de leurs centromères respectifs.
Soit un croisement entre deux souches P1 et P2 de génotypes respectifs ab+ et a+b.
Etude séparée des 2 couples d'allèles
Dans la descendance de ce croisement la ségrégation de chaque couple d'allèles sera une
ségrégation 1-1. De plus pour chaque couple d'allèles on observera 6 types d'asques : 2 pré-
réduits et 4 post réduits. Ainsi :
- pour le couple d'allèles a+/a la fréquence de post-réduction sera x et la fréquence de pré-
réduction 1-x ;
- pour le couple d'allèles b+/b la fréquence de post-réduction sera y et la fréquence de pré-
réduction 1-y.
Etude des 2 couples d'allèles pris simultanément
Considérons maintenant simultanément les 2 COUples d'allèles, on aboutit dans ce cas à 36
types d'asques différents qui sont présentés dans le tableau ci-après.
Les associations d'allèles
En considérant le croisement parental ab+ x a+b il apparaît que 4 associations d'allèles
peuvent exister ab+ et a+b qui sont des associations parentales ; ab et a+b+ qui sont des
associations recombinées.
Les 3 types d'Asques
Sur la base des associations d'allèles qu'ils renferment, on distingue dans la descendance 3

31
types d'asques :
- ceux qui ne renferment que les 2 associations ab+ et a+b parentales : ils sont appelés Ditype
Parental DP ;
- ceux qui ne renferment que les 2 associations ab et a+b+ recombinées : ils sont appelés
Ditype Recombiné DR ;
- ceux enfin qui renferment les 2 associations ab+ et a+b parentales et les 2 associations ab et
ab et a+b+ recombinés : ils sont appelés Tétratype TT.
ab+ x a+b
Pré réduction de b+/b Post réduction de b
+/b
b b b
+
b+
b+
b+
b b
b b
+
b b
+
b+
b b
+
b
b+
b b b
+
b b
+
b+
b
Pré réduction de a
+/a
a a a
+
a+
DR a b ab
a+b
+
a+b
+
DP ab
+
ab+
a+b
a+b
TT ab ab
+
a+b
a+b
+
TT ab
+
ab a
+b
+
a+b
TT a b
+
ab a
+b
a+b
+
TT ab ab
+
a+b
+
a+b
a+
a+
a a
DP a
+b
a+b
a b+
a b+
DR a
+b
+
a+b
+
ab ab
TT a
+b
a+b
+
ab ab
+
TT a
+b
+
a+b
ab+
ab
TT a
+b
+
a+b
ab ab
+
TT a
+b
a+ b
+
ab+
ab
Post réduction
de a+/a
a a
+
a a
+
TT ab a
+b
ab+
a+b
+
TT ab
+
a+b
+
ab a
+b
DR ab
a+b
+
ab a
+b
+
DP ab
+
a+b
ab+
a+b
TT ab
+
a+b
ab a
+b
+
TT ab
a+b
+
a b+
a+b
a+
a a
+
a
TT a
+b
a b a
+b
+
a b+
TT a
+b
+
ab+
a+ b
ab
DP a
+b
ab+
a+b
ab+
DR a
+b
+
ab a
+ b
+
ab
TT a
+b
+
ab a
+b
ab+
TT a
+b
ab+
a+b
+
ab a
+
a a a
+
TT a
+b
a b a b
+
a+ b
+
TT a
+b
+
ab+
ab a
+b
TT a
+b
ab+
ab a
+b
+
TT a
+b
+
ab ab
+
a+b
DR a
+b
+
ab ab
a+b
+
DP a
+b
ab+
ab+
a+b
a a
+
a+
a
TT a b a
+b
a+b
+
ab+
TT ab
+
a+b
+
a+b
ab
TT ab
a+b
+
a+b
ab+
TT ab
+
a+b
a+b
+
ab
DP ab
+
a+b
a+b
ab+
DR ab
a+b
+
a+b
+
ab
Schéma 21 : Formation des DP,DR et TT dans le cas d’asques ordonnés
Origines des DP, DR et TT
Quels sont les événements à l'origine de ces 3 types d'asques.
L'analyse de la descendance montre que :
- les DP et les DR ont les mêmes origines. On les obtient lorsque les deux couples d'allèles sont
pré-réduits ou post-réduits.
- les TT s'obtiennent après qu'un couple d'allèles soit pré-réduit et l'autre post-réduit ou après

32
que les 2 couples d'allèles soient post-réduits.
Fréquences des DP, DR et TT
Maintenant que nous connaissons les événements à l'origine de ces 3 types d'asques, nous
allons déterminer leurs fréquences respectives.
fr DP = [(1-x)/2 * (1-y)/2]* 2 + [x/4* y/4]* 4 = 1/2 (1-x) (1-y) + 1/4 xy
fr DR = [(1-x)/2 * (1-y)/2]* 2 + [x/4* y/4]* 4 = 1/2 (1-x) (1-y) + 1/4 xy
fr TT = [x/4 *(1-y)/2] * 8 +[y/4*(1-x)/2] * 8 + (x/4 * y/4) * 8 = x(1-y) + y(1-x) + 1/2 xy
II ressort que fr DP = fr DR quelles que soient les fréquences de post réduction x et y des 2
couples d'allèles.
La fr TT dépend essentiellement des post-réductions respectives des 2 couples d'allèles.
Ainsi, lorsque les ségrégations de 2 couples d'allèles sont indépendantes, les fréquences des
DP et DR sont équivalentes et réciproquement.
Etude de quelques cas particuliers
Ces cas sont les mêmes que ceux décrits pour les tétrades non ordonnées. Les conclusions
dégagées sont également les mêmes. Avec le tableau à 36 cases on arrive à mieux visualiser
les situations décrites précédemment.
3 - Analyse de spores en vrac
Qu'il utilise Neurospora ou Ascobolus comme matériel d'étude, l'expérimentateur peut
récolter seulement une population de spores en vrac.
Nous avons vu que dans les cas de tétrades ordonnées et non ordonnées lorsque 2 couples
d'allèles ont une ségrégation indépendante l'un de l'autre la fréquence des DP est égale à celle
des DR, avec une fréquence de TT qui varie de 0 à 2/3
Si on réalise un croisement ab+ x a+b en analysant des tétrades on aurait eu des DP, DR et TT.
Les DP contiennent 4 spores ab+ et 4 spores a+b
Les DR contiennent 4 spores ab et 4 spores a+b+
Les TT contiennent 2 spores ab+; 2 spores a+b ; 2 spores ab ; 2 spores a+b+
Les spores de type parental étant ab+ et a+b et les spores de type recombiné étant ab et a+b+
du fait de l'égalité DP = DR et de la composition des TT, il apparaît que la fréquence des spores
parentales est équivalente à celle des spores recombinées.
La seule difficulté au niveau de l'analyse de spores en vrac réside parfois dans le fait que les 4
génotypes attendus dans le cas de 2 couples d'allèles indépendants ne conduisent pas à 4
phénotypes différents. Les exemples ci-après illustrent ces cas particuliers
Les 2 couples d'allèles contrôlent le même caractère
On connaît chez Ascobolus plusieurs gènes contrôlant la coloration des spores ; ils sont
appelés bl1, bl2 etc...Si on effectue un croisement entre 2 souches mutantes à spores blanches
:bl1bl2+ x bl1 +bl2, on obtiendra :
- 3/4 de spores blanches bl1bl2+ et bl1
+bl2 et bl1bl2
- et 1/4 de spores noires bl1+ bl2
+

33
L'expression d'un couple d'allèles est masquée par celle de l'autre
On connaît chez Ascobolus des souches mutantes présentant des spores dont le pigment est
reparti en sphérules et non uniformément sur la paroi de la spore : ces spores sont dites
granuleuses.
Appelons gr+/gr le couple d'allèles qui est responsable du caractère granuleux et bl+/bl celui
qui est responsable de la coloration.
Si on effectue un croisement entre une souche à spores granuleuses et une souche à spores
blanches : bl+gr x blgr+ on obtient :
- 1/4 de spores granuleuses bl+gr
- 1/4 de spores noires bl+gr+
- et 1/2 de spore blanche blgr+, blgr
L’explication réside dans le fait qu’en présence de l’allèle bl le pigment n’est pas synthétisé,
l’expression de l’allèle gr est masquée et les spores de génotype blgr sont blanches.

34
CHAPITRE III - SEGREGATION DE DEUX COUPLES D'ALLELES LOCALISES SUR LA
MEME PAIRE DE CHROMOSOMES
1- Cas des organismes à tétrades non ordonnés
Analyse des résultats d'un croisement chez Ascobolus entre un mutant à spores rondes et un
mutant à spores granuleuses
Schéma 22 : Descendance composée de DP, DR et TT dans le cas de deux couples d’allèles
localisés sur la même paire de chromosomes

Ce croisement fait intervenir 2 caractères : la forme de la spore et coloration de la spore
Pour le caractère forme, dans chaque asque on a 4 spores rondes et 4 spores ovales,
c'est à dire une ségrégation 1-1 qui permet de conclure que le caractère est sous la
dépendance d'un couple d'allèles r+ /r.
Pour le caractère coloration, dans chaque asque on a 4 spores noires et 4 spores
granuleuses, c'est à dire une ségrégation 1-1 qui permet de conclure que le caractère
est sous la dépendance d'un couple d'allèles g+/g.
Si les 2 couples d'allèles étaient indépendants on aurait dans la descendance autant de
DP que
de DR. Ce n'est pas le cas, DP est supérieur à DR. Comment expliquer cette nouvelle
situation?
Faisons abstraction des asques et intéressons nous uniquement aux spores. En
totalisant toutes les spores présentant le même phénotype sans tenir compte du type
d'asque d'origine, ce qui revient à procéder à une analyse en vrac, on obtient les
résultats suivants :
3174 spores rondes noires rg+
3174 spores ovales granuleuses r+g
842 spores rondes granuleuses rg
842 spores ovales noires r+g+
Les spores de type parental ont un effectif supérieur à celui des spores de type
recombiné.
Tout se passe comme s'il existait une force contraignant les allèles à conserver
préférentiellement les associations qu'ils avaient dans les souches parentales. On dit
que les 2 couples d'allèles sont liés. Par opposition à l'état d'indépendance, on parle
de liaison (en anglais linkage).
La liaison de 2 caractères a été observée par les anglais Bateson et Punnett, dès
1906, dans des expériences sur les pois de senteur.
C'est Morgan qui en 1910 a postulé à partir d’expériences chez la drosophile que les
couples d'allèles qui n’ont pas une ségrégation indépendante sont physiquement
liées du fait de leur localisation sur la même paire de chromosomes. Il a imaginé
aussi que les produits recombinés pouvaient cependant apparaître grâce aux
crossing-over se produisant dans l'intervalle qui sépare les 2 couples d'allèles
considérés.
Du fait de la faible fréquence de crossing-over dans une population de méioses, les
types recombinés auront une fréquence inférieure à celle des types parentaux.
On se souvient que dans l'analyse des spores en vrac dans le cas d'une indépendance
P = R de sorte que le % de recombinaison est égale à 50 %. Dans le cas de la liaison
on voit que le % recombinaison sera inférieur à 50 % .Dans notre exemple le calcul de
ce pourcentage de recombinaison est le suivant :
% recombinaison = x 100 =20,9%

36
La distance entre r+/r et g+/g est donc de 20,9 UR.
Maintenant que nous savons que les 2 couples d'allèles r+/r et g+/g sont portés par la
même paire de chromosomes, déterminons l'origine des différents types d'asques et
calculons la distance qui sépare r+/r de g+/g.
Le croisement réalisé est : rg+ x r+ g et le zygote formé a pour génotype r+g/rg+
Dans une population de méioses on peut avoir les événements suivants :
- aucun crossing-over ne se produit entre r+/r et g+/g, dans ce cas on aboutit à 4
produits parentaux : 2 produits rg+ et 2 produits r+ g. Les asques sont des DP.
- Schéma 23 : Formation de DP résultant de l’abscence de crossing-over entre les deux couples
d’allèles
- un crossing-over se produit entre r+r et g+/g
Dans ce cas on aboutit à 2 produits parentaux rg+ et r+g et à 2 produits recombinés
r+g+ et rg. Les 4 produits sont différents. Les asques sont des TT.
- Schéma 24 : Formation de TT résultant de l’existence d’un crossing-over entre les deux couples
d’allèles
- deux crossing-over se produisent entre r+/r et g+/g
Lorsque 1 crossing-over se produit, le second peut se produire de manière aléatoire
selon 4 modalités. On aboutit donc à 4 situations équiprobables.
- les 2 crossing-over touchent 2 chromatides et les 4 remaniements
chromatidiques aboutissent à 4 produits parentaux : 2 produits rg+ et 2 produits
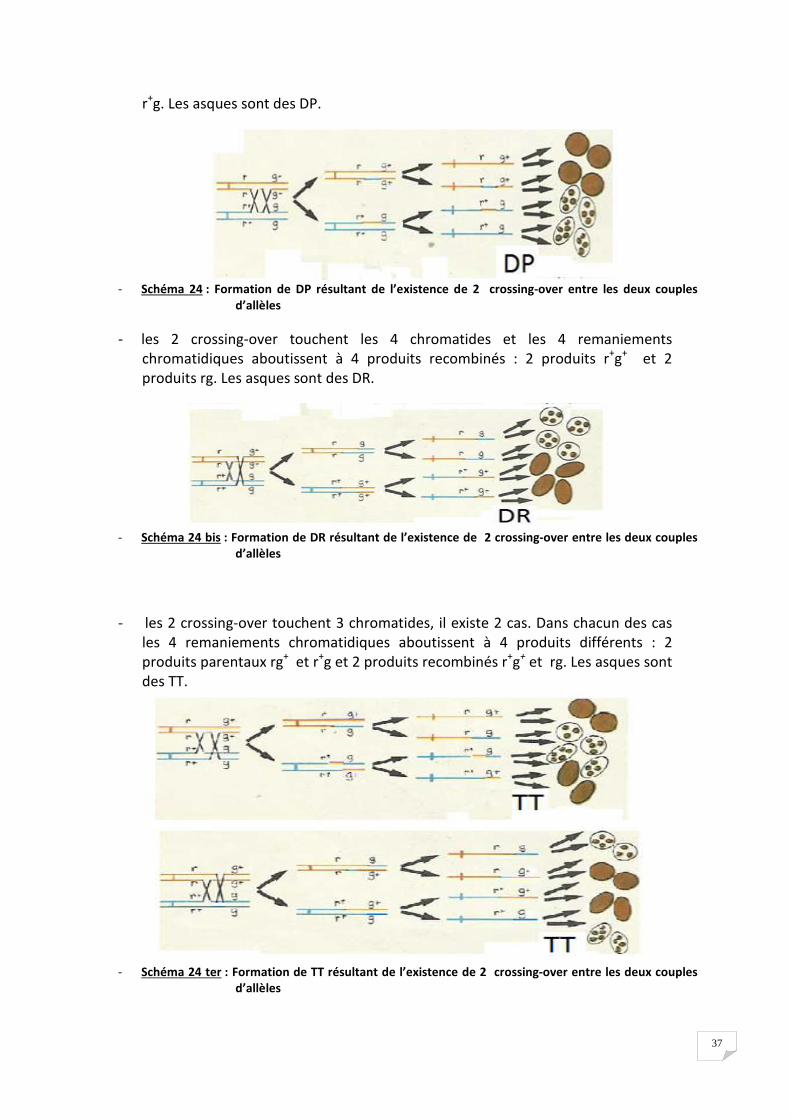
37
r+g. Les asques sont des DP.
- Schéma 24 : Formation de DP résultant de l’existence de 2 crossing-over entre les deux couples
d’allèles
- les 2 crossing-over touchent les 4 chromatides et les 4 remaniements
chromatidiques aboutissent à 4 produits recombinés : 2 produits r+g+ et 2
produits rg. Les asques sont des DR.
- Schéma 24 bis : Formation de DR résultant de l’existence de 2 crossing-over entre les deux couples
d’allèles
- les 2 crossing-over touchent 3 chromatides, il existe 2 cas. Dans chacun des cas
les 4 remaniements chromatidiques aboutissent à 4 produits différents : 2
produits parentaux rg+ et r+g et 2 produits recombinés r+g+ et rg. Les asques sont
des TT.
- Schéma 24 ter : Formation de TT résultant de l’existence de 2 crossing-over entre les deux couples
d’allèles

38
Ainsi lorsqu'il se produit 2 crossing-over entre r+/r et g+/g on aboutit à :
1/4 de DP, 1/4 de DR et 1/2 TT
Pour une population de méioses, lorsqu'on considère les 3 événements 0, 1, 2
crossing-over compte tenu de la fréquence faible de 1 crossing-over, à fortiori de 2
CO, il est clair que :
fr DP > fr DR.
Ainsi, lorsque 2 couples d'allèles sont liés, la fréquence des DP est supérieure à
celle des DR et réciproquement.
Calcul de la distance génétique
dg = x 100 (UR)
dg= [ 2TT1co+4DP2co+4DR+4 TT2co]/4(DP+DR+TT) x 100 (UR)
Les DP dûs à 2 CO et les DP dûs à 0 CO sont confondus.
Il en est de même des TT dûs à à 1 CO et à 2 CO.
Cependant on peut facilement estimer les fréquences des DP2c.o , des TT2c.o, et
TT1c.o.
Nous savons que lorsque 2 crossing-over se produisent on aboutit à 1/4 de DP ; 1/4
de DR et 1/2de TT. Puisque les DR ne résultent que de cet évènement on peut
conclure que :
DP2c.o , = DR2c.o = DR
TT1c.o. = 2 DR2c.o = 2 DR
II s'en suit que TT1C.O =TT- 2 DR
Ainsi la distance devient :
dg = [2(TT-2DR)+4DR+4DR+4(2DR)]/4(DP+DR+TT)x100 (UR)
dg = (2TT+ 12 DR)/ 4(DP+DR+TT)x 100 (UR)
dg = 2TT/4(DP+DR+TT) x100 + 12 DR/4(DP+DR+TT) x100 (UR)
dg = 1/2 %TT + 3 % DR
Dans notre exemple
dg =1/2x[331/(628+331+45)]x100 + 3 x [45/(628+331+45)] x 100 (UR)
dg = 29,7 UR
REMARQUES

39
- Le calcul de la distance à partir des recombinés de la descendance de spores en
vrac était de 20,9 UR ; dans ce cas il n'a pas évidemment été possible d'effectuer une
correction. On a donc une sous estimation qui est dans cet exemple de 9 UR ce qui
représente presque 30 % d'erreur.
- Lorsque 2 couples d'allèles sont liés, nous avons vu que les asques DR
n'apparaissent qu'après que 2 crossing-over interviennent dans l'intervalle qui
sépare ces 2 couples.
-si fr DR = 0 cela signifiera que dans l'intervalle considéré il ne peut se produire
qu'un seul crossing-over ; dans ce cas : dg = 1/2 % TT. II en ressort que les
fréquences des DR et des TT seront d'autant plus faibles que les 2 couples
d'allèles seront rapprochés.
- si les 2 couples d'allèles sont éloignés et que des crossing-over multiples sont
possibles, les 2 couples d’allèles subissent une ségrégation indépendante l’un de
l’autre. Dans ces conditions nous avons fr DP = fr DR. On dit dans ce cas que les 2
couples d'allèles sont physiquement liés parce que portés par la même paire de
chromosome mais indépendants génétiquement.
2- Cas des organismes à tétrades ordonnées
Tout ce qui vient d'être dit à propos de 2 couples d'allèles liés chez Ascobolus
immersus s'applique chez les autres champignons ascomycètes tels que Neurospora
et Sordaria chez lesquels on distingue également des DP DR et TT. Chez ces
champignons l'ordre des spores dans les asques permettra de préciser les relations
gène-centromère.
Lorsque les 2 couples d'allèles sont liés physiquement et indépendants
génétiquement il apparait que la fr DP = fr DR = 1/6 et fréquence TT = 4/6 (voir ce
calcul au niveau du tableau à 36 cases relatif à 2couples d’allèles indépendants)
3- Cas de spores en vrac
L’analyse porte sur une population de spores récoltées à partir de tétrades
ordonnées ou non ordonnées.
Lorsque la descendance est en tétrades on a montré que la fréquence des DP est
nettement supérieure à la fréquence des DR.
Lorsque les 2 couples d’allèles sont liés compte tenu de la composition des différents
types d’asques, on en déduit que le nombre de spores parentales est également
supérieur au nombre de spores recombinées. La distance entre les 2 couples d’allèles
est donc égale au pourcentage de spores recombinées.

40
CHAPITRE IV : LES CARTES FACTORIELLES
1- Définition
La constance de la fréquence de post-réduction pour chaque couple d'allèles et la
constance de la fréquence de recombinaison entre 2 couples d'allèles liés, montrent
que chaque gène occupe, sur un chromosome, une place fixe : son locus.
Ce fait permet d'ordonner les gènes les uns par rapport aux autres et de dresser des
cartes génétiques ou cartes factorielles.
Les distances sur les cartes factorielles sont mesurées par les fréquences de
recombinaison.
Groupe de liaison
On définit un groupe de liaison comme un ensemble de gènes qui apparaissent liés
entre eux et indépendants de tous les autres gènes. Le support matériel d'un groupe
de liaison est un chromosome.
Chez les organismes qui sont très étudiés et chez lesquels on connaît de nombreux
gènes, le nombre de groupes de liaison qu'on peut mettre en évidence est égal au
nombre haploïde de chromosomes caractéristique de l'espèce. Chez Neurospora
crassa où n = 7 on constate l'existence de 7 groupes de liaison.
2- Etablissement d'une carte factorielle
Croisement bifactoriel
Supposons que nous ayons à ordonner 3 gènes A, B et C appartenant au même
groupe de liaison.
Pour le faire on réalise des croisements qui implique pour chacun, à la fois 2 gènes
on parle de croisement bi factoriel. Si les croisements réalisés montrent que la
distance entre A et B est de 5 UR et que celle entre A et C est de 7 UR, il se
présentera deux possibilités pour ranger les gènes.
Si le croisement impliquant à la fois les gènes B et C montre qu'ils sont séparés de 12 UR
on retient l'ordre BAC. Par contre si la distance est de 2 UR on retient l'ordre ABC.
Cette méthode simple qui permet d'ordonner linéairement les gènes est basée sur
l'additivité des distances. Elle n'est valable que si les gènes sont relativement

41
proches. Lorsque les gènes sont très proches cette méthode ne permettra pas de les
ordonner du fait des fluctuations statistiques des distances calculées.
Lorsque les gènes sont très éloignés, les distances calculées ne seront plus additives.
Elles sont sous-estimées car les fréquences de recombinaison ne traduisent pas en
totalité les remaniements chromatidiques du fait de l'existence de crossing-over
multiples.
Il ressort donc que les croisements bi-factoriels sont insuffisants pour établir sans
ambiguïté l'ordre des gènes le long du chromosome.
3- Croisement tri-factoriel ou test à 3 points
La méthode qui permet d'établir les ordres respectifs des gènes est un croisement
impliquant à la fois 3 gènes appelé croisement tri-factoriel ou test à 3 points.
Pour trois couples d’allèles liés a+/a, b+/b et c+/c, un croisement tri-factoriel sera un
croisement du type abc x a+b+c+ ou encore ab+c x a+bc+ ou toute autre combinaison
génotypique. Il suffit que la cellule diploïde qui subit la méiose soit hétérozygote
pour les 3 couples d'allèles.
Faisons l’hypothèse que les 3 couples d’allèles sont liés selon l’ordre suivant l a+/a -
b+/b - c+/c qui équivaut également à l’ordre c+/c - b+/b - a+/a. On dira simplement
que l'ordre est tel que le couple d’allèles b+/b est en position médiane et que les
couples d’allèles a+/a et c+/c sont en positions extrêmes.
Soit le croisement abc x a+b+c+.
La cellule diploïde qui subit la méiose est hétérozygote abc//a+b+c+
Un tel hétérozygote produira 8 gamètes après les évènements suivants :
- aucun CO ne se produit entre a+/a – b+/b et entre b+/b – c+/c ; on obtient 2 gamètes
de type parental abc et a+b+c+ ;
- 1 CO se produit entre a+/a – b+/b et aucun CO ne se produit entre b+/b – c+/c ; on
obtient 2 gamètes de type recombiné ab+c+ et a+bc
- aucun CO ne se produit entre a+/a – b+/b et 1 CO se produit entre b+/b – c+/c, on
obtient 2 gamètes de type recombiné abc+ et a+b+c

42
- 1 CO se produit entre a+/a – b+/b et 1 CO se produit entre b+/b – c+/c on obtient 2
gamètes de type recombiné ab+c et a+bc+
La descendance du croisement abc x a+b+c+ sera composée comme suit :
CLASSE I : a b c et a+b+c+ parentaux
CLASSE II : ab+c et abc simple recombinés
CLASSE III : a b c+ et a+b+c simple recombinés
CLASSE IV : ab+c et a+bc+ double recombinés
Chaque classe comporte 2 types de gamètes complémentaires qui ont des fréquences ou
des effectifs voisins ; les différences n'étant dûes qu'à des fluctuations
d'échantillonnage.
Les 4 classes ont des fréquences ou des effectifs différents. Compte tenu des
événements à l'origine de la formation de ces gamètes il apparaît que :
- la classe I des parentaux : abc, a+b+c+ est la plus fréquente ;
- la classe IV des doubles recombinées : ab+c, a+bc+ est 1 moins fréquente
Principe de l'établissement de l'ordre des gènes
L'établissement de l'ordre des gènes est basé sur le fait que les gènes en positions
extrêmes présentent les mêmes associations d'allèles dans les classes des parentaux
et des doubles recombinés. Dans l'exemple, les 2 couples d'allèles a+/a et c+/c
présentent les mêmes associations ac et a+c+ dans les classes I et IV. On en déduit
qu'ils sont en positions extrêmes.
Calcul des distances génétiques
distance entre a /a et b /b = fr ab+c+ + fr a+bc + fr ab+ c + fr a+bc+ x 100.
distance entre b /b et c /c = fr abc+ + fr a+ b+c + fr ab+ c + fr a+bc + x 100
distance entre a/a et c/c = fr ab+c+ + fr a+bc + abc+ + fr a+b+c + 2 fr ab+c + 2 fr a+bc+ x
100.
Il en ressort que les distances deviennent additives du fait de la correction apportée
dans le calcul de la distance entre les 2 couples d'allèles extrêmes.
4- Notion d'interférence
De nombreuses expériences réalisées chez la plupart des organismes bien étudiés,
ont révélé que les crossing-over ne sont pas des événements indépendants.
Lorsqu'un crossing-over se produit en un point, très souvent, il diminue les chances
d'apparition d'un second dans son voisinage : ce phénomène est appelé
l'interférence chromosomique.
Le résultat tangible de cette interférence est que le nombre de double crossing-over
observés dans un test à 3 points est inférieur à celui que l'on peut attendre compte
tenu des distances génétiques et de l'hypothèse de l'indépendance des événements
de crossing-over.
L'existence de l'interférence ne modifie pas les distances génétiques puisque les loci
des gènes sont fixes.
Par contre les fréquences des simples recombinés dûs à un crossing-over seront
augmentées d'autant que les fréquences des doubles recombinés et des parentaux
sont diminuées.

43
On évalue l'interférence grâce aux relations suivantes :
- Interférence + coïncidence = 1
- Coincidence= % doubles crossing-over observés/% doubles crossing-over calculés
Exemple
On considère que sur ce segment de 30 UR l'interférence est de 40 %.
Quelles sont les fréquences des différents génotypes obtenus dans la descendance
d'un croisement a+b+c+ x abc ?
-------------------------
L'interférence étant égale à 40 %, cela signifie que 40 % des doubles crossing-over
attendus apparaîtront sous forme de simples crossing-over. Ainsi 0,20 x 0,10 x 0,40 =
0,008 ou 0,8 % des double crossing-over attendus, apparaîtront sous forme de
simples crossing-over.
Comme l'interférence est égale à 40 % on en déduit que la coïncidence est égale à 1-
0,40 = 0,60 ou 60 %. Cela signifie que 60 % des doubles crossing-over attendus sont
observés.
Ainsi 0,20 x 0,10 x 0,60 = 0,012 ou 1,2 % de doubles crossing-over, seront observés.
Calculons les fréquences des différents génotypes de la descendance.
- doubles recombinés a+bc+ et ab+c : 0,20 x 0,10 x 0,60 = 0,012 ; a+bc+ = 0,006 ab+c
= 0,006
- simples recombinés a+bc et ab+c+ : 0,20 - 0,012 = 0,188 ; a+bc = 0,094 ab+c+ =
0,094
- simples recombinés a+b+c et abc : 0,10 - 0,012 = 0,088 : a+b+c = 0,044 abc''" =
0,044
- parentaux a+b+c+ et abc : 1-(0,188 + 0,088 + 0,012) = 0,712 a+b+c+ = 0,356 abc =
0,356
Si l'interférence était nulle c'est à dire que la coïncidence était égale à on aurait les
fréquences suivantes pour les différents génotypes :
- doubles recombinés a+bc+ et ab+c : 0,20 x 0,10 = 0,020 ; a+bc+ = 0,01 ab+c = 0,01
- simples recombinés a+bc et ab+c+ : 0,20 x 0,90 = 0,180; a+bc = 0,09 ab+c+ = 0,09
- simples recombinés a+b+c et abc : 0,80 x 0,10 = 0,080 : a+b+c = 0,04 abc''" = 0,04
- parentaux a+b+c+ et abc : 0,80 x 0,90 = 0,720 : a+b+c+ = 0,36 abc = 0,36

44
Lorsque l'on compare les fréquences obtenues dans les 2 cas il apparaît que quand
l'interférence est égale à 40% les fréquences des simples recombinés augmentent
d'un facteur 0,008 alors que les doubles recombinés et les parentaux diminuent de
ce même facteur. Ce facteur correspond à la fréquence des doubles recombinés
attendus qui apparaissent sous forme de simples recombinés dûs à 1 crossing-over
du fait de l'interférence de 40 % : 0,20 x 0,10 x 0,40 = 0,008.
Ainsi les fréquences des simples recombinés dûs à un crossing-over sont augmentées
d'autant que les fréquences des doubles recombinés et des parentaux sont
diminuées.
Exercice d’application
Chez Neurospora, la souche sauvage de référence est prototrophe à spores noires et
ovales. Le croisement entre 2 souches mutantes a donné une descendance
composée de :
373 spores noires ovales tyr-
362 spores blanches rondes tyr+
83 spores blanches ovales tyr+
72 spores noires rondes tyr-
57 spores noires rondes tyr+
48 spores blanches ovales tyr-
4 spores noires ovales tyr+
1 spore blanche rondes tyr-
Comment interprétez ces résultats ?
Solution
Ce croisement fait intervenir 3 caractères : la coloration de la spore, la forme de la
spore et l'auxotrophie par rapport à la tyrosine. L'analyse réalisée est celle des
spores en vrac.
Analyse caractàre par caractère
Pour le caractère coloration de la spore, la descendance se compose de :
Classes I= 0 et
C=1
I= 0,40 et
C=0,60
Différence
entre I=0
et I=0,40
Doubles recombinés
a+bc+ et ab+c
0,020 0,012 - 0,008
Simples recombinés
a+bc et ab+c+
0,180 0,188 + 0,008
Simples recombinés
a+b+c et abc
0,080 0,088 + 0,008
Parentaux
a+b+c+ et abc
0,720 0,712 - 0,008

45
[blanche] : 362 + 83 + 48 + 1 = 494
[noire] : 373 + 72 + 57 + 4 = 506
II y a autant de spores blanches que de spores noires c'est à dire une ségrégation 1-1
qui permet de conclure que ce caractère est gouverné par un couple d'allèles b+/b.
Pour le caractère forme de la spore, la descendance se compose de :
[ovale] : 373 + 83 + 48 + 4 = 508
[ronde] : 362 +72+57+1 = 492
II y a autant de spores ovales que de spores rondes, c'est à dire une ségrégation 1-1
qui permet de conclure que ce-caractère est gouverné par un couple d'allèles r+/r.
Pour le caractère auxotrophie par rapport à la tyrosine, la descendance se compose
de :
[tyr+] = 362 + 83 + 57 + 4 = 506
[tyr-] = 373 +72+ 48 + 1 = 494
II y a autant de spores tyr+ que de spores tyr-, c'es à dire une ségrégation 1-1 qui
permet de conclure que ce caractère est gouverné par un couple d'allèles tyr+ /tyr-
Trois couples d'allèles interviennent donc dans ce croisement. Si ces 3 couples
étaient indépendants, nous aurions eu une descendance composée de 8 types de
spores ayant la même
fréquence 1/8 donc le même effectif c’est à dire 1000 x 1/8 = 125
Cette descendance est composée de 4 classes sur la base des effectifs. Nous pouvons
donc envisager l'hypothèse de 3 couples d'allèles liés ; dans cette hypothèse le
croisement réalisé serait un test à 3 points.
Les effectifs 373 et 362 sont ceux des phénotypes de la classe des parentaux. Les
effectifs 4 et 1 sont ceux des phénotypes de la classe des boubles-recombinés.
Ordre des gènes
Déterminons l'ordre des gènes à partir de la classe des parentaux et de celle des
doubles recombinés. On a:
373 b+ r+ tyr-
362 b r tyr+
4 b+ r+tyr+
1 b r tyr-
II apparaît que les 2 couples d'allèles b+/b et r+/r présentent les mêmes associations
d'allèles b+r+ et br dans les 2 classes (parentaux et doubles recombinés) ; ces 2
couples d'allèles sont donc en positions extrêmes, le couple tyr /tyr est en position
médiane.
L'ordre est donc : b+/b - tyr+/tyr- - r+/r
En tenant compte de cet ordre la descendance est la suivante:
373 b+ tyr-r+
362 b tyr+r
83 b tyr+r+
72 b+tyr-r
57 b+tyr+r

46
48 b+tyr+r+
4 b+tyr+r+
1 b tyr-r
Les 2 souches mutantes parentales sont donc : b+ tyr-r+ et b tyr+r
On a réalisé le croisement b+ tyr-r+ x b tyr+r.
Calculons les distances respect ives qui séparent le 3 couples d'allèles.
Distance entre b+/b et tyr+/tyr d = [(57+48+4+1)/1000] x 100 (UR) =
11 UR
distance entre tyr+/tyr et r+/r d = [(83+72+4+1)/1000] x 100 (UR) = 16
UR
distance entre b+ /b et r+ /r d =
[(57+48+83+72+2(4+1))/1000] x100 (UR) = 27 UR
Les 3 couples d'allèles sont donc liés selon la carte factorielle :
Existe-t-il une interférence chromosomique ?
On sait que :
- Coïncidence = % doubles crossing-overobservés/% doubles crossing-
overobservés
- Coïncidence + Interférence = 1
- Interférence = 1 - Coïncidence
Coincidence = [(4+1)/1000]/0,11 x0,16 = 28,4 %
Donc Interférence = 1 - 0,284 = 0,716
Interférence = 71 ,6 %
Ainsi 71,6 % des doubles crossing-over attendus compte tenu des distances génétiques,
ne sont pas observés, ils apparaissent sous forme de simples crossing-over


















