
Génétique évolutive et des...
34
Génétique évolutive et des populations Nicolas Lartillot 16 janvier 2012 Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 1 / 34
Transcript of Génétique évolutive et des...

Génétique évolutive et des populations
Nicolas Lartillot
16 janvier 2012
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 1 / 34

1 Génétique évolutive : traitement déterministeCas haploideCas diploide
2 Dynamique adaptative
3 Génétique quantitative des traits complexes
4 Génétique des populations. La dérive génétiquele modèle de Wright et FisherCoalescent et diversité
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 2 / 34

Génétique évolutive : traitement déterministe
Génétique évolutive
Objet généralrendre compte de l’évolution phénotypique en termes dechangement de fréquences des gènes et génotypes dans lespopulationsen premier lieu, modèles déterministes (larges populations)calculer l’évolution de la fréquence de plusieurs allèles au coursdes générations, en fonction de la valeur sélective de chacunutiliser ces modèles pour étudier l’évolution d’une espèce, ou deplusieurs espèces en interaction
Ronald A. Fisher, Sewall Wright, John B.S. Haldane, ...John Maynard Smith. Evolutionary Genetics, 2d edition (1998)
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 3 / 34

Génétique évolutive : traitement déterministe
Sélection naturelle = compétition intraspécifique
R. Butler, copyright mangabey.com
Sélection requiertvariation phénotypique héritablecompétition
entre individus d’une même espèce(pousser plus haut que ses congénères)
entre allèles en un même locus
Attention aux arguments fallacieuxsélection de groupe(selection ne travaille pas pour le bien de l’espèce)
arguments panglossiens(tout n’est pas dû à de l’adaptation)
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 4 / 34

Génétique évolutive : traitement déterministe
Sélection directionnelle : mélanisme industriel
Biston bitulariaWikimedia commons
genet.univ-tours.fr
Test de l’hypothèse industrielle (Kettlewell, 1955)zone non polluée (Dorsey) zone polluée (Birmingham)carbonaria typica carbonaria typica
relâchés 473 496 154 64recapturés 30 62 82 16proportion rec. 6,3 % 12,5 % 52,3 % 25,0 %
genet.univ-tours.fr (excellent site pédagogique génétique/évolution)Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 5 / 34

Génétique évolutive : traitement déterministe
Fitness et coefficient de sélectionFitness ou valeur sélective d’un allèle
contribution moyenne à la génération suivante des individusporteurs de l’allèle.sélection de viabilité : fitness = taux de survieforme sauvage (typica) : w1
forme mutante (carbonaria) : w1 ' 0,5w1 en zone non polluée,w2 ' 2w1 en zone polluée.
Coefficient de sélectiondifférence relative entre mutant et sauvage s = (w2 − w1)/w1
s = −0.5 en zone non polluée, s = 1 en zone polluées < 0 : sélection négative, ou purificatrices > 0 : sélection positive, ou directionnelles = 0 : régime neutre (pas de différence sélective entre 2 allèles)
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 6 / 34

Génétique évolutive : traitement déterministe Cas haploide
Sélection directionnelle, cas haploide : formalisationHypothèses
population haploide, asexuéegénérations discrètes (t = 1..T )
fluctuations stochastiques négligées2 allèles A1 et A2 au locus considéréfitness allèle sauvage w1, mutant w2 = w1(1 + s), s > 0
Position du problèmeallèle sauvage : N1(t), allèle mutant N2(t), (N1(t) + N2(t) = N(t)).proportion allèle sauvage : p(t) = N1(t)/N.proportion allèle mutant : q(t) = N2(t)/N (p(t) + q(t) = 1).calculer q(t) en fonction de t et s(variation de fréquence de l’allèle mutant due à la sélection)
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 7 / 34
Génétique évolutive : traitement déterministe Cas haploide
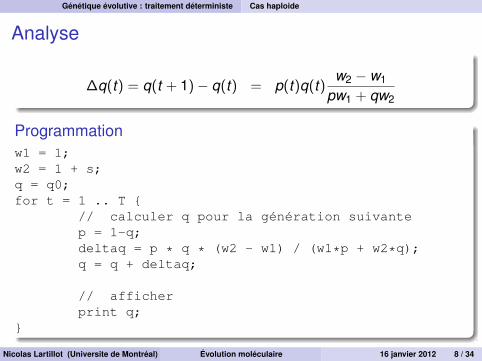
Analyse
∆q(t) = q(t + 1)− q(t) = p(t)q(t)w2 − w1
pw1 + qw2
Programmationw1 = 1;w2 = 1 + s;q = q0;for t = 1 .. T {
// calculer q pour la génération suivantep = 1-q;deltaq = p * q * (w2 - w1) / (w1*p + w2*q);q = q + deltaq;
// afficherprint q;
}
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 8 / 34
Génétique évolutive : traitement déterministe Cas haploide
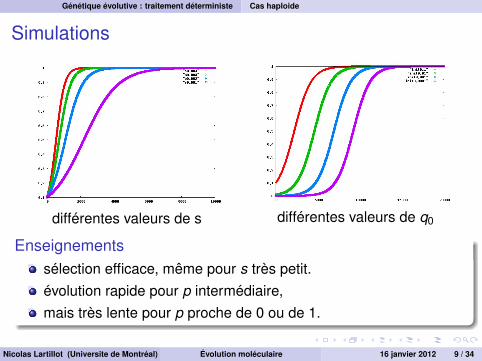
Simulations
différentes valeurs de s différentes valeurs de q0
Enseignementssélection efficace, même pour s très petit.évolution rapide pour p intermédiaire,mais très lente pour p proche de 0 ou de 1.
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 9 / 34

Génétique évolutive : traitement déterministe Cas haploide
approximation en sélection faible (s << 1)générations discrètes
∆q = spq = sq(1− q)
limite continue :
dqdt
= sq(1− q)
solution :
p(t) =1
1− p(0)q(0)e
−st
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 10 / 34

Génétique évolutive : traitement déterministe Cas diploide
Population diploïde sexuée
Sélection chez les diploidesA11, fréquence P11, fitness w11 = 1A12, fréquence P12, fitness w12 = 1 + hsA22, fréquence P22, fitness w22 = 1 + s
h : coefficient de dominance
différents modes de dominanceh = 0 : allèle A2 récessifh = 1/2 : allèle A2 codominanth = 1 : allèle A2 dominanth > 1 : allèle surdominant (avantage hétérozygote)
tous les intermédiaires sont possibles
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 11 / 34

Génétique évolutive : traitement déterministe Cas diploide
Équilibre de Hardy-Weinberg
Principeallèles A1, A2, fréquences p + q = 1fréquences des individus A1A1 (P11), A1A2 (P12), A2A2 (P22),P11 + P12 + P22 = 1.en panmixie :
P11 = p2
P12 = 2pqP22 = q2
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 12 / 34

Programmationw11 = 1;w12 = 1 + h*s;w22 = 1 + s;
// initialiser qq = q0;
for t = 0.. T {
p = 1 - q;
// fréquences des génotypes (Hardy Weinberg)P11 = p * p;P12 = 2 * p * q;P22 = q * q;
// fréquences des génotypes après sélectionPP11 = P11 * w11;PP12 = P12 * w12;PP22 = P22 * w22;
// récupérer fréquences alléliques après sélectionpp2 = 2*PP11 + PP12;qq2 = 2*PP22 + PP12;
// normalisertotal = pq2 + qq2;q = qq2 / total;
// afficherprint q;
}
Génétique évolutive : traitement déterministe Cas diploide
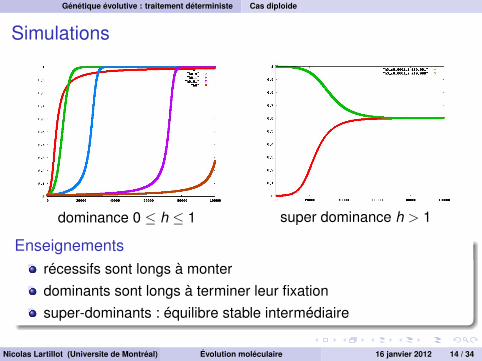
Simulations
dominance 0 ≤ h ≤ 1 super dominance h > 1
Enseignementsrécessifs sont longs à monterdominants sont longs à terminer leur fixationsuper-dominants : équilibre stable intermédiaire
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 14 / 34

Dynamique adaptative
Dynamique adaptative (analyse par invasion évolutive)Principe
une population initialeun allèle mutant apparaîtcet allèle modifie un trait phénotypique ou comportementaldéterminer si cet allèle peut envahir la population (se fixer)états évolutivement stables (qu’aucun mutant ne peut envahir)
Applicationsévolution du sex-ratio, de l’histoire de viemodèles d’évolution altruisme / tricheursinteractions proie/prédateur ou hôte/parasiteévolution de la parthénogénèse
John Maynard Smith (Evolution and the theory of games)
Richard Dawkins (The selfish gene, le gène égoiste)
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 15 / 34

Dynamique adaptative
Sélection diversifiante
Cepaea nemoralisTaylor, J. W. (1894-1914) : Monograph of the land and freshwater Mollusca of the British Isles
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 16 / 34

Dynamique adaptative
Sélection sexuelle
Teleopsis
compétition entre individus du même sexe en vue de lareproductionconflits entre mâleschoix par la femelle (good-gene and sexy-son hypotheses)
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 17 / 34

Dynamique adaptative
L’hypothèse de la reine rouge (L. van Valen, 1973)
"In this place it takes all therunning you can do, to keep in
the same place"(Lewis Carroll, Through the looking glass)
John Tenniel, 1871, Alice et la Reine rouge.
Course aux armements perpétuelle entrehotes-parasitesproies-prédateursespèces concurrentes
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 18 / 34
Génétique quantitative des traits complexes
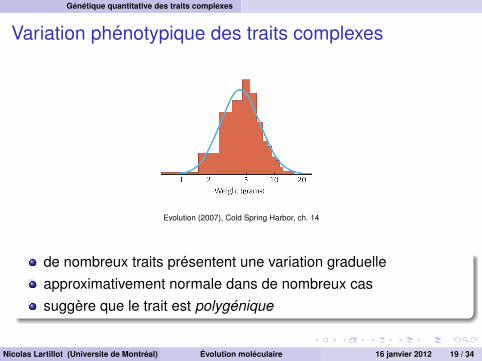
Variation phénotypique des traits complexes
Evolution (2007), Cold Spring Harbor, ch. 14
de nombreux traits présentent une variation graduelleapproximativement normale dans de nombreux cassuggère que le trait est polygénique
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 19 / 34
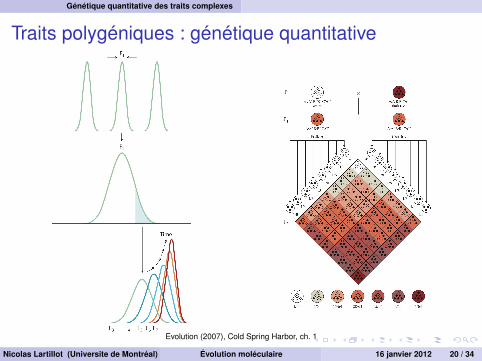
Génétique quantitative des traits complexes
Traits polygéniques : génétique quantitative
Evolution (2007), Cold Spring Harbor, ch. 1
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 20 / 34

Génétique quantitative des traits complexes
Génétique quantitative
Objetcomprendre comment la sélection modifie un trait quantitatifpolygénique, sans connaître les détails moléculairesdiscriminer entre variation génétique et environnementaleestimer le nombre de locus déterminant le trait, et leur importancerespectivedéterminer efficacité des schémas de sélection artificielle
Méthodesmesurer corrélations phénotypiques entre individus apparentésmesurer réponse des populations à de la sélection artificielleexpliquer ces mesures par des modèles statistiques multi-locus
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 21 / 34

Génétique des populations. La dérive génétique
Effets stochastiques
Génétique des populationscomprendre les mécanismes de l’évolution à l’échelleintraspécifiqueen population finie, effets stochastiques (dérive génétique) sontimportantsgénétique des populations nécessite des outils probabilistesétude de l’évolution des fréquences alléliques en fonction de lasélection et de la dérive (et de la mutation)mesurer l’importance relative des effets stochastiques et de lasélection
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 22 / 34

Le modèle de Wright et Fisher : cas neutre
Time
Hypothèsespopulation panmictique et de taille constanteà chaque génération, tirage binomial du nombre de descendantsde chaque individu
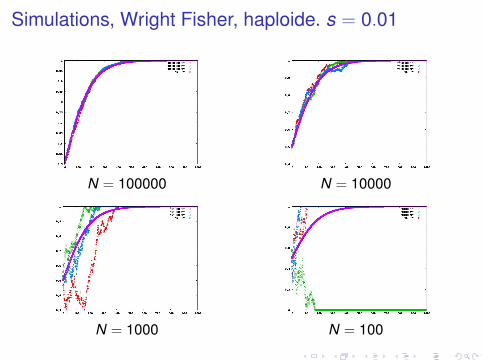
Simulations, Wright Fisher, haploide. s = 0.01
N = 100000 N = 10000
N = 1000 N = 100
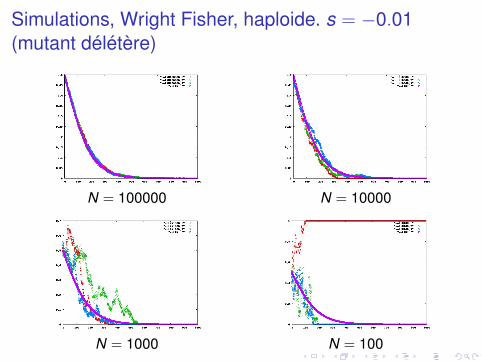
Simulations, Wright Fisher, haploide. s = −0.01(mutant délétère)
N = 100000 N = 10000
N = 1000 N = 100

Génétique des populations. La dérive génétique le modèle de Wright et Fisher
La dérive génétique
large population : loi des grands nombres, fluctuations sontmoyennéespetites populations : fluctuations très importantesen petite population, un allèle avantageux peut être perdusurtout, en petite population, un allèle délétère peut se fixer
Probabilité de fixation
pfix (s,N,q0)
dépend de la sélection, la taille efficace, et la fréquence de départnouveau mutant : q0 = 1/N en haploide, 1/2N en diploide
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 26 / 34

Génétique des populations. La dérive génétique le modèle de Wright et Fisher
Fixation d’un allèle neutre initialement à 1 copie
www.coalescent.dk
haploide
pfix (0) =1N
diploide
pfix (0) =1
2N
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 27 / 34
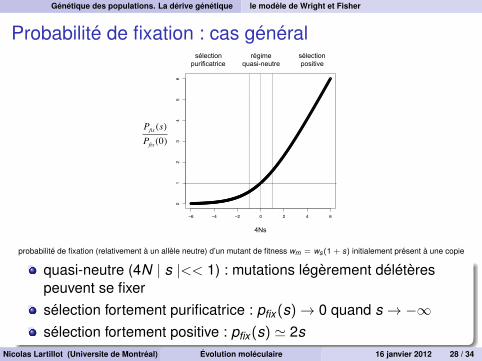
Génétique des populations. La dérive génétique le modèle de Wright et Fisher
Probabilité de fixation : cas général
!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!
!!!!!!!!!!!!!!!!!!!
!!!!!!!!!!!!!
!!!!!!!!!!
!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!!
−6 −4 −2 0 2 4 6
01
23
45
6
S
P
régime quasi-neutre
sélection positive
sélection purificatrice
4Ns
!
Pfix (s)Pfix (0)
probabilité de fixation (relativement à un allèle neutre) d’un mutant de fitness wm = ws(1 + s) initialement présent à une copie
quasi-neutre (4N | s |<< 1) : mutations légèrement délétèrespeuvent se fixersélection fortement purificatrice : pfix (s)→ 0 quand s → −∞sélection fortement positive : pfix (s) ' 2s
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 28 / 34

Génétique des populations. La dérive génétique Coalescent et diversité
Inferer taille efficace N à partir de donnéeshaplotypiques
Hein, Shierup and Wiuf, 2005
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 29 / 34

Retracer le dernier ancêtre commun
www.coalescent.dk
prob. de coalescence à la génération précédente 1/(2N)
temps moyen depuis l’ancêtre T = 2N.
Génétique des populations. La dérive génétique Coalescent et diversité
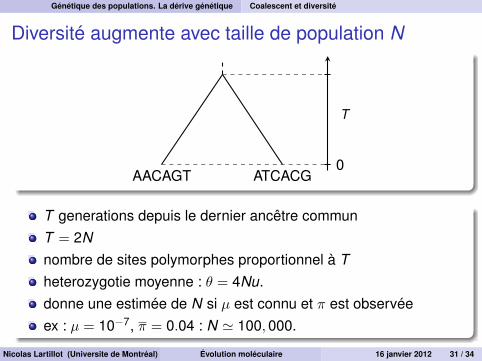
Diversité augmente avec taille de population N
AACAGT ATCACG0
T
T generations depuis le dernier ancêtre communT = 2Nnombre de sites polymorphes proportionnel à Theterozygotie moyenne : θ = 4Nu.donne une estimée de N si µ est connu et π est observéeex : µ = 10−7, π = 0.04 : N ' 100,000.
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 31 / 34
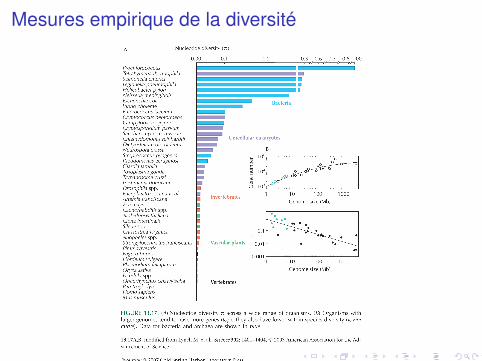
Mesures empirique de la diversité

Génétique des populations. La dérive génétique Coalescent et diversité
Taille efficace 6= taille réellevariations démographiques (goulets d’étranglement)structure de populationgrande variance reproductive (males dominants)
Intérêt du concept de taille efficacetaille efficace donne une mesure de l’importance relative deseffets stochastiques (de la dérive génétique) par rapport à lasélection.régime évolutif dépend avant tout de la taille efficace, et non de lataille réelle.
Différents modes adaptatifs pour différents types d’organismesbactéries ont une sélection plus efficace que les eukaryoteslevures ont une sélection plus efficace que les animauxinsectes ont une sélection plus efficace que les vertébrés
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 33 / 34

Génétique des populations. La dérive génétique Coalescent et diversité
Bilan
adaptation peut être très rapidesélection très efficace en grande populationdérive génétique (fluctuations) limite l’efficacité de la sélection, enparticulier en petite population.dérive génétique a des conséquences très importantes : limite lacapacité d’adaptation des génomes.
Plusieurs domaines en sciences de l’évolutiondynamique adaptativegénétique quantitativegénétique des populationsévolution moléculaire
Nicolas Lartillot (Universite de Montréal) Évolution moléculaire 16 janvier 2012 34 / 34


















