
Déroulement et contrôle de la fermentation. · Par contre, c’est dès le début de la phase de...
29
Déroulement et contrôle de la fermentation. J.M. Sablayrolles, J.M. Salmon, INRA, UMR Sciences pour L'œnologie, 2 place Viala, 34060 Montpellier cedex 1, France. E-mail: [email protected] INTRODUCTION La fermentation alcoolique est une étape clé dans l'élaboration du vin rosé. Au cours de ce processus, les sucres sont, pour l’essentiel, convertis en éthanol et dioxyde de carbone. Mais de très nombreux autres produits sont aussi synthétisés parmi lesquels certains jouent un rôle important sur les propriétés sensorielles du vin final. La souche de levure joue un rôle majeur mais la composition du moût et le mode de conduite de la fermentation sont aussi très importants. En effet, le déroulement de la fermentation est le résultat des interactions entre la souche de levure, la disponibilité des éléments nutritifs dans le moût et le mode de contrôle de la fermentation. Optimiser le contrôle des fermentations alcooliques consiste à les sécuriser, en assurant l’épuisement en sucres, mais aussi à optimiser la qualité du produit, ce qui constitue un objectif très complexe. De plus, ce contrôle doit être mené dans le cadre de fortes contraintes législatives qui limitent les interventions possibles. Enfin, les vinifications en rosé présentent certaines difficultés spécifiques. En effet, elles s’apparentent avant tout aux vinifications en blanc, avec les contraintes et les caractéristiques des fermentations en phase liquide, mais elles présentent également certains points en commun avec les vinifications en rouge, notamment en ce qui concerne la maîtrise de la couleur. Malgré tout, les connaissances actuelles permettent de disposer de moyens d’action de plus en plus efficaces. C’est ce que nous nous proposons de récapituler dans ce chapitre en évoquant aussi – mais de façon plus succincte - la fermentation malo-lactique qui constitue, dans certains cas, une autre étape importante réalisée par des microorganismes. 1 DEROULEMENT DE LA FERMENTATION ALCOOLIQUE Ce chapitre décrit le déroulement d’une fermentation alcoolique dans des conditions de vinification en rosé « standards », c’est à dire en phase liquide, avec ensemencement par une souche commerciale et régulation de la température à une valeur constante. Dans ces conditions, la fermentation alcoolique peut être divisée en trois phases. 1.1 Phase de latence La phase de latence, qui a lieu en tout début de fermentation, correspond à la saturation progressive du milieu en gaz carbonique (environ 1,5 g/L de CO 2 ). A la fin de cette phase, lorsque le gaz carbonique (CO 2 ) commence à se dégager, la population levurienne atteint environ 10 7 cellules/ml, ce qui correspond à 2-3 générations dans les conditions habituelles d’ensemencement. Pendant cette phase, la composition du milieu est très peu modifiée, à l’exception notable de la consommation de la thiamine. En effet, Bataillon et al. (1996) ont montré que, dans un moût inoculé à 10 6 cellules/ml, la thiamine est épuisée en quelques heures. La durée de la phase de latence peut varier de quelques heures à plusieurs jours. Elle est surtout fonction de la température, du taux d’inoculation et de la présence d’inhibiteurs, principalement le dioxyde de soufre (SO 2 ). Ce dernier peut causer des phases de latence très longues, comme le montre le Tableau 1. Ce tableau indique aussi que l’effet du SO 2 est très différent suivant les souches : en présence d’une faible concentration en SO 2 , la phase de latence est plus courte avec la souche B, mais c’est le phénomène inverse qui est observé
Transcript of Déroulement et contrôle de la fermentation. · Par contre, c’est dès le début de la phase de...

Déroulement et contrôle de la fermentation. J.M. Sablayrolles, J.M. Salmon, INRA, UMR Sciences pour L'œnologie, 2 place Viala, 34060
Montpellier cedex 1, France. E-mail: [email protected]
INTRODUCTION La fermentation alcoolique est une étape clé dans l'élaboration du vin rosé. Au cours de ce processus, les sucres sont, pour l’essentiel, convertis en éthanol et dioxyde de carbone. Mais de très nombreux autres produits sont aussi synthétisés parmi lesquels certains jouent un rôle important sur les propriétés sensorielles du vin final. La souche de levure joue un rôle majeur mais la composition du moût et le mode de conduite de la fermentation sont aussi très importants. En effet, le déroulement de la fermentation est le résultat des interactions entre la souche de levure, la disponibilité des éléments nutritifs dans le moût et le mode de contrôle de la fermentation. Optimiser le contrôle des fermentations alcooliques consiste à les sécuriser, en assurant l’épuisement en sucres, mais aussi à optimiser la qualité du produit, ce qui constitue un objectif très complexe. De plus, ce contrôle doit être mené dans le cadre de fortes contraintes législatives qui limitent les interventions possibles. Enfin, les vinifications en rosé présentent certaines difficultés spécifiques. En effet, elles s’apparentent avant tout aux vinifications en blanc, avec les contraintes et les caractéristiques des fermentations en phase liquide, mais elles présentent également certains points en commun avec les vinifications en rouge, notamment en ce qui concerne la maîtrise de la couleur. Malgré tout, les connaissances actuelles permettent de disposer de moyens d’action de plus en plus efficaces. C’est ce que nous nous proposons de récapituler dans ce chapitre en évoquant aussi – mais de façon plus succincte - la fermentation malo-lactique qui constitue, dans certains cas, une autre étape importante réalisée par des microorganismes. 1 DEROULEMENT DE LA FERMENTATION ALCOOLIQUE Ce chapitre décrit le déroulement d’une fermentation alcoolique dans des conditions de vinification en rosé « standards », c’est à dire en phase liquide, avec ensemencement par une souche commerciale et régulation de la température à une valeur constante. Dans ces conditions, la fermentation alcoolique peut être divisée en trois phases. 1.1 Phase de latence La phase de latence, qui a lieu en tout début de fermentation, correspond à la saturation progressive du milieu en gaz carbonique (environ 1,5 g/L de CO2). A la fin de cette phase, lorsque le gaz carbonique (CO2) commence à se dégager, la population levurienne atteint environ 107 cellules/ml, ce qui correspond à 2-3 générations dans les conditions habituelles d’ensemencement. Pendant cette phase, la composition du milieu est très peu modifiée, à l’exception notable de la consommation de la thiamine. En effet, Bataillon et al. (1996) ont montré que, dans un moût inoculé à 106 cellules/ml, la thiamine est épuisée en quelques heures. La durée de la phase de latence peut varier de quelques heures à plusieurs jours. Elle est surtout fonction de la température, du taux d’inoculation et de la présence d’inhibiteurs, principalement le dioxyde de soufre (SO2). Ce dernier peut causer des phases de latence très longues, comme le montre le Tableau 1. Ce tableau indique aussi que l’effet du SO2 est très différent suivant les souches : en présence d’une faible concentration en SO2, la phase de latence est plus courte avec la souche B, mais c’est le phénomène inverse qui est observé

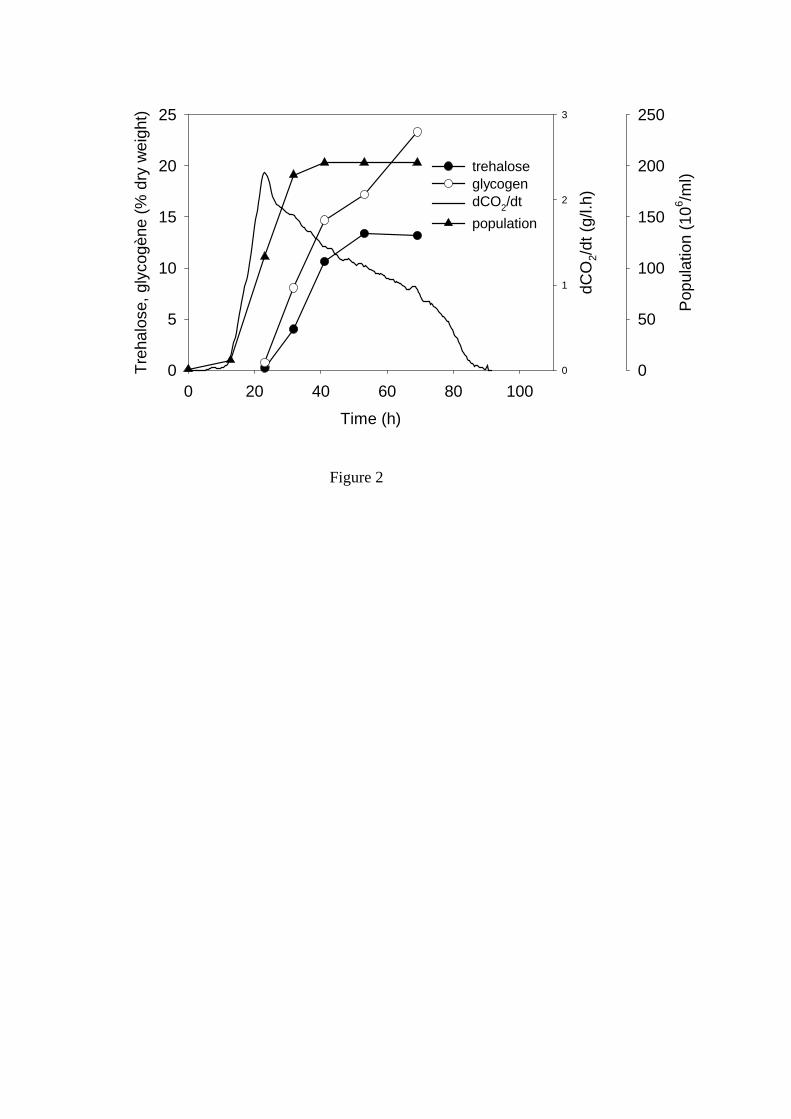
lorsque la concentration est plus élevée. La concentration en acétaldéhyde est, elle aussi, importante et le Tableau 1indique que la concentration en cet intermédiaire du métabolisme levurien, qui possède la caractéristique de combiner le SO2, est identique, quelle que soit la souche, lorsque débute la phase de croissance. 1.2 Phase de croissance La phase de croissance dure jusqu’à l’obtention de la population maximale, soit généralement jusqu’à 20 à 30 % de la fermentation. C’est à la fin de cette phase qu’est atteinte la vitesse maximale de fermentation (Figure 1). C’est donc à cet instant que le dégagement de CO2, ainsi que les besoins en frigories pour réguler la température de fermentation, sont maximums. Par contre, c’est dès le début de la phase de croissance qu’est atteint le maximum de la vitesse spécifique de fermentation qui rend compte de l’activité fermentaire moyenne de chaque levure, activité qui ne cesse par la suite de diminuer. La population atteinte est très variable suivant les moûts. D’après Bely et al. (1990a), elle est comprise entre 50 et 250×106 cellules/ml. Dans la plupart des cas, la multiplication cellulaire prend fin suite à l’épuisement du moût en azote assimilable. De nombreux auteurs, notamment Henschke et Jiranek (1993), ont étudié l’assimilation de l’azote et montré la complexité du transport des différentes sources d’azote assimilable (principalement acides aminés et azote ammoniacal) à l’intérieur des levures. On peut cependant retenir que l’azote ammoniacal est assimilé rapidement et en même temps que les acides aminés. A noter que cette consommation d’azote conduit à une baisse du pH qui atteint sa valeur minimum à la fin de la phase de croissance. 1.3 Phase stationnaire C’est pendant la phase stationnaire que l’essentiel du sucre est fermenté, ce qui constitue une particularité des fermentations œnologiques par rapport aux fermentations brassicoles, par exemple. Pendant cette phase, les levures ne se multiplient plus et leur activité diminue régulièrement même si leur viabilité reste généralement supérieure à 80-90 %, excepté dans les cas d’arrêts de fermentation. Cette baisse d’activité correspond à une diminution régulière de la vitesse de transport des sucres (Salmon et al., 1993). Plusieurs mécanismes d’inhibition sont impliqués, mettant en jeu les acides gras à moyenne chaîne, les toxines killer, etc., mais c’est l’éthanol qui joue le rôle essentiel. Ansanay et al. (2001) ont étudié l’effet d’additions d’éthanol pendant la phase stationnaire et ont montré que, pour les souches commerciales, il y avait généralement une corrélation directe entre la concentration en éthanol dans le milieu et la vitesse de fermentation. Tout au long de la phase stationnaire, la taille des levures augmente et donc leur poids sec, principalement à cause de la synthèse de tréhalose et glycogène, comme le montre la Figure 2. Le glycogène peut être considéré comme un sucre de réserve (Parou et al., 1999) alors que le tréhalose joue un rôle plus complexe. Il est impliqué dans les mécanismes de réponse au stress, de résistance à la pression osmotique (qui atteint plus de 2000 mOsm/kg selon Roustan et Sablayrolles, 2004) mais aussi de régulation fine de la glycolyse (Novo et al., 2005). Les fermentations « languissantes » et les arrêts de fermentation sont caractérisés par de très faibles vitesses finales causées par une forte mortalité des levures. Insa et al. (1995) considèrent qu’il est important de les distinguer des fermentations « lentes » dont la vitesse est faible tout au long de la phase stationnaire mais pendant lesquelles la viabilité reste bonne (Figure 3). Ces points sont discutés plus en détail dans les paragraphes relatifs aux rôles respectifs de l’azote assimilable et de l’oxygène. 2 FERMENTATION MALOLACTIQUE

La fermentation malolactique (FML), qu’elle soit spontanée ou induite par inoculation de bactéries lactiques, conduit à une conversion de l’acide L(-) malique du vin en acide (L-) ou D(-) lactique, suivant les espèces de bactéries lactiques mises en jeu et les substrats dégradés. Pour chaque gramme d’acide malique métabolisé, 0,67 gramme d’acide lactique et 0,33 gramme (165 ml) de CO2 sont ainsi produits. Les bactéries lactiques peuvent également dégrader en parallèle 0,3 à 2 g/L de sucres résiduels pour former environ 100 mg/L d’acide D(-) lactique (Krieger et al., 2000). Cette conversion est accompagnée par toute une évolution des caractéristiques physico-chimiques et sensorielles du vin, qui peuvent donner à l’œnologue un outil supplémentaire pour modifier le style final du vin produit. 2.1 Déroulement de la fermentation malolactique Les bactéries lactiques doivent atteindre un certain seuil de population avant de commencer la FML proprement dite. Ce niveau a été estimé autour de 105 cellules/ml. En pratique, le stress induit par l’environnement du vin sur les bactéries lactiques réduit cette limite, et les inoculations sont généralement réalisées à des teneurs plus élevées. L’inhibition des bactéries lactiques en milieu vinique n’implique pas forcément une mort cellulaire, mais bien souvent une dormance, qui peut être levée quand les conditions environnementales sont plus favorables. Ceci explique notamment l’augmentation soudaine de la fermentation malolactique en cave lorsque la température moyenne ré-augmente au printemps. Une FML spontanée (à partir des bactéries lactiques indigènes) peut donc demander une phase de latence très longue (de quelques semaines à plusieurs mois) avant de démarrer, phase rallongée lorsque les valeurs de pH du vin sont basses. Aux valeurs basses de pH, c’est Oenococcus oeni qui sera l’espèce favorisée (Bousbouras et Kunkee, 1971), alors qu’aux hautes valeurs de pH, ce seront les genres Lactobacillus et Pediococcus qui domineront (Costello et al., 1983). Pendant la FML, la population bactérienne atteint souvent 106 à 107 cellules/ml, alors qu’elle ne représente que 103 à 104 cellules/ml en fin de fermentation alcoolique. Après cette croissance exponentielle, la population bactérienne décline rapidement (de fortes températures ou de fortes concentrations en SO2 accroissent ce phénomène). La phase d’adaptation des bactéries lactiques associée aux fermentations malolactiques spontanées présente donc un risque élevé de développement de micro-organismes indésirables, en particulier dans des vins à faible teneur en SO2 actif, ou à forte acidité volatile. C’est dans ce contexte que s’est développé l’utilisation de levains lactiques, déjà adaptés au milieu vinique (Krieger et al., 2000 ; Palacios et al, 2004), aussi bien en début, au cours ou en fin de fermentation alcoolique. De plus, certaines fermentations malolactiques spontanées menées sur des vins à haute valeur de pH et à faible teneur en SO2, peuvent parfois produire des goûts et odeurs déplaisantes, caractérisées par des nez sulfatés, des odeurs phénoliques ou de moisissure, et des arrière-goûts amers et huileux. Ces défauts ne sont jamais observés après inoculation de bactéries sélectionnées. 2.2 Effets sur les caractéristiques des vins La FML, par son rôle désacidifiant, conduit généralement à une réduction d’acidité titrable variant de 1,0 à 4,6 g/L (en équivalent d’acide tartrique), et à une augmentation du pH du vin pouvant varier de 0,1 à 0,45 unités (en général 0,1 à 0,25 unités pH). Ces valeurs élevées de pH peuvent être une cause d’instabilité microbienne mais, à l’inverse, la FML peut également être considérée favorable vis à vis de la stabilité microbiologique du vin. En effet, de nombreuses espèces bactériennes de contamination sont susceptible de consommer l’acide malique. De plus, les bactéries lactiques utilisent des nutriments pendant leur croissance (acides aminés, peptides, vitamines) et réduisent donc leur bio-disponibilité. Les bactéries malolactiques produisent également des sous produits susceptibles d’influer directement sur la perception sensorielle, ou d’interagir avec les substances amères et/ou

astringentes déjà présentes dans le vin. Ces effets sont extrêmement dépendant des souches de bactéries lactiques utilisées et peuvent, selon les cas, jouer un rôle négatif de masquage des caractères fruités ou variétaux ou, au contraire, en révéler de nouveaux (Krieger et al., 2000; Palacios et al, 2004). Le diacétyle est un sous-produit de la métabolisation de l’acide citrique produit par de nombreuses bactéries lactiques, qui possède un attribut sensoriel considéré généralement comme négatif. Toutefois, ce composé est généralement ultérieurement réduit par les levures (même lors de l’élevage sur lies) ou les bactéries lactiques en acétoine et en 2,3-butanediol, molécules qui n’ont quasiment pas d’impact sensoriel sur le vin aux concentrations normales. 2.3 Interactions levures-bactéries – conséquences technologiques Des interactions entre levure de vinification (Saccharomyces cerevisiae) et Oenococcus oeni peuvent poser des problèmes de réalisation de FML. En effet, Saccharomyces cerevisiae produit de l’éthanol, du dioxyde de soufre (SO2) et des acides gras à courte et moyenne chaîne qui peuvent inhiber O. oeni. De plus, l’utilisation des acides aminés par la levure comme source d’azote pendant les premières étapes de la fermentation alcoolique peut compliquer la croissance ultérieure des bactéries lactiques (Costello et al, 1983). L’inhibition des bactéries lactiques décroît fortement en fin de fermentation alcoolique, vraisemblablement du fait de la mortalité levurienne. Toutefois, il peut être intéressant d’inoculer les bactéries lactiques avant la fin de la fermentation alcoolique de façon à profiter d’un environnement plus favorable, car moins riche en éthanol, et contenant plus de nutriments. Un achèvement de la FML avant la fin de la fermentation alcoolique permet effectivement une addition beaucoup plus précoce de SO2 pour stabiliser le vin. 3 IMPORTANCE DE L’AZOTE ASSIMILABLE 3.1 Rôle et variabilité de l’azote assimilable. L’azote assimilable est généralement le nutriment le plus limitant dans les moûts pour les levures et joue, donc, un rôle essentiel sur la cinétique fermentaire. Bely et al. (1990b) ont montré qu’il existait une relation directe entre les concentrations en azote assimilable et les vitesses maximales de production de CO2 (proportionnelles aux vitesses de fermentation). Les carences en azote assimilable peuvent donc être considérées comme les causes essentielles des fermentations « lentes ». La concentration en azote assimilable est très variable suivant les moûts. Cette variabilité a été décrite par Bely et al. (1990b) qui ont étudié une centaine de moûts issus des principales régions viticoles françaises. Sur cet échantillonnage, les teneurs en azote assimilable variaient de 53 à 444 mg/L. En conditions standardisées, cela conduisait à des vitesses maximales de production de CO2 (proportionnelles aux vitesses de fermentation) comprises entre 0,4 et 2,8 g/L.h et à des populations levuriennes de 50 à 220×106 cellules/ml. Si d’un point de vue microbiologique, on peut considérer que l’azote était quasiment toujours limitant, l’effet de la carence était surtout sensible lorsque sa concentration initiale était inférieure à 150-180 mg/L. C’est la raison pour laquelle ces auteurs proposent un seuil de « carence technologique » dans cette zone. Cette variabilité dans le contenu azoté des moûts est due à de nombreux facteurs, parmi lesquels le cépage et la situation géographique. C’est ainsi que Nicolini et al. (2004) ont mesuré une teneur moyenne en azote de 136 mg/L dans 600 moûts italiens alors que Butzkee (1998) a obtenu une concentration moyenne de 213 mg/L, avec un échantillonnage de 1500 moûts californiens. Le niveau de maturité affecte aussi de façon importante le contenu azoté des moûts avec une baisse de l’azote ammoniacal au cours du temps, accompagnée d’une

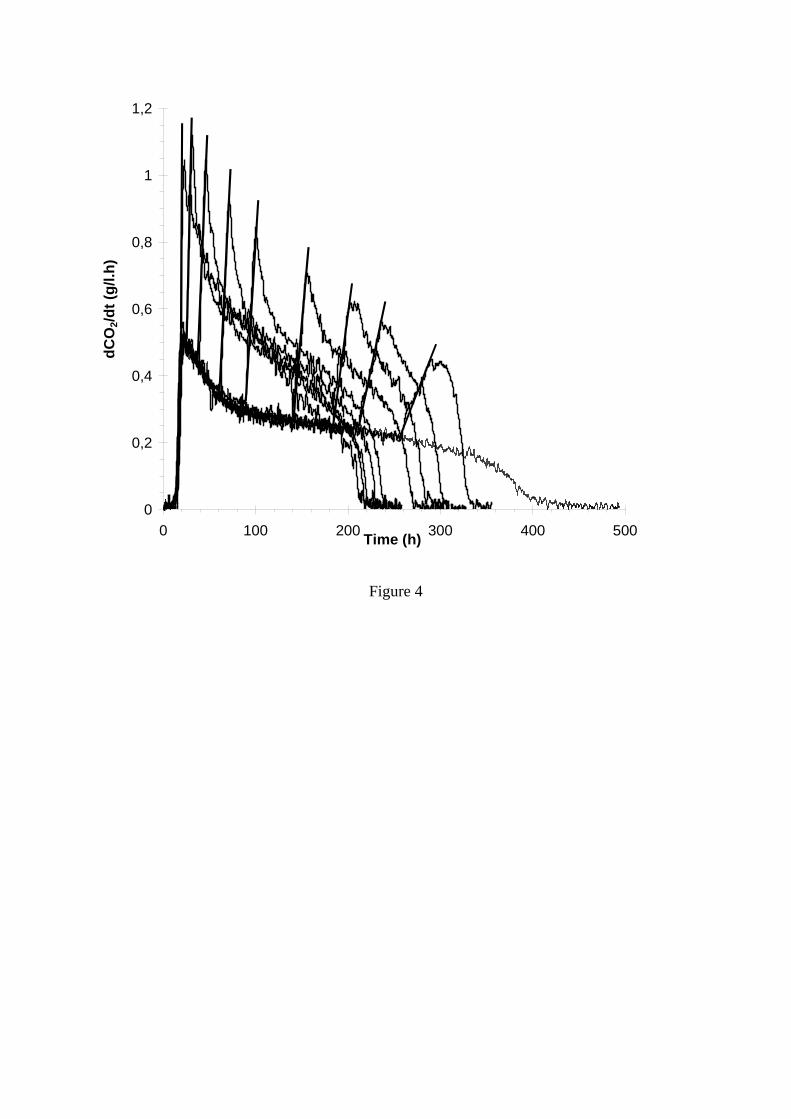
évolution plus complexe des acides aminés (Dubois et al., 1996). Il est à noter que ces observations ont été faites pour différents types de moûts, parmi lesquels certains ont été prélevés lors de vinifications en rosé. 3.2 Effet des ajouts d’azote sur la cinétique fermentaire Dans le cas des milieux fortement carencés (moins de 150 mg/L d’azote assimilable), l’addition de sels d’ammonium (phosphate diammonique (DAP) ou sulfate diammonique) accélère de façon spectaculaire la vitesse de fermentation (Bely et al., 1990b) et, par voie de conséquence raccourcit fortement les fermentations. Mais le moment d’ajout est aussi très important. Si l’azote est ajouté au moment de l’inoculation, il conduit à une augmentation de la population levurienne, alors que s’il est ajouté pendant la phase stationnaire (vers la mi-fermentation), il augmente l’activité des levures, grâce, notamment, à une réactivation des systèmes de transport des sucres (Bely et al., 1994). Dans le cas des fermentations « lentes », caractérisées avant tout par une carence en azote, les apports d’azote ont une efficacité comparable pendant toute la première partie de la fermentation (Figure 4). Dans le cas des fermentations « languissantes », qui sont caractérisées par d’autres types de carences, principalement en oxygène et composés lipidiques, une addition de DAP au moment de l’inoculation peut au contraire avoir un effet néfaste. En effet, cette addition augmente la population et la vitesse maximale de fermentation, mais conduit à une plus forte mortalité finale à cause d’une dilution excessive des réserves lipidiques des levures (Sablayrolles, 1996). 3.3 Effets additionnels des ajouts d’azote Si les additions d’azote ont un impact très fort sur les vitesses de fermentation, elles ont aussi des conséquences sur la synthèse de sous produits du métabolisme et aussi, parfois, sur les caractéristiques organoleptiques du produit final. Bell et Henschke (2005) insistent, à ce sujet, sur la nécessité de maîtrise des ajouts de sorte à rester dans une fourchette acceptable d’azote assimilable (initial plus ajouté) disponible pour les levures. Des quantités trop faibles conduisent souvent à des productions excessives de composés soufrés indésirables, en particulier de sulfure d’hydrogène (H2S) et d’alcools supérieurs alors qu’elles limitent les synthèses d’esters. A l’inverse, les quantités excessives d’azote peuvent augmenter les concentrations en acétate d’éthyle et en acide acétique. Elles augmentent aussi les risques de troubles protéiques, de synthèse de molécules indésirables telles que le carbamate d’éthyle et, surtout d’instabilité microbiologique, avec des conséquences possibles en terme de vieillissement atypique par exemple. Ces risques liés aux quantités excessives d’azote doivent être d’autant plus pris en considération que l’ajout maximum autorisé a été augmenté, ces dernières années, de 300 mg/L à 1 g/L de DAP (soit de 63mg/L à 210 mg/L d’azote). La gestion des apports azotés en cours de fermentation alcoolique peut aussi avoir des conséquences sur le déroulement de la fermentation malolactique. Cependant, il ne faut pas en exagérer l’importance, car les acides aminés sont utilisés par les bactéries lactiques plus comme source d’énergie que comme substrat de croissance proprement dit. De plus, toutes les bactéries lactiques sont capables de produire des protéases extracellulaires, ce qui explique l’augmentation relative des concentrations en acides aminés au cours de la fermentation malolactique. Enfin, la croissance de certaines bactéries lactiques est plus stimulée par certains peptides que par les acides aminés proprement dit (Alexandre et al, 2004). 4 IMPORTANCE DE L’OXYGENE ET DES LIPIDES 4.1 Rôle de l’oxygène et des composés lipidiques

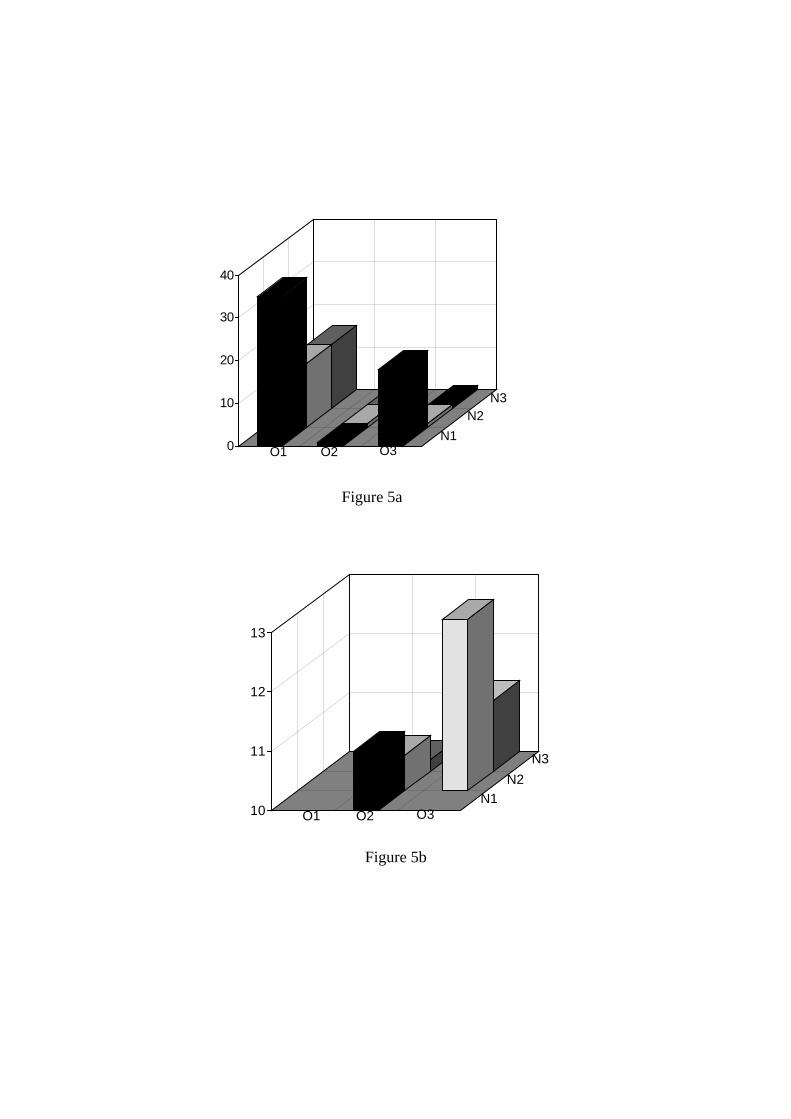
L’oxygène est nécessaire pour la croissance des levures et, surtout, pour le maintien d’une bonne viabilité en fin de fermentation. Il est connu depuis longtemps que ce rôle positif est lié avant tout à la synthèse de composés lipidiques, qui sont des constituants essentiels de la membrane cytoplasmique. Plus récemment, il a été montré (Salmon, 2006) que cette consommation est beaucoup plus dédiée à la synthèse de stérols (jusqu’à 75% de l’oxygène consommé) qu’à celle des acides gras insaturés. Les levures utilisées lors de l’inoculation sous forme de levures sèches, contiennent de grandes quantités de composés lipidiques, soit incorporés dans les membranes, soit stockés sous forme de réserves (triglycérides, stérols, etc.). L’aptitude des levures à diluer ces réserves leur permet plusieurs générations en anaérobiose mais, dans ces conditions, les levures atteignent en fin de croissance des teneurs très faibles en ces composés, ce qui les rend plus fragiles, en particulier vis à vis de l’effet toxique de l’éthanol. Le taux d’inoculation a donc un impact direct en tendant à limiter la dilution de ces réserves. Les composés lipidiques présents dans le moût, principalement les phytostérols, jouent aussi un rôle important et limitent les besoins en oxygène. Ces composés sont présents sous forme assimilable dans les cires cuticulaires (Higgins et Peng, 1976) et dans la pellicule. Ils sont très apolaires et restent adsorbés sur les structures telles que les flocs pectiques. Ils sont donc, au moins partiellement, éliminés lors des différentes étapes préfermentaires de clarification. Certaines solutions commerciales d’activateurs de fermentation contiennent des levures inactivées susceptibles de relarguer des composés lipidiques. Depuis ces dernières années, il existe des préparations de levures inactivées à rajouter aux levures en cours de réhydratation. Il a été montré (Soubeyrand et al., 2005) que, dans ces conditions, il existe effectivement un transfert de stérols vers les levures sèches (LSA), qui leur permet d’être plus performantes, ultérieurement en cours de fermentation. 4.2 Importance et contrôle de l’oxygénation En pratique, les carences en oxygène sont parmi les causes essentielles des arrêts de fermentation et l’ajout d’au moins 10 mg/L d’oxygène est généralement nécessaire. Le moment le plus favorable pour l’oxygénation est la fin de la phase de croissance (vers le quart de la fermentation) avec une efficacité encore très grande à mi-fermentation. A l’inverse, les ajouts initiaux sont à la fois peu efficaces pour les levures et susceptibles de conduire à une oxydation du moût. Le contrôle de l’oxygénation est délicat dans la mesure où celui-ci doit avoir lieu en pleine fermentation, c’est à dire lorsque les levures le consomment à une vitesse supérieure à la vitesse d’apport. Pour quantifier l’oxygène transféré, il est dès lors préférable de réaliser un étalonnage avant fermentation (Blateyron et al., 1998). En l’absence de calibration, ces auteurs considèrent qu’avec un diffuseur performant, produisant de fines bulles, au moins la moitié de l’oxygène injecté dans le milieu est effectivement transféré sous forme d’oxygène dissous, qui est la seule forme utilisable par les levures. 4.3 Efficacité de l’oxygénation et des ajouts combinés oxygène-azote assimilable Buescher et al (2001) ont montré qu’une forte oxygénation pouvait permettre d’obtenir un vin à 20,95 % d’éthanol à partir d’un mout de raisin concentré. Il s’agit là de conditions extrêmes qui illustrent l’efficacité potentielle d’ajouts d’oxygène. En conditions œnologiques, cette efficacité a été étudiée par Blateyron et al. (2000), en couplant l’oxygénation à des ajouts de DAP (300 mg/L). Dans un premier temps, ces auteurs ont confirmé qu’il était essentiel d’effectuer ces ajouts combinés, non pas en début de fermentation mais au début de la phase stationnaire, un peu avant la mi-fermentation (Figure 5). Ils ont ensuite testé l’efficacité de cet ajout combiné sur 72 moûts donnant lieu à des arrêts de fermentation ou des fermentations languissantes. Dans tous les cas, ces ajouts se sont révélés efficaces conduisant

systématiquement à un épuisement des sucres et à une baisse moyenne de 44 % de la durée de fermentation. Cet effet s’est révélé indépendant du cépage, de l’origine du moût et de la souche de levure. Il est à noter que de telles additions ont aussi été testés sur des moûts ne conduisant pas à des problèmes fermentaires et, dans ce cas, la durée de fermentation a aussi été fortement abaissée (35% en moyenne). Cela indique que cette addition conjointe d’azote et d’oxygène est quasiment toujours efficace, même lorsqu’elle n’est pas indispensable. 4.4 Effets de l’oxygénation et des lipides sur les caractéristique du vin L’addition d’oxygène peut aussi avoir un impact sur le métabolisme levurien et les concentrations en alcools supérieurs et esters, notamment. Les conséquences de l’oxygénation sur la synthèse et la conservation des arômes variétaux sont encore mal comprises ; les données actuellement disponibles concernant avant tout l’effet de l’oxygène pendant l’élevage et le stockage. D’un point de vue sensoriel, Blateyron et al. (2000) n’ont détecté aucun impact pour des ajouts de 5 mg/L d’oxygène en début de phase stationnaire, alors que cet impact était très négatif, aussi bien au niveau de la couleur que des arômes, lorsque la quantité ajoutée était très excessive (50 mg/L). Les lipides, quand à eux, ont un impact avéré sur l’acidité volatil dont ils limitent la production de façon très sensible (Delfini et Costa, 1993). L’effet de l’oxygène sur le déroulement de la fermentation malolactique, reste, lui aussi, mal défini car, si les bactéries lactiques sont généralement micro-aérophiles, l’impact favorable des conditions de micro-aérobiose sur le développement des bactéries lactiques en conditions œnologiques n’est pas systématique (Kelly et al, 1989). 5 IMPORTANCE DES AUTRES NUTRIMENTS 5.1 Sels minéraux Même si l’azote assimilable et l’oxygène peuvent être considérés comme les nutriments principaux en conditions œnologiques, ils ne sont pas les seuls susceptibles d’influer sur le déroulement des fermentations. La concentration en magnésium peut, dans certaines conditions, aussi avoir un effet sur la croissance et la dégradation des sucres (Birch et al., 2003). En effet, les ions Mg2+, avec les phospholipides, jouent un rôle important pour stabiliser la membrane cytoplasmique et ainsi participer à la protection des cellules vis à vis de différents facteurs de stress, dont principalement l’éthanol. A l’inverse, la perte de magnésium couplée à une entrée excessive de calcium accroît la sensibilité des levures à l’éthanol. 5.2 Vitamines Les concentrations en vitamines ne sont généralement pas limitantes dans les moûts. C’est ainsi que Sablayrolles et Salmon (2001) n’ont constaté qu’un effet très faible de l’addition d’un mélange de biotine, thiamine, méso-inositol, acide pantothénique, acide nicotinique et pyridoxine dans 17 moûts différents. Cependant, la thiamine peut, dans certaines situations, être limitante. Ceci s’explique par le fait que cette vitamine est très rapidement consommée par les levures (Bataillon et al., 1996), y compris par la plupart des populations indigènes. Cela peut conduire à une carence induite des moûts au moment de l’inoculation et être la cause de fermentations difficiles. Dans ces conditions, l’addition de thiamine permet de restaurer une cinétique de fermentation normale. Dans la pratique, la thiamine est souvent ajoutée sous forme de solutions commerciales complexes, contenant généralement aussi du DAP et des levures inactivées, ce qui rend difficile la différenciation de l’effet de chaque nutriment. Au delà de cet impact sur le déroulement des fermentations, la thiamine est connue pour sa capacité à se combiner avec différentes molécules, dont le SO2. Dans certains cas, ceci peut se

traduire par une baisse d’efficacité de celui-ci et avoir, donc, des conséquences technologiques. 6 INFLUENCE DE LA SOUCHE DE LEVURE Saccharomyces cerevisiae est la principale espèce de levures utilisée en œnologie. Lors des fermentations spontanées, d’autres espèces peuvent intervenir et jouer un rôle significatif mais de façon complexe et souvent imprévisible. Lors des 30 dernières années, l’utilisation des levures sélectionnées, sous forme de levures sèches actives (LSA), s’est très largement répandue. C’est ainsi qu’en France environ ¾ des fermentations sont actuellement ensemencées avec des LSA. D’autre part, le nombre de souches commercialisées a continuellement augmenté pour atteindre un nombre d’environ 200. Cette augmentation spectaculaire a fait suite à une évolution dans les critères de choix des souches. Initialement, c’est la notion de fiabilité des fermentations qui était essentielle alors qu’actuellement les arguments mis en évidence sont surtout d’ordre qualitatif (qualité et spécificités aromatiques, etc…). 6.1 Influence des souches de levures sur la cinétique fermentaire Colombie et al. (2005) ont caractérisé les cinétiques de fermentation obtenues avec 20 souches de levures choisies au hasard parmi celles disponibles sur le marché français. Les résultats obtenus, sur un milieu synthétique simulant un moût de raisin facilement fermentescible, mettent en évidence des différences relativement faibles avec, respectivement : des vitesses maximales de production de CO2 comprises entre 1,85 et 2,52 g/L.h (moyenne: 2,33, écart type: 0,18), des durées de fermentation de 80 à 109 heures (moyenne: 90, écart type: 6,8) et des populations comprises entre 114 et 188 ×106 cellules/ml. Des différences beaucoup plus importantes sont rapportées par Blateyron et al. (2000), qui ont comparé les performances de 13 souches (choisies au hasard) dans un moût très difficile à fermenter et donc beaucoup plus discriminant. 3 souches ont conduit à des arrêts de fermentation, avec des concentrations résiduelles en sucres parfois considérables (10 à 56 g/L), alors que les 10 autres ont permis l’épuisement des sucres avec des durées de fermentation variant de 119 à 170 heures. Plusieurs travaux ont montré que ces différences étaient principalement causées par des besoins différents en azote assimilable et oxygène. Julien et al. (2000) ont quantifié les besoins en azote en pilotant automatiquement des ajouts de DAP et en comparant les ajouts d’azote (ammoniacal) nécessaires pour réguler la vitesse de fermentation à un niveau prédéfini. Dans ces conditions, ces ajouts se sont échelonnés entre 48 et 110 mg/L d’azote, soit entre 230 et 525 mg/L de DAP. De leur coté, Beltran et al. (2007) ont montré que ces besoins en azote variaient très sensiblement avec la température. Pour estimer et comparer les besoins en oxygène, Julien et al. (2000) ont mesuré les cinétiques fermentaires obtenues avec un milieu très difficile à fermenter (milieu synthétique sans facteurs anaérobies (lipides) et désaéré). Il ont observé des différences très importantes et montré que les souches qui se multipliaient le plus, et conduisaient aux valeurs les plus élevées de vitesses maximales de fermentation étaient, à l’inverse, celles qui avaient les plus mauvaises viabilités finales, certainement à cause d’une dilution excessive de leurs réserves lipidiques. 6.2 Influence des souches de levures sur les caractéristiques du vin De plus en plus, les vinificateurs effectuent le choix de la souche de levure à utiliser en fonction de critères spécifiques, en lien avec le type de vin souhaité.

La concentration en éthanol de vins est un paramètre important, d’autant plus que la richesse moyenne en sucres des moûts a fortement augmenté au cours des 2 dernières décennies. Certains vinificateurs souhaiteraient pouvoir abaisser le degré final, de 2% environ, en diminuant le rendement en éthanol de la fermentation alcoolique. Est-ce possible avec des souches actuellement commercialisées ? Pour répondre à cette question, Ortiz-Julien (2007, communication personnelle) a comparé 55 souches, en conditions parfaitement standardisées, et n’a obtenu que des différences très faibles dans les concentrations finales en éthanol : 11,72 à 12,09 %, soit une moyenne de 11,95 % et un écart type de 0,08 %. Ces résultats indiquent clairement qu’il est illusoire de rechercher des différences en éthanol final supérieures à 0,5 % en changeant de souche commerciale. Ceci est peu surprenant dans la mesure où (i) il faut dévier des quantités très importantes de sucres pour obtenir une baisse significative du rendement (16,8 g/L de sucres pour un abaissement d’1 % d’éthanol et (ii) il faut éviter l’accumulation de sous produits indésirables. S’il n’est donc pas possible d’espérer obtenir des baisses significatives du rendement éthanol avec les souches commerciales actuelles, il est envisageable d’obtenir des souches modifiées (pas nécessairement des OGM) qui auraient effectivement ces caractéristiques. Plusieurs approches sont actuellement testées en ce sens. L’impact de la levure sur les composé aromatiques est de plus en plus pris en compte, mais il s’agit d’une caractéristique difficile à estimer. En effet, la synthèse de ces molécules d’intérêt est toujours influencée, non seulement par la souche, mais aussi par la composition du moût et le mode de conduite de la fermentation. Cela donne lieu à des mécanismes d’interactions, souvent complexes, qui peuvent conduire à des observations contradictoires suivant les conditions. C’est plus particulièrement le cas lorsque l’on souhaite faire intervenir des notions très complexes et mal définies telles que la typicité. Malgré ces difficultés, il existe des données de plus en plus fiables sur les capacités de certaines souches : - A augmenter le caractère fruité des vins. L’aptitude à produire des vins fruités, avec de fortes teneurs en esters, a été un critère de sélection important dans les années 1980. Dans la décennie suivante, plusieurs souches cryotolérantes ont été sélectionnes, pour des fermentations à très basse température (10-12°C) (Castellari et al. 1992, Massoutier et al. 1998). Dans de telles conditions, ces souches produisent des quantités très importantes de 2-phenyl éthanol et de 2-phenyl acétate. Il est à noter que la capacité à produire des arômes fermentaires d’intérêt est particulièrement importante dans le cas des vins rosés. - Pour augmenter ou moduler le caractère variétal de certains cépages. Le Sauvignon est l’exemple le plus étudié avec la description de thiols volatils très fortement impliqués dans son arôme caractéristique (Dubourdieu et al. 2006) mais, même dans ce cas, les mécanismes mis en jeu en cours de fermentation (molécules précurseurs, voies métaboliques impliquées, effet des variables opératoires) ne sont encore que partiellement compris (Swiegers et Pretorius 2007, Subileau et al 2008). L’aptitude des levures à libérer des arômes variétaux deviendra certainement un critère de plus en plus important et de mieux en mieux mesurable. C’est vrai en premier lieu pour les vins blancs mais aussi pour les vins rosés dans lesquels ces mêmes thiols ont été notamment mis en évidence. - Pour limiter la production d’acides organiques ou de composés indésirables tels que les phénols volatils (Shinohara et al. 2000) ou le H2S. C’est ainsi qu’a été récemment décrite une souche qui ne produit quasiment plus de H2S (Swiegers, 2008). Plusieurs auteurs ont aussi mis en évidence l’intérêt de certaines souches spécifiques pour produire des mannoprotéines (Moine-Ledoux & Dubourdieu 2002). 6.3 Influence des levures sur la couleur Il n’existe que peu d’études relatives à l’adsorption des polyphénols sur les levures au cours de la fermentation alcoolique, particulièrement en conditions de vinification en rosé. Il est cependant connu que les levures peuvent modifier la couleur des vins par l’établissement

d’interactions faibles et réversibles entre leur parois et les anthocyanes (Morata et al. 2003) ou par l’intervention d’une activité béta-glucosidasique périplasmique pouvant dégrader la partie 3-O-glucoside des anthocyanes (Delcroix, Günata, Sapis, Salmon & Bayonove, 1994). Une étude récente a montré que très peu de composés phénoliques monomèriques restent en fait adsorbés à la surface des levures, et que ces mêmes levures ne présentent pas d’adsorption préférentielle pour les tannins de bas ou haut poids moléculaire (Mazauric & Salmon, 2005). Les tanins condensés restant dans le vin contiennent moins de résidus épigallocatéchine que les tanins initiaux, indiquant que les tanins condensés polaires sont préférentiellement adsorbés par les levures. Inversement, l’efficacité d’adsorption des anthocyanes par les levures n’est pas relié à leur polarité (Mazauric & Salmon, 2006). La polymérisation des polyphénols au cours de la fermentation peut également faire intervenir l’acétaldéhyde, qui est considéré comme un composant clef de leur polymérisation oxydative. Pendant la fermentation, les levures produisent de l’acétaldéhyde comme sous-produit de la fermentation. La complexation lente des anthocyanes avec cet acétaldéhyde conduit à la formation de composés de couleur jaunes, qui peuvent à leur tour se condenser lentement avec d’autres polyphénols (Ben Abdeljalil et al., 2000). La forte variabilité existant dans la production d’acétaldéhyde entre souches de levures en fonction des conditions de fermentation peut donc être un facteur influant sur la production plus ou moins grande de ces composés colorés. 7 IMPORTANCE DE LA TEMPERATURE 7.1 Effet du régime de température sur la cinétique fermentaire La température a un effet primordial sur la cinétique fermentaire. Entre 15 et 25°C, la vitesse de fermentation est doublée lorsque la température est augmentée d’environ 8°C (Bely et al., 1990a). Le profil de température est, lui aussi, important et une augmentation de plusieurs degrés en cours de fermentation modifie profondément la courbe de fermentation. Le Tableau 2 illustre l’impact du régime de température sur la durée de fermentation ainsi que sur la vitesse maximale de fermentation. Dans tous les cas, la température est comprise entre 15°C et 22°C, soit une gamme d’intérêt pour les fermentations en rosé. Les valeurs indiquées dans ce tableau ont été calculées grâce à un simulateur de la fermentation récemment développé (Sablayrolles et Gerland, 2007) et basé sur le modèle cinétique de Malherbe et al. (2003) et sur le modèle thermique de Colombie et al. (2007). Ces modèles ont été validés dans de nombreuses situations et les erreurs maximales peuvent être estimées à 10% (Colombie et al., 2005). De façon logique, les fermentations menées en non isothermie (anisothermie ??) ont des valeurs intermédiaires entre les 2 isothermes à 15°C et 22°C, avec des fermentations plus rapides lorsque la température augmente dès le début. On peut cependant noter l’effet très significatif de l’augmentation de température pendant la deuxième partie de la fermentation. En effet, comparativement à l’isotherme à 15°C, ce régime de température (qui correspond à une pratique courante) permet de raccourcir la fermentation alcoolique de 7 jours. Par ailleurs, il faut noter qu’une augmentation de température autour de 20°C est quasiment indispensable dans le cas où l’on souhaite réaliser une fermentation malolactique. 7.2 Effet du régime de température sur les besoins énergétiques Le régime de température a aussi un impact très important sur les besoins énergétiques nécessaires pour la régulation de température. Dans le Tableau 2, sont reportés les estimations (i) de la puissance maximale (Pmax) utilisée (au moment où la vitesse de fermentation est maximale) et (ii) de l’énergie totale (Etot) dépensée pendant la fermentation. Ces paramètres présentent un grand intérêt technologique car ils sont proportionnels, respectivement, à la puissance du groupe frigorifique et à la consommation électrique. Par rapport à l’isothermie à

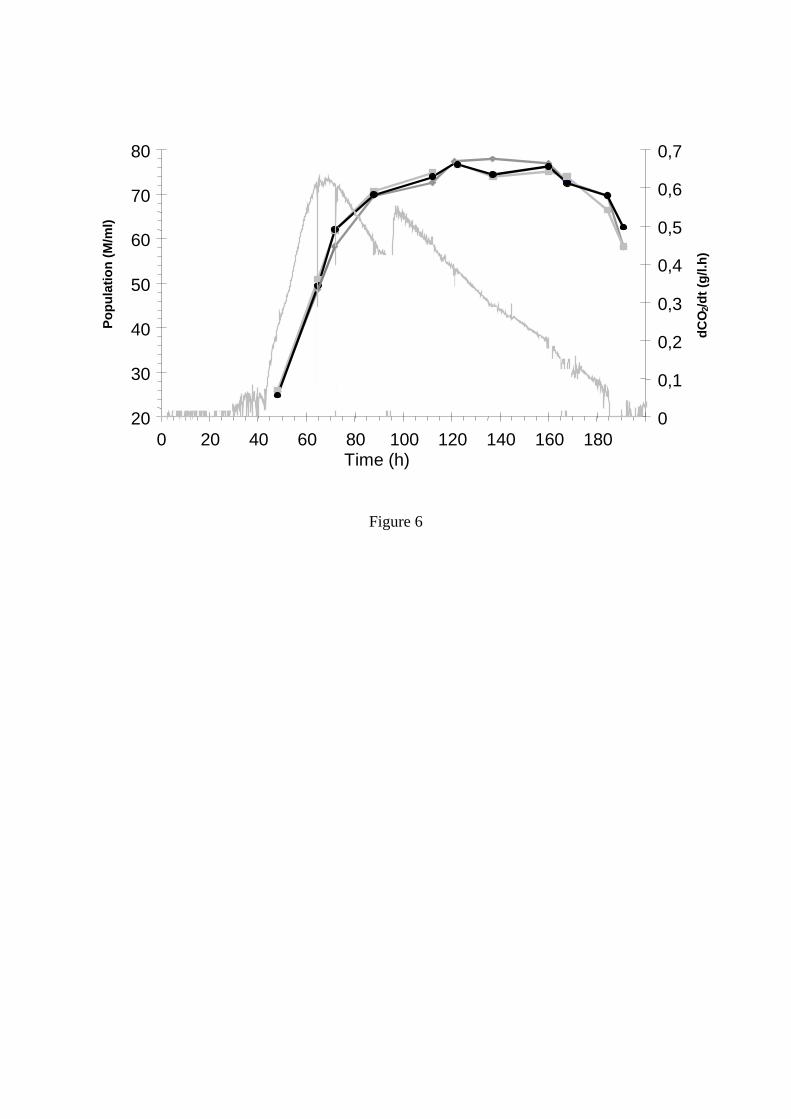
15°C (Etot = 1411 103 kcal), le fait d’augmenter la température jusqu’à 22°C permet des économies d’énergie très importantes de respectivement 43% (Etot = 811 103 kcal) et 34% (Etot = 935 103 kcal). L’augmentation précoce de température permet une économie légèrement supérieure, mais l’augmentation plus tardive (à mi-fermentation) présente l’intérêt d’avoir, en outre, une valeur modérée de Pmax. En effet, dans ce cas, la vitesse maximale de fermentation est la même que celle obtenue lors de la fermentation à 15°C. 7.3 Effet de la température sur les caractéristiques organoleptiques du vin. De façon générale, les très basses températures (10 à 15°C) favorisent la production d’arômes fermentaires tels que les esters (Cotrell and MC Lellan, 1986; Torija et al., 2003). De telles températures peuvent donc être utilisées pour augmenter le caractère fruité des vins. Mais, dans ce cas, les risques de fermentations languissantes, voire d’arrêt de fermentation sont fortement augmentés et une bonne gestion de la fermentation est indispensable. Elle inclue, en particulier, le choix d’une levure suffisamment cryotolérante et une augmentation de température en fin de fermentation. Des travaux récents ont aussi mis en évidence l’importance de la température vis à vis de la libération d’arômes variétaux, plus particulièrement de thiols (Howell et al. 2004, Masneuf et al. 2006). Ils ont montré que, quelle que soit la souche utilisée, leur production est plus importante à température relativement élevée (20°C) qu’à température basse (13°C). Le choix de la température peut donc être considéré comme un moyen de favoriser soit les arômes fermentaires, soit les arômes variétaux. 8 IMPORTANCE DES CONDITIONS HYDRODYNAMIQUES Lors des vinifications en rosé, la fermentation alcoolique est effectuée en phase liquide à basse température sur un moût clarifié. Quelles sont donc les conséquences au niveau hydrodynamique (homogénéité, agitation, turbidité…) et pourquoi est-il important de maîtriser ces pratiques ? Voici quelques éléments de réponse en ce qui concerne la fermentation alcoolique. 8.1 Homogénéité des cuves en fermentation L’homogénéité du moût en fermentation est importante pour favoriser les échanges entre les levures et le milieu. Dans les cuves œnologiques, le mélange est assuré par la production de CO2. Garcia et al. (1994) ont proposé un modèle pour calculer la puissance de cette agitation et estimer un facteur d’hétérogénéité. En pratique, ces auteurs considèrent que, dans la plupart des fermentations en phase liquide, le milieu est homogène. Malherbe (2003) a mesuré les principaux paramètres à différents endroits d’une cuve de 110 hl lors d’une fermentation en phase liquide, à 20°C. Elle a observé que la température ainsi que les concentrations en sucres et éthanol étaient parfaitement identiques aux différents points de mesure, quel que soit le stade de la fermentation. La population levurienne, quand à elle, baissait de 30% en fin de fermentation, suite à une sédimentation. Ce phénomène n’est pas surprenant mais il est intéressant de remarquer que : - la sédimentation des levures ne commence que lorsque la vitesse de production de CO2 devient inférieure à 0,15 g/L.h (soit environ 0,3 g/L.h de consommation de sucres), ce qui représente une vitesse très faible. Ceci a été, par la suite, validé lors de plusieurs fermentations réalisées à l’échelle pilote (Aguera et Sablayrolles, 2005) - de façon surprenante, cette population décroît exactement de la même façon dans toute la cuve (Figure 6). Ceci indique que la phase liquide reste parfaitement homogène jusqu’à la fin de la fermentation, alors que la sédimentation ne s’effectue seulement que sur une fine épaisseur, de quelques centimètres, en bas de cuve. Si l’on mesure des paramètres plus fins

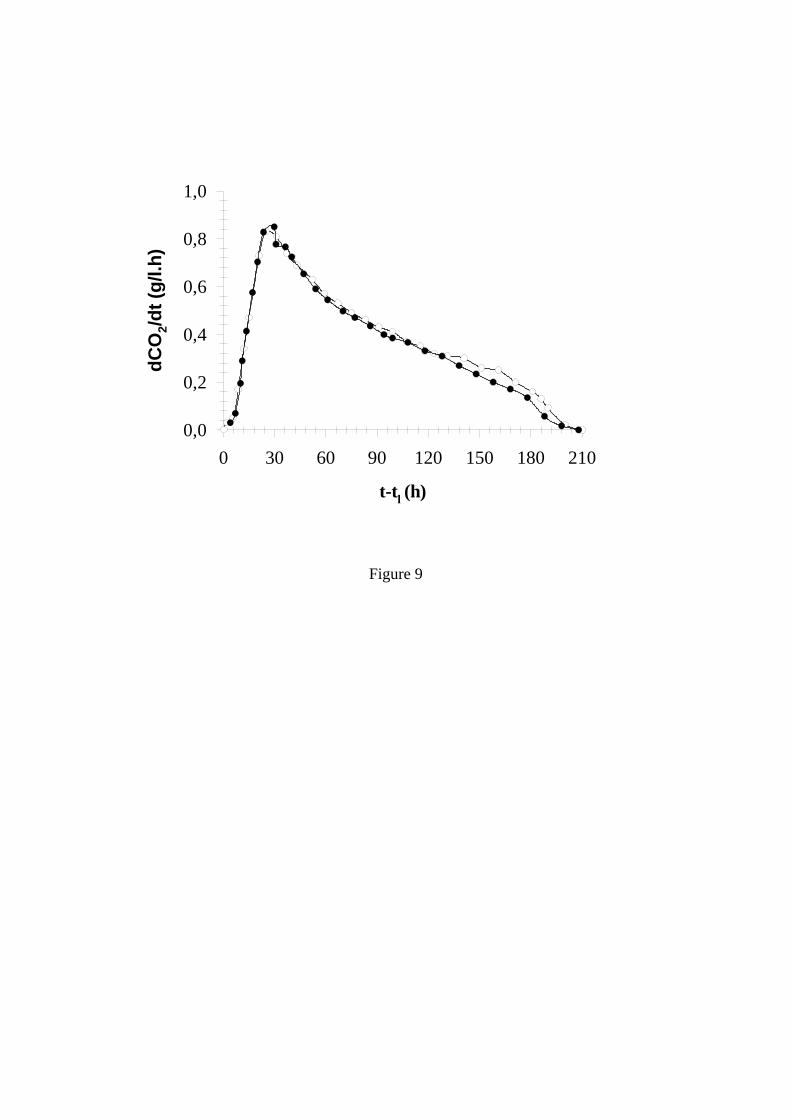
tels que la viabilité cellulaire ou la morphologie des levures, on retrouve bien cette homogénéité au sein de la population en suspension, mais aussi une différenciation vis à vis de celle qui a sédimenté. Cela est illustré sur la Figure 7 qui décrit la distribution par classes de tailles, c’est à dire une information sur la morphologie des levures. Cette figure montre clairement que les levures prélevées à différents endroits de la cuve ont exactement la même morphologie alors qu’elles sont bien différentes de celles qui sont en bas de la cuve (Aguera and Sablayrolles, 2005); ces dernières étant plus petites avec un taux de mortalité plus élevé. Vlassides et Block (2000) ont étudié les profils de concentration cellulaire en tout début de fermentation. Ils ont observé, juste après inoculation, une sédimentation des levures, puis un démarrage de la croissance levurienne à partir du bas de la cuve et enfin une homogénéisation. Ces auteurs considèrent que le gradient initial pourrait être dû, en partie, à l’hydratation incomplète des levures sèches initialement ajoutées. 8.2 Effet de la turbidité La présence de particules solides est essentielle non seulement parce que celles ci contiennent des nutriments, principalement des lipides (cf &3.) mais aussi car elles jouent un rôle, purement physique, de nucléation du CO2, aidant ainsi à la formation de bulles et au dégagement du CO2. La Figure 8 illustre le très fort impact des bourbes sur la cinétique fermentaire : sur cet exemple, le fait de rajouter 0,4 et 3% de bourbes à un moût extrêmement clarifié (moins de 10 NTU) a permis d’augmenter les vitesses maximales de fermentation de, respectivement, 58 et 141 %, avec un effet maximum atteint dans le deuxième cas. Pour ne pas avoir de limitation de la cinétique fermentaire par une trop grande clarification des moûts, on peut donc considérer qu’il faut 2-3% de bourbes fines, ce qui correspond à des niveaux de turbidité de 100-150 NTU. Plouy (2000) a montré l’existence d’une interaction entre agitation et turbidité. En effet, en comparant les cinétiques obtenues, à petite échelle, avec et sans agitation du milieu, il a observé des différences importantes, particulièrement en fin de fermentation, dans le cas d’un moût très clarifié. Par contre, cette différence était beaucoup moins marquée avec un moût à la turbidité élevée (200 NTU). Ce phénomène est certainement, lui aussi, à relier au rôle des particules solides vis à vis du dégagement de CO2 et, par voie de conséquence, de son pouvoir d’agitation et d’homogénéisation. 8.3 Effet de la taille de la cuve Les cuves peuvent être de taille très différentes, de quelques hl à plusieurs centaines d’hl. Est-ce de nature à modifier le déroulement des fermentations ? Malherbe (2003) a comparé les courbes de fermentation obtenues, en conditions rigoureusement identiques de fermentation en phase liquide, dans des cuves de 1 hl (taille pilote) et 110 hl. De son coté, Bely (1990a) a comparé l’échelle pilote avec l’échelle laboratoire (Figure 9). Dans les 2 cas, les courbes d’évolution de la vitesse de fermentation, qui sont plus discriminantes que les courbes d’évolution de la densité, étaient superposables, ce qui montre (i) que dans la gamme de taille utilisée dans la pratique, il n’y a certainement pas d’effet échelle, et (ii) qu’il est tout à fait possible de reproduire à petite échelle les cinétiques fermentaires observées à la taille industrielle. Ce dernier point doit cependant être modulé: - dans le cas des fermentations languissantes et des arrêts de fermentation. En effet, les phénomènes de sédimentation qui interviennent dans les cuves lorsque le niveau de vitesse devient très faible sont très difficiles à reproduire dans un petit fermenteur, - lorsque l’on souhaite décrire les caractéristiques organoleptiques du produit. Vila et al. (2000) ont montré qu’il existait des différences sensorielles significatives entre des échantillons fermentés, en conditions identiques, à l’échelle laboratoire et à l’échelle pilote.

9 QUELQUES PERSPECTIVES De plus en plus, l’industrie viticole doit s’adapter aux goûts et exigences des consommateurs. En quoi de nouvelles approches, au niveau de la fermentation en particulier, pourraient aider à obtenir des produits les plus proches possibles de ceux souhaités ? Voici quelques pistes concernant plus particulièrement la fermentation alcoolique. 9.1 De nouvelles levures Des innovations sont fortement probables dans le domaine des levures œnologiques. En effet, les connaissances ne cessent de progresser, avec un rythme qui s’accélère, grâce, notamment, à l’utilisation des techniques génomiques et post génomiques qui (i) fournissent un nombre considérable d’informations sur la génétique et le métabolisme des levures, et (ii) permettent d’avoir une vision globale de l’ensemble du métabolisme et non, comme auparavant, de se limiter à quelques réactions particulières. Pour les praticiens, l’enjeu est une meilleure compréhension des principaux caractères d’intérêt technologique, avec pour conséquence, (i) une possibilité de reconnaissance de ces caractères et d’aide pour une « sélection assistée », (ii) des perspectives d’amélioration ciblées des souches. Il est à noter que les techniques de croisement sont à nouveau à l’ordre du jour mais, là encore, avec une possibilité d’ « assistance » par des marqueurs moléculaires de façon à rendre cette approche beaucoup plus performante. C’est ainsi qu’a été proposé, par exemple, une souche combinant de bonnes capacités fermentaires avec un très faible niveau de production de composés volatiles indésirables (Marullo et al., 2006). Plusieurs équipes travaillent actuellement sur l’amélioration des souches, avec parmi les objectifs technologiques principaux: la libération des thiols variétaux et la baisse du rendement en éthanol. L’utilisation d’OGM ouvrirait encore la gamme des objectifs technologiques possibles. De nombreuses souches obtenues par génie génétique ont effectivement été décrites (résumées par Dequin, 2001 et Swieger et al., 2005) mais leur utilisation n’est envisageable, pour l’instant, ni en Europe, ni dans l’Hemisphère Sud pour des raisons d’acceptabilité par les consommateurs. Cependant, il faut noter la présence sur le marché Nord Américain de 2 levures de ce type permettant d’effectuer la fermentation malolactique et d’abaisser la production de carbamate d’éthyle. Une autre perspective concerne l’utilisation de levures non S. cerevisiae. En effet, si l’on souhaite élargir la gamme et la complexité aromatique des vins, une possibilité consiste à ensemencer à la fois des levures S. cerevisiae, pour assurer la fiabilité des la fermentation, associées à d’autres espèces telles que Candida stellata, Torulospora delbrukii, Debaryomyces vanrijae ou Hanseniaspora valbyensis pour obtenir des quantités plus importantes d’arômes fermentaires . Pour maîtriser de telles cultures, des stratégies telles que l’inoculation séquentielle ont été proposées (Soden et al., 2000) mais la dissémination des cultures mixtes ne pourra avoir lieu que lorsque seront mieux compris les mécanismes complexes d’interaction entre souches (Cheraiti et al., 2005, Ciani et al., 2006). 9.2 Un contrôle optimisé des fermentations Malgré toutes les avancées en terme de contrôle des fermentations effectuées au cours des dernières décennies, on peut considérer qu’il existe encore une marge considérable, en particulier pour les vinificateurs qui se placent dans l’optique d’élaborer des vins avec des caractéristiques pré-définies. Une perspective prometteuse consiste à adapter le mode de contrôle (température, ajouts de nutriments …) en fonction des caractéristiques du moût utilisé. Cela passe par un suivi très fin et automatique du déroulement de la fermentation, afin de connaître à tout instant, non seulement les concentrations en sucres et alcool mais aussi la vitesse de fermentation. Ce

paramètre, qui n’est pas accessible avec des mesures manuelles, est intéressant à plusieurs titres. En effet, comme nous l’avons vu précédemment, il est directement proportionnel d’une part à l’activité des levures, d’autre part aux besoins instantanées en puissance frigorifique. Il s’agit donc d’une information très pertinente pour améliorer la fiabilité de la fermentation et l’utilisation des ressources de la cave (cuverie, groupes frigorifiques), en ajoutant des nutriments seulement si nécessaire et au meilleur moment, par exemple. Cette vitesse est aussi proportionnelle aux pertes de composés aromatiques dans la phase gaz et représente donc une information certainement aussi pertinente que la température. C’est particulièrement vrai lorsqu’il s’agit d’optimiser la production de ces composés en réalisant des fermentations à basse température (et donc à faible vitesse). L’intérêt potentiel du suivi en ligne est discuté depuis la fin des années 80, mais à cette époque la technologie nécessaire (ordinateurs, capteurs) était certainement incompatible avec le contexte œnologique. Par contre, ce contexte a bien évolué, comme en attestent de nombreux essais industriels actuellement en cours à travers le monde, et les premiers dispositifs commerciaux permettant ce type d’approche ne vont pas tarder à apparaître sur le marché. Au delà du suivi et du contrôle de la cinétique fermentaire, on peut penser que de nouvelles fonctionnalités sont envisageables (Sablayrolles, 2008), telles que le suivi (si possible en ligne) et la prise en compte dans la stratégie de contrôle de « molécules marqueurs » ayant un impact organoleptique, permettant ainsi de piloter directement la fermentation en fonction de paramètres reliés à la qualité du produit. De façon générale, les fermentations devraient, dans les années à venir, être contrôlées de plus en plus finement grâce à l’apparition de nouveaux outils et à la mise en œuvre de nouveaux modes de contrôle. Mais ces évolutions ne seront effectives que si elles sont perçues par les vinificateurs comme de nouveaux moyens mis à leur disposition et venant en complément – et non à la place – de leur expertise.

REFERENCES Aguera E, Sablayrolles JM (2005) Vinificatiuons à l’échelle pilote (100l). II Caractérisation -
intéret. Wine Internet Technical Journal n°7 (www.vinidea.net) Alexandre H, Costello PJ, Remize F, Guzzo J, Guilloux-Benatier M (2004) Saccharomyces
cerevisiae-Oenococcus oeni interactions in wine : current knowledge and perspectives. International Journal of Food Microbiology 93(2):141-154
Ansanay-Galéote V, Blondin B, Dequin S, Sablayrolles JM (2001) Stress effect of ethanol on fermentation kinetics by stationary-phase cells of Saccharomyces cerevisiae. Biotechnology Letters 23:677-681
Bataillon M, Rico A, Sablayrolles JM, Salmon JM, Barre P (1996) Early thiamine assimilation by yeasts under enological conditions : impact on fermentation kinetics. Journal of Fermentation and Bioengineering 82(2):145-150
Bell SJ, Henschke PA (2005) Implications of nitrogen nutrition for grapes, fermentation and wine. Australian Journal of Grape and Wine Research 11(3):242-295
Beltran G, Rozes N, Mas A, Guillamon JM (2007). Effect of low-temperature fermentation on yeast nitrogen metabolism. World Journal of Microbiology and Biotechnology 23(6):809-815
Bely M, Sablayrolles JM, Barre P (1990b) Automatic control of assimilable nitrogen addition during alcoholic fermentation in enological conditions. Journal of Fermentation and Bioengineering 70:1-6
Bely M, Salmon JM, Barre P (1994) Assimilable nitrogen addition and hexose transport system activity during enological fermentations. Journal of the Institute of Brewing 100:279-282
Bely M., Sablayrolles JM, Barre P (1990a) Description of alcoholic fermentation kinetics: its variability and interest. American Journal of Enology and Viticulture, 41 (4), 319-324.
Ben Abdeljalil, C., Cheynier, V., Fulcrand, H., Hakiki, A., Mosaddak, M., & Moutounet, M. (2000). Mise en évidence de nouveaux pigments formés par réaction des anthocyanes avec des métabolites de levures. Sciences des Aliments, 20, 203-220.
Birch RM, Ciani M, Walker GM (2003) Magnesium, calcium and fermentative metabolism in wine yeasts. Journal of Wine Research 14(1):3-15
Blateyron L, Aguera E, Dubois C, Sablayrolles JM (1998) Control of oxygen additions during alcoholic fermentations. Wein-Wissenschaft, 53(3):131-135
Blateyron L, Julien A, Sablayrolles JM (2000) Stuck fermentations - O2 and nitrogen requirements - importance of optimizing their addition. Lallemand Research Meeting, Vienne, Autriche
Bousbouras, G.E., and Kunkee, R.E. (1971). Effect of pH on malolactic fermentation in wine. Am. J. Enol. Vitic. 22, 121-126.
Buescher WA, Siler CE, Morris JR, Threlfall RT, Main GL, Cone GC (2001) High alcohol wine production from grape juice concentrates. American Journal of Enology and Viticulture 52(4):345-350
Butzke CE (1998) Study of yeast assimilable N status in musts from California, Oregon and Washington. American Journal of Enology and Viticulture 49(2):220-224
Castellari L, Pacchioli G, Zambonelli C, Tini V, Grazia L (1992) Isolation and initial characterization of cryotolerant Saccharomyces strains. Italian Journal of Food Science 4:179-186
Cheraiti N, Guezenec S, Salmon JM (2005) Redox interactions between Saccharomyces cerevisiae and Saccharomyces uvarum in mixed culture under enological conditions. Applied and Environmental Microbiology 71(1):255-260

Ciani M, Beco L, Comitini F (2006) Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. International Journal of Food Microbiology 108 (2):239-245
Colombié S, Malherbe S, Sablayrolles JM (2005) Modeling alcoholic fermentation in enological conditions : feasibility and interest. Amerian Journal of Enology and Viticulture 56:238-245
Colombié S, Malherbe S, Sablayrolles JM (2007) Modeling of heat transfer in tanks during wine-making fermentation. Food control 18:953-960
Costello, P.J., Morrison, R.H., Lee, R.H., and Fleet, G.H. (1983). Numbers and species of lactic acid bacteria in wines during vinification. Food Technol. Aust. 35, 14-18.
Cotrell THE, Mc Lellan MR (1986) The effect of fermentation temperature on chemical and sensory characteristics of wines from seven white grape cultivars grown in New York State. American Journal of Enology and Viticulture 37(3):190–194
Delcroix, A., Günata, Z., Sapis, J. C., Salmon, J. M. & Bayonove, C. (1994). Glycosidase activities of three enological yeast strains during winemaking: effect on the terpenol content of muscat wine. American Journal of Enology and Viticulture, 45, 291-296.
Delfini C, Costa A (1993) Effects of the grape must lees and insoluble materials on the alcoholic fermentation rate and the production of acetic acid, pyruvic acid, and acetaldehyde. American Journal of Enology and Viticulture 44(1):86-92
Dequin S (2001) The potential of genetic engineering for improving brewing, wine-making and baking yeasts, Applied Microbiology and Biotechnology, 56(5-6), 577-588
Dubois C, Manginot C, Roustan JL, Sablayrolles JM, Barre P (1996) Effect of variety, year, and grape maturity on the kinetics of alcoholic fermentation. American Journal of Enology and Viticulture 47(4):363-368
Dubourdieu D, Tominaga T, Masneuf I, Peyrot des Gachons C, Murat ML (2006) Role of yeasts in grape flavour development during fermentation: the example of Sauvignon Blanc. American Journal of Enology and Viticulture 57(1):81-88
Garcia A, Garcia LA, Diaz M (1994) Mechanisms for mixing and homogeneisation in beer fermentation. Bioprocess engineering 10:179-184
Henschke PA, Jiranek V (1993) Yeast-Metabolism of nitrogen compounds. In: Wine. Microbiology and Biotechnology (Fleet. G. H., Eds.). Harwood Academic Publishers, Chur, Switzerland, pp 77-165
Higgins PA, Peng AC (1976) Lipid composition of Concord grapes. Am J Enol Vitic 27:32–35
Howell KS, Swiegers J H, Elsey GM, Siebert TE, Bartowsky EJ, Fleet GH, Pretorius IS, Barros Lopes MA (2004) Variation in 4-mercapto-4-methyl-pentan-2-one release by Saccharomyces cerevisiae commercial wine strains. FEMS Microbiology Letters 240(2):125-129
Insa G, Sablayrolles JM, Douzal V (1995) Alcoholic fermentation under enological conditions. Use of a combination of data analysis and neural networks to predict sluggish and stuck fermentations. Bioprocess Engineering 13:171-176
Kelly W.J., Asmundson R.V., Hopcroft D.H. (1989). Growth of Leuconostoc oenos under anaerobic conditions. Am J Enol Vitic. 40, 277-282.
Krieger, S., Trioli, G., and Dulau, L. (2000). "Bacteria and Wine Quality - State of the Art," www.lallemandwine.com.
Malherbe S (2003) Modelling of alcoholic fermentation in winemaking conditions. PhD Thesis. University of Montpellier II
Malherbe S, Fromion V, Hilgert N, Sablayrolles JM (2004) Modeling the effects of assimilable nitrogen and temperature on fermentation kinetics in enological conditions. Biotechnology and Bioengineering 86:261-272

Marullo P, Bely M, Masneuf-Pomarede I, Pons M, Aigle M, Dubourdieu D (2006) Breeding strategies for combining fermentative qualities and reducing off-flavor production in a wine yeast model, FEMS Yeast Research, 6(2), 268-279
Masneuf-Pomarede I, Mansour C, Murat ML, Tominaga T, Dubourdieu D (2006) Influence of fermentation temperature on volatile thiols concentrations in Sauvignon blanc wines. International Journal of Food Microbiology 108(3):385-390
Massoutier C, Alexandre H, Feuillat M, Charpentier C (1998) Isolation and characterization of cryotolerant Saccharomyces strains. Vitis 37(1):55-59
Mazauric, J.P., & Salmon, J.M. (2005). Interactions between yeast lees and wine polyphenols during simulation of wine aging : I. Analysis of remnant polyphenolic compounds in the resulting wines. Journal of Agricultural and Food Chemistry, 53, 5647-5653.
Mazauric, J.P., & Salmon, J.M. (2006). Interactions between yeast lees and wine polyphenols during simulation of wine aging : II. Analysis of desorbed polyphenol compounds from yeast lees. Journal of Agricultural and Food Chemistry, 54(11), 3876-3881.
Moine-Ledoux V, Dubourdieu D (2002) Role yeast mannoproteins with regard to tartaric stabilisation of wines, Bulletin de l'OIV 75(857/858):471-482
Morata, A., Gomez-Cordoves, M. C., Suberviola, J., Bartolome, B., Colomo, B., Suarez, J. A. (2003). Adsorption of anthocyanins by yeast cell walls during the fermentation of red wines. Journal of Agricultural and Food Chemistry, 51, 4084-4088.
Nicolini G, Larcher R, Versini G (2004) Status of yeast assimilable nitrogen in Italian grape musts and effects of variety, ripening and vintage. Vitis 43(2):89-96
Novo MT, Beltran G, Torija MJ, Poblet M, Rozes N, Guillamon JM, Mas A (2003) Changes in wine yeast storage carbohydrate levels during preadaptation, rehydration and low temperature fermentations. International Journal of Food Microbiology 86(1-2):153-161
Palacios A., Suárez C., Krieger S., Theodore D., Otaño L., Peña F. (2004) Percepción del consumidor bien informado de defectos organolépticos del vino, provocados por fermentación maloláctica sin control. Viticultura/Enología Profesional, 94, 29-38 .
Parrou JL, Enjalbert B, Plourde L, Bauche A, Gonzalez B, Francois JM (1999) Dynamic responses of reserve carbohydrate metabolism under carbon and nitrogen limitations in Saccharomyces cerevisiae. Yeast 15(3):191-203
Plouy D (2000) Characterisaton of winemaking fermentors at the pilot scale. Master of Science. Fachhochschule Giessen-Friedberg. Germany
Roustan JL, Sablayrolles JM (2004) Role of trehalose and glycogen in alcoholic fermentation in wine-making conditions. Journal of Wine Research 15:189-202
Sablayrolles J.M., 2008. Kinetics of yeast fermentation during wine production. In Yeasts in the Production of Wine, Romano P. (ed.), Springer-Verlag (sous presse)
Sablayrolles J.M.,Ciosi O., Gerland C., 2008. SOFA : nouveau logiciel pour la simulation de la fermentation alcoolique. 15th International Enology Symposium, Trèves
Sablayrolles JM (1996) Sluggish and stuck fermentations. Effectiveness of ammoniacal nitrogen and oxygen additions. Viticulture and Enology Science 51(3):147-151
Sablayrolles JM, Salmon JM (2001) Vitamins : importance in alcoholic fermentation. Lallemand Research Meeting, Perugia, Italie
Salmon J.M., 2006 - Interactions between yeast, oxygen and polyphenols during alcoholic fermentations : practical implications. LWT-Food Science and Technology, 39(9), 959-965.
Salmon JM, Vincent O, Mauricio JC, Bely M, Barre P (1993) Sugar transport inhibition and apparent loss of activity in Saccharomyces cerevisiae as a major factor of enological fermentations. American Journal of Enology and Viticulture 44(1):56-64
Shinohara T, Kubodera S, Yanagida F (2000) Distribution of phenolic yeasts and production of phenolic off-flavors in wine fermentation. Journal of Bioscience and Bioengineering, 90(1):90-97

Soden A, Francis IL, Oakey H, Henschke PA (2000) Effects of co-fermentation with Candida stellata and Saccharomyces cerevisiae on the aroma and composition of Chardonnay wine. Australian Journal of Grape and Wine Research 6 (1):21-30
Soubeyrand V, Luparia V, Williams P, Doco T, Vernhet A, Ortiz-Julien A, Salmon JM (2005) Formation of micella containing solubilized sterols during rehydration of active dry yeasts improves their fermenting capacity. Journal of Agricultural and Food Chemistry 53(20):8025-8032
Subileau M, Schneider R, Salmon JM, Degryse E (2008) Nitrogen catabolite repression modulates the production of aromatic thiols characteristic of Sauvignon Blanc at the level of precursor transport. FEMS Yeast Research, 8 (5):771-780
Swiegers H., 2008. Développement de souches de levures commerciales ne produisant pas de sulfure d’hydrogène detectable. 5th International Enology Symposium, Trèves
Swiegers JH, Bartowsky EJ, Henschke PA, Pretorius S (2005) Yeast and bacterial modulation of wine aroma and flavour. Australian Journal of Grape and Wine Research 11(2):139-173
Swiegers JH, Pretorius IS (2007) Modulation of volatile sulfur compounds by wine yeast. Applied Microbiology and Biotechnology 74(5):954-960
Torija MJ, Beltran G, Novo M, Poblet M, Guillamon JM, Mas A, Rozes N (2003) Effects of fermentation temperature and Saccharomyces species on the cell fatty acid composition and presence of volatile compounds in wine. International Journal of Food Microbiology 85(1-2):127-136
Vila I., Sablayrolles J.M., Gerland C., Baumes R., Bayonove C., Barre P., 2000. Comparison of ‘aromatic’ and ‘neutral’ strains. Influence of vinification conditions. Vitic. Enol. Sc., 55 (2), 59-66.
Vlassides S, Block D E (2000) Evaluation of cell concentration profiles and mixing in unagitated wine fermentors. American Journal of Enology and Viticulture 51(1):73-80
Zhao (2005) Effect of sulfur dioxide on fermentation kinetics. Master of Sciences. University of Montpellier II. France
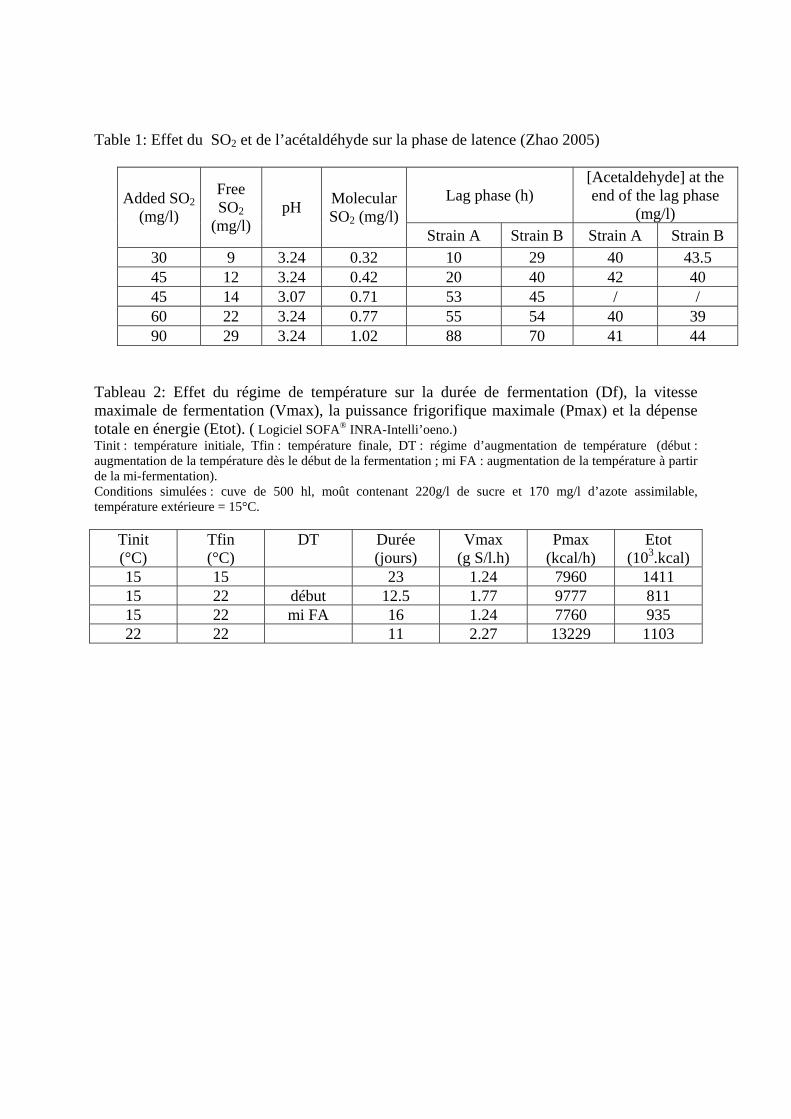
Table 1: Effet du SO2 et de l’acétaldéhyde sur la phase de latence (Zhao 2005)
Lag phase (h) [Acetaldehyde] at the end of the lag phase
(mg/l) Added SO2
(mg/l)
Free SO2
(mg/l) pH Molecular
SO2 (mg/l)Strain A Strain B Strain A Strain B
30 9 3.24 0.32 10 29 40 43.5 45 12 3.24 0.42 20 40 42 40 45 14 3.07 0.71 53 45 / / 60 22 3.24 0.77 55 54 40 39 90 29 3.24 1.02 88 70 41 44
Tableau 2: Effet du régime de température sur la durée de fermentation (Df), la vitesse maximale de fermentation (Vmax), la puissance frigorifique maximale (Pmax) et la dépense totale en énergie (Etot). ( Logiciel SOFA® INRA-Intelli’oeno.) Tinit : température initiale, Tfin : température finale, DT : régime d’augmentation de température (début : augmentation de la température dès le début de la fermentation ; mi FA : augmentation de la température à partir de la mi-fermentation). Conditions simulées : cuve de 500 hl, moût contenant 220g/l de sucre et 170 mg/l d’azote assimilable, température extérieure = 15°C.
Tinit (°C)
Tfin (°C)
DT
Durée (jours)
Vmax (g S/l.h)
Pmax (kcal/h)
Etot (103.kcal)
15 15 23 1.24 7960 1411 15 22 début 12.5 1.77 9777 811 15 22 mi FA 16 1.24 7760 935 22 22 11 2.27 13229 1103

FIGURES
Figure 1. Description d’une fermentation cinétique ‘standard’. Evolution de la population levurienne, du dégagement de CO2 (proportionnel à l’éthanol), de la vitesse de consommation du sucre et de la concentration résiduelle en azote assimilable.
Figure 2. Evolution de la teneur en tréhalose et glycogène dans les levures. Suivi de la population et de la vitesse de production de CO2. (Roustan et Sablayrolles, 2004)
Figure 3. Evolution de la vitesse de dégagement de CO2. Différence entre une fermentation ‘lente’ (1) et une fermentation ‘languissante’ (2) (Insa et al., 1995)
Figure 4. Effet du moment d’ajout de DAP sur la vitesse de production de CO2 (Malherbe 2004).
Figure 5 Effet d’ajouts combinés d’oxygène (5 mg/l) et DAP (300 mg/l) à différents moments de la fermentation. 1 : inoculation, 2 : fin de la phase de croissance, 3 : mi-fermentation (Sablayrolles, 1996)
Figure 6: Déroulement d’une fermentation dans une cuve de 110 hl. Evolution de la vitesse de dégagement de CO2 et de la population levurienne en haut, milieu et bas de cuve. (Malherbe, 2003).
Figure 7: Comparaison des histogrammes de classes de tailles des levures. Figure 7a : levures prélevées à différents endroits de la cuve (haut, bas, milieu, bord). Figure 7b : comparaison entre des levures en suspension (rouge) et des levures sédimentées (bleu) (Aguera et Sablayrolles, 2005).
Figure 8: Effet de la turbidité sur la vitesse de production de CO2. Comparaison de différents pourcentages de bourbes : 0, 0.4 et 3%. (Aguera et Sablayrolles, 2005).
Figure 9: Comparaison entre les cinétiques obtenues à l’échelle laboratoire (1.2 l) et à l’échelle pilote (500 l) (Bely 1990a)
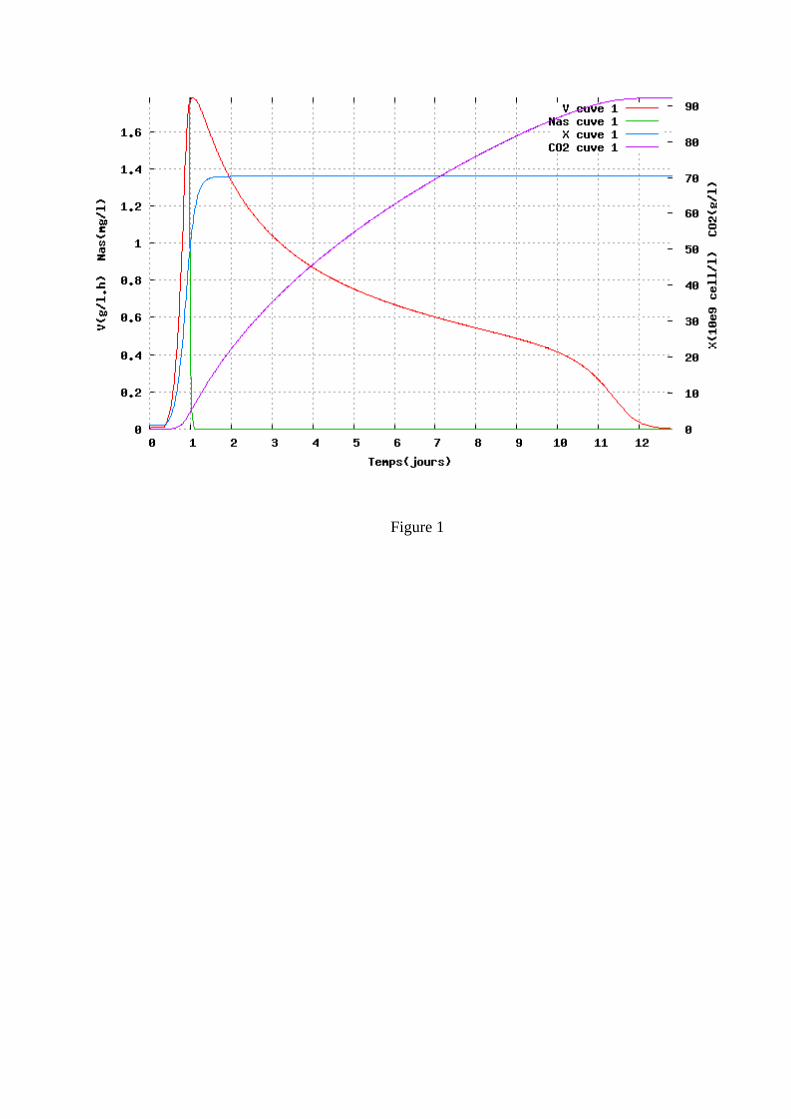
Figure 1
Time (h)
0 20 40 60 80 100
Treh
alos
e, g
lyco
gène
(% d
ry w
eigh
t)
0
5
10
15
20
25
dCO
2/dt (
g/l.h
)
0
1
2
3
Pop
ulat
ion
(106 /m
l)
0
50
100
150
200
250
trehaloseglycogendCO2/dtpopulation
Figure 2
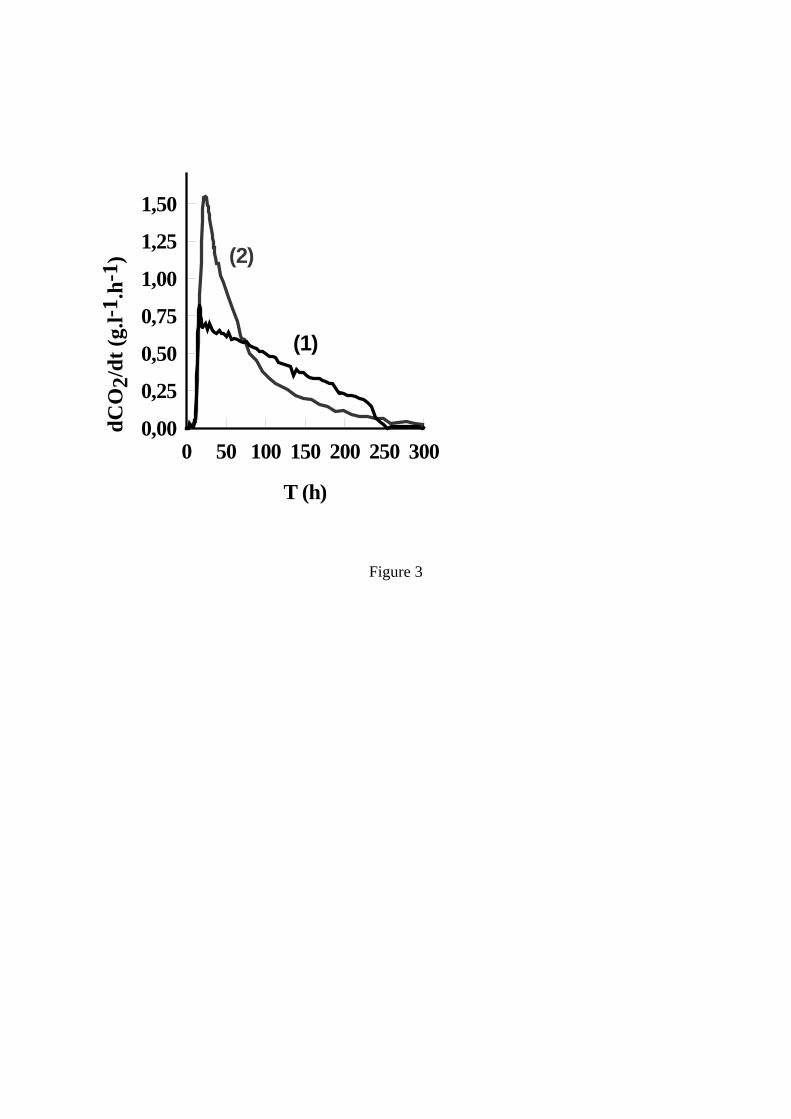
Figure 3
T (h)
0 50 100 150 200 250 3000,00
0,25
0,50
0,75
1,00
1,25
1,50
dCO
2/dt
(g.l-
1 .h-
1 )
(1)
(2)
0
0,2
0,4
0,6
0,8
1
1,2
0 100 200 300 400 500Time (h)
dCO
2/dt (
g/l.h
)
Figure 4
0
10
20
30
40
N1N2
N3
O1 O2 O3
Figure 5a
10
11
12
13
N1N2
N3
O1 O2 O3
Figure 5b
20
30
40
50
60
70
80
0 20 40 60 80 100 120 140 160 180 Time (h)
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Popu
latio
n (M
/ml)
dCO
2/dt (
g/l.h
)
Figure 6
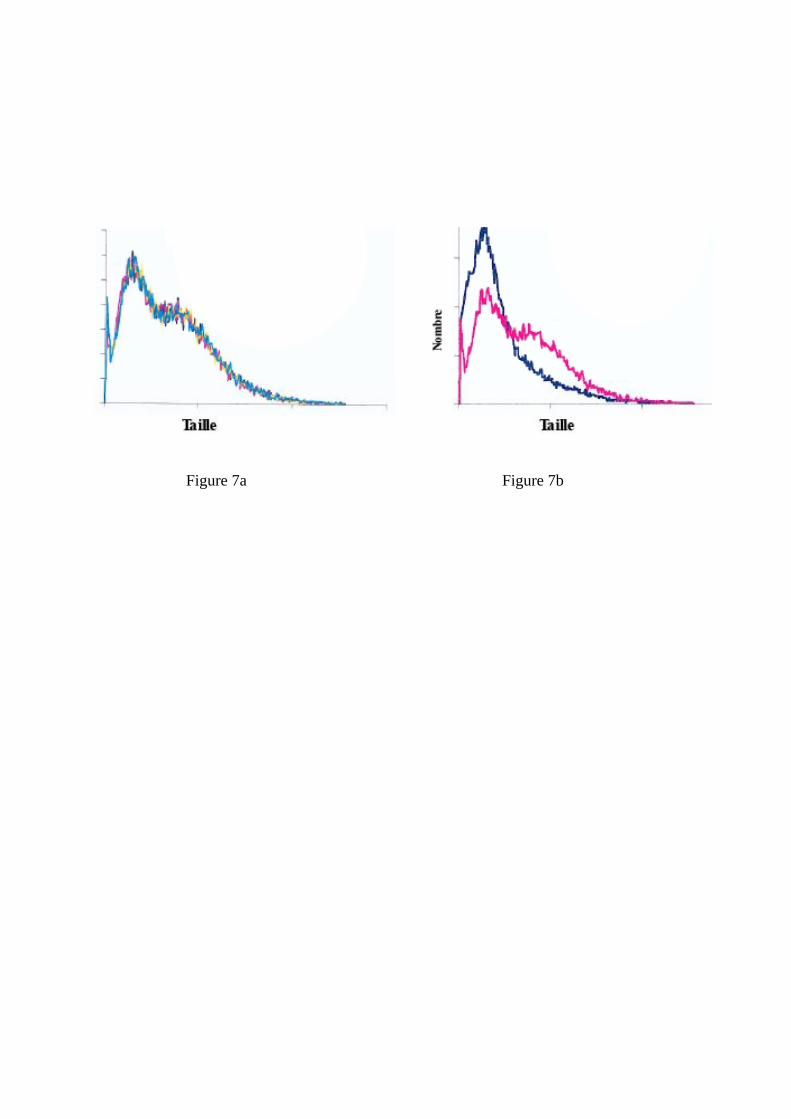
Figure 7a Figure 7b

Figure 8
Temps (h)
Vinifications en blancVinifications en blanc
dCO
2/dt (
g/l.h
)
t-tl (h)
0 30 60 90 120 150 180 210
dCO
2/dt (
g/l.h
)
0,0
0,2
0,4
0,6
0,8
1,0
Figure 9


















