







Langages
Pages
Légal


1
UNIVERSITE de MEDECINE de CRETEIL
(PARIS 12)
Année 2010
THESE
Pour le Diplôme de
DOCTEUR en MEDECINE
Spécialité : OPHTALMOLOGIE
Par
Monsieur Alexis CHASSIGNOL
Né le 29 Août 1979 à Montpellier (Hérault)
Présentée et soutenue publiquement le 22 Octobre 2010
INTERACTIONS CELLULAIRES IN VITRO DE CELLULES
CONJONCTIVALES EN LIGNEE ET DE LYMPHOCYTES ISSUS DU
SANG CIRCULANT. MODELE D’ETUDE DE LA SURFACE
OCULAIRE, IMMUNOLOGIE ET TOXICOLOGIE.
JURY :
MONSIEUR LE PROFESSEUR CHRISTOPHE BAUDOUIN (DIRECTEUR DE THÈSE)
MONSIEUR LE PROFESSEUR LAURENT LAROCHE (PRÉSIDENT DU JURY)
MONSIEUR LE PROFESSEUR BAHRAM BODAGHI
MADAME LE DOCTEUR FRANÇOISE BRIGNOLE-BAUDOUIN
MADAME LE DOCTEUR EMMANUELLE BRASNU

2
Au président du jury de cette thèse, Monsieur le Professeur Laurent Laroche, pour m‟avoir
fait l‟honneur d‟accepter de présider la soutenance de cette thèse,
A mon directeur de thèse et Maître, Monsieur le Professeur Christophe Baudouin, pour votre
confiance, votre soutien et votre disponibilité sans faille,
Aux membres du jury de cette thèse, Monsieur le Professeur Bahram Bodaghi, Madame le
Docteur Françoise Baudouin et Madame le Docteur Emmanuelle Brasnu, pour m‟avoir fait
l‟honneur d‟accepter de siéger dans ce jury,
Veuillez accepter ce travail comme l‟expression de mon profond respect.
A ma femme, ma famille, mes amis et mes collègues,
Merci pour votre soutien, vos conseils, vos encouragements et votre confiance.

3
TABLES DES MATIERES
I. INTRODUCTION...............................................................................................................5
II. TOXICOLOGIE OCULAIRE : EXEMPLE DU CHLORURE DE
BENZALKONIUM.............................................................................................................7
1. LES CONSERVATEURS UTILISÉS DANS LES COLLYRES EN OPHTALMOLOGIE................7
2. LE CHLORURE DE BENZALKONIUM.............................................................................8
3. CONSEQUENCES CLINIQUES : LA SURFACE OCULAIRE DES PATIENTS
GLAUCOMATEUX......................................................................................................10
III. MODES D’ETUDES DE LA TOXICITE DU CHLORURE DE BENZALKONIUM
SUR LA SURFACE OCULAIRE....................................................................................14
1. MODELES ANIMAUX.................................................................................................14
2. ETUDES CLINIQUES ..................................................................................................18
3. CHEZ L‟HOMME........................................................................................................20
4. MODÈLES CELLULAIRES...........................................................................................23
a. LES CULTURES PRIMAIRES............................................................................23
b. LES LIGNEES CELLULAIRES ..........................................................................24
c. LES MODÈLES D‟ÉPITHÉLIUMS RECONSTITUES EN 3 DIMENSIONS.................25
IV. ETUDE DES INTERACTIONS CELLULAIRES DE CELLULES
CONJONCTIVALES EN LIGNEES AVEC DES LYMPHOCYTES ISSUS DU
SANG CIRCULANT.........................................................................................................27
1. INTRODUCTION ........................................................................................................27
2. MATERIELS ET METHODES.......................................................................................28
a. LA LIGNÉE DE CELLULES CONJONCTIVALES .................................................28
b. LES LYMPHOCYTES ISSUS DU SANG CIRCULANT...........................................29
c. LES TRAITEMENTS CELLULAIRES..................................................................29
d. LA COCULTURE............................................................................................30
e. LA CYTOMÉTRIE DE FLUX.............................................................................31
f. LA MICROTITRATION CYTOFLUORIMÉTRIQUE ADAPTÉE AUX
MICROPLAQUES............................................................................................31
g. ANALYSE DE LA MORPHOLOGIE CELLULAIRE...............................................32
h. LA MICROSCOPIE ÉLECTRONIQUE À BALAYAGE............................................32
i. LES RÉACTIFS...............................................................................................33

4
3. RÉSULTATS..............................................................................................................33
a. LES INTERACTIONS CELLULAIRES.................................................................33
b. L‟APOPTOSE.................................................................................................34
c. MODIFICATIONS DES CELLULES ÉPITHÉLIALES.............................................34
d. MODIFICATIONS DES CELLULES LYMPHOCYTAIRES......................................35
e. TEST TOXICOLOGIQUE SUR LE MODÈLE........................................................35
4. DISCUSSION ET CONCLUSION...................................................................................47
V. CONCLUSION ET PERSPECTIVES..............................................................................54
VI. ANNEXES .........................................................................................................................56
VII. REFERENCES BIBLIOGRAPHIQUES..................................................................69
VIII. TITRE ET RESUME EN ANGLAIS...........................................................................82

5
I. INTRODUCTION.
L‟étude de la toxicité des médicaments ou cosmétiques sur la surface oculaire se développa au
cours du 20ème
siècle avec l‟essor de l‟industrie pharmaceutique et la survenue de
complications liées à l‟instillation de certains collyres ou l‟utilisation de certains produits
cosmétiques. Il devint alors indispensable de développer des méthodes permettant d‟étudier la
toxicité oculaire de ces nouveaux produits ou des produits incriminés dans la survenue
d‟évènements indésirables (1). En effet, en 1933, Mc Cally et al. (2) relatent le cas de 17
personnes atteintes de lésions oculaires graves provoquées par l‟utilisation du p-
phénylènediamine dans un produit de maquillage. Ce produit ne sera retiré du marché que
cinq ans plus tard. Latvenar et Molitor (1939) sont parmi les premiers à proposer l‟utilisation
du lapin pour prédire l‟irritation oculaire chez l‟homme, mais ce n‟est qu‟en 1944 que
Friedenwald et al. (3) décrivent les effets des acides et des bases sur l‟œil de lapin et mettent
en place pour la première fois un système de cotation subjective pour qualifier les effets sur la
cornée, la conjonctive et l‟iris. Ainsi, John Draize et al. (4) publient, la même année, les
résultats de leur étude de la toxicité de produits et améliorent le système de cotation décrit par
Friedenwald en standardisant la méthode et en simplifiant les cotations. La toxicité oculaire
est ainsi déterminée par l‟observation des lésions de la cornée, de l‟iris et de la conjonctive,
tout cela regroupé sous le terme « test de Draize ». Mais rapidement, pour des raisons
éthiques évidentes, la mise en place de méthodes alternatives aux modèles animaux s‟imposa
avec le développement de modèles In vitro et In vivo d‟étude de la toxicologie oculaire. En
1978, le terme d‟« alternative » fut exploité par le physiologiste David Smyth (5) qui l‟utilisa
pour décrire tout changement à partir des procédures de l‟époque visant à remplacer les
animaux, à en réduire leur nombre utilisé et à diminuer leur douleur et leur stress.
De nos jours, un des grands axes de recherche en toxicologie oculaire est l‟étude de l‟effet des
conservateurs contenus dans les différentes formulations ophtalmologiques (collyres, produits
de conservations des lentilles). En effet, la Pharmacopée actuelle recommande la présence
d‟un agent antimicrobien dans chaque flacon de collyre afin de limiter la prolifération
d‟agents infectieux une fois que la bouteille de collyre a été ouverte. Les conservateurs en
ophtalmologie appartiennent à différentes familles de composés chimiques incluant l‟EDTA,
les dérivés mercuriels, l‟alcool, le paraben, les biguanides chlorés (comme la chlorhexidine)
et les ammoniums quaternaires (comme le chlorure de Benzalkonium). Ces derniers, du fait
de leur faible pouvoir allergisant et de leur apparente innocuité, se sont rapidement imposés
comme les conservateurs les plus largement utilisés en ophtalmologie. Cependant, depuis un

6
passé récent, l‟accumulation de preuves cliniques et expérimentales sur l‟implication des
traitements anti-glaucomateux au long cours dans l‟apparition d‟une inflammation chronique
de la surface oculaire mettent en exergue le potentiel toxique des conservateurs de ces
collyres (et principalement du chlorure de benzalkonium) et leur nocivité pour l‟œil.
La première partie de cette thèse sera consacrée à la description des conservateurs dans les
collyres en ophtalmologie en s‟attardant sur leur chef de file, le chlorure de benzalkonium,
pour détailler ensuite les conséquences cliniques de leur utilisation prolongée. Dans la
seconde partie, nous nous intéresserons plus particulièrement aux différents modèles d‟études
de la toxicité oculaire de ces conservateurs. En effet, l‟essor des méthodes « alternatives »,
afin de diminuer l‟utilisation des modèles animaux, tant à privilégier maintenant les modèles
In vitro, tels que l‟utilisation des lignées cellulaires. Les lignées cellulaires sont un outil
intéressant car peu coûteux, reproductible et applicable à différentes techniques d‟analyse.
Cependant, l‟utilisation de ces lignées monocellulaires dans l‟étude des effets toxiques des
collyres sur la surface oculaire présente comme principal inconvénient de ne pas correspondre
parfaitement à la réalité anatomique de la conjonctive ou de la cornée. En effet, la cornée et la
conjonctive sont des épithéliums au sein desquels coopèrent différents types cellulaires
associés aux cellules épithéliales. Extrapoler les résultats d‟études de toxicité des
conservateurs sur des lignées monocellulaires de cellules épithéliales de conjonctive pourrait
paraître réducteur. Ainsi, l‟objectif du travail faisant l‟objet de cette thèse fut d‟étudier le
comportement et les interactions cellulaires d‟une coculture de lignée de cellules épithéliales
de conjonctive (Wong Kilbourne Derivative of Chang) et de lymphocytes issus du sang
circulant, puis d‟effectuer des tests de toxicité avec le chlorure de benzalkonium. Les résultats
de cette étude seront exposés dans la troisième partie de cette thèse.

7
II. TOXICOLOGIE OCULAIRE : EXEMPLE DU CHLORURE DE BENZALKONIUM.
1. Les conservateurs utilisés dans les collyres en ophtalmologie.
La contamination des flacons de collyres se fait essentiellement par le contact avec les mains
du patient lors de la manipulation du flacon ou lors de l‟utilisation du collyre. Un contact de
l‟embout du flacon peut se produire avec les cils, les paupières, les larmes, la conjonctive ou
la peau du visage. Une transmission interhumaine est également possible lorsque le même
flacon est utilisé pour différents patients, situation qui peut se présenter au sein d‟une
structure hospitalière ou d‟une structure privée de soins ou au sein d‟une famille. Dans les
années 60, dans les suites d‟un incident de ce type à l‟hôpital de Birmingham, les autorités
britanniques avaient alerté l‟industrie pharmaceutique sur la nécessité de développer des
préparations de collyres sous la forme d‟unidoses et de limiter la durée d‟ouverture d‟un
flacon (6). Ce n‟est que dans les années 1970 qu‟il a été décidé de rendre obligatoire la
présence d‟agents antimicrobiens (conservateurs) dans les flacons multidoses de collyre. La
pharmacopée américaine basait l‟efficacité d‟un conservateur sur le « test du pouvoir
conservateur ». Ce test consistait, In vitro, à inoculer 1x106 Colony Forming Units (CFU)/mL
à l‟instant T=0 de divers micro-organismes tels que des bactéries (Staphylococcus aureus,
Pseudomonas aeruginosa, Escherichia coli) ou des champignons (Aspergillus niger et
candida albicans). L‟efficacité prouvée du conservateur se base sur les résultats suivants :
réduction de 1.0 Log au 7ème
jour, réduction de 3.0 Log au 14ème
jour sans prolifération des
survivants, pas de prolifération des survivants au 28ème
jour (7). Cependant, de tels effets
cytotoxiques des conservateurs sur les micro-organismes ne peuvent être obtenus sans une
certaine toxicité, même minimale, sur les tissus sur lesquels la goutte de collyre a été
appliquée. Les autorités de santé sont, de nos jours, particulièrement sensibilisées à ce
problème, de telle façon que, récemment, l‟Agence Européenne de Médecine (EMEA,
European Medicine Agency) a statué sur la nécessité pour l‟industrie pharmaceutique et tous
les partenaires de santé de supprimer l‟utilisation des conservateurs pour « les patients ne
tolérant pas les collyres conservés » et ceux traités au long cours, en utilisant « la
concentration minimale efficace qui satisfasse la fonction antimicrobienne du conservateur »
et en promulguant « de nouvelles préparations ophtalmiques ne contenant pas de
conservateurs à base de mercure » (8).

8
2. Le chlorure de benzalkonium.
De nos jours, le conservateur le plus communément utilisé dans les préparations
ophtalmologiques est le chlorure de benzalkonium (BAC). Le BAC, aussi connu sous le nom
de chlorure d'alkyldiméthylbenzylammonium et ADBAC, est un mélange de chlorures
d'alkylbenzyldiméthylammonium avec des chaînes carbonées de longueur variable. Ce
produit est un agent de surface cationique de la famille des ammoniums quaternaires.
FIG 1. FORMULE CHIMIQUE DU CHLORURE DE BENZALKONIUM
Le chlorure de benzalkonium est facilement soluble dans l'éthanol et l'acétone. Bien que sa
dissolution dans l'eau soit lente, les solutions aqueuses sont plus faciles d'emploi et sont plus
largement utilisées. Les applications sont très variées, allant de la formulation de désinfectants
à l'inhibition de corrosion microbienne dans les huiles en utilisant des propriétés d‟agent de
transfert de phase et de surfactant cationique. Les ammoniums quaternaires sont des
composés bipolaires hautement hydrosolubles. En pharmacologie, l'activité biocide la plus
importante est associée aux dérivés alkylés en C12 à C14. On pense que le mécanisme
bactéricide est dû à la disruption des interactions intermoléculaires. Ceci peut causer la
dissociation des lipides dans la membrane cellulaire, ce qui compromet la perméabilité de la
cellule et induit une fuite de son contenu. Les solutions de BAC sont des agents bactéricides à
action rapide et de durée modérément longue. Les spores des bactéries sont considérées
comme résistantes et les bactéries à Gram positif (Staphylococcus) sont généralement plus
sensibles que les Gram négatifs (Pseudomonas aeruginosa). L'activité n'est pas grandement
influencée par le pH, mais augmente aux températures élevées et avec la durée d'exposition.

9
De nouvelles formulations utilisant du benzalkonium mélangé à d'autres ammoniums
quaternaires peuvent être utilisées pour étendre le spectre biocide et augmenter l'efficacité du
désinfectant. Par exemple, l‟action combinée du BAC avec l‟EDTA 0.1% augmente l‟action
bactéricide contre les bactéries Gram négatif. De plus, le BAC a aussi une activité spermicide.
Enfin, les concentrations de BAC le plus généralement utilisées vont de 0.004 à 0.025%.
Le BAC a été largement utilisé pour son bon rapport efficacité/sécurité apparent. Il a été
décrit comme modérément allergisant (9, 10) mais possédant d‟importantes propriétés
antimicrobiennes. En effet, une étude a montré que parmi 5 conservateurs différents
(BAC/EDTA, paraben, chlorobutanol, complexe argent-chloride et le complexe stabilisé
purite oxychloré), seul le BAC/EDTA remplissait les critères de validité d‟activité
antimicrobienne (11).
De plus, il a été montré que les substituts lacrymaux conditionnés en unidoses sans
conservateur étaient à risque de contamination, après plusieurs utilisations au-delà de 10
heures d‟ouverture (12). Le risque de contamination est augmenté chez les patients les plus
âgés et en cas de manipulation inappropriée avec les doigts. Cependant, il faut noter que le
taux de contamination des unidoses de collyres ne dépasse pas les 2%, même dans les pires
cas de manipulations répétées, alors qu‟il a été répertorié près de 35% de taux de
contamination des flacons de collyres préservés avec le BAC après 15 jours d‟utilisation (13),
mettant en exergue la « relative » sécurité apportée par l‟utilisation du BAC en tant que
conservateur antimicrobien. De plus, il a été décrit que certaines souches de Pseudomonas
aeruginosa seraient résistantes à l‟action antimicrobienne du BAC et pourraient proliférer
dans des solutions contenant du BAC (14). Cette résistance serait corrigée par l‟addition
d‟EDTA au BAC, mais pourrait augmenter alors grandement la toxicité sur les tissus
avoisinants.
L‟un des principaux arguments de l‟utilisation du BAC comme conservateur des collyres
utilisés en ophtalmologie, est l‟amélioration de la pénétration du principe actif au niveau de la
chambre antérieure, du fait de l‟action de disruption de la barrière hydrophobe de l‟épithélium
cornéen par le BAC. L‟augmentation de concentration du principe actif dans l‟humeur
aqueuse pourrait permettre une meilleure efficacité du collyre. La pénétration cornéenne de
cyclosporine A appliquée en topique serait améliorée par la présence de BAC en solution dans
le collyre (15), et le couple BAC/EDTA permettrait une augmentation de la pénétration
transcornéenne d‟aciclovir (16) ou de certains antibiotiques (17). Cependant une autre étude
n‟a pas trouvé de différence dans la concentration de l‟analogue des prostaglandines
Tafluprost dans l‟humeur aqueuse après instillation d‟une solution conservée et d‟une solution

10
non conservée (18). Il apparaît tout de même évident que les différentes formulations
(hydrophilie vs. lipophilie, solutions vs. suspension ou émulsion, prodrogue vs. principe actif)
de la multitude de variétés de produits actuellement disponibles, entraînent des différences
majeures de pharmacocinétique, alors qu‟il n‟y a souvent pas de corrélation réelle entre la
pharmacocinétique d‟un produit et son efficacité. Et, de plus, hormis le fait que les
modifications histologiques majeures de la barrière cornéenne, liées à l‟utilisation de collyres
conservés permettant l‟amélioration de la pénétration d‟un produit, peuvent être considérées
comme des effets secondaires toxiques (19), il a fréquemment été démontré, à quelques
exceptions près, une équivalence totale en terme d‟efficacité des collyres antiglaucomateux
conservés et non conservés.
3. Conséquences cliniques : la surface oculaire des patients glaucomateux.
Le glaucome est une maladie chronique qui concerne près d‟un million de personnes en
France et environ 30 millions dans le monde. Il s„agit initialement d‟une dégénérescence du
trabéculum, conduisant à une raréfaction cellulaire progressive, à un remodelage matriciel et à
une hyperpression oculaire qui entraîne secondairement la perte des cellules ganglionnaires
rétiniennes et donc, de facto, la perte de la fonction visuelle. Le troisième tissu touché par le
glaucome, et qui nous importe ici, est la surface oculaire, qui reçoit pendant des années ces
traitements locaux toxiques et/ou proinflammatoires, stimulant à la fois l‟apoptose épithéliale
et le système immunitaire. Cliniquement, il est possible d‟observer que, par exemple, suite à
l‟instillation chronique au long cours de ces collyres conservés, des symptômes de
conjonctivite allergique ou de syndrome sec oculaire, une pseudophemphigoïde cicatricielle
ou encore une augmentation de l‟échec de la chirurgie filtrante peuvent survenir.
Les symptômes d‟allergie oculaire sont dominés par le prurit, le chemosis, l‟œdème palpébral,
le larmoiement, la rougeur oculaire et la photophobie. Dans certains cas, il est noté jusqu‟à la
présence d‟une conjonctivite giganto-papillaire, voire de sévères réactions allergiques de type
IV. L‟ensemble de ces manifestations d‟allergie oculaire est donc très varié dans sa
présentation ainsi que le délai de survenue, certaines réactions retardées pouvant même mimer
une blépharite chronique avec une inflammation à bas bruit. Par conséquent, le traitement
simple de l‟allergie oculaire visant à soustraire l‟allergène en arrêtant le traitement n‟est que

11
rarement envisagé. Une méta-analyse de 28 essais cliniques randomisés relatait de 0% à 16%
seulement d‟arrêt du traitement pour des manifestations d‟allergie oculaire (20).
Les manifestations de sécheresse oculaire rassemblent un cortège de symptômes aspécifiques
mais particulièrement fréquents tels que l‟ « inconfort » lors de l‟instillation du collyre, une
sensation de brûlure ou de picotement, une sensation de corps étranger, une sensation d‟ « œil
sec », un larmoiement ou des démangeaisons palpébrales. Une enquête épidémiologique
conduite en 2002 sur 4107 patients glaucomateux a évalué le taux de manifestations oculaires
d‟œil sec en pratique clinique de routine et trouvé une prévalence des symptômes très élevée
pour les patients utilisant des collyres conservés. Cette fréquence de signes et de symptômes
oculaires augmentait avec le nombre de collyres conservés utilisés (21). Sur le même
principe, l‟analyse des données provenant de 9658 patients retrouvait une incidence des
symptômes oculaires de sécheresse comprise entre 30 et 50% (22). Les symptômes de
sécheresse oculaires surviennent plus fréquemment lorsque le glaucome est plus sévère,
lorsque trois (ou plus) collyres sont utilisés, et cette fréquence augmente avec la durée
d‟évolution du glaucome (23). Il a par ailleurs été montré que ces effets secondaires oculaires
liés à l‟instillation des collyres avaient un effet néfaste non négligeable sur la qualité de vie
des patients (24) et étaient la deuxième cause de non observance ou de changement de
traitement (25).
Les manifestations de pseudopemphigoïde cicatricielle conjonctivale surviennent chez les
patients glaucomateux traités de longue date. Elles sont liées à une augmentation de la densité
fibroblastique au niveau de la substantia propria sous-conjonctivale en rapport avec une
augmentation de la présence de cellules inflammatoires (26). L‟instillation de collyres
conservés serait de loin la première cause de pemphigoïde cicatricielle (27).
L‟échec de la chirurgie filtrante pratiquée chez les patients glaucomateux, telle que la
trabéculectomie, serait initialement lié à une réponse fibroblastique conjonctivale excessive
au niveau de la bulle de filtration dans le premier mois post-opératoire. Cette inflammation
conjonctivale avec prolifération fibroblastique et dépôts au niveau de la matrice
extracellulaire aboutit à un blocage de l‟évacuation de l‟humeur aqueuse dans l‟espace sous
conjonctival fibrosé. Par conséquent, tous les facteurs favorisant la fibrose post opératoire au
niveau de la bulle de filtration de la trabéculectomie sont des facteurs de risque d‟échec de la
chirurgie. L‟utilisation au long cours de collyres conservés chez les patients glaucomateux
induit à terme une inflammation conjonctivale avec pour principale conséquence, une
augmentation de la fibrose post opératoire. Différentes études cliniques viennent confirmer
que le taux de succès de la trabéculectomie est influencé négativement par l‟utilisation au

12
long cours de collyres conservés (28) et qu‟il y a une relation entre le taux d‟échec de la
chirurgie et le nombre de collyres conservés utilisés et leur durée d‟utilisation (29).
A un stade infra clinique, il a été clairement démontré que même sans la présence de
symptômes oculaires évidents comme ceux précédemment décrits, l‟utilisation de collyres
anti-glaucomateux conservés était responsable de la présence anormale d‟inflammation au
niveau de l‟épithélium conjonctival et de la substantia propria. Il a en effet été noté une
augmentation du nombre de macrophages, lymphocytes, mastocytes et fibroblastes et une
diminution des cellules à mucus chez les patients opérés après de longues années de
traitement en comparaison à ceux opérés en thérapeutique primaire. Ces résultats ont été
observés lors d‟études immunohistologiques faites sur la conjonctive et la capsule de Tenon
des patients, obtenues par biopsies conjonctivales réalisées au moment de la chirurgie (30).
Notre équipe a réalisé des travaux similaires et a montré, par le biais d‟études
immunohistologiques, une expression positive de marqueurs d‟inflammation et d‟apoptose sur
les cellules conjonctivales des patients traités au long cours par les collyres conservés, les
cellules conjonctivales ayant été obtenues par biopsies (26). Les empreintes conjonctivales
permettent de collecter de manière rapide, non invasive et indolore, la couche la plus
superficielle de l‟épithélium conjonctival comprenant les cellules épithéliales, les cellules à
mucus et les cellules inflammatoires infiltrant la conjonctive (31). Les empreintes
conjonctivales sont donc un outil indispensable d‟étude des manifestations infra cliniques de
la surface oculaire, en particulier des manifestations de sécheresse oculaire. Initialement, les
empreintes conjonctivales avait permis de mettre en évidence des changements de la surface
oculaire (augmentation du degré de métaplasie, diminution de la densité en cellules à
mucus…) (32), mais récemment, différentes techniques immunohistologiques appliquées aux
empreintes conjonctivales ont été développées pour évaluer plus précisément l‟inflammation
conjonctivale au cours des maladies de la surface oculaire. Notre équipe a ainsi été capable de
quantifier l‟expression d‟HLA-DR sur les empreintes conjonctivales. HLA-DR n‟est
normalement pas exprimé sur les cellules conjonctivales, sauf en cas de processus
inflammatoire. HLA-DR est alors apparu comme le marqueur fiable pour démontrer que
l‟épithélium est impliqué dans une pathologie inflammatoire de la surface oculaire, mais
également pour quantifier cette inflammation, montrant son intérêt principalement lorsque la
pathologie n‟est cliniquement pas ou peu inflammatoire. La première étude
immunocytologique réalisée par notre équipe, incluant 107 yeux de 55 patients glaucomateux
montrait une absence totale de réactivité pour l‟anticorps monoclonal HLA-DR dans le
groupe non traité et à l‟inverse une positivité de l‟expression de HLA-DR pour 43/88 yeux

13
dans le groupe traité (33). De plus, notre équipe a développé la technique de cytométrie de
flux appliquée aux empreintes conjonctivales, permettant d‟évaluer de façon fiable et
reproductible l‟expression du marqueur HLA-DR ou de cytokine proinflammatoires au sein
de l‟épithélium conjonctival chez les patients glaucomateux même asymptomatiques
cliniquement (34, 35, 36). Enfin, des travaux plus récents utilisant les mêmes techniques de
cytométrie de flux sur empreintes conjonctivales ont permis d‟étudier l‟expression des
marqueurs CCR5 et CCR4, deux marqueurs associés aux profils d‟inflammation de
lymphocytes T helper Th1 et Th2 respectivement, Th2 étant plutôt impliqué dans les réactions
allergiques et Th1 étant plutôt impliqué dans les réactions de toxicité. Il a été noté, chez des
patients glaucomateux traités au long cours, une expression élevée simultanée des deux
marqueurs, illustrant toute la complexité des mécanismes mis en œuvre au cours du
glaucome, incluant des réactions allergiques et toxiques, une stimulation directe de cellules
inflammatoires, une déficience du film lacrymal, une destruction des cellules épithéliales.
Toutes ces modifications infra cliniques surviennent très rapidement après l‟instauration d‟un
traitement anti-glaucomateux, puisque l‟expression du marqueur HLA-DR aurait été mise en
évidence dès le deuxième mois de traitement (37).

14
III. MODES D’ETUDES DE LA TOXOCITE DU CHLORURE DE BENZALKONIUM
SUR LA SURFACE OCULAIRE
1. Modèles Animaux
Les produits chimiques, les produits cosmétiques et les produits pharmaceutiques doivent être
testés avant toute utilisation chez l‟homme, du fait de leur pouvoir irritant potentiel et du
risque encouru pour l‟œil humain qui serait en contact avec l‟un de ces produits. Cependant,
jusqu‟à aujourd‟hui encore, la seule méthode admise et validée mondialement par les autorités
compétentes pour l‟étude du pouvoir irritant potentiel d‟un composé, est le test de Draize (4),
qui d‟ores et déjà est critiqué et condamné par les associations de défense et de protection de
animaux.

15
Des alternatives au classique test de Draize ont été proposées, comme le test de Draize à
« faible volume » (M) qui serait plus prédictif et moins douloureux pour l‟animal, le volume
du produit instillé étant réduit à 0.01ml au lieu de 0.1ml (38). Une autre alternative au test de
Draize, se rapprochant encore plus des conditions du vivo sans sacrifier d‟animaux, est la
technique de « l‟œil énucléé », utilisant des yeux isolés de lapin, de bœuf ou de poulet
provenant d‟abattoirs, permettant de classer les irritants oculaires sévères selon trois
paramètres : l‟opacification cornéenne, la prise de fluorescéine et surtout l‟œdème cornéen
qui est le paramètre le plus objectif et le plus discriminant (39).
Par ailleurs, l‟utilité du test de Draize, sa fiabilité et sa précision sont également critiquées par
les équipes de recherche du fait de l‟importante variabilité des résultats, et donc de l‟absence
de prévisibilité de la réponse humaine au toxique préalablement testé sur l‟animal (40, 41). En
effet, le test de Draize est principalement basé sur les changements macroscopiques observés
sur les cornées, conjonctives et iris de lapin. Différents scores de réponse au collyre peuvent
être utilisés. Cependant, quelque soit le score mis en pratique, le modèle d‟œil de lapin montre
ses limites principalement lors de l‟utilisation de tests d‟irritation peu sévère et pour des
substances toxiques de faibles puissance.
Afin d‟améliorer le qualité du modèle d‟œil de lapin, il a été développé, en addition aux
mesures cliniques et histologiques post mortem, la méthode de l‟analyse de la surface oculaire
de l‟œil de lapin in vivo, non traumatique par le biais de l‟utilisation de la microscopie
cornéenne confocale in vivo (IVCM). Cet outil permet d‟examiner de façon reproductible,
sensible et sûre la surface oculaire des animaux de laboratoire en réalisant des images de
coupe de tissus vivants sans sacrifier les animaux et sans préparation préalable, avec une très
bonne corrélation entre les images obtenues et l‟histologie. L‟intérêt de la microscopie
confocale réside également dans son pouvoir à mettre en évidence de minimes altérations de
la surface oculaire après exposition à des agents de toxicité faible ou modérée, et d‟étudier
leur réversibilité dans le temps.
La première étude toxicologique de ce type fut menée par Ichijima et al. (42), en s‟appuyant
sur le développement d‟un modèle de stress aigu de la surface oculaire pouvant mimer les
effets à long terme d‟un toxique de faible puissance instillé sur une courte période de temps.
Les auteurs étudièrent les changements induits par l‟application du BAC sur la cornée de
lapin en utilisant une méthode d‟investigation combinant le microscope confocal in vivo et la
microscopie électronique à balayage. L‟application de 15 gouttes de BAC à la concentration
de 2.10−2
%, 10−2
%, et 5.10−3
% à 5 minutes d‟intervalles entraînait la desquamation des
cellules les plus superficielles de l‟épithelium cornéen, visible au microscope confocal. Cette

16
desquamation augmentait avec la concentration de BAC instillée. De plus, une heure après la
dernière goutte appliquée, des cellules inflammatoires apparaissaient à la surface de la cornée.
Toutes ces données furent confirmées par l‟analyse en microscopie électronique, alors que
l‟application de collyres sans conservateur ne provoquait, quant à eux, aucune toxicité sur
l‟épithélium cornéen.
Notre équipe a également utilisé le modèle d‟oeil de lapin à des fins d‟investigation
toxicologique à l‟échelon histologique toujours par le biais de l‟utilisation du microscope
confocal in vivo. L‟apport du récent module cornéen adapté pour le tomographe rétinien de la
marque Heidelberg (Rostock Corneal Module of the Heidelberg Retina Tomograph -
HRT/RCM - Heidelberg Engineering, Heidelberg, Allemagne) (Figure1) a permis d‟obtenir
une résolution de près de 1 µm, et trouvant ainsi des applications dans un bon nombre de
pathologies humaines (43, 44, 45). En plus de l‟analyse des altérations cornéennes induites
par la toxicité du BAC, la microscopie confocale in vivo a permis d‟analyser la périphérie
cornéenne, le limbe et même la conjonctive, structures qui jusqu‟alors n‟étaient accessibles
sur le plan histologique que post-mortem.
Figure 1 : « Rostock Cornea Module »
(Baudouin et al. Apport de la microscopie confocale in vivo pour l‟exploration de la surface oculaire. 2006)

17
Les travaux récents de notre équipe ont abouti à la mise en évidence in vivo des post mortem
(conjunctiva-associated lymphoid tissue) et de leur infiltration inflammatoire après activation
par le BAC (46). Cette infiltration de cellules inflammatoires en transit dans le CALT
semblerait liée à la concentration de BAC appliquée, du fait de l‟absence de changement
observé au niveau du CALT après utilisation de collyres sans conservateur, alors qu‟une
importante infiltration de cellules inflammatoires au sein du CALT étaient observée,
augmentant avec la concentration de BAC utilisée.
Le modèle d‟œil de lapin est également un bon support pour l‟utilisation des empreintes
conjonctivales comme outils d‟étude de la surface oculaire (47). En effet, cette technique
permet d‟effectuer de multiples manipulations sur un même animal avant son sacrifice,
réduisant ainsi le nombre d‟animaux utilisés à des fins d‟expérimentations scientifiques.
Le modèle de rat est un modèle utile d‟étude de la surface oculaire car plus adapté pour la
manipulation et pour l‟hébergement que le lapin. Le modèle de rat a déjà été utilisé pour
démontrer la toxicité du BAC, en confirmant que l‟application de collyres antiglaucomateux
contenant du BAC provoquait des changements histologiques au niveau de la surface oculaire
des rats, même ceux qui étaient en bonne santé. Ces changements n‟étaient pas observés
lorsque le collyre sans conservateur était appliqué (26). Cependant, même si, sur un plan
purement pratique, l‟utilisation du rat paraît plus simple que celle du lapin, les modèles de rat
ne sont pas si souvent que cela utilisés pour les études de toxicologie oculaire, du fait de
l‟extrême variabilité de leur comportement oculaire et des modifications observables au
niveau de la surface oculaire de rat. L‟utilisation de la microscopie confocale in vivo a
toutefois permis d‟améliorer l‟utilité de ces modèles de rat en se couplant à l‟analyse
biomicroscopique et à l‟étude histologique post-mortem. Ainsi, notre équipe a réalisé une
série d‟investigations de la toxicité du BAC sur l‟œil de rat après instillations répétées et à des
concentrations croissantes. Les effets du BAC furent mesurés tout d‟abord cliniquement par
l‟utilisation du test de Draize, montrant une irritation cliniquement significative seulement
pour les concentrations de BAC les plus élevées (0.5% et 0.25%), puis confirmés par la
mesure des changements à l‟échelon histologique par l‟utilisation du microscope confocal
HRT-II, montrant une altération de la cornée de rat en relation dose-dépendante avec le BAC.
Les concentrations les plus élevées de BAC induisaient une totale désépithélialisation de la
cornée avec altération en profondeur du stroma cornéen jusqu‟à l‟infiltration inflammatoire, la
néovascularisation et la fibrose endothéliale de la cornée (48, 49). les plus faibles
concentrations de BAC n‟entraînaient des dommages que pour les zones les plus
superficielles de l‟épithélium cornéen. Ces dommages auraient pu passer inaperçus sans

18
l‟évaluation des critères les plus sensibles. C‟est ainsi que notre équipe a développé une grille
de cotation des altérations de la surface oculaire observées en microscopie confocale in vivo
après exposition à un toxique (50). Cette grille s‟appuyant sur l‟observation de différents
paramètres que sont la taille et la forme des cellules, leur mode de réflectivité, l‟inflammation,
la néovascularisation et la mesure de l‟épaisseur cornéenne. Son intérêt est de permettre la
standardisation de l‟évaluation de l‟atteinte cornéenne à l‟échelle cellulaire même après
exposition à des faibles doses d‟agents irritants.
2. Etudes Cliniques
Très peu d‟études prospectives se sont posées la question du rôle délétère de la présence d‟un
conservateur dans les collyres. Cela est principalement dû au manque actuel de solution
proposée parmi les collyres sans conservateur, et dans une plus large mesure dû au fait que la
surface oculaire subit trop rapidement les effets toxiques du conservateur après l‟instauration
du traitement. Cependant, une étude sur volontaires sains a trouvé que l‟instillation de timolol
conservé causait plus d‟instabilité du film lacrymal qu‟après instillation de timolol non
conservé (51). Notre équipe a trouvé des résultats similaires en mesurant le break-up time
chez des volontaires sains, le trouvant significativement plus court après instillation de
carteolol contenant du BAC (52). Il faut noter que ces deux études cliniques ont été réalisées
chez des volontaires sains, jeunes ayant une surface oculaire parfaitement normale au stade
initial. Cela permet de mieux comprendre pourquoi il est observé autant de manifestations de
sécheresse oculaire chez les patients glaucomateux, qui généralement accumulent de longues
années de traitement, traités par de multiples collyres, et ces patients sont donc beaucoup plus
sujets à présenter une surface oculaire altérée et affaiblie. Deux autres études prospectives ont
analysé les conséquences de l‟application de BAC chez le volontaire sain. La première
conclut que même de petites concentrations de BAC induisent une perte en cellules à mucus
ainsi qu‟une augmentation du rapport nucléo-cytoplasmique, qui sont deux caractéristiques du
syndrome sec oculaire (53). La deuxième montre une diminution significative du test de
Schirmer chez les volontaires ayant reçu des instillations BAC comparativement aux
volontaires n‟ayant pas reçu de traitement (54).
Plusieurs enquêtes comparatives observationnelles ont été menées récemment afin toujours
d‟évaluer le caractère toxique et délétère de la présence du conservateur dans les collyres anti-
glaucomateux. Yalvac et al. trouvèrent que le test de Schirmer et la mesure du Break up time
étaient significativement diminués dans le groupe traité versus le groupe placebo (55).

19
D‟autres auteurs montrèrent, en utilisant l‟analyse In vivo au microscope confocal de la
surface oculaire, une réduction de la densité en cellules épithéliales superficielles et une
augmentation de la densité des cellules basales avec activation kératocytaire stromale dans les
groupes traités par des collyres conservés quelle que soit leur classe (bêtabloquants,
prostaglandines ou association fixe de ces deux classes) en comparaison du groupe placebo
(56). Il a également été mis en évidence une diminution de la sensibilité cornéenne, basée sur
l‟étude de l‟esthésiométrie cornéenne, dans les tous groupes traités par les collyres conservés
par le BAC. En effet, il est connu que le BAC peut entraîner des manifestations de
dénervation dans d‟autres systèmes que le système oculaire, tels que le plexus soléaire gastro-
intestinal ou le tractus génito-urinaire (57). Ces propriétés toxiques du BAC sur les nerfs
cornéens, avec diminution de la sensibilité cornéenne, peuvent participer au « relatif » bon
confort qu‟éprouvent bon nombre de patients traités au long cours par des collyres contenant
du BAC. Mais en rapport avec cette notion de baisse de la sensibilité cornéenne, cela poserait
plutôt le problème de l‟aspect faussement bon de la tolérance de ces collyres.
Certains travaux ont pris le parti d‟analyser les conséquences du changement de traitement
pris par le patient afin de tenter de mettre en évidence une amélioration des symptômes
lorsque le BAC était exclu des formulations des collyres. Ainsi, Pisella et al. ont observé les
effets cliniques de la permutation du traitement de 349 patients qui présentaient des signes et
symptômes d‟irritation de la surface oculaire (tels que l‟allergie oculaire, la blépharite ou le
syndrome sec oculaire) et qui étaient traités par des collyres conservés. Après le quatrième
mois, une importante et significative diminution de tous les signes et symptômes fonctionnels
a été retrouvée (21). Une plus large étude à l‟échelon européen a confirmé ces résultats (22).
D‟autres travaux montrèrent, après le troisième mois de changement de traitement pour des
collyres anti-glaucomateux sans BAC, une nette diminution des symptômes d‟œil sec, de
sensation de corps étranger, d‟hyperhémie conjonctivale et de kératite ponctuée superficielle
(58). Campagna et al. ont corroboré ces données cliniques avec l‟étude à l‟échelon cellulaire
des conséquences de ce changement de traitement. Ils rapportent une diminution significative
des processus inflammatoires au niveau de la conjonctive par l‟analyse d‟empreintes
conjonctivales deux mois après le « switch », en observant principalement une augmentation
du nombre de cellules à mucus et de cellules épithéliales (59). Ces modifications à l‟échelon
cellulaire étaient corrélées à une amélioration clinque avec diminution de tout le cortège de
symptômes accompagnant l‟irritation de la surface oculaire.

20
3. Chez l‟homme
En théorie, le meilleur moyen d‟explorer la surface oculaire chez l‟homme est la biopsie
conjonctivale qui permet d‟obtenir des cellules stromales, épithéliales, inflammatoires et des
cellules à mucus ex vivo, accessibles en microscopie conventionnelle, confocale ou
électronique autorisant l‟utilisation de techniques d‟immunohistochimie ou de biologie
moléculaire. Elle peut ainsi permettre l‟identification et le comptage de cellules
inflammatoires, l‟analyse de marqueurs cellulaires membranaires, de molécules intra
cytoplasmiques ou de composants de la matrice extracellulaire. La biopsie est facilement
réalisable sous anesthésie locale, au niveau de la conjonctive bulbaire adjacente au limbe sans
qu‟aucune suture ne soit nécessaire. Malgré leur simplicité de réalisation apparente, les
biopsies conjonctivales sont des examens invasifs et doivent être réservées à des fins
diagnostiques ou thérapeutiques lorsqu‟un bénéfice est attendu pour le patient. La biopsie
conjonctivale est l'outil de choix pour le diagnostic de certaines maladies inflammatoires,
infectieuses auto-immunes dépendantes des anticorps et du complément au niveau de la
membrane basale conjonctivale. La réalisation de techniques d'immunomarquage sur les
biopsies conjonctivales permet par exemple de détecter des complexes immuns formés in situ
dans la conjonctive (60) au cours des maladies bulleuses auto-immunes comme la
pemphigoïde cicatricielle, l'épidermolyse huileuse acquise ou la pemphigoïde bulleuse, et de
différencier ces maladies des pseudopemphigoïdes iatrogènes, liées par exemple au BAC. Des
biopsies conjonctivales pratiquées chez des patients glaucomateux traités au long cours, lors
de leur intervention chirurgicale, ont également apporté beaucoup de renseignements sur l'état
inflammatoire des yeux glaucomateux multi traités (61,62,63,64).
Plus simples à réaliser et plus utiles pour le diagnostic des atteintes allergiques, les examens
de cytologie permettent l'examen des cellules conjonctivales desquamées et surtout
l'identification de cellules inflammatoires dont certaines sont parfois évocatrices du
diagnostic. Ces examens cytologiques sont appelés « brossages conjonctivaux » et les cellules
recueillies peuvent être analysées à l‟aide de techniques d‟immunomarquage et de cytométrie
de flux. Après instillation de collyres anesthésiants, les cellules conjonctivales sont récoltées
par grattage au niveau de la conjonctive bulbaire à l‟aide d‟une brosse puis remise en
suspension dans un tampon de PBS (« phosphate buffer saline ») (65). La présence
d'éosinophiles et de basophiles suggère ainsi une réaction allergique, mais n'est pas
spécifique, puisque ces cellules, bien qu'en nombre plus faible peuvent aussi être retrouvées

21
dans des syndromes purement toxiques (66). Des aspects de kératinisation et de
dégénérescence épithéliale, accompagnées d'infiltrats de lymphocytes et de polynucléaires
neutrophiles, sont plus évocateurs de causes toxiques, mais toutes les associations sont
possibles (67,68).
Les premières empreintes conjonctivales ont été réalisées vers la fin des années 1970
notamment dans le cadre de la sécheresse oculaire (69,70). L‟empreinte conjonctivale
représente un moyen simple, non traumatique, exsangue et indolore de recueil des cellules
épithéliales de la surface oculaire. Les renseignements obtenus diffèrent cependant de ceux
fournis par une biopsie conjonctivale car cette dernière permet également l‟étude du chorion
sous-jacent. Cependant, l‟empreinte conjonctivale présente deux avantages majeurs par
rapport aux brossages conjonctivaux ; d‟une part, elle permet de prélever un tapis cellulaire
homogène gardant son architecture et ses liaisons intercellulaires, et elle est pratiquement
indolore. L‟empreinte conjonctivale permet de réaliser un instantané photographique de
l‟épithélium conjonctival avec l‟identification des trois populations cellulaires qui le
composent : les cellules épithéliales, les cellules à mucus et les cellules immunocompétentes
représentées par les cellules de Langerhans. L‟étude de ces cellules par des colorations
histologiques standards a permis initialement d‟appréhender la souffrance conjonctivale dans
l‟œil sec dont la traduction principale est la diminution des cellules à mucus et la diminution
progressive du rapport nucléo-cytoplasmique des cellules épithéliales, jusqu‟à un état de
kératinisation étendue (71). D‟autre part, l‟utilisation de marqueurs d‟inflammation comme le
marqueur HLA-DR a permis de caractériser les cellules épithéliales positives pour de
l‟inflammation conjonctivale présente à un stade infraclinique dans certaines situations
pathologiques comme l‟œil sec ou la pathologie iatrogène (72,73). La technique de recueil
cellulaire par empreintes conjonctivales repose sur l‟application, après instillation d‟un
collyre anesthésiant, d‟un filtre opaque d‟acétate de cellulose (type MilliporeGSWP ou
Gellman Supor; diamètre 13 mm, pores de 0,2 µm) ou transparent (type Biopore),
préalablement coupé en deux à l‟aide d‟une pince, sur la conjonctive bulbaire supérieure ou la
conjonctive palpébrale, après avoir soulevé la paupière supérieure, en demandant au patient
de regarder vers le bas. Un simple contact de quelques secondes sans pression suffit. Le
devenir de cette membrane chargée de matériel cellulaire dépend alors de la technique
d‟analyse envisagée, la membrane pouvant être fixée dans du formol, de l‟éthanol ou du
paraformaldéhyde pour l‟utilisation de colorations cytologiques standards. L‟utilisation en
immunohistochimie nécessite un transfert des cellules sur support transparent, obtenu en
appliquant fermement la membrane sur une lame de verre. Récemment développée,

22
l‟utilisation de méthodes de quantification cellulaire, comme la cytométrie de flux, nécessite
le maintien des cellules recueillies sur leur support, en suspension liquide, avant technique
d‟extraction cellulaire. Les tubes sont alors conservés à 4°C et sont analysés dans la semaine
qui suit leur prélèvement. L‟extraction des cellules se fait manuellement par agitation pendant
20 minutes suivie d‟une centrifugation à 1600 tours/minutes pendant 5 minutes. Au total, les
cellules recueillies par empreintes conjonctivales peuvent être analysées de différentes
manières : soit en cytologie standard, qui consiste à colorer de façon classique les cellules
(avec l‟acide périodique Schiff, le bleu alcian, l‟hématoxyline, la coloration de Gill par
Papanicolaou ou le May-Grünwald-Giemsa) et à les examiner en microscopie
conventionnelle. Cela permet d‟analyser la taille, la forme des cellules épithéliales, leur
rapport nucléo-cytoplasmique, la densité des cellules à mucus ainsi que les modifications de
la chromatine (74, 75). Soit en immunofluorescence standard, permettant un marquage et un
examen direct des cellules, sans qu‟aucun transfert sur support ne soit nécessaire réduisant la
perte et la détérioration cellulaires (76). Cette technique s‟est beaucoup développée avec
l‟utilisation de la microscopie confocale, apportant des renseignements importants non plus
seulement sur la morphologie cellulaires mais également sur leur structure et leur
fonctionnement. Cependant, il n‟est pas possible, en immunofluorescence, de quantifier de
manière fiable et reproductible les différents marqueurs testés comme cela est possible avec la
cytométrie de flux (CMF). La CMF consiste à faire défiler en « file indienne » les cellules
isolées une par une devant un système d‟illumination. Le guidage des cellules en suspension
se fait à l‟aide d‟un flux laminaire assurant leur défilement. Le système de détection doit être
suffisamment performant pour permettre d‟effectuer l‟analyse d‟une cellule donnée dans un
temps très court, de l‟ordre de quelques millisecondes. L‟utilisation de la CMF pour l‟analyse
des cellules recueillies par empreintes conjonctivales a été développée dans les années 1990
(77). Cela nécessite l‟extraction manuelle des cellules préalablement fixées dans du
paraformaldéhyde (PFA). Les applications possibles de cette techniques sont, entre autres,
l‟étude de l‟apoptose avec des marqueurs tels que le pic SubG1, l‟Apo 2.7, ou la mise en
évidence et la quantification de marqueurs inflammatoires divers par les cellules épithéliales
tels que l‟expression de l‟antigène de classe II HLA-DR ou le marqueurs Fas (78). La dernière
façon d‟analyser les cellules d‟empreintes conjonctivales est d‟utiliser des techniques de
biologie moléculaire telle que la « reverse transcriptase polymerase chain reaction » (RT-
PCR) permettant d‟identifier certains marqueurs inflammatoires (Il-6, Il-8, HLA-DR, ICAM-
1…) entre autres.

23
4. Modèles cellulaires
De tous les moyens d‟étude de la surface oculaire que nous venons de décrire, la biopsie
conjonctivale est le meilleur car elle permet de recueillir tous les types cellulaires formant la
conjonctive (cellules stromales, épithéliales, inflammatoires et à mucus) sous un format « ex
vivo ». Cependant, le matériel humain étant difficile à obtenir et les prélèvements étant
invasifs, les brossages conjonctivaux et les empreintes conjonctivales ont été développés,
permettant de réaliser des études toxicologiques mais aussi immunologiques et
physiopathologiques de la surface oculaire. Malgré cela, le recueil de cellules conjonctivales
par ces deux méthodes étant très limité, les recherches sur la surface oculaire se sont tournées
vers l‟utilisation des cultures cellulaires, permettant d‟étudier de façon fiable et reproductible
l‟influence de divers métabolites, médiateurs inflammatoires ou thérapeutiques sur la
conjonctive ou la cornée In vitro.
a. Les cultures primaires
Le principe d‟une culture primaire de cellules conjonctivales consiste à prélever plusieurs
biopsies de 1 cm2 de conjonctive au niveau bulbaire chez des patients sains, puis de les
stocker dans un milieu spécifique. Les cellules conjonctivales de l‟explant seront dissociées
par incubation avec une protéase, puis recueillies, rincées et mises en culture dans des
flasques contenant un milieu spécifique changé tous les 2 ou 3 jours (79). ce type de culture
est un outils intéressant pour les études physiopathologiques de la surface oculaire en raison
de ses similitudes phénotypiques avec l‟épithélium conjonctival humain normal. Les
caractéristiques des cultures primaires de cellules conjonctivales sont répertoriées dans les
Tableaux 2 et 3. Cependant, bien que les cultures primaires se rapprochent
phénotypiquement parlant de la conjonctive normale, elles ne sont que peu utilisées en
pratique du fait de trois inconvénients majeurs : d‟une part ce type de biopsie nécessite la
réalisation d‟un large prélèvement conjonctival chez le patient, d‟autre part, la lourdeur des
techniques de culture et le nombre limité de passages cellulaires, et enfin l‟importante
variabilité de la qualité de la culture rendant aléatoire la probabilité d‟obtenir suffisamment de
cellules conjonctivales pour mettre en œuvre les expérimentations.

24
b. Les lignées cellulaires
Les lignées cellulaires sont très utilisées actuellement en toxicologie car peu coûteuses,
reproductibles, indépendantes de tout prélèvement oculaire avec une croissance cellulaire
rapide, fiable et stable dans le temps.
Toutes les lignées ne sont pas identiques et il est très important de bien connaître leurs
caractéristiques de croissance, leurs caractères phénotypiques et leurs propriétés
fonctionnelles pour pouvoir interpréter les résultats. Il faudra également tenir compte du
caractère plus ou moins indifférencié des cellules en lignées dépendant en partie du nombre
de passages cellulaires déjà effectués jusqu‟alors.
Certaines lignées cellulaires ont été immortalisées par transfection, telle que la lignée HC0597
(transformée par infection par un plasmide RSV-T codant pour l‟oncogène T de SV40, virus
simien) qui a déjà été utilisée pour l‟étude des effets de la thymosine bêta 4 sur
l‟inflammation oculaire et de l‟apoptose induite par le BAC, ou pour étudier les effets de TNF
et IFN sur la sécrétion de molécules chimiotactiques comme l‟IL-8 (80, 81).
Une autre lignée cellulaire conjonctivale, la lignée de WKD (Wong Kilbourne derivative) de
Chang, celle que nous avons utilisée pour nos travaux, a été issue de cellules conjonctivales
humaines, spontanément immortalisées. Cette lignée présente plusieurs caractéristiques
communes avec les cellules épithéliales conjonctivales comme la présence de desmosomes, le
développement de microvillosités, la présence du récepteur à l‟EGF et des marqueurs
muciniques (79). Cependant, elle présente également des différences morphologiques et
fonctionnelles en comparaison avec des cellules en culture primaires et des cellules
épithéliales de conjonctive (tableau 2 et 3). Par exemple, les cellules de Chang ont un aspect
plutôt fibroblastique, avec une expression élevée de vimentine sans expression de la
cytokératine 4 ni de la E-cadhérine, rendant compte d‟un aspect plus mésenchymateux
qu‟épithélial. Toutefois, les test de stimulations au TNF et à l‟IFN semblent les rendre
comparables aux cultures primaires et utilisables pour des études de toxicologie in vitro, mais
aussi pour des études fonctionnelles comme la production de cytokines en réponse à des stress
oxydant, d‟inflammation et d‟apoptose (82, 83).

25
Cultures
Primaires
Lignée de
Chang
Normal IFN gamma TNF alpha normal IFN gamma TNF alpha
HLA DR +/- - +/- -
CD40 ++ + =
CD44 ++ = +++ = =
ICAM-1 +++ +++
CD63 ++ ++ =
CD80 - - - - - -
CD86 - - - - - -
CD95/Fas + + =
E-Cadhérine + = = - - -
Pancytokératine +++ = = ++ = =
Cytokératine 4 ++ = = - - -
Vimentine ++ = = +++ =
Muc 1 ++ = + =
Muc 5 AC - NE NE - NE NE
Tableau 2 :
Effets de l’IFN et du TNF sur l’expression d’antigènes dans les cultures primaires
d’épithélium conjonctival ou sur les lignées de Chang *d‟après De Saint Jean M, et al. Exp Eye Res 2002
CD : Cluster de différenciation. HLA-DR : Human leukocyte antigen-D locus related protein. ICAM : Inter-
Cellular Adhesion Molecule. IFN Interferon. NE : non étudié. TNF : Tumor Necrosis Factor
c. Les modèles d‟épithéliums reconstitués en 3 dimensions
Il existe plusieurs types de modèles d‟épithéliums cornéens reconstitués en trois dimensions
(3D), le principal étant le modèle SkinEthic. Il est réalisé à partir de cellules épithéliales
cornéennes (lignées ou cultures primaires) qui, cultivées dans certaines conditions, peuvent se

26
stratifier et se différencier en des structures non kératinisées après ensemencement sur un
substrat spécifique.
L‟avantage des modèles 3D par rapport aux cultures cellulaires en deux dimensions est de
pouvoir reproduire, in vitro, les caractéristiques structurelles de la cornée. L‟effet du toxique
pourra être étudié sur toute l‟épaisseur de l‟épithélium. En revanche, cet outil est moins
accessible à des techniques nécessitant la dissociation des cellules comme pour la cytométrie
de flux.

27
IV. ETUDE DES INTERACTIONS CELLULAIRES DE CELLULES
CONJONCTIVALES EN LIGNEES AVEC DES LYMPHOCYTES ISSUS DU SANG
CIRCULANT (ANNEXE)
1. Introduction
Le BAC est le conservateur le plus couramment utilisé dans les collyres en ophtalmologie,
principalement pour le traitement de maladies oculaires chroniques telles que le glaucome, ou
l‟allergie oculaire. Au niveau oculaire, le renouvellement du BAC est très lent, et il a été
montré que des traces de BAC pouvaient être retrouvées jusqu‟à 7 jours après l‟instillation
d‟une seule goutte (84). La toxicité oculaire du BAC a été démontrée à plusieurs reprises par
de nombreux travaux de notre équipe et d‟autres auteurs, en utilisant des modèles in vivo et in
vitro (35, 85, 86, 87). Plusieurs études ont montré l‟implication du BAC dans de nombreuses
pathologies oculaires telles que l‟allergie, la fibrose, l‟inflammation et le syndrome sec
oculaire (88, 89, 90, 91, 92), ainsi que son rôle pro nécrotique, pro-apoptotique et pro-oxydant
sur les cellules conjonctivales in vitro et in vivo (93, 94, 95, 96). Cependant, la conjonctive
humaine n‟est pas seulement composée de cellules épithéliales conjonctivales, mais inclus
d‟autres types cellulaires tels qu‟une variété de cellules inflammatoires et de cellules à mucus,
en contact étroit avec les vaisseaux sanguins et donc les lymphocytes extra oculaires (97, 98,
99, 100). Tous ces types cellulaires participent au maintien physiologique de la surface
oculaire, et seraient impliqués directement dans la survenue d‟une pathologie inflammatoire
épithéliale de la surface oculaire (73, 82).
Pour évaluer la toxicité du BAC ou d‟autres xénobiotiques sur l‟épithélium conjonctival, la
méthode la plus courante est basée sur l‟utilisation de modèles in vitro offrant la possibilité
d‟étudier l‟influence de métabolites, médiateurs ou médicaments, tout cela se faisant dans un
environnement contrôlé, stable et fiable. Actuellement sont disponibles deux principales
lignées de cellules épithéliales conjonctivales, continues, non transfectées, qui sont les
cellules de « Wong-Kilbourne derivative of Chang » (WKD) (101) et les cellules « IOBA-
NHC » (102). La lignée de cellules WKD a été isolée à partir d‟un épithélium conjonctival
humain normal, puis spontanément immortalisée (103). Mais cette lignée diffère de
l‟épithélium conjonctival normal et d‟une culture primaire de cellules épithéliales
conjonctivales par ces caractéristiques phénotypiques ainsi que ses réponses à certaines
cytokines pro inflammatoire (79) et, généralement, la réserve émise pour valider l‟utilisation
d‟une lignée cellulaire comme un équivalent de tissu humain est la ressemblance

28
phénotypique de cette lignée avec son homologue précurseur non transformé. Les cellules
WKD sont ainsi largement utilisées comme modèle in vitro unicellulaire fiable d‟étude de la
surface oculaire, et ont déjà permis à de nombreux travaux de prouver les effets toxiques du
BAC (78, 104), mais toutes ces études in vitro se sont basées sur l‟utilisation des cellules
WKD prises isolement, sans tenir compte de la composante inflammatoire qui existe en
permanence au niveau de l‟épithélium conjonctival normal. Des interactions cellulaires in
vitro entre des cellules épithéliales et des cellules inflammatoires ont déjà été analysées dans
des modèles de peau ou de poumon (105, 106, 107, 108). De tels travaux de coculture ont été
menés également au niveau conjonctival en mettant en interaction une culture primaire de
cellules épithéliales de conjonctive avec des lymphocytes issus du sang circulant (Peripheral
Blood Lymphocytes, PBL), retrouvant des modifications de caractéristiques à la fois sur les
lymphocytes et sur les cellules épithéliales (109). En effet, les auteurs ont montré
principalement que le contact direct, ou par le biais de médiateurs solubles, entre ces deux
types cellulaires permettait l‟expression du marqueur HML-1/CD103 à la surface des
lymphocytes après sept jours de coculture, rendant compte de leurs interactions in vitro et des
conséquences moléculaires qui en découlent. Mais, en comparaison avec une culture primaire,
une lignée cellulaire telles que les cellules WKD offre une meilleure reproductibilité, une
acquisition plus rapide des résultats et est indépendante de la disponibilité ou non des cellules
épithéliales issues des biopsies conjonctivales. C‟est pourquoi les cellules de WKD ont été
choisies à de nombreuses reprises par le passé pour effectuer des études de toxicologie sur la
surface oculaire (78, 94, 104). Une nouvelle étape serait donc d‟associer cette lignée cellulaire
avec des cellules inflammatoires et d‟étudier ainsi leurs interactions. Ainsi, pour mieux
répondre aux travaux de Gomes et al. (109), nous avons commencé par associer la lignée de
cellules WKD, déjà validée sur le plan toxicologique, avec des PBL. Par conséquent,
l‟objectif de cette étude fut d‟évaluer les interactions entre les cellules WKD et les PBL en
vue de développer et de valider in vitro le potentiel du nouveau modèle dans le but d‟étudier
les pathologies de la surface oculaire et d‟avancer plus loin dans la recherche toxicologique.
2. Matériels et Méthodes
a. La lignée de cellules conjonctivales :
Les cellules conjonctivales de WKD (clone 1 to 5c-4 American Type Culture Collection
[ATCC, Manassas, VA]-certified cell line [CCL], 20.2) ont été cultivées selon un protocole

29
standardisé, avec des conditions normalisées (atmosphère humidifiée à 37 °C avec 5% de
CO2), en flasque de 75 cm2, dans un milieu de culture “Dulbecco” minimum essentiel à leur
croissance (Dulbecco‟s minimum essential medium, DMEM) supplémenté en serum de veau
foetal à 10% (fetal bovine serum, FBS), en glutamine à 1%, avec 50 UI/ml de pénicilline et 50
UI/ml de streptomycine.
Les cellules étaient repiquées une à deux fois par semaine par trypsination. Le développement
normal des cellules était contrôlé quotidiennement avec un microscope inversé a contraste de
phase (Leica, DMIRB). Ainsi, après trypsination à confluence et comptage, pour les test de
cytométrie de flux, les cellules étaient incubées dans des plaques 6 puits (Corning, Schiphol-
Rijk, the Netherlands) et sur des lamelles (Lab-Tek™ II chambered coverglass; Nunc
International, Naperville, IL) pour l‟analyse en immunofluorescence standard et en
microscopie confocale. Les cultures cellulaires étaient maintenues en incubation à 37°C
pendant 24 heures puis, à sub-confluence (une fois que 70% de la surface du puit de culture
était couverte), étaient exposées à différentes concentration de BAC.
b. Les lymphocytes issus du sang périphérique :
En partenariat avec l‟agence française du sang (AFS), les lymphocytes étaient obtenus à partir
du sang périphérique de donneurs sanguin anonymes et volontaires sains. L‟AFS réalisait
l‟extraction plaquettaire de l‟échantillon de sang de ces donneurs puis nous fournissait les
reliquats de cette procédure, à savoir le culot de cytaphérèse, dans l‟objectif que nous
séparions les lymphocytes périphériques des globules rouges. Ces deux types cellulaires,
contenus dans les culots de cytaphérèse étaient séparés par centrifugation à 400g pendant 30
minutes selon la technique du gradient de Ficoll (Sigma Chemical Inc, St Louis, MO, USA)
permettant d‟isoler les cellules mononuclées. Les PBL étaient lavés dans un tampon de
solution phosphate salée (phosphate buffer saline, PBS), puis centrifugés à nouveau à 300g
pendant 15 minutes, lavés dans du PBS, puis centrifugés une dernière fois à 200g pendant 10
minutes. Les PBL ainsi isolés étaient maintenus en suspension dans un milieu RPMI 1640
(Roswell Park Memorial Institute medium) supplémenté en FBS à 10% puis mis en culture
selon des conditions standard (atmosphère humidifié avec 5% de CO2 et à 37 °C) pendant 12
heures dans des flasques de 75 cm2. Le surnageant était prélevé et lavé deux fois.
c. Les traitements cellulaires :

30
Pour les expériences d‟activation lymphocytaire in vitro, les lymphocytes étaient incubés dans
leur milieu standard de RPMI 1640 avec 2.5 ng/ml de phorbol 12-myristate 13-acetate (PMA,
Sigma, St Louis, MO, USA) et 250 ng/ml de Ionomycine (Sigma) à 37°C. Après stimulation,
les cellules étaient lavées trois fois dans du RPMI 1640. Les cellules de WKD étaient traitées
avec de la Trichostatine A (Trichostatin A, Ready Made Solution 5mM in DMSO [0.2μm-
filtered], from Streptomyces sp., Sigma
) à 0.33M pendant 24 heures sous conditions
standards, dans un milieu de culture “Dulbecco” minimum essentiel à leur croissance
(Dulbecco‟s minimum essential medium, DMEM) supplémenté en serum de veau feotal à
10% (fetal bovine serum, FBS), en glutamine à 1%, avec 50 UI/ml de pénicilline et 50 UI/ml
de streptomycine, avec pour objectif de faire exprimer la E-Cadhérine à la surface des WKD
(110).
En accord avec de précédents travaux sur cette même lignée cellulaire (94), les cellules de
WKD, une fois arrivées à sub-confluence (surface de plaque couverte à 70%) étaient alors
exposées à différentes concentrations de BAC (10−2
%, 5.10−3
%, 10−3
%, 10−4
%, et 10−5
%) ou
de PBS pendant 15 minutes, suivi de 24 heures de récupération en milieu complet, afin
d‟évaluer la viabilité cellulaire et les effets toxiques du BAC sur ce modèle. Les
concentrations de BAC choisies sont équivalentes ou inférieures à celles rencontrées dans la
plupart des collyres disponibles actuellement sur le marché. Les concentrations de 10−1
% et
supérieures n‟ont pas été testé car de toxicité trop importante sur les cellules conjonctivales in
vitro entraînant une lyse cellulaire immédiatement après l‟application du traitement. Le temps
d‟incubation du traitement appliqué aux cellules de WKD (et lors de leur coculture avec les
PBL) était de 15 minutes suivi de 24 heures de récupération. Cette période de 24 heures de
récupération a été testé en vue de s‟approcher au plus près des conditions cliniques au cours
desquelles la conjonctive peut être protégée généralement après l‟instillation de la goutte de
collyre.
d. La coculture :
1 ml de PBL, à la concentration de 106
cellules/ml, était mis en coculture avec des cellules de
WKD arrivées à confluence. Les PBL étaient disposés, soit en contact direct avec les cellules
épithéliales, soit séparés des cellules de WKD par le biais d‟une membrane poreuse appelée
Transwell (Cell culture inserts, translucent PET membrane, 0.4m, BD Falcon, BD
Biosciences), qui permettait d‟éviter tous contacts direct entre les types cellulaires, mais
laissait librement la place aux échanges entre les deux milieux de culture. Après 1 ou 7 jours

31
de coculture, les lymphocytes étaient collectés et remis en suspension dans 2 ml de milieu
standard (RPMI 1640 supplémenté avec du FBS à 10%), puis analysés en cytométrie de flux.
e. La Cytométrie de flux :
Le marqueur d‟inflammation HLA DR (Human leucocyte Antigen Class II), le récepteur lié à
l‟apoptose Fas (APO-1, CD95), le marqueur de co-stimulation CD86, l‟antigène HML-1
(Human Leucocyte Antigen-1) et la molécule d‟adhésion E-cadhérine ont été testé sur les
cellules conjonctivales de WKD. Toutes ces mesures se sont déroulées en utilisant le
cytomètre Cytomics FC500-CXP (Beckman Coulter, Miami, FL), équipé d‟un laser Argon
émettant à 488 nm. L‟anticorps anti-IgG de souris conjugué au FITC/PC5 a été utilisé comme
contrôle négatif pour estimer la fluorescence de fond non spécifique.
La dégradation de l‟ADN cellulaire a été mesurée à l‟aide de la détermination du pic sub-G1
en cytométrie de flux par le biais d‟un histogramme avec pic en avant du pic sub-G1, sur l‟axe
des intensités de fluorescence, qui caractérise le processus d‟apoptose tardive. Après avoir été
mis en coculture, les PBL et les cellules de WKD, préalablement traitées par le BAC ou le
PBS, ont été récupérés en utilisant de l‟acide ethylenediaminetetraacetique (EDTA) à la
concentration de 1mM, puis mis en suspension dans 1ml de PBS et, enfin, fixés dans du
paraformaldehyde (PFA) à 0.5% pour une conservation à 4 °C pendant 24 heures. Finalement,
les échantillons sont lavés dans du PBS froid et les cellules perméabilisées grâce à la saponine
à la concentration de 0.1%, colorées avec 50μg/ml de iodure de propidium (PI, Interchim,
Montluçon, France), et analysées au cytomètre.
f. La microtitration cytofluorimétrique adaptée aux microplaques :
Pour la réalisation de nos tests de cytotoxicité par cytofluorimétrie en microplaques, nous
avons utilisé un spectrofluorimètre Safire (Tecan Instruments, Lyon, France), composé de
deux monochromateurs pouvant chacun balayer une bande spectrale de 250 À 1000 nm. Ce
spectrophotomètre permet d‟enregistrer des spectres de fluorescence en excitation et en
émission. Au total, la microtitration cytofluorimétrique est une technique sensible (pg-fg/ml),
reproductible et spécifique permettant la mise au point de méthodes standardisées d‟analyse
de la cytotoxicité et du stress oxydant, préconisée notamment lors d‟études multicentriques
pour l‟évaluation de méthodes alternatives en toxicologie cellulaire (111, 112, 113).

32
Le colorant Hoechst33342 a été utilisé pour évaluer l‟apoptose de cellules de WKD, et le
colorant rouge neutre (Neutral Red, NR, Fluka, Ronkonkoma, NY) a été utilisé pour évaluer
leur intégrité membranaire, paramètre de très près corrélé à la viabilité cellulaire. Dans toutes
ces expériences, le PBS a été manipulé comme contrôle. Les résultats de microtitration
cytofluorimétrique adaptée aux microplaques ont été exprimés en unité de fluorescence
comme un pourcentage par rapport au contrôle du PBS. Les résultats de la coloration au
Hoechst33342 ont été rapportés aux résultats de la coloration par le rouge neutre afin de
corréler ces derniers à la viabilité cellulaire.
g. Analyse de la morphologie cellulaire :
Nous avons réalisé des expériences d‟immunofluorescence standard en utilisant différentes
substances fluorescentes telles que la phalloïdine (F-actin detection, 200U/ml; Alexa Fluor
488 Phalloidin, Sigma, St Louis, MO, USA), le PKH67 et le PKH26, deux colorants
lipophiles émettant dans le vert et le rouge respectivement. Ces colorations ont été analysées
au microscope confocal à épifluorescence (E800, PCM 2000; Nikon, Tokyo, Japan) dans le
but d‟évaluer les modifications morphologiques des deux types cellulaires étudiés lors de la
coculture. Après 30 minutes d‟incubation avec la phalloïdine, les cellules sont lavées dans du
PBS avant d‟être colorées au iodure de propidium puis examinées directement au microscope.
Les cellules de WKD ont été colorées avec le PKH67 (Green Fluorescent Cell Linker Kit,
Sigma), et les PBL avec le PKH26 (Red Fluorescent Cell Linker Kit, Sigma). Pour la
coloration avec ces substances, les cellules de WKD et les PBL (20 millions de cellules pour
chaque type) ont été lavés soit dans du milieu DMEM supplémenté en glutamine à 1% avec
50 UI/ml de pénicilline et 50 UI/ml de streptomycine sans FBS pour les cellules de WKD,
soit dans un milieu de RPMI 1640 pour les PBL, à 400g pendant 5 minutes puis mis en
contact avec le colorant PKH à la concentration de 4M pendant 4 minutes. Pour finir, les
cellules colorées ont été lavées 3 fois dans le PBS puis remises en suspension dans leur propre
milieu de culture complet.
h. La microscopie électronique à balayage :
La microscopie électronique à balayage a été spécifiquement utilisée pour évaluer les
interactions cellulaires entre les cellules de WKD et les PBL. Pour ce faire, les cellules ont
d‟abord été fixées dans le glutaraldehyde à 2.5% (Electron Microscopy Science, Hatfield,

33
USA) pendant 45 minutes, puis dans l‟osmium OsO4 réduit à la concentration de 2%
(Electron Microscopy Science, Hatfield, USA) pendant 45 minutes également. Après ces deux
étapes de fixation, les cellules ont été déshydratées dans l‟éthanol, nettoyées à l‟oxyde de
propylène et saturées par l‟Hexamethyldisilane (Sigma). Puis, une fois passé le dernier
processus de saturation avec le gold-palladium à l‟aide du saturateur JEOL FINE COAT ion
sputter JFC-1100, les cellules ont été examinées avec le microscope électronique à balayage
JEOL 35CF (Hitachi
, Japan).
i. Les réactifs :
Le iodure de propidium, le PBS, les colorants PKH26 et PKH67 ont été obtenus auprès de
Sigma (St Louis, MO, USA).
L‟anticorps monoclonal de souris anti-CD103 humain (Mucosa Lymphocyte Antigen, clone
Ber-ACT8, FITC-conjugated), l‟anticorps de chèvre conjugué au FITC anti-souris et
l‟anticorps de lapin conjugué au phycoérythrine (PE) anti-souris ont été fournis par DAKO
(Glostrup, Denmark).
Les anticorps monoclonaux de souris anti-HLA-DR humain (Clone Immu-357, FITC-
conjugated), anti-CD86 (Clone HA5.2B7, PE-conjugated) et anti-CD95 (Clone 7C11, PE-
conjugated) ont été fournis par la société Coulter (Miami, FL, USA).
Les anticorps de contrôle (IgG1 de souris purifié et conjugué au FITC) ont été acquis auprès
d‟Immunotech (Marseille, France).
La phalloïdine (Alexa Fluor 488-conjugated) a été obtenu auprès de la société Molecular
Probes (Leiden, The Netherlands).
Tous les réactifs ont été utilisés selon les recommandations du fournisseur.
3. Résultats
a. Les interactions cellulaires :
Des échanges de membranes plasmiques entre les deux types cellulaires ont été observés
après un jour de coculture entre les cellules de WKD et les PBL en contact direct (Figure 2).
Cette intégration de membrane plasmique lymphocytaire au sein des cellules épithéliales de
WKD débute rapidement lors de la mise en contact des deux types cellulaires. Un pic a été
obtenu après 90 minutes de coculture (figure 3). Ces contacts rapprochés relevés entre les

34
cellules épithéliales et les lymphocytes, ressemblant à une fusion de membrane cellulaire, ont
également été observés en microscopie électronique à balayage (Figure 4), et ce phénomène
pourrait s‟apparenter à une forme de pinocytose. Les PBL, lors de la coculture avec les
cellules épithéliales, se sont disposés entre les tapis de cellules de WKD (Figure 5) en
montrant, après coloration avec la phalloïdine et analyse au microscope à fluorescence, la
formation d‟expansions membranaires cytoplasmiques (Figure 6).
b. L‟apoptose :
Les résultats de mort cellulaire ont été exprimés en pourcentage de survenue de l‟évènement
sub-G1 rapporté au nombre total de la population cellulaire. Après un jour de coculture,
l‟apoptose des cellules WKD cultivées en coculture avec des PBL non activés ne variait pas.
D‟un autre côté, comme attendu, l‟apoptose des cellules de WKD a augmenté lorsqu‟elles
étaient en contact avec des PBL activés par le PMA et l‟Ionomycine. Ces résultats de
cytotoxicité de la présence des PBL activés sur les cellules de WKD, après un jour de
coculture, étaient similaires que la coculture soient réalisée en contact direct ou que les deux
types cellulaires soient séparés l‟un de l‟autre lors de la coculture par une membrane poreuse
(appelée Transwell). Ensuite, une altération du contenu en ADN cellulaire des cellules de
WKD, préalablement traitées pendant 15 minutes avec du BAC à la concentration de 5.10-3
%,
suivies de 24 heures de récupération, a été observée lorsque cette période de récupération a
été accompagnée par la présence dans le milieu de culture de PBL non activés (Figure 7)
Cette potentialisation de l‟effet cytotoxique de la présence des PBL non activés n‟a pas
seulement été notée lorsque la coculture était en contact direct mais également lorsque les
deux types cellulaires étaient séparés par un Transwell. La concentration de BAC à 5.10-3
%
que nous avons utilisée, telle qu‟elle a déjà été décrite (114), correspond à la concentration
seuil de BAC induisant une toxicité significative sur les cellules de WKD. En effet, plus la
concentration de BAC augmente, plus l‟intégrité des membranes cellulaires des cellules de
WKD diminue. La cytotoxicité la plus importante était observée pour les concentrations de
BAC les plus élevées, supérieures à 5.10-3
%. Et cette concentration seuil de BAC est la
concentration inférieure ou égale à celle retrouvée dans la grande majorité des collyres utilisés
en ophtalmologie.
c. Modifications des cellules épithéliales :

35
Que ce soit de façon constitutive ou après coculture avec des PBL, nous n‟avons jamais pu
retrouver l‟expression membranaire de la E-Cadhérine à la surface des cellules de WKD,
après analyses faites en cytométrie de flux et en immunofluorescence standard. Cependant, le
traitement des cellules de WKD par la Trichostatine A (TSA) a induit une faible expression
membranaire de l‟E-Cadhérine, détectée en cytométrie de flux (Figure 8) et en
immunofluorescence standard (Figure 9).
Le marqueur d‟inflammation HLA-DR a vu son expression augmenter sur les cellules de
WKD après 1 jour de coculture en contact direct avec des PBL non activés, ainsi qu‟après 7
jours de coculture sans contact direct (cellules séparées par un Transwell) avec des PBL
activés (Figure 10).
Le marqueur d‟apoptose CD95/Fas ainsi que le marqueur de costimulation CD86 ont été
exprimés sur les cellules de WKD après 1 jour de coculture en contact direct avec des PBL
activés. De plus, il a été retrouvé un aspect spiculé des cellules de WKD, compatible avec une
souffrance cellulaire, augmentant avec le temps de la coculture, sur un marquage à la
phalloïdine et une coloration au iodure de propidium.
d. Modifications des cellules lymphocytaires :
Dans notre étude, nous n‟avons pas pu mettre en évidence d‟expression du marqueur
CD103/HML-1 après coculture entre les PBL et les cellules de WKD. Cependant, la
réalisation d‟une coculture entre des cellules de WKD prétraitées par la TSA avec les PBL a
montré une expression modérée mais significative du marqueur CD103 à la surface des PBL
(Figure 11).
e. Test toxicologique sur le modèle :
L‟apoptose des cellules de WKD traitées 15 minutes par le BAC augmente avec la
concentration du BAC, échelonnée de 5.10-4
% à 5.10-2
%, sans rapport avec la présence de
PBL dans le milieu de culture pendant les 24 heures de récupération post-traitement.
Cependant, pour les concentrations de BAC les plus faibles (inférieures à 10-2
%), nous avons
pu constater un effet potentialisant sur l‟apoptose des cellules de WKD de la présence des
PBL en contact direct dans le milieu de récupération, corroborant les résultats trouvés lors des
tests d‟apoptose au sub-G1. A l‟inverse, nous avons noté que pour les concentrations de BAC

36
les plus élevées (supérieures à 5.10-3
%), la présence des PBL en contact direct dans le milieu
de récupération induisait une diminution de l‟apoptose de cellules de WKD (Figure 12).

37
Figure 2.
Analyse de la morphologie cellulaire et des interactions cellulaires au cours d‟une coculture
entre des cellules de WKD et des PBL. Les cellules de WKD sont colorées en vert par le
PKH67 et les PBL sont colorés en rouge par le PKH26. Les photos A et B représentent la
même zone photographiée au microscope à fluorescence, soit en utilisant un filtre vert soit en
utilisant un filtre rouge. Les cellules de WKD ont été cultivées en coculture avec des PBL en
contact direct pendant 24 heures. Des échanges de membranes plasmiques entre les deux
types cellulaires ont été observés après un jour de coculture exclusivement si les cellules de
WKD étaient en contact direct avec les lymphocytes. Nous pouvons noter ici que les cellules
de WKD apparaissent à la fois vertes et rouge, signifiant qu‟un transfert de colorant entre les
deux types cellulaires s‟est produit. Les expériences contrôles (photos non fournies) faites
avec soit les cellules de WKD cultivées seules soit avec les cellules de WKD cultivées en
coculture avec des PBL mais séparées par une membrane PET translucide présentant des
pores de 0.4m de diamètre (pouvant laisser passer les colorant PKH), montre des cellules de
WKD restant exclusivement verte, sans aucun transfert de colorant rouge par les PBL. Ici, sur
les photos, l‟aspect de coloration jaune est lié à une surexposition à la fluorescence du
microscope.
A B
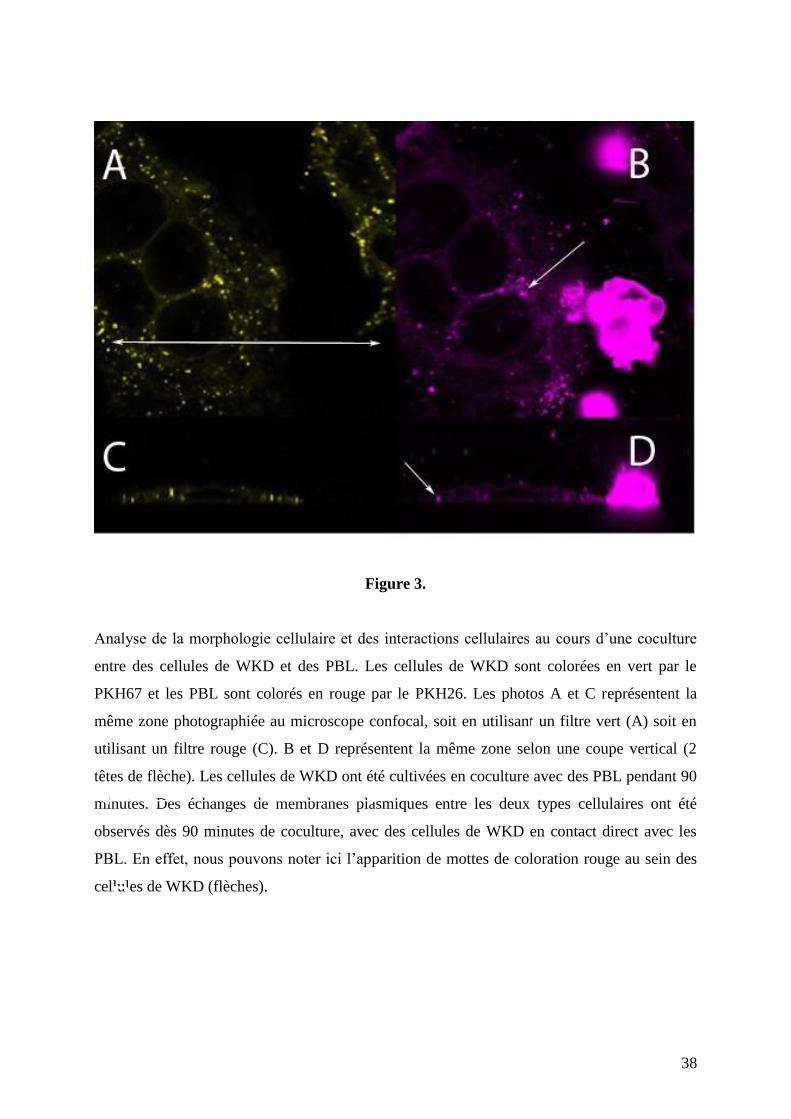
38
Figure 3.
Analyse de la morphologie cellulaire et des interactions cellulaires au cours d‟une coculture
entre des cellules de WKD et des PBL. Les cellules de WKD sont colorées en vert par le
PKH67 et les PBL sont colorés en rouge par le PKH26. Les photos A et C représentent la
même zone photographiée au microscope confocal, soit en utilisant un filtre vert (A) soit en
utilisant un filtre rouge (C). B et D représentent la même zone selon une coupe vertical (2
têtes de flèche). Les cellules de WKD ont été cultivées en coculture avec des PBL pendant 90
minutes. Des échanges de membranes plasmiques entre les deux types cellulaires ont été
observés dès 90 minutes de coculture, avec des cellules de WKD en contact direct avec les
PBL. En effet, nous pouvons noter ici l‟apparition de mottes de coloration rouge au sein des
cellules de WKD (flèches).
A
B
C
D

39
Figure 4.
Analyse en microscopie électronique à balayage de la morphologie cellulaire et des
interactions cellulaires au cours d‟une coculture entre des cellules de WKD et des PBL. Nous
pouvons noter ici les connections rapprochées entre les PBL (petite flèche) et les cellules de
WKD, ressemblant à une fusion de membranes cellulaires (grandes flèches).
Figure 5.
Analyse de la morphologie cellulaire et des interactions cellulaires au cours d‟une coculture
entre des cellules de WKD et des PBL. Les cellules de WKD sont colorées en vert par le
PKH67 et les PBL sont colorés en rouge par le PKH26. Les photos A, B et C représentent la
même zone photographiée au microscope confocal, soit en utilisant un filtre vert (A) soit en
utilisant un filtre rouge (C) soit en utilisant la fonction de « fusion » des deux images
précédentes. Les cellules de WKD ont été cultivées en coculture avec des PBL en contact
direct pendant 24 heures. Nous pouvons noter ici que les PBL prennent place entre les cellules
de WKD, dans le but probable de former des connections rapprochées avec ces dernières.

40
Figure 6.
Analyses au microscope à fluorescence de la morphologie cellulaire après coloration à la
phalloïdine des cellules de WKD et des PBL. Les deux types cellulaires ont été cultivé
ensemble en coculture pendant 24 heures. Le cytosquelette (F-actine) est coloré en vert par la
phalloïdine. Nous pouvons noter ici que les PBL développent des expansions membranaires
cytoplasmiques, en connections rapprochées avec les cellules de WKD.

41
Figure 7.
Analyse de l‟altération du contenu en ADN des cellules de WKD. L‟altération de l‟ADN
cellulaire a été mesurée par l‟étude en cytométrie de flux du pic sub-G1 sur les cellules de
WKD. Successivement, les cellules de WKD ont été cultivées seules dans du milieu standard
complet, puis en coculture pendant 24 heures avec des PBL séparées par un Transwell, puis
avec des PBL en coculture en contact direct. Ensuite, les mêmes expériences furent réalisées
en traitant au préalable les cellules de WKD par du BAC à la concentration de 5.10-3
%
pendant 15 minutes suivi de 24 heures de récupération seules, ou en présence sans contact de
PBL, ou en présence en contact direct de PBL. Nous pouvons noter ici la potentialisation de
l‟effet toxique des PBL en contact direct sur les cellules de WKD (flèche, p<0.001).
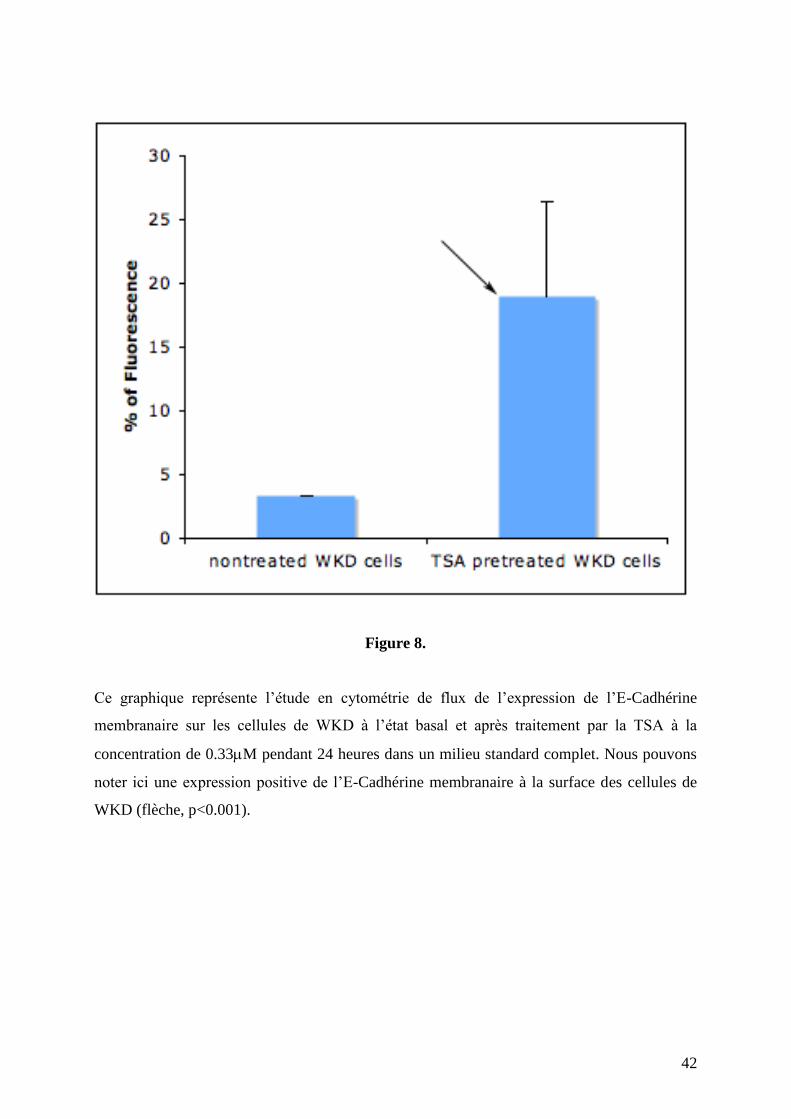
42
Figure 8.
Ce graphique représente l‟étude en cytométrie de flux de l‟expression de l‟E-Cadhérine
membranaire sur les cellules de WKD à l‟état basal et après traitement par la TSA à la
concentration de 0.33M pendant 24 heures dans un milieu standard complet. Nous pouvons
noter ici une expression positive de l‟E-Cadhérine membranaire à la surface des cellules de
WKD (flèche, p<0.001).

43
Figure 9.
Analyse en immunomarquage au microscope confocal de l‟expression d‟E-Cadhérine à la
surface des cellules de WKD. Les noyaux ont été contre colorés au DAPI. Sur la photo de
gauche, les cellules de WKD sont à leur état basal et sur la photo de droite, les cellules de
WKD ont été traitées pendant 24 heures avec la TSA. Nous pouvons noter ici une expression
positive d‟E-Cadhérine après traitement par le TSA (flèche).

44
Figure 10.
Ce graphique représente l‟étude en cytométrie de flux de l‟expression du marqueur
d‟inflammation HLA-DR sur la membrane des cellules de WKD à l‟état basal et après 1 ou 7
jours de coculture, en contact direct ou séparés par un Transwell, avec des PBL, activés par le
PMA et l‟Ionomycine (B) ou non (A). Les moyennes d‟intensité de fluorescence ont été
relevées et les résultats ont été exprimés en pourcentage par rapport au contrôle. Les PBL
activés ont été incubés avec 2.5 ng/ml de PMA et 250 ng/ml of Ionomycine à 37°C pendant 2
heures. Nous pouvons noter ici une expression positive du marqueur HLA-DR sur les cellules
de WKD après 24 heures de coculture en contact direct avec des PBL non activés, et après 7
jours de coculture sans contact direct avec des PBL activés (flèche, p<0.03).
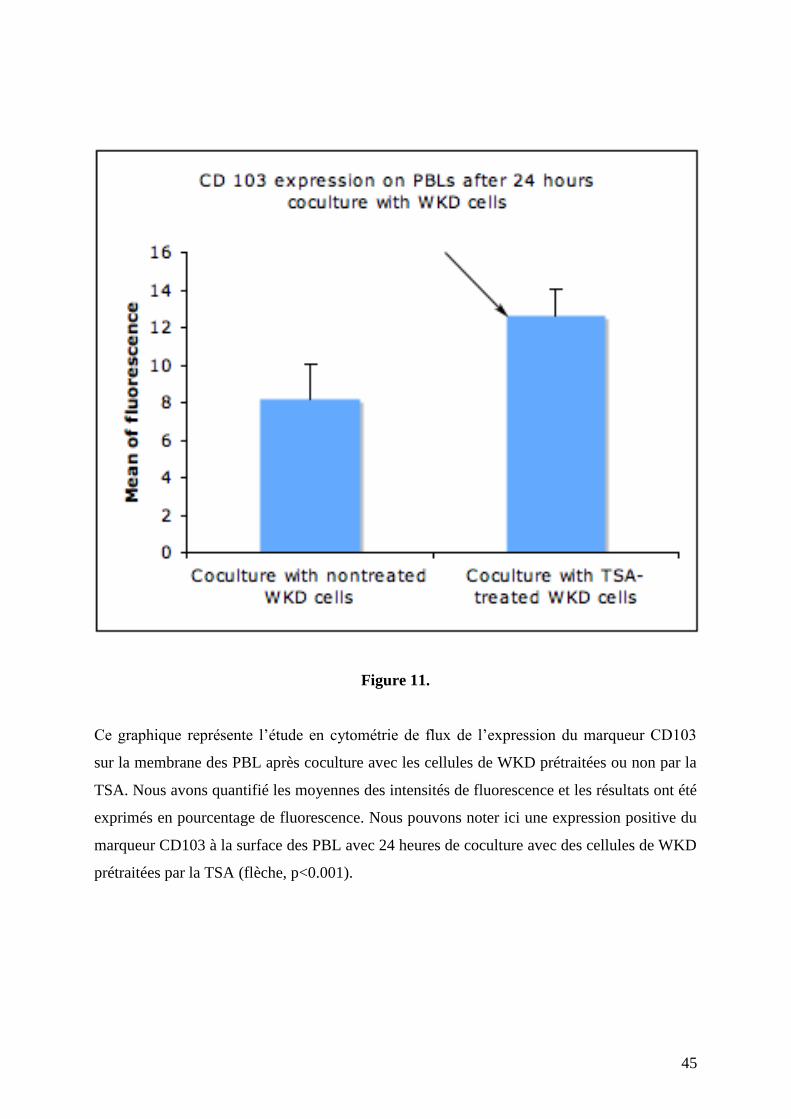
45
Figure 11.
Ce graphique représente l‟étude en cytométrie de flux de l‟expression du marqueur CD103
sur la membrane des PBL après coculture avec les cellules de WKD prétraitées ou non par la
TSA. Nous avons quantifié les moyennes des intensités de fluorescence et les résultats ont été
exprimés en pourcentage de fluorescence. Nous pouvons noter ici une expression positive du
marqueur CD103 à la surface des PBL avec 24 heures de coculture avec des cellules de WKD
prétraitées par la TSA (flèche, p<0.001).

46
Figure 12.
Ce graphique représente les tests d‟évaluation de la toxicité du BAC sur les cellules de WKD,
mesurée à l‟aide des test HOESCHT et Rouge neutre en cytofluorimétrie sur microplaques.
Les résultats ont été exprimés par le rapport du test d‟apoptose HOESCHT au test
d‟évaluation de l‟intégrité des membranes cellulaires à l‟aide du Rouge neutre . Les cellules
de WKD ont été traitées par différentes concentration de BAC allant de 5.10-4
% à 5.10-2
%
pendant 15 minutes suivies de 24 heures de récupération dans un milieu complet, seules ou en
coculture avec des PBL non activés. Nous pouvons noter ici que pour les plus fortes
concentrations de BAC (supérieures à 5.10-3
%), la présence des PBL en contact direct dans le
milieu de récupération induisait une diminution de l‟apoptose de cellules de WKD, en
comparaison avec l‟apoptose induite par le BAC sur les cellules WKD seules (flèches,
p<0.001).

47
4. Discussion
In vivo, la conjonctive joue un rôle central dans le système immunitaire de la surface oculaire
et peut être basiquement décrite comme une étroite coopération de différents types cellulaires
comprenant les cellules immunitaires (telles que les cellules dendritiques, les lymphocytes…)
et les cellules épithéliales, toutes ces cellules participant de façon active à tous les processus
inflammatoires se déclenchant au niveau de la surface oculaire (115). Une forme
d‟investigation des pathologies de la surface oculaire et de la conjonctive en particulier est
l‟utilisation de modèles animaux. Cependant, les cultures cellulaires sont considérées comme
une parfaite alternative à l‟utilisation d‟animaux pour les expérimentations de recherche en
physiopathologie humaine. Les cellules épithéliales jouent le rôle central dans le
fonctionnement de la surface oculaire, et de précédents travaux réalisés par notre équipe se
sont déjà portés sur l‟utilisation de différentes lignées cellulaires pour étudier, par exemple,
les relations entre apoptose et processus inflammatoires dans le cadre d‟investigations en
toxicologie oculaire (73, 82, 95, 96). La culture cellulaire, et plus spécifiquement les lignées
cellulaires permettent de réaliser une croissance en cellules rapide et peu coûteuse en
comparaison avec les cultures primaires de cellules épithéliales conjonctivales. De plus,
reproductibilité et stabilité sont les deux autres avantages majeurs de l‟utilisation des lignées
cellulaires comme outils d‟étude de la surface oculaire. En effet, la lignée cellulaire de WKD,
la première lignée continue, non transfectée issue de cellules épithéliales de conjonctive
humaine, a déjà été très fréquemment utilisée pour des travaux d‟expérimentations portant
principalement sur l‟évaluation de la toxicité des conservateurs utilisés dans les collyres anti-
glaucomateux et anti-allergiques. Mais nous savons que la muqueuse conjonctivale fait
également partie du « système immunitaire muqueux » (116, 117, 118). Cette notion est
validée par la présence, au sein de la conjonctive et de son chorion, de « tissus lymphoïde
associé à la conjonctive » (conjunctiva-associated lymphoid tissue, CALT) composé d‟un
agglomérat de cellules lymphatiques, permettant au CALT de participer à l‟induction de la
production par les lymphocytes B d‟IgA spécifiquement dirigées contre des antigènes
exogènes à la conjonctive (119). Pourtant, cette « part » immunitaire de la conjonctive n‟a
jamais été prise en compte lors de tous les précédents travaux de toxicologie oculaire réalisés
in vitro, ces derniers se focalisant exclusivement sur les cellules conjonctivales seules. En
effet, les modèles in vitro utilisés jusqu‟alors se basaient simplement sur un système binaire
incluant les cellules épithéliales conjonctivales d‟une part et le médicament potentiellement
cytotoxique de l‟autre. Malheureusement, en dépit de leur utilité et de leur simplicité, ces

48
modèles ne semblent pas être entièrement adaptés aux études toxicologiques in vitro, car pas
assez proches de la réalité in vivo de la surface oculaire.
Différentes études par le passé se sont portées sur la coculture de lymphocytes et de cellules
cornéennes, donc en incluant cette part immunitaire de la conjonctive, pour les relations
possibles entre les différents types cellulaires (120, 121). Iwata et al. (122) utilisèrent des PBL
en coculture avec des cellules épithéliales de cornées humaines (human corneal epithelial
cells, HCE cells) pour déterminer le rôle de l‟épithélium dans l‟induction de la reaction
allogénique et dans le processus d‟activation lymphocytaire. Ils démontrèrent qu‟un
traitement préalable par l‟Interferon gamma entraînait l‟expression de l‟antigène HLA de
classe II sur les cellules de HCE, produisant une activation des lymphocytes allogéniques
fraîchement isolés. De plus, Gomes et al. (109) ont décrit un intéressant nouveau modèle de
coculture consistant en la mise en interaction de cellules épithéliales de conjonctive ou de
cornée en culture primaire avec des lymphocytes issus du sang périphérique. Ils étudièrent sur
ce modèle l‟expression du marqueur HML-1/CD103 à la surface des PBL après contact direct
ou indirect avec les cellules épithéliales de la surface oculaire, et ils montrèrent que
l‟expression du marqueur HML-1 à la surface des lymphocytes CD8+ n‟était pas corrélée au
statut d‟activation lymphocytaire.
Partant de là, notre objectif fut d‟étudier les interactions cellulaires entre des cellules
épithéliales en lignée et des lymphocytes issus du sang circulant, dans le cadre d‟un modèle
de coculture in vitro, puis de valider ce modèle comme une tentative de produire un meilleur
modèle d‟étude toxicologique que les cellules épithéliales en monocouche. Ce travail rentre
dans le cadre de la recherche et du développement d‟alternative aux modèles animaux. Cet
important champs de recherche est justifié dans le cadre tant du programme REACH
(Registration, Evaluation, Authorisation and Restriction of chemical substances, REACH,
2006) que du 7ième amendement de 1976 portant sur les directives “cosmétiques”, qui
baniera d‟ici 2013 tous les tests d‟ingrédients cosmétiques sur les animaux (123).
Dans notre étude, nous avons d‟abord mis en évidence la survenue d‟interactions rapprochées
in vitro entre les cellules de WKD et les PBL, telles que des échanges de membranes, une
fusion de membranes cellulaires pouvant s‟apparenter à une forme de macropinocytose. La
macropinocytose est un processus qui permet la capture d‟une grande quantité de matériel,
ressemblant à une forme d‟endocytose, et pouvant réguler, au sein de la cellule, l‟exocytose
ou d‟autres propriétés membranaires. Dans de précédents travaux, il a été montré que la
macropinocytose pouvait être soit un phénomène constitutif de la cellule, comme pour les
cellules phagocytaires professionnels, telles que les cellules dendritiques ou les macrophages

49
(124), soit un phénomène induit comme pour les cellules épithéliales lorsqu‟elles sont
stimulées par des facteurs de croissance (125). Dans notre travail, les cellules de WKD ont
englouti et digéré des fragments de lymphocytes selon un mode temps-dépendent similaire à
ce qui pourrait être observé chez les phagocytes professionnels. Mais ces macropinosomes
n‟ont pas été redirigé vers la fusion avec les lysosomes comme cela aurait pu être le cas, au
regard de la façon dont ces pinosomes sont traités dans les macrophages. En effet, le matériel
internalisé, dans nos travaux, a été recyclé à la membrane, en moins de 24 heures. Au niveau
des muqueuses, une telle activité de macropinocytose des cellules épithéliales a déjà été
décrite comme support de la fonction de nettoyage des phagocytes résidents professionnels.
En effet, il a été démontré que la macropinocytose induite par l‟EGF au sein des cellules
épithéliales permettait la capture et l‟internalisation de matériel qui était alors très rapidement
recyclé en retour, à la membrane de la cellule (126, 127). Ce phénomène met en lumière un
des rôles possibles de traitement et de présentation d‟antigène endogènes et exogènes de la
cellule épithéliale au sein du système immunitaire muqueux. De plus, dans notre modèle de
coculture, les interactions très rapprochées entre les deux types cellulaires ont conduit à la
fusion des membranes cellulaires.
De nombreuses hypothèses concernant la physiopathologie des principaux troubles de la
surface oculaire ont été émises suites à des travaux réalisés in vitro sur une monocouche de
cellules épithéliales en lignée de WKD. Principalement, il a été démontré que l‟Interferon
gamma induisait le programme de mort cellulaire et entraînait une surexpression du marqueur
HLA-DR et de l‟antigène CD95/Fas à la surface des cellules épithéliales (83). Dans notre
étude, l‟expression de HLA-DR sur les cellules de WKD était augmentée après 7 jours de
coculture soit en contact direct avec des PBL non activés, soit sans contact direct (séparés par
un Transwell) avec des PBL activés. Le marqueur d‟apoptose CD95/Fas était exprimé sur les
cellules de WKD seulement après 1 jour de coculture en contact direct avec des PBL activés.
En rapport avec ses résultats, nous pouvons émettre l‟hypothèse que dans notre système de
coculture, en présence de PBL, l‟activation cellulaire, l‟inflammation et l‟apoptose pourraient
éventuellement être médiées par le ou les mêmes stimuli et les mêmes voies de transduction,
liés à des interactions rapprochées entre les deux types cellulaires, via un contact cellulaire
direct ou par le biais de médiateurs solubles. La stimulation allogénique qui s‟en suit peut
alors parfaitement résulter initialement de la libération d‟Interferon gamma. Par exemple,
L‟interferon gamma pourrait permettre l‟expression du marqueur HLA-DR sur les cellules
épithéliales, et par la suite induire la réponse allogénique lymphocytaire par le biais du
complexe majeur d‟histocompatibilité (121). Cependant, dans d‟autres modèles de réactions

50
primaires lymphocytaires, la réaction allogénique des lymphocytes ne fut pas
systématiquement induite même en la présence de l‟Interferon gamma ou du marqueur HLA-
DR (128, 129), pour des raisons qui demeurent encore actuellement obscures. Il a été suggéré
que l‟expression d‟HLA-DR était alors nécessaire mais pas forcement suffisante. De plus,
Gomes et al. (130) ont étudié les changements induits sur l‟expression des antigènes de
surface des lymphocytes activés et non activés provenant des mêmes donneurs mis en
coculture avec des cellules épithéliales de cornée provenant de donneurs différents. Ils ont
observé que la réaction allogénique causée par la présentation d‟antigènes par les cellules
épithéliales entraînait une expression très variable du marqueur d‟activation CD25 à la surface
des lymphocytes, en concluant que l‟utilisation de lymphocytes allo réactifs dans un modèle
de coculture avec des cellules épithéliales n‟influencerait pas particulièrement l‟expression
d‟antigène de surface sur les deux types cellulaires. L‟absence d‟utilisation de l‟Interferon
gamma au cours de leurs expérimentations pourrait expliquer la variabilité d‟expression du
marqueur d‟activation lymphocytaire. En transposant le même raisonnement à nos résultats, la
surexpression des marqueurs HLA-DR et CD95/Fas induite par la présence des PBL n‟a très
probablement pas été influencée par la réaction allogénique. La surexpression de HLA-DR a
plutôt été induite par un effet pro inflammatoire sur les cellules de WKD lié à la présence de
cellules immunitaires.
Le système immunitaire muqueux inclus de façon traditionnelle la présence de lymphocytes
intra épithéliaux (Intra Epithelial Lymphocytes, IEL) qui sont représentés en majorité par une
sous population spécialisées de lymphocytes T CD8+ présentant un récepteur T αβ positif
(131, 132, 133). Plus de 90% de ces IEL expriment le récepteur HML-1/CD103, contre
seulement 2% des PBL (134, 135). Cette population de lymphocytes T pourrait jouer un rôle
spécial de défense immunologique au niveau de la surface oculaire. Le marqueur HML-
1/CD103 est une molécule d‟adhésion appartenant à la famille des intégrines, composée d‟une
sous unité 7 en association avec une sous unité E (136), permettant une adhésion
spécifique des IEL avec les cellules épithéliales muqueuses. Cette adhésion des IEL aux
cellules épithéliales se fait grâce au récepteur HML-1/CD103 sur les IEL se liant à une
molécule d‟adhésion de distribution non ubiquitaire appelée E-Cadhérine (137, 138, 139,
140). Les cadhérines constituent une famille de molécules d‟adhésion de surface impliquées
dans l‟adhésion cellulaire via une interaction moléculaire spécifique homophilique calcium-
dépendante (139). L‟E-cadhérine est nécessaire pour la formation des jonctions adhérentes
entre les cellules épithéliales. Gomes et al. (109) ont retrouvé une expression de l‟intégrine
E7 (CD103/HML-1) sur les PBL après sept jours de coculture avec des cellules épithéliales

51
en culture primaire, indiquant un probable changement de phénotypes de PBL en IEL. Sept
jours de coculture étaient le délai d‟incubation qui semblait optimal pour l‟expression du
marqueur HML-1 sur les lymphocytes activés (140). Dans notre étude, l‟expression du
marqueur CD103/HML-1 n‟a pas été induite sur les PBL après 1 ou 7 jours de coculture avec
des cellules de WKD. La principale différence entre les cellules épithéliales en culture
primaire et la lignée de cellules de WKD réside dans la perte de l‟expression de l‟E-cadhérine
à la surface des cellules de WKD (79). D‟ailleurs, bien que la plus fréquente des réserves
émises sur les lignées cellulaires pour les accepter en tant qu‟équivalent de tissu humain fiable
soit la ressemblance phénotypique de la lignée immortalisée avec son homologue précurseur
non transformé, les cellules de WKD partageraient de nombreuses caractéristiques
morphologiques et fonctionnelles communes avec les cellules épithéliales humaines in vivo.
Par exemple, Les cellules de WKD montrent un développement de microvilli, la présence
d‟un récepteur à l‟EGF, des desmosomes, une sécrétion de mucine… A l‟inverse, les cellules
de WKD présentent également des caractéristiques qui font d‟elles des cellules immortalisées,
dédifférenciées avec un phénotype mésenchymateux d‟apparence fibroblastique, telles que
l‟expression élevée de la vimentine, la perte de la E-cadhérine, l‟expression de la cytokératine
4 et la présence d‟anomalies caryotypiques (79, 101, 103). Toutefois, l‟utilisation de la
Trichostatine A (TSA), un inhibiteur des histones désacétylases (HDCACs inhibitor) nous a
permis de retrouver une expression faible mais positive d‟E-Cadhérine à la surface des
cellules de WKD. En effet, selon des travaux antérieurs, il avait été montré que la régulation
négative à long terme de l‟expression d‟E-Cadhérine était liée à un processus transcriptionnel,
utilisant des facteurs répresseurs de la transcription tels que les protéines en “doigt de zinc” et
les histones désacétylases (141, 142, 143). Nous avons donc choisi d‟utiliser un inhibiteur des
histones désacétylases afin de permettre la suppression de la répression de transcription de
l‟E-cadhérine au sein des cellules de WKD. Ainsi, dans notre travail, nous avons pu observer
une fine expression du marqueur CD103/HML-1 à la surface des PBL après 1 et 7 jours de
coculture avec des cellules de WKD positives pour l‟E-cadhérine, c‟est à dire ayant été
prétraitées par la TSA. D‟autres expérimentations seraient tout de même maintenant
nécessaires afin de définitivement confirmer la possibilité de la transformation des PBL en un
phénotype d‟IEL au contact de cellules épithéliales positives pour l‟E-cadhérine.
D‟anciennes études toxicologiques utilisant le Hoechst33342 et le rouge neutre en
cytofluorimétrie sur microplaques avaient évalué les effets proapoptotiques et pronécrotiques
du BAC sur les cellules de WKD. Cet effet toxique du BAC augmentait avec la concentration
de ce dernier (114). Dans notre modèle, les PBL semblent montrer un rôle protecteur sur les

52
cellules de WKD pour les concentrations de BAC les plus élevées (celles supérieures à 5.10-
3%), concentration de BAC pour lesquelles l‟apoptose des cellules épithéliales est maximale,
comme si une mort excessive des cellules épithéliales pouvaient alors envoyer un message de
demande de survie aux PBL. A l‟inverse, pour les plus faibles concentrations de BAC (celles
inférieures à 10-3
%), les PBL semblent alors avoir un rôle pro-apoptotique sur les cellules de
WKD. Des expériences complémentaires sont donc nécessaires dans un premier temps pour
confirmer définitivement cet effet bipolaire de la présence de PBL sur les cellules de WKD en
fonction de la concentration de BAC, puis dans un second temps pour définitivement valider
ce modèle séduisant mais complexe pour de nouvelles études toxicologiques. Quoi qu'il en
soit, ces résultats permettent déjà la mise en lumière des interactions entre les cellules
immunitaires et les cellules épithéliales de la surface oculaire.
En conclusion, cette étude constitue une première étape pour mieux comprendre les relations
et les interactions dans un modèle in vitro entre des cellules épithéliales de la conjonctive et
des lymphocytes. Des connexions rapprochées et des interactions cellulaires par le biais de
médiateurs solubles ou par contact direct entre les deux types de cellules ont été observées
dans ce modèle, avec des effets proinflammatoires et proapoptotiques, en rapport avec
l‟expression des marqueurs HLA-DR et CD95/Fas ou le profil du pic sub-G1. Une réaction
allogénique entre une lignée cellulaire d‟un donneur et des cellules immunitaires issues d‟un
autre donneur ne pourrait expliquer à elle seule tous ces résultats. Une voie alternative de
recherche serait de collecter les cellules conjonctivales d‟un patient, de les cultiver, selon la
méthode de culture primaire, et de renouveler la procédure de coculture avec les PBL issus de
ce même patient. Mais cette manœuvre semble très compliquée à mettre en oeuvre et
techniquement très difficile, manquant de reproductibilité du fait d‟une grande variabilité liée
aux cultures primaires de l‟épithélium conjonctival et des quantités extrêmement faiblesde
tissu disponible auprès des donneurs vivants. Une expression de la E-cadhérine sur les
cellules de WKD a été observée dans notre étude suite au traitement par la TSA, avec en
retour une discrète expression du marqueur CD103/HML-1 sur les PBL, démontrant un
potentiel réel de commutation des PBL en un phénotype intra épithélial lié à la coculture. En
outre, un double effet, à savoir de protection et pro-apoptotique, de la présence des PBL sur
les cellules de WKD en fonction de la concentration de BAC appliquée au préalable sur les
cellules de WKD a été trouvé, apportant des éléments prometteurs à la compréhension du
comportement des xénobiotiques sur la surface oculaire ainsi que sur les interactions entre les
cellules immunitaires, les cellules épithéliales dans le carde d‟un stress environnemental. Ces
résultats nous amènent à croire que ce modèle pourrait effectivement constituer un pas en

53
avant, comme une nouvelle perspective d‟étude de la physiopathologie et de la toxicologie in
vitro dans le cadre des affections de la surface oculaire. La prochaine étape pourrait consister
à utiliser une approche similaire pour explorer davantage l'interaction entre la lignée de
cellules de WKD et des populations spécifiques immunitaire, telle qu‟une sous population de
lymphocytes ou une population de cellules dendritiques. Nous utiliserions un système de
coculture in vitro, mettant en contact des lignées cellulaires établies à la fois de cellules
épithéliales et de monocytes / macrophages afin de définitivement surmonter les problèmes de
variabilité extrinsèque liée aux cultures primaires et de variabilité intrinsèque liée aux
donateurs individuels.

54
V. CONCLUSION ET PERSPECTIVES
La lignée de cellules conjonctivales de Chang, première lignée spontanément immortalisée, a
été beaucoup utilisée au cours de ces dix dernières années dans le cadre d‟études
toxicologiques pour illustrer les effets toxiques des conservateurs utilisés dans les collyres, en
particulier le rôle du chlorure de benzalkonium, soulignant que l‟enjeu actuel de la
pharmacopée en ophtalmologie est de pouvoir développer des collyres qui n‟en contiennent
pas. En effet, la lignée de Chang présente les avantages d‟être facile à cultiver, d‟être stables
et peu coûteuses. Cependant, utiliser cette lignée dans un système monocellulaire pour tester
la toxicité d‟un conservateur ou d‟un collyre, revient à considérer la conjonctive comme étant
constituée exclusivement de cellules conjonctivales. Cependant, les mécanismes complexes
impliqués dans la toxicité de la surface oculaire font intervenir d‟autres types cellulaires
comme les mucocytes et les cellules inflammatoires (lymphocytes, monocytes, cellules
dendritiques) avec de nombreuses interactions entre ces différents types cellulaires. Une
approche pertinente et innovante pour la compréhension de la physiopathologie de la
conjonctive est donc de développer un modèle de coculture faisant intervenir des cellules
épithéliales (culture primaire ou lignées) et des cellules inflammatoires (lymphocytes par
exemple) afin de se rapprocher de la réalité anatomique et physiologique de la conjonctive.
Basés à la fois sur l‟expérience de notre équipe dans l‟utilisation de la lignée de cellules
épithéliales de Chang, et sur un travail publié mettant en coculture des lymphocytes
périphériques avec des cellules conjonctivales et cornéennes en culture primaire, nous avons
décidé d‟étudier les interactions entre les cellules de Chang et les lymphocytes périphériques,
et d‟éprouver ce modèle de coculture comme outils d‟étude de la surface oculaire sur le plan
toxicologique. Les interactions manifestes entre les deux types cellulaires et les conséquences
de ces interactions en termes d‟inflammation, d‟apoptose, de mort et de survie cellulaire, que
nous avons mis en évidence, n‟occultent cependant pas le fait que la reaction allogénique
déclenchée par le contact direct ou par le biais de médiateurs solubles entre des cellules
épithéliales et des lymphocytes alloréactifs doit être considérée dans l‟interprétation des
résultats et dans leur reproductibilité. Par ailleurs, certains paramètres de dédifférenciation des
cellules de la lignée de Chang, comme la perte de l‟E-cadhérine, pourraient être un obstacle
aux bonnes interactions cellules épithéliales/cellules inflammatoires qui caractérisent ce tissu
complexe qu‟est la conjonctive. La suite intéressante à donner à ce travail serait donc
d‟étudier les interactions entre des cellules épithéliales d‟un donneur et des cellules
dendritiques, obtenues à partir de lymphocytes périphériques provenant du même donneur,

55
réduisant au maximum les effets collatéraux d‟une éventuelle réaction allogénique et laissant
libre cours aux interactions cellulaires.

56
VI. ANNEXE
In vitro interactions between peripheral blood lymphocytes and Wong-Kilbourne
Derivative of Chang conjunctival cells
A. Chassignol 1,2
, E. Brasnu 1,2
, C. Baudouin 1,2,4
, L. Riancho 2,3
, J.-M. Warnet 2,3
, and F.
Brignole-Baudouin 2,3
.
1- Department of Ophthalmology III, Quinze-Vingts National Ophthalmology Hospital, Paris, France.
2- INSERM, U968, - UPMC Univ Paris 06, UMR_S 968, Institut de la Vision, CNRS, UMR_7210
Paris F-75012, France.
3- Department of Toxicology, Faculty of Biological and Pharmacological Sciences, Paris, France.
4- Ambroise Paré Hospital, APHP, University of Versailles, France.
Correspondence to: Christophe Baudouin, Department of Ophthalmology III, Quinze-Vingts National
Ophthalmology, Hospital, 28 rue de Charenton, 75012, Paris, France; Phone: 33 1 40 02 13 01; FAX:
33 1 40 02 13 99; email: [email protected]
ABSTRACT The Chang cells (WKD) are used as a monocellular in vitro model to assess the toxic effects of
Benzalkonium Chloride. However, conjunctival epithelium is composed with epithelial cells but
includes also inflammatory cells. We therefore assessed the interactions between WKD cells and
lymphocytes (PBL) to validate in vitro this new model. PBL were cocultured with WKD cells using
cell-contact or transwell cultures. Morphological analyses were performed using plasma membrane
staining. Sub-G1 peak, HLA DR, E-cadherin and CD103 expression on cell types were assessed using
flow cytometry (FCM). BAC effects were assessed by microplate cytofluorometer assays. Coculture
allowed membrane exchanges, increased HLA DR on WKD, and CD103 on PBL and seemed to
reduce apoptosis after BAC treatment. Such results lead us to believe that this model of WKD / PBL
coculture could actually constitute a step forward, as a new prospect for toxicological in vitro studies
on the ocular surface.
INTRODUCTION Benzalkonium Chloride (BAC) is the main preservative used in ophthalmic solutions such as
antiglaucoma or anti-allergic eye drops. In the eye, BAC turnover is very low, and this molecule was
shown to be retained in ocular tissues up to 7 days after a single drop administration (1). Its ocular
cytotoxicity was demonstrated in many in vivo and in vitro models (2). Several studies showed the role
of BAC in ocular surface inflammation, allergy, fibrosis, and dry eye syndrome (3), and its
pronecrotic, pro-apoptotic and pro-oxidative effects on conjunctival cells in vitro and in vivo (4).
However, human conjunctival epithelium is not only composed with conjunctival epithelial cells, but
includes other cell types such as a variety of inflammatory cells and goblet cells, in close connections
with blood vessels and extraocular lymphocytes (5). All these cell types contribute to maintain a
healthy ocular surface, and there is a large body of evidence on the direct role of the conjunctival
epithelium in inflammatory diseases of the ocular surface (6, 7).
To assess BAC and other xenobiotic toxicity on the conjunctival epithelium, one of the mostly used
methods was based on in vitro systems that offer the possibility of studying the influence of
metabolites, mediators or drugs in a controlled stable and reliable environment. There are currently
two main continuous, untransfected epithelial cell lines available, the Wong-Kilbourne derivative of
Chang cells, WKD (8) and the IOBA-NHC cells (9). The WKD cell line has been immortalized
spontaneously from normal human conjunctival epithelial cells (10) but differs from normal tissue and
primary cultures in its phenotypic characteristics and in its response to inflammatory cytokines (11),
and one reserve for accepting cell lines as an equivalent of human tissue is their phenotypic
resemblance with their non-transformed precursors. WKD cell line, spent as a monocellular and
reliable nonclinical model, has been already used in several studies to assess the cytotoxic effects of

57
BAC (12, 13), but all these in vitro studies used epithelial cell lines isolated without taking into
account the inflammatory components existing in the normal human conjunctiva. In vitro interactions
between both inflammatory and epithelial cell types have already been studied in skin and lung models
(14-17). Such a study has been performed on conjunctiva as well, using a coculture model between
human primary cultured conjunctival cells and human peripheral blood lymphocytes (PBL), that
showed modifications of both T cells' and epithelial cells‟ characteristics (18). But in comparison with
primary cultures, an established cell line like the WKD cell line offers better reproducibility and
rapidity for acquiring data and has the advantage of being independent from conjunctival biopsy
availability. This is why this cell line was chosen as model for previous toxicological studies tackling
the ocular surface (4, 12, 13). A further step forward was hence to associate this cell line with
inflammatory cells and study between-cell interactions. To best fit the study by Gomes et al. (18), we
started by mixing our well-validated epithelial cell line with PBL. Therefore, the aim of this study was
to assess the interactions between WKD and PBL in order to develop and validate in vitro the potential
of such new model for investigating ocular surface disorders and further extend toxicological research.
RESULTS Cellular interactions:
Plasma membrane exchanges between both cell types were observed after 1 day of coculture in the
direct-contact coculture model between epithelial cells and PBL (Figure 1). An obvious integration of
lymphocyte membranes in epithelial cellular membrane rapidly occurred, in a time-dependent manner.
A peak was obtained after 90 minutes of coculture (Figure 2). These tight connections between both
cell types, resembling cell membrane fusion, have been observed in SEM as well (Figure 3), and this
phenomenon seemed to be similar to pinocytosis. Lymphocytes that were disposed between epithelial
cell layers (Data not shown) showed cytoplasmic expansions after phalloidin staining and fluorescence
microscopy analysis (Figure 4).
Figure 1
Figure 2
A B
A
B
C
D

58
Figure 3
Figure 4
Apoptosis:
Results were expressed in percentage of sub-G1 events compared to the total cell population. After a
1-day coculture, apoptosis of WKD cocultured with non-activated PBL remained unchanged. On the
other hand, as expected, apoptosis of WKD increased when PBL were activated by PMA and
ionomycin. These cytotoxicity profiles obtained on WKD after 1 day of coculture with activated PBL
were similarly obtained in direct contact or when cells were separately cultured using a Transwell.
Then, alteration of DNA content of WKD pre-treated with 5.10-3
% BAC for 15 min followed by 24 h
of cell recovery was observed in the presence of non-activated PBL (Figure 5). This potentiated
cytotoxic effect by the presence of PBL was noted only when WKD were cocultured in direct contact
with PBL, but not when they were separated by a Transwell. We used, as previously described (19),
the 5.10-3
% BAC threshold concentration that induced a significant toxicity on WKD. Indeed,
membrane integrity decreased in a BAC concentration-dependent manner. The highest cytotoxicity
was observed with BAC concentrations higher than 5.10-3
%. This BAC concentration is equivalent to
or lower than the concentrations of most available eye drops.
Figure 5

59
Epithelial cell behavior:
Either constitutively or after coculture with PBL, no E-cadherin expression on WKD was observed in
flow cytometry and in standard immunofluorescence. Nevertheless, the Trichostatin A pre-treatment
of WKD induced a weak E-cadherin expression detected with flow cytometry (Figure 6) and standard
immunofluorescence staining (Figure 7). HLA DR expression increased on WKD after 1 day of direct-
contact coculture with non-activated PBL, and after 7 days of non-contact coculture with activated
PBL (Figure 8). CD95/Fas antigen was expressed on WKD after 1 day of direct-contact coculture with
activated PBL. Surprisingly, the CD86 costimulating marker was expressed on WKD after 1 day of
coculture with activated PBL. Furthermore, WKD shrinkage increased in a similar manner after 1 or 7
days of coculture using phalloidin and propidium iodide stainings (data not shown).
Figure 6
Figure 7
Figure 8

60
Lymphocyte behavior:
In our study, no CD103 lymphocyte expression was obtained after WKD / PBL coculture. However,
TSA pre-treated WKD / PBL coculture showed a slight increase of CD103 expression on PBL (Figure
9).
Figure 9
Cytotoxicity evaluation:
WKD cells‟ apoptosis increased in a BAC concentration-dependent manner (from 5.10-4
% to 5.10-2
%),
no matter of the presence of PBL during the recovery time. Furthermore, for the BAC lowest
concentrations (less than 10-2
%), the presence of PBL, in coculture during the recovery time, increased
apoptosis after 1 day of direct contact, as a potentiating of the apoptotic effect of BAC, consistent with
the results obtained on subG1peak with a 5.10-3
% concentration. Conversely, for higher concentration
of BAC (more than 5.10-3
%), the presence of PBL in direct contact coculture during the 24h recovery
time induced a decrease of apoptosis (Figure 10).
Figure 10
DISCUSSION
In vivo, conjunctiva plays a central role in the immune system of the ocular surface and is basically
characterized by a tight cooperation between immune cells, like dendritic cells, lymphocytes, and
epithelial cells, all cell types actively participating in inflammatory processes (20). One way to
investigate ocular surface and conjunctiva is to use animal models. Nonetheless, cell cultures are
generally considered to be a good alternative to animal studies for investigating the pathophysiology
of human diseases. Epithelial cells play a central role in ocular surface functioning, and previous
studies of our team concerned toxicological investigations and assessment of relationship between
apoptosis and inflammatory process in epithelial cells using various cell lines (4, 6, 7). Cell cultures,
and especially cell lines, allow quick and cheap cell growth in comparison with primary cultures.
Moreover, reproducibility and stability are two main advantages of the utilization of cell lines for
investigating the ocular surface. Indeed, the Wong-Kilbourne derivative of Chang conjunctival

61
(WKD) cell line, the first established continuous, untransfected epithelial cell line from normal human
conjunctiva, has been widely used in previous experiments, particularly for the assessment of the
toxicity of preservatives and preserved antiglaucoma or anti-allergic eye drops. But we know that the
conjunctival mucosa is also part of the mucosal immune system (21, 22). This is supported by the
presence of conjunctiva-associated lymphoid tissue (CALT) composed of aggregated lymphatic cells,
as CALT participates to the induction of an antigen-specific IgA production by B cells, against
exogenous antigen to conjunctiva (23). Thus, this immune counterpart of the conjunctiva was not
considered in previous in vitro toxicological ocular surface studies performed on conjunctival cells
alone. Indeed, these studies were performed using a binary system including epithelial conjunctiva
cells on one side and a potentially cytotoxic drug on the other side. Unfortunately, despite its
usefulness and easiness, this model seems to be not fully suitable for toxicological in vitro studies,
because not close enough to the in vivo reality.
Previous experimental models involving coculture of lymphocytes and corneal cells, therefore
including the immune part of conjunctiva, have been used to study such relations (24, 25). Iwata et al.
(26) used PBL cocultured with human corneal epithelial (HCE) cells to determine the role of
epithelium and allogeneic lymphocyte interaction on the lymphocytic activation process. They
demonstrated that IFN-gamma-treated, HLA class II-bearing HCE cells stimulated freshly isolated
allogeneic lymphocytes. Moreover, Gomes et al. (18) described a new interesting coculture model
consisting of primary cultures of human corneal or conjunctival epithelial cells with peripheral blood
lymphocytes. They studied HML-1 expression on PBL induced by direct and indirect interactions with
ocular surface epithelial cells, showing that this HML-1 expression on CD8+ lymphocytes was not
correlated to the PBL activation status.
Hence, our purpose was to study epithelial cell-lymphocyte interactions in an in vitro coculture model
using a conjunctival epithelial cell line, and then to validate this model for toxicological investigations
of the ocular surface, as an attempt to provide a better toxicological model than single epithelial
monolayers. This work is part of the development of alternative methods to animal testing. This
important field of research is justified in the context of both the REACH program (Registration,
Evaluation, Authorisation and Restriction of chemical substances, REACH, 2006) and the 7th
amendment to the 1976 cosmetics directive, which will ban testing of cosmetic ingredients on animals
by 2013 (27).
First, we observed tight in vitro interactions between WKD and PBL, such as membrane exchanges,
cellular membrane fusion and macropinocytosis, a process that allows the capture of large material, as
a form of endocytosis, which could regulate exocytosis and other membrane properties. In previous
studies, it has been shown that macropinocytosis can be either constitutive, as in professional
phagocytes, like dendritic cells or macrophages (28), or induced as in epithelial cells when stimulated
by growth factors (29). In our study, WKD engulfed and digested fragments of lymphocytes in a time-
dependent manner similar to that of professional phagocytes. But, these macropinosomes did not
undergo fusion with lysosomes like they used to do in macrophages. Indeed, the internalized material
was recycled back to the cell membrane, in less than 24 hours. In mucosal tissues, such an activity in
epithelial cells has been reported as a support for the scavenging activity of resident macrophages.
Indeed, it has been shown that EGF-induced macropinocytosis in epithelial cells allowed the capture
and internalization of the material that was, however, rapidly recycled back to the cell surface (30, 31).
This phenomenon shed in light a possible role in processing and presenting self and foreign antigens
by epithelial cells to the mucosal immune system. Moreover, in our coculture model, tight in vitro
interactions between both cell types also led to cell membrane fusion that was observed using SEM.
Many hypotheses about pathophysiology of ocular surface disorders have been raised from WKD in
vitro studies conducted in a monolayer system. Principally, it has been demonstrated that IFN-gamma
induced programmed cell death and a dose-dependent up-regulation of HLA DR and CD95/Fas
antigens (32). In our study, HLA DR expression increased on WKD after 7 days of coculture either in
direct-contact with non-activated PBL, or without any contact but with activated PBL. The apoptosis-
related marker CD95/Fas was expressed on WKD after only 1 day of direct-contact coculture with
activated PBL. According to these results, we can hypothesize that, in our system in presence of PBL,
cell activation and apoptosis could possibly be mediated by the same stimuli and the same
transduction pathways, with tight between-cell interactions, through direct cell contact or via soluble
mediators. An allogeneic stimulation could also be hypothesized, through IFN-gamma. For example,

62
IFN-gamma could lead to HLA DR antigen expression on epithelial cells, so as to induce allogeneic
lymphocyte responses by MHC-dependent mechanism (25). Nevertheless, in other models of primary
mixed lymphocyte reactions, allogeneic lymphocytes were not always induced, even in presence of
IFN-gamma or HLA DR expression (33,34) for reasons that remain unclear. It has been suggested that
HLA DR expression is necessary but may not be hardly sufficient. Moreover, Gomes et al. (35)
studied the changes induced on surface antigens of non-activated lymphocytes from the same donors
cocultured with corneal epithelial cells from different donors. They observed that any allogeneic
response caused by antigen presentation resulted in a variability of CD25 activation on PBL and they
concluded that using allogeneic lymphocytes for coculture with epithelial cells did not particularly
influence the expression of surface antigens on both cell types. This could be explained by the lack of
IFN-gamma in their experiments. In the same way, in the present study, the up-regulation of HLA DR
and CD95/Fas on WKD induced by the presence of PBL was unlikely to have been influenced by an
allogeneic reaction. The up-regulation of HLA DR may have been rather induced by a
proinflammatory effect on WKD related to the presence of immune cells.
The mucosal immune system traditionally includes intraepithelial lymphocytes (IELs) that are
predominantly a specialized subpopulation of CD8+T cells with a αβ positive T-cell receptor
(36,37,38). More than 90% of these IEL express the human mucosal lymphocyte antigen (HML-1,
CD103), against only 2% of PBL (39, 40). This T-cell population might play a special role in
immunological defense of the ocular surface. The HML-1 is an adhesion molecule belonging to the
integrin family, composed of the 7 subunit in association with the E subunit (41) and it mediates
specific interaction between IEL and mucosal epithelial cells thanks to a tissue-restricted adhesion
molecule named E-cadherin (40, 41, 42, 43). Cadherins constitute a family of cell surface adhesion
molecules that are involved in calcium-dependent homophilic cell-to-cell adhesion (44). E-cadherin is
required for the formation of adherent junctions between epithelial cells. Gomes et al. (18) found an
expression of the integrin E7 (CD103/HML-1) on PBL after 7 days of coculture with epithelial
cells in primary culture indicating a transformation of PBL into IEL. Seven days seemed to be the
optimal incubation time for HML-1 expression on activated lymphocytes (45). In our study,
CD103/HML-1 expression was not induced on PBL after 1 to 7 days of coculture with WKD. The
main difference between epithelial cells in primary culture and WKD is the loss of E-cadherin
expression on WKD surface (11). Although, one of the most frequent reserve for accepting the cell
line as a valuable equivalent of human tissue is the phenotypic resemblance of immortalized cells with
regard to their non-transformed precursors, WKD share common morphological and functional
characteristics with human conjunctival cells in vivo, such as the development of microvilli, the
presence of EGF receptor and desmosomes or mucin secretion. Conversely, WKD also show
characteristics related to an immortalized, dedifferentiated and mesenchymal cell phenotype with
fibroblastic appearance, vimentin-high expression, loss of E-cadherin, cytokeratine-4 expression, and
presence of kariotypic abnormalities (8, 10, 11). Nevertheless, using Trichostatin A, a histone
deacetylase (HDCACs) inhibitor, we observed a weak E-cadherin expression on WKD. Indeed,
according to previous studies, it has been shown that long-term E-cadherin down-regulation could be
transcriptional, using « zinc-finger » protein repressor factors and HDCACs (46, 47, 48). Thus, in our
study, a slight CD103/HML-1 expression was observed on PBL after 1 to 7 days of coculture with E-
cadherin-positive WKD. Further experiments are thus required to definitively assess the ability of PBL
to develop an IEL phenotype after coculture with WKD.
Previous toxicological studies using the Hoechst33342 and Neutral Red assays have shown the
proapoptotic and pronecrotic effects of BAC on WKD with a BAC concentration-dependent increase
(19). In this model, PBL tended to show a protective effect on WKD against BAC for the highest BAC
concentrations (more than 5.10-3
% BAC), concentrations where WKD apoptosis was maximal, as if
excessive epithelial cell death would send a survival signal to PBL. Conversely, for lower
concentrations of BAC, PBL seemed to have a pro-apoptotic role on WKD. Further experiments are
therefore needed to definitively confirm such bipolar effects of the presence of PBL on WKD
depending on the BAC concentration, and then definitively validate this appealing but complex model
for further toxicological studies. Anyway, these results already shed light on the interactions between
immune and epithelial cells in the ocular surface.
In conclusion, this study is a first step for better understanding the relationships and interactions
between conjunctival epithelial cells and lymphocytes in an in vitro model. Tight connections and cell

63
interactions through soluble mediators or through direct cell contact have been observed in this model,
with pro-inflammatory and pro-apoptotic effects, according to HLA DR expression, CD95/Fas or sub-
G1 profile. This might not be explained only by the allogeneic reaction between cell lines from one
donor and immune cells issued from another one. An alternative way of research might be to collect
and culture conjunctival cells, for primary culture, and peripheral blood cells from the same person,
but it appears highly challenging and technically difficult, lacking reproducibility, due to the high
variability of primary cultures of conjunctival epithelium and the tiny amounts of tissues usually
available from a live donor. Expression of E-cadherin on WKD was observed in our model, with a
trend toward CD103/HML-1 expression on PBL demonstrating a potential switch to an intra-epithelial
phenotype. Moreover, a dual, namely protective or proapoptotic, effect of PBL on WKD depending on
the BAC concentrations, was found, opening up very promising findings on the effects of xenobiotics
on the ocular surface and possible interactions between immune cells, epithelial cells and
environmental stresses. Such evidence leads us to believe that this model could actually constitute a
step forward, as a new prospect for pathophysiological and toxicological in vitro studies on the ocular
surface. The next step could be to use a similar approach to further explore the interaction between
conjunctival cell lines and specific immune populations like lymphocyte subpopulations or dendritic
cells in an in vitro coculture system, using established cell lines of both epithelial and
monocyte/macrophage lineages, in order to definitively overcome the issues of primary cultures and
variability intrinsic to individual donors.
METHODS
Conjunctival cell line:
The Wong-Kilbourne derivative of Chang conjunctival cells (clone 1 to 5c-4 American Type Culture
Collection [ATCC, Manassas, VA]-certified cell line [CCL], 20.2) were cultured under standard
conditions (humidified atmosphere of 5% CO2 at 37 °C) in Dulbecco‟s minimum essential medium
(DMEM) supplemented with 10% fetal bovine serum (FBS), 1% glutamine, 50 UI/ml penicillin, and
50 UI/ml streptomycin.
Cells were removed by gentle trypsin incubation at confluence and counted. Then, they were seeded
into 6-well cell culture plates (Corning, Schiphol-Rijk, the Netherlands) for cytometric analysis, and
on slides (Lab-Tek™ II chambered coverglass; Nunc International, Naperville, IL) for standard
immunofluorescence and confocal microscopy. Cultures were kept at 37 °C for 24 h. Subconfluent
cells (culture surface covering nearly 70%) were then exposed to the different BAC concentrations.
Peripheral blood lymphocytes (PBL):
In partnership with the French Blood Agency (FBA), lymphocytes were obtained from the peripheral
blood of healthy blood donors. FBA performed plateletpheresis on these donors and the remains of
this procedure (cytapheresis concentrate) were used in order to separate peripheral blood lymphocytes
from red blood cells. These two cellular types were separated by centrifugation at 400g for 30 minutes
on a Ficoll density gradient (Sigma Chemical Inc, St Louis, MO, USA) to isolate mononuclear cells.
PBL were washed in phosphate buffer saline (PBS), centrifuged at 300g for 15 minutes, and washed
again in PBS, then centrifuged at 200g for 10 minutes. PBL were maintained in RPMI (Roswell Park
Memorial Institute medium) 1640 medium supplemented with 10% FBS and cultured under standard
conditions (humidified atmosphere of 5% CO2 at 37 °C) for 12 hours. Supernatant was removed and
washed twice.
Cell treatments:
For the in vitro activation experiments, lymphocytes were incubated with 2.5ng/ml of phorbol 12-
myristate 13-acetate (PMA, Sigma, St Louis, MO, USA) and 250ng/ml of Ionomycin (Sigma) at 37°C
for 2 hours. After stimulation, cells were washed three times with RPMI. WKD were treated with
Trichostatin A (Trichostatin A, Ready Made Solution 5mM in DMSO [0.2μm-filtered], from
Streptomyces sp., Sigma) at 0.33M during 24 hours under standard conditions (humidified
atmosphere of 5% CO2 at 37°C) in complete culture medium (DMEM supplemented with 10% FBS,
1% glutamine, 50 UI/ml penicillin, and 50 UI/ml streptomycin) in order to induce E-cadherin
expression on WKD cells (49).
According to previous toxicological studies in the same cell line (50), subconfluent cells (culture
surface covering nearly 70%) were then exposed to five different concentrations of BAC (10−2
%,
5.10−3
%, 10−3
%, 10−4
%, and 10−5
%) or PBS during 15 minutes followed by 24 hours of cell recovery

64
in complete culture medium for evaluation of cell viability and cytotoxic effect of BAC in the
coculture model. These BAC concentrations are equivalent to or lower than the concentrations of most
available eye drops. Concentrations of 10−1
% and higher were not tested as they are known to be
excessively cytotoxic on conjunctival cells in vitro, inducing cell lysis immediately after treatment.
The incubation time applied to the WKD (or in coculture with PBL) was 15 min followed by 24 h of
cell recovery in complete medium. The 24-h cell recovery period was tested to approach the clinical
conditions in which the conjunctival tissue may be covered after eye drop instillation.
Coculture:
PBL (1ml of 106cells/ml) were cocultured with confluent epithelial WKD. PBL were cultured either in
direct contact with the epithelial WKD or separated from the epithelial cells by a Transwell (Cell
culture inserts, translucent PET membrane, 0.4m, BD Falcon, BD Biosciences), which prevents
direct contact between cells but allows free interchanges of culture media between the epithelial cells'
and lymphocytes' compartments. On day 1 and 7, the lymphocytes were collected and suspended in
2ml of culture medium (RPMI supplemented with 10% FBS) and analyzed by flow cytometry.
Flow cytometry:
The inflammatory marker HLA DR (Human Leucocyte Antigen Class II antigen), the apoptosis-
related receptor Fas (APO-1, CD95), the co-stimulating marker CD86, the HML-1/CD103 lymphocyte
antigen and the adhesion molecule E-cadherin were tested on Chang conjunctival cells. All
experiments were performed using a Cytomics FC500-CXP flow cytometer (Beckman Coulter,
Miami, FL) equipped with an argon laser emitting at 488 nm. Isotype matched IgG mouse conjugated
to FITC/PE/PC5 were used as negative control on human leucocytes to determine non-specific
fluorescence background.
Alteration of DNA content was measured through the sub-G1 peak flow cytometric analysis on a FL4-
fluorescence histogram to characterize the late apoptosis process. After coculture between
lymphocytes and epithelial cells pre-treated by either BAC or PBS (phosphate buffer saline), cells
were collected using ethylenediaminetetraacetic acid (EDTA) at 1mM, collected, and suspended in
1ml PBS, then fixed with 0.5% paraformaldehyde (PFA) in PBS at 4 °C for 24 h. Then, samples were
washed with cold PBS, permeabilized in 0.1% saponin, stained with 50μg/ml propidium iodide (PI,
Interchim, Montluçon, France), and analyzed on the flow cytometer. The sub-G1 region was
determined by a cursor defined in the controls.
Microplate cytofluorometry:
Microplate cytofluorometry was performed on a Safire® microplate cytometer (Tecan Instruments,
Lyon, France). Hoechst33342 dye was used to evaluate WKD apoptosis, and Neutral Red (NR) dye
(Fluka, Ronkonkoma, NY) was used to evaluate membrane integrity, closely correlated with cellular
viability. In all experiments, sterile PBS was used as the control. Microplate cytofluorometry results
were obtained in fluorescence units and were expressed as a percentage of the control PBS.
Hoechst33342 results were expressed as a ratio of the results of this assay to those of the NR assay in
order to correlate them to cellular viability.
Cell morphology analyses:
Standard immunofluorescence using fluorescent phalloidin (F-actin detection, 200U/ml; Alexa Fluor
488 Phalloidin, Sigma, St Louis, MO, USA) and lipophilic green-emitting dye PKH67 and lipophilic
red-emitting dye PKH26 stainings under a confocal epifluorescence microscope (E800, PCM 2000;
Nikon, Tokyo, Japan) were performed to assess morphologic patterns of cells. After 30-min
incubation with phalloidin, cells were washed in PBS before PI staining and microscopy examination.
WKD were stained with PKH67 (Green Fluorescent Cell Linker Kit, Sigma), and PBL with PKH26
(Red Fluorescent Cell Linker Kit, Sigma). For PKH stainings, WKD and PBL (20 millions each) were
washed either in DMEM supplemented 1% glutamine, 50 UI/ml penicillin, and 50 UI/ml streptomycin
culture medium without FBS for WKD cells, or in RPMI1640 for PBL, at 400g for 5 minutes and then
stained during 4 minutes with PKH at 4M. Then, stained cells were washed 3 times in D-PBS and
maintained in their own complete culture medium.
Scanning Electron Microscopy:
A specific method was performed to assess cellular interactions using Scanning Electron Microscopy
(SEM). Cells were fixed in 2.5% glutaraldehyde (Electron Microscopy Science, Hatfield, USA) for 45
minutes, then in 2% reduced osmium OsO4 (Electron Microscopy Science, Hatfield, USA) for 45
minutes as well. After the two fixation processes, cells were dehydrated in ethanol, cleared in

65
propylene oxide and embedded in Hexamethyldisilane (Sigma). Then, cells were sputter-coated with
gold-palladium using JEOL FINE COAT ion sputter JFC-1100 and were examined with a JEOL 35CF
Scanning Electron Microscope (Hitachi
, Japan).
Reagents:
Propidium iodide, PBS, PKH26 and 67 stainings were provided by Sigma (St Louis, MO, USA).
Monoclonal mouse anti-human CD103 (Mucosa Lymphocyte Antigen, clone Ber-ACT8, FITC-
conjugated), FITC-conjugated goat anti-mouse and phycoerythrin (PE)-conjugated rabbit anti-mouse
antibodies were obtained from DAKO (Glostrup, Denmark).
Monoclonal mouse anti-HLA-DR (Clone Immu-357, FITC-conjugated) and monoclonal mouse anti-
CD86 (Clone HA5.2B7, IgG2b purified) were provided by Coulter Corporation (Miami, FL, USA).
Control antibodies (purified and FITC-conjugated mouse IgG1) were purchased from Immunotech
(Marseilles, France).
The fluorescent probe phalloidin Alexa Fluor 488-conjugated was provided by Molecular Probes
(Leiden, The Netherlands).
All reagents were used as recommended by suppliers.
DISCLOSURE
A.Chassignol, none; E. Brasnu, none, ;C. Baudouin, none ; L. Riancho, none ;J.-M. Warnet, none
and F. Brignole-Baudouin, none.
REFERENCES 1. Green, K., Chapman, J. M., Cheeks, L., Clayton, R. M., Wilson, M., Zehir, A., Detergent penetration
into young and adult rabbit eyes: comparative pharmacokinetics. Toxicol Cut Ocul Toxicol. 1987 ;6 :89-
107.
2. Baudouin, C., Labbé, A., Liang, H., Pauly, A., Brignole-Baudouin, F., Preservatives in eyedrops : The
good, the bad and the ugly. Prog Retin Eye Res. 2010 Jul ;29(4) :312-34.
3. Mietz, H., Niesen, U., Krieglstein, G. K., The effects of preservatives and antiglaucomatous medication
on histopathology of the conjunctiva. Graefes Arch Clin Exp Ophthalmol. 1994 ;232 :56l-565.
4. De Saint Jean, M., Brignole, F., Bringuier, A. F., Bauchet, A., Feldmann, G., Baudouin, C., Effects of
benzalkonium chloride on growth and survival of Chang conjunctival cells. Invest Ophthalmol Vis Sci.
1999 ; 40 :619-30.
5. El Annan, J., Chauhan, S. K., Ecoiffier, T., Zhang, Q., Saban, D. R., Dana, R., Characterization of
effector T cells in dry eye disease. Invest Ophthalmol Vis Sci. 2009 Aug ;50(8) :3802-7.
6. Brignole, F., De Saint Jean, M., Becquet, F., Goldschild, M., Goguel, A., Baudouin, C., Expression of
Fas-Fas ligand antigens and apoptotic marker APO2·7 by the human conjunctival epithelium. Positive
correlation with class II HLA DR expression in inflammatory conditions. Exp. Eye Res. 1998 ;67 :687–
697
7. De Saint Jean, M., Brignole, F., Goguel, A., Baudouin, C., Interferon gamma induces apoptosis and
expression of inflammationrelated proteins in Chang conjunctival cells. Invest Ophthalmol Vis Sci.
1999 ;40 :2199–2212.
8. Lavappa, K. S., Macy, M. L., Shannon, J. E., Examination of ATCC stocks for HeLa marker
chromosomes in human cell lines. Nature 1976 ;259(5540) :211-213
9. Diebold, Y., et al., Characterization of a spontaneously immortalized cell line (IOBA-NHC) from
normal human conjunctiva. Invest Ophthalmol Vis Sci. 2003 ;44: 4263-74.
10. Chang, R. S., Continuous subcultivation of epithelial-like cells from normal human tissues. Proc Soc
Exp Biol Med. 1954 ;87(2) :440-3.
11. De Saint Jean, M., et al., Comparison of morphological and functional characteristics of primary-
cultured human conjunctival epithelium and of Wong-Kilbourne derivative of Chang conjunctival cell
line. Exp Eye Res. 2004 ;78 :257-74.
12. Guenoun, J. M., C. Baudouin, Rat, P., Pauly, A., Warnet, J. M., Brignole-Baudouin, F., In vitro study of
inflammatory potential and toxicity profile of latanoprost, travoprost, and bimatoprost in conjunctiva-
derived epithelial cells. Invest Ophthalmol Vis Sci. 2005; 46(7) :2444-50.
13. Pisella, P. J., et al., Conjunctival proinflammatory and proapoptotic effects of latanoprost and preserved
and unpreserved timolol: an ex vivo and in vitro study. Invest Ophthalmol Vis Sci. 2004 ;45(5) :1360-8.
14. Ryan, C. A., Kimber, I., Basketter, D. A., Pallardy, M., Gildea, L. A., Gerberick, G. F., Dendritic cells
and skin sensitization: biological roles and uses in hazard identification. Toxicol Appl Pharmacol. 2007
Jun 15 ;221(3) :384-94.

66
15. Fayol-Messaoudi, D., Coconnier-Polter, M. H., Moal, V. L., Atassi, F., Berger, C. N., Servin, A.L., The
Lactobacillus plantarum strain ACA-DC287 isolated from a Greek cheese demonstrates antagonistic
activity in vitro and in vivo against Salmonella enterica serovar Typhimurium. J Appl Microbiol. 2007
Sep ;103(3) :657-65
16. Upham, J. W., Stick, S. M., Interactions between airway epithelial cells and dendritic cells: implications
for the regulation of airway inflammation. Curr Drug Targets. 2006 May ;7(5) :541-5
17. Wang, Y., et al., BRS-3 activation transforms the effect of human bronchial epithelial cells from PGE2
mediated inhibition to TGF-beta1 dependent promotion on proliferation and collagen synthesis of lung
fibroblasts. Cell Biol Int. 2007 Dec ;31(12) :1495-1500.
18. Gomes, J. A., Dua, H. S., Rizzo, L. V., Nishi, M., Joseph, A., Donoso, L. A., Ocular surface epithelium
induces expression of human mucosal lymphocyte antigen (HML-1) on peripheral blood lymphocytes.
Br J Ophthalmol. 2004 ;88(2) :280-5.
19. Brasnu, E., Brignole-Baudouin, F., Riancho, L., Warnet, J. M., Baudouin, C., Comparative study on the
cytotoxic effects of benzalkonium chloride on the Wong-Kilbourne derivative of Chang conjunctival
and IOBA-NHC cell lines. Mol Vis. 2008 ;14 :394-402.
20. Dua, H. S., Gomes, J. A., Donoso, L. A., Laibson, P. R., The ocular surface as part of the mucosal
immune system: conjunctival mucosa-specific lymphocytes in ocular surface pathology. Eye. 1995 ;9(pt
3) :261-7.
21. Dua, H. S., et al., Mucosa specific lymphocytes in the human conjunctiva, corneoscleral limbus and
lacrimal gland. Curr Eye Res. 1994 Jan ;13(1) :87-93.
22. Sacks, E. H., Wieczorek, R., Jakobiec, F. A., Knowles, D. M., Lymphocytic subpopulations in the
normal human conjunctiva. A monoclonal antibody study. Ophthalmology. 1986 Oct ;93(10) :1276-83.
23. Franklin, R. M., Remus, L. E., Conjunctival-associated lymphoid tissue: evidence for a role in the
secretory immune system. Invest Ophthalmol Vis Sci. 1984 Feb ;25(2) :181-7.
24. Iwata, M., Kiritoshi, A., Roat, M. I., Yagihashi, A., Thoft, R. A., Regulation of HLA class II antigen
expression on cultured corneal epithelium by interferon-gamma. Invest Ophthalmol Vis Sci. 1992 Aug
;33(9) :2714-21.
25. Shams, N. B., Huggins, E. M. Jr., Sigel, M. M., Interleukin-1 regulates the proliferation of leukocytes in
human corneal cell-peripheral blood leukocyte cocultures. Cornea. 1994 Jan ;13(1) :9-15.
26. Iwata, M., Yagihashi, A., Roat, M. I., Zeevi, A., Iwaki, Y., Thoft, R. A., Human leukocyte antigen-class
II-positive human corneal epithelial cells activate allogeneic T cells. Invest Ophthalmol Vis Sci. 1994
Nov ;35(12) :3991-4000.
27. Wanner, R., et al., Classification of sensitizing and irritative potential in a combined in-vitro assay.
Toxicol Appl Pharmacol. 2010 ;245 :211-8.
28. Swanson, J. A. & Watts, C., Macropinocytosis. Trends Cell Biol. 1995 ;5(11) :424-8.
29. Racoosin, E. L., & Swanson, J. A., M-CSF-induced macropinocytosis increases solute endocytosis but
not receptor-mediated endocytosis in mouse macrophages. J Cell Sci. 1992 ;102 :867-80.
30. Fiorentini, C., et al., Activation of rho GTPases by cytotoxic necrotizing factor 1 induces
macropinocytosis and scavenging activity in epithelial cells. Mol Biol Cell. 2001 ;12(7) :2061-73.
31. Hewlett, L.J., Prescott, A. R., Watts, C., The coated pit and macropinocytotic pathways serve distinct
endosome populations. J Cell Biol. 1994 ;124(5) :689-703.
32. De Saint Jean, M., et al., Fas- and interferon gamma-induced apoptosis in Chang conjunctival cell line.
Further investigations. Invest Ophthalmol Vis Sci 2000 ;41 :2531–2543.
33. Geppert, T. D., Lipsky, P. E., Antigen presentation by interferon-gamma-treated endothelial cells and
fibroblasts: differential ability to function as antigen-presenting cells despite comparable Ia expression.
J Immunol. 1985 Dec ;135(6) :3750-62.
34. Young, E., Stark, W. J., In vitro immunological function of human corneal fibroblasts. Invest
Ophthalmol Vis Sci. 1988 Sep ;29(9) :1402-6.
35. Gomes, J. A., et al. Mechanism of expression of the human mucosal lymphocyte antigen (HML-1) in
conjunctiva associated lymphoid tissue (CALT). Invest Ophthalmol Vis Sci. 1995 ;36 :S840.
36. Poussier, P., & Julius, M., Thymus independent T cell development and selection in the intestinal
epithelium. Annu Rev Immunol. 1994 ;12 :521-53.
37. Brandtzaeg, P., et al. Lymphoepithelial interactions in the mucosal immune system. Gut. 1988 ;29(8)
:1116-30.
38. Jarry, A., Cerf-Bensussan, N., Brousse, N., Selz, F., Guy-Grand, D., Subsets of CD3+ (T cell receptor
alpha/beta or gamma/delta) and CD3- lymphocytes isolated from normal human gut epithelium display
phenotypical features different from their counterparts in peripheral blood. Eur J Immunol. 1990 ;20(5)
:1097-103.

67
39. Cerf-Bensussan, N., Jarry, A., Brousse, N., Lisowska-Grospierre, B., Guy-Grand D., Griscelli, C., A
monoclonal antibody (HML-1) defining a novel membrane molecule present on human intestinal
lymphocytes. Eur J Immunol. 1987 ;17(9) :1279-85.
40. Cepek, K. L., et al., Adhesion between epithelial cells and T lymphocytes mediated by E-cadherin and
the alpha E beta 7 integrin. Nature. 1994 ;372(6502) :190-3.
41. Karecla, P. I., Green, S. J., Bowden, S. J., Coadwell, J., Kilshaw, P. J., Identification of a binding site
for integrin alphaEbeta7 in the N-terminal domain of E-cadherin. J Biol Chem. 1996 ;271(48) :30909-
15.
42. Parker, C. M., et al., A family of beta 7 integrins on human mucosal lymphocytes. Proc Natl Acad Sci
USA. 1992 ;89(5) :1924-8.
43. Higgins, J. M., et al., Direct and regulated interaction of integrin alphaEbeta7 with E-cadherin. J Cell
Biol. 1998 ;140(1) :197-210.
44. Takeichi, M., Cadherins: a molecular family important in selective cell-cell adhesion. Annu Rev
Biochem. 1990 ;59 :237-52.
45. Schieferdecker, H. L., et al., The HML-1 antigen of intestinal lymphocytes is an activation antigen. J
Immunol. 1990 ;144 :2541–9.
46. Lu, Z., Ghosh, S., Wang, Z., Hunter, T., Downregulation of caveolin-1 function by EGF leads to the
loss of E-cadherin, increased transcriptional activity of beta-catenin, and enhanced tumor cell invasion.
Cancer Cell. 2003 ;4(6) :499-515.
47. Verschueren, K., et al., SIP1, a novel zinc finger/homeodomain repressor, interacts with Smad proteins
and binds to 5'-CACCT sequences in candidate target genes. J Biol Chem. 1999 ;274(29) :20489-98.
48. Chinnadurai, G., CtBP, an unconventional transcriptional corepressor in development and oncogenesis.
Mol Cell. 2002 ;9(2) :213-24.
49. Ohira, T., et al., WNT7a induces E-cadherin in lung cancer cells. Proc Natl Acad Sci USA. 2003
;100(18) :10429-34.
50. Debbasch, C., Brignole, F., Pisella, P. J., Warnet, J. M., Rat, P., Baudouin, C., Quaternary ammoniums
and other preservatives' contribution in oxidative stress and apoptosis on Chang conjunctival cells.
Invest Ophthalmol Vis Sci. 2001 ;42 :642-52.
FIGURE LEGENDS
Figure 1. Cellular morphology and interaction analyses in cocultures using cell membrane staining.
Epithelial cells were stained in green with PKH67 and lymphocytes in red with PKH26. This figure
shows the same area using either green filter or red filter under fluorescence microscopy. WKD cells
were cocultured with PBL in direct contact during 24 hours. Plasmic membrane exchanges between
both cell types were observed after 1 day of coculture only if epithelial cells were in direct contact
with peripheral blood lymphocytes, because we noted that WKD cells appeared green and red,
meaning a staining transfer between both cellular types occurred. Controls (data not shown), either
WKD cells alone or WKD cells cocultured with PBL separated by a 0.4m hole translucent PET
membrane, show WKD cells exclusively green, without any transfer of red staining by PBL to WKD
cells. Yellow staining is due to an overexposure to fluorescence microscopy. WKD, Wong Kilbourne-
Derivative of Chang; PBL, Peripheral Blood Lymphocytes.
Figure 2. Cellular morphology and interaction analyses in coculture using cell membrane staining.
Epithelial cells were stained with PKH67 and lymphocytes with PKH26. This figure shows the same
area (A and C) using either green filter (A) or red filter (C) under Confocal microscopy. B and D show
the same area in a vertical section (Two-head arrow). WKD cells were cocultured with PBL in direct
contact during 90 minutes. Plasmic membrane exchanges between both cell types were observed after
90 minutes coculture while epithelial cells were in direct contact with peripheral blood lymphocytes.
Indeed, note that red staining started to appear in WKD cells (arrows).
Figure 3. Cellular morphology and interaction analyses in coculture using scanning electron
microscopy. Note that tight connections between PBL (little arrow) and WKD cells looked like cell
membrane fusion (arrows).
Figure 4. Cell morphology analysis in coculture using cell membrane staining. Standard
immunofluorescence staining on WKD cells and PBL cells. Phalloidin staining shows the morphology

68
of the two cell types after 24 hours coculture. Cytoskeleton (F-actin) was stained in green by
phalloidin. Note that PBL showed cytoplasmic expansions (arrow), in contact with WKD cells.
Figure 5. Alteration of DNA content analysis on WKD cells. The alteration of DNA content was
measured through the sub-G1 peak flow analysis (flow cytometry) on WKD cells. Successively, WKD
cells were cultured alone in normal complete medium, then cocultured 24 hours with PBL separated
by a Transwell, and with PBL in direct contact. Next, WKD cells pre-treated with 5.10-3% BAC for
15 minutes followed by 24 h of cell recovery were cultured alone in normal complete medium, then
cocultured 24 hours with PBL separated by a Transwell or in direct contact. Note the potentiated
apoptotic effect of contact with PBL on pre-treated WKD cells (arrow : P < 0.001).
Figure 6. Graph representing a flow cytometric analysis of expression of E-cadherin at baseline and
after Trichostatin-A treatment. Mean fluorescence intensities were quantified and expressed as % of
fluorescence. WKD cells were treated by Trichostatin A (TSA) at 0.33M during 24 hours under
standard conditions in complete culture medium. Note a positive expression of E-cadherin on WKD
cells after TSA treatment ( arrow : P < 0.001).
Figure 7. Laser scanning confocal micrographs showing immunolabeling pattern for E-cadherin on
WKD cells. Nuclei were counterstained with DAPI. (photo on left) WKD cells at baseline cultured in
normal medium. (photo on right) Trichostatin-A Pre-treated WKD cells. Note the increased expression
of E-cadherin on TSA pre-treated WKD cells (arrow).
Figure 8. Graph representing a flow cytometric analysis of expression of HLA-DR inflammatory
marker on WKD cells at baseline and after 24 hours or 7 days of coculture with PBL, pre treated by
PMA and Ionomycin (B) or not (A), and by Transwell or in direct contact. Mean fluorescence
intensities were quantified and expressed as % of fluorescence report to control. Lymphocytes were
incubated with 2.5 ng/ml of PMA and 250 ng/ml of Ionomycin at 37°C for 2 hours. Note the
significantly positive expression of HLA-DR on WKD cells after 24 hours coculture with non-
activated PBL in direct contact and after 7 days coculture with activated PBL in Transwell ( arrow : P
< 0.03). PMA, phorbol 12-myristate 13-acetatein.
Figure 9. Graph representing a flow cytometric analysis of expression of CD103 on PBL after 24
hours coculture with WKD cells, pre-treated by TSA or not. Mean fluorescenceintensities were quan
tified and expressed as % of fluorescence. Note the trend to a positive expression of CD 103 on PBL
after 24 hours coculture with TSA pre-treated WKD cells ( arrow : P < 0.001).
Figure 10. Toxicity evaluation using HOECHST and neutral red tests on WKD cells. This figure
shows the toxicity evaluation using the apoptotic test HOECHST report to the integrity membrane
evaluation test called neutral red test (microplate cytofluorometry) on WKD cells after 15 min of
treatment with different BAC concentrations followed by 24 h of cell recovery in complete medium or
in coculture with non activated PBL. Note that toxicity increased on WKD cells in coculture with PBL
for lower concentration of BAC (less than 10-2
%) and decreased on WKD cells in coculture with PBL
for higher concentrations of BAC, in comparison with toxicity on WKD cells cultured alone (arrow : P
< 0.001).

69
VII. REFERENCES BIBLIOGRAPHIQUES
1. Vinardell MP, Mitjans M. Alternative methods for eye and skin irritation tests: an
overview. J Pharm Sci. 2008 Jan ; 97(1) : 46-59.
2. Mc Cally AW, Farmer AG, Loomis EC. Corneal ulceration following use of lash lure.
J Am Med Assoc. 1933 ; 101 : 1160-1161.
3. Friedenwald JS, Hughes W, Herrman H. Acid-base tolerance of the cornea. Arch
Ophthalmol. 1944 ; 31 : 279-283.
4. Draize JH, Woodard G, Calvery HO. Methods for the study of irritation and
toxicology of substances applied topically to the skin and mucous membranes. J
Pharmacol Exp Ther. 1944 ; 83 : 377-390.
5. Smyth DH. Alternatives to Animal Experiments. Cambridge : Scolar Press. 1978.
6. Chibret H. Conservateurs et préparation ophtalmiques : réalités et perspectives. Ann
Pharmaceutiques Françaises. 1997 ; 55 : 228e231.
7. Rosenthal RA, Buck SL, Henry CL, Schlech BA. Evaluation of the preserving
efficacy of lubricant eyedrops with a novel preservative system. J Ocul Pharmacol
Ther. 2006 ; 22 : 440e448.
8. EMEA. Public statement on antimicrobial preservatives in ophthalmic preparations for
human use. London 8 December 2009 ; EMEA/622721/2009.
9. Liu H, Routley I, Teichmann KD. Toxic endothelial cell destruction from intraocular
benzalkonium chloride. J Cataract Refract Surg. 2001 Nov ; 27(11) : 1746-50.
10. Oiso N, Fukai K, Ishii M. Irritant contact dermatitis from benzalkonium chloride in
shampoo. Contact Dermatitis. 2005 Jan ; 52(1) : 54.
11. Charnock, C. Are multidose over-the-counter artificial tears adequately preserved ?
Cornea. 2006 ; 25 : 432e437.
12. Kim MS, Choi CY, Kim JM, Chang HR, Woo HY. Microbial contamination of
multiply used preservative-free artificial tears packed in reclosable containers. Br J
Ophthalmol. 2008 ; 92 : 1518e1521.
13. Tasli H, Cosar G. Microbial contamination of eyedrops. Cent Eur J Public Health.
2001 ; 9 : 162e164.
14. Pawar PK, Majumdar DK. Effect of formulation factors on in vitro permeation of

70
moxifloxacin from aqueous drops through excised goat, sheep, and buffalo corneas.
AAPS Pharm Sci Tech. 2006 ; 7 : E13.
15. Yenice I, Mocan MC, Palaska E, Bochot A, Bilensoy E, Vural I, Irkec M, Hincal AA.
Hyaluronic acid coated poly-epsilon-caprolactone nanospheres deliver high
concentrations of cyclosporine A into the cornea. Exp Eye Res. 2008 ; 87 : 162e167.
16. Majumdar S, Hippalgaonkar K, Repka MA. Effect of chitosan, benzalkonium chloride
and ethylenediaminetetraacetic acid on permeation of acyclovir across isolated rabbit
cornea. Int J Pharm. 2008 ; 348 : 175e178.
17. Rathore MS, Majumdar DK. Effect of formulation factors on in vitro trans-corneal
permeation of gatifloxacin from aqueous drops. AAPS Pharm Sci Tech. 2006 ; 7 : 57.
18. Pellinen P, Lokkila J. Corneal penetration into rabbit aqueous humoris comparable
between preserved and preservative-free tafluprost. Ophthalmic Res. 2009 ; 41 :
118e122.
19. De Jong C, Stolwijk T, Kuppens E, De Keizer R, Van Best J. Topical timolol with and
without benzalkonium chloride : epithelial permeability and autofluorescence of the
cornea in glaucoma. Graefes Arch Clin Exp Ophthalmol. 1994 ; 232 : 221e224.
20. Van der Valk R, Webers CA, Schouten JS, Zeegers MP, Hendrikse F, Prins MH.
Intraocular pressure-lowering effects of all commonly used glaucoma drugs: a meta-
analysis of randomized clinical trials. Ophthalmology. 2005 ; 112 : 1177e1185.
21. Pisella PJ, Pouliquen P, Baudouin C. Prevalence of ocular symptoms and signs with
preserved and preservative-free glaucoma medication. Br J Ophthalmol. 2002 ; 86 :
418e423.
22. Jaenen N, Baudouin C, Pouliquen P, Manni G, Figueiredo A, Zeyen T. Ocular
symptoms and signs with preserved and preservative-free glaucoma medications. Eur
J Ophthalmol. 2007 ; 17 : 341e349.
23. Erb C, Gast U, Schremmer D. German register for glaucoma patients with dry eye. I.
Basic outcome with respect to dry eye. Graefes Arch Clin Exp. Ophthalmol. 2008 ;
246 : 1593e1601.
24. Dry Eye Work Shop. The definition and classification of dry eye disease : report of the
Definition and Classification Subcommittee of the International Dry Eye Work Shop.
Ocul Surf. 2007 ; 5 : 75e92.

71
25. Zimmerman TJ, Hahn SR, Gelb L, Tan H, Kim EE. The impact of ocular adverse
effects in patients treated with topical prostaglandin analogs : changes in prescription
patterns and patient persistence. J Ocul Pharmacol Ther. 2009 ; 25 : 145e152.
26. Baudouin C, Pisella PJ, Fillacier K, Goldschild M, Becquet F, De Saint Jean M,
Bechetoille A. Ocular surface inflammatory changes induced by topical antiglaucoma
drugs : human and animal studies. Ophthalmology. 1999 ; 106 : 556e563.
27. Thorne JE, Anhalt GJ, Jabs DA. Mucous membrane pemphigoid and pseudo
pemphigoid. Ophthalmology. 2004 ; 111 : 45e52.
28. Lavin MJ, Wormald RP, Migdal CS, Hitchings RA. The influence of prior therapy on
the success of trabeculectomy. Arch Ophthalmol. 1990 ; 108 : 1543e1548.
29. Broadway DC, Grierson I, O‟Brien C, Hitchings RA. Adverse effects of topical
antiglaucoma medication. II. The outcome of filtration surgery. Arch. Ophthalmol.
1994 ; 112 : 1446e1454.
30. Sherwood MB, Grierson I, Millar L, Hitchings RA. Long-term morphologic effects of
antiglaucoma drugs on the conjunctiva and Tenon‟s capsule in glaucomatous patients.
Ophthalmology. 1989 ; 96 : 327e335.
31. Brignole-Baudouin F, Ott AC, Warnet JM, Baudouin C. Flow cytometry in
conjunctival impression cytology : a new tool for exploring ocular surface pathologies.
Exp Eye Res. 2004 ; 78 : 473e481.
32. Brandt JD, Wittpenn JR, Katz LJ, Steinmann WN, Spaeth GL. Conjunctival
impression cytology in patients with glaucoma using long-term topical medication.
Am J Ophthalmol. 1991 ; 112 : 297e301.
33. Baudouin C, Garcher C, Haouat N, Bron A, Gastaud P. Expression of inflammatory
membrane markers by conjunctival cells in chronically treated patients with glaucoma.
Ophthalmology. 1994 ; 101 : 454e460.
34. Baudouin C, Hamard P, Liang H, Creuzot-Garcher C, Bensoussan L, Brignole F.
Conjunctival epithelial cell expression of interleukins and inflammatory markers in
glaucoma patients treated over the long term. Ophthalmology. 2004 ; 111 :
2186e2192.
35. Baudouin C, Liang H, Bremond-Gignac D, Hamard P, Hreiche R, Creuzot- Garcher C,
Warnet JM, Brignole-Baudouin F. CCR4 and CCR5 expression in conjunctival

72
specimens as differential markers of T(H)1/T(H)2 in ocular surface disorders. J
Allergy Clin Immunol. 2005 ; 116 : 614e619.
36. Baudouin C, Liang H, Hamard P, Riancho L, Creuzot-Garcher C, Warnet JM,
Brignole-Baudouin F. The ocular surface of glaucoma patients treated over the long
term expresses inflammatory markers related to both T-helper1 and T-helper2
pathways. Ophthalmology. 2008 ; 115 : 109e115.
37. Rodrigues M de L, Felipe Crosta DP, Soares CP, Deghaide NH, Duarte R, Sakamoto
FS, Furtado JM, Paula JS, Donadi EA, Soares EG. Immunohistochemical expression
of HLA-DR in the conjunctiva of patients under topical prostaglandin analogs
treatment. J Glaucoma. 2009 ; 18 : 197e200.
38. Calvin G. New approaches to the assessment of eye and skin irritation. Toxicol Lett.
1992 ; 64-65 Spec No : 157-64.
39. Prinsen MK, Koëter HB. Justification of the enucleated eye test with eyes of
slaughterhouse animals as an alternative to the Draize eye irritation test with rabbits.
Food Chem Toxicol. 1993 ; 31(1) : 69-76.
40. Curren RD, Harbell JW. Ocular safety : a silent (in vitro) success story. Altern Lab
Anim. 2002 ; 30(Suppl.2) : 69e74.
41. Wilhelmus KR. The Draize eye test. Surv Ophthalmol. 2001 ; 45 : 493e515.
42. Ichijima H, Petroll WM, Jester JV, Cavanagh HD. Confocal microscopic studies of
living rabbit cornea treated with benzalkonium chloride. Cornea1. 1992 ; 1 : 221e225.
43. Labbe A, Dupas B, Hamard P, Baudouin C. In vivo confocal microscopy study of
blebs after filtering surgery. Ophthalmology. 2005 ; 112 : 1979e1986.
44. Labbe A, Nicola RD, Dupas B, Auclin F, Baudouin C. Epithelial basement membrane
dystrophy : evaluation with the HRT II Rostock Cornea Module. Ophthalmology.
2006 ; 113 : 1301e1308.
45. Labbe A, Niaudet P, Loirat C, Charbit M, Guest G, Baudouin C. In vivo confocal
microscopy and anterior segment optical coherence tomography analysis of the cornea
in nephropathic cystinosis. Ophthalmology. 2009 ; 116 : 870e876.
46. Liang H, Baudouin C, Dupas B, Brignole-Baudouin F.Live conjunctiva-associated
lymphoid associated tissue (CALT) analysis in rabbit using in vivo confocal
microscopy (IVCM) under inflammatory stimuli. Invest Ophthalmol Vis Sci. 2009 ;

73
(Epubaheadofprint).
47. Liang H, Baudouin C, Pauly A, Brignole-Baudouin F. Conjunctival and corneal
reactions in rabbits following short- and repeated exposure to preservative-free
tafluprost, commercially available latanoprost and 0.02%benzalkonium chloride. Br J
Ophthalmol. 2008 ; 92 : 1275e1282.
48. Labbe A, Pauly A, Liang H, Brignole-Baudouin F, Martin C, Warnet JM, Baudouin C.
Comparison of toxicological profiles of benzalkonium chloride and polyquaternium-1
: an experimental study. J Ocul Pharmacol Ther. 2006 ; 22 : 267e278.
49. Pauly A, Brignole-Baudouin F, Labbe A, Liang H, Warnet JM, Baudouin C. New
tools for the evaluation of toxic ocular surface changes in the rat. Invest Ophthalmol
Vis Sci. 2007 ; 48 : 5473e5483.
50. Pauly A, Labbe A, Baudouin C, Liang H, Warnet JM, Brignole-Baudouin F. In vivo
confocal microscopic grading system for standardized corneal evaluation: application
to toxic-induced damage in rat. Curr Eye Res. 2008 ; 33 : 826e838.
51. Ishibashi T, Yokoi N, Kinoshita S. Comparison of the short-term effects on the human
corneal surface of topical timolol maleate with and without benzalkonium chloride. J
Glaucoma. 2003 ; 12 : 486e490.
52. Baudouin C, de Lunardo C. Short-term comparative study of topical 2% carteolol with
and without benzalkonium chloride in healthy volunteers. Br J Ophthalmol. 1998 ;
82 : 39e42.
53. Rolando M, Brezzo V, Giordano G, Campagna P, Burlando S, Calabria G. The effect
of different benzalkonium chloride concentrations on human normal ocular surface. In
: Van Bijsterveld O, Lemp M, Spinelli D. The Lacrimal System (EDS.). 1991. Kugler
and Ghedini Publications, Amsterdam, Berkely, Milano.
54. Nuzzi R, Finazzo C, Cerruti A. Adverse effects of topical antiglaucomatous
medications on the conjunctiva and the lachrymal. Int Ophthalmol. 1998 ; 22 : 31e35.
55. Yalvac IS, Gedikoglu G, Karagoz Y, Akgun U, Nurozler A, Koc F, Kasim R, Duman
S. Effects of antiglaucoma drugs on ocular surface. Acta Ophthalmol Scand. 1995 ;
73 : 246e248.
56. Martone G, Frezzotti P, Tosi GM, Traversi C, Mittica V, Malandrini A, Pichierri P,
Balestrazzi A, Motolese PA, Motolese I, Motolese E.An in vivo confocal microscopy

74
analysis of effects of topical antiglaucoma therapy with preservative on corneal
innervation and morphology. Am J Ophthalmol. 2009 ; 147 : 725e735.
57. Rolle U, Chertin B, Puri P. Effects of benzalkonium chloride treatment on the intra
mural innervation of the upper urinary tract. B J U Int. 2003 ; 91 : 683e686.
58. Bron A, Chiambaretta F, Pouliquen P, Rigal D, Rouland JF. Efficacy and safety of
substituting a twice-daily regimen of timolol with a single daily instillation of non
preserved beta-blocker in patients with chronic glaucoma or ocular hypertension. J Fr
Ophtalmol. 2003 ; 26 : 668e674.
59. Campagna P, Macri A, Rolando M, Calabria G. Chronic topical eye preservative-free
beta-blocker therapy effect on the ocular surface in glau- comatous patients. Acta
Ophthalmol Scand Suppl. 1997 ; 224 : 53.
60. Foster Cs. Cicatricial pemphigoid. Trans Am Ophthalmol Soc 1986 ; 84 : 527-663.
61. Sherwood MB, Grierson I, Millar L, Hitchings RA. Long-term morphologic effects of
antiglaucoma drugs on the conjunctiva and Tenon‟s capsule in glaucomatous patients.
Ophthalmology. 1989 ; 96 : 327-335.
62. Broadway DC, Grierson I, Hitchings RA. Adverse effects of topical antiglaucoma
medications on the conjonctiva. Br J Ophtalmol. 1993 ; 77 : 590-596.
63. Broadway DC, Grierson I, O‟Brien C, Hitchings RA. Adverse effects of topical
antiglaucoma medications. I. The conjunctival cell profile. Arch Ophthalmol. 1994 ;
112 : 1437-1445.
64. Broadway DC, Grierson I, O‟Brien C, Hitchings RA. Adverse effects of topical
antiglaucoma medications. II. The outcome of filtration surgery. Arch Ophthalmol.
1994 ; 112 : 1446-1454.
65. Fujihara T, Takeuchi T, Saito K, Kitajima Y, Kobayashi TK, Tsubota K. Evaluation of
human conjunctival epithelium by a combination of brush cytology and flow
cytometry: an approach to the quantitative technique. Diagn Cytopathol 1997;17:456-
460.
66. Wilson FM. Adverse external ocular effects of topical ophthalmic medications. Survey
Ophthalmol. 1979 ; 24 : 57-88.
67. Rolando M, Barabino S, Mingari C, Moretti S, Giuffrida S, Calabria G. Distribution of
conjunctival HLA-DR expression and the pathogenesis of damage in early dry eyes.

75
Cornea. 2005 ; 24 : 951-954.
68. Tsubota K, Takamura E, Hasegawa T, Kobayashi T. Detection by brush cytology of
mast cells and eosinophils in allergic and vernal conjunctivitis. Cornea. 1991 ; 10 :
525-531.
69. Egbert PR, Lauber S, Maurice DM. A simple conjunctival biopsy. Am J Ophthalmol.
1977 ; 84 : 798-801.
70. Thatcher RW, Darougar S, Jones BR. Conjunctival impression cytology. Arch
Ophthalmol. 1977 ; 95 : 678-681.
71. Liotet S, Van Bijsterveld OP, Bletry O, Chomette G, Moulias R, Arrata M. L‟œil sec.
Rapport de la Soc Fr Ophtalmol. Masson, Paris, 1987.
72. Labbé A, Brignole-Baudouin F, Baudouin C. Ocular surface investigations in dry eye
J Fr Ophtalmol. 2007 ; 30(1) : 76-97.
73. Brignole F, De Saint-Jean M, Goldschild M, Becquet F, Goguel A, Baudouin C.
Expression of Fas-Fas ligand antigens and apoptotic marker APO2.7 by the human
conjunctival epithelium. Positive correlation with class II HLA DR expression in
inflammatory ocular surface disorders. Exp Eye Res. 1998 Dec ; 67(6) : 687-97.
74. Nelson JD. Impression cytology. Cornea 1988 ; 7 : 71-81
75. Tseng SC. Staging of conjunctival squamous metaplasia by impression cytology.
Ophthalmology. 1985 ; 92 : 728-733.
76. Calonge M, Diebold Y, Saez V, et al. Impression cytology of the ocular surface : a
review. Exp Eye Res. 2004 ; 78 : 457-472.
77. Baudouin C, Brignole F, Becquet F, Pisella PJ, Goguel A. Flow cytometry in
impression cytology specimens. A new method for evaluation of conjunctival
inflammation. Invest Ophthalmol Vis Sci. 1997 ; 38(7) : 1458-64.
78. Pisella PJ, Debbasch C, Hamard P, Creuzot-Garcher C, Rat P, Brignole F, Baudouin
C. Conjunctival proinflammatory and proapoptotic effects of latanoprost and
preserved and unpreserved timolol: an ex vivo and in vitro study. Invest Ophthalmol
Vis Sci. 2004 ; 45(5) : 1360-8.
79. De Saint Jean M, Baudouin C, Di Nolfo M, Roman S, Lozato P, Warnet JM, Brignole
F. Comparison of morphological and functional characteristics of primary-cultured
human conjunctival epithelium and of Wong-Kilbourne derivative of Chang

76
conjunctival cell line. Exp Eye Res. 2004 ; 78 : 257-74.
80. Smit EE, Sra SK, Grabowski LR, Ward SL, Trocme SD. Modulation of IL-8 and
RANTES release in human conjunctival epithelial cells: primary cells and cell line
compared and contrasted. Cornea. 2003 May ; 22(4) : 332-7.
81. Sosne G, Albeiruti AR, Hollis B, Siddiqi A, Ellenberg D, Kurpakus-Wheater M.
Thymosin beta4 inhibits benzalkonium chloride-mediated apoptosis in corneal and
conjunctival epithelial cells in vitro. Exp Eye Res. 2006 ; 83(3) : 502-7.
82. De Saint Jean M, Brignole F, Goguel A, Baudouin C. Interferon gamma induces
apoptosis and expression of inflammationrelated proteins in Chang conjunctival cells.
Invest Ophthalmol Vis Sci. 1999 ; 40 : 2199–2212.
83. De Saint Jean M, Debbasch C, Rahmani M, Brignole F, Feldmann G, Warnet JM,
Baudouin C. Fas- and interferon gamma-induced apoptosis in Chang conjunctival
cells: further investigations. Invest Ophthalmol Vis Sci. 2000 ; 41(9) : 2531-43.
84. Green K, Chapman JM, Cheeks L, Clayton RM, Wilson M, Zehir A. Detergent
penetration into young and adult rabbit eyes: comparative pharmacokinetics. Toxicol
Cut Ocul Toxicol. 1987 ; 6 : 89 -107.
85. Grant RL, Acosta D. Delayed toxicity of benzalkonium chloride and sodium dodecyl
sulfate evaluated in primary cultures of rabbit corneal epithelial cells. Toxicol
Methods. 1994 ; 4 : 259-273.
86. Samples JR, Binder S, Nayak S. The effects of epinephrine and benzalkonium
chloride on cultured corneal and trabecular mesh- work cells. Exp Eye Res. 1989 ; 49 :
1-12.
87. Baudouin C, Labbé A, Liang H, Pauly A, Brignole-Baudouin F. Preservatives in
eyedrops: The good, the bad and the ugly. Prog Retin Eye Res. 2010 ; 29(4) : 312-34.
88. Mietz H, Niesen U, Krieglstein GK. The effects of preservatives and antiglaucomatous
medication on histopathology of the conjunctiva. Graefes Arch Clin Exp Ophthalmol.
1994 ; 232 : 56l-565.
89. Fisher AA. Allergic contact dermatitis and conjunctivitis from benzalkonium chloride.
Cutis. 1987 ; 39 : 381-383.
90. Goh CL. Contact sensitivity to topical antimicrobials, II: sensitizing potentials of some
topical antimicrobials. Contact Dermatitis. 1989 ; 21 : l66-171.

77
91. Kuppens EV, de Jong CA, Stolwijk TR, de Kreizer RJ, van Best JA. Effects of timolol
with and without preservative on the basal tear turnover in glaucoma. Br J
Ophthalmol. 1995 ; 79 : 339-342.
92. Gobbels M, Spitznas M. Corneal epithelial permeability of dry eyes before and after
treatment with artificial tears. Ophthalmology. 1992 ; 99(6) : 873-878.
93. De Saint Jean M, Brignole F, Bringuier AF, Bauchet A, Feldmann G, Baudouin C.
Effects of benzalkonium chloride on growth and survival of Chang conjunctival cells.
Invest Ophthalmol Vis Sci. 1999 ; 40 : 619-30.
94. Debbasch C, Brignole F, Pisella PJ, Warnet JM, Rat P, Baudouin C. Quaternary
ammoniums and other preservatives' contribution in oxidative stress and apoptosis on
Chang conjunctival cells. Invest Ophthalmol Vis Sci. 2001 ; 42 : 642-52.
95. Debbasch C, Pisella PJ, De Saint Jean M, Rat P, Warnet JM, Baudouin C.
Mitochondrial activity and glutathione injury in apoptosis induced by unpreserved and
preserved betablockers on Chang conjunctival cells. Invest Ophthalmol Vis Sci. 2001 ;
42 : 2525-33.
96. Guenoun JM, Baudouin C, Rat P, Pauly A, Warnet JM, Brignole-Baudouin F. In vitro
comparison of cytoprotective and antioxidative effects of latanoprost, travoprost, and
bimatoprost on conjunctiva-derived epithelial cells. Invest Ophthalmol Vis Sci. 2005 ;
46 : 4594-9.
97. Srinivasan BD, Worgul BW, Iwamoto T, Merriam GR Jr. The conjunctival
epithelium. II. Histochemical and ultrastructural studies on human and rat conjunctiva.
Ophthalmic Res. 1977 ; 9 : 65–79.
98. Greiner JV, Kenyon KR, Henriquez AS, Korb DR, Weidman TA, Allansmith MR.
Mucus secretory vesicles in conjunctival epithelial cells of wearers of contact lenses.
Arch Ophthalmol. 1980 ; 98 : 1843–1846.
99. El Annan J, Chauhan SK, Ecoiffier T, Zhang Q, Saban DR, Dana R. Characterization
of effector T cells in dry eye disease. Invest Ophthalmol Vis Sci. 2009 ; 50(8) : 3802-
7.
100. Chauhan SK, El Annan J, Ecoiffier T, Goyal S, Zhang Q, Saban DR, Dana R.
Autoimmunity in dry eye is due to resistance of Th17 to Treg suppression. J Immunol.
2009 ; 182(3) : 1247-52.

78
101. Lavappa KS, Macy ML, et al. Examination of ATCC stocks for HeLa marker
chromosomes in human cell lines. Nature. 1976 ; 259(5540) : 211-3.
102. Diebold Y, Calonge M, Enriquez de Salamanca A, Callejo S, Corrales RM,
Saez V, Siemasko KF, Stern ME. Characterization of a spontaneously immortalized
cell line (IOBA-NHC) from normal human conjunctiva. Invest Ophthalmol Vis Sci
2003 ; 44 : 4263-74.
103. Chang RS. Continuous subcultivation of epithelial-like cells from normal
human tissues. Proc Soc Exp Biol Med. 1954 ; 87(2) : 440-3.
104. Guenoun JM, Baudouin C, et al. In vitro study of inflammatory potential and
toxicity profile of latanoprost, travoprost, and bimatoprost in conjunctiva-derived
epithelial cells. Invest Ophthalmol Vis Sci. 2005 ; 46(7) : 2444-50.
105. Ryan CA, Kimber I, Basketter DA, Pallardy M, Gildea LA, Gerberick GF.
Dendritic cells and skin sensitization: biological roles and uses in hazard
identification. Toxicol Appl Pharmacol. 2007 ; 15;221(3) : 384-94.
106. Fayol-Messaoudi D, Coconnier-Polter MH, Moal VL, Atassi F, Berger CN,
Servin AL. The Lactobacillus plantarum strain ACA-DC287 isolated from a Greek
cheese demonstrates antagonistic activity in vitro and in vivo against Salmonella
enterica serovar Typhimurium. J Appl Microbiol. 2007 ; 103(3) : 657-65.
107. Upham JW, Stick SM. Interactions between airway epithelial cells and
dendritic cells: implications for the regulation of airway inflammation. Curr Drug
Targets. 2006 ; 7(5) : 541-5.
108. Wang Y, Zhang M, Tan Y, Xiang Y, Liu H, Qu F, Qin L, Qin X. BRS-3
activation transforms the effect of human bronchial epithelial cells from PGE2
mediated inhibition to TGF-beta1 dependent promotion on proliferation and collagen
synthesis of lung fibroblasts. Cell Biol Int. 2007 ; 31(12) : 1495-1500.
109. Gomes JA, Dua HS, et al. Ocular surface epithelium induces expression of
human mucosal lymphocyte antigen (HML-1) on peripheral blood lymphocytes. Br J
Ophthalmol. 2004 ; 88(2) : 280-5.
110. Ohira T, Gemmill RM, et al. WNT7a induces E-cadherin in lung cancer cells.
Proc Natl Acad Sci U S A. 2003 ; 100(18) : 10429-34.
111. Blein OR, Adolphe M. Cross contamination associated with the use of

79
multiwell culture plates for cytoxicity assessment of volatile chemicals.
Cytotechnology. 1991 ; 6 : 79-82.
112. Peloux A, Federici C, Bichet N, Gouy D, Cano J. Hepatocytes in primary
culture: an alternative to LD 50 testing? Validation of a predictive model by
multivariate analysis. ATLA. 1992 ; 28 : 8-26.
113. Rat P, Osseni R, Christen M, Thevenin M, Warnet J-M, Adolphe M.
Microtitration fluorimetric assays on living cells (MiFALC tests): new tools for
screening in cell pharmacotoxicology. In : Van Zutphen LFM BM, ed. Animal
Alternatives, Welfare and Ethics. Paris: Elsevier 1997 : 813-825.
114. Brasnu E, Brignole-Baudouin F, et al. Comparative study on the cytotoxic
effects of benzalkonium chloride on the Wong-Kilbourne derivative of Chang
conjunctival and IOBA-NHC cell lines. Mol Vis. 2008 ; 14 : 394-402.
115. Dua HS, Gomes JA, et al. The ocular surface as part of the mucosal immune
system: conjunctival mucosa-specific lymphocytes in ocular surface pathology. Eye.
1995 ; 9 ( Pt 3) : 261-7.
116. Dua HS, Gomes JA, Jindal VK, Appa SN, Schwarting R, Eagle RC Jr, Donoso
LA, Laibson PR. Mucosa specific lymphocytes in the human conjunctiva,
corneoscleral limbus and lacrimal gland. Curr Eye Res. 1994 ; 13(1) : 87-93.
117. Chandler JW, Gillette TE. Immunologic defense mechanisms of the ocular
surface. Ophthalmology. 1983 Jun ; 90(6) : 585-91.
118. Sacks EH, Wieczorek R, Jakobiec FA, Knowles DM 2nd. Lymphocytic
subpopulations in the normal human conjunctiva. A monoclonal antibody study.
Ophthalmology. 1986 ; 93(10) : 1276-83.
119. Franklin RM, Remus LE. Conjunctival-associated lymphoid tissue: evidence
for a role in the secretory immune system. Invest Ophthalmol Vis Sci. 1984 Feb ;
25(2) : 181-7.
120. Iwata M, Kiritoshi A, Roat MI, Yagihashi A, Thoft RA.Regulation of HLA
class II antigen expression on cultured corneal epithelium by interferon-gamma.Invest
Ophthalmol Vis Sci. 1992 ; 33(9) : 2714-21.
121. Shams NB, Huggins EM Jr, Sigel MM. Interleukin-1 regulates the proliferation
of leukocytes in human corneal cell-peripheral blood leukocyte cocultures. Cornea.

80
1994 ; 13(1) : 9-15.
122. Iwata M, Yagihashi A, Roat MI, Zeevi A, Iwaki Y, Thoft RA. Human
leukocyte antigen-class II-positive human corneal epithelial cells activate allogeneic T
cells. Invest Ophthalmol Vis Sci. 1994 ; 35(12) : 3991-4000.
123. Wanner R, Sonnenburg A, Quatchadze M, Schreiner M, Peiser M, Zuberbier T,
Stahlmann R. Classification of sensitizing and irritative potential in a combined in-
vitro assay. Toxicol Appl Pharmacol. 2010 ; 245 : 211-8.
124. Swanson JA, Watts C. Macropinocytosis. Trends Cell Biol. 1995 ; 5(11) : 424-
8.
125. Racoosin EL, Swanson JA. M-CSF-induced macropinocytosis increases solute
endocytosis but not receptor-mediated endocytosis in mouse macrophages. J Cell Sci.
1992 ; 102 : 867-80.
126. Fiorentini C, Falzano L, et al. Activation of rho GTPases by cytotoxic
necrotizing factor 1 induces macropinocytosis and scavenging activity in epithelial
cells. Mol Biol Cell. 2001 ; 12(7) : 2061-73.
127. Hewlett LJ, Watts C, et al. The coated pit and macropinocytotic pathways
serve distinct endosome populations. J Cell Biol. 1994 ; 124(5) : 689-703.
128. Geppert TD, Lipsky PE. Antigen presentation by interferon-gamma-treated
endothelial cells and fibroblasts: differential ability to function as antigen-presenting
cells despite comparable Ia expression. J Immunol. 1985 ; 135(6) : 3750-62.
129. Young E, Stark WJ. In vitro immunological function of human corneal
fibroblasts. Invest Ophthalmol Vis Sci. 1988 ; 29(9) : 1402-6.
130. Gomes JA, Schwarting R, Rizzo L, et al. Mechanism of expression of the
human mucosal lymphocyte antigen (HML-1) in conjunctiva associated lymphoid
tissue (CALT). Invest Ophthalmol Vis Sci 1995 ; 36 : S840.
131. Poussier P, Julius M. Thymus independent T cell development and selection in
the intestinal epithelium. Annu Rev Immunol. 1994 ; 12 : 521-53.
132. Brandtzaeg P, Sollid ML, et al. Lymphoepithelial interactions in the mucosal
immune system. Gut. 1988 ; 29(8) : 1116-30.
133. Jarry A, Cerf-Bensussan N, et al. Subsets of CD3+ (T cell receptor alpha/beta
or gamma/delta) and CD3- lymphocytes isolated from normal human gut epithelium

81
display phenotypical features different from their counterparts in peripheral blood. Eur
J Immunol. 1990 ; 20(5) : 1097-103.
134. Cerf-Bensussan N, Jarry A, et al. A monoclonal antibody (HML-1) defining a
novel membrane molecule present on human intestinal lymphocytes. Eur J Immunol.
1987 ; 17(9) : 1279-85.
135. Cepek KL, Shaw SK, et al. Adhesion between epithelial cells and T
lymphocytes mediated by E-cadherin and the alpha E beta 7 integrin." Nature. 1994 ;
372(6502) : 190-3.
136. Karecla PI, Green SJ, et al. Identification of a binding site for integrin
alphaEbeta7 in the N-terminal domain of E-cadherin. J Biol Chem. 1996 ; 271(48) :
30909-15.
137. Parker CM, Cepek KL, et al. A family of beta 7 integrins on human mucosal
lymphocytes. Proc Natl Acad Sci U S A. 1992 ; 89(5) : 1924-8.
138. Higgins JM, Mandlebrot DA, et al. Direct and regulated interaction of integrin
alphaEbeta7 with E-cadherin. J Cell Biol. 1998 ; 140(1) : 197-210.
139. Takeichi M. Cadherins : a molecular family important in selective cell-cell
adhesion. Annu Rev Biochem. 1990 ; 59 : 237-52.
140. Schieferdecker HL, Ullrich R, Weiss-Breckwoldt AN, et al. The HML-1
antigen of intestinal lymphocytes is an activation antigen. J Immunol. 1990 ; 144 :
2541–9.
141. Lu Z, Ghosh S, et al. Downregulation of caveolin-1 function by EGF leads to
the loss of E-cadherin, increased transcriptional activity of beta-catenin, and enhanced
tumor cell invasion. Cancer Cell. 2003 ; 4(6) : 499-515.
142. Verschueren K, Remacle JE, et al. SIP1, a novel zinc finger/homeodomain
repressor, interacts with Smad proteins and binds to 5'-CACCT sequences in candidate
target genes. J Biol Chem. 1999 ; 274(29) : 20489-98.
143. Chinnadurai G. CtBP, an unconventional transcriptional corepressor in
development and oncogenesis. Mol Cell. 2002 ; 9(2) : 213-24.

82
VIII. TITRE ET RESUME EN ANGLAIS.
In vitro interactions between peripheral blood lymphocytes and Wong-Kilbourne
Derivative of Chang conjunctival cells. Towards a model of study of the ocular surface,
immunological and toxicological assessment.
During the past ten years, the effects of the toxicant in eye drops have been the subject of
many toxicological studies on ocular surface. The leader of these products is the
benzalkonium chloride (BAC), belonging to the quaternary ammonium family, used as a
preservative in eye drops for its bactericidal properties. But the BAC is also responsible in the
occurrence of clinical symptoms such as allergy or dry eye syndrom when used chronically,
such as in glaucoma disease. Several in vivo and in vitro tools are relevant in this aera of
knowledge. Up to now, the main in vitro studies used the Chang conjunctiva cell line as a
monocellular model that does not yet match perfectly the in vivo reality, in which
inflammatory cells are also involved. In the work that is reported here, we studied the cellular
interactions between Chang cells and peripheral blood lymphocytes, then used this model for
toxicological assessment. Cell morphology, apoptosis, inflammation and BAC-induced cell
death have been studied using standard immunofluorescence, microplate cytofluorometry and
flow cytometry, showing cells‟ fusion, increase of inflammation and epithelial cells apoptosis
due to the presence of lymphocytes, and sometimes decrease of epithelial cells apoptosis
under certain conditions. However, the allogeneic reaction induced by this coculture model
could compromise a reliable interpretation of the results. Thus, new cells model like coculture
of epithelial cells and dendritic cells taking from the same donor may be extremely relevant
for understanding and studying the ocular surface physiopathology.

83
ANNEE : 2010
Thèse soutenue le 22 Octobre 2010 par Monsieur Alexis CHASSIGNOL
PRESIDENT : Monsieur le Professeur LAROCHE
DIRECTEUR : Monsieur le Professeur BAUDOUIN
INTERACTIONS CELLULAIRES IN VITRO DE CELLULES
CONJONCTIVALES EN LIGNEE ET DE LYMPHOCYTES ISSUS DU
SANG CIRCULANT. MODELE D’ETUDE DE LA SURFACE
OCULAIRE, IMMUNOLOGIE ET TOXICOLOGIE.
Ces dernières années, les effets des toxiques contenus dans les collyres ont fait l‟objet de
nombreuses études toxicologiques de la surface oculaire. Le chef de file de ces produits
toxiques est le chlorure de benzalkonium (BAC), agent de la famille des ammoniums
quaternaires, utilisé comme conservateur dans les collyres pour ces propriétés bactéricides.
Mais le BAC est aussi responsable d‟un cortège de symptômes cliniques tels que l‟allergie ou
le syndrome sec oculaires lorsqu‟il est utilisé de façon chronique, comme chez les patients
glaucomateux. Plusieurs outils in vivo et in vitro sont utilisables dans ce domaine pour mettre
en évidence l‟effet toxique du conservateur. Les principales études in vitro réalisées jusqu‟à
présent utilisaient comme modèle la lignée de cellules conjonctivales de Chang, dans un
système unicellulaire qui ne reproduisait pas les conditions du vivo, au sein duquel
interviennent également des cellules inflammatoires. Au cours de ce travail, nous avons étudié
les interactions cellulaires entre des cellules de Chang et des lymphocytes issus du sang
circulant, puis tester ce modèle sur le plan toxicologique. Morphologie cellulaire, apoptose,
inflammation et mort cellulaire induite par le BAC ont été étudiées en utilisant des techniques
d‟immunofluorescence standard, de microtitration fluorimétrique en microplaques, et de
cytométrie de flux, mettant en évidence des phénomènes de fusion cellulaire, de
potentialisation de l‟inflammation et de l‟apoptose des cellulaires épithéliales liée à la
présence des lymphocytes, et parfois de survie des cellules épithéliales dans certaines
conditions. Cependant, la reaction allogénique induite par ce modèle de coculture peut
parasiter l‟interprétation des résultats. Ceci ouvre la voie vers un autre axe de recherche qui
serait le développement d‟un modèle de coculture entre des cellules épithéliales et des cellules
dendritiques provenant du même donneur permettant de progresser vers la constitution d‟un
modèle d‟étude de la surface oculaire au plus près du vivo.
MOTS CLES :
Benzalkonium, composés, Alternatives à l'utilisation d'animaux - Lignées cellulaires,
Techniques de coculture, Conjonctive.
DISCIPLINE ;
Médecine (Spécialité OPHTALMOLOGIE)
UFR :
Université Paris Est Créteil
8 rue de général Sarrail
94010 CRETEIL
Top Related