
Spécialisations membranaires des épithéliums - … membranaires.pdf · chaque membrane plasmique...
9
Spécialisations membranaires des épithéliums Traduction: Young et al: Wheater's Functional Histology 5Ed Les membranes cytoplasmiques des cellules épithéliales présentent une grande variété de structures spécialisées qui leur permettent de remplir leur fonction de barrière à perméabilité sélective. Dans certains cas, la barrière épithéliale est imperméable, par exemple, l'épithélium transitionnel de la vessie, tandis que d'autres cas tels que l'épithélium de la muqueuse de l'intestin grêle ou des tubes contournés du rein favorise le mouvement de certains ions et molécules à travers l'épithélium. Surfaces intercellulaires Les surfaces latérales des cellules épithéliales adjacentes sont liées par des jonctions cellulaires de telle sorte que l'épithélium forme une couche continue homogène. Les jonctions cellulaires fonctionnent également comme des canaux de communication qui régissent des fonctions telles que la croissance et la division cellulaire. Les différents types de jonction se composent de protéines transmembranaires qui interagissent avec des protéines analogues de cellules adjacentes et sont liés à des structures intracellulaires du côté cytoplasmique. Les jonctions cellulaires sont de trois types fonctionnels: • Jonctions serrées (jonctions occlusives) bloquent le passage des molécules entre les cellules adjacentes. Elles sont situées immédiatement au-dessous de la surface luminale d'épithélium simple (par exemple muqueuse intestinale). Elles scellent les espaces intercellulaires de sorte que le contenu luminal ne peut pas pénétrer entre les cellules de la paroi. Chaque jonction serrée forme une bande circonférentielle continue ou zonule autour de la cellule, d’ou son nom, zonula occludens. • Zonula adherens (jonction adhérente) et desmosomes confèrent la force à l’épithélium en reliant les cytosquelettes de cellules adjacentes. Les jonctions adhérentes se lient à l'actine du cytosquelette de la cellule. Dans l'épithélium cylindrique simple, la zonula adherens forme une bande continue autour de la cellule sous la zonula occludens pour former une bande circonférentielle de contraction à proximité de la surface apicale de la cellule. Les desmosomes (ou macula adherens) fournissent également un lien solide entre les cellules adjacentes. Ils relient les filaments intermédiaires (kératines) pour former un solide cadre architectural pour la totalité de l’épithélium. Les desmosomes sont disposés de manière circonférentielle sous la jonction adhérente. L’association des zonula occludens, adherens, et des desmosomes est appelé complexe jonctionnel. Ce dernier divise la membrane cytoplasmique de la cellule en surfaces apicale et basolatérale. Les desmosomes sont largement dispersés ailleurs dans interfaces intercellulaires des épithéliums. • Jonctions communicantes, également appelées gap, permettent le passage de petites molécules directement entre les cellules adjacentes. Ces molécules de signalisation coordonnent et synchronisent les fonctions de l'épithélium. Les jonctions adhérentes et communicantes ne sont pas exclusives aux épithéliums et sont également présents dans le muscle cardiaque et des viscères où ils assurent des fonctions similaires. Surfaces luminales Les surfaces luminales ou apicale des cellules épithéliales peuvent intégrer trois types principaux de spécialisation: cils, microvillosités et stéréocils. Les cils sont des structures activement mobiles qui peuvent être observés par microscopie optique. En revanche, les microvillosités sont de courtes projections de la membrane plasmique qui ne peuvent être vus individuellement par microscopie optique. Une seule cellule peut avoir des milliers de microvillosités ou seulement quelques-uns. Les stéréocils sont de très longues microvillosités que l'on trouve habituellement seuls ou en petits nombres; les stéréocils sont pas mobiles. Surfaces basales L'interface entre l'épithélium et les tissus de support sous-jacent est délimitée par une structure non cellulaire appelée membrane basale, qui fournit un support architectural pour l'épithélium et constitue une barrière sélective pour le passage des substrats entre l'épithélium et le tissu de support . Les hémidesmosomes, une variante de desmosomes, lient la base de la cellule à la membrane basale sous-jacente par un réseau de filaments intermédiaires.
Transcript of Spécialisations membranaires des épithéliums - … membranaires.pdf · chaque membrane plasmique...

Spécialisations membranaires des épithéliums Traduction: Young et al: Wheater's Functional Histology 5Ed Les membranes cytoplasmiques des cellules épithéliales présentent une grande variété de structures spécialisées qui leur permettent de remplir leur fonction de barrière à perméabilité sélective. Dans certains cas, la barrière épithéliale est imperméable, par exemple, l'épithélium transitionnel de la vessie, tandis que d'autres cas tels que l'épithélium de la muqueuse de l'intestin grêle ou des tubes contournés du rein favorise le mouvement de certains ions et molécules à travers l'épithélium. Surfaces intercellulaires Les surfaces latérales des cellules épithéliales adjacentes sont liées par des jonctions cellulaires de telle sorte que l'épithélium forme une couche continue homogène. Les jonctions cellulaires fonctionnent également comme des canaux de communication qui régissent des fonctions telles que la croissance et la division cellulaire. Les différents types de jonction se composent de protéines transmembranaires qui interagissent avec des protéines analogues de cellules adjacentes et sont liés à des structures intracellulaires du côté cytoplasmique. Les jonctions cellulaires sont de trois types fonctionnels: • Jonctions serrées (jonctions occlusives) bloquent le passage des molécules entre les cellules adjacentes.
Elles sont situées immédiatement au-dessous de la surface luminale d'épithélium simple (par exemple muqueuse intestinale). Elles scellent les espaces intercellulaires de sorte que le contenu luminal ne peut pas pénétrer entre les cellules de la paroi. Chaque jonction serrée forme une bande circonférentielle continue ou zonule autour de la cellule, d’ou son nom, zonula occludens.
• Zonula adherens (jonction adhérente) et desmosomes confèrent la force à l’épithélium en reliant les cytosquelettes de cellules adjacentes. Les jonctions adhérentes se lient à l'actine du cytosquelette de la cellule. Dans l'épithélium cylindrique simple, la zonula adherens forme une bande continue autour de la cellule sous la zonula occludens pour former une bande circonférentielle de contraction à proximité de la surface apicale de la cellule. Les desmosomes (ou macula adherens) fournissent également un lien solide entre les cellules adjacentes. Ils relient les filaments intermédiaires (kératines) pour former un solide cadre architectural pour la totalité de l’épithélium. Les desmosomes sont disposés de manière circonférentielle sous la jonction adhérente. L’association des zonula occludens, adherens, et des desmosomes est appelé complexe jonctionnel. Ce dernier divise la membrane cytoplasmique de la cellule en surfaces apicale et basolatérale. Les desmosomes sont largement dispersés ailleurs dans interfaces intercellulaires des épithéliums.
• Jonctions communicantes, également appelées gap, permettent le passage de petites molécules directement entre les cellules adjacentes. Ces molécules de signalisation coordonnent et synchronisent les fonctions de l'épithélium.
Les jonctions adhérentes et communicantes ne sont pas exclusives aux épithéliums et sont également présents dans le muscle cardiaque et des viscères où ils assurent des fonctions similaires. Surfaces luminales Les surfaces luminales ou apicale des cellules épithéliales peuvent intégrer trois types principaux de spécialisation: cils, microvillosités et stéréocils. Les cils sont des structures activement mobiles qui peuvent être observés par microscopie optique. En revanche, les microvillosités sont de courtes projections de la membrane plasmique qui ne peuvent être vus individuellement par microscopie optique. Une seule cellule peut avoir des milliers de microvillosités ou seulement quelques-uns. Les stéréocils sont de très longues microvillosités que l'on trouve habituellement seuls ou en petits nombres; les stéréocils sont pas mobiles. Surfaces basales L'interface entre l'épithélium et les tissus de support sous-jacent est délimitée par une structure non cellulaire appelée membrane basale, qui fournit un support architectural pour l'épithélium et constitue une barrière sélective pour le passage des substrats entre l'épithélium et le tissu de support . Les hémidesmosomes, une variante de desmosomes, lient la base de la cellule à la membrane basale sous-jacente par un réseau de filaments intermédiaires.
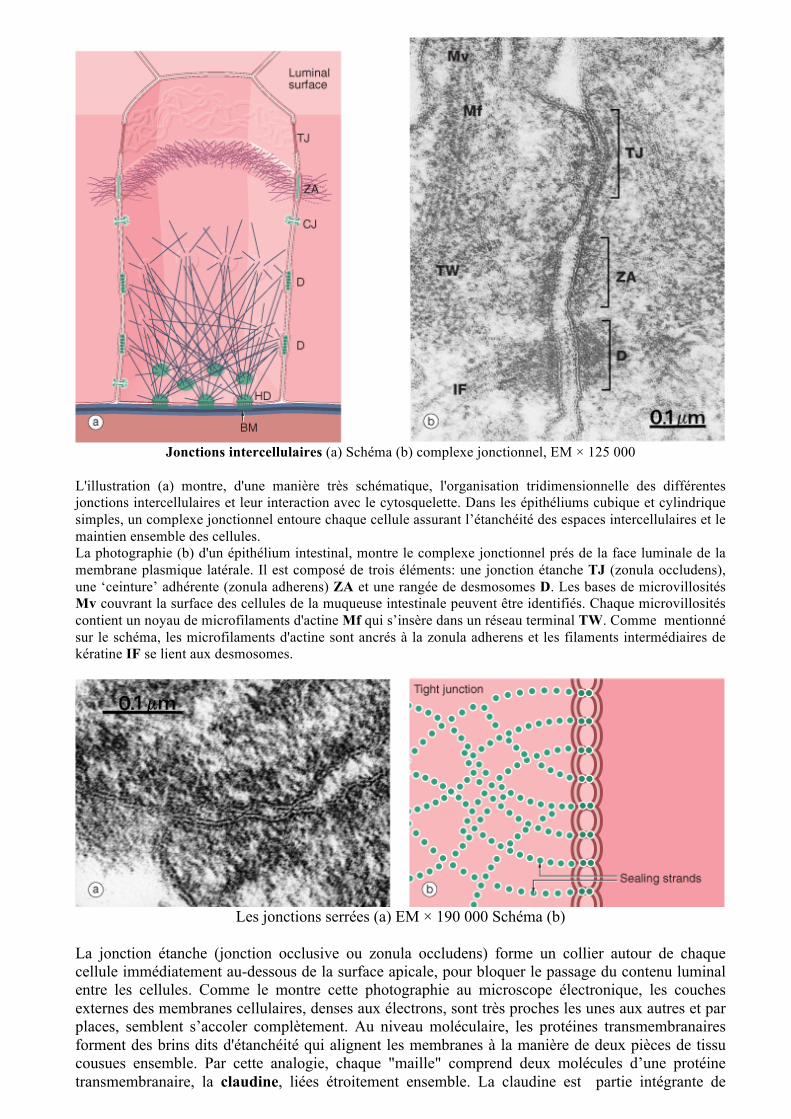
Jonctions intercellulaires (a) Schéma (b) complexe jonctionnel, EM × 125 000
L'illustration (a) montre, d'une manière très schématique, l'organisation tridimensionnelle des différentes jonctions intercellulaires et leur interaction avec le cytosquelette. Dans les épithéliums cubique et cylindrique simples, un complexe jonctionnel entoure chaque cellule assurant l’étanchéité des espaces intercellulaires et le maintien ensemble des cellules. La photographie (b) d'un épithélium intestinal, montre le complexe jonctionnel prés de la face luminale de la membrane plasmique latérale. Il est composé de trois éléments: une jonction étanche TJ (zonula occludens), une ‘ceinture’ adhérente (zonula adherens) ZA et une rangée de desmosomes D. Les bases de microvillosités Mv couvrant la surface des cellules de la muqueuse intestinale peuvent être identifiés. Chaque microvillosités contient un noyau de microfilaments d'actine Mf qui s’insère dans un réseau terminal TW. Comme mentionné sur le schéma, les microfilaments d'actine sont ancrés à la zonula adherens et les filaments intermédiaires de kératine IF se lient aux desmosomes.
Les jonctions serrées (a) EM × 190 000 Schéma (b)
La jonction étanche (jonction occlusive ou zonula occludens) forme un collier autour de chaque cellule immédiatement au-dessous de la surface apicale, pour bloquer le passage du contenu luminal entre les cellules. Comme le montre cette photographie au microscope électronique, les couches externes des membranes cellulaires, denses aux électrons, sont très proches les unes aux autres et par places, semblent s’accoler complètement. Au niveau moléculaire, les protéines transmembranaires forment des brins dits d'étanchéité qui alignent les membranes à la manière de deux pièces de tissu cousues ensemble. Par cette analogie, chaque "maille" comprend deux molécules d’une protéine transmembranaire, la claudine, liées étroitement ensemble. La claudine est partie intégrante de
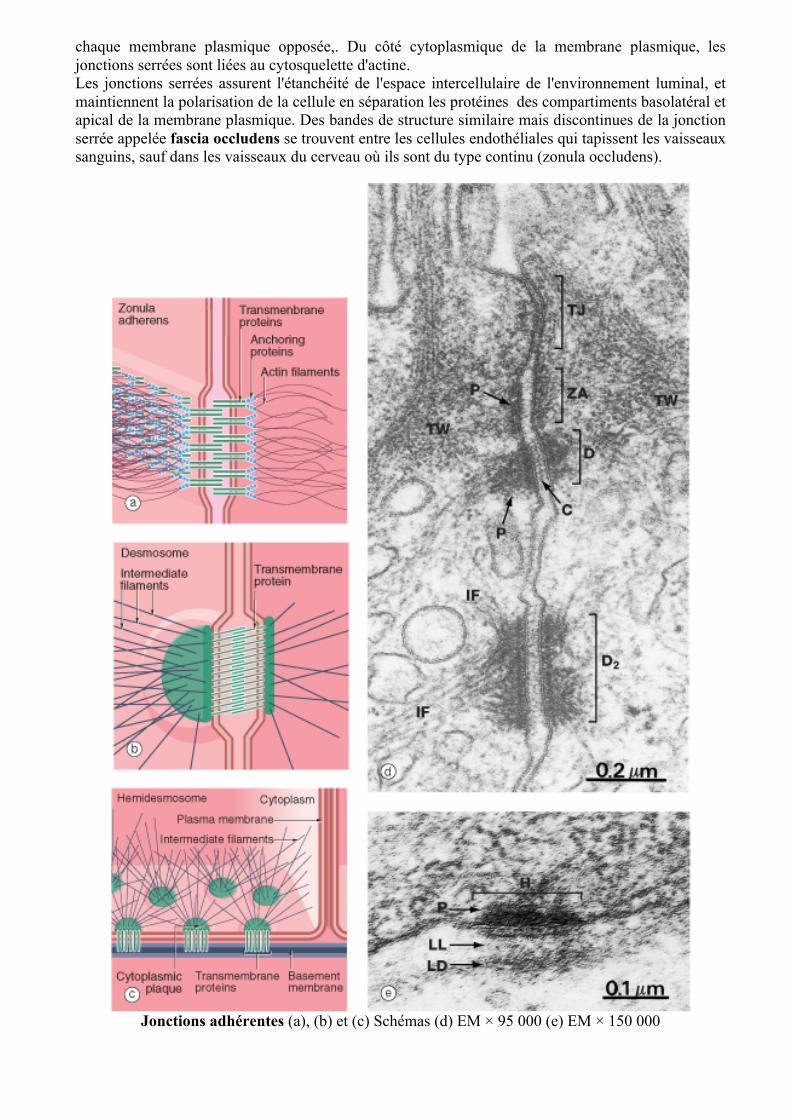
chaque membrane plasmique opposée,. Du côté cytoplasmique de la membrane plasmique, les jonctions serrées sont liées au cytosquelette d'actine. Les jonctions serrées assurent l'étanchéité de l'espace intercellulaire de l'environnement luminal, et maintiennent la polarisation de la cellule en séparation les protéines des compartiments basolatéral et apical de la membrane plasmique. Des bandes de structure similaire mais discontinues de la jonction serrée appelée fascia occludens se trouvent entre les cellules endothéliales qui tapissent les vaisseaux sanguins, sauf dans les vaisseaux du cerveau où ils sont du type continu (zonula occludens).
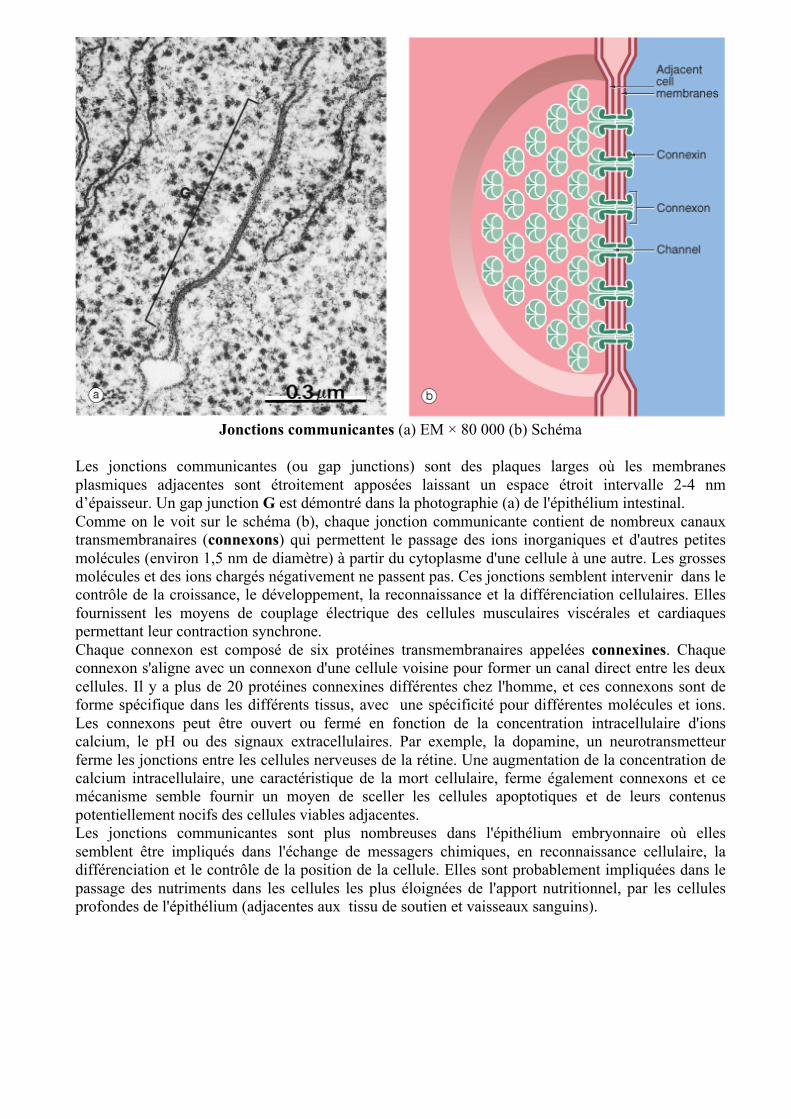
Jonctions adhérentes (a), (b) et (c) Schémas (d) EM × 95 000 (e) EM × 150 000

Les jonctions adhérentes fournissent des points d'ancrage pour les éléments du cytosquelette, reliant les cytosquelettes des cellules individuelles en un solide réseau transcellulaire. Ces jonctions sont de trois types: la ceinture d'adhérence (zonula adherens) et les desmosomes (macula adherens) lient les cellules adjacentes et les hemidesmosomes qui lient la cellule à la membrane basale sous-jacente. La zonula adherens forme une bande continue située à la profondeur de jonction étanche de l'extrémité luminale des membranes plasmiques latérales de l'épithélium cylindrique. Sous la zonula adherens il y a un anneau de desmosomes, la troisième composante du complexe jonctionnel. De grands desmosomes sont également dispersés sur les surfaces intercellulaires de toutes les cellules épithéliales. Les jonctions adhérentes sont constitués de trois composants: des protéines transmembranaires qui se lient à des protéines similaires des cellules adjacentes (zonula adherens et desmosomes) ou à la matrice extracellulaire (hémidesmosomes), des protéines d'ancrage sur le côté cytoplasmique de la jonction qui relient les protéines transmembranaires à la troisième composante, le cytosquelette. Dans La zonula adherens (schéma : a) les protéines transmembranaires sont les cadhérines. Les cadhérines couvrent les membranes plasmiques des cellules et se lient aux cadhérines identiques sur les cellules adjacentes. Les queues cytoplasmiques des cadhérines se lient aux protéines d'ancrage (caténines, vinculine et α-actinine) qui se lient à leur tour aux molécules d'actine. La composante intracellulaire de la zonula adherens apparaît comme une petite plaque P dense aux électrons sur le côté cytoplasmique de la membrane plasmique. Les cadhérines constituent également les protéines transmembranaires des desmosomes (schéma : b). Les segments des molécules de cadhérines qui se chevauchent dans l'espace intercellulaire forment une ligne C, dense aux électrons. Sur le versant cytoplasmique, les protéines d'ancrage (desmoplakine et plakoglobine) se lient aux filaments intermédiaires IF formant une plaque P dense aux électrons sur la face interne du desmosome. Les desmosomes sont plus nombreux dans l'épithélium pavimenteux stratifié, qui doit supporter plus de stress mécanique. Les hémidesmosomes (schéma : c) sont des desmosomes modifiés qui se trouvent à la face basale de la cellule. Les protéines transmembranaires sont les intégrines, dont les composants extracellulaires se lient aux laminines de la membrane basale. Le composant intracellulaire des intégrines se lie à la protéine d'ancrage, la plectine et par elle, aux filaments intermédiaires, la kératine. Le composant intracellulaire apparaît comme une plaque dense aux électrons P. La micrographie (d) de la muqueuse intestinale illustre un complexe jonctionnel comprenant une jonction serrée TJ, la zonula adherens ZA et un desmosome D. À un niveau plus profond, un plus grand desmosome D2 est observé. Notez la petite plaque P denses aux électrons de la zonula adherens et la plus grande plaque P des desmosomes. La ligne dense aux électrons créée par le chevauchement des molécules de cadhérine C est également visible dans les desmosomes. La micrographie (e) illustre un hémidesmosome H le long de la membrane basale d'une cellule épithéliale. La plaque P est sur la face cytoplasmique de la membrane plasmique. La lamina densa LD sous-jacente est épaisse, dense aux électrons, de même que lamina lucida LL, qui contient une ligne dense aux électrons. Cet aspect est le résultat de la liaison du composant extracellulaire des intégrines aux laminines de la membrane basale.
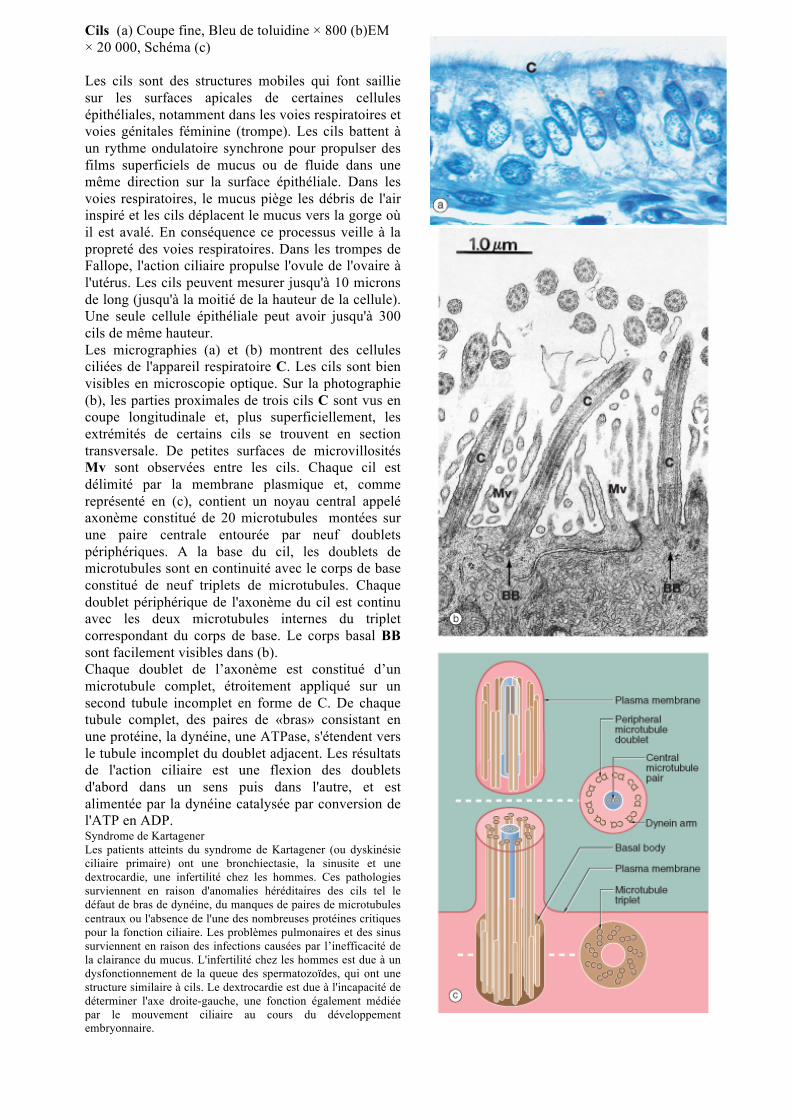
Jonctions communicantes (a) EM × 80 000 (b) Schéma
Les jonctions communicantes (ou gap junctions) sont des plaques larges où les membranes plasmiques adjacentes sont étroitement apposées laissant un espace étroit intervalle 2-4 nm d’épaisseur. Un gap junction G est démontré dans la photographie (a) de l'épithélium intestinal. Comme on le voit sur le schéma (b), chaque jonction communicante contient de nombreux canaux transmembranaires (connexons) qui permettent le passage des ions inorganiques et d'autres petites molécules (environ 1,5 nm de diamètre) à partir du cytoplasme d'une cellule à une autre. Les grosses molécules et des ions chargés négativement ne passent pas. Ces jonctions semblent intervenir dans le contrôle de la croissance, le développement, la reconnaissance et la différenciation cellulaires. Elles fournissent les moyens de couplage électrique des cellules musculaires viscérales et cardiaques permettant leur contraction synchrone. Chaque connexon est composé de six protéines transmembranaires appelées connexines. Chaque connexon s'aligne avec un connexon d'une cellule voisine pour former un canal direct entre les deux cellules. Il y a plus de 20 protéines connexines différentes chez l'homme, et ces connexons sont de forme spécifique dans les différents tissus, avec une spécificité pour différentes molécules et ions. Les connexons peut être ouvert ou fermé en fonction de la concentration intracellulaire d'ions calcium, le pH ou des signaux extracellulaires. Par exemple, la dopamine, un neurotransmetteur ferme les jonctions entre les cellules nerveuses de la rétine. Une augmentation de la concentration de calcium intracellulaire, une caractéristique de la mort cellulaire, ferme également connexons et ce mécanisme semble fournir un moyen de sceller les cellules apoptotiques et de leurs contenus potentiellement nocifs des cellules viables adjacentes. Les jonctions communicantes sont plus nombreuses dans l'épithélium embryonnaire où elles semblent être impliqués dans l'échange de messagers chimiques, en reconnaissance cellulaire, la différenciation et le contrôle de la position de la cellule. Elles sont probablement impliquées dans le passage des nutriments dans les cellules les plus éloignées de l'apport nutritionnel, par les cellules profondes de l'épithélium (adjacentes aux tissu de soutien et vaisseaux sanguins).
Cils (a) Coupe fine, Bleu de toluidine × 800 (b)EM × 20 000, Schéma (c) Les cils sont des structures mobiles qui font saillie sur les surfaces apicales de certaines cellules épithéliales, notamment dans les voies respiratoires et voies génitales féminine (trompe). Les cils battent à un rythme ondulatoire synchrone pour propulser des films superficiels de mucus ou de fluide dans une même direction sur la surface épithéliale. Dans les voies respiratoires, le mucus piège les débris de l'air inspiré et les cils déplacent le mucus vers la gorge où il est avalé. En conséquence ce processus veille à la propreté des voies respiratoires. Dans les trompes de Fallope, l'action ciliaire propulse l'ovule de l'ovaire à l'utérus. Les cils peuvent mesurer jusqu'à 10 microns de long (jusqu'à la moitié de la hauteur de la cellule). Une seule cellule épithéliale peut avoir jusqu'à 300 cils de même hauteur. Les micrographies (a) et (b) montrent des cellules ciliées de l'appareil respiratoire C. Les cils sont bien visibles en microscopie optique. Sur la photographie (b), les parties proximales de trois cils C sont vus en coupe longitudinale et, plus superficiellement, les extrémités de certains cils se trouvent en section transversale. De petites surfaces de microvillosités Mv sont observées entre les cils. Chaque cil est délimité par la membrane plasmique et, comme représenté en (c), contient un noyau central appelé axonème constitué de 20 microtubules montées sur une paire centrale entourée par neuf doublets périphériques. A la base du cil, les doublets de microtubules sont en continuité avec le corps de base constitué de neuf triplets de microtubules. Chaque doublet périphérique de l'axonème du cil est continu avec les deux microtubules internes du triplet correspondant du corps de base. Le corps basal BB sont facilement visibles dans (b). Chaque doublet de l’axonème est constitué d’un microtubule complet, étroitement appliqué sur un second tubule incomplet en forme de C. De chaque tubule complet, des paires de «bras» consistant en une protéine, la dynéine, une ATPase, s'étendent vers le tubule incomplet du doublet adjacent. Les résultats de l'action ciliaire est une flexion des doublets d'abord dans un sens puis dans l'autre, et est alimentée par la dynéine catalysée par conversion de l'ATP en ADP. Syndrome de Kartagener Les patients atteints du syndrome de Kartagener (ou dyskinésie ciliaire primaire) ont une bronchiectasie, la sinusite et une dextrocardie, une infertilité chez les hommes. Ces pathologies surviennent en raison d'anomalies héréditaires des cils tel le défaut de bras de dynéine, du manques de paires de microtubules centraux ou l'absence de l'une des nombreuses protéines critiques pour la fonction ciliaire. Les problèmes pulmonaires et des sinus surviennent en raison des infections causées par l’inefficacité de la clairance du mucus. L'infertilité chez les hommes est due à un dysfonctionnement de la queue des spermatozoïdes, qui ont une structure similaire à cils. Le dextrocardie est due à l'incapacité de déterminer l'axe droite-gauche, une fonction également médiée par le mouvement ciliaire au cours du développement embryonnaire.
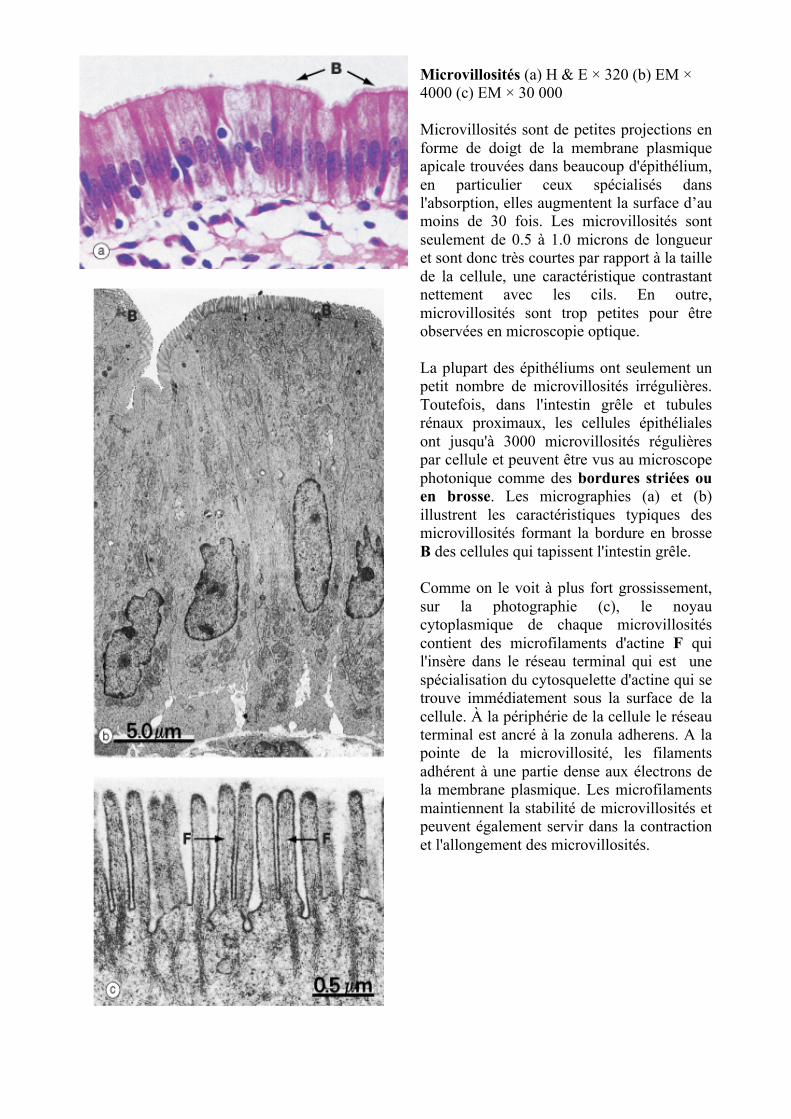
Microvillosités (a) H & E × 320 (b) EM × 4000 (c) EM × 30 000 Microvillosités sont de petites projections en forme de doigt de la membrane plasmique apicale trouvées dans beaucoup d'épithélium, en particulier ceux spécialisés dans l'absorption, elles augmentent la surface d’au moins de 30 fois. Les microvillosités sont seulement de 0.5 à 1.0 microns de longueur et sont donc très courtes par rapport à la taille de la cellule, une caractéristique contrastant nettement avec les cils. En outre, microvillosités sont trop petites pour être observées en microscopie optique. La plupart des épithéliums ont seulement un petit nombre de microvillosités irrégulières. Toutefois, dans l'intestin grêle et tubules rénaux proximaux, les cellules épithéliales ont jusqu'à 3000 microvillosités régulières par cellule et peuvent être vus au microscope photonique comme des bordures striées ou en brosse. Les micrographies (a) et (b) illustrent les caractéristiques typiques des microvillosités formant la bordure en brosse B des cellules qui tapissent l'intestin grêle. Comme on le voit à plus fort grossissement, sur la photographie (c), le noyau cytoplasmique de chaque microvillosités contient des microfilaments d'actine F qui l'insère dans le réseau terminal qui est une spécialisation du cytosquelette d'actine qui se trouve immédiatement sous la surface de la cellule. À la périphérie de la cellule le réseau terminal est ancré à la zonula adherens. A la pointe de la microvillosité, les filaments adhérent à une partie dense aux électrons de la membrane plasmique. Les microfilaments maintiennent la stabilité de microvillosités et peuvent également servir dans la contraction et l'allongement des microvillosités.
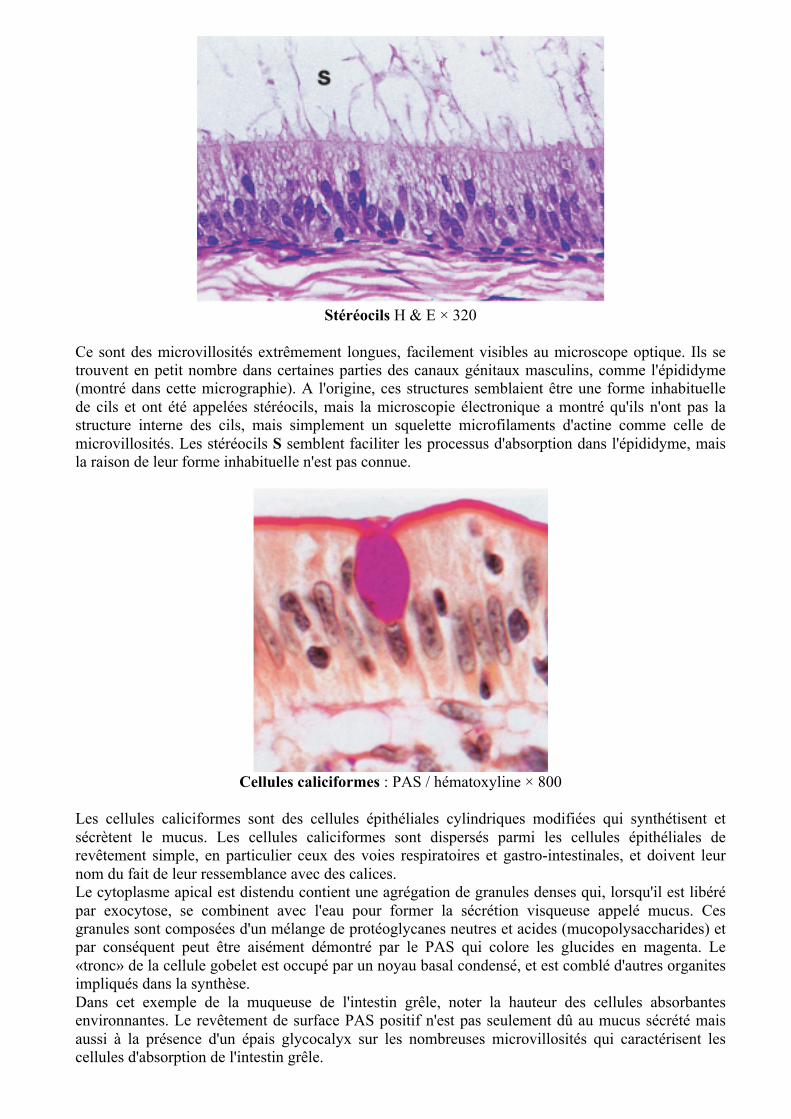
Stéréocils H & E × 320
Ce sont des microvillosités extrêmement longues, facilement visibles au microscope optique. Ils se trouvent en petit nombre dans certaines parties des canaux génitaux masculins, comme l'épididyme (montré dans cette micrographie). A l'origine, ces structures semblaient être une forme inhabituelle de cils et ont été appelées stéréocils, mais la microscopie électronique a montré qu'ils n'ont pas la structure interne des cils, mais simplement un squelette microfilaments d'actine comme celle de microvillosités. Les stéréocils S semblent faciliter les processus d'absorption dans l'épididyme, mais la raison de leur forme inhabituelle n'est pas connue.
Cellules caliciformes : PAS / hématoxyline × 800
Les cellules caliciformes sont des cellules épithéliales cylindriques modifiées qui synthétisent et sécrètent le mucus. Les cellules caliciformes sont dispersés parmi les cellules épithéliales de revêtement simple, en particulier ceux des voies respiratoires et gastro-intestinales, et doivent leur nom du fait de leur ressemblance avec des calices. Le cytoplasme apical est distendu contient une agrégation de granules denses qui, lorsqu'il est libéré par exocytose, se combinent avec l'eau pour former la sécrétion visqueuse appelé mucus. Ces granules sont composées d'un mélange de protéoglycanes neutres et acides (mucopolysaccharides) et par conséquent peut être aisément démontré par le PAS qui colore les glucides en magenta. Le «tronc» de la cellule gobelet est occupé par un noyau basal condensé, et est comblé d'autres organites impliqués dans la synthèse. Dans cet exemple de la muqueuse de l'intestin grêle, noter la hauteur des cellules absorbantes environnantes. Le revêtement de surface PAS positif n'est pas seulement dû au mucus sécrété mais aussi à la présence d'un épais glycocalyx sur les nombreuses microvillosités qui caractérisent les cellules d'absorption de l'intestin grêle.
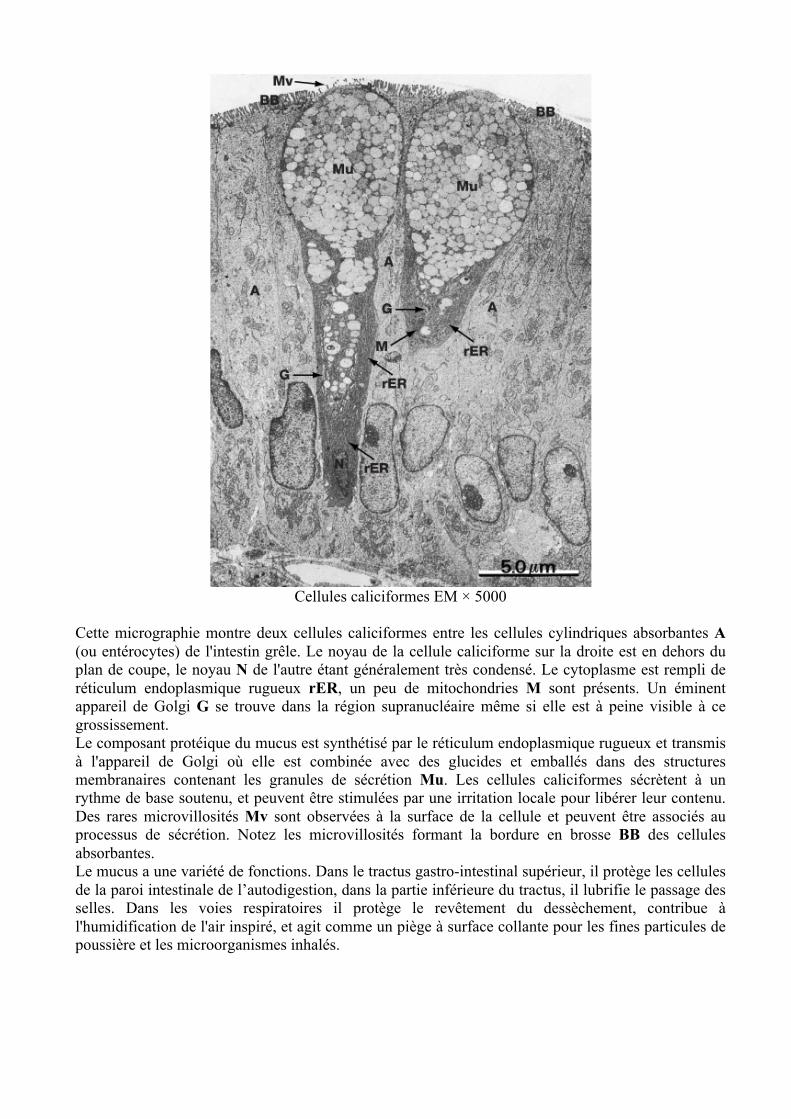
Cellules caliciformes EM × 5000
Cette micrographie montre deux cellules caliciformes entre les cellules cylindriques absorbantes A (ou entérocytes) de l'intestin grêle. Le noyau de la cellule caliciforme sur la droite est en dehors du plan de coupe, le noyau N de l'autre étant généralement très condensé. Le cytoplasme est rempli de réticulum endoplasmique rugueux rER, un peu de mitochondries M sont présents. Un éminent appareil de Golgi G se trouve dans la région supranucléaire même si elle est à peine visible à ce grossissement. Le composant protéique du mucus est synthétisé par le réticulum endoplasmique rugueux et transmis à l'appareil de Golgi où elle est combinée avec des glucides et emballés dans des structures membranaires contenant les granules de sécrétion Mu. Les cellules caliciformes sécrètent à un rythme de base soutenu, et peuvent être stimulées par une irritation locale pour libérer leur contenu. Des rares microvillosités Mv sont observées à la surface de la cellule et peuvent être associés au processus de sécrétion. Notez les microvillosités formant la bordure en brosse BB des cellules absorbantes. Le mucus a une variété de fonctions. Dans le tractus gastro-intestinal supérieur, il protège les cellules de la paroi intestinale de l’autodigestion, dans la partie inférieure du tractus, il lubrifie le passage des selles. Dans les voies respiratoires il protège le revêtement du dessèchement, contribue à l'humidification de l'air inspiré, et agit comme un piège à surface collante pour les fines particules de poussière et les microorganismes inhalés.


















