organisation Cellulaire Et Moléculaire Du Monde...
66
Organisation cellulaire et moléculaire du monde vivant Pierre DIEUMEGARD, le 5 avril 2008
Transcript of organisation Cellulaire Et Moléculaire Du Monde...

Organisation cellulaire et moléculaire du monde vivant
Pierre DIEUMEGARD,le 5 avril 2008
Organisation anatomique
vue microscopique
ultrastructure (microscopie électronique à transmission)
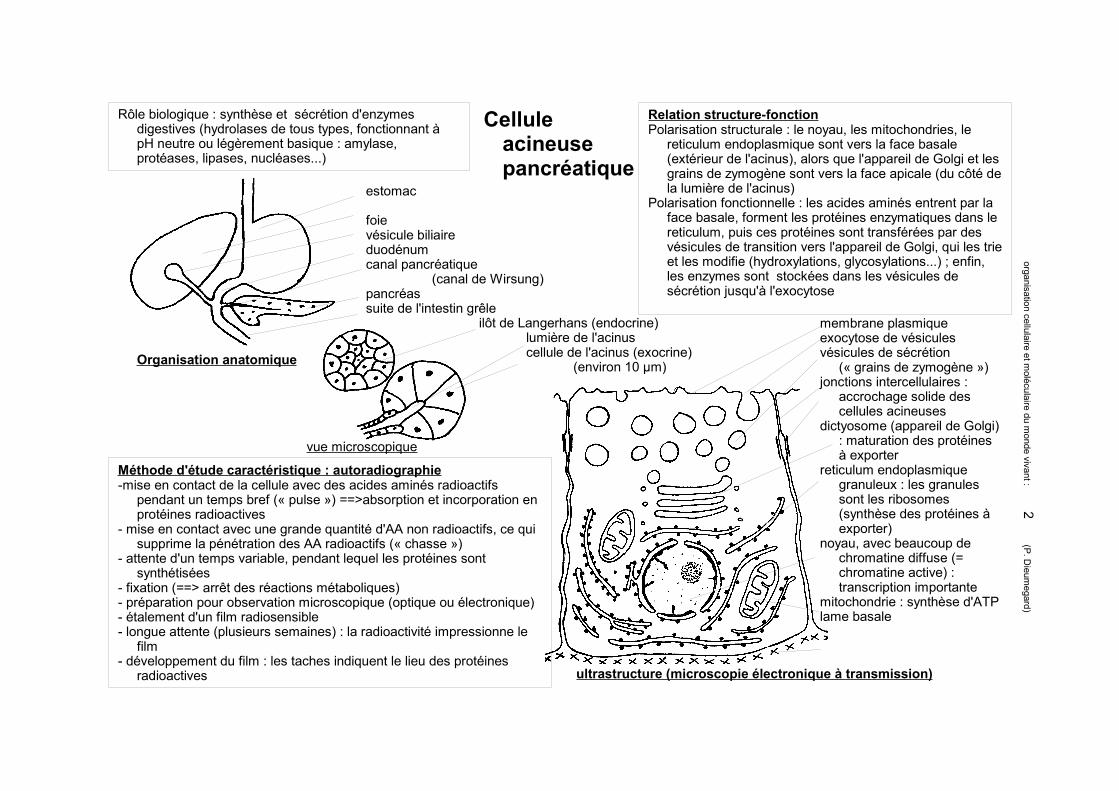
estomac
foievésicule biliaireduodénumcanal pancréatique
(canal de Wirsung)pancréassuite de l'intestin grêle
ilôt de Langerhans (endocrine)lumière de l'acinuscellule de l'acinus (exocrine)
(environ 10 µm)
Cellule acineuse pancréatique
Rôle biologique : synthèse et sécrétion d'enzymes digestives (hydrolases de tous types, fonctionnant à pH neutre ou légèrement basique : amylase, protéases, lipases, nucléases...)
Méthode d'étude caractéristique : autoradiographie-mise en contact de la cellule avec des acides aminés radioactifs
pendant un temps bref (« pulse ») ==>absorption et incorporation en protéines radioactives
- mise en contact avec une grande quantité d'AA non radioactifs, ce qui supprime la pénétration des AA radioactifs (« chasse »)
- attente d'un temps variable, pendant lequel les protéines sont synthétisées
- fixation (==> arrêt des réactions métaboliques)- préparation pour observation microscopique (optique ou électronique)- étalement d'un film radiosensible- longue attente (plusieurs semaines) : la radioactivité impressionne le
film- développement du film : les taches indiquent le lieu des protéines
radioactives
membrane plasmiqueexocytose de vésiculesvésicules de sécrétion
(« grains de zymogène »)jonctions intercellulaires :
accrochage solide des cellules acineuses
dictyosome (appareil de Golgi) : maturation des protéines à exporter
reticulum endoplasmique granuleux : les granules sont les ribosomes (synthèse des protéines à exporter)
noyau, avec beaucoup de chromatine diffuse (= chromatine active) : transcription importante
mitochondrie : synthèse d'ATPlame basale
Relation structure-fonctionPolarisation structurale : le noyau, les mitochondries, le
reticulum endoplasmique sont vers la face basale (extérieur de l'acinus), alors que l'appareil de Golgi et les grains de zymogène sont vers la face apicale (du côté de la lumière de l'acinus)
Polarisation fonctionnelle : les acides aminés entrent par la face basale, forment les protéines enzymatiques dans le reticulum, puis ces protéines sont transférées par des vésicules de transition vers l'appareil de Golgi, qui les trie et les modifie (hydroxylations, glycosylations...) ; enfin, les enzymes sont stockées dans les vésicules de sécrétion jusqu'à l'exocytose
organisation cellulaire et moléculaire du m
onde vivant : 2 (P. D
ieumegard)

Compartimentation de la cellule acineuse pancréatique
Compartimentation = division en compartiments
Chaque compartiment a une structure, une composition et un fonctionnement qui lui sont propres
Les compartiments sont délimités par des membranes
Compartiment cytosol = hyaloplasme
C'est un « carrefour métabolique » : pour passer d'un compartiment à un autre, il faut passer par le cytosol.
Rôles : glycolyse, dégradation des acides aminés, synthèse des protéines, structuration de la cellule grâce au cytosquelette.
Le Ca2+ est un effecteur important des protéines cytosoliques.
Compartiment reticulum endoplasmique RE (+ espace intermembranaire nucléaire)
Récupération des protéines comportant un signal d'adressage au RE : les granules du RE granuleux sont les ribosomes.
Rôles : stockage de Ca2+, métabolisme des lipides, N-glycosylation de protéines, hydroxylation et solubilisation de molécules toxiques et des hormones stéroïdes...
Compartiments de l'appareil de Golgi
Les divers saccules contiennent des enzymes différentes, et ont donc des rôles différents. La face cis est du côté du noyau, la face trans est du côté de la lumière.
Rôles : O-glycosylation des protéines et allongement des chaines glycosidiques ; synthèse de sphingolipides ; tri et maturation des protéines.
Compartiments vésiculairesVésicules de sécrétion provenant de bourgeonnement de
l'appareil de GolgiEndosomes = phagosomes : vésicules provenant d'endocytoseLysosomes : vésicules contenant des enzymes d'hydrolyses
actives à pH acide. Les lysosomes primaires se forment par bourgeonnement de saccules golgiens, les lysosomes secondaires sont forméspar fusion d'endosomes et de lysosomes primaires.Rôles : hydrolyse de proies ingérées par phagocytose (hétérophagie) ou de vieux organites de la cellule (autophagie).
Caveosomes
Peroxysomes : vésicules contenant des enzymes de peroxydation.Rôles : production et dégradation de peroxyde d'hydrogène (H2O2), oxydation de diverses molécules
Compartiment mitochondriesLes mitochondries sont limitées par une double
membrane : la membrane interne est très spécifique (imperméable, avec molécules de la chaîne respiratoire et ATP-synthases)
Rôles : synthèse d'ATP, cycle de Krebs, synthèse d'AA, de stérols, de phospholipides, β-oxydation des acides gras...
Compartiment noyauIl est en communication avec le compartiment hyaloplasme :
pendant l'interphase les pores nucléaires permettent l'exportation des ARN, l'importation des protéines ribosomales, l'exportation des sous-unités ribosomales ; pendant la mitose, l'enveloppe nucléaire disparaît.
Rôles : stockage d'information génétique (ADN), transcription en ARN et maturation de ces ARN, assemblage des sous-unités des ribosomes dans le nucléole.
organisation cellulaire et moléculaire du m
onde vivant : 3 (P. D
ieumegard)
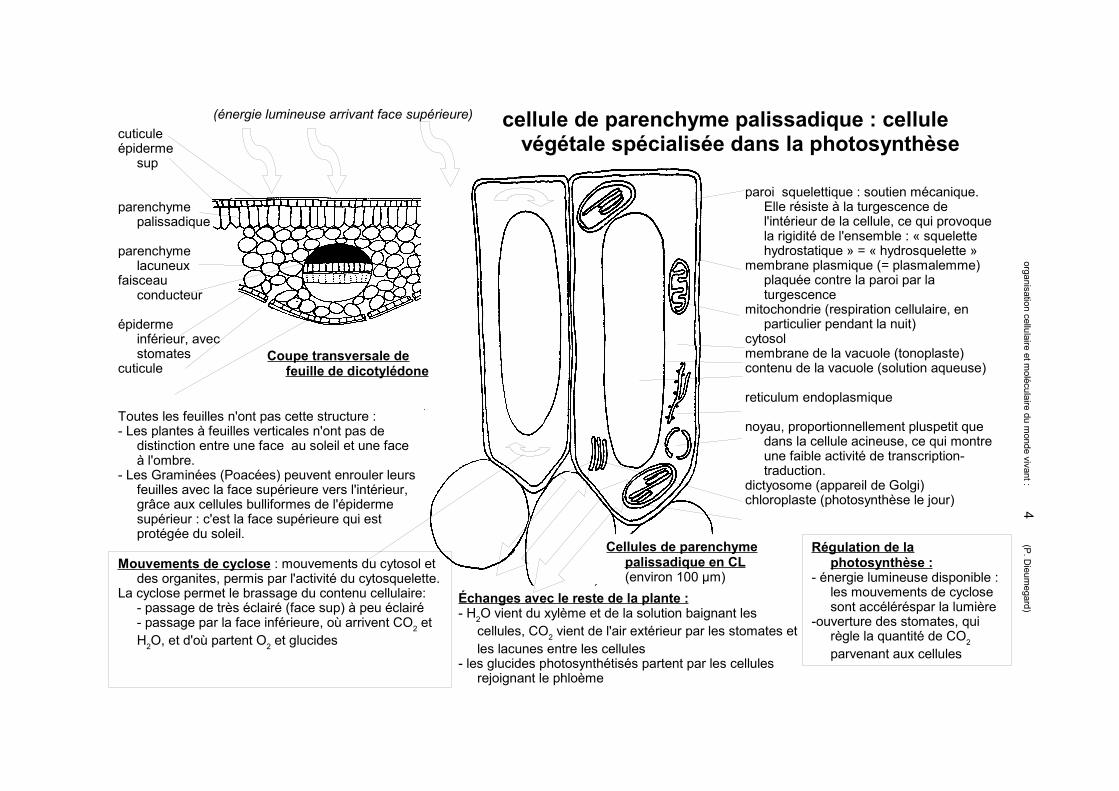
cellule de parenchyme palissadique : cellule végétale spécialisée dans la photosynthèse
(énergie lumineuse arrivant face supérieure)cuticuleépiderme
sup
parenchyme palissadique
parenchyme lacuneux
faisceau conducteur
épiderme inférieur, avec stomates
cuticuleCoupe transversale de
feuille de dicotylédone
Toutes les feuilles n'ont pas cette structure :- Les plantes à feuilles verticales n'ont pas de
distinction entre une face au soleil et une face à l'ombre.
- Les Graminées (Poacées) peuvent enrouler leurs feuilles avec la face supérieure vers l'intérieur, grâce aux cellules bulliformes de l'épiderme supérieur : c'est la face supérieure qui est protégée du soleil.
paroi squelettique : soutien mécanique. Elle résiste à la turgescence de l'intérieur de la cellule, ce qui provoque la rigidité de l'ensemble : « squelette hydrostatique » = « hydrosquelette »
membrane plasmique (= plasmalemme) plaquée contre la paroi par la turgescence
mitochondrie (respiration cellulaire, en particulier pendant la nuit)
cytosolmembrane de la vacuole (tonoplaste)contenu de la vacuole (solution aqueuse)
reticulum endoplasmique
noyau, proportionnellement pluspetit que dans la cellule acineuse, ce qui montre une faible activité de transcription-traduction.
dictyosome (appareil de Golgi)chloroplaste (photosynthèse le jour)
Échanges avec le reste de la plante :- H2O vient du xylème et de la solution baignant les
cellules, CO2 vient de l'air extérieur par les stomates et les lacunes entre les cellules
- les glucides photosynthétisés partent par les cellules rejoignant le phloème
Régulation de la photosynthèse :
- énergie lumineuse disponible : les mouvements de cyclose sont accéléréspar la lumière
-ouverture des stomates, qui règle la quantité de CO2 parvenant aux cellules
Mouvements de cyclose : mouvements du cytosol et des organites, permis par l'activité du cytosquelette.
La cyclose permet le brassage du contenu cellulaire:- passage de très éclairé (face sup) à peu éclairé- passage par la face inférieure, où arrivent CO2 et H2O, et d'où partent O2 et glucides
Cellules de parenchyme palissadique en CL(environ 100 µm)
organisation cellulaire et moléculaire du m
onde vivant : 4 (P. D
ieumegard)

Escherichia coli (colibacille) : une Eubactérie
bacille = bactérie en forme de bâtonnet
coli- = du côlon = de l'intestin
classification phylogénétique (d'après la séquence d'ARNr)
EubactériesEucaryotes
Archébactéries« Procaryotes »
« Procaryotes » :- pas de noyau limité par une enveloppe
nucléaire ; l'ADN est sous forme de « chromosome » circulaire, c'est à dire fermé sur lui-même.
- pas de flux endomembranaire- pas de mitose, de méiose ni de
fécondation (pas de sexualité) ; on les considère comme haploïdes (un seul exemplaire de chaque gène.
- division par étranglement, après réplication du matériel génétique.
Autres caractères des Eubactéries par rapport aux Eucaryotes :
- pas de stérols dans les membranes, mais des cardiolipides (même rôle de stabilisation)
- ribosomes plus petits- gènes sans introns ni exons- gènes souvent groupés en opérons
Nutrition par diffusion (pas de phagocytose) : sécrétion d'enzymes hydrolysant les molécules, et absorption des petites molécules.
Paroi utile pour lutter contre l'éclatement par turgescence (la pénicilline, le premier antibiotique connu, agit en luttant contre la synthèse de paroi).
Schéma de colibacille (environ 1 µm)
nucléoïde (région contenant l'ADN, mais non limitée par une enveloppe nucléaire)
ARN messager, avec ribosomes (synthèse de protéines)
plasmide (ADN différent du chromosome principal, portant quelques gènes), en nombre variable
membrane plasmique
paroi de peptidoglycane
(espace périplasmique)
membrane externe (hémimembrane interne riche en lipoprotéines liées à la paroi ; hémimembrane externe riche en lipopolysaccharides et en porines)
Deux catégories traditionnelles d'Eubactéries :- Gram + : bien colorées par méthode de Gram, car la paroi
de peptidoglycanes est épaisse, et il n'existe pas de membrane externe (exemple Staphylococcus aureus)
- Gram - : mal colorées par la coloration de Gram, car la paroi est fine et il existe une membrane externe (exemple Escherichia coli)
organisation cellulaire et moléculaire du m
onde vivant : 5 (P. D
ieumegard)
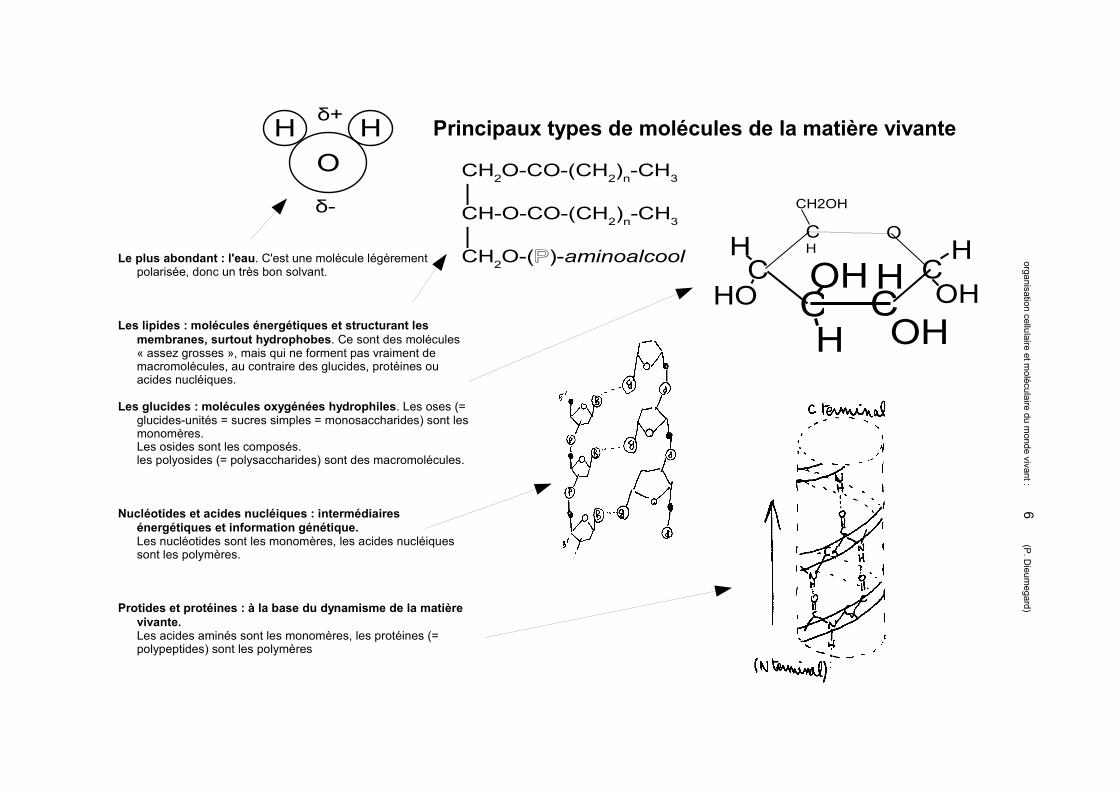
Principaux types de molécules de la matière vivante
Le plus abondant : l'eau. C'est une molécule légèrement polarisée, donc un très bon solvant.
Les lipides : molécules énergétiques et structurant les membranes, surtout hydrophobes. Ce sont des molécules « assez grosses », mais qui ne forment pas vraiment de macromolécules, au contraire des glucides, protéines ou acides nucléiques.
Les glucides : molécules oxygénées hydrophiles. Les oses (= glucides-unités = sucres simples = monosaccharides) sont les monomères.Les osides sont les composés.les polyosides (= polysaccharides) sont des macromolécules.
Nucléotides et acides nucléiques : intermédiaires énergétiques et information génétique. Les nucléotides sont les monomères, les acides nucléiques sont les polymères.
Protides et protéines : à la base du dynamisme de la matière vivante.Les acides aminés sont les monomères, les protéines (= polypeptides) sont les polymères
HO
Hδ+
δ-
CH2O-CO-(CH2)n-CH3
│CH-O-CO-(CH2)n-CH3
│CH2O-(PP)-aminoalcool
CH
C CC C
O
CH2OH
H
OHHOH
OH
HHO
H organisation cellulaire et moléculaire du m
onde vivant : 6 (P. D
ieumegard)

L'eau, H2O,une petite molécule polarisée
M+
Quantitativement importante :60% en masse du corps humain (80% dans les
muscles, 20% dans les os).Cette proportion diminue avec l'âge : 90% dans un foetus de trois mois, 70% dans un nouveau -né.
Chez l'humain, une déshydratation de 10% peut provoquer la mort.
Les organismes en vie ralentie sont déshydratés (une graine sèche contient environ 10% d'eau).
Pôle positif vers les hydrogènes, pôle négatif vers l'oxygène
On peut même considérer qu'une petite proportion de l'eau est cassée en un ion H+ (= H3O+) et un ion OH- ; normalement, dans l'eau pure, [H+] = [OH-] = 10-7.
Cette polarisation permet la formation de liaisons hydrogène :Liaisons entre les molécules d'eauL'attirance entre les molécules d'eau est à l'origine de plusieurs
phénomènes importants :- forte chaleur de vaporisation (2,3 kJ/mole), puisque la
vaporisation est la rupture des liaisons entre les molécules liquides, qui se séparent à l'état de gaz.
- forte chaleur de fusion (6 kJ/mole)
Liaisons entre les molécules d'eau et les solutés-Les substances hydrosolubles sont hydrophiles, soit
polarisées (groupes -OH, avec H positif et O négatif), soit ionisées.
- Les petits ions minéraux (K+, Na+, etc.)sont en fait entourés d'une couronne d'eau, qui les rend beaucoup plus volumineux que lorsqu'ils sont anhydres. C'est pourquoi ils diffusent moins vite à travers les membranes que l'eau elle-même. Exemples d'ions hydratés (= solvatés) : (K (H2O)4)+
Liaisons entre l'eau et les molécules biologiques- les membranes sont formées de phospholipides associés à
des protéines : l'extérieur de la membrane est hydrophile (partie polaire des lipides, résidus polaires ou ionisés des protéines), alors que l'extérieur est hydrophobe (partie apolaire des lipides, résidus hydrophobes des protéines).
OH H
δ+
δ-Les grosses molécules biologiques sont détruites
par hydrolyse« hydrolyse » = « eau - cassure » : les grosses
molécules sont cassées par l'eau, mais l'eau est elle-même cassée pendant la réaction : un H part d'un côté, et le OH restant de l'autre.
Cette hydrolyse est normalement exoénergétique (spontanée thermodynamiquement). La réaction inverse est la condensation : 2 petites molécules s'unissent pour en former une plus grosse, avec perte de H2O.
Exemples :- hydrolyse des lipides en glycérol et en acides gras- hydrolyse des protéines en acides aminés- hydrolyse des acides nucléiques en nucléotides
monophosphates- hydrolyse des glucides complexes en oses
(R1)-O-(R2)----->(R1)-OH + HO-(R2)
Hydrolyse d'une molécule biologique
organisation cellulaire et moléculaire du m
onde vivant : 7 (P. D
ieumegard)
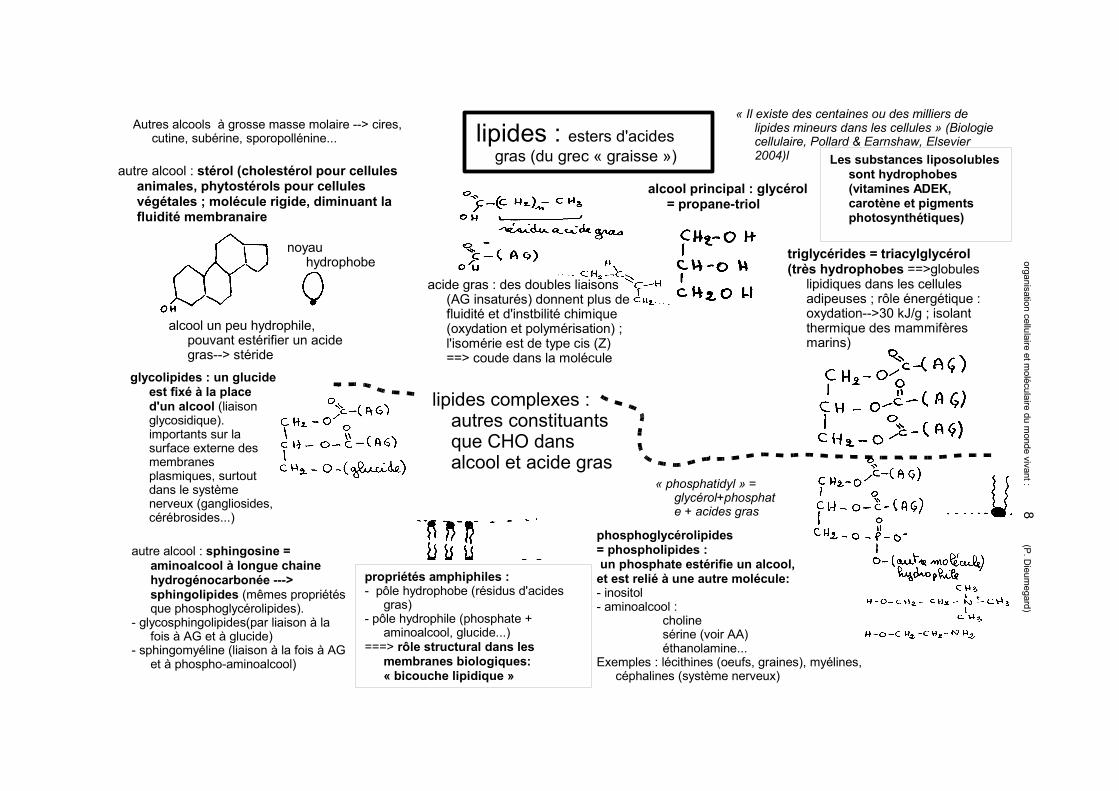
lipides : esters d'acides gras (du grec « graisse »)
« Il existe des centaines ou des milliers de lipides mineurs dans les cellules » (Biologie cellulaire, Pollard & Earnshaw, Elsevier 2004)l
acide gras : des doubles liaisons (AG insaturés) donnent plus de fluidité et d'instbilité chimique (oxydation et polymérisation) ; l'isomérie est de type cis (Z) ==> coude dans la molécule
alcool principal : glycérol = propane-triol
triglycérides = triacylglycérol(très hydrophobes ==>globules
lipidiques dans les cellules adipeuses ; rôle énergétique : oxydation-->30 kJ/g ; isolant thermique des mammifères marins)
Autres alcools à grosse masse molaire --> cires, cutine, subérine, sporopollénine...
autre alcool : stérol (cholestérol pour cellules animales, phytostérols pour cellules végétales ; molécule rigide, diminuant la fluidité membranaire
noyau hydrophobe
alcool un peu hydrophile, pouvant estérifier un acide gras--> stéride
lipides complexes : autres constituants que CHO dans alcool et acide gras
glycolipides : un glucide est fixé à la place d'un alcool (liaison glycosidique). importants sur la surface externe des membranes plasmiques, surtout dans le système nerveux (gangliosides, cérébrosides...)
autre alcool : sphingosine = aminoalcool à longue chaine hydrogénocarbonée ---> sphingolipides (mêmes propriétés que phosphoglycérolipides).
- glycosphingolipides(par liaison à la fois à AG et à glucide)
- sphingomyéline (liaison à la fois à AG et à phospho-aminoalcool)
propriétés amphiphiles :- pôle hydrophobe (résidus d'acides
gras) - pôle hydrophile (phosphate +
aminoalcool, glucide...)===> rôle structural dans les
membranes biologiques: « bicouche lipidique »
phosphoglycérolipides = phospholipides : un phosphate estérifie un alcool, et est relié à une autre molécule:- inositol- aminoalcool :
cholinesérine (voir AA)éthanolamine...
Exemples : lécithines (oeufs, graines), myélines, céphalines (système nerveux)
« phosphatidyl » = glycérol+phosphate + acides gras
Les substances liposolubles sont hydrophobes (vitamines ADEK, carotène et pigments photosynthétiques)
organisation cellulaire et moléculaire du m
onde vivant : 8 (P. D
ieumegard)
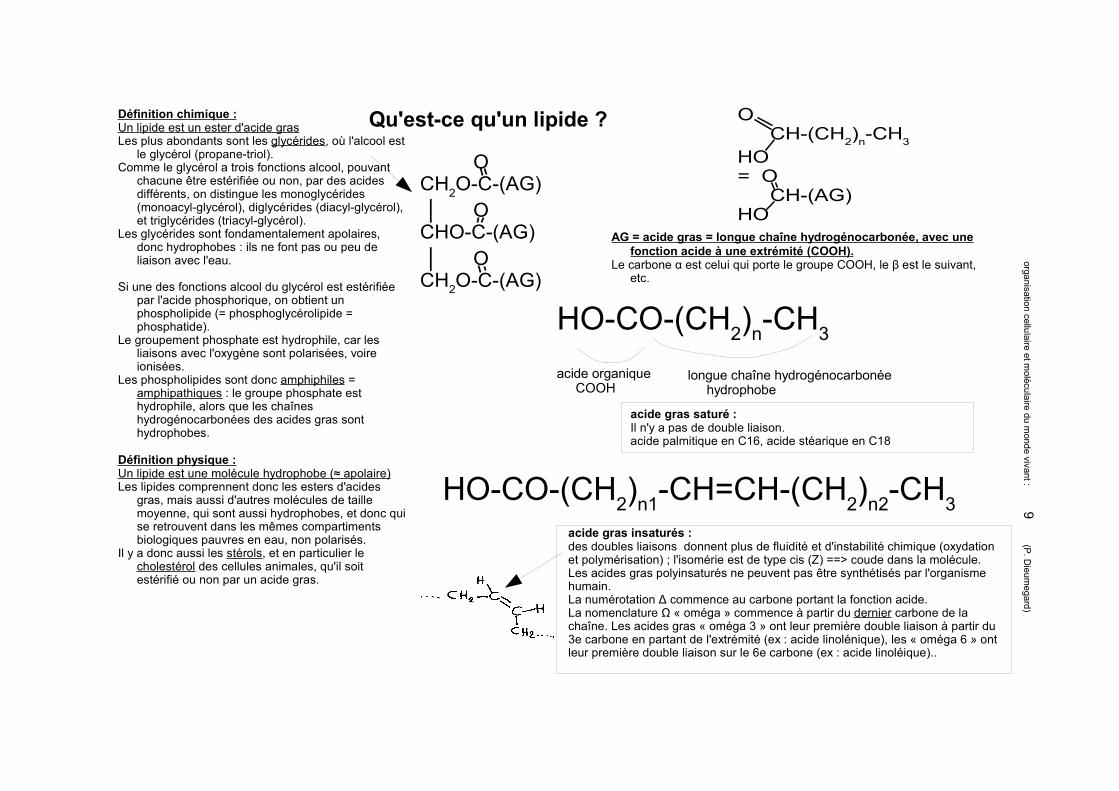
Qu'est-ce qu'un lipide ?Définition chimique :Un lipide est un ester d'acide grasLes plus abondants sont les glycérides, où l'alcool est
le glycérol (propane-triol).Comme le glycérol a trois fonctions alcool, pouvant
chacune être estérifiée ou non, par des acides différents, on distingue les monoglycérides (monoacyl-glycérol), diglycérides (diacyl-glycérol), et triglycérides (triacyl-glycérol).
Les glycérides sont fondamentalement apolaires, donc hydrophobes : ils ne font pas ou peu de liaison avec l'eau.
Si une des fonctions alcool du glycérol est estérifiée par l'acide phosphorique, on obtient un phospholipide (= phosphoglycérolipide = phosphatide).
Le groupement phosphate est hydrophile, car les liaisons avec l'oxygène sont polarisées, voire ionisées.
Les phospholipides sont donc amphiphiles = amphipathiques : le groupe phosphate est hydrophile, alors que les chaînes hydrogénocarbonées des acides gras sont hydrophobes.
Définition physique :Un lipide est une molécule hydrophobe (≈ apolaire)Les lipides comprennent donc les esters d'acides
gras, mais aussi d'autres molécules de taille moyenne, qui sont aussi hydrophobes, et donc qui se retrouvent dans les mêmes compartiments biologiques pauvres en eau, non polarisés.
Il y a donc aussi les stérols, et en particulier le cholestérol des cellules animales, qu'il soit estérifié ou non par un acide gras.
AG = acide gras = longue chaîne hydrogénocarbonée, avec une fonction acide à une extrémité (COOH).
Le carbone α est celui qui porte le groupe COOH, le β est le suivant, etc.
acide gras insaturés : des doubles liaisons donnent plus de fluidité et d'instabilité chimique (oxydation et polymérisation) ; l'isomérie est de type cis (Z) ==> coude dans la molécule.Les acides gras polyinsaturés ne peuvent pas être synthétisés par l'organisme humain.La numérotation Δ commence au carbone portant la fonction acide.La nomenclature Ω « oméga » commence à partir du dernier carbone de la chaîne. Les acides gras « oméga 3 » ont leur première double liaison à partir du 3e carbone en partant de l'extrémité (ex : acide linolénique), les « oméga 6 » ont leur première double liaison sur le 6e carbone (ex : acide linoléique)..
acide gras saturé :Il n'y a pas de double liaison.acide palmitique en C16, acide stéarique en C18
OCH2O-C-(AG) │ OCHO-C-(AG)
│ OCH2O-C-(AG)
HO-CO-(CH2)n-CH3
acide organique COOH
longue chaîne hydrogénocarbonée hydrophobe
HO-CO-(CH2)n1-CH=CH-(CH2)n2-CH3
OCH-(CH2)n-CH3
HO= O
CH-(AG)HO
organisation cellulaire et moléculaire du m
onde vivant : 9 (P. D
ieumegard)
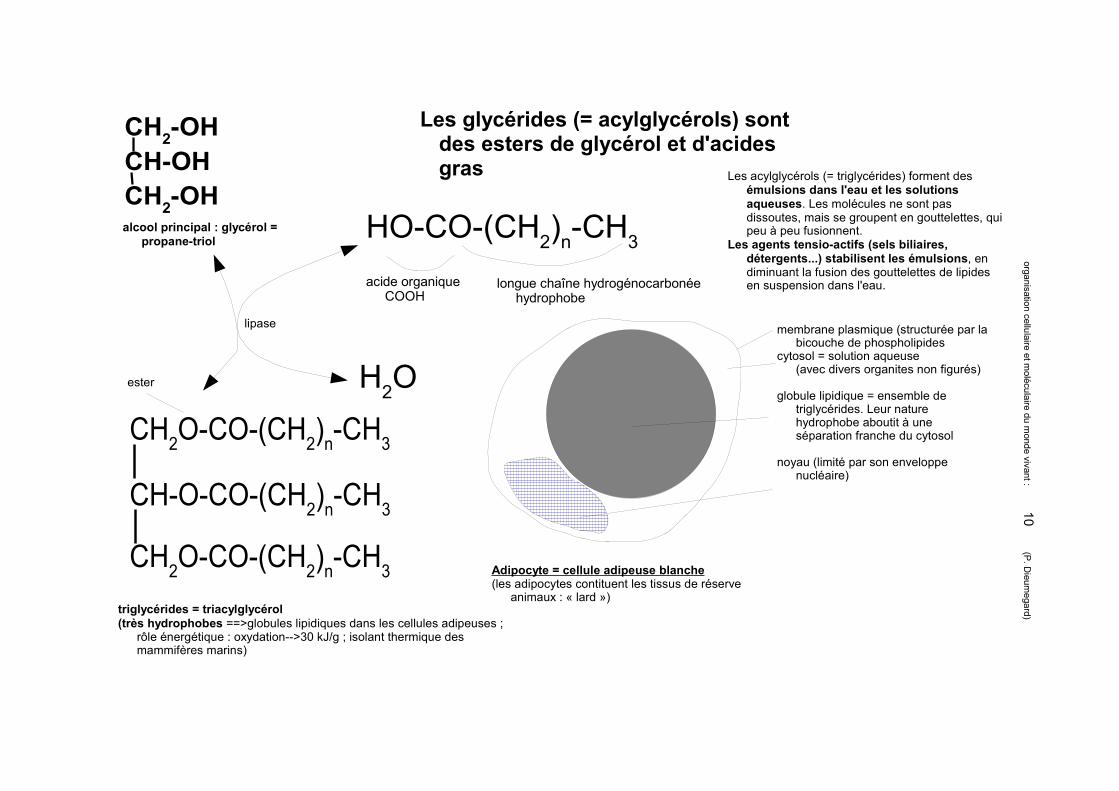
Les glycérides (= acylglycérols) sont des esters de glycérol et d'acides gras
alcool principal : glycérol = propane-triol
CH2-OHCH-OH
CH2-OH
triglycérides = triacylglycérol(très hydrophobes ==>globules lipidiques dans les cellules adipeuses ;
rôle énergétique : oxydation-->30 kJ/g ; isolant thermique des mammifères marins)
HO-CO-(CH2)n-CH3
acide organique COOH
longue chaîne hydrogénocarbonée hydrophobe
CH2O-CO-(CH2)n-CH3
CH-O-CO-(CH2)n-CH3
CH2O-CO-(CH2)n-CH3
H2O
lipase
ester
Les acylglycérols (= triglycérides) forment des émulsions dans l'eau et les solutions aqueuses. Les molécules ne sont pas dissoutes, mais se groupent en gouttelettes, qui peu à peu fusionnent.
Les agents tensio-actifs (sels biliaires, détergents...) stabilisent les émulsions, en diminuant la fusion des gouttelettes de lipides en suspension dans l'eau.
Adipocyte = cellule adipeuse blanche(les adipocytes contituent les tissus de réserve
animaux : « lard »)
membrane plasmique (structurée par la bicouche de phospholipides
cytosol = solution aqueuse(avec divers organites non figurés)
globule lipidique = ensemble de triglycérides. Leur nature hydrophobe aboutit à une séparation franche du cytosol
noyau (limité par son enveloppe nucléaire)
organisation cellulaire et moléculaire du m
onde vivant : 10 (P. D
ieumegard)
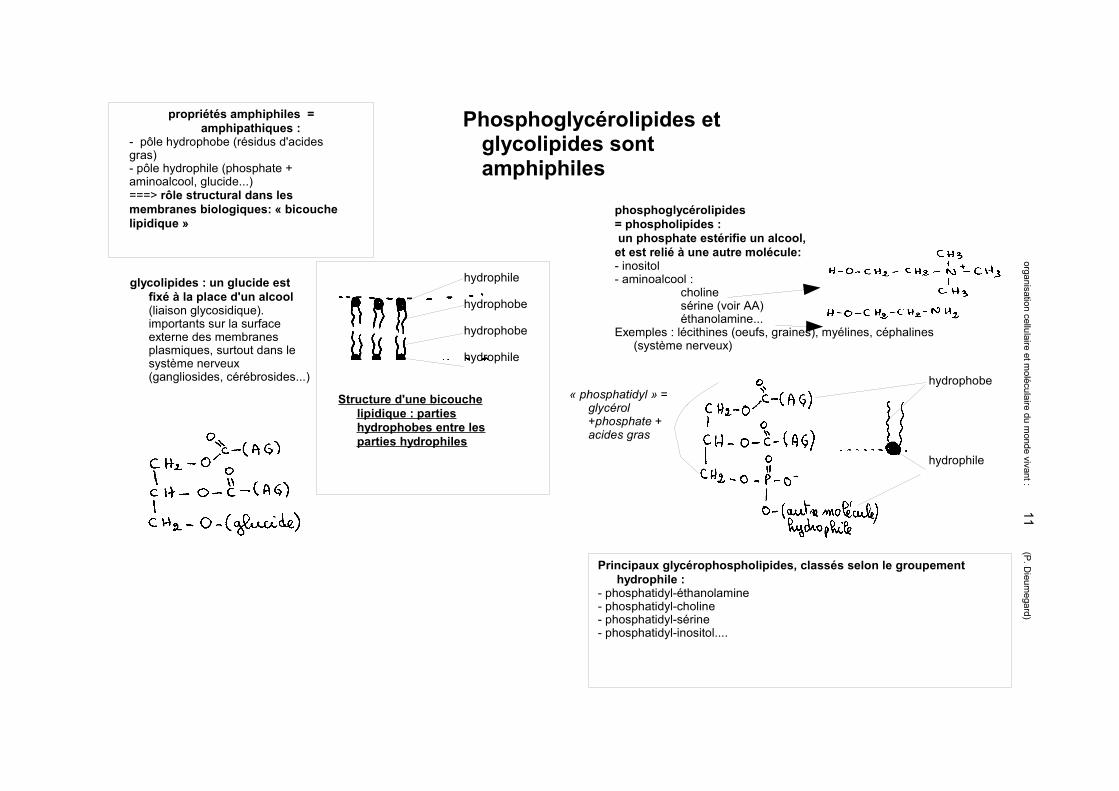
Phosphoglycérolipides et glycolipides sont amphiphiles
phosphoglycérolipides = phospholipides : un phosphate estérifie un alcool, et est relié à une autre molécule:- inositol- aminoalcool :
cholinesérine (voir AA)éthanolamine...
Exemples : lécithines (oeufs, graines), myélines, céphalines (système nerveux)
« phosphatidyl » = glycérol +phosphate + acides gras
glycolipides : un glucide est fixé à la place d'un alcool (liaison glycosidique). importants sur la surface externe des membranes plasmiques, surtout dans le système nerveux (gangliosides, cérébrosides...)
propriétés amphiphiles = amphipathiques :
- pôle hydrophobe (résidus d'acides gras) - pôle hydrophile (phosphate + aminoalcool, glucide...)===> rôle structural dans les membranes biologiques: « bicouche lipidique »
hydrophile
hydrophobe
hydrophobe
hydrophile
Structure d'une bicouche lipidique : parties hydrophobes entre les parties hydrophiles
Principaux glycérophospholipides, classés selon le groupement hydrophile :
- phosphatidyl-éthanolamine- phosphatidyl-choline- phosphatidyl-sérine- phosphatidyl-inositol....
hydrophobe
hydrophile
organisation cellulaire et moléculaire du m
onde vivant : 11 (P. D
ieumegard)

autre alcool : stérol (cholestérol pour cellules animales, phytostérols pour cellules végétales) ; molécule rigide, diminuant la fluidité membranaire
noyau hydrophobe
alcool un peu hydrophile, pouvant estérifier un acide gras--> stéride
autre alcool : sphingosine = aminoalcool à longue chaine hydrogénocarbonée ---> sphingolipides (mêmes propriétés que phosphoglycérolipides).
- glycosphingolipides(par liaison à la fois à AG et à glucide)
- sphingomyéline (liaison à la fois à AG et à phospho-aminoalcool)
Autres alcools à grosse masse molaire --> cires, cutine, subérine, sporopollénine... (cuticule des cellules végétales)
« Il existe des centaines ou des milliers de lipides mineurs dans les cellules » (Biologie cellulaire, Pollard & Earnshaw, Elsevier 2004)
lipides complexes : autres constituants que CHO dans alcool et acide gras
Autres lipides...organisation cellulaire et m
oléculaire du monde vivant : 12 (P
. Dieum
egard)

glucides = « hydrates de carbone »formule brute du type Cn(H2O)n'
Monomères : les oses sont des polyalcools avec un groupe carbonyle (aldéhyde ou cétone)(exemple : le glucose)
Oligomères : les oligosides = oses liés par des liaisons osidiquesExemples :- saccharose = glucose - fructose- maltose = glucose – glucose
Polymères : les polyosidesExemples :- amidon et glycogène (rôle énergétique)- cellulose (rôle structural)
Propriétés optiques : pouvoir rotatoireLes solutions d'oses et osides dévient le plan de vibration de la
lumière polarisée. Cette propriété provient des carbones asymétriques (ayant 4 liaisons avec 4 groupements différents).
Le pouvoir rotatoire est proportionnel à la concentration de la solution de glucide :
Loi de Biot : α = α0 C loù α est l'angle de rotation, α0 est le pouvoir rotatoire spécifique, C
est la concentration et l est la longueur de solution traversée.
Glucides (du grec « sucre »)« hydrates de carbone » : formule (H-C-OH)n
CH2OH
O
OHOH
HO
OH
α glucose CH2OH
HOCH2 O
HO
HO
O
CH2OH
O
OHHO
OH
saccharose = α glucose 1-4 β fructose
CH2OHCH2OHO
OHOH
OHO
O
OHHO
OH
maltose = α glucose 1-4 α glucose
OH
OH
O
OH
HOO
OHOH
OH
OH
O
lactose = β galactose 1-4 glucose
organisation cellulaire et moléculaire du m
onde vivant : 13 (P. D
ieumegard)

Glucides (du grec « sucre »)« hydrates de carbone » : for mule (H-C-
OH)
Oses = polyalcools non ramifiés, dont une fonction alcool est oxydée en carbonyle (C=O, aldéhyde ou cétone). Souvent cyclisés ==> disparition du carbonyle (« semi-aldéhyde » ou « semi-cétone »).
Nombreux isomères.
glucose (αD glucopyranose) (aldose)
si CH2OH en bas : forme L (non naturelle)
si OH de C2 ou C3 ailleurs :
mannose ou autres isomères
si OH en haut : galactose
si OH en haut : β
si COO- au lieu de CH2OH : acides uroniques (glycuronique pour glucose, galacturonique pour galactose...)
Osides : liaison osidique avec perte de H 2O
condensationhydrolyse
fructose : β D fructofuranose (cétose)
(voir glycolyse et fructose 1-6 diphosphate)
trioses : glycéraldéhydeetdihyroxyacétone(voir glycolyse)
pentoses : ribose et désoxyribose (voir
acides nucléiques)ribulose 1-5 diphosphate : voir
photosynthèse
β D ribofuranose (aldose)
diholosides : exemple maltose (α glucose 1-4 αglucose)
lactose : α galactose 1-4 glucosesaccharose = sucrose : αglucose 1-1 βfructose
polyosides : nommés d'après l'ose majoritaire : arabanes, xylanes, glucosanes, fructosanes, galactanes...
polyosides simples :amidon (amyloplastes dans cellules végétales) ≈ glycogène
(cellules animales + Mycètes) : α glucose 1-4 + ramifications en 1-6 (forme enroulée en hélice)
cellulose : β glucose 1-6 dans paroi végétale(forme en fibres droites, liées les unes aux autres )
polyosides hétérogènes : (oses, acides uroniques,
osamines...)- agarose
- polyosides de la paroi bactérienne
- glycosaminoglycannes (= mucopolysaccharides) = protéoglycannes : matrice extracellulaire (exemple chondroïtine sulfate du cartilage)
si -OH remplacé par -NH2 : osaminessi -OH remplacé par -NH-CO-CH3 :
acétyl-osamines (--->chitine des Arthropodes)
organisation cellulaire et moléculaire du m
onde vivant : 14 (P. D
ieumegard)

La molécule de glucose, et différentes façons de la figurer
Le glucose est un aldohexose : 6 atomes de carbone, portant chacun une fonction alcool (C-OH) sauf le carbone 1 qui porte une fonction aldéhyde.
Mais, comme c'est normal pour un hexose, la molécule est plus stable de façon cyclisée. Le cycle est à 6 côtés : l'oxygène du carbone 4 fait un pont avec le carbone 1, qui se retrouve ainsi sans double liaison avec l'oxygène : on parle de fonction « semi-aldéhyde », car les propriétés du groupe aldéhyde ont fortement disparu.
Ce cycle peut exister sous les formes « chaise » et « bateau », mais on y accorde peu d'importance en biologie. Le cycle est représenté sous la forme d'un hexagone régulier. On suppose voir cet hexagone par dessus (c'est pourquoi la liaison entre les carbones 2 et 3, qui est la plus proche de nous, est figurée parfois en traits plus épais), l'oxygène étant au loin et à droite.
La nature « glucose » est donnée par la position des groupes -OH des carbones 2, 3 et 4.
α glucose
Les oses naturels sont de la série « D » : le carbone 6 est au dessus du plan de la molécule.
La cyclisation fait que le carbone 1 est asymétrique. Le glucose α a le OH du carbone 1 au dessous du plan de la molécule, alors que le glucose β a le OH au dessus.
β glucose
dérivés du glucose :- glucosamine : le carbone 2 porte
un -NH2 au lieu du OH.acétylglucosamine : le carbone 2 porte -NH-CO-CH3
- acide glycuronique ou glucuronique : le carbone 6 est un acide (COOH) et non un alcool (CH2-OH)
- glucose 6 sulfate : le carbone 6 est un ester minéral CH2OSO3H
NH2
COOH
C2C3C4C5C6 OHH
H
OHHOHHHHOOHH
C1OH
CH2OH
O
OH
OH
HO
OH Pour simplifier l'écriture : on figure les groupes OH par un simple trait, les carbones sont simplement les angles entre les liaisons, et on ne figure pas les hydrogènes.
Numérotation des carbones à partir de l'extrémité portant la fonction carbonyle C=O en forme linéaire (à droite de l'oxygène en forme cyclisée).
organisation cellulaire et moléculaire du m
onde vivant : 15 (P. D
ieumegard)
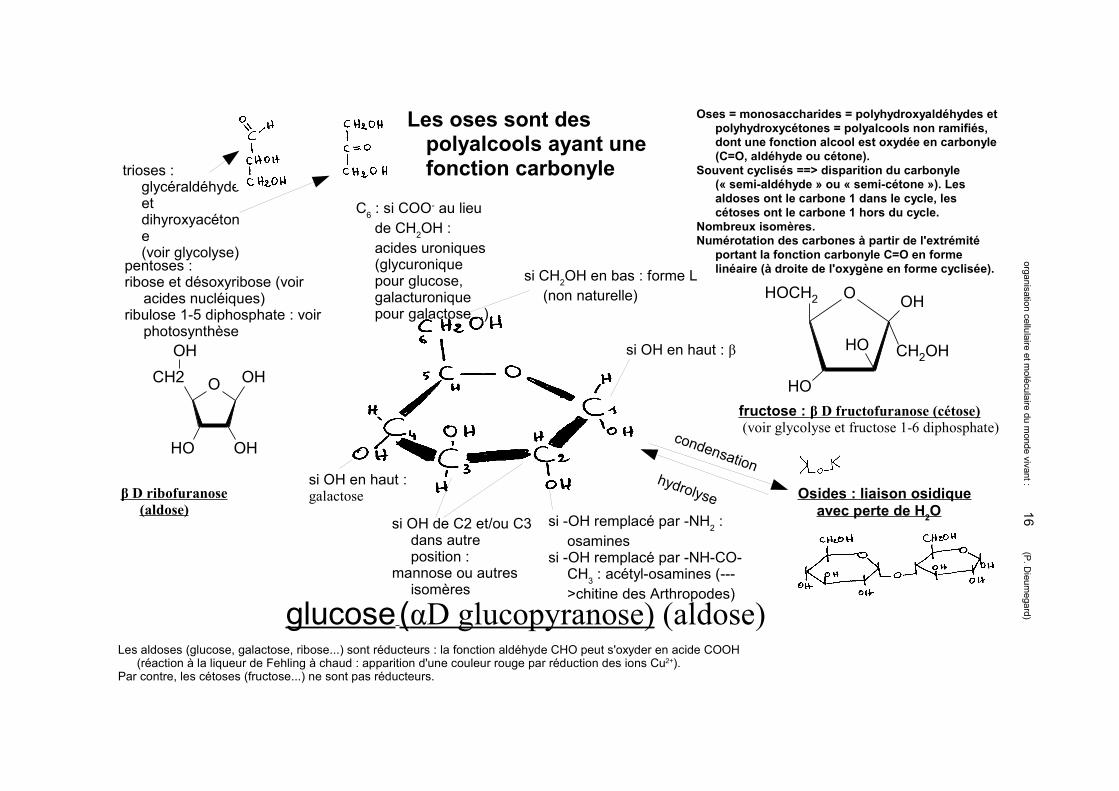
Les oses sont des polyalcools ayant une fonction carbonyle
si CH2OH en bas : forme L (non naturelle)
si OH en haut : β
C6 : si COO- au lieu de CH2OH : acides uroniques (glycuronique pour glucose, galacturonique pour galactose...)
si OH de C2 et/ou C3 dans autre position :
mannose ou autres isomères
si OH en haut : galactose
si -OH remplacé par -NH2 : osamines
si -OH remplacé par -NH-CO-CH3 : acétyl-osamines (--->chitine des Arthropodes)
condensationhydrolyse Osides : liaison osidique
avec perte de H 2O
glucose (αD glucopyranose) (aldose)
β D ribofuranose (aldose)
trioses : glycéraldéhydeetdihyroxyacétone(voir glycolyse)
pentoses : ribose et désoxyribose (voir
acides nucléiques)ribulose 1-5 diphosphate : voir
photosynthèse
Oses = monosaccharides = polyhydroxyaldéhydes et polyhydroxycétones = polyalcools non ramifiés, dont une fonction alcool est oxydée en carbonyle (C=O, aldéhyde ou cétone).
Souvent cyclisés ==> disparition du carbonyle (« semi-aldéhyde » ou « semi-cétone »). Les aldoses ont le carbone 1 dans le cycle, les cétoses ont le carbone 1 hors du cycle.
Nombreux isomères.Numérotation des carbones à partir de l'extrémité
portant la fonction carbonyle C=O en forme linéaire (à droite de l'oxygène en forme cyclisée).
fructose : β D fructofuranose (cétose) (voir glycolyse et fructose 1-6 diphosphate)
Les aldoses (glucose, galactose, ribose...) sont réducteurs : la fonction aldéhyde CHO peut s'oxyder en acide COOH (réaction à la liqueur de Fehling à chaud : apparition d'une couleur rouge par réduction des ions Cu2+).
Par contre, les cétoses (fructose...) ne sont pas réducteurs.
CH2OH
HOCH2 O
HO
HO
OH
O OH
OHHO
CH2OH
organisation cellulaire et moléculaire du m
onde vivant : 16 (P. D
ieumegard)
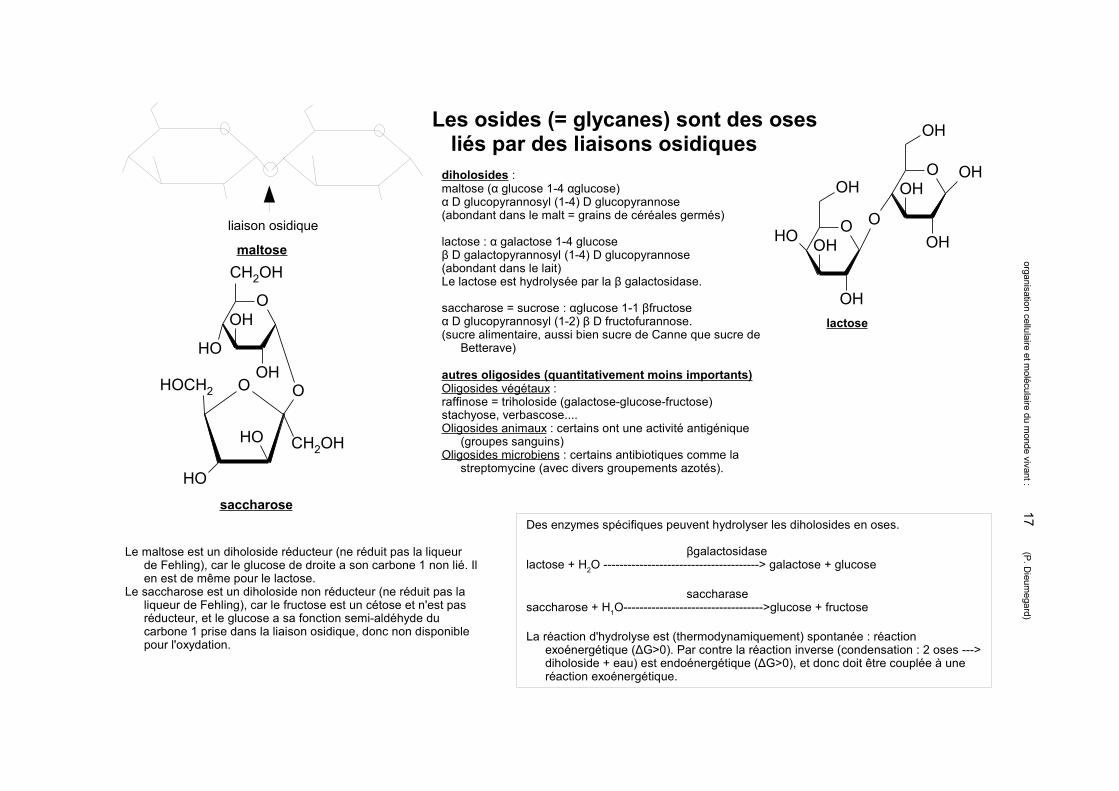
Les osides (= glycanes) sont des oses liés par des liaisons osidiques
diholosides : maltose (α glucose 1-4 αglucose)α D glucopyrannosyl (1-4) D glucopyrannose(abondant dans le malt = grains de céréales germés)
lactose : α galactose 1-4 glucoseβ D galactopyrannosyl (1-4) D glucopyrannose(abondant dans le lait)Le lactose est hydrolysée par la β galactosidase.
saccharose = sucrose : αglucose 1-1 βfructoseα D glucopyrannosyl (1-2) β D fructofurannose.(sucre alimentaire, aussi bien sucre de Canne que sucre de
Betterave)
autres oligosides (quantitativement moins importants)Oligosides végétaux :raffinose = triholoside (galactose-glucose-fructose)stachyose, verbascose....Oligosides animaux : certains ont une activité antigénique
(groupes sanguins)Oligosides microbiens : certains antibiotiques comme la
streptomycine (avec divers groupements azotés).
liaison osidique
maltose OH
OH
O
OH
HOO
OHOH
OH
OH
O
lactose
CH2OH
HOCH2 O
HO
HO
O
CH2OH
O
OHHO
OH
saccharoseDes enzymes spécifiques peuvent hydrolyser les diholosides en oses.
βgalactosidaselactose + H2O ---------------------------------------> galactose + glucose
saccharasesaccharose + H1O----------------------------------->glucose + fructose
La réaction d'hydrolyse est (thermodynamiquement) spontanée : réaction exoénergétique (ΔG>0). Par contre la réaction inverse (condensation : 2 oses ---> diholoside + eau) est endoénergétique (ΔG>0), et donc doit être couplée à une réaction exoénergétique.
Le maltose est un diholoside réducteur (ne réduit pas la liqueur de Fehling), car le glucose de droite a son carbone 1 non lié. Il en est de même pour le lactose.
Le saccharose est un diholoside non réducteur (ne réduit pas la liqueur de Fehling), car le fructose est un cétose et n'est pas réducteur, et le glucose a sa fonction semi-aldéhyde du carbone 1 prise dans la liaison osidique, donc non disponible pour l'oxydation.
organisation cellulaire et moléculaire du m
onde vivant : 17 (P. D
ieumegard)

polyosides : nommés d'après l'ose majoritaire : arabanes, xylanes, glucosanes, fructosanes, galactanes...
Les polyosides ne sont pas réducteurs : la seule fonction semi-aldéhyde disponible est en bout de chaîne, et est quantitativement très minoritaire.
polyosides hétérogènes : (oses, acides uroniques,
osamines...)- agarose
- polyosides de la paroi bactérienne
- glycosaminoglycannes (= mucopolysaccharides) = protéoglycannes : matrice extracellulaire.- Chondroïtine sulfate du cartilage : il existe des groupes CH2OSO3H en position 6, ainsi que groupes acétylamine en position 2- Acide hyaluronique du tissu conjonctif
polyosides homogènes :amidon (grains d'amidon dans les
plastes des cellules végétales) ≈ glycogène (cellules animales + Mycètes) : α glucose 1-4 + ramifications en 1-6 (forme enroulée en hélice). L'amidon est une molécule de réserve végétale, le glycogène est une molécule de réserve animale.
cellulose : β glucose 1-6 dans paroi végétale(forme en fibres droites, liées les unes aux autres par de nombreuses liaisons H, ce qui donne une grande rigidité). La cellulose est le constituant de base des parois squelettiques végétales.
chitine = polymère de N-acétylglucosamine (constituant de la cuticule des Arthropodes et des Némathelminthes)
pectine = polymère d'acide galacturonique (constituant des parois végétales, surtout la lamelle moyenne, donnant une consistance gélifiée).
Polyholosides : nombreux oses reliés par de nombreuses liaisons osidiques
Polyosides à rôle structural : cellulose, pectine, chitine...
Polyosides à rôle énergétique : amidon, glycogène, inuline...
O
OH
OH
OHH2C
OO
OH
OH
OHH2C
O
O
OH
OH
OHH2C
O
O
OH
OH
OHH2C
...O O...
Molécule de cellulose : β-glucose en liaison 1-4
Schéma de cellules végétales, montrant divers polyosides à rôle structural et de réserve
(cellule 1) (cellule 2)
paroi de cellule 1 (cellulose)
paroi de cellule 2 (cellulose)
lamelle moyenne (pectine)
grain d'amidon
membranes du plaste
thylakoïdes du plaste
cytosols des cellules
organisation cellulaire et moléculaire du m
onde vivant : 18 (P. D
ieumegard)

Amidon et glycogène : α glucose en liaison 1-4 (ramifié en 1-6)
O
CH2OHCH2OHCH2OH
OH
O
OH
O
O OOHOH
OOH
OH
OH
...OO..
O
OH
CH2OH
CH2OH
O
OH
OHO
...O
OH
CH2OH
O
CH2OH
O
O O
O
OH
OHOO
OHO
O
OH
OH
OH
O..
OH
OH
CH2OH CH2
OH
CH2OH
..O
Amylose = chaînes linéaires d'α-glucose polymérisé en 1-4
Amylopectine : chaînes d'α-glucose polymérisé en 1-4, avec des ramifications en 1-6
Amidon = forme de réserve glucidique des Végétaux
- dans les grains d'amidon (amyloplastes, et un peu dans les chloroplastes très éclairés)
- poudre blanche, insoluble à froid dans l'eau (la suspension d'amidon dans l'eau s'appelle « lait d'amidon »)
- soluble à chaud (à partir de 70°C), en formant une solution colloïdale, l'empois d'amidon.
- coloration bleu-violet à froid avec l'eau iodée
L'amidon naturel est un mélange de deux molécules de structure voisine :
- l'amylose est une molécule linéaire (= non ramifiée), constituée de 200 à 500 résidus de glucose liés en 1-4 par un oxygène (liaison osidique). En réalité les cycles de glucose ne sont pas dans le même plan, et la chaîne forme une hélice.
- l'amylopectine est une molécule ramifiée, de même structure que l'amylose, mais avec des ramifications en 1-6. L'amylopectine peut être constituée de plus de 1000 résidus de glucose.
L'amidon est hydrolysable soit à chaud en milieu acide, soit à froid par des enzymes spécialisées, les amylases (= α-glucosidases).
Les exoamylases coupent à partir de l'extrémité de la chaîne, en libérant surtout des maltoses.
Les endoamylases coupent à l'intérieur de la chaîne, en libérant des osides de plus en plus courts, les dextrines (colorables en rouge, puis en jaune par l'eau iodée).
Le glycogène est un polymère animal, de structure semblable à l'amylopectine (en plus ramifié, et beaucoup plus court : seulement quelques dizaines de résidus glucose). Il est colorable en brun-rouge par l'eau iodée.
Ces α-glucosides ont une structure en hélice : les cycles de glucose successifs ne sont pas dans le même plan, et l'ensemble forme une hélice où peut venir se loger l'iode de l'eau iodée, ce qui provoque une coloration caractéristique.
organisation cellulaire et moléculaire du m
onde vivant : 19 (P. D
ieumegard)
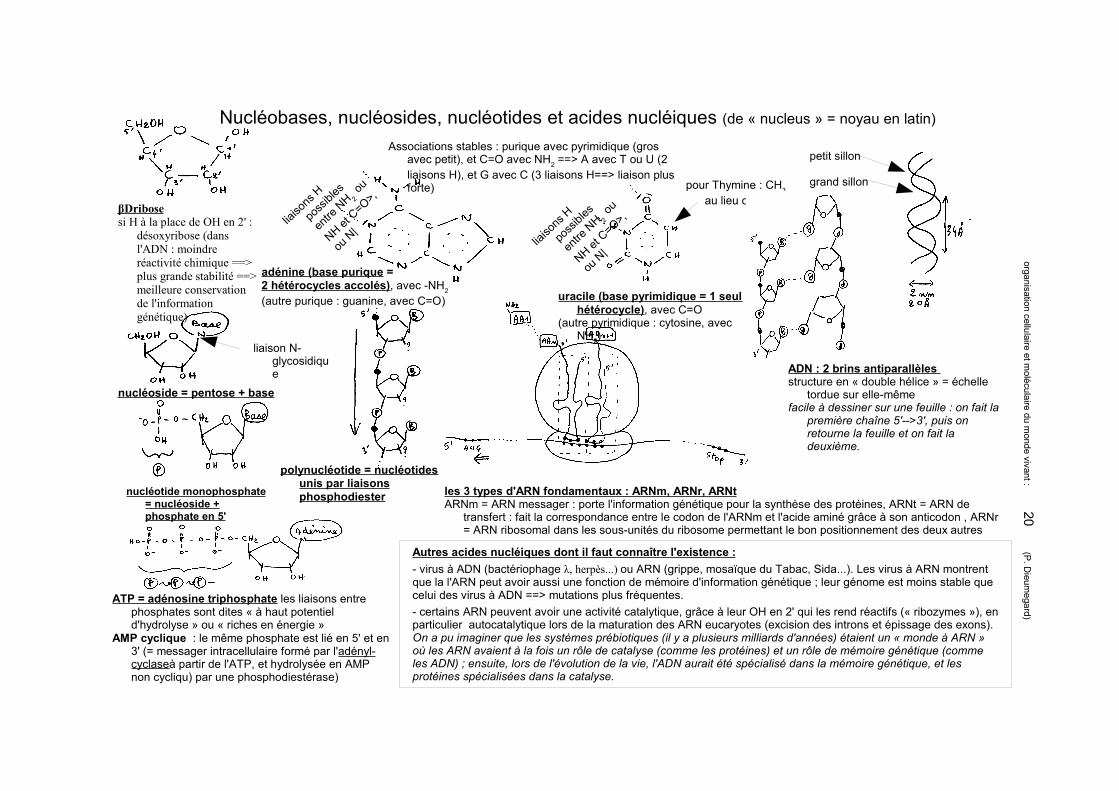
Nucléobases, nucléosides, nucléotides et acides nucléiques (de « nucleus » = noyau en latin)
βDribosesi H à la place de OH en 2' :
désoxyribose (dans l'ADN : moindre réactivité chimique ==> plus grande stabilité ==> meilleure conservation de l'information génétique)
adénine (base purique = 2 hétérocycles accolés), avec -NH2 (autre purique : guanine, avec C=O) uracile (base pyrimidique = 1 seul
hétérocycle), avec C=O(autre pyrimidique : cytosine, avec
NH2)
liaiso
ns H
poss
ibles
entre
NH 2
ou
NH et C
=O>,
ou N
|lia
isons
H
poss
ibles
entre
NH 2
ou
NH et C
=O>,
ou N
|
Associations stables : purique avec pyrimidique (gros avec petit), et C=O avec NH2 ==> A avec T ou U (2 liaisons H), et G avec C (3 liaisons H==> liaison plus forte) pour Thymine : CH3
au lieu de H
liaison N-glycosidique
nucléoside = pentose + base
nucléotide monophosphate = nucléoside + phosphate en 5'
ATP = adénosine triphosphate les liaisons entre phosphates sont dites « à haut potentiel d'hydrolyse » ou « riches en énergie »
AMP cyclique : le même phosphate est lié en 5' et en 3' (= messager intracellulaire formé par l'adényl-cyclaseà partir de l'ATP, et hydrolysée en AMP non cycliqu) par une phosphodiestérase)
polynucléotide = nucléotides unis par liaisons phosphodiester
ADN : 2 brins antiparallèles structure en « double hélice » = échelle
tordue sur elle-mêmefacile à dessiner sur une feuille : on fait la
première chaîne 5'-->3', puis on retourne la feuille et on fait la deuxième.
petit sillon
grand sillon
les 3 types d'ARN fondamentaux : ARNm, ARNr, ARNtARNm = ARN messager : porte l'information génétique pour la synthèse des protéines, ARNt = ARN de
transfert : fait la correspondance entre le codon de l'ARNm et l'acide aminé grâce à son anticodon , ARNr = ARN ribosomal dans les sous-unités du ribosome permettant le bon positionnement des deux autres
Autres acides nucléiques dont il faut connaître l'existence :- virus à ADN (bactériophage λ, herpès...) ou ARN (grippe, mosaïque du Tabac, Sida...). Les virus à ARN montrent que la l'ARN peut avoir aussi une fonction de mémoire d'information génétique ; leur génome est moins stable que celui des virus à ADN ==> mutations plus fréquentes.- certains ARN peuvent avoir une activité catalytique, grâce à leur OH en 2' qui les rend réactifs (« ribozymes »), en particulier autocatalytique lors de la maturation des ARN eucaryotes (excision des introns et épissage des exons). On a pu imaginer que les systèmes prébiotiques (il y a plusieurs milliards d'années) étaient un « monde à ARN » où les ARN avaient à la fois un rôle de catalyse (comme les protéines) et un rôle de mémoire génétique (comme les ADN) ; ensuite, lors de l'évolution de la vie, l'ADN aurait été spécialisé dans la mémoire génétique, et les protéines spécialisées dans la catalyse.
organisation cellulaire et moléculaire du m
onde vivant : 20 (P. D
ieumegard)
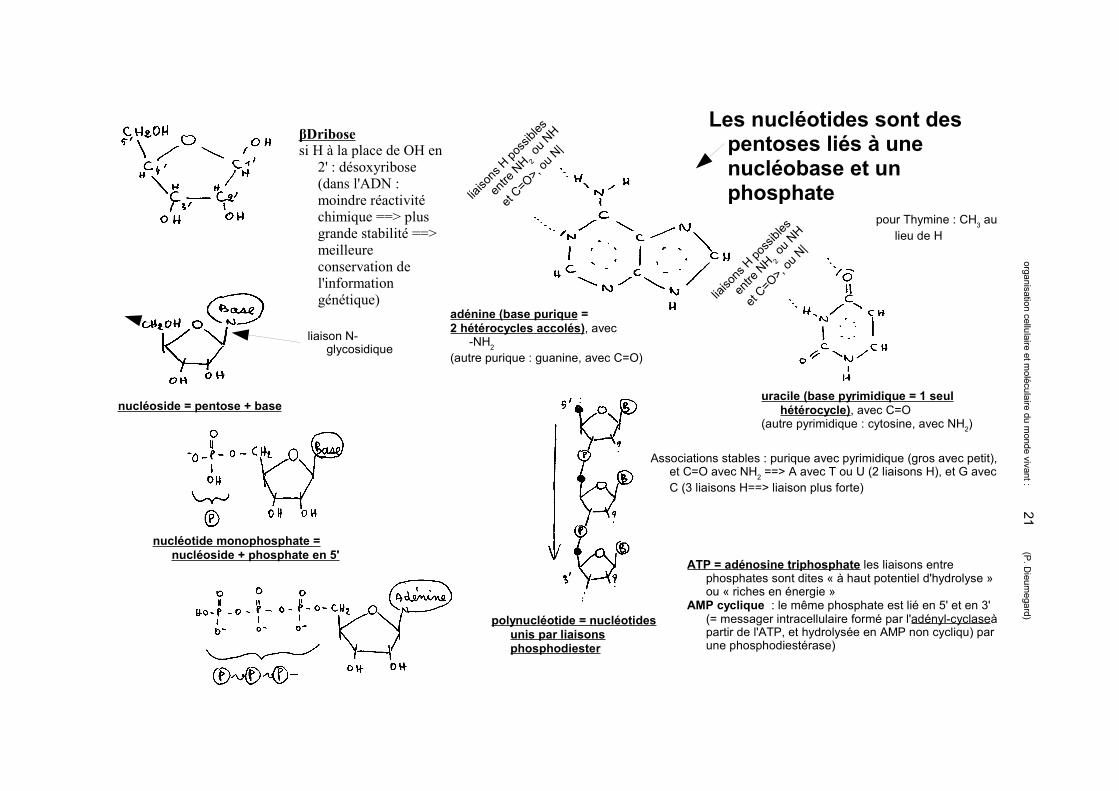
βDribosesi H à la place de OH en
2' : désoxyribose (dans l'ADN : moindre réactivité chimique ==> plus grande stabilité ==> meilleure conservation de l'information génétique)
adénine (base purique = 2 hétérocycles accolés), avec
-NH2 (autre purique : guanine, avec C=O)
uracile (base pyrimidique = 1 seul hétérocycle), avec C=O
(autre pyrimidique : cytosine, avec NH2)
liaiso
ns H
poss
ibles
entre
NH 2
ou N
H
et C=O
>, ou N
|
Associations stables : purique avec pyrimidique (gros avec petit), et C=O avec NH2 ==> A avec T ou U (2 liaisons H), et G avec C (3 liaisons H==> liaison plus forte)
pour Thymine : CH3 au lieu de H
liaison N-glycosidique
nucléoside = pentose + base
nucléotide monophosphate = nucléoside + phosphate en 5'
ATP = adénosine triphosphate les liaisons entre phosphates sont dites « à haut potentiel d'hydrolyse » ou « riches en énergie »
AMP cyclique : le même phosphate est lié en 5' et en 3' (= messager intracellulaire formé par l'adényl-cyclaseà partir de l'ATP, et hydrolysée en AMP non cycliqu) par une phosphodiestérase)
polynucléotide = nucléotides unis par liaisons phosphodiester
liaiso
ns H
poss
ibles
entre
NH 2
ou N
H
et C=O>, o
u N|
Les nucléotides sont des pentoses liés à une nucléobase et un phosphate
organisation cellulaire et moléculaire du m
onde vivant : 21 (P. D
ieumegard)
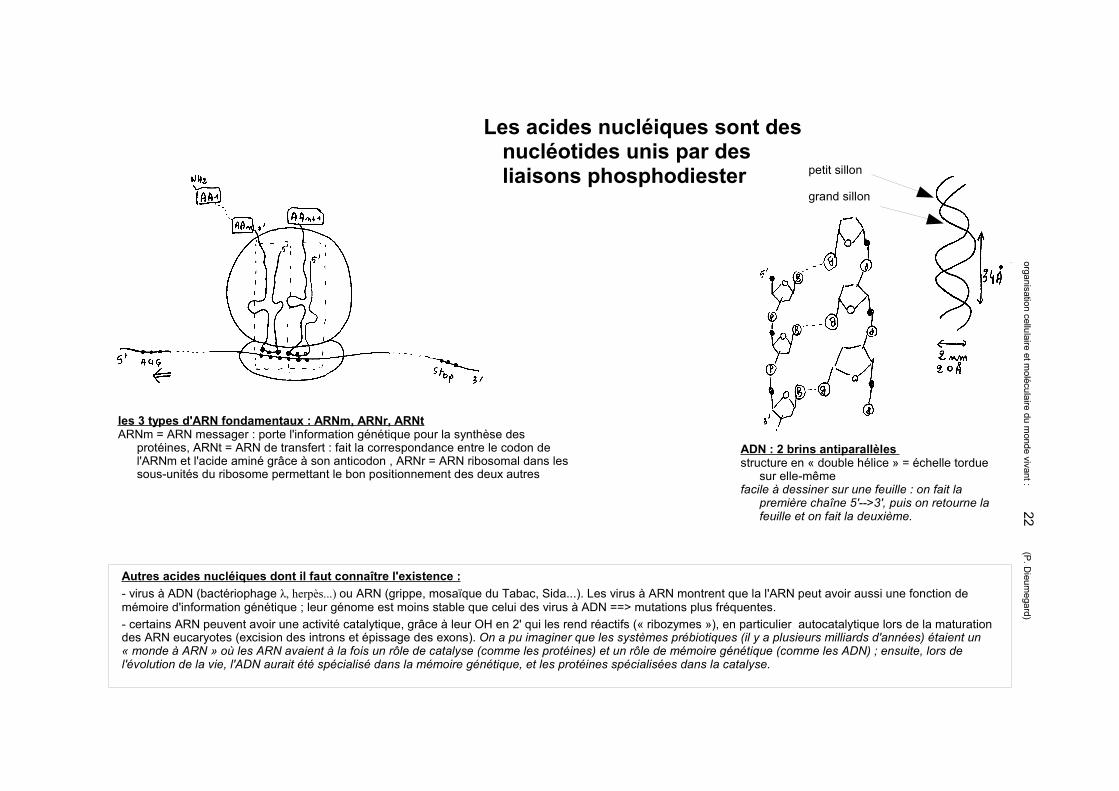
Les acides nucléiques sont des nucléotides unis par des liaisons phosphodiester
Autres acides nucléiques dont il faut connaître l'existence :- virus à ADN (bactériophage λ, herpès...) ou ARN (grippe, mosaïque du Tabac, Sida...). Les virus à ARN montrent que la l'ARN peut avoir aussi une fonction de mémoire d'information génétique ; leur génome est moins stable que celui des virus à ADN ==> mutations plus fréquentes.- certains ARN peuvent avoir une activité catalytique, grâce à leur OH en 2' qui les rend réactifs (« ribozymes »), en particulier autocatalytique lors de la maturation des ARN eucaryotes (excision des introns et épissage des exons). On a pu imaginer que les systèmes prébiotiques (il y a plusieurs milliards d'années) étaient un « monde à ARN » où les ARN avaient à la fois un rôle de catalyse (comme les protéines) et un rôle de mémoire génétique (comme les ADN) ; ensuite, lors de l'évolution de la vie, l'ADN aurait été spécialisé dans la mémoire génétique, et les protéines spécialisées dans la catalyse.
ADN : 2 brins antiparallèles structure en « double hélice » = échelle tordue
sur elle-mêmefacile à dessiner sur une feuille : on fait la
première chaîne 5'-->3', puis on retourne la feuille et on fait la deuxième.
petit sillon
grand sillon
les 3 types d'ARN fondamentaux : ARNm, ARNr, ARNtARNm = ARN messager : porte l'information génétique pour la synthèse des
protéines, ARNt = ARN de transfert : fait la correspondance entre le codon de l'ARNm et l'acide aminé grâce à son anticodon , ARNr = ARN ribosomal dans les sous-unités du ribosome permettant le bon positionnement des deux autres
organisation cellulaire et moléculaire du m
onde vivant : 22 (P. D
ieumegard)

Propriétés physiques des acides nucléiquesSolubilitéL'ADN est soluble dans l'eau
(surtout sous forme de sels de sodium), en formant une solution assez visqueuse.
L'ADN est insoluble dans l'alcool, et forme un précipité. Cette propriété peut être utilisée pour séparer l'ADN des autres constituants cellulaires.
Absorption de la lumière UV à 260 nmLes hétérocycles des nucléobases provoquent
une absorption des rayons ultra-violets à 260 nm. C'est une des raisons de la génotoxicité des rayons UV.
Effet hypochrome de l'ADN en double hélice
L'ADN en double brin absorbe moins les UV que la même quantité de nucléotides séparés en solution, à cause des liaisons hydrogène entre les nucléobases complémentaires, qui perturbent l'absorption.
Dénaturation thermiqueLe chauffage d'ADN double-brin provoque une
augmentation de la DO260 (densité optique à 260 nm) au delà qu'une certaine température, nommée « température de fusion ».
C'est dû à la séparation des deux brins. Parallèlement, il y a diminution de la viscosité de la solution.
Renaturation et hybridation des acides nucléiquesDes acides nucléiques chauffés deviennent
monocaténaires. Si on les refroidit rapidement, ils restent monocaténaires.Si on les refroidit lentement, les portions homologues
s'associent, et les doubles brins réapparaissent.A partir d'un mélange d'acides nucléiques chauffés, puis
refroidis lentement, on peut obtenir des acides nucléiques hybrides : ADN d'origine différente, ADN et ARN, etc.
DO260
température
« température de fusion » = température de demi-dissociation
augmentation de l'absorption des UV lors du chauffage
Cette « température de fusion » varie selon la composition de l'ADN.
Elle augmente avec la richesse en paires G=C, car celles-ci ont 3 liaisons hydrogène, alors que les paires A=T n'en ont que 2, et sont donc moins solides (moins résistantes à la rupture, donc la dénaturation a lieu à température plus basse).
organisation cellulaire et moléculaire du m
onde vivant : 23 (P. D
ieumegard)
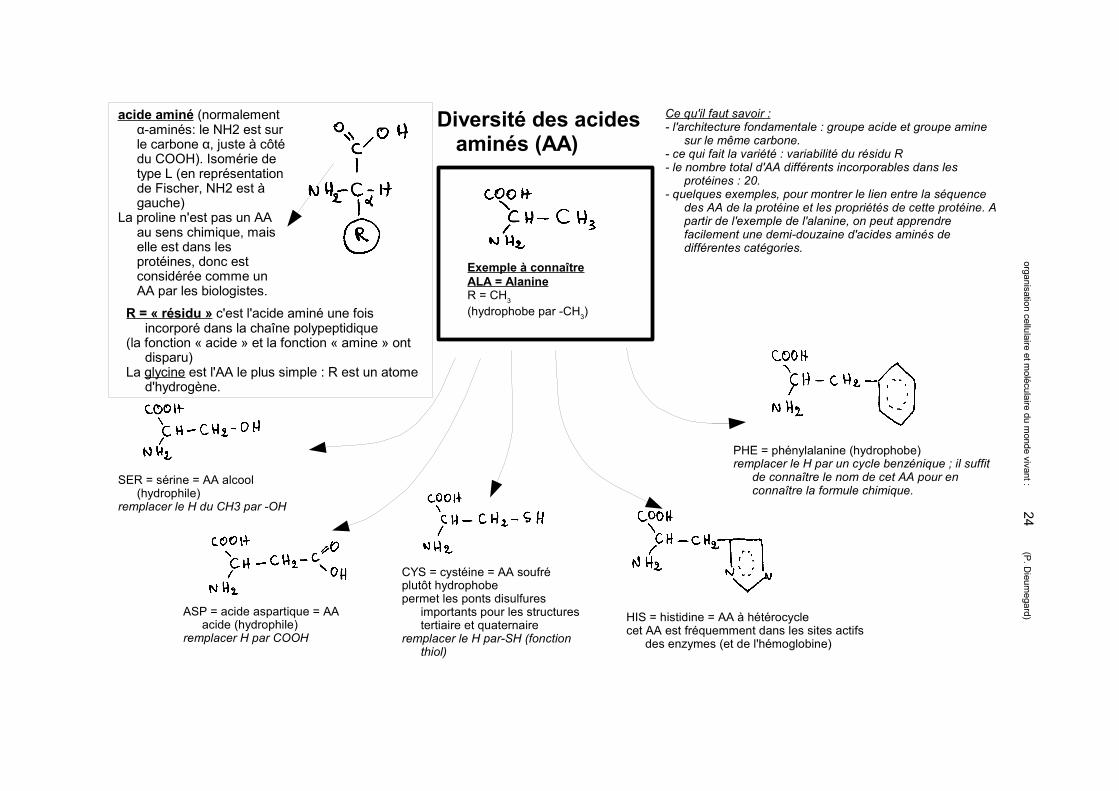
Diversité des acides aminés (AA)
acide aminé (normalement α-aminés: le NH2 est sur le carbone α, juste à côté du COOH). Isomérie de type L (en représentation de Fischer, NH2 est à gauche)
La proline n'est pas un AA au sens chimique, mais elle est dans les protéines, donc est considérée comme un AA par les biologistes.
Exemple à connaîtreALA = AlanineR = CH3
(hydrophobe par -CH3)
SER = sérine = AA alcool (hydrophile)
remplacer le H du CH3 par -OH
ASP = acide aspartique = AA acide (hydrophile)
remplacer H par COOH
CYS = cystéine = AA soufré plutôt hydrophobepermet les ponts disulfures
importants pour les structures tertiaire et quaternaire
remplacer le H par-SH (fonction thiol)
PHE = phénylalanine (hydrophobe)remplacer le H par un cycle benzénique ; il suffit
de connaître le nom de cet AA pour en connaître la formule chimique.
HIS = histidine = AA à hétérocyclecet AA est fréquemment dans les sites actifs
des enzymes (et de l'hémoglobine)
R = « résidu » c'est l'acide aminé une fois incorporé dans la chaîne polypeptidique
(la fonction « acide » et la fonction « amine » ont disparu)
La glycine est l'AA le plus simple : R est un atome d'hydrogène.
Ce qu'il faut savoir :- l'architecture fondamentale : groupe acide et groupe amine
sur le même carbone.- ce qui fait la variété : variabilité du résidu R- le nombre total d'AA différents incorporables dans les
protéines : 20. - quelques exemples, pour montrer le lien entre la séquence
des AA de la protéine et les propriétés de cette protéine. A partir de l'exemple de l'alanine, on peut apprendre facilement une demi-douzaine d'acides aminés de différentes catégories.
organisation cellulaire et moléculaire du m
onde vivant : 24 (P. D
ieumegard)
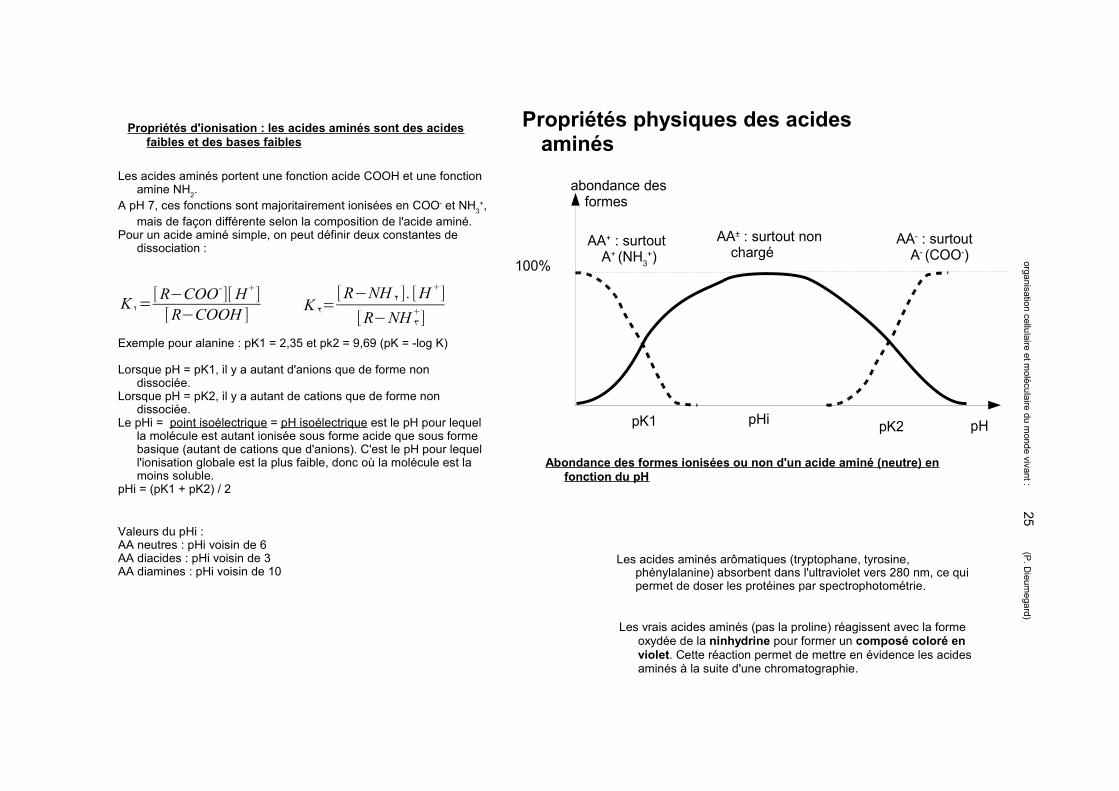
Propriétés physiques des acides aminés
Les acides aminés portent une fonction acide COOH et une fonction amine NH2.
A pH 7, ces fonctions sont majoritairement ionisées en COO- et NH3+,
mais de façon différente selon la composition de l'acide aminé.Pour un acide aminé simple, on peut définir deux constantes de
dissociation :
K١=[R−COO-][H +][R−COOH ] K ٢=
[R−NH ٢ ]. [H+]
[R−NH ٣+]
Exemple pour alanine : pK1 = 2,35 et pk2 = 9,69 (pK = -log K)
Lorsque pH = pK1, il y a autant d'anions que de forme non dissociée.
Lorsque pH = pK2, il y a autant de cations que de forme non dissociée.
Le pHi = point isoélectrique = pH isoélectrique est le pH pour lequel la molécule est autant ionisée sous forme acide que sous forme basique (autant de cations que d'anions). C'est le pH pour lequel l'ionisation globale est la plus faible, donc où la molécule est la moins soluble.
pHi = (pK1 + pK2) / 2
Les vrais acides aminés (pas la proline) réagissent avec la forme oxydée de la ninhydrine pour former un composé coloré en violet. Cette réaction permet de mettre en évidence les acides aminés à la suite d'une chromatographie.
Propriétés d'ionisation : les acides aminés sont des acides faibles et des bases faibles
Valeurs du pHi : AA neutres : pHi voisin de 6AA diacides : pHi voisin de 3AA diamines : pHi voisin de 10
Abondance des formes ionisées ou non d'un acide aminé (neutre) en fonction du pH
Les acides aminés arômatiques (tryptophane, tyrosine, phénylalanine) absorbent dans l'ultraviolet vers 280 nm, ce qui permet de doser les protéines par spectrophotométrie.
pHi
100%
AA± : surtout non chargé
AA+ : surtout A+ (NH3
+)
pK1 pK2
AA- : surtout A- (COO-)
pH
abondance des formes
organisation cellulaire et moléculaire du m
onde vivant : 25 (P. D
ieumegard)

Quatre niveaux structuraux des protéinesliaisons H entre N-H et
C=O ==> structure secondaire.
Exemple à connaître l'hélice α (forme de cylindre, la chaîne polypeptidique est enroulée en hélice droite, avec les N-H en face des C=O ; 3,6 résidus par tour)
Autre exemple , moins facile à dessiner : le feuillet β
liaisons entre protomères ==> structure quaternaire (protéines oligomériques)
Exemple à connaître : hémoglobine (tétramère de 2 chaines α et 2 chaînes β)
hème = groupement prosthétique
(non constitué d'acides aminés)
(site régulateur)site actif
liaison peptidique ==> structure primaire La structure primaire des protéines (la séquence de leurs résidus) est
déterminée par la séquence des nucléotides sur l'ARN messager (un codon est un triplet de nucléotides qui détermine un AA)
liaison entre résidus ===> structure tertiaire (assemblage dans l'espace ==> forme générale de la molécule)
Comme ces résidus sont déterminés par la séquence, on peut dire que la structure primaire détermine la structure tertiaire.
Mais souvent la correspondance n'est pas absolue: les chaperonines = protéines-chaperons veillent à la bonne conformation des protéines
la numérotation des résidus commence à l'extrémité N-terminale (-NH2 libre)
le dernier résidu est celui à l'extrémité C-terminale (COOH libre)
(forme globale de la protéine)
chaîne polypeptidique, parfois disposée en feuillets β (2 brins antiparallèles),ou en hélice α
organisation cellulaire et moléculaire du m
onde vivant : 26 (P. D
ieumegard)
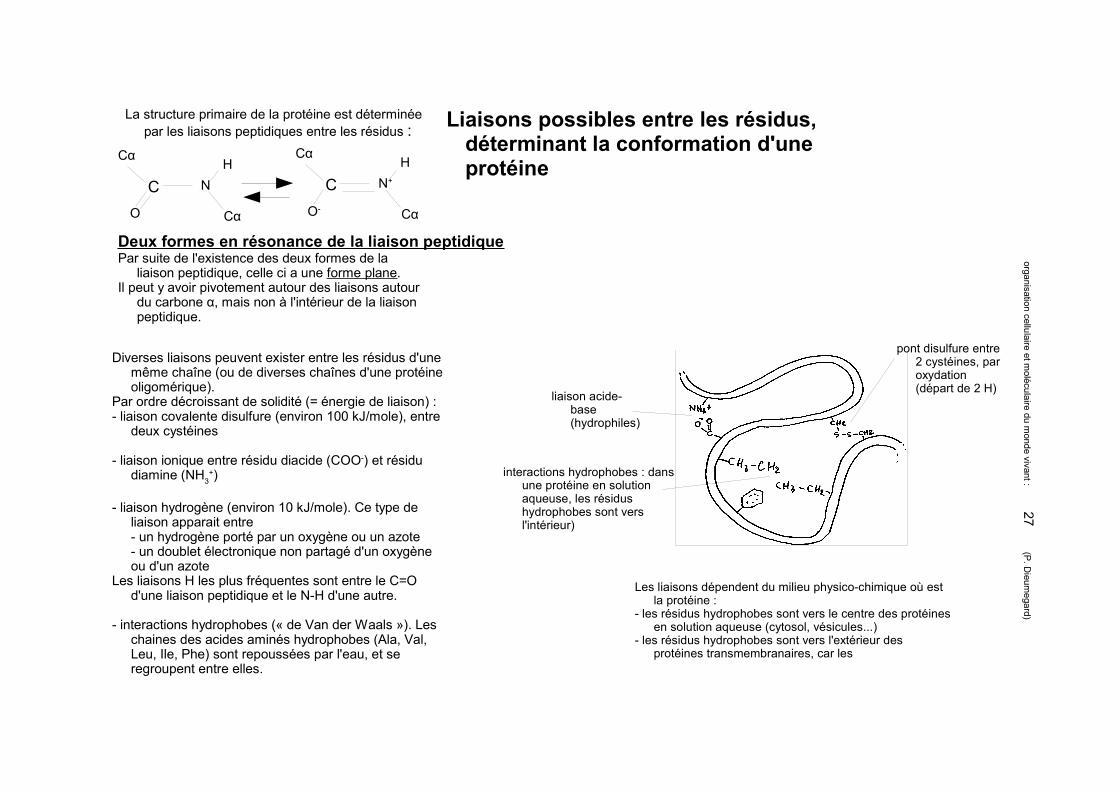
Liaisons possibles entre les résidus, déterminant la conformation d'une protéine
Deux formes en résonance de la liaison peptidique
C N
O
Cα
Cα
HC N+
O-
Cα
Cα
H
Par suite de l'existence des deux formes de la liaison peptidique, celle ci a une forme plane.
Il peut y avoir pivotement autour des liaisons autour du carbone α, mais non à l'intérieur de la liaison peptidique.
Diverses liaisons peuvent exister entre les résidus d'une même chaîne (ou de diverses chaînes d'une protéine oligomérique).
Par ordre décroissant de solidité (= énergie de liaison) :- liaison covalente disulfure (environ 100 kJ/mole), entre
deux cystéines
- liaison ionique entre résidu diacide (COO-) et résidu diamine (NH3
+)
- liaison hydrogène (environ 10 kJ/mole). Ce type de liaison apparait entre - un hydrogène porté par un oxygène ou un azote- un doublet électronique non partagé d'un oxygène ou d'un azote
Les liaisons H les plus fréquentes sont entre le C=O d'une liaison peptidique et le N-H d'une autre.
- interactions hydrophobes (« de Van der Waals »). Les chaines des acides aminés hydrophobes (Ala, Val, Leu, Ile, Phe) sont repoussées par l'eau, et se regroupent entre elles.
La structure primaire de la protéine est déterminée par les liaisons peptidiques entre les résidus :
liaison acide-base (hydrophiles)
interactions hydrophobes : dans une protéine en solution aqueuse, les résidus hydrophobes sont vers l'intérieur)
Les liaisons dépendent du milieu physico-chimique où est la protéine :
- les résidus hydrophobes sont vers le centre des protéines en solution aqueuse (cytosol, vésicules...)
- les résidus hydrophobes sont vers l'extérieur des protéines transmembranaires, car les
pont disulfure entre 2 cystéines, par oxydation (départ de 2 H)
organisation cellulaire et moléculaire du m
onde vivant : 27 (P. D
ieumegard)

Les deux structures secondaires fondamentales : hélice α et feuillet β
Structure de l'hexokinase (concours G2E 2006) :
3 hélices α, et 5 feuillets β
NH
CαCO C
O
CαNH
NH
NH
CαCO C
O
CαNH
NH
NH
CαCO C
O
CαNH
NH
Ces deux structures sont déterminées par les liaisons hydrogène entre C=O et N-H.
Hélice alpha : la chaîne polypeptidique est enroulée en une hélice droite, et stabilisée par les liaisons H entre deux tours successifs de l'hélice.
Cα-C-N-O
H
Cα -C-N-
O
HCα-C-N-
O
HC
α -C-N-
O
HO
Cα-N-C-
HO
Cα-N-C-
H
OCα -N-C-H
OCα -N-C-H
chaîne polypeptidique
ici, les deux chaînes polypeptidiques sont antiparallèles
Exemple : la fibroïne (soie)feuillet beta : deux chaînes polypeptidiques
sont reliées par des liaisons H.
Symbolisme :- les hélices α sont figurées par
des hélices ou des cylindres- les feuillets β sont figurés par
des flèches, dont la point est vers l'extrémité C terminale
Ces deux structures ne sont pas universelles :- il existe d'autres structures stables, moins fréquentes (exemple : triple hélice du collagène)- lorsqu'aucune structure n'est clairement reconnaissable, on parle de « pelote statistique »
Déterminisme de la structure :- certains résidus favorisent la structure en hélice, d'autres la structure en feuillets- le milieu extérieur joue aussi un rôle : certaines portions de chaîne polypeptidique peuvent être tantôt en hélice,
tantôt en feuillet. Les résidus ne sont pas figurés ; ils sont à
l'extérieur de l'hélice.Diamètre : 1 nm3,6 résidus par tour d'hélice.Exemple : la kératine (poils, ongles...)
Les résidus ne sont pas figurés : ils sont à l'extérieur de la pliure du feuillet (déterminée par les Cα)
organisation cellulaire et moléculaire du m
onde vivant : 28 (P. D
ieumegard)

Détermination de la masse molaire des protéines
La masse molaire des protéines peut varier entre 10 000 et 106. Il est important de pouvoir la déterminer, en particulier pour savoir si une
protéine est constituée d'une seule sous-unité, ou bien de plusieurs.Sachant que la masse molaire d'un acide aminé moyen est 100, on peut
calculer le nombre d'AA d'une protéine à partir de sa masse molaire : n = M/100
Filtration sur gel de dextrane (Séphadex)(chromatographie d'exclusion = tamisage moléculaire)La solution de protéines traverse une colonne pleine de
billes de gel de dextranes (glucides). Ces billes ont une certaine porosité.
Les plus grosses molécules sont trop grosses pour entrer dans les pores et passent donc rapidement.
Les petites molécules passent dans les pores, et y sont piégées momentanément.
On commence par étalonner le gel en faisant passer des protéines de masse molaire connue, puis on fait passer la protéine inconnue, et on compare les temps de rétention.
Électrophorèse en gel de polyacrylamide, après dénaturation par SDS
Le SDS est un agent dénaturant qui se fixe sur les protéines en leur conférant une charge négative. Ainsi, dans un champ électrique, les protéines migreront à une vitesse dépendant de leur taille, et non de leur richesse en résidus acides ou amines.
Là aussi, la détermination de la masse molaire n'est possible que par comparaison avec des protéines connues.
flux de liquide(mélange de
petites et grosses
molécules)
les grosses ne pénètrent pas et passent rapidement
les petites pénètrent et sont ralenties
bille poreuse de dextrane
dépôt d'un mélange de protéines chargées (-) par le SDS
(-)
(+)
les grosses protéines sont retenues par les fibres du gel et migrent lentement
les petites molécules migrent plus vite
gel de polyacrylamide :
le pôle (+) attire les molécules chargées (-)
organisation cellulaire et moléculaire du m
onde vivant : 29 (P. D
ieumegard)
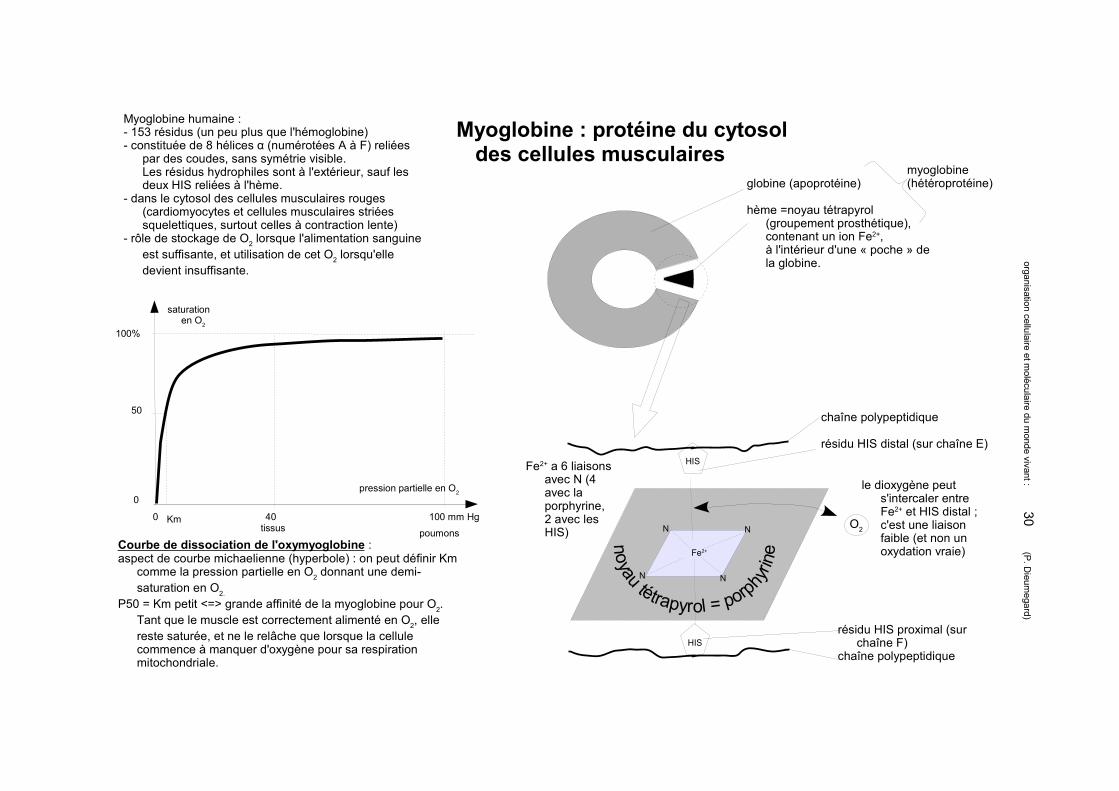
Myoglobine : protéine du cytosol des cellules musculaires
Fe2+
N N
N N
noyau tétrapyrol = porphyrin
e
HIS
HIS
globine (apoprotéine)
hème =noyau tétrapyrol (groupement prosthétique), contenant un ion Fe2+,à l'intérieur d'une « poche » de la globine.
myoglobine(hétéroprotéine)
Myoglobine humaine : - 153 résidus (un peu plus que l'hémoglobine)- constituée de 8 hélices α (numérotées A à F) reliées
par des coudes, sans symétrie visible.Les résidus hydrophiles sont à l'extérieur, sauf les deux HIS reliées à l'hème.
- dans le cytosol des cellules musculaires rouges (cardiomyocytes et cellules musculaires striées squelettiques, surtout celles à contraction lente)
- rôle de stockage de O2 lorsque l'alimentation sanguine est suffisante, et utilisation de cet O2 lorsqu'elle devient insuffisante.
chaîne polypeptidique
résidu HIS distal (sur chaîne E)
Fe2+ a 6 liaisons avec N (4 avec la porphyrine, 2 avec les HIS)
O2
le dioxygène peut s'intercaler entre Fe2+ et HIS distal ; c'est une liaison faible (et non un oxydation vraie)
résidu HIS proximal (sur chaîne F)
chaîne polypeptidique
Courbe de dissociation de l'oxymyoglobine : aspect de courbe michaelienne (hyperbole) : on peut définir Km
comme la pression partielle en O2 donnant une demi-saturation en O2.
P50 = Km petit <=> grande affinité de la myoglobine pour O2. Tant que le muscle est correctement alimenté en O2, elle reste saturée, et ne le relâche que lorsque la cellule commence à manquer d'oxygène pour sa respiration mitochondriale.
100%
saturation en O2
0
0 40 100 mm Hg
pression partielle en O2
tissus poumons
50
Km
organisation cellulaire et moléculaire du m
onde vivant : 30 (P. D
ieumegard)
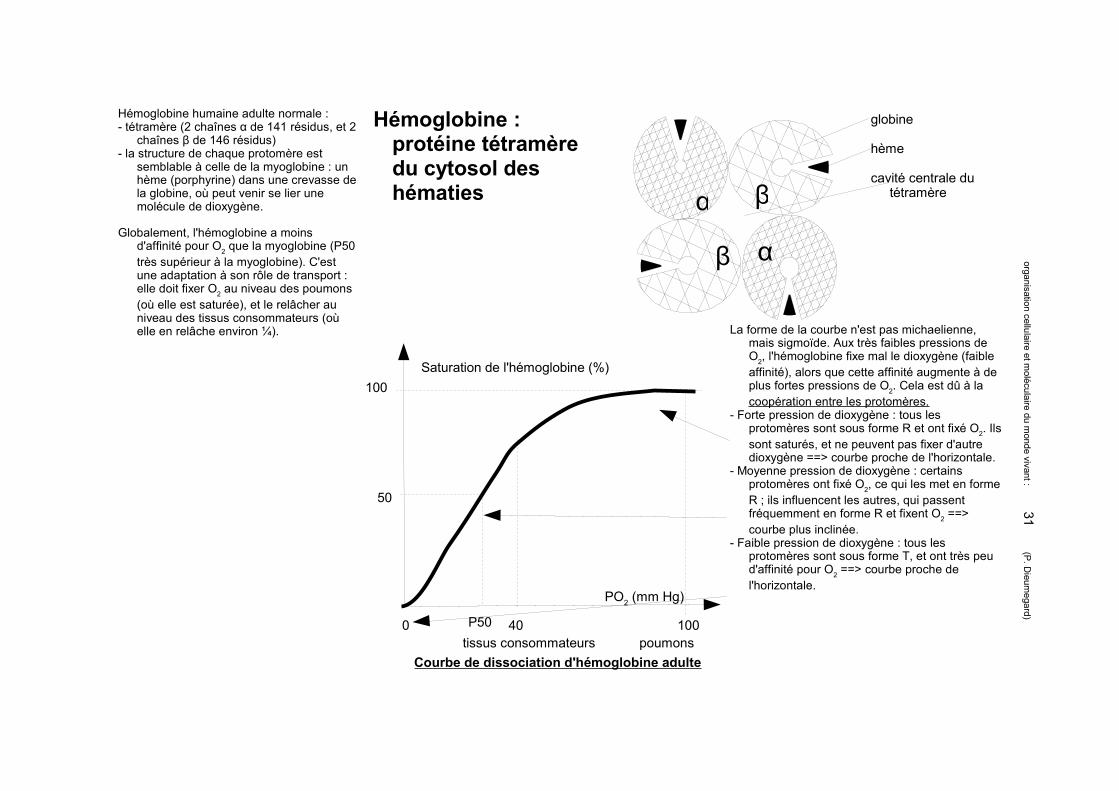
Hémoglobine : protéine tétramère du cytosol des hématies
Hémoglobine humaine adulte normale :- tétramère (2 chaînes α de 141 résidus, et 2
chaînes β de 146 résidus)- la structure de chaque protomère est
semblable à celle de la myoglobine : un hème (porphyrine) dans une crevasse de la globine, où peut venir se lier une molécule de dioxygène. α
α
β
β
globine
hème
cavité centrale du tétramère
Globalement, l'hémoglobine a moins d'affinité pour O2 que la myoglobine (P50 très supérieur à la myoglobine). C'est une adaptation à son rôle de transport : elle doit fixer O2 au niveau des poumons (où elle est saturée), et le relâcher au niveau des tissus consommateurs (où elle en relâche environ ¼).
Saturation de l'hémoglobine (%)100
PO2 (mm Hg)
0 40 100
Courbe de dissociation d'hémoglobine adultetissus consommateurs poumons
50
P50
La forme de la courbe n'est pas michaelienne, mais sigmoïde. Aux très faibles pressions de O2, l'hémoglobine fixe mal le dioxygène (faible affinité), alors que cette affinité augmente à de plus fortes pressions de O2. Cela est dû à la coopération entre les protomères.
- Forte pression de dioxygène : tous les protomères sont sous forme R et ont fixé O2. Ils sont saturés, et ne peuvent pas fixer d'autre dioxygène ==> courbe proche de l'horizontale.
- Moyenne pression de dioxygène : certains protomères ont fixé O2, ce qui les met en forme R ; ils influencent les autres, qui passent fréquemment en forme R et fixent O2 ==> courbe plus inclinée.
- Faible pression de dioxygène : tous les protomères sont sous forme T, et ont très peu d'affinité pour O2 ==> courbe proche de l'horizontale.
organisation cellulaire et moléculaire du m
onde vivant : 31 (P. D
ieumegard)

Changements de conformation de l'hémoglobine par effets allostériques coopératifs
L'hémoglobine non oxygénée est rouge sombre ; l'hémoglobine oxygénée est rouge clair (« rouge vif »).
Ces changements de couleur sont dûs à des changements de propriétés optiques de la molécule, eux-mêmes dûs à des changements dans la structure de la molécule, provoqués par la fixation de ligands
Forme oxygénée = forme R (Relâchée) = oxyhémoglobine
- cavité centrale plus petite- forte affinité pour O2
α
α
β
β
Forme non oxygénée = forme T (Tendue) = désoxyhémoglobine
- cavité centrale plus grande- faible affinité pour O2
α
α
β
β
dioxygène O2 fixé entre l'hème et HIS distal
site actif inaccessible pour O2 dans la forme T
Allostérie par interaction entre les diverses sous-unités (= divers protomères) :La fixation d'une molécule de O2 sur un hème déforme la globine correspondante. Comme cette globine
est elle-même reliée aux autres, la déformation est transmise aux autres protomères, qui accueillent plus facilement le dioxygène.
Inversement, le départ de O2 d'un hème provoque la déformation de la globine correspondante, donc des autres globines, et diminue l'affinité de ces autres protomères pour le dioxygène.
organisation cellulaire et moléculaire du m
onde vivant : 32 (P. D
ieumegard)

Effecteur allostérique : ligand pouvant agir sur une protéine, en modifiant (légèrement) sa conformation, donc en modifiant son activité.
Changements de conformation de l'hémoglobine par effets de ligands extérieurs
Effet du CO 2 : diminution de l'affinité pour O 2Le CO2 provoque le relâchement du dioxygène par l'hémoglobine.Le dioxyde de carbone peut se fixer réversiblement sur les résidus aminés
(plutôt chargés (+) en formant des résidus carbaminés (plutôt chargés (-)).
Ce changement des propriétés physiques des résidus concernés provoque un changement de conformation de l'hémoglobine, qui provoque le relâchement de O2.
C'est une bonne adaptation au transport des gaz respiratoires :- au niveau des poumons, CO2 part dans l'air alvéolaire, sa pression partielle
diminue dans le sang, donc les résidus carbaminés perdent leur CO2, et l'affinité de l'hémoglobine pour O2 augmente, ce qui permet de capter le maximm d'O2 possible.
- au niveau des tissus consommateurs (muscles, viscères, etc), la respiration mitochondriale dégage beaucoup de CO2, qui pénètre dans les hématies, et se fixe sur les résidus amines de l'hémoglobine. Donc l'affinité pour O2 diminue, donc davantage de dioxygène est relâché, qui pourra diffuser jusqu'aux cellules consommatrices.
Saturation de l'hémoglobine (%)100
PO2 (mm Hg)0 40 100
Courbe de dissociation selon la pression partielle du sang en CO 2
tissus consommateurs poumons
50
ppCO2 forte
ppCO2 faible
Effet du BPG = DPG (2-3 diphosphoglycérate) : diminution de l'affinité pour O 2Le DPG est produit par la glycolyse.En présence de DPG, la courbe de saturation est déplacée vers la droite, ce qui
signifie que l'affinité de l'hémoglobine pour O2 est diminuée.La molécule de DPG se fixe dans la cavité centrale du tétramère d'hémoglobine, entre
les chaînes β. Ceci déforme la molécule, et diminue l'affinité (favorise la forme tendue T).
Effet de l'acidité : diminution de l'affinité pour O 2Les ions H+ modifient aussi la conformation de la globine, en diminuant l'affinité. Ceci
favorise aussi la libération du dioxygène au niveau des tissus consommateurs d'oxygène, qui libèrent des acides organiques ou de l'acide carbonique lorsqu'ils sont actifs. Ceci permet encore une bonne libération de dioxygène au niveau des tissus. En physiologie, cette propriété a reçu le nom d'effet Bohr.
Courbes de dissociation de l'hémoglobine en présence de divers effecteurs (sujet ENS Lyon 1992)
organisation cellulaire et moléculaire du m
onde vivant : 33 (P. D
ieumegard)
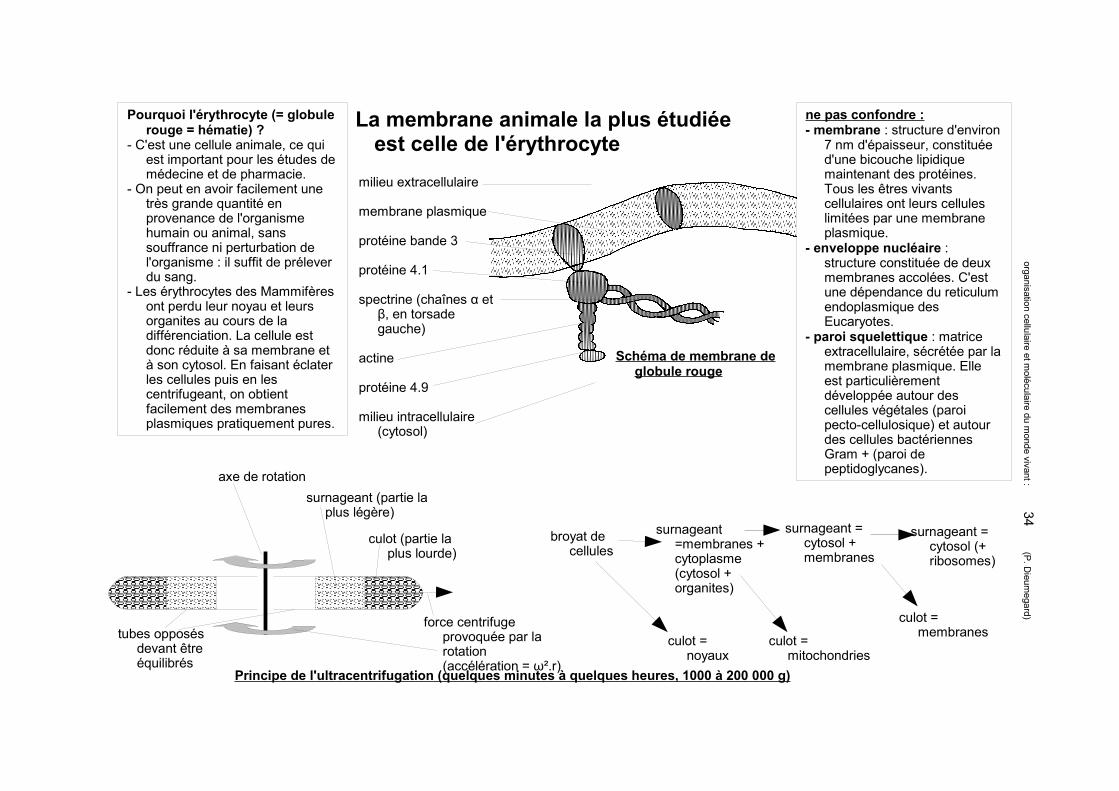
La membrane animale la plus étudiée est celle de l'érythrocyte
Pourquoi l'érythrocyte (= globule rouge = hématie) ?
- C'est une cellule animale, ce qui est important pour les études de médecine et de pharmacie.
- On peut en avoir facilement une très grande quantité en provenance de l'organisme humain ou animal, sans souffrance ni perturbation de l'organisme : il suffit de prélever du sang.
- Les érythrocytes des Mammifères ont perdu leur noyau et leurs organites au cours de la différenciation. La cellule est donc réduite à sa membrane et à son cytosol. En faisant éclater les cellules puis en les centrifugeant, on obtient facilement des membranes plasmiques pratiquement pures.
milieu extracellulaire
membrane plasmique
protéine bande 3
protéine 4.1
spectrine (chaînes α et β, en torsade gauche)
actine
protéine 4.9
milieu intracellulaire (cytosol)
Schéma de membrane de globule rouge
ne pas confondre :- membrane : structure d'environ
7 nm d'épaisseur, constituée d'une bicouche lipidique maintenant des protéines. Tous les êtres vivants cellulaires ont leurs cellules limitées par une membrane plasmique.
- enveloppe nucléaire : structure constituée de deux membranes accolées. C'est une dépendance du reticulum endoplasmique des Eucaryotes.
- paroi squelettique : matrice extracellulaire, sécrétée par la membrane plasmique. Elle est particulièrement développée autour des cellules végétales (paroi pecto-cellulosique) et autour des cellules bactériennes Gram + (paroi de peptidoglycanes).
Principe de l'ultracentrifugation (quelques minutes à quelques heures, 1000 à 200 000 g)
axe de rotationsurnageant (partie la
plus légère)
culot (partie la plus lourde)
force centrifuge provoquée par la rotation (accélération = ω².r)
tubes opposés devant être équilibrés
broyat de cellules
culot = noyaux
surnageant =membranes + cytoplasme (cytosol + organites)
surnageant = cytosol + membranes
culot = mitochondries
surnageant = cytosol (+ ribosomes)
culot = membranes
organisation cellulaire et moléculaire du m
onde vivant : 34 (P. D
ieumegard)

Modèle de Singer et Nicholson (1972) = « modèle en mosaïque fluide »
(face cytosolique)
(face extracellulaire)
La membrane de base = membrane unitaire
(épaisseur environ 7 nm = 70 Ǻ)
Description valable pour les membranes des Eucaryotes, sauf pour la membrane interne des mitochondries et des plastes (et les membranes des thylakoïdes)
Les glucides sont uniquement sur la face qui n'est pas en contact avec le cytosol : face extracellulaire pour la membrane plasmique, face luminale pour les endomembranes
En masse : 50% de protéines, 50% de lipides (et un peu de glucides...)
En nombre de molécules : grosse majorité de lipides, car les protéines sont des molécules beaucoup plus grosses que les lipides.
Attention à l'aspect trifeuilleté en microscopie électronique à transmission !!!
Après fixation et coloration à l'acide osmique (OsO4), la membrane apparaît avec deux feuillets sombres encadrant un feuillet clair, mais c'est un artefact, dû à la préparation.
Cet aspect hétérogène est en particulier observable en MET après cryofracture et cryodécapage. « mosaïque fluide » :
- mosaïque, car les molécules sontdisposées selon un plan, le plan de la membrane
- fluide, car les déplacements sont faciles dans le plan de la membrane, simplement par l'agitation thermique des molécules.La fluidité est augmentée par la température, et par l'abondance des doubles liaisons (acides gras insaturés). La fluidité est diminuée par le cholestérol.
Par contre, le passage des molécules d'une hémimembrane à une autre (« flip-flop ») est très difficile. Les lipides membranaires peuvent effectuer ce passage grâce à des enzymes particulières, les flippases.
protéines : - certaines sont intrinsèques (faisant
partie intégrante de la membrane), souvent avec des hélices α transmembranaires
- d'autres sont périphériques, liées assez faiblement à des constituants membranaires (lipides ou autres protéines)
glucides, liés aux protéines ou aux lipides.
Ils forment le glycocalyx = revêtement fibreux = cell-coat
stérol (stabilise la structure de la membrane)
phospholipide (donne la structure fondamentale de la membrane en bicouche lipidique)
organisation cellulaire et moléculaire du m
onde vivant : 35 (P. D
ieumegard)
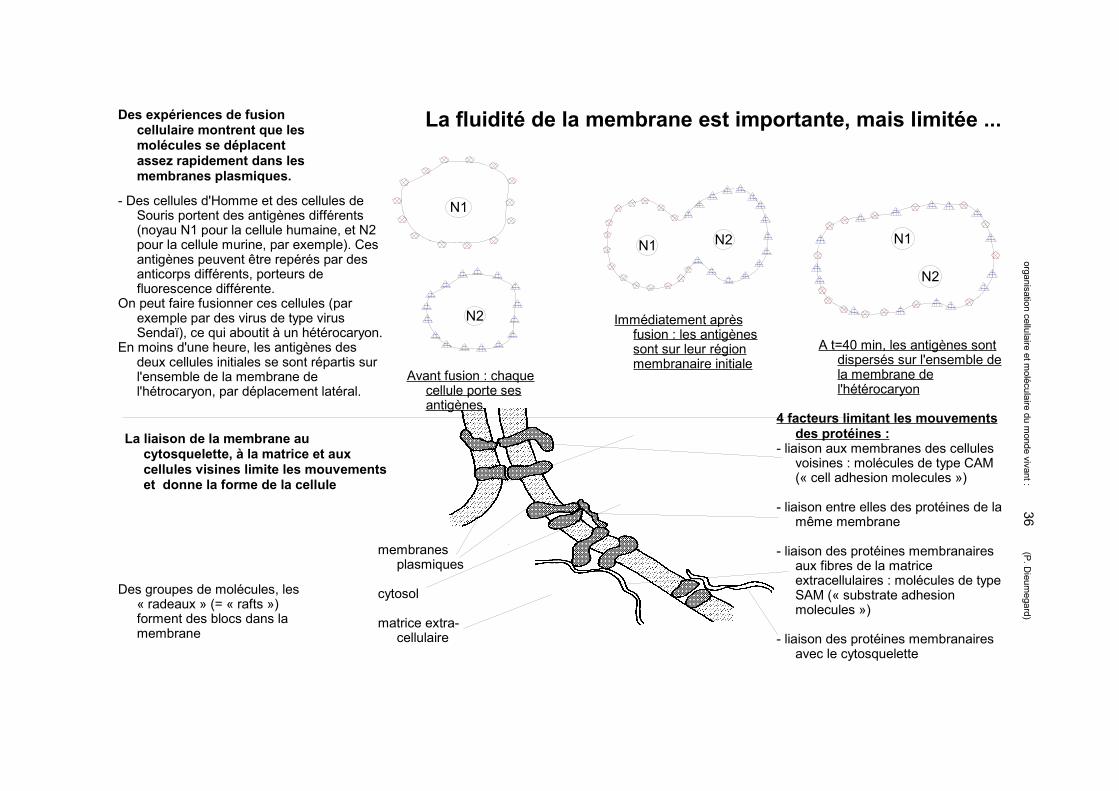
La fluidité de la membrane est importante, mais limitée ...
Des groupes de molécules, les « radeaux » (= « rafts ») forment des blocs dans la membrane
La liaison de la membrane au cytosquelette, à la matrice et aux cellules visines limite les mouvements et donne la forme de la cellule
4 facteurs limitant les mouvements des protéines :
- liaison aux membranes des cellules voisines : molécules de type CAM (« cell adhesion molecules »)
- liaison entre elles des protéines de la même membrane
- liaison des protéines membranaires aux fibres de la matrice extracellulaires : molécules de type SAM (« substrate adhesion molecules »)
- liaison des protéines membranaires avec le cytosquelette
membranes plasmiques
cytosol
matrice extra-cellulaire
N1
N2
N1 N2 N1
N2
Des expériences de fusion cellulaire montrent que les molécules se déplacent assez rapidement dans les membranes plasmiques.
- Des cellules d'Homme et des cellules de Souris portent des antigènes différents (noyau N1 pour la cellule humaine, et N2 pour la cellule murine, par exemple). Ces antigènes peuvent être repérés par des anticorps différents, porteurs de fluorescence différente.
On peut faire fusionner ces cellules (par exemple par des virus de type virus Sendaï), ce qui aboutit à un hétérocaryon.
En moins d'une heure, les antigènes des deux cellules initiales se sont répartis sur l'ensemble de la membrane de l'hétrocaryon, par déplacement latéral.
Avant fusion : chaque cellule porte ses antigènes
Immédiatement après fusion : les antigènes sont sur leur région membranaire initiale
A t=40 min, les antigènes sont dispersés sur l'ensemble de la membrane de l'hétérocaryon
organisation cellulaire et moléculaire du m
onde vivant : 36 (P. D
ieumegard)
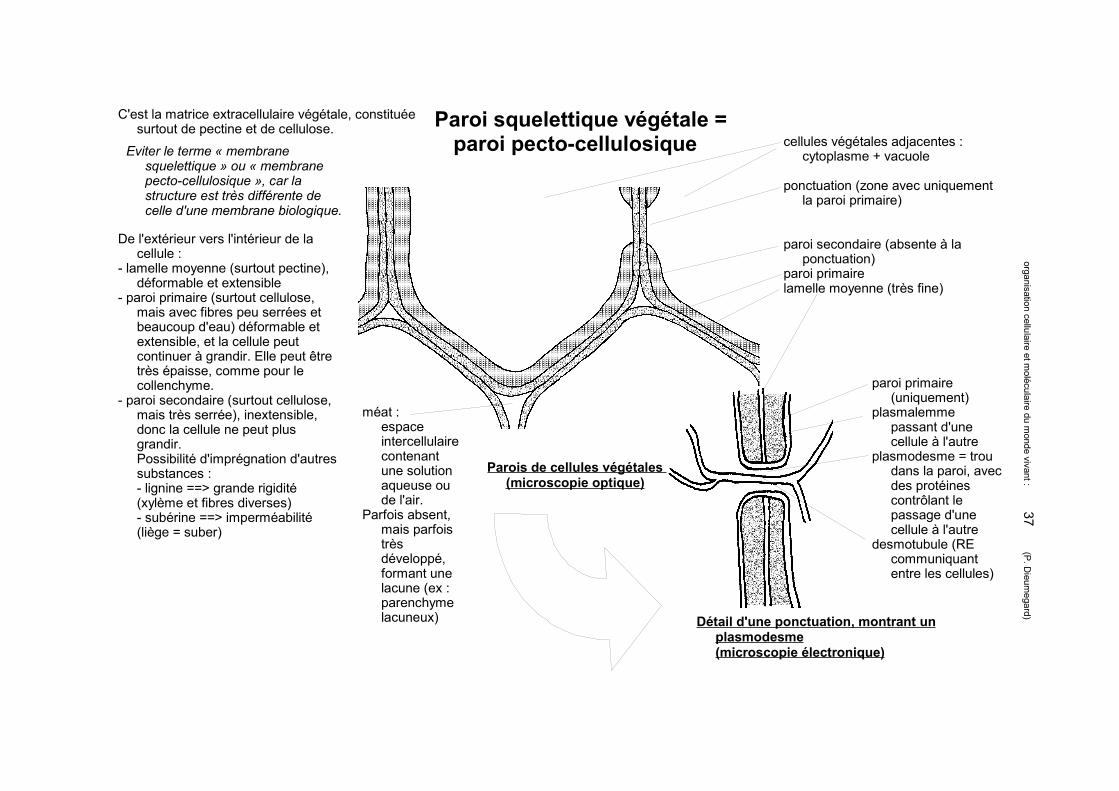
Paroi squelettique végétale = paroi pecto-cellulosique
paroi primaire (uniquement)
plasmalemme passant d'une cellule à l'autre
plasmodesme = trou dans la paroi, avec des protéines contrôlant le passage d'une cellule à l'autre
desmotubule (RE communiquant entre les cellules)
Eviter le terme « membrane squelettique » ou « membrane pecto-cellulosique », car la structure est très différente de celle d'une membrane biologique.
De l'extérieur vers l'intérieur de la cellule :
- lamelle moyenne (surtout pectine), déformable et extensible
- paroi primaire (surtout cellulose, mais avec fibres peu serrées et beaucoup d'eau) déformable et extensible, et la cellule peut continuer à grandir. Elle peut être très épaisse, comme pour le collenchyme.
- paroi secondaire (surtout cellulose, mais très serrée), inextensible, donc la cellule ne peut plus grandir.Possibilité d'imprégnation d'autres substances :- lignine ==> grande rigidité (xylème et fibres diverses)- subérine ==> imperméabilité (liège = suber)
C'est la matrice extracellulaire végétale, constituée surtout de pectine et de cellulose.
cellules végétales adjacentes : cytoplasme + vacuole
ponctuation (zone avec uniquement la paroi primaire)
paroi secondaire (absente à la ponctuation)
paroi primairelamelle moyenne (très fine)
méat : espace intercellulaire contenant une solution aqueuse ou de l'air.
Parfois absent, mais parfois très développé, formant une lacune (ex : parenchyme lacuneux)
Parois de cellules végétales (microscopie optique)
Détail d'une ponctuation, montrant un plasmodesme(microscopie électronique)
organisation cellulaire et moléculaire du m
onde vivant : 37 (P. D
ieumegard)
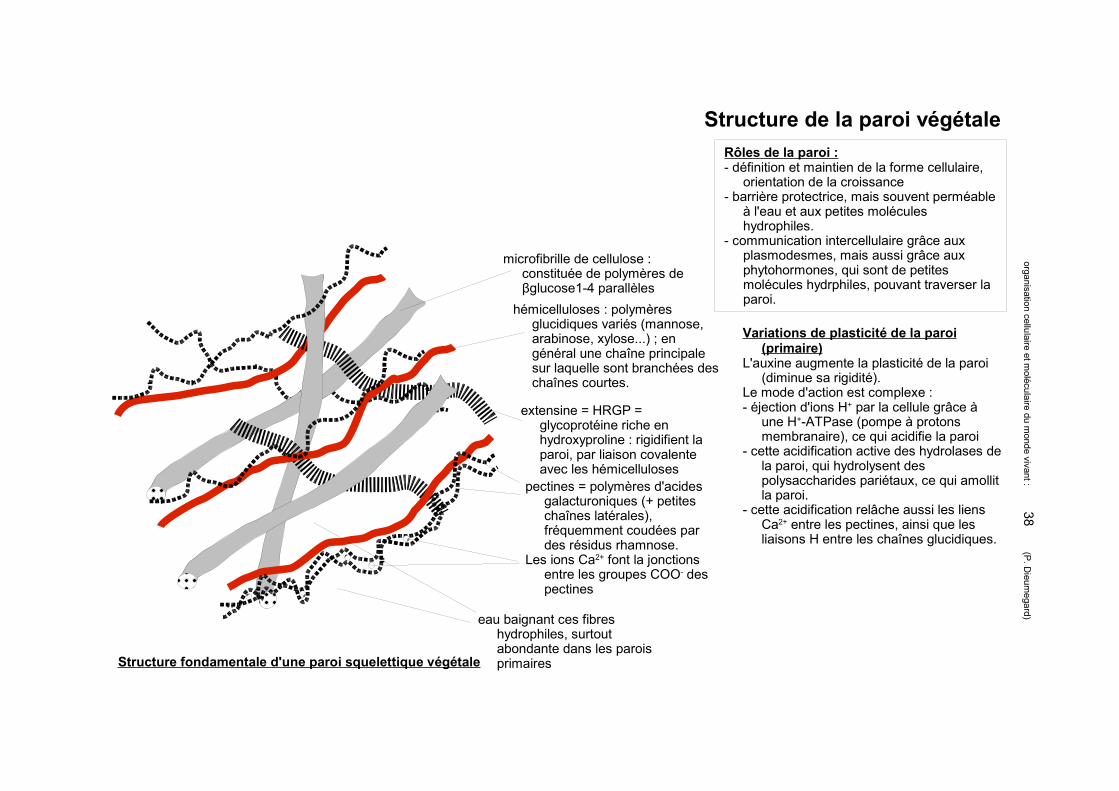
Structure de la paroi végétale
microfibrille de cellulose : constituée de polymères de βglucose1-4 parallèles
hémicelluloses : polymères glucidiques variés (mannose, arabinose, xylose...) ; en général une chaîne principale sur laquelle sont branchées des chaînes courtes.
extensine = HRGP = glycoprotéine riche en hydroxyproline : rigidifient la paroi, par liaison covalente avec les hémicelluloses
pectines = polymères d'acides galacturoniques (+ petites chaînes latérales), fréquemment coudées par des résidus rhamnose.
Les ions Ca2+ font la jonctions entre les groupes COO- des pectines
eau baignant ces fibres hydrophiles, surtout abondante dans les parois primairesStructure fondamentale d'une paroi squelettique végétale
Rôles de la paroi :- définition et maintien de la forme cellulaire,
orientation de la croissance- barrière protectrice, mais souvent perméable
à l'eau et aux petites molécules hydrophiles.
- communication intercellulaire grâce aux plasmodesmes, mais aussi grâce aux phytohormones, qui sont de petites molécules hydrphiles, pouvant traverser la paroi.
Variations de plasticité de la paroi (primaire)
L'auxine augmente la plasticité de la paroi (diminue sa rigidité).
Le mode d'action est complexe :- éjection d'ions H+ par la cellule grâce à
une H+-ATPase (pompe à protons membranaire), ce qui acidifie la paroi
- cette acidification active des hydrolases de la paroi, qui hydrolysent des polysaccharides pariétaux, ce qui amollit la paroi.
- cette acidification relâche aussi les liens Ca2+ entre les pectines, ainsi que les liaisons H entre les chaînes glucidiques.
organisation cellulaire et moléculaire du m
onde vivant : 38 (P. D
ieumegard)
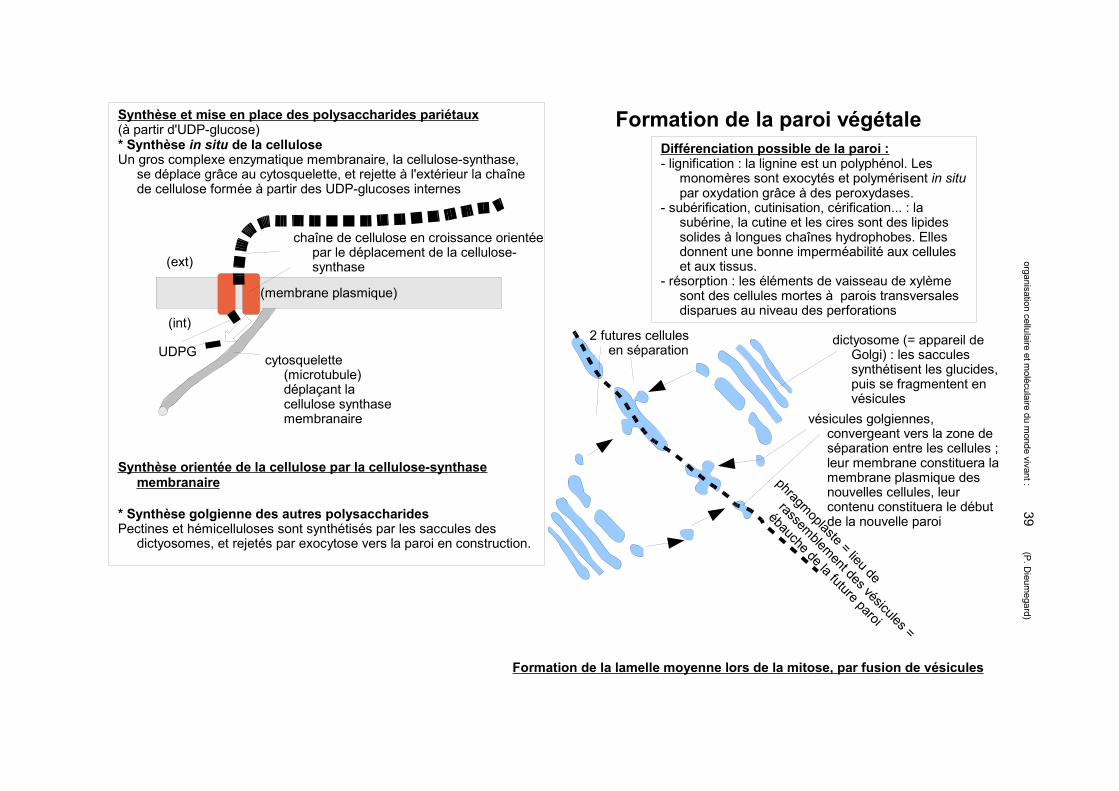
Différenciation possible de la paroi :- lignification : la lignine est un polyphénol. Les
monomères sont exocytés et polymérisent in situ par oxydation grâce à des peroxydases.
- subérification, cutinisation, cérification... : la subérine, la cutine et les cires sont des lipides solides à longues chaînes hydrophobes. Elles donnent une bonne imperméabilité aux cellules et aux tissus.
- résorption : les éléments de vaisseau de xylème sont des cellules mortes à parois transversales disparues au niveau des perforations
Synthèse et mise en place des polysaccharides pariétaux(à partir d'UDP-glucose)* Synthèse in situ de la celluloseUn gros complexe enzymatique membranaire, la cellulose-synthase,
se déplace grâce au cytosquelette, et rejette à l'extérieur la chaîne de cellulose formée à partir des UDP-glucoses internes
* Synthèse golgienne des autres polysaccharides Pectines et hémicelluloses sont synthétisés par les saccules des
dictyosomes, et rejetés par exocytose vers la paroi en construction.
(membrane plasmique)
(ext)
(int)
UDPG
chaîne de cellulose en croissance orientée par le déplacement de la cellulose-synthase
cytosquelette (microtubule) déplaçant la cellulose synthase membranaire
Synthèse orientée de la cellulose par la cellulose-synthase membranaire
Formation de la paroi végétale
Formation de la lamelle moyenne lors de la mitose, par fusion de vésicules
2 futures cellules en séparation
dictyosome (= appareil de Golgi) : les saccules synthétisent les glucides, puis se fragmentent en vésicules
vésicules golgiennes, convergeant vers la zone de séparation entre les cellules ; leur membrane constituera la membrane plasmique des nouvelles cellules, leur contenu constituera le début de la nouvelle paroi
phragmoplaste = lieu de
rassemblement des vésicules =
ébauche de la future paroi
organisation cellulaire et moléculaire du m
onde vivant : 39 (P. D
ieumegard)

Paroi bactérienneAutrefois (XIXe siècle et début du XXe siècle), les bactéries étaient rattachées aux « Végétaux », et étudiées dans les cours de « botanique ». Ce classement est caduc depuis le développement des classifications phylogénétiques, mais il correspond quand même à une réalité physique : comme les cellules végétales, les cellules bactériennes sont entourées par une paroi plus ou moins rigide, leur conférant une résistance à la turgescence cellulaire.
Par contre, la nature chimique de la paroi bactérienne est complètement différente de la paroi végétale.
Schéma de colibacille (Bactérie Gram -) : 1 µm
chromosome = ADN circulaire
ARNm et ribosomes plasmide
membrane externe (avec lipopolysaccharides, n'existe que chez les bactéries Gram-)
membrane plasmique
« capsule » = « glycocalyx » : gangue d'épaisseur très variable, constituée de polysaccharides et glycoprotéines
paroi bactérienne au sens strict : mince chez les bactéries gram-, épaisse chez les bactéries gram+.
La paroi bactérienne est surtout constituée de peptidoglycaneLes peptidoglycanes sont des molécules complexes : polysachharides (avec
de l'acide acétyl-muramique) pontés par de petits polypeptides, ce qui réalise un ensemble rigide.
organisation cellulaire et moléculaire du m
onde vivant : 40 (P. D
ieumegard)

Matrice extracellulaire animale : les protéoglycanes forment la substance fondamentale
Différences entre glycoprotéines et protéoglycanes :
- les glycoprotéines ont la partie glucidique représentant moins de 50% de la masse ; les chaînes glucidiques sont courtes (une dizaine de résidus) et souvent ramifiées.
- les protéoglycanes ont des chaînes glucidiques longues (80 résidus en moyenne), linéaires, et la partie glucidique peut représenter 95% de la masse de l'ensemble. protéine centrale
= « core-protein »
GAG = glycosaminoglycanes
protéoglycane
structure d'un protéoglycane
Les glycosaminoglycanes (GAG) peuvent être considérés comme des polymères de disaccharides :
- l'un des oses est un acide uronique (donc ionisé en COO- aux pH biologiques)
- l'autre est acétyl-aminé (N-acétyl-glucosamine, par exemple), souvent sulfaté, donc aussi ionisé négativement.
Les sucres sont de type β, et soudés en 1-4 ou en 1-3. L'isomérie β leur donne une forme droite (comme la cellulose), et plutôt rigide
Ces molécules très chargées négativement attirent fortement l'eau, et constituent des gels très hydratés.
Les protéoglycanes, bien que constitués majoritairement de glucides, peuvent être considérés comme des protéines qui ont subi une glycosylation intense au niveau de l'appareil de Golgi (O-glycosylation). organisation cellulaire et m
oléculaire du monde vivant : 41 (P
. Dieum
egard)
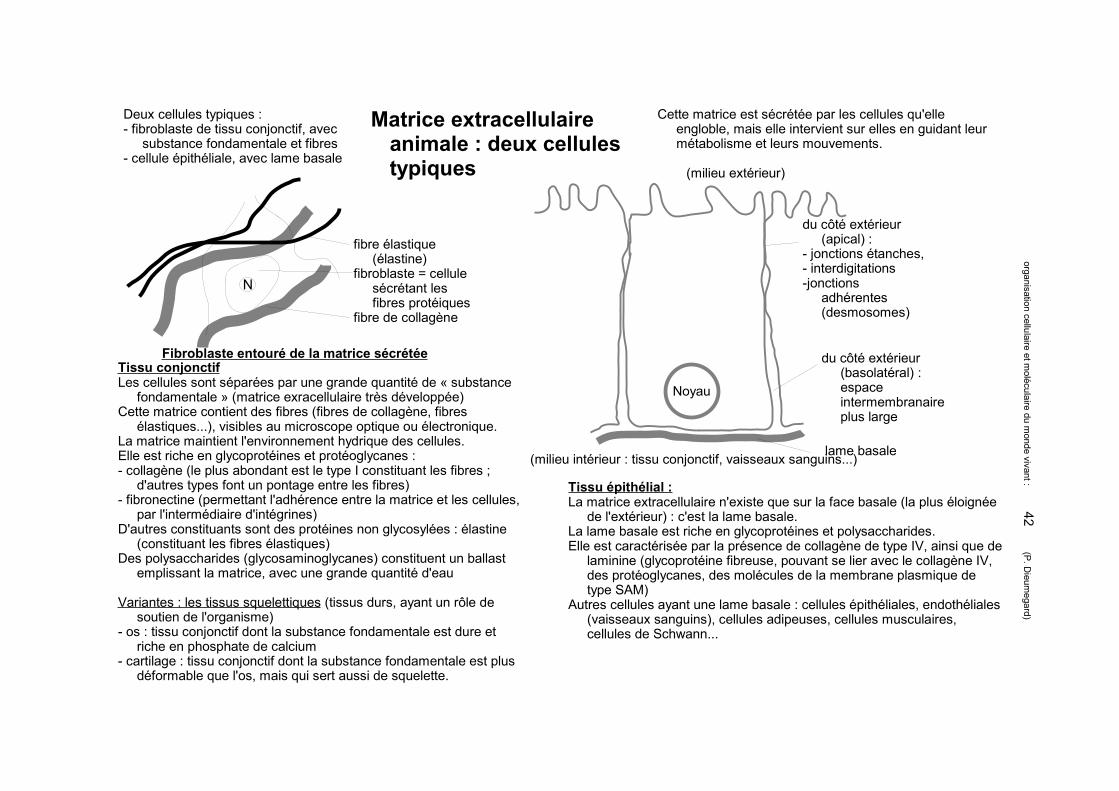
Matrice extracellulaire animale : deux cellules typiques
Deux cellules typiques :- fibroblaste de tissu conjonctif, avec
substance fondamentale et fibres- cellule épithéliale, avec lame basale
Tissu conjonctifLes cellules sont séparées par une grande quantité de « substance
fondamentale » (matrice exracellulaire très développée)Cette matrice contient des fibres (fibres de collagène, fibres
élastiques...), visibles au microscope optique ou électronique.La matrice maintient l'environnement hydrique des cellules.Elle est riche en glycoprotéines et protéoglycanes :- collagène (le plus abondant est le type I constituant les fibres ;
d'autres types font un pontage entre les fibres)- fibronectine (permettant l'adhérence entre la matrice et les cellules,
par l'intermédiaire d'intégrines)D'autres constituants sont des protéines non glycosylées : élastine
(constituant les fibres élastiques)Des polysaccharides (glycosaminoglycanes) constituent un ballast
emplissant la matrice, avec une grande quantité d'eau
Variantes : les tissus squelettiques (tissus durs, ayant un rôle de soutien de l'organisme)
- os : tissu conjonctif dont la substance fondamentale est dure et riche en phosphate de calcium
- cartilage : tissu conjonctif dont la substance fondamentale est plus déformable que l'os, mais qui sert aussi de squelette.
Tissu épithélial :La matrice extracellulaire n'existe que sur la face basale (la plus éloignée
de l'extérieur) : c'est la lame basale.La lame basale est riche en glycoprotéines et polysaccharides. Elle est caractérisée par la présence de collagène de type IV, ainsi que de
laminine (glycoprotéine fibreuse, pouvant se lier avec le collagène IV, des protéoglycanes, des molécules de la membrane plasmique de type SAM)
Autres cellules ayant une lame basale : cellules épithéliales, endothéliales (vaisseaux sanguins), cellules adipeuses, cellules musculaires, cellules de Schwann...
Cette matrice est sécrétée par les cellules qu'elle engloble, mais elle intervient sur elles en guidant leur métabolisme et leurs mouvements.
N
fibre élastique (élastine)
fibroblaste = cellule sécrétant les fibres protéiques
fibre de collagène
Fibroblaste entouré de la matrice sécrétée
(milieu extérieur)
(milieu intérieur : tissu conjonctif, vaisseaux sanguins...)
Noyau
du côté extérieur (apical) :
- jonctions étanches,- interdigitations-jonctions
adhérentes (desmosomes)
du côté extérieur (basolatéral) : espace intermembranaire plus large
lame basale
organisation cellulaire et moléculaire du m
onde vivant : 42 (P. D
ieumegard)
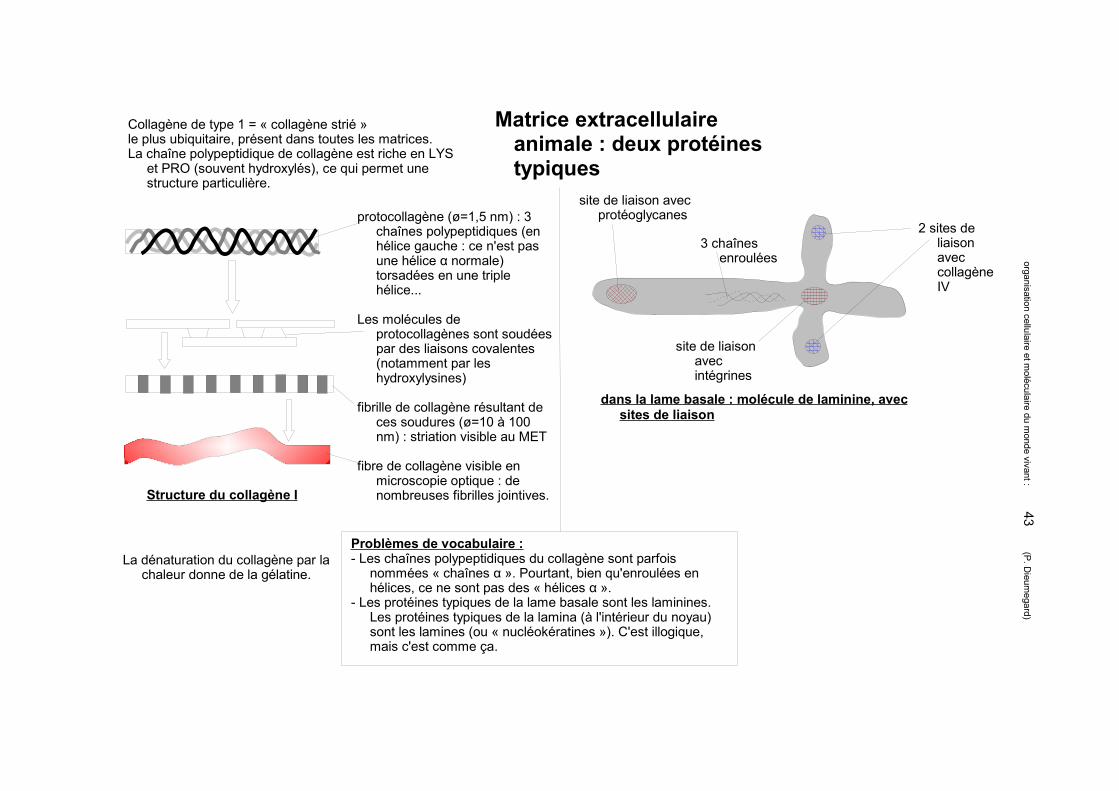
Matrice extracellulaire animale : deux protéines typiques
Collagène de type 1 = « collagène strié »le plus ubiquitaire, présent dans toutes les matrices.La chaîne polypeptidique de collagène est riche en LYS
et PRO (souvent hydroxylés), ce qui permet une structure particulière.
protocollagène (ø=1,5 nm) : 3 chaînes polypeptidiques (en hélice gauche : ce n'est pas une hélice α normale) torsadées en une triple hélice...
Les molécules de protocollagènes sont soudées par des liaisons covalentes (notamment par les hydroxylysines)
fibrille de collagène résultant de ces soudures (ø=10 à 100 nm) : striation visible au MET
fibre de collagène visible en microscopie optique : de nombreuses fibrilles jointives.Structure du collagène I
La dénaturation du collagène par la chaleur donne de la gélatine.
Problèmes de vocabulaire :- Les chaînes polypeptidiques du collagène sont parfois
nommées « chaînes α ». Pourtant, bien qu'enroulées en hélices, ce ne sont pas des « hélices α ».
- Les protéines typiques de la lame basale sont les laminines. Les protéines typiques de la lamina (à l'intérieur du noyau) sont les lamines (ou « nucléokératines »). C'est illogique, mais c'est comme ça.
site de liaison avec protéoglycanes
3 chaînes enroulées
site de liaison avec intégrines
2 sites de liaison avec collagène IV
dans la lame basale : molécule de laminine, avec sites de liaison
organisation cellulaire et moléculaire du m
onde vivant : 43 (P. D
ieumegard)

Endomembranes et formation de la matrice extracellulaire
Règle générale :Lors de l'exocytose, la membrane de la vésicule se transforme en
une portion du plasmalemme (= de la membrane plasmique), et son contenu, déversé à l'extérieur de la cellule, peut constituer de la matrice extracellulaire. ex
cytosol
Flux
mem
bran
aire
di
rect
Flux membranaire direct : du reticulum vers l'extérieur
ribosomes : synthèse des protéines.
Les protéines à destination du RE commencent par une « séquence-signal » qui provoque l'accolement des ribosomes à la membrane du RE (REG = reticulum endoplasmique rugueux). REL = reticulum endoplasmique
lisse : synthèse des lipides membranaires, et intégration à la membrane par les flippases
Dictyosome (appareil de Golgi) :
- O-glycosylation des protéines
-
RE : N-glycosylation des protéines
La « séquence-signal » est une séquence de 10 à 25 résidus hydrophobes, à l'extrémité N-terminale du polypeptide. Elle est reconnue par un complexe ribonucléoprotéique, la SRP (« Signal Recognition Particle »).
La membrane du RE contient un récepteur à la SRP. Ce récepteur capte le complexe ribosome-ARNm-SRP, et grâce à l'énergie d'hydrolyse de GTP, provoque le passage de la nouvelle protéine à travers le translocon (complexe protéique de la membrane du RE).
La séquence-signal est excisée par une peptidase.Dans la lumière du REG, la protéine est mise en forme par des protéines-chaperons (BIP = « BInding-protein), qui aident aussi à la pénétration du
polypeptide dans la lumière du reticulum.Quelle est la différence entre les protéines qui passent dans la lumière du RE et celles qui restent dans la membrane du RE ? Ces dernières ont une
séquence hydrophobe qui permet de rester ancré dans la membrane, et qui bloque donc la translocation complète vers la lumière du RE.À retenir : la séquence polypeptidique comporte des « étiquettes » qui provoquent l'adressage vers un compartiment cellulaire ou un autre.
N-glycosylation des protéines (dans le RE) : des oligosaccharides initialement construits sur des lipides de la face luminale sont transférés sur le groupement amine d'un résidu asparagine, d'où le nom N-glycosylation, puisque c'est l'azote N qui fait la liaison. Ultérieurement,
O-glycosylation des protéines : fixation de glucides sur un résidu alcool (sérine)
organisation cellulaire et moléculaire du m
onde vivant : 44 (P. D
ieumegard)

Jonctions intermembranaires mécaniques des cellules animales en microscopie électronique
Les animaux (Métazoaires) sont constitués d'un grand nombre de cellules, qui sont bien sûr liées entre elles. Les cellules du mésoderme sont souvent liées indirectement par une matrice extracellulaire importante (tissu conjonctif, tissu osseux, tissu cartilagineux...). Par contre, les cellules épithéliales sont directement liées entre elles par des jonctions intermembranaires mécaniquement importantes.
D'autres cellules sont reliées par des jonctions laculaires (= jonctions gap), qui sont importantes pour la communication d'informations entre les cellules.
Entérocyte, cellule typique des jonctions intercellulaires
(lumière intestinale)(microvillosités sur la face luminale)jonction serrée (« tight junction » =
« zonula occludens »). Les deux membranes sont pratiquement accolées, il n'y a pas de circulation de fluides entre les cellules : pas de circulation paracellulaire.
zonula adherens : bande avec des molécules fibreuses d'accrochage, caractéristique des cellules épithéliales ; aspect semblable aux desmosomes.
desmosome = (« macula adherens »)interdigitations = engrènement :
accrochage mécanique entre les cellules
(la région basolatérale a peu de jonctions)
hémidesmosome sur la face basale :accrochage de la cellule à la matrice extracellulaire (lame basale).
jonction serrée : membranes jointives
protéines de jonction (claudines et connexines : protéines à 4 domaines transmembranaires, comme les connexines des jonctions gap)
tonofilaments = filaments intermédiaires de kératine (ou autres protéines, selon le type cellulaire)
plaque intracellulaire (desmoplakine)
plaque intercellulaire, formée par l'association des cadhérines transmembranaires (CAM : desmocollines et desmogléines)
membranes plasmiques
desmosome : ancrage de deux cellules
hémidesmosome : ancrage d'une cellule à la matrice
microfilaments d'actine (c'est une différence avec les desmosomes)
membrane plasmique
intégrines (autre différence avec les desmosomes
fibres de la lame basale
organisation cellulaire et moléculaire du m
onde vivant : 45 (P. D
ieumegard)
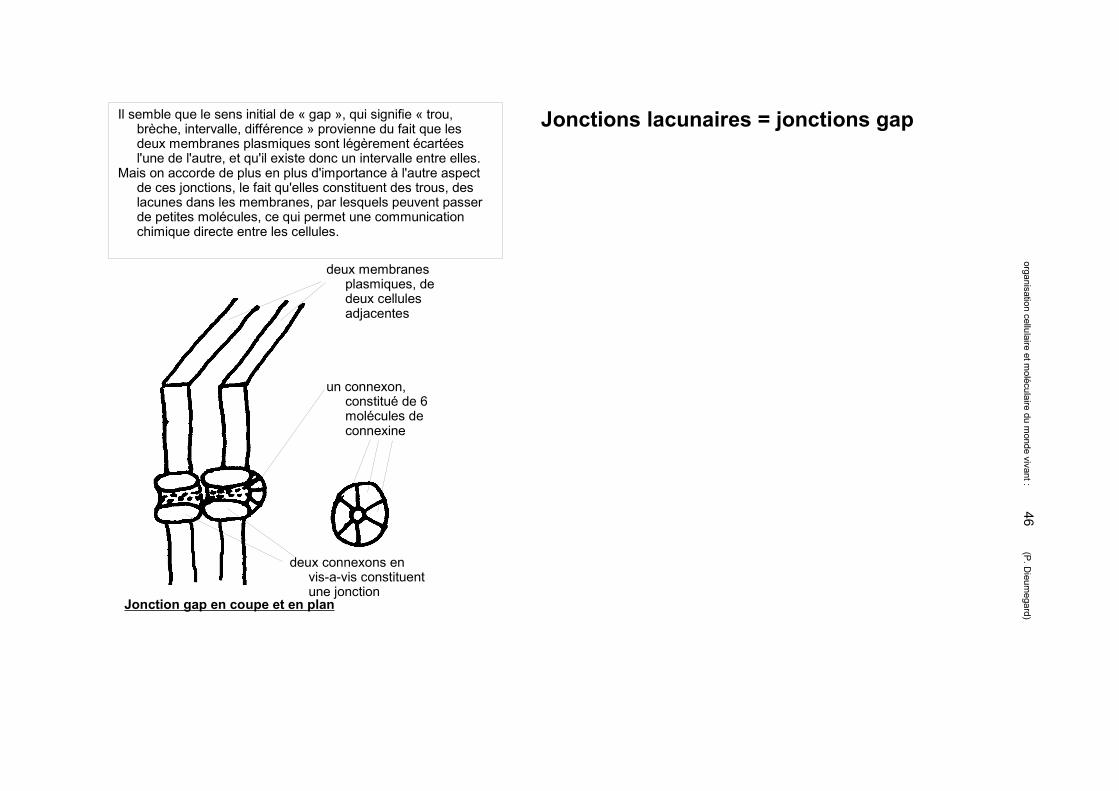
Jonctions lacunaires = jonctions gapIl semble que le sens initial de « gap », qui signifie « trou, brèche, intervalle, différence » provienne du fait que les deux membranes plasmiques sont légèrement écartées l'une de l'autre, et qu'il existe donc un intervalle entre elles.
Mais on accorde de plus en plus d'importance à l'autre aspect de ces jonctions, le fait qu'elles constituent des trous, des lacunes dans les membranes, par lesquels peuvent passer de petites molécules, ce qui permet une communication chimique directe entre les cellules.
Jonction gap en coupe et en plan
deux membranes plasmiques, de deux cellules adjacentes
un connexon, constitué de 6 molécules de connexine
deux connexons en vis-a-vis constituent une jonction
organisation cellulaire et moléculaire du m
onde vivant : 46 (P. D
ieumegard)
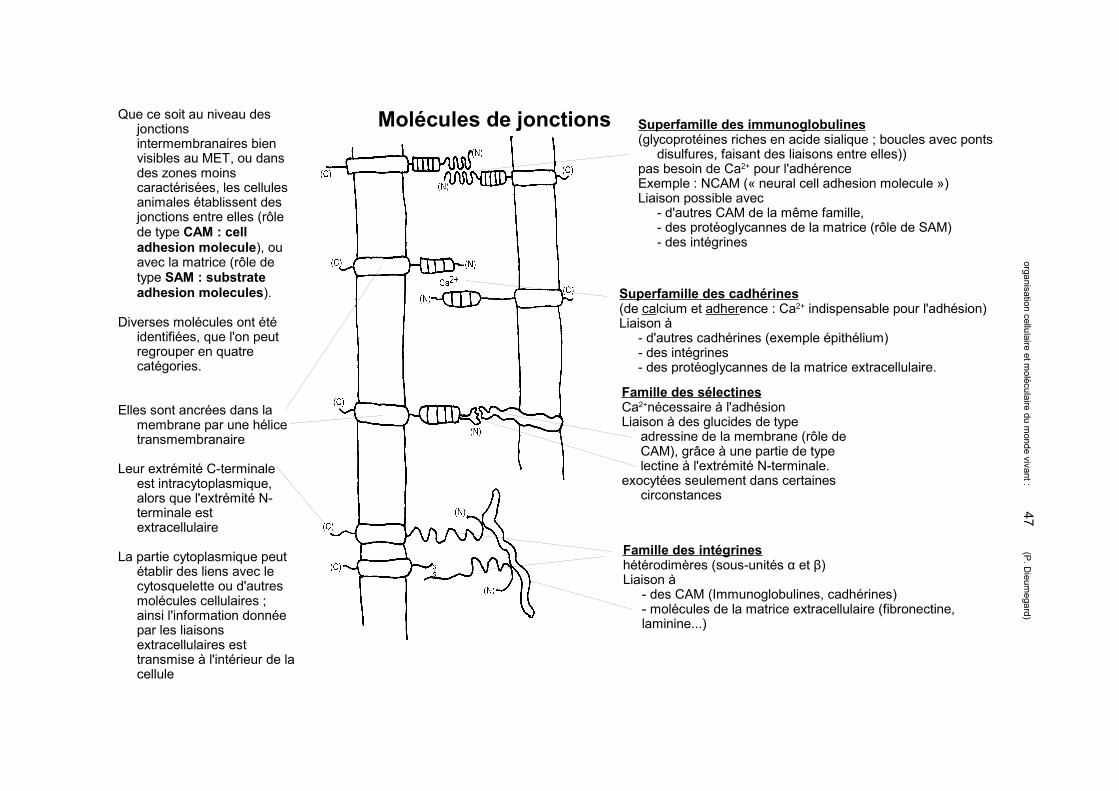
Molécules de jonctionsQue ce soit au niveau des jonctions intermembranaires bien visibles au MET, ou dans des zones moins caractérisées, les cellules animales établissent des jonctions entre elles (rôle de type CAM : cell adhesion molecule), ou avec la matrice (rôle de type SAM : substrate adhesion molecules).
Diverses molécules ont été identifiées, que l'on peut regrouper en quatre catégories.
Elles sont ancrées dans la membrane par une hélice transmembranaire
Leur extrémité C-terminale est intracytoplasmique, alors que l'extrémité N-terminale est extracellulaire
La partie cytoplasmique peut établir des liens avec le cytosquelette ou d'autres molécules cellulaires ; ainsi l'information donnée par les liaisons extracellulaires est transmise à l'intérieur de la cellule
Superfamille des immunoglobulines(glycoprotéines riches en acide sialique ; boucles avec ponts
disulfures, faisant des liaisons entre elles))pas besoin de Ca2+ pour l'adhérenceExemple : NCAM (« neural cell adhesion molecule »)Liaison possible avec
- d'autres CAM de la même famille, - des protéoglycannes de la matrice (rôle de SAM)- des intégrines
Superfamille des cadhérines(de calcium et adherence : Ca2+ indispensable pour l'adhésion)Liaison à
- d'autres cadhérines (exemple épithélium)- des intégrines- des protéoglycannes de la matrice extracellulaire.
Famille des sélectinesCa2+nécessaire à l'adhésionLiaison à des glucides de type
adressine de la membrane (rôle de CAM), grâce à une partie de type lectine à l'extrémité N-terminale.
exocytées seulement dans certaines circonstances
Famille des intégrineshétérodimères (sous-unités α et β)Liaison à
- des CAM (Immunoglobulines, cadhérines)- molécules de la matrice extracellulaire (fibronectine, laminine...)
organisation cellulaire et moléculaire du m
onde vivant : 47 (P. D
ieumegard)

Transports transmembranaires
compartiment 1 : concentration C1
du soluté étudié
compartiment 2 :concentration C2
du soluté
membrane
Transports perméatifs : sans déformation d'ensemble de la membrane
Transports cytotiques : avec déformation de la membrane
Transports passifs : ils ne nécessitent pas d'énergie extérieure, et sont possibles avec des cellules mortes, ou des membranes artificielles. Ils correspondent à une diminution de la différence de potentiel électrochimique du soluté considéré.
Transports actifs : ils correspondent à une augmentation de la différence de potentiel électrochimique du soluté considéré. Ils nécessitent de l'énergie extérieure, et ne sont donc possibles que pour des cellules vivantes, ayant une source d'énergie (ATP, DDP électrique, ou autre).
Ils sont toujours actifs, car la déformation de la membrane nécessite de l'énergie (hydrolyse d'ATP et déformation du cytosquelette, par exemple).
Ce sont les mouvements de molécules d'un côté de membrane à l'autre.Ces molécules peuvent être :- le solvant (l'eau pour les cellules vivantes)- les solutés (substances dissoutes : glucose, acides aminés, Na+, K+, H+, etc)
Il y a deux conditions pour qu'un transport passif se produise réellement :
- condition thermodynamique : la substance doit être en déséquilibre. Pour les molécules non chargées, il faut une différence de concentration ; pour les ions, il faut une différence de potentiel électrochimique.
- condition cinétique : la membrane doit être perméable à la substance considérée. Les molécules hydrophobes passent à travers la bicouche de lipides. L'eau et les petites molécules hydrophiles passent plutôt par les protéines, soit quelconques, soit spécialisées (aquaporines, perméases, canaux ioniques...). Ces protéines spécialisées sont soumises à une régulation, ce qui permet à la cellule de contrôler le flux à travers la membrane.
organisation cellulaire et moléculaire du m
onde vivant : 48 (P. D
ieumegard)
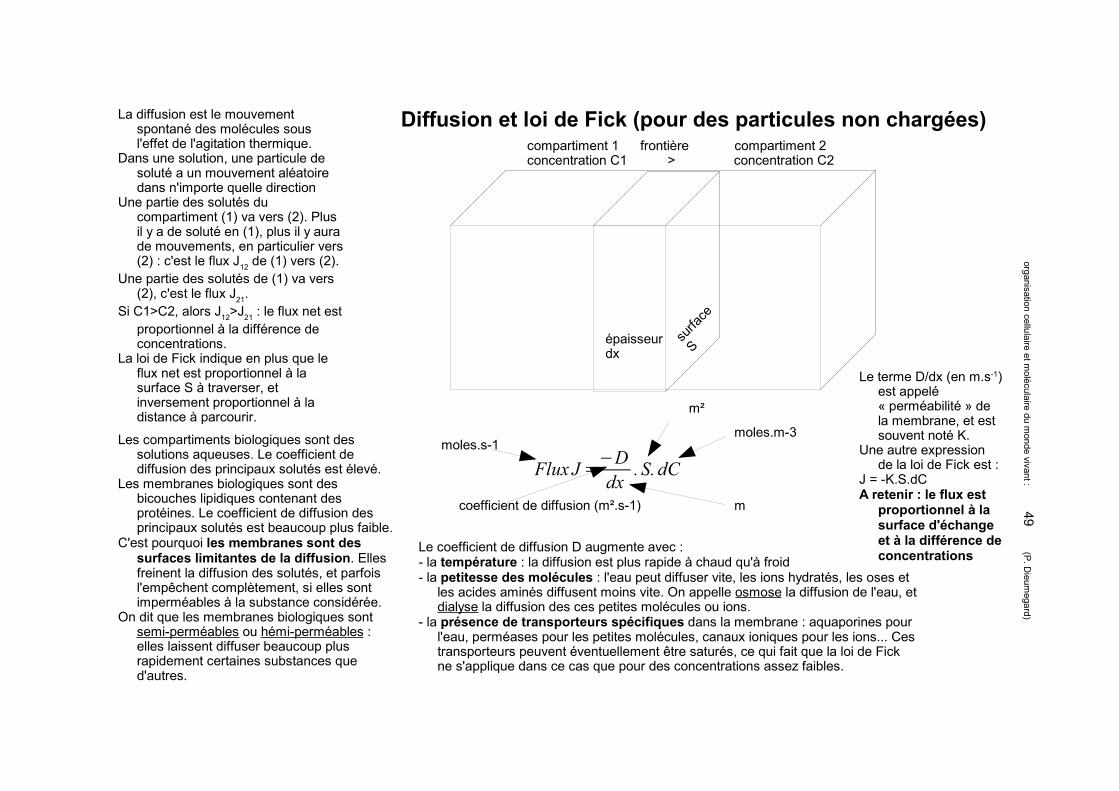
Diffusion et loi de Fick (pour des particules non chargées)La diffusion est le mouvement spontané des molécules sous l'effet de l'agitation thermique.
Dans une solution, une particule de soluté a un mouvement aléatoire dans n'importe quelle direction
Une partie des solutés du compartiment (1) va vers (2). Plus il y a de soluté en (1), plus il y aura de mouvements, en particulier vers (2) : c'est le flux J12 de (1) vers (2).
Une partie des solutés de (1) va vers (2), c'est le flux J21.
Si C1>C2, alors J12>J21 : le flux net est proportionnel à la différence de concentrations.
La loi de Fick indique en plus que le flux net est proportionnel à la surface S à traverser, et inversement proportionnel à la distance à parcourir.
épaisseur dx
compartiment 1 frontière compartiment 2concentration C1 > concentration C2
surfa
ce
S
Flux J=−Ddx. S. dC
moles.m-3
m
moles.s-1
m²
coefficient de diffusion (m².s-1)
Les compartiments biologiques sont des solutions aqueuses. Le coefficient de diffusion des principaux solutés est élevé.
Les membranes biologiques sont des bicouches lipidiques contenant des protéines. Le coefficient de diffusion des principaux solutés est beaucoup plus faible.
C'est pourquoi les membranes sont des surfaces limitantes de la diffusion. Elles freinent la diffusion des solutés, et parfois l'empêchent complètement, si elles sont imperméables à la substance considérée.
On dit que les membranes biologiques sont semi-perméables ou hémi-perméables : elles laissent diffuser beaucoup plus rapidement certaines substances que d'autres.
Le coefficient de diffusion D augmente avec :- la température : la diffusion est plus rapide à chaud qu'à froid- la petitesse des molécules : l'eau peut diffuser vite, les ions hydratés, les oses et
les acides aminés diffusent moins vite. On appelle osmose la diffusion de l'eau, et dialyse la diffusion des ces petites molécules ou ions.
- la présence de transporteurs spécifiques dans la membrane : aquaporines pour l'eau, perméases pour les petites molécules, canaux ioniques pour les ions... Ces transporteurs peuvent éventuellement être saturés, ce qui fait que la loi de Fick ne s'applique dans ce cas que pour des concentrations assez faibles.
Le terme D/dx (en m.s-1) est appelé « perméabilité » de la membrane, et est souvent noté K.
Une autre expression de la loi de Fick est :
J = -K.S.dC A retenir : le flux est
proportionnel à la surface d'échange et à la différence de concentrations
organisation cellulaire et moléculaire du m
onde vivant : 49 (P. D
ieumegard)

Traversée passive des membranes par les ions : principe du potentiel électrochimique
La loi de Fick (flux = k.Δc) indique que les particules dissoutes vont avoir un mouvement qui aboutit à l'équilibration des concentrations, ce qui correspond à la loi physique générale : les phénomènes aboutissent à une diminution de l'énergie potentielle, et finalement à un équilibre.
Ceci n'est valable que pour les molécules non chargées. En plus d'être soumis à la force de diffusion causée par la différence de concentration, les ions sont soumis à la force électrique : ils sont attirés par le compartiment de charge opposée.
Pour un ion donné, on peut définir le potentiel électrochimique d'un compartiment. Si les deux compartiments ont le même potentiel électrochimique, l'ion va être en équilibre. Sinon, l'ion va être en déséquilibre, et traversera la membrane de façon à aboutir à l'équilibre.
Soit un dispositif de deux compartiments séparés par une membrane semi-perméable, et soumis à des potentiels électriques différents :
Le glucose n'est pas chargé. Il sera soumis seulement à la force de diffusion, et ira de la gauche vers la droite.
Le sodium Na+ est attiré vers la droite à la fois par la force de diffusion ([Na+] est plus grande à gauche) et par la force électrique (potentiel négatif à droite). Il ira de la gauche vers la droite.
Le potassium K+ est attiré vers la gauche par la force de diffusion ([K+] est plus faible à gauche) mais est attiré vers la droite par la force électrique, qui l'attire vers le compartiment de charge opposée (-). Son flux net n'est pas facile à prédire, et il faut utiliser la loi de Nernst.
potentiel électrochimique (J/mole) μ = μ0 + R.T.Ln(c) + z.V.Fpour un ion et un compartiment donnésμ0 est le potentiel électrochimique de référenceR est la constante des gaz parfaits (8,314 J/mole)T est la température absoluec est la concentration de l'ion dans le compartimentz est la charge de l'ion (1 pour K+ et Na+, 2 pour Ca2+, -1 pour Cl-, etc)V est le potentiel électrique du compartimentF est la constante de Faraday (charge d'une mole d'électrons = 96500 coulombs)
+ -
potentiel électrique V1 (+) potentiel électrique V2 (-)
Na+Na+
K+ K+
glucose glucose
Les deux compartiments (1) et (2) seront en équilibre pour l'ion considéré s'ils ont la même énergie, c'est à dire si μ1 = μ2, c'est à dire si μ1 – μ2 = 0
μ1 – μ2 = R.T. (Ln(c1) – Ln(c2)) – z. F (V1 – V2)= R.T.Ln(c1/ c2) - z.F(V1 – V2)
Pour que l'ion soit en équilibre, il faut que z.F(V1 – V2) = R.T.Ln(c1 / c2) donc
V ١−V ٢=RTz F
ln c١−c٢
organisation cellulaire et moléculaire du m
onde vivant : 50 (P. D
ieumegard)

Osmose : traversée des membranes par l'eauGlobalement, les mouvements passifs d'eau
aboutissent au même résultat que tous les mouvements de diffusion : équilibration des concentrations.
En d'autres termes, l'eau va du compartiment où elle est « proportionnellement la plus abondante » vers le compartiment où elle est « proportionnellement la plus rare ».
Donc l'eau va du compartiment le plus concentré en soluté vers le compartiment le moins concentré en soluté : c'est l'osmose.
On définit le potentiel hydrique d'un compartiment comme la capacité de ce compartiment à donner de l'eau aux autres.
Le potentiel hydrique a plusieurs constituants :- potentiel osmotique, si les concentrations sont
différentes- potentiel hydrostatique, si les pressions sont
différentes (l'eau va vers le compartiment de pression plus faible)
- potentiel d'imbibition, si un compartiment aspire l'eau par capillarité
- etc.
Par rapport à la cellule ou l'organisme de référence, on dit que le milieu extérieur est
isotonique s'il est aussi concentré ; le flux moyen d'eau par osmose est nul.
hypertonique s'il est plus concentré ; le flux moyen d'eau par osmose va vers le milieu extérieur, et l'organisme ou la cellule se déshydrate.
hypotonique s'il est moins concentré ; le flux moyen d'eau par osmose va vers la cellule ou l'organisme, qui a tendance à gonfler.
La loi fondamentale de l'osmose : loi de Van T'Hoff
Na+Na+
K+ K+
glucose glucose
concentration totale C1
concentration totale C2
Π.V = - n. R. Toù Π est le potentiel osmotiqueV le volume du compartimentn le nombre de particules
dissoutes (ions ou molécules)
R est la constante des gaz parfaits
T est la température absolue.
milieu isotonique milieu hypotonique milieu hypertonique
(les flèches figurent le mouvement d'eau)
donc entre les deux compartiments :ΔΠ = - R. T. ΔC, si ΔC est la différence de concentration (tous solutés confondus) entre
les deux compartiments.
A retenir : les milieux très concentrés attirent l'eau par osmose (potentiel hydrique très négatif)
organisation cellulaire et moléculaire du m
onde vivant : 51 (P. D
ieumegard)

Perméases : protéines de perméabilité aux solutés, avec liaisons faibles
C'est ici l'aspect cinétique de la perméabilité : il ne suffit pas qu'un soluté soit en déséquilibre, il faut que la composition de la membrane lui permette de passer.
différence de concentration
flux global
Mise en évidence : étude de la pénétration du glucose dans l'hématie
Grâce à du glucose marqué isotopiquement, on peut étudier la pénétration du glucose dans l'hématie (la concentration externe est constante, et égale à la glycémie ; le glucose est consommé par le métabolisme de la cellule).
Le glucose pénètre très rapidement dans l'hématie, mais le flux n'est pas proportionnel à la différence de concentration.
La courbe (flux entrant)= f(différence de concentration) ressemble à une hyperbole, donc à une courbe de Michaelis, mais l'asymptote n'est pas horizontale.
Avec un inhibiteur compétitif, la pénétration est beaucoup moins rapide, et suit la loi de Fick.
En faisant la différence entre les deux, on obtient une courbe qui est de type michaelien ; on suppose donc que la pénétration naturelle est la somme de deux types de pénétration, la diffusion simple et la diffusion facilitée, qui se ferait grâce à une protéine fonctionnant comme une enzyme, nommée la glucose-perméase.
En faisant l'analogie avec une réaction enzymatique :
glucose ext. + perméase <==>glucose-perméase ---->perméase + glucose intérieur
C'est une protéine spécialisée, qui fait passer le D-glucose et le D-galactose, mais non les formes L.
On peut définir une vitesse maximale (de pénétration) et une constante de Michaelis ([glucose] pour la moitié de Vmax)
Km=[ perméase seule ]. [glucose ext][ perméase−glucose]
V=Vmax . [S ]Km[S ]
Modèle moléculaire :Les perméases sont dans l'épaisseur de la
membrane, fixent le glucose d'un côté, se déforment et libèrent le glucose de l'autre côté.
Lorsqu'il y a beaucoup de glucose, la perméase peut être saturée : son site actif est encombré par les molécules en train d'être transférées, et il n'y a pas de place pour des molécules supplémentaires.
À retenir:- c'est un phénomène passif- il y a des liaisons faibles entre la perméase et la
molécule transportée, ce qui donne une grande spécificité à la perméase. La glucose-perméase est spécialisée dans le transport du glucose, et ne transporte pas d'autres oses ou oligosides.
- la vitesse de transfert est relativement lente, car le nombre de perméases dans la membrane est limité, et qu'il faut un certain temps pour que l'ensemble fixation-déformation-libération se déroule.
fixation du substrat déformation => pénétration
libérationdéformation => retour
à l'état initial
organisation cellulaire et moléculaire du m
onde vivant : 52 (P. D
ieumegard)
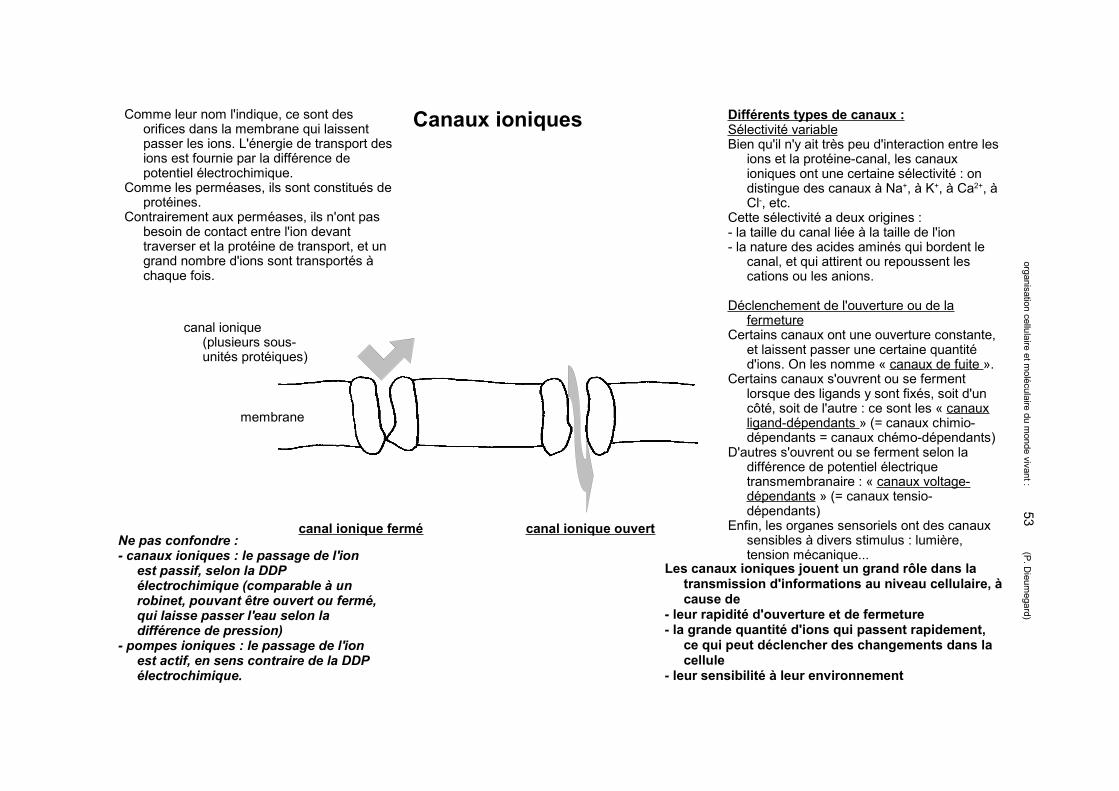
Canaux ioniquesComme leur nom l'indique, ce sont des orifices dans la membrane qui laissent passer les ions. L'énergie de transport des ions est fournie par la différence de potentiel électrochimique.
Comme les perméases, ils sont constitués de protéines.
Contrairement aux perméases, ils n'ont pas besoin de contact entre l'ion devant traverser et la protéine de transport, et un grand nombre d'ions sont transportés à chaque fois.
membrane
canal ionique (plusieurs sous-unités protéiques)
canal ionique fermé canal ionique ouvert
Différents types de canaux :Sélectivité variableBien qu'il n'y ait très peu d'interaction entre les
ions et la protéine-canal, les canaux ioniques ont une certaine sélectivité : on distingue des canaux à Na+, à K+, à Ca2+, à Cl-, etc.
Cette sélectivité a deux origines :- la taille du canal liée à la taille de l'ion- la nature des acides aminés qui bordent le
canal, et qui attirent ou repoussent les cations ou les anions.
Déclenchement de l'ouverture ou de la fermeture
Certains canaux ont une ouverture constante, et laissent passer une certaine quantité d'ions. On les nomme « canaux de fuite ».
Certains canaux s'ouvrent ou se ferment lorsque des ligands y sont fixés, soit d'un côté, soit de l'autre : ce sont les « canaux ligand-dépendants » (= canaux chimio-dépendants = canaux chémo-dépendants)
D'autres s'ouvrent ou se ferment selon la différence de potentiel électrique transmembranaire : « canaux voltage-dépendants » (= canaux tensio-dépendants)
Enfin, les organes sensoriels ont des canaux sensibles à divers stimulus : lumière, tension mécanique...
Les canaux ioniques jouent un grand rôle dans la transmission d'informations au niveau cellulaire, à cause de
- leur rapidité d'ouverture et de fermeture- la grande quantité d'ions qui passent rapidement,
ce qui peut déclencher des changements dans la cellule
- leur sensibilité à leur environnement
Ne pas confondre :- canaux ioniques : le passage de l'ion
est passif, selon la DDP électrochimique (comparable à un robinet, pouvant être ouvert ou fermé, qui laisse passer l'eau selon la différence de pression)
- pompes ioniques : le passage de l'ion est actif, en sens contraire de la DDP électrochimique.
organisation cellulaire et moléculaire du m
onde vivant : 53 (P. D
ieumegard)

Méthode d'étude des canaux ioniques : technique du patch-clamp
(pas encore fait !)
organisation cellulaire et moléculaire du m
onde vivant : 54 (P. D
ieumegard)

Transports actifs primaires et secondaires
Le transport actif d'un soluté nécessite de l'énergie pour faire passer ce soluté d'un côté à l'autre de la membrane (il augmente la DDP électrochimique de ce soluté : ΔG>0)
Cette énergie provient du couplage avec une autre réaction physique ou chimique libérant de l'énergie (ΔG<0) :- hydrolyse d'ATP ou autre réaction chimique pour les transports primaires. Exemple : pompe à Na+/K+ des cellules animales.- transport passif d'un soluté en fort déséquilibre (grande DDP électrochimique) pour les cotransports.
organisation cellulaire et moléculaire du m
onde vivant : 55 (P. D
ieumegard)

La pompe à Na+/K+ des cellules animales
3 Na+
3 Na+
2 K+
2 K+
ATPADP
P
PP
ext
membrane
int
fixation de 3 Na + à l'intérieur
hydrolyse d'ATP, phosphorylation, déformation, départ des Na + à l'extérieur
arrivée de 2 K + à l'extérieurdéphosphorylation,
déformation, et départ des 2 K + à l'extérieur
Flux sortant de 24Na
temps(quelques heures)(action d'un inhibiteur métabolique :
arrêt de formation d'ATP)
(injection d'ATP)
pente naturelle : peu à peu, 24Na devient moins abondant dans l'axone
Expérience fondamentale de mise en évidence :Un axone géant de Calmar est incubé dans un milieu contenant du 24Na
(radioactif) : peu à peu le Na qu'il contient devient du 24Na, puisque le 23Na normal initial est remplacé par 24Na.
On le place ensuite dans un milieu contenant du 23Na (normal) ; on mesure l'efflux de Na+ en mesurant l'apparition de radioactivité dans le milieu : elle est due à la sortie de l'isotope radioactif 24Na.
axone géant de Calmar, contenant 24Na (radioactif)
milieu extracellulaire, avec 23Na (non radioactif)
Pendant cette expérience, on fait agir sur la cellule un inhibiteur métabolique (DNP, cyanure...) qui bloque la formation d'ATP par les mitochondries. On constate que l'efflux de Na+ diminue très fortement. C'est donc qu'il y a besoin d'ATP pour faire sortir Na+ de la cellule.
Confirmation : en injectant directement de l'ATP dans l'axone, on rétablit transitoirement l'efflux de Na+, et lorsqu'on cesse de faire agir l'inhibiteur, l'efflux de Na+ reprend.
Conclusion : il existe une « pompe métabolique », qui expulse Na+ en utilisant l'énergie d'hydrolyse de l'ATP.
Par ailleurs, on peut montrer que l'efflux de Na+ diminue lorsqu'il n'y a pas du tout de K+ dans le milieu. Donc l'efflux de Na+ est couplé à un influx de K+.
Parmi les protéines membranaires, on a trouvé une ATP-ase qui est activée par la présence de K+ et de Na+ : « ATP-ase Na+/K+ dépendante ». Cette molécule est la pompe en question.
Elle est constituée de 4 sous unités, de deux types, α et β
Modèle moléculaire de la pompe à sodium-potassium
organisation cellulaire et moléculaire du m
onde vivant : 56 (P. D
ieumegard)
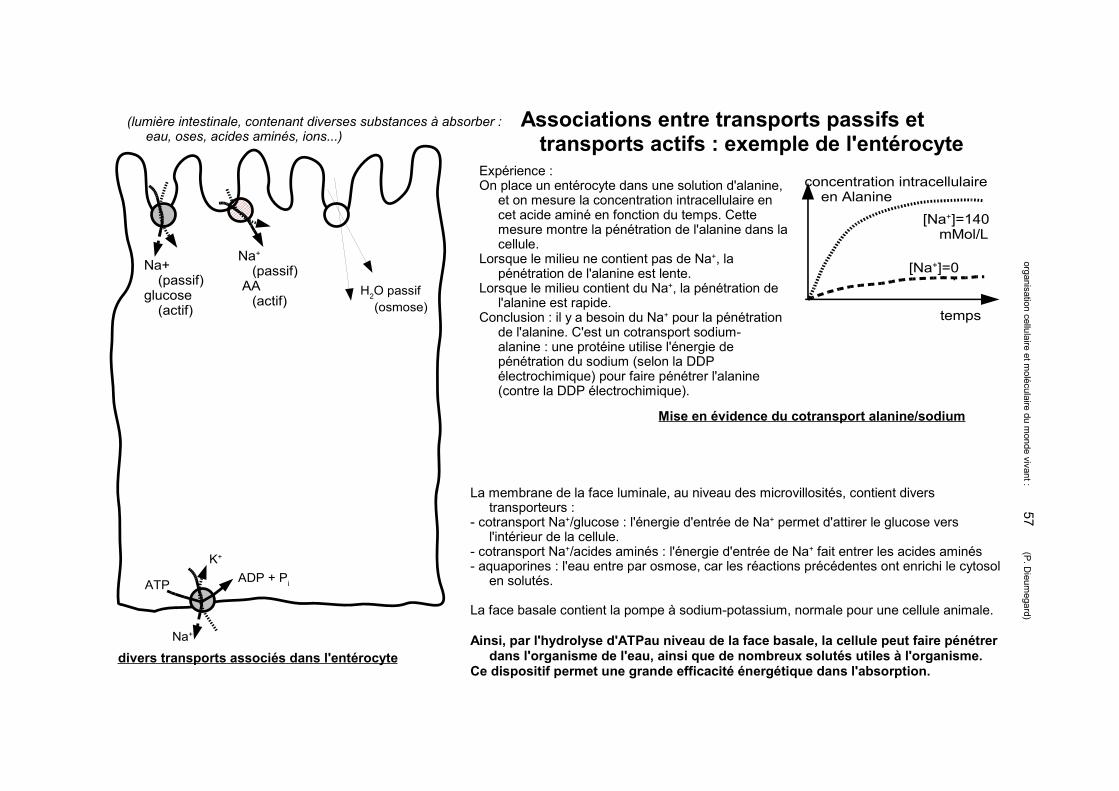
Na+ (passif)
glucose (actif)
Na+
(passif) AA
(actif)
ATP ADP + Pi
H2O passif (osmose)
K+
Na+
Associations entre transports passifs et transports actifs : exemple de l'entérocyte
(lumière intestinale, contenant diverses substances à absorber : eau, oses, acides aminés, ions...)
La membrane de la face luminale, au niveau des microvillosités, contient divers transporteurs :
- cotransport Na+/glucose : l'énergie d'entrée de Na+ permet d'attirer le glucose vers l'intérieur de la cellule.
- cotransport Na+/acides aminés : l'énergie d'entrée de Na+ fait entrer les acides aminés- aquaporines : l'eau entre par osmose, car les réactions précédentes ont enrichi le cytosol
en solutés.
La face basale contient la pompe à sodium-potassium, normale pour une cellule animale.
Ainsi, par l'hydrolyse d'ATPau niveau de la face basale, la cellule peut faire pénétrer dans l'organisme de l'eau, ainsi que de nombreux solutés utiles à l'organisme.
Ce dispositif permet une grande efficacité énergétique dans l'absorption.
temps
concentration intracellulaire en Alanine
[Na+]=0
[Na+]=140 mMol/L
Expérience :On place un entérocyte dans une solution d'alanine,
et on mesure la concentration intracellulaire en cet acide aminé en fonction du temps. Cette mesure montre la pénétration de l'alanine dans la cellule.
Lorsque le milieu ne contient pas de Na+, la pénétration de l'alanine est lente.
Lorsque le milieu contient du Na+, la pénétration de l'alanine est rapide.
Conclusion : il y a besoin du Na+ pour la pénétration de l'alanine. C'est un cotransport sodium-alanine : une protéine utilise l'énergie de pénétration du sodium (selon la DDP électrochimique) pour faire pénétrer l'alanine (contre la DDP électrochimique).
Mise en évidence du cotransport alanine/sodium
divers transports associés dans l'entérocyte
organisation cellulaire et moléculaire du m
onde vivant : 57 (P. D
ieumegard)

Mouvements cytotiques : endocytose, exocytose, bourgeonnement...
Les macromolécules, par exemple les protéines, ne peuvent pas traverser les membranes. Il existe donc des mécanismes leur permettant de passer de l'intérieur de la cellule à l'extérieur (ou inversement) grâce à des mouvements de déformation de la membrane.
Définitions :Endocytose : une partie de l'espace
extracellulaire est transférée à l'intérieur de la cellule (phénomène d'internalisation). Les vésicules ainsi formées sont les endosomes. Exemple : endocytose des bactéries étrangères par les leucocytes.
Exocytose : exportation de matériel intracellulaire (externalisation), sans exportation de membrane plasmique. Exemple : exocytose des enzymes digestives par la cellule acineuse pancréatique.
Transcytose : traversée d'une cellule par endocytose d'un côté, traversée de la cellule par la vésicule, et exocytose de l'autre côté (ex : endothéliums et épithéliums polarisés).
Bourgeonnement : exportation de matériel intracellulaire, avec exportation de membrane plasmique. Exemple : bourgeonnement de virions du VIH (virus du SIDA).Il existe aussi divers types de bourgeonnement de vésicules à l'intérieur du cytoplasme (par exemple bourgeonnement du RE)
(milieu extérieur)
membrane plasmique
invagination ==> endocytose
vésicule d'endocytose
fusion de vésicules
fusion d'une vésicule avec la membrane ==> exocytose
Principe général d'endocytose et exocytose
(cytosol)
principe du bourgeonnement de la membrane plasmique
particule issue du bourgeonnement
(entourée par une membrane)
membrane plasmique
milieu extracellulaire
cytosol
organisation cellulaire et moléculaire du m
onde vivant : 58 (P. D
ieumegard)

Divers types d'endocytose
On a coutume de définir 3 types d'endocytose selon la taille des vésicules et le mécanisme :
pinocytose (pino = « boire » en grec ») : les vésicules sont de faible volume, diamètre inférieur à 150 nm ; pas de revêtement sur la face cytosolique visible au microscope électronique ;les molécules à internaliser sont concentrées par le revêtement fibreux glucidique (cell-coat = glycocalyx). C'est un phénomène permanent.
endocytose par récepteurs ; les vésicules sont aussi de faible volume ; l'endocytose ne se fait que lorsque des récepteurs spécialisés ont fixé leur ligand.
phagocytose (phago = « manger » en grec) ; volume important, diamètre supérieur à 1µm.
Divers types d'endocytose par récepteurs :
Avec un manteau de clathrine et de protéines associées du côté cytosolique
Avec un revêtement de caveoline du côté cytosolique
(sans revêtement observable au MET...)
organisation cellulaire et moléculaire du m
onde vivant : 59 (P. D
ieumegard)

Un exemple de mécanisme d'endocytose : internalisation et recyclage des récepteurs aux LDL
Rôle biologique : fourniture à la cellules de molécules du milieu extérieur, mais qui n'y existent qu'en faible concentration.
organisation cellulaire et moléculaire du m
onde vivant : 60 (P. D
ieumegard)

Endocytose de cavéoles = vésicules à caveoline
organisation cellulaire et moléculaire du m
onde vivant : 61 (P. D
ieumegard)
(pas encore fait)

Biosynthèse et renouvellement de la membrane plasmique
La membrane plasmique est une structure permanente de la cellule, mais ses constituants sont renouvelés rapidement.
Dans une cellule animale, les molécules membranaires ne persistent que quelques jours (demi-vie des glycoprotéines : 2 à 3 jours ; demi-vie des lipides : 3 à 5 jours)
De la nouvelle membrane apparait par exocytose de vésicules golgiennes.
Les molécules de la membrane disparaissent par endocytose, mais aussi par destruction directe des molécules in situ.
Il est fréquent que le plasmalemme des cellules soit déchiré, en particulier lorsqu'elles sont soumises à des contraintes mécaniques fortes (cellules musculaires).
La mise en communication du cytosol et du milieu extracellulaire provoque l'annulation de la DDP transmembranaire et des mouvements d'eau et de solutés qui pourraient rapidement provoquer la mort de toute la cellule.
Il est donc indispensable que la membrane soit rapidement cicatrisée.
Cette cicatrisation se fait par fusion de vésicules diverses (une sorte d'exocytose commune), déclenchée par l'entrée d'ions Ca2+ à partir de l'extérieur (puisqu'à l'état normal, il y a très peu d'ions Ca2+ dans le cytosol alors que cet ion est en fort déséquilibre électrochimique).
organisation cellulaire et moléculaire du m
onde vivant : 62 (P. D
ieumegard)
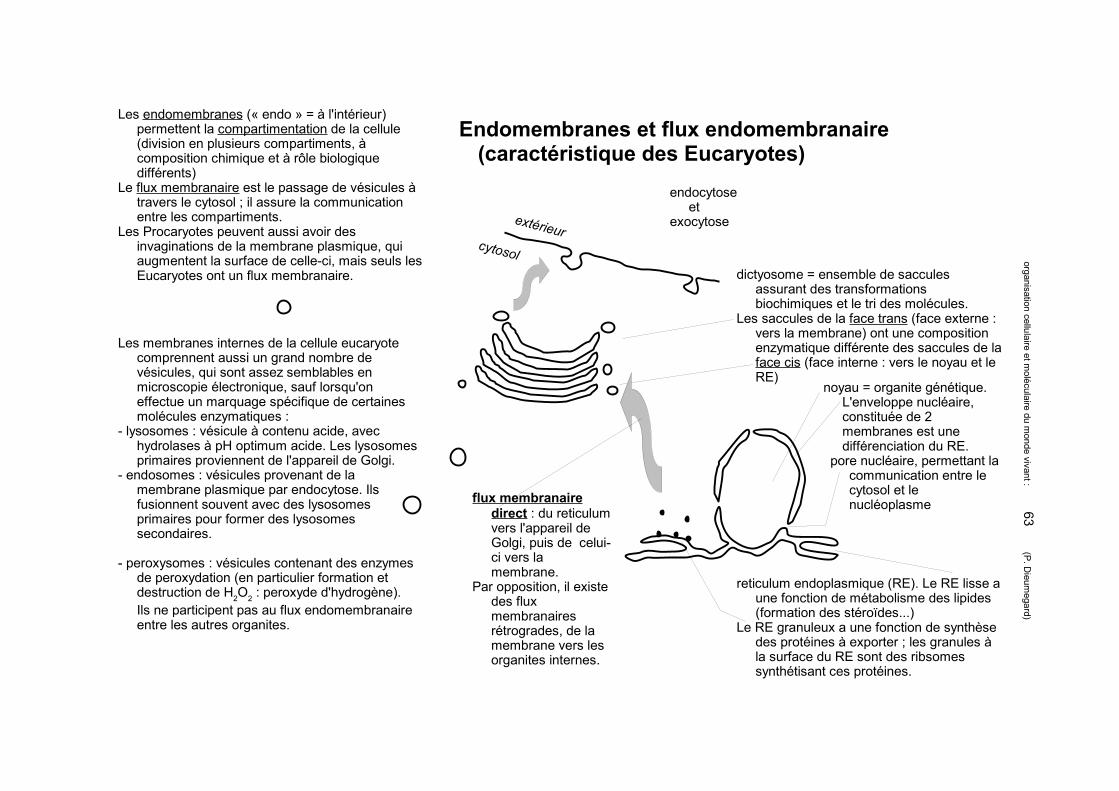
Endomembranes et flux endomembranaire (caractéristique des Eucaryotes)
extérieurcytosol
Les endomembranes (« endo » = à l'intérieur) permettent la compartimentation de la cellule (division en plusieurs compartiments, à composition chimique et à rôle biologique différents)
Le flux membranaire est le passage de vésicules à travers le cytosol ; il assure la communication entre les compartiments.
Les Procaryotes peuvent aussi avoir des invaginations de la membrane plasmique, qui augmentent la surface de celle-ci, mais seuls les Eucaryotes ont un flux membranaire.
endocytoseet
exocytose
dictyosome = ensemble de saccules assurant des transformations biochimiques et le tri des molécules.
Les saccules de la face trans (face externe : vers la membrane) ont une composition enzymatique différente des saccules de la face cis (face interne : vers le noyau et le RE)
noyau = organite génétique. L'enveloppe nucléaire, constituée de 2 membranes est une différenciation du RE.
reticulum endoplasmique (RE). Le RE lisse a une fonction de métabolisme des lipides (formation des stéroïdes...)
Le RE granuleux a une fonction de synthèse des protéines à exporter ; les granules à la surface du RE sont des ribsomes synthétisant ces protéines.
organisation cellulaire et moléculaire du m
onde vivant : 63 (P. D
ieumegard)
pore nucléaire, permettant la communication entre le cytosol et le nucléoplasme
Les membranes internes de la cellule eucaryote comprennent aussi un grand nombre de vésicules, qui sont assez semblables en microscopie électronique, sauf lorsqu'on effectue un marquage spécifique de certaines molécules enzymatiques :
- lysosomes : vésicule à contenu acide, avec hydrolases à pH optimum acide. Les lysosomes primaires proviennent de l'appareil de Golgi.
- endosomes : vésicules provenant de la membrane plasmique par endocytose. Ils fusionnent souvent avec des lysosomes primaires pour former des lysosomes secondaires.
- peroxysomes : vésicules contenant des enzymes de peroxydation (en particulier formation et destruction de H2O2 : peroxyde d'hydrogène). Ils ne participent pas au flux endomembranaire entre les autres organites.
flux membranaire direct : du reticulum vers l'appareil de Golgi, puis de celui-ci vers la membrane.
Par opposition, il existe des flux membranaires rétrogrades, de la membrane vers les organites internes.

organisation cellulaire et moléculaire du m
onde vivant : 64 (P. D
ieumegard)
Mitochondries et plastes ne sont pas des endomembranes comme les autres
mitochondrie (organite énergétique)La membrane externe
est assez semblable aux autres membranes cellulaires.
La membrane interne est très différente : grande surface à cause des replis (--> crêtes mitochondriales), pas de stérols mais des cardiolipides ; beaucoup de protéines, en particulier des protéines de transport et de la chaîne respiratoire.
Chloroplaste et plastes divers (végétaux uniquement)Eux aussi ont une grosse différence entre membrane interne
et membrane externe.Les thylakoïdes sont des sacs aplatis,
équivalents aux crêtes mitochondriales ; ils contiennent les pigments photosynthétiques, et sont très nombreux dans les chloroplastes différenciés et fonctionnels, qui sont les organites de la photosynthèse.
Les proplastes sont des plastes non différenciés (très peu de thylakoïdes, pas de pigments photosynthétiques)
Les amyloplastes sont différenciés en contenant un gros grain d'amidon, mais pas ou peu de thylakoïdes..
Mitochondries (pour tous les Eucaryotes) et plastes (pour les végétaux) sont limités par une double membrane.
Le noyau cellulaire aussi est limité par une enveloppe constituée d'une double membrane, mais l'enveloppe nucléaire correspond avec le reticulum endoplasmique, et disparait à la prophase pour se reconstituer à la télophase de la mitose.
Au contraire, les deux membranes des mitochondries et des plastes sont permanentes, et sont bien différentes l'une de l'autre.
ADN mitochondrial :- forme circulaire, avec des gènes organisés
comme les gènes bactériens (non en mosaïque)
- petite taille <=> très peu de gènes ; ces gènes codent en particulier pour des gènes de la chaîne respiratoire.
ribosomes mitochondriaux = mitoribosomes
- plus petits que les ribosomes du cytosol (comme les ribosomes bactériens)
- sensibles aux mêmes inhibiteurs métaboliques que les ribosomes bactériens
ribosomes plastidiaux (ou chloroplastiques)
Ils sont aussi différents des ribosomes du cytosol.
ADN chloroplastique :- forme circulaire (comme ADN
bactérien)- taille plus grande que l'ADN
mitochondrial (plus de gènes ; en particulier gènes de la photosynthèse)
Ces divers caractères m
ontrent une similitude entre l'organisation des
mitochondries et des plastes et l'organisation d'une bactérie.
Cette sim
ilitude est à la base de la théorie endosymbiotique des m
itochondries et des plastes :
On im
agine que (au cours de l'Archéen), des procaryotes capables de respiration seraient venus vivre en sym
biose dans le cytoplasme d'eucaryotes incapables
de respiration, et peu à peu seraient devenus les mitochondries actuelles.
De m
ême, des procaryotes photosynthétiques seraient venus vivre en sym
biose dans le cytoplasm
e d'Eucaryotes, et seraient devenus les chloroplaste actuels.
Mitochondries et plastes sont des organites semi-autonomes
- semi-autonomie génétique : ils ont du matériel génétique, et des ribosomes pour faire la traduction en protéine, mais cet ADN n'est pas suffisant pour la synthèse de tous leurs constituants.
- semi-autonomie énergétique : ils réalisent respiration et photosynthèse.
Acide aminé...20, 22, 24 sv, 29,57Acide aspartique......................24Acide glycuronique.................15Acide gras...................................8Acides aminés. 2 sv, 6 sv, 24 sv,27, 48 sv, 53, 57Acides nucléiques. .6 sv, 14, 16,20, 22 svAcylglycérols...........................10Adénine................................20 svAdénosine............................20 svAdipocyte ................................10Adressine..................................47Agarose.....................................14Alanine..........................24 sv, 57Aldoses.....................................16Allostérie..................................32Amidon...............................14, 19Amidon ....................................13AMP cyclique..........................20Amphipathiques.........................9Amylopectine...........................19Amyloplastes.....................14, 19Amylose....................................19Anticodon..........................20, 22Aquaporine...................48 sv, 57Archébactéries...........................5ARN de transfert.....................22ARN messager.......5, 20, 22, 26ARN ribosomal.................20, 22ATP............................................20Autoradiographie.......................2Auxine.......................................38Axone géant de Calmar..........56Bactéries .......................5, 40, 58Base pyrimidique....................20BPG...........................................33Cadhérines.........................45, 47
Canaux ioniques.....48 sv, 53 svCardiolipides..............................5Cartilage......................14, 18, 42Caveoline...........................59, 61Cellule épithéliale...................42Cellules ......................................8Cellules acineuses.....................2Cellules bulliformes..................4Cellules de Schwann..............42Cellulose.....13 sv, 18, 37 sv, 41Cétoses......................................16Chaleur de vaporisation............7Chaperonines...........................26Chitine................................14, 16Chloroplaste..................4, 19, 64Cholestérol............................8 svCholine......................................11Chondroïtine sulfate...............14Chromatine diffuse....................2Chromatographie..............25, 29Clathrine...................................59Claudine....................................45Codon...........................20, 22, 26Colibacille..................................5Collagène......................28, 42 svCollenchyme............................37Compartimentation...................3Connexine...........................45 svConnexon.................................46Constante de Michaelis .........52Cotransports.............................55Courbe michaelienne..............30Cryodécapage..........................35Cryofracture.............................35Cuticule.......................................4Cutine.............................8, 12, 39Cyclose........................................4Cystéine..............................24, 27Cytosol........................................3
Cytosquelette3 sv, 36, 39, 47 svDesmosome.......................42, 45Désoxyribose.........14, 16, 20 svDictyosome........2, 4, 39, 44, 63Dictyosomes.............................39Diffusion.........................5, 49 svDiholosides...............................17Dihyroxyacétone.....................14DPG....................................33, 39Eau, H2O....................................7Effecteur allostérique.............33Effet Bohr.................................33Élastine.....................................42Électrophorèse.........................29Endoamylases..........................19Endocytose......................3, 58 svEndomembranes...35, 44, 63 svEndosomes....................3, 58, 63Entérocyte..........................45, 57Enveloppe nucl...3, 5, 34, 63 svÉpiderme ....................................4Érythrocyte...............................34Escherichia coli.........................5Éthanolamine...........................11Eubactéries.................................5Eucaryotes...5, 20, 22, 34 sv, 63svExoamylases............................19Exocytose....2, 39, 44, 58, 62 svExons.......................5, 20, 22, 46Feuillet beta..............................28Fibroblaste................................42Fibroïne.....................................28Fluidité membranaire................8Flux membranaire............44, 63Fructose..............................13, 16Galactose.......................13 sv, 16Globule rouge..........................34Glucides................................6, 13
Glucosamine............................15Glucose..13 sv, 38 sv, 48, 50 sv,57Glycanes...................................17Glycéraldéhyde........................14Glycérides................................10Glycérol....................................10Glycine......................................24Glycocalyx..................35, 40, 59Glycogène................................14Glycolipides.............................11Glycoprotéines......40 sv, 47, 62Glycosaminoglycannes....14, 18Glycosylation.............................3Golgi.......2 sv, 39, 41, 44, 62 svGram............................................5Guanine................................20 svHélice alpha.............................28Hématie...................31, 33 sv, 52Hémicellulose.....................38 svHémoglobine.........24, 26, 30 svHétérocaryon............................36Hexose......................................15Histidine...................................24Hyaloplasme...............................3Hybridation .............................23Hydrophobes..............................8Hydrosquelette...........................4Hypertonique...........................51Hypotonique.............................51Immunoglobuline....................47Inhibiteur compétitif...............52Inositol......................................11Intégrine..................42 sv, 45, 47Interdigitations..................42, 45Introns............................5, 20, 22Ionisation..................................25Isotonique.................................51Jonction gap.............................46
organisation cellulaire et moléculaire du m
onde vivant : 65 (P. D
ieumegard)

Jonction serrée...........................45Jonctions étanches....................42Jonctions lacunaires..................46Lactose............................13 sv, 17Lame basale...............2, 42 sv, 45Lamelle moyenne........18, 37, 39Laminine.........................42 sv, 47Lécithine.....................................11Liaison hydrogène....................27Liaison osidique........................16Liaison peptidique...............26 svLiaisons osidiques.....................17Liaisons peptidiques.................27Lignine.................................37, 39Lipides......................................6, 8Liposolubles.................................8Liqueur de Fehling...................16Loi de Fick......................49 sv, 52Lysosomes.............................3, 63Maltose.......................................17Matrice extracellulaire18, 37, 41sv, 47Membrane. 2 sv, 10 sv, 18, 34 sv,39 sv, 42, 44 sv, 55 sv, 62 svMembrane plasmique 2, 4 sv, 10,34 sv, 39, 42, 44 sv, 58, 62 svMembrane semiperméable......50Mésoderme.................................45Métazoaires................................45Michaelis....................................52Microtubule................................39Microvillosités....................45, 57Mitochondries 2 sv, 34 sv, 56, 64Mitose..........................3, 5, 39, 64Modèle de Singer et Nicholson......................................................35
Mosaïque fluide.........................35Mouvements cytotiques...........58Mucopolysaccharides........14, 18Myélines.....................................11Myoglobine...........................30 svNernst..........................................50Noyau 2 sv, 8, 10, 12, 20, 30, 34,36, 42 sv, 63 svNoyau tétrapyrol.......................30Nucléobases...............................20Nucléoïde.....................................5Nucléosides................................20Nucléotides............6 sv, 20 sv, 26Oligosides............................13, 17Oméga 3........................................9Opérons.........................................5Osamines....................................18Oses.............................................13Osides.........................................17Osmose..........................49, 51, 57Pancréas........................................2Parenchyme .................................4Paroi bactérienne................14, 40Paroi pecto-cellulosique..........37Paroi squelettique.....4, 34, 37 svParoi végétale................14, 38 svPatch-clamp...............................54Pectine........................18 sv, 37 svPentoses...............................14, 16Peptidoglycane...............5, 34, 40Perméabilité...........37, 39, 49, 52Perméase....................48 sv, 52 svPeroxysomes.........................3, 63Phagocytose......................3, 5, 59Phagosomes..................................3Phénylalanine ...........................24
Phloème........................................4Phosphodiestérase................20 svPhosphoglycérolipides.........8, 11Phospholipides.................8 sv, 11Photosynthèse..............................4Phytostérols..................................8Pinocytose..................................59Plasmalemme...........4, 37, 44, 62Plasmide.......................................5Plasmodesme........................37 svPlastes................14, 18 sv, 35, 64Plasticité de la paroi.................38Point isoélectrique....................25Polarisation fonctionnelle..........2Polarisation structurale...............2Polyholosides.............................18Polynucléotide......................20 svPolyosides............................13, 18Polysaccharides. .5 sv, 14, 18, 38sv, 42Pompe à sodium-potassium....56svPompes ioniques.......................53Ponctuation................................37Potentiel électrochimique 48, 50,53Potentiel hydrique.....................51Potentiel osmotique..................51Pouvoir rotatoire.......................13Procaryotes.......................5, 63 svProline.............................24 sv, 38Protéines.....2 sv, 5 sv, 20, 22, 24sv, 29, 34 sv, 40 sv, 47 sv, 52 sv,56, 58 sv, 62 svProtéines oligomériques..........26Protéoglycannes......14, 18, 41 sv
Protides.........................................6Protocollagène...........................43Protomères......................26, 31 svPulse..............................................2Purique........................................20Pyrimidique................................20Reticulum.......2 sv, 34, 44, 63 svRibofuranose ............................16Ribose...................................14, 16Ribosome.2 sv, 5, 20, 22, 34, 40,44, 64Ribozymes...........................20, 22Ribulose 1-5 diphosphate........16Saccharose......................13 sv, 17SDS.............................................29Sélectine.....................................47Semi-aldéhyde...........................15Séquence-signal........................44Sérine.........................8, 11, 24, 44Sphingosine................................12Sporopollénine.............................8SRP..............................................44Stérol...........3, 5, 8 sv, 12, 35, 64Stomates.......................................4Structure tertiaire......................26Subérine....................8, 12, 37, 39Température de fusion..............23Thymine................................20 svTissu conjonctif............18, 42, 45Transcytose................................58Transports actifs .........48, 55, 57Transports cytotiques...............48Transports passifs ..............48, 57Transports perméatifs ..............48Transports transmembranaires48Triacylglycérol.............................8
Triglycérides................................8Trioses.........................................14Turgescence ................................4Ultracentrifugation...................34Uracile...................................20 svVIH..............................................58Virus à ADN .......................20, 22Virus à ARN .......................20, 22Vitamines......................................8Xylème............................4, 37, 39 organisation cellulaire et m
oléculaire du monde vivant : 66 (P
. Dieum
egard)