
Métabolisme bioénergétique et grands cycles de la matière...
130
Métabolisme bioénergétique et grands cycles de la matière métabolisme de l'Oxygène
Transcript of Métabolisme bioénergétique et grands cycles de la matière...

Métabolisme bioénergétiqueet
grands cycles de la matière
métabolisme de l'Oxygène

intérieur
ADP
ATP
H+
ATPasered ox
Chémio osmose – la façon la plus efficace de faire la bioénergétique
N
extérieur
H+
P
N

0
-400H2 / 2H+
HCO2-
/CO2
HS - / SO4 2-
succinate/fumaratelactate/pyruvate
-800
-1200Chl/Chl *
NAD(P)/NAD(P)H
0
+400
+800
+1200
NO2- / NO3
-
H2O / O2
Fe2+ / Fe3+
arsénite/arséniate
N2 / NO3-
N2O / NO
succinate/fumarate
Chl+/Chl
des couples rédox de la bioénergétique

0
-400H2 / 2H+
HCO2-
/CO2
HS - / SO4 2-
succinate/fumaratelactate/pyruvate
-800
-1200Chl/Chl *
L’oxygène et la lumièreabsorbé par une moléculede chlorophylle sont unesource d’énergie puissantepour les êtres vivants.
Leur disponibilitén’est pas lié à unenvironnement spécifique.
NAD(P)/NAD(P)H
0
+400
+800
+1200
NO2- / NO3
-
H2O / O2
Fe2+ / Fe3+
arsénite/arséniate
N2 / NO3-
N2O / NO
succinate/fumarate
Chl+/Chl
L’abondance d’énergiequ’ils mettent à dispositiona permis l’apparitionde la vie multicellulaire
des couples rédox de la bioénergétique

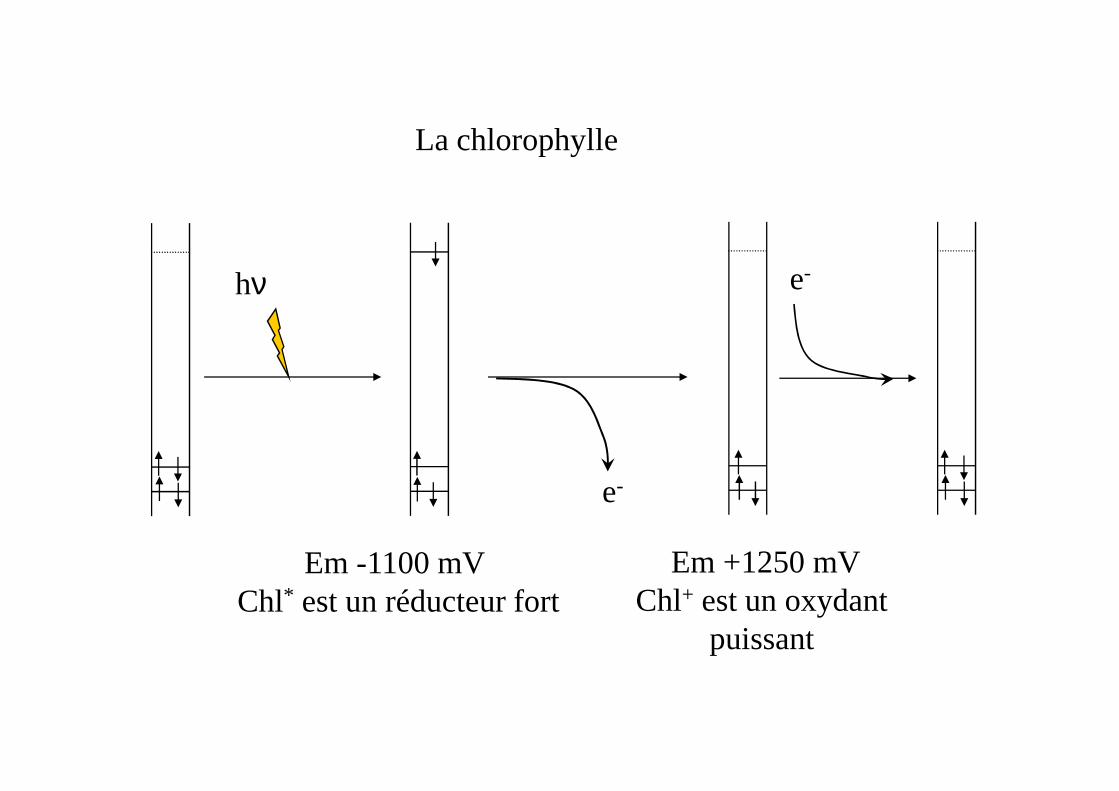
La chlorophylle
hν e-
Em -1100 mV Chl* est un réducteur fort
Em +1250 mVChl+ est un oxydant
puissant
e-

- Chl a- Chl b- BChla- BChl b- BChl c- BChl d- BChl e
les spectres d’absorbance des chlorophylles
Caroteoid

RCII bc1
Q
in
Utilisation de la lumière: centre réactionnel type II
Bactéries pourpres
Chloroflexaceae
photosynthèse anoxygénique
Q
QH2
out
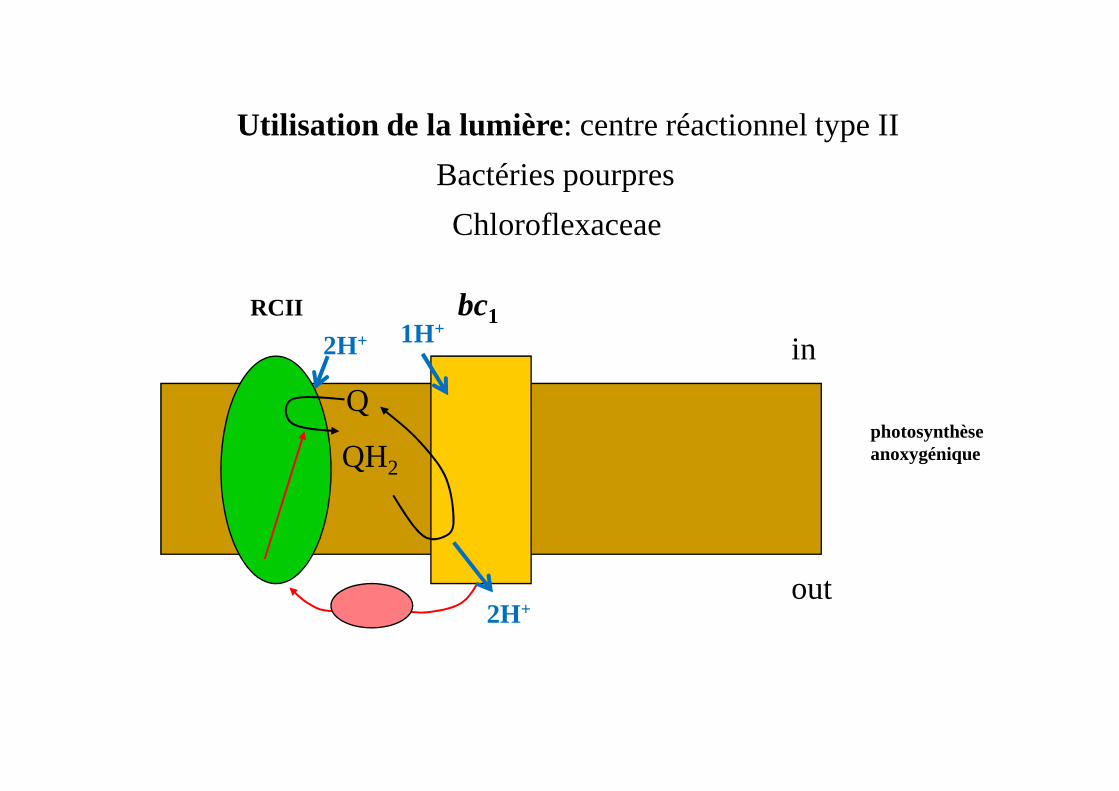
RCII bc1
Q
in
Utilisation de la lumière: centre réactionnel type II
Bactéries pourpres
Chloroflexaceae
2H+ 1H+
photosynthèse anoxygénique
Q
QH2
out2H+

b6cc
Q
in
Utilisation de la lumière: centre réactionnel type I
Héliobactéries
Bactéries vertes-sulfureuses
RCINADPH
photosynthèse anoxygénique
Q
QH2
out
Complex I
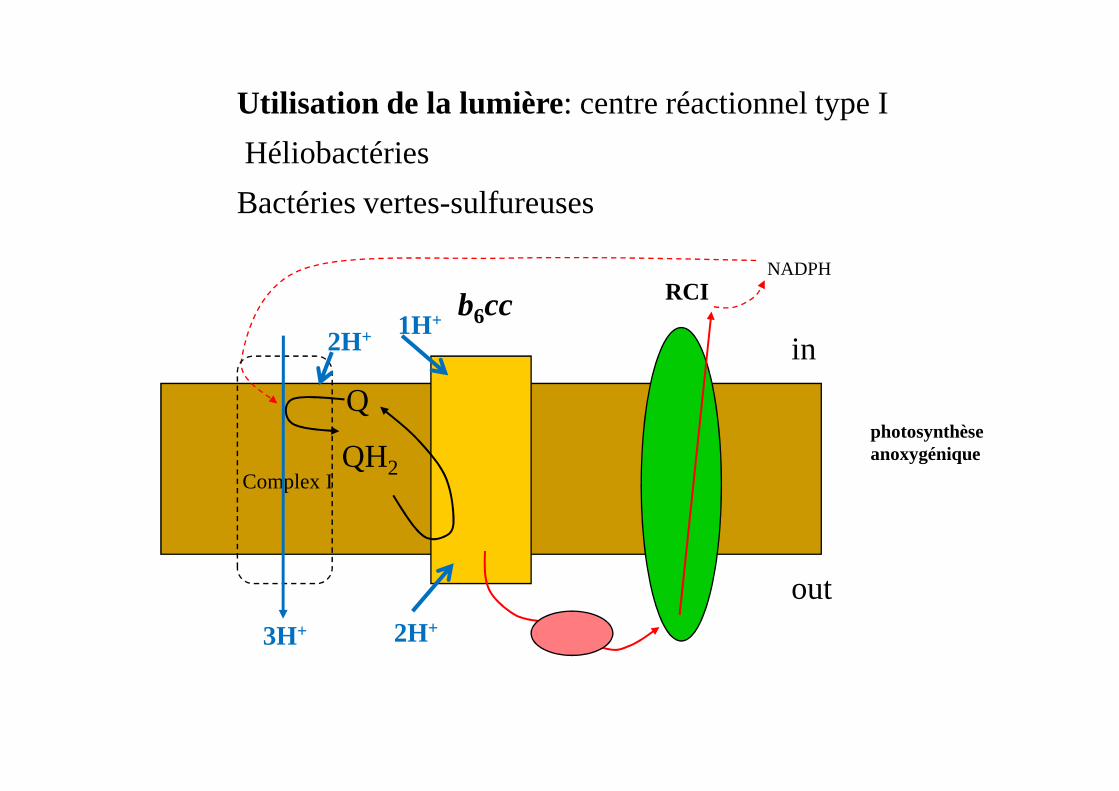
b6cc
Q
in
Utilisation de la lumière: centre réactionnel type I
Héliobactéries
Bactéries vertes-sulfureuses
RCINADPH
2H+ 1H+
photosynthèse anoxygénique
Q
QH2
out
Complex I
2H+3H+

PSII b6fPSI
Q
in
NADPH
Utilisation de la lumière et de l’eau:
Photosynstèmes I et II
Cyanobacteries
Chloroplasts
photosynthèse oxygénique
H2O 1/2O2 +
Q
QH2
out

PSII b6fPSI
Q
in
NADPH
2H+ 1H+
Utilisation de la lumière et de l’eau:
Photosynstèmes I et II
Cyanobacteries
Chloroplasts
photosynthèse oxygénique
H2O 1/2O2 + 2H+
Q
QH2
out
2H+
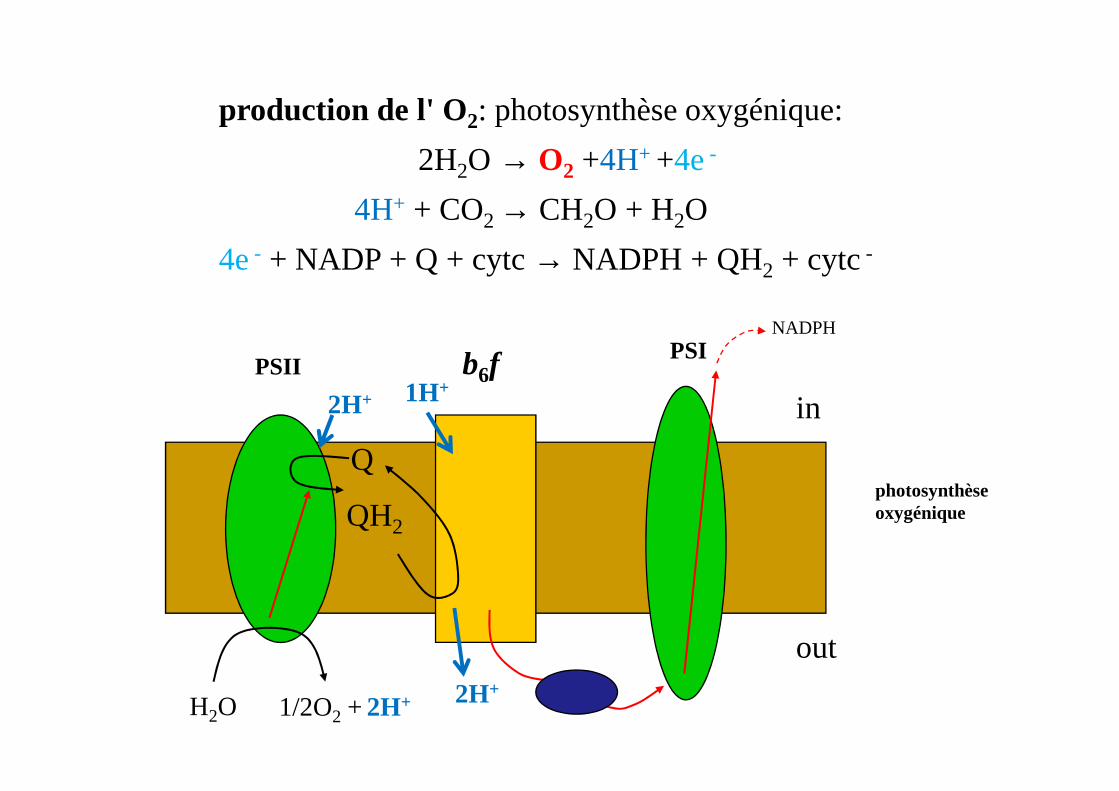
PSII b6fPSI
in
NADPH
2H+ 1H+
production de l' O2: photosynthèse oxygénique:
2H2O → O2 +4H+ +4e-
4H+ + CO2 → CH2O + H2O
4e- + NADP + Q + cytc→ NADPH + QH2 + cytc-
photosynthèse oxygénique
H2O 1/2O2 + 2H+
Q
QH2
out
in2H+
2H+
1H+

0
-200
-400
-600
-800
-1000
-1200
-1400
(m
V)
PSII Chl*/Chl+ (-1100 mV)
NAD(P)+/NAD(P)H(-320 mV)
PSI Chl*/Chl+ (-1250 mV)
ν
hν
1200
1000
800
600
400
200
0
Em (
mV
)
O2/H2O (+820 mV)
PSII Chl+/Chl (+1250 mV)
PQ/PQH2 (+100 mV)
PSI Chl+/Chl (+400 mV)
hν

l‘oxygène
génération de l'oxygène- 100%biologique- une seule source : l'eau- un seul mécanisme, un seul enzyme: le PSII

La chlorophylle
hν e-
Em -1100 mV Chl* est un réducteur fort
Em +1250 mVChl+ est un oxydant
puissant
e-
les propriétés rédox de la chlorophylle
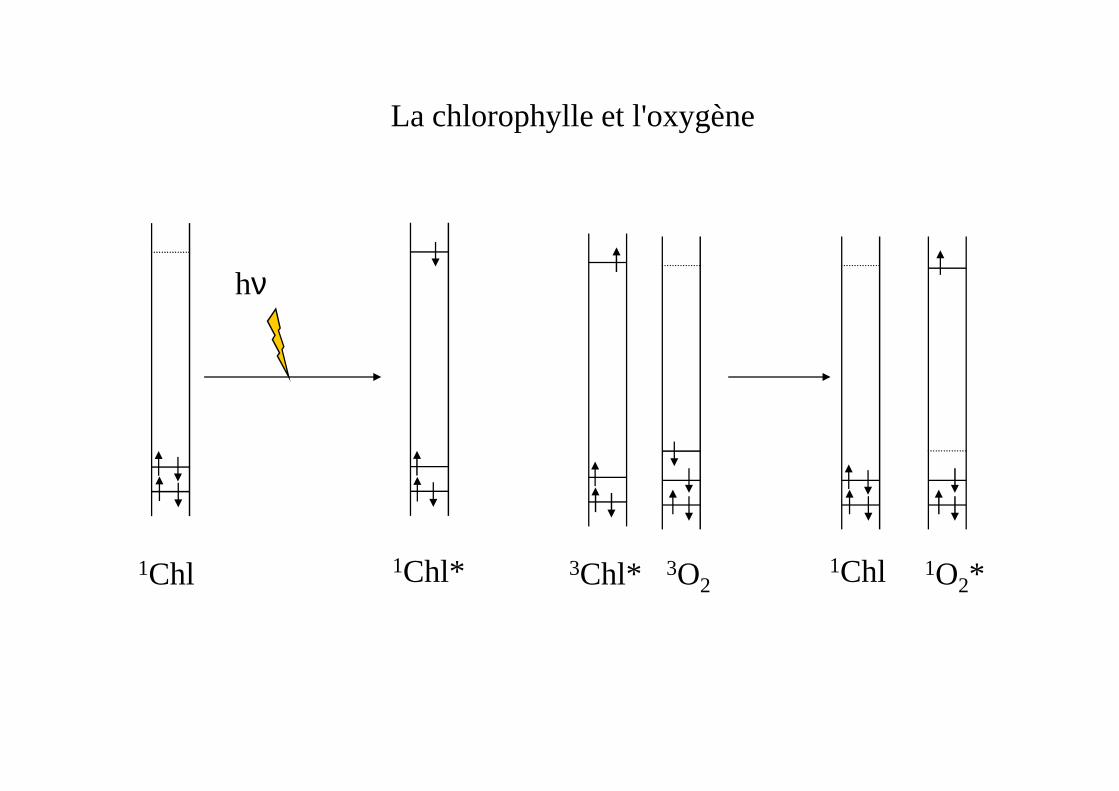
La chlorophylle et l'oxygène
hν
3Chl*1Chl*1Chl 1Chl3O21O2*

La chlorophylle et le carotenoïde
3Chl* 1Chl1Car 3Car* 3O23Car*

H2O ⇔ OH* + e- + H+ E° +2.31V
OH* + H2O ⇔ H2O2 + e- + H+ E° +380 mV
H2O2 ⇔ O2- + e- + 2H+ E° +890 mV
O2- ⇔ O2 + e- E° -330 mV
2H O ⇔ O +4e-+4H+ E =+815 mV2H2O ⇔ O2+4e-+4H+ E0 =+815 mV
O2 + 2 e- + 2H+ → H2O2 E° +281mV
H2O2 + 2 e- + 2H+ → 2H2O E° +1.35V

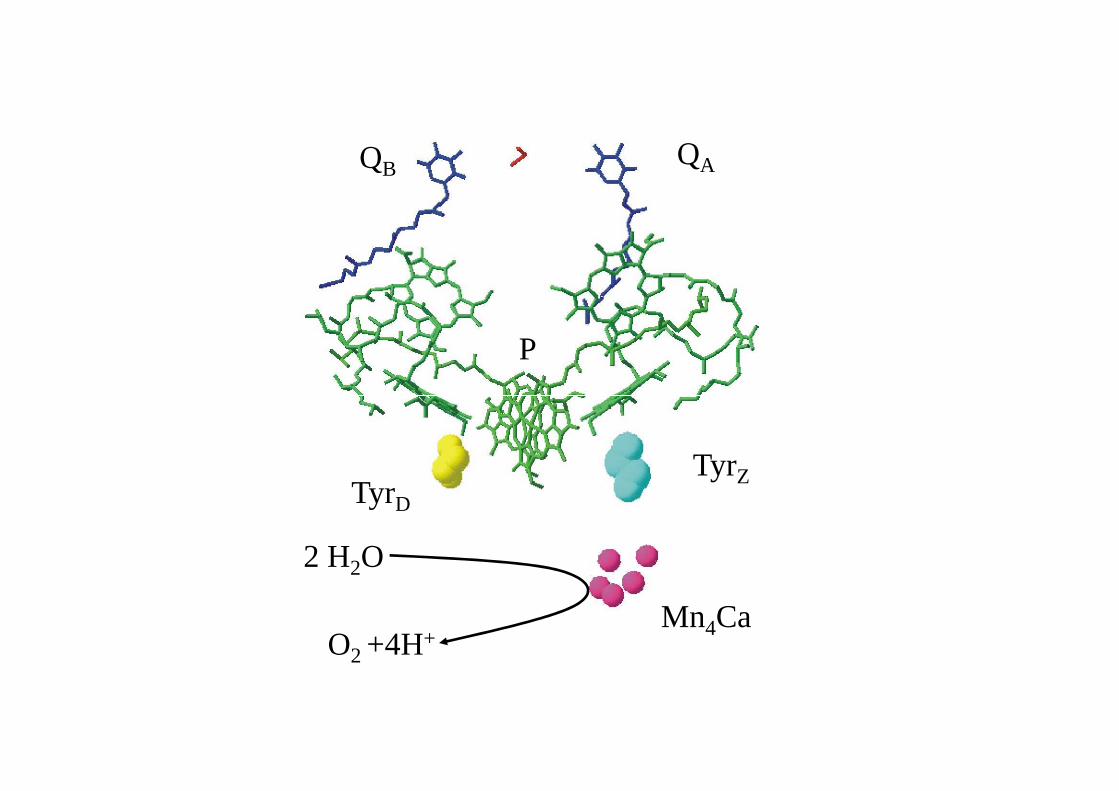
D2D1
D2

PSII RCI/PSIRCII
vue perpendiculaire à la membrane sur les hélices α des centres photosynthétiquesil y a deux fois cinq hélices qui fixent les cofacteursil y a six hélices additionnelles qui fixent des chlorophylles supplémentaires

La chlorophylle
hν e-
Em -1100 mV Chl* est un réducteur fort
Em +1250 mVChl+ est un oxydant
puissant
e-

QAQB
Fx
FA
FB
NAD(P)
QHQH2
PSII PSI/RCIRCII
Mn4Ca
TyrZTyrD
P
P
III
P
QH2
H2O
cytcPc, cytc

P
QAQB
Mn4Ca
TyrZTyrD
Crystal structure, Zouni et al.2001

Phe
QAQB
Phe
cytb
PQH2
PQ
P680
ChlChl
Tyr ZTyr D
Mn4ClCa
cytb559cytb559
2H2O O2 + 4H+ Rutherford 1989

co-facteurs du PSII

in
out
Structure du PSII de Thermosynechocossus elongatus à 3.2Å, 2003

Le mécanisme moléculaire des réactions rédox du photosystème II

enlèvement de 4H+ du H2O
expulsion de 4 H+
transfert de 4 e- vers la chaîne de transfert d'électrons
Production de l'oxygène
formation d'une liaison O-O
expulsion de l'O2
2H2O ↔ O2+4e-+4H+ E0 =+815 mV
pour oxyder l‘H2O il faut un oxydant puissant
La chlorophylle
hν e-
Em -1100 mV Chl* est un réducteur fort
Em +1250 mVChl+ est un oxydant
puissant
e-
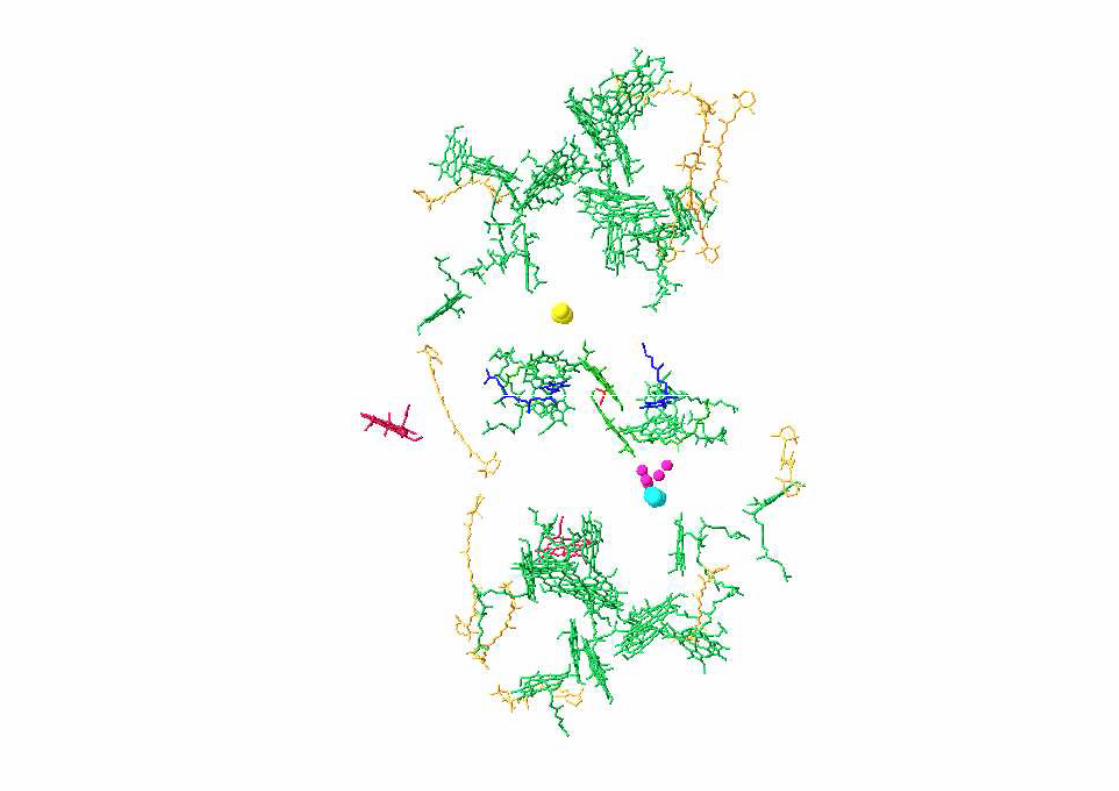
P
QAQB
Mn4Ca
TyrZTyrD
2 H2O
O2 +4H+

P
QAQB
Mn4Ca
TyrZTyrD
2 H2O
O2 +4H+


SO/S1
transfert d'e- en 30 µs
H2O
H2O
Mn4 (III, III, IV, IV)
*
H+

S1/S2
transfert d'e- en 100 µs
Mn4 (III, IV, IV, IV)
*
HO
HO
H+

S2/S3
transfert d'e- en 350 µs
HO
HO
Mn4 (III, IV, IV, IV) Tyr Z*Mn4 (IV, IV, IV, IV)
*
H+
-OH peut encore êtreéchangé avec D2O

S3/S4/S0transfert d'e- en 1300 µs
précédé par un transfert de protonsenviron 200 µs
formation de la liaison O-Odégagement de l'Oen 1300 µs
Mn4 (III, III, III, IV)
*dégagement de l'O2 en 1300 µs
O2
H+
4e-

H+
H+
e-
e-
1st photon
2nd photon
4th photon
So
S1
S2 350µs
100µs30µs
pH, H/D
H+
H+
e-
e- 3rd photon
proton-first
S3
S4
S4’
1.3ms
200µs
350µs
O2
pH, H/D
1ms

H2OCa
O2
Le cluster de manganèse dans la structure de 1.9A résolution (Umena et al. 2011)
Mn
O2

Structure at 1.9Å resolution 2011

Le cluster de manganèse dans la structure de 1.9A résolution (Umena et al. 2011)et son environnement protéique

P
QAQB
Mn4Ca
TyrZTyrD
2 H2O
O2 +4H+

Guskov et al. 2009
la recherche des canaux d’acheminement d’eau et d’évacuation d’oxygène est en cours
canaux putativesà protons
canauxputativesàH2O etO2

Gabdulkhakov et al. 2009

deux quinones et un canal bifurqué ont été trouvés dans la structure ils pourront assurer un échange rapide du quinol contre une quinone

Umena et al. 2011


Tout l'oxygène sur terre provient de la photosynthèse (PSII)
Presque toutes les formes de la vie multicellulaire obtiennent leur énergie de ces réactions de reduction de l'oxygène
La photosynthèse est présente uniquement dans le règne des Bactéries et chez leurs descendants, les chloroplasts.Elle n'était pas présente à l'origine de la vie

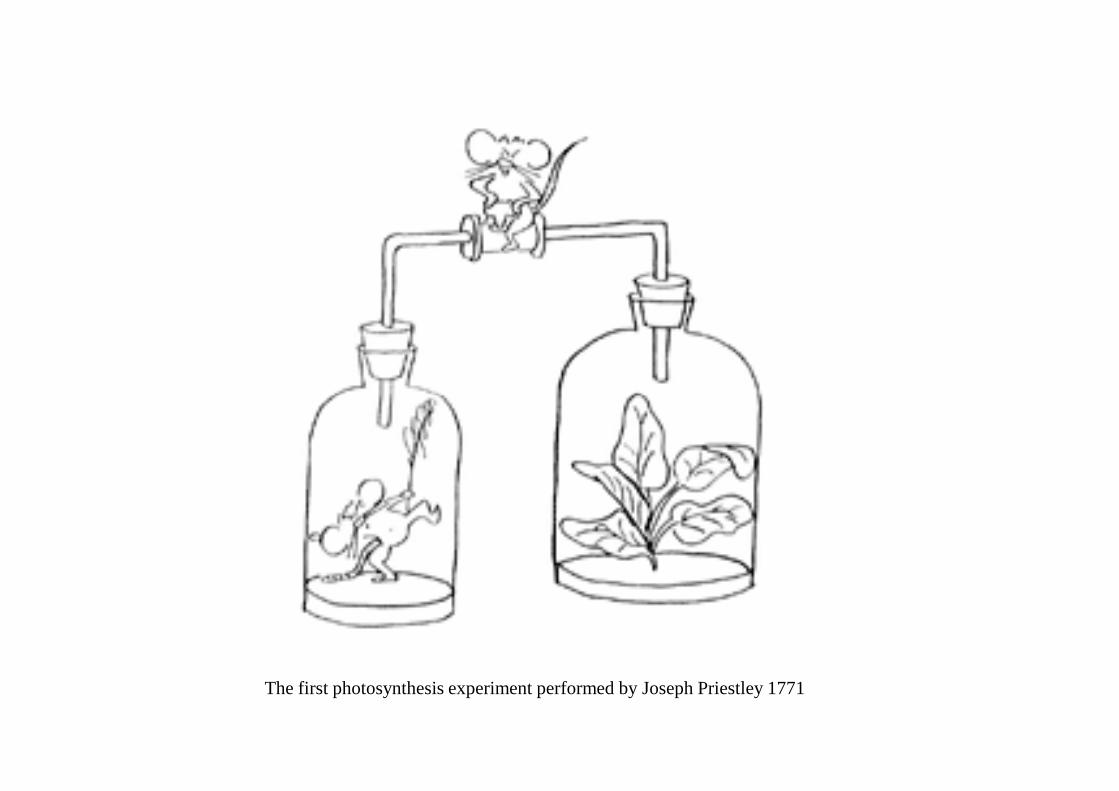
The first photosynthesis experiment performed by Joseph Priestley 1771

Les grands cycles de la matière
métabolisme de l'Oxygène
génération de l'oxygène- 100%biologique- une seule source : l'eau- un seul mécanisme, un seul enzyme: le PSII
consommation de l'oxygène- 90%par des réactions bioénergétiques
différents types d'oxydases

O2 + e- → O2- E° -330 mV flavins and semiquinones
O2- + e- + 2H+ → H2O2 E° +890 mVcatalysed by free Fe3+
H2O2 + e- + H+ → H2O + OH* E° +380 mVcatalysed by free Fe2+ and Cu+
OH* + e- + H+ → H2O E° +2.31V
O2 + 2 e- + 2H+ → H2O2 E° +281mV
H2O2 + 2 e- + 2H+ → 2H2O E° +1.35V
O2 + 4H+ + 4 e- → 2H2O E° +815 mV(for 1Atm=1.2mM)
le radical libre OH* peut demarrer
une réaction en chaîne de radicaux libres
=>oxygénation des protéines, de l'ADN et des lipides
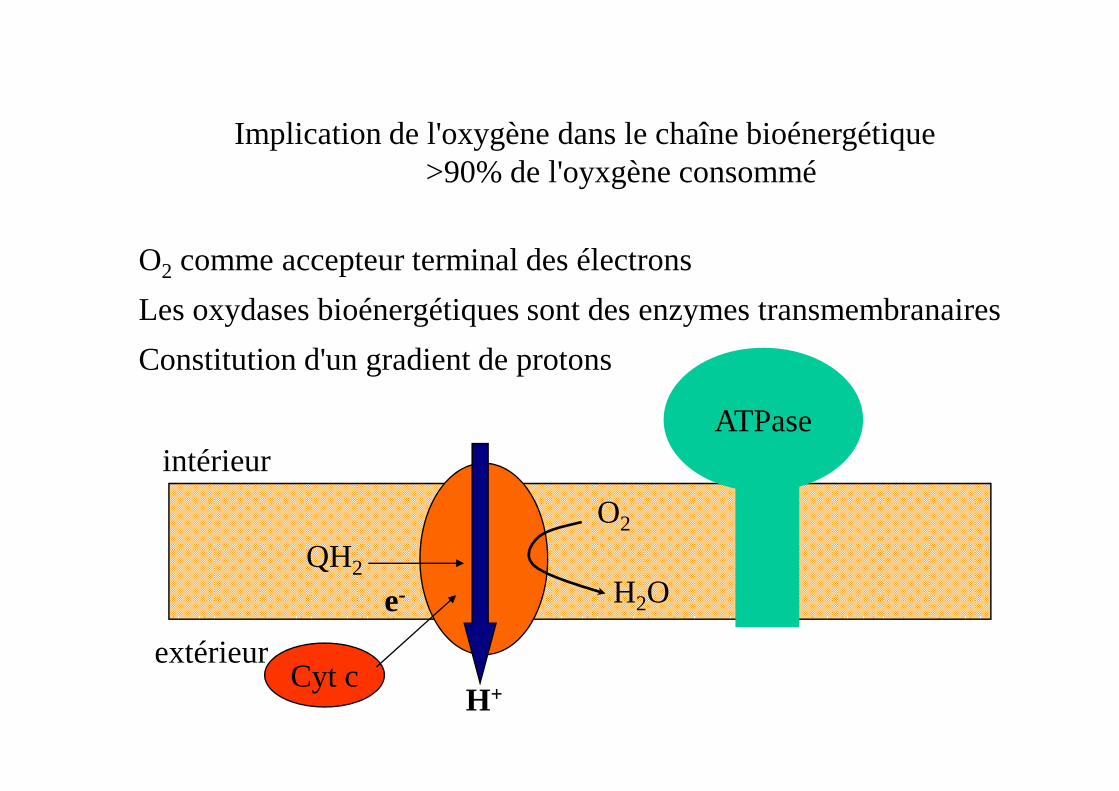
Implication de l'oxygène dans le chaîne bioénergétique>90% de l'oyxgène consommé
O2 comme accepteur terminal des électrons
Les oxydases bioénergétiques sont des enzymes transmembranaires
Constitution d'un gradient de protons
H2O
O2
extérieur
intérieurATPase
H+Cyt c
QH2
e-

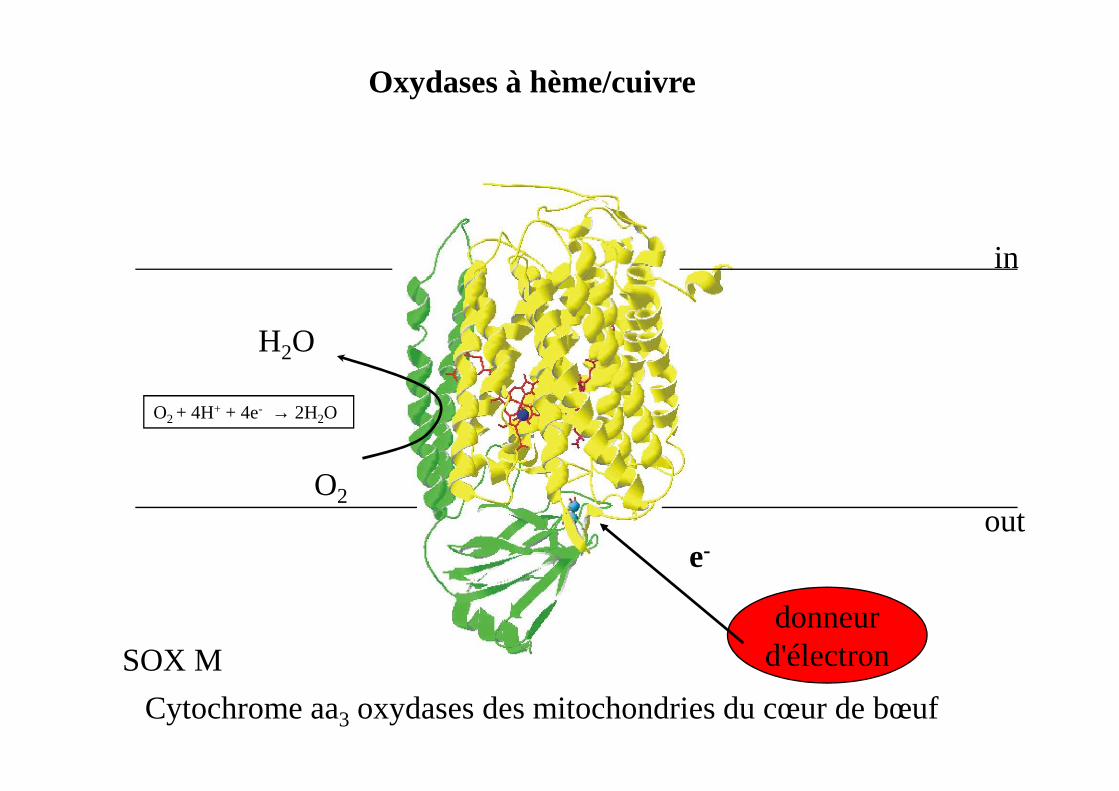
Oxydases à hème/cuivre
H2O
in
donneurd'électron
e-
O2
Cytochrome aa3 oxydases des mitochondries du cœur de bœuf
out
SOX M
O2 + 4H+ + 4e- → 2H2O

Le mécanisme moléculaire des réactions rédox
des oxydases à hème-cuivredes oxydases à hème-cuivre

90°
hème low spin
centre binucléairehème/cuivre

Les hèmes

- Cys - X - X - Cys -
Les hèmes /1
hème b hème c

CH3 OH
Les hèmes /2
OH
hème o →→→→ hème ahème b
hème d
hème bhèmeahèmeo hème d
O2

His Met His
Les hèmes /3
hème bas spin hème haut spin

Fe3+/4+
Les réactions se font au centre binucléaire: O2 + 4e - + 4H+ ⇔ 2 H2O
Cu1+/2+
FeFe2+/4+
l’hème et le cuivre peuvent mettre trois électrons à disposition pour la réaction

O2
Fe2+
Tyr-OH
Cu+
Fe2+
l’O2 se fixe au site actif complètement réduit

Fe4+
Tyr-O*
O2-
Cu2+
Fe4+ O2-
OH
les quatre électrons et le proton de la tyrosine sont transférés à l’O2
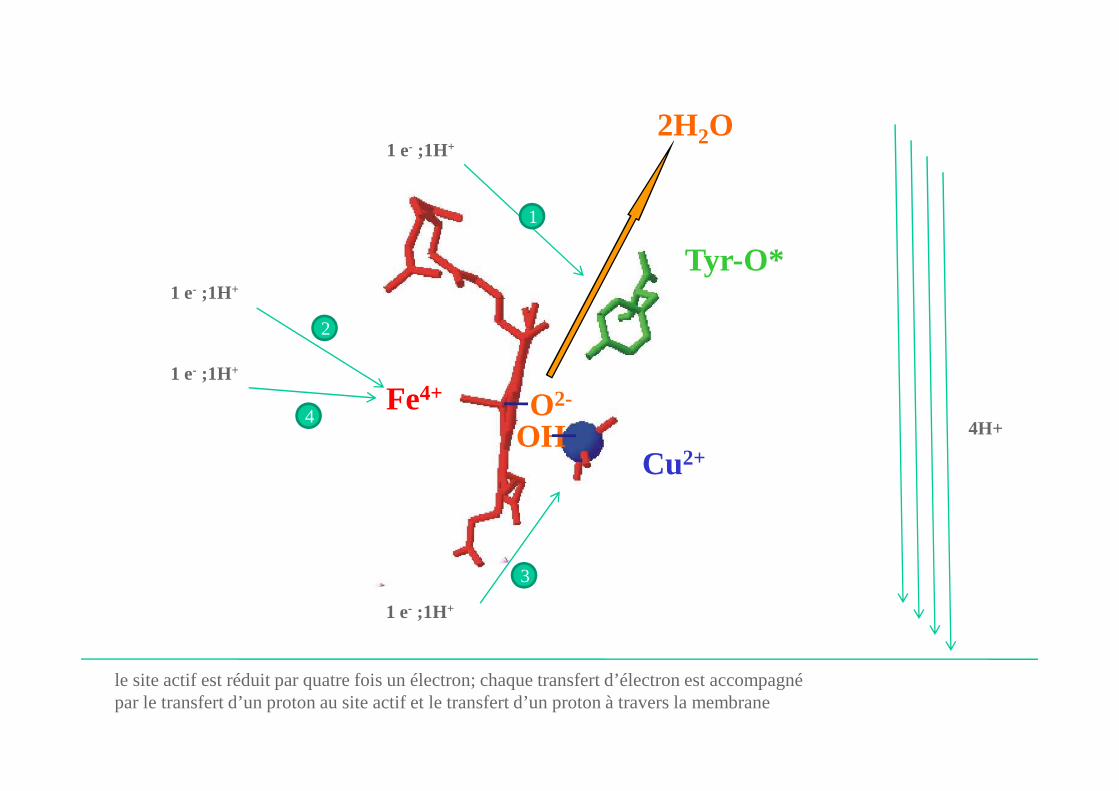
Fe4+
Tyr-O*
O2-
1 e- ;1H+
1 e- ;1H+
1 e- ;1H+
1
2
2H2O
Cu2+
Fe4+ O2-
OH
1 e- ;1H+
3
44H+
le site actif est réduit par quatre fois un électron; chaque transfert d’électron est accompagnépar le transfert d’un proton au site actif et le transfert d’un proton à travers la membrane
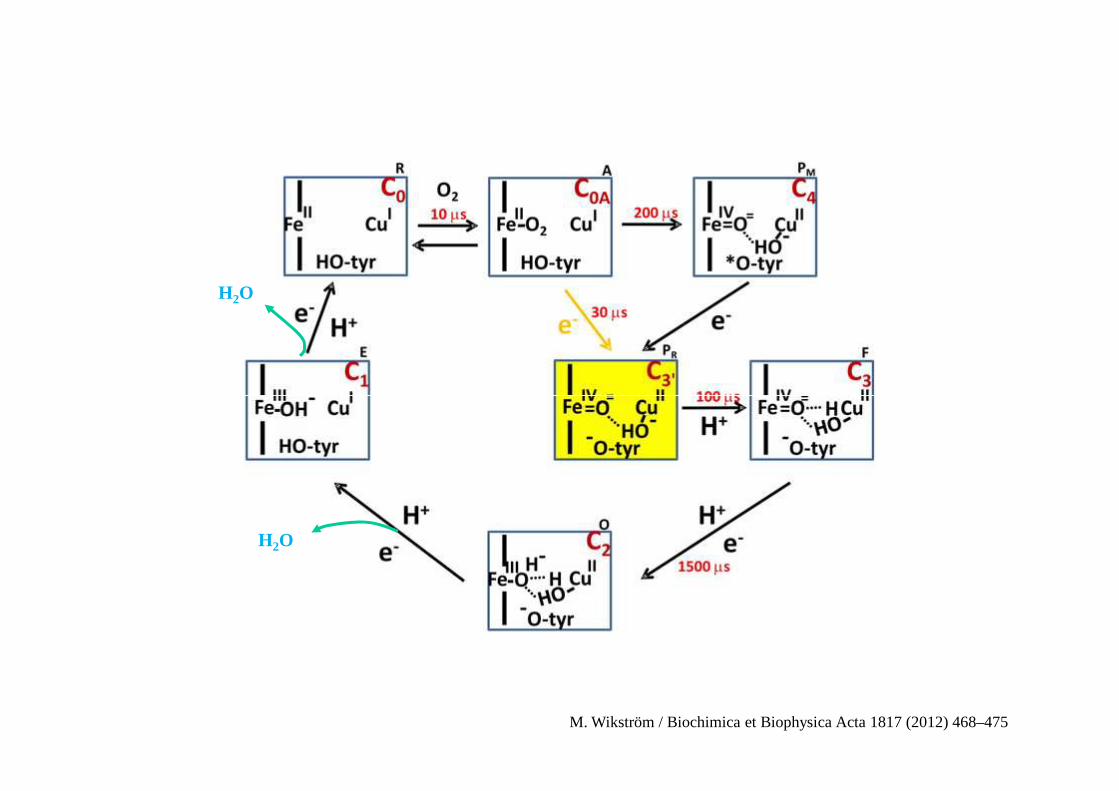
H2O
M. Wikström / Biochimica et Biophysica Acta 1817 (2012) 468–475
H2O

Chemins de transfert de protons de la cytochrome oxydase aa3 des mitochondries du cœur de bœuf
in
out

Glu Asp
incanal Dcanal K
Arg
Tyr
out
dans l’oxydase aa3 des bactéries pourpres deux canaux à proton ont été identifiés

in
H+
canal Dcanal K
out
pK=5
H

in
H+
canal Dcanal K
out
e-
pK=9
H
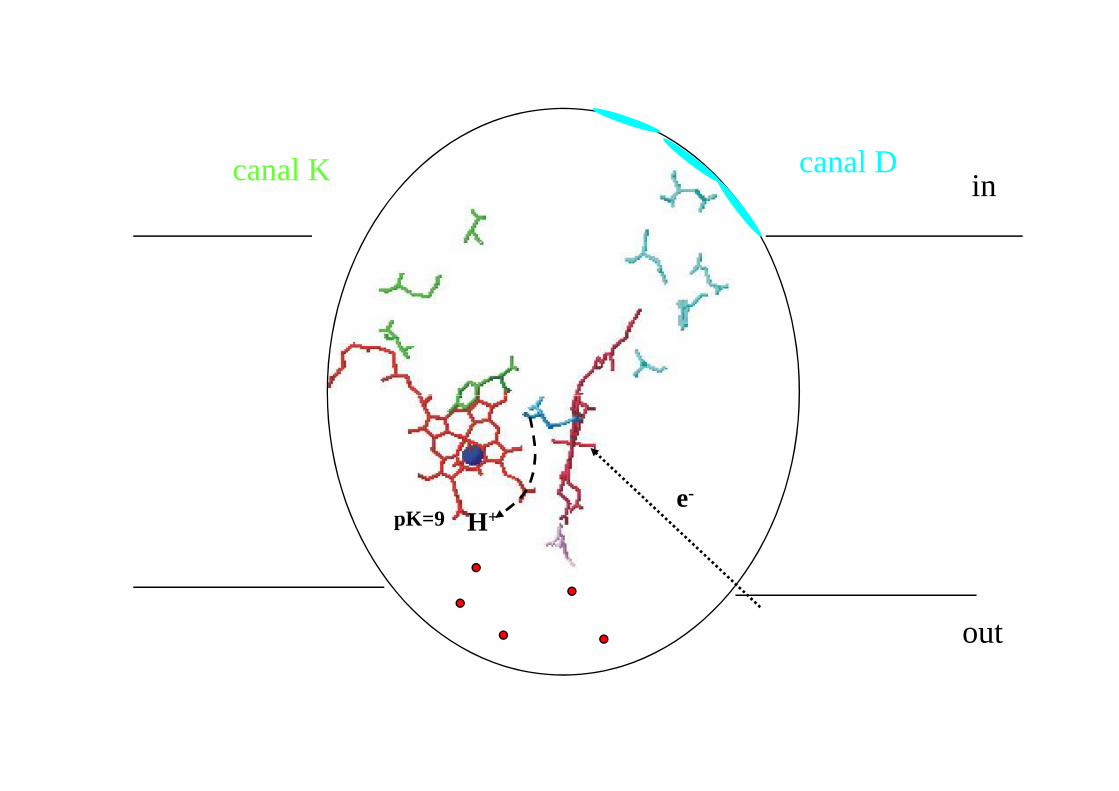
incanal Dcanal K
out
e-
pK=9 H+
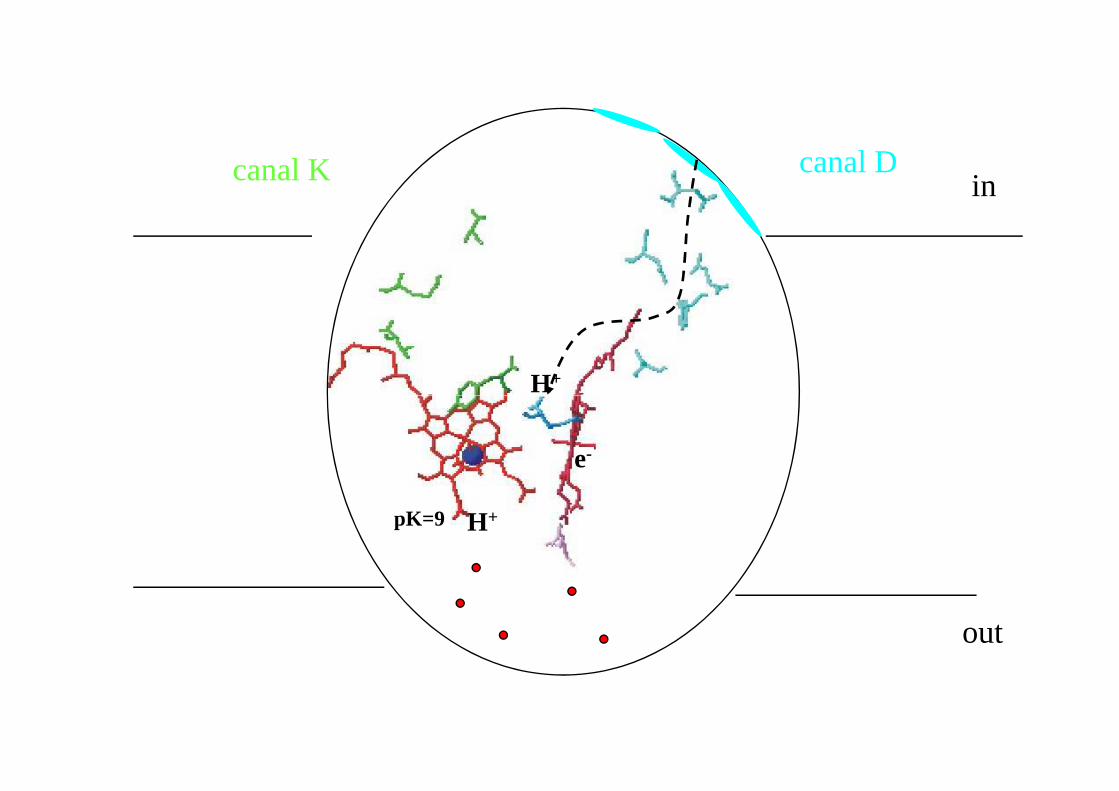
in
H+
canal Dcanal K
out
e-
pK=9 H+
H

in
H+
canal Dcanal K
out
e-
pK=11 H+
H

in
H+
canal Dcanal K
out
e-
pK=5 H+
H

in
H+
canal Dcanal K
out
e-
pK=5 H+

canal Dcanal K in
out
pK=5
H+

O2 + 4H+ + 4 e- → 2H2O E° +800 mV ([O2]=120 uM)
cyt c2+ + 1 e- → cyt c3+ Em =+300 mV ([cyt c2+]/[cyt c3+]=1)
4 cyt c2+ + 4H+ + O2 → 4 cyt c3+ +2 H2O : +2000 mV
Quelques chiffres
Potentiel transmembranaire dans des mitochondries actives:+170mV à +220 mV
Transfert de 8H+ :1360 à1760 mV
=> efficacité de 70 à >80%
Transfert de 1500e-/sec => ca 2200H+/sec par le canal D

M. Radzi Noor, T. Soulimane / Biochimica et Biophysica Acta 1817 (2012) 638–649

in
O2
oxydase ba3 de Thermus thermophilus
out
O2Xe
H2O
des molécules de Xénon marquent une cavité par laquelle l’O2 peut accéder au site actif

in
oxydase aa3 de Paracoccus denitrificans
out
l’arrivé d’un canal au site actif est rétrécie par la présence d’une Phe et une Trp
Oxydases à hème/cuivre
H2O
in
donneurd'électron
e-
O2
Cytochrome aa3 oxydases des mitochondries du cœur de bœuf
out
SOX M
O2 + 4H+ + 4e- → 2H2O

Oxydases à hème/cuivre
H2O
in
Cytochrome caa3 oxydase (SOX M) de Thermus thermophilusSoulimaneet al. 2012
donneurd'électron
e-
O2out

e-
H2O
QH
in
e
O2
QH2
quinole oxydase d'Escherichia coli
out
SOX M
O2 + 4H+ + 4e- → 2H2O

in
O2 + 4H+ + 4e- → 2H2O
H2O
Cytochrome ba3 oxydase de Thermus thermophilus
outdonneurd'électrone-
SOX B
O2
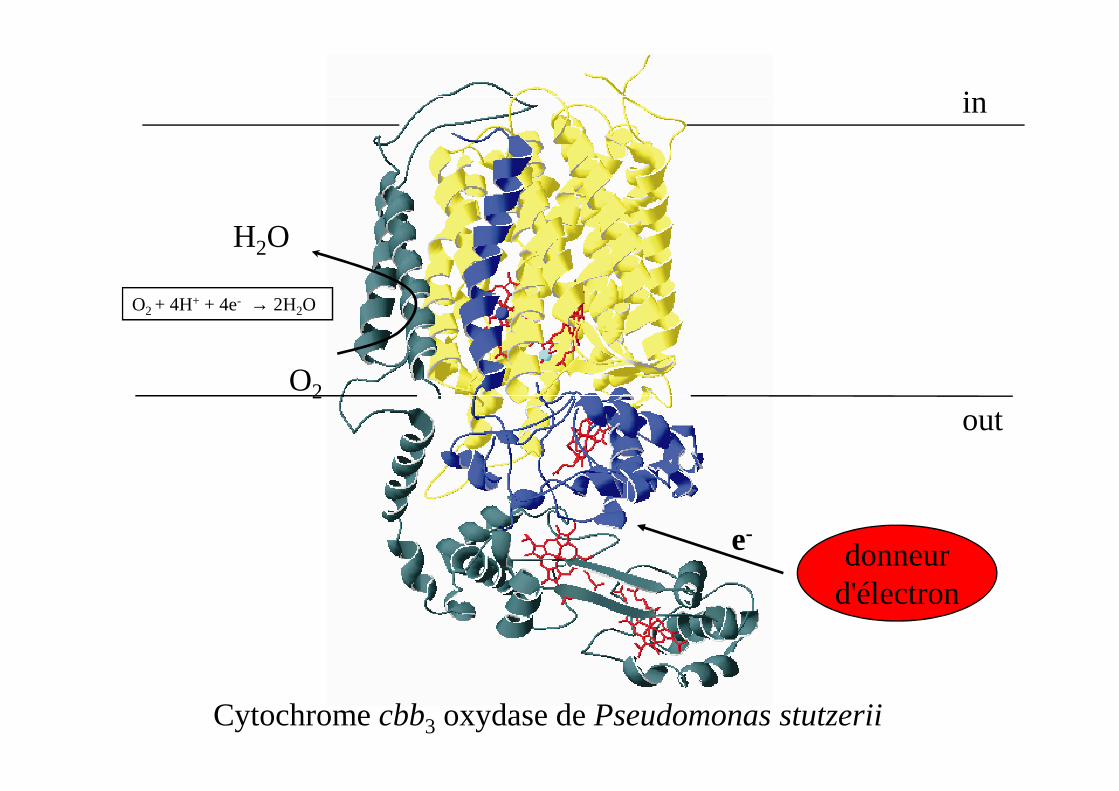
in
O2 + 4H+ + 4e- → 2H2O
H2O
O2
Cytochrome cbb3 oxydase de Pseudomonas stutzerii
out
donneurd'électron
e-
O2

SoxB/M
cbb3
le quatrième électron qui est nécessaire pour réduire l’O2 vient d’une tyrosine à proximité

in
2NO + 2H+ + 2e- → N2 + H2O
N2 +H2O
2NO
Cytochrome NO-réductase de Pseudomonas aeruginosa
out
donneurd'électron
e-
2NO
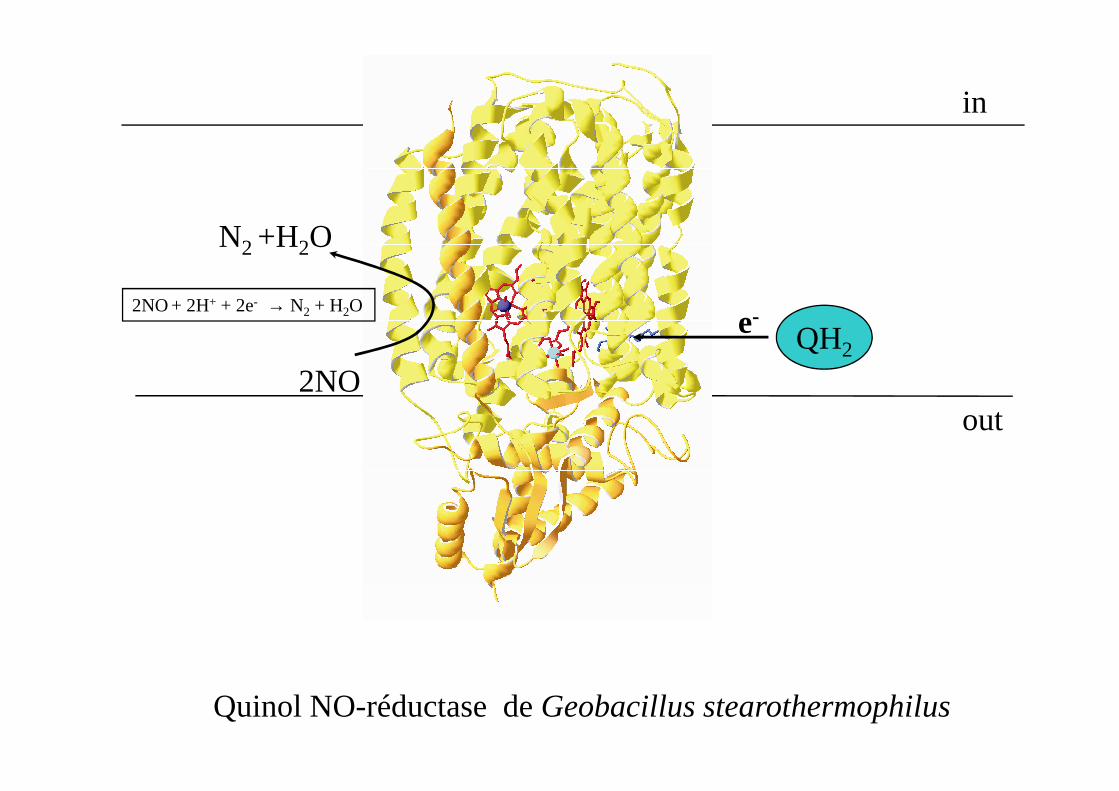
in
2NO + 2H+ + 2e- → N2 + H2O
N2 +H2O
2NO
e-QH2
Quinol NO-réductase de Geobacillus stearothermophilus
out2NO

NO-reductase O2-reductase
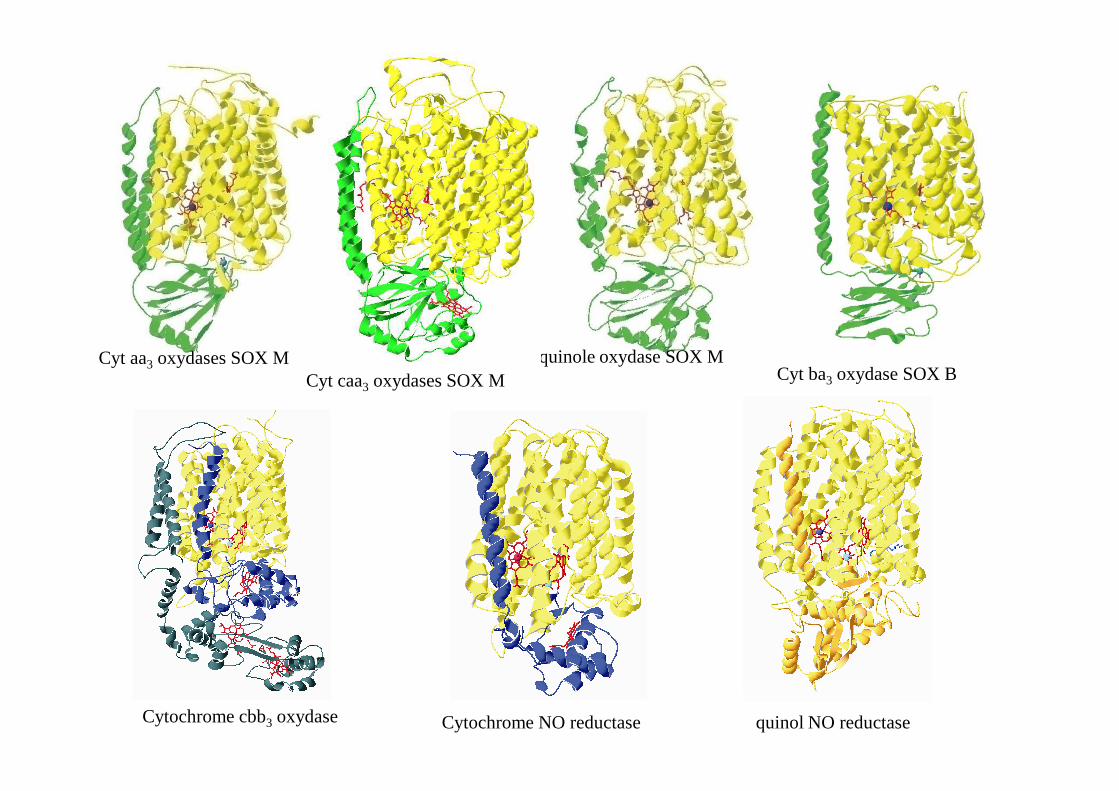
Cyt aa3 oxydases SOX M quinole oxydase SOX MCyt ba3 oxydase SOX BCyt caa3 oxydases SOX M
Cytochrome cbb3 oxydase Cytochrome NO reductase quinol NO reductase
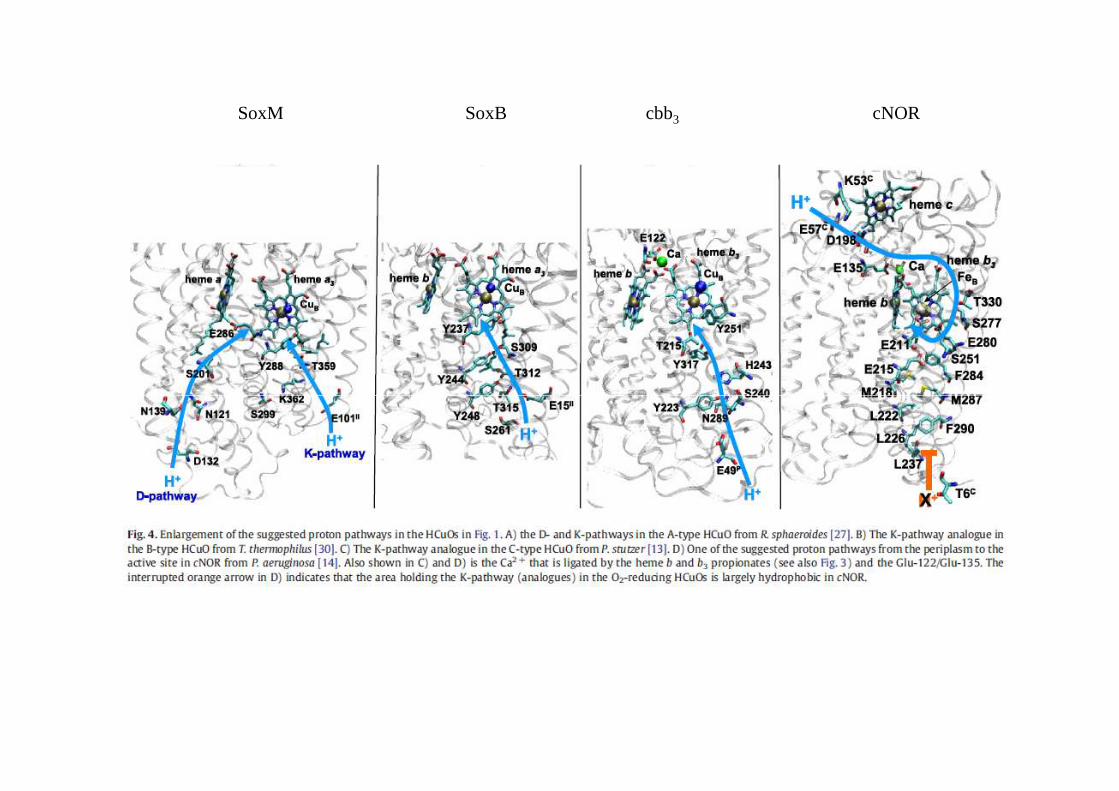
SoxM SoxB cbb3 cNOR

bd-oxydase
in
b595dQH
H+
=> rôle dans la transition anaerobie/aerobie?O2 comme accepteur d'électrons des bactéries 'nanoaerobes'
outb558
b595dQH2
Q
H+
O2
H2O

type bd SoxM / A-type SoxB / b-type cbb3 / C- cNor qNor
substrateaffinity*
B H B B B/H B
proton channels
D et K K K -/K K
H+/e- nd 2 1-2 1 0/nd nd
Subunits A, B I, II, III, IV I, II I(N), O, P/R I, O I (=I+O)
SUII 8TM helices 2TM helices+Cu domain(+cytc)
1TM helix+ Cu domain
1TM helix+cytc
1TM helix+cytc
Domain: 2TM helices+cytc fold(+cytc) +cytc fold
co-factors I:2 hèmes HSHème LS
I:CuHème HSHème LSTyrII: 2Cu(hème)
I: CuHème HSHème LSTyr
I: CuHème HSHème LSTyrII: 2Cu
I: CuHème HSHème LSTyr
I: CuHème HSHème LSTyrO: 1 cytcP: 2 cytcR: 1 cytc
I: FeHème HSHème LS
O: 1 cytc
FeHème HSHème LS
substrate O2 O2 O2 O2 NO NO
e-- domors Q Cyt c Q Cyt c Q Cyt c Cyt c Q
* B: basse affinité (5nm-100nm); H: haute affinité (uM)


b
a oair-sufficient cells
Bacillus PS3
air-sufficient cells
air-limited cells

air-sufficient cells
550 nm: hèmes c560 nm: hèmes b604 nm: hèmes aair-limited cells 604 nm: hèmes a

highly aerated cells
slightly air-limited cells
a o
a o
two protein preparations that showidentical subunit compositions


L'évolution du métabolisme de l'oxygène
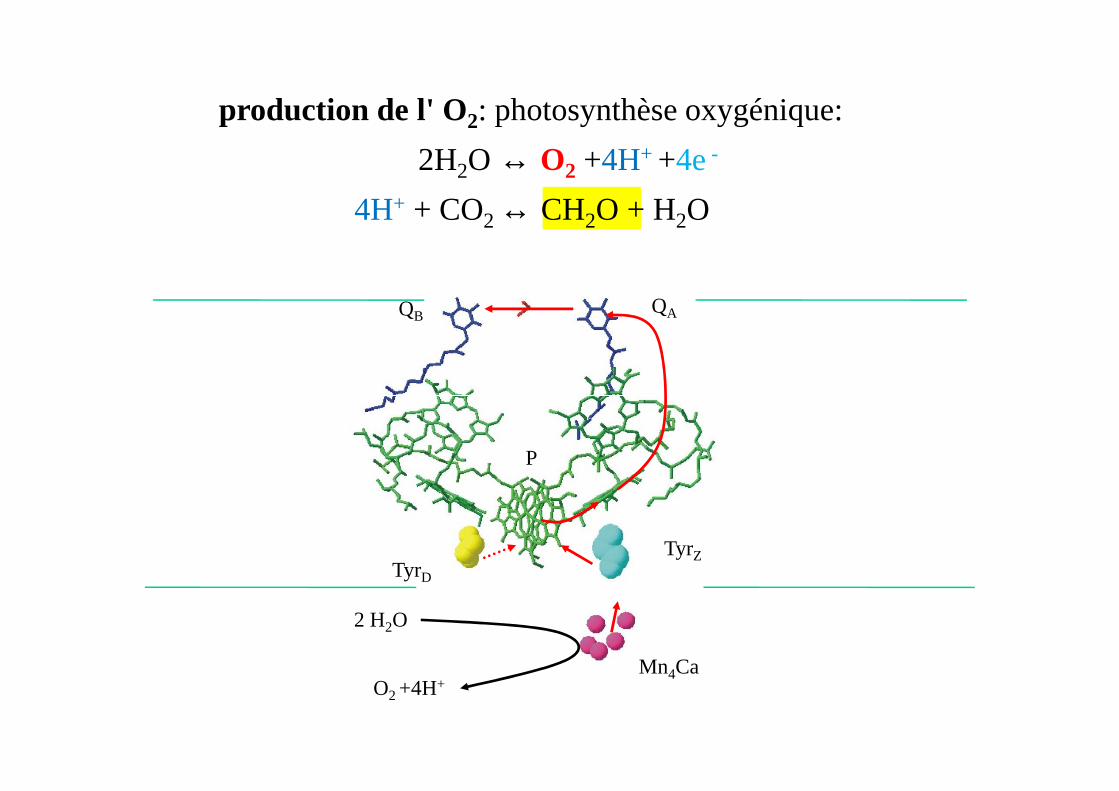
production de l' O2: photosynthèse oxygénique:
2H2O ↔ O2 +4H+ +4e-
4H+ + CO2 ↔ CH2O + H2O
QAQB
Mn4Ca
TyrZTyrD
P
2 H2O
O2 +4H+

Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue

Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
mic
rob
ial
foss
ils
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
carb
on
fix
ati
on
bio
log
ica
l

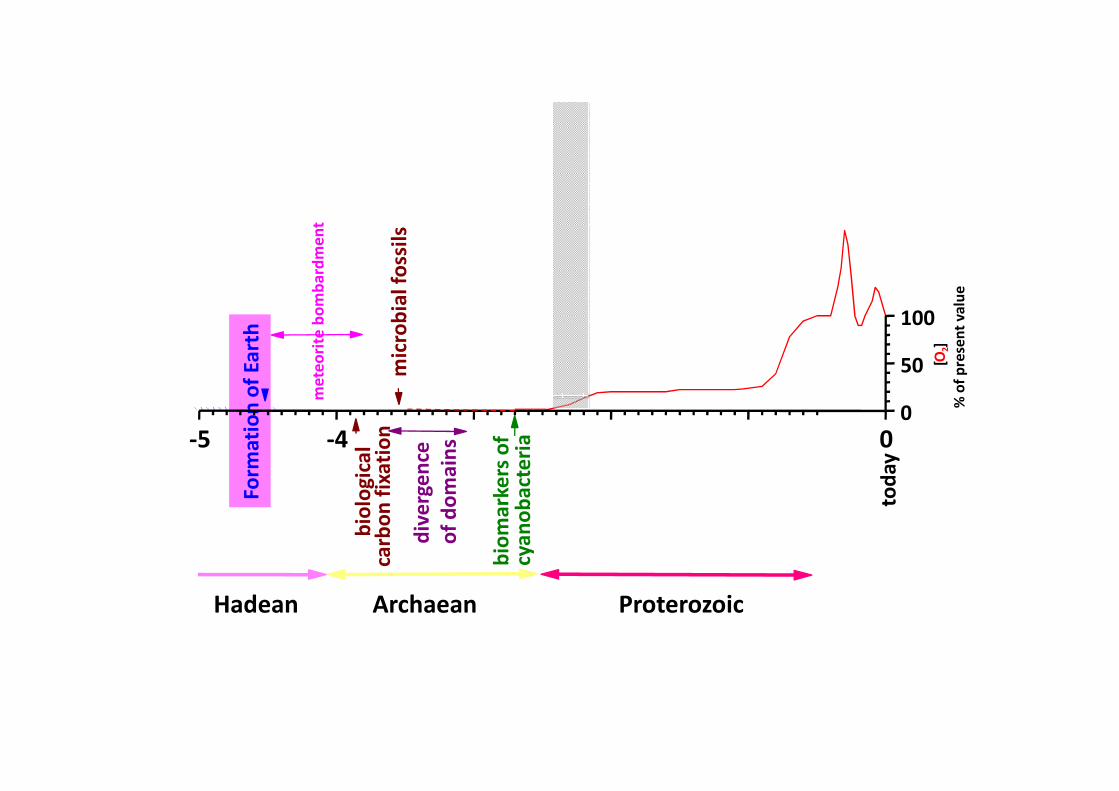
dans l'Archaean 20% N2, 80% CO2 4-5 atm
2.2*109 yrs CH4 peut-être500 fois plus élévé
qu'aujourd'hui
traces d'oxygène et de peroxyde d'hydrogène par la photolyse de l'eautraces d'oxygène et de peroxyde d'hydrogène par la photolyse de l'eau
présence de NO par volcanisme et réaction de NO avec CO2 (hν)
depuis l'origine de la vie la substance organique est immobilisée

Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
mic
rob
ial
foss
ils
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
carb
on
fix
ati
on
bio
log
ica
l
div
erg
en
ce
of
do
ma
ins
bio
ma
rke
rs o
fcy
an
ob
act
eri
a
Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
mic
rob
ial
foss
ils
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
carb
on
fix
ati
on
bio
log
ica
l
div
erg
en
ce
of
do
ma
ins
bio
ma
rke
rs o
fcy
an
ob
act
eri
a

Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
mic
rob
ial
foss
ils
abundant stromatolites
cyanobacteria
dominate the
biosphere
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
carb
on
fix
ati
on
bio
log
ica
l
div
erg
en
ce
of
do
ma
ins
bio
ma
rke
rs o
fcy
an
ob
act
eri
a

stromatholites
fossilisés
vivants

Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
mic
rob
ial
foss
ils
abundant stromatolites
cyanobacteria
dominate the
biosphere
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
carb
on
fix
ati
on
bio
log
ica
l
div
erg
en
ce
of
do
ma
ins
bio
ma
rke
rs o
fcy
an
ob
act
eri
a

Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
mic
rob
ial
foss
ils
abundant stromatolites
cyanobacteria
dominate the
biosphere
lan
dp
lan
ts
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
carb
on
fix
ati
on
bio
log
ica
l
div
erg
en
ce
of
do
ma
ins
bio
ma
rke
rs o
fcy
an
ob
act
eri
a
alg
ae

Form
ati
on
of
Ea
rth
50
100
[O2]
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
mic
rob
ial
foss
ils
abundant stromatolites
cyanobacteria
dominate the
biosphere
lan
dp
lan
ts
coa
l fo
rest
s
ProterozoicArchaeanHadean
Form
ati
on
of
Ea
rth
-5 -4 00
tod
ay
% o
f p
rese
nt
va
lue
me
teo
rite
bo
mb
ard
me
nt
carb
on
fix
ati
on
bio
log
ica
l
div
erg
en
ce
of
do
ma
ins
bio
ma
rke
rs o
fcy
an
ob
act
eri
a
alg
ae

aujourd'hui 99.9% de la matière organique recombine
par la respiration ou se décompose avec l'O2,
moins de CH4, présence d'O2 et d'une couche d'O3 ,
trés peu d'échappement d'H dans l'espace
H2O est piégé dans le froid de l'atmosphère
l'atmosphère d'aujourd'huil'atmosphère d'aujourd'hui
78% N2, 20.9% O2, ca 4% H2O, 0.9% Ar, 0.034% CO2
carbone organique immobilisé dans les sédiments et les roches
(26000 fois le carbone du vivant, la plupart dans le grès)
et il y a toujours de l'eau sur terre!
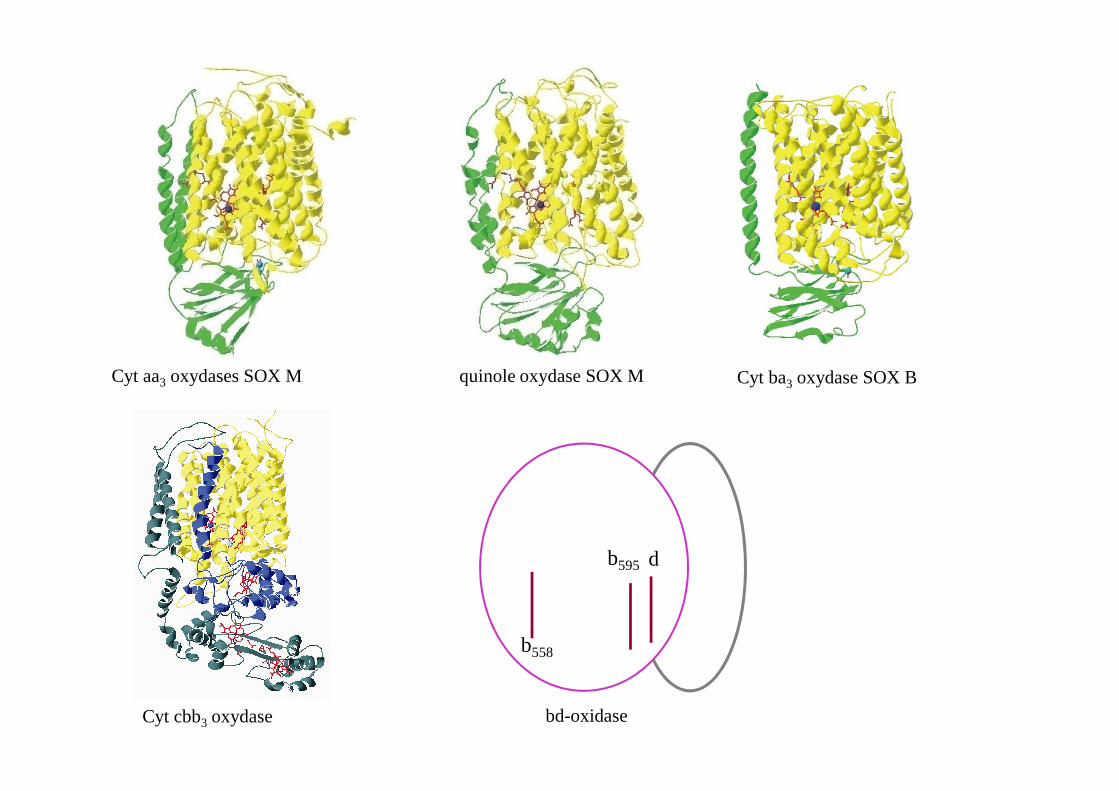
Cyt aa3 oxydases SOX M quinole oxydase SOX M Cyt ba3 oxydase SOX B
Cyt cbb3 oxydase bd-oxidase
b558
b595 d

un peu de phylogénie

CLUSTAL X (1.8) multiple sequence alignment
dt --MEINFREUNDSA-GT-ANNAICH----LIEBE-DICHnd --MIJNFRIENDZE-GT-ANNAIK-----HOUVAN-JE-eg --MY--FRIENDSA-YS-ANNAI------LOVE--YOU-dn --MIN-V--EN-SI-GERANNAJEG---ELSKER-DIG-sv --MIN-V-AEN-SAEGERANNAJAG--AELSKAR-DIG-
Comparer les séquences
sv --MIN-V-AEN-SAEGERANNAJAG--AELSKAR-DIG-fr --MON-AMI---DI--T-ANNAJE-T-A---IME-----cat LAMEVAAMIGA-DIU---ANNA---T--ESTIMO-----es --MI--AMIGO-DI-CE-ANNAYO-TEA----MO-----it --MIO-AMIGO-DI-CE-ANNA---TIA----MO-----oc --MON-AMIGA-DI-TZ-ANNA---T-A---IMI-----
* . ****
ug ABARATOMAZTMONDJASZERTLEKANNA

fr
oc
774
cat
esit
835743
Calculer un arbre phylogénique
Bactéria
0.1
eg
dt
nd
518
694
dn
sv 980
989 Archaea

fr
oc
774
cat
esit
835743
Trouver la racine d'un arbre = déduire l'histoire évolutive
Bactéria
0.1
eg
dt
nd
518
694
dn
sv 980
989 Archaea

"maman a du fenouil
la tante doit du foin"
multiples versions d'une protéine
étaient présent dans l'ancêtre commun
des organismes étudiés

1994 Castresana J, Lübben M, Saraste M, Higgins DG.
‘Evolution of cytochrome oxidase, an enzyme older than atmospheric oxygen’
1994 Saraste M, Castresana J
‘Cytochrome oxidase evolved by tinkering with denitrification enzymes’‘Cytochrome oxidase evolved by tinkering with denitrification enzymes’

Cyt aa3 oxydases SOX M quinole oxydase SOX M Cyt ba3 oxydase SOX B
Cyt cbb3 oxydase bd-oxidase
b558
b595 d

Cyt aa3 oxydases SOX M quinole oxydase SOX MCyt ba3 oxydase SOX BCyt caa3 oxydases SOX M
Cytochrome cbb3 oxydase Cytochrome NO reductase quinol NO reductase
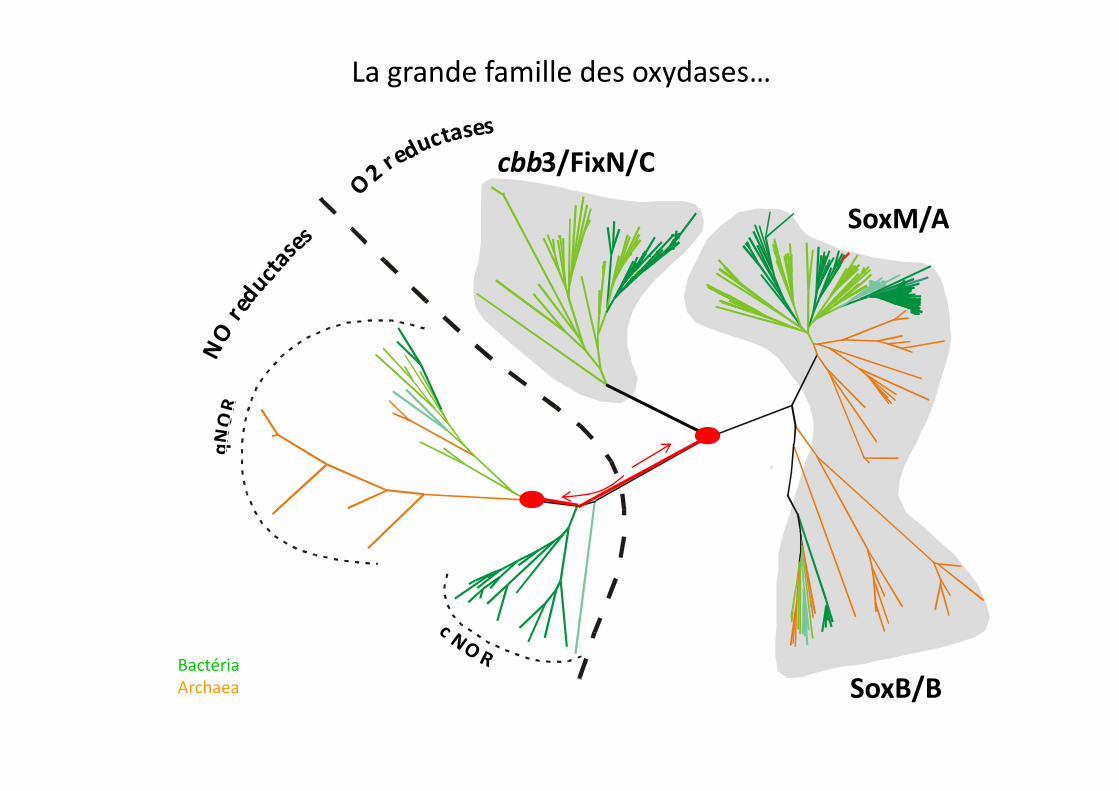
La grande famille des oxydases…
cbb3/FixN/C
SoxM/A
SoxB/BBactéria
Archaea


CO2 /CH3CO2
-
CO2/CH4
H2S/SO4
2-
H2 /H+
low Emquinones
early Archaeanpresent
HS - /SO4
2-
H2 / H+CO2 /
CH3CO2-
CO2/CH4
low Emquinones
organicsubstrates
hυυυυ
-600
-800
-1000
-400
-200
0
?
As3+/As5+
As3+/As5+
high Emquinones
O2/H2O
+1000
+800
+600
+400
+200
0

CO2 /CH3CO2
-
CO2/CH4
H2S/SO4
2-
H2 /H+
low Emquinones
early Archaeanpresent
As3+/As5+As3+/
HS - /SO4
2-
H2 / H+CO2 /
CH3CO2-
CO2/CH4
low Emquinones
high Emquinones
organicsubstrates
hυυυυ
-600
-800
-1000
-400
-200
0
?
AsAs /As5+quinones
O2/H2O+1000
+800
+600
+400
+200 NO 2-/
NO
NO3- /
NO2-
NO/N2O
N2O/N2+1200
NO/N2O
NO 2-/
NO
NO3- /
NO2-
NO/N2O
N2O/N2

Quinone 'bas-potentiel':
Ménaquinone
Présentes chez la plusparts de phyla des Archaebactéries et des Bactéries étudiées
Quinones 'haut poteniel':Présentes chez quelques phyla archébactériens et bactériens répandus aujourd'hui:
Ubiquinonechez le protéobactéries (et les mitochiondries)
PlastoquinonePlastoquinonechez les cyanobactéries (et les chloroplasts)
Caldarinellaquinonechez les Sulfolobales
Ces quinones 'haut-potentiel' sont apparus plusieurs fois de façon indépendantes dans l'évolution
en réponse de l'organisme à une augmentation du potentiel ambiant
suite à l'accumulation de l'oxygène sur terre

GSB
Hélio
cyanos
RCI
RCI
PSI+PSII
pourpres
RCII
Chloroflexaceae
RCII
Ancêtre commun des êtres vivants
Archaebactéries
Bactéries
RCIPSI+PSII

GSB
Hélio
cyanos
RCI
RCI
PSI+PSII
pourpres
RCII
Chloroflexaceae
RCII
Ancêtre commun des êtres vivants
Archaebactéries
Bactéries
RCIPSI+PSII
MK

GSB
Hélio
cyanos
RCI
RCI
PSI+PSII
pourpres
RCII
Chloroflexaceae
RCII
UQ
Ancêtre commun des êtres vivants
Archaebactéries
Bactéries
RCIPSI+PSII
PQCQMK

CO2 /CH3CO2
-
CO2/CH4
H2S/SO4
2-
H2 /H+
low Emquinones
early Archaeanpresent
As3+/As5+
As3+/
HS - /SO4
2-
H2 / H+CO2 /
CH3CO2-
CO2/CH4
low Emquinones
high Emquinones
organicsubstrates
hυυυυ
-600
-800
-1000
-400
-200
0
?
As5+As /As5+quinones
O2/H2O+1000
+800
+600
+400
+200 NO /NO2
-
NO2- /
NO3-
NO/N2O
N2O/N2+1200
NO/N2O

Le diapanorama sous format pdf se trouve
sur notre site web:
http://bip.cnrs-mrs.fr/bip09/http://bip.cnrs-mrs.fr/bip09/
dans la rubrique ‘Teaching’









![SERVIS KVM KVMエクステンダー USBデバイス延長モデル ...[FE-1600CWU、FE-3100CXU、FE-4000CXU、FE-4500CXU] 1. コンソールのみを延長する場合 ③送信ユニット(LOCAL)ではなくPCに接続する機種](https://static.fdocuments.fr/doc/165x107/60c7344248c2b568c26f13a5/servis-kvm-kvmffff-usbffefff-fe-1600cwufe-3100cxufe-4000cxufe-4500cxu.jpg)








