Cours 5 Régulation de la glycémie, diabète et cétogenèse
16
Page 1 sur 16 Ronéo 3 – Cours 5 UE8 UE8 : Nutrition Pr Cavé Lundi 9 Octobre 2017 de 13h30 à 15h30 Ronéotypeur : Camille PAVEC Ronéoficheur : Cécile PAUCHET Cours 5 – Régulation de la glycémie, diabète et cétogenèse La professeure a indiqué qu’il était très important de connaître et de comprendre l’étiologie du diabète de type II.
Transcript of Cours 5 Régulation de la glycémie, diabète et cétogenèse

Page 1 sur 16 Ronéo 3 – Cours 5 UE8
UE8 : Nutrition
Pr Cavé
Lundi 9 Octobre 2017 de 13h30 à 15h30
Ronéotypeur : Camille PAVEC
Ronéoficheur : Cécile PAUCHET
Cours 5 – Régulation de la glycémie, diabète
et cétogenèse
La professeure a indiqué qu’il était très important de connaître et de comprendre l’étiologie du
diabète de type II.

Page 2 sur 16 Ronéo 3 – Cours 5 UE8
SOMMAIRE
I – Rappels sur la glycémie
II – Système digestif et régulation de la glycémie
A) Les incrétines
B) Synthèse du glucagon et des peptides dérivés comme le GLP-1
C) Mécanismes d’action de la GLP-1 dans l’organisme
D) Régulation de la GLP-1
III- Pathologies de la régulation de la glycémie
A) Le diabète
1) Généralités
2) Le diabète des enfants
B) Le diabète de type I
1) Altération du métabolisme glucidique
2) Altération du métabolisme lipidique
C) Diabète monogénique
D) Diabète de type II
1) Epidémiologie
2) Le syndrome métabolique
3) Génétique du diabète de type II
E) Complications du diabète à long terme
F) Liens entre obésité, diabète et cancers

Page 3 sur 16 Ronéo 3 – Cours 5 UE8
I – Rappels sur la glycémie
Il est important de rappeler que les valeurs de glycémie normale sont :
- A jeûn 0,7 à 1,1g/L soit 4 à 6 mmol/L
- En post prandial (2h) inférieur à 1,4g/L soit 8mmol/L
N.B. : 1g/L = 5,5 mmol /L
La glycémie doit être maintenue dans un certain intervalle à tout moment car :
- le glucose est utilisé pour produire de l’ATP ou comme substrat de biosynthèses ;
- il est la source exclusive ou quasi-exclusive de certains tissus tels que le système nerveux
central (corps cétoniques utilisables) ou les globules rouges et la source d’énergie
d’urgence des tissus ;
- l’excès ou le défaut de glucose entraîne des effets délétères sur les tissus périphériques.
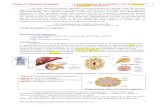
Dans le cours précédent (UE1 Cours n°3), nous avons vu que le foie et le pancréas sont des
organes majeurs de la régulation de la glycémie. Le foie pour son activité métabolique et le pancréas
pour son rôle de synthèse hormonale. Apparait alors un 3ème
organe dont on connait maintenant
l’importance dans la régulation de la glycémie : le système digestif.
II – Système digestif et régulation de la glycémie
A) Les incrétines
En post-prandial, on observe un pic d’insuline qui fait écho à
l’augmentation de la glycémie. En effet, la sécrétion d’insuline est
régulée par le glucose qui joue le rôle de messager au niveau de la
cellule β pancréatique. Chez un sujet normal, à mêmes quantités de
glucose administrées on remarque une insulinémie 2 à 3 fois plus
importante par voie orale (ou per os) que par voie intraveineuse (IV).
Cela indique une différence de réponse qui a permis de mettre en
évidence l’existence de peptides sécrétés par le tractus gastro-
intestinal : les incrétines. Ces peptides permettent une amplification
de la réponse à l’insuline suite à l’absorption de glucose : c’est
l’effet incrétine. Cela permet un retour rapide de la glycémie à la
normale après un repas. Une molécule participe majoritairement à
l’effet incrétine : c’est le peptide GLP-1 (Glucagon Like Peptid 1).
N.B. : Les incrétines ont un effet systémique.
B) Synthèse du glucagon et des peptides dérivés comme le GLP-1
La synthèse du glucagon est réalisée dans les cellules α pancréatiques. Le glucagon est synthétisé
à partir d’un précurseur, une pro-protéine le pro-glucagon qui subit des clivages protéolytiques par
des enzymes appelées les pro-hormones convertases (PC1 spécifique du pancréas).

Page 4 sur 16 Ronéo 3 – Cours 5 UE8
Au niveau de l’intestin et du
cerveau ce pro-glucagon peut aussi
subir des clivages différents réalisés
par la PC2 qui aboutit à la création du
GLP-1 une incrétine. Le GLP-1 est
ensuite sécrété par les cellules L de
l’iléon.
On a donc une synthèse
parallèle de glucagon et GLP-
1 médiée par la distribution
tissulaire des PC dans l’organisme.
Le GLP-1 et le glucagon ont un mécanisme d’action similaire, ils se fixent sur des récepteurs à 7
domaines transmembranaires couplés aux protéines G et induisent la synthèse d’AMPc puis
l’activation de la PKA. Cependant le rôle n’est physiologiquement pas le même.
N.B. : Là s’arrête la ressemblance entre le glucagon et le GLP-1 qui n’ont pas les mêmes rôles
physiologiques. /!\ il ne faut pas être induit en erreur par la similarité des noms entre la synthèse de
glucagon et des incrétines.
C) Mécanismes d’action de la GLP-1 dans l’organisme
L’effet incrétine va se différencier en
2 types de mécanismes selon les
organes touchés qui sont
principalement l’estomac, le pancréas
endocrine avec les ilots de Langerhans
et le cerveau.
Au niveau de l’estomac la vidange
gastrique va être ralentie. Ce
ralentissement a un effet satiétogène
(je signale à mon organisme que j’ai
mangé et que je n’ai pas besoin d’une
autre prise alimentaire).
Au niveau du cerveau, le GLP-1 agit comme un neuromédiateur qui permet d’avoir un effet
satiétogène (rétrocontrôle négatif) qui s’ajoute à l’effet sur l’estomac.
Mais la régulation de la glycémie par les incrétines intervient surtout au niveau du pancréas avec
les cellules β et les cellules α des ilots de Langerhans. Le contrôle du GLP-1 peut être distingué en 2
actions sur les cellules β :
- un rôle sur la production et sécrétion de l’insuline. Les incrétines potentialisent la
sécrétion d’insuline par une augmentation du blocage des canaux K+ ATP dépendant
provoquant une sécrétion par exocytose plus importante d'insuline ;

Page 5 sur 16 Ronéo 3 – Cours 5 UE8
- un rôle sur la cellule β elle-même de type « facteur de croissance » à plus long terme. Le
GLP-1 augmente la prolifération des cellules β tout en inhibant l’apoptose qui favorise le
maintien en vie des cellules β pancréatiques.
En parallèle sur les cellules α, on va avoir un effet d’inhibition de la sécrétion de glucagon. En
effet, pendant une prise alimentaire une hormone hyperglycémiante n’est pas utile pour éviter un effet
antagoniste du glucagon.
D) Régulation de la GLP-1
La régulation de la glycémie par les
incrétines est très importante mais aussi très
transitoire après une prise alimentaire. La
durée de vie du GLP-1 est très courte (< 2
minutes). En effet, cette incrétine est
rapidement clivée par la dipeptidyl
peptidase IV (DPP-4) la rendant inactive.
Dans certaines pathologies de la régulation de la glycémie comme le diabète de type II, l’effet
incrétine est diminué et devient insuffisant par rapport à l’effet chez un sujet normal. Le différentiel
(voie orale et voie IV) est extrêmement amoindri chez un sujet diabétique de type II. Cela signifie
qu’après une prise alimentaire le sujet diabétique n’est pas capable de façon aussi importante que le
sujet normal de sécréter de l’insuline pour réguler sa glycémie.
N.B. : Le diabète de type II entraîne un défaut de la DPP-4 qui clive trop rapidement la GLP-1. Elle
n’est alors plus capable d’augmenter l’insulinémie.
Cette voie d’inactivation de la GLP-1 est exploitable en pathologie. Deux axes de recherche
pharmacologiques sont étudiés pour traiter le diabète de type II : des incrétino-mimétiques
(analogues des incrétines) qui résisteraient à la DPP-4 ou des inhibiteurs pharmacologiques de la
DPP-4. Ces deux approches permettent une action prolongée de la GLP-1 sur la sécrétion d’insuline.
III- Pathologies de la régulation de la glycémie
A) Le diabète
1) Généralités
Le diabète sucré (à différencier du diabète insipide) est une maladie métabolique caractérisée
par 2 grands éléments :
- une surproduction de glucose par le foie;
- une sous-utilisation du glucose par les cellules.
Paradoxe du diabète : excès de glucose dans le sang et manque de glucose au niveau des
organes

Page 6 sur 16 Ronéo 3 – Cours 5 UE8
Les critères diagnostiques s’appuient sur l’hyperglycémie provoquée :
- une glycémie à jeûn supérieure à 7mM à 2 reprises (éviter les erreurs de laboratoire) ;
- une glycémie supérieure à 11mM après administration de glucose par voie orale appelé
test de charge ou tolérance (en cas de diagnostic incertain on pousse l’organisme dans ses
retranchements en lui administrant du glucose).
La prévalence du diabète en France est plus élevée chez les hommes que chez les femmes.
L’augmentation de la prévalence est concomitante à l’âge avec un début vers 40 ans un pic de
fréquence chez les sujets âgés (70-75 ans).
Il n’existe pas un mais 4 types de diabète avec des étiologies et des fréquences très différentes :
- Les diabètes secondaires suite à un traitement, une infection (non traité dans le cours) ;
- Le diabète de type I ou diabète insulino-dépendant (DID) relativement rare (10 à 15%
des diabètes et incidence de 0,38% en France) et se déclare souvent dans l’enfance. Il met
en cause l’auto-immunité (destruction des cellules β et non sécrétion d’insuline), on parle
alors de maladie immune -> cause immunologique
- Le diabète de type II ou diabète non insulino dépendant (DNID) le plus fréquent (85 à
90% des diabètes et incidence en France de 4%). Il est aussi appelé diabète du sujet âgé ou
diabète du sujet obèse car l’âge et le surpoids sont des facteurs favorisant son apparition. Il
est lié à une insulino-résistance et une baisse de l’insulino-sécrétion étudiées
ultérieurement -> cause métabolique
- Le diabète monogénique relativement rare (1 à 2% des cas de diabète) mais en
augmentation. Il est d’origine génétique, avec une forme familiale transmissible et se
déclare avant 25ans -> cause génétique
2) Le diabète des enfants
90% des diabètes pédiatriques sont des cas
d’auto-immunité (cas principal DID). Quelques
pourcents des diabètes sont à cause génétique
(diabètes monogéniques). D’encore plus rares cas de
diabètes avec obésité sont observés chez les enfants
(nombre de cas en augmentation).
B) Diabète de type I
Dans le diabète de type I, l’anomalie de l’homéostasie du glucose est due à l’absence de sécrétion
d’insuline. L’auto-immunité dirigée contre les cellules β des ilots de Langerhans les a détruits : ils ne
sont donc plus capables de sécréter de l’insuline. L’absence d’insuline va se traduire par l’altération de
3 types de métabolisme : glucidique en 1er lieu puis lipidique.

Page 7 sur 16 Ronéo 3 – Cours 5 UE8
1) Altération du métabolimse glucidique
Les symptômes du diabète
sont la polyurie, l’amaigrissement,
la fatigue, des douleurs abdominales
donc une dégradation de l’état
général.
Comment relier ces
symptômes avec l’absence
d’insuline ?
L’insuline a une action sur 2
cibles différentes : le foie et les
tissus périphériques (muscle et
tissus adipeux)
- Au niveau du foie, la carence en insuline entraîne une surproduction du glucose
(hyperglycémie) par l’absence d’inhibition de la formation de glucose ;
- Les tissus périphériques (rôle majeur mais pas exclusif du muscle) ne sont pas capables
d’absorber l’excédent de glucose produit (pas de translocation de GLUT-2 à la
membrane).
- Le glucose est normalement réabsorbé au niveau rénal mais l’hyperglycémie est telle que
ses capacités de réabsorption sont rapidement dépassées donnant lieu à une perte urinaire
de glucose : la glycosurie (en situation physiologique il n’y a pas de glucose dans les
urines.
- Cette glycosurie va alors s’accompagner d’une perte d’eau : la polyurie et la perte
d’électrolytes dans un contexte de diurèse osmotique, de façon à éliminer le glucose.
- Cette perte d’eau provoque une sensation de soif : la polydipsie qui provoque à terme une
déshydratation pouvant évoluer en coma hyperosmolaire (un des signes de déclaration
du diabète).
2) Altération du métabolisme lipidique
Pour rappel, l’insuline est une hormone de mise en réserve qui permet la lipogénèse au niveau des
tissus adipeux. En son absence c’est la réaction inverse qui se produit la lipolyse. On a une production
importante d’acides gras (AG) qui formeront des acides cétoniques. Ces acides entraînent une baisse
du pH et donc une acidose pouvant aboutir à un coma acido-cétosique (un des signes de déclaration
du diabète).

Page 8 sur 16 Ronéo 3 – Cours 5 UE8
Formation et utilisation des corps cétoniques
La source majeure de production d’énergie provient des AG au niveau hépatique. Les AG vont
être dégradés par la β-oxydation en Acétyl-CoA qui va se combiner à un Oxaloacétate pour entrer dans
le cycle de Krebs et produire de l’énergie. Il faut donc un pool suffisamment abondant en oxaloacétate
disponible dans la cellule pour prendre en charge l’Acétyl-CoA. Chez un sujet en post-prandial, on a
tout en abondance donc la β-oxydation et l’entrée cycle de Krebs se font de manière harmonieuse.
Lors d’un jeûne, le foie met en œuvre 2 métabolismes pour compenser cette hypoglycémie qui
sont la glycogénolyse et la néoglucogenèse. En cas de jeûne prolongé, la néoglucogénèse prédomine.
Le substrat de cette néoglucogénèse est l’oxaloacétate qui est donc dévié de son utilisation par le cycle
de Krebs. Son pool s’appauvrit alors dans les cellules hépatiques et les AG absorbés ne peuvent plus
être pris en charge par le cycle de l’acide citrique. Dans ce cas, l’organisme met en place une voie
d’utilisation secondaire de l’Acétyl-CoA : c’est la cétogénèse.
L’Acétyl-CoA produit par la dégradation des AG n’entre dans le cycle de Krebs que si les
dégradations des lipides et des glucides sont équilibrées, d’où l’expression « les graisses ne
brûlent que sur le feu des sucres ».
Qu’est-ce que la cétogénèse ?
La cétogénèse est la production de corps cétoniques au nombre de 3 : l’acétoacétate, l’acétone et le
D-3-hydroxybutyrate. Ces corps cétoniques sont exclusivement synthétisés dans le foie au niveau de la
mitochondrie.
N.B. :Le D-3-hydroxybutyrate n’est pas une cétone. On a regroupé les 3 molécules sous le nom de
corps cétoniques pour témoigner de leur origine commune.
Il existe des interconversions entre ces corps cétoniques :
- l’acétoacétate peut spontanément se carboxyler en acétone ;
- l’acétoacétate peut dans la mitochondrie par action de la D-3 hydroxybutyrate
déshydrogénase se transformer en D-3-hydroxybutyrate en utilisant un proton provenant
du NADH,H+.

Page 9 sur 16 Ronéo 3 – Cours 5 UE8
1. Sous l’effet de la β-Cétothiolase (Acétyl-CoA-Acyl-Transférase) 2 molécules d’Acétyl-CoA
provenant de la β-oxydation vont se condenser en Acéto-Acétyl-CoA à 4 carbones.
2. S’ajoute un nouveau Acétyl-CoA à l’Acéto-Acétyl-CoA grâce à une HMG-CoA Synthétase
mitochondriale donnant le 3-Hydroxy-3-Méthyl-Glutaryl-CoA (HMG-CoA).
3. Se produit alors le départ d’un Acétyl-CoA d’une HMG-CoA grâce à une HMG-CoA Lyase
donnant l’Acétoacétate.
4. Ce dernier peut être hydrogéné en D-3-hydroxybutyrate ou spontanément déshydrogéné en
Acétone.
Le foie ne pouvant pas utiliser ces corps cétoniques, l’Acéto-Acétate, le D-3-Hydroxybutyrate et
l’Acétone sont exportés vers les tissus périphériques (tissus autres que le foie) : c’est la conversion
périphérique des corps
cétoniques.
1. Le β-Hydroxybutyrate (D-3-hydroxybutyrate) est transformé en Acétoacétate sous l’action de
la β-Hydroxybutyrate déshydrogénase (DH).
2. Cet Acétoacétate en présence de Succinyl-CoA se convertit en Acéto-Acétyl-CoA et du
Succinate. Ce transfert du CoA est réalisé par l’Acéto-Acétyl-CoA transférase qui n’est pas
exprimée dans le foie (c’est pour cette raison que les corps cétoniques ne sont pas utilisables
par le foie).

Page 10 sur 16 Ronéo 3 – Cours 5 UE8
3. Enfin une β-cétothiolase clive l’Acéto-Acétyl-CoA en 2 Acétyl-CoA pouvant rentrer dans le
cycle de Krebs de ces tissus périphériques qui ne font pas la néoglucogénèse.
C’est un métabolisme qui fait sens dans l’organisme entier, le foie d’un côté, le sang pour la
communication et les tissus hépatiques de l’autre. On a une production hépatique et une
consommation extra-hépatique des corps cétoniques.
Exemple de la régulation de la glycémie après une nuit de jeûne
Utilisons alors toute nos
connaissances sur le sujet pour
comprendre ce qui se passe pendant
une nuit de jeûne. Pendant une nuit de
jeûne, différents phénomènes vont
s’enclencher les uns à la suite des
autres de façon à garder une glycémie
normale.
Le premier phénomène mis en jeu
le plus rapidement est la glycogénolyse hépatique, pendant les 14 premières heures. (avec en parallèle
une diminution progressive de l’insuline.) C’est le moyen le plus rapide d’obtenir du glucose. On
utilise alors les réserves de glycogène réalisées la journée. Si le jeûne se prolonge ces réserves vont
progressivement diminuer pour au bout de 60 h être quasiment réduites à néant. En même temps la
néoglucogenèse se met en œuvre modestement et prend le pas sur la glycogénolyse au bout de 22h de
jeûne pour assurer la quasi-totalité des apports en glucose par la suite. (à l’origine de 90% de la
production de glucose après 60h). Elle est faite essentiellement par le foie et utilise le glycérol de la
lipolyse, les AA glycogéniques obtenus par la fonte musculaire (alanine et glutamine). Cette
néoglucogenèse est plus puissante que la glycogénolyse mais dépend du muscle ce qui peut avoir des
effets délétères sur l’organisme à long terme.
L’organisme essaye alors de se protéger d’une protéolyse trop importante et met en place la
cétogenèse qui va fournir de l’énergie aux muscles. L’objectif physiologique est d’épargner. Les corps
cétoniques produits peuvent être aussi utilisés par le système nerveux (en particulier le SNC). Les
muscles et le système nerveux sont alors protégés en cas de jeûne prolongé.
Dans les conditions physiologiques la production de corps cétoniques est toujours modérée et
contrôlée par une petite sécrétion basale d’insuline. Mais en cas de diabète de type 1 (en absence
d’insuline) cette production de corps cétoniques est extraordinairement amplifiée et aboutit à un coma
acido-cétosique. On a un emballement de cette cétogenèse qui s’ajoute à un taux de glucose abondant
mais parfaitement inutilisable : Une lipolyse importante se met en place sous l’effet de l’activation des
lipases hormono-sensibles. On a un passage dans le sang d’un nombre important d’AG libres qui
rejoignent le foie. Au niveau du foie la néoglucogenèse continue sans cesse puisqu’elle n’est pas
stoppée par l’insuline. Le cycle de Krebs quant à lui ne fonctionne pas correctement pour dégrader les
AG. On a donc une production de corps cétoniques à partir d’acétyl-CoA après β-oxydation. Dans un
premier temps ces corps cétoniques vont atteindre les tissus extra-hépatiques et permettre au cerveau
de fonctionner correctement mais dans un second temps ils vont avoir un effet toxique sur le SNC par
la diminution du PH. C’est ainsi que le manque d’insuline va se traduire par un coma acido-cétosique.
(à l’origine d’un danger mortel pour les enfants).

Page 11 sur 16 Ronéo 3 – Cours 5 UE8
N.B. : Au niveau du sang on peut
avoir une perte de ces corps
cétoniques, l’acétone étant volatil.
Cette acétone va avoir tendance à
être éliminée en particulier par le
poumon (à l’origine de la
mauvaise haleine après une nuit
de jeune : on souffle de l’acétone).
On a aussi l’élimination de corps
cétoniques dans les urines.
L’injection d’insuline plusieurs fois par jour permet de traiter le diabète de type 1.
C) Diabète monogénique
Appelé pendant longtemps le MODY (Maturity Onset Diabet in Young : le diabète du sujet jeune)
on l’appelle maintenant diabète monogénique. On a découvert que derrière ce diabète se cachait une
cause génétique : une mutation mendélienne de gènes contrôlant la production et la sécrétion
d’insuline, ou le bon développement du pancréas. Les cibles de ses mutations sont diverses et
variées : facteurs de transcription comme la glukokinase, SUR1 du canal K+ ATP-dépendant (qui
quand il est muté, ne répond plus à l’ATP et n’entraine pas d’exocytose de l’insuline. (Voir cours n°3
en l’UE1)), KIR.6, ect…
Ce schéma regroupe toutes les protéines qui peuvent être mutés dans ce diabète monogénique
(représentés par les étoiles)

Page 12 sur 16 Ronéo 3 – Cours 5 UE8
D) Diabète de type 2
1) Epidémiologie
Le diabète de type 2 a une base physiopathologie complètement différente des deux autres
types de diabète traités. C’est un diabète qui a une origine métabolique qui se caractérise par une
résistance à l’insuline.
L’obésité reste la particularité épidémique et biologique importance de ce diabète. L’obésité
dans le monde a augmenté durant les 20 dernières années et le surpoids touche de nos jours 80% de la
population mondiale. En parallèle une augmentation du nombre de diabète de type 2 apparaît ce qui
nous amène à parler de « diabésité » compte tenu du lien étroit ente obésité et diabète de type 2. Le
diabète de type 2 est une pathologie en constante progression qui touche en 2014, 4% de la population
française et 9% de la pop mondiale. Notons aussi que l’obésité touche de plus en plus les enfants et est
donc à l’origine de l’apparition du diabète du sujet âgé chez les enfants.
2) Le syndrome métabolique
Le syndrome métabolique (ou syndrome X) est un nouveau syndrome pré-diabétique qui
précède souvent la survenue de pathologies comme le diabète 2 et les complications cardio
vasculaires. C’est un syndrome apparu suite à l’augmentation de l’obésité. Il associe plusieurs
anomalies métaboliques : l’obésité abdominale, l’hypertriglycéridémie, le HDL-cholestérol bas,
l’hyperglycémie à jeun. Cette hyperglycémie à jeûn témoigne d’une insulino-résistance.
N.B. : Ce syndrome a une définition qui varie puisque c’est quelque chose de pas encore
totalement établi.
Qu’est-ce qu’est l’insulino-résistance ?
Chez un sujet insulino-résistant, on observe de façon concomitante des choses qui ne devraient
pas arriver en même temps : on a une glycémie à jeàn anormalement augmentée par une surproduction
hépatique excessive de glucose en dépit de taux élevés d’insuline. (Normalement on a soit un manque
d’insuline et alors la une glycémie trop forte ou une glycémie basse par sécrétion de taux élevé
d’insuline) : L’insuline n’est alors pas capable de normaliser la glycémie. Ce qui définit la résistance a
l’insuline, c’est la réduction de la réponse des tissus à des concentrations physiologiques normales
d’insuline. Pour comprendre comment cela fonctionne il faut se rappeler de la manière dont l’insuline
envoie son signal aux cellules.
L’insuline se fixe sur son
récepteur à tyrosine kinase et va
ainsi effectuer une transduction du
signal par le biais d’un substrat qui
va être phosphorylé par le récepteur
à l’insuline : l’IRS. Cet IRS
phosphorylé au niveau des tyrosines,
va ensuite médié un signal via la
voie MAP-kinases, responsable des
actions mitogènes, et la voie PI3K,
responsable de l’ensemble des
actions métaboliques (comme la

Page 13 sur 16 Ronéo 3 – Cours 5 UE8
translocation de GLUT4, qui favorise l’entrée du glucose dans les cellules). Chez les patients atteints
de DNID, on a un nombre diminué de récepteurs à l’insuline avec une activité diminuée de ce
récepteur par rapport à la normale. Ceci est lié au faite qu’on a un nombre plus faible d’IRS qui sont
majoritairement déphosphorylés et donc une diminution de l’activité de la voie PI3K.
Qu’est ce qui se passe chez des patiens insulino-résistants ?
Ces patients en surpoids ont une
quantité plus importante de tissus adipeux
avec une composition différente du tissu
adipeux. On observe une infiltration de
macrophages à hauteur de 40% alors qu’elle
est observée physiologiquement à hauteur de
10%. Ces cellules vont libérer des cytokines
inflammatoires, les TNF-α qui induisent un
état permanent de pro-inflammation. De plus,
du fait de sa représentation plus importante,
chez le sujet obèse on aura une libération plus
importante d’AG plasmatiques. On remarque
de plus un équilibre modifié des adipokines
(pas plus détaillé dans le cours).
Il faut d’abord rappeler la situation physiologique pour comprendre le mécanisme de la
pathologie. L’insuline en se fixant sur son récepteur active différentes voies de signalisations telles
que MAPK et PI3K qui vont entraîner le scénario de leur propre extinction en même temps que
l’action liée à l’insuline. En effet, MAPK et PI3K activent des Sérines Kinases (IKKβ et JNK)
capables de phosphoryler le complexe IRS et ainsi découpler le récepteur à l’insuline de l’IRS. Ce
mécanisme permet l’extinction physiologique du signal à l’insuline. Se rajoute l’action des Tyrosines
Phosphatases (PTP1β) qui enlève les phosphores des Tyrosines de l’IRS et parfait l’extinction du
signal.
Dans le diabète de type II, il y a des interférences sur ce mécanisme d’extinction du signal :
- 1ère
interférence liée aux AG. Les
AGL sont en quantités plus
importantes chez un sujet obèse. Ils
sont capables d’entrer dans la cellule
et d’agir comme des messagers et
d’activer une PKC qui va
phosphoryler IRS au niveau de
Sérines inactivatrices comme le
font les kinases stimulées
normalement dans l’extinction du
signal. Via cet excès d’acyl-CoA
(provenant de la dégradation des
AG) on va favoriser le découplage du

Page 14 sur 16 Ronéo 3 – Cours 5 UE8
récepteur à l’insuline et de l’IRS et donc le récepteur fonctionne moins bien.
- 2ème
interférence liée à l’infiltration des tissus adipeux par les macrophages qui génèrent
une sécrétion continue de cytokines (TNF-α). Le TNF-α est capable via ses récepteurs
membranaires d’activer IKKβ et donc la phosphorylation d’IRS qui favorise le
découplage du récepteur et du complexe.
Ces deux mécanismes vont interférer avec l’extinction physiologique du signal. Quand ces
éléments sont augmentés la signalisation est moins efficace : c’est l’origine de la résistance à
l’insuline.
- 3ème
interférence
liée aux protéines. En situation obèse on a une
présence importante d’AA soit par les protéines
alimentaires qui sont ingérées en excès soit à
cause d’un catabolisme protéique accéléré. Ces
AA vont venir activer mTOR qui
physiologiquement active des Serines kinases
(S6K1 et 4EBP) responsables de la
phosphorylation de l’IRS. mTOR est activé
physiologiquement par PKB/AKT et exerce
donc un rétro-contrôle négatif. Les AA
potentialisent encore plus le découplage du
récepteur à l’insuline et de l’IRS par
l’activation de mTOR et donc une
augmentation du rétro-contrôle négatif.
L’organisme a donc besoin de quantités en insuline plus importantes que la normale pour fonctionner.
Les différentes étapes de l’installation lente du diabète de type II :
1. Résistance des tissus à l’insuline qui ne se voit pas car l’organisme compense.
2. Mauvaise réponse à l’insuline : absence de capture du glucose, production exagérée de
glucose par le foie.
3. Hyperglycémie qui va augmenter la sécrétion d’insuline = sursécrétion d’insuline pour
normaliser la glycémie.
4. L’organisme n’arrive plus à compenser les cellules β pancréatiques s’épuisent (plus
capables de sécréter suffisamment d’insuline.
3) Génétique du diabète de type II
Le diabète de type II est soumis à des facteurs génétiques mais n’est pas transmis sur un mode
mendélien. C’est une maladie polygénique c’est-à-dire déterminée par plusieurs facteurs ayant chacun
un effet faible. Il faut donc prendre en compte un certain nombre de variants génétiques et les effets de
l’environnement sur le développement de la pathologie. Pour étudier ces variants on utilise des études
d’associations pangénomiques (GWAS ou Genome Wide Association Studies) entre des variants
communs du génome (SNP) et le développement de la maladie. Ce sont des études de cohorte qui ne
peuvent se faire que sur des populations importantes. On prend pour cela deux groupes appariés sur
le plus de facteurs possibles. On va étudier l’ensemble du génome pour essayer de voir si il y a des

Page 15 sur 16 Ronéo 3 – Cours 5 UE8
variants nucléotidiques qui sont plus fréquemment retrouvés chez les patients atteints de diabète par
rapport au groupe contrôle non atteint = fréquence allélique.
Les premières études ont permis d’identifier 44 polymorphismes dans les régions codantes ou
non dont la présence était plus importante chez les patients atteints par rapport au groupe témoin non
atteint. Ces variants ne sont pas placés n’importe où dans le
génome, ils se situent à proximité de gènes impliqués dans
la sécrétion de l’insuline par les cellules β du pancréas.
Par exemple, KCJN11 qui code pour KIR6.2, le port du
canal potassique ATP dépendant des cellules β. Ce gène est
parfois retrouvé muté dans des cancers monogéniques mais
on met surtout en évidence un variant (E23K) qui rend plus
susceptible le développement de la maladie. Il faut
cependant prendre en compte les facteurs environnementaux.
Montre le rôle central des capacités de sécrétion mises en jeu dans le diabète de type 2.
Plus on a de variants qui permettent de sécréter normalement de l’insuline plus on va pallier
les effets de l’obésité mais plus on a de variants de susceptibilité de développement de la
maladie et qu’on pousse l’organisme dans ses retranchements on a développement de la
maladie.
E) Les complications du diabète à long terme
- Rétinopathie diabétique : 1ère
cause de cécité en France chez les moins de 50 ans, 40%
des patients après 10 ans de diabète. Après 15 ans de diabète : 2% d’aveugles et 10% de
malvoyants. Importance surveillance et dépistage du diabète.
- Neuropathie diabétique : 28 à 45%
- Athérome : manifestations ischémiques chroniques et épisodes aigus thrombotiques
- Néphropathie diabétique : 13% des insuffisances rénales en France (>30% aux USA).
Plus de 50% des diabétiques de type II sont touchés et 30% des types I entre la 10ème
et la
25ème
année.
F) Liens entre obésité, diabète et cancers
On s’aperçoit, par des études d’épidémiologie, que lorsqu’il y a un syndrome métabolique de
diabète de type II et/ou un syndrome d’obésité que le risque de développer un cancer augmente. L’un
des grand champs de la recherche est d’observer à quel point les modes de vie influencent le risque et
la gravité des cancers.
Le facteur de croissance IGF est capable de signaler sa présence à la cellule par un récepteur à
activité tyrosine kinase qui est très proche de celui de l’insuline. L’insuline est capable de reconnaître
le récepteur à l’IGF et réciproquement. Si l’insuline est en quantité normale, elle se signale
essentiellement à son récepteur et assure ses effets métaboliques avec peu d’effets mitogènes.

Page 16 sur 16 Ronéo 3 – Cours 5 UE8
Dans le diabète de type II, l’insuline est en quantité plus importante et se fixe sur le récepteur à
l’IGF (présent dans beaucoup de tissus du corps humain) ce qui augmente les effets mitogènes et
facilite la prolifération de cellules. Cela ne suffit pas à faire un cancer mais est un élément qui a été
montré comme augmentant le risque d’en développer un.
Dédicace :
- La team Valide Thon Année ou Life Comes Back (peu importe le nom les meilleurs y sont)
avec Tomy, Jiji, Jeannou, Jo, Mili, Lulu et nous CamK et Cec ! Même en P2 on pense à
vous les épicières et la masseuse.
- Gros big up au Bungalow des BG du WEI on vous trash fort !
- Love et câlins à tous les petits P2 qui sont arrivés jusqu’à cette page.