Les membranes biologiques Membrane plasmique et membranes internes ( endomembranes )

Chapitre III
Fonctions de la membrane plasmique:C/ perméabilité cellulaire
Dr A. DEKAR
2014- 2015
Médecine dentaire

Objectifs pédagogiques
Objectif 3: Citer les caractéristiques du transport perméatif passif
Objectif 5: Indiquer les classes de molécules qui empruntent la diffusion facilitée
Objectif 6: Comparer les caractères structuraux des perméases et des canaux
Objectif 1: Citer les caractères distinctifs entre le transport perméatif et le transport cytotique
Objectif 2: Enumérer les variété de transports perméatifs
Objectif 4: Donner les caractères physico-chimiques de molécules qui répondent à la diffusion simple
Objectif 7: Comparer les modalités de transport du glucose dans 3 types cellulaires: Les érythrocytes, les adipocytes et les entérocytes.

Objectif 14: Citer les pathologies humaines liées aux dysfonctionnement des systèmes de transport perméatif et cytotique.
Objectif 8: Donner les spécificités du transport de l’eau dans les cellules rénales
Objectif 9: Comparer les modalités de transport des ions dans les cellules musculaires et les cellules nerveuses.
Objectif 10: Citer les caractéristiques du transport perméatif actif.
Objectif 11: Comparer, par des exemples cellulaires les modalités des transports actifs primaire et secondaire.
Objectif 12: Citer les caractéristiques du transport cytotique.
Objectif 13: Enumérer les variétés du transport cytotique.
Objectif 14: Expliquer, a l’aide de schémas, les modalités des endocytoses.

La perméabilité cellulaire= transports membranaire
Transports perméatifs …...
Transports cytotiques
la membrane assure destransports spécifiques
A l’échelle moléculaire
Non visibles à l’observation de la cellule au ME
A l’échelle cellulaire
Visibles en ME

La taille et les propriétés chimiques des molécules sont déterminantes pour leur passage ou non à travers la bicouche lipidique
La membrane assure un transport spécifique et sélectif
A travers une membrane artificielle purement lipidique
Membrane non perméable à tout type de molécules !!!
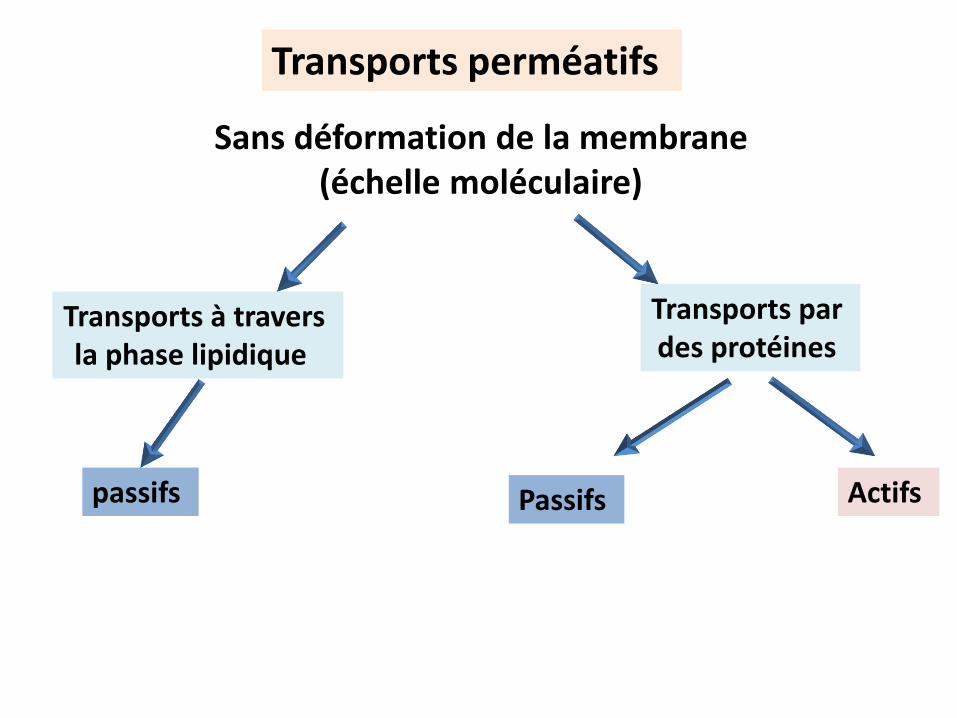
Transports perméatifs
Transports à travers la phase lipidique
Transports par des protéines
Sans déformation de la membrane(échelle moléculaire)
passifs Actifs Passifs
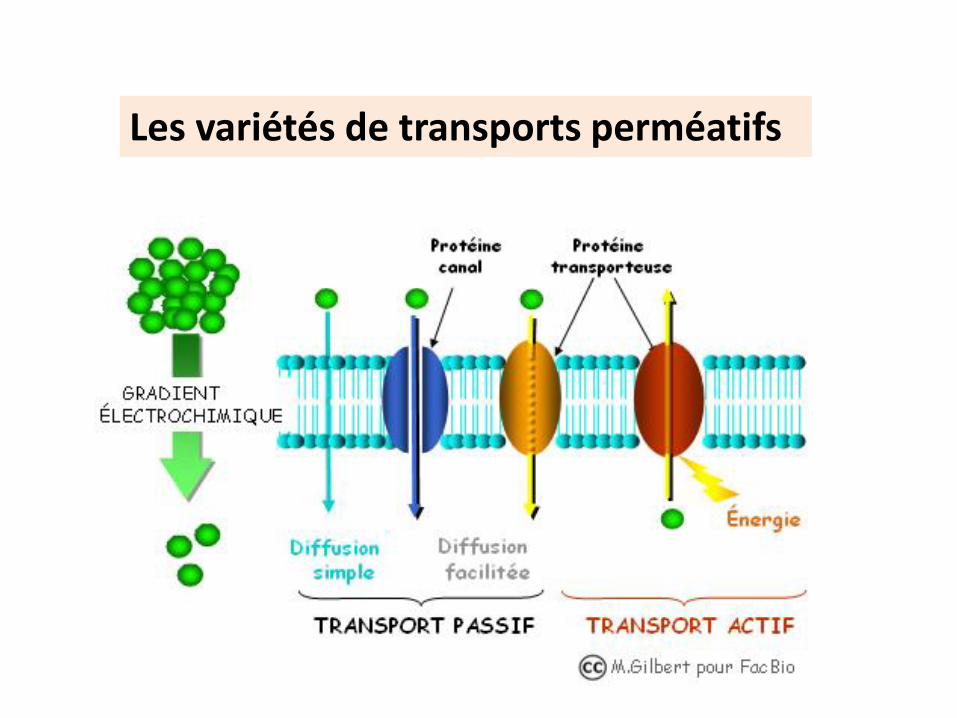
Les variétés de transports perméatifs
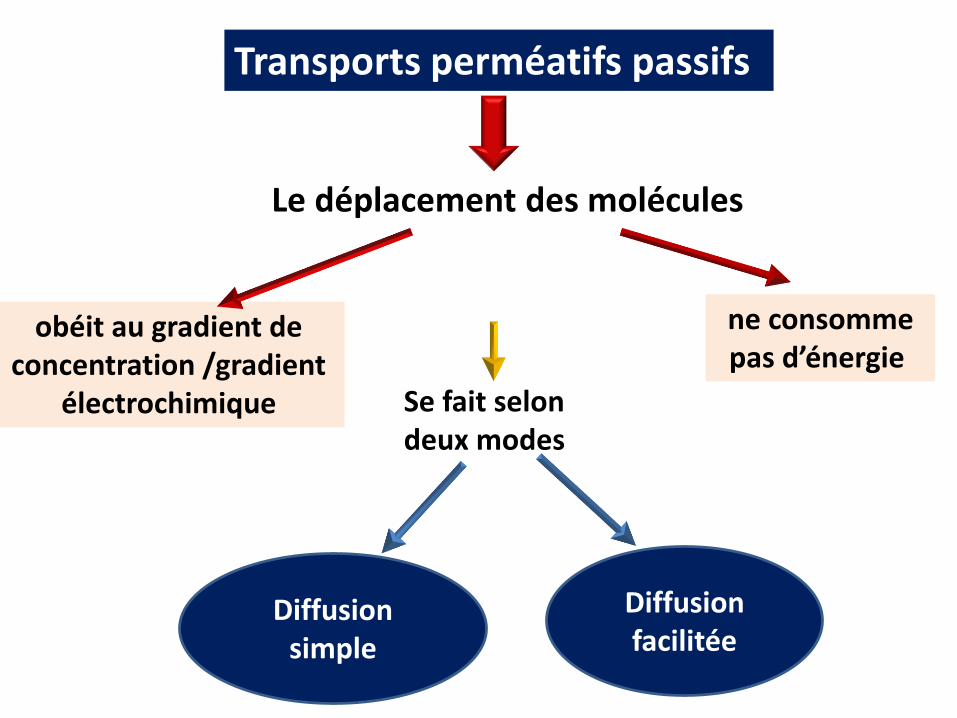
Transports perméatifs passifs
Le déplacement des molécules
ne consomme pas d’énergie
obéit au gradient de concentration /gradient
électrochimique Se fait selon deux modes
Diffusion simple
Diffusion facilitée

Le gradient de concentration: le transport se fait du Du milieu riche vers le milieu pauvre Jusqu’à équilibre des concentrations
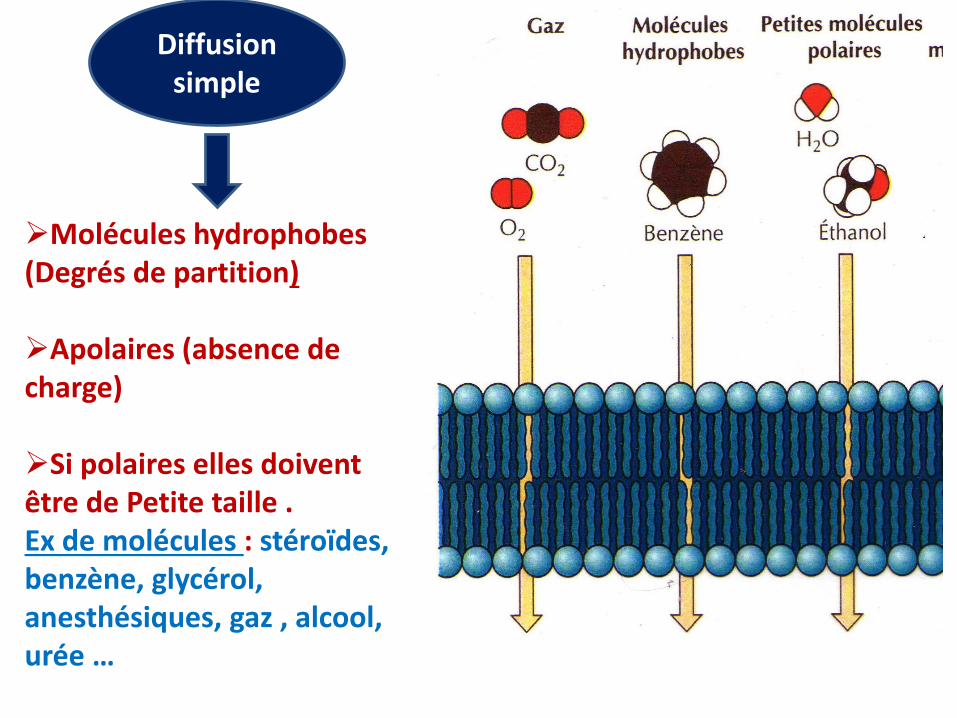
Molécules hydrophobes (Degrés de partition)
Apolaires (absence de charge)
Si polaires elles doivent être de Petite taille .Ex de molécules : stéroïdes, benzène, glycérol, anesthésiques, gaz , alcool, urée …
Diffusion simple

Molécules polaires de grande taille: sucres,acides aminés…Petites molécules chargées: Ions: Na+, K+,Cl-
Diffusion facilitée

Glycoprotéines transmembranaires
perméases Protéines poreuses
Canaux ioniques
Connexons
osesEau
GLUT (AQP)
Aquaporines
Diffusion facilitée par transporteurs
Ions
Petites moléculessignalétiques

Les perméases

Les perméases
Glycoprotéines, transmembranaires porteusescaractérisées par:
Présence de site(s) de fixation pour la oules molécule(s) à transporter
Son changement conformationnel assure le déplacement de(s) molécule(s) d’un milieu vers l’autre.
sa position dans la membrane indique l’orientation du / dessite (s) et détermine le sens du transport.
Son fonctionnement nécessite ou pas de l’énergie cellulaire

Les protéines porteuses
Transport passif Transport actif
Deux modes de fonctionnement des perméases

Transport passif
Suivent le gradient deconcentration /
gradient électrochimique
Sans consommationd’énergie
Transport actif
Avec consommationd’énergie
Contre le gradient deconcentration /
gradient électrochimique
Deux modes de fonctionnement des perméases
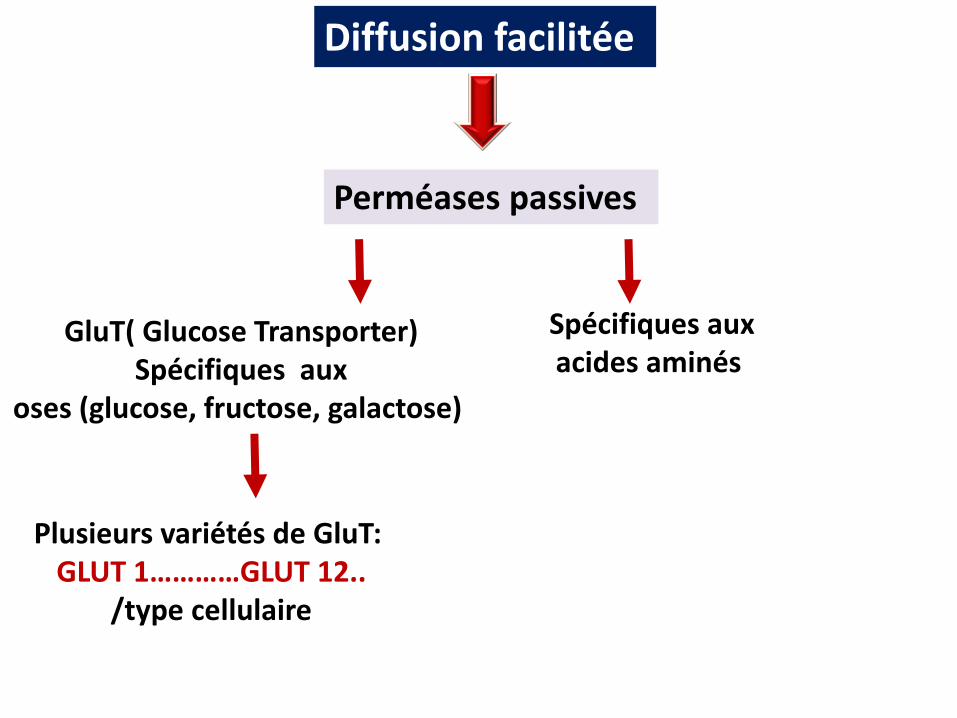
Diffusion facilitée
Plusieurs variétés de GluT: GLUT 1…………GLUT 12..
/type cellulaire
Perméases passives
GluT( Glucose Transporter)Spécifiques aux
oses (glucose, fructose, galactose)
Spécifiques auxacides aminés

Type de Glut Distribution tissulaire Fonction
Glut1 +++ érythrocytes, Transport du glucose basal
Glut 2 Cellules polariées:Pole basal des Entérocytes et cell. Rénales, faces latérales des hépatocytes
Transport bidirectionnel du glucose: sortie vers la circulation
Glut 4 Cell. Insulino-dépendantes: Adipocytes, muscle cardiaque et squelettique
Transport régulé du glucose
Principales variétés des Glut

Perméases du glucose
Diffusion facilitée du glucose par les GLUT
GLUT 1 GLUT 2
Entrée du Glucose dansles Erythrocytes
Sortie du glucose des cellules
vers la circulation : cas des cell. orientées
Entrée du glucose dans les adipocytes
GLUT 4
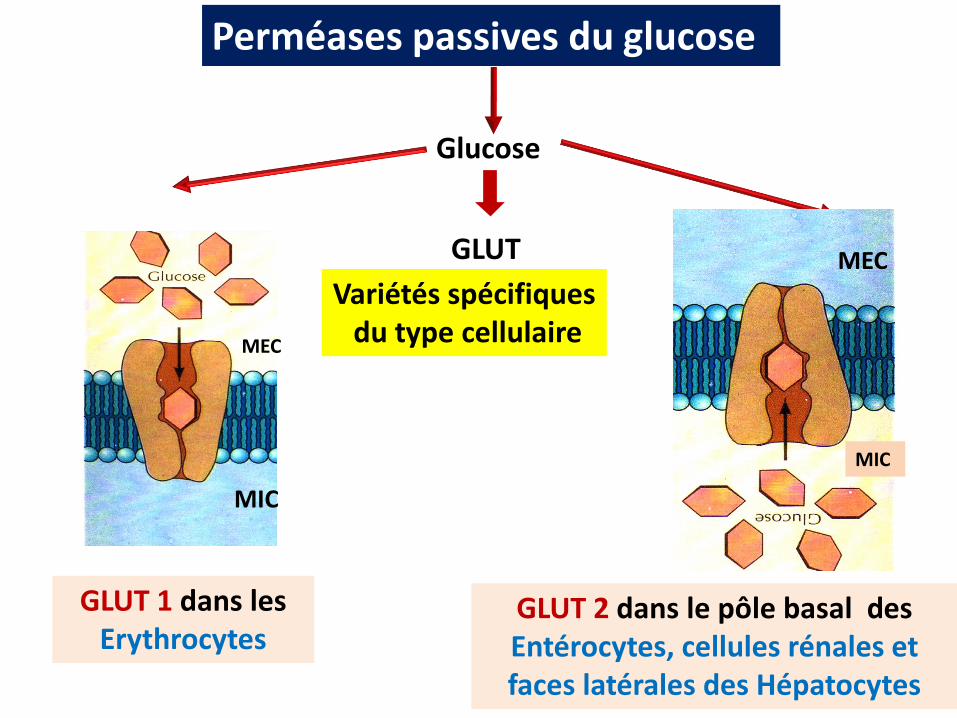
Perméases passives du glucose
GLUT
Glucose
Variétés spécifiquesdu type cellulaire
GLUT 1 dans les Erythrocytes
GLUT 2 dans le pôle basal des Entérocytes, cellules rénales et faces latérales des Hépatocytes
MEC
MIC
MEC
MIC

Transport du glucose

les GLUT 1 assurent l’entrée du glucose dans l’érythrocyte et la majorité des types cellulaires en 3 étapes (voir planche)
1Liaison &
Formation du complexe
2Changement de conformation
3 Libération &
Retour à la position initiale

La diffusion facilitée du glucose par les Glut 1 est contrôlée par leur disponibilité: la courbe de
déplacement présente un plateau de saturation.

Les GUT 2 assurent la sortie du glucose de l’entérocyte vers la circulation ( voir planche )
Glut 2 au pôle basal de l’entérocyte
Glucose
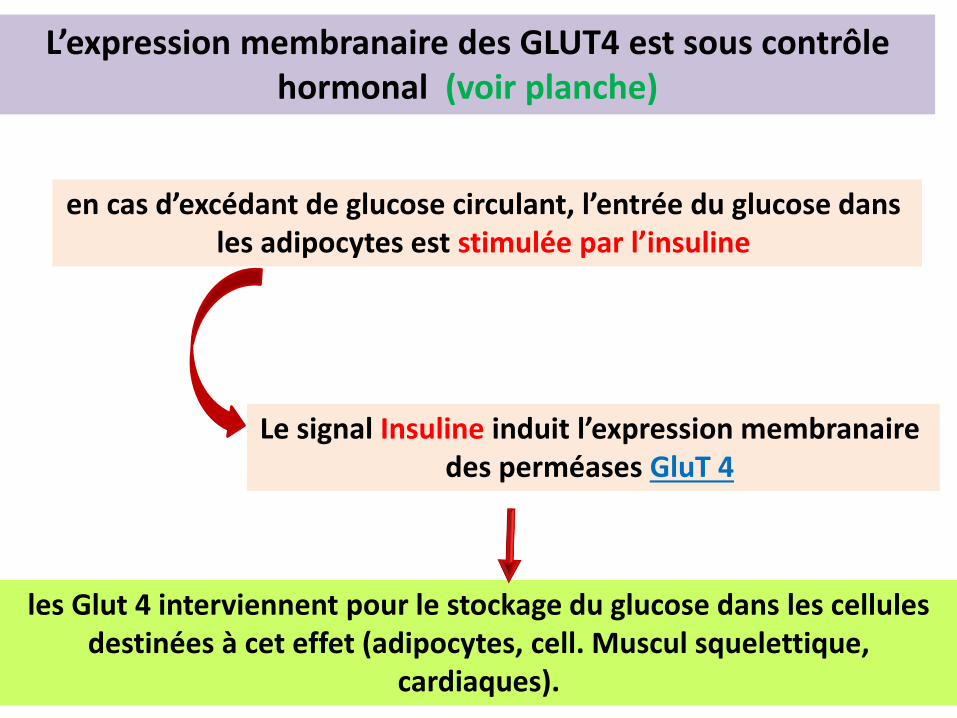
Le signal Insuline induit l’expression membranaire des perméases GluT 4
en cas d’excédant de glucose circulant, l’entrée du glucose dans les adipocytes est stimulée par l’insuline
les Glut 4 interviennent pour le stockage du glucose dans les cellules destinées à cet effet (adipocytes, cell. Muscul squelettique,
cardiaques).
L’expression membranaire des GLUT4 est sous contrôle hormonal (voir planche)

Entrée du glucose dans les adipocytes ( selon3 étapes comme par les Glut1)
Conditions et mode d’expression des Glut 4 au niveau des adipocytes
En cas d’hyperglycémie
Stimulation des cellules β du pancréas
Libération de l’insuline par exocytose
Fixation de l’insuline à ses récepteurs membranaires portés par les adipocytes
Cascade de signalisation intracellulaire induite par le complexe insuline- récepteur (voir cours communication cellulaire)
Activation de l’exocytose de vésicules intracellulaires videsdont les membranes sont riches en Glut 4

Expression des Glut 4 membranaires
Libération et action de l’insuline sur ses cellules cibles(cas de la cellule musculaire)

Mécanisme de transport du glucose dans les cellules insulino- dépendantes: ex l’ Adipocyte
REG
Golgi
Vésicule À Glut 4
Signaux moléculaires
Migration
exocytose
Expression membranaire
des Glut 4
Synthèse et modificationdes Glut 4
Glucose
MEC
Endosome
insuline
1
3Endocytose
Dissociation
2
7
4
5
68
9
A. Dekar

Expression des Glut 4 au niveau membranaire
Transport cytotique des perméases Glut 4
L’entrée du glucose dans les adipocytesSe fait en deux phases
Diffusion facilitée du glucose
Entrée du glucose du MEC vers le MIC
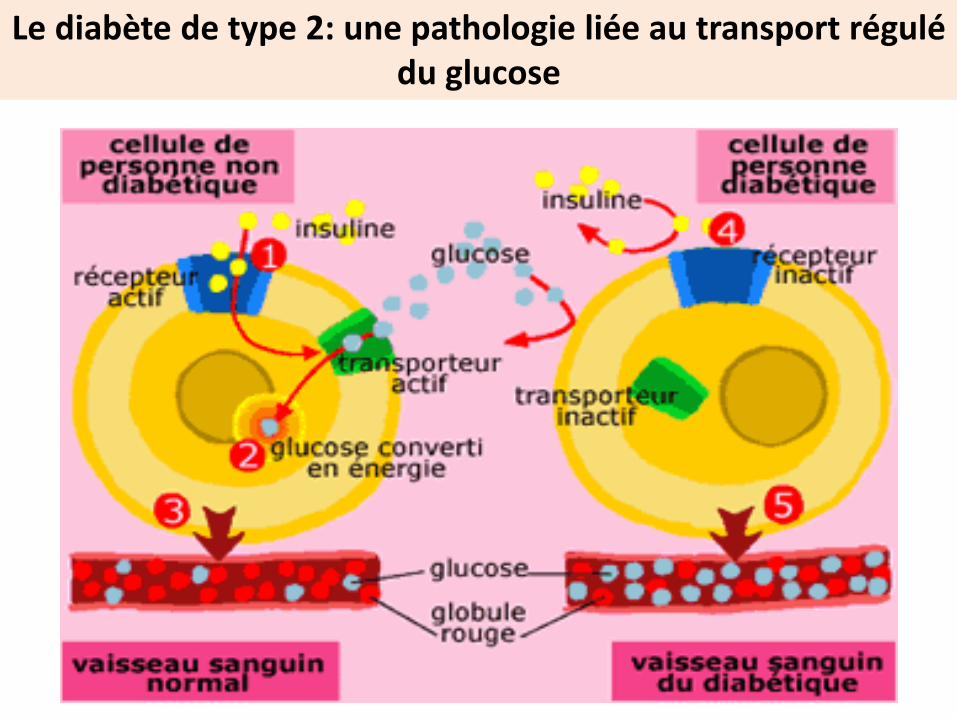
Le diabète de type 2: une pathologie liée au transport régulé du glucose

R-insulineabsents
R- insuline présents mais
non fonctionnels
Pas d’expression des Glut 4
Le glucose reste en circulation
Causes possibles
Pas de sécrétion d’Insuline
Diabète

Les protéines poreuses=
Les canaux
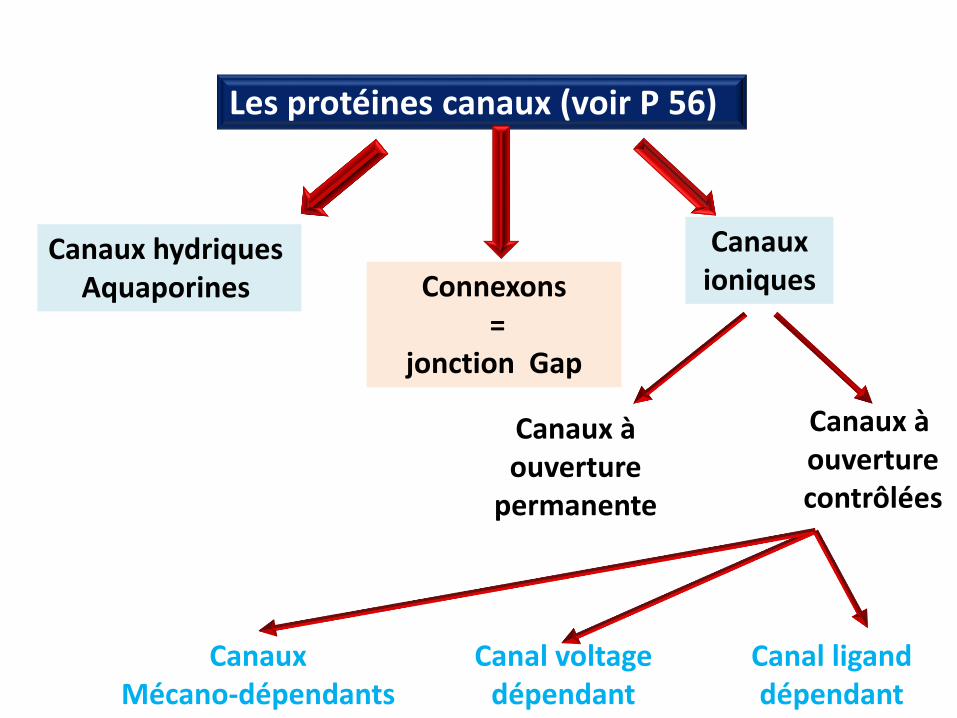
Les protéines canaux (voir P 56)
Canaux hydriques Aquaporines
Canaux Mécano-dépendants
Canal voltage dépendant
Canal ligand dépendant
Connexons=
jonction Gap
Canauxioniques
Canaux à ouverture
permanente
Canaux à ouverturecontrôlées

Les aquaporines

De grands volumes d'eau circulent dans notre corps de cellules en
cellules. La diffusion des molécules d'eau au travers des
membranes cellulaires n'explique pas la circulation de telles
quantités d'eau.
En effet, ces transports sont assurés par des protéines membranaires
poreuses (canal) qui transportent spécifiquement l’eau: ce sont les
aquaporines (AQP)
Découverte

L’aquaporine 1 humaine est exprimée dans de très nombreux tissus : érythrocytes, tubules rénaux, coeur, rein, muscle squelettique, foie,
pancréas, ...
Actuellement, on dénombre 13 aquaporines chez les mammifères:AQP O, AQP1……AQP 12
Les aquaporines ont été découvertes par Peter Agre(Prix Nobel 2003).
les variétés d’AQP ont une distribution tissulaire spécifique:

Répartition des aquaporines et des canaux ioniques dans la membrane

Chaque AQP est un tétramères qui délimite un pore. Le diamètre du pore permet le passage des
molécules d’eau une à une à la manière d’un sablier
Diffusion facilitée de l’eau
La structure de l’AQP:

transport de l’eau dans les cellules rénales fait intervenirplusieurs types d’AQP (AQP 2, AQP 3 et AQP 4)

L’expression des AQP 2 au pôle apical répond au signal vasopressine,
(hormone impliquée dans le contrôle de l’hydratation des tissus)
dont les récepteurs sont situés au pôle basal.

L’eau qui rentre du pôle apical par les AQP2 est libérée dans la circulation par les faces latéro-basales grâce aux AQP 3 et AQP4

Les protéines canaux (voir P 56)
Canaux hydriques Aquaporines
Canaux Mécano-dépendants
Canal voltage dépendant
Canal ligand dépendant
Connexons=
jonction Gap
Canauxioniques
Canaux à ouverture
permanente
Canaux à ouverturecontrôlées
Les connexons (voir jonctions intercellulaires )
Connexons

Les canaux ioniques

Canaux ioniques
Canal voltage
dépendant
Canal ligand
dépendant
Canaux à ouverture permanente
Canaux à ouverture Contrôlée
Canal Mécano-
dépendants

Le gradient de concentration / gradient électrochimique
Gradient Cl-
Gradient K+

Le passage des ions par les canaux suitle gradient électrochimique spécifique à chaque ion
+++++++++++++++++++++++
--------------------------------------

La concentraction des ions dans le MIC et le MEC est différente. Ce gradient électrochimique détermine le potentiel de membrane
et le sens du transport de chaque ion
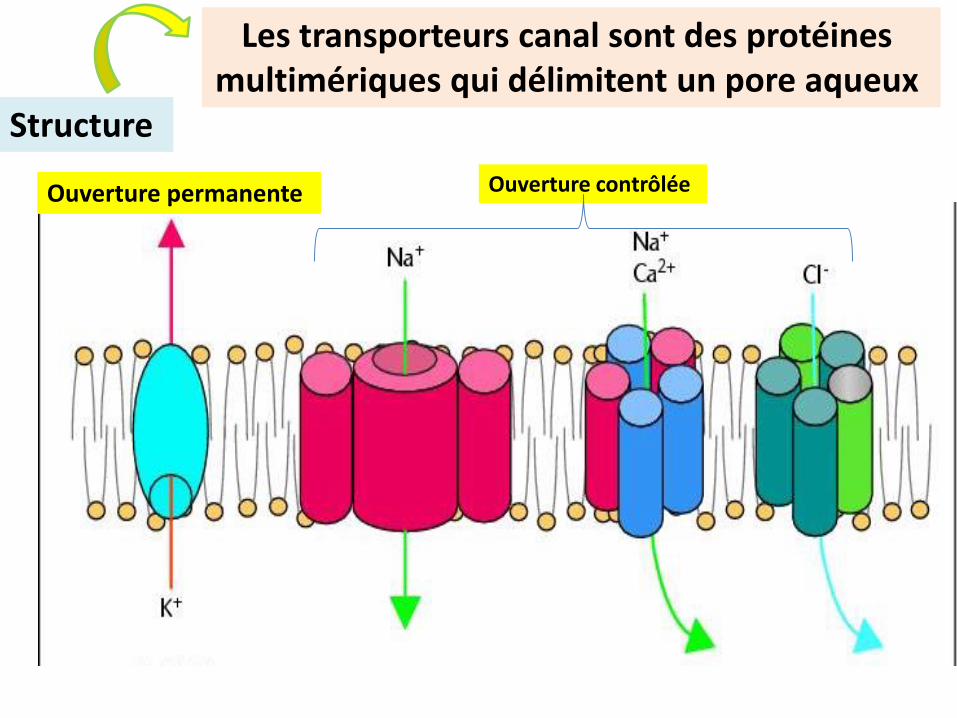
Les transporteurs canal sont des protéines multimériques qui délimitent un pore aqueux
Ouverture permanente
Structure
Ouverture contrôlée

Reconstitution tridimensionnelle des canaux ioniques
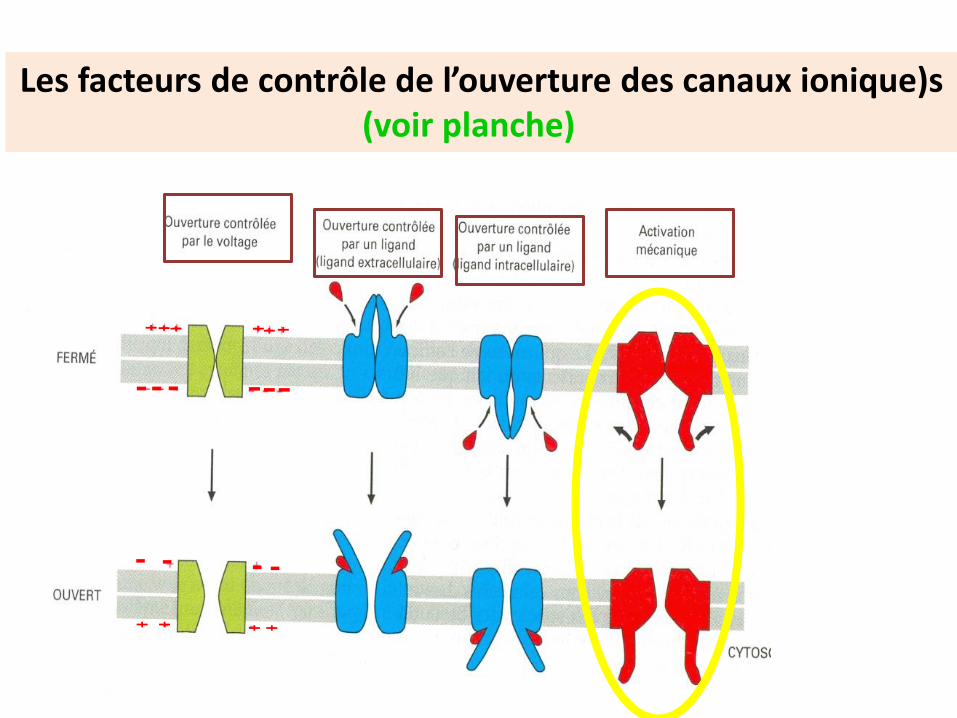
Les facteurs de contrôle de l’ouverture des canaux ionique)s(voir planche)

Cas des canaux mécano-dépendants
Répondent à la déformation membranaire. Ex: cerveau , oreille interne
N’est pas à retenir

Le pore du canal est sélectif pour chaque ion. les ions passent suivant leur gradient électrochimique

Canal à ouverture contrôlée par un ligand Le ligand peut être externe (neurotransmetteur) ou interne
(messagers secondaires )

Les Canaux à ouverture contrôlée par un ligand sont perméables à des cations ( Rn-Ach) ou des anions (R- GABA)
Ligand c’est l’ Ach, /nicotine& l’Ion c’est le Na+
Ligand c’est le GABA, & l’Ion c’est le Cl-
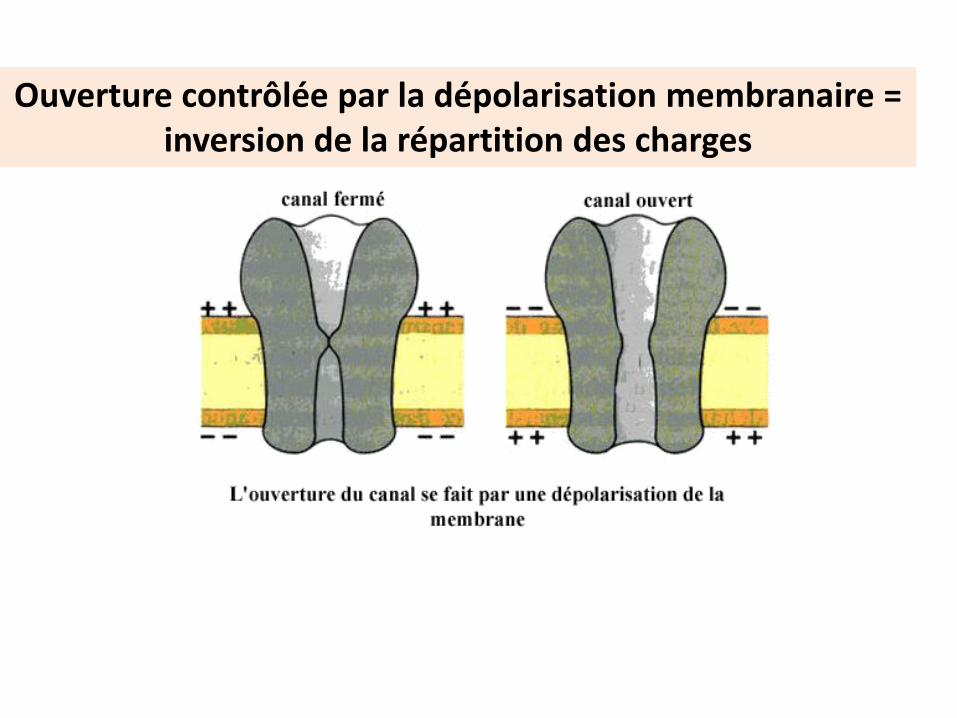
Ouverture contrôlée par la dépolarisation membranaire =inversion de la répartition des charges

Intervention des canaux ioniques dans le déclenchement de la contraction musculaire
(Voir COMMUNICATION CELLULAIRE )

Dans la synapse neuro-musculaire , les canaux ligands dépendants et les canaux voltage dépendants sont activés progressivement
Tubule T

l’ouverture séquentielle des canaux ioniques assure la propagationde l’onde de dépolarisation de la membrane
neuronale à la membrane musculaire

Mécanisme électrique de la contraction musculaire

La fixation de L’Ach sur son récepteur ouvre un canal Na+et dépolarise localement la membrane

L’inversion locale des charges induit l’ouverture des canauxvoltage dépendants d’où propagation de l’influx
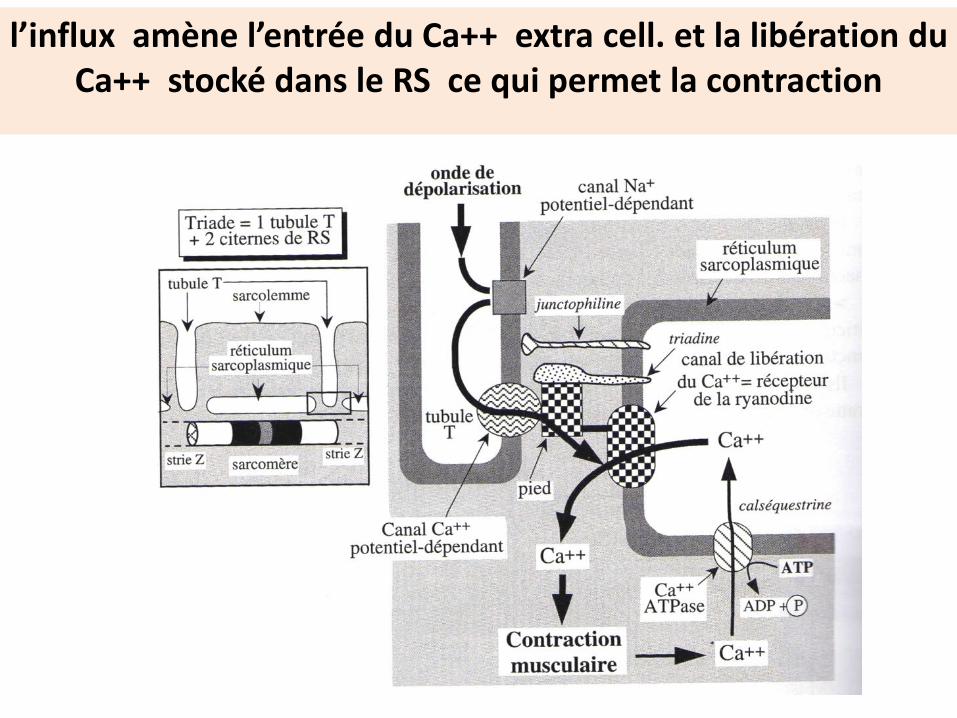
l’influx amène l’entrée du Ca++ extra cell. et la libération du Ca++ stocké dans le RS ce qui permet la contraction

Les transports perméatifs actifs
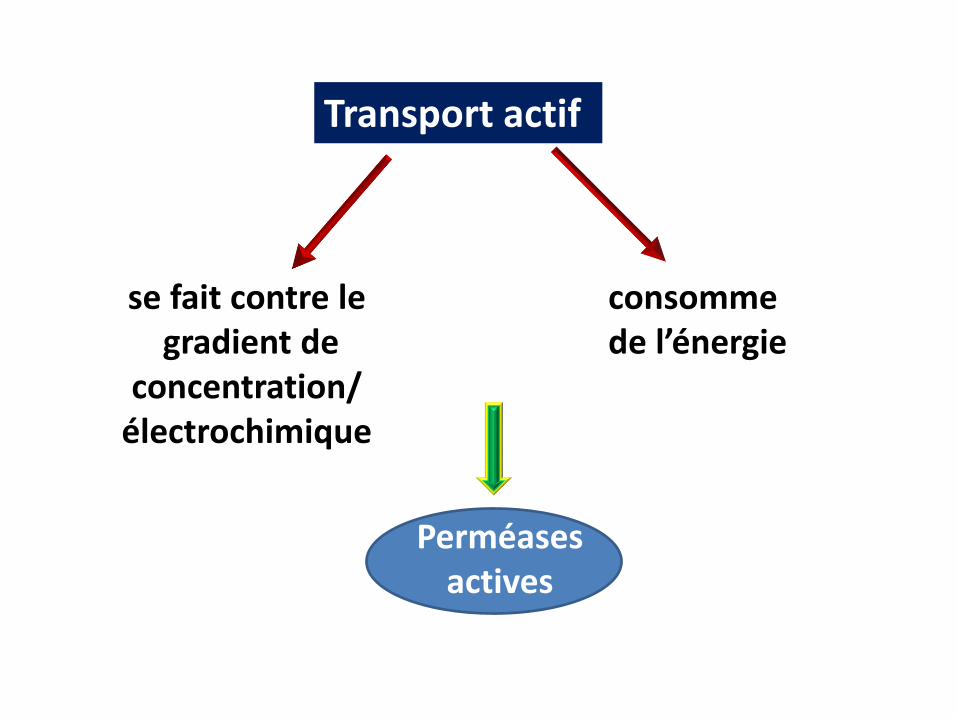
Transport actif
se fait contre legradient de
concentration/ électrochimique
consomme de l’énergie
Perméases actives

Perméases actives
Transport actif Primaire
Transport actif Secondaire
protéines enzymes =ATPases = Pompes
ATPase Ca++ ATPase H+
Energie ATP
Perméases deco-transport
Energie Gradient Na+
Symport Antiport
Ex: Glucose /Na+ATPase Na+/K+Ca++/Na+
Dekar

Transports actifs Iaire et IIaire

Transport actif primaire

Cas de la pompe ou ATP ase Na+ / K+ dépendante
Structure (voir P 83)
Site d’hydrolyse d’ATP site de phosphorylation 3 Sites de fixation de Na + 2 sites de fixation de K+ /site de fixation d’un inhibiteur des sites K+ :l’Ouabaïne

L’ATP ase contrôle les canaux de fuite pour le maintien de la différence de répartition des charges
responsable du potentiel de repos membranaire

La phosphorylation de l’ATP ase induitla libération de 3Na+ dans le MEC
Na+
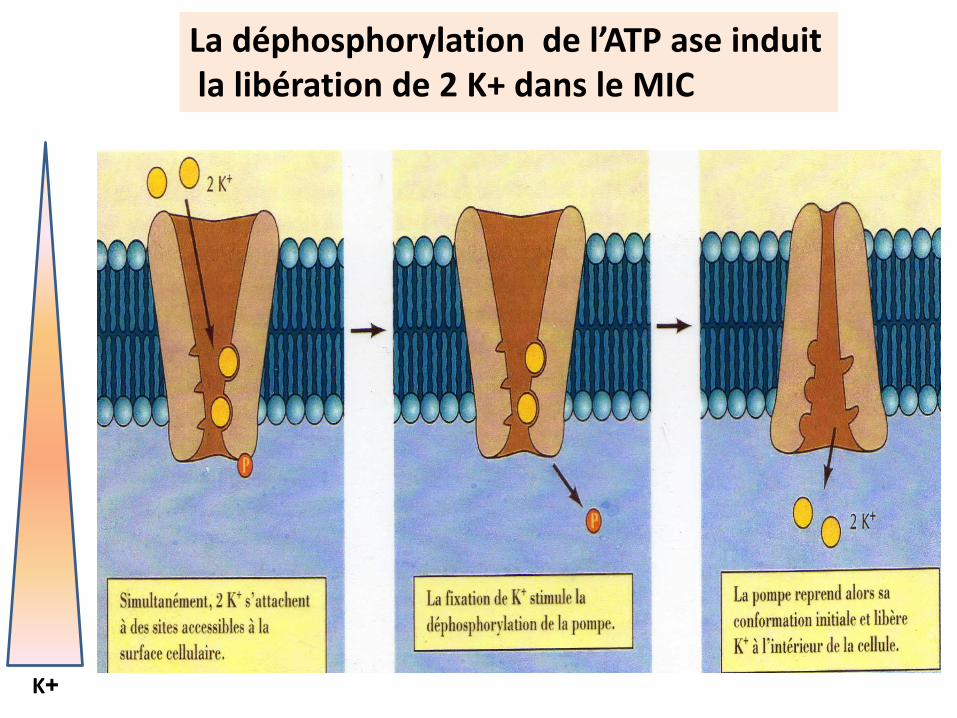
La déphosphorylation de l’ATP ase induitla libération de 2 K+ dans le MIC
K+

L’OUABAINE bloque l’entrée du K+ et donc le fonctionnement de la pompe voir (P.83)

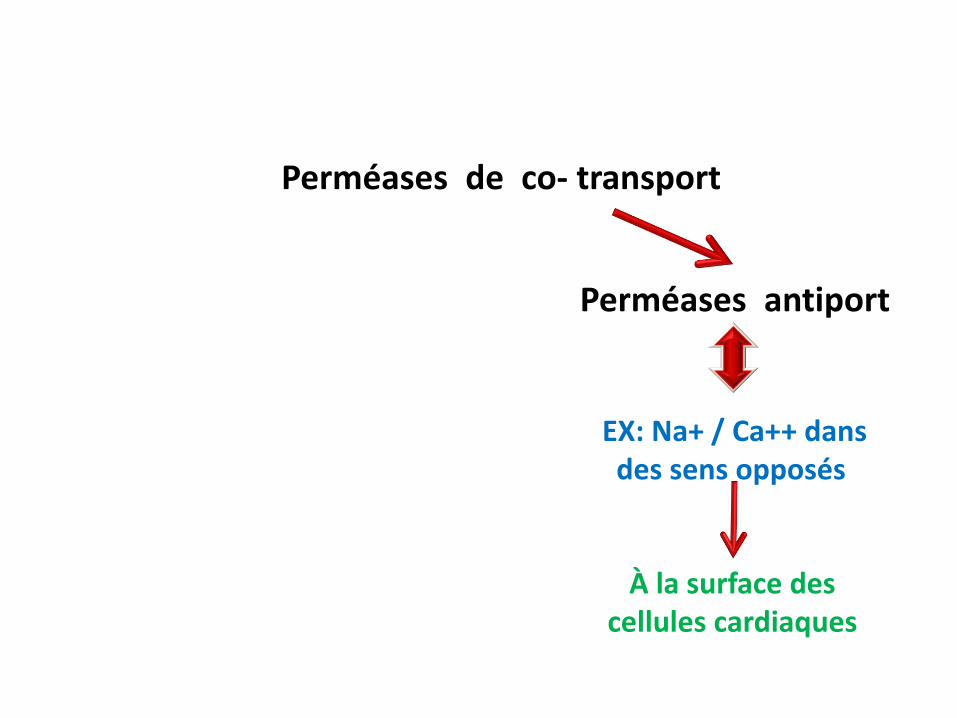
transport actif secondaire
Perméases de co- transport
Perméases - symport Perméases antiport
EX: Na+ / Glucose dansle même sens
EX: Na+ / Ca++ dansdes sens opposés
Pôle apical des entérocytes et des cellules rénales
À la surface des cellules cardiaques
Dekar

Perméases de co- transport
Perméases - symport
EX: Na+ / Glucose dansle même sens
Pôle apical des entérocytes et des cellules rénales

l’entrée du glucose de la lumière vers les cellules intestinales (gauche) et rénales (droite) est active

Entrée du glucose par le pôlle apical en utilisant unsystème de co-transport symport:
la Sglut Na=/Glucose (voir Planche )

Na+
Glucose
MEC
MIC

Compartimentation des transporteurs du glucose Dans les cellules épithéliales à microvillosités
Perméases de co- transport
Perméases antiport
EX: Na+ / Ca++ dansdes sens opposés
À la surface des cellules cardiaques

La régulation de la force de contraction des cellules cardiaques est assurée par la perméase de co transport antiport Na+/ Ca++

Une des thérapies de l’insuffisance cardiaque est le blocage du Co-transport Na+/Ca++ à travers l’inhibition de la pompe Na+/K+
Ouabaine ATPase
Arrêt du transport actif II aire
Pas de gradient Na+
Pas de sortie
du Ca++
Disponibilité intracellulaire
du Ca++
Augmentation de la force de contraction du muscle cardiaque
l’insuffisance cardiaque Traitement cardiotonique
Dekar

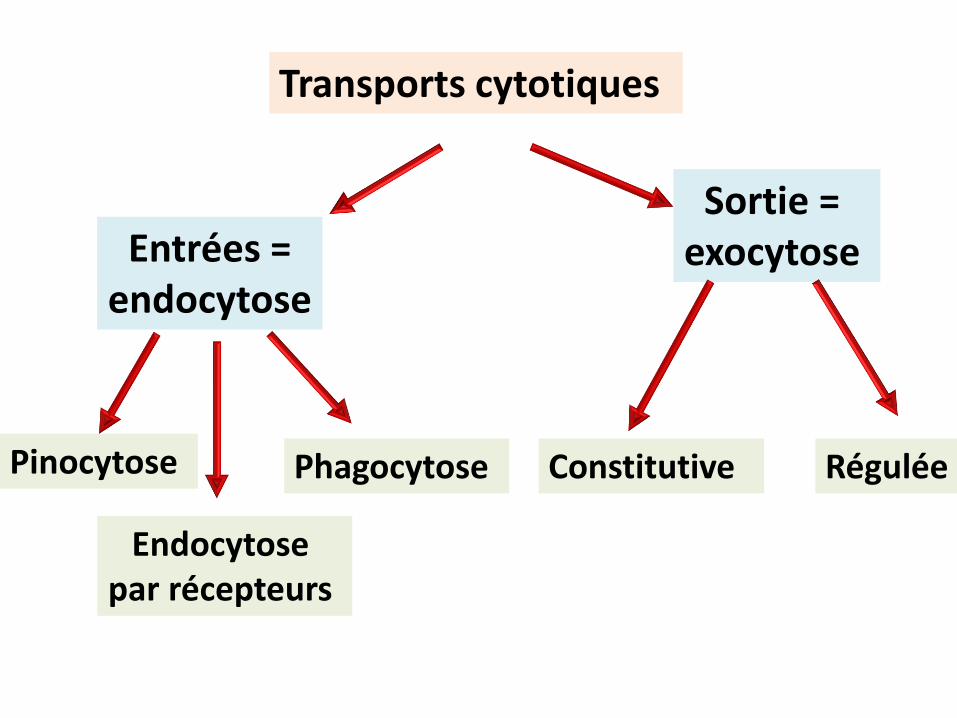
Transports cytotiques

Transports cytotiques
déforment la membrane utilisent le cytosquelette consomment de l’énergie emballage des produits dans des compartiments membranaires ( vésicules lisses/ recouvertes) ou vacuoles
caractéristiques
Transports cytotiques
Phagocytose Pinocytose
Sortie = exocytoseEntrées =
endocytose
Endocytosepar récepteurs
Régulée Constitutive

Endocytose & exocytose

Endocytose

Phagocytose Pinocytose
entrées =endocytoses
Endocytosepar récepteurs

La pinocytose concerne l’entrée de petites molécules dissoutes dans le liquide extracellulaire
(Voir Schéma 18, P 59)

4étapes Piégeage et invagination
de la mb
Pincement des bordsde la mb
Formation d’une vésicule lisse
(100 nm de Ø)
fusion avec l’endosome

Notion d’endosome
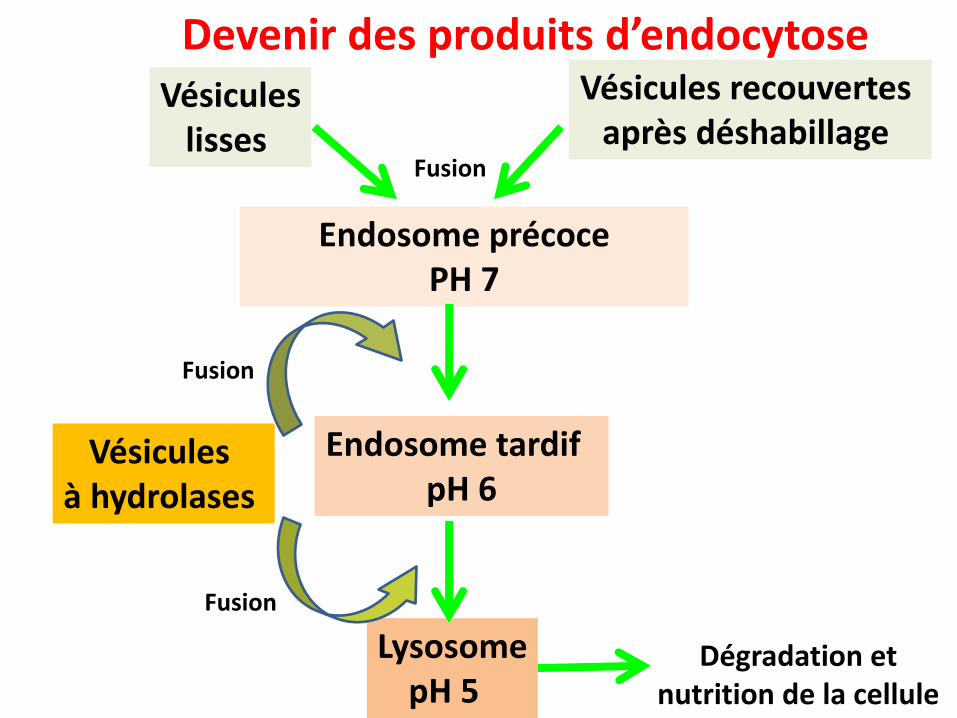
Vésicules à hydrolases
Endosome précoce PH 7
Vésiculeslisses
Vésicules recouvertes après déshabillage
LysosomepH 5
Endosome tardif pH 6
Fusion
Fusion
Fusion
Devenir des produits d’endocytose
Dégradation et nutrition de la cellule

Les vesicules à hydrolases sont produites par le réseau trans Golgi et destinées aux endosomes

La phagocytose

La Phagocytose concerne l’entrée de complexes moléculaires solides dans les cellules phagocytaires
(Voir Schéma 19, P 59)
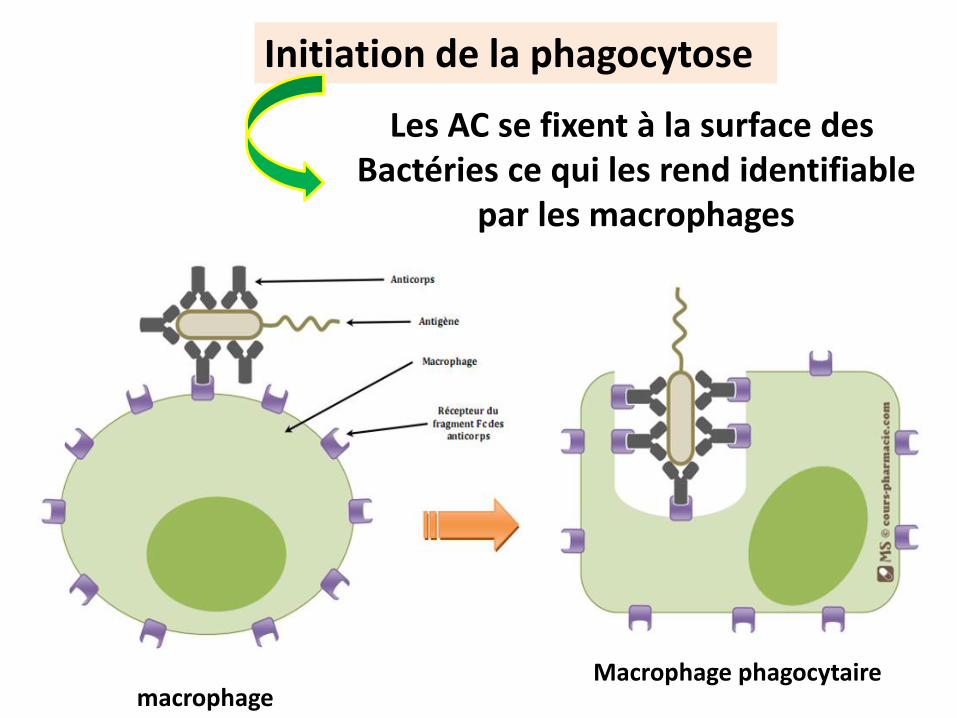
macrophage
Initiation de la phagocytose
Les AC se fixent à la surface des Bactéries ce qui les rend identifiable
par les macrophages
Macrophage phagocytaire

Étapes de de la phagocytose
SéquestrationVoiles hyaloplasmiques
Hydrolyse acidedu contenu
Exocytose des déchets

Le phagosome = vacuole de phagocytose

Endocytose par récepteursCaractéristiques
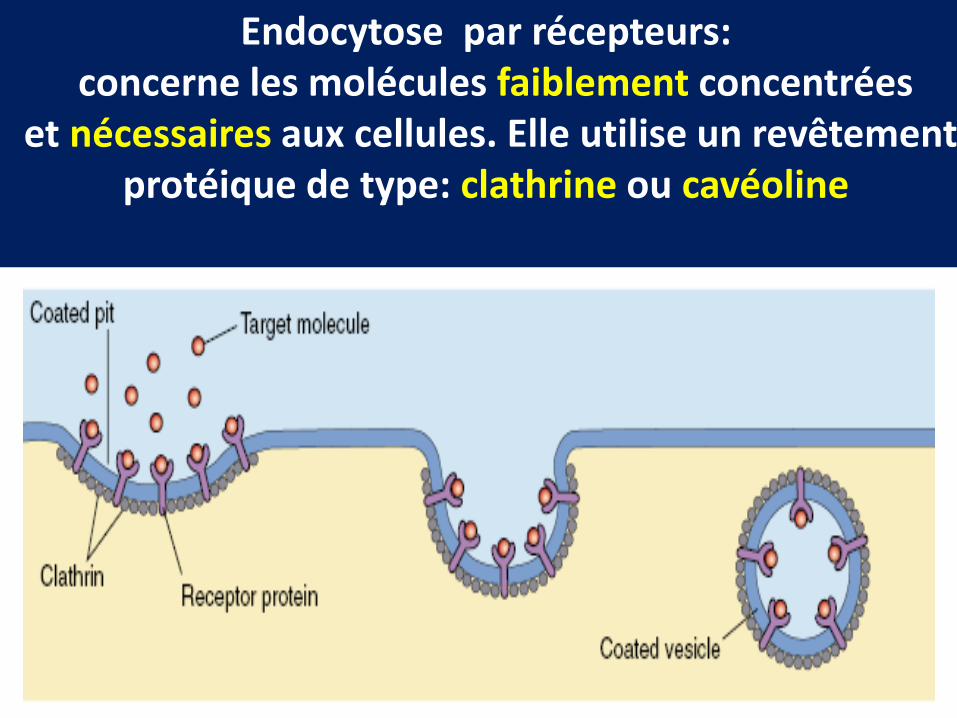
Endocytose par récepteurs:concerne les molécules faiblement concentrées
et nécessaires aux cellules. Elle utilise un revêtementprotéique de type: clathrine ou cavéoline
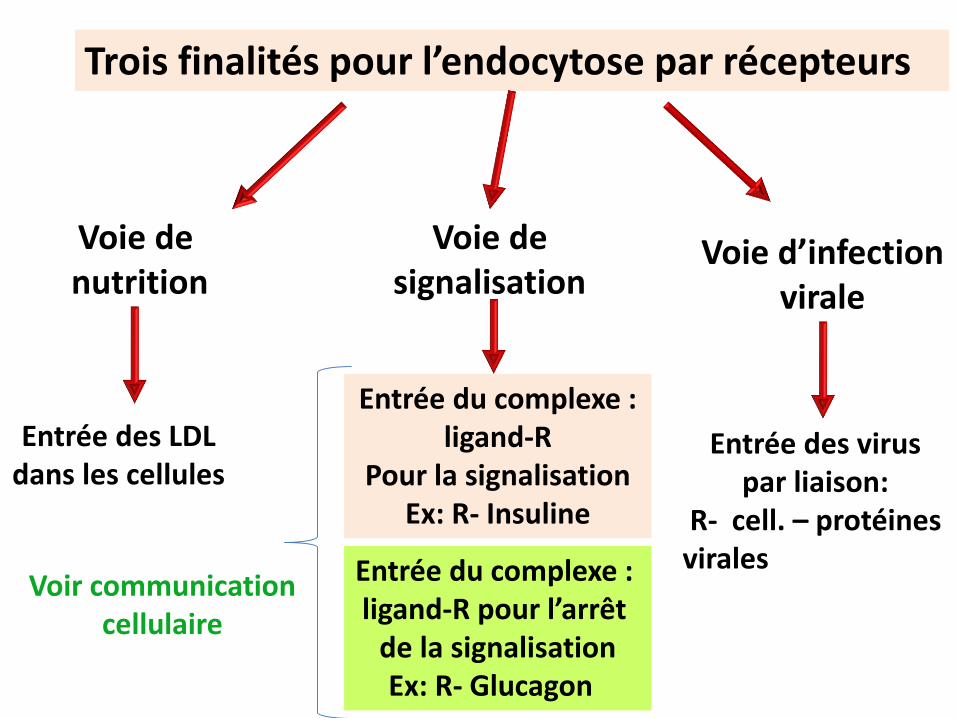
Trois finalités pour l’endocytose par récepteurs
Voie de nutrition
Voie designalisation
Voie d’infection virale
Entrée des LDL dans les cellules
Entrée des virus par liaison:
R- cell. – protéines virales
Entrée du complexe : ligand-R
Pour la signalisationEx: R- Insuline
Entrée du complexe : ligand-R pour l’arrêt
de la signalisationEx: R- Glucagon
Voir communicationcellulaire
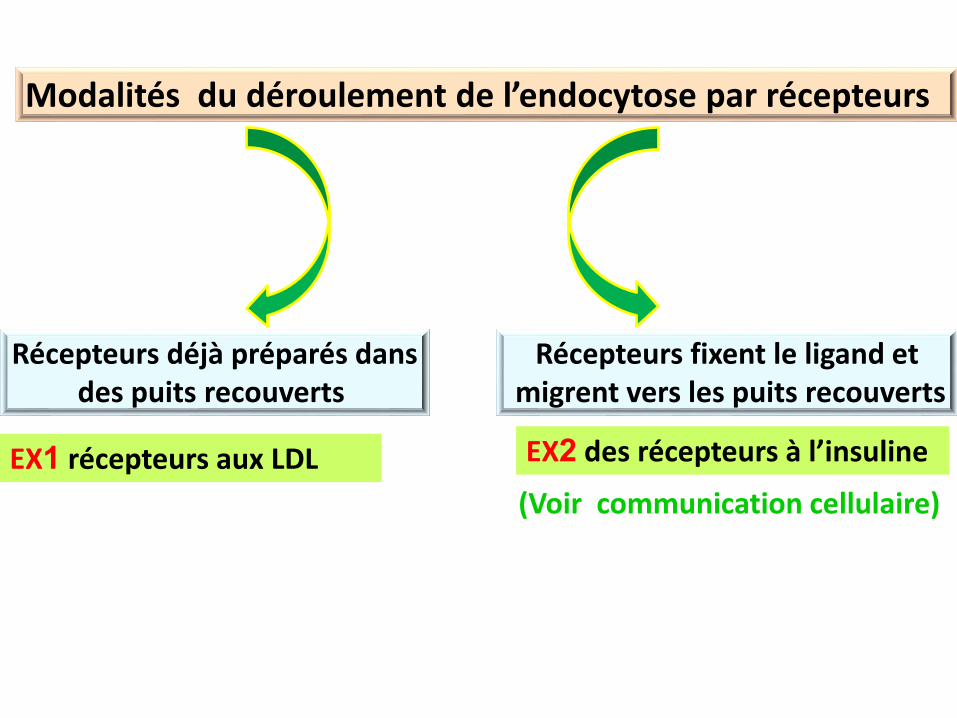
Modalités du déroulement de l’endocytose par récepteurs
Récepteurs déjà préparés dansdes puits recouverts
Récepteurs fixent le ligand etmigrent vers les puits recouverts
EX1 récepteurs aux LDL EX2 des récepteurs à l’insuline
(Voir communication cellulaire)
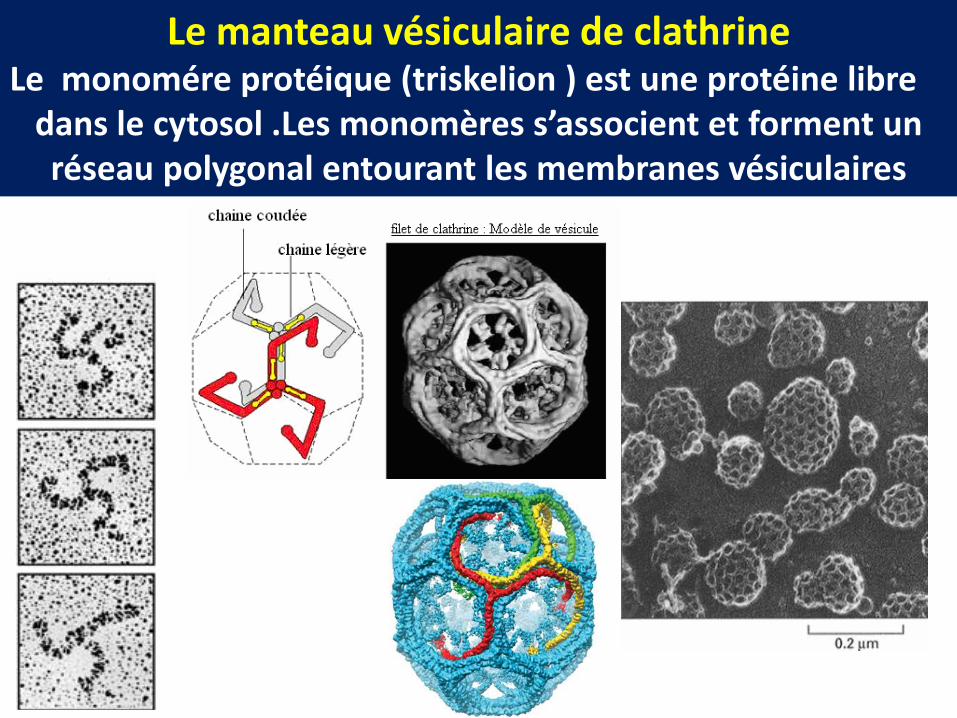
Le manteau vésiculaire de clathrineLe monomére protéique (triskelion ) est une protéine libre
dans le cytosol .Les monomères s’associent et forment un réseau polygonal entourant les membranes vésiculaires
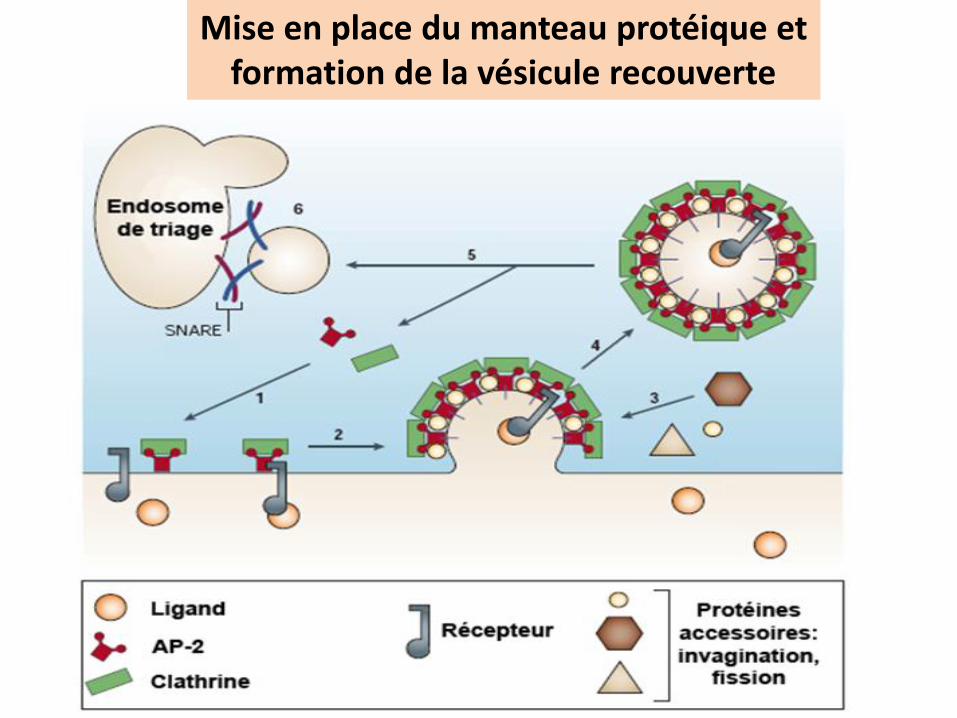
Mise en place du manteau protéique etformation de la vésicule recouverte

Etapes de l’internalisation des vésicules recouvertes
1
2
3
4

La fixation du ligandentraine la migration du complexe R-ligand dans des dépressions de la membrane:
les puits recouverts

Les complexes récepteur- ligand migrent vers les puits recouverts EX2

EX: endocytose des particules de LDL: : entrée du cholestérol dans les
différents types cellulaires (Voir planche tirage)
Endocytose par récepteurs :voie de nutrition
EX 1

Composition moléculaire de la particule de LDL
APOB
Esters de cholestérol
Monocouche de phospholipides
Molécule de cholestérol
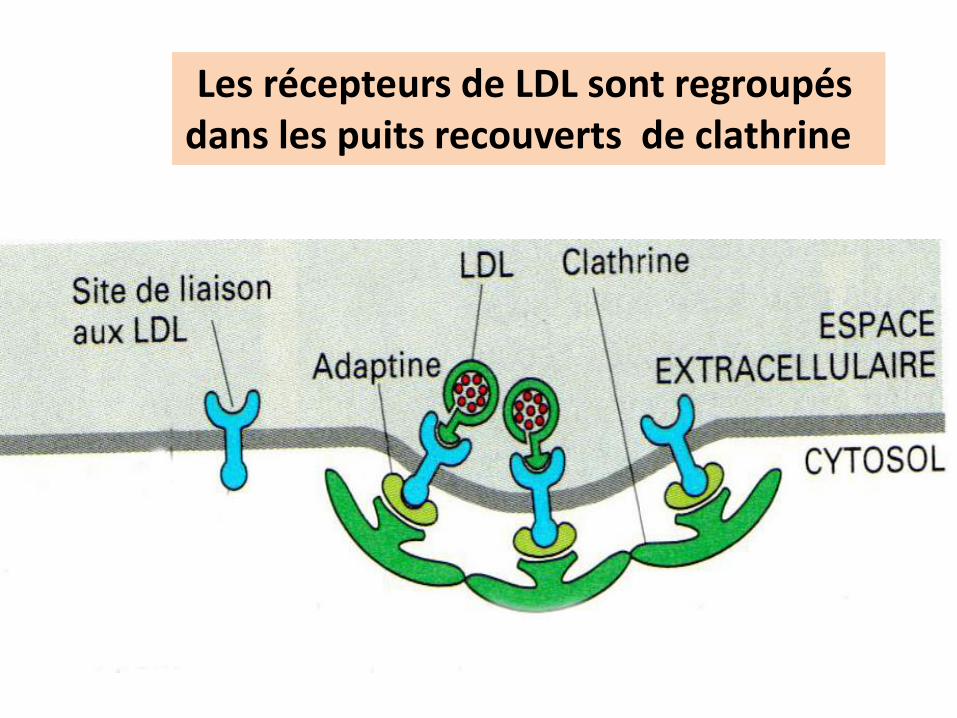
Les récepteurs de LDL sont regroupés dans les puits recouverts de clathrine

Particule de LDLMEC
MIC
Vésicule recouverte De clathrine
Vésicule lisse
Vésicule de recyclagedes R-LDL
Endosome tardif Découplage R-LDL
pH= 6
ATPase H+
Endosome précoce pH= 7
Perte de clathrine
Exocytose
Lysosome: égradation des LDL et libération du choléstérol
Vésicules àhydrolases
Détail d’une vésicules à hydrolases
Récepteur LDL
Cholésterol
pH= 5Perméased’entréeH+
H+
Acidesaminés
Oses
Mécanisme de l’’internalisation des particules de LDL: utilisation du cholestérol par tous les types cellulaires

l’hypercholestérolemie familiale: une pathologie liée à l’altération des récepteurs de LDL

Devenir des molécules internalisées dans la cellule

L’endocytose par récepteurs: Voie de signalisation
Voir communication intercellulaire
Entrée du complexe : ligand-R
Pour la signalisationEx: R- Insuline
Entrée du complexe : ligand-R pour l’arrêt de la signalisation
Ex: R- Glucagon

L’endocytode par récepteurs: voie d’infection virale

Endocytose par récepteurs indépendante de la clathrine/Dépendante de la cavéoline


EXOCYTOSE

Exocytose

Exocytose = Sortie
Régulée Constitutive
Permanente Compensatrice de lapinocytose
A la demande Obéit à un signal

Deux voies d’exocytose: constitutive & régulée

Fusion membranaire dans l’exocytose
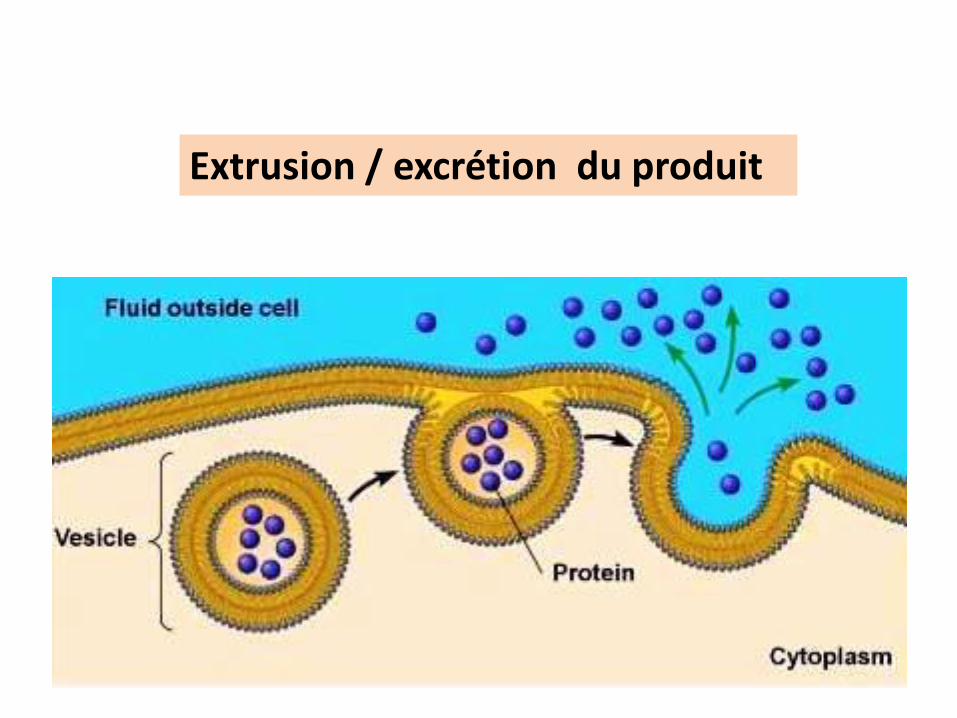
Extrusion / excrétion du produit


Fin