Anatomie de La Rétine 01
14
Anatomie de la rétine F. Behar-Cohen, L. Kowalczuk, N. Keller, M. Savoldelli, F. Azan, J.-C. Jeanny La neurorétine est une unité fonctionnelle du système nerveux central assurant la conversion d’un signal lumineux en un influx nerveux. La rétine mature de mammifère est composée de deux tissus : la rétine neurale et l’épithélium pigmentaire de la rétine. La neurorétine est un tissu d’origine neuroectodermique, dérivé du diencéphale, stratifié et composé de six types de cellules neuronales (deux types de photorécepteurs : cônes et bâtonnets, cellules horizontales, bipolaires, amacrines et ganglionnaires) et de trois types de cellules gliales (cellules gliales de Müller, astrocytes et cellules microgliales). L’existence des barrières hématorétiniennes interne et externe et des jonctions intrarétiniennes rend compte de la finesse de régulation des échanges de la rétine avec la circulation et au sein de la rétine elle-même. Les données anatomiques de la rétine humaine sont encore imparfaitement décrites. Des études structurales sont indispensables pour compléter les analyses moléculaires et fonctionnelles. © 2009 Elsevier Masson SAS. Tous droits réservés. Mots clés : Rétine ; Anatomie de la rétine ; Macula ; Glie ; Photorécepteurs ; Vaisseaux de la rétine Plan ¶ Généralités 1 ¶ Origine embryologique de la rétine 2 Développement de la rétine 2 Contrôle de la détermination et différenciation des progéniteurs de la rétine 3 Cellules souches de la rétine 3 ¶ Quelques notions d’anatomie comparée 4 Photorécepteurs 4 Existence d’une zone rétinienne centrale différenciée 5 Tapetum lucidum 5 ¶ Anatomie macroscopique 5 Ora serrata 5 Épaisseur rétinienne 6 Papille optique 6 Zones anatomiques rétiniennes distinctes 7 ¶ Vascularisation rétinienne 7 Système vasculaire rétinien 7 Système vasculaire choroïdien 8 ¶ Anatomie microscopique 9 Structure histologique générale 9 Épithélium pigmentaire de la rétine 9 Photorécepteurs 10 Couche plexiforme externe 11 Couche nucléaire interne 11 Couche plexiforme interne 12 Couche des cellules ganglionnaires 12 Couche des fibres optiques 12 Membrane limitante interne 12 Cellules gliales de la rétine 12 ¶ Conclusion 13 ■ Généralités La neurorétine est une unité fonctionnelle du système nerveux central (SNC), qui permet la conversion d’un signal lumineux en un influx nerveux. La rétine mature de mammifère est composée de deux tissus : la rétine neurale et l’épithélium pigmentaire de la rétine (EPR). Ce dernier est constitué d’une monocouche de cellules épithé- liales et se caractérise par la présence de mélanosomes impliqués dans la synthèse et le stockage des pigments de mélanine. La neurorétine est un tissu d’origine neuroectodermique, dérivé du diencéphale, stratifié et composé de six types de cellules neuronales (deux types de photorécepteurs : cônes et bâtonnets, cellules horizontales, bipolaires, amacrines et ganglionnaires) et de trois types de cellules gliales (cellules gliales de Müller, astrocytes et cellules microgliales). Tous sont issus des mêmes progéniteurs neuronaux et chacun de ces types cellulaires est localisé dans une couche rétinienne particulière (Fig. 1). La neurorétine est ainsi organisée en couches de noyaux cellulaires alternant avec des couches de synapses et de prolongements de cellules gliales. • La couche nucléaire externe est exclusivement composée des corps cellulaires des photorécepteurs (cônes et bâtonnets) (Fig. 2). • La couche plexiforme externe est le lieu de formation des connexions synaptiques entre les photorécepteurs et les cellules bipolaires. • La couche nucléaire interne est composée des noyaux de trois types de neurones, les cellules horizontales, les cellules bipolaires et les cellules amacrines, et des noyaux de cellules gliales de Müller. • La couche plexiforme interne est le lieu de formation des connexions synaptiques entre les axones des cellules bipolai- res et les dendrites des cellules ganglionnaires. • La couche des cellules ganglionnaires, la plus interne, contient les noyaux des cellules ganglionnaires. ¶ 21-003-C-40 1 Ophtalmologie
-
Upload
liliana-urse-praja -
Category
Documents
-
view
22 -
download
2
description
Anatomie de La Rétine
Transcript of Anatomie de La Rétine 01

Anatomie de la rétine
F. Behar-Cohen, L. Kowalczuk, N. Keller, M. Savoldelli, F. Azan, J.-C. Jeanny
La neurorétine est une unité fonctionnelle du système nerveux central assurant la conversion d’un signallumineux en un influx nerveux. La rétine mature de mammifère est composée de deux tissus : la rétineneurale et l’épithélium pigmentaire de la rétine. La neurorétine est un tissu d’origine neuroectodermique,dérivé du diencéphale, stratifié et composé de six types de cellules neuronales (deux types dephotorécepteurs : cônes et bâtonnets, cellules horizontales, bipolaires, amacrines et ganglionnaires) et detrois types de cellules gliales (cellules gliales de Müller, astrocytes et cellules microgliales). L’existence desbarrières hématorétiniennes interne et externe et des jonctions intrarétiniennes rend compte de la finessede régulation des échanges de la rétine avec la circulation et au sein de la rétine elle-même. Les donnéesanatomiques de la rétine humaine sont encore imparfaitement décrites. Des études structurales sontindispensables pour compléter les analyses moléculaires et fonctionnelles.© 2009 Elsevier Masson SAS. Tous droits réservés.
Mots clés : Rétine ; Anatomie de la rétine ; Macula ; Glie ; Photorécepteurs ; Vaisseaux de la rétine
Plan
¶ Généralités 1
¶ Origine embryologique de la rétine 2Développement de la rétine 2Contrôle de la détermination et différenciation des progéniteursde la rétine 3Cellules souches de la rétine 3
¶ Quelques notions d’anatomie comparée 4Photorécepteurs 4Existence d’une zone rétinienne centrale différenciée 5Tapetum lucidum 5
¶ Anatomie macroscopique 5Ora serrata 5Épaisseur rétinienne 6Papille optique 6Zones anatomiques rétiniennes distinctes 7
¶ Vascularisation rétinienne 7Système vasculaire rétinien 7Système vasculaire choroïdien 8
¶ Anatomie microscopique 9Structure histologique générale 9Épithélium pigmentaire de la rétine 9Photorécepteurs 10Couche plexiforme externe 11Couche nucléaire interne 11Couche plexiforme interne 12Couche des cellules ganglionnaires 12Couche des fibres optiques 12Membrane limitante interne 12Cellules gliales de la rétine 12
¶ Conclusion 13
■ Généralités
La neurorétine est une unité fonctionnelle du systèmenerveux central (SNC), qui permet la conversion d’un signallumineux en un influx nerveux.
La rétine mature de mammifère est composée de deux tissus :la rétine neurale et l’épithélium pigmentaire de la rétine (EPR).Ce dernier est constitué d’une monocouche de cellules épithé-liales et se caractérise par la présence de mélanosomes impliquésdans la synthèse et le stockage des pigments de mélanine.
La neurorétine est un tissu d’origine neuroectodermique,dérivé du diencéphale, stratifié et composé de six types decellules neuronales (deux types de photorécepteurs : cônes etbâtonnets, cellules horizontales, bipolaires, amacrines etganglionnaires) et de trois types de cellules gliales (cellulesgliales de Müller, astrocytes et cellules microgliales). Tous sontissus des mêmes progéniteurs neuronaux et chacun de ces typescellulaires est localisé dans une couche rétinienne particulière(Fig. 1). La neurorétine est ainsi organisée en couches denoyaux cellulaires alternant avec des couches de synapses et deprolongements de cellules gliales.• La couche nucléaire externe est exclusivement composée des
corps cellulaires des photorécepteurs (cônes et bâtonnets)(Fig. 2).
• La couche plexiforme externe est le lieu de formation desconnexions synaptiques entre les photorécepteurs et lescellules bipolaires.
• La couche nucléaire interne est composée des noyaux de troistypes de neurones, les cellules horizontales, les cellulesbipolaires et les cellules amacrines, et des noyaux de cellulesgliales de Müller.
• La couche plexiforme interne est le lieu de formation desconnexions synaptiques entre les axones des cellules bipolai-res et les dendrites des cellules ganglionnaires.
• La couche des cellules ganglionnaires, la plus interne,contient les noyaux des cellules ganglionnaires.
¶ 21-003-C-40
1Ophtalmologie

■ Origine embryologiquede la rétine
Développement de la rétine
Au cours du développement embryonnaire, les différentesstructures oculaires découlent d’interactions entre trois tissus : leneuroectoderme, l’ectoderme et le mésenchyme, ce dernierétant dérivé des crêtes neurales et s’interposant entre les deux
premiers. La rétine est un dérivé purement ectoblastique,d’origine neuroectodermique. Aux environs du jour 22 dedéveloppement, quand l’embryon présente huit paires desomites et qu’il mesure de l’ordre de 2 mm, deux évaginationslatérales du diencéphale, prédéterminées lors de la gastrulation,vont donner naissance aux vésicules optiques, qui mettent encontact le neuroépithélium encéphalique et l’ectoderme de latête. Des interactions entre ces deux tissus naît un épaississe-ment de l’ectoderme, la placode cristallinienne, qui s’invagineen vésicule cristallinienne. Dans le même temps, les vésiculesoptiques s’invaginent en doigt de gant pour former les cupulesoptiques constituées de deux feuillets. Après 6 semaines dedéveloppement, tandis que le feuillet externe des cupulesoptiques évolue en rétine pigmentaire, le feuillet interne vafournir les différentes couches de la neurorétine ou rétinesensorielle (Fig. 3).
Trois phases, dont les transitions se chevauchent dans letemps en fonction des types cellulaires, sont responsables de lamise en place de l’architecture de la rétine.
Étape de prolifération des précurseursneuroectodermiques
Les cellules précurseurs, en division symétrique puis progres-sivement asymétrique, sont localisées dans la région la plusexterne de la rétine, la zone ventriculaire, adjacente au futurEPR. Après leur mitose terminale, les précurseurs migrent vers laface interne (côté vitré) pour y prendre leur position définitiveet se différencier en neurones.
Étape de migration des cellules en différenciation
Lors de cette étape, les différentes couches cellulaires appa-raissent. La première couche à s’individualiser est celle descellules ganglionnaires, dont les précurseurs sont les premiers àcesser de se diviser. Viennent ensuite les cellules horizontales etamacrines, suivies par les cellules bipolaires et les cellules glialesde Müller. Cet arrêt de la prolifération, couplé à la migration età la différenciation des cellules précurseurs, s’effectue schémati-quement selon deux gradients : un gradient antéropostérieur, de
Figure 1. Image en microscopie optique d’une coupe semi-fine derétine de souris adulte. Coloration au bleu de toluidine. Ch : choroïde ;CCG : couche des cellules ganglionnaires ; CNE : couche nucléaire ex-terne ; CNI : couche nucléaire interne ; CPE : couche plexiforme externe ;CPI : couche plexiforme interne ; EPR : épithélium pigmenté de la rétine ;FN : fibres nerveuses ; S : sclère ; SE : segments externes des photorécep-teurs ; SI : segments internes des photorécepteurs.
Figure 2. Coupe semi-fine de la région périmaculaire d’une rétinehumaine. Coloration au bleu de toluidine. Cette coupe montre la couchedes noyaux des photorécepteurs cônes (C) et bâtonnets (B), ainsi que lamembrane limitante externe (2) et la couche des fibres de Henle (FH). 1.Couche nucléaire externe ; 2. limitante externe ; 3. segments internes ; 4.segments internes.
1
2
3
A
1
2
B
5
4
6
C
89
10
7
D
12
13141516
11
E
Figure 3. Schéma simplifié de la formation de l’œil au cours du déve-loppement embryonnaire.A. Évagination de la vésicule optique (1) à partir du diencéphale.B, C. Mise en place successivement de la placode cristallinienne (2) et dela cupule optique (6).D, E. Formation du cristallin (14) et de la rétine (rétine neurale : 13 etépithélium pigmentaire de la rétine : 12). 3. Neuroderme ; 4. coucheexterne ; 5. couche interne ; 7. tractus optique ; 8. vésicule cristalli-nienne ; 9. future cornée ; 10. espace intrarétinien ; 11. futur nerf opti-que ; 15. cornée ; 16. corps vitré. D’après Furelaud G et Bonnet G. L’œil :structure, origine, propriétés physiques.
21-003-C-40 ¶ Anatomie de la rétine
2 Ophtalmologie

la face interne (cellules ganglionnaires) vers la face externe(photorécepteurs), et un gradient centropériphérique, du nerfoptique vers l’iris.
Étape de différenciation terminale
Elle comprend la formation des synapses, des segmentsinternes puis externes des photorécepteurs. Globalement, cesétapes se retrouvent dans le même ordre chronologique chez laplupart des espèces, même si elles sont imbriquées les unes dansles autres et qu’il existe des différences notables dans l’appari-tion et la durée de la synaptogenèse (Fig. 3, 4). Les premièrescellules à apparaître sont les cellules ganglionnaires, puis ce sontles cellules horizontales et les cônes, ensuite les bâtonnets etcellules bipolaires. Les cellules gliales de Müller sont lesdernières à apparaître (Fig. 4) [1].
Contrôle de la déterminationet différenciation des progéniteursde la rétine
Pendant longtemps, une question essentielle a été de savoirsi l’apparition progressive et séquentielle dans le temps desdifférents types cellulaires rétiniens résultait de facteurs envi-ronnementaux et/ou des capacités intrinsèques des progéniteurs.Il apparaît à présent que la régulation temporelle du développe-ment rétinien (prolifération des progéniteurs, neurogenèse etgliogenèse) s’opère à la fois par des facteurs extrinsèques, quipeuvent être des facteurs de croissance, et par des facteursintrinsèques (généralement des facteurs de transcription). Eneffet, les progéniteurs passent successivement par différents étatsde compétence, ces derniers étant définis par la capacité qu’ontles progéniteurs à répondre à différents signaux externes pourproduire un ou des types cellulaires particuliers.
Chaque état de compétence est provisoire et caractérisé parl’expression d’une combinaison de différents facteurs detranscription intrinsèques limitant les capacités de différencia-tion des cellules. La progression d’un état de compétence ausuivant peut être due à des facteurs extrinsèques et/ou à unprogramme intrinsèque [2, 3]. Les régulateurs intrinsèques de ladifférenciation cellulaire rétinienne sont des gènes basic helix-loop-helix (bHLH) [4], qui sont séparés d’un point de vuefonctionnel en deux groupes : les répresseurs et les activateurs.Les bHLH répresseurs tels que Hes1 et Hes5 sont exprimés parles progéniteurs communs et inhibent la différenciation neuro-nale. Ils assurent le maintien des progéniteurs dans la rétine
embryonnaire et la différenciation gliale dans la rétine postna-tale. Quand les bHLH répresseurs sont diminués, les bHLHactivateurs, tels que Mash1 et Math5, favorisent la différencia-tion neuronale.
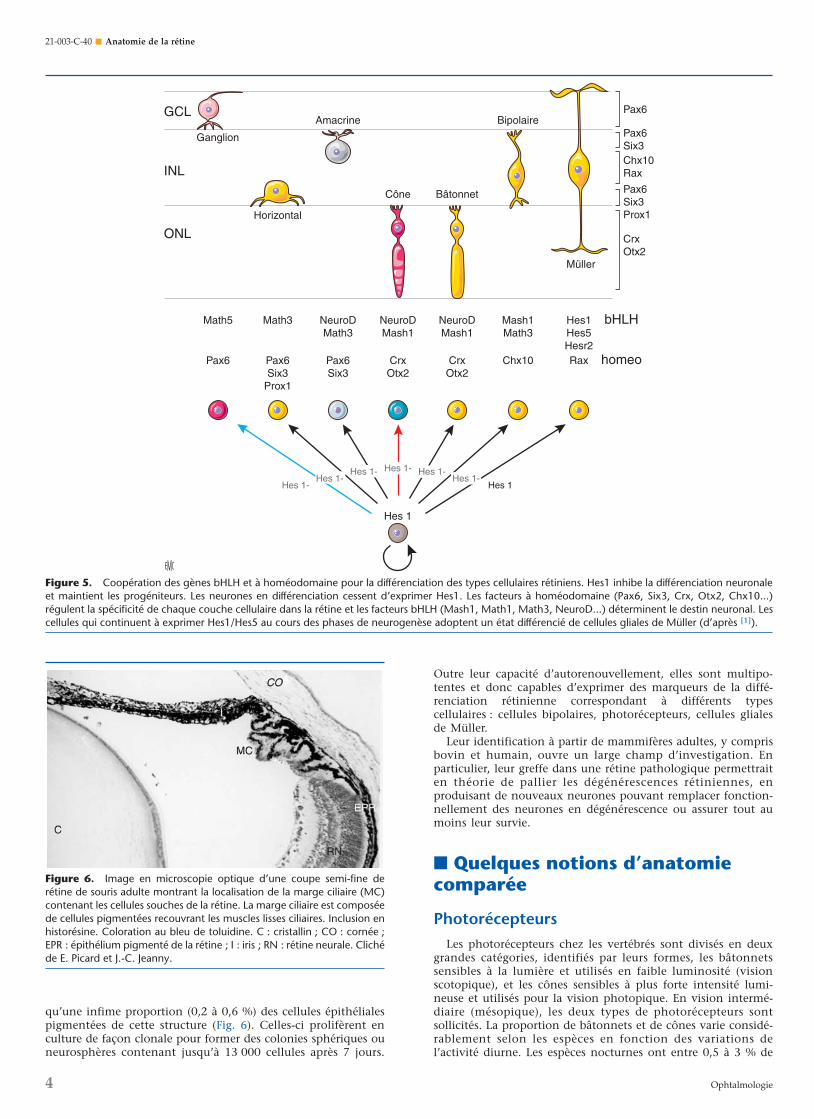
Concernant la neurogenèse, ce sont les gènes bHLH activa-teurs qui interviennent, mais ils nécessitent également l’inter-vention de gènes à homéodomaine. À titre d’exemple, ladifférenciation des cellules bipolaires nécessite l’intervention dugène à homéodomaine Chx10, qui dirige les cellules vers lacouche nucléaire interne, et des facteurs de transcription bHLHMash1 et Math3, qui leur confèrent les caractéristiques decellules bipolaires. Pour chaque type cellulaire rétinien, il y aune combinatoire de gènes bHLH et à homéodomaine spécifi-que [1] (Fig. 5).
Cellules souches de la rétine
La rétine contient également des cellules souches multipo-tentes, qui persistent à l’état adulte. Elles ont été caractérisées,il y a déjà très longtemps, chez certains vertébrés inférieurscomme les poissons téléostéens et les amphibiens. Localiséesprincipalement dans la zone marginale ciliaire (Fig. 6), ellesdonnent continuellement naissance à de nouveaux neurones etcellules gliales de Müller, et permettent ainsi une croissancecontinue de la rétine et sa réparation en cas de blessure [5]. Chezle poisson, il existe également des progéniteurs de type bâton-net, localisés dans la couche nucléaire interne, et qui ontconservé la capacité de reconstituer une rétine entière tout aulong de la vie de cette espèce [6]. La rétine des amphibienspossède aussi la capacité de se régénérer après rétinectomie, àpartir de l’EPR. Ce processus appelé « transdifférenciation » meten jeu successivement une dédifférenciation des cellules del’EPR, leur prolifération et leur redifférenciation pour formerune nouvelle neurorétine complète et fonctionnelle. Cettetransdifférenciation de l’EPR s’observe également chez le poulet,après ablation de la neurorétine, mais elle n’est possible quependant une brève période du développement embryonnaire(jusqu’à E5), et si on injecte simultanément du fibroblast growthfactor (FGF)-2 dans le vitré [7]. Ces données suggèrent donc unecertaine plasticité des cellules de l’EPR, qui toutefois s’amenuiseen fonction de l’âge et plus on monte dans l’échelle zoologique.L’existence de cellules souches ou progénitrices de la rétine n’aété démontrée chez les vertébrés supérieurs, comme les mam-mifères adultes, qu’en 2000 [8]. Elles sont localisées au niveau del’épithélium pigmentaire de la marge ciliaire et ne constituent
E10 12 14 16 18 B P1 3 5 7 9 11
Cônes
Bâtonnets
Cellules bipolaires
Cellules gliales de Müller
Cellules amacrines
Cellules horizontales
Cellules ganglionnaires
Figure 4. Développement de la rétine. Apparition des sept types cellulaires rétiniens chez la souris. Ceux-ci se différencient selon un ordre conservé entre lesdifférentes espèces : d’abord les cellules ganglionnaires, suivies par les cellules horizontales, les cônes, les cellules amacrines, les bâtonnets, les cellules bipolaireset les cellules gliales de Müller. E : âge embryonnaire (en jours) ; B : naissance ; P : âge après la naissance (en jours). D’après [1].
Anatomie de la rétine ¶ 21-003-C-40
3Ophtalmologie
qu’une infime proportion (0,2 à 0,6 %) des cellules épithélialespigmentées de cette structure (Fig. 6). Celles-ci prolifèrent enculture de façon clonale pour former des colonies sphériques ouneurosphères contenant jusqu’à 13 000 cellules après 7 jours.
Outre leur capacité d’autorenouvellement, elles sont multipo-tentes et donc capables d’exprimer des marqueurs de la diffé-renciation rétinienne correspondant à différents typescellulaires : cellules bipolaires, photorécepteurs, cellules glialesde Müller.
Leur identification à partir de mammifères adultes, y comprisbovin et humain, ouvre un large champ d’investigation. Enparticulier, leur greffe dans une rétine pathologique permettraiten théorie de pallier les dégénérescences rétiniennes, enproduisant de nouveaux neurones pouvant remplacer fonction-nellement des neurones en dégénérescence ou assurer tout aumoins leur survie.
■ Quelques notions d’anatomiecomparée
PhotorécepteursLes photorécepteurs chez les vertébrés sont divisés en deux
grandes catégories, identifiés par leurs formes, les bâtonnetssensibles à la lumière et utilisés en faible luminosité (visionscotopique), et les cônes sensibles à plus forte intensité lumi-neuse et utilisés pour la vision photopique. En vision intermé-diaire (mésopique), les deux types de photorécepteurs sontsollicités. La proportion de bâtonnets et de cônes varie considé-rablement selon les espèces en fonction des variations del’activité diurne. Les espèces nocturnes ont entre 0,5 à 3 % de
ONL
INL
GCL
Ganglion
Amacrine
Horizontal
Cône Bâtonnet
Müller
Bipolaire
Math5 NeuroDMath3
Math3 NeuroDMash1
NeuroDMash1
Hes1Hes5Hesr2
bHLHMash1Math3
Pax6 Pax6Six3
Pax6Six3
Prox1
CrxOtx2
Hes 1
Hes 1-
CrxOtx2
Rax homeoChx10
Pax6
Pax6Six3Chx10Rax
Pax6Six3Prox1
CrxOtx2
Hes 1-Hes 1- Hes 1-
Hes 1- Hes 1
Hes 1-
Figure 5. Coopération des gènes bHLH et à homéodomaine pour la différenciation des types cellulaires rétiniens. Hes1 inhibe la différenciation neuronaleet maintient les progéniteurs. Les neurones en différenciation cessent d’exprimer Hes1. Les facteurs à homéodomaine (Pax6, Six3, Crx, Otx2, Chx10...)régulent la spécificité de chaque couche cellulaire dans la rétine et les facteurs bHLH (Mash1, Math1, Math3, NeuroD...) déterminent le destin neuronal. Lescellules qui continuent à exprimer Hes1/Hes5 au cours des phases de neurogenèse adoptent un état différencié de cellules gliales de Müller (d’après [1]).
Figure 6. Image en microscopie optique d’une coupe semi-fine derétine de souris adulte montrant la localisation de la marge ciliaire (MC)contenant les cellules souches de la rétine. La marge ciliaire est composéede cellules pigmentées recouvrant les muscles lisses ciliaires. Inclusion enhistorésine. Coloration au bleu de toluidine. C : cristallin ; CO : cornée ;EPR : épithélium pigmenté de la rétine ; I : iris ; RN : rétine neurale. Clichéde E. Picard et J.-C. Jeanny.
21-003-C-40 ¶ Anatomie de la rétine
4 Ophtalmologie

cônes, les espèces crépusculaires entre 2 et 10 % de cônes, et lesmammifères diurnes ont une plus grande proportion de cônesentre 8 et 95 %. Cependant, la plupart des espèces diurnes ontune majorité de bâtonnets (10 à 20 % de cônes). Seules certai-nes espèces diurnes sont connues pour leurs taux très élevés decônes, comme la musaraigne (95 %) ou les écureuils (85 %).
Les densités maximales de bâtonnets se trouvent chez desmammifères nocturnes comme le rat d’Afrique, qui possèdeenviron 730 000 bâtonnets/mm2.
Contrairement aux idées communément admises, les donnéesrécentes montrent que toutes les rétines possèdent des bâton-nets et des cônes, même si ceux-ci y sont en proportionsvariables [9].
Existence d’une zone rétinienne centraledifférenciée
Les animaux possèdent des différenciations rétiniennesadaptées à leur mode de vie. Chez les animaux diurnes, l’acuitédépend en partie de l’intensité lumineuse stimulante, mettanten jeu, lorsqu’elle est élevée, un plus grand nombre de cônes.La richesse cellulaire de la rétine en photorécepteurs n’est pasla même selon la zone topographique. Chez les espèces dont larétine possède à la fois cônes et bâtonnets, les cônes sont plusnombreux au centre qu’à la périphérie, même quand l’animalne possède pas de macula.
On distingue trois niveaux de différenciation de la rétinecentrale, l’area centralis, la macula et la fovéa.
L’area centralis est une zone médiotemporale de plus granderichesse cellulaire. Elle correspond à un épaississement rétinienparfois macroscopiquement visible sous l’aspect d’une tacheplus foncée.
Il existe une area centralis chez les sélaciens (sous-classe depoissons cartilagineux à laquelle appartient la raie), les amphi-biens, quelques sauriens, tel l’alligator, et chez les carnivores,chiens et chats ; chez les ruminants, elle a une dispositionhorizontale allongée qui la fait qualifier de striaeformis.
La macula est une zone avasculaire de plus grande richesse encônes. Elle existe chez les primates, macroscopiquement visibleà l’examen du fond d’œil. La fovéa, uniquement constituée decônes, présente en outre un amincissement dû à la raréfactionou à la disparition de certaines couches rétiniennes.
Chez les téléostéens, qui regroupent la plupart des poissonsosseux, la fovéa est rudimentaire et constituée de cellulesamincies, sans raréfaction des couches rétiniennes. Les requinspossèdent un tapetum lucidum et une rétine riche en cônes etbâtonnets, mais ils ne possèdent pas de macula.
Chez les lacertiliens (reptiles à pattes, comme les lézards),existe tantôt une area, tantôt une fovéa, comme chez lecaméléon.
Les oiseaux de basse-cour n’ont qu’une area ; en revanche, lesrapaces ont deux fovéas : une fovéa centrale et une latérale.Dans la fovéa centrale, seule la couche des cellules ganglionnai-res fait défaut, les couches nucléaires étant amincies maisprésentes. La fovéa des oiseaux prédateurs compte 1 500 cônescontre 200 chez l’homme. L’acuité visuelle des oiseaux de proieest ainsi de loin la plus développée. Elle serait sept à huit foissupérieure à celle de l’homme.
L’homme et les primates possèdent une macula et une fovéa,mais le développement de la fovéa diffère chez l’homme et leprimate. Ainsi, la fovéa est différenciée dès la 12e semainepostnatale chez le singe alors qu’elle parvient à cette maturationvers 11-15 mois chez l’homme, n’atteignant sa maturationstructurale définitive qu’à l’âge de 5 ans [10].
Tapetum lucidum
Une autre particularité de la rétine animale est le tapetumlucidum, structure de réflexion, présente chez certains vertébréset permettant d’augmenter l’acuité visuelle en basse luminosité.Cette structure est située soit dans les couches rétiniennesprofondes, soit dans la choroïde, et sa composition est variable
selon les animaux diurnes qui la possèdent. Elle agit comme unréflecteur de photons vers les photorécepteurs. C’est le tapetumlucidum qui est responsable de l’impression d’œil lumineux,observé par exemple chez les chats, mais présent aussi chez descarnivores, des crocodiles, les vaches, moutons, chevaux et chezcertains poissons (requin). Les variations anatomiques et decompositions structurales sont un exemple d’adaptation spécia-lisée de la rétine neurosensorielle à des conditions de lumino-sités variables selon les conditions de vie des différentes espècesanimales [11].
Principales espèces animales utiliséespour une modélisation expérimentale
Les différences anatomiques observées entre les différentesespèces et en particulier parmi les espèces animales utiliséescomme modèles expérimentaux de pathologies rétiniennesdoivent être reconnues et prises en compte dans l’extrapolationà la pathologie humaine. En particulier, il faut noter que lavascularisation de la rétine du lapin diffère considérablement decelle de l’homme. Chez le lapin, utilisé pour les études pharma-cocinétiques, il existe un réseau vasculaire rétinien limité à larégion centrale péripapillaire, la majorité de la rétine interneétant vascularisée par le réseau choroïdien. Chez les rongeurs,habituellement utilisés comme modèles expérimentaux transgé-niques, la vascularisation de la rétine est proche de celle del’homme avec deux réseaux distincts : l’un rétinien comportantune barrière hématorétinienne, l’autre choriocapillaire nourris-sant la rétine externe par l’intermédiaire de la couche descellules de l’épithélium pigmentaire. Cependant, bien que larégion centrale de la rétine soit plus riche en cônes que la rétinepériphérique, les rongeurs ne possèdent pas de macula.
Seuls les primates possèdent une anatomie rétinienne compa-rable à celle de l’homme, tant sur le plan de la vascularisationque du point de vue de la différenciation de la rétine centrale.
Le poulet fréquemment utilisé dans les études sur la myopieprésente une rétine essentiellement composée de cônes etdépourvue de vascularisation rétinienne.
■ Anatomie macroscopique
La rétine est une fine tunique transparente in vivo, laissantapparaître la vascularisation choroïdienne. C’est cette transpa-rence qui donne son aspect rose orangé au fond d’œil. La rétineest transparente car elle est dans un état de relative déshydra-tation. En cas de décollement, et très rapidement après la mort,la rétine devient blanchâtre.
Ora serrata
La rétine s’étend de l’ora serrata en avant jusqu’au nerfoptique au pôle postérieur du globe oculaire. L’ora serrata est lazone où la limite antérieure de la rétine est en continuité avecl’épithélium non pigmenté de la pars plana (Fig. 6, 7). Danscette zone de la rétine, les bâtonnets sont absents et remplacéspar des cônes dont la morphologie est altérée comparée auxautres cônes rétiniens [12]. Les couches nucléaires fusionnent etla couche plexiforme externe disparaît. Les couches des cellulesganglionnaires et les fibres nerveuses s’interrompent environ0,5 mm en arrière de l’ora serrata. Des travaux récents ontmontré l’existence de jonctions adhérentes et de jonctionsserrées focales entre les membranes des cellules de l’épithéliumciliaire et les cellules adjacentes chez le lapin à ce niveau [13].
L’ora serrata est dentelée dans la région nasale et beaucoupplus lisse dans la région temporale. On compte entre 17 à34 indentations, réparties de façon irrégulière avec parfois delarges zones dépourvues de toute indentation (Fig. 8A). Leszones qui séparent les dents de l’ora sont appelées les baies del’ora serrata. Les dents de l’ora peuvent se projeter en avant dela rétine de 0,5 mm jusqu’à 2,5 mm. On peut trouver des zonesoù les baies sont encerclées laissant une zone de rétine centrale.
Anatomie de la rétine ¶ 21-003-C-40
5Ophtalmologie
En avant de l’ora serrata, la pars plana est une zone de 3 mmde long en nasal et de 4,5 mm en temporal. Les sclérectomiespostérieures sont réalisées dans la pars plana antérieure en avantde l’ora serrata et de l’insertion de la base du vitré (Fig. 8B).Chez le patient myope, l’ora serrata peut être déplacée posté-rieurement, proportionnellement à l’importance de la longueuraxiale, ce qui doit être pris en compte dans la chirurgie dudécollement de rétine chez ces patients.
Le diamètre transverse interne de l’œil emmétrope adulte estd’environ 22 mm et son diamètre externe de 24 mm, la sclèreet la choroïde faisant environ 1 mm. La sclère mesure 500 à800 µm d’épaisseur au limbe, 500 µm à l’équateur, 1 mm auniveau du nerf optique et 300 µm en arrière de l’insertion desmuscles extraoculaires.
La rétine couvre environ 15-16 mm du diamètre interne del’œil et 72 % de la surface de la sphère oculaire.
Épaisseur rétinienne
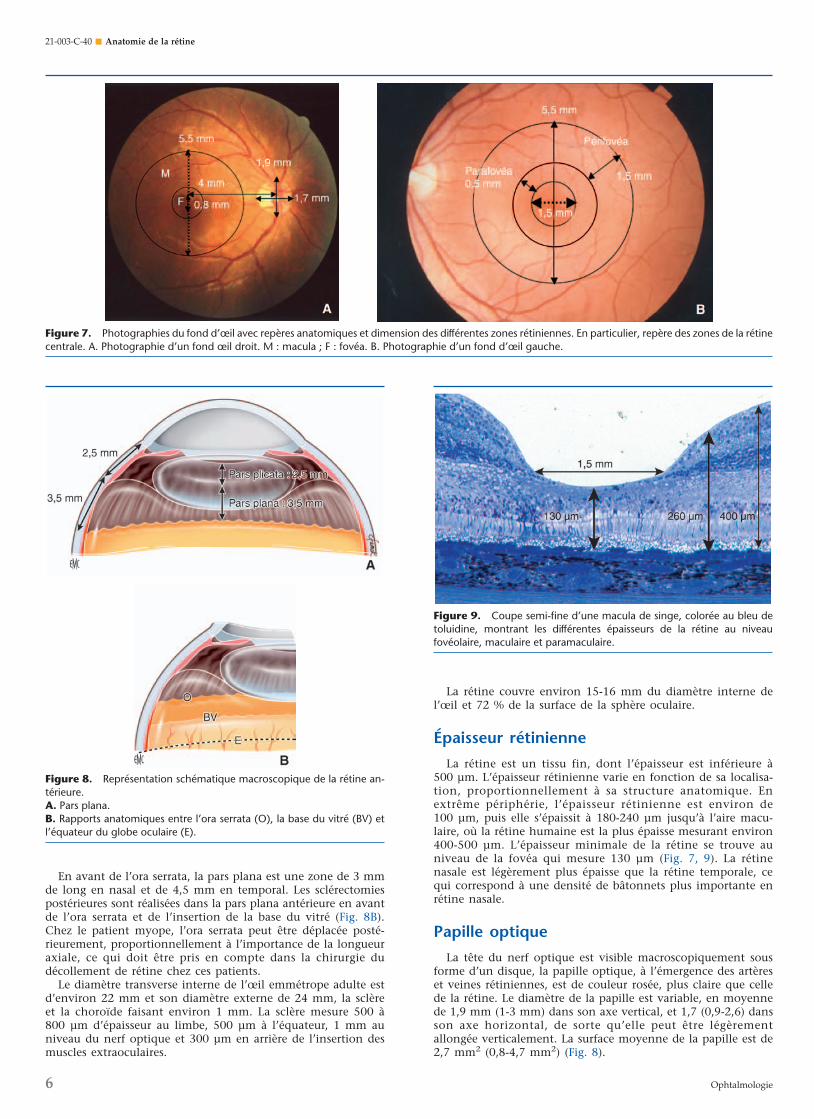
La rétine est un tissu fin, dont l’épaisseur est inférieure à500 µm. L’épaisseur rétinienne varie en fonction de sa localisa-tion, proportionnellement à sa structure anatomique. Enextrême périphérie, l’épaisseur rétinienne est environ de100 µm, puis elle s’épaissit à 180-240 µm jusqu’à l’aire macu-laire, où la rétine humaine est la plus épaisse mesurant environ400-500 µm. L’épaisseur minimale de la rétine se trouve auniveau de la fovéa qui mesure 130 µm (Fig. 7, 9). La rétinenasale est légèrement plus épaisse que la rétine temporale, cequi correspond à une densité de bâtonnets plus importante enrétine nasale.
Papille optique
La tête du nerf optique est visible macroscopiquement sousforme d’un disque, la papille optique, à l’émergence des artèreset veines rétiniennes, est de couleur rosée, plus claire que cellede la rétine. Le diamètre de la papille est variable, en moyennede 1,9 mm (1-3 mm) dans son axe vertical, et 1,7 (0,9-2,6) dansson axe horizontal, de sorte qu’elle peut être légèrementallongée verticalement. La surface moyenne de la papille est de2,7 mm2 (0,8-4,7 mm2) (Fig. 8).
Figure 7. Photographies du fond d’œil avec repères anatomiques et dimension des différentes zones rétiniennes. En particulier, repère des zones de la rétinecentrale. A. Photographie d’un fond œil droit. M : macula ; F : fovéa. B. Photographie d’un fond d’œil gauche.
Figure 8. Représentation schématique macroscopique de la rétine an-térieure.A. Pars plana.B. Rapports anatomiques entre l’ora serrata (O), la base du vitré (BV) etl’équateur du globe oculaire (E).
Figure 9. Coupe semi-fine d’une macula de singe, colorée au bleu detoluidine, montrant les différentes épaisseurs de la rétine au niveaufovéolaire, maculaire et paramaculaire.
21-003-C-40 ¶ Anatomie de la rétine
6 Ophtalmologie

Zones anatomiques rétiniennes distinctes
La macula (Fig. 9) est une zone de la rétine postérieure quimesure 5,5 mm de diamètre et dont le centre est situé à 4 mmen temporal et 0,8 mm inférieurement au centre de la papille.On repère facilement cette zone par la présence d’un pigmentjaunâtre, plus visible chez le sujet jeune, et par la raréfaction desvaisseaux rétiniens en son centre (Fig. 9).
La macula est divisée en plusieurs zones concentriquescorrespondant à la structure microscopique. Le centre de lamacula est une zone de 1,5 mm de diamètre (ou un diamètrepapillaire) formant une dépression. Son centre (0,35 mm dediamètre) est la fovéola. Elle est la zone la plus mince de larétine (environ 130 µm), très riche en cônes et en pigmentsmaculaires (lutéine et zéaxanthine). Son fond est reconnucomme le point central ou « umbo ». La fovéola et sa périphérie(une zone d’environ 0,5 mm de diamètre) sont dépourvues decapillaires rétiniens. On appelle cette zone, la « zone avasculairecentrale ».
Autour de la fovéa, une zone de 500 µm de diamètre corres-pond à la région où la rétine est la plus épaisse, du fait d’uneplus grande densité de cellules ganglionnaires, de la nucléaireinterne et de la couche des fibres de Henle, qui correspond auxaxones déportés de cônes et de prolongements de cellules glialesde Müller. Cette zone est appelée la zone parafovéolaire. Enfin,la dernière zone concentrique la plus externe correspond à unezone où la rétine s’amincit, c’est la zone périfovéolaire, quimesure 1,5 mm de diamètre (Fig. 9).
La zone de rétine postérieure ou proche périphérie rétinienneest une zone de 1,5 mm située autour de la région maculaire.
La rétine de moyenne périphérie correspond à une zone de3 mm en arrière.
La rétine d’extrême périphérie s’étend sur 9-10 mm du côténasal et sur environ 16 mm du côté temporal jusqu’à l’oraserrata.
Au niveau de l’extrême périphérie de la rétine, le vitré estadhérent sur 2 à 6 mm. Cette zone d’adhérence est appelée labase du vitré.
Le bord antérieur de la base du vitré est situé à 5 mm dulimbe sur tous les méridiens. Son adhérence dans la pars planaest donc plus antérieure du côté temporal que du côté nasal. Laréalisation de sclérotomie postérieure à moins de 5 mm dulimbe permet d’éviter une pénétration dans la base du vitré etle risque de traction sur des lésions préexistantes.
La base du vitré s’étend postérieurement à l’ora serrata de2 mm en temporal et de 3 mm en région nasale. Cette zoned’insertion postérieure n’est pas toujours bien définie, ni fixe, etpeut évoluer avec une myopisation du globe et avec l’âge,expliquant une migration graduelle postérieure possible de labase du vitré.
La base du vitré ne peut pas être disséquée de la rétineinterne sous-jacente, car il n’y a pas de plan de dissection, ni declivage entre le vitré et la rétine et la pars plana.
■ Vascularisation rétinienne
La rétine est vascularisée par deux systèmes vasculairesdifférents sans connexions anatomiques en conditions physio-logiques : le réseau capillaire rétinien, qui assure la vascularisa-tion directe des couches rétiniennes internes, et le réseauchoroïdien, qui assure, quant à lui, la vascularisation de larétine de façon indirecte, puisqu’il n’y a pas de capillaires dansla rétine externe (Fig. 10A).
Système vasculaire rétinien
L’artère centrale de la rétine émerge directement de l’artèreophtalmique dans seulement 50 % des cas. Dans les autres cas,elle émerge de l’artère ciliaire longue médiale (38 % des cas) etplus rarement d’une autre artère orbitaire, musculaire inférieureou ciliaire longue latérale [14]. Dans tous ces cas, l’artère centrale
de la rétine dérive primitivement de la carotide interne. Lecalibre de l’artère centrale de la rétine est compris entre 80 et100 µm. La veine centrale de la rétine a un calibre légèrementsupérieur de 100 à 125 µm. L’artère pénètre la face inférieure dunerf optique en moyenne 5 à 15 mm en arrière du globeoculaire. Puis l’artère suit un trajet intraneural pour émerger àla papille où elle se divise en ses quatre branches terminales :temporales et nasales, supérieures et inférieures. Les artèresterminales se divisent en collatérales, lesquelles se divisent enterminales qui comportent des collatérales et ainsi de suite defaçon dichotomique jusqu’à former un réseau en grillagecouvrant pour chacune des artères un quadrant de la rétineinterne. Les capillaires rétiniens sont issus de ces vaisseauxcollatéraux.
Dans 10 à 20 % des cas, une artère ciliorétinienne, branchedirecte de l’artère ciliaire, émerge du bord temporal de la papilleet vient suppléer la vascularisation de la région maculaire. Cettesuppléance joue un rôle majeur en cas d’occlusion de l’artèrecentrale de la rétine.
Le calibre des artérioles diminue au fur et à mesure de leurdivision en périphérie où leur calibre est compris entre 10 et20 µm. Les artères et artérioles sont formées d’une couche decellules endothéliales reposant sur une membrane basale, d’unemédia formée de cellules musculaires et d’une adventice. Lesadventices des artères et des veines rétiniennes sont communes.Le drainage de la rétine est assuré par la veine rétinienne qui sedéverse dans la veine ophtalmique supérieure.
Les artères et les veines rétiniennes sont localisées dans lacouche des fibres optiques (Fig. 10A). Les capillaires sontorganisés en une ou deux rangées en périphérie de la rétine et
10
9
11
12
1
3
2
4
5
6
7
9
A
8
Figure 10. Vascularisation de la rétine.A. Localisation des vaisseaux rétiniens et choroïdiens par rapport auxcouches de la rétine. 1. Couche des fibres optiques ; 2. couche des cellulesganglionnaires ; 3. couche plexiforme interne ; 4. couche nucléaire in-terne ; 5. couche plexiforme externe ; 6. couche nucléaire externe ; 7.cellules épithéliales pigmentaires ; 8. choroïde ; 9. réseau rétinien ; 10.choriocapillaires ; 11. artérioles et veinules.B. Montage à plat d’une rétine de singe et marquage de la vascularisationpar une lectine couplée à la fluorescéine, mettant en évidence la zoneavasculaire centrale (ZAC).
Anatomie de la rétine ¶ 21-003-C-40
7Ophtalmologie
en trois ou quatre rangées dans la partie la plus centrale de larétine. Les capillaires rétiniens font 5 à 6 µm de diamètre etsont formés de cellules endothéliales à jonctions serrées sur uneépaisse membrane basale. Des péricytes et des cellules micro-gliales entourent les capillaires. Au cours de la transition enveinules et artérioles, les péricytes sont remplacés par descellules musculaires lisses.
Une zone de 850 µm de diamètre, centrée par l’umbo, estdépourvue de capillaires rétiniens, c’est la zone avasculairecentrale (ZAC) (Fig. 10B).
Système vasculaire choroïdien
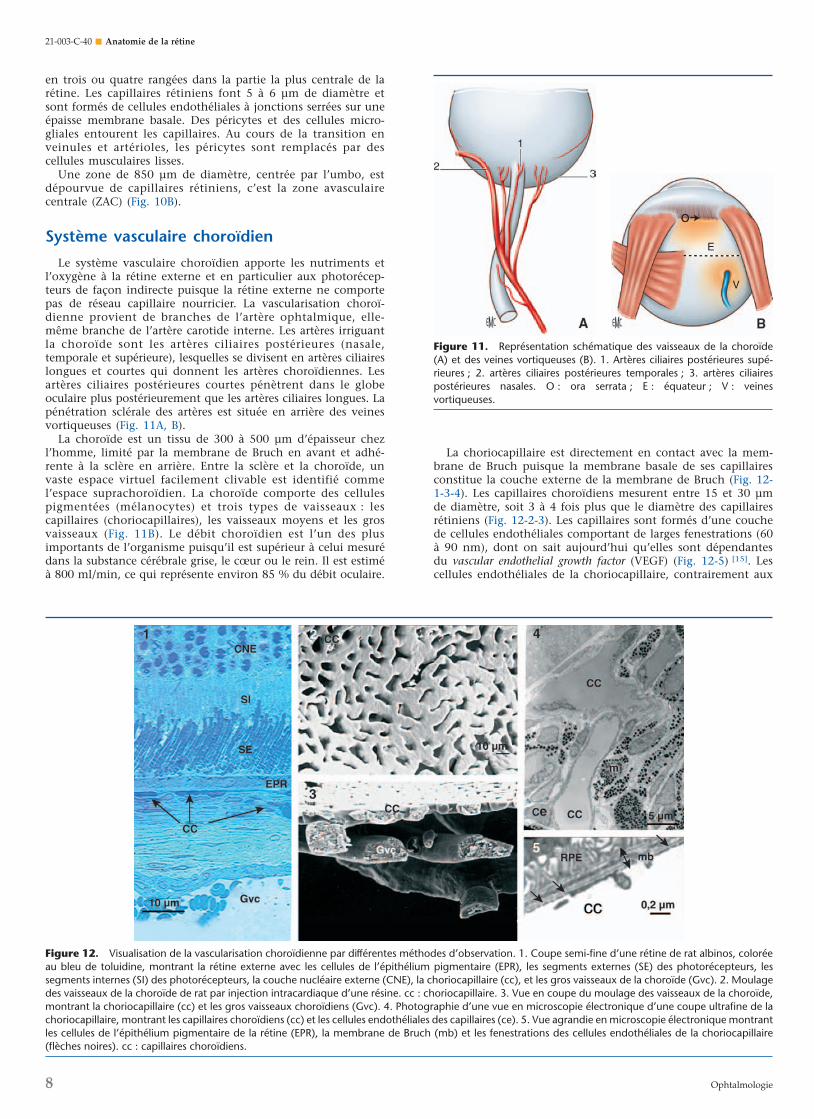
Le système vasculaire choroïdien apporte les nutriments etl’oxygène à la rétine externe et en particulier aux photorécep-teurs de façon indirecte puisque la rétine externe ne comportepas de réseau capillaire nourricier. La vascularisation choroï-dienne provient de branches de l’artère ophtalmique, elle-même branche de l’artère carotide interne. Les artères irriguantla choroïde sont les artères ciliaires postérieures (nasale,temporale et supérieure), lesquelles se divisent en artères ciliaireslongues et courtes qui donnent les artères choroïdiennes. Lesartères ciliaires postérieures courtes pénètrent dans le globeoculaire plus postérieurement que les artères ciliaires longues. Lapénétration sclérale des artères est située en arrière des veinesvortiqueuses (Fig. 11A, B).
La choroïde est un tissu de 300 à 500 µm d’épaisseur chezl’homme, limité par la membrane de Bruch en avant et adhé-rente à la sclère en arrière. Entre la sclère et la choroïde, unvaste espace virtuel facilement clivable est identifié commel’espace suprachoroïdien. La choroïde comporte des cellulespigmentées (mélanocytes) et trois types de vaisseaux : lescapillaires (choriocapillaires), les vaisseaux moyens et les grosvaisseaux (Fig. 11B). Le débit choroïdien est l’un des plusimportants de l’organisme puisqu’il est supérieur à celui mesurédans la substance cérébrale grise, le cœur ou le rein. Il est estiméà 800 ml/min, ce qui représente environ 85 % du débit oculaire.
La choriocapillaire est directement en contact avec la mem-brane de Bruch puisque la membrane basale de ses capillairesconstitue la couche externe de la membrane de Bruch (Fig. 12-1-3-4). Les capillaires choroïdiens mesurent entre 15 et 30 µmde diamètre, soit 3 à 4 fois plus que le diamètre des capillairesrétiniens (Fig. 12-2-3). Les capillaires sont formés d’une couchede cellules endothéliales comportant de larges fenestrations (60à 90 nm), dont on sait aujourd’hui qu’elles sont dépendantesdu vascular endothelial growth factor (VEGF) (Fig. 12-5) [15]. Lescellules endothéliales de la choriocapillaire, contrairement aux
Figure 12. Visualisation de la vascularisation choroïdienne par différentes méthodes d’observation. 1. Coupe semi-fine d’une rétine de rat albinos, coloréeau bleu de toluidine, montrant la rétine externe avec les cellules de l’épithélium pigmentaire (EPR), les segments externes (SE) des photorécepteurs, lessegments internes (SI) des photorécepteurs, la couche nucléaire externe (CNE), la choriocapillaire (cc), et les gros vaisseaux de la choroïde (Gvc). 2. Moulagedes vaisseaux de la choroïde de rat par injection intracardiaque d’une résine. cc : choriocapillaire. 3. Vue en coupe du moulage des vaisseaux de la choroïde,montrant la choriocapillaire (cc) et les gros vaisseaux choroïdiens (Gvc). 4. Photographie d’une vue en microscopie électronique d’une coupe ultrafine de lachoriocapillaire, montrant les capillaires choroïdiens (cc) et les cellules endothéliales des capillaires (ce). 5. Vue agrandie en microscopie électronique montrantles cellules de l’épithélium pigmentaire de la rétine (EPR), la membrane de Bruch (mb) et les fenestrations des cellules endothéliales de la choriocapillaire(flèches noires). cc : capillaires choroïdiens.
E
V
OO
Figure 11. Représentation schématique des vaisseaux de la choroïde(A) et des veines vortiqueuses (B). 1. Artères ciliaires postérieures supé-rieures ; 2. artères ciliaires postérieures temporales ; 3. artères ciliairespostérieures nasales. O : ora serrata ; E : équateur ; V : veinesvortiqueuses.
21-003-C-40 ¶ Anatomie de la rétine
8 Ophtalmologie

cellules endothéliales des capillaires rétiniens, ne comportentpas de jonctions serrées. Ainsi la barrière hématorétinienneexterne n’est-elle pas une barrière endothéliale. La choriocapil-laire est organisée en lobules fonctionnels irrigués par desartérioles indépendantes, sans anastomoses évidentes. Ces unitésfonctionnelles ont une taille de 300 µm à 1 mm, les lobules lesplus grands étant localisés en rétine périphérique, les plus petitsau niveau du pôle postérieur.
Les vaisseaux moyens comprennent les artérioles (10-20 µm)et les veinules (15-30 µm).
Les gros vaisseaux sont les artères et les veines choroïdiennes.Les artères ont un calibre de 50 à 100 µm. Elles sont forméesd’un endothélium non fenestré, d’une membrane basale, d’unelame élastique et de cellules musculaires lisses. Les artériolesperdent leurs cellules musculaires lisses et leur lame élastique.Chaque artère ciliaire courte postérieure irrigue un territoiretriangulaire à base externe, expliquant certaines formes d’isché-mie triangulaire. La macula est irriguée par 8 à 16 branches del’artère ciliaire courte postérieure temporale.
Les veines choroïdiennes convergent vers les golfs vortiqueuxà l’équateur du globe oculaire (Fig. 11). Les veines vortiqueusesfont environ un demi-millimètre de diamètre et sont visibles àleur sortie trans-sclérale du globe oculaire, 2 à 3 mm en arrièrede l’équateur (Fig. 11B).
■ Anatomie microscopique
Structure histologique généraleLa structure histologique de la rétine varie selon que l’on se
trouve proche de l’ora serrata ou bien dans la région maculaire.En dehors de ces zones spécifiques, l’anatomie microscopique dela rétine varie peu sinon du point de vue du nombre de couchesde noyaux de photorécepteurs.
Classiquement, on distingue dix couches de l’extérieur versl’intérieur du globe oculaire (Fig. 1) :• la couche des cellules de l’EPR ;• les segments externes et internes des photorécepteurs ;• la membrane limitante externe, lieu de connexion des
cellules gliales de Müller avec les segments internes desphotorécepteurs et des photorécepteurs entre eux par dessystèmes de jonctions adhérentes et serrées ;
• la couche de noyaux des photorécepteurs ou couche nucléaireexterne ;
• la couche plexiforme externe formée par les synapses entreles cellules bipolaires et les photorécepteurs et avec lescellules horizontales. Ces dernières modulent le messagenerveux transmis par la voie directe ;
• la couche des noyaux des cellules horizontales, bipolaires,amacrines et des cellules gliales de Müller, ou couchenucléaire interne. La région externe contient les corpscellulaires des cellules horizontales, la partie centrale lesnoyaux des cellules bipolaires et des cellules gliales de Müller,la partie la plus interne les cellules amacrines. Mais cetteorganisation n’est pas constante particulièrement pour ce quiconcerne les cellules amacrines, qui peuvent être déplacées ;
• la couche plexiforme interne constituée par les dendrites descellules ganglionnaires et les axones des cellules bipolaires ;
• la couche des cellules ganglionnaires (contenant des cellulesamacrines déplacées) ;
• la couche des fibres nerveuses est constituée par les axonesdes cellules ganglionnaires qui forment le nerf optique reliéau cerveau ;
• la membrane limitante interne est une expansion membra-neuse composée des pieds internes des cellules gliales deMüller.Nous débutons la description de la rétine par les couches
cellulaires les plus externes.En dedans de la sclère, la choroïde constitue l’uvée posté-
rieure, qui comporte comme nous l’avons vu plusieurs niveauxde vaisseaux mais aussi des cellules telles que des fibroblastes,des mélanocytes, des plasmocytes et des cellules dendritiques.
La membrane basale de la choriocapillaire constitue la couchela plus externe de la membrane de Bruch, qui comporte cinqcouches différenciées (Fig. 12-4). De l’intérieur vers l’extérieur,on distingue : la membrane basale de l’EPR, une couche internede fibres collagènes, une couche élastique, une couche externede fibres collagènes et la membrane basale de la choriocapillaire.La couche élastique et la membrane basale de la choriocapillaireprésentent des zones de discontinuité, mais qui ne s’affrontentpas. L’épaisseur de la membrane de Bruch varie du pôle posté-rieur à la périphérie. Elle fait 2-4 µm autour du nerf optique et1-2 µm en périphérie de la rétine. Elle est formée d’élastine, decollagène de types I, II, III, IV, V, XI et à un degré moindre, decollagène IX et XII. Elle contient aussi de la laminine, de lafibronectine, des héparanes sulfates protéoglycanes et dunidogène-entactine.
Épithélium pigmentaire de la rétineLa monocouche d’EPR est un élément majeur dans la rétine,
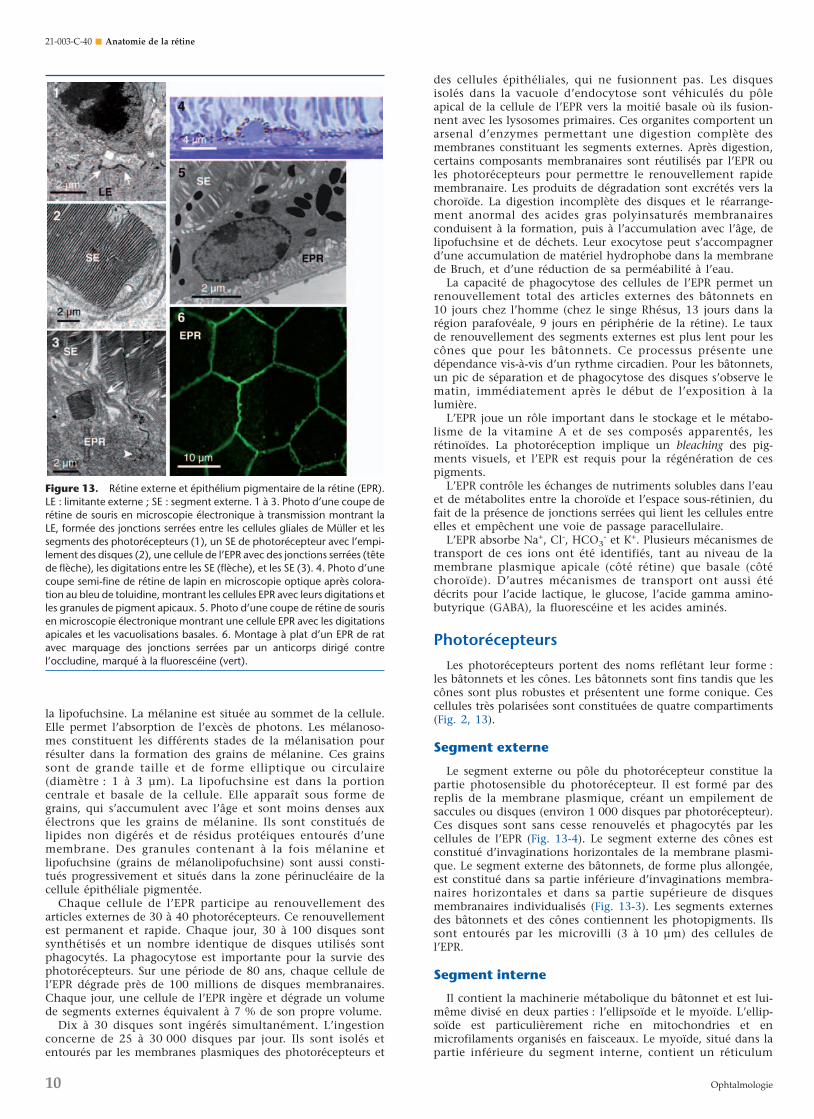
non seulement en raison de ses propriétés physiques de barrièrehématorétinienne, de ses propriétés optiques, mais surtout, enraison de ses nombreuses activités métaboliques. Elle occupeune situation stratégique entre les photorécepteurs et la chorio-capillaire et son intégrité est fondamentale pour le fonctionne-ment normal de la rétine, en particulier des photorécepteurs(Fig. 13-1). Si l’EPR est détruit mécaniquement ou chimique-ment, photorécepteurs et choriocapillaires s’atrophient.
Elle est responsable de l’adhésion de la rétine neurale, à lafois par l’interdigitation de ses villosités avec les photorécepteursmais encore plus en raison de l’existence d’une pompe ionique(Fig. 13-4 à 6). Elle est directement impliquée dans le systèmevisuel par son activité métabolique au niveau de la transductionvisuelle, mais aussi par la phagocytose des segments externes« usagés » des photorécepteurs.
Elle est le site de synthèse ou de stockage de nombreuxfacteurs de croissance (FGF, VEGF et transforming growth factor[TGF]b) et interagit avec les types cellulaires avoisinants, enparticulier les photorécepteurs.
La monocouche d’EPR est constituée de cellules de formehexagonale (4 à 6 millions, mais de nombre variable selon l’âgeet leur localisation périphérique ou maculaire ; 5 000 cellules/mm2 dans la fovéa, 2 000 cellules/mm2 en périphérie). Au pôlepostérieur, les cellules sont hautes (14 µm) et longues (10-14 µm). À l’ora serrata, elles sont plus plates et plus larges(60 µm) (Fig. 13-3). Certaines sont binucléées. La liaison descellules entre elles s’opère par jonctions serrées (Fig. 13-4).
La membrane plasmique de la cellule épithéliale pigmentéeprésente de nombreuses villosités aux pôles apical et basal, quipermettent une augmentation de la surface d’échange, deséchanges métaboliques entre EPR et choriocapillaire et untransport ionique actif au travers de l’EPR (Fig. 13-6).
La face latérale de la membrane plasmique est régulière. Elleest pourvue de zonulae occludentes (jonctions intercellulairesserrées, fusion de deux membranes cellulaires adjacentes) etzonulae adherentes (espace intercellulaire 200 Å) pour constituerune barre terminale à l’extrémité supérieure de la cellule. Elleempêche le passage passif des macromolécules entre choroïde etphotorécepteurs. Il existe aussi parfois des desmosomes basaux(matériel intercellulaire associé à des tonofilaments) (Fig. 13-3).Sur la face interne, il y a un contact intime des cellules de l’EPRavec les segments externes des photorécepteurs (Fig. 13-4).
Une cellule de l’EPR est en contact avec 30 à 45 segmentsexternes de photorécepteurs. Deux types de villosités sontimpliqués dans la phagocytose : des longues (5 à 7 µm, exten-sion entre les segments externes) et des courtes (3 à 4 µm,manchon autour de l’extrémité des segments externes). Lessegments externes des cônes n’atteignent pas la surface apicalede l’EPR, mais les microvilli. On note aussi la présence dephagosomes dans la portion apicale de la cellule de l’EPR,ceux-ci contenant des inclusions lamellaires de membranes desphotorécepteurs, à différents stades de leur dégradation.Différents types de grains de pigments sont enfin présents dansles cellules épithéliales. Les deux principaux sont la mélanine et
Anatomie de la rétine ¶ 21-003-C-40
9Ophtalmologie
la lipofuchsine. La mélanine est située au sommet de la cellule.Elle permet l’absorption de l’excès de photons. Les mélanoso-mes constituent les différents stades de la mélanisation pourrésulter dans la formation des grains de mélanine. Ces grainssont de grande taille et de forme elliptique ou circulaire(diamètre : 1 à 3 µm). La lipofuchsine est dans la portioncentrale et basale de la cellule. Elle apparaît sous forme degrains, qui s’accumulent avec l’âge et sont moins denses auxélectrons que les grains de mélanine. Ils sont constitués delipides non digérés et de résidus protéiques entourés d’unemembrane. Des granules contenant à la fois mélanine etlipofuchsine (grains de mélanolipofuchsine) sont aussi consti-tués progressivement et situés dans la zone périnucléaire de lacellule épithéliale pigmentée.
Chaque cellule de l’EPR participe au renouvellement desarticles externes de 30 à 40 photorécepteurs. Ce renouvellementest permanent et rapide. Chaque jour, 30 à 100 disques sontsynthétisés et un nombre identique de disques utilisés sontphagocytés. La phagocytose est importante pour la survie desphotorécepteurs. Sur une période de 80 ans, chaque cellule del’EPR dégrade près de 100 millions de disques membranaires.Chaque jour, une cellule de l’EPR ingère et dégrade un volumede segments externes équivalent à 7 % de son propre volume.
Dix à 30 disques sont ingérés simultanément. L’ingestionconcerne de 25 à 30 000 disques par jour. Ils sont isolés etentourés par les membranes plasmiques des photorécepteurs et
des cellules épithéliales, qui ne fusionnent pas. Les disquesisolés dans la vacuole d’endocytose sont véhiculés du pôleapical de la cellule de l’EPR vers la moitié basale où ils fusion-nent avec les lysosomes primaires. Ces organites comportent unarsenal d’enzymes permettant une digestion complète desmembranes constituant les segments externes. Après digestion,certains composants membranaires sont réutilisés par l’EPR oules photorécepteurs pour permettre le renouvellement rapidemembranaire. Les produits de dégradation sont excrétés vers lachoroïde. La digestion incomplète des disques et le réarrange-ment anormal des acides gras polyinsaturés membranairesconduisent à la formation, puis à l’accumulation avec l’âge, delipofuchsine et de déchets. Leur exocytose peut s’accompagnerd’une accumulation de matériel hydrophobe dans la membranede Bruch, et d’une réduction de sa perméabilité à l’eau.
La capacité de phagocytose des cellules de l’EPR permet unrenouvellement total des articles externes des bâtonnets en10 jours chez l’homme (chez le singe Rhésus, 13 jours dans larégion parafovéale, 9 jours en périphérie de la rétine). Le tauxde renouvellement des segments externes est plus lent pour lescônes que pour les bâtonnets. Ce processus présente unedépendance vis-à-vis d’un rythme circadien. Pour les bâtonnets,un pic de séparation et de phagocytose des disques s’observe lematin, immédiatement après le début de l’exposition à lalumière.
L’EPR joue un rôle important dans le stockage et le métabo-lisme de la vitamine A et de ses composés apparentés, lesrétinoïdes. La photoréception implique un bleaching des pig-ments visuels, et l’EPR est requis pour la régénération de cespigments.
L’EPR contrôle les échanges de nutriments solubles dans l’eauet de métabolites entre la choroïde et l’espace sous-rétinien, dufait de la présence de jonctions serrées qui lient les cellules entreelles et empêchent une voie de passage paracellulaire.
L’EPR absorbe Na+, Cl-, HCO3- et K+. Plusieurs mécanismes de
transport de ces ions ont été identifiés, tant au niveau de lamembrane plasmique apicale (côté rétine) que basale (côtéchoroïde). D’autres mécanismes de transport ont aussi étédécrits pour l’acide lactique, le glucose, l’acide gamma amino-butyrique (GABA), la fluorescéine et les acides aminés.
Photorécepteurs
Les photorécepteurs portent des noms reflétant leur forme :les bâtonnets et les cônes. Les bâtonnets sont fins tandis que lescônes sont plus robustes et présentent une forme conique. Cescellules très polarisées sont constituées de quatre compartiments(Fig. 2, 13).
Segment externe
Le segment externe ou pôle du photorécepteur constitue lapartie photosensible du photorécepteur. Il est formé par desreplis de la membrane plasmique, créant un empilement desaccules ou disques (environ 1 000 disques par photorécepteur).Ces disques sont sans cesse renouvelés et phagocytés par lescellules de l’EPR (Fig. 13-4). Le segment externe des cônes estconstitué d’invaginations horizontales de la membrane plasmi-que. Le segment externe des bâtonnets, de forme plus allongée,est constitué dans sa partie inférieure d’invaginations membra-naires horizontales et dans sa partie supérieure de disquesmembranaires individualisés (Fig. 13-3). Les segments externesdes bâtonnets et des cônes contiennent les photopigments. Ilssont entourés par les microvilli (3 à 10 µm) des cellules del’EPR.
Segment interne
Il contient la machinerie métabolique du bâtonnet et est lui-même divisé en deux parties : l’ellipsoïde et le myoïde. L’ellip-soïde est particulièrement riche en mitochondries et enmicrofilaments organisés en faisceaux. Le myoïde, situé dans lapartie inférieure du segment interne, contient un réticulum
Figure 13. Rétine externe et épithélium pigmentaire de la rétine (EPR).LE : limitante externe ; SE : segment externe. 1 à 3. Photo d’une coupe derétine de souris en microscopie électronique à transmission montrant laLE, formée des jonctions serrées entre les cellules gliales de Müller et lessegments des photorécepteurs (1), un SE de photorécepteur avec l’empi-lement des disques (2), une cellule de l’EPR avec des jonctions serrées (têtede flèche), les digitations entre les SE (flèche), et les SE (3). 4. Photo d’unecoupe semi-fine de rétine de lapin en microscopie optique après colora-tion au bleu de toluidine, montrant les cellules EPR avec leurs digitations etles granules de pigment apicaux. 5. Photo d’une coupe de rétine de sourisen microscopie électronique montrant une cellule EPR avec les digitationsapicales et les vacuolisations basales. 6. Montage à plat d’un EPR de ratavec marquage des jonctions serrées par un anticorps dirigé contrel’occludine, marqué à la fluorescéine (vert).
21-003-C-40 ¶ Anatomie de la rétine
10 Ophtalmologie

endoplasmique rugueux abondant et un appareil de Golgi trèsdéveloppé. Les segments internes des cônes sont plus larges queceux des bâtonnets, ils ont une forme plus allongée dans larégion fovéolaire. Les segments internes sont séparés entre euxpar des prolongements de cellules gliales de Müller.
Le segment interne et le segment externe sont reliés par uncil connecteur. Neuf tubules ciliaires émanent du myoïde ets’engagent à des niveaux variables dans le segment externe. Lessegments externes des cônes se raccourcissent en périphérie,alors que ceux des bâtonnets gardent une longueur constante.
Membrane limitante externe
Ce n’est pas une véritable membrane mais plutôt une zone dedensification (Fig. 13-1) située entre la couche nucléaire externeet les segments internes des photorécepteurs. Ces densificationscorrespondent à des jonctions adhérentes entre les photorécep-teurs et les cellules gliales de Müller. Il apparaît de plus en plusclairement que ces zones d’adhérence ont des composantsspécifiques au niveau des cônes. Au cours du développement, lalimitante externe se forme avant l’apparition des segmentsexternes. La membrane limitante externe s’étend de l’ora serrataau bord de la papille optique. Elle suit la courbe postérieure duglobe sauf au niveau de la région maculaire, où les cônesfovéaux s’allongent. La description ultrastructurale précise de lamembrane limitante externe revient à Spitznas qui, en 1970,montre la présence de zonula adherentes (ou barre terminale)entre les cellules gliales de Müller et les photorécepteurs (côneset bâtonnets), entre les cellules de Müller, et plus rarement entreles photorécepteurs.
Couche nucléaire externe
Elle contient les corps cellulaires où est situé le noyau. Lesnoyaux des cônes sont légèrement plus gros que ceux desbâtonnets et contiennent moins d’hétérochromatine, ce quiexplique leur plus faible réactivité aux colorants nucléaires(Fig. 2).
À proximité du bord nasal de la papille, la couche nucléaireexterne mesure 45 µm et contient huit à neuf rangées denoyaux. L’épaisseur en temporal de la papille est de 22 µm avecseulement quatre rangées de noyaux, puis augmente jusqu’àatteindre 50 µm et dix rangées de noyaux au niveau fovéolaire.Sur le reste de la surface rétinienne, à l’exception de l’oraserrata, la couche nucléaire externe mesure 27 µm, contenantune rangée unique de noyaux de cônes à proximité de lamembrane limitante externe et quatre rangées de noyaux debâtonnets en dedans. Les noyaux de bâtonnets mesurent5,5 µm de diamètre alors que celui des cônes est légèrementplus grand, de 6 à 7 µm. Au niveau de la fovéa, les noyaux decônes peuvent être déplacés en dehors de la membrane limi-tante externe. Les cônes bleus (environ 10 à 15 % des cônes)peuvent être distingués morphologiquement par un segmentinterne un peu plus long et de petites extrémités synaptiques.
Le pédicule synaptique contient des faisceaux de microfila-ments, des microtubules et des rubans synaptiques entourés denombreuses vésicules. Ce pédicule, appelé aussi pôle neuro-transmetteur, libère des neurotransmetteurs et principalement leglutamate.
Dans la rétine de mammifères, il existe un seul type debâtonnet et quatre types de cônes spécifiques de chaque couleurdétectée (rouge, bleu, vert et violet). Environ 50 à 55 % descônes sont sensibles à la lumière verte et 33 à 35 % à la lumièrerouge.
Les bâtonnets sont responsables de la vision périphérique, ladétection de mouvement et la vision nocturne (ou visionscotopique). Ils ont la plus grande sensibilité puisqu’un seulphoton est capable d’exciter un bâtonnet. Les cônes, moinsnombreux, sont responsables de la vision diurne (ou visionphotopique) et permettent de différencier les couleurs. La rétine
humaine contient en moyenne 4,6 millions de cônes (4,08-5,29, données recueillies sur huit rétines humaines) avec unedensité fovéolaire maximale d’environ 190 000 cônes/mm2
(mais avec d’importantes variations possibles). Le diamètrehorizontal de la zone fovéolaire dépourvue de bâtonnets estd’environ 350 µm. Les bâtonnets sont en moyenne au nombrede 92 millions (77,9 - 107,3, données recueillies sur huit rétineshumaines) et leur densité maximale est observée autour du nerfoptique. La densité des bâtonnets est plus importante en rétinenasale qu’en rétine temporale, et en rétine inférieure qu’enrétine supérieure [16]. Les cônes centraux sont plus fins qu’unbâtonnet et leurs segments externes sont très allongés. Auniveau fovéolaire, il n’y a pas de cônes bleus et l’arrangementdes cônes forme une mosaïque régulière ; les segments internesdes cônes étant séparés de moins de 2,5 µm. En périphérie dela fovéa, les bâtonnets commencent à apparaître avec unedensité qui atteint 100 000/ mm2 à une distance de 1,5 mm ducentre de la fovéa.
De façon intéressante, des études morphologiques ont montréque la densité des cônes est très stable au cours de la vie et nedécroît pas avec l’âge contrairement à la densité des bâtonnetscentraux qui diminue de plus de 30 % dans les 3 mm centrauxà la neuvième décennie de la vie. Ceci suggérerait une plusgrande fragilité des bâtonnets [17].
Spécificité morphologique des cônes fovéolaires
Le diamètre fovéolaire dépourvu de bâtonnets mesure envi-ron 570 µm, contenant 35 000 cônes. La fovéola contient 2 500cônes régulièrement compactés et orientés verticalement. Lescônes fovéaux mesurent 80 µm de long, leurs segments internes20 à 30 µm et leurs segments externes 45 µm. Les noyaux decônes fovéaux mesurent 7 µm de diamètre. L’arrangement descônes fovéolaires forme une mosaïque régulière, les segmentsinternes étant séparés de 0,3 µm par des prolongements decellules gliales de Müller. Les mitochondries sont régulièrementdispersées dans les segments internes. Sur le bord de la fovéa,on compte dix rangées de noyaux, mais la couche nucléaireexterne s’amincit considérablement au centre de la fovéa, où onne compte plus que deux ou trois rangées de noyaux.
Les photorécepteurs sont des neurones très spécialisés, seulscapables de phototransduction. La phototransduction, effectuéepar les photorécepteurs, englobe les étapes qui vont de ladétection du photon lumineux jusqu’à l’ébauche d’une réponsesensorielle. Il s’agit de la transformation d’une énergie électro-magnétique purement physique, en une énergie métaboliquequi correspond à la genèse d’un signal électrophysiologique. Lesétapes purement biochimiques de la phototransduction sontbien décrites au niveau du bâtonnet alors qu’elles restenthypothétiques au niveau du cône.
Couche plexiforme externeElle est formée des synapses entre les photorécepteurs et les
cellules bipolaires et horizontales, lesquelles forment unecouche plus épaisse, la « couche des fibres de Henle », dans larégion maculaire (50 µm). Dans les autres régions de la rétine,la plexiforme externe ne fait que quelques micromètres et serétrécit en rétine périphérique. Les prolongements des cellulesgliales de Müller se trouvent également dans cette couche.
Couche nucléaire interneElle contient quatre types de cellules, les cellules horizontales,
les cellules bipolaires, les cellules amacrines et les cellules glialesde Müller.
Certaines cellules amacrines peuvent se trouver dans lacouche des cellules ganglionnaires, on les qualifie de « dépla-cées ». Des cellules interplexiformes ont leurs noyaux dans lacouche nucléaire interne et des prolongements synaptiques à lafois dans les plexiformes interne et externe.
Au niveau fovéolaire, la couche nucléaire internes’interrompt.
Anatomie de la rétine ¶ 21-003-C-40
11Ophtalmologie

Couche plexiforme interne
Elle est le siège des synapses entre les cellules bipolaires etamacrines et les cellules ganglionnaires. Son épaisseur varie de18 à 36 µm et elle est absente au niveau de la fovéa. Uneorganisation ultrastructurale en trois rangées est décrite.
Couche des cellules ganglionnaires
Les cellules ganglionnaires sont les neurones qui collectentl’information visuelle pour la transmettre au système nerveuxcentral. Leurs noyaux sont localisés dans la couche nucléaire laplus interne de la rétine, dénommée la couche des cellulesganglionnaires, mais on peut également trouver plus rarementdes noyaux de cellules ganglionnaires dans la plexiformeinterne et dans la nucléaire interne. Les cellules ganglionnairessont de grosses cellules d’environ 10 à 20 µm qui forment dessynapses, par l’intermédiaire de leur dendrite, avec les cellulesbipolaires et les cellules amacrines dans la couche plexiformeinterne. Les signaux des photorécepteurs sont en effet relayésvers les cellules ganglionnaires par un réseau d’interneurones.Chaque type d’interneurones rétiniens (cellules horizontales,bipolaires, amacrines) joue un rôle spécifique dans la mise enforme du signal des photorécepteurs transmis à travers la rétine.
Les axones des cellules ganglionnaires convergent vers lapapille pour former le nerf optique. Les cellules ganglionnairesforment une monocouche sauf dans la région parafovéolaire, oùelles sont empilées en six-sept couches. Les cellules ganglion-naires forment une monocouche à l’extrémité de la zonepérifovéolaire, qui correspond également à la limite postérieurede la rétine centrale. Au niveau de la rétine centrale (5-6 mmcentraux, correspondant à un champ visuel central de 18°), lespigments sont plus concentrés dans les cellules ganglionnaireset parfois également dans les cellules amacrines, particulière-ment la lutéine et la zéaxanthine.
Les afférences d’une cellule ganglionnaire sont originaires desphotorécepteurs de voisinage dans une aire circonscrite de larétine, le champ récepteur de cette cellule. Les champs récep-teurs des cellules ganglionnaires possèdent deux importantespropriétés. En premier, l’exploration de la rétine à l’aide d’unstimulus en forme de petite tache lumineuse montre que leschamps récepteurs sont globalement circulaires. Deuxièmement,pour la plupart des cellules ganglionnaires, le champ récepteurest divisé en deux parties : une zone circulaire au centre appeléele centre du champ récepteur, et la partie restante, appeléepériphérie du champ récepteur. Les cellules ganglionnairesrépondent surtout à l’illumination différentielle des partiescentrale et périphérique de leurs champs récepteurs. Les cellulesganglionnaires centre-on et centre-off sont présentes en quan-tités égales et tous les photorécepteurs projettent sur les deuxtypes. Les cellules ganglionnaires sont ainsi spécialisées dans ladétection de contrastes et les changements rapides de l’imagevisuelle.
Couche des fibres optiques
La couche des fibres optiques a une épaisseur maximaleautour de la papille où les fibres convergent. À ce niveau, sonépaisseur mesurée en tomographie à cohérence optique in vivosur des sujets sains est d’environ 107 ± 20 µm [18]. Histologique-ment, l’épaisseur moyenne de la couche des fibres optiques surun diamètre de 3,5 mm autour du nerf optique est de 60,3± 19,5 µm (n = 17), avec des variations en rétine supérieure,75,3 ± 26,5 µm par rapport à la rétine inférieure 69,4 ± 22,4 µmet en rétine nasale 48,1 ± 15 µm par rapport à la rétine tempo-rale 49,2 ± 26,4 µm [19].
La taille des axones varie de 0,6 à 2 µm selon le type decellules ganglionnaires dont ils émergent. Les axones de cellulesganglionnaires forment des fibres entrelacées avec des prolon-gements des cellules gliales de Müller. Les fibres convergent aunerf optique avec une orientation radiaire, sauf pour ce quiconcerne les fibres maculaires qui forment un réseau rectiligne
interpapillomaculaire. Le faisceau temporal correspond à toutesles fibres situées en dehors de la papille et le faisceau nasal àcelles situées en dedans de la papille. Le faisceau temporal étantmoins dense, le bord temporal de la couche des fibres optiquesest moins charnu que le côté nasal. Les fibres optiques peuventrarement être myelinisées, mais ceci n’est pas la règle, et lesfibres nerveuses de la rétine de lapin sont une exception à cetterègle.
Membrane limitante interneL’existence de la membrane limitante interne a été longtemps
contestée, mais les études ultrastructurales ont pu démontrerqu’une membrane individualisable de 1 à 2 µm existe à l’inter-face de la rétine et du cortex vitréen. Elle est continue sur toutela surface de la rétine et s’interrompt au niveau des bords de lapapille. Les pieds des cellules gliales de Müller reposent sur cettemembrane basale. D’autres cellules gliales sont en contact avecelle (en particulier les astrocytes et des cellules microgliales). Ducollagène vitréen est adhérent à la face interne de la membranetandis que la face externe est irrégulière et suit les prolonge-ments de cellules gliales de Müller.
Cellules gliales de la rétineLa glie rétinienne comme la glie neuronale dans le système
nerveux central joue de multiples rôles dans l’homéostasieneuronale : transport de nutriments, régulation hydro-ionique,immunomodulation, production de facteurs neurotrophiques.
Les astrocytes et la microglie sont localisés en situationphysiologique basale dans les couches rétiniennes internes. Lamicroglie rétinienne, véritable macrophage spécialisé de larétine, entoure les capillaires rétiniens. Les astrocytes sont situésaussi autour des vaisseaux rétiniens, sous la membrane limitanteinterne et sont très nombreux à l’émergence de la tête du nerfoptique.
Les cellules gliales de Müller ont leurs noyaux dans la partiemédiane de la couche nucléaire interne, mais leurs prolonge-ments cytoplasmiques s’étendent de la membrane limitanteinterne jusqu’aux segments internes des photorécepteurs. Iln’est pas exclu que leurs prolongements puissent affronter lesmicrovilli des cellules de l’EPR.
Cellules gliales de Müller
Les cellules de Müller sont les principales cellules gliales de larétine des vertébrés.
Comme noté dans les paragraphes précédents, elles sontprésentes dans toutes les couches de la rétine car elles traversentradialement toute l’épaisseur de la rétine et sont en contact avectous les neurones rétiniens (Fig. 14). Elles sont essentielles dansla création et le maintien de l’architecture rétinienne, servant desupport et substrat pour la migration des neurones postmitoti-ques et la croissance de leurs axones, et supportent la différen-ciation et la survie neuronale.
Leur corps cellulaire est localisé au niveau de la couchenucléaire interne et elles mesurent environ 100 µm de long.L’extrémité interne des cellules gliales de Müller forme un pied,apposé sur la membrane limitante interne, qui constitue leurmembrane basale. Des prolongements des cellules gliales deMüller forment des manchons autour des capillaires rétiniens.
Les cellules gliales de Müller sont ainsi impliquées dans lecontrôle de l’angiogenèse et la régulation du flux sanguinrétinien et contribuent à la formation et au maintien de labarrière hématorétinienne.
Au niveau de la couche nucléaire externe, les prolongementsdes cellules gliales de Müller entourent les noyaux des photoré-cepteurs et forment des jonctions adhérentes avec les photoré-cepteurs au niveau de la partie interne de leurs segmentsinternes et entre elles. Ces jonctions forment la membranelimitante externe.
Au niveau de la fovéa, les cellules de Müller ont une formeconique dont l’apex tronqué est situé au niveau de la limitante
21-003-C-40 ¶ Anatomie de la rétine
12 Ophtalmologie

externe, dans une zone dénuée de noyaux. La base des cellulesde Müller-cône est située au niveau de la limitante interne.Cette morphologie conique a laissé penser pendant longtemps,à tort, qu’il n’y avait pas de cellules gliales de Müller au niveaude la fovéa, comme on peut le trouver rapporté dans certainstraités d’anatomie [20].
Les liens étroits qui existent entre les prolongements decellules gliales de Müller et les cônes fovéolaires ont été àprésent bien établis.
Les cellules gliales de Müller constituent un lien anatomiqueentre les neurones rétiniens et les compartiments avec lesquelsils échangent des molécules, à savoir les vaisseaux rétiniens, levitré et l’espace sous-rétinien. Ce lien n’est pas exclusivementanatomique mais aussi fonctionnel, incluant une symbiosemétabolique et le traitement et l’acheminement de l’informa-tion visuelle. Les cellules de Müller sont dotées de nombreuxcanaux ioniques différents, de récepteurs pour des ligands, demolécules de transport transmembranaire et d’enzymes. Ellesexpriment en particulier l’enzyme, glutamine synthétase, quiconvertit les molécules de glutamate internalisées en glutamine.Elles sont responsables du maintien de l’homéostasie du milieuextracellulaire rétinien (ions, eau, neurotransmetteurs et pH).Une de leurs principales caractéristiques est la conductanceélevée vis-à-vis du K+ de leur membrane plasmique. Elles sontimpliquées dans le métabolisme du glucose rétinien, apportantaux neurones lactate/pyruvate pour leur métabolisme oxydatifet l’élimination des déchets métaboliques.
En outre, les cellules gliales de Müller possèdent deux autrespropriétés récemment mises en évidence, la possibilité de servir
de progéniteurs neuronaux [21] et un rôle important dans laconduction des photons jusqu’aux photorécepteurs. Les cellulesgliales de Müller joueraient en effet le rôle de véritables fibresoptiques naturelles au niveau de la rétine [22].
Astrocytes et microglie
Les astrocytes de la rétine sont de deux types. Des cellulesdeux fois plus longues que larges, disposées le long des fais-ceaux de fibres optiques, sans contact vasculaire, et une autrepopulation de cellules astrogliales étoilées qui traversent lacouche des fibres optiques établissent des contacts non spécia-lisés avec les vaisseaux rétiniens. La plus grande densité decellules astrogliales est observée autour du nerf optique et lelong des faisceaux de fibres [23].
■ ConclusionLa structure de la rétine reflète sa complexité fonctionnelle et
l’extraordinaire spécialisation topographique de ce tissu. Denombreuses analyses structurales de la rétine humaine, utilisantdes méthodes dédiées in vivo et ex vivo, sont encore nécessairespour préciser la morphologie de la rétine normale d’une part etles variations physiologiques fonctionnelles d’autre part. Lerecours aux méthodes d’imagerie in vivo permettent desmesures d’épaisseur, dont la corrélation histologique reste àétablir. Enfin, l’identification de marqueurs spécifiques dedifférents types cellulaires reste nécessaire pour une analysemorphologique plus précise.
■ Références[1] Hatakeyama J, Kageyama R. Retinal cell fate determination and bHLH
factors. Semin Cell Dev Biol 2004;15:83-9.[2] Cepko CL, Austin CP, Yang X, Alexiades M, Ezzeddine D. Cell fate
determination in the vertebrate retina. Proc Natl Acad Sci USA 1996;93:589-95.
[3] Belliveau MJ, Cepko CL. Extrinsic and intrinsic factors control thegenesis of amacrine and cone cells in the rat retina. Development 1999;126:555-66.
[4] Vetter ML, Brown NL. The role of basic helix-loop-helix genes invertebrate retinogenesis. Semin Cell Dev Biol 2001;12:491-8.
[5] Hollyfield JG. Elimination of egg pigment from developing oculartissues in the frog Rana pipiens. Dev Biol 1973;30:115-28.
[6] Levine EM, Schechter N. Homeobox genes are expressed in the retinaand brain of adult goldfish. Proc Natl Acad Sci USA 1993;90:2729-33.
[7] Park CM, Hollenberg MJ. Basic fibroblast growth factor induces retinalregeneration in vivo. Dev Biol 1989;134:201-5.
[8] Tropepe V, Coles BL, Chiasson BJ, Horsford DJ, EliaAJ, McInnes RR,et al. Retinal stem cells in the adult mammalian eye. Science 2000;287:2032-6.
[9] Peichl L. Diversity of mammalian photoreceptor properties: adaptationto habitat and lifestyle? Anat Rec A Discov Mol Cell Evol Biol 2005;287:1001-12.
[10] HendricksonA.Amorphological comparison of foveal development inman and monkey. Eye 1992;6(Pt2):136-44.
[11] Ollivier FJ, Samuelson DA, Brooks DE, Lewis PA, Kallberg ME,Komáromy AM. Comparative morphology of the tapetum lucidum(among selected species). Vet Ophthalmol 2004;7:11-22.
[12] Hogan MJ, Alvarado JA, Weddell JE. Histology of the human eye: anatlas and textbook. Philadelphia: WB Saunders; 1971 (402p).
[13] Nobeschi L, Freymuller E, Smith RL. Intercellular junctions in rabbiteye ora serrata. Anat Histol Embryol 2006;35:287-92.
[14] DucasseA, Delattre JF, SegalA, BuretteA. L’artère centrale de la rétine.Aspects macroscopiques. À propos de 50 dissections. Bull Mem Soc FrOphtalmol 1983;95:574-7.
[15] MaharajAS, D’Amore PA. Roles for VEGF in the adult. Microvasc Res2007;74:100-13.
[16] Curcio CA, Sloan KR, Kalina RE, Hendrickson AE. Humanphotoreceptor topography. J Comp Neurol 1990;292:497-523.
[17] Curcio CA, Millican CL, Allen KA, Kalina RE. Aging of the humanphotoreceptor mosaic: evidence for selective vulnerability of rods incentral retina. Invest Ophthalmol Vis Sci 1993;34:3278-96.
Figure 14. Représentation des cellules gliales de Müller.A. Coupe d’une rétine de souris en semi-fine marquée au bleu de toluidineet représentation schématique en regard d’une cellule gliale de Müller(qui n’est pas à l’échelle) montrant les rapports entre les différentes partiesde la cellule et les couches de la rétine. EPR : épithélium pigmentaire de larétine ; SE : segments externes ; SI : segments internes ; LE : limitanteexterne ; CNE : couche nucléaire externe ; CNI : couche nucléaire interne.1. Pied ; 2. prolongements internes ; 3. corps ; 4. prolongements exter-nes ; 5. microvilli.B. Marquage de cellules gliales de Müller sur une coupe de rétine de rat,par un anticorps dirigé contre la protéine gliale fibrillaire, marqué à lafluorescéine (GFAP).
.
Anatomie de la rétine ¶ 21-003-C-40
13Ophtalmologie

[18] Hsu SY, Tsai RK. Analysis of retinal nerve fiber layer and macularthickness measurements in healthy Taiwanese individuals using opticalcoherence tomography (Stratus OCT). J Glaucoma 2008;17:30-5.
[19] Cohen MJ, Kaliner E, Frenkel S, Kogan M, Miron H, Blumenthal EZ.Morphometric analysis of human peripapillary retinal nerve fiber layerthickness. Invest Ophthalmol Vis Sci 2008;49:941-4.
[20] Gass JD. Müller cell cone, an overlooked part of the anatomy of thefovea centralis. Arch Ophthalmol 1999;117:821-3.
[21] Ohta K, Ito A, Tanaka H. Neuronal stem/progenitor cells in thevertebrate eye. Dev Growth Differ 2008;50:253-9.
[22] Franze K, Grosche J, Skatchkov SN, Schinkinger S, Foja C, Schild D,et al. Müller cells are living optical fibers in the vertebrate retina. ProcNatl Acad Sci USA 2007;104:8287-92.
[23] Ogden TE. Nerve fiber layer astrocytes of the primate retina:morphology, distribution, and density. Invest Ophthalmol Vis Sci1978;17:499-510.
F. Behar-Cohen ([email protected]).Service d’ophtalmologie, Hôtel-Dieu de Paris, 1, place du Parvis-Notre-Dame, 75004 Paris, France.UMR S872, Physiopathologie des maladies oculaires : innovations thérapeutiques, Centre de recherches des cordeliers, 15, rue de l’École-de-Médecine,75006 Paris, France.
L. Kowalczuk.N. Keller.UMR S872, Physiopathologie des maladies oculaires : innovations thérapeutiques, Centre de recherches des cordeliers, 15, rue de l’École-de-Médecine,75006 Paris, France.
M. Savoldelli.F. Azan.Service d’ophtalmologie, Hôtel-Dieu de Paris, 1, place du Parvis-Notre-Dame, 75004 Paris, France.
J.-C. Jeanny.UMR S872, Physiopathologie des maladies oculaires : innovations thérapeutiques, Centre de recherches des cordeliers, 15, rue de l’École-de-Médecine,75006 Paris, France.
Toute référence à cet article doit porter la mention : Behar-Cohen F., Kowalczuk L., Keller N., Savoldelli M., Azan F., Jeanny J.-C. Anatomie de la rétine. EMC(Elsevier Masson SAS, Paris), Ophtalmologie, 21-003-C-40, 2009.
Disponibles sur www.em-consulte.com
Arbresdécisionnels
Iconographiessupplémentaires
Vidéos /Animations
Documentslégaux
Informationau patient
Informationssupplémentaires
Auto-évaluations
21-003-C-40 ¶ Anatomie de la rétine
14 Ophtalmologie