






Langages
Pages
Légal


UMR7242 CNRS – UnistraBSC ESBS, 300 boulevard Sébastien Brant, BP 10413 F-67412 Illkirch Cedex, FranceTel . 03 68 85 47 10http://bsc.unistra.fr/

Jérôme Wagner Modifications Post Traductionnelles et Cancérogenèse UMR 7242 Biologie et Signalisation Cellulaire (BSC) E-mail [email protected]
Fidélité de la Réplication, Dommages à L’ADN, Réparation par Réversion Directe et Mutagenèse
Principale source pour les illustrations: “DNA Repair and Mutagenesis ”, 1st and 2nd Ed., E. Friedberg et al., ASM Press
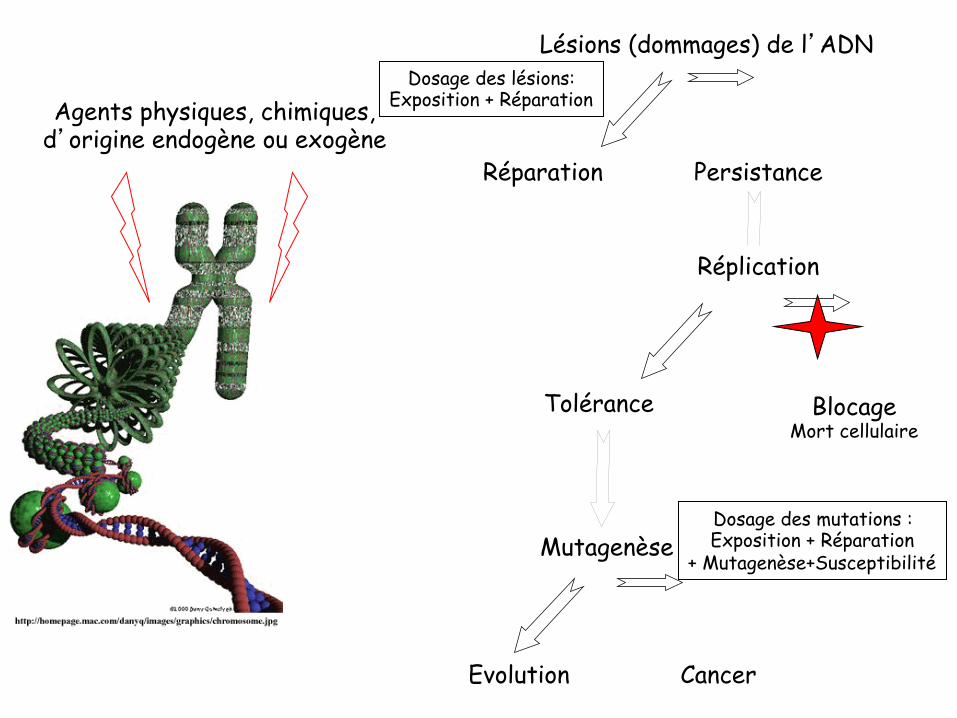
Agents physiques, chimiques, d’origine endogène ou exogène
Lésions (dommages) de l’ADN
Réparation
Tolérance
Mutagenèse
Cancer Evolution
Blocage Mort cellulaire
Persistance
Réplication
Dosage des lésions: Exposition + Réparation
Dosage des mutations : Exposition + Réparation
+ Mutagenèse+Susceptibilité

II. Origines et diversité des dommages dans l ’ADN Détection des lésions
III. Mécanismes de tolérances des lésions, définitions relatives à la mutagenèse
Détection des mutations
Notion de réparation par réversion directe
I. Bases de la fidélité de la réplication
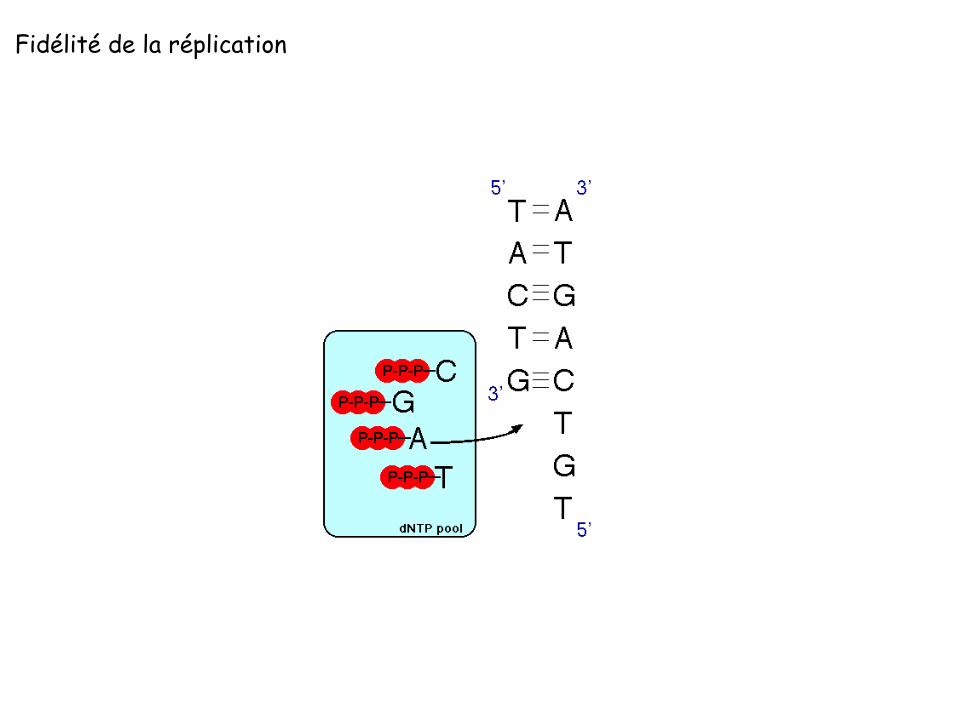
Fidélité de la réplication

La vitesse de réplication peut atteindre plus de1000 nucléotides par seconde chez la bactérie,
50 chez les humains.

Bases de la fidélité de la réplication: Appariement des bases (Watson Crick)
ΔG (correcte/incorrecte) 2-3 kcal/mol
Fidélité de la réplication
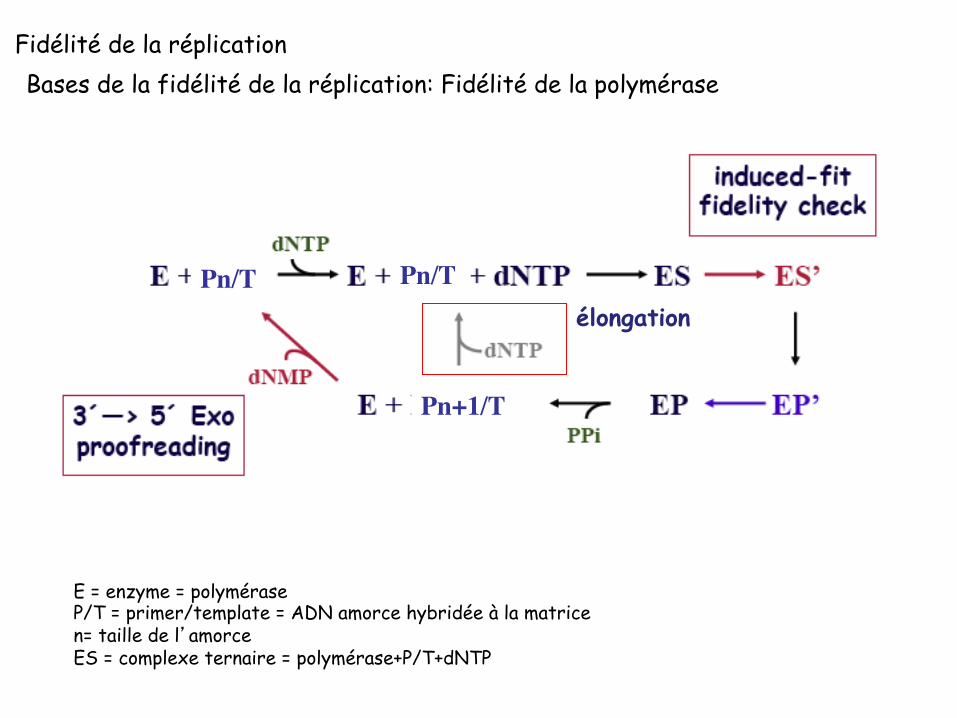
Bases de la fidélité de la réplication: Fidélité de la polymérase
E = enzyme = polymérase P/T = primer/template = ADN amorce hybridée à la matrice n= taille de l’amorce ES = complexe ternaire = polymérase+P/T+dNTP
Pn/T Pn/T
Pn+1/T
élongation
Fidélité de la réplication
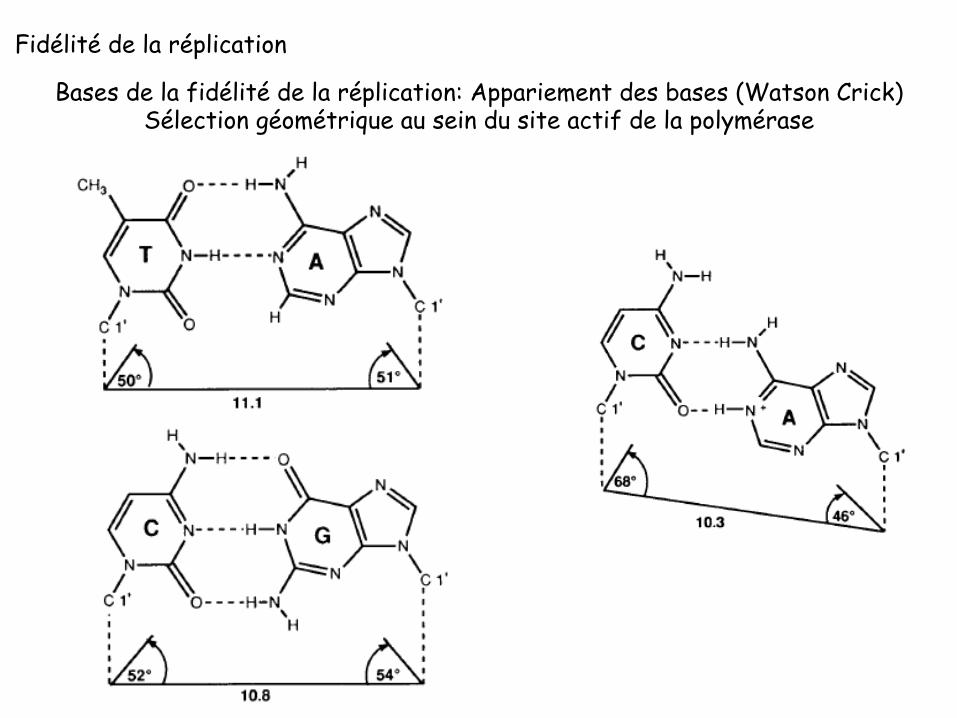
Bases de la fidélité de la réplication: Appariement des bases (Watson Crick) Sélection géométrique au sein du site actif de la polymérase
Fidélité de la réplication

en noir, valeurs pour un nucléotide correcten rouge, valeurs pour un nucléotide incorrect
Fidélité de la réplication
Aspect cinétique de polymérisation

Correction des mésappariements ou «mismatch repair (MMR)
Corrige les erreurs des ADN polymérases => augmente la fidélité de la réplication au moins 100 fois
Contrôle la recombinaison homologue en empêchant l’échange de brins entre séquences divergentes
4 étapes
1: recrutement de facteurs qui reconnaissent le mésappariements 2: recherche d’un signal discriminant le brin néosynthétisé 3: dégradation de la portion d’ADN contenant la mauvaise base 4: re-synthèse de la portion d’ADN excisé
Chez l’homme, un défaut dans MMR est responsable de la maladie HNPCC: cancer colorectal sans polypose (2% des cas de cancer colorectal; env. 750 nouveaux cas par an en France)
Fidélité de la réplication

Correction des mésappariements ou «mismatch repair (MMR) Fidélité de la réplication
Polδ Polδ
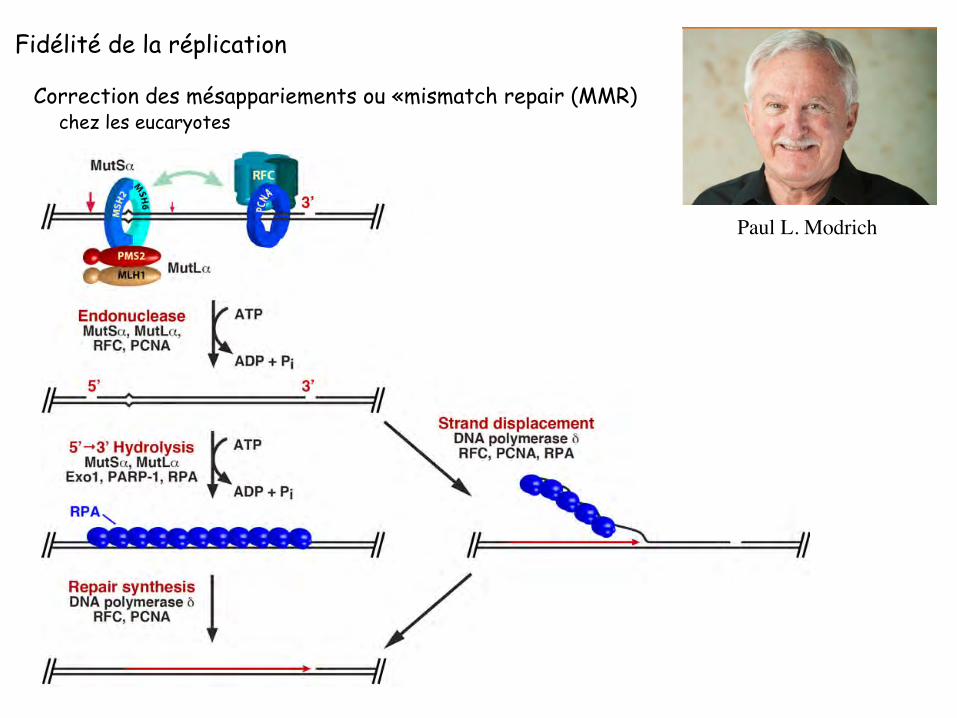
Correction des mésappariements ou «mismatch repair (MMR) chez les eucaryotes
Fidélité de la réplication
Paul L. Modrich

Aspect quantitatif
Mécanisme Facteur de fidélité Appariement des bases (W.C.) 101-102 Fidélité de l’ADN polymérase réplicative 105- 106 (proofreading inclus)
Protéines accessoires (SSB…) 107
Correction des mésappaiements 1010
(Mismatch Repair)
Fidélité de la réplication
1 erreur sur 1010 nucléotides incorporés

Facteurs pouvant affecter la fidélité de la réplication
Inactivation des mécanismes de réparation des mésappariements
Modification de la balance du pool des nucléotides
Inactivation de l ’activité de correction de lecture
Mutations dans le site actif de la polymérase
Lésions de l ’ADN
Modifications de bases dans le pool des nucléotides (mésappariements possibles: dA/8-oxo-dGTP substitution de base)
Contexte de séquence (séquences répétées décalage du cadre de lecture)
AAF
10-1 x 107 ATGG 5 ’
TACCGCGGACA 3 ’ 5 ’
ATGGCCTGT 5 ’ TACCGGACA 3 ’ 5 ’ 10-8
Fréquence de mutation

II. Origines et diversité des dommages dans l ’ADN Détection des lésions
III. Mécanismes de tolérances des lésions, définitions relatives à la mutagenèse
Détection des mutations
Notions de réparation par réversion directe
I. Bases de la fidélité de la réplication

Agents endommageant d’origine endogène
Altérations spontanées des bases de l’ADN
Dommages oxydatifs

Altérations spontanées des bases de l’ADN Tautomérisation des bases
G(e) / T
A(i) / C
C(i) / A
T(e) / G
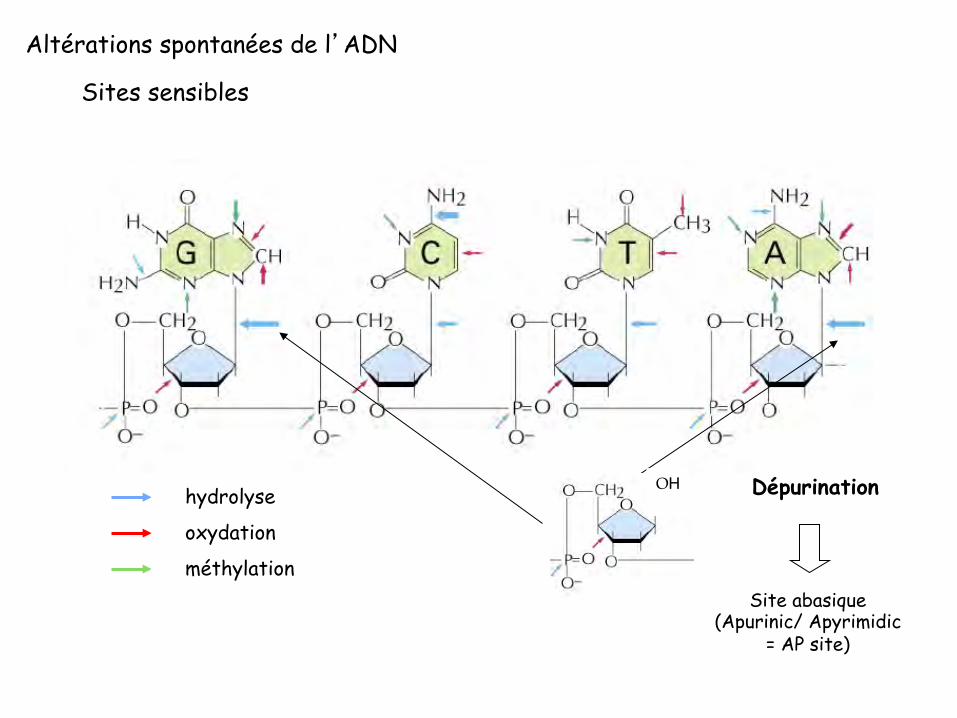
Altérations spontanées de l’ADN
Sites sensibles
hydrolyse
oxydation
méthylation Site abasique
(Apurinic/ Apyrimidic = AP site)
Dépurination OH
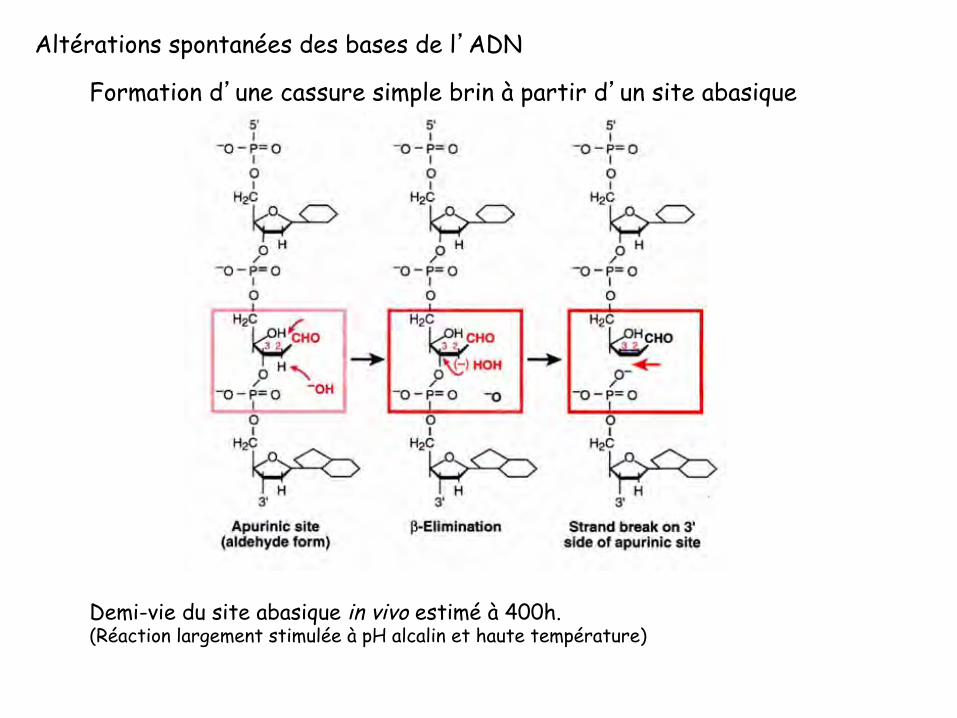
Formation d’une cassure simple brin à partir d’un site abasique
Altérations spontanées des bases de l’ADN
Demi-vie du site abasique in vivo estimé à 400h. (Réaction largement stimulée à pH alcalin et haute température)
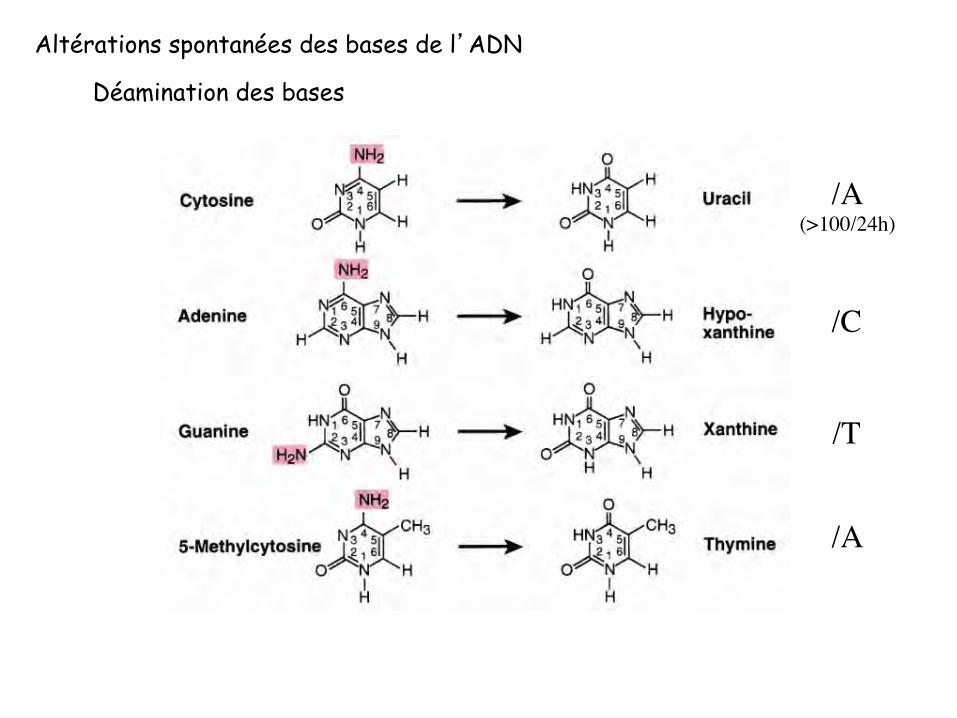
Altérations spontanées des bases de l’ADN
Déamination des bases
/A(>100/24h)
/C
/A
/T
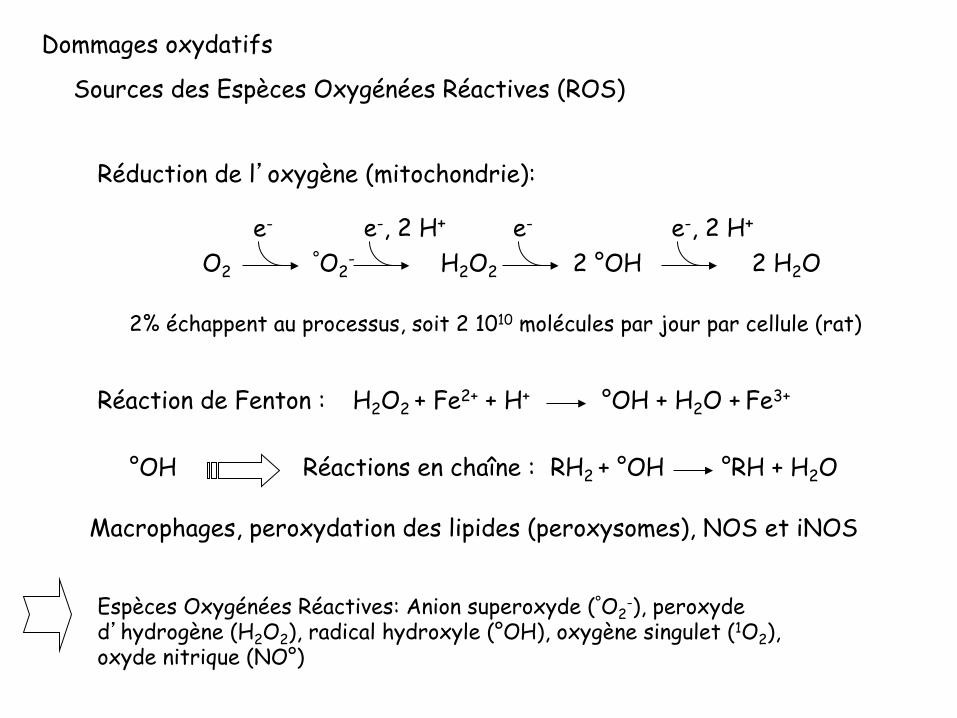
Dommages oxydatifs
Sources des Espèces Oxygénées Réactives (ROS)
Réduction de l’oxygène (mitochondrie):
Espèces Oxygénées Réactives: Anion superoxyde (°O2-), peroxyde
d’hydrogène (H2O2), radical hydroxyle (°OH), oxygène singulet (1O2), oxyde nitrique (NO°)
O2 °O2- 2 °OH 2 H2O
e-, 2 H+
H2O2
e- e- e-, 2 H+
2% échappent au processus, soit 2 1010 molécules par jour par cellule (rat)
Macrophages, peroxydation des lipides (peroxysomes), NOS et iNOS
RH2 + °OH °RH + H2O Réactions en chaîne :
Réaction de Fenton : H2O2 + Fe2+ + H+ °OH + H2O + Fe3+
°OH

Dommages oxydatifs
Compartimentalisation : Noyau/mitochondrie/peroxysome
Défenses
Séquestration des métaux de transition (Fe2+ par ferritine)
SuperOxyde Dismutase (SOD): 2 O2° + 2 H+ 2 H2O2 + O2
Catalase: 2H2O2 2 H2O + O2
Glutathion peroxydase: H2O2 + 2 GSH 2 H2O + GS-SG
Systèmes enzymatiques
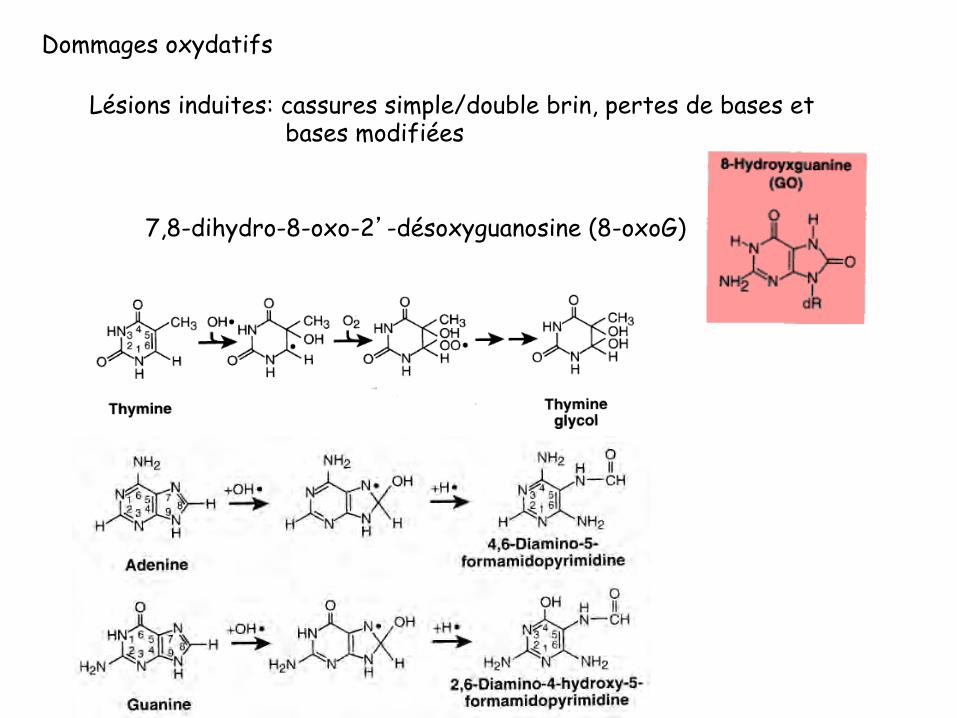
Dommages oxydatifs
Lésions induites: cassures simple/double brin, pertes de bases et bases modifiées
7,8-dihydro-8-oxo-2’-désoxyguanosine (8-oxoG)
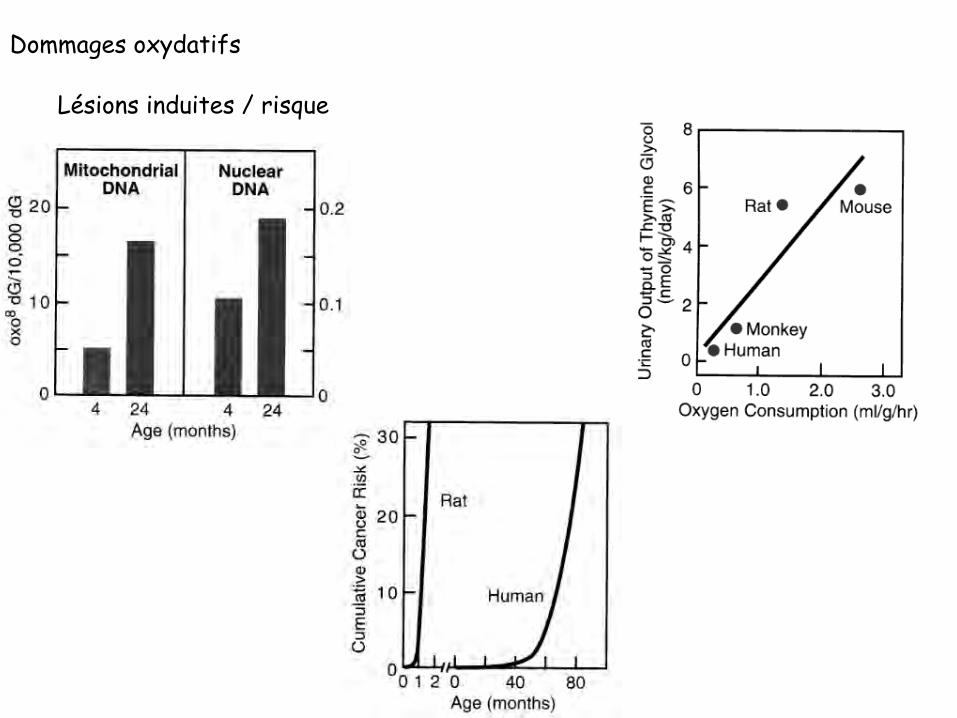
Dommages oxydatifs
Lésions induites / risque

Altérations spontanées des bases de l’ADN Aspect quantitatif
> 30 000 lésions spontanées / jour/ cellule
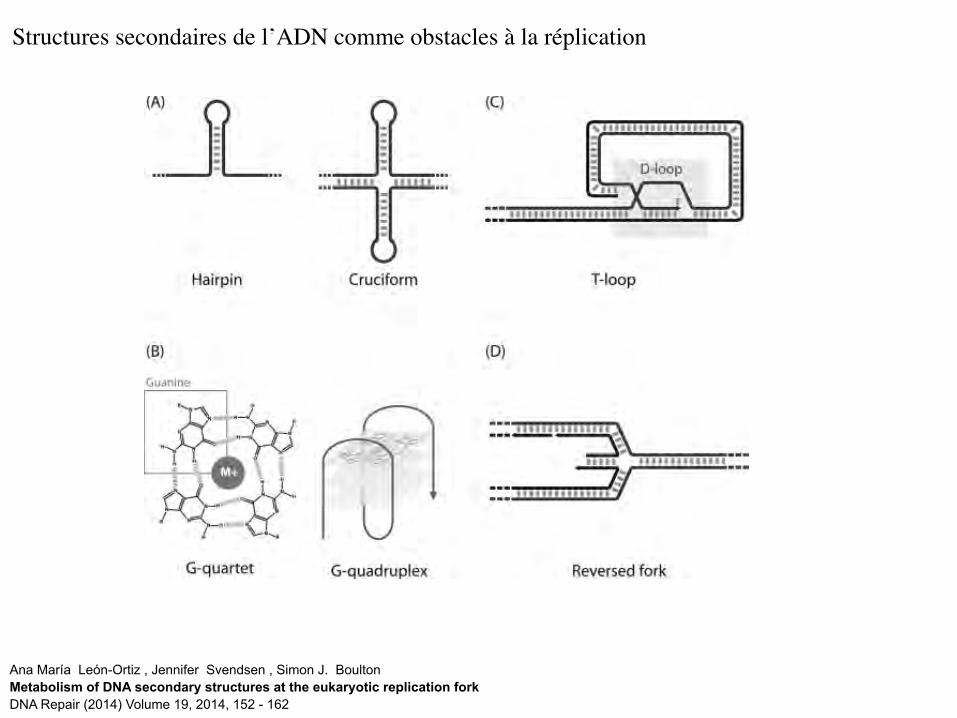
Ana María León-Ortiz , Jennifer Svendsen , Simon J. Boulton Metabolism of DNA secondary structures at the eukaryotic replication fork DNA Repair (2014) Volume 19, 2014, 152 - 162
Structures secondaires de l’ADN comme obstacles à la réplication

Agents endommageant d’origine exogène
Radiations ionisantes
Rayonnement UV
Agents chimiques
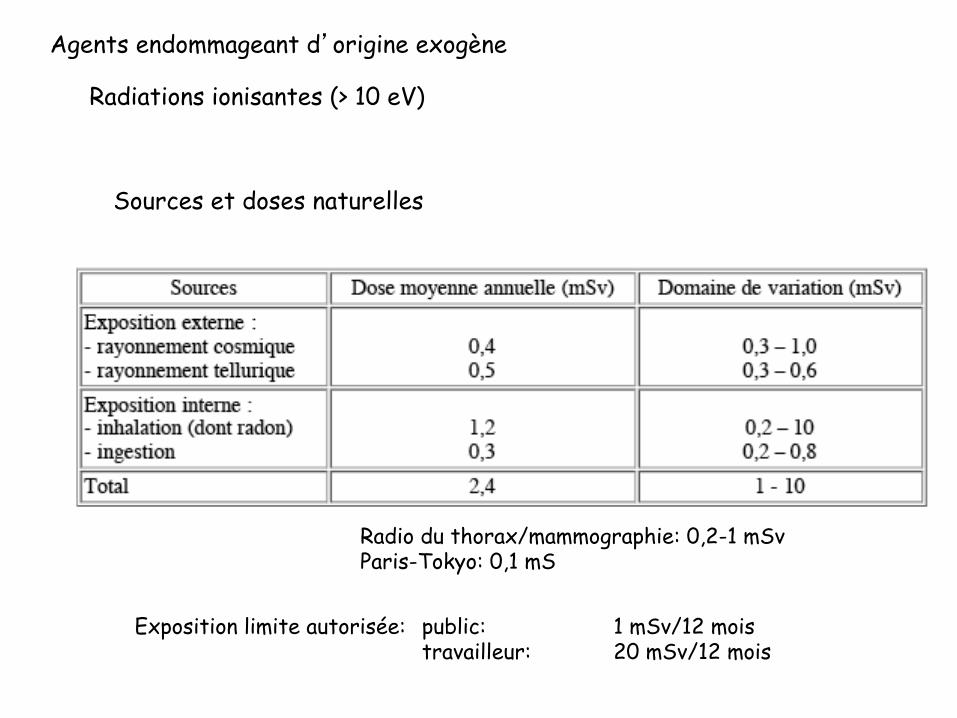
Agents endommageant d’origine exogène
Radiations ionisantes (> 10 eV)
Sources et doses naturelles
Radio du thorax/mammographie: 0,2-1 mSv Paris-Tokyo: 0,1 mS
Exposition limite autorisée: public: 1 mSv/12 mois travailleur: 20 mSv/12 mois

Agents endommageant d’origine exogène
Radiations ionisantes
Interaction de l’énergie de la radiation avec l’ADN
ou
Interaction de l’énergie de la radiation avec des composants cellulaires autre que l’ADN qui vont eux réagir avec la macromolécule
H2O H2O+ + e-
H2O+ + H2O+ °OH + H3O+
°OH + °OH H2O2
Radiolyse de l’eau :
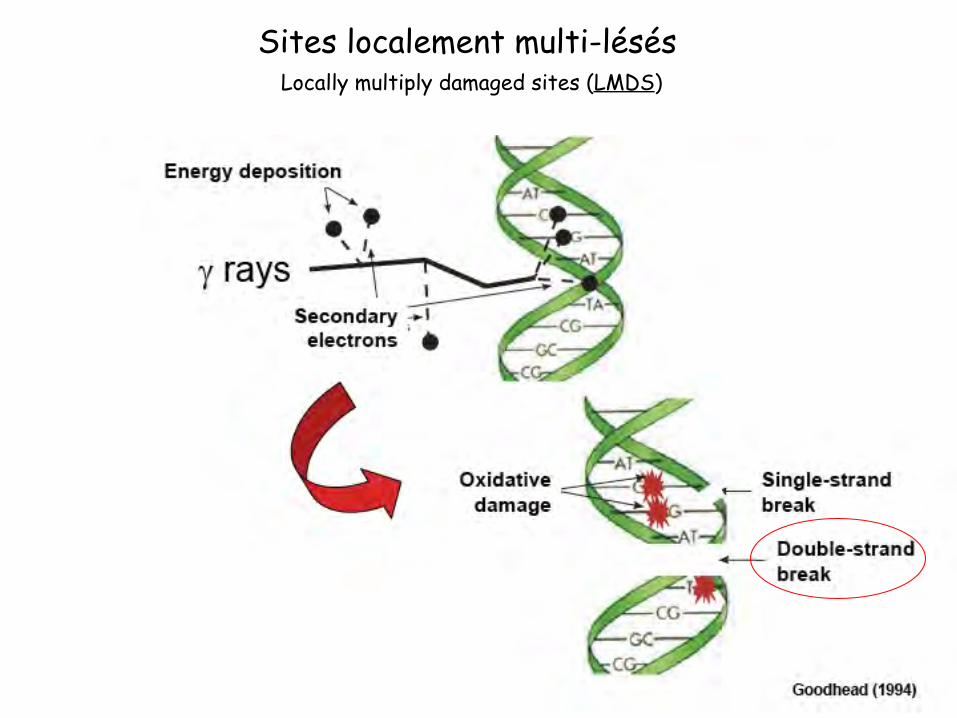
Locally multiply damaged sites (LMDS) Sites localement multi-lésés

Radiations ionisantes
a. Altération du squelette sucre-phosphate
Perte de base Altération du sucre
Cassure simple ou double brin Altération du groupement phosphate
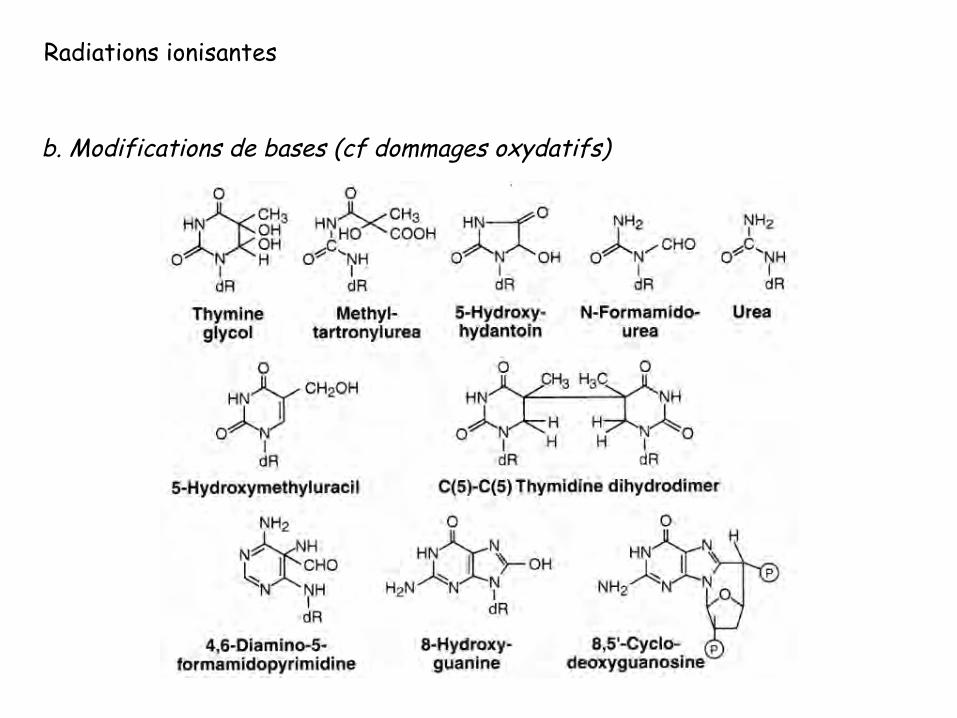
Radiations ionisantes
b. Modifications de bases (cf dommages oxydatifs)
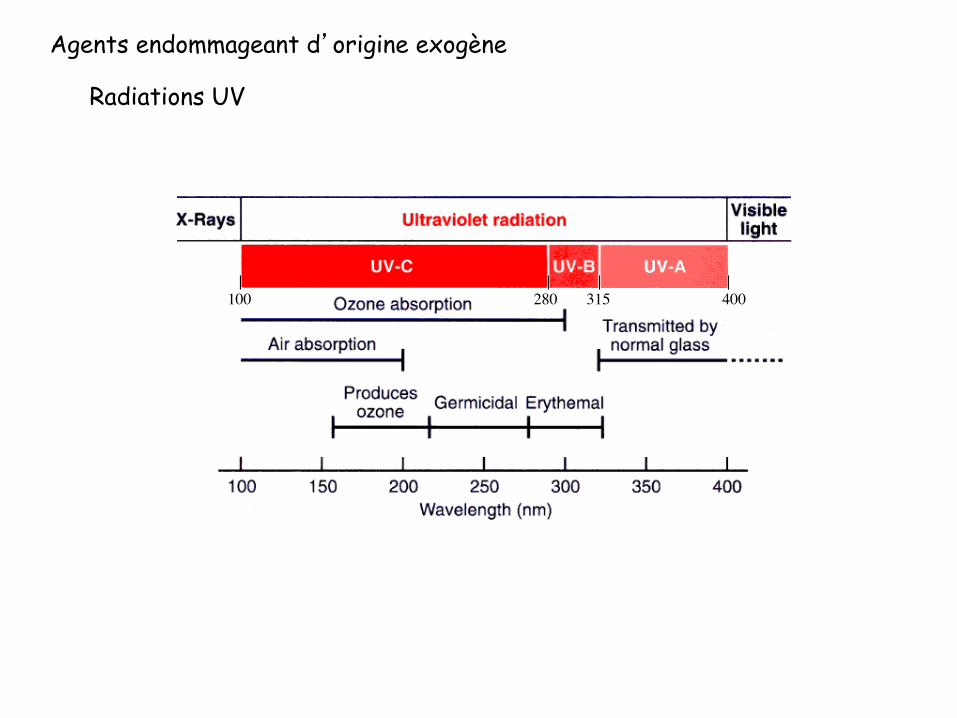
Agents endommageant d’origine exogène
Radiations UV
100 280 315 400
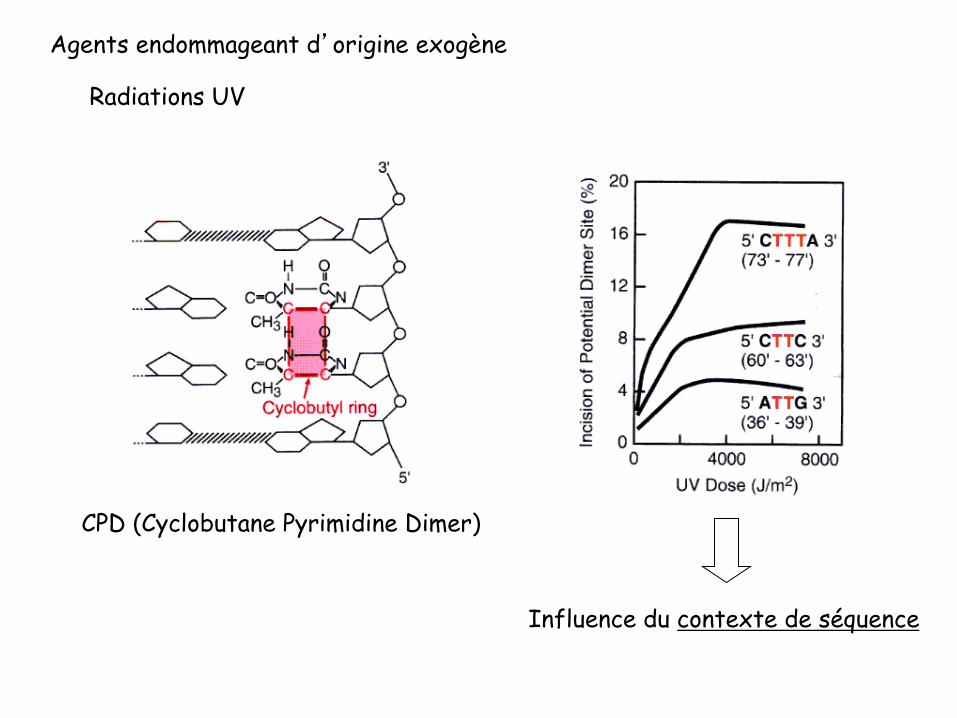
CPD (Cyclobutane Pyrimidine Dimer)
Influence du contexte de séquence
Agents endommageant d’origine exogène
Radiations UV

Agents endommageant d’origine exogène
Radiations UV
Photoproduit Pyrimidine-Pyrimidone (6-4) en moyenne 3 fois moins nombreux que les CPDs
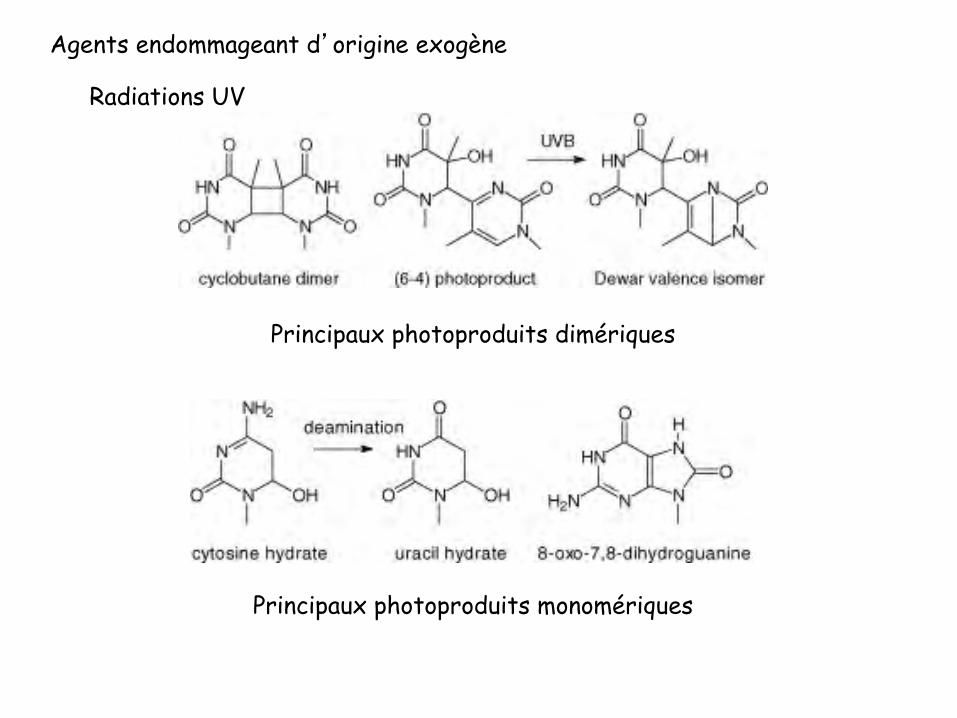
Principaux photoproduits dimériques
Principaux photoproduits monomériques
Agents endommageant d’origine exogène
Radiations UV

Agents endommageant d’origine exogène
Agents chimiques
Agents Alkylants (MMS, MNU, MNNG) Agents de pontage (Azote Moutarde, cis-Platine)
Intercalants (Psoralène)
Agents chimiques nécessitant une activation métabolique
Amines aromatiques (AAF) PHA (Hydrocarbures Polycycliques Aromatiques; B(a)P)
Aflatoxines

Agents endommageant d’origine exogène
Agents alkylants
Centres nucléophiles des bases

Agents endommageant d’origine exogène
Agents alkylants
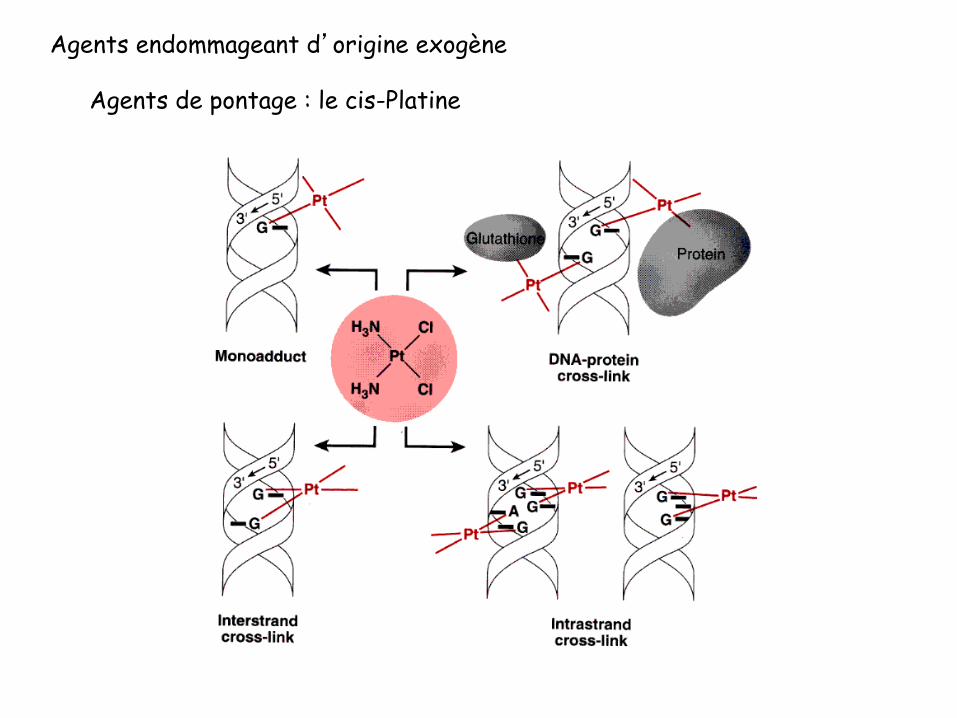
Agents endommageant d’origine exogène
Agents de pontage : le cis-Platine
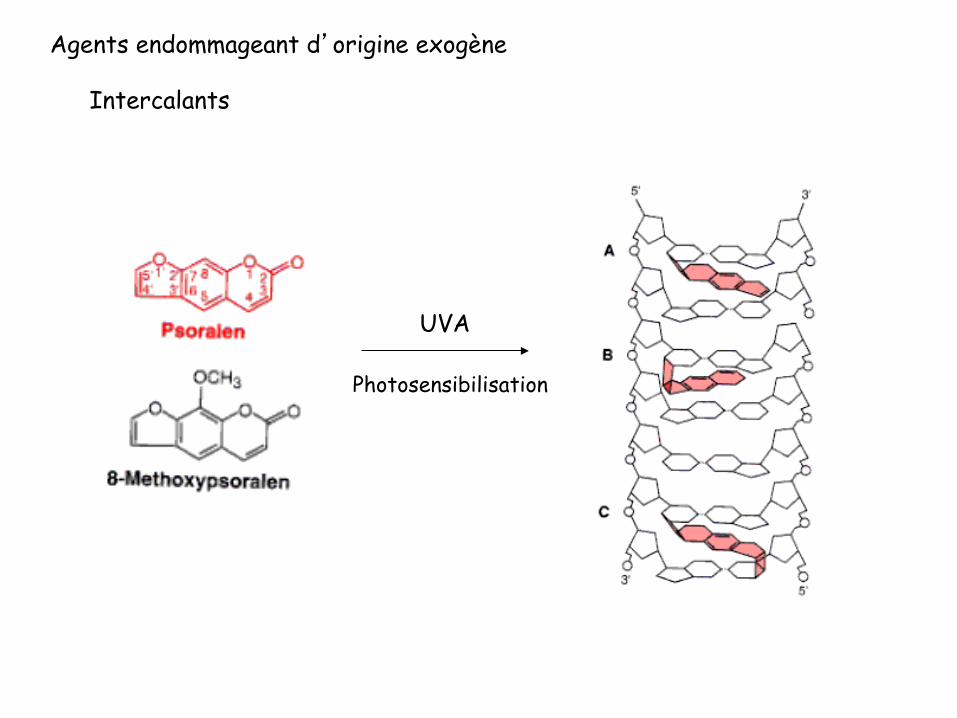
Agents endommageant d’origine exogène
Intercalants
UVA
Photosensibilisation

Agents endommageant d’origine exogène
Déformation de l’hélice d’ADN par un pontage par le psoralène
46.5 ° “Unwinding“
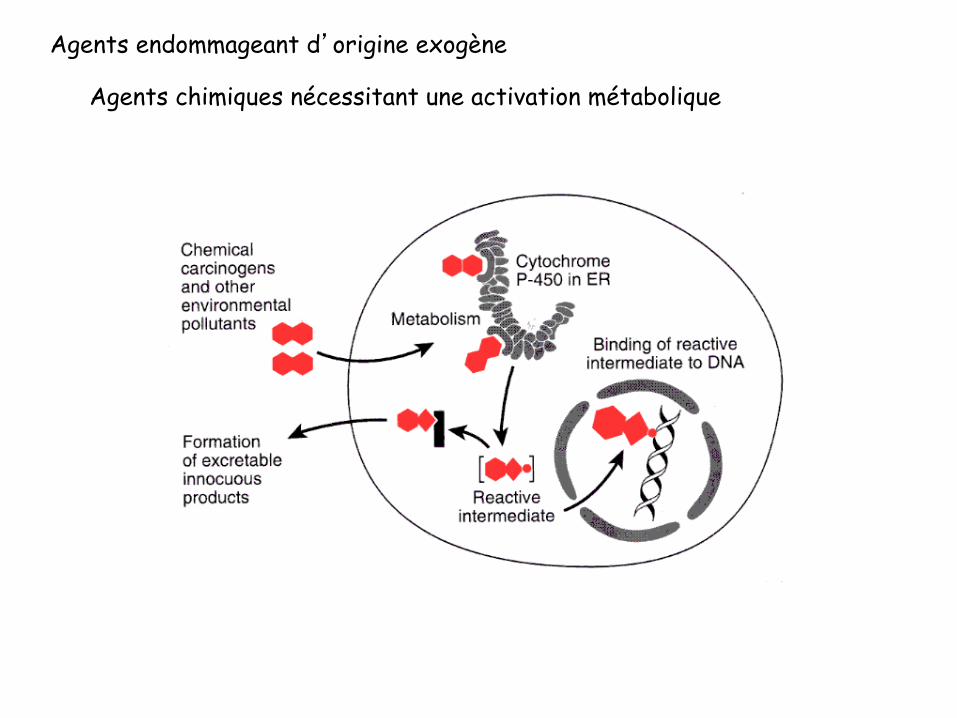
Agents endommageant d’origine exogène
Agents chimiques nécessitant une activation métabolique
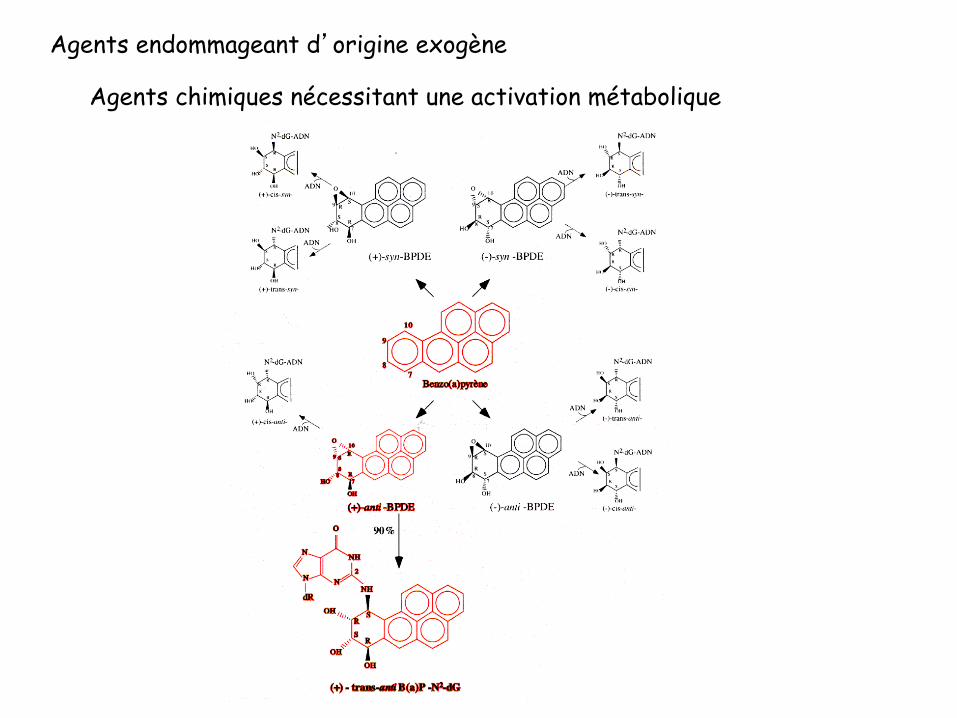
Agents endommageant d’origine exogène
Agents chimiques nécessitant une activation métabolique
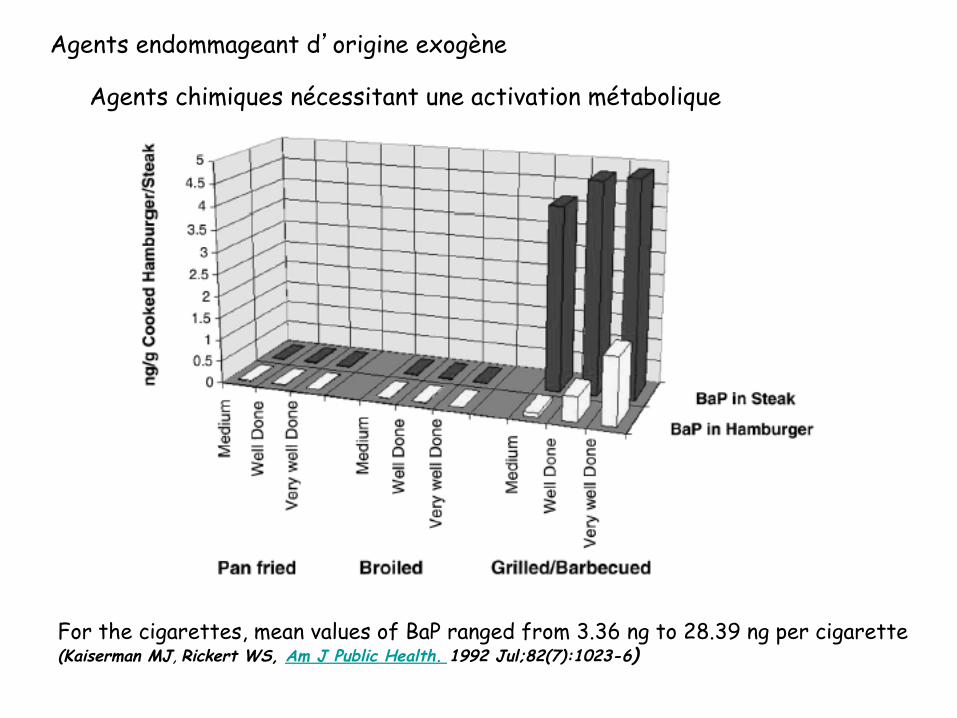
Agents endommageant d’origine exogène
Agents chimiques nécessitant une activation métabolique
For the cigarettes, mean values of BaP ranged from 3.36 ng to 28.39 ng per cigarette (Kaiserman MJ, Rickert WS, Am J Public Health. 1992 Jul;82(7):1023-6)
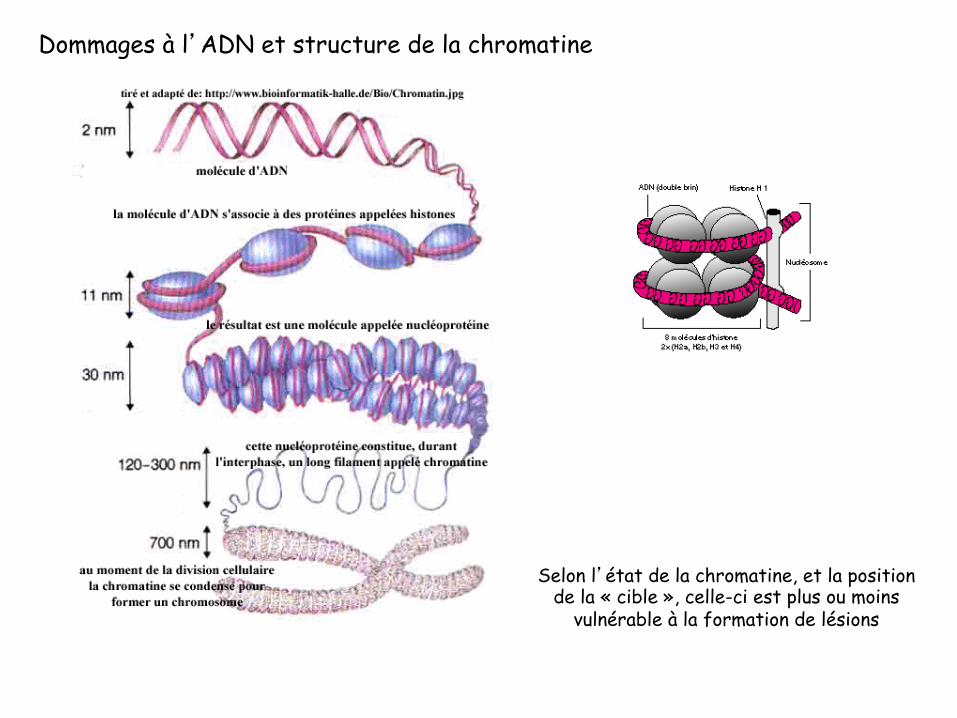
Dommages à l’ADN et structure de la chromatine
Selon l’état de la chromatine, et la position de la « cible », celle-ci est plus ou moins
vulnérable à la formation de lésions

Dommages à l’ADN et structure de la chromatine

Copyright ©2005 by the National Academy of Sciences
Besaratinia, Ahmad et al. (2005) Proc. Natl. Acad. Sci. USA 102, 10058-10063
Quantifying CPDs in the genome by immuno-dot blot assay
Détection des Dommages à l’ADN
Détection de lésions UV par dosage immunochimique
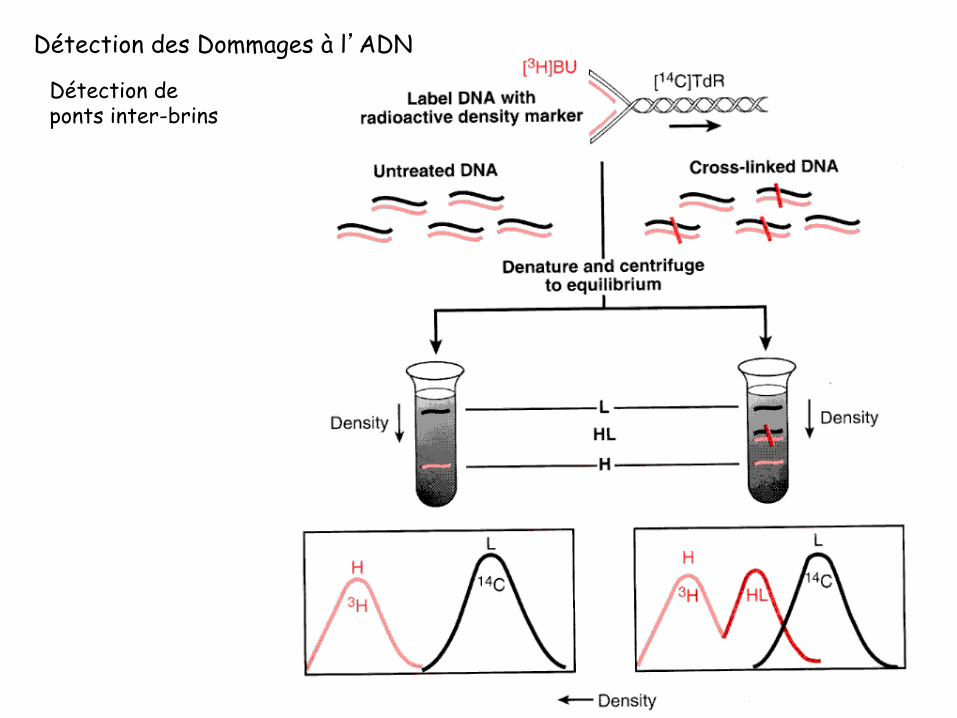
Détection des Dommages à l’ADN
Détection de ponts inter-brins
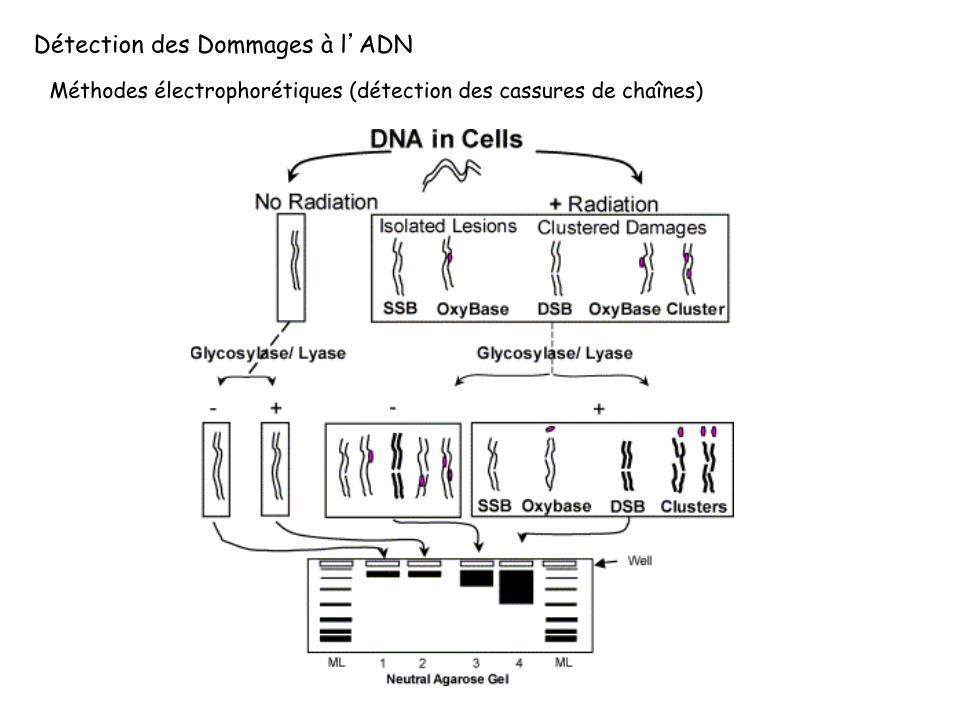
Détection des Dommages à l’ADN
Méthodes électrophorétiques (détection des cassures de chaînes)

Détection des Dommages à l’ADN
Principe de l’essai comète (Single Cell Gel electrophoresis, SCG)
Densité et taille de la queue proportionnelle à la quantité
de dommages
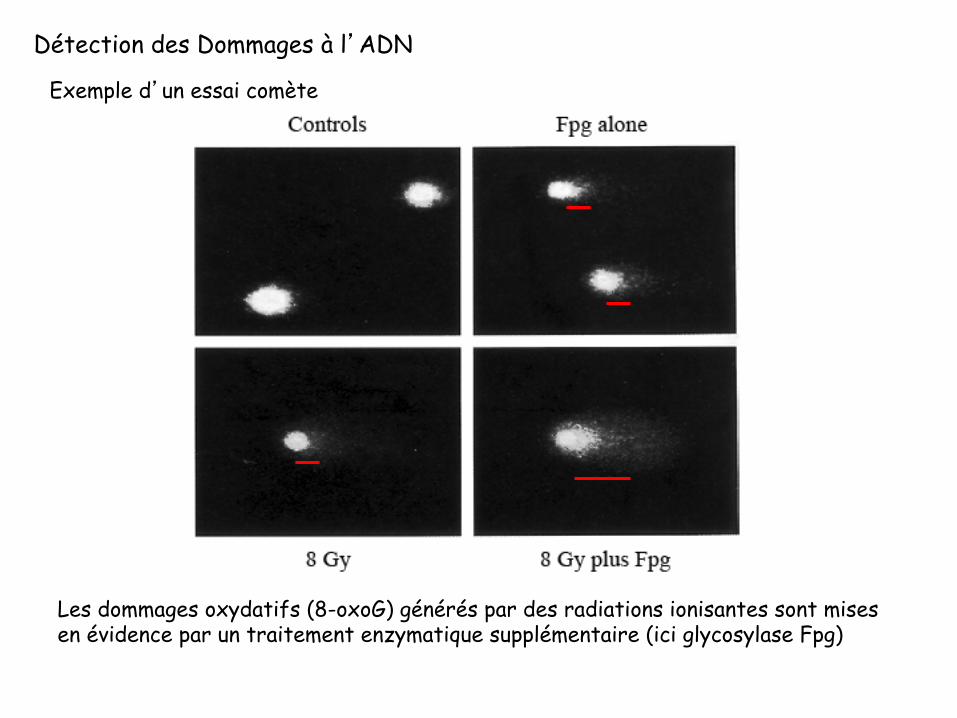
Détection des Dommages à l’ADN
Les dommages oxydatifs (8-oxoG) générés par des radiations ionisantes sont mises en évidence par un traitement enzymatique supplémentaire (ici glycosylase Fpg)
Exemple d’un essai comète

HPLC–MS/MS elution profile of the enzymatic digest of DNA of human keratinocytes that have been exposed either to UVB radiation (left panel) or successively to UVB and UVA radiations (right panel).
Détection des Dommages à l’ADN
Méthodes chromatographiques (détection des bases ou nucléotides modifiés)
ADN dNMPs Nucléases
Chromatographie liquide haute perf. (HPLC) + Spectrométrie de masse (MS)

Mais que fait la cellule ?
« Réverse » le dommage: restaure l’ADN sans excision
Enlève le dommage: Base Excision Repair (BER) et Nucleotide Excision Repair (NER) (implique l’incision de l’ADN et re-synthèse) Recombinaison
Tolère le dommage: Synthèse Translésionnelle (TLS; potentiellement mutagène) Contourne le dommage (mécanismes type recombinaison)
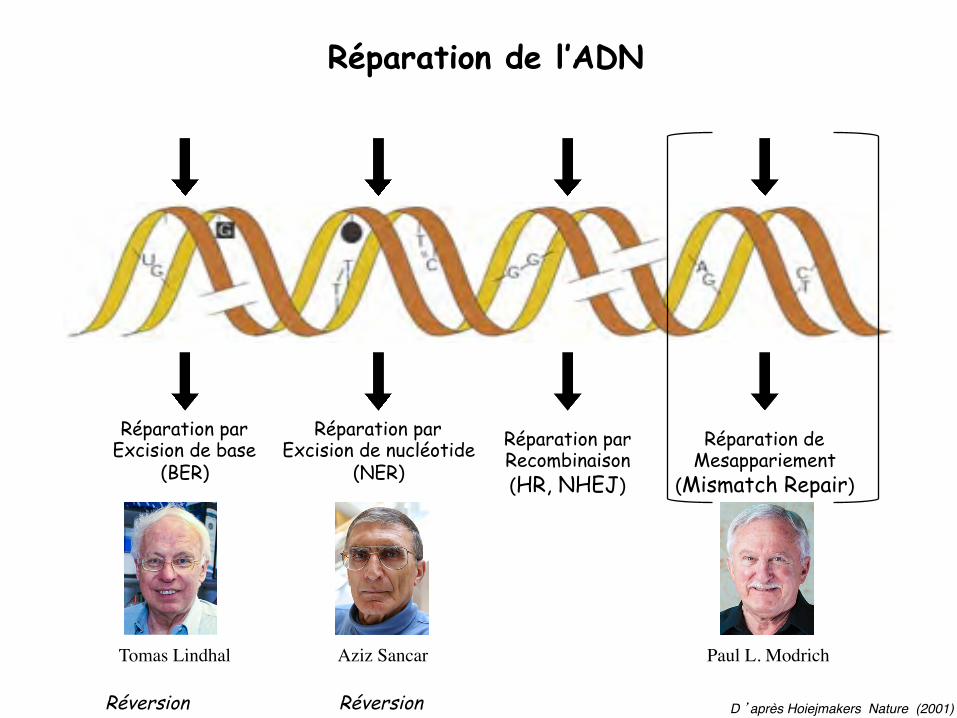
Réparation de l’ADN
D ’après Hoiejmakers Nature (2001)
Réparation par Excision de base
(BER)
Réparation par Excision de nucléotide
(NER)
Réparation par Recombinaison (HR, NHEJ)
Réparation de Mesappariement
(Mismatch Repair)
Réversion Réversion
Paul L. ModrichAziz SancarTomas Lindhal
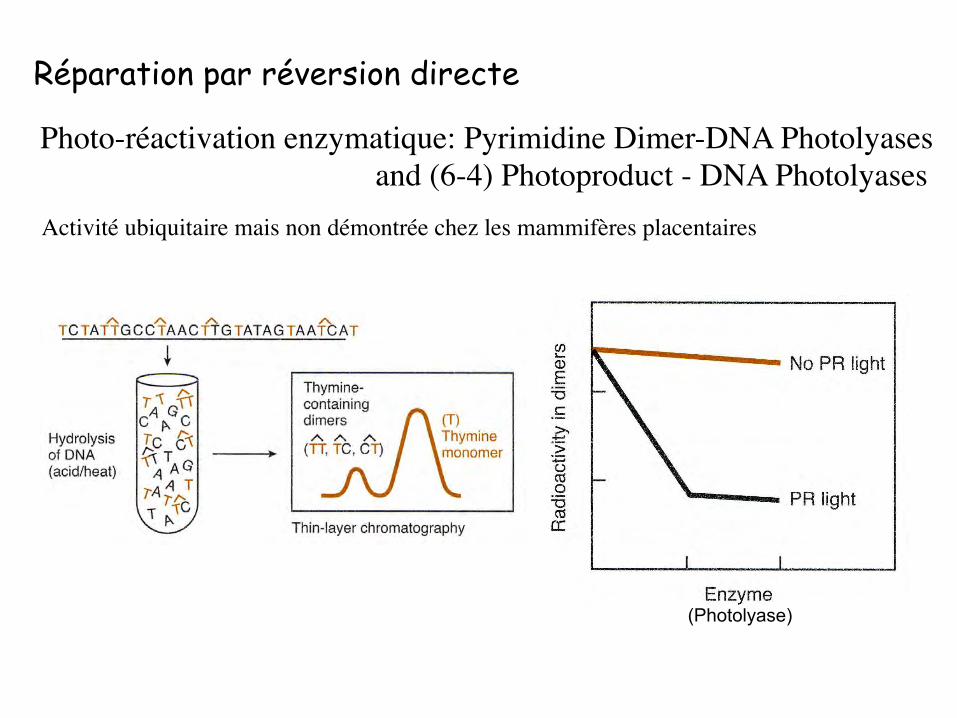
Photo-réactivation enzymatique: Pyrimidine Dimer-DNA Photolyases and (6-4) Photoproduct - DNA Photolyases
Réparation par réversion directe
Activité ubiquitaire mais non démontrée chez les mammifères placentaires
(Photolyase)
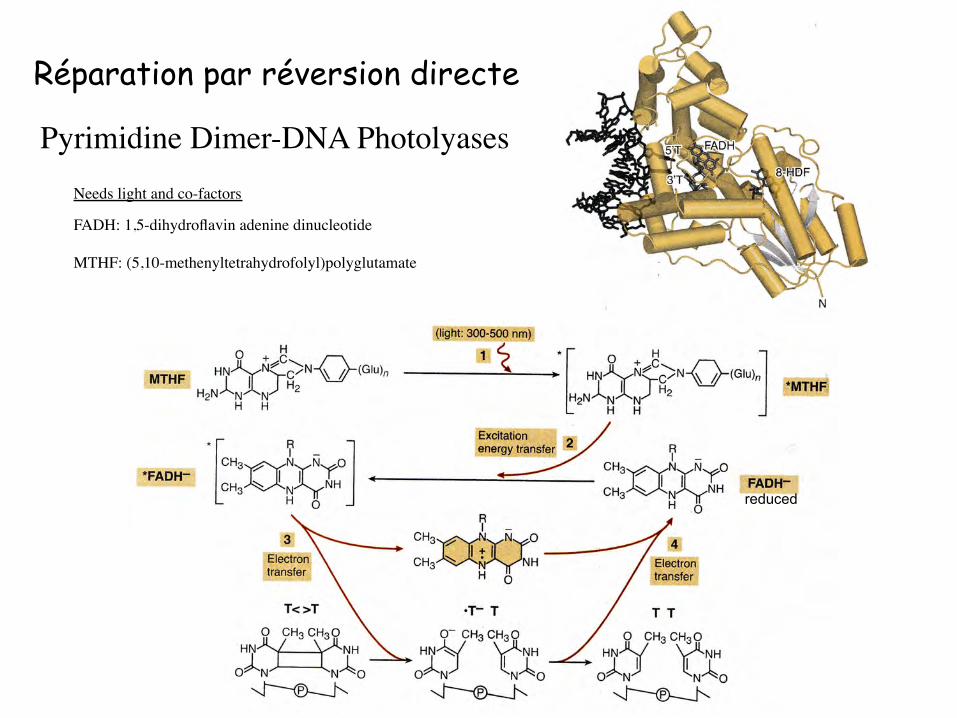
Pyrimidine Dimer-DNA Photolyases
Réparation par réversion directe
Needs light and co-factors
FADH: 1,5-dihydroflavin adenine dinucleotide
MTHF: (5,10-methenyltetrahydrofolyl)polyglutamate
reduced
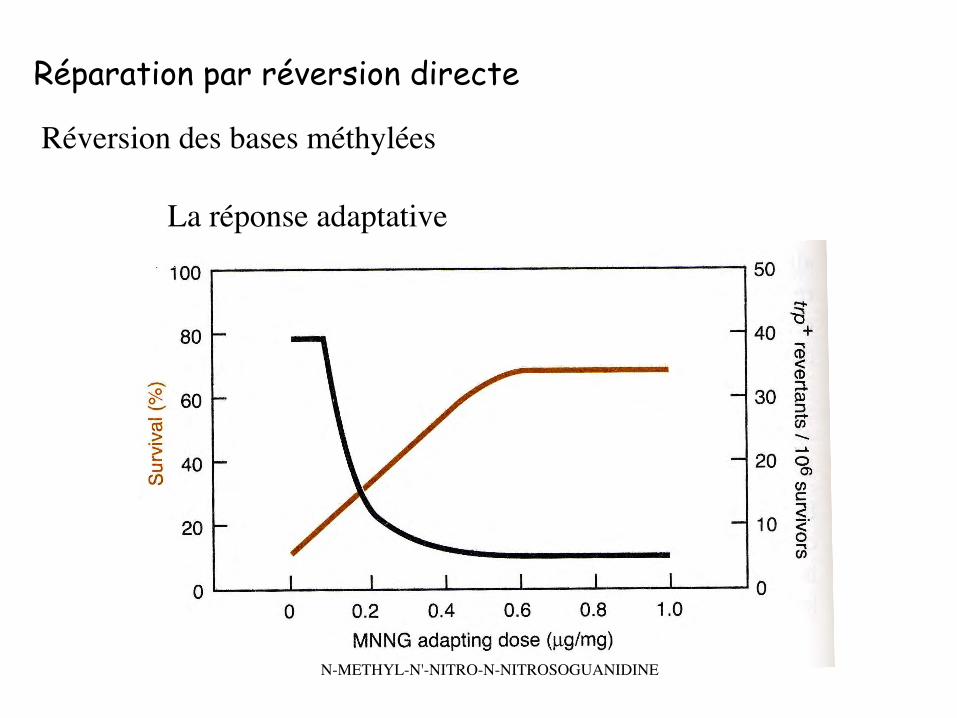
Réparation par réversion directe
Réversion des bases méthylées
La réponse adaptative
N-METHYL-N'-NITRO-N-NITROSOGUANIDINE

Réparation par réversion directe
Réversion des bases méthyléesProtéines impliquées
O6-methylguanine, O4-methylthymine: Ada (O6-AGT I) , O6-AGT II; O6-MGT
Enzymes« suicide »
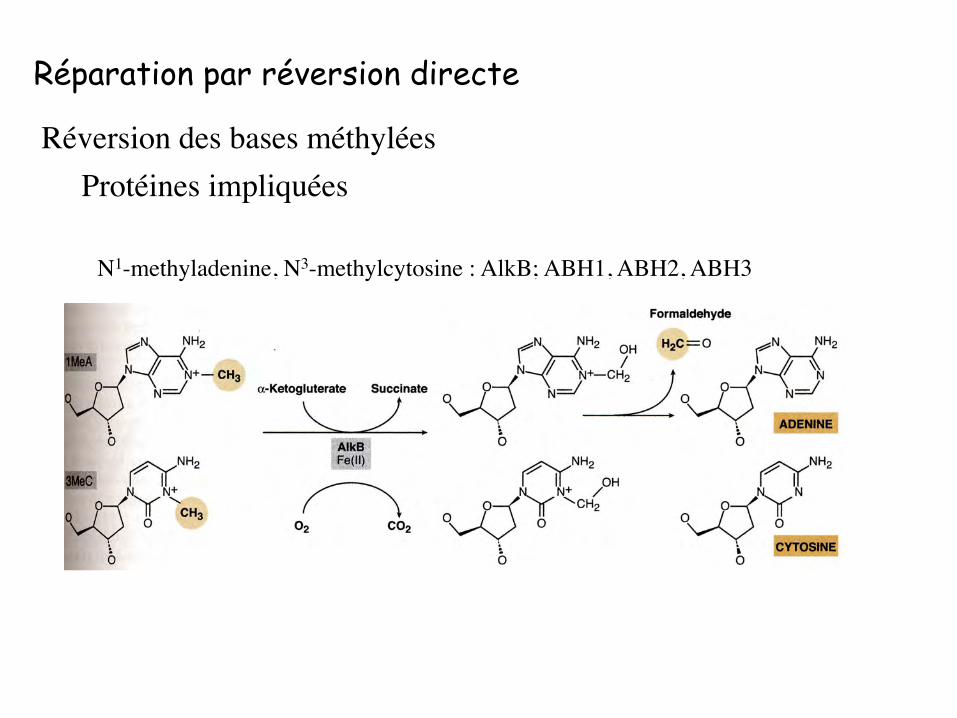
N1-methyladenine, N3-methylcytosine : AlkB; ABH1, ABH2, ABH3
Réparation par réversion directe
Réversion des bases méthyléesProtéines impliquées
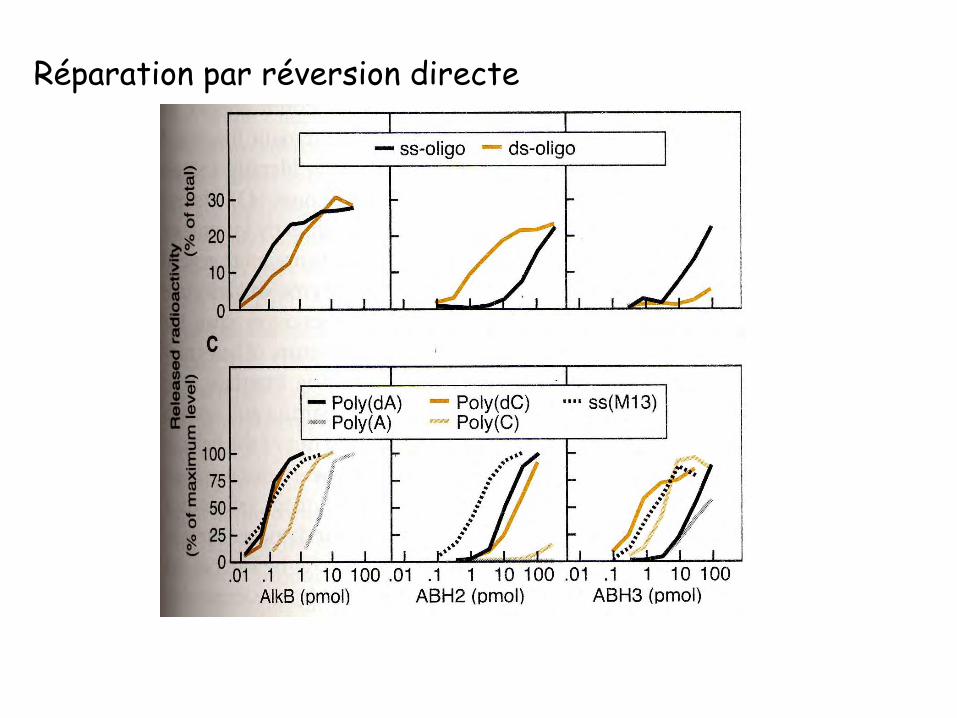
Réparation par réversion directe
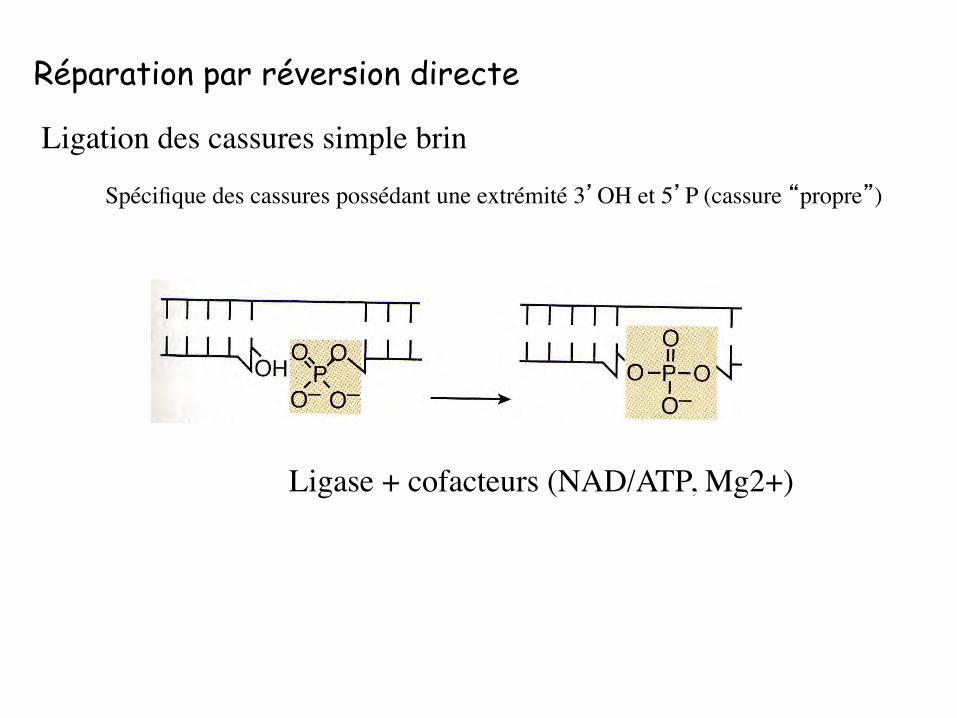
Réparation par réversion directe
Ligase + cofacteurs (NAD/ATP, Mg2+)
Ligation des cassures simple brin
Spécifique des cassures possédant une extrémité 3’OH et 5’P (cassure “propre”)

Réparation par réversion directe
Conclusions
Le produit d’un seul gène est requis: économie de l’informationgénétique. Le cas des O6-AGTs/MGT est énergétiquement coûteux
Cinétiquement avantageux car à priori plus rapide qu’un mécanismeen plusieurs étapes (BER, NER)
Mécanismes fidèles car hautement spécifiques et ne nécessitant pasde synthèse d’ADN post-excision

II. Origines et diversité des dommages dans l ’ADN Détection des lésions
III. Mécanismes de tolérances des lésions, définitions relatives à la mutagenèse
Détection des mutations
Notions de réparation par réversion directe
I. Bases de la fidélité de la réplication

Définitions relatives à la mutagenèse
Mutagenèse spontanée
Mutagenèse induite (Mécanismes de tolérances des lésions, Synthèse Translésionnelle)
Méthodes d’analyses

Quelques définitions:
Mutation: Changement, génétiquement transmissible, dans le génome d’un organisme ≠ lésion !
Génotype: Ensemble de l’information génétique portée par le génome d’un organisme On ne précise en général que les variations par rapport au génotype “sauvage“
Phénotype: Ensemble des caractéristiques observables d’un organisme (manifestation phénotypique d’une mutation)
Mutant: Un organisme qui contient une ou plusieurs mutations dans son génome par extension, on parle aussi de séquence mutante, de protéine mutante
Mutagène: Toute substance qui provoque une augmentation de la fréquence d’apparition de mutations
Mutagenèse: Le processus par lequel les mutations sont générées Mutagenèse spontanée: qui a lieu en absence de traitement exogène Mutagenèse induite: qui a lieu suite à un traitement exogène
Mutation conditionnelle: phénotype exprimé uniquement dans certaines conditions (exemples: mutation thermo-sensible, tissu/organe spécifique)
Mutation auxotrophe: qui cause un défaut dans la synthèse d’un métabolite essentiel
Mutation létale: qui rend l’organisme non viable

Les différentes classes de mutations
Mutations ponctuelles:
n’affectent qu’une ou quelques paires de bases: Substitutions de bases Décalage du cadre de lecture
Mutations impliquant des réarrangements plus importants:
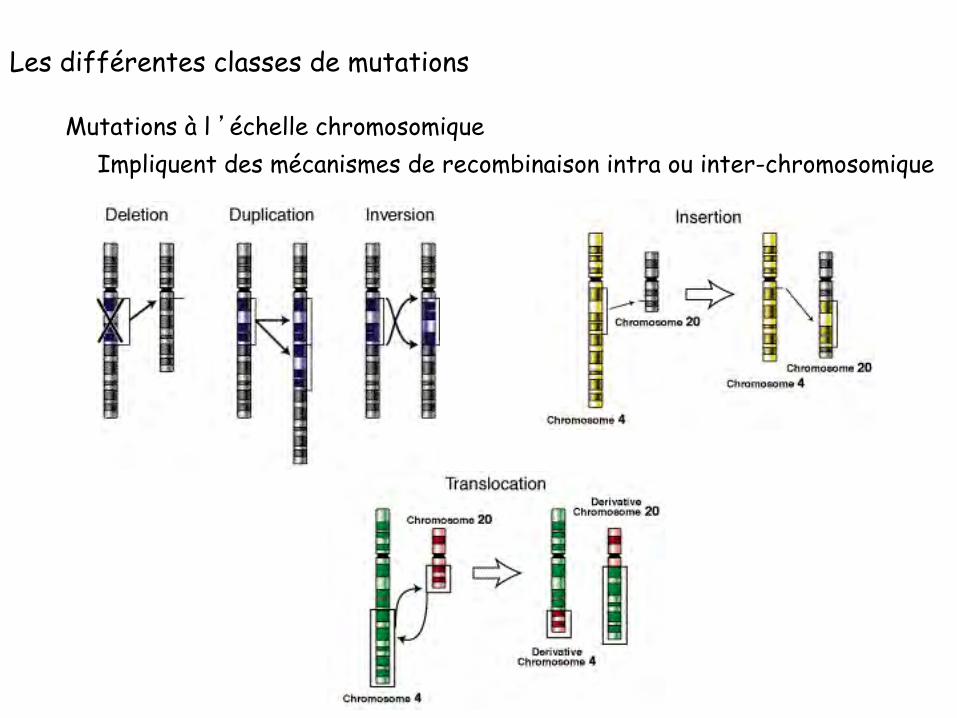
Mutations à l ’échelle chromosomique
Les différentes classes de mutations
Impliquent des mécanismes de recombinaison intra ou inter-chromosomique
A:T T:A
C:G G:C
TRANSITIONS
A:T T:A
C:G G:C
TRANSVERSIONS
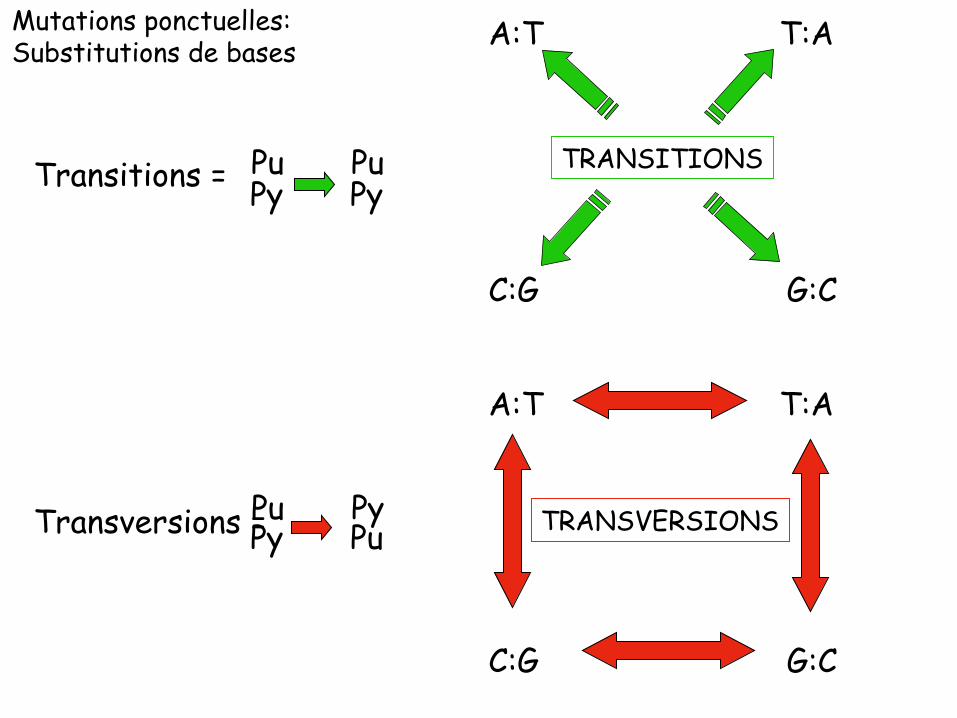
Mutations ponctuelles: Substitutions de bases
Pu Pu Py Py Transitions =
Pu Py Py Pu Transversions =

Mutations ponctuelles: Décalage du cadre de lecture (Frameshift)
= insertion ou délétion de (3n ± 1) nucléotide(s)
ADN : 5’ GCT GCT GCT GCT GCT GCT GCT GAT 3’
ARN : 5’ GCU GCU GCU GCU GCU GCU GCU GCU 3’
Protéine : Ala Ala Ala Ala Ala Ala Ala Asp 3’
GC A
ARN : 5’ GCU GCU GCU GCG CUG CUG CUG CUG 3’
ARN : 5’ GCU GCU GCU GCU CUG CUG CUG AU 3’
ARN : 5’ GCU GCU GCU GCU GCU GAC UGC UGA 3’
Protéine : Ala Ala Ala Ala Leu Leu Leu Leu 3’
Protéine : Ala Ala Ala Ala Leu Leu Leu 3’
Protéine : Ala Ala Ala Ala Ala Asp Cys STOP 3’
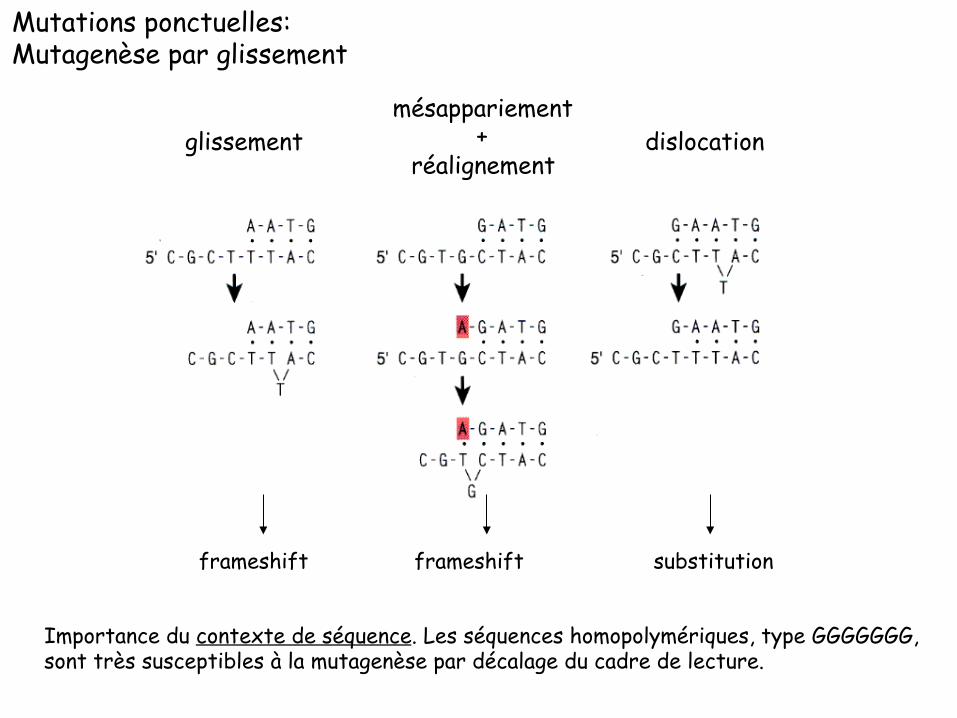
Mutations ponctuelles: Mutagenèse par glissement
glissement mésappariement
+ réalignement
dislocation
frameshift frameshift substitution
Importance du contexte de séquence. Les séquences homopolymériques, type GGGGGGG, sont très susceptibles à la mutagenèse par décalage du cadre de lecture.
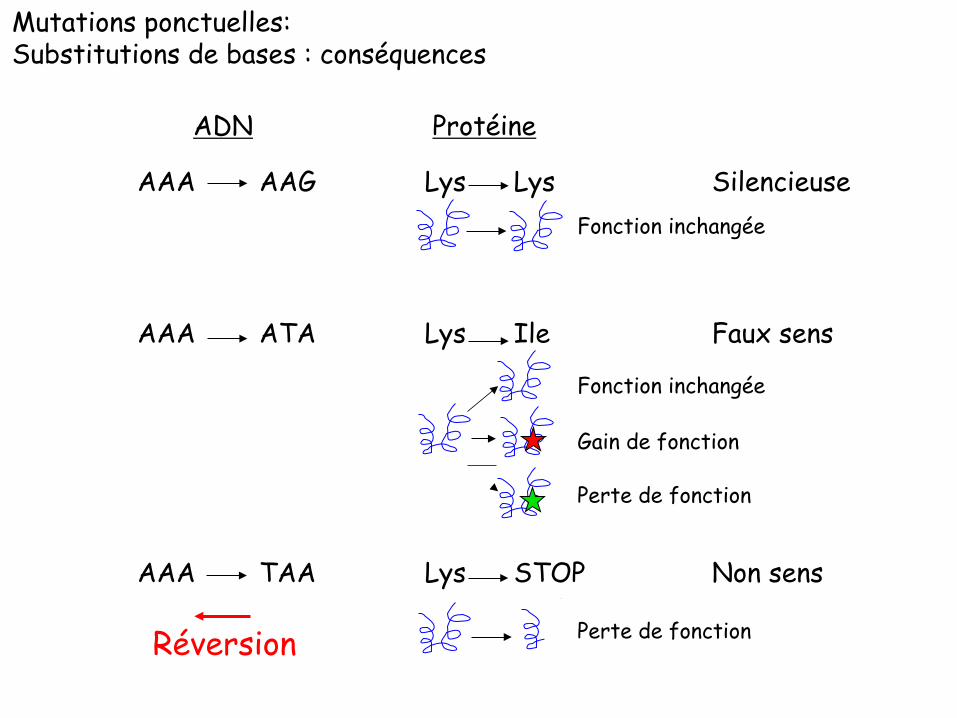
Mutations ponctuelles: Substitutions de bases : conséquences
AAA AAG Lys Lys Silencieuse
AAA ATA Lys Ile Faux sens
AAA TAA Lys STOP Non sens
Gain de fonction
Perte de fonction
Perte de fonction
Fonction inchangée
Fonction inchangée
ADN Protéine
Réversion
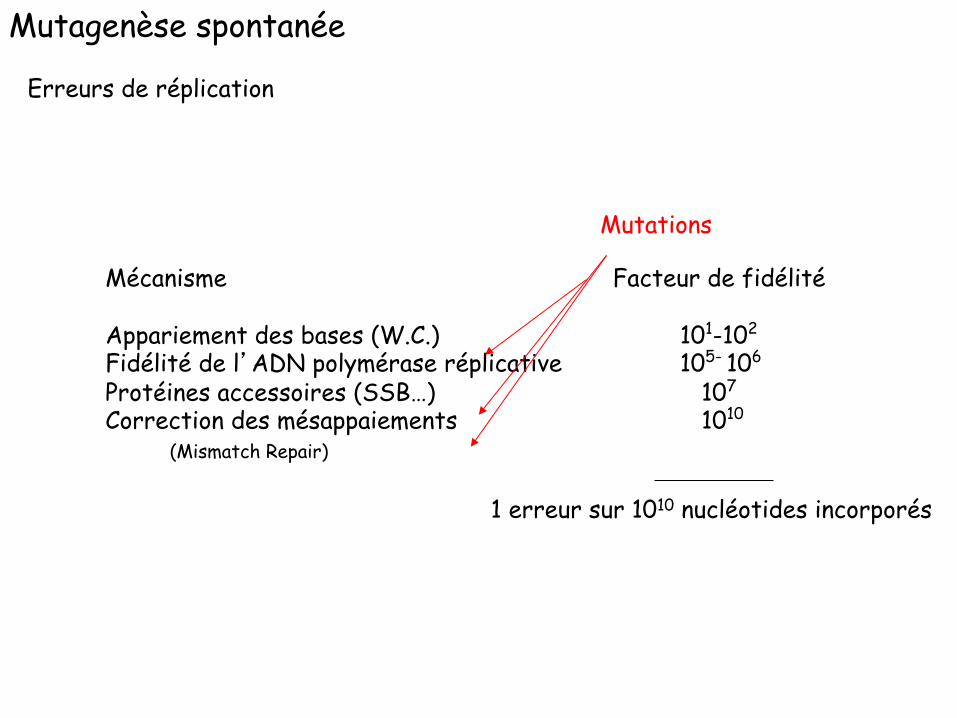
Mutagenèse spontanée
Mécanisme Facteur de fidélité Appariement des bases (W.C.) 101-102 Fidélité de l’ADN polymérase réplicative 105- 106
Protéines accessoires (SSB…) 107
Correction des mésappaiements 1010
(Mismatch Repair)
Erreurs de réplication
1 erreur sur 1010 nucléotides incorporés
Mutations

Mutagenèse spontanée
Mutations générées par des répétitions directes
5’ …AGTCGTCAGTACGCCGTACCTGAATTTT…TTTTTTAGTACGCCGTAC… 3’
Formation de structures secondaires (Hairpin)
Maladies à triplets: découlent de l’instabilité de séquences répétées (type microsatellite)
X fragile (CGG)n 6< n <53 : normal 60< n <230: malade
5’ AGTCGTCAGTA TAC… 3’
TG
G
C
CCC
AAC
C
G
GC
T
G
..........
......
Délétions, Extensions

Mutagenèse induite (par des lésions)
anneau de processivité
Les lésions peuvent provoquer un blocage de la fourche de
réplication
Mort cellulaire
Levée du blocage =
Tolérance

Les mécanismes de tolérance des lésions Régression de
fourche
« Réparation » par recombinaison
Evitement de la lésion
Fidèle
Mutagène
Synthèse translésionnelle
(TLS)
M

Méthodes d’analyses
In vivo : spectres de mutagenèse
potentiel mutagène et génotoxique, génétique de la mutagenèse
utilisation d’ADN mono-modifiés
In vitro : extension d’amorces (analyse fine des mécanismes moléculaires de la mutagenèse)
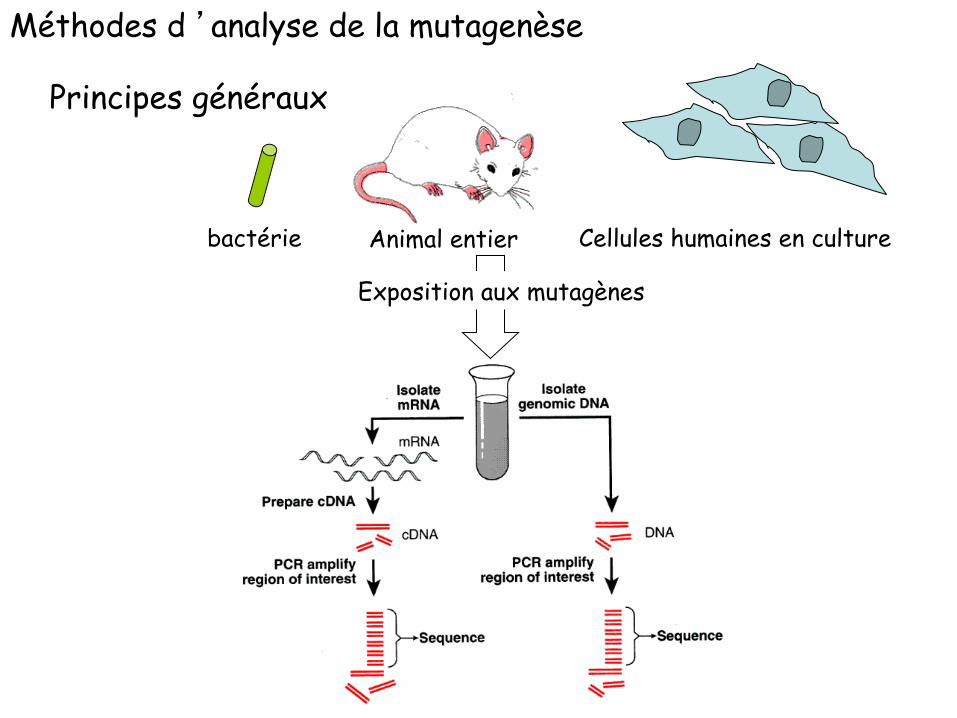
Animal entier bactérie Cellules humaines en culture
Méthodes d ’analyse de la mutagenèse
Principes généraux
Exposition aux mutagènes

Méthodes d ’analyse de la mutagenèse
Principes généraux
ADN cible : l’apparition d’une mutation permet la sélection du mutantExemples : lacI (seuls les mutants poussent en présence d’un analogue du galactose)
lacZ (sélection phénotypique sur milieu indicateur)rif (seuls les mutants poussent en présence de l’antibiotique Rifampicine)HPRT (seuls les mutants poussent en présence de l’analogue 6-thio-guanine)
Modification : Exposition in vitro ou in vivo à l’agent mutagène; notion de doses; activation métabolique potentiellement requise
Réplication : in vitro ou in vivo; choix de l'hôte (fond génétique)
Analyse: détermination du spectre de mutagenèse, du pouvoir mutagène et potentiellement toxique de l’agent testé, étude génétique
Mutagenèse en avant : on ne sélectionne pas, à priori, de type particulier de mutations
Mutagenèse par réversion : seules les mutations d’un type donné sont observées
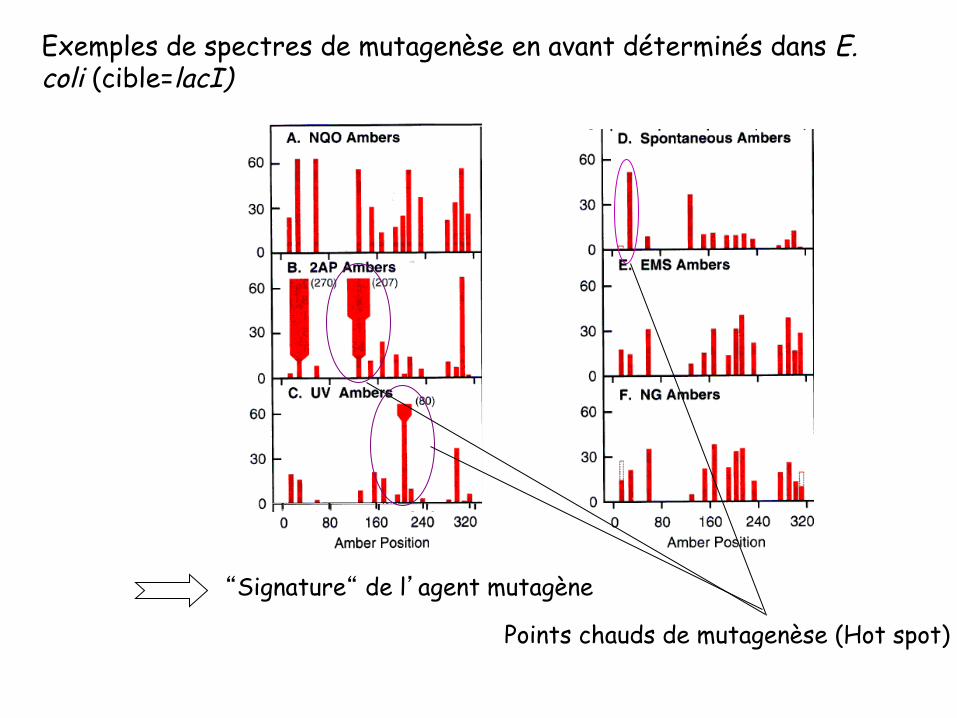
Points chauds de mutagenèse (Hot spot)
Exemples de spectres de mutagenèse en avant déterminés dans E. coli (cible=lacI)
“Signature“ de l’agent mutagène

Exemples de cibles pour l’étude de la mutagenèse par tests de réversion (Claire Cupples and Jeffrey Miller)

Maladies associées à des gènes de réparation
Syndrome Type de voie de instabilité Types de réparation affectée génétique cancers associés
Xeroderma NER (± TCR) Mutations Cancer de Pigmentosum « classique » ponctuelles la peau (UV)
Ataxia Réparation de Aberrations telangiectasia (AT) cassure double-brin chromosomiques Lymphomes Syndrome de Nijmegen Recombinaison Aberrations Lymphomes
homologue (?) chromosomiques BRCA1/BRCA2 RH Aberrations Cancer du
chromosomiques sein et ovaire Syndrome de Werner RH / TLS Aberrations variés
chromosomiques Syndrome de Bloom Réparation de Aberrations Leucémies
cassure double-brin chromosomiques Lymphomes HNPCC Réparation des Mutations Cancer
mesappariements ponctuelles colorectal
Xeroderma Synthèse Mutations Cancer de Pigmentosum variant translésionnelle (TLS) ponctuelles la peau (UV)
Top Related