







Langages
Pages
Légal


Stochastic models for gene expression
Ovidiu Radulescu, DIMNP UMR 5235, Univ. of Montpellier 2
Colloque franco-roumain mathématiques appliquées, Poitiers 29/08/10

Summary
Motivation : fluctuations in gene networks
Stochastic models for gene expression
Application to biology: fluctuome

1. Fluctuations in gene networks
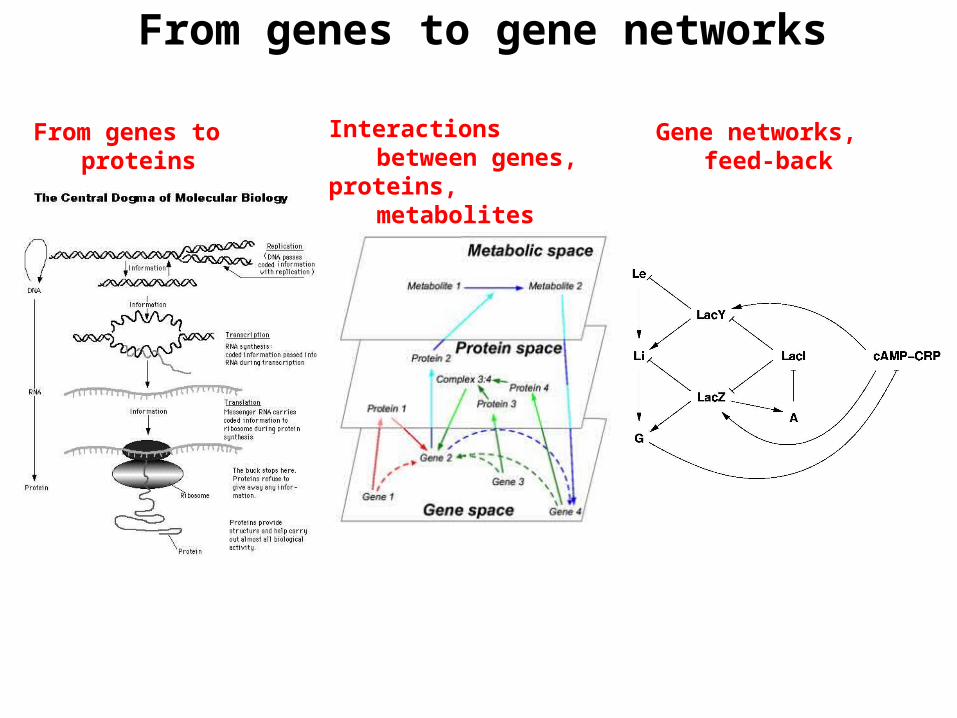
From genes to gene networks
From genes to proteins Interactions between genes,proteins, metabolites
Gene networks, feed-back
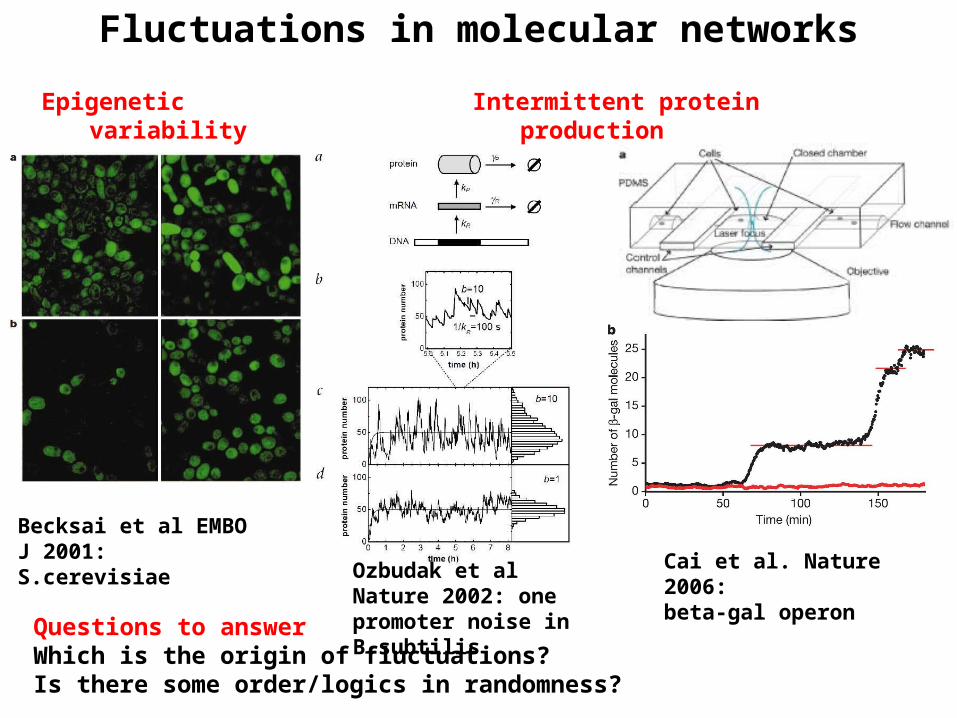
Fluctuations in molecular networks
Epigenetic variability
Which is the origin of fluctuations?Is there some order/logics in randomness?
Questions to answer
Becksai et al EMBO J 2001: S.cerevisiae
Ozbudak et al Nature 2002: one promoter noise in B.subtilis
Cai et al. Nature 2006:beta-gal operon
Intermittent protein production
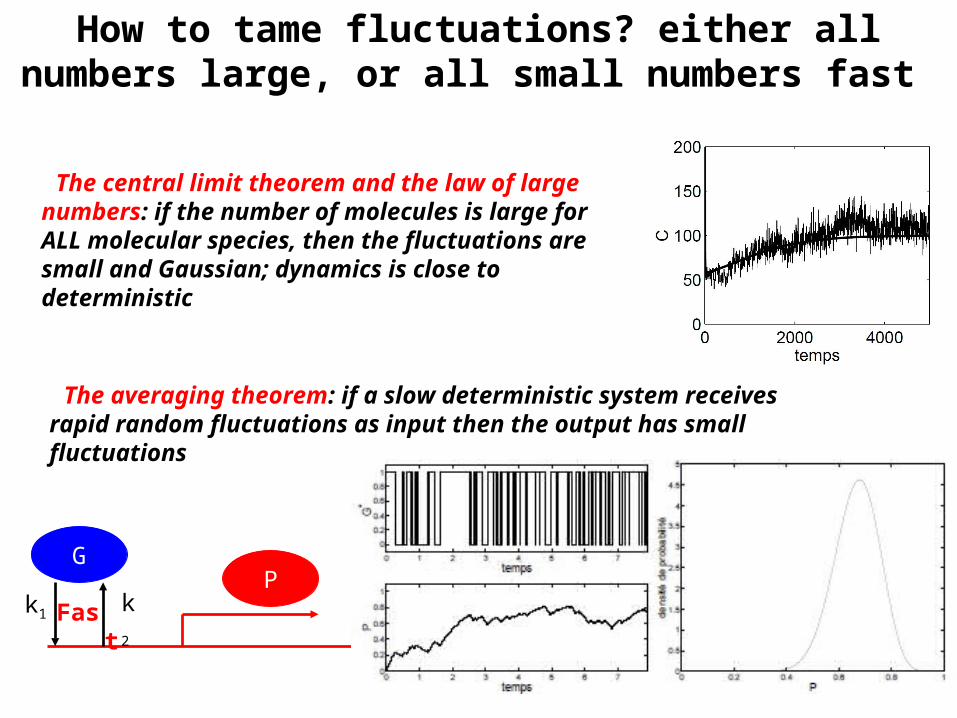
How to tame fluctuations? either all numbers large, or all small numbers fast
The central limit theorem and the law of large numbers: if the number of molecules is large for ALL molecular species, then the fluctuations are small and Gaussian; dynamics is close to deterministic
The averaging theorem: if a slow deterministic system receives rapid random fluctuations as input then the output has small fluctuations
PG
k1 k2Fast

Noise in multiscale networks with inverted time hierarchy
Hybrid noise : discrete variations of low numbers species, continuous variation punctuated by jumps or switching of large numbers species
Low and large numbers : a broad distribution of abundances from a few to 104 per cell
Fast and slow processes : from 10-3 to 104 s
Multiscale
Inverted time hierarchy: some processes involving low numbers are slow
PG
k1 k2Slow

2. Stochastic models for gene expression

Delbrück-Rényi-Bartolomay approachTwo main assumptions:
Reactions are independent; Transport is instantaneous.
Dynamics variables are numbers of molecules of different species All species evolve by discrete jumps separated by random waiting times
Which reaction?
Which time?Generate exponential variable of
parameter (k1A+k2B)
Gillespie’s algorithm
Disadvantages:
Time costly Analytical solutions of theChemical Master Equation are rarelyavailable

Continuous time Markov jump processes
nr
iXiXi
iiXqXqX
1
(.)])((.))([,.)(
are n chemical species
jump vectorrninZiii ,1,
nininini AAAA ...... 1111
nAA ,...,1
nr
iii XVXVX
1
)]()([)(
nr
jjjii XVXVXVXq
1
)]()([)/()(
intensity
distribution of jumps
biochemical reaction
jump probability
nZX is the state
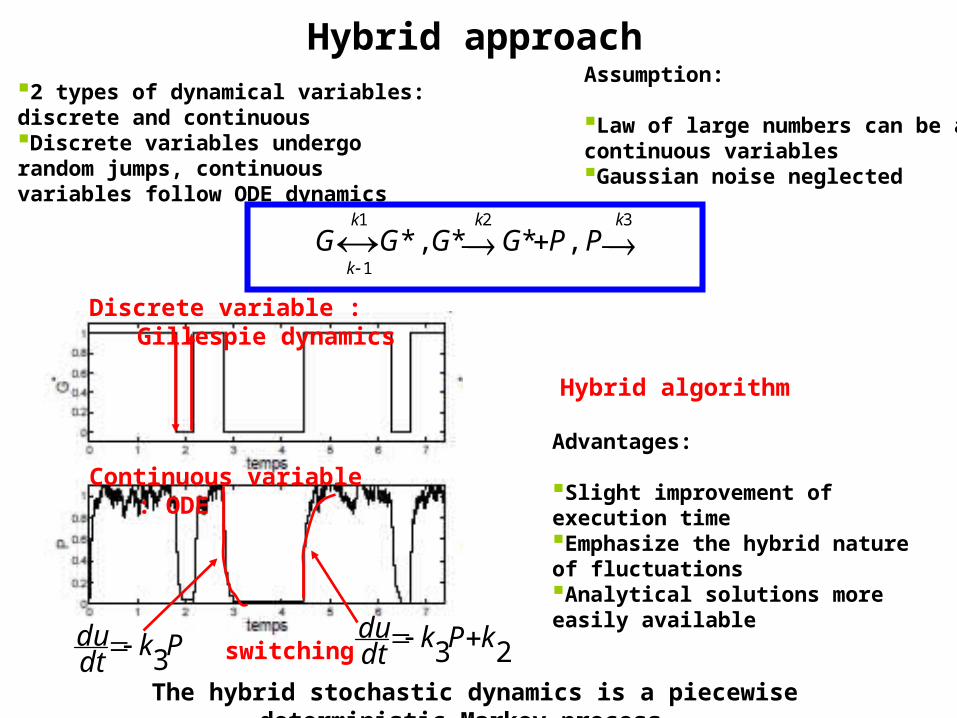
Hybrid approach
The hybrid stochastic dynamics is a piecewise deterministic Markov process
Assumption:
Law of large numbers can be applied to continuous variablesGaussian noise neglected
2 types of dynamical variables: discrete and continuousDiscrete variables undergo random jumps, continuous variables follow ODE dynamics
Hybrid algorithm
Advantages:
Slight improvement of execution timeEmphasize the hybrid nature of fluctuations Analytical solutions more easily available
Continuous variable : ODE
Discrete variable : Gillespie dynamics
23 kPkdtdu Pk
dtdu
3
321
1
,***,kkk
k
PPGGGG
switching

Partial fluid approximation:partitionGiven a pure jump model, find its hybrid approximation
Species partition: discrete, continuous),( CD XXCDCD RRR Reaction partition
Discrete transitions
Contributions to continuous flow
coupling intrinsic
321
1
,***,kkk
k
PPGGGG
Coupling between discrete and continuous only if super-reactions of type 1: fast, change mode XC, rates depend on XD
switching

Partial fluid approximation : expansion)/,(
~),( VXxXXXX CCDCD Rescaling
1st order Taylor expansion in
) , , ( ) , ( ) ,~
( )~
( ) ,~
(t x X p x X V t X p X t Xtpc D
R i
c D iDi
D
Di
) , / , ( ) / , (t V x X p V x X VCi
DC
Cic D
R i
c D i
) , / , ( ) / (t V x X p V x VCi
C
Cic D
R i
c i
VCi/
) , , ( ) , (t x X p x Xfc D c D xc
V x V V x X V x Xfdtdx
C
Ci
DC
Ci
R i
c i
R i
c D i c Dc
)/ ( )/ , ( ) , (
Chemical Master Equation
Hybrid Master/Fokker-Planck equation
Switching
Drift
of a PDP with switching
Coupling if super-reactions of type 1
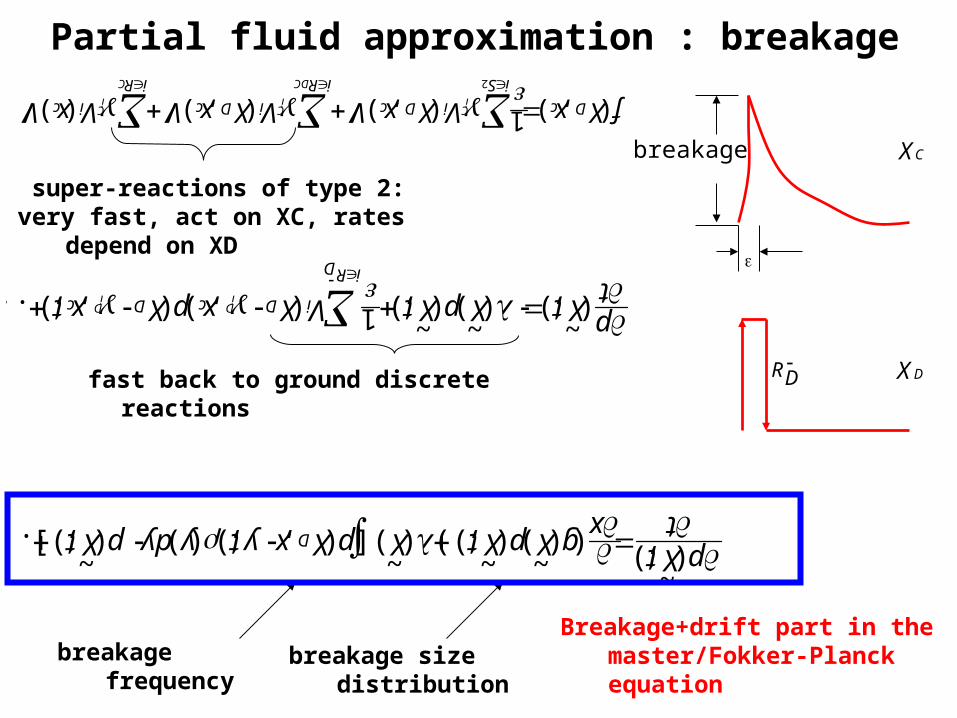
Partial fluid approximation : breakage
Breakage+drift part in the master/Fokker-Planck equation
V x V V x X V V x X V x XfC
Ci
DC
Ci
Ci
R i
c i
R i
c D i
S i
c D i c D)/ ( )/ , ( )/ , ( 1 ) , (2
super-reactions of type 2:very fast, act on XC, rates depend on XD
breakage
... ) , , ( ) , ( 1 ) ,~
( )~
( ) ,~
(
t x X p x X V t X p X t Xtpc D
DR i
c D iDi
Di
fast back to ground discrete reactions DR
DX
CX... )] ,
~( ) ( ) , , ( )[
~( )) ,
~( )
~( ( ) ,
~(
t X p dy y t y x X p X t X p X gx t
t X pD
breakage frequency breakage size distribution
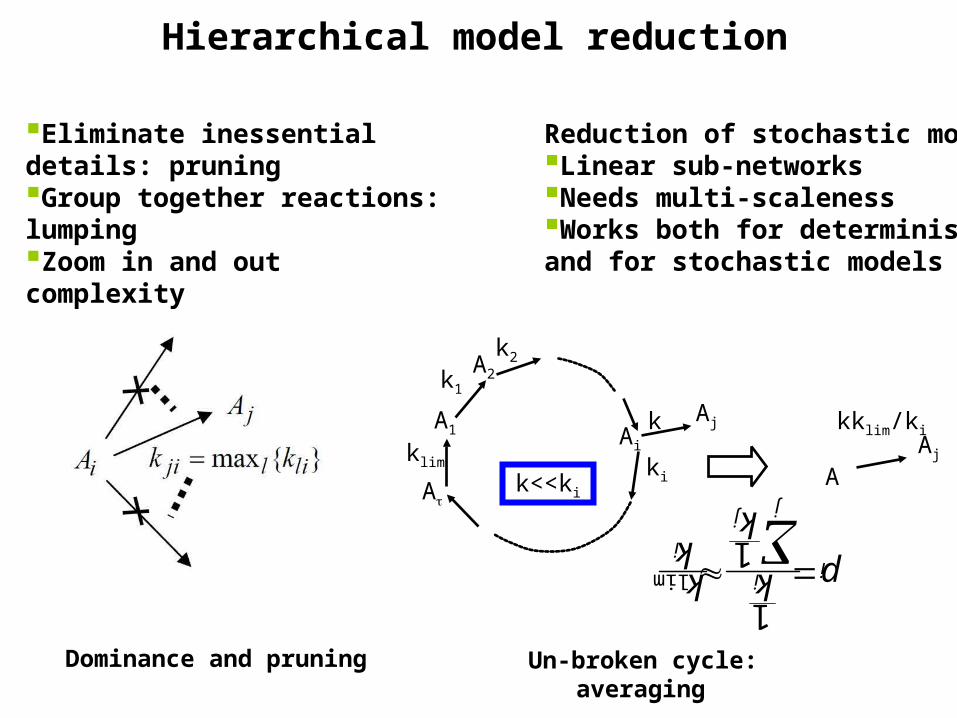
Hierarchical model reduction
Reduction of stochastic models:Linear sub-networksNeeds multi-scalenessWorks both for deterministicand for stochastic models
Eliminate inessential details: pruningGroup together reactions: lumpingZoom in and out complexityDrastic decrease of simulation time
Dominance and pruning Un-broken cycle: averaging
A1
A2
Ai Aj
A
k2
k1
klim ki
k
k<<ki
Aj
A
kklim/ki
i
jj
ii
kk
k
k plim
1
1
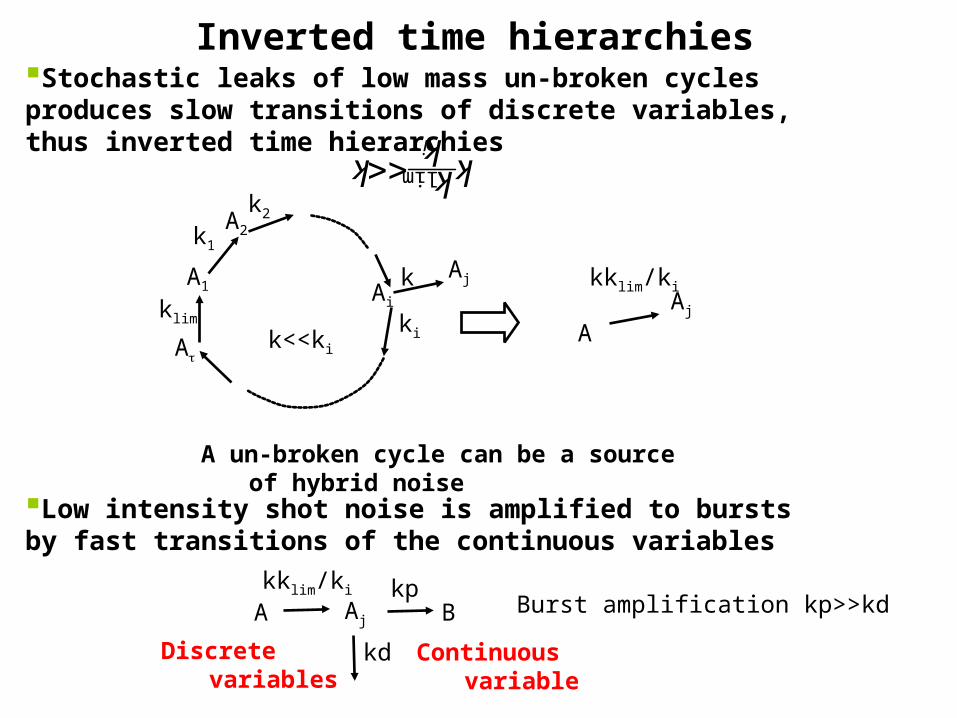
Inverted time hierarchiesStochastic leaks of low mass un-broken cycles produces slow transitions of discrete variables, thus inverted time hierarchies
A un-broken cycle can be a source of hybrid noise
A1
A2
Ai Aj
A
k2
k1
klim ki
k
k<<ki
Aj
A
kklim/ki
kkk ki
lim
Low intensity shot noise is amplified to bursts by fast transitions of the continuous variables
AjA Bkklim/ki
kd
kpBurst amplification kp>>kd
Continuous variableDiscrete variables
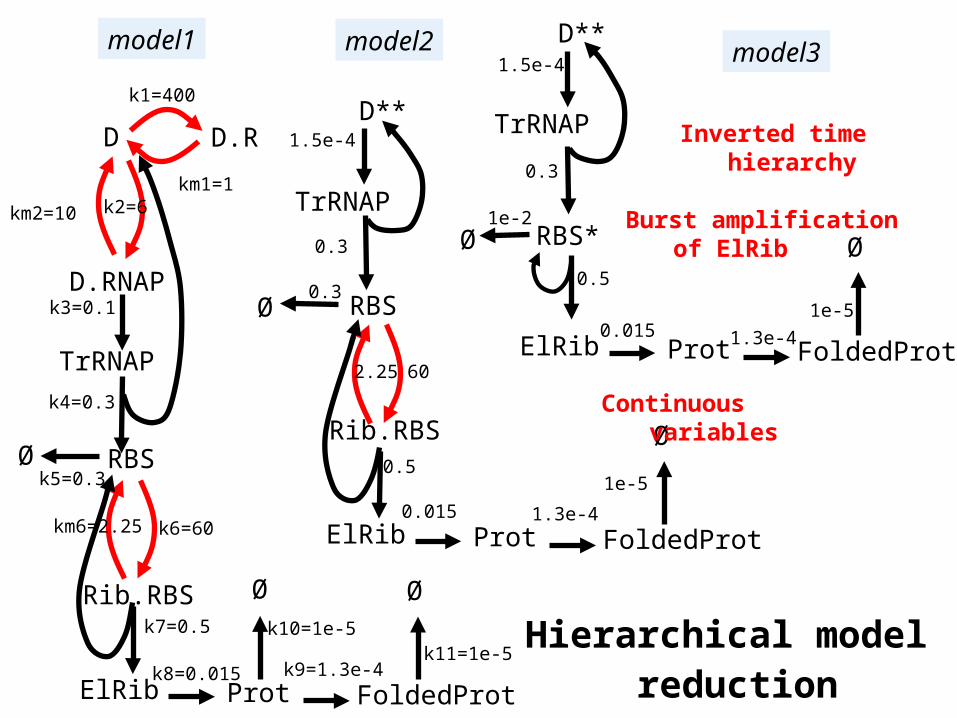
Prot FoldedProt
D D.R
D.RNAP
TrRNAP
RBS
Rib.RBS
ElRib
Ø
k1=400
km1=1
km2=10 k2=6
k3=0.1
k4=0.3
k6=60km6=2.25
k7=0.5
k8=0.015
k10=1e-5k11=1e-5
k9=1.3e-4
Ø
Øk5=0.3
model1
Prot FoldedProt
D**
TrRNAP
RBS
Rib.RBS
ElRib
0.3
602.25
0.5
0.015
1e-5
1.3e-4
Ø
1.5e-4
Prot FoldedProt
RBS*
ElRib
D**
TrRNAP
0.5
0.0151e-5
1.3e-4
Ø
model2 model31.5e-4
0.3
Ø
Ø
0.3
1e-2
Hierarchical model reduction
Inverted time hierarchy
Continuous variables
Burst amplification of ElRib
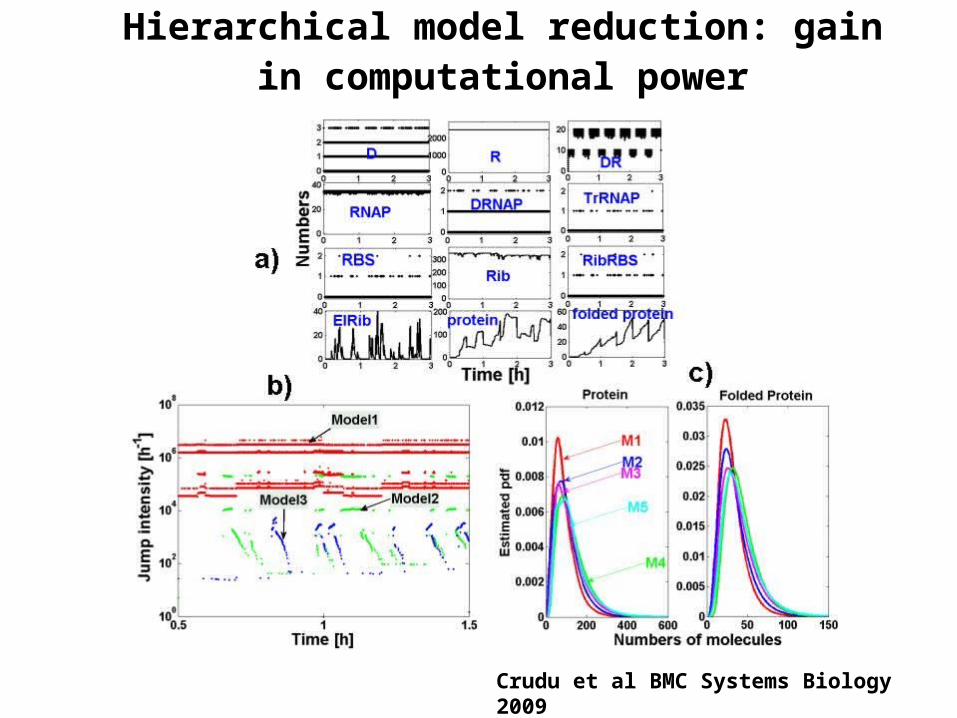
Hierarchical model reduction: gain in computational power
Crudu et al BMC Systems Biology 2009
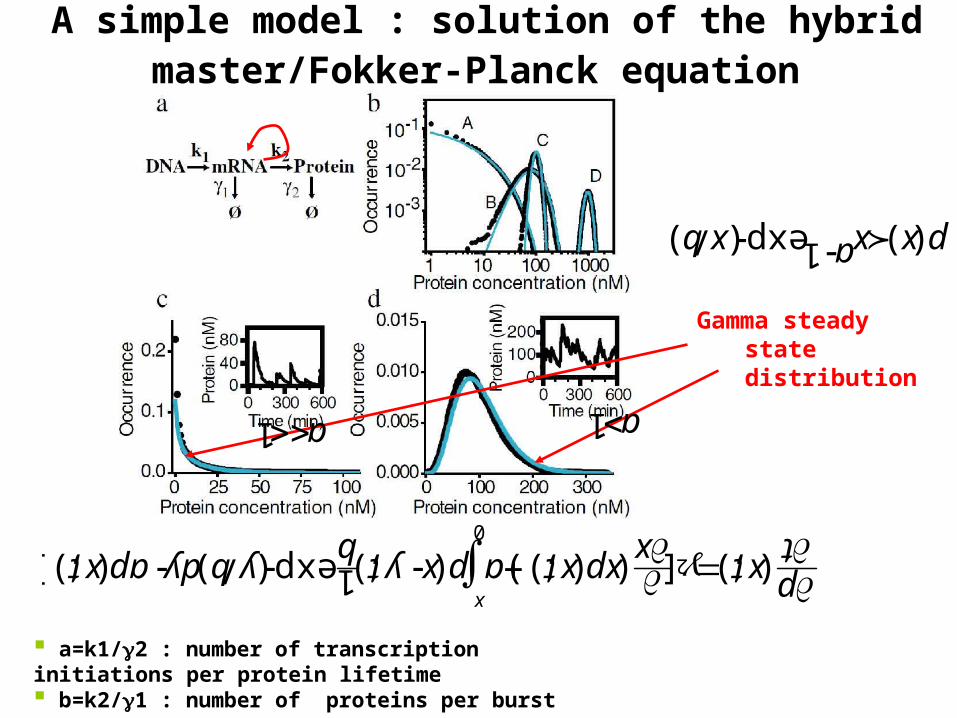
A simple model : solution of the hybrid master/Fokker-Planck equation
a=k1/2 : number of transcription initiations per protein lifetime b=k2/1 : number of proteins per burst
)] , ( ) / exp( 1) , ( )) , ( ( [ ) , (0
2t x ap dy b ybt y x p a t x xp
xt x
tp
x
) / exp( 1 ) (b x ax x p
Gamma steady state distribution
1 a 1a

3. Application to biology: fluctuome
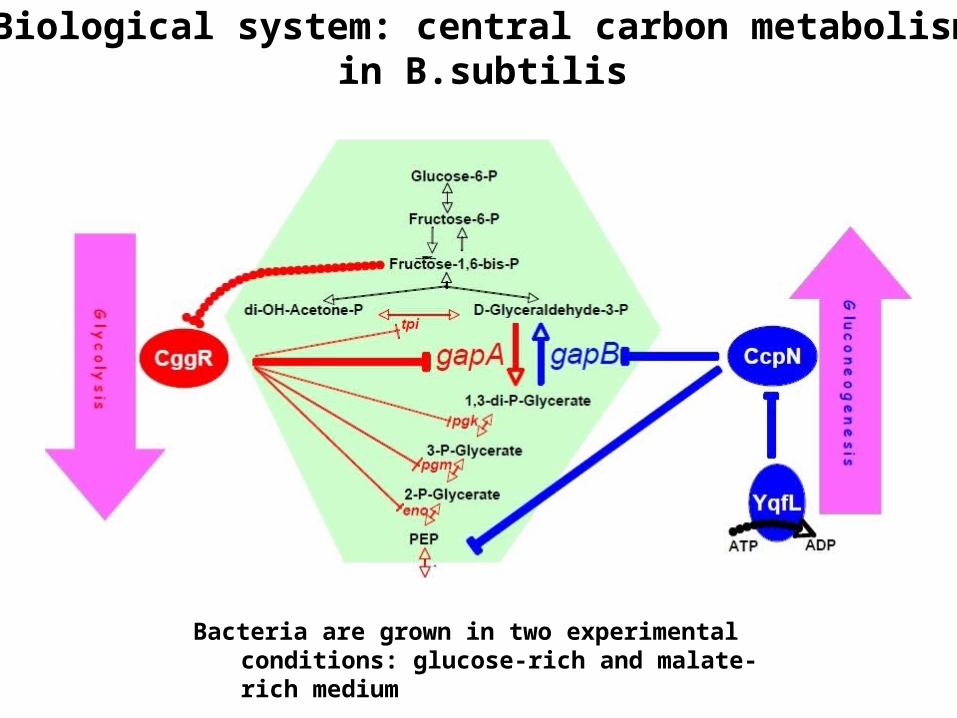
Biological system: central carbon metabolism in B.subtilis
Bacteria are grown in two experimental conditions: glucose-rich and malate-rich medium
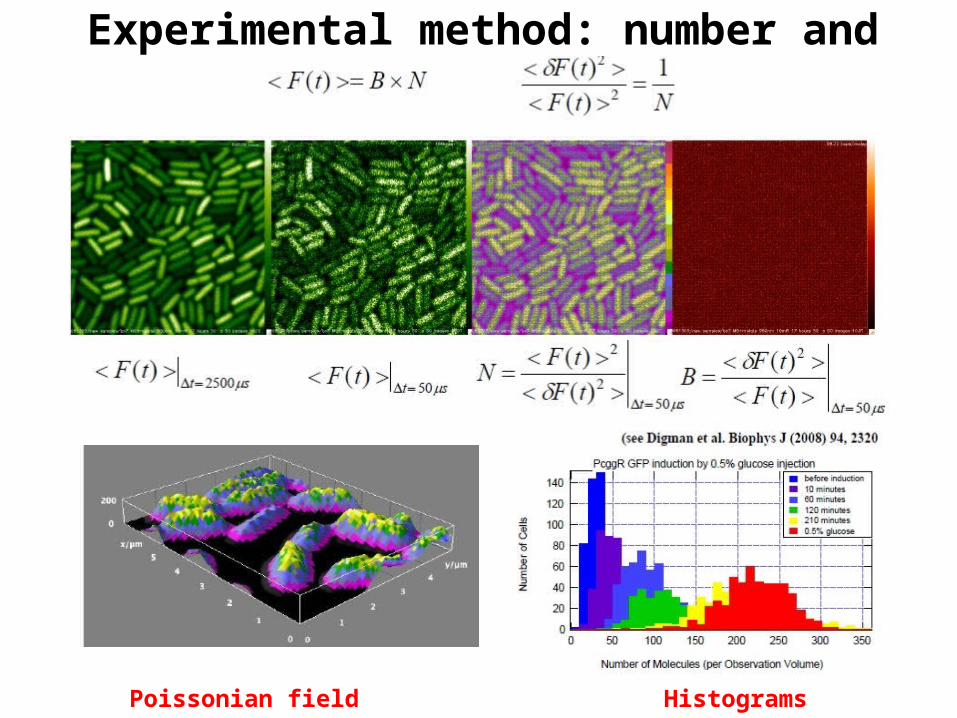
Experimental method: number and brightness
Poissonian field F map Histograms
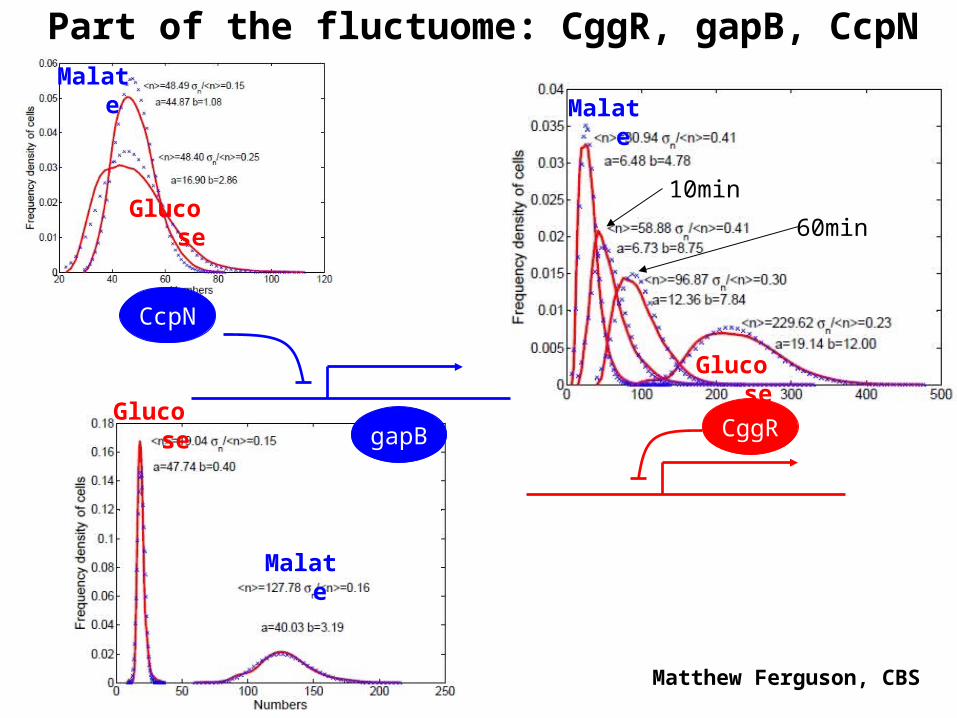
Part of the fluctuome: CggR, gapB, CcpN
CggRgapB
CcpN
Glucose
Malate
Glucose
Malate
Glucose
Malate
Matthew Ferguson, CBS
10min
60min
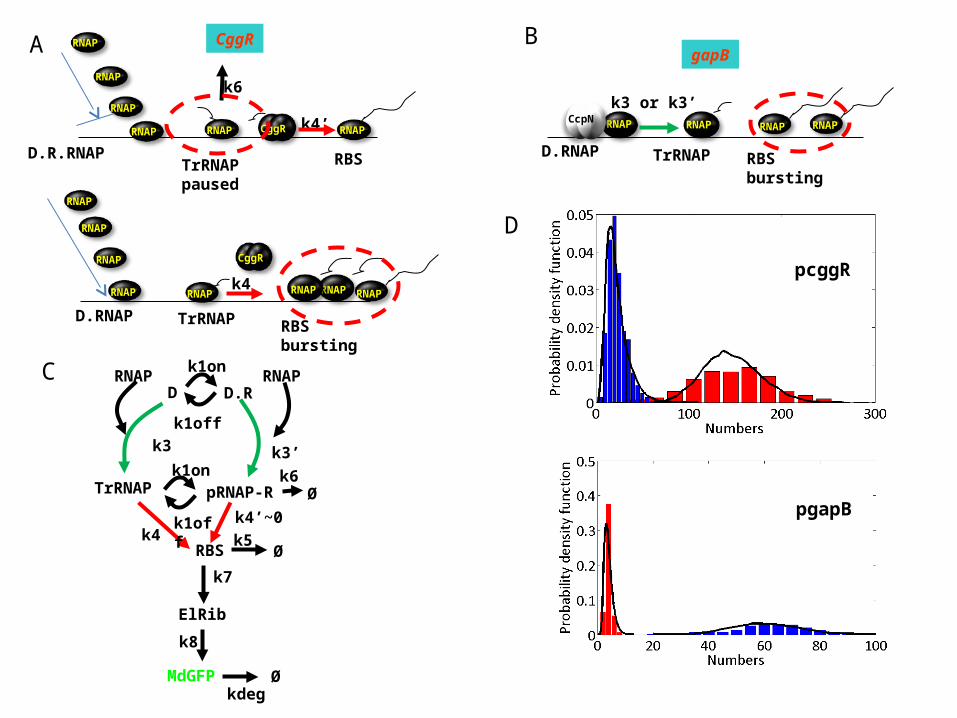
MdGFP
D D.R
RBS
ElRib
k3
k7
k8
kdegØ
Øk5
k3’
TrRNAP
k4
k1on
k1off
k4’~0Ø
k6
RNAP
pRNAP-R
RNAP
k1off
k1on
CggR
CggR
TrRNAPpaused
D.R.RNAP
k6
RBS
k4’RNAP RNAP RNAP
RNAP
RNAP
RNAP
TrRNAPD.RNAP RBSbursting
RNAP RNAP
RNAPRNAP
CcpNk3 or k3’
gapBA
C
TrRNAPD.RNAPRBSbursting
CggR
k4
RNAP
RNAP
RNAP
RNAP RNAP
RNAPRNAP
B
D
pcggR
pgapB
RNAP

Take home message
The origin of noise in multiscale molecular network is the inverted time hierarchy; noise is hybrid
Hierarchical model reduction unravels functional structure: unbroken cycles, burst amplifiers, integrators
Noise carries information about interactions : fluctuome could be an important tool

Acknowledgements
Nathalie Declerck, Matthew Ferguson, Catherine RoyerCBS Montpellier
Alina Crudu, Arnaud Debussche, Aurelie MullerIRMAR Rennes, ENS Cachan
Alexander GorbanUniversity of Leicester
Top Related