Recherches biochimiques sur l'oogenèse
8
BIOCHIMIE, 1972, 54, 1189-1196. Recherches biochimiqucs sur l'oogenbse. V - Comparaison entre le RNA 5 S somatique et le RNA 5 S des oocytes de Xenopus laevis (*). Herman DENIS, Mauriee W~GNF.Z, et Robert WILLEM. Laboratoire de Biochimie, Unioersit~ de Liege, place Delcour, 17, B-#O00 Libge, Belgique. (8/6/1972). Summary. -- 5 S RNA from oocytes of Xenopus laeois differs from somatic 5 S RNA by the degree of phosphorylation of its 5'-end. There is in somatic 5 S RNA a higher proportion of molecules carrying 2 or 3 phosphate groups on the 5'-terminal nucleotide. Somatic 5 S RNA was retarded by columns of Sephadex G-100 slightly less than oocyte 5 S RNA. 5 S RNA from somatic ribosomes was eluted from columns of methylated albumin-Kieselguhr far behind 5 S RNA from whole oocytes and from oocyte ribosomes. Ribosomes from early embryos were found to contain 2 types of 5 S RNA : [1] newly made 5 S RNA which is of somatic type and [2] bulk 5 S RNA which is of the ooeyte type. Somatic 5 S RNA migrated in 7.4 p. cent polyacrylamide gels slightly ahead of oocyte 5 S RNA. This difference of electrophoretic mobility was suppressed neither by removal of the terminal phosphates by alkaline phosphatase nor by heat or urea denaturation. Electrophoresis in 12.3 p. cent gels revealed the existence of 2 distinct forms in somatic 5 S RNA. One of these had the same mobility as oocyte 5 S RNA. The other one migrated faster. Somatic and oocyte 5 S RNA's reacted differently to heat denaturation: after cooling both native forms of somatic 5 S RNA were replaced by a third one, which migrated faster, whereas most of the oocyte RNA remained in native form. When heated in saline, oocyte 5 S RNA shorted a slightly higher hyperchromieity than somatic 5 S BNA. These results suggest that both the primary and the secondary structures are different in somatic and in oocyte 5 S RNA's. INTRODUC~ON. PrOs de la moiti~ du RNA pr6sent dans les oocytes du x6nope en petit aeeroissement (dia- mbtre : 50 h 150 ~m) est form6e par du RNA 5 S [1, 2, 3]. Dans ee travail, nous eomparons les pro- pri6t6s du RNA 5 S des ooeytes avee eelles du RNA 5 S somatique. Nous 6tudions le eomporte- ment de ees deux types de RNA sur eolonnes d'hy- droxyapatite, de Sephadex G-100 et d'albumine m6thyl6e, leur mobilit6 61ectrophor6tique en gel de polyaerylamide, ainsi que leur profil d'hyperchro- mieit6. Abrdoiations : tRNA : RNA de transfert. Tris : tris(hydroxym~thyl)-aminom~thane. EDTA : acide ~thyl6nedinitrilot~traae~tique, sel di- sodique. SSC : NaC1 150 raM, citrate de sodium 15 raM. SDS : doddeylsulfate de sodium. (*) Pour les premiers articles de eette s~rie, voir les r~f6renees [I, 2 et 6]. M,ETHODES. 1) PURIFICATION DU RNA 5 S. Le RNA 5 S somatique est puriti6 soit h partir de eellules r6nales eultiv6es in vitro en pr6senee d'uridine [3HJ et de guanosine [3Hi [4], soit h partir de ribosomes de foie, soit encore h partir d'embryons entiers marqu6s au 14CO 2 ou au azp [I]. Le RNA est extrait par le proe6d6 au SDS- ph6nol froid [5], puis pass6 sur colonne de Sepha- dex G-100, eomme d6erit pr6e~demment [13. Le RNA 5 S est 61u6 juste apr6s le RNA de poids mo- 16eulaire 61ev6. Le RNA 5 S d'ooeytes est extrait h partir d'ovaires entiers de femelles immatures ou de ribosomes de femelles mflres. Comme nous l'avons montr6 pr6e6demment, la quasi-totalit6 du RNA d'ovaires entiers provient des ooeytes, et non des eellules follieuleuses qui les entourent [13. La ma- jeure partie du RNA 5 S des ovaires immatures
Transcript of Recherches biochimiques sur l'oogenèse

BIOCHIMIE, 1972, 54, 1189-1196.
Recherches biochimiqucs sur l'oogenbse. V - Comparaison entre le R N A 5 S somat ique et le R N A 5 S des oocytes
de Xenopus laevis (*).
H e r m a n DENIS, M a u r i e e W~GNF.Z, et R o b e r t WILLEM.
Laboratoire de Biochimie, Unioersit~ de Liege, place Delcour, 17, B-#O00 Libge, Belgique.
(8/6/1972).
Summary. - - 5 S RNA f rom oocytes of Xenopus laeois differs f rom somat ic 5 S RNA by the degree of phosphory la t ion of i ts 5'-end. There is in somat ic 5 S RNA a h igher p ropor t ion of molecules car ry ing 2 or 3 phospha te groups on the 5 ' - t e rmina l nucleotide.
Somatic 5 S RNA was re tarded by columns of Sephadex G-100 s l ight ly less t h a n oocyte 5 S RNA. 5 S RNA f rom somatic r ibosomes was eluted f rom columns of me thy la t ed a lbumin-Kiese lguhr fa r beh ind 5 S RNA f rom whole oocytes and f rom oocyte r ibosomes. Ribosomes f rom ear ly embryos were found to con ta in 2 types of 5 S RNA : [1] newly made 5 S RNA which is of somat ic type and [2] bu lk 5 S RNA which is of the ooeyte type.
Somatic 5 S RNA migra ted in 7.4 p. cent po lyacry lamide gels s l ight ly ahead of oocyte 5 S RNA. This difference of e lectrophoret ic mobi l i ty was suppressed ne i the r by removal of the t e r m i n a l phospha tes by a lka l ine phospha tase no r by hea t or u rea dena tu ra t ion . Electrophoresis in 12.3 p. cent gels revealed the existence of 2 d is t inc t fo rms in somatic 5 S RNA. One of these had the same mobi l i ty as oocyte 5 S RNA. The o ther one migrated faster . Somatic and oocyte 5 S RNA's reacted different ly to hea t d e n a t u r a t i o n : a f te r cooling bo th nat ive forms of somat ic 5 S RNA were replaced by a th i rd one, which migra ted fas ter , whereas most of the oocyte RNA remained in na t ive form.
W h e n hea ted in saline, oocyte 5 S RNA shorted a s l ight ly h igher hyperchromie i ty t h a n somatic 5 S BNA.
These resul ts suggest t ha t bo th the p r ima ry and the secondary s t ruc tures are different in somatic and in oocyte 5 S RNA's.
I N T R O D U C ~ O N .
PrOs de la m o i t i ~ d u RNA p r 6 s e n t d a n s les o o c y t e s d u x 6 n o p e e n p e t i t a e e r o i s s e m e n t (d ia - m b t r e : 50 h 150 ~m) es t f o r m 6 e p a r d u R N A 5 S [1, 2, 3]. D a n s ee t r a v a i l , n o u s e o m p a r o n s les p r o - p r i 6 t 6 s d u RNA 5 S des o o e y t e s a v e e ee l l es d u RNA 5 S s o m a t i q u e . Nous 6 t u d i o n s le e o m p o r t e - m e n t de ees d e u x t y p e s de R N A s u r e o l o n n e s d ' h y - d r o x y a p a t i t e , de S e p h a d e x G-100 et d ' a l b u m i n e m 6 t h y l 6 e , l e u r m o b i l i t 6 6 1 e c t r o p h o r 6 t i q u e e n gel de p o l y a e r y l a m i d e , a i n s i que l e u r p r o f i l d ' h y p e r c h r o - m ie i t 6 .
Abrdoiations : tRNA : RNA de t rans fe r t . Tris : t r i s (hydroxym~thy l ) - aminom~thane . EDTA : acide ~thyl6nedini t r i lo t~traae~t ique, sel di-
sodique. SSC : NaC1 150 raM, c i t ra te de sodium 15 raM. SDS : doddeylsulfate de sodium. (*) Pou r les p remiers art icles de eette s~rie, voir
les r~f6renees [I, 2 et 6].
M,ETHODES.
1) PURIFICATION DU R N A 5 S.
Le RNA 5 S s o m a t i q u e es t p u r i t i 6 so i t h p a r t i r de ee l l u l e s r 6 n a l e s e u l t i v 6 e s in vitro e n p r 6 s e n e e d ' u r i d i n e [3HJ et de g u a n o s i n e [3Hi [4], so i t h p a r t i r de r i b o s o m e s de fo ie , so i t e n c o r e h p a r t i r d ' e m b r y o n s e n t i e r s m a r q u 6 s au 14CO 2 o u au azp [ I ] . Le RNA est e x t r a i t p a r le p r o e 6 d 6 au SDS- p h 6 n o l f r o i d [5] , p u i s p a s s 6 s u r c o l o n n e de S e p h a - dex G-100, e o m m e d 6 e r i t p r 6 e ~ d e m m e n t [13. Le RNA 5 S es t 61u6 jus t e a p r 6 s le RNA de p o i d s mo- 16eu la i re 61ev6.
Le RNA 5 S d ' o o e y t e s es t e x t r a i t h p a r t i r d ' o v a i r e s e n t i e r s de f e m e l l e s i m m a t u r e s o u de r i b o s o m e s de f eme l l e s mfl res . C o m m e n o u s l ' a v o n s m o n t r 6 p r 6 e 6 d e m m e n t , la q u a s i - t o t a l i t 6 d u R N A d ' o v a i r e s e n t i e r s p r o v i e n t de s o o e y t e s , e t n o n de s ee l lu l e s f o l l i e u l e u s e s qu i les e n t o u r e n t [13. L a m a - j e u r e p a r t i e d u RNA 5 S des o v a i r e s i m m a t u r e s
1 1 9 0 H. Denis , M. W e g n e z el R. W i l l e m .
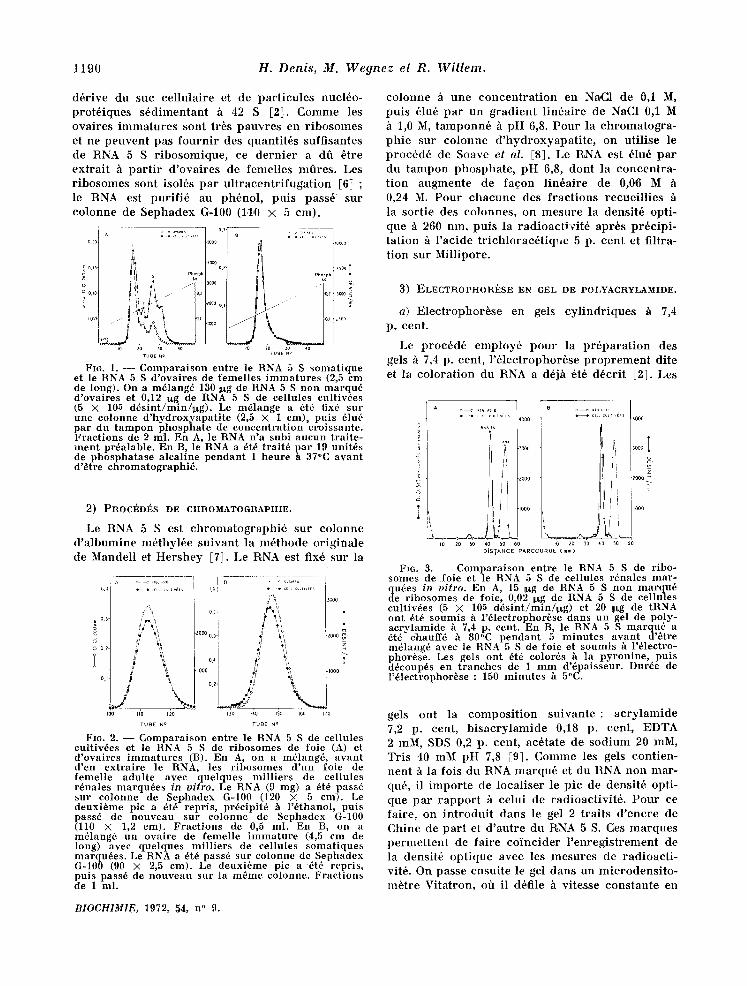
d 6 r i v e d u suc c e l l u l a i r e et de p a r t i c u l e s nuc l6o - p r o t 6 i q u e s s 6 d i m e n t a n t h 42 S [2]. C o m m e les o v a i r e s i m m a t u r e s s o n t t r6s p a u v r e s en r i b o s o m e s et n e p e u v e n t p a s f o u r n i r des q u a n t i t 6 s s u f f i s a n t e s de RNA 5 S r i b o s o m i q u e , ce d e r n i e r a dO 6 t re e x t r a i t h p a r t i r d ' o v a i r e s de f eme l l e s m o r e s . Les r i b o s o m e s s o n t iso16s p a r u l t r a c e n t r i f u g a t i o n [6] ; le RNA est p u r i f i 6 au p h 6 n o l , p u t s p a s s 6 s u r c o l o n n e de S e p h a d e x G-100 (110 × 5 cm) .
1 ,o zo ~o ~o ~
ruse ~o ru~ m°
FI6. 1. - - - Compara i son entre le RNA 5 S somat ique et le RNA 5 S d 'ovaires de femelles i m m a t u r e s (2,5 em de long). On a m6lang6 130 :lxg de RNA 5 S non marqu~ d 'ovaires et 0,12 ~xg de RNA 5 S de eellules eultiv6es (5 X 105 d~s in t /min /~g ) . Le m~lange a 6t6 fix6 sur une eolonne d 'hydroxyapa t i t e (2,5 X 1 crn), puis 61u6 par du t ampon phospha te de concen t ra t ion croissante. Frac t ions de 2 ml. En A, le RNA n 'a subi aucun t ra i te - men t pr6alable. En B, le RNA a 6t6 t ra i t6 pa r 19 unitds de phospha tase a leal ine pendan t 1 heure h 37°C avan t d 'e t re chromatographi6 .
2) PROCI~.D~S DE CHROMATOGRAPHIE.
Le RNA 5 S es t c h r o m a t o g r a p h i 6 s u r c o l o n n e d ' a l b u m i n e m 6 t h y l 6 e s u i v a n t la m 6 t h o d e o r i g i n a l e de M a n d e l l et H e r s h e y [7]. Le RNA est fix6 s u r la
~,1~ : : Y ? : ; . . . . . . . . . . ' ° ..........
I ,?\ .... 02 !
z
c o l o n n e ~ u n e c o n c e n t r a t i o n e n Na.Gl de 0,1 M, p u t s 61u6 p a r u n g r a d i e n t l i n 6 a i r e de NaC1 0,1 M
1,0 M, t a m p o n n 6 h p H 6,8. P o u r la c h r o m a t o g r a - p h i e su r c o l o n n e d ' h y d r o x y a p a t i t e , o n u t i l i s e le p r o c 6 d 6 de Soave et al. [8]. Le RNA est 61u6 p a r d u t a m p o n p h o s p h a t e , p H 6,8, d o n t la c o n c e n t r a - t i o n a u g m e n t e de f a c o n l i n 6 a i r e de 0,06 M 0,24 M. P o u r c h a c u n e des f r a c t i o n s r e c u e i l l i e s h la s o r t i e des c o l o n n e s , o n m e s u r e la d e n s i t 6 op t i - que h 260 nm, p u t s l a r a d i o a c t i v i t 6 a p r 6 s p r 6 c i p i - t a t i o n h l ' a c i d e t r i c h l o r a c 6 t i q , , e 5 p. c e n t e t f i l t r a - t i o n s u r Mi l l i po re .
3) ELECTROPHORESE E.N GEL DE POLYACRYLAMIDE.
a) E l e c t r o p h o r ~ s e e n gels c y l i n d r i q u e s h 7,4 p. c en t .
Le p r o c 6 d 6 e m p l o y 6 p o u r la p r 6 p a r a t i o n des gels "h 7,4 p. cen t , l ' 6 1 e c t r o p h o r 6 s e p r o p r e m e n t d i t e et la c o l o r a t i o n du RNA a d6 jh 6t6 d 6 c r i t [2]. Les
A
i
6
d
Io ~o 50 40 50
DIS
4000
5000
2000
I000
,ooo!
~ooo ~
60 ~0 20 30 40 50 60 ~ N C E PARCOUF~UE Cmm)
Fro. 3. - - Compara l son entre le RNA 5 S de r ibo- somes de fo ie et le RNA 5 S de eellules r6nales mar - qu6es in vitro. En A, 15 I~g de RNA 5 S non marqu6 de r ibosomes de foie, 0,02 10g de RNA 5 S de cellules cultiv6es (5 × 105 d6sint/min/,~xg) et 20 ~xg de tRNA out 6t6 soumis h l '61ectrophor6se dans un gel de poly- acry lamide h 7,4 p. cent. En B, le RNA 5 S marqu6 a 6t6 chauff6 h 80°C pendan t 5 minu tes avan t d 'dtre m61ang6 avee le RNA 5 S de foie et soumis h l'61ectro- phorSse. Les gels ont 6td color6s h la pyronine , puts d6coup6s en t ranches de 1 m m d'6paisseur. Dur~e de l '6lectrophor6se : 150 minu te s h 5°C.
100 II0 120 150 ~40 150 160 I?0
TUBE N ° TUBE N °
Fro. 2. - - Compara i son entre le RNA 5 S de cellules cultiv6es et le RNA 5 S de r ibosomes de foie (A) et d 'ovaires immatu re s (B). En A, on a m61ang6, avan t d 'en ext ra i re le RNA, les r ibosomes d 'un foie de femelle adul te avec quelques mi l l ie rs de eellules r~nales marqu6es in vitro. Le RNA (9 mg) a ~t~ pass6 sur colonne de Sephadcx G-100 (120 X 5 cm). Le deuxi6me p i c a ~t~ repris, pr6cipit~ h l '6 thanol , puts pass~ de nouveau sur colonne de Sephadex G-100 (110 × 1,2 cm). Frac t ions de 0,5 ml. En B, on a m~lang~ un ovaire de femelle i m m a t u r e (4,5 cm de long) avec quelqucs mil l iers de cellules somat iques marquees . Le RNA a 8t6 pass6 sur colonne de Sephadex G-100 (90 × 2,5 cm). Le deuxi6me p i c a ~t~ repris, puts pass~ de nouveau sur la mSme colonne. Fract ions de 1 ml.
BIOCH!M1E, 1972, 54, n ° 9.
gels o n t la c o m p o s i t i o n s u i v a n t e : a c r y l a m i d e 7,2 p. cen t , b i s a c r y l a m i d e 0,18 p. c en t , E!DTA 2 raM, SDS 0,2 p. cen t , a c6 t a t e de s o d i u m 20 raM, T r i s 40 mM p H 7,8 [9]. C o m m e les gels c o n t i e n - n e n t h la fo is d u RNA m a r q u 6 et d u RNA n o n m a r - qu6, il i m p o r t e de l o c a l i s e r le p i c de d e n s i t 6 op t i - que p a r r a p p o r t h ce lu i de r a d i o a c t i v i t 6 . P o u r ce fa i re , o n i n t r o d u i t d a n s le gel 2 t r a i t s d ' e n c r e de C h i n e de p a r t e t d ' a u t r e d u RNA 5 S. Ces m a r q u e s p e r m e t t e n t de f a i r e c o i n c i d e r l ' e n r e g i s t r e m e n t de la d e n s i t 6 o p t i q u e a v e c les m e s u r e s de r a d i o a c t i - vi t6. On p a s s e e n s u i t e le gel d a n s u n m i c r o d e n s i t o - m 6 t r e V i t a t r o n , off il d6,file ~ v i t e s se c o n s t a n t e e n
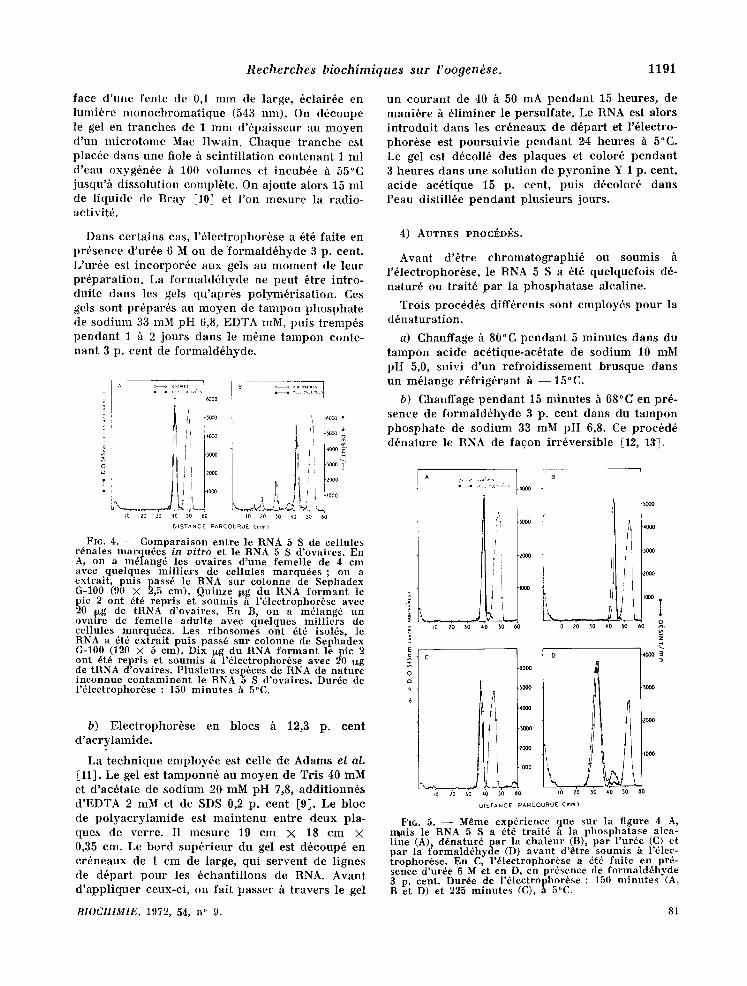
R e c h e r c h e s b i o c h i m i q u e s s u r l 'oogendse. 1191
face d ' u n e f e n t e de 0,1 m m de l a rge , 6c l a i r6e e n l u m i 6 r e m o n o c h r o m a t i q u e (543 n m ) . On d 6 c o u p e le gel en t r a n c h e s de 1 m m d ' 6 p a i s s e u r au m o y e n d ' u n m i c r o t o m e Mac I l w a i n . C h a q u e t r a n c h e es t p l a c 6 c d a n s u n e f iole h s c i n t i l l a t i o n c o n t e n a n t 1 m l d ' e a u o x y g 6 n 6 e h 100 v o l u m e s et i n c u b 6 e h 55°C jusqu"h d i s s o l u t i o n c o m p l b t e . On a jou te a lo r s 15 m l de l i q u i d e de B r a y [10] et l ' o n m e s u r e la r a d i o - ac t iv i t6 .
D a n s c e r t a i n s cas, l ' 6 1 e c t r o p h o r b s e a 6t6 f a i t e e n p r 6 s e n c e d ' u r 6 e 6 M ou de f o r m a l d 6 h y d e 3 p. cen t . L ' u r 6 e esl i n c o r p o r 6 e aux gels au m o m e n t de l e u r p r 6 p a r a t i o n . La f o r m a l d 6 h y d e n e p e u t 6 t re i n t r o - d n i t e d u n s les gels q u ' a p r b s p o l y m 6 r i s a t i o n . Ces gels s o n t p r 6 p a r 6 s au m o y e n de t a m p o n p h o s p h a t e de s o d i u m 33 mM p H 6,8, E D T A raM, p u t s t r e m p 6 s p e n d a n t 1 "h 2 j o u r s d a n s le m 6 m e t a m p o n c o n t e - n a n t 3 p. c e n t de f o r m a l d 6 h y d e .
CELL CULTiVEE~
5O0O 6000 '~
~ 4000 5000
{ 4ooo ~" 30OO -~
? I I .~ooo
1000 i .1000 ,I
10 ~0 50 40 50 60 ~0 ~0 30 40 50 50
DISTANCE PA~COU~UE (mm)
FIG. 4. - - Compara ison ent re le RNA 5 S de cellules r6nales marqu6es in vitro et le RNA 5 S d 'ovaires. En A, on a m61ang6 les ovaires d 'une femelle de 4 cm avec quelques mi l l ie rs de cellules marqudes ; on a extrai t , puts pass6 le RNA sur colonne de Sephadex G-100 (90 × 2,5 cm). Quinze ~g du RNA f o r m a n t le pic 2 ont 6t6 repr is et soumis h l '61ectrophor6se avec 20 .~g de tRNA d'ovaires. En B, on a mdlang6 un ova~re de femelle adul te avec quelques mi l l ie rs de cellules marqudes. Les r ibosomes ont 6t6 isol6s, le RNA a 6t6 ex t ra i t puts pass6 sur colonne de Sephadex G-100 (120 X 5 em). Dix ~o.g du RNA f o r m a n t le pic 2 ont 6t6 repr is et soumis ~ l '61eetrophor6se avec 20 ug de tRNA d'ovaires. P lus ieurs esp6ces de RNA de na ture inconnue con t amin en t le RNA 5 S d 'ovaires. Dur6e de l '61ectrophordse : 150 minu te s h 5°C.
b) E l e c t r o p h o r b s e e n b l o c s h 12,3 p. c e n t d'aclD, l a m i d e .
La t e c h n i q u e e m p l o y 6 e es t ce l le de A d a m s et al. [11]. Le gel est t a m p o n n 6 a u m o y e n de T r i s 40 m M et d ' a c 6 t a t e de s o d i u m 2,0 m M p H 7,8, a d d i t i o n n 6 s d ' E D T A 2 mM et de SDS 0,2 p. c e n t [9]. Le b l o c de p o l y a c r y l a m i d e est m a i n t e n u e n t r e d e u x p la - ques de v e r r e . I1 m e s u r e 19 c m × 18 c m X 0,35 cm. Le b o r d s u p 6 r i e u r d u gel es t d 6 c o u p 6 e n c r 6 n e a u x de 1 c m de l a rge , q u i s e r v e n t de l i g n e s de d 6 p a r t p o u r les 6 c h a n t i l l o n s de RNA. A v a n t d ' a p p l i q u e r ceux-c i , o n fa i r p a s s e r h t r a v e r s le gel
BIOCHIMIE, 1972, 54, n ° 9.
u n c o u r a n t de 40 /t 50 m A p e n d a n t 15 h e u r e s , de m a n i 6 r e h 61 t ra iner le p e r s u l f a t e . Le RNA est a l o r s i n t r o d u i t d u n s les c r 6 n e a u x de d 6 p a r t et l '61ect ro- p h o r 6 s e es t p o u r s u i v i e p e n d a n t 24 h e u r e s h 5°C. Le gel es t d6co l l6 des p l a q u e s et co lo r6 p e n d a n t 3 h e u r e s d a n s u n e s o l u t i o n de p y r o n i n e Y 1 p. c en t , a c i d e a c 6 t i q u e 15 p. cen t , p u t s d 6 c o l o r 6 d a n s l ' e a u d i s t i l l 6e p e n d a n t p l u s i e u r s j ou r s .
4) AUTRES PROC~DI~S.
A v a n t d ' 6 t r e c h r o m a t o g r a p h i 6 ou s o u m i s h l ' 6 1 e c t r o p h o r b s e , le RNA 5 S a 6t6 q u e l q u e f o i s d6- n a t u r 6 ou t r a i t 6 p a r la p h o s p h a t a s e a l ea l i ne .
T r o i s p r o e 6 d 6 s d i f f 6 r e n t s s o n t e m p l o y 6 s p o u r ]a d 6 n a t u r a t i o n .
a) C h a n f f a g e h 80°G p e n d a n t 5 m i n u t e s d a n s d u t a m p o n a c i d e a c 6 t i q u e - a c 6 t a t e de s o d i u m 10 mM p H 5,0, s u i v i d ' n n r e f r o i d i s s e m e n t b r u s q u e d a n s u n m d l a n g e r 6 f r i g d r a n t h - - 1 5 ° C .
b) C h a u f f a g e p e n d a n t 15 m i n u t e s h 68°C e n p r6 - s e n c e de f o r m a l d 6 h y d e 3 p. c e n t d a n s d u t a m p o n p h o s p h a t e de s o d i u m 33 m M p H 6,8. Ce p r o c 6 d 6 d 6 n a t u r e le RNA de f a c o n i r r 6 v e r s i b l e [12, 13].
; j
8 i
I I
¸4000
¸3OOO
i .~000
ICO0
10 20 30 40 50 60 ~ ,
z
• ~
600O
5000 • .~
• 2
l0 20 30 60 10 20 50 40 50 60
DISTANCE OARCOURUE (.mm)
D j4000 3
,1
I I
Fro. 5. - - M~me exp6rience que sur la figure 4 A, mats le RNA 5 S a 6t6 t ra i t6 h la phospha tase alca- l ine (A), d6natur6 p a r la cha leur (B), par l 'ur6e (C) et pa r la formald6hyde (D) avan t d '6tre soumis /t l'61ec- t rophor6se. En C, l '61ectrophor6se a 6t6 fai te en pr6- sence d 'ur6e 6 M e t en D, en pr6sence de fo rmaldehyde 3 cent. Dur6e de l 'dlectrophor6se : 150 minu tes (A, B Pt D) et 225 minu tes (C), h 5°C.
8 1
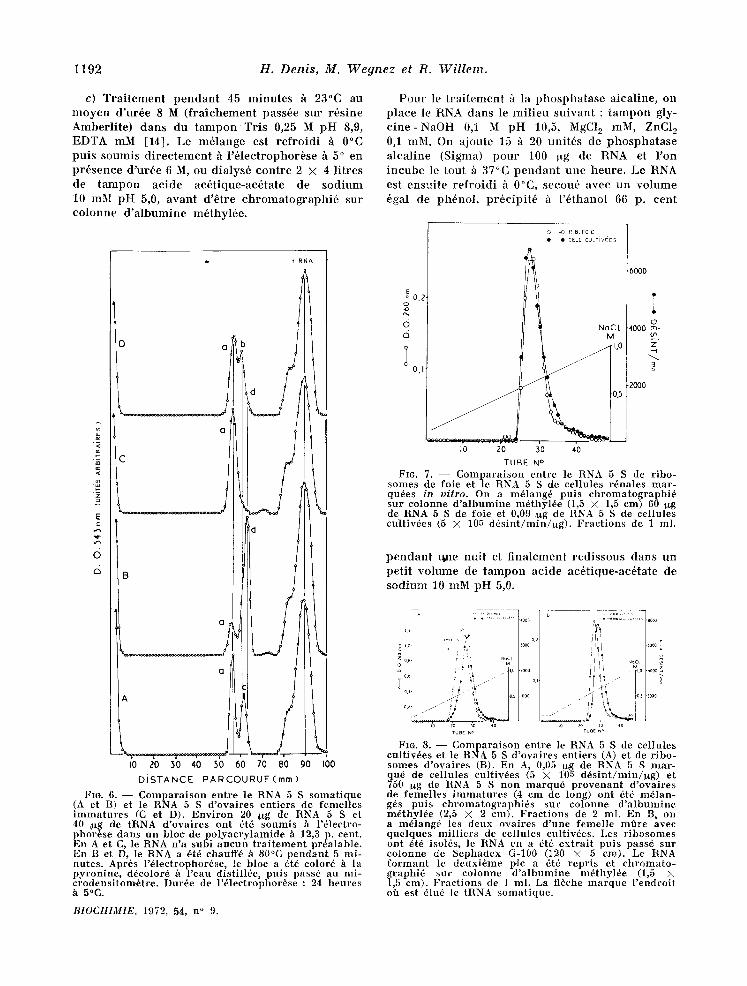
1192 H. Denis, M. Wegnez et R. Willem.
c) Traitement pendant 45 minutes h 23°C au m o y e n d'ur6e 8 M (fraiehement pass6e sur r6sine Amberlite) dans du tampon Tris 0,25 M pH 8,9, EDTA mM [14]. Le m61ange est refroidi h 0°C puis soumis d irectement h l'61ectrophorbse h 5 ° en pr6sence d'ur6e 6 M, ou dialys6 contre 2 X 4 l itres de tampon acide ac6tique-ac6tate de sod ium 10 mM pH 5,0, avant d'6tre chromatographi6 sur colonne d'albumine m6thy16e.
z
E
6 d
Pour le t ra i tement '~ la phosphatase aicaline, on place le RNA dans le mi l ieu sn ivant : t ampon gly- c i n e - N a O H 0,1 M pH 10,5, MgC12 raM, ZnCI2 0,1 mM. On ajoute 15 h 20 unit6s de phosphatase alcal ine (Sigma) pour 100 ~tg de RNA et l 'on incube le tout fi 37°C pe nda n t une heure. Le RNA est ensuite refroidi h 0°C, secou6 avec un volume 6gal de ph6nol, pr6cipit6 "h l '6thanol 66 p. cent
I ~ Ria. FO[E I I~ l l CELLCULT[VEES
E ~0,2'
g r~ o c5
0,1-
10 20 50 40
TUBE N °
6000
0 NaCl 4000 ra-
M w. 1,0 z
2000 0,5
FIG. 7. - - C o m p a r a i s o n ent re le RNA 5 S de r ibo- somes de foie et le RNA 5 S de eellules r6nales m a r - qn~es in oitro. On a mdlang6 pu i s e h r o m a t o g r a p h i ~ su r eo lonne d ' a l b u m i n e mdthyl~e (1,5 X 1,5 em) 60 ~g de RNA 5 S de foie et 0,09 lug de RNA 5 S de eellnles cultiv~es (5 × 105 d ~ s i n t / m i n / ~ g ) . F rac t ions de 1 ml.
pendan t t~ne nui t et f inalement redissous duns un petit volume de tampon aeide ae~tique-ae6tate de sodium 10 mM pH 5,0.
o ~-
I0 20 30 40 50 60 70 80 90 I00
DISTANCE PARCOURUE (mm)
Fig. 6. - - C o m p a r a i s o n ent re le RNA 5 S s o m a t i q u e ( A e t B) et le RNA 5 S d 'ova i res en t ie rs de femel les i m m a t u r e s (C et D). E n v i r o n 20 ~g de RNA 5 S e t 40 !ttg de tRNA d 'ova i res ont 6t6 soumis h l 'dlectro- phor6se dans un bloc de p o l y a c r y l a m i d e h 12,3 p. cent. En A et C, le RNA n 'a subi a u c u n t r a i t e m e n t pr6alable . En B e t D, le RNA a 6t~ ehauff~ h 80°C p e n d a n t 5 mi- nutes . Apr6s l 'd leetrophor6se, le bloe a 6t6 colord h la py ron ine , ddeolor~ h l 'eau distillde, pu i s pass6 au mi- crodensitom,~tre. Dur6e de l 'd lec t rophor6se : 24 heures h 5°C.
BIOCH1MIE, 1972, 54, n ° 9.
FIG. 8. - - C o m p a r a i s o n ent re ]e RNA 5 S de cel]ules cult iv6es et le RNA 5 S d 'ova i res en t ie rs (A) et de r ibo- somes d 'ova i res (B). En A, 0,05 ~g de RNA 5 S m a r - qu6 de eel lules cult iv6es (5 X 105 d6s in t /m in /ug ) et 750 ug de RNA 5 S n o n m a r q u 6 p r o v e n a n t d 'ova i res de femel les i m m a t u r e s (4 cm de long) on t 6t6 m61an- g6s puis ch ro ina tog raph i6s su r colonne d ' a l b u m i n e m6thyl6e (2,5 X 2 cm). F rac t ions de 2 ml. En B, on a m61ang6 les deux ovaires d 'une femelle mfire avec que lques mi l l i e r s de eellules cultiv6es. Les r i b o s o m e s ou t ~t~ isol~s, le RNA en a ~t6 ex t r a i t pu i s pass~ s u r co lonne de Sephadex G-100 (120 × 5 era). Le RNA f o r m a n t le deuxi~me pie a dt6 repr i s et c h r o m a t o - graphi(~ su r colonne d ' a l b u m i n e mdthyl~e (1,5 × 1,5 em). F rac t ions de l ml. La fl~che m a r q u e l ' endroi t off est 61u6 le tRNA somat ique .
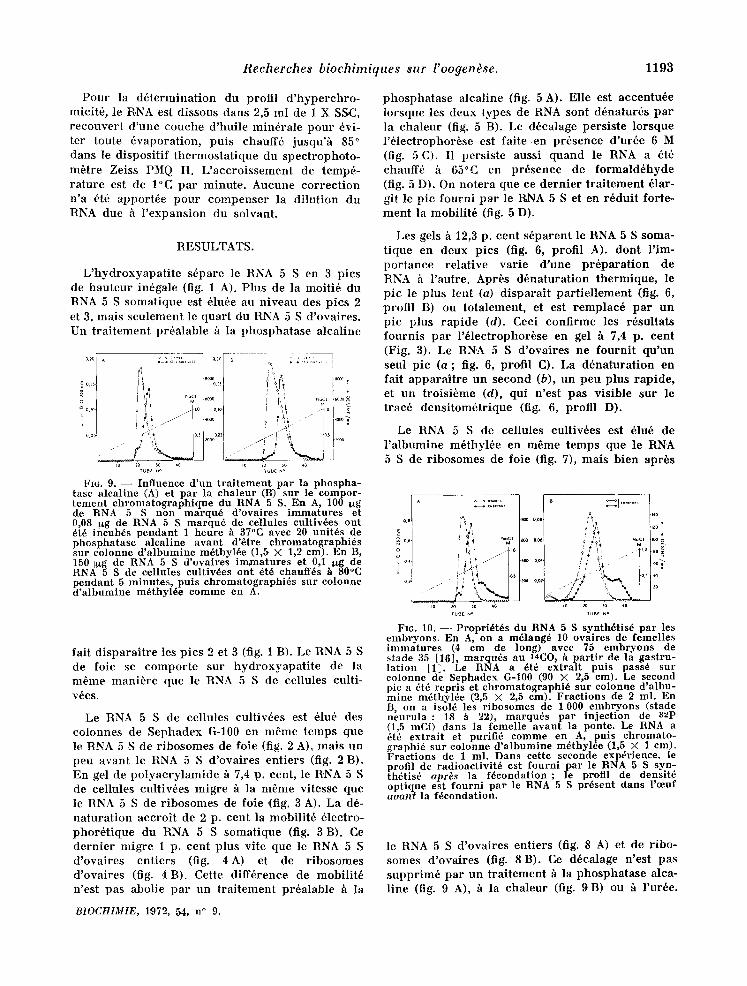
Recherches biochimiques sur l'oogen~se. 1193
Pour la d6terminat ion du profil d ' hype reh ro - micit6, le RNA est dissous dans 2,5 ml de 1 X SSC, reeouver t d 'une couehe d 'hui le min6rale pour 6vi- ter toute 6vaporat ion, puts chauff6 jusqu'h 85 ° dans le disposi t i f thermosta t ique du spectrophoto- m~tre Zeiss PMQ H. L 'accro i ssement de temp6- ra ture est de I °C par minute. Aucune cor rec t ion n 'a 0 6 apport6e pour compenser la di lut ion du RNA due h l ' expans ion du solvant.
RESULTATS.
L 'hydroxyapa t i t e s6pare le RNA 5 S e n 3 pies de hauteur in6gale (fig. 1 A). Plus de la moiti6 du RNA 5 S somat ique est 61u6e au niveau des pies 2 et 3, mats seulement le quart du RNA 5 S d 'ovaires. Un t ra i tement pr6alable h la phosphatase alcal ine
o:0 I ...... .....
io to ~o 40
Fro. 9. - - Influence d'un traitement par la phospha- tase alcaline (A) et par la chaleur (B) sur le compor- tement ehromatographique du RNA 5 S. En A, 100 ug de RNA 5 S non marqu6 d'ovaires immatures et 0,08 I~g de RNA 5 S marqu6 de cellules cultiv6es oat 6t6 incub6s pendant 1 heure h 37°C avec 20 unit6s de phosphatase alcaline avant d'etre chromatographi6s snr colonne d'albumine m6thyl6e (1,5 × 1,2 cm). En B, 150 I~g de RNA 5 S d'ovaires imxtaatures et 0,1 ltg de RNA 5 S de c ellules eultiv6es ont 6t6 chauff6s h 80°C pendant 5 minutes, puts chromatographi~s sur colonne d'albumine m6thyl6e comme en A.
fair d ispara i t re les pies 2 et 3 (fig. 1 B). Le RNA 5 S de foie se compor te sur hydroxyapa t i t e de la m6me mani+re que le RNA 5 S de cellules culti- v6es.
Le RNA 5 S de cellules cultiv6es est 61u6 des colonnes de Sephadex G-100 en m6me temps que le RNA 5 S de r ibosomes de foie (fig. 2 A), mats un peu avant le RNA 5 S d 'ovai res ent iers (fig. 2 B). En gel de po lyac ry l amide h 7,4 p. cent, le RNA 5 S de cellules cultiv6es migre h la m6me vitesse que le RNA 5 S de r ibosomes de foie ,(fig. 3 A). La d6- natura t ion accroi t de 2 p. cent la mobil i t6 61ectro- phor6t ique du RNA 5 S somat ique (fig. 3 B). Ce dern ie r migre 1 p. cent plus vite que le RNA 5 S d 'ovai res ent iers (fig. 4A) et de r ibosomes d 'ovai res (fig. 4 B). Cette diff6rence de mobili t6 n 'est pas abolie par un t ra i tement pr6alable h la
phosphatase alcal ine (fig. 5 A). Elle est accentu6e lorsque les deux types de RNA sont d6nalur6s par la chaleur (fig. 5 B). Le d6ealage persis te lorsque l '61ectrophorbse est faite e n pr6sence d 'ur6e 6 M (fig. 5 C). II persis te aussi quand le RNA a 6t6 chauff6 h 65°C en pr6senee de formald6hyde (fig. 5 D). On notera que ce dern ie r t ra i tement 61ar- git le p ic fourni pa r le RNA 5 S e t en r6duit forte- ment la mobili t6 (fig. 5 D).
Les gels ~ 12,3 p. cent s6parent le I~NA 5 S soma- t ique en deux pies (fig. 6, pro411 A). dont l ' im- por t ance relat ive var ie d 'une pr6para t ion de RNA h l 'autre. Apr~s d6naturat ion thermique , le p ic le plus lent (a) d ispara l t pa r t i e l l ement (fig. 6, profil B) ou totalement , et est remplac6 pa r un p ic plus rap ide (d). Ceci confi rme les r6sultats fournis pa r l '61ectrophorbse en gel h 7,4 p. cent (Fig. 3). Le RNA 5 S d 'ovai res ne fourni t qu 'un seul p ie ( a ; fig. 6, prafil C). La d6naturat ion en fair appara l t re un second (b), un peu plus rap ide , et un t rois ibme (d), qui n'est pas vis ible sur le trac6 dens i tom6tr ique (fig. 6, profil D).
Le RNA 5 S de cellules cultiv6es est 61u6 de l ' a lbumine m6thyl6e en m6me temps que le RNA 5 S de r ibosomes de foie (fig. 7), mats bien apr6s
H ~ ' . . . . .
o0 ,' i!'. ~[,.0
~UBE N °
140
i~ '°I , ,
4 i"" . . . . . . 7
io ~o ~o 4 0
Tua r NO
Fro. 10. - - Propri6t6s du RNA 5 S synth6tis6 par les embryons. En A, on a m61ang6 10 ovaires de femelles immatures (4 cm de long) avec 75 embryons de stade 35 [IS], marquis an ~4CO~ h I?artir de la gastru- lation Ell. Le RNA a 6t6 extra]t puts pass6 sur colonne de Sephadex G-100 (90)< 2,5 cm). Le second pie a 6t6 repris et chromatographi6 sur eolonne d'albu- mine m6thyl6e (2,5 × 2,5 era). Fractions de 2 ml. En B, on a isol6 les ribosomes de 1 000 embryons (stade neurula : 18 h 22), marqu6s par injection de 32p (1,5 mCi) dans la femelle avant la ponte. Le RNA a 6t6 extrait et purifi6 comme en A, puts chromato- graphi6 sur colonne d'albumine m6thyl6e (1,5 X 1 cm). Fractions de 1 ml. Dans cette seeonde exp6rienee, le profil de radioactivit6 est fourni par le RNA 5 S syn- th6tis6 apr~s la f6condation; le profit de densit6 optique est fourni par le RNA 5 S pr6sent dans l'ceuf avant la f6condation.
le RNA 5 S d 'ovai res entiers (fig. 8 A) et de r ibo- somes d 'ovai res (fig. 8 B). Ce d6ealage n'est pas suppr im6 par un t ra i tement h la phosphatase alea- l ine (fig. 9 A), h la chaleur (fig. 9B) ou h l 'ur6e.
BIOCHIMIE, 1972, 54, n ° 9.
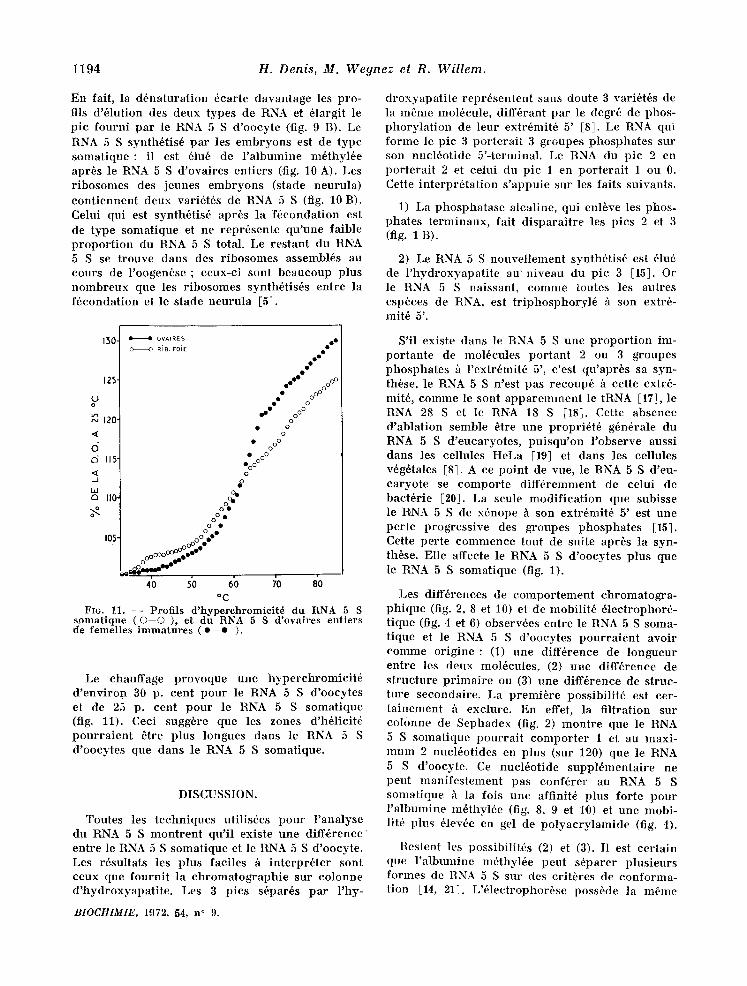
1194 H. Denis, M. Wegnez el R. Willem.
En fait, la d6natura t ion 6carte davantage les pro- ills d'61ution des deux types de RNA et 61argit le pic fourni par le R~NA 5 S d 'oocyte (fig. 9 B). Le RNA 5 S synth6tis6 par les e lnbryons est de type so lna t ique : il est 61u6 de l ' a lbumine m~thyl6e apr6s le RNA 5 S d 'ovaires entiers (fig. 10 A). Les r ibosomes des jeunes embryons (stade neurula) con t i ennen t deux vari6t6s de RNA 5 S (fig. 10 B). Celui qui est synth6tis6 apr6s la f6condat ion est de type somatique et ne repr6sente qu 'une faible p ropor t ion du RNA 5. S total. Le restant du RN~A 5 S se trouve dans des r ibosomes asselnbl6s au cours de l 'oogen6se ; ceux-ci sont beaucoup plus nombreux que les r ibosomes synth6tis6s entre la f6condation et le stade neuru la [5].
130-
125-
O o
120- <
d ,N 115" < J kd a 110-
~ OVAiRES
C ~ RiB. FOiE ~OOQ IQQ
: '" oooO°°° e $ O ..- ooO
• 0 0
• 0 oo
• oo %0 ° 0 .o
o°° 0 •
ms ooO°O ** oooO~_°... "
o o ° ,,,
40 50 6'0 o C
~0 8'0
FrG. 11. - - P r o f i l s d ' h y p e r c h r o m i c i t 6 d u R N A 5 S s o m a t i q u e ( O - - O ) , e t d u R N A 5 S d ' o v a i r e s e n t i e r s de f e m e l l e s i m m a t u r e s ( e - • ).
Le chauffage provoque une hyperchromic i t6 d ' env i ron 30 p. cent pour le RNA 5 S d'oocytes et de 25 p. cent pour le RNA 5 S somatique (fig. 11). Ceci sugg6re que les zones d'h61icit6 pour ra i en t 6tre plus longues dans le RNA 5 S d'oocytes que dans le RNA 5 S solnatique.
DISCUSSION.
Toutes les techniques utilis6es pour l 'analyse du RNA 5 S mon t ren t qu ' i l existe une d i f f6 rence entre le RNA 5 S somatique et le RNA 5 S d'oocyte. Les r6sultats les plus faciles ~ in terpr6ter sont eeux que fourni t la chromatographie sur colonne d 'hydroxyapat i te . Les 3 pics s6par6s par l 'hy-
droxyapat i te repr6sentent sans doute 3 vari6t6s de la m6me mol6cule, diff6rant par le degr6 de phos- phory la t ion de leur extr61nit6 5' [8]. Le RNA qui forlne le pic 3 por tera i t 3 groupes phosphates sur son nucl6otide 5 '- terminal. Le !~NA du pic 2 en porterai t 2 et celui du pic 1 en por tera i t 1 ou 0. Cette in terpr6ta t ion s 'appuie sur les faits suivants.
1) La phosphatase alcaline, qui enlbve les phos- phates ter lninaux, fait d isparal t re les pics 2 et 3 (fig. 1 B).
2) Le RNA 5 S nouvel le lnent synth6tis6 est 61u6 de l 'hydroxyapat i te a u niveau du pic 3 E15]. Or le RNA 5 S naissant, comnle routes les autres espbces de RNA, est tr iphosphory16 fi son extr6- 1nit6 5'.
S'il existe dans le RNA 5 S une propor t ion iln- por tante de 1nol6cules por tan t 2 ou 3 groupes phosphates "h l 'extr6mit6 5', c'est qu 'aprbs sa syn- th~se, le RNA 5 S n 'est pas recoup6 "h cettc extr6- mit~, colnme le sont apparemlnent le tRNA [17], le RNA 28 S et Ie RNA 18 S [183. Cette absence d 'abla t ion selnble 6tre une propri6t6 g6n6rale du RNA 5 S d 'eucaryotes, pu i squ 'on l 'observe aussi clans ]es cellules HeLa [19] et dans les cellules v6g6tales [8]. A ce point de vue, le RNA 5 S d'eu- caryote se comporte diff6relnlnent de celui de bact6rie [20]. La seule modif ica t ion que subisse le R~NA 5 S de x6nope ~ son extr6mit6 5' est une perte progressive des groupes phosphates [15]. Cette perte commence tout de suite aprbs la syn- lh6se. Elle affecte le RNA 5 S d'oocytes plus que le RNA 5 S somatique (fig. 1).
Les diff6rences de compor tement chromatogra- phique (fig. 2, 8 et 10) et de 1nobilit6 61ectrophor6- t ique (fig. 4 et 6) observ6es entre le RNA 5 S solna- tique et le RNA 5 S d'oocytes pour ra ien t avoir colnlne origine : (1) une diff6rence de longueur entre les deux mol6cules, (2) une diff6rence de s t ructure pr i lna i re ou (3) une diff6rence de struc- ture secondaire. La prelni6re possibili t6 est cer- ta ine lnent ix exclure. En effet, la fi l tration sur col0nne de Sephadex (fig. 2) mont re que le RNA 5 S somatique pour ra i t colnporter 1 et au maxi- 1nuln 2 nucl6otides en plus (sur 120) que le RNA 5 S d'oocyte. Ce nucl6otide supp161nentaire ne peut 1nanifestelnent pas conf6rer au RNA 5 S solnatique h la fois une affinit6 plus forte pour l ' a lbuinine In6thyl6e (fig. 8, 9 et 1~0) et une mobi- lit6 plus 61ev6e en gel de polyacryla ln ide (fig. 4).
Restent les possibilit6s (2) et (3). II est cer ta in que l ' a lbumine 1n6thyl6e peot s~parer plusieurs forlnes de RNA 5 S sur des crit6res de conforlna- l i o n [14, 21 ] . L'61eetrophor6se possbde la 1n~1ne
BIOCHIM1E, 1~972, 54, n ° 9.

R e c h e r c h e s b i o c h i m i q u e s sur l'oogenOse. 1195
p r o p r i 6 t 6 p u i s q u e les d e u x f o r m e s de RNA 5 S s o m a t i q u e s6par6es p a r les gels c o n c e n t r 6 s (fig. 6 A) on t la m 6 m e s t r u c t u r e p r i m a i r e [22]. On est tent6 d ' e n d 6 d u i r e que les d i v e r s e s var i6 t6s de RNA 5 S, et en p a r t i c u l i e r le RNA 5 S s o m a t i q u e et le RNA 5 S d ' o o c y t e s , ne d i f f6 r en t l ' u n e de l ' a u t r e que p a r l eu r s t r u c t u r e s e e o n d a i r e . Cette con c l u - s ion est c o m p a t i b l e avec les r6su l t a t s des exp6- r i e n c e s de d 6 n a t u r a t i o n t h e r m i q u e (fig. 11). Toute - tots , le RNA 5 S s o m a t i q u e c o n t i n u e h m i g r e r en gel de p o l y a c r y l a m i d e un p e u p lus vi te que le RNA 5 S d ' o o e y t e s , d a n s des c o n d i t i o n s qui s o n t cens6es a b o l i r rou te s t r u c t u r e s e c o n d a i r e (fig. 5 C et D). P a r a i l l eurs , si les deux t y p e s de RNA poss6- d a i e n t la rn6me s t r u c t u r e p r i m a i r e , on p o u r r a i t s ' a t t e n d r e h ce qu ' i l s a d o p t e n t , ap r6s d 6 n a t u r a t i o n , une c o n f i g u r a t i o n i d e n t i q u e . Or, il n ' e n est r i e n : les p r o p r i 6 t 6 s du I~NA 5 S d ' o o c y t e s et du RNA 5 S s o m a t i q u e s o n t p lus d i f f6 r en t e s a p r b s la d 6 n a t u r a - t ion q u ' a v a n t (fig. 4 et 5 B, 6, 8 et 9 B). I1 est d o n c p o s s i b l e que le RNA 5 S d ' o o c y t e s et le RNA 5 S s o m a t i q u e d i f f b r e n t h la fo is p a r l eu r s t r u c t u r e p r i m a i r e et p a r l eu r s t r u c t u r e s e c o n d a i r e . P o u r p r o u v e r qu ' i l en est b i en a ins i , il f a u d r a 6 t ab l i r la s 6 q u e n c e nuc16ot id ique c o m p l O e des d e u x mol6- cules . Ce t r ava i l es t en c o u r s ~ l ' h e u r e actuel le .
Remerciements.
M. Wegnez est boursier de l ' Ins t i tu t pour l 'Encou- ragement de la Recherche Scientiflque dans l ' Industr ie et l 'Agriculture. Ce travail a 6td rendu possible par l 'aide finanei6re du Fonds National de la Recherche Scientifique. Nous remercions le Professeur R. Monier d 'avoir bien voulu lire et er i t iquer le manuscri t .
R~suxi~.
Le RNA 5 S des oocytes de Xenopus laeois diffSre du RNA 5 S somatique par le degr6 de phosphoryla t ion de son extr6mit6 5' : le RNA 5 S somatique contient une plus forte proport ion de mol6cules por tant 2 ou 3 groupes phosphates sur leur nucl~otide 5 '- terminal .
Le RNA 5 S somatique est re tards par le Sephadex G-100 un peu moins que le RNA 5 S d'oocytes. Le RNA 5 S provenant de r ibosomes somatiques est 61u6 de l ' a lbumine mSthyl6e bien apr6s le RNA 5 S d'oocytes entiers et de r ibosomes d'oocytes. Les ribo- somes des jeunes embryons eont iennent deux types de RNA 5 S. Celui qui est synth~tis~ apr~s la f6con- dation est de type somatique et ne repr6sente qu 'une faible propor t ion du RNA 5 S total. Le res tant du RNA 5 S embryonnai re se trouve dans des r ibosomes assemblds au cours de l'oogen6se.
Le RNA 5 S somatique migre dans les gels de poly- acrylamide h 7,4 p. cent un peu plus r i te que le RNA 5 S d'oocytes. Ni l 'enl6vement des groupes phos- phates te rminaux au moyen de la phosphatase alca- line, ni la d6naturat ion par la ehaleur ou l 'ur6e ne suppr iment cette diff6rence de mobilit6. L'61eetropho- r6se en gel de polyacrylamide h 12,3 p. cent rdv~le l 'existence de deux formes distinctes de RNA 5 S somatique. L'une d 'entre elles a la m6me mobilit6 que
le RNA 5 S d'oocytes. L'autre forme migre plus vite. Le RNA 5 S d'oocytes et le RNA 5 S somatique rdagissent diffdremment h la d6naturat ion thermique : la plus grande pat t ie du RNA 5 S d'oocytes conserve, apr6s refroidissement , la m6me mobilit6 qu 'avant , tandis que le RNA 5 S somatique adopte une forme nouvelle, qui migre plus vite que toutes les autres.
Quand on le chauffe en solution saline, le RNA 5 S d'oocytes prdsente une hyperchromiei t6 un peu plus forte que le RNA 5 S somatique.
Ces rdsultats sugg6rent que le RNA 5 S somatique et le RNA 5 S d'oocytes poss6dent /~ la fois une struc- ture pr imaire et une structure secondaire diff6rentes.
ZUSAMMENFASSUNG.
Die RNS der Oozyten des Xenopus laevis unterschei- det sieh v o n d e r somatischen 5 S RNS durch den Phos- phorylat ionsgrad ihres 5' Endes : die somatisebe 5 S RNS enthhl t einen st/irkeren Anteil yon Molekiilen, die 2 oder 3 Phosphatgruppen auf ih rem 5'-Endnu- kleotid tragen.
Die 5 S RNS der Oozyten wird durch Sephadex G-100 ein wenig st/irker zurfickgehalten als die soma- tische 5 S RNS. Die aus den somatischen Ribosomeu s tammende 5 S RNS wird aus e iner S~iule methyl ier ten Albumin - Kieselgurs wesentl ieh sp~iter eluiert als 5 S RNS der ganzen Oozyten und der Ribosomen der Oozyten. Die Ribosomen der jungen Embryos enthal ten zwei Typen der 5 S RNS. Die eine wird nach der Befruchtung synthet is ier t , sie ist vom somatischen Typ und stellt nur einen schwachen Anteil der gesamten 5 S RNS dar. Die iibrige 5 S RNS des Embryos befindet sich in den Ribosomen~ die sich zum Zeitpunkt der Oogenese verbinden.
Die somatische 5 S RNS wander t in Polyacrylamidgel bet 7,4 p. cent sehneller als die 5 S RNS der Oozyten. Diese elektrophoret ische Beweglichkeit wird weder durch Abtrennung der Phosphatendgruppen mit tels der alkalischen Phosphatase noch durch Denaturierung durch W/irme oder Harnstoff beeintr/ichtigt. Die Elek- t rophorese in Polyacrylainidgel bet 12,3 p. cent enthii l l t die Existenz zweier untersehiedl icher Formen der somatischen 5 S RNS. Die eine davon hat dieselbe Beweglichkeit wie die 5 S RNS der Oozyten. Die andere Form wander t wesent l ich schneller. Die 5 S RNS der Oozyten und die somatische 5 S RNS reagieren in verscbiedener VCeise auf thermische Dena- tur ierung : der gr6sste Tell der 5 S RNS der Oozyten beh~ilt nach Abkiihlung dieselbe Beweglichkeit wie zuvor w~ihrend die somatische 5 S RNS eine neue Form annimmt , die schneller als alle andereu wandert .
Bet Erw~irmung in Salzl6sung zeigt die 5 S RNS eine st/irkere Hyperchromizit~it als die somatische 5 S RNS.
Die Ergebnisse legen es nahe, dass die somatische 5 S RNS und die 5 S RNS der Oozyten zugleieh eine untersehiedliche prim~ire und sekund~ire Struktur aufweisen.
BIBLIOGRAPHIE.
1. Mairy, M. & Denis, H. (1971) Develop. Biol., 24, 143. 2. Denis, H. & Mairy, M. (1972) Eur. J. Biochem., 25,
524. 3. Ford, P. 3. (1971) Nature, 233, 561. 4. Wegnez, M. a Denis, H. (1972) Biochimie, XX, 20. 5. Brown, D. D. ~ Littna, E. (1964) J. Mol. Biol., 8,
669.
BIOCH1MIE, 1972, 54, n ° 9.

1 1 9 6 H . D e n i s , M . W e g n e z e t R . W i l l e m .
6. Mairy, M. a Denis, H. (1972) Enr. J. Biochem., 25, 535.
7. Mandell, J. D. a Hershey, A. D. (1960) Anal. Bio- chem., 1, 66.
8. Soave, C., Galante, E. a Torti , G. (1970) Bull. Soc. Chim. Biol., 52, 857.
9. Loening, U. E. (1967) Biochem. J., 101, 251. 10. Bray, G. A. (19.60) Anal. Biochem., 1, 279. 11. Adams, J. M., Jeppesen, P. G. N., Sanger, F.
Barrell , B. G. (1969) Nature, 223, 1009. 12. Boedtker, H. (1967) Biochemis try , 6, 2718. 13. Cox, R. A. a Canaga l ingam, K. (1967) Biochem. g.,
103, 749. 14. Aubert , M., Scott, J. F., Reynier , M. a Monier, R.
(19.68) Proc. Natl. Acad. Sci. U.S.A., 61, 292. 15. Denis, H., Wegnez, M. & Wil lem, R., m a n u s c r i t
en prfiparation. 16. Nieuwkoop, P. D. s~ Faber, J. (1967) (( Normal Table
of Xenopus laeois (Daudin) >>, North Holland Pub l i sh ing Co.
17. Al tman, S. a Smith, J. D. (1971) Nature New Biol., 233, 35.
18. Maden, B. E. H. (1971) Progress in Biophysics and Molecular Biology, 22, 127.
19. Hatlen, L., Amaldi , F. ~ Attardi, G. (1969) Bio- chemistry , 8, 4989.
26. .Jordan, B. R., Feun teun , J. ~ Monier, R. (1970) J. Mot. Biol., 50~ 605.
21. Forget, B. G. ~: W e i s s m a n , S. M. (1967) Nature, 213, 878.
22. Wegnez, M. ,~ Denis, H., experiences in6dites.
Note ajoutde sur dpreuoe. - - La s t ruc ture p r imai re complete du RNA 5 S soma t ique et du RNA 5 S d 'ooeytes a 6t6 r6eemment d6termin6e (Wegnez, M., Monier, R. ~ Denis, H. (1972) FEBS Letters, 25, 13). Le3 deux types de RNA 5 S possbdent 6 r6sidus diff6- rents sur 120.
BIOCH1M1E, 1972, 54, n ° 9.