






Langages
Pages
Légal


BIOCHIMIE, 1972, 54, 1189-1196.
Recherches biochimiqucs sur l'oogenbse. V - Comparaison entre le R N A 5 S somat ique et le R N A 5 S des oocytes
de Xenopus laevis (*).
H e r m a n DENIS, M a u r i e e W~GNF.Z, et R o b e r t WILLEM.
Laboratoire de Biochimie, Unioersit~ de Liege, place Delcour, 17, B-#O00 Libge, Belgique.
(8/6/1972).
Summary. - - 5 S RNA f rom oocytes of Xenopus laeois differs f rom somat ic 5 S RNA by the degree of phosphory la t ion of i ts 5'-end. There is in somat ic 5 S RNA a h igher p ropor t ion of molecules car ry ing 2 or 3 phospha te groups on the 5 ' - t e rmina l nucleotide.
Somatic 5 S RNA was re tarded by columns of Sephadex G-100 s l ight ly less t h a n oocyte 5 S RNA. 5 S RNA f rom somatic r ibosomes was eluted f rom columns of me thy la t ed a lbumin-Kiese lguhr fa r beh ind 5 S RNA f rom whole oocytes and f rom oocyte r ibosomes. Ribosomes f rom ear ly embryos were found to con ta in 2 types of 5 S RNA : [1] newly made 5 S RNA which is of somat ic type and [2] bu lk 5 S RNA which is of the ooeyte type.
Somatic 5 S RNA migra ted in 7.4 p. cent po lyacry lamide gels s l ight ly ahead of oocyte 5 S RNA. This difference of e lectrophoret ic mobi l i ty was suppressed ne i the r by removal of the t e r m i n a l phospha tes by a lka l ine phospha tase no r by hea t or u rea dena tu ra t ion . Electrophoresis in 12.3 p. cent gels revealed the existence of 2 d is t inc t fo rms in somatic 5 S RNA. One of these had the same mobi l i ty as oocyte 5 S RNA. The o ther one migrated faster . Somatic and oocyte 5 S RNA's reacted different ly to hea t d e n a t u r a t i o n : a f te r cooling bo th nat ive forms of somat ic 5 S RNA were replaced by a th i rd one, which migra ted fas ter , whereas most of the oocyte RNA remained in na t ive form.
W h e n hea ted in saline, oocyte 5 S RNA shorted a s l ight ly h igher hyperchromie i ty t h a n somatic 5 S BNA.
These resul ts suggest t ha t bo th the p r ima ry and the secondary s t ruc tures are different in somatic and in oocyte 5 S RNA's.
I N T R O D U C ~ O N .
PrOs de la m o i t i ~ d u RNA p r 6 s e n t d a n s les o o c y t e s d u x 6 n o p e e n p e t i t a e e r o i s s e m e n t (d ia - m b t r e : 50 h 150 ~m) es t f o r m 6 e p a r d u R N A 5 S [1, 2, 3]. D a n s ee t r a v a i l , n o u s e o m p a r o n s les p r o - p r i 6 t 6 s d u RNA 5 S des o o e y t e s a v e e ee l l es d u RNA 5 S s o m a t i q u e . Nous 6 t u d i o n s le e o m p o r t e - m e n t de ees d e u x t y p e s de R N A s u r e o l o n n e s d ' h y - d r o x y a p a t i t e , de S e p h a d e x G-100 et d ' a l b u m i n e m 6 t h y l 6 e , l e u r m o b i l i t 6 6 1 e c t r o p h o r 6 t i q u e e n gel de p o l y a e r y l a m i d e , a i n s i que l e u r p r o f i l d ' h y p e r c h r o - m ie i t 6 .
Abrdoiations : tRNA : RNA de t rans fe r t . Tris : t r i s (hydroxym~thy l ) - aminom~thane . EDTA : acide ~thyl6nedini t r i lo t~traae~t ique, sel di-
sodique. SSC : NaC1 150 raM, c i t ra te de sodium 15 raM. SDS : doddeylsulfate de sodium. (*) Pou r les p remiers art icles de eette s~rie, voir
les r~f6renees [I, 2 et 6].
M,ETHODES.
1) PURIFICATION DU R N A 5 S.
Le RNA 5 S s o m a t i q u e es t p u r i t i 6 so i t h p a r t i r de ee l l u l e s r 6 n a l e s e u l t i v 6 e s in vitro e n p r 6 s e n e e d ' u r i d i n e [3HJ et de g u a n o s i n e [3Hi [4], so i t h p a r t i r de r i b o s o m e s de fo ie , so i t e n c o r e h p a r t i r d ' e m b r y o n s e n t i e r s m a r q u 6 s au 14CO 2 o u au azp [ I ] . Le RNA est e x t r a i t p a r le p r o e 6 d 6 au SDS- p h 6 n o l f r o i d [5] , p u i s p a s s 6 s u r c o l o n n e de S e p h a - dex G-100, e o m m e d 6 e r i t p r 6 e ~ d e m m e n t [13. Le RNA 5 S es t 61u6 jus t e a p r 6 s le RNA de p o i d s mo- 16eu la i re 61ev6.
Le RNA 5 S d ' o o e y t e s es t e x t r a i t h p a r t i r d ' o v a i r e s e n t i e r s de f e m e l l e s i m m a t u r e s o u de r i b o s o m e s de f eme l l e s mfl res . C o m m e n o u s l ' a v o n s m o n t r 6 p r 6 e 6 d e m m e n t , la q u a s i - t o t a l i t 6 d u R N A d ' o v a i r e s e n t i e r s p r o v i e n t de s o o e y t e s , e t n o n de s ee l lu l e s f o l l i e u l e u s e s qu i les e n t o u r e n t [13. L a m a - j e u r e p a r t i e d u RNA 5 S des o v a i r e s i m m a t u r e s
1 1 9 0 H. Denis , M. W e g n e z el R. W i l l e m .
d 6 r i v e d u suc c e l l u l a i r e et de p a r t i c u l e s nuc l6o - p r o t 6 i q u e s s 6 d i m e n t a n t h 42 S [2]. C o m m e les o v a i r e s i m m a t u r e s s o n t t r6s p a u v r e s en r i b o s o m e s et n e p e u v e n t p a s f o u r n i r des q u a n t i t 6 s s u f f i s a n t e s de RNA 5 S r i b o s o m i q u e , ce d e r n i e r a dO 6 t re e x t r a i t h p a r t i r d ' o v a i r e s de f eme l l e s m o r e s . Les r i b o s o m e s s o n t iso16s p a r u l t r a c e n t r i f u g a t i o n [6] ; le RNA est p u r i f i 6 au p h 6 n o l , p u t s p a s s 6 s u r c o l o n n e de S e p h a d e x G-100 (110 × 5 cm) .
1 ,o zo ~o ~o ~
ruse ~o ru~ m°
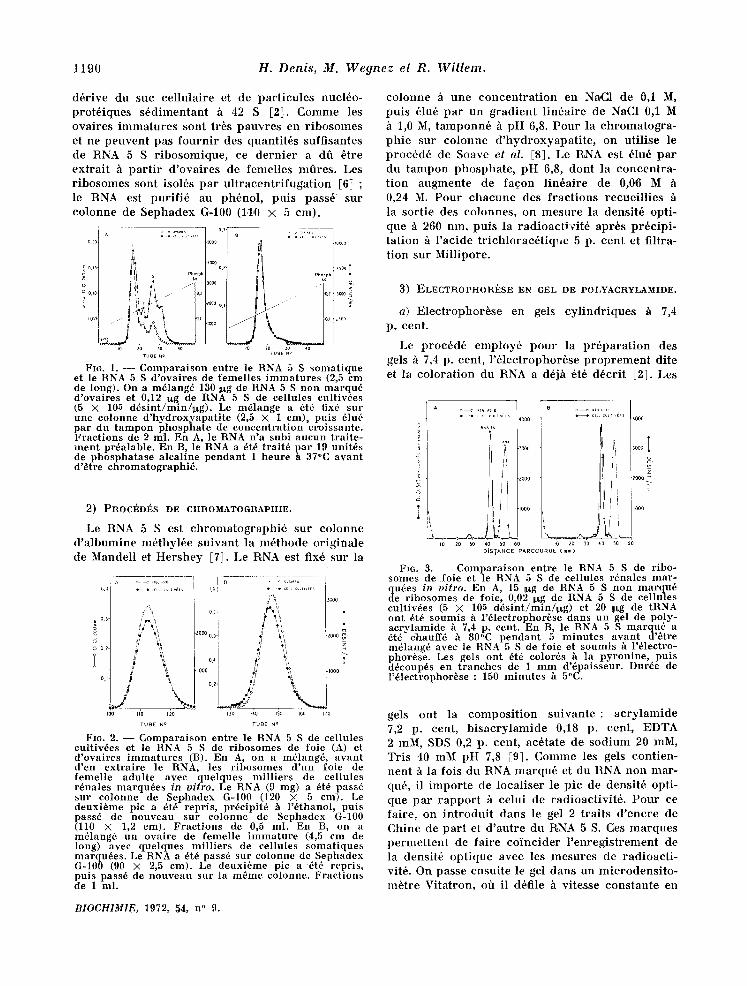
FI6. 1. - - - Compara i son entre le RNA 5 S somat ique et le RNA 5 S d 'ovaires de femelles i m m a t u r e s (2,5 em de long). On a m6lang6 130 :lxg de RNA 5 S non marqu~ d 'ovaires et 0,12 ~xg de RNA 5 S de eellules eultiv6es (5 X 105 d~s in t /min /~g ) . Le m~lange a 6t6 fix6 sur une eolonne d 'hydroxyapa t i t e (2,5 X 1 crn), puis 61u6 par du t ampon phospha te de concen t ra t ion croissante. Frac t ions de 2 ml. En A, le RNA n 'a subi aucun t ra i te - men t pr6alable. En B, le RNA a 6t6 t ra i t6 pa r 19 unitds de phospha tase a leal ine pendan t 1 heure h 37°C avan t d 'e t re chromatographi6 .
2) PROCI~.D~S DE CHROMATOGRAPHIE.
Le RNA 5 S es t c h r o m a t o g r a p h i 6 s u r c o l o n n e d ' a l b u m i n e m 6 t h y l 6 e s u i v a n t la m 6 t h o d e o r i g i n a l e de M a n d e l l et H e r s h e y [7]. Le RNA est fix6 s u r la
~,1~ : : Y ? : ; . . . . . . . . . . ' ° ..........
I ,?\ .... 02 !
z
c o l o n n e ~ u n e c o n c e n t r a t i o n e n Na.Gl de 0,1 M, p u t s 61u6 p a r u n g r a d i e n t l i n 6 a i r e de NaC1 0,1 M
1,0 M, t a m p o n n 6 h p H 6,8. P o u r la c h r o m a t o g r a - p h i e su r c o l o n n e d ' h y d r o x y a p a t i t e , o n u t i l i s e le p r o c 6 d 6 de Soave et al. [8]. Le RNA est 61u6 p a r d u t a m p o n p h o s p h a t e , p H 6,8, d o n t la c o n c e n t r a - t i o n a u g m e n t e de f a c o n l i n 6 a i r e de 0,06 M 0,24 M. P o u r c h a c u n e des f r a c t i o n s r e c u e i l l i e s h la s o r t i e des c o l o n n e s , o n m e s u r e la d e n s i t 6 op t i - que h 260 nm, p u t s l a r a d i o a c t i v i t 6 a p r 6 s p r 6 c i p i - t a t i o n h l ' a c i d e t r i c h l o r a c 6 t i q , , e 5 p. c e n t e t f i l t r a - t i o n s u r Mi l l i po re .
3) ELECTROPHORESE E.N GEL DE POLYACRYLAMIDE.
a) E l e c t r o p h o r ~ s e e n gels c y l i n d r i q u e s h 7,4 p. c en t .
Le p r o c 6 d 6 e m p l o y 6 p o u r la p r 6 p a r a t i o n des gels "h 7,4 p. cen t , l ' 6 1 e c t r o p h o r 6 s e p r o p r e m e n t d i t e et la c o l o r a t i o n du RNA a d6 jh 6t6 d 6 c r i t [2]. Les
A
i
6
d
Io ~o 50 40 50
DIS
4000
5000
2000
I000
,ooo!
~ooo ~
60 ~0 20 30 40 50 60 ~ N C E PARCOUF~UE Cmm)
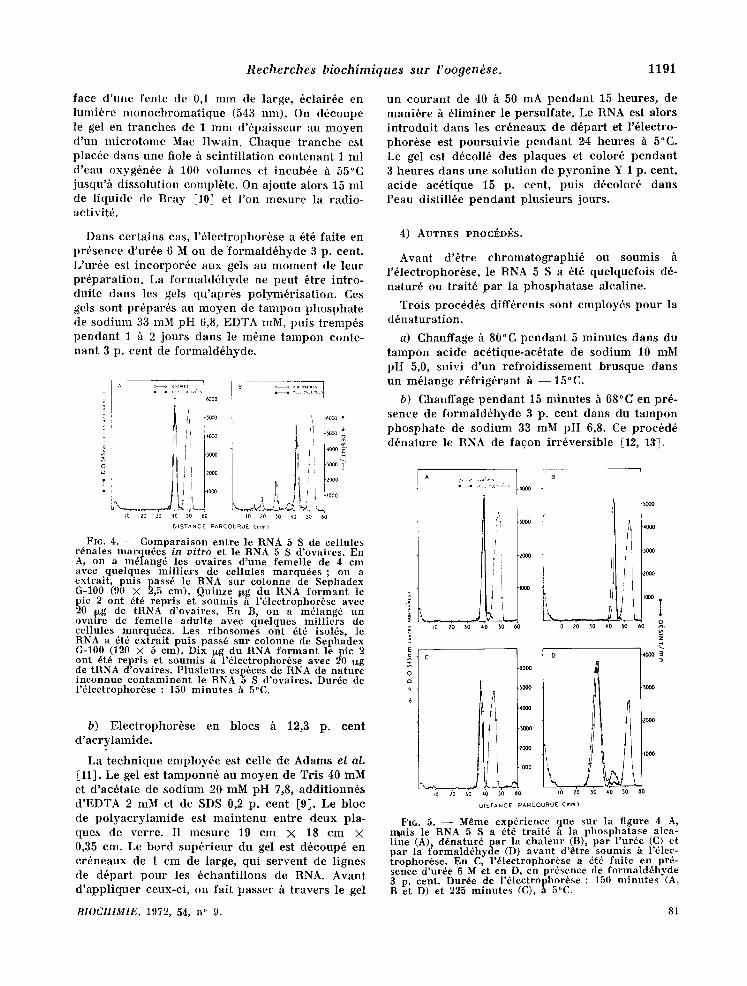
Fro. 3. - - Compara l son entre le RNA 5 S de r ibo- somes de fo ie et le RNA 5 S de eellules r6nales mar - qu6es in vitro. En A, 15 I~g de RNA 5 S non marqu6 de r ibosomes de foie, 0,02 10g de RNA 5 S de cellules cultiv6es (5 × 105 d6sint/min/,~xg) et 20 ~xg de tRNA out 6t6 soumis h l '61ectrophor6se dans un gel de poly- acry lamide h 7,4 p. cent. En B, le RNA 5 S marqu6 a 6t6 chauff6 h 80°C pendan t 5 minu tes avan t d 'dtre m61ang6 avee le RNA 5 S de foie et soumis h l'61ectro- phorSse. Les gels ont 6td color6s h la pyronine , puts d6coup6s en t ranches de 1 m m d'6paisseur. Dur~e de l '6lectrophor6se : 150 minu te s h 5°C.
100 II0 120 150 ~40 150 160 I?0
TUBE N ° TUBE N °
Fro. 2. - - Compara i son entre le RNA 5 S de cellules cultiv6es et le RNA 5 S de r ibosomes de foie (A) et d 'ovaires immatu re s (B). En A, on a m61ang6, avan t d 'en ext ra i re le RNA, les r ibosomes d 'un foie de femelle adul te avec quelques mi l l ie rs de eellules r~nales marqu6es in vitro. Le RNA (9 mg) a ~t~ pass6 sur colonne de Sephadcx G-100 (120 X 5 cm). Le deuxi6me p i c a ~t~ repris, pr6cipit~ h l '6 thanol , puts pass~ de nouveau sur colonne de Sephadex G-100 (110 × 1,2 cm). Frac t ions de 0,5 ml. En B, on a m~lang~ un ovaire de femelle i m m a t u r e (4,5 cm de long) avec quelqucs mil l iers de cellules somat iques marquees . Le RNA a 8t6 pass6 sur colonne de Sephadex G-100 (90 × 2,5 cm). Le deuxi6me p i c a ~t~ repris, puts pass~ de nouveau sur la mSme colonne. Fract ions de 1 ml.
BIOCH!M1E, 1972, 54, n ° 9.
gels o n t la c o m p o s i t i o n s u i v a n t e : a c r y l a m i d e 7,2 p. cen t , b i s a c r y l a m i d e 0,18 p. c en t , E!DTA 2 raM, SDS 0,2 p. cen t , a c6 t a t e de s o d i u m 20 raM, T r i s 40 mM p H 7,8 [9]. C o m m e les gels c o n t i e n - n e n t h la fo is d u RNA m a r q u 6 et d u RNA n o n m a r - qu6, il i m p o r t e de l o c a l i s e r le p i c de d e n s i t 6 op t i - que p a r r a p p o r t h ce lu i de r a d i o a c t i v i t 6 . P o u r ce fa i re , o n i n t r o d u i t d a n s le gel 2 t r a i t s d ' e n c r e de C h i n e de p a r t e t d ' a u t r e d u RNA 5 S. Ces m a r q u e s p e r m e t t e n t de f a i r e c o i n c i d e r l ' e n r e g i s t r e m e n t de la d e n s i t 6 o p t i q u e a v e c les m e s u r e s de r a d i o a c t i - vi t6. On p a s s e e n s u i t e le gel d a n s u n m i c r o d e n s i t o - m 6 t r e V i t a t r o n , off il d6,file ~ v i t e s se c o n s t a n t e e n
R e c h e r c h e s b i o c h i m i q u e s s u r l 'oogendse. 1191
face d ' u n e f e n t e de 0,1 m m de l a rge , 6c l a i r6e e n l u m i 6 r e m o n o c h r o m a t i q u e (543 n m ) . On d 6 c o u p e le gel en t r a n c h e s de 1 m m d ' 6 p a i s s e u r au m o y e n d ' u n m i c r o t o m e Mac I l w a i n . C h a q u e t r a n c h e es t p l a c 6 c d a n s u n e f iole h s c i n t i l l a t i o n c o n t e n a n t 1 m l d ' e a u o x y g 6 n 6 e h 100 v o l u m e s et i n c u b 6 e h 55°C jusqu"h d i s s o l u t i o n c o m p l b t e . On a jou te a lo r s 15 m l de l i q u i d e de B r a y [10] et l ' o n m e s u r e la r a d i o - ac t iv i t6 .
D a n s c e r t a i n s cas, l ' 6 1 e c t r o p h o r b s e a 6t6 f a i t e e n p r 6 s e n c e d ' u r 6 e 6 M ou de f o r m a l d 6 h y d e 3 p. cen t . L ' u r 6 e esl i n c o r p o r 6 e aux gels au m o m e n t de l e u r p r 6 p a r a t i o n . La f o r m a l d 6 h y d e n e p e u t 6 t re i n t r o - d n i t e d u n s les gels q u ' a p r b s p o l y m 6 r i s a t i o n . Ces gels s o n t p r 6 p a r 6 s au m o y e n de t a m p o n p h o s p h a t e de s o d i u m 33 mM p H 6,8, E D T A raM, p u t s t r e m p 6 s p e n d a n t 1 "h 2 j o u r s d a n s le m 6 m e t a m p o n c o n t e - n a n t 3 p. c e n t de f o r m a l d 6 h y d e .
CELL CULTiVEE~
5O0O 6000 '~
~ 4000 5000
{ 4ooo ~" 30OO -~
? I I .~ooo
1000 i .1000 ,I
10 ~0 50 40 50 60 ~0 ~0 30 40 50 50
DISTANCE PA~COU~UE (mm)
FIG. 4. - - Compara ison ent re le RNA 5 S de cellules r6nales marqu6es in vitro et le RNA 5 S d 'ovaires. En A, on a m61ang6 les ovaires d 'une femelle de 4 cm avec quelques mi l l ie rs de cellules marqudes ; on a extrai t , puts pass6 le RNA sur colonne de Sephadex G-100 (90 × 2,5 cm). Quinze ~g du RNA f o r m a n t le pic 2 ont 6t6 repr is et soumis h l '61ectrophor6se avec 20 .~g de tRNA d'ovaires. En B, on a mdlang6 un ova~re de femelle adul te avec quelques mi l l ie rs de cellules marqudes. Les r ibosomes ont 6t6 isol6s, le RNA a 6t6 ex t ra i t puts pass6 sur colonne de Sephadex G-100 (120 X 5 em). Dix ~o.g du RNA f o r m a n t le pic 2 ont 6t6 repr is et soumis ~ l '61eetrophor6se avec 20 ug de tRNA d'ovaires. P lus ieurs esp6ces de RNA de na ture inconnue con t amin en t le RNA 5 S d 'ovaires. Dur6e de l '61ectrophordse : 150 minu te s h 5°C.
b) E l e c t r o p h o r b s e e n b l o c s h 12,3 p. c e n t d'aclD, l a m i d e .
La t e c h n i q u e e m p l o y 6 e es t ce l le de A d a m s et al. [11]. Le gel est t a m p o n n 6 a u m o y e n de T r i s 40 m M et d ' a c 6 t a t e de s o d i u m 2,0 m M p H 7,8, a d d i t i o n n 6 s d ' E D T A 2 mM et de SDS 0,2 p. c e n t [9]. Le b l o c de p o l y a c r y l a m i d e est m a i n t e n u e n t r e d e u x p la - ques de v e r r e . I1 m e s u r e 19 c m × 18 c m X 0,35 cm. Le b o r d s u p 6 r i e u r d u gel es t d 6 c o u p 6 e n c r 6 n e a u x de 1 c m de l a rge , q u i s e r v e n t de l i g n e s de d 6 p a r t p o u r les 6 c h a n t i l l o n s de RNA. A v a n t d ' a p p l i q u e r ceux-c i , o n fa i r p a s s e r h t r a v e r s le gel
BIOCHIMIE, 1972, 54, n ° 9.
u n c o u r a n t de 40 /t 50 m A p e n d a n t 15 h e u r e s , de m a n i 6 r e h 61 t ra iner le p e r s u l f a t e . Le RNA est a l o r s i n t r o d u i t d u n s les c r 6 n e a u x de d 6 p a r t et l '61ect ro- p h o r 6 s e es t p o u r s u i v i e p e n d a n t 24 h e u r e s h 5°C. Le gel es t d6co l l6 des p l a q u e s et co lo r6 p e n d a n t 3 h e u r e s d a n s u n e s o l u t i o n de p y r o n i n e Y 1 p. c en t , a c i d e a c 6 t i q u e 15 p. cen t , p u t s d 6 c o l o r 6 d a n s l ' e a u d i s t i l l 6e p e n d a n t p l u s i e u r s j ou r s .
4) AUTRES PROC~DI~S.
A v a n t d ' 6 t r e c h r o m a t o g r a p h i 6 ou s o u m i s h l ' 6 1 e c t r o p h o r b s e , le RNA 5 S a 6t6 q u e l q u e f o i s d6- n a t u r 6 ou t r a i t 6 p a r la p h o s p h a t a s e a l ea l i ne .
T r o i s p r o e 6 d 6 s d i f f 6 r e n t s s o n t e m p l o y 6 s p o u r ]a d 6 n a t u r a t i o n .
a) C h a n f f a g e h 80°G p e n d a n t 5 m i n u t e s d a n s d u t a m p o n a c i d e a c 6 t i q u e - a c 6 t a t e de s o d i u m 10 mM p H 5,0, s u i v i d ' n n r e f r o i d i s s e m e n t b r u s q u e d a n s u n m d l a n g e r 6 f r i g d r a n t h - - 1 5 ° C .
b) C h a u f f a g e p e n d a n t 15 m i n u t e s h 68°C e n p r6 - s e n c e de f o r m a l d 6 h y d e 3 p. c e n t d a n s d u t a m p o n p h o s p h a t e de s o d i u m 33 m M p H 6,8. Ce p r o c 6 d 6 d 6 n a t u r e le RNA de f a c o n i r r 6 v e r s i b l e [12, 13].
; j
8 i
I I
¸4000
¸3OOO
i .~000
ICO0
10 20 30 40 50 60 ~ ,
z
• ~
600O
5000 • .~
• 2
l0 20 30 60 10 20 50 40 50 60
DISTANCE OARCOURUE (.mm)
D j4000 3
,1
I I
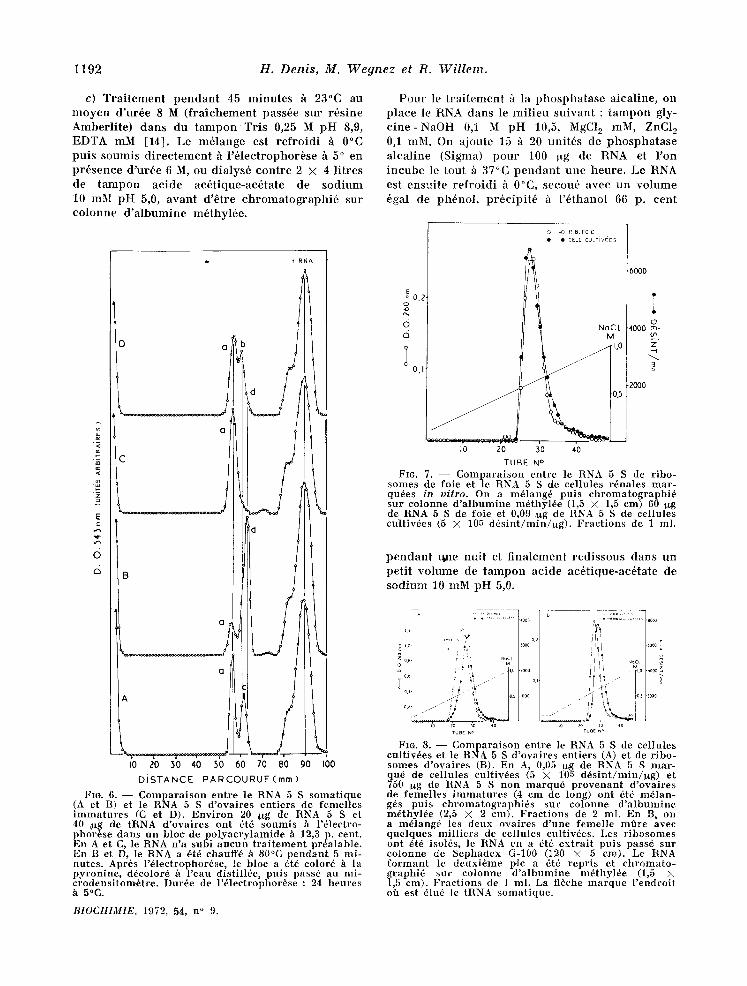
Fro. 5. - - M~me exp6rience que sur la figure 4 A, mats le RNA 5 S a 6t6 t ra i t6 h la phospha tase alca- l ine (A), d6natur6 p a r la cha leur (B), par l 'ur6e (C) et pa r la formald6hyde (D) avan t d '6tre soumis /t l'61ec- t rophor6se. En C, l '61ectrophor6se a 6t6 fai te en pr6- sence d 'ur6e 6 M e t en D, en pr6sence de fo rmaldehyde 3 cent. Dur6e de l 'dlectrophor6se : 150 minu tes (A, B Pt D) et 225 minu tes (C), h 5°C.
8 1
1192 H. Denis, M. Wegnez et R. Willem.
c) Traitement pendant 45 minutes h 23°C au m o y e n d'ur6e 8 M (fraiehement pass6e sur r6sine Amberlite) dans du tampon Tris 0,25 M pH 8,9, EDTA mM [14]. Le m61ange est refroidi h 0°C puis soumis d irectement h l'61ectrophorbse h 5 ° en pr6sence d'ur6e 6 M, ou dialys6 contre 2 X 4 l itres de tampon acide ac6tique-ac6tate de sod ium 10 mM pH 5,0, avant d'6tre chromatographi6 sur colonne d'albumine m6thy16e.
z
E
6 d
Pour le t ra i tement '~ la phosphatase aicaline, on place le RNA dans le mi l ieu sn ivant : t ampon gly- c i n e - N a O H 0,1 M pH 10,5, MgC12 raM, ZnCI2 0,1 mM. On ajoute 15 h 20 unit6s de phosphatase alcal ine (Sigma) pour 100 ~tg de RNA et l 'on incube le tout fi 37°C pe nda n t une heure. Le RNA est ensuite refroidi h 0°C, secou6 avec un volume 6gal de ph6nol, pr6cipit6 "h l '6thanol 66 p. cent
I ~ Ria. FO[E I I~ l l CELLCULT[VEES
E ~0,2'
g r~ o c5
0,1-
10 20 50 40
TUBE N °
6000
0 NaCl 4000 ra-
M w. 1,0 z
2000 0,5
FIG. 7. - - C o m p a r a i s o n ent re le RNA 5 S de r ibo- somes de foie et le RNA 5 S de eellules r6nales m a r - qn~es in oitro. On a mdlang6 pu i s e h r o m a t o g r a p h i ~ su r eo lonne d ' a l b u m i n e mdthyl~e (1,5 X 1,5 em) 60 ~g de RNA 5 S de foie et 0,09 lug de RNA 5 S de eellnles cultiv~es (5 × 105 d ~ s i n t / m i n / ~ g ) . F rac t ions de 1 ml.
pendan t t~ne nui t et f inalement redissous duns un petit volume de tampon aeide ae~tique-ae6tate de sodium 10 mM pH 5,0.
o ~-
I0 20 30 40 50 60 70 80 90 I00
DISTANCE PARCOURUE (mm)
Fig. 6. - - C o m p a r a i s o n ent re le RNA 5 S s o m a t i q u e ( A e t B) et le RNA 5 S d 'ova i res en t ie rs de femel les i m m a t u r e s (C et D). E n v i r o n 20 ~g de RNA 5 S e t 40 !ttg de tRNA d 'ova i res ont 6t6 soumis h l 'dlectro- phor6se dans un bloc de p o l y a c r y l a m i d e h 12,3 p. cent. En A et C, le RNA n 'a subi a u c u n t r a i t e m e n t pr6alable . En B e t D, le RNA a 6t~ ehauff~ h 80°C p e n d a n t 5 mi- nutes . Apr6s l 'd leetrophor6se, le bloe a 6t6 colord h la py ron ine , ddeolor~ h l 'eau distillde, pu i s pass6 au mi- crodensitom,~tre. Dur6e de l 'd lec t rophor6se : 24 heures h 5°C.
BIOCH1MIE, 1972, 54, n ° 9.
FIG. 8. - - C o m p a r a i s o n ent re ]e RNA 5 S de cel]ules cult iv6es et le RNA 5 S d 'ova i res en t ie rs (A) et de r ibo- somes d 'ova i res (B). En A, 0,05 ~g de RNA 5 S m a r - qu6 de eel lules cult iv6es (5 X 105 d6s in t /m in /ug ) et 750 ug de RNA 5 S n o n m a r q u 6 p r o v e n a n t d 'ova i res de femel les i m m a t u r e s (4 cm de long) on t 6t6 m61an- g6s puis ch ro ina tog raph i6s su r colonne d ' a l b u m i n e m6thyl6e (2,5 X 2 cm). F rac t ions de 2 ml. En B, on a m61ang6 les deux ovaires d 'une femelle mfire avec que lques mi l l i e r s de eellules cultiv6es. Les r i b o s o m e s ou t ~t~ isol~s, le RNA en a ~t6 ex t r a i t pu i s pass~ s u r co lonne de Sephadex G-100 (120 × 5 era). Le RNA f o r m a n t le deuxi~me pie a dt6 repr i s et c h r o m a t o - graphi(~ su r colonne d ' a l b u m i n e mdthyl~e (1,5 × 1,5 em). F rac t ions de l ml. La fl~che m a r q u e l ' endroi t off est 61u6 le tRNA somat ique .
Recherches biochimiques sur l'oogen~se. 1193
Pour la d6terminat ion du profil d ' hype reh ro - micit6, le RNA est dissous dans 2,5 ml de 1 X SSC, reeouver t d 'une couehe d 'hui le min6rale pour 6vi- ter toute 6vaporat ion, puts chauff6 jusqu'h 85 ° dans le disposi t i f thermosta t ique du spectrophoto- m~tre Zeiss PMQ H. L 'accro i ssement de temp6- ra ture est de I °C par minute. Aucune cor rec t ion n 'a 0 6 apport6e pour compenser la di lut ion du RNA due h l ' expans ion du solvant.
RESULTATS.
L 'hydroxyapa t i t e s6pare le RNA 5 S e n 3 pies de hauteur in6gale (fig. 1 A). Plus de la moiti6 du RNA 5 S somat ique est 61u6e au niveau des pies 2 et 3, mats seulement le quart du RNA 5 S d 'ovaires. Un t ra i tement pr6alable h la phosphatase alcal ine
o:0 I ...... .....
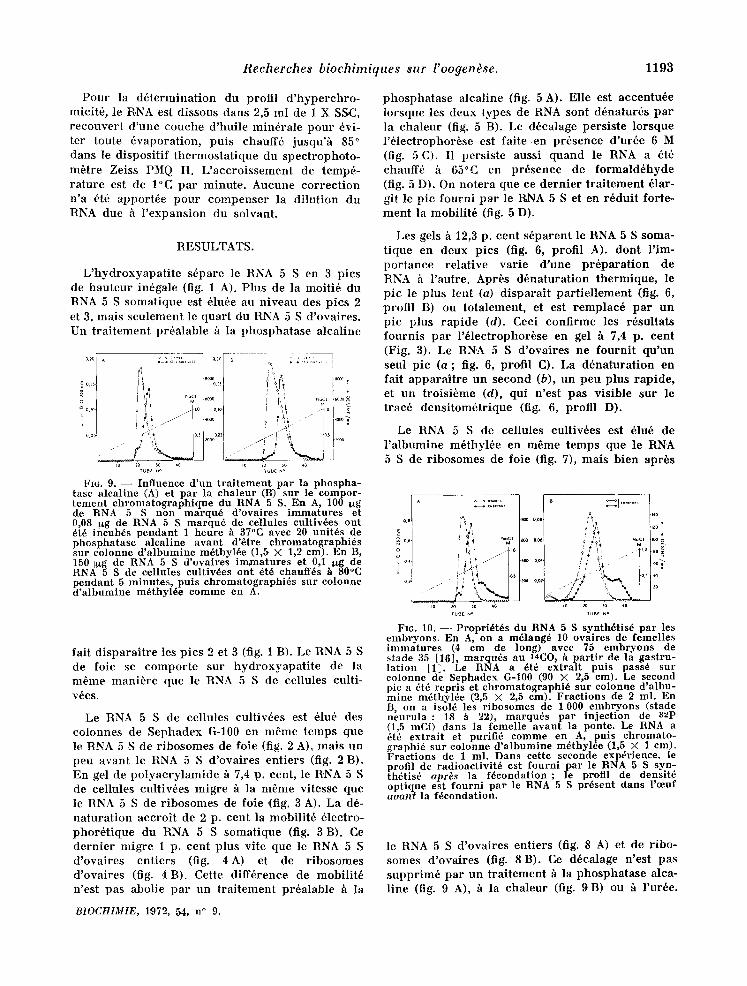
io to ~o 40
Fro. 9. - - Influence d'un traitement par la phospha- tase alcaline (A) et par la chaleur (B) sur le compor- tement ehromatographique du RNA 5 S. En A, 100 ug de RNA 5 S non marqu6 d'ovaires immatures et 0,08 I~g de RNA 5 S marqu6 de cellules cultiv6es oat 6t6 incub6s pendant 1 heure h 37°C avec 20 unit6s de phosphatase alcaline avant d'etre chromatographi6s snr colonne d'albumine m6thyl6e (1,5 × 1,2 cm). En B, 150 I~g de RNA 5 S d'ovaires imxtaatures et 0,1 ltg de RNA 5 S de c ellules eultiv6es ont 6t6 chauff6s h 80°C pendant 5 minutes, puts chromatographi~s sur colonne d'albumine m6thyl6e comme en A.
fair d ispara i t re les pies 2 et 3 (fig. 1 B). Le RNA 5 S de foie se compor te sur hydroxyapa t i t e de la m6me mani+re que le RNA 5 S de cellules culti- v6es.
Le RNA 5 S de cellules cultiv6es est 61u6 des colonnes de Sephadex G-100 en m6me temps que le RNA 5 S de r ibosomes de foie (fig. 2 A), mats un peu avant le RNA 5 S d 'ovai res ent iers (fig. 2 B). En gel de po lyac ry l amide h 7,4 p. cent, le RNA 5 S de cellules cultiv6es migre h la m6me vitesse que le RNA 5 S de r ibosomes de foie ,(fig. 3 A). La d6- natura t ion accroi t de 2 p. cent la mobil i t6 61ectro- phor6t ique du RNA 5 S somat ique (fig. 3 B). Ce dern ie r migre 1 p. cent plus vite que le RNA 5 S d 'ovai res ent iers (fig. 4A) et de r ibosomes d 'ovai res (fig. 4 B). Cette diff6rence de mobili t6 n 'est pas abolie par un t ra i tement pr6alable h la
phosphatase alcal ine (fig. 5 A). Elle est accentu6e lorsque les deux types de RNA sont d6nalur6s par la chaleur (fig. 5 B). Le d6ealage persis te lorsque l '61ectrophorbse est faite e n pr6sence d 'ur6e 6 M (fig. 5 C). II persis te aussi quand le RNA a 6t6 chauff6 h 65°C en pr6senee de formald6hyde (fig. 5 D). On notera que ce dern ie r t ra i tement 61ar- git le p ic fourni pa r le RNA 5 S e t en r6duit forte- ment la mobili t6 (fig. 5 D).
Les gels ~ 12,3 p. cent s6parent le I~NA 5 S soma- t ique en deux pies (fig. 6, pro411 A). dont l ' im- por t ance relat ive var ie d 'une pr6para t ion de RNA h l 'autre. Apr~s d6naturat ion thermique , le p ic le plus lent (a) d ispara l t pa r t i e l l ement (fig. 6, profil B) ou totalement , et est remplac6 pa r un p ic plus rap ide (d). Ceci confi rme les r6sultats fournis pa r l '61ectrophorbse en gel h 7,4 p. cent (Fig. 3). Le RNA 5 S d 'ovai res ne fourni t qu 'un seul p ie ( a ; fig. 6, prafil C). La d6naturat ion en fair appara l t re un second (b), un peu plus rap ide , et un t rois ibme (d), qui n'est pas vis ible sur le trac6 dens i tom6tr ique (fig. 6, profil D).
Le RNA 5 S de cellules cultiv6es est 61u6 de l ' a lbumine m6thyl6e en m6me temps que le RNA 5 S de r ibosomes de foie (fig. 7), mats bien apr6s
H ~ ' . . . . .
o0 ,' i!'. ~[,.0
~UBE N °
140
i~ '°I , ,
4 i"" . . . . . . 7
io ~o ~o 4 0
Tua r NO
Fro. 10. - - Propri6t6s du RNA 5 S synth6tis6 par les embryons. En A, on a m61ang6 10 ovaires de femelles immatures (4 cm de long) avec 75 embryons de stade 35 [IS], marquis an ~4CO~ h I?artir de la gastru- lation Ell. Le RNA a 6t6 extra]t puts pass6 sur colonne de Sephadex G-100 (90)< 2,5 cm). Le second pie a 6t6 repris et chromatographi6 sur eolonne d'albu- mine m6thyl6e (2,5 × 2,5 era). Fractions de 2 ml. En B, on a isol6 les ribosomes de 1 000 embryons (stade neurula : 18 h 22), marqu6s par injection de 32p (1,5 mCi) dans la femelle avant la ponte. Le RNA a 6t6 extrait et purifi6 comme en A, puts chromato- graphi6 sur colonne d'albumine m6thyl6e (1,5 X 1 cm). Fractions de 1 ml. Dans cette seeonde exp6rienee, le profil de radioactivit6 est fourni par le RNA 5 S syn- th6tis6 apr~s la f6condation; le profit de densit6 optique est fourni par le RNA 5 S pr6sent dans l'ceuf avant la f6condation.
le RNA 5 S d 'ovai res entiers (fig. 8 A) et de r ibo- somes d 'ovai res (fig. 8 B). Ce d6ealage n'est pas suppr im6 par un t ra i tement h la phosphatase alea- l ine (fig. 9 A), h la chaleur (fig. 9B) ou h l 'ur6e.
BIOCHIMIE, 1972, 54, n ° 9.
1194 H. Denis, M. Wegnez el R. Willem.
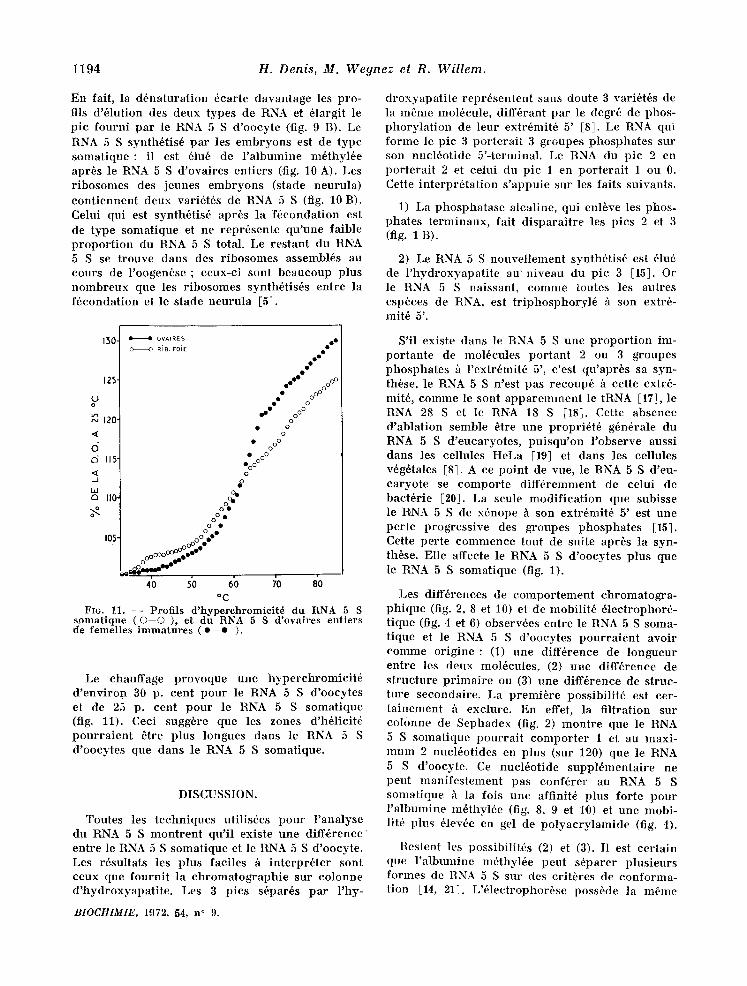
En fait, la d6natura t ion 6carte davantage les pro- ills d'61ution des deux types de RNA et 61argit le pic fourni par le R~NA 5 S d 'oocyte (fig. 9 B). Le RNA 5 S synth6tis6 par les e lnbryons est de type so lna t ique : il est 61u6 de l ' a lbumine m~thyl6e apr6s le RNA 5 S d 'ovaires entiers (fig. 10 A). Les r ibosomes des jeunes embryons (stade neurula) con t i ennen t deux vari6t6s de RNA 5 S (fig. 10 B). Celui qui est synth6tis6 apr6s la f6condat ion est de type somatique et ne repr6sente qu 'une faible p ropor t ion du RNA 5. S total. Le restant du RN~A 5 S se trouve dans des r ibosomes asselnbl6s au cours de l 'oogen6se ; ceux-ci sont beaucoup plus nombreux que les r ibosomes synth6tis6s entre la f6condation et le stade neuru la [5].
130-
125-
O o
120- <
d ,N 115" < J kd a 110-
~ OVAiRES
C ~ RiB. FOiE ~OOQ IQQ
: '" oooO°°° e $ O ..- ooO
• 0 0
• 0 oo
• oo %0 ° 0 .o
o°° 0 •
ms ooO°O ** oooO~_°... "
o o ° ,,,
40 50 6'0 o C
~0 8'0
FrG. 11. - - P r o f i l s d ' h y p e r c h r o m i c i t 6 d u R N A 5 S s o m a t i q u e ( O - - O ) , e t d u R N A 5 S d ' o v a i r e s e n t i e r s de f e m e l l e s i m m a t u r e s ( e - • ).
Le chauffage provoque une hyperchromic i t6 d ' env i ron 30 p. cent pour le RNA 5 S d'oocytes et de 25 p. cent pour le RNA 5 S somatique (fig. 11). Ceci sugg6re que les zones d'h61icit6 pour ra i en t 6tre plus longues dans le RNA 5 S d'oocytes que dans le RNA 5 S solnatique.
DISCUSSION.
Toutes les techniques utilis6es pour l 'analyse du RNA 5 S mon t ren t qu ' i l existe une d i f f6 rence entre le RNA 5 S somatique et le RNA 5 S d'oocyte. Les r6sultats les plus faciles ~ in terpr6ter sont eeux que fourni t la chromatographie sur colonne d 'hydroxyapat i te . Les 3 pics s6par6s par l 'hy-
droxyapat i te repr6sentent sans doute 3 vari6t6s de la m6me mol6cule, diff6rant par le degr6 de phos- phory la t ion de leur extr61nit6 5' [8]. Le RNA qui forlne le pic 3 por tera i t 3 groupes phosphates sur son nucl6otide 5 '- terminal. Le !~NA du pic 2 en porterai t 2 et celui du pic 1 en por tera i t 1 ou 0. Cette in terpr6ta t ion s 'appuie sur les faits suivants.
1) La phosphatase alcaline, qui enlbve les phos- phates ter lninaux, fait d isparal t re les pics 2 et 3 (fig. 1 B).
2) Le RNA 5 S nouvel le lnent synth6tis6 est 61u6 de l 'hydroxyapat i te a u niveau du pic 3 E15]. Or le RNA 5 S naissant, comnle routes les autres espbces de RNA, est tr iphosphory16 fi son extr6- 1nit6 5'.
S'il existe dans le RNA 5 S une propor t ion iln- por tante de 1nol6cules por tan t 2 ou 3 groupes phosphates "h l 'extr6mit6 5', c'est qu 'aprbs sa syn- th~se, le RNA 5 S n 'est pas recoup6 "h cettc extr6- mit~, colnme le sont apparemlnent le tRNA [17], le RNA 28 S et Ie RNA 18 S [183. Cette absence d 'abla t ion selnble 6tre une propri6t6 g6n6rale du RNA 5 S d 'eucaryotes, pu i squ 'on l 'observe aussi clans ]es cellules HeLa [19] et dans les cellules v6g6tales [8]. A ce point de vue, le RNA 5 S d'eu- caryote se comporte diff6relnlnent de celui de bact6rie [20]. La seule modif ica t ion que subisse le R~NA 5 S de x6nope ~ son extr6mit6 5' est une perte progressive des groupes phosphates [15]. Cette perte commence tout de suite aprbs la syn- lh6se. Elle affecte le RNA 5 S d'oocytes plus que le RNA 5 S somatique (fig. 1).
Les diff6rences de compor tement chromatogra- phique (fig. 2, 8 et 10) et de 1nobilit6 61ectrophor6- t ique (fig. 4 et 6) observ6es entre le RNA 5 S solna- tique et le RNA 5 S d'oocytes pour ra ien t avoir colnlne origine : (1) une diff6rence de longueur entre les deux mol6cules, (2) une diff6rence de s t ructure pr i lna i re ou (3) une diff6rence de struc- ture secondaire. La prelni6re possibili t6 est cer- ta ine lnent ix exclure. En effet, la fi l tration sur col0nne de Sephadex (fig. 2) mont re que le RNA 5 S somatique pour ra i t colnporter 1 et au maxi- 1nuln 2 nucl6otides en plus (sur 120) que le RNA 5 S d'oocyte. Ce nucl6otide supp161nentaire ne peut 1nanifestelnent pas conf6rer au RNA 5 S solnatique h la fois une affinit6 plus forte pour l ' a lbuinine In6thyl6e (fig. 8, 9 et 1~0) et une mobi- lit6 plus 61ev6e en gel de polyacryla ln ide (fig. 4).
Restent les possibilit6s (2) et (3). II est cer ta in que l ' a lbumine 1n6thyl6e peot s~parer plusieurs forlnes de RNA 5 S sur des crit6res de conforlna- l i o n [14, 21 ] . L'61eetrophor6se possbde la 1n~1ne
BIOCHIM1E, 1~972, 54, n ° 9.

R e c h e r c h e s b i o c h i m i q u e s sur l'oogenOse. 1195
p r o p r i 6 t 6 p u i s q u e les d e u x f o r m e s de RNA 5 S s o m a t i q u e s6par6es p a r les gels c o n c e n t r 6 s (fig. 6 A) on t la m 6 m e s t r u c t u r e p r i m a i r e [22]. On est tent6 d ' e n d 6 d u i r e que les d i v e r s e s var i6 t6s de RNA 5 S, et en p a r t i c u l i e r le RNA 5 S s o m a t i q u e et le RNA 5 S d ' o o c y t e s , ne d i f f6 r en t l ' u n e de l ' a u t r e que p a r l eu r s t r u c t u r e s e e o n d a i r e . Cette con c l u - s ion est c o m p a t i b l e avec les r6su l t a t s des exp6- r i e n c e s de d 6 n a t u r a t i o n t h e r m i q u e (fig. 11). Toute - tots , le RNA 5 S s o m a t i q u e c o n t i n u e h m i g r e r en gel de p o l y a c r y l a m i d e un p e u p lus vi te que le RNA 5 S d ' o o e y t e s , d a n s des c o n d i t i o n s qui s o n t cens6es a b o l i r rou te s t r u c t u r e s e c o n d a i r e (fig. 5 C et D). P a r a i l l eurs , si les deux t y p e s de RNA poss6- d a i e n t la rn6me s t r u c t u r e p r i m a i r e , on p o u r r a i t s ' a t t e n d r e h ce qu ' i l s a d o p t e n t , ap r6s d 6 n a t u r a t i o n , une c o n f i g u r a t i o n i d e n t i q u e . Or, il n ' e n est r i e n : les p r o p r i 6 t 6 s du I~NA 5 S d ' o o c y t e s et du RNA 5 S s o m a t i q u e s o n t p lus d i f f6 r en t e s a p r b s la d 6 n a t u r a - t ion q u ' a v a n t (fig. 4 et 5 B, 6, 8 et 9 B). I1 est d o n c p o s s i b l e que le RNA 5 S d ' o o c y t e s et le RNA 5 S s o m a t i q u e d i f f b r e n t h la fo is p a r l eu r s t r u c t u r e p r i m a i r e et p a r l eu r s t r u c t u r e s e c o n d a i r e . P o u r p r o u v e r qu ' i l en est b i en a ins i , il f a u d r a 6 t ab l i r la s 6 q u e n c e nuc16ot id ique c o m p l O e des d e u x mol6- cules . Ce t r ava i l es t en c o u r s ~ l ' h e u r e actuel le .
Remerciements.
M. Wegnez est boursier de l ' Ins t i tu t pour l 'Encou- ragement de la Recherche Scientiflque dans l ' Industr ie et l 'Agriculture. Ce travail a 6td rendu possible par l 'aide finanei6re du Fonds National de la Recherche Scientifique. Nous remercions le Professeur R. Monier d 'avoir bien voulu lire et er i t iquer le manuscri t .
R~suxi~.
Le RNA 5 S des oocytes de Xenopus laeois diffSre du RNA 5 S somatique par le degr6 de phosphoryla t ion de son extr6mit6 5' : le RNA 5 S somatique contient une plus forte proport ion de mol6cules por tant 2 ou 3 groupes phosphates sur leur nucl~otide 5 '- terminal .
Le RNA 5 S somatique est re tards par le Sephadex G-100 un peu moins que le RNA 5 S d'oocytes. Le RNA 5 S provenant de r ibosomes somatiques est 61u6 de l ' a lbumine mSthyl6e bien apr6s le RNA 5 S d'oocytes entiers et de r ibosomes d'oocytes. Les ribo- somes des jeunes embryons eont iennent deux types de RNA 5 S. Celui qui est synth~tis~ apr~s la f6con- dation est de type somatique et ne repr6sente qu 'une faible propor t ion du RNA 5 S total. Le res tant du RNA 5 S embryonnai re se trouve dans des r ibosomes assemblds au cours de l'oogen6se.
Le RNA 5 S somatique migre dans les gels de poly- acrylamide h 7,4 p. cent un peu plus r i te que le RNA 5 S d'oocytes. Ni l 'enl6vement des groupes phos- phates te rminaux au moyen de la phosphatase alca- line, ni la d6naturat ion par la ehaleur ou l 'ur6e ne suppr iment cette diff6rence de mobilit6. L'61eetropho- r6se en gel de polyacrylamide h 12,3 p. cent rdv~le l 'existence de deux formes distinctes de RNA 5 S somatique. L'une d 'entre elles a la m6me mobilit6 que
le RNA 5 S d'oocytes. L'autre forme migre plus vite. Le RNA 5 S d'oocytes et le RNA 5 S somatique rdagissent diffdremment h la d6naturat ion thermique : la plus grande pat t ie du RNA 5 S d'oocytes conserve, apr6s refroidissement , la m6me mobilit6 qu 'avant , tandis que le RNA 5 S somatique adopte une forme nouvelle, qui migre plus vite que toutes les autres.
Quand on le chauffe en solution saline, le RNA 5 S d'oocytes prdsente une hyperchromiei t6 un peu plus forte que le RNA 5 S somatique.
Ces rdsultats sugg6rent que le RNA 5 S somatique et le RNA 5 S d'oocytes poss6dent /~ la fois une struc- ture pr imaire et une structure secondaire diff6rentes.
ZUSAMMENFASSUNG.
Die RNS der Oozyten des Xenopus laevis unterschei- det sieh v o n d e r somatischen 5 S RNS durch den Phos- phorylat ionsgrad ihres 5' Endes : die somatisebe 5 S RNS enthhl t einen st/irkeren Anteil yon Molekiilen, die 2 oder 3 Phosphatgruppen auf ih rem 5'-Endnu- kleotid tragen.
Die 5 S RNS der Oozyten wird durch Sephadex G-100 ein wenig st/irker zurfickgehalten als die soma- tische 5 S RNS. Die aus den somatischen Ribosomeu s tammende 5 S RNS wird aus e iner S~iule methyl ier ten Albumin - Kieselgurs wesentl ieh sp~iter eluiert als 5 S RNS der ganzen Oozyten und der Ribosomen der Oozyten. Die Ribosomen der jungen Embryos enthal ten zwei Typen der 5 S RNS. Die eine wird nach der Befruchtung synthet is ier t , sie ist vom somatischen Typ und stellt nur einen schwachen Anteil der gesamten 5 S RNS dar. Die iibrige 5 S RNS des Embryos befindet sich in den Ribosomen~ die sich zum Zeitpunkt der Oogenese verbinden.
Die somatische 5 S RNS wander t in Polyacrylamidgel bet 7,4 p. cent sehneller als die 5 S RNS der Oozyten. Diese elektrophoret ische Beweglichkeit wird weder durch Abtrennung der Phosphatendgruppen mit tels der alkalischen Phosphatase noch durch Denaturierung durch W/irme oder Harnstoff beeintr/ichtigt. Die Elek- t rophorese in Polyacrylainidgel bet 12,3 p. cent enthii l l t die Existenz zweier untersehiedl icher Formen der somatischen 5 S RNS. Die eine davon hat dieselbe Beweglichkeit wie die 5 S RNS der Oozyten. Die andere Form wander t wesent l ich schneller. Die 5 S RNS der Oozyten und die somatische 5 S RNS reagieren in verscbiedener VCeise auf thermische Dena- tur ierung : der gr6sste Tell der 5 S RNS der Oozyten beh~ilt nach Abkiihlung dieselbe Beweglichkeit wie zuvor w~ihrend die somatische 5 S RNS eine neue Form annimmt , die schneller als alle andereu wandert .
Bet Erw~irmung in Salzl6sung zeigt die 5 S RNS eine st/irkere Hyperchromizit~it als die somatische 5 S RNS.
Die Ergebnisse legen es nahe, dass die somatische 5 S RNS und die 5 S RNS der Oozyten zugleieh eine untersehiedliche prim~ire und sekund~ire Struktur aufweisen.
BIBLIOGRAPHIE.
1. Mairy, M. & Denis, H. (1971) Develop. Biol., 24, 143. 2. Denis, H. & Mairy, M. (1972) Eur. J. Biochem., 25,
524. 3. Ford, P. 3. (1971) Nature, 233, 561. 4. Wegnez, M. a Denis, H. (1972) Biochimie, XX, 20. 5. Brown, D. D. ~ Littna, E. (1964) J. Mol. Biol., 8,
669.
BIOCH1MIE, 1972, 54, n ° 9.

1 1 9 6 H . D e n i s , M . W e g n e z e t R . W i l l e m .
6. Mairy, M. a Denis, H. (1972) Enr. J. Biochem., 25, 535.
7. Mandell, J. D. a Hershey, A. D. (1960) Anal. Bio- chem., 1, 66.
8. Soave, C., Galante, E. a Torti , G. (1970) Bull. Soc. Chim. Biol., 52, 857.
9. Loening, U. E. (1967) Biochem. J., 101, 251. 10. Bray, G. A. (19.60) Anal. Biochem., 1, 279. 11. Adams, J. M., Jeppesen, P. G. N., Sanger, F.
Barrell , B. G. (1969) Nature, 223, 1009. 12. Boedtker, H. (1967) Biochemis try , 6, 2718. 13. Cox, R. A. a Canaga l ingam, K. (1967) Biochem. g.,
103, 749. 14. Aubert , M., Scott, J. F., Reynier , M. a Monier, R.
(19.68) Proc. Natl. Acad. Sci. U.S.A., 61, 292. 15. Denis, H., Wegnez, M. & Wil lem, R., m a n u s c r i t
en prfiparation. 16. Nieuwkoop, P. D. s~ Faber, J. (1967) (( Normal Table
of Xenopus laeois (Daudin) >>, North Holland Pub l i sh ing Co.
17. Al tman, S. a Smith, J. D. (1971) Nature New Biol., 233, 35.
18. Maden, B. E. H. (1971) Progress in Biophysics and Molecular Biology, 22, 127.
19. Hatlen, L., Amaldi , F. ~ Attardi, G. (1969) Bio- chemistry , 8, 4989.
26. .Jordan, B. R., Feun teun , J. ~ Monier, R. (1970) J. Mot. Biol., 50~ 605.
21. Forget, B. G. ~: W e i s s m a n , S. M. (1967) Nature, 213, 878.
22. Wegnez, M. ,~ Denis, H., experiences in6dites.
Note ajoutde sur dpreuoe. - - La s t ruc ture p r imai re complete du RNA 5 S soma t ique et du RNA 5 S d 'ooeytes a 6t6 r6eemment d6termin6e (Wegnez, M., Monier, R. ~ Denis, H. (1972) FEBS Letters, 25, 13). Le3 deux types de RNA 5 S possbdent 6 r6sidus diff6- rents sur 120.
BIOCH1M1E, 1972, 54, n ° 9.
Top Related