
Physicochimie de Macromolécules - lism.cnrs-mrs.fr · Evolution dirigée. But du programme ... –...
237
Physicochimie de Macromolécules Evolution et physicochimie James Sturgis Jean-Pierre Duneau
-
Upload
truongduong -
Category
Documents
-
view
215 -
download
0
Transcript of Physicochimie de Macromolécules - lism.cnrs-mrs.fr · Evolution dirigée. But du programme ... –...

Physicochimie de Macromolécules
Evolution et physicochimie
James SturgisJean-Pierre Duneau

Sommaire
But du programme Organization des
enseignements L'examen Les resources
Rappels– L'évolution– La génétique– Les mutations
Evolution dirigée

But du programme
Donner une introduction– A la physicochimie des
protéines– A l'analyse de fonction– Aux relations structure-
fonction
Voir comment mettre en evidence les liens structure-fonction.
Renforcer votre anglais scientifique.

Organisation des Enseignements
Les Cours (JS) Essentiellement une
seule exemple (hémoglobine) dans les cours...
Les TD (JPD) Analyse des données
– Pratiquer la conversion des mesures experimentales en valeurs utiles.
– Comment lire un article scientifique.

Sommaire des cours
Chapitre 1 : Introduction et rappels. Chapitre 2 : Les hémoglobines. Chapitre 3 : Variations entre éspèces :
des oies et des poissons Chapitre 4 : Variations intra-éspèce :
hémoglobines humaines Chapitre 5 : Découverte de fonction :
l'hémoglobine d'ascaris. Chapitre 6 : Hémoglobines de signalisation

Organisation de l'examen
Deux parties... Exercise d'analyse
– Une serie de questions basées sur un extrait d'article.
Sujet plus rédactionelle– Souvent un choix– Souvent demandant une
synthèse.
Les deux parties sont important et peuvent porter sur...– le contenu des cours– le contenu des TD.

Resources
Les transparents – Site web du
département.
Les TD– Site web du
département.
Les articles pour approfondissement.
http://biologie.univmrs.fr/

Rappels sur l'évolution

Introduction
Rappels sur...– L'évolution– La génétique– Les mutations et la
variation.– La sélection et l'absence
de la sélection.– Quelques nouvautés!
Une exemple d'évolution dirigée.
Variants du papillon Biston betularia

Evolution
Variabilité individuelle Transmission de cette
variabilité aux enfants Sélection des individus
les plus adaptés à l'environnement
Darwin avait 31 ans quand il élaborait sa théorie.
Aujourd'hui nous pouvons examiner les bases moléculaires de ces trois points.

Variabilité – les mutations
Changements des séquences d'ADN – Codants et – Non-codants
Réorganisations d'ADN– Délétion– Transposition– Insertion Erreurs de reproduction,
mutations pontuelles.
Plus facile a comprendre au niveau de la protéine.
Variabilité individuelle Transmission de cette
variabilité aux enfants Sélection des individus
les plus adaptés à l'environnement

Variabilité – les mutations
Changements des séquences d'ADN – Codants et – Non-codants
Réorganisations d'ADN– Délétion– Transposition– Insertion Transposons et virus,
systèmes de réparation
Variabilité individuelle Transmission de cette
variabilité aux enfants Sélection des individus
les plus adaptés à l'environnement

Variabilité – les mutations
Les mutations ponctuelles peuvent etre prédites
Vitesse de mutation:– Jukes Cantor (1969)
– Kimura (K2P-1980)
– Plus complexe – notamment dépendant de contexte.
GA
C T
GA
C T

Variabilité – les mutations
Si les probabilités de différentes mutations peuvent être prédites leurs conséquences physiologiques ne peuvent pas facilement ...

Transmission – la génétique
Les procaryotes sont simples...– Une copie d'un simple
chromosomeparfois deux parfois des plasmides
– Qui est répliqué – et une copie distribuée à
chaque cellule fille.Transmission d'une seule copie de l'ensemble du patrimoine génétique par l'unique cellule génitrice aux deux cellules filles.
Variabilité individuelle Transmission de cette
variabilité aux enfants Sélection des individus
les plus adaptés à l'environnement

Variabilité individuelle Transmission de cette
variabilité aux enfants Sélection des individus
les plus adaptés à l'environnement
Transmission – la génétique
Transmission d'une copie (ou plusieurs copies) du patrimoine génétique par chaqun de deux progenitors aux enfants.
La reproduction sexué chez les eukaryotes est plus complexe...– Plusieurs copies de
plusieurs chromosomes– Qui sont répliqués– Réorganisés (linkage)– La moitié des
chromosomes hérités de chaque parent.
Variabilité individuelle Transmission de cette
variabilité aux enfants Sélection des individus
les plus adaptés à l'environnement

Sélection
Une fois qu'une mutation est introduite le nouvel allèle peut être:– Perdu– Fixé (transmis dans la
population).
On parle de la penetration d'une allèle dans la population.
Variabilité individuelle Transmission de cette
variabilité aux enfants Sélection des individus
les plus adaptés à l'environnement

Sélection
Le sort d'une allèle dans la population dépend de– La pression de sélection – La dérive génétique
(chance)
Nota:– On considère ici des
allèles dans la population et pas
– les mutations dans une génome.
– Une changement d'échelle.

Même dans l'absence de sélection la pénétration d'un allèle dans la population varie de generation en generation.
Avec une petite population c'est parfois très rapide
F(0): p(bleu) = 0,5
Dérive génétique et sélection
La moitié de la population ont des enfants

F(0): p = 0,5
F(1): p = 0,7
Dérive génétique et sélection
Même dans l'absence de sélection la pénétration d'un allèle dans la population varie de generation en generation.
Avec une petite population c'est parfois très rapide
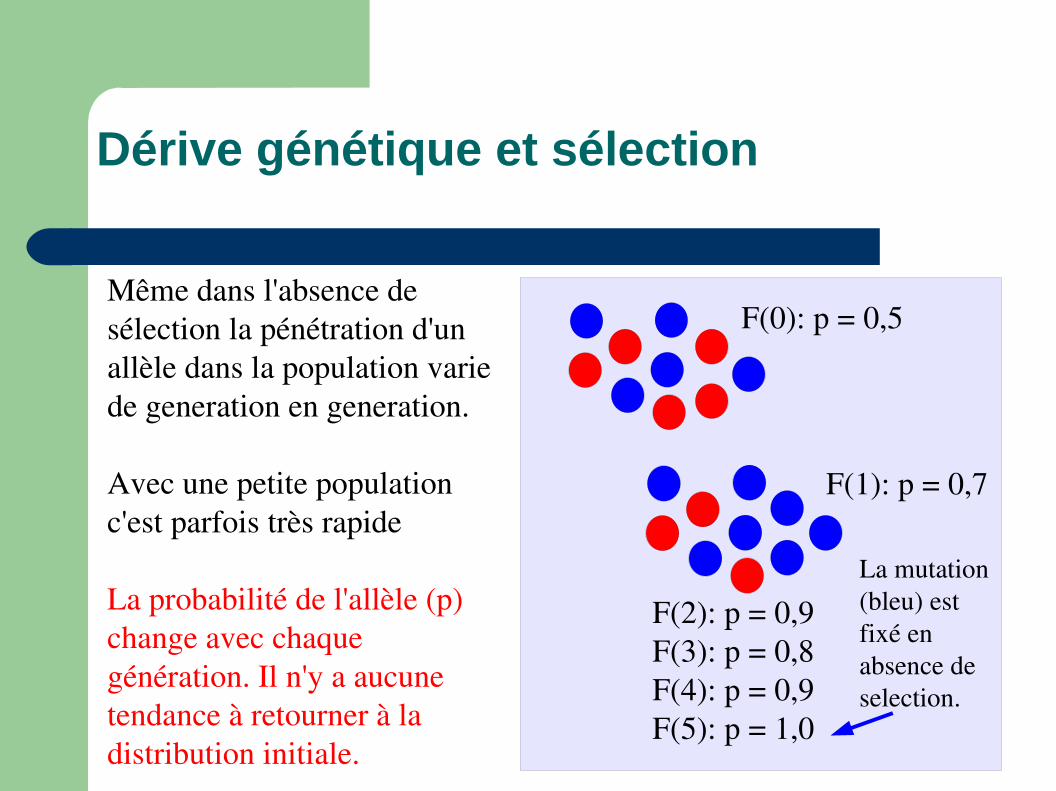
Même dans l'absence de sélection la pénétration d'un allèle dans la population varie de generation en generation.
Avec une petite population c'est parfois très rapide
La probabilité de l'allèle (p) change avec chaque génération. Il n'y a aucune tendance à retourner à la distribution initiale.
F(0): p = 0,5
F(1): p = 0,7
F(2): p = 0,9F(3): p = 0,8F(4): p = 0,9F(5): p = 1,0
Dérive génétique et sélection
La mutation (bleu) est fixé en absence de selection.

Dérive génétique et sélection
Dans une population la dérive génétique s'assure que même dans l'absence de toute sélection une nouvelle allele sera eventuellement fixée (p = 1) ouperdue (p = 0)
et l'allele initialeperdue (p=0) oufixée (p=1).
Dans une population stable en moyenne cela prend 4N generations
Probabilité environ 1/N
Une derive plus vite dans des petites populations.

Dérive génétique et sélection
La dérive génétique accumule avec le temps.– Elle cause une perte de
variabilité dans une population
– Elle augmente la variabilité entre deux populations
La vitesse de la dérive est plus grande pour des petites populations.
Les mutations accumule avec les temps– Elles augmentent la
variabilité entre populations et dans une population.

Dérive génétique et sélection
La selection :– modifie le temps pour la
fixation d'une mutation – modifie le probabilité de
fixation d'une mutation.
Une pression de sélection peut augmenter la vitesse d'une dérive génétique– en diminuant la population
– sans que l'allèle ait la moindre influence sur la survie!!
– a cause des liens génétiques
Nota: fixation en depit d'une sélection négative est possible, seulement moins probable que 1/N et moins rapide que ~4N génerations en moyenne

Dérive génétique et sélection
les mutations augmentent la variabilité génique
la sélection et la dérive génétique diminuent la variabilité génétique.
Tous les deux modifient les séquences des gènes et des protéines.
Quelle est l'importance rélative de la sélection et de la dérive génétique dans la détermination de la séquence d'une protéine?
Tous les différences ne sont pas dû a la sélection.

Le Paradigme (Biologie Moléculaire)
Séquence du Géne
Séquence de la Protéine
Structure de la Protéine
Fonction de la Protéine
Variabilité individuelle Transmission de cette
variabilité aux enfants Séléction des individus
les plus adaptés à l'environnement

Le Paradigme (Evolution Moléculaire)
Séquence du Génome
ARN messagerSéquence, Quantité et Patron d'expression
ProtéineSéquence, Quantité et Patron d'expression
Structure
FonctionBiochimique Cellulaire Physiologique
Résistance à la séléction

Le Paradigme (Evolution Moléculaire)
Séquence du Génome
ARN messagerSéquence, Quantité et Patron d'expression
ProtéineSéquence, Quantité et Patron d'expression
Structure
FonctionBiochimique Cellulaire Physiologique
Résistance à la séléction

Le Paradigme (Evolution Moléculaire)
Pour le comprendre il faut examiner des systèmes modeles un peu plus simples...
Les plus grandes difficultés sont d'établir les liens en structure – fonction et ecologie.
Séquence du Génome
ARN messagerSéquence, Quantité et Patron d'expression
ProtéineSéquence, Quantité et Patron d'expression
Structure
FonctionBiochimique Cellulaire Physiologique
Résistance à la séléction

Etude de l'evolution moléculaire
Comment éclaircir les liens entre séquence et niche écologique?
Manipuler d'une façon expérimentale la variabilité et la pression de sélection.
– Evolution dirigée...
– Mutagénèse dirigée...
Etudier la variation naturelle.
Facile a comprendre les resultats... difficile d'en avoir beaucoup.
Maintenant!
Facile d'avoir beaucoup de resultats... dificle de les comprendre!
Les autres cours!

Evolution dirigée
Comment éclaircir les liens entre séquence et niche écologique?
Manipuler d'une façon expérimentale la variabilité et la pression de sélection.
– Evolution dirigée...
– Mutagénèse dirigée...
Etudier la variation naturelle.
Evolution d'une fucosidase à partir d'une glucosidase.
Protocol de DNAshuffling et sélection un exemple d'évolution dirigée.
Zhang JH, Dawes G. and Stemmer W.P. 1997. Directed evolution of a fucosidase from a galactosidase by DNA shuffling and screening. Proc. Natl Acad. Sci. USA 94: 45044509.

DNA–shuffling
Fragmentation aléatoire
Réassemblage par PCR (mutagénique et recombinogénique)
Criblage (hydrolyse du nouveau substrate)
Plusieurs génomes

DNA–shuffling
Fragmentation aléatoire Réassemblage par
PCR:– mutagène et– recombinatoire
Criblage
Population de fragmentsavec mutations

DNA–shuffling
Fragmentation aléatoire Réassemblage par
PCR:– mutagène et– recombinatoire
Criblage
Nouvelle sequence différente des originals

DNA–shuffling
Fragmentation aléatoire Réassemblage par
PCR:– mutagène et– recombinatoire
Criblage
Nouvelle sequence différente des originals
De plus introduction de mutations

DNA–shuffling
Fragmentation aléatoire Réassemblage par
PCR:– mutagène et– recombinatoire
Criblage

DNA–shuffling
Ce protocol est capable après 8 cycles de convertir une glucosidase en fucosidase (presque)...
Comment savoir?– k
cat = vitesse maximale de la réaction (plus c'est grande
mieux c'est).
– KM = affinité pour le substrat (plus c'est basse mieux
c'est).
– kcat
/KM = mesure d'efficacité (grande est mieux).
– specificité = rapport de kcat
/KM pour différentes substrats.

DNA–shuffling
Glucosidase FucosidasePNPG k
cat (s1) 268 31
KM
(mM) 0,04 0,18
kcat
/KM
6700 172
PNPF kcat
(s1) 209 97K
M (mM) 31 1,5
kcat
/KM
6,7 64,4
Specificity 1000 2,7

DNA–shuffling
Quelles sont les modifications de séquence...
t/cg/a g/a
t/c
c/t a/g a/g a/g a/g a/g
g/a
g/ac/t
D908
NN604
S
Q573
R
P511
SQ135
RV9IV/P
G/S
13 mutations ponctuelles – 6 changements dans la séquence protéique de la glucosidase – 5 changements silencieux
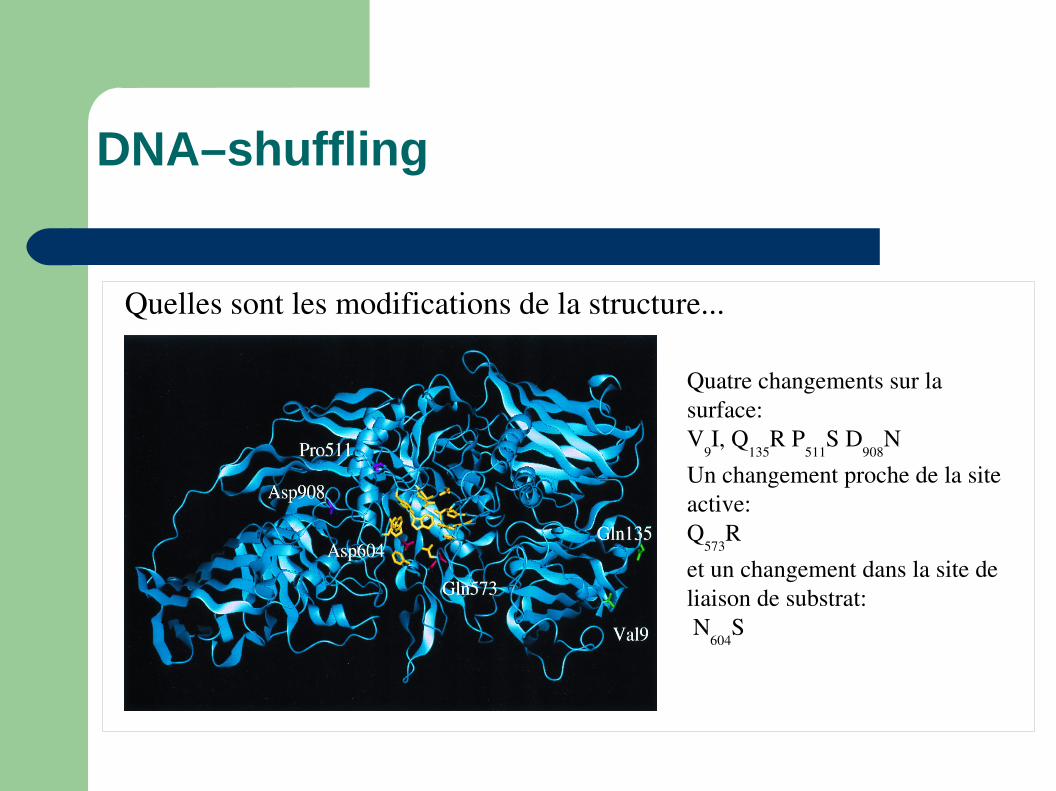
DNA–shuffling
Quelles sont les modifications de la structure...
Quatre changements sur la surface:V
9I, Q
135R P
511S D
908N
Un changement proche de la site active:Q
573R
et un changement dans la site de liaison de substrat: N
604S

DNA–shuffling
Quelles sont les modifications de séquence...
t/cg/a g/a
t/c
c/t a/g a/g a/g a/g a/g
g/a
g/ac/t
D908
NN604
S
Q573
R
P511
SQ135
RV9IV/P
G/S
Résidu dans la site actif
1 à 3 sur 13 (723%) mutations probablement important – résultat de sélection positive et les autres 11 d'une dérive génétique.
Modification du niveau d'expression?

Evolution dirigée
Il est possible à diriger l'évolution moléculaire et construire des nouvelles protéines.
Avec un contrôle précise sur– la variabilié et – la sélection.
Le plupart des modifications selectionés ne donnent pas une sélection positive.
Rôle important de – la petite population et – la forte taux de mutation.

Liens séquence – ecologie
Nous arrivons souvent, avec l'aide de la structure d'une protéine, – A comprendre les liens entre séquence et fonction
biochimique.Mutation N
604S et activité et affinité pour substrat
– Meme si nous pouvons pas toujours les prédire avec précision.
Nos connaissances ne suffisent pas normalement a voir les relations entre une séquence protéique et la niche écologique d'une espèce.

Evolution moléculaire
Un bon modèle: l'hémoglobine.

Evolution Moléculaire
Trouver un système modele simple:– Facile a étudier– Beaucoup
d'information– Possible à voir les
liens entre fonction et écologie.
L'Hémoglobine

Evolution Moléculaire
Rôle physiologique important (transport d'oxygène)
Patron d'expression simple (erythrocytes)
Structure et fonction biochimique bien établies.

La famile des hémoglobines
Nous allons parler de toute la famille des hémoglobines et pas uniquement l'hémoglobine du sang humain.
La structure de la famille La diversité de la famille L'hémoglobine sanguine
– Cooperativité et structure.
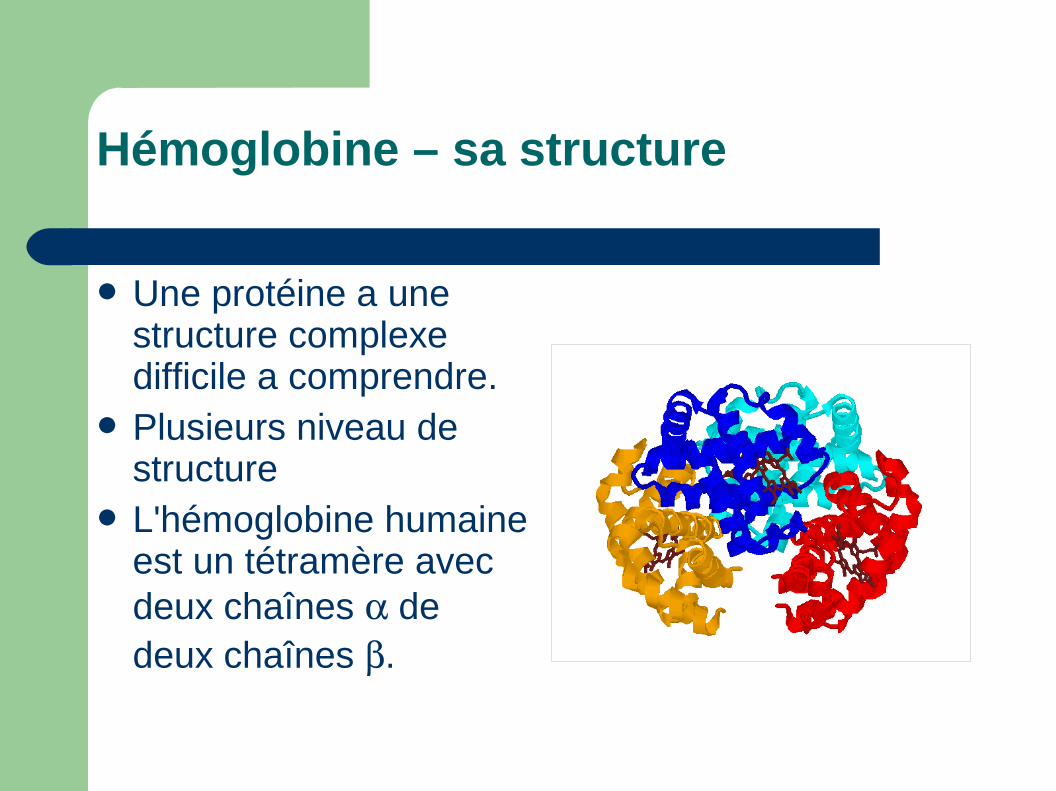
Hémoglobine – sa structure
Une protéine a une structure complexe difficile a comprendre.
Plusieurs niveau de structure
L'hémoglobine humaine est un tétramère avec deux chaînes de deux chaînes .
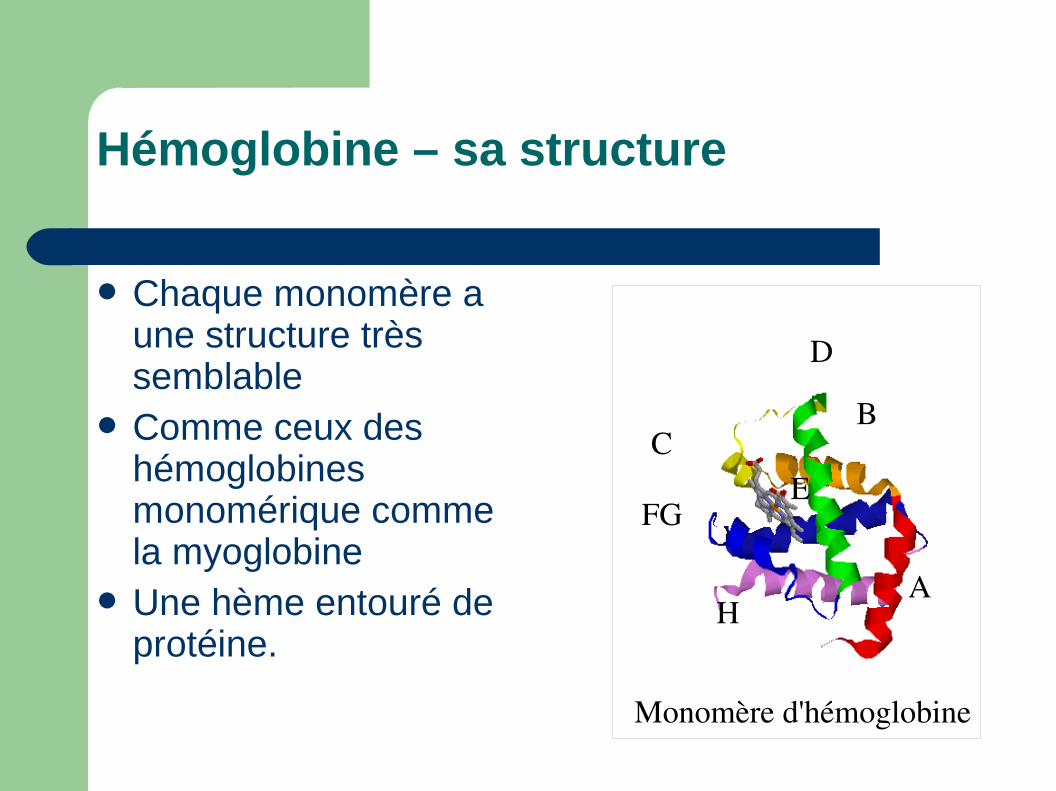
Monomère d'hémoglobine
A
BC
D
EFG
H
Hémoglobine – sa structure
Chaque monomère a une structure très semblable
Comme ceux des hémoglobines monomérique comme la myoglobine
Une hème entouré de protéine.
Monomère d'hémoglobine
A
BC
D
EFG
H
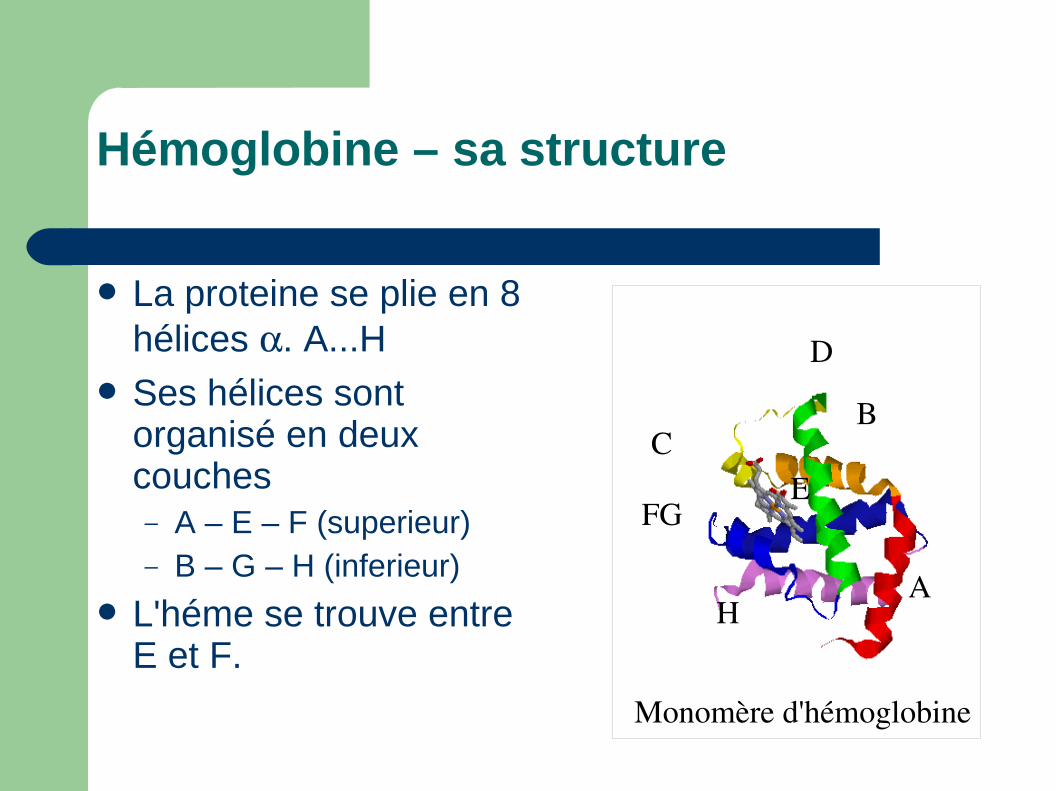
Hémoglobine – sa structure
La proteine se plie en 8 hélices . A...H
Ses hélices sont organisé en deux couches– A – E – F (superieur)– B – G – H (inferieur)
L'héme se trouve entre E et F.
HF8HE6
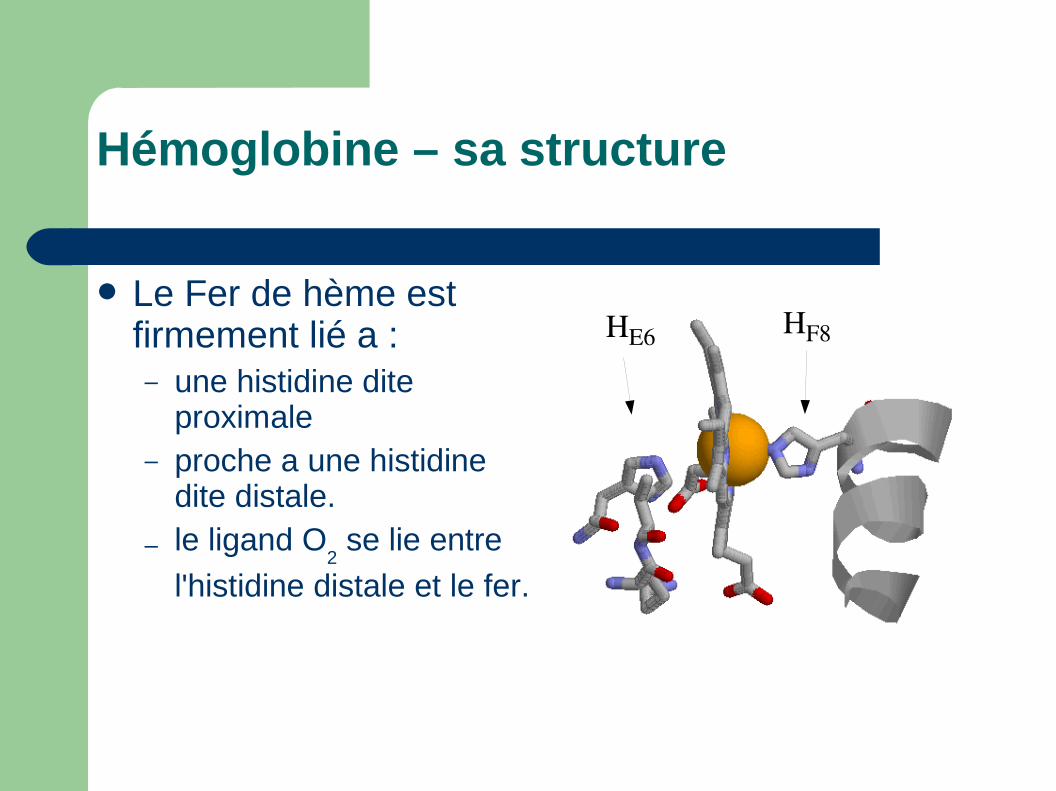
Hémoglobine – sa structure
Le Fer de hème est firmement lié a : – une histidine dite
proximale– proche a une histidine
dite distale.
– le ligand O2 se lie entre
l'histidine distale et le fer.
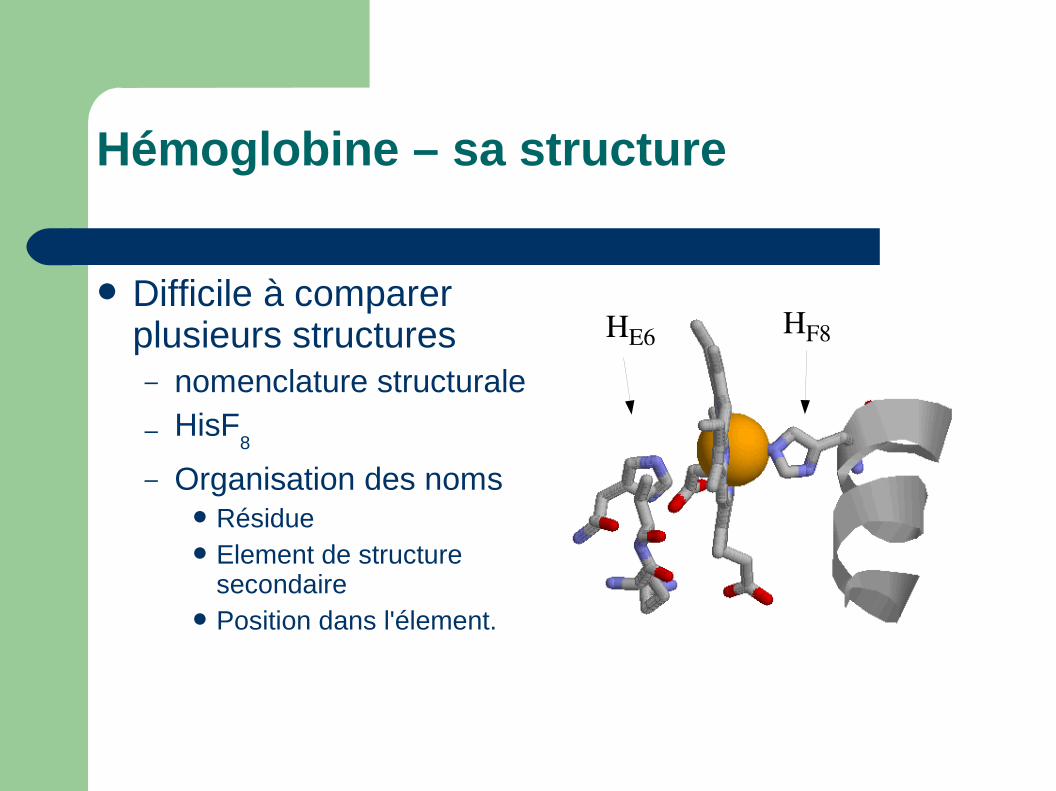
Hémoglobine – sa structure
Difficile à comparer plusieurs structures– nomenclature structurale
– HisF8
– Organisation des nomsRésidueElement de structure
secondairePosition dans l'élement.
HF8HE6

Evolution archaique
On pense que ces protéines ont une origine achaique avant l'accumulation d'O
2 vers -2GYa.
Plusieurs protéines ont une repliement similaire comme:– phycocyanines, colicines, la toxine dipthérique,
hémocyanine ou le cytochrome b2.
Avec l'accumulation d'oxygène elles ont prise un rôle important dans le liaison d'oxygène.
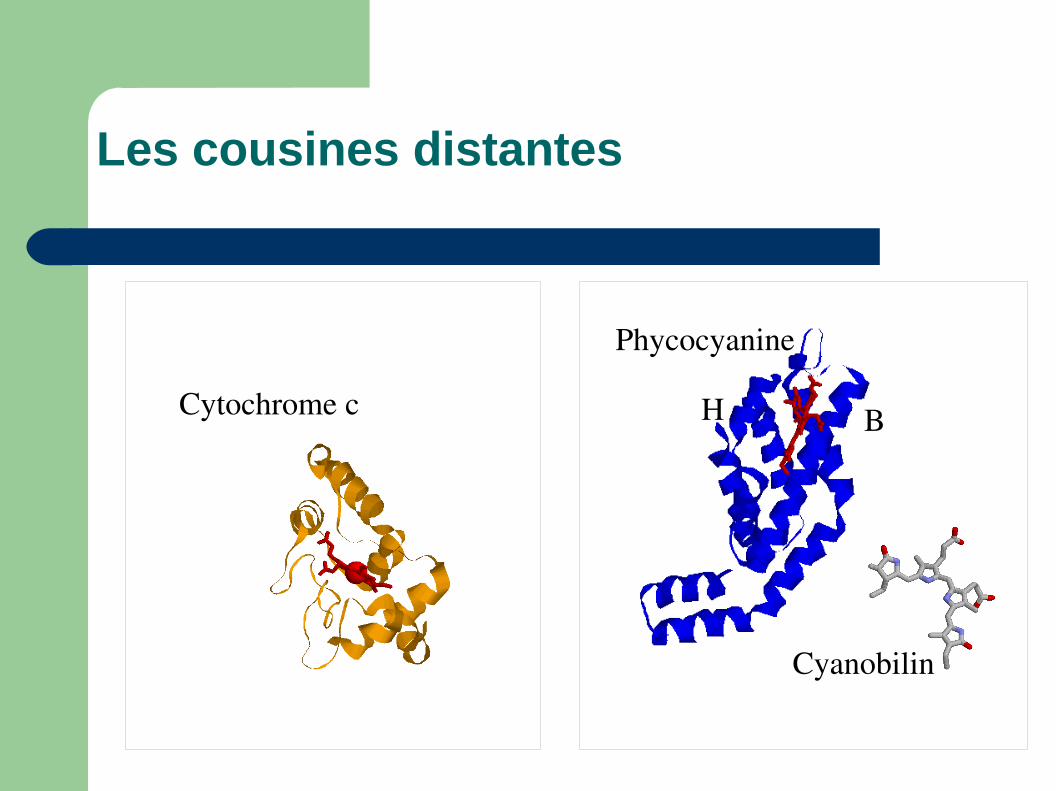
Les cousines distantes
Cytochrome c
Phycocyanine
BH
Cyanobilin

Une famille diverse
Hémoglobines – à basse affinité– à haute affinité
Dissociation lenteAssociation rapide
Fusions
Hémoglobines – d'animale, – de plante, – de bactérie
Hémoglobines– monomérique, – dimérique et plus.
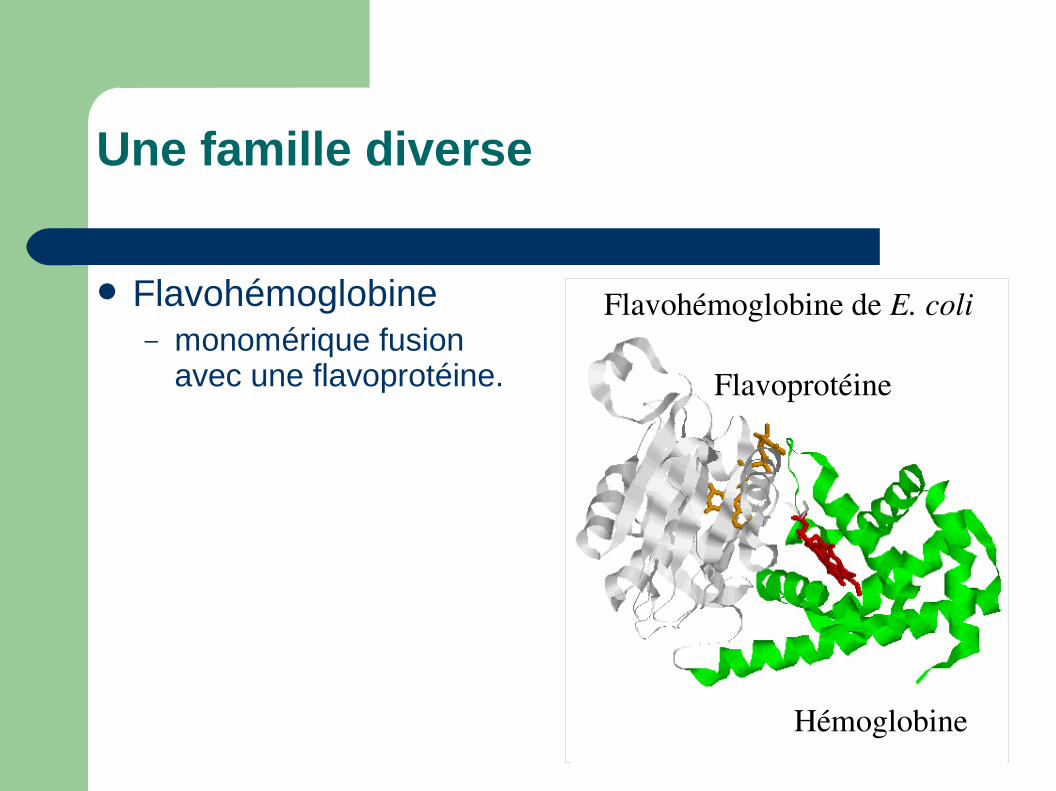
Flavoprotéine
Hémoglobine
Flavohémoglobine de E. coli
Une famille diverse
Flavohémoglobine– monomérique fusion
avec une flavoprotéine.
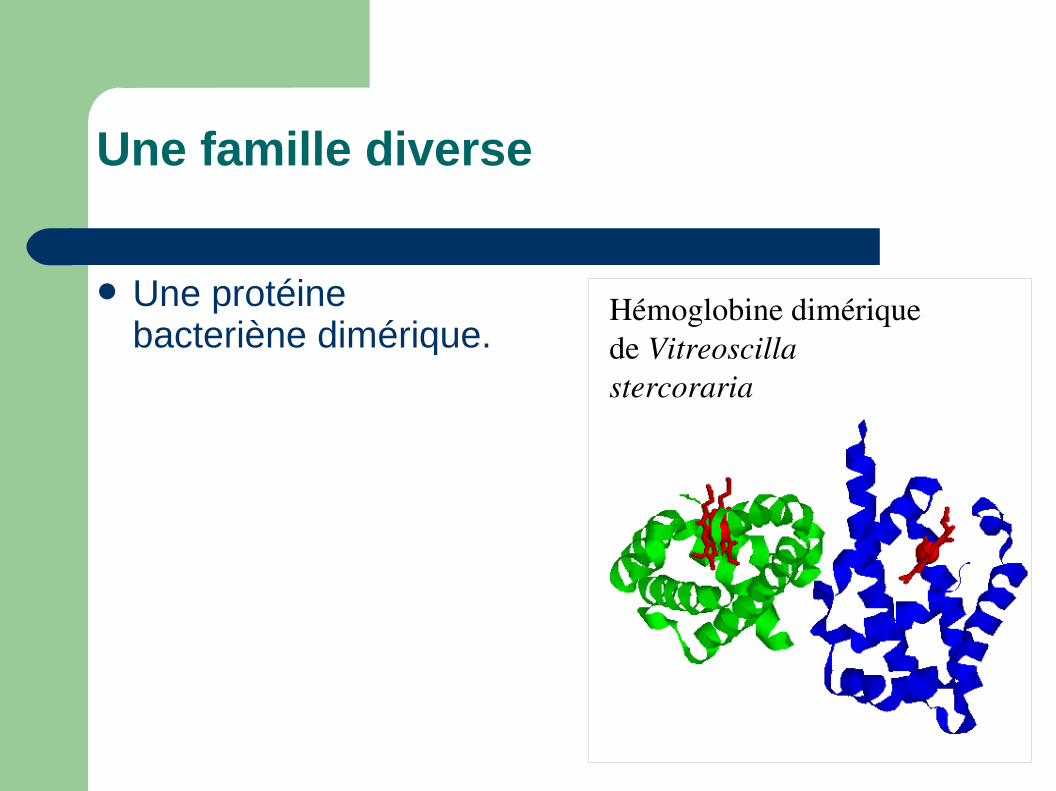
Une famille diverse
Hémoglobine dimérique de Vitreoscilla stercoraria
Une protéine bacteriène dimérique.
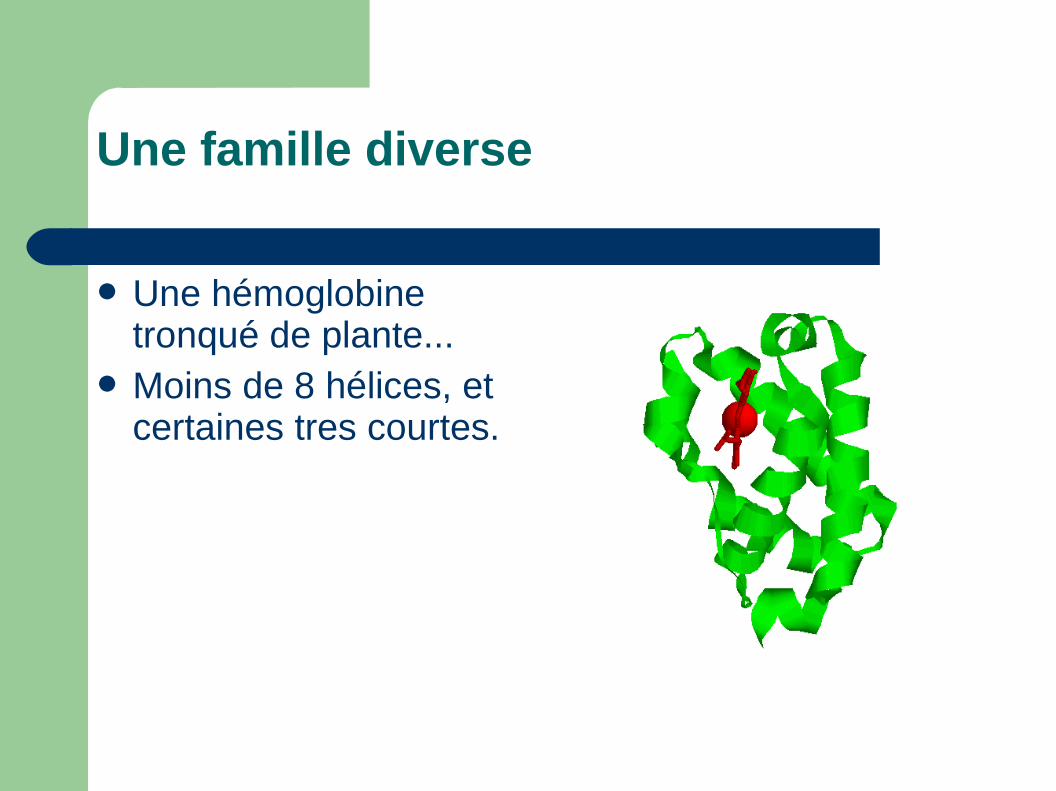
Une famille diverse
Une hémoglobine tronqué de plante...
Moins de 8 hélices, et certaines tres courtes.
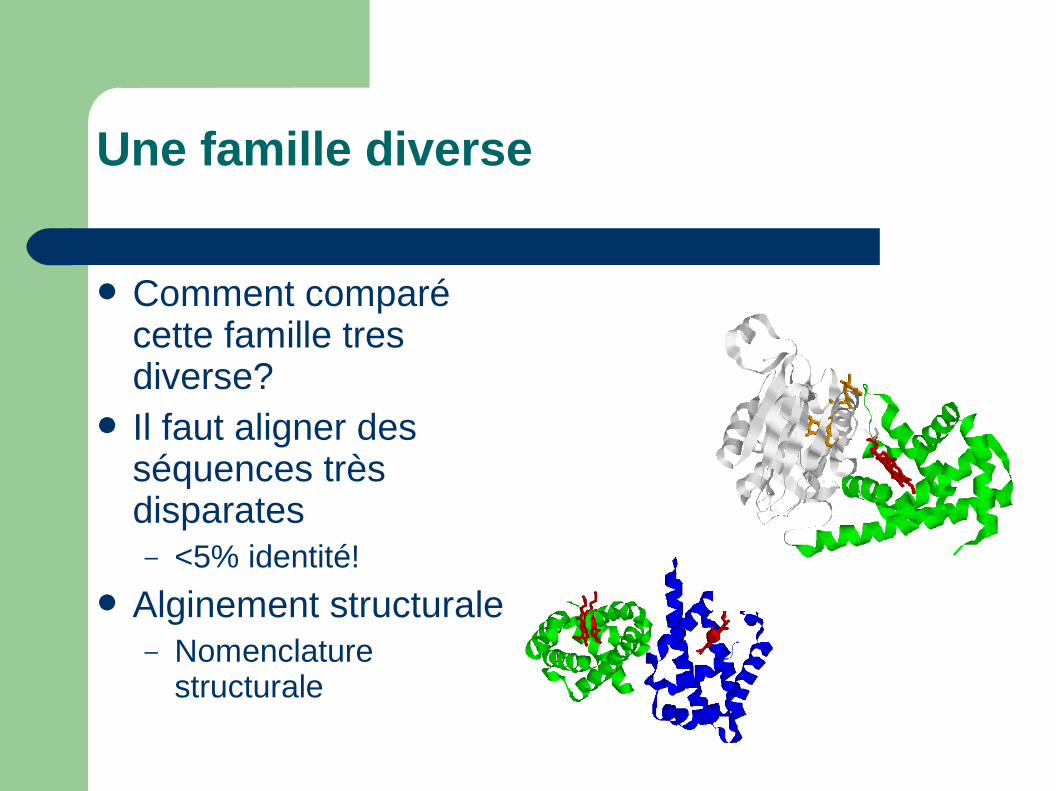
Une famille diverse
Comment comparé cette famille tres diverse?
Il faut aligner des séquences très disparates– <5% identité!
Alginement structurale– Nomenclature
structurale

Construction d'un arbre
Un “gabarit”– Il faut aligner des
séquences très disparates.
Evolution de fonction.
C'est quoi un gabarit?– Un patron qui donne les
différentes possibilites aux differentes positions...
Il existe depuis longtemps un gabarit basé sur l'alignement de protéines des vertebrés (Bashford, Chothia et Lesk) mais il est mal adapté aux hémoglobines des non-vertebrés....
Moens et al. (1996) Mol. Biol. Evol. 13: 324333

Construction d'un arbre
Alignement structural...– Il faut aligner les
structures plutot que simplement les séquences.
– Il faut definir les possibilités
C'est quoi un gabarit?– Un patron qui donne les
differentes possibilites aux differentes positions...
Il existe depuis longtemps un gabarit basé sur l'alignement de protéines des vertebrés (Bashford, Chothia et Lesk) mais il est mal adapté aux hémoglobines des non-vertebrés....

Construction d'un arbre
Le gabarit– Il n'est pas tres exigeant,
beaucoup de possibilites a certains positions.
– Seulement deux positions requises CD1 = F et F8 = H.
– S = surface, i = interne et req = requis.

Construction d'un arbre
Le gabarit– Les listes et penalties ne
renforcent pas toujours nos conceptions des groupes d'acide amines.
– C2 – P – G – ST – AQ– E7 – HQ – VL– G12 – VILMF – T– (RKDENQHGASCT)P

Construction d'un arbre
Hemoglobins
Plantes et metazoaires
Ciliates, Chlamydomonas et Nostoc
Levures et Bactéries
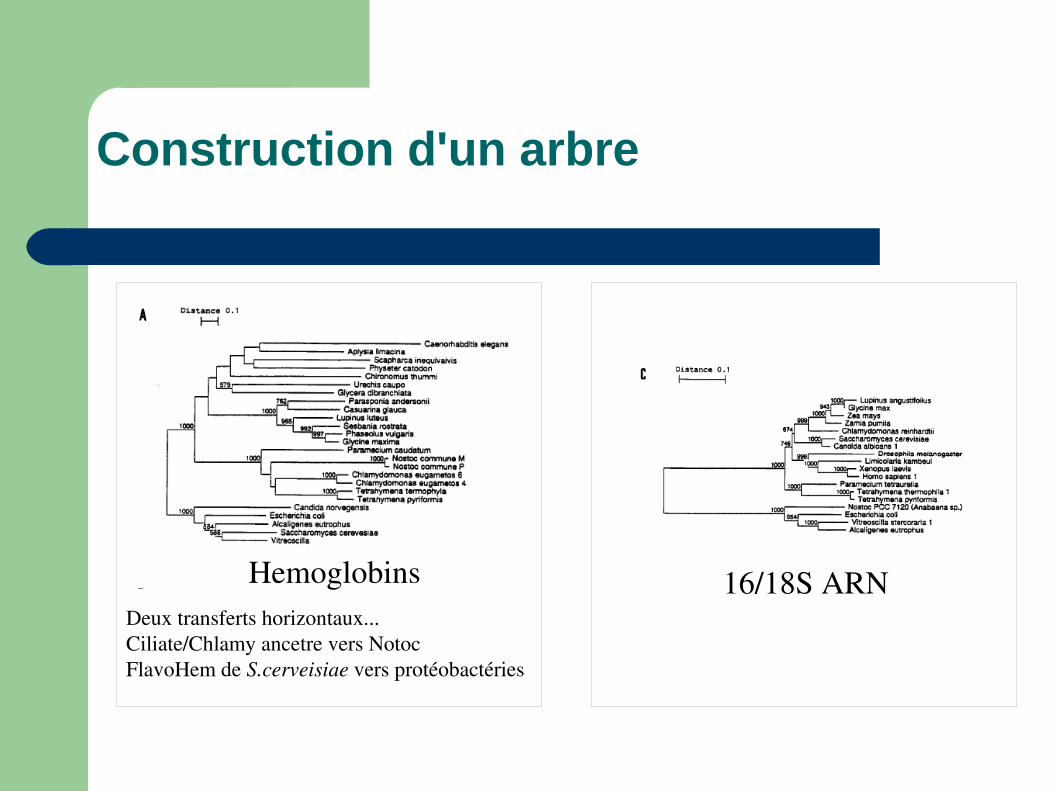
Construction d'un arbre
Hemoglobins 16/18S ARNDeux transferts horizontaux...Ciliate/Chlamy ancetre vers NotocFlavoHem de S.cerveisiae vers protéobactéries
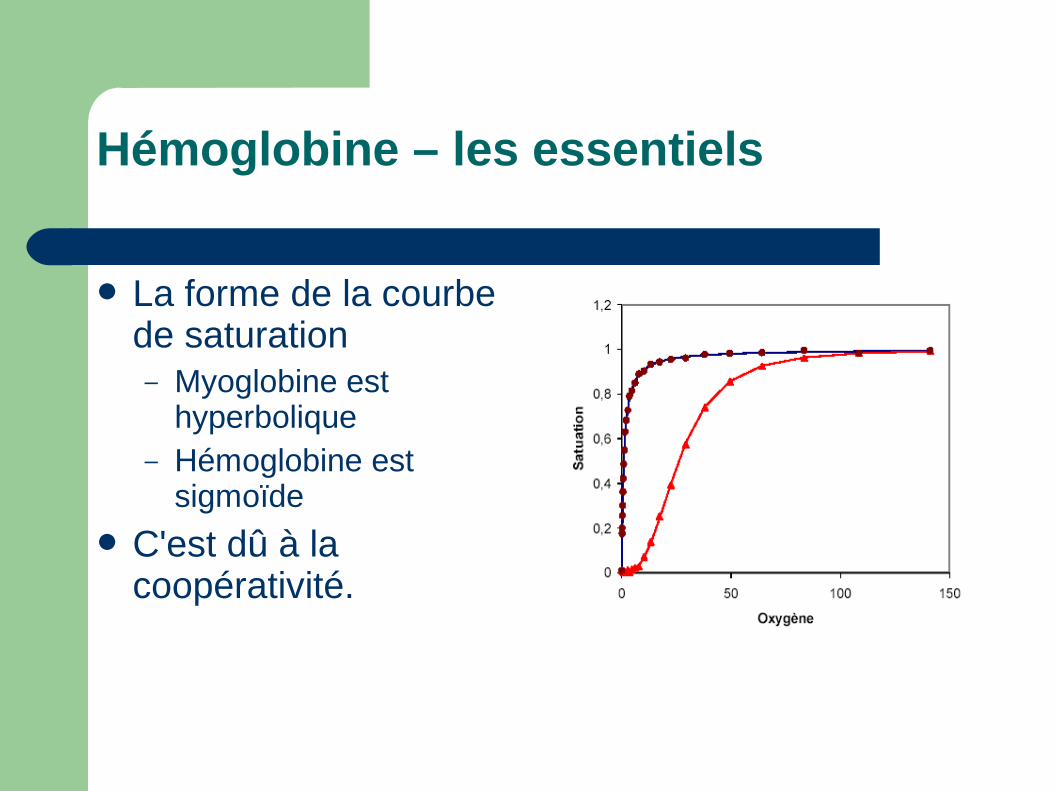
La forme de la courbe de saturation– Myoglobine est
hyperbolique– Hémoglobine est
sigmoïde
C'est dû à la coopérativité.
Hémoglobine – les essentiels

Hémoglobine – les essentiels
Explication moléculaire de la cooperativité...
Il faut plusieurs sites de fixation– L'hémoglobine est un
tétramère.

Hémoglobine – les essentiels
Explication moléculaire de la cooperativité...
Il faut plusieurs sites de fixation
Les différentes sites qui interagissent– A courte distance ou– à longue distance
(changements de structure)

Hémoglobine – les essentiels
Explication moléculaire de la cooperativité...
Il faut plusieurs sites de fixation
Les différentes sites qui interagissent
Deux structures– Tendue (T) et – Relachée (R)

Liaison de l'oxygène facilite une changement de conformation.– L'information est transduit de l'interieur de la protéine
vers l'exterieur.
Les deux conformations ont une affinité pour l'oxygène differente– T (basse affinité) / R (haute affinité)
Hémoglobine – les essentiels
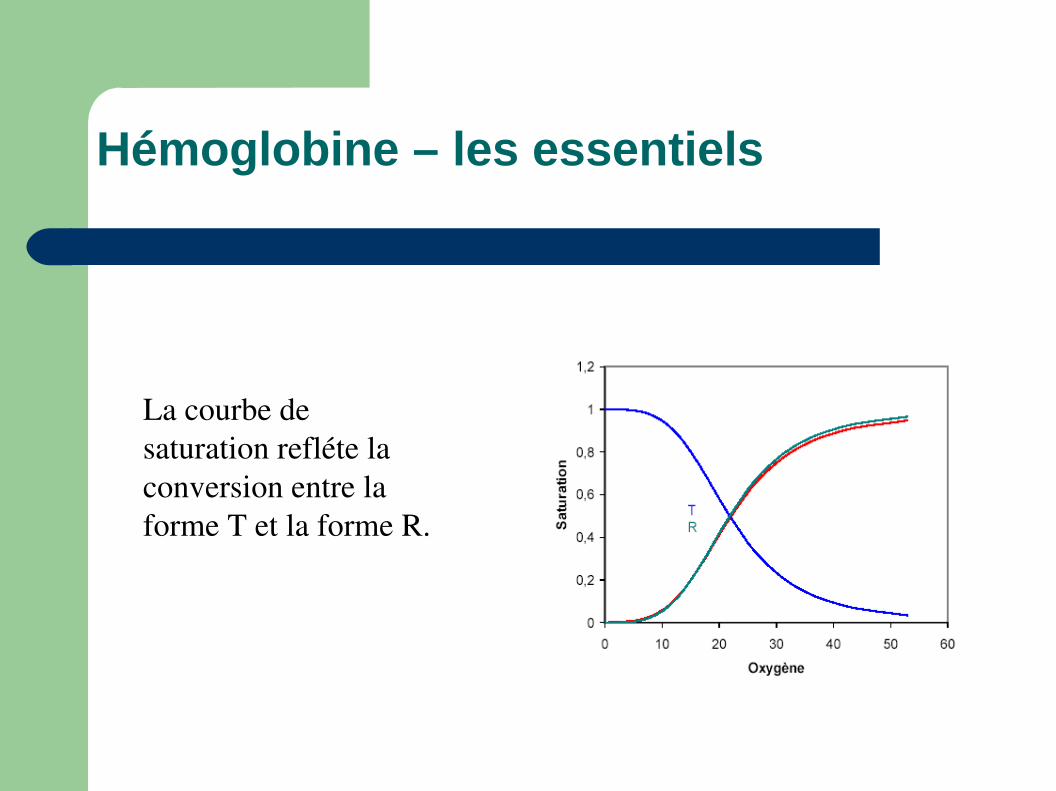
La courbe de saturation refléte la conversion entre la forme T et la forme R.
Hémoglobine – les essentiels

Des importants modifications de conformation des C-terminaux...
Une liaison hydrogène entre le tyrosine HC2 et la chaine peptidique (F8) est brisée.
Helice F
YHC2
HF8HE6
Hémoglobine – les essentiels

H146
pont salin avec D94
pont salinavec
Que font les C-terminaux des sous-unitées?
Hémoglobine – les essentiels
Mécanique

Que font les C-terminaux des sous-unitées? R141
pont salin pont salin D126
Hémoglobine – les essentiels

T RL
0
c4L0
Les oxygènes destabilisent les ponts salins et ainsi rendent la transition TR plus facile.
c=K R
KT KT
4 KR
4
Donc: c > 1K
R > K
T, comme ils sont des
constants d'association ca veut dire que la transition TR (L0) augment l'affinité.
Hémoglobine – les essentiels
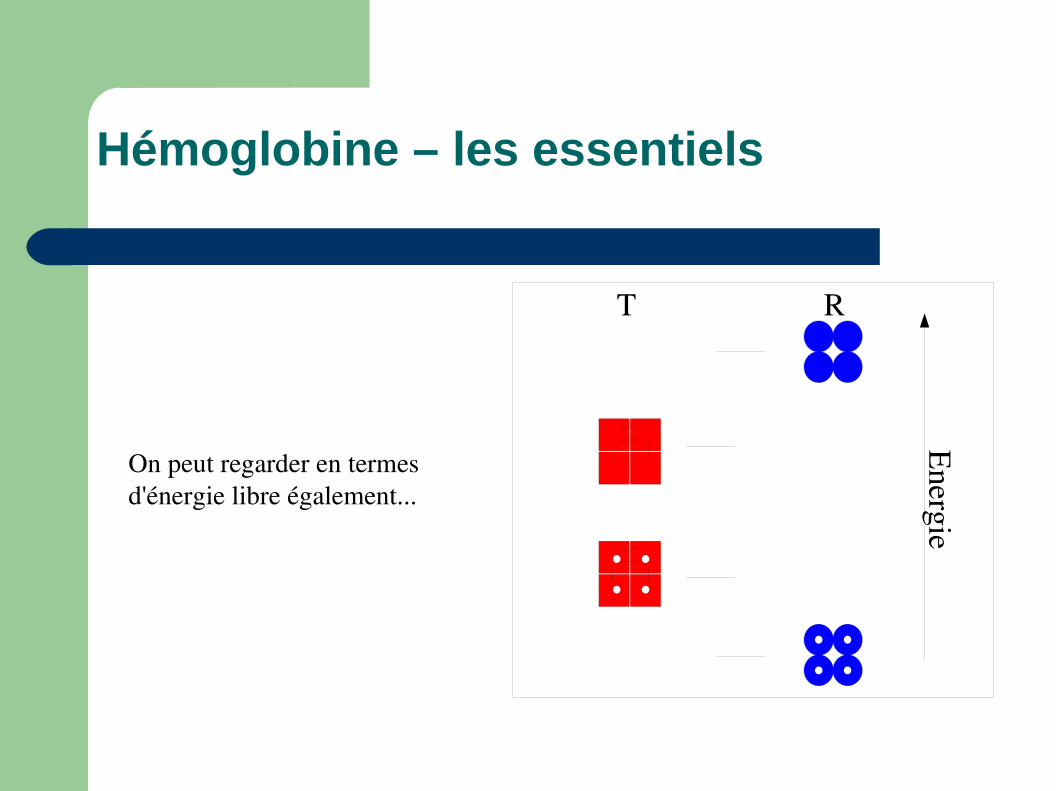
T R
Energie
On peut regarder en termes d'énergie libre également...
Hémoglobine – les essentiels

Plusieurs modulateurs changent l'affinité.– H+
– CO2
– 23BPG
Ils modifient l'equilibre TR.
Hémoglobine – les essentiels
TR
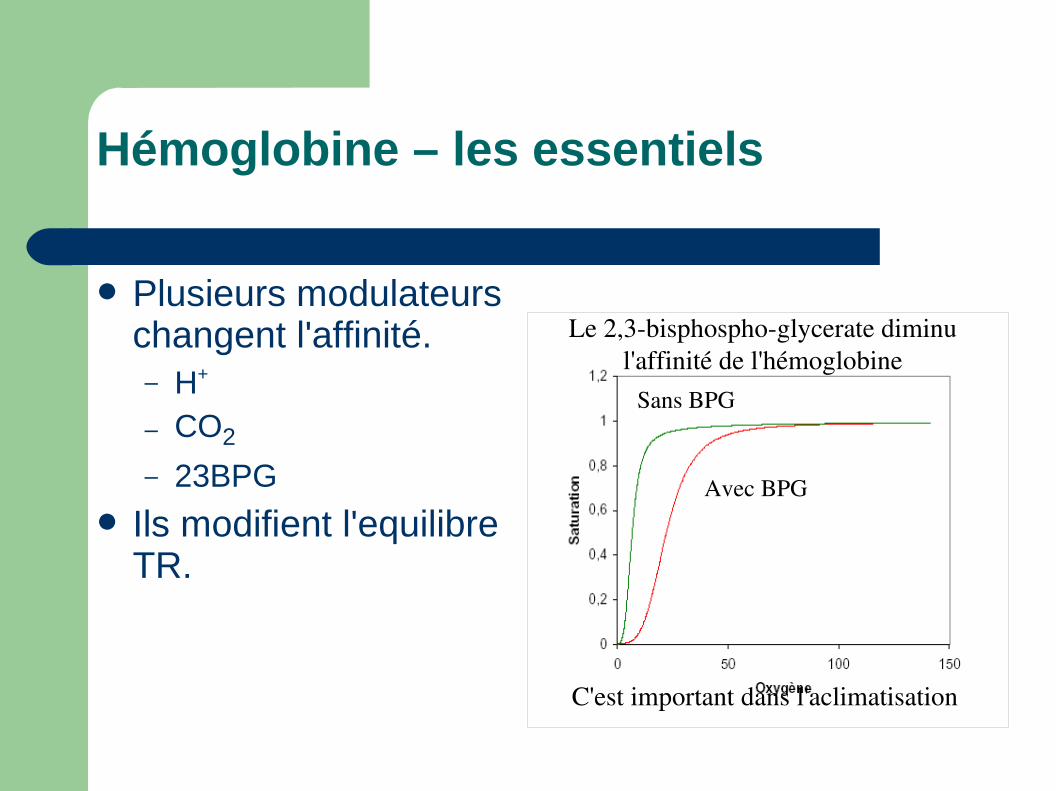
C'est important dans l'aclimatisation
Avec BPG
Sans BPG
Le 2,3bisphosphoglycerate diminu l'affinité de l'hémoglobine
Hémoglobine – les essentiels
Plusieurs modulateurs changent l'affinité.– H+
– CO2
– 23BPG
Ils modifient l'equilibre TR.

Il se lie entre les deux sousunités dans la forme T – il n'y a pas d'espace dans la
forme R
Hémoglobine – les essentiels
Plusieurs modulateurs changent l'affinité.– H+
– CO2
– 23BPG
Ils modifient l'equilibre TR.

Hémoglobine – les essentiels
TRUne forme T de basse affinité et
une forme R de haute affinité. Le P
50 est déterminé par la facilité
d'échange entre ces deux formes.
Ponts salins23BPG

Hémoglobine – les essentiels
La structure générale de l'hémoglobine
La variabilité de l'hémoglobine
Deux formes T et R et l'origine de la cooperativité.– Différences de structure– Différences d'affinité
Rôle des modulateurs

Variation de séquence I
Variations entres espèces et les liens entre séquence physicochimie et ecologie

Les exemples
Deux espèces d’oies apparentées, l’une qui vol d’Inde vers le Tibet à 7-9000m.
L'effet Root des hémoglobines de poisson.

Les oies indiennes
Les oies Anser anser (oie cendrée) habite à basse altitude, mais Anser indicus (oie à tête barrée) fait une migration annuelle à travers l’Himalaya (volant jusqu ’à 9000m).

Les oies indiennes
Les oies Anser anser (oie cendrée) habitent a basse altitude, mais les oies Anser indicus (oie à tête barrée) font une migration annuelle à travers l'Himalaya (volant jusqu'à 9000m)
Le sang d'Anser indicus a une très haute affinité pour l'oxygène, et l'animale est résistante à une stress hypoxique (une nécessité pour la migration).

Les oies indiennes
Quelles sont les origines de cette difference? Il y à que 4 différences de séquence entre les
séquences d’hémoglobine des deux espèces.
Gly18α →SerAla63α →ValPro119α →AlaGlu125β →Asp
Lesquelles de ses mutations sont responsables de la haute affinité et comment?
Certaines de ces mutations sont elles neutres?

Les oies indiennes
Quelles sont les origines de cette difference?
Il y à que 4 différences de séquence entre les séquences d’hémoglobine des deux espèces.
Gly18α →SerAla63α →ValPro119α →AlaGlu125β →Asp

Les oies indiennes
Quelles sont les origines de cette difference? Il y à que 4 différences de séquence entre les
séquences d’hémoglobine des deux espèces.
Gly18α →SerAla63α →ValPro119α →AlaGlu125β →Asp

Les oies indiennes
Quelles sont les origines de cette difference?
Il y à que 4 différences de séquence entre les séquences d’hémoglobine des deux espèces.
Gly18α →SerAla63α →ValPro119α →AlaGlu125β →Asp
Gly18α est externe sur le surface.
Ala63α est également à l’extérieur.
Glu125β a sa chaîne latérale dans le solvant.
Pro119α est dans le région de contact α
1β
1 et touche la Leu55β

Les oies indiennes
Quelles sont les origines de cette difference?– Résidues de surface
probablement peu important (pourquoi?)
– Aucune modification dans la site de liaison, pourtant modification de l'affinité.

Les oies indiennes
Suppression de deux CH2
Proline
CCH
O
N
H2C
CH2 CH2
Alanine
CCH
O
N
CH3
H
La mutation Pro119
Ala
crée un trou entre les sous-unités et .
Pas très bon pour la stabilité!
Quelle effect sur l'affinité?

Les oies indiennes
La mutation Pro119
Ala
crée un trou entre les sous-unités et .
Pas très bon pour la stabilité!
Quelle effect sur l'affinité?
Suppression de deux CH2
Pro119
Leu55
DesoxyHémoglobine

Les oies indiennes
La mutation Pro119
Ala
crée un trou entre les sous-unités et .
Pas très bon pour la stabilité!
Quelle effect sur l'affinité?
Suppression de deux CH2
Hb P50 mmHg Log P50 log(P50)HbA 5,8 0,76HbA Pro119A 3,3 0,53 0,23Anser anser 2,8 0,45Anser indica 2,0 0,30 0,15
Nota: G = RT ln(Keq)G ∝ ln(Keq) ∝ log(Keq/Keq)
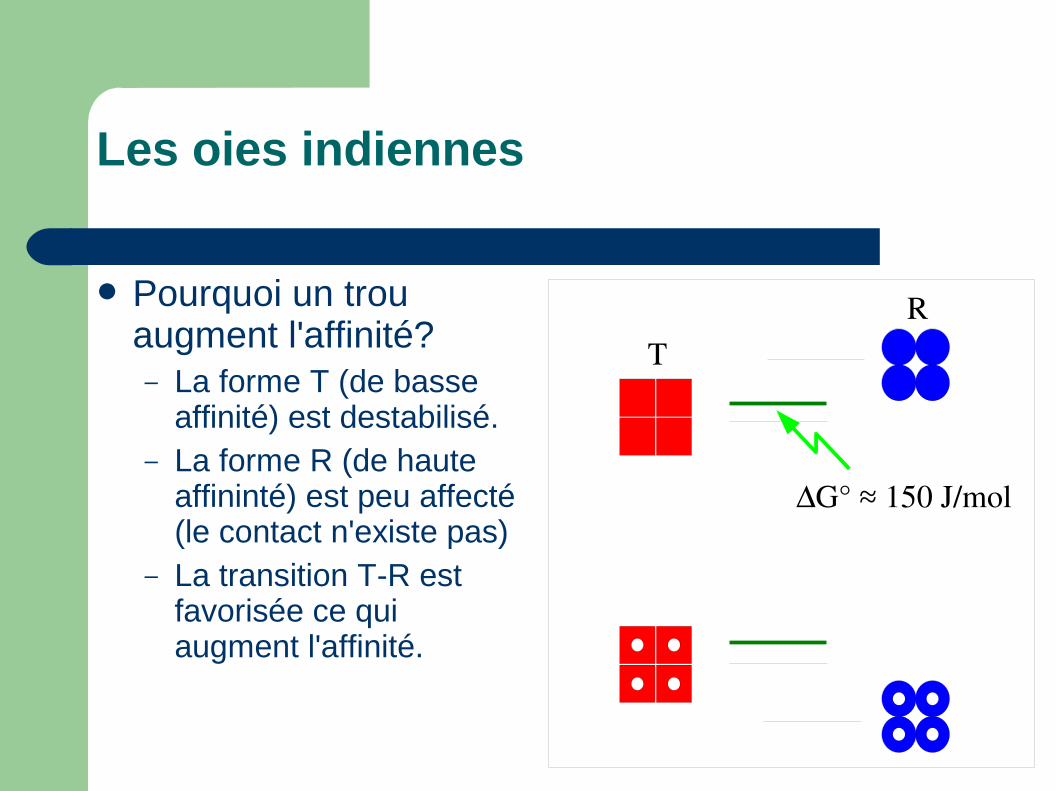
Les oies indiennes
TR
G° ≈ 150 J/mol
Pourquoi un trou augment l'affinité?– La forme T (de basse
affinité) est destabilisé.– La forme R (de haute
affininté) est peu affecté (le contact n'existe pas)
– La transition T-R est favorisée ce qui augment l'affinité.

Les oies indiennes
TR
G° ≈ 150 J/mol
La taille de l'effet est petit!– L'energie d'interaction de
deux CH2 est de
150J/mol– environ 25% d'une
liaison H.– mais cela modifie
l'affinité

Les oies indiennes
Une augmentation de l'affinité de l'hémoglobine de 0,8 mmHg (2,8 contre 2,0) est elle suffisante pour expliquer la modification d'habitat?
pas vraiment mais...
Dans le sang (grace aux modulateurs) les chiffres sont differentes:

Les oies indiennes
Anser anser 50mM Hg 8 000m
Anser indica 35mM Hg 12 000m
Meme effet energetique 150 J/mol ( 2*1,4 = 2,8 et 35 * 1,4 = 49 )
Dans le sang (grace aux modulateurs) les chiffres sont differentes:

Les oies andiennes
Dans une autre oie, l'oie andienne (Chloephaga melanoptera) une example frappante!
Cette oie qui vit a altitude a 16 modifications dans sa hémoglobine contre cela de l'oie cendrée.
Hiebl I, Braunitzer G, Schneeganss D.(1987)Biol Chem Hoppe Seyler. 368:15591569.

Les oies andiennes
Parmi les modifications il y a une qui touche le contact Leu
55-Pro
119
Cette fois c'est une mutation Leu
55Ser

Les oies andiennes
Suppression de deux CH3
Leucine Serine
CCH
O
N
CH2
H
OH
CCH
O
N
H3CCH
CH2
CH3
H
Encore 2 groupements methyl perdus.
Encore le même effet sur l'affinité d'oxygène!
Encore le même effet sur l'ecologie.

Les oies indiennes
L'étude de proches cousins peut donner des informations sur les liens séquence – écologie.
Un petit effet energétique (150J/mol) peut avoir des conséquences importants.
La majorité (75%-90%) des modifications ne sont pas important– ni pour la fonction de la
protéine – ni pour l'ecologie de
l'espèce.– ils sont neutres.

L'effet de Root
Les poissons téléosténiens ont une vessie natatoire - qu’ils remplissent avec l’oxygène.
Cet oxygène vient des globules rouges et de l’hémoglobine.
Comment les poissons des profondeurs peuvent remplir leur vessie natatoire contre une pression hydrostatique qui arrive à des centaines de atmosphères.
L’effet Root (variant de l’effet de Bohr).
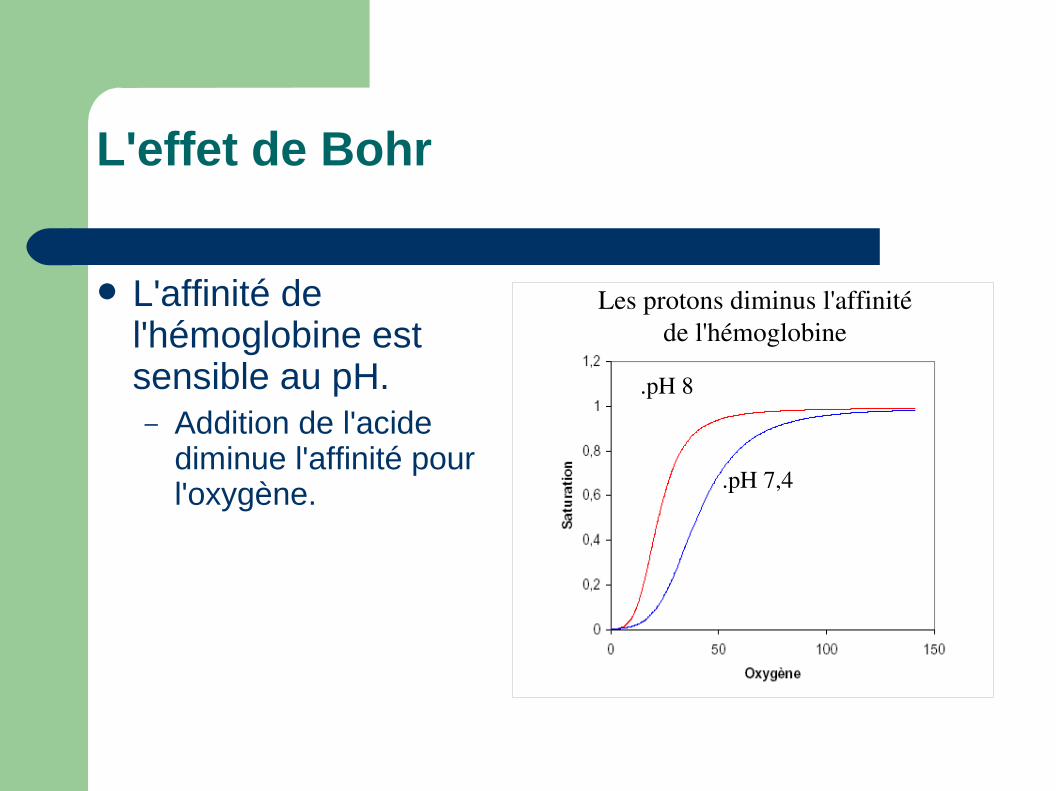
L'effet de Bohr
L'affinité de l'hémoglobine est sensible au pH.– Addition de l'acide
diminue l'affinité pour l'oxygène.
Les protons diminus l'affinité de l'hémoglobine
.pH 8
.pH 7,4

L'effet de Bohr.
les protons diminue l'affinité pour l'oxygène
reciproquement l'oxygène diminue l'affinité pour des protons.
Deux équilibres couplées. Pour Hb humaine 0,3H+
par O2.
Hb + 4O2
Hb1,24O2 + 1,2H+
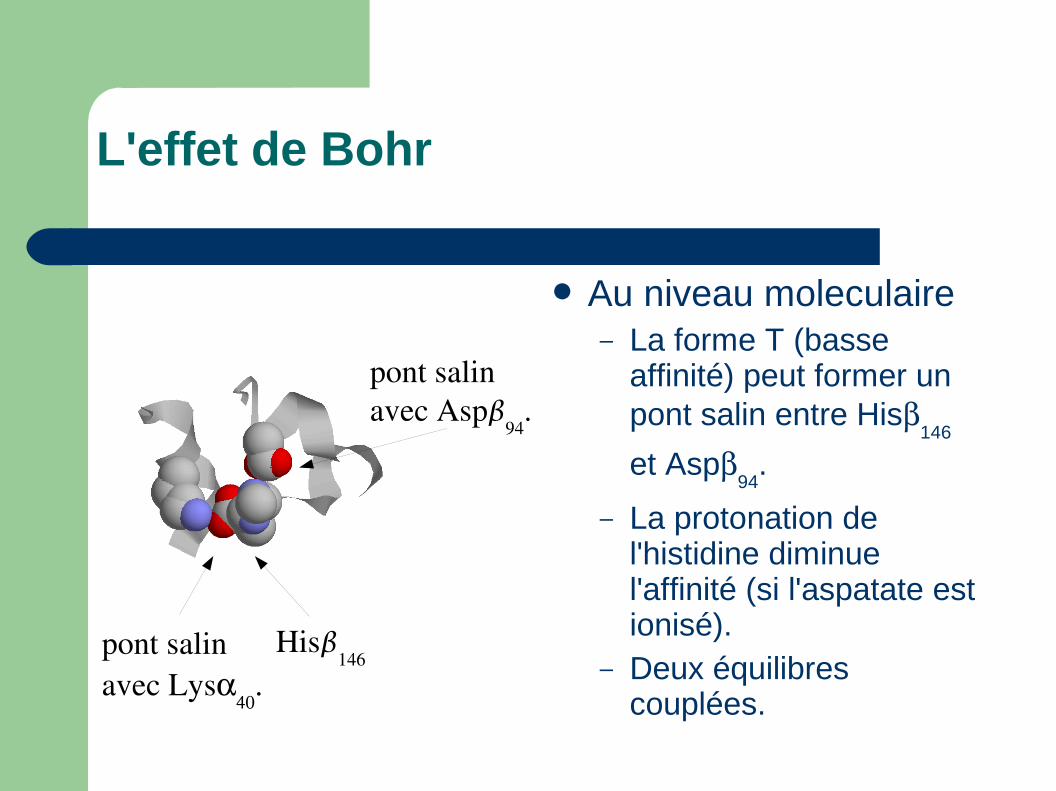
L'effet de Bohr
Au niveau moleculaire– La forme T (basse
affinité) peut former un pont salin entre His
146
et Asp94
.
– La protonation de l'histidine diminue l'affinité (si l'aspatate est ionisé).
– Deux équilibres couplées.
pont salinavec Lys
His146
pont salin avec Asp
94.
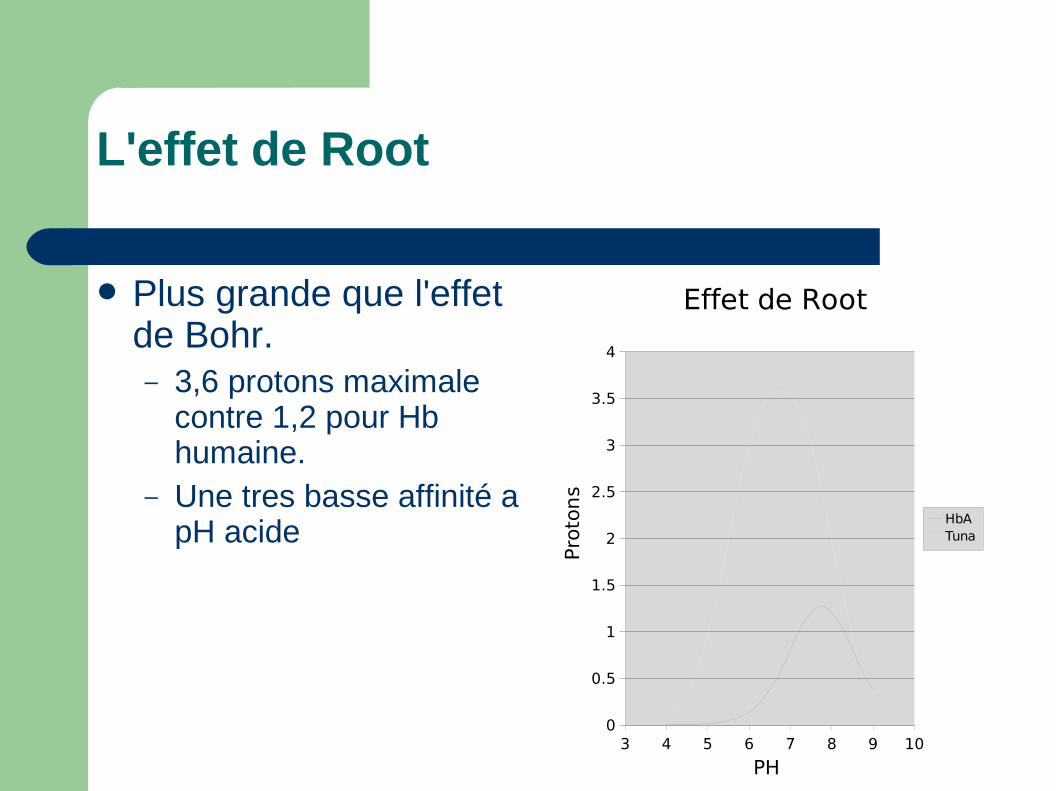
L'effet de Root
3 4 5 6 7 8 9 100
0.5
1
1.5
2
2.5
3
3.5
4
Effet de Root
HbATuna
PH
Pro
ton
s
Plus grande que l'effet de Bohr.– 3,6 protons maximale
contre 1,2 pour Hb humaine.
– Une tres basse affinité a pH acide
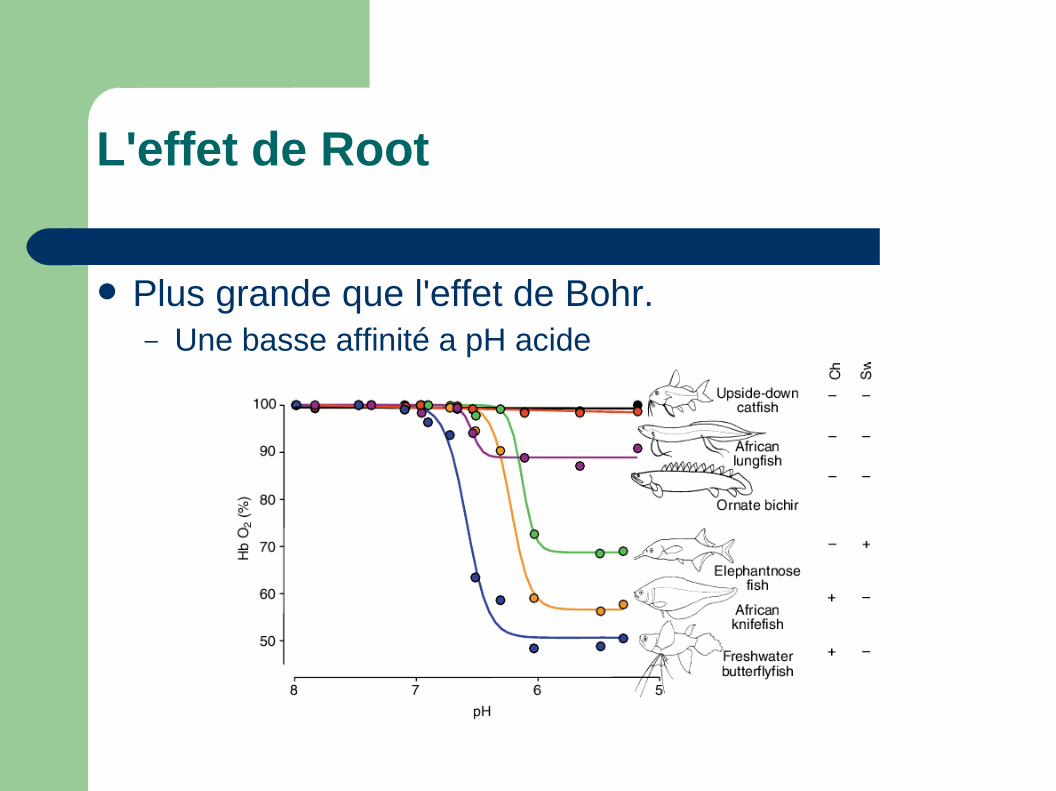
L'effet de Root
Plus grande que l'effet de Bohr.– Une basse affinité a pH acide

L'effet de Root
Plus grande que l'effet de Bohr.– Evolution vers 300 MYa

L'effet de Root
Les poissons pompent de l’oxygène avec de l’acide lactique.
Ils ont une structure spéciale (la rete) à côté de leur vessie natatoire ou ils accumulent suffisamment d’acide lactique pour remplir leur vessie!
Hb + 4 O2 ⇔ Hb.4O
2 + 3,6 H+
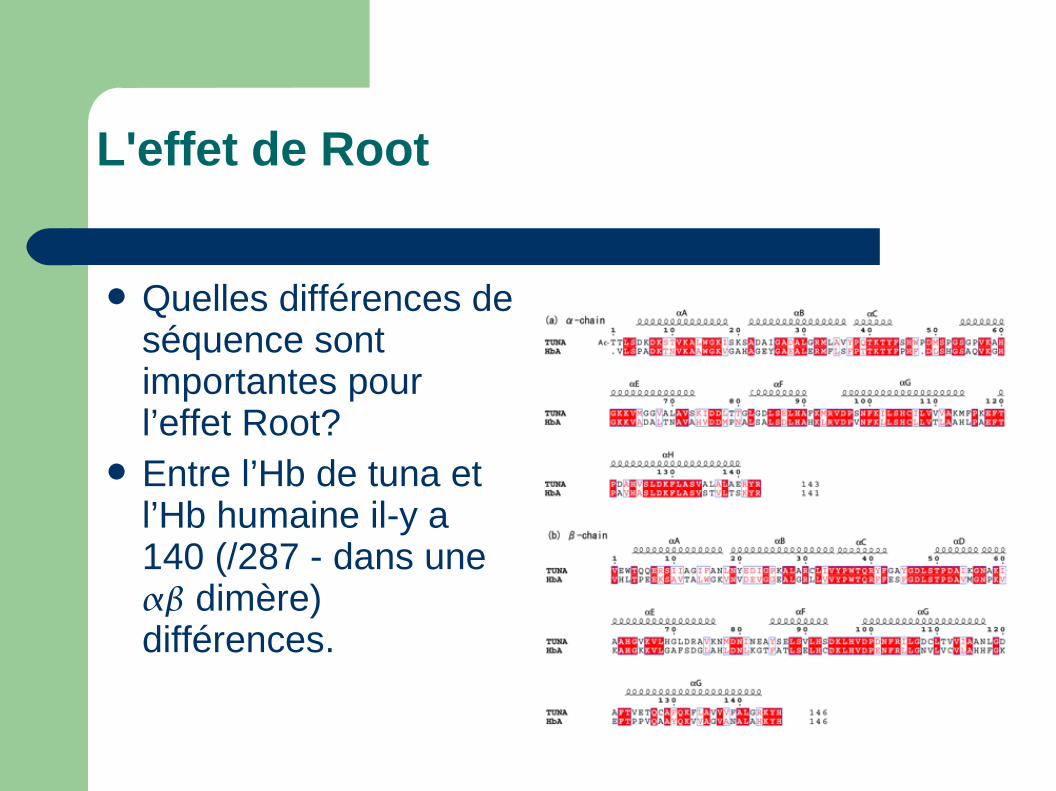
L'effet de Root
Quelles différences de séquence sont importantes pour l’effet Root?
Entre l’Hb de tuna et l’Hb humaine il-y a 140 (/287 - dans une dimère) différences.

L'effet de Root
Quels residus sont important pour l'effet de Root?
1ère idée : la substitution Cβ F9S (Pourquoi?)

L'effet de Root
Serine
CCH
O
N
CH2
H
OH
Cysteine
CCH
O
N
CH2
H
SH
Quels residus sont important pour l'effet de Root?
1ère idée : la substitution Cβ F9S (Pourquoi?)
Modification d'une atome S – O.

L'effet de Root
Serine
CCH
O
N
CH2
H
O
Cysteine
CCH
O
N
CH2
H
S
H H
NO
H
CH
His146 CH2
CO⊖
O
H
CH
His146 CH2
CO⊖
N
Différences entre S et O...– Angle optimale CSH
~180° et COH ~104°– O plus acide que S– Ser plus de liaisons H
qui sont plus forts.
Ces différences font que His
146 mieux lié à
la structure avec Ser.

L'effet de Root
Pourquoi cela modifie l'affinité et l'effet des protons?– Dans la forme T (basse
affinité) His146
peut faire une
pont salin avec Asp94
.
– Trois réactions liées:conformation T/R et affinité
protonation His146
et le pont
salin.
liaisons H His146
et Ser/CysF9
.
pont salinavec Lys
His146
pont salin avec Asp
94.

L'effet de Root
Pourquoi cela modifie l'affinité et l'effet des protons?– Dans la forme T (basse
affinité) His146
peut faire une
pont salin avec Asp94
.
– Trois réactions liées:conformation T/R et affinité
protonation His146
et le pont
salin.
liaisons H His146
et Ser/CysF9
.
A pH acide le His146
protoné
peut plus facilement trouver l'Asp
94 et basculer
l'oxyhémoglobine en forme T qui est de basse affinité et perd l'oxygène.

L'effet de Root
Test Ser/Cys modification

L'effet de Root
Un échange très conservateur (C→S) d’acide aminé dans une site clé peut changer, d'une façon importante, le comportement physico-chimique d’une protéine.
Malheureusement il n'est pas suffisant pour donner un effet de root a l'hemoglobine humaine.

L'effet de Root
Quels residus sont important pour l'effet de Root?
1ère idée : la substitution Cβ F9S (Pourquoi?)
2ème idée : un pont salin additionelle en forme T.– His
69-Asp
72.
– Encore plus d'effet de pH.

L'effet de Root
Test extra salt bridge His-Asp

Quels residus sont important pour l'effet de Root?
1ère idée : la substitution Cβ F9S
2ème idée : un pont salin additionelle en forme T.
L'effet de Root
3ème idée : peut-être en plus des changements subtils dans la site de liaison d'oxygène.
4ème idée : plusieurs mechanismes avec roles variables selon l'espece.

Stabilisation de la position de His
146 dans la forme T.
Diminution du nombre d'histidines de surface.
Destabilisation de la forme R (groupes de charge).
Liaison H additionnelles dans la forme T.– Asp de surface .
L'effet de Root

Conclusions
L'effet de Root moins claire que l'histoire des oies.– 140 modifications à
analyser.
Des petits changements peuvent être importants– Ser/Cys– Residus similaires
Souvent plusieurs modifications synergiques sont impliquées.

Variation de séquence II
Les variations au sein d'une espèce

Variations au sein d'une espèce
L'espèce humaine, car plus d'information.
Deux types de variation...– Les variations au cours
du développement.– Les differentes alleles
des gènes et la variabilité naturelle.

Variations dans l'espèce
22
22
22
22
22
22
Au cour du développement normal 6 différentes hémoglobines sont fabriquées
chaqu'une a un moment différent dans le développement.

Hémoglobine foetale
P50 Nom chaines 4 Torr Gower I 22
12 Torr Gower II 22
6 Torr Portland 22
20 Torr Foetal 22
26 Torr Adult 22
Les différentes formes ont des propriétés différentes.
L'embryon et le foetus ont une grande besoin en oxygène.

Hémoglobine foetale
VHLTPEEKSA VTALWGKVNV DEVGGEALGR GHFTEEDKAT ITSLWGKVNV EDAGGETLGR
LLVVYPWTQR FFESFGDLST PDAVMGNPKV LLVVYPWTQR FFDSFGNLSS ASAIMGNPKV
KAHGKKVLGA FSDGLAHLDN LKGTFATLSE KAHGKKVLTS LGDAIKHLDD LKGTFAQLSE
LHCDKLHVDP ENFRLLGNVL VCVLAHHFGK LHCDKLHVDP ENFKLLGNVL VTVLAIHFGK
EFTPPVQAAY QKVVAGVANA LAHKYH EFTPEVQASW QKMVTAVASA LSSRYH
Entre les chaines et il y a 40 différences de séquence.

Hémoglobine foetale
Entre les chaines et il y a 40 différences de séquence.– la moitié sont des
modifications sur la surface (peu important?)
VHLTPEEKSA VTALWGKVNV DEVGGEALGR GHFTEEDKAT ITSLWGKVNV EDAGGETLGR
LLVVYPWTQR FFESFGDLST PDAVMGNPKV LLVVYPWTQR FFDSFGNLSS ASAIMGNPKV
KAHGKKVLGA FSDGLAHLDN LKGTFATLSE KAHGKKVLTS LGDAIKHLDD LKGTFAQLSE
LHCDKLHVDP ENFRLLGNVL VCVLAHHFGK LHCDKLHVDP ENFKLLGNVL VTVLAIHFGK
EFTPPVQAAY QKVVAGVANA LAHKYH EFTPEVQASW QKMVTAVASA LSSRYH

Hémoglobine foetale
Entre les chaines et il y a 40 différences de séquence.– la moitié sont des
modifications sur la surface (peu important?)
– deux mutations dans les 4 résidus qui interagissent avec le modulateur 2,3-BPG V
1
et H143
.
VHLTPEEKSA VTALWGKVNV DEVGGEALGR GHFTEEDKAT ITSLWGKVNV EDAGGETLGR
LLVVYPWTQR FFESFGDLST PDAVMGNPKV LLVVYPWTQR FFDSFGNLSS ASAIMGNPKV
KAHGKKVLGA FSDGLAHLDN LKGTFATLSE KAHGKKVLTS LGDAIKHLDD LKGTFAQLSE
LHCDKLHVDP ENFRLLGNVL VCVLAHHFGK LHCDKLHVDP ENFKLLGNVL VTVLAIHFGK
EFTPPVQAAY QKVVAGVANA LAHKYH EFTPEVQASW QKMVTAVASA LSSRYH

Hémoglobine foetale
La liaison de 2,3-BPG dépend des interactions ioniques avec:– NH
3
+-Val1
– His2
+
– Lys82
+
– His143
+ - Ser

Hémoglobine foetale
Normalement le 2,3-BPG lie à l'hémoglobine et diminue sa affinité pour l'oxygène.
La HbF lie le 2,3-BPG moins bien.
Ce qui stabilise la forme R (haute affinité).
Il se lie entre les deux sousunités dans la forme T (basse affinité) mais
pas dans la forme R
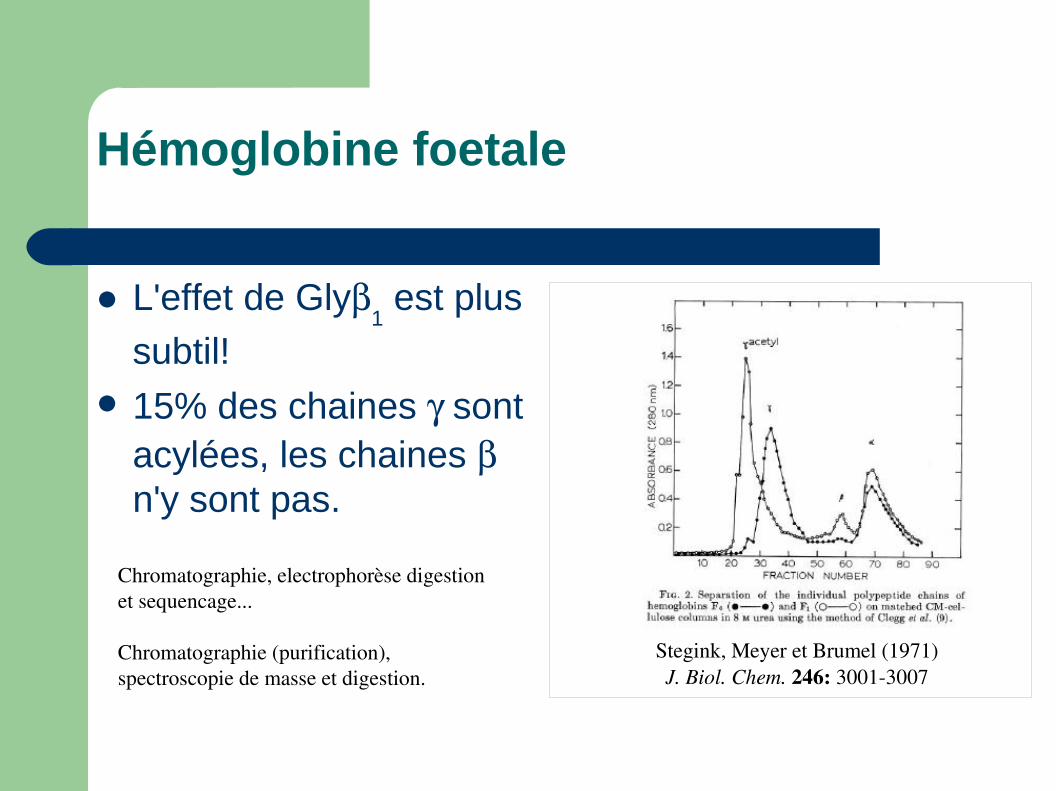
Hémoglobine foetale
Stegink, Meyer et Brumel (1971) J. Biol. Chem. 246: 30013007
Chromatographie, electrophorèse digestion et sequencage...
Chromatographie (purification), spectroscopie de masse et digestion.
L'effet de Gly1 est plus
subtil! 15% des chaines sont
acylées, les chaines n'y sont pas.

Hémoglobine foetale
Acetyl-Gly1 n'est plus
chargé. Val est moins flexible et
évite l'acetylation. Exemple d'une
différence qui agisse par les modifications post-translationelles

Hémoglobine foetale
Normalement 3% de hemoglobine (HbA) en forme CO et specificity (CO/O2) environ 250.Pour hème sans protéine specificité est vers 10,000Pour HbF la spécicitité est diminuée vers 80 et malgré ca +10% en forme CO.
Polarité de poche de liaisonLiaison H avec Histidine

Variations dans l'espèce
Environ 400 mutations ponctuelles
HbS (β G6→V)
Hb Rayleigh (β V1→A)
Hb Hiroshima (β H146
→D)
Hb Rouen (α Y140
→H)
Une grand nombre de mutations ponctuelles sont connues.

Hb S
Peut-etre la premiere maladie associé avec une protéine mutante.
Une allele avec une grande pénetration grace aux avantages de la mutation.
Associé avec une résistance au paludisme

Maladié génétique
Décrite par James Herrick en 1904.
Les globules rouges d'une forme particulière “falciformes”.
Cause le “drépanocytose”

Maladie génétique
Les malades (homozygotes) peuvent avoir environ 50% d'erythrocytes falciformes dans leur sang veineux, mais pas arterial.
Pour les porteurs (heterozygotes) seulement environ 1% d'erythrocytes falciformes dans leur sang veineux.
Grande pénétration du gène dans certains populations, environ 11%.– 1% homozygotes et 10% héterozygotes.

Distribution d'HbS

Paludisme et HbS
Le paludisme est peut-être le plus grande influence sur l'évolution humaine!
Les liens entre HbS, la physicochimie et le paludisme.
D'autres mutations.

Paludisme
Le paludisme tue encore plus de 1M personnes par an, essentiellement les enfants.
Très important dans l'évolution de l'espece humaine.
Ils existent plusieurs mutations qui protègent partiellement– Thallesimies– Drepanocytose et autres
mutations ponctuelles– Glucose-6-phosphate
deshydrogenase.
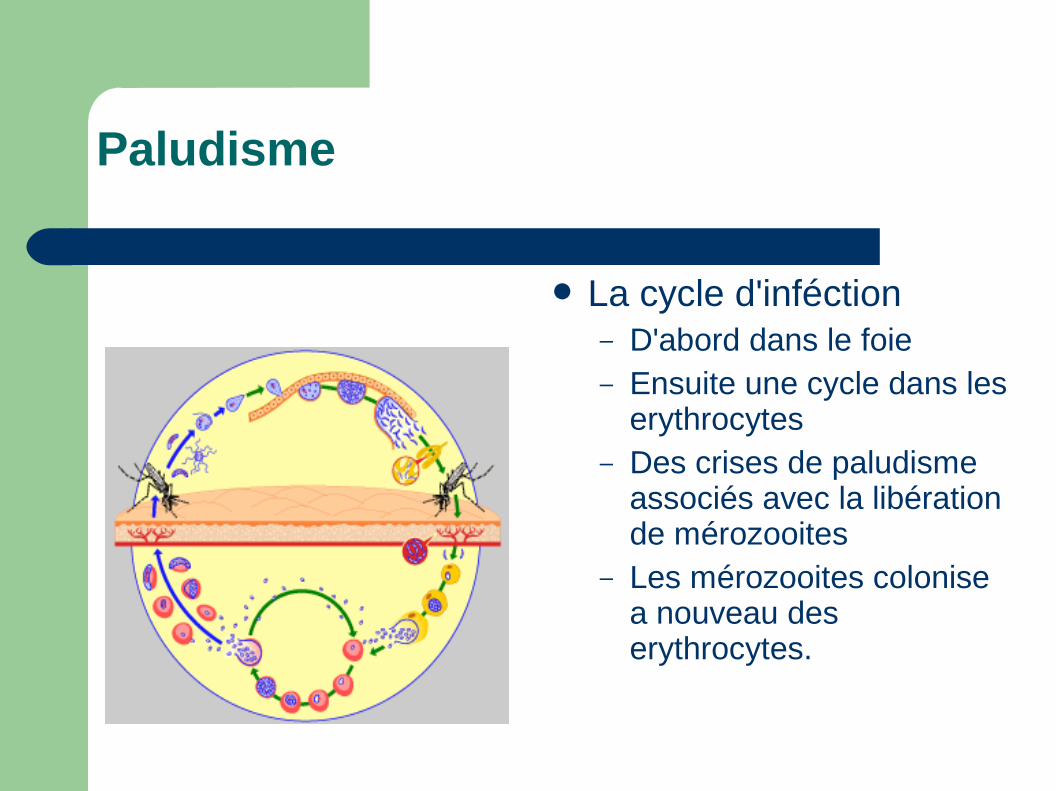
Paludisme
La cycle d'inféction– D'abord dans le foie– Ensuite une cycle dans les
erythrocytes– Des crises de paludisme
associés avec la libération de mérozooites
– Les mérozooites colonise a nouveau des erythrocytes.
Paludisme
La maturation des mérozooites dans les erythrocytes se passent dans les tissus pour eviter l'immunosurveillance de la rate.

Hb S
Quelle est la mutation? Comment la mutation
engendre t'elle le phénotype?
Quelle est l'origine de l'avantage contre le paludisme?

Hb S
Quelle est la mutation?– Ca change la charge de
la protéine.– Les deux gènes sont
exprimés dans des héterozygote.
Origine
HbS
HbA+
Norm
ale
Porteur
Malade
Linus Pauling en 1949 à trouvé une différence de pI entre l ’HbA et l ’HbS

Hb S
Quelle est la mutation?– Ca change la charge de
la protéine.– Les deux gènes sont
exprimés dans des héterozygote.
– Glu6Val
HbA ValHisLeuThrProGluGluLysHbS ValHisLeuThrProValGluLys
Glutamate Valine
CCH
O
N
H3CCH
CH3
HC
CH
O
N
O
CH2
CH2
C
H
O⊖
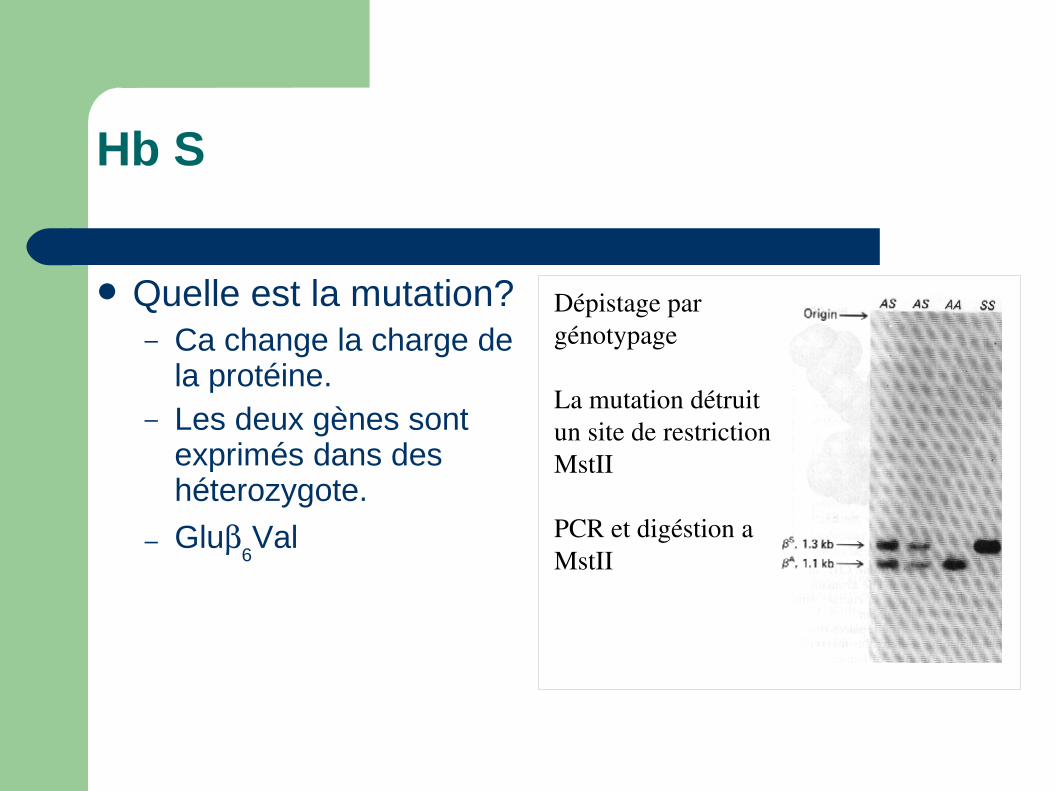
Hb S
Quelle est la mutation?– Ca change la charge de
la protéine.– Les deux gènes sont
exprimés dans des héterozygote.
– Glu6Val
Dépistage par génotypage
La mutation détruit un site de restriction MstII
PCR et digéstion a MstII

Hb S
Quelle est la mutation?– Ca change la charge de
la protéine.– Les deux gènes sont
exprimés dans des héterozygote.
– Glu6Val
– Sur la surface de la protéine.
Une mutation de surface (peu importante)

HbS
Comment la mutation cause les erythrocytes falciformes?– L'Hb est tres concenté dans
des globules rouges. (5mM ou 320mg/ml)
– L'HbS est beaucoup moins soluble en particulier en forme desoxygène (35mg/ml)
– La solubilité dépend de la surface de la protéine.

HbS
Comment la mutation cause les erythrocytes falciformes?– Problème de solubilité
de la forme desoxy.– Formation de fibres.
From G. Rykes, R.H. Crepeau, and S.J. Edelstein. Nature 272(1978):509.

HbS
Comment la mutation cause les erythrocytes falciformes?– Problème de solubilité
de la forme desoxy.– Formation de fibres.– Les fibres déforment les
cellules dans la circulation veineux.
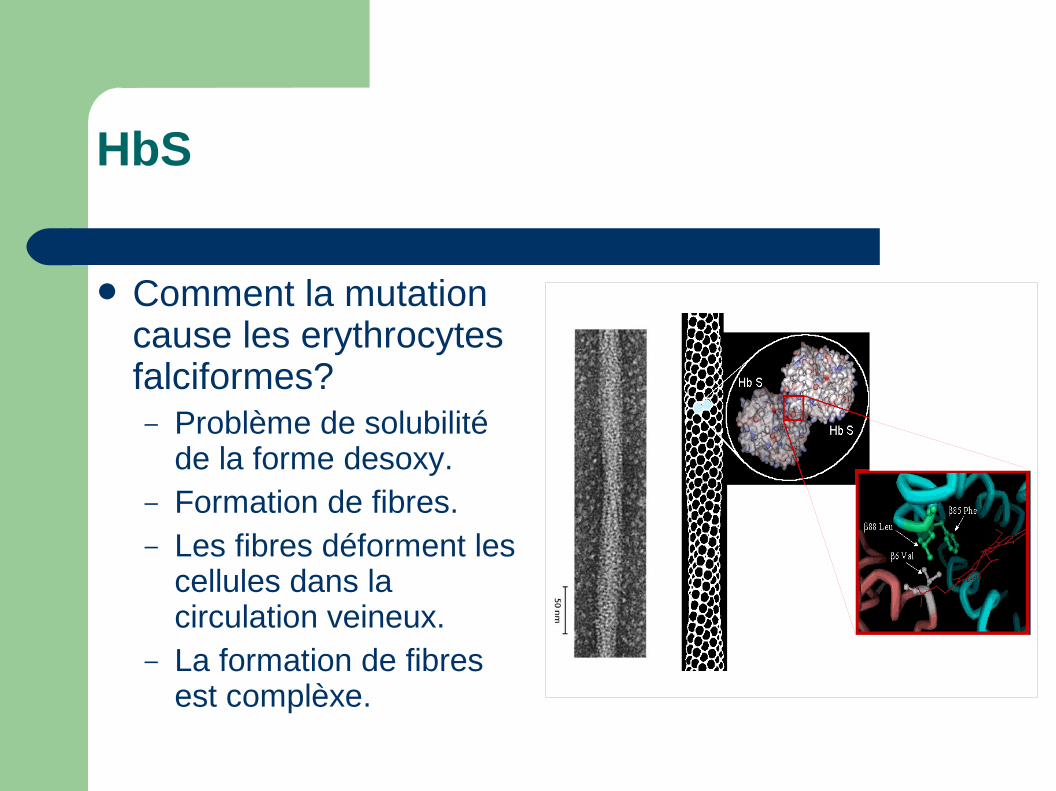
HbS
Comment la mutation cause les erythrocytes falciformes?– Problème de solubilité
de la forme desoxy.– Formation de fibres.– Les fibres déforment les
cellules dans la circulation veineux.
– La formation de fibres est complèxe.
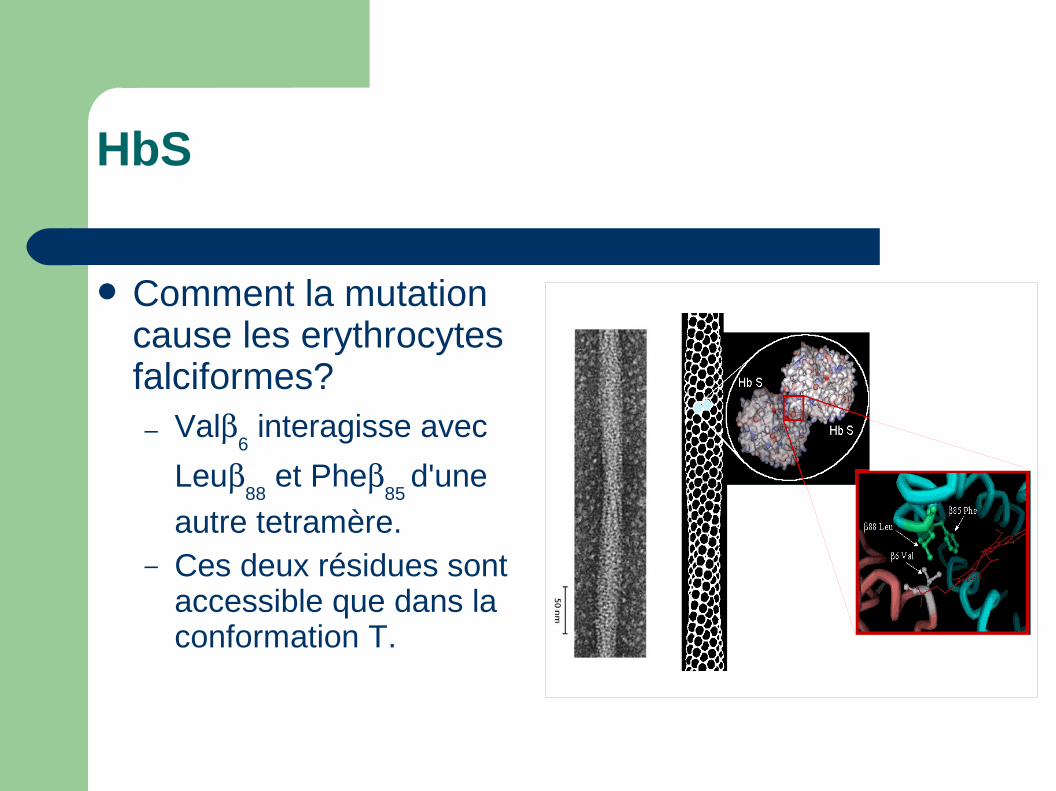
HbS
Comment la mutation cause les erythrocytes falciformes?– Val
6 interagisse avec
Leu88
et Phe85
d'une
autre tetramère.– Ces deux résidues sont
accessible que dans la conformation T.
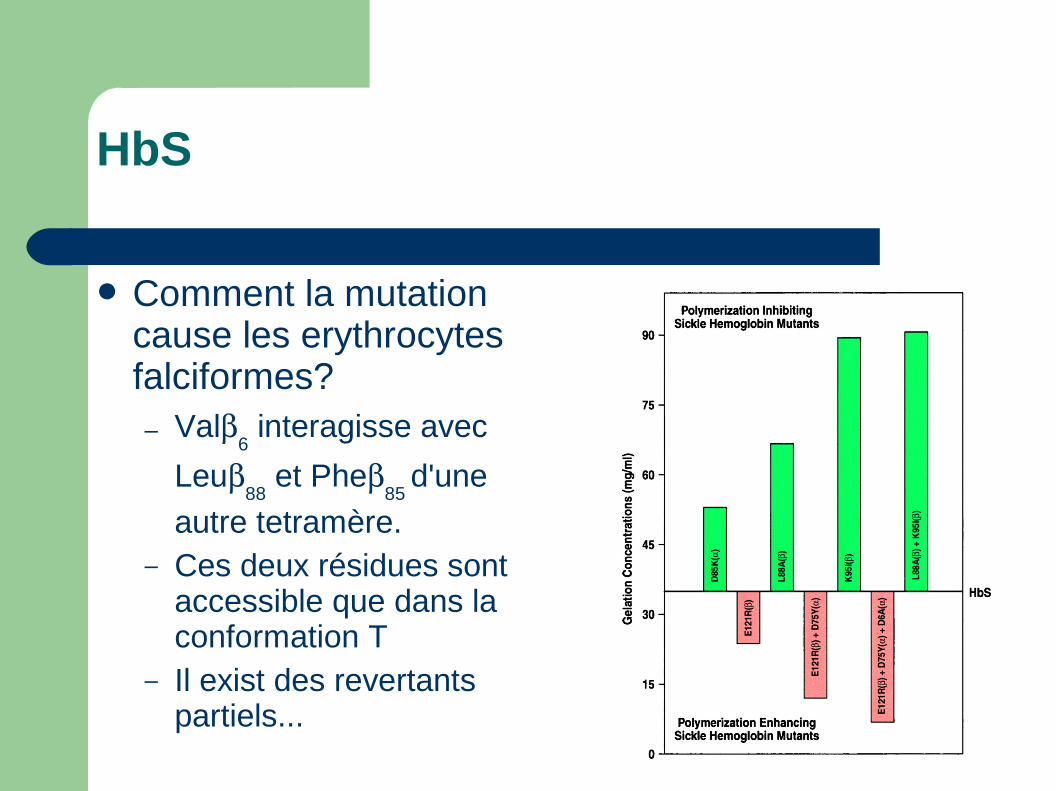
HbS
Comment la mutation cause les erythrocytes falciformes?– Val
6 interagisse avec
Leu88
et Phe85
d'une
autre tetramère.– Ces deux résidues sont
accessible que dans la conformation T
– Il exist des revertants partiels...
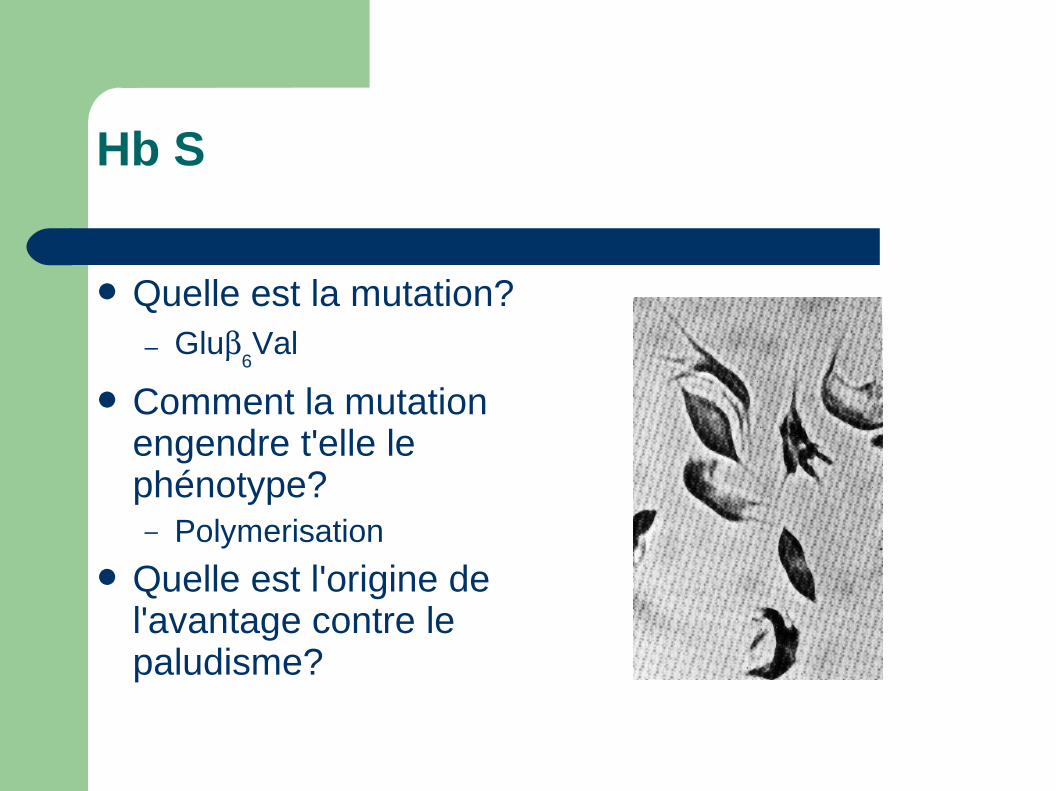
Hb S
Quelle est la mutation?– Glu
6Val
Comment la mutation engendre t'elle le phénotype?– Polymerisation
Quelle est l'origine de l'avantage contre le paludisme?

Hb S
Quelle est l'origine de l'avantage contre le paludisme?– A une basse
concentration d'oxygène l'HbS empeche la survie de la parasite.
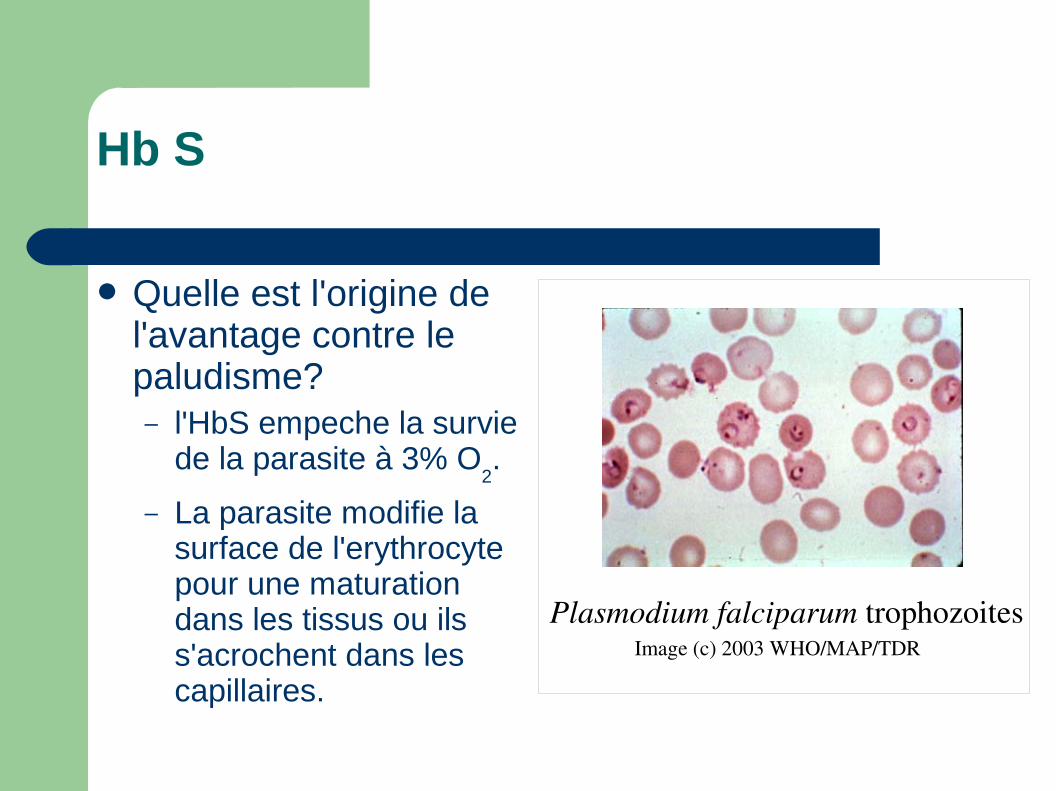
Hb S
Quelle est l'origine de l'avantage contre le paludisme?– l'HbS empeche la survie
de la parasite à 3% O2.
– La parasite modifie la surface de l'erythrocyte pour une maturation dans les tissus ou ils s'acrochent dans les capillaires.
Image (c) 2003 WHO/MAP/TDR
Plasmodium falciparum trophozoites

HbS
Une erythrocyte infectée s'accroche dans une tissue.
La basse concentration d'O2 cause l'hémoglobine à
se desoxygèner. L'HbS n'est plus soluble et elle forme des fibres
– ce qui entraine encore plus de desoxygèneation et polymerisation (equilibres couplées)
La deformation des membranes cause une perte de K+ intracellulaire
Ce qui tue les parasites.

HbS
Dans homozygote la deformation est rapide et donc est visible dans le sang.
Dans l'héterozygote, le processus est plus lente et donc est limité SAUF dans les ertythrocytes accrochés dans les tissus.

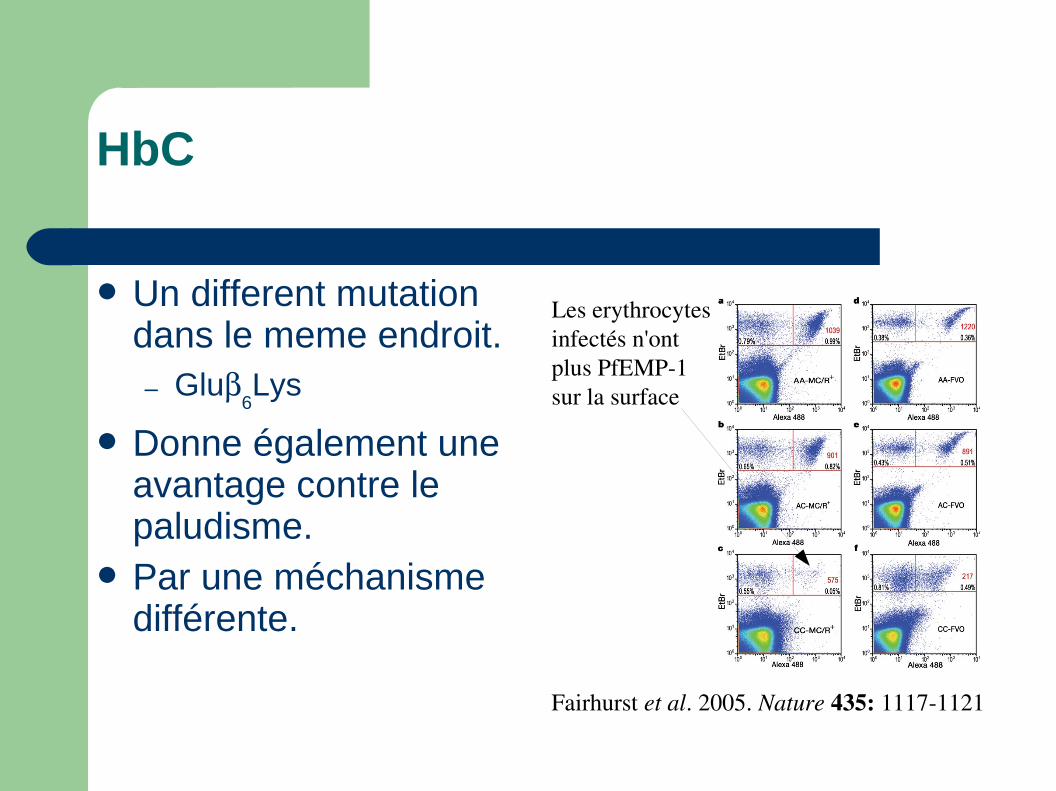
HbC
Un different mutation dans le meme endroit.– Glu
6Lys
Donne également une avantage contre le paludisme.
Par une méchanisme différente.
Fairhurst et al. 2005. Nature 435: 11171121
Les erythrocytes infectés ne collent plus dans les tissus...
HbC
Un different mutation dans le meme endroit.– Glu
6Lys
Donne également une avantage contre le paludisme.
Par une méchanisme différente.
Fairhurst et al. 2005. Nature 435: 11171121
Les erythrocytesinfectés n'ont plus PfEMP1 sur la surface

HbC
Donne également une avantage contre le paludisme.– Les erythrocytes ne
montrent plus PfEMP-1 correctement sur la surface.
– Ils ne peuvent plus s'accrocher dans les tissus.
Fairhurst et al. 2005. Nature 435: 11171121
Les erythrocytesinfectés n'ont plus PfEMP1 sur la surface...
et les heterozygotes ont une morphologie différente.

HbC
Par une méchanisme différente.– Les erythrocytes viellissent plus vite, parceque
l'hémoglobine est moins stable.– L'hémoglobine dénaturé largue son héme qui
s'accrochent sous le membane.– Ce qui
diminue le temps de vie des erythrocytes.modifie l'organisation de PfEMP-1 en diminuant son efficacité et
augmentant son antigénicité.

Conclusions
Effets de mutation parfois indirecte (modification).
Multiplicité de fonctions d'une protéine O
2 et
CO. Les modifications de
surface ne sont pas tous anodin.
Divers aspets de la physicochimie sont important pour lié la séquence et la physiologie.– solubilité/precipitation– stabilité/denaturation– flexibilité/rigidité– charge et polarité

Variations de séquence III
Le hémoglobine d'Ascaris suum

Ascaris suum
Hémoglobine P50
(mmHg)
Ascaris suum 0,0072Hbα 0,82HbA 21,4Mb 15,5
Certaines hémoglobines ont une énorme affinité pour l'oxygène.– Quelles sont les origines
moléculaires?– Pourquoi une telle
affinité?

Ascaris suum
Hémoglobine P50
(mmHg)
Ascaris suum (O2) 0,0072
Ascaris suum (CO) 0,35Mb (O
2) 15,5
Mb (CO) 0,55
La haute affinité est spécifique à l'O
2
– L'affinité pour CO n'est guerre changée.

Ascaris suum
La haute affinité est spécifique à l'O
2
– L'affinité pour CO n'est guerre changée.
– Elle est du en grande partie a une dissociation très lente (1000x)
Hémoglobine kon
(M1s1) koff
(s1)
Ascaris suum 2,8 0,013Mb 15,0 14,0HbA 40 50

Ascaris suum
Myoglobin GLSDGEWQLVLNVWGKVEA-Ascaris ---MATACVKSLESVQCGTC
Myoglobin ---DIPGHGQEVLIRLFKGHAscaris EKTIANGTEFYALLFDKHPD
Myoglobin PETLEKFDKFKHLKSEDEMKAscaris LRHYFKGNENLTGADVKKSD
Myoglobin ASEDLKKHGATVLTALGGILAscaris HFKKQGQRLLLACHVLAHLE
Myoglobin KKKGH---HEAEIKPLAQSHAscaris NDPASFKAYAREI---VDPH
Myoglobin AT-KHKIPVKYLEFISECIIAscaris LRMSVHLEPKLWSEFWPIWL
Myoglobin QVLQSKHPGDFGADAQGAMNAscaris DYLSTKES-V-DDATKNAWL
Myoglobin KALELFRKDMASNYKELGFQGAscaris ALGKKFSDECLDHLKNLGQPH
Une chaîne unique (comme Mb - myoglobine)
Tellement de différences de séquence que :– une alignement tres
difficile (8% d'identité)– l'algnement nous aide
peu.

Ascaris suum
Comme monomérique pas de problème d'allostérie...– Il faut regarder dans la
site de liaison pour l'explication moléculaire de la haute affinité.

Une approche biophysique
Le spectra d'absorption de l'hémoglobine– Bandes Soret, et – Formes oxy (Fe2+),
desoxy et oxydée (Fe3+)– Formes haut-spin et bas
spin.
Le spectre d'absorption des hémoglobines contient plusieurs pics. La forme du spectre nous renseigne sur le Fer et son environnement.

Une approche biophysique
Le spectra d'absorption de l'hémoglobine– Bandes Soret, et – Formes oxy (Fe2+),
desoxy et oxydée (Fe3+)– Formes haut-spin et bas
spin.
Le hémoglobine peut exister dans deux états d'oxydation chacun avec diverses ligands.

Une approche biophysique
Le spectra d'absorption de l'hémoglobine– Bandes Soret, et – Formes oxy (Fe2+),
desoxy et oxydée (Fe3+)– Formes haut-spin et bas
spin.
Selon les ligands le fer peut se trouver dans une état basse spin (ligands forts doublet) ou haut spin (ligands faibles singlet).
Haut spin 4/2 Bas spin (0)Eg
Tg
Approche Biophysique
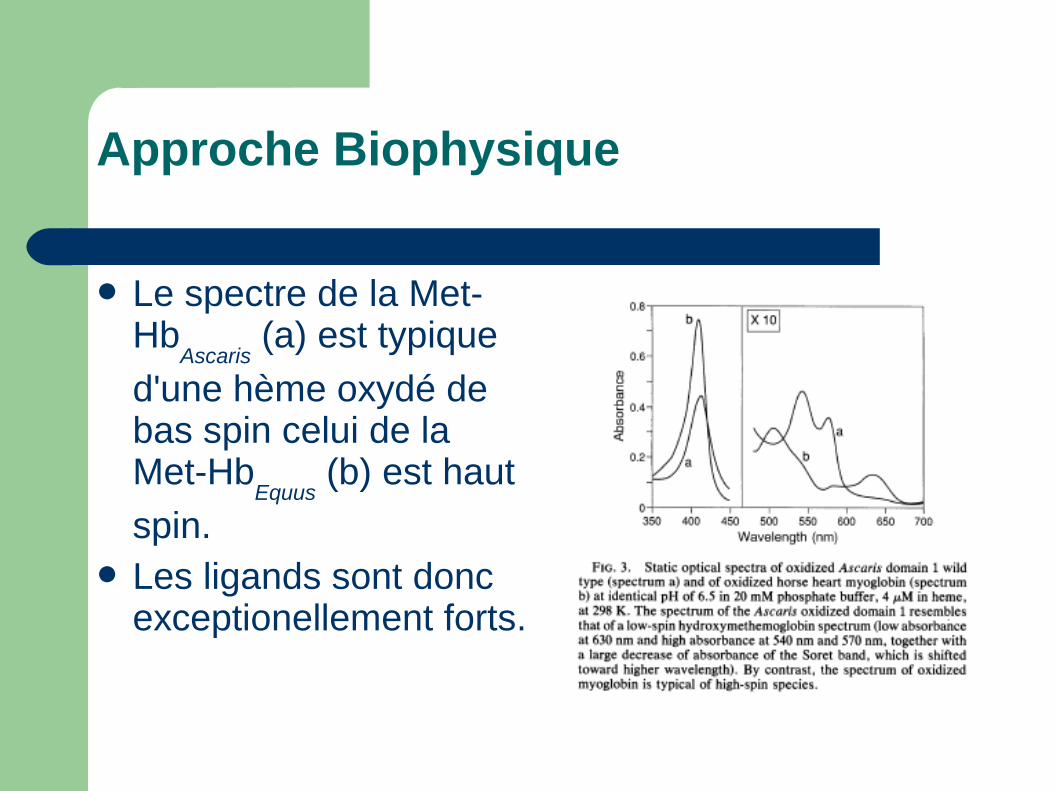
Le spectre de la Met-Hb
Ascaris (a) est typique
d'une hème oxydé de bas spin celui de la Met-Hb
Equus (b) est haut
spin. Les ligands sont donc
exceptionellement forts.

Une hypothèse
Il faut chercher des différences autour de la site de liaison qui vont augmenter la force de liaison du ligand axiale...
Myoglobin GLSDGEWQLVLNVWGKVEA-Ascaris ---MATACVKSLESVQCGTC
Myoglobin ---DIPGHGQEVLIRLFKGHAscaris EKTIANGTEFYALLFDKHPD
Myoglobin PETLEKFDKFKHLKSEDEMKAscaris LRHYFKGNENLTGADVKKSD
Myoglobin ASEDLKKHGATVLTALGGILAscaris HFKKQGQRLLLACHVLAHLE
Myoglobin KKKGH---HEAEIKPLAQSHAscaris NDPASFKAYAREI---VDPH
Myoglobin AT-KHKIPVKYLEFISECIIAscaris LRMSVHLEPKLWSEFWPIWL
Myoglobin QVLQSKHPGDFGADAQGAMNAscaris DYLSTKES-V-DDATKNAWL
Myoglobin KALELFRKDMASNYKELGFQGAscaris ALGKKFSDECLDHLKNLGQPH
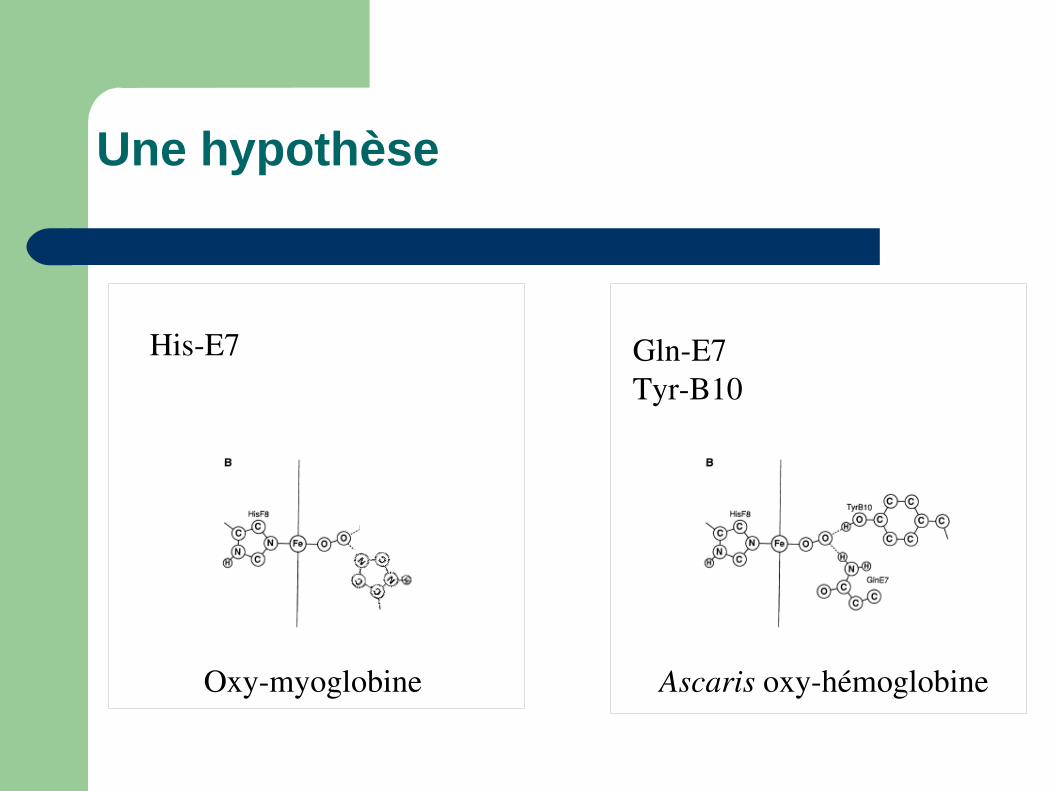
Une hypothèse
HisE7 GlnE7TyrB10
Oxymyoglobine Ascaris oxyhémoglobine

Une hypothèse
Le résidu TyrB10 estil important?Mutagenèse dirigée:
Hémoglobine P50
(mmHg) kon
(M1s1) koff
(s1)
Ascaris suum 0,0072 2,8 0,013B10 YL 8,2 9,0 5,0B10 YF 0,6 40,3 2
Oui, ce résidu est très important pour l'affinité.

Cristallographie– Très haute resolution
avec Rayons X ou neutron crysallographie.
– Tres longue et souvent couteux (difficile d'avoir des cristaux adequate et des neutrons)
– Peu d'information– Interpretation facile.
Liaisons Hydrogènes
Comment voir des liaisons hydrogène et éstimer leur importance?– Cristallographie– Spectroscopie
vibrationelle– Echange H/D ou H/T

Spectroscopie vibrationelle– La frèquence des
vibrations dépend des liaisons H.
– Spectres compliques– Interpretation difficile– Information sur la force
des liaisons.
Liaisons Hydrogènes
Comment voir des liaisons hydrogène et éstimer leur importance?– Cristallographie– Spectroscopie
vibrationelle– Echange H/D ou H/T

Echange H/D (rmn)– Les liaisons hydrogène
ralentissent l'échange H/D.
– Information structurale simple
– Calibration difficile en force.
– Assignation des resonances difficile.
Liaisons Hydrogènes
Comment voir des liaisons hydrogène et éstimer leur importance?– Cristallographie– Spectroscopie
vibrationelle– Echange H/D ou H/T

Spectroscopie vibrationelle
Mesures les fréquences de vibration d'une molécule.– Depend de la masse des
atomesplus lourde plus lente
– Depend de la force du liaison.plus forte plus rapide
C O

Spectroscopie vibrationelle
Mesures les fréquences de vibration d'une molécule.– Beaucoup de frequences
3n6 fréquences ou n = nombre d'atomes dans la molécule (H inclus) pour une haemoglobine monomerique environ 8000!

Spectroscopie vibrationelle
Mesures les fréquences de vibration d'une molécule.– Beaucoup de frequences– Simplification
Spéctoscopie differentielleMarquage IsotopiqueSpectroscopies Raman
– CARRS ou– Raman de résonance.
Compare deux spectres similaires par exemple une enzyme sans et avec substrat.
Seul les choses qui changent sont visibles par exemple la site active.

Spectroscopie vibrationelle
Mesures les fréquences de vibration d'une molécule.– Beaucoup de frequences– Simplification
Spéctoscopie differentielleMarquage IsotopiqueSpectroscopies Raman
– CARRS ou– Raman de résonance.
Mettre une ou des atomes de masse different dans des endroits clés : D; O18 ou N14. Ce qui donneront des fréquences specifiques
On peut coupler ca avec la spectroscopie differentielle (compare O16 et O18).

Spectroscopie vibrationelle
Mesures les fréquences de vibration d'une molécule.– Beaucoup de frequences– Simplification
Spéctoscopie differentielleMarquage IsotopiqueSpectroscopies Raman
– CARRS ou– Raman de résonance.
Spectroscopie Raman (prix Nobel 1930)

Spectroscopie vibrationelle
Mesures les fréquences de vibration d'une molécule.– Beaucoup de frequences– Simplification
Spéctoscopie differentielleMarquage IsotopiqueSpectroscopies Raman
– CARRS ou– Raman de résonance.
Spectroscopie Raman diffusion inélastique des photons
Absorption IR
Diffusion Raman
ExcitationDiffusion

Spectroscopie vibrationelle
Mesures les fréquences de vibration d'une molécule.– Beaucoup de frequences– Simplification
Spéctoscopie differentielleMarquage IsotopiqueSpectroscopies Raman
– CARRS ou– Raman de résonance.
Spectroscopie Raman Sélection possible...
Absorption IR
Diffusion Raman
ExcitationDiffusion
Exhaltation de l'effet (106x) si :●proche d'une surface●excitation correspond à une absorption●ou certains autres conditions

Methode choisie
Dans les exemples qui suit les auteurs ont utilisé de la spectroscopie Raman de résonance différentielle avec marquage isotopique!!! Pour regarder l'importance des liaisons H dans l'hemoglobine d'Ascaris suum.
Une spectroscopie vibrationelle qui permet d'observer la fréquence de vibration de certaines liaisons.
Une méthode de sélection qui permet de ne regarder que les vibrations associées avec un chromophore (hème ou tyrosine)
Une méthode de sélection qui permet de ne regarder que les vibrations sensibles à un changement (par exemple de ligand)
Une méthode de sélection qui permet d'attribuer des vibrations à certains atomes ou groupes

Ascaris suum – Raman spectroscopy
Avec une laser UV on peut regarder spécifiquement les tyrosines et tryptophanes.– Les différentes bandes W16 ou
Y9a sont attribué a des vibrations spécifique de chaque résidue
– Les spectres avec “rien”, O2 et
CO sont comparés

Ascaris suum – Raman spectroscopy
Avec une laser UV on peut regarder spécifiquement les tyrosines et tryptophanes.– Les spectres de différence
indiquent une interaction spécifique entre O2 et une tyrosine dans la région Y8a/Y8b
– Cette alteration de spectre est typique de la formation d'une liaison hydrogène.

Ascaris suum – Raman spectoscopie
Avec une laser Krypton (413nm) on peut regarder l'héme...– Nous pouvons se
concentrer sur la liaison Fe-LigandBasse fréquence (Fe lourd)Sensible a une marquage
isotopique (12C16O ou 13C180)Double band 515 et 543 cm-1
se déplace à 500 et 531 cm-1.
12C16O 13C180

Ascaris suum – Raman spectoscopie
Avec une laser Krypton (413nm) on peut regarder l'héme...– Nous pouvons se
concentrer sur la liaison Fe-LigandSensible a la mutation B
10Y/F
Sensible a une marquage isotopique (12C16O ou 13C180)
Simple band 520 cm-1 se déplace à 504 cm-1.

Ascaris suum – Raman spectoscopie
Nous pouvons se concentrer sur la liaison Fe-Ligand– Dans le mutant B
10Y/F une
seule site de liaison CO.– Dans la type sauvage deux
modes de liaison liaison forte ou liaison faible.

Ascaris suum – Raman spectoscopie
Nous pouvons se concentrer sur la liaison CO...– Dans le mutant B
10Y/F une
seule site de liaison CO1928/1840
– Dans la type sauvage deux modes de liaison1909/18231948/1863.

Ascaris suum – Raman spectoscopie
Les fréquences CO et FeC montent normalement une correlation...– Dans le mutant B
10Y/F une
seule type de liaison COavec des liaison H “moyenne”
– Dans la type sauvage deux modes de liaisonLiaisons H “forte”Liaisons H “faibles” (quasi
absents)
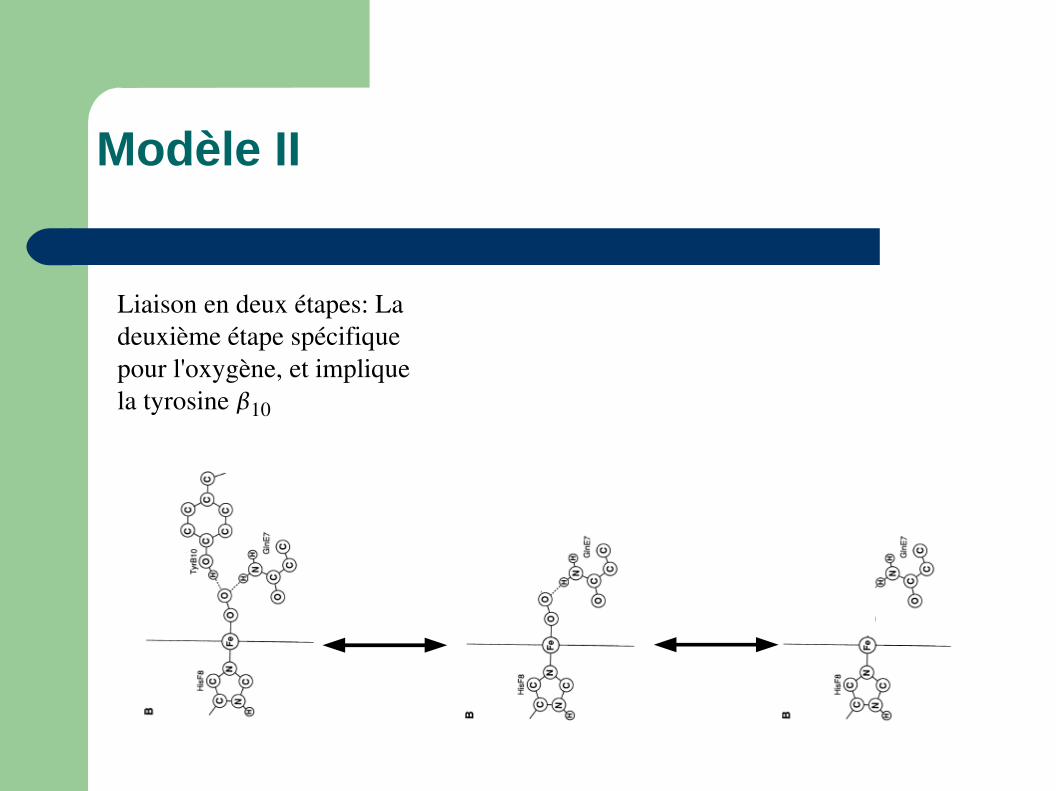
Modèle II
Liaison en deux étapes: La deuxième étape spécifique pour l'oxygène, et implique la tyrosine 10
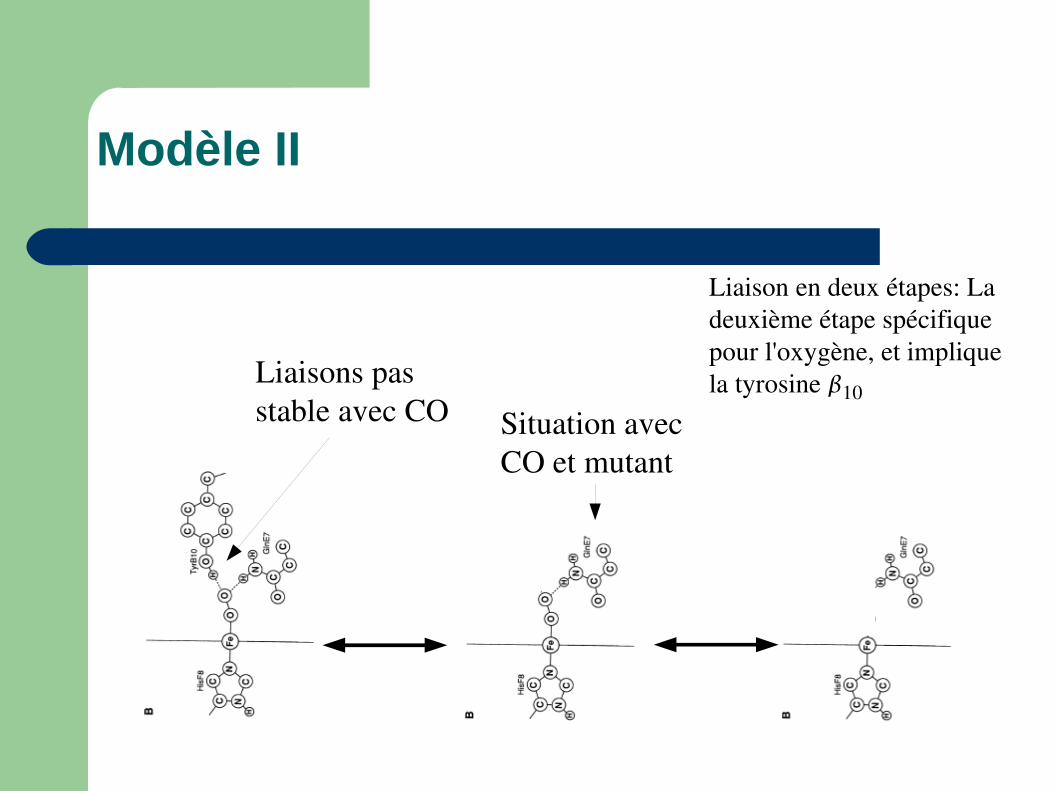
Modèle II
Liaison en deux étapes: La deuxième étape spécifique pour l'oxygène, et implique la tyrosine 10
Situation avec CO et mutant
Liaisons pas stable avec CO

Ascaris suum
Hémoglobine P50
(mmHg)
Ascaris suum 0,0072Hbα 0,82HbA 21,4Mb 15,5
Certaines hémoglobines ont une énorme affinité pour l'oxygène.– Quelles sont les origines
moléculaires?Liaisons H de B
10Y
specifique à O2
– Pourquoi une telle affinité?

Ascaris suum
Avec une telle affinité et k
off si lente une fois une
oxygène accroche elle reste!– Pas une système
dynamique– C'est une enzyme! une
desoxygènase
O2 + NO + e
NO3
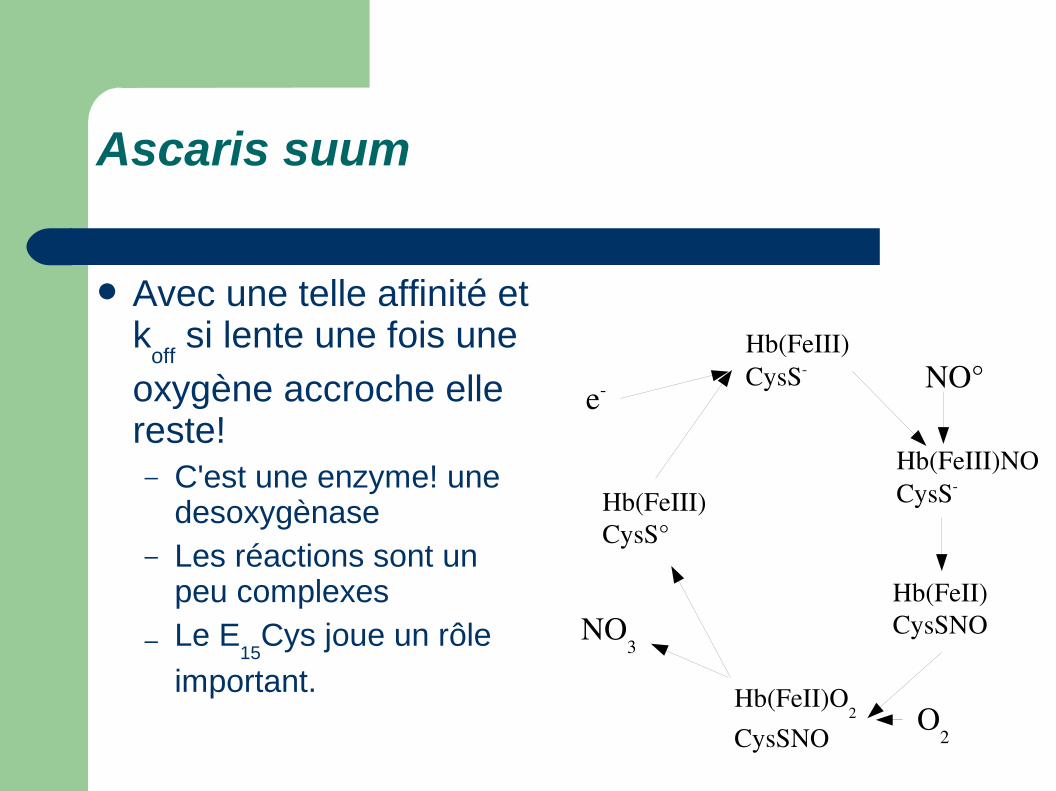
Ascaris suum
Avec une telle affinité et k
off si lente une fois une
oxygène accroche elle reste!– C'est une enzyme! une
desoxygènase– Les réactions sont un
peu complexes
– Le E15
Cys joue un rôle
important.
Hb(FeIII)CysS
Hb(FeIII)NOCysS
Hb(FeII)CysSNO
NO°
Hb(FeII)O2
CysSNO
Hb(FeIII)CysS°
O2
NO3
e

Conclusions
Les liaisons H sont important en biologie pas facile a étudier.
Les très fortes affinités indiquent un système statique et peu interessant biologiquement!

Nouveux membres de la famille
Hemoglobines et signalisation.

Nouveaux membres
Les efforts de séquencage ont mise en evidence des nouveux hémoglobines:– Neuroglobine (humain et
autres)– Cytoglobine (humain et
autres)– HemAT (bacterien)
Changement de paradigme– Anterieurement “cherche
la protéine résponsable d'une fonction observé”.
– Aujourdhui “cherche la fonction d'une ORF identifiée”

Neuroglobine et Cytoglobine
Récemment deux nouveaux hémoglobines ont été découvertes: la neuroglobine et la cytoglobine
Identification du patron d'expression par immunohistochimie.
La cytoglobuline est ubiquitaireLa neuroglobine se trouve dans les neurones mais aussi dans des cellules β des islets de pancreas.
Neuroglobine
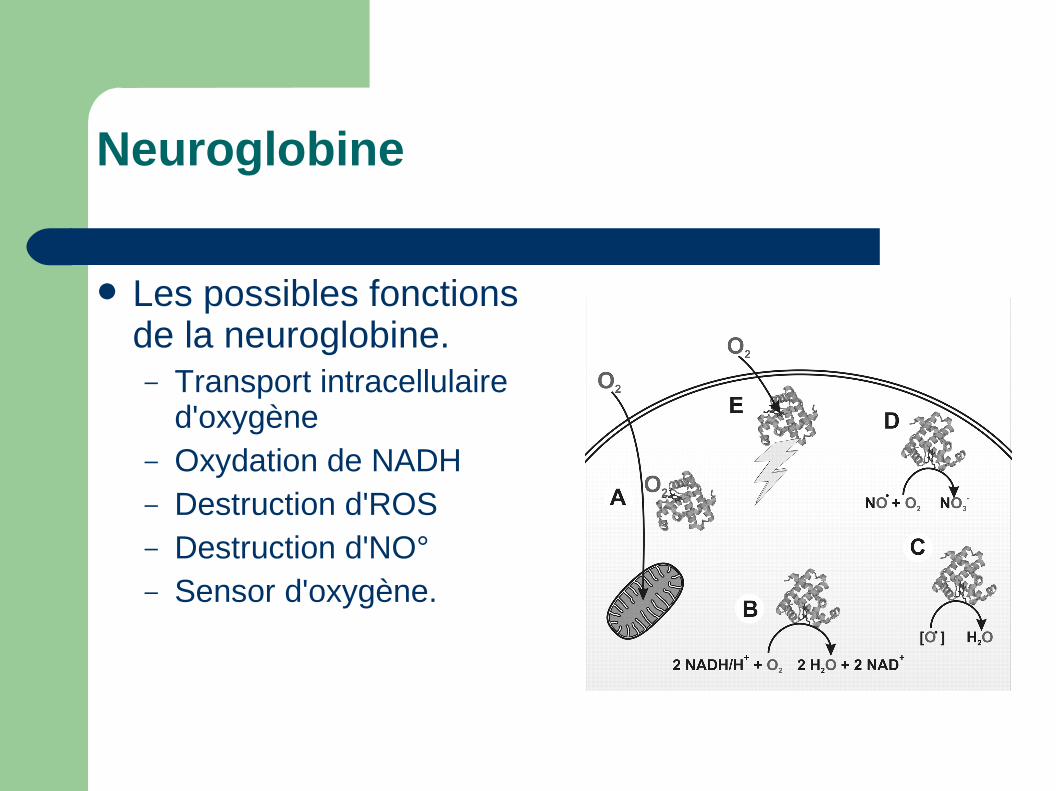
Les possibles fonctions de la neuroglobine.– Transport intracellulaire
d'oxygène– Oxydation de NADH– Destruction d'ROS– Destruction d'NO°– Sensor d'oxygène.

Neuroglobine
Alignement des hemoglobines humaines : NGB, neuroglobine; CYGB, cytoglobine; MB, myoglobine; HBA, HBB, HBD, HBE, HBG, and HBZ, chaînes d'hemoglobine.Des résidus de Cys pourraient jouer une rôle important.

Neuroglobine
Les Cys pourraient agir comme dans l'hémoglobine d'Ascaris
L'hémoglobine pourrait etre une oxygènase.
Les Cys peuvent réagir avec NO°
Aucune evidence d'activité.
Aucune interêt connue pour la cellule.
Neuroglobine
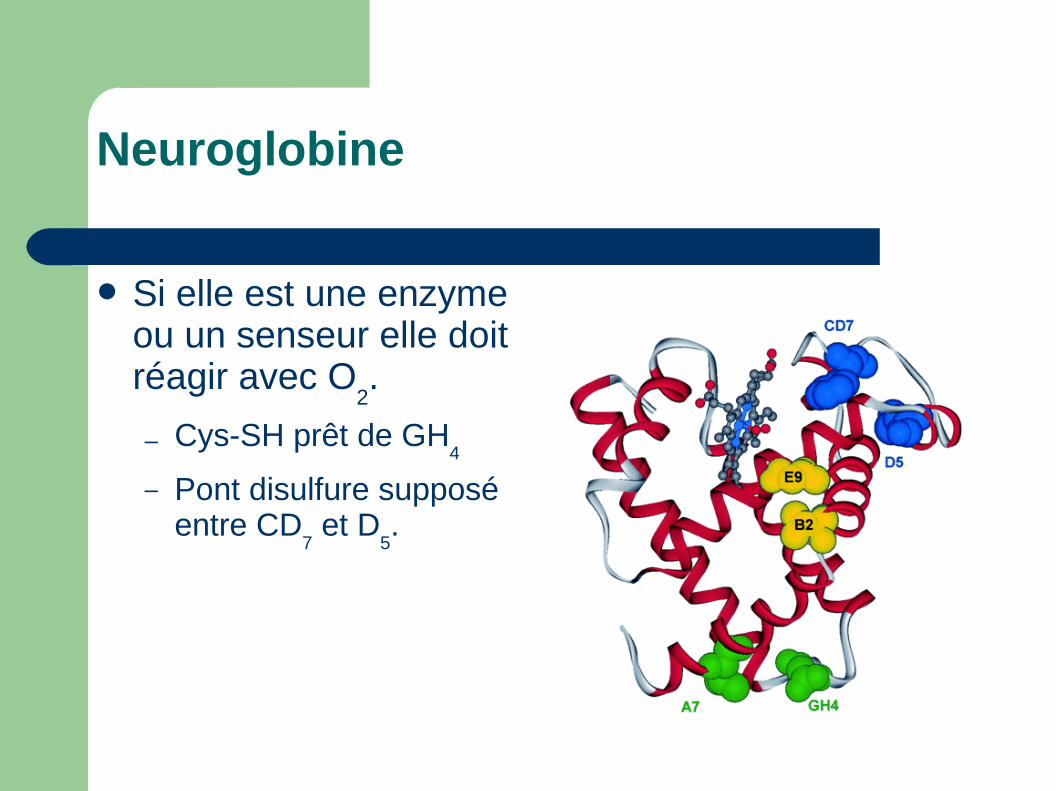
Si elle est une enzyme ou un senseur elle doit réagir avec O
2.
– Cys-SH prêt de GH4
– Pont disulfure supposé entre CD
7 et D
5.

Neuroglobine
Si elle est une enzyme ou un senseur elle doit réagir avec O
2.
– Cys-SH prêt de GH4
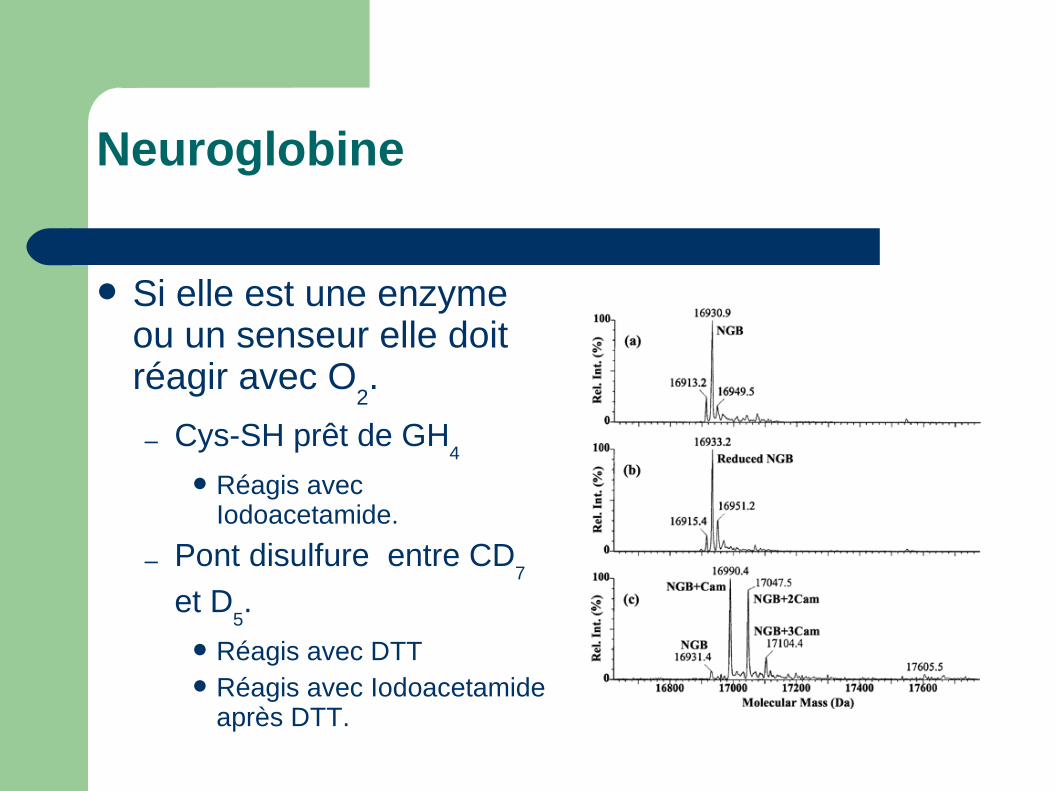
Réagis très bien avec DNTB.
– Pont disulfure entre CD7
et D5.
Protégé contre DNTB
Neuroglobine
Si elle est une enzyme ou un senseur elle doit réagir avec O
2.
– Cys-SH prêt de GH4
Réagis avec Iodoacetamide.
– Pont disulfure entre CD7
et D5.
Réagis avec DTTRéagis avec Iodoacetamide
après DTT.
Neuroglobine
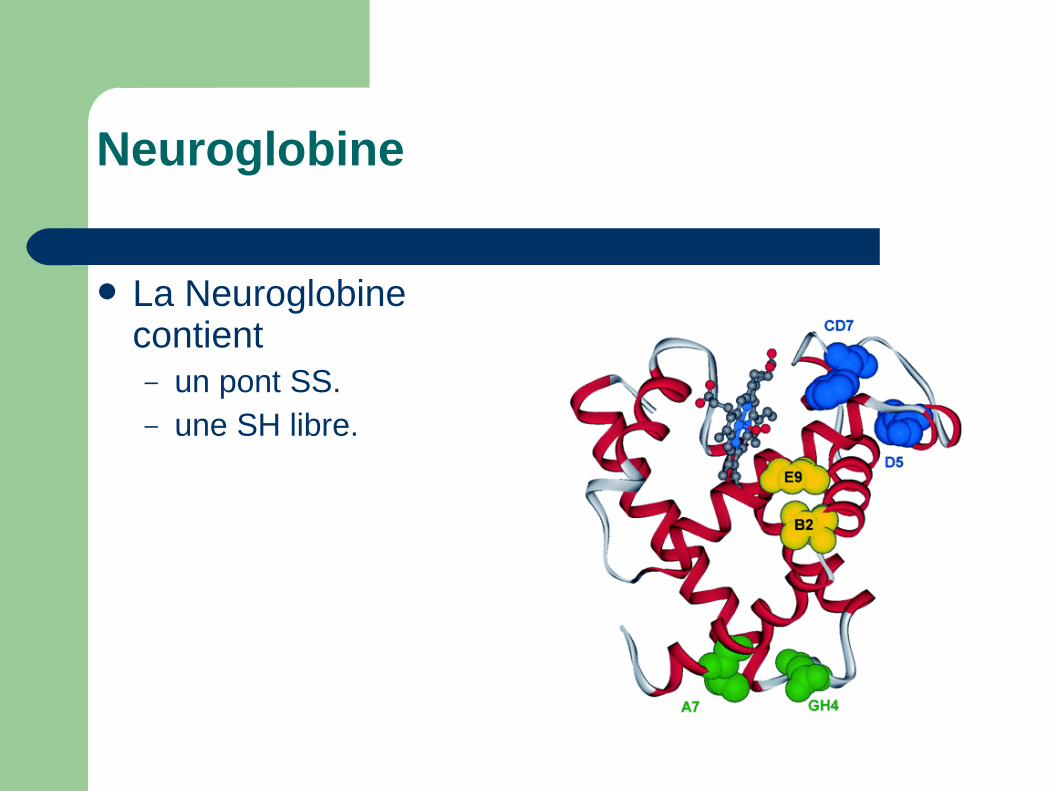
La Neuroglobine contient– un pont SS.– une SH libre.

Neuroglobine
Examination des cinétiques de réassociation de CO.
Methode classique pour les hémoglobines...
HbCO Hb + COint
Hb + COext
Lumière

Neuroglobine
Examination des cinétiques de réassociation de CO.– Utilisation de representation log-
log.– Réassociation bi-phasique
deux phases relativement lents.
– Réassociation sensible au pont SS.Phase rapide accelerée et plus grandePhase lente retardée.

Neuroglobine
Examination des cinétiques de réassociation de CO.– Utilisation de representation
log-log.– Réassociation bi-phasique– Typique des hemoglobines
hexa-coordinéVitesse limité par ouverture.Ce qui est sensible au SS
HisFeHis+ CO
int
HisFeCOHis

Neuroglobine
Ces resultats cinétiques petmettent a envisager.– Lien entre oxygenation
et état redox.– Possible d'envisager
dans une cycle catalytique ou de signalisation.
– Peu commun de garder une bonne affinité pour oxygène.

Neuroglobine
Aucune evidence
Un peu d'evidenceLie l'oxygène, important pour la survie de cellules en anoxie ou avec ROS.
Les possibles fonctions de la neuroglobine.– Transport intracellulaire
d'oxygène– Oxydation de NADH– Destruction d'ROS– Destruction d'NO°– Sensor d'oxygène.

Neuroglobine
Une rôle comme senseur...– Similitude à la
myoglobine– Similitude a des
domaines RGSRégulateurs de la
Signalisation par des protéines G.
– Similitude sur la surface.
Neuroglobine
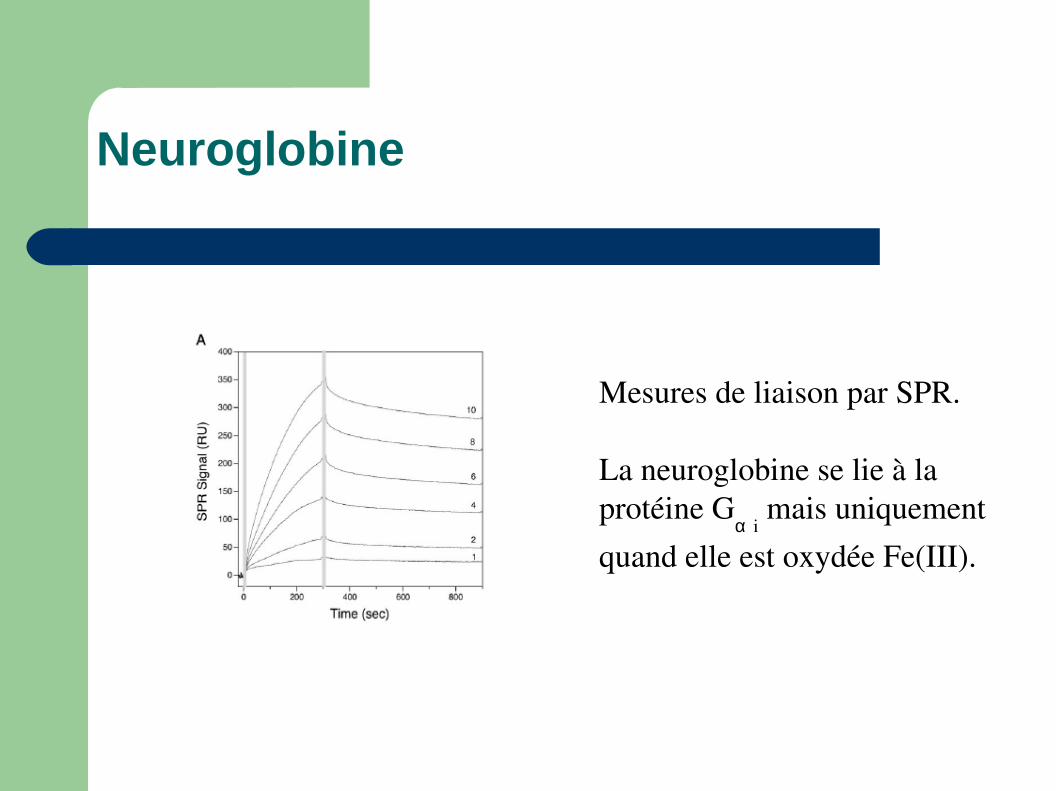
Mesures de liaison par SPR.
La neuroglobine se lie à la protéine G
α i mais uniquement
quand elle est oxydée Fe(III).
Neuroglobine
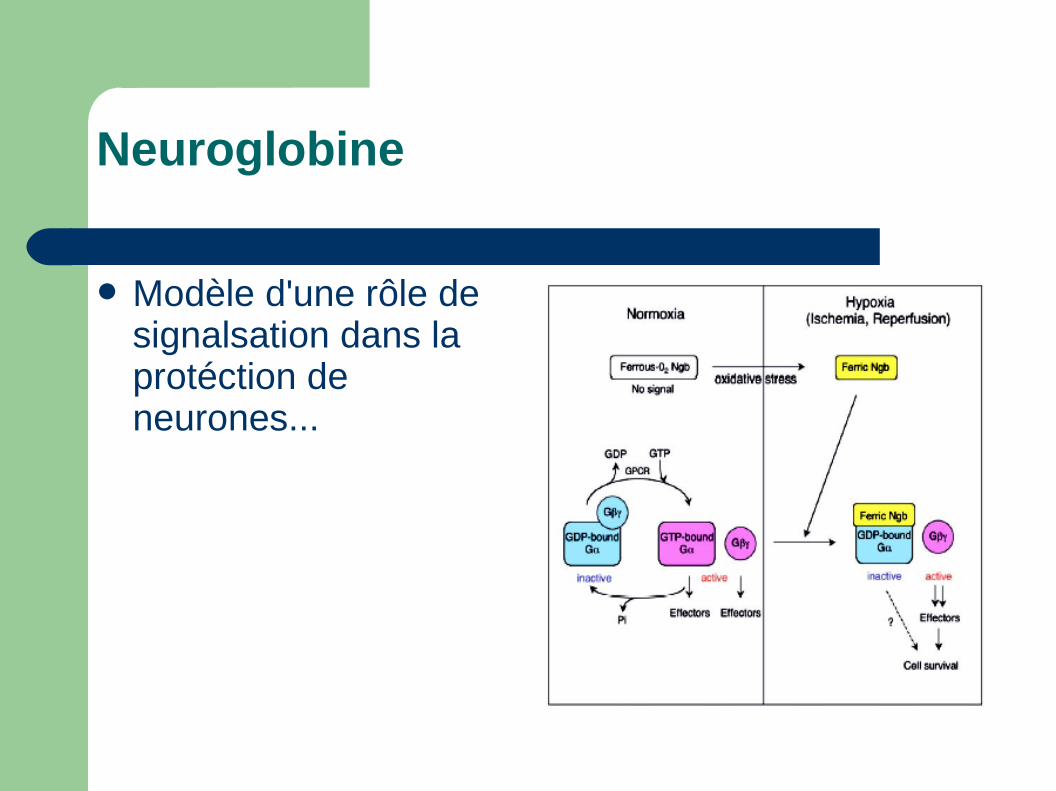
Modèle d'une rôle de signalsation dans la protéction de neurones...

Neuroglobine
Après 7 ans...– Peut-etre signalisation
Pas une bonne demonstation dans une cellule encore.
– Peut-etre ROS destruction.Pareil....
Peut liée Gi
Structure connue Hexacoodoné en
absence d'O2.
Bonne affinité pour oxygène et CO.
Protége contre ROS et anoxie in vivo.

HemAT
Protéines d'aerotaxie des bactéries et d'archea
Hou et al. (2000) Nature 403: 540-544.
Hou et al. (2001) PNAS 98: 9353-9358
Zhang & Phillips (2003) Structure 11: 1097-1110
Deux protéines HemATHs et HemATBs.
Identifié en Halobacterium salinarum (un archae) et Bacilus subtilis (un bactérie).
Des protéines de 489 (et 423) résidus avec deux parties....

HemAT
1 184 222 489
Domaine 1: 16% identité avec la myoglobine.
Domaine 2: 30% identité avec les domaines de signalisation de certains MCP. (Protéines de chemotaxie acceptant des methyls).
Protéine bacterien a deux domains.
Lie t'elle des ligands? Quelle est sa rôle
physiologique?
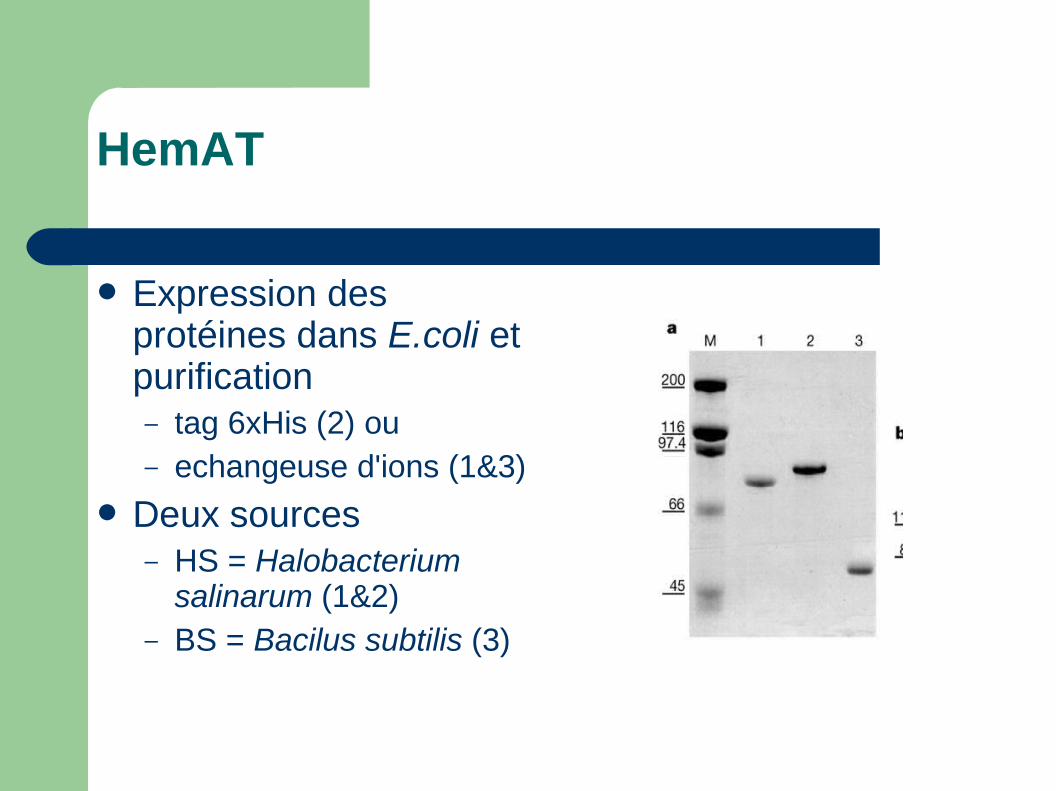
HemAT
Expression des protéines dans E.coli et purification– tag 6xHis (2) ou– echangeuse d'ions (1&3)
Deux sources– HS = Halobacterium
salinarum (1&2)– BS = Bacilus subtilis (3)

HemAT
Après expression hétérologue et purification examination du spectre d'absorption
Similaire a l'oxymyoglobine.– Elle lie l'héme et– l'héme lie l'oxygène.
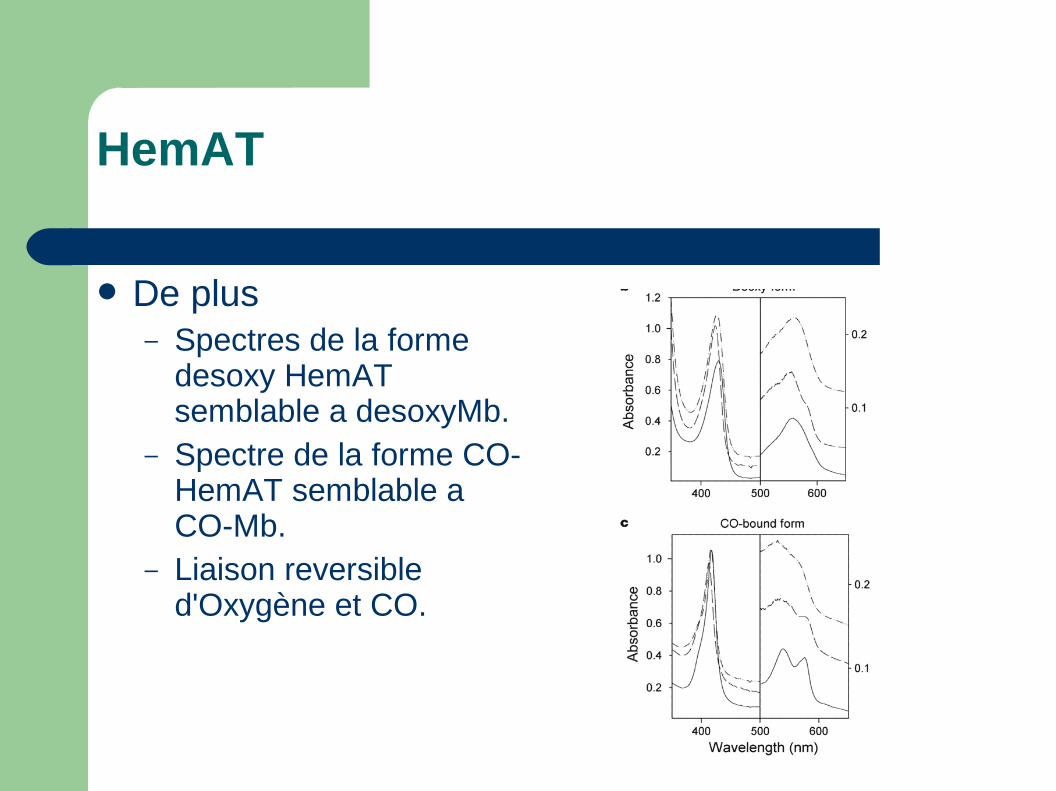
HemAT
De plus– Spectres de la forme
desoxy HemAT semblable a desoxyMb.
– Spectre de la forme CO-HemAT semblable a CO-Mb.
– Liaison reversible d'Oxygène et CO.

HemAT
Mesure de l'affinité et cinétique de liaison d'oxygène
Affinité et cinétiques semblable à d'autres hémoglobines
Cohérent avec une rôle dans la detection d'oxygène.

HemAT
Pense a une rôle dans la signalisation– Une domaine detecteur
d'O2
– Une domaine de chimiotaxie.
– Peut-etre une senseur pour l'aerotaxie?
Comment le demontré?

HemAT
Pense a une rôle dans la signalisation– Une domaine detecteur
d'O2
– Une domaine de chimiotaxie.
– Peut-etre une senseur pour l'aerotaxie?
Comment le demontré? Génétique moléculaire
– Construire des mutants de déletion
– Complementation héterologue
– Analyse de mutants

HemAT
Comment le demontré? Génétique moléculaire
– Construire des mutants de déletion
– Complementation héterologue
– Analyse de mutants
Un peu complexe comme plusieurs systèmes.

HemAT
Crystallographie– Que une seule domaine
(la domaine hemoglobine) visible dans la structure!
– Une dimère...Es vrai pour la molecule
entière?Quelle role?

HemAT
Structure de la domaine hémoglobine typique. mais avec une hélice N terminale de plus - Z

HemAT
Les deux molécules dans le dimère sont différentes!
Notamment au niveau de leur site de liaison– Une seule monomère
avec ligand (CN-)– Une rotation de Y70.
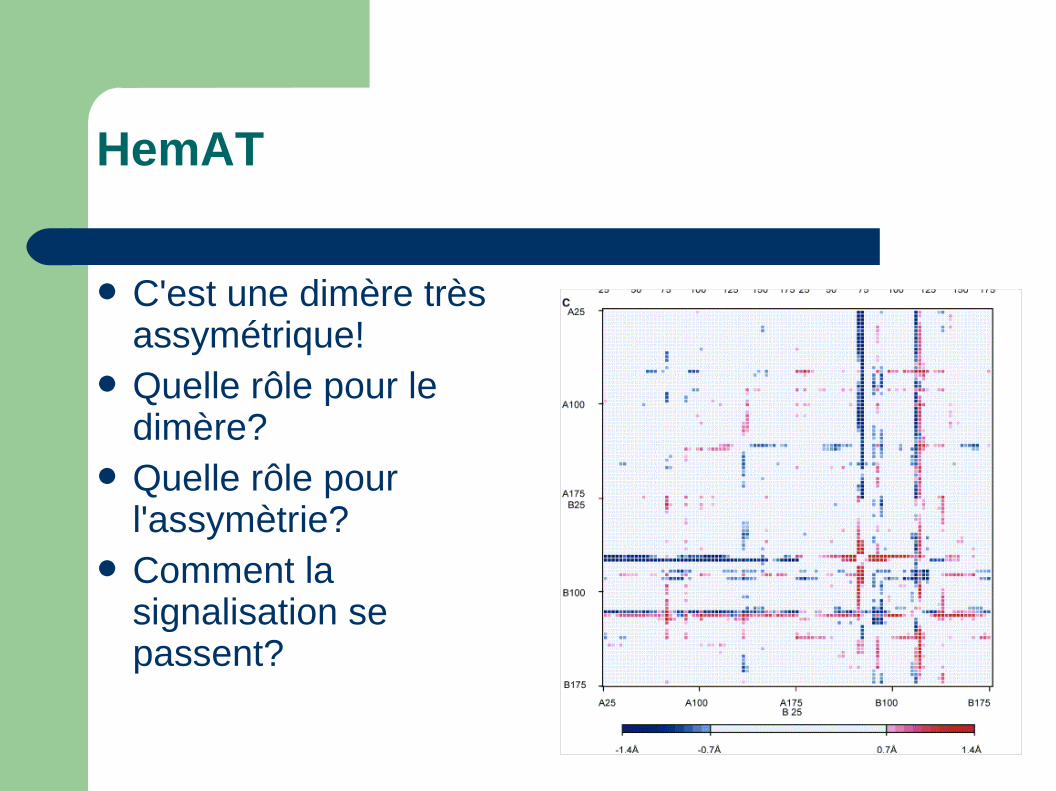
HemAT
C'est une dimère très assymétrique!
Quelle rôle pour le dimère?
Quelle rôle pour l'assymètrie?
Comment la signalisation se passent?
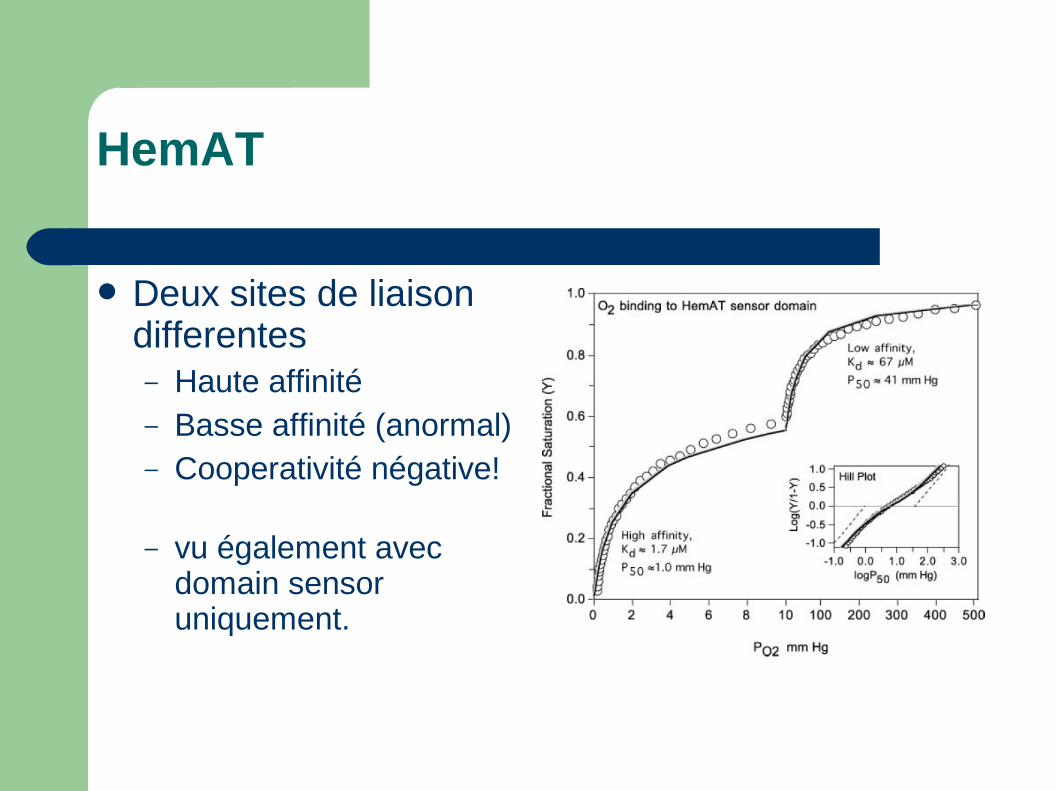
HemAT
Deux sites de liaison differentes– Haute affinité– Basse affinité (anormal)– Cooperativité négative!
– vu également avec domain sensor uniquement.

HemAT
Reste à éclaircir entre autre...– Comment liaison du
ligand est transmis a la domaine MCP?
– Pourquoi une cooperativité negative?
Cooperativité negative et deux sites est vu dans plusieurs senseurs...– Tar / Tsr– HemAT
Probablement important! pourquoi?

Conclusions
Deux facons de gagner une fonction de signalisation.– Evolution convergente
(Neuroglobuline)– Ajoute de domaines
(HemAT)
Démontré une fonction cellulaire et biochimique n'est pas toujours simple.

Fin


















