
Mycorhizes VAM et Geosiphon pyriformis (Kützing) … · ordre des Gigasporineae), ils sont...
17
Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille 1 LES GLOMEROMYCOTA - Mycorhizes VAM et Geosiphon pyriformis (Kützing) Wettstein - Jean-Pierre GAVÉRIAUX 14, résidence Les Hirsons, rue Eugène Mordacque - 62800 LIÉVIN [email protected] Les Ascomycota et les Basidiomycota sont les champignons les plus connus du grand public, ce sont eux qui sont presque exclusivement étudiés par les membres de nos associations mycologiques. Le règne fongique n’est toutefois pas limité à ces deux phylums, il en regroupe plusieurs autres, parmi lesquels celui des Glomeromycota, qui ne renferme qu’un peu plus de 200 espèces différentes, pratiquement inconnues des mycologues, sans lesquelles pourtant, le fonctionnement de notre planète ne serait pas possible. Les Glomeromycota permettent (entre autres) à plus de 80% des plantes herbacées (et des arbres en régions tropicales) de s’approvisionner en eau et en éléments minéraux (phosphore en particulier), puisés dans le sol grâce à des mycorhizes particulières, les VAM - abréviation de Vesicular-Arbuscular Mycorrhizas - mycorhizes à vésicules et arbuscules -parfois encore appelées endomycorhizes vésiculo-arbusculaires, par opposition aux ectomycorhizes engendrées par les lactaires, russules, bolets ou cortinaires… dans nos écosystèmes forestiers tempérés et arctiques. Chytridiomycota Descendants actuels des plus anciens Mycota apparus la Terre ; ils ont des cellules mobiles munies d’un flagelle postérieur. Entomophtoromycotina Parasites (pf. saprophytes), présence de zoospores uniflagellées, avec une alternance de générations haploïdes et diploïdes. Zoopagomycotina Parasites de champignons ou d’amibes (autrefois dans les zygomycètes (quelques dizaines d’espèces). Kickxellomycotina Les hyphes possèdent au niveau de leurs cloisons des pores lenticulaires (ce sont des parasites ou des commensaux) Mucormycotina Correspondent à la plupart des espèces autrefois placées dans les Zygomycètes. Neocallimastigomycota Organismes anaérobies (dépourvus de mitochondries), vivant dans le système digestif des grands mammifères herbivores. Microsporidia Plus de 1100 espèces, des parasites intracellulaires des animaux (y compris l’homme), posant de nombreux problèmes aux éleveurs. Glomeromycota Hyphes avec ébauches de cloisons, pas de reproduction sexuée, formation d’endomycorhizes vésiculo-arbusculaires. Ascomycota Hyphes septées, corps de Woronin au niveau des cloisons, présence d’une dicaryophase, production d’endospores dans des asques. Basidiomycota Hyphes septées, dolipores au niveau des cloisons, présence d’une dicaryophase, production d’exospores sur des basides. Principales divisions actuelles du règne fongique 1 Ces champignons symbiotiques étaient autrefois classés dans l’ordre des Glomales parmi les Zygomycètes. 1 La présentation des divers phylums de champignons inférieurs a fait l’objet d’une conférence d’une heure, le samedi 26 décembre, à la faculté de pharmacie de Lille, dans le cadre des activités du second semestre, de la Société Mycologique du Nord de la France.
Transcript of Mycorhizes VAM et Geosiphon pyriformis (Kützing) … · ordre des Gigasporineae), ils sont...

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
1
LES GLOMEROMYCOTA - Mycorhizes VAM et Geosiphon pyriformis (Kützing) Wettstein -
Jean-Pierre GAVÉRIAUX 14, résidence Les Hirsons, rue Eugène Mordacque - 62800 LIÉVIN
[email protected] Les Ascomycota et les Basidiomycota sont les champignons les plus connus du grand public, ce sont eux qui sont presque exclusivement étudiés par les membres de nos associations mycologiques. Le règne fongique n’est toutefois pas limité à ces deux phylums, il en regroupe plusieurs autres, parmi lesquels celui des Glomeromycota, qui ne renferme qu’un peu plus de 200 espèces différentes, pratiquement inconnues des mycologues, sans lesquelles pourtant, le fonctionnement de notre planète ne serait pas possible. Les Glomeromycota permettent (entre autres) à plus de 80% des plantes herbacées (et des arbres en régions tropicales) de s’approvisionner en eau et en éléments minéraux (phosphore en particulier), puisés dans le sol grâce à des mycorhizes particulières, les VAM - abréviation de Vesicular-Arbuscular Mycorrhizas - mycorhizes à vésicules et arbuscules -parfois encore appelées endomycorhizes vésiculo-arbusculaires, par opposition aux ectomycorhizes engendrées par les lactaires, russules, bolets ou cortinaires… dans nos écosystèmes forestiers tempérés et arctiques.
Chytridiomycota Descendants actuels des plus anciens Mycota apparus la Terre ; ils ont des cellules mobiles munies d’un flagelle postérieur.
Entomophtoromycotina Parasites (pf. saprophytes), présence de zoospores uniflagellées, avec une alternance de générations haploïdes et diploïdes.
Zoopagomycotina Parasites de champignons ou d’amibes (autrefois dans les zygomycètes (quelques dizaines d’espèces).
Kickxellomycotina Les hyphes possèdent au niveau de leurs cloisons des pores lenticulaires (ce sont des parasites ou des commensaux)
Mucormycotina Correspondent à la plupart des espèces autrefois placées dans les Zygomycètes.
Neocallimastigomycota Organismes anaérobies (dépourvus de mitochondries), vivant dans le système digestif des grands mammifères herbivores.
Microsporidia Plus de 1100 espèces, des parasites intracellulaires des animaux (y compris l’homme), posant de nombreux problèmes aux éleveurs.
Glomeromycota Hyphes avec ébauches de cloisons, pas de reproduction sexuée, formation d’endomycorhizes vésiculo-arbusculaires.
Ascomycota Hyphes septées, corps de Woronin au niveau des cloisons, présence d’une dicaryophase, production d’endospores dans des asques.
Basidiomycota Hyphes septées, dolipores au niveau des cloisons, présence d’une dicaryophase, production d’exospores sur des basides.
Principales divisions actuelles du règne fongique 1 Ces champignons symbiotiques étaient autrefois classés dans l’ordre des Glomales parmi les Zygomycètes.
1 La présentation des divers phylums de champignons inférieurs a fait l’objet d’une conférence d’une heure, le samedi 26 décembre, à la faculté de pharmacie de Lille, dans le cadre des activités du second semestre, de la Société Mycologique du Nord de la France.

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
2
Les études phylogénétiques récentes ont conduit à la création d’un phylum bien individualisé, celui des Glomeromycota, clade monophylétique dans lequel une espèce, Geosiphon pyriforme, présente une symbiose de type différent en s’associant, non pas avec des photosymbiotes eucaryotes mais avec des photosymbiotes procaryotes, des cyanobactéries (Nostoc). Dans les Glomeromycota on distingue actuellement quatre ordres, les Glomerales (autrefois appelées Glomales), les Diversisporales, les Paraglomerales, et les Archaeosporales (ex-Geosiphonales), parmi lesquelles se trouve la famille des Geosiphonaceae, monospécifique, contenant le Geosiphon (autrefois considéré comme étant une espèce lichénisée). I. Étapes de la formation de la mycorhize à arbuscules et vésicules 1. Développement des hyphes dans le sol Les premières hyphes ont deux origines possibles, elles sont issues de la germination d’une spore ou proviennent de racines mycorhizées qui se trouvent déjà dans le sol. Les hyphes issues de la germination de spores ont une durée de vie très limitée, elles dépérissent si elles ne rencontrent pas rapidement une racine compatible ; ce champignon est en effet un biotrophe obligatoire, il ne peut pas se développer sans son hôte. Les hyphes d’origine racinaire sont des filaments résultant de l’activité d’une mycorhization antérieure qui développe son réseau mycélien dans le but de rechercher des éléments nutritifs ou de partir à la conquête d’autres racines. Ces hyphes ne sont pas cloisonnées, elles ont une structure cœnocytique, les noyaux sont dans un même cytoplasme commun ; en fonction de leur activité principale, on peut distinguer plusieurs types d’hyphes extra-racinaires : - Les hyphes d’absorption, très ramifiées et minces, elles prélèvent les molécules du sol. - Les hyphes conductrices ayant un diamètre plus important et un cytoplasme peu abondant. - Les hyphes d’infection qui peuvent coloniser de nouvelles racines. - Les hyphes sporogènes qui donneront naissances aux spores. 2. Rencontre avec la racine Lorsqu’une racine rencontre au cours de sa croissance, les hyphes du sol, issues d’une germination récente (ou en survie suite à une diminution de leur activité métabolique), les hyphes sont comme attirées, elles viennent se placer le long de la racine, des contacts s’établissent, de petits gonflements apparaissent ; ils correspondent à des appressoria (du l atin apprimere) à partir desquels les hyphes vont pénétrer dans la partie externe de la racine en passant entre les cellules de l’épiderme et du cortex ; les hyphes vont alors pouvoir se développer, se ramifier, coloniser la racine.

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
3
Cette communication entre l’hyphe et la racine est possible via la libération de substances chimiques par la plante, les plus courantes étant les molécules de strigolactone (le 5-deoxy-strigol) qui stimulent l’activité des mitochondries du champignon qui reprend son activité métabolique. Ces messagers chimiques interviennent à des concentrations très faibles, de l’ordre du picogramme (unité de masse égale à un millième de milliardième de gramme). 3. La prolifération intra-racinaire et la formation des arbuscules À partir de l’appressorium, l’hyphe va essayer de passer dans les divers espaces intercellulaires disponibles ; deux types de propagation ont été définis (Smith & Smith, 1997) : - la propagation linéaire où le développement des hyphes s’effectue principalement dans les espaces intercellulaires longitudinaux ; - la propagation en enroulement dans lequel les hyphes ont tendance à s’enrouler, à suivre un cheminement tortueux lorsque l’espace intercellulaire disponible est insuffisant. Quelques jours après l’entrée dans le cortex, les hyphes vont se ramifier par dichotomie en donnant des hyphes ayant un diamètre de plus en plus petit ; à partir d’une hyphe initiale ayant 10 µm de Ø, on arrive dans les dernières ramifications à moins de 1 µm de Ø. L’ensemble de ces ramifications prend la forme d’un petit arbre, d’où le terme d’arbuscule attribué à cette structure (Galland, 1905) qui est un haustorium particulier. L’arbuscule provoque une déformation de la cellule proche dans laquelle il finit par entrer ; toutefois, il n’y a jamais pénétration dans le cytoplasme dont la membrane cytoplasmique souple s’est invaginée. Il y a ainsi une augmentation considérable de la surface de contact entre l’hyphe et le cytoplasme de la cellule végétale et apparition d’une vaste zone de transfert entre les deux partenaires de la symbiose. La mycorhize à arbuscules est devenue fonctionnelle, le champignon obtient les sucres (glucides) qu’il ne sait pas fabriquer. Ces arbuscules vivent peu de temps, en général quelques jours (quelques mois dans le sous-ordre des Gigasporineae), ils sont absorbés par le cytoplasme de la cellule racinaire qu is’approvisionne ainsi en eau, sels minéraux, facteurs de croissance…
Prolifération des hyphes, formation des arbuscules et des vésicules. (d’après un schéma trouvé sur le site : http://mycorrhizas.info/resource.html#photos)
4. La formation des Vésicules Après le développement des arbuscules, des hyphes commencent à se renfler à certains endroits de leur parcours ou à leur extrémité pour donner des espaces de stockage qui permettront la mise en réserve de diverses substances : lipides (corps gras), tri-acylglycérol (TAG)... Ces structures appelées vésicules peuvent être inter- ou intracellulaires, elles

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
4
contiennent de nombreux noyaux et leur paroi est souvent mince mais peut être épaisse, dans le cortex des racines âgées (absentes toutefois chez les Gigasporinae). Remarque : Les modalités d’organisation de ces VAM sont très variables. Les arbuscules peuvent être diversement ramifiés, présenter des hyphes de diamètres différents, l’ensemble de la ramification paraître sombre ou clair. Les vésicules sont également caractérisées par leurs formes, leur taille, leur répartition, leur ± grande abondance (ex. chez le poireau, Allium porrum, Glomus intraradices donne plus de 200 vésicules par mm de racine, tandis que Glomus monosporum n’en forme que quelques-unes). 5. Les cellules auxiliaires Ces structures ne se rencontrent que dans le sous-ordre des Gigasporinae leur surface est épineuse dans le genre Gigaspora, presque lisse dans le genre Scutellospora ; elles se forment sur les hyphes extra-racinaires sporogènes au moment de la sporulation, parfois sur les hyphes germinatives avant l’entrée dans les racines. Leur durée de vie est courte et leur rôle n’est pas encore connu avec précision mais on pense qu’elles fourniraient des molécules carbonées. II. Le fonctionnement de la mycorhize : la symbiose mutualiste La mycorhize est réalisée par les deux partenaires de la symbiose, chacun des associés essaie d’obtenir le maximum mais globalement les échanges se produisent avec bénéfices réciproques et chaque symbiote tire avantage de cette vie en commun. Avantages pour le champignon Le mycosymbiote hétérotrophe ne sait pas fabriquer de glucides à partir de l’eau et du dioxyde de carbone, les Glomeromycota ne sont pas n’est pas capables de dégrader la cellulose et la lignine, c’est la plante autotrophe qui va lui fournir ses molécules carbonées, des photosynthétats, principalement des hexoses (sucres en C6 comme le glucose par exemple) qui ont trois utilisations principales : - être dégradés pour fournir de l’énergie aux cellules fongiques, - être utilisés comme squelettes carbonés pour la synthèse des composés cellulaires. - être stockés pour une utilisation ultérieure, sous forme de glycogène dans les hyphes extra-racinaires, sous forme de tri-acylglycérol dans les vésicules (Bago, et al., 2000). L’apport n’est pas limité aux molécules glucidiques, des molécules complexes sont également élaborées par la plante, certaines d’ente elles stimulent la croissance des hyphes. Avantages pour la plante-hôte - Accroissement du volume de sol exploitable Les possibilités d’échanges entre les racines d’une plante et le sol sont limitées au volume de sol accessible aux racines ; ce volume correspond à la rhizosphère. Lorsque les racines sont prolongées par des hyphes fongiques, le volume de sol exploitable augmente, il y a création d’une mycorhizosphère qui amplifie de façon très significative la zone dans laquelle la plante peut extraire de l’eau et des éléments nutritifs et peut explorer des couches humides situées à

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
5
grande profondeur. Chez certaines plantes de déserts, les hyphes extra-racinaires peuvent aller extraire l’eau à plusieurs dizaines de mètres de profondeur.
Les hyphes augmentent de façon considérable le volume de sol exploitable.
- Amélioration de la nutrition minérale [P / N / oligoéléments] Dans le sol, les ions phosphates sont retenus dans les colloïdes ou immobilisés sous forme de phosphates de fer ou d’aluminium ; bien que présent, la majorité du phosphate est donc indisponible pour les plantes. L’équipement enzymatique des champignons est très vaste, des phosphatases sont présentes et sous leur action accompagnées au niveau des membranes fongiques de mécanismes de transport actif, le phosphore se retrouve dans les hyphes extra-racinaires. Les ions phosphates absorbés ont trois devenirs possibles : - une partie est utilisée par le champignon, - une partie cédée à la plante via l’interface arbusculaire, - le reste mis en réserve dans les vacuoles des cellules fongiques sous forme de polyphosphates. Pour l’azote, les mécanismes n’ont pas encore été étudiés en détail et seuls quelques résultats sont actuellement disponibles. Chez certaines Glomales, les hyphes extra-racinaires assimilent l’azote sous forme de nitrates, nitrites, ammonium du sol ; ces molécules sont transformées par le champignon et stockées dans les cellules du mycélium extra-racinaire sous forme d’arginine. Cet acide aminé est ensuite transféré au mycélium intra-racinaire, dégradé en ammonium qui est ensuite transféré à la plante au niveau de l’arbuscule. Les besoins des plantes en sels minéraux ne sont pas limités au phosphore et à l’azote ; d’autres éléments comme le soufre ou des oligoéléments comme le cuivre, le zinc, le fer, le manganèse… sont également indispensables. Ces substances difficilement mobilisables bénéficient également de l’aide fongique surtout suite à une meilleure exploitation du sol par les hyphes extra-racinaires (Vinayak et Bagyaraj, 1990). Dans pratiquement tous les cas on note que la quantité prélevée est très supérieure à celle qui serait absorbée par l’ensemble racine, radicelles et poils absorbants. - Amélioration de l’alimentation en eau et meilleure résistance au stress hydrique La circulation de l’eau dans une plante est due à deux moteurs, les mécanismes d’absorption au niveau des racines et les mécanismes de transpiration au niveau des feuilles. Chez les plantes mycorhizées les hyphes du réseau extra-racinaire, dépourvues de cloisons transversales et à cytoplasme peu abondant, facilitent le transport de l’eau vers les racines.

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
6
Ces racines supportent mieux le dessèchement du sol et récupèrent plus rapidement leur turgescence après un apport d’eau. Les mécanismes sont toutefois complexes, il y a une grande variété de comportements métaboliques. On note toutefois une augmentation de la transpiration, de la conduction racinaire, et lors de périodes de stress hydrique, la turgescence des feuilles est maintenue par un ajustement des pressions osmotiques des vacuoles suite à une augmentation des concentrations en ions organiques et/ou minéraux. - Production de messagers chimiques (hormones) Des phytohormones, des auxines qui agissent en quantité infinitésimale, stimulent le développement de racines dans lesquelles de jeunes hyphes pourront entrer facilement. Le développement du système racinaire est ainsi facilité. - Tolérance aux excès de certains éléments minéraux Par exemple sur les terrils, au bout de quelques années de passage à la friche, la partie superficielle du sol présente un excès de magnésium ; presque toutes les plantes herbacées disparaissent ; une des rares plantes à résister, qui donne alors des peuplements pratiquement monospécifique, est l’avoine élevée (Arrhenaterum elatius), des études ont montré que cette plante était protégée par des Glomus qui stockaient l’excès de magnésium dans des mycorhizes VAM. - Protection phytosanitaire Dans le sol, tout autour de la racine, dans la rhizosphère, de nombreux micro-organismes sont présents et cherchent à établir des relations avec la racine. Si certains sont symbiotiques ou commensaux, d’autres se comportent parfois en dangereux ravageurs, parasites ou sont phytopathogènes. L’association des capacités de défense des hyphes et des capacités de défense de la plante permet souvent une lutte beaucoup plus efficace contre les bactéries et champignons pathogènes et même parfois contre les nématodes. De nombreux champignons sont également capables de produire des substances antibiotiques qui protègent la plante-hôte. Dans 1 gramme de sol de mycorhizosphère, il est possible d’avoir 12 mètres d’hyphes de champignons endomycorhizogènes. Outre les avantages apportés à la plante-hôte, ce réseau permet la communication entre les diverses plantes d’une même culture.
Plantes concernées par les mycorhizes VAM La mycorhization faisant intervenir des VAM est l'une des symbioses les plus importantes sur la terre, elle concerne plus de 80% des plantes vasculaires, mais se rencontre également chez certaines Ptéridophytes. Ces mycorhizes VAM sont la quasi-généralité chez les plantes cultivées, essentiellement dans des groupes que nous connaissons tous : - Poaceae (ex-Graminées) : graminées de nos prairies, graminées cultivées : blé, riz, maïs…, - Fabaceae (ex-Papilionacées) : soja, haricot, pois, lentilles, fèves, tournesol, luzerne…, - cultures légumières : pommes de terre, tomates, oignons, poireaux, concombres, - cultures horticoles : œillets, géraniums, rosiers, géraniums…, - cultures fruitières : fraisiers, framboisiers, mûrier, pommiers, pruniers, cerisiers… Par contre dans quelques familles la plupart des plantes ne sont jamais mycorhizées : - Brassicaceae (ex-Crucifères) : chou, colza, navet, radis, - Chenopodiaceae : épinards, betteraves, chénopode…

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
7
- Caryophyllaceae : œillet… Remarques : - On recense actuellement un peu plus de 150 espèces de Glomeromycota entretenant des endomycorhizes VAM avec près de 400 000 espèces végétales distinctes. Ces espèces fongiques peuvent donc s’associer avec plusieurs plantes différentes et possèdent obligatoirement des capacités génétiques leur permettant de faire face à des exigences extrêmement diversifiées. - Les endomycorhizes VAM, symbiotes de plantes qui nous sont particulièrement utiles, font l’objet de nombreuses études en agronomie, horticulture et foresterie. Dans les régions arides, où les plantes sont souvent en condition de stress hydrique, on constate que les arbres des forêts tropicales, capables de résister à la sécheresse, sont tous endomycorhiziques. - De nombreux programmes de recherches sont actuellement en cours, une collection de tous les Glomeromycota disponibles a été réalisée ; cette banque mondiale, l’IBG (International Bank of Glomeromycota) a été créée en 2002 ; elle est gérée en France par l’INRA (Institut National de la Recherche Agronomique) de Dijon (http://www.dijon.inra.fr/mychintec/) ; elle est à la disposition des chercheurs du monde entier. Ces champignons commencent enfin à être connus et plusieurs organismes commerciaux proposent, depuis quelque temps, les premiers plants inoculés. III. Reproduction des Glomeromycota Le suivi de gènes accompagnés de marqueurs moléculaires n’a pas permis de mettre en évidence des recombinaisons génétiques (Tommerup et Sivasithamparam, 1990). Il est donc actuellement admis par la majorité des mycologues que ces champignons sont dépourvus de reproduction sexuée et qu’ils forment des spores asexuées (mitospores) selon un processus de multiplication végétative bien que, la question de savoir, si les noyaux du mycélium cœnocytique sont génétiquement identiques ou non, fasse l’objet de nombreux débats (Kuhn & al., 2001 ; Pawlowska & Taylor, 2004 ; Hijri & Sanders, 2005). Les microspores se forment lorsque l’association symbiotique commence à passer en phase de sénescence ou lorsque le champignon commence à réutiliser les nutriments préalablement stockés dans les racines. Cette élaboration peut se faire à partir des hyphes de la racine ou des hyphes du sol.
hyphe sporogène renflement plurinucléé individualisation de la spore
Certaines hyphes mycéliennes cœnocytiques, appelées hyphes sporogènes, forment des renflements qui contiennent du cytoplasme, de nombreux noyaux, des réserves, principalement sous forme de lipides ; ces renflements s’entourent ensuite d’une paroi ± épaisse, ± pluristratifiée (mitosporanges), ce qui permet l’individualisation des spores. Parfois la spore est portée par un petit pédicelle (ex. chez les Archaeospora) ou se forme à l’extrémité de l’hyphe sporogène (ex. chez les Glomus) ; dans quelques cas, les mitospores sont regroupées, forment des agglomérats, les mitosporomes (souvent nommés sporocarpes bien

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
8
que ce terme soit étymologiquement incorrect, les champignons ne donnant pas de fruits) enveloppés d’une couche périphérique d’hyphes serrées constituant le péridium. Ces spores globuleuses sont très grandes, de 20 µm pour les plus petites à parfois presque 2000 µm de diamètre pour les plus grandes (Gigaspora), elles contiennent de très nombreux noyaux (jusqu’à plusieurs centaines). Elles permettent la dissémination de l’espèce, mais elles servent également de structures de stockage ou sont parfois considérées comme des phases de repos dans le cycle biologique. Elles contiennent, outre les constituants cellulaires classiques, de nombreux cytosymbiotes, en particulier de nombreuses bactéries et autres champignons (Bianciotto & al., 2000 ; Hijri & al., 2002) ; ces divers microorganismes dont le rôle n’est pas encore connu avec précision, jouent très certainement un rôle très important au niveau de la rhizosphère. Chaque spore est entourée d’une paroi qui provient de l’hyphe sporogène avec laquelle elle reste parfois en continuité. Pendant la croissance de la spore, la paroi différencie plusieurs couches. Lorsque la croissance de la spore est terminée, toutes les couches sont formées, mais peuvent encore subir des transformations qui affectent la couleur, la rigidité. La morphologie des spores a été le premier critère retenu pour l’identification des diverses espèces (appelées parfois morpho-espèces) ; on utilisait la taille, la couleur, l’ornementation, les modalités de groupement dans les sporomes, mais surtout la structure de la paroi. En 1983, Walker a standardisé la technique de description des parois et de nouvelles contributions taxonomiques et systématiques sont venues compléter la monographie des Endogonaceae de Gerdemann & Trappe (1974). Actuellement on complète ces données morphologiques par d’autres techniques pour délimiter les taxons. Remarques : - Dans tous les genres, sauf Gigaspora, certaines espèces produisent des spores intraracinaires. - La colonisation des racines de la plante-hôte peut également se faire à partir de fragments d’hyphes du sol ou directement à partir de mycosymbiotes issus des racines d'une plante voisine. IV. Quelques techniques permettant d’observer ces structures 1. Observation des hyphes, arbuscules et vésicules Sur des coupes minces de racines mycorhizées, l’observation microscopique directe des hyphes n’est pas possible ; les composants internes des cellules racinaires et les pigments localisés dans les parois masquent les structures fongiques. Un traitement préalable est indispensable. Celui que nous proposons est extrait du livre de Brundrett M., N. Bougher, Dell B., T. Grove et Malajczuk N., publié en 1996 par le Centre Australien de Recherches Internationales en Agriculture : Working with Mycorrhizas in Forestry and Agriculture (chapitre 4.2, p. : 179-183).

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
9
- 1e étape : Rendre les racines mycorhizées transparentes Les racines débarrassées de toutes particules de terre sont traitées avec une solution de KOH à 10% afin de détruire le cytoplasme des cellules végétales, ses inclusions et la majorité des pigments pariétaux. La durée du traitement dépend de la température. - À la température ambiante, 2 à 3 jours sont nécessaires, - au bain-marie 60-90°C, 2 à 4 heures, - dans un four (ou mieux un autoclave) à 120°C, 5 à 20 minutes suffisent. Des essais sont toutefois nécessaires, la durée doit être augmentée si les racines sont plus âgées ou riches en dérivés phénoliques. Avant traitement, les fragments de racines à observer peuvent être conservés dans l’éthanol (ou le méthanol) à 60%. - 2e étape : Faire agir un colorant pour mettre en évidence les hyphes Le colorant recommandé est le noir de chlorazol en solution dans le lactoglycérol à 0,03%. Préparation : Dissoudre 0,03 g de noir de chlorazol [CBE = Chlorazol Black E (Direct Black 38 CI n° 30235)] dans 33,3 g d’eau bidistillée, ajouter 33,3 g d’acide lactique puis la même quantité de glycérol. D’autres colorants peuvent être utilisés, le bleu trypan à 0,05% dans lactoglycérol (1/1/1 - acide lactique / glycérol / d'eau), le bleu lactique, la fuschine acide, l’encre noire… les contrastes sont toutefois nettement moins prononcés et les structures fongiques se décolorent assez rapidement. Les coupes colorées peuvent être conservées dans le lactophénol ou dans le glycérol aqueux à 50%. 2. Observation des spores L’extraction des spores n’est pas difficile, mais elle nécessite la mise en œuvre d’un matériel qui n’est disponible que dans certains laboratoires (en particulier une colonne de tamis et une centrifugeuse). La technique présentée est proposée depuis avril 2001 par le Laboratoire de Microbiologie agricole, Université agricole de Huazhong, Wuhan, Chine. Ce protocole a été mis au point en collaboration avec le Laboratoire de Phytoparasitologie de l’INRA-CMSE, Dijon, France. L’intégralité des textes et des schémas est disponible à l’URL suivant : http://www.dijon.inra.fr/mychintec - À l’aide d’un emporte-pièce de 1-2 cm de Ø, prélever une portion de sol contenant les racines de la plante-hôte accompagnées d’un échantillon de sol de la rhizosphère. - À l’aide d’un broyeur, que l’on fait agir quelques secondes, fragmenter la terre et les racines afin de détacher les spores de leurs hyphes extra- et intra-racinaires ; ne pas prolonger ce broyage afin de ne pas abîmer les spores. - Préparer la colonne de tamisage. Cette colonne est constituée d’une série de tamis à mailles rigoureusement calibrées ; seuls deux tamis sont indispensables, le tamis à mailles de 710 µm qui sera posé sur le tamis à mailles de 45 µm, il est inutile de descendre en dessous, les spores étant très grandes. - Le broyat (surtout la partie argileuse, la fraction sableuse ayant sédimenté dans le broyeur) est placé dans le tamis supérieur et lavé à l’eau courante pendant quelques instants. Le tamis supérieur retient les morceaux de racines (et les spores dépassant 710 µm ce qui est exceptionnel), le tamis inférieur contient des particules terreuses et les spores mesurant entre 45 et 710 µm ; les spores plus petites sont perdues, mais elles sont souvent très peu nombreuses.

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
10
On fait passer de l’eau (lavage) sur lebroyat placé dans le tamis supérieur.
Le tamis inférieur contient les éléments dontla taille est comprise entre 0,7 mm et 0,045 mm.
Les spores inférieures à 45 µm de Ø sont perdues mais elles sont très peu nombreuses.
- Transvaser le contenu du tamis de 45 µm dans un petit bécher. Agiter et transvaser rapidement dans des tubes à centrifuger de 50 mL, en ne remplissant les tubes qu’à moitié.
- Compléter avec une solution de saccharose à 60% injectée au fond du tube à l’aide d’une seringue prolongée d’un tuyau. On doit voir la séparation entre les 2 solutions, la phase aqueuse au-dessus, la phase sucrée en dessous. Fermer les tubes. - Centrifuger les tubes fermés pendant 2 à 3 minutes. Les spores viennent se placer à l’interface phase aqueuse / phase sucrée. - Prélever les spores à l’aide de la seringue prolongée par le tuyau en commençant par la partie supérieure et en effectuant un mouvement circulaire. Nettoyer le tamis de 45 µm.
- Déposer le contenu de la seringue sur le tamis de 45 µm ; laver pour éliminer toute trace de saccharose. - Le contenu du tamis de 45 µm est finalement transféré dans une boîte de Pétri en verre. À ce stade, les spores peuvent être stockées à 4°C pendant plusieurs semaines (il est toutefois nécessaire de faire une vérification tous les 3-4 jours pour éliminer immédiatement les spores parasitées). - Observer à la loupe binoculaire, séparer les divers morphotypes avec une pince fine. Ces morphotypes peuvent faire l’objet d’une expertise taxonomique, être inoculés à des plantes, envoyés dans un laboratoire pour étude de l’ADN… - Le montage des spores pour examen au microscope se fait dans le lacto-glycérol-polyvinylique, directement et après séjour dans le Melzer. Les deux montages étant situés côte à côte sur une même lame (mettre environ 10 spores par montage). Au bout d’une trentaine de secondes, appuyer légèrement sur la lamelle pour casser quelques parois sporiques. IV. Caractères et principaux genres du phylum des Glomeromycota 1. Caractères des Glomeromycota Ces champignons ne possèdent plus de cellules flagellées, mais leurs autres caractères sont ceux de champignons peu évolués. On peut citer : - des hyphes non ou très peu cloisonnées (ou lorsqu’elles sont âgées avec uniquement quelques ébauches de cloisons), - une structure cœnocytique, avec de très nombreux noyaux dans le même cytoplasme,

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
11
- l’absence de reproduction sexuée, mais la présence d’anastomoses (comme chez les Dikarya) permettent des fusions cytoplasmiques assurant un mélange de noyaux pour obtenir des hétérocaryons (mécanisme très important chez des champignons dépourvus de reproduction sexuée), - des spores de grande taille munies de parois généralement de structure complexe, - ce sont des symbiotes obligatoires formant des arbuscules (souvent accompagnées de vésicules), - ils sont peu spécifiques, une même espèce pouvant vivre en symbiose avec de nombreuses plantes différentes. 2. Un peu d’histoire d’après Walker and Schüßler [2002] L’historique détaillé des Glomales (sensus lato) est donné dans la publication de Walker et Schüßler ; nous en résumons ci-dessous quelques étapes. - En 1809, le mycologue allemand, Link crée le genre Endogone dans lequel il regroupe les champignons donnant des mycorhizes à arbuscules (et quelques zygomycètes ECM). - En 1844, les frères Tulasne font la description du genre Glomus dans lequel ils distinguent 2 espèces, Glomus microcarpum et Glomus macrocarpum. - En 1875, Berkeley et Broome décrivent le genre Sclerocystis. - En 1922, Thaxter place tous les Glomus dans le genre Endogone, qu’il place dans la famille des Endogoneae avec le genre Sclerocystis. - La même année, Bucholtz (1922) place les Endogonaceae dans le Mucorales (affinité des Endogone avec les Mortierellaceae) au sein des Zygomycètes. - En 1953, Moreau place les Endogonaceae dans leur propre ordre, celui des Endogonales, et ses travaux sont confirmés par Benjamin (1979). - En 1974, Gerdemann et Trappe font éclater le genre Endogone et le remplacent par 4 genres Glomus, Sclerocystis, Acaulospora et Gigaspora dans leur monographie des Endogonaceae. - En 1983, Walker standardise la technique de description des parois sporales afin de délimiter les taxons. - En 1990, deux nouveaux genres Scutellospora et Entrophospora, sont créés, trois familles (Glomaceae, Gigasporaceae et Acaulosporaceae) reconnues et Morton et Benny créent l’ordre des Glomales où ils placent ces champignons AM (arbuscular Micorrhizes). - En 1995, le genre Glomites est créé par Taylor et al. pour nommer les champignons fossiles dont les caractéristiques concordent étroitement avec celles des Glomus actuels. - Fin XXe / début du XXIe siècle, les données moléculaires commencent à arriver et de profonds remaniements commencent. Des nouveaux genres sont créés, d’autres sont renommés. Pour identifier les genres 4 jeux de caractères morphologiques sont utilisés : 1. La structure des divers constituants de la mycorhize (hyphes, arbuscules, vésicules, cellules auxiliaires) quand la racine existe, ce champignon ne se développant pas en absence de la racine de la plante-hôte. 2. Les modalités de formation des spores, terminale, intercalaire, dans l’axe de l’hyphe ou latéralement, sur un pédicelle ou non, continuité ou non avec les couches de la paroi… 3. La structure de la paroi qui comprend souvent plusieurs couches qui peuvent être mucilagineuses, granuleuses, membraneuses, gonflées dans l’acide, permanentes ou évanescentes, amorphes, coriaces, flexibles ou rigides, ± épaisses, colorées ou non par le Melzer, munies de petites excroissances…

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
12
4. Le mode de germination, par le lumen, par un flexible, par percement direct de la paroi… Sur le site Web de l’INVAM International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi : http://invam.caf.wvu.edu/index.html
Une importante documentation actualisée est disponible. Elle présente les carac-tères morphologiques, les techniques d’étude au laboratoire, des clés de détermination des genres. Pour de très nombreuses espèces il y a une description complète de toutes les structures rencontrées, accompagnées de nombreuses photographies en couleur avec des gros plans dévoilant le détail des couches de la paroi sporale. http://invam.caf.wvu.edu/index.html Plusieurs publications récentes sont consacrées à la systématique actuelle des Mycota. - 2006, Phylogeny of the Glomeromycota de Redecker & Raab P., - 2007, A higher-level phylogenetic classification of the Fungi de Hibbett D.-S. & al., 4 ordres et 10 genres sont retenus au sein du phylum des Glomeromycota. - 2010, The Glomeromycota, a species list with new families and new genera, de A. Schüßler et C. Walker. - 2011, Advances in Glomeromycota taxonomy and classification de F. Oehl, E. Sieverding , J. Palenzuela, K. Ineichen et GA de Silva. Dans cette dernière publication, sont acceptées 3 classes, 5 ordres, 14 familles et 29 genres pour environ 230 espèces. Le tableau ci-dessous, adapté de cette publication, place les 29 genres admis en 2011, dans les familles et ordres respectifs.
ORDRES FAMILLES GENRES Archaeosporaceae Archaeospora, Intraspora Geosiphonaceae Geosiphon Archaeosporales Ambisporaceae Ambispora (= Appendicispora) Acaulosporaceae Acaulospora, Kuklospora
Diversisporacea Diversispora, Tricispora, Otospora, Redeckera
Sacculosporaceae Sacculospora Diversisporales
Pacisporaceae Pacispora Gigasoporaceae Gigaspora Dentiscutataceae Dentiscuta, Quatunica, Fuscutata Racocetraceae Racocetra, Cetraspora Gigasporales Scutellosporaceae Scutellospora, Orbispora
Glomeraceae Glomus, Septoglomus, Funelliformis, Simiglomus Glomerales Entrophosporaceae
(= Claroideoglomeraceae) Claroideoglomus, Viscospora, Entrophospora, Albahypha
Paraglomerales Paraglomeraceae Paraglomus Glomus est le genre contenant le plus grand nombre d’espèces avec plus de 70 morphotypes différents. Les spores prennent naissance à l’extrémité de l’hyphe sporogène et restent

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
13
souvent attachées à celle-ci à maturité ; leurs parois sont complexes et possèdent plusieurs couches. Gigaspora et Scutellospora ne forment pas de vésicules, mais présentent des cellules auxiliaires à surface épineuse dans le genre Gigaspora, presque lisse dans le genre Scutellospora ;ils forment leurs spores sur un petit bulbe. Acaulospora et Entrophospora forment leurs spores au niveau d'un saccule sporifère, situé dans l’axe de l’hyphe sporogène chez Entrophospora, latéral chez Acaulospora. Ces deux genres présentent une paroi interne flexible. Pacispora genre récent créé pour regrouper des espèces voisines des Glomus mais ayant des parois internes élastiques. Diversispora genre récent créé pour des Glomus du groupe C ayant plus d’affinités avec les Acaulosporaceae qu’avec les Glomeraceae ; fait établi à la suite d’études moléculaires (Walker et Schüßler, 2004). Archaeospora le saccule sporifère est porté par un court pédicelle qui le sépare de l’hyphe sporogène ; les couches flexibles de sa paroi ne sont pas colorées par le Melzer. Paraglomus possèdent des acides gras particuliers qui ne se trouvent pas chez les autres Glomeromycota, forment des petites spores hyalines mais ne présentent pas de caractères morphologiques permettant de les distinguer des Glomus ; les données moléculaires les placent toutefois dans une branche séparée de celle des Glomus. Geosiphon forme des spores glomoïdes, mais il est le seul membre du phylum à entretenir un autre type de symbiose (Schüßler et 1994 al.) - voir ci-dessous. Cas particulier du géosiphon : Geosiphon pyriformis (Kütz.) Wettstein On parle beaucoup de ce champignon, mais en réalité il est très rare ; le géosiphon n’a été signalé que 5 fois en Allemagne, trouvé une fois près de Munich en 1970 ; actuellement il n’y a qu’une seule station mondiale connue, près de Bieber dans les montagnes de Spessart en Allemagne. Schéma simplifé d’après un original de Shlüssler et Wolf, 2005
- Cette espèce a été décrite pour la première fois par Kützing en 1849 et nommée Botrydium pyriforme. - Von Wettstein, en 1915, le présente comme étant une algue siphonnée multinucléée donnant des petites vésicules en forme de poires inversées contenant des cyanobactéries

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
14
(algues bleues à cette époque) endosymbiotiques, une symbiose entre 2 algues, il note cependant la présence de chitine (substance présente dans la paroi des hyphes fongiques), il change le nom de genre et le nom scientifique devient Geosiphon pyriforme. La description était valide mais l’épithète pyriforme inexact, l’orthographe correcte étant pyriformis. - En 1933, Knapp reconnaît la structure fongique et fait adopter par la communauté scientifique, le géosiphon, comme étant un cyanolichen (lichen à cyanobactéries), le champignon étant placé dans les phycomycètes (terme actuellement obsolète). - Ce n’est que très récemment, fin du XXe / début du XXIe siècle, que cette lichénisation et le statut taxonomique de cette espèce ont été remis en cause, les études moléculaires ayant montré que ce champignon appartenait au phylum des Glomeromycota, ce qui était tout à fait inattendu. Le géosiphon vit dans la partie superficielle d’un sol humide, pauvre en nutriments ; il présente :
- une partie hypogée, un mycélium tubulaire cœnocytique, d’où le nom de genre Geosiphon [du grec geo = terre et siph, siphonis = tubes] qui lui a été attribué ; - une partie épigée formée par des vésicules obpyriformes qui se forment lorsque les hyphes rencontrent des Nostoc pouvant vivre librement dans le sol (Nostoc punctiforme) d’où le terme de pyriformis attribué à cette espèce [du latin pyrus = poire et forma = forme].
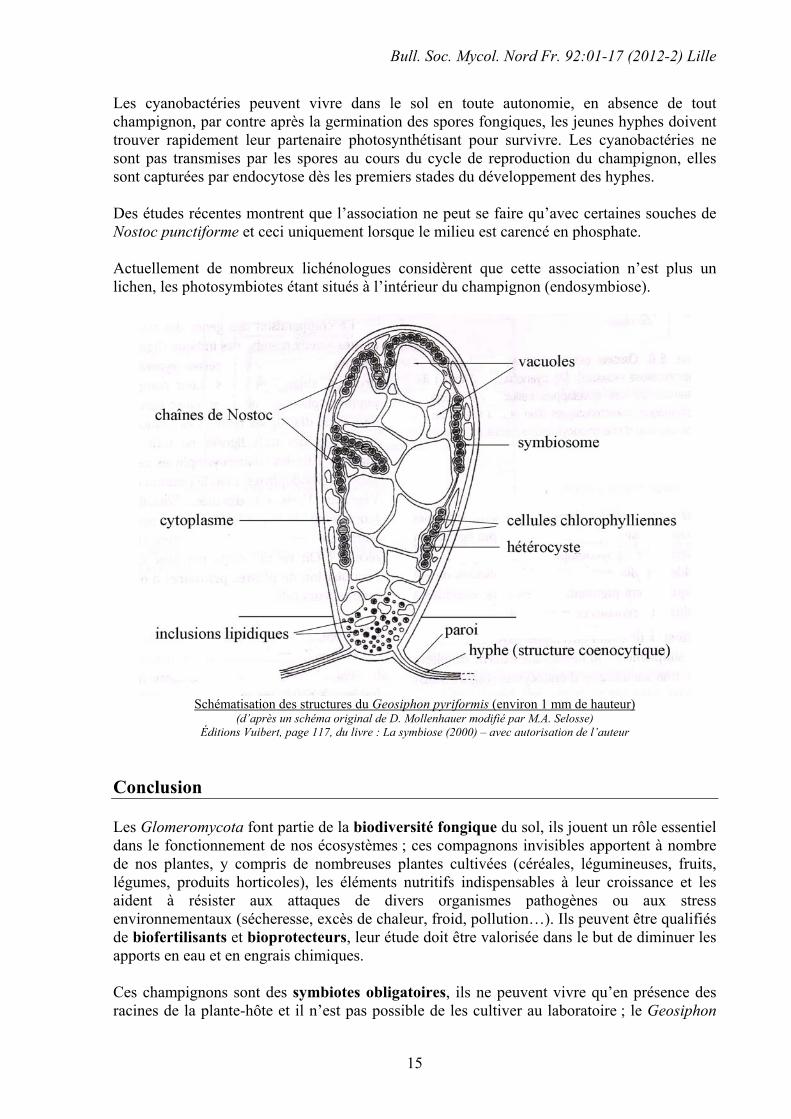
Photo issue du site web : http://www.tu-darmstadt.de/fb/bio/bot/schuessler/geosiphon/geosiphon_home.html Ces vésicules sont des renflements unicellulaires de 1-2 mm de hauteur, ils vivent en moyenne 6 mois (dans des cultures de laboratoires) ; la partie basale en contact avec le sol contient de nombreuses gouttelettes lipidiques, la partie centrale renferme de nombreuses vacuoles, la partie apicale périphérique est occupée par les cyanobactéries (chaînes de petites cellules ayant environ 0,5 µm de Ø) incluses dans des invaginations de la membrane cytoplasmique du renflement pyriforme ; l’ensemble de toutes les inclusions contenant les cyanobactéries forme le symbiosome, dans lequel la structure des cyanobactéries est profondément modifiée ; elles gardent cependant la double autotrophie, elles absorbent le dioxyde de carbone (CO2) mais également l’azote atmosphérique (N2) au niveau des hétérocystes. La zone d’échanges entre le champignon et la cyanobactérie semble comparable à celle que l’on observe au niveau de l’interface endomycorhize / plante.
Les hyphes n’entourent pas les cellules du photosymbiote (cas des champignons lichénisés), elles se déforment pour le faire pénétrer à l’intérieur de leur cytoplasme (endosymbiose)
(d’après Yu-Ting Wu, Yen-Zeng Wang et Chiang-Her Yen, 2004 réalisé à partir d’un schéma de Mollenhauer de 1996). Documents disponibles sur le Net en fichier PDF (il suffit de taper Geosiphon dans les moteurs de recherche
Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
15
Les cyanobactéries peuvent vivre dans le sol en toute autonomie, en absence de tout champignon, par contre après la germination des spores fongiques, les jeunes hyphes doivent trouver rapidement leur partenaire photosynthétisant pour survivre. Les cyanobactéries ne sont pas transmises par les spores au cours du cycle de reproduction du champignon, elles sont capturées par endocytose dès les premiers stades du développement des hyphes. Des études récentes montrent que l’association ne peut se faire qu’avec certaines souches de Nostoc punctiforme et ceci uniquement lorsque le milieu est carencé en phosphate. Actuellement de nombreux lichénologues considèrent que cette association n’est plus un lichen, les photosymbiotes étant situés à l’intérieur du champignon (endosymbiose).
Schématisation des structures du Geosiphon pyriformis (environ 1 mm de hauteur)
(d’après un schéma original de D. Mollenhauer modifié par M.A. Selosse) Éditions Vuibert, page 117, du livre : La symbiose (2000) – avec autorisation de l’auteur
Conclusion Les Glomeromycota font partie de la biodiversité fongique du sol, ils jouent un rôle essentiel dans le fonctionnement de nos écosystèmes ; ces compagnons invisibles apportent à nombre de nos plantes, y compris de nombreuses plantes cultivées (céréales, légumineuses, fruits, légumes, produits horticoles), les éléments nutritifs indispensables à leur croissance et les aident à résister aux attaques de divers organismes pathogènes ou aux stress environnementaux (sécheresse, excès de chaleur, froid, pollution…). Ils peuvent être qualifiés de biofertilisants et bioprotecteurs, leur étude doit être valorisée dans le but de diminuer les apports en eau et en engrais chimiques. Ces champignons sont des symbiotes obligatoires, ils ne peuvent vivre qu’en présence des racines de la plante-hôte et il n’est pas possible de les cultiver au laboratoire ; le Geosiphon

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
16
pyriformis, qui vit en association avec les mousses (Anthoceros et Blasia) et dont la lignée évolutive est incluse dans celle des Glomeromycota doit faire l’objet de recherches complémentaires ; cette espèce, que l’on sait cultiver au laboratoire, étant très certainement capable de s’engager dans des associations endomycorhiziennes avec d’autres plantes et pourrait constituer un excellent modèle de laboratoire. Enfin, il ne faut pas oublier de rappeler que les Glomeromycota sont les descendants actuels des premiers Mycota apparus sur notre planète ; c’est dans ce groupe que l’on trouve les plus anciens fossiles de champignons. Des fossiles d’arbuscules ont été trouvés dans des terrains du Dévonien (-400 MA) et des spores typiques du phylum dans des roches de l’Ordovicien (-460 MA) ; les Glomeromycota étaient donc présents lors des premiers stades de colonisation des terres émergées par les plantes. Ces stratégies symbiotiques ont donc très certainement joué un rôle déterminant lors de la sortie des eaux et directement contribué à la survie et au développement des végétaux sur les continents. Bibliographie Seuls sont référencés les tirés à part et livres personnels utilisés pour écrire ces quelques pages d’introduction à l’étude des Glomeromycota. Ahmadjlan V., 1993. The Lichen Symbiosis, New York, 250 pages.. Brundrett M., N. Bougher, Dell B., T. Grove et Malajczuk N., 1996. Working with
Mycorrhizas in Forestry and Agriculture, Centre Australien de Recherches Internationales en Agriculture (ACIAR), 383 pages.
Dworkin M., Falkow S., Rosenberg E., Schleifer K.-H., et al., 2006. Prokaryotes : A Handbook on the Biology of Bacteria : Symbiotic Associations, Biotechnology, Applied Microbiology, 959 pages.
Gerdemann J.-W. & Trappe J.-M., 1974. Endogonceae in the Pacific Northwest. Mycologia, Mem. 5:1-76.
Godfroy O., 2008. Études génétique et moléculaire de deux gènes de Medicago truncatula, DMI3 et RPG, contrôlant l'établissement de symbioses racinaires, Thèse de Doctorat, université Paul Sabatier de Toulouse, 232 pages.
Hibbett D.-S. & al., 2007. A higher-level phylogenetic classification of the Fungi. Mycological, Research, 111(5): 509-547.
Le Tacon F. et M.-A. Selosse, 1997. Le rôle des mycorhizes dans la colonisation des continents et la diversification des écosystèmes terrestres, Rev. For. Fr. XLIX - nunéro spécial, pages 15 à 24.
Oehl F., E. Sieverding , J. Palenzuela, K. Ineichen et GA de Silva, 2011. Advances in Glomeromycota taxonomy and classification.
Redecker D. & Raab P., 2006. Phylogeny of the Glomeromycota (arbuscular mycorrhizal ungi) : recent developments and new gene markers, Mycologia 98(6):885-895.
Schüßler A., D. Mollenhauer, Ε. Schnepf and Μ. Kluge, 1993. Geosiphon pyriforme, an Endosymbiotic Association of Fungus and Cyanobacteria : the spore structure resembles that of arbuscular mycorrhizal (AM) Fungi, Botanica acta, 10 pages.
Schüßler A., Schwarzott D. and C. Walk, 2002. Glomeromycota rRNA genes, the diversity of myths, Mycorrhiza, Vol. 13, 4:233-236.

Bull. Soc. Mycol. Nord Fr. 92:01-17 (2012-2) Lille
17
Schüßler A. et C. Walker, 2010., The Glomeromycota, a species list with new families and new genera.
Selosse M.-A. & Le Tacon F., 2000. La symbiose, structures et fonctions, rôle écologique et évolutif, Editions Vuibert, 154 pages.
Selosse M.-A., 2001. Les stratégies symbiotiques de conquête du milieu terrestre par les végétaux, éditions scientifiques et médicales Elsevier SAS, Année Biol. 40(2001)3-20.
Sjöberg J., 2005. Arbuscular Mycorrhizal Fungi Occurrence in Sweden and Interaction with a Plant Pathogenic Fungus in Barley, Doctoral thesis, Swedish University of Agricultural Sciences, Uppsala, 56 pages.
Strullu, D.G. 1991. Les mycorhizes des arbres et plantes cultivées. Collection Tec. & Doc., Lavoisier, Paris, 250 pages.
Walker C., 1983. Taxonomic concepts in the Endogonaceae : spore wall characteristics in species descriptions. Mycotaxon 18:443-455.
Wu Yu-Ting, Yen-Zeng Wang and Chiang-Her Yen, 2004. Geosiphon pyriforme is evidenced to be an ancestral member of the Glomales by morphological identification and SSU rRNA analysis, 12 pages.
Appel à contributions pour notre prochain bulletin semestriel
Les pages de notre bulletin sont ouvertes à tous les membres de la SMNF et celui-ci ne peut être élaboré sans vos contributions. Nous espérons donc vos propositions d’articles, même très courts (1 ou 2 pages), pour notre prochain bulletin semestriel. Sachez que les responsables du bulletin, vous aideront pour la rédaction et la mise en page éventuelle de vos photos.


















