
Modélisation des pertes de rendement des cultures de...
151
Thèse présentée en vue de l’obtention du grade de Docteur en sciences et gestion de l’environnement par Christophe MACKELS Jury : Pr.C.Périlleux Université de Liège, sciences de la vie Président Pr.L.Hoffmann CRP Gabriel Lippmann, Environnement et agro-biotechnologies Rapporteur Dr.P.Delfosse CRP Gabriel Lippmann, Environnement et agro-biotechnologies Rapporteur Dr.M.O. Bancal Institut National Agronomique de Paris-Grignon Examinateur Dr.B.Tychon Université de Liège, sciences et gestion de l’environnement Examinateur Dr.M.El Jarroudi Université de Liège, sciences et gestion de l’environnement Examinateur Département des Sciences et Gestion de l’Environnement Modélisation des pertes de rendement des cultures de blé d’hiver au Grand-Duché de Luxembourg sur base de l’étude des surfaces photosynthétiquement actives.
Transcript of Modélisation des pertes de rendement des cultures de...

Thèse présentée en vue de l’obtention du grade de Docteur en sciences et gestion de l’environnement
par
Christophe MACKELS
Jury :
Pr.C.Périlleux Université de Liège, sciences de la vie Président
Pr.L.Hoffmann CRP Gabriel Lippmann, Environnement et agro-biotechnologies Rapporteur
Dr.P.Delfosse CRP Gabriel Lippmann, Environnement et agro-biotechnologies Rapporteur
Dr.M.O. Bancal Institut National Agronomique de Paris-Grignon Examinateur
Dr.B.Tychon Université de Liège, sciences et gestion de l’environnement Examinateur
Dr.M.El Jarroudi Université de Liège, sciences et gestion de l’environnement Examinateur
Département des Sciences et Gestion de l’Environnement
Modélisation des pertes de rendement des cultures de blé d’hiver au Grand-Duché de
Luxembourg sur base de l’étude des surfaces photosynthétiquement actives.

2
SOMMAIRE
INTRODUCTION GENERALE................................................................................................................................... 4 Historique de la quantification et de la modélisation des pertes de récoltes. ............................................................... 6 Effets des pathogènes foliaires sur la photosynthèse à l’échelle de la feuille. ............................................................. 8 Historique des études concernant la relation entre la surface verte et le rendement .................................................. 10 Effets des fongicides sur la surface verte et gains en rendement. .............................................................................. 14 Elaboration des rendements et cycles des pathogènes présents sur les feuilles de blé au Grand-Duché de Luxembourg............................................................................................................................................................... 15
PROBLÉMATIQUE ET APPROCHE ENVISAGÉE. ............................................................................................. 19 Hypothèses de travail. ................................................................................................................................................ 19 Approche envisagée. .................................................................................................................................................. 20 Structure du travail..................................................................................................................................................... 21
PARTIE I : METHODOLOGIE D’ACQUISITION DES SURFACES ET DES SURFACES FOLIAIRES VERTES. ....................................................................................................................................................................... 22
CHAPITRE I : Acquisition des surfaces ....................................................................................................................24
I.1. Acquisition des surfaces par l’analyse d’images et validation de la méthode. .................................................... 24 Matériel et méthodes.................................................................................................................................................. 24 Résultats et discussion. .............................................................................................................................................. 33 I.2. Acquisition du pourcentage de surface verte par la méthode d’estimation visuelle et corrélations avec les données issues de la méthode d’estimation par l’analyse d’images........................................................................... 39 Introduction................................................................................................................................................................ 39 Matériel et méthodes.................................................................................................................................................. 41 Résultats et discussion ............................................................................................................................................... 41
CHAPITRE II : Obtention du LAI. ............................................................................................................................ 45
II.1. Obtention et validation d’un modèle de détermination du LAI à partir de valeurs de PAI obtenues à l’aide d’une méthode indirecte............................................................................................................................................. 45 Introduction................................................................................................................................................................ 45 Matériel et méthodes.................................................................................................................................................. 47 Résultats et discussion ............................................................................................................................................... 51 II.2. Estimation de la relation existant entre la fraction du sol couverte par la canopée verte, le LAI et la surface verte foliaire des trois dernières feuilles du blé d’hiver. ............................................................................................ 54 Introduction................................................................................................................................................................ 54 Matériel et méthodes.................................................................................................................................................. 55 Résultats et discussion ............................................................................................................................................... 56
Conclusion générale de la partie I ............................................................................................................................... 62
PARTIE II : MODELISATION DES PERTES DE RENDEMENT. ..................................................................... 64
CHAPITRE I : Validation d’un modèle d’estimation des rendements du blé d’hiver au Grand Duché de Luxembourg. ................................................................................................................................................................. 66
I.1. Validation du modèle de calcul du HAA............................................................................................................. 66 Matériel et méthodes. ............................................................................................................................................ 66 Résultats et discussion........................................................................................................................................... 69
I.2. Validation du modèle de calcul de la matière sèche. ........................................................................................... 75 Matériel et méthodes. ............................................................................................................................................ 75 Résultats et discussion........................................................................................................................................... 78

3
CHAPITRE II : Simplification et amélioration d’un m odèle d’estimation des rendements du blé d’hiver au Grand Duché de Luxembourg..................................................................................................................................... 85
II.1. Simplification du modèle du calcul du HAA. .................................................................................................... 85 II.2. Simplification du modèle de calcul de la matière sèche..................................................................................... 86 II.3. Analyse des biais introduits par l’utilisation des estimations visuelles du pourcentage de surface verte........... 87
Matériel et méthodes. ............................................................................................................................................ 88 Résultats et discussion........................................................................................................................................... 88
II.4. Amélioration de l’application du modèle de la matière sèche à la culture du blé d’hiver sur le site expérimental d’Everlange................................................................................................................................................................ 93
Matériel et méthodes. ............................................................................................................................................ 93 Résultats et discussion........................................................................................................................................... 97 Conclusion .......................................................................................................................................................... 105
Conclusion générale de la partie II ........................................................................................................................... 109
PARTIE III : APPLICATION DU MODELE EN CONDITIONS REELLES..... ............................................... 111
Matériel et méthodes................................................................................................................................................ 112 Résultats et discussion. ............................................................................................................................................ 116 Conclusion. .............................................................................................................................................................. 126
DISCUSSION GENERALE ET PERSPECTIVES ................................................................................................. 129 Contexte et domaine de l’étude................................................................................................................................ 130 Approches envisagées aux différentes étapes. ......................................................................................................... 130 Validité de l’étude.................................................................................................................................................... 132 Synthèse des résultats issus de l’acquisition des données........................................................................................ 133 Synthèse des résultats issus de la validation, la simplification et l’amélioration d’un modèle d’estimation des rendements. .............................................................................................................................................................. 135 Synthèse des résultats issus de la proposition d’un modèle de simulation des risques de perte de rendement. ....... 136 Conclusion et perspectives....................................................................................................................................... 137
BIBLIOGRAPHIE ..................................................................................................................................................... 142

4
INTRODUCTION GENERALE
La politique agricole commune (PAC) et celle du Luxembourg ont pour principaux objectifs
d'accroître la productivité de l'agriculture, d'assurer un niveau de vie équitable à la population
agricole, de stabiliser les marchés, de garantir la sécurité des approvisionnements et d’assurer des
prix raisonnables aux consommateurs.
L'Union européenne s'attache ainsi à promouvoir une agriculture de qualité, respectueuse de
l'environnement et des normes sanitaires1.
Dans ce contexte, l’estimation des pertes de récoltes pour différents scenarii (selon les variétés, les
précédents culturaux, les traitements phytosanitaires, les accidents climatiques) en cours de saison
agricole est intéressante car elle permettrait d’appliquer des produits phytosanitaires de manière
plus raisonnée, de façon à améliorer à la fois le rendement économique des cultures et la
préservation de l’environnement. Sur de grands territoires, la prévision des rendements avant les récoltes intéresse également les
agences gouvernementales, les entreprises et les producteurs, de manière à anticiper leurs actions
(Boote et al., 1996). Les informations précoces sur les rendements et les volumes de production
peuvent aider ces institutions dans la planification des activités de transport, le marketing des
produits agricoles ou la planification des importations de denrées alimentaires.
A l’échelle mondiale, les prix des marchés agricoles sont influencés par l’information sur l’offre
ou la consommation de denrées alimentaires (Marcus and Heitkemper, 1994). L’ajustement des
prix du marché ou les changements de l’offre agricole dans un coin du monde provoquent souvent
un ajustement des prix dans d’autres territoires éloignés. A l’échelle Européenne, c’est la Direction
Générale de l’Agriculture (DG Agri) de l’Union Européenne qui est responsable de la mise en
œuvre et du contrôle des différentes politiques en agriculture. Pour gérer ces politiques, la DG Agri
a besoin d’informations détaillées sur les surfaces cultivées, les rendements des cultures et les
volumes de production (De Winne, 1994). Pour aider la DG Agri dans l’estimation des rendements
attendus, un modèle de croissance des cultures a été combiné à une carte détaillée des sols, des
paramètres pour les différentes cultures et des informations spatialisées sur les cultures pour créer
le Crop Growth Simulation System (CGMS). Ce système, comprend le modèle WOFOST qui
utilise des observations météorologiques pour estimer les statuts des cultures (par ex. stress
hydrique, production de biomasse, etc…) au cours de la saison de croissance et le rendement à la
fin de la saison. D’autres approches ont également été envisagées, au niveau européen, pour 1 Site « europa » de la Commission européenne ; http://euramis.net/pol/agr/index_fr.htm.

5
effectuer des estimations du rendement. L’étude de la tendance statistique des rendements des
années passées est par exemple prise en compte au dernier niveau des sorties de CGMS2. D’autre
part, la télédétection est également mise au service d’un suivi de l’agriculture et complémente le
modèle CGMS dans son estimation des rendements au travers du projet MARS (Monitoring
Agriculture with Remote Sensing techniques).
Parallèlement au travail de recherche mené par l’Union Européenne, des équipes de chercheurs
explorent d’autres approches pour l’estimation et la prévision des rendements sur certaines cultures
dans leur pays. Plus particulièrement, certaines approches sont issues d’équipes étudiant la relation
existant entre les pertes de rendement et le développement des principales maladies et pestes
affectant une culture. Dans le cas présent, la culture étudiée est le blé d’hiver, et les principales
maladies affectant son rendement sont la septoriose, causée par la forme conidienne Septoria tritici
[Robbins et Desmazières], téléomorphe Mycosphaerella graminicola [Fuckel et Schroeter], et la
rouille brune causée par Puccinia triticina (forme asexuée). Ces maladies s’attaquent aux surfaces
foliaires et affectent le rendement au travers des effets qu’elles ont sur la cinétique de dégradation
des surfaces photosynthétiques (Shaw et Royle, 1989b ; Scholes, 1992). Le Modèle Proculture, validé pour la culture du blé tendre d’hiver au Grand-Duché de
Luxembourg (El Jarroudi et al., 2002) permet de faire des avertissements concernant l’application
de produits phytosanitaires par un suivi et une prévision du développement de la septoriose au
cours de la saison de culture. Ce modèle ne présente pas de module adapté à l’estimation des
rendements, or l’estimation des rendements pourrait contribuer à la construction d’un indicateur
plus solide de déclenchement des traitements. Un module « rendement » basé sur une connaissance
des risques de pertes de rendement en fonction du nombre de traitements motiverait l’agriculteur
dans son choix d’appliquer un ou deux traitements.
Ce travail vise à compléter le modèle Proculture par un module d’estimation des pertes de
rendement. Pour atteindre cet objectif, l’impact des stress biotiques et abiotiques sur l’interception
du rayonnement photosynthétiquement actif par les surfaces foliaires vertes du blé tendre d’hiver
au Grand-Duché de Luxembourg va être étudié. L’étude des surfaces vertes et du rayonnement
intercepté par celles-ci pour expliquer et estimer les risques de pertes de rendement est une
démarche qui a été choisie sur base de la connaissance de l’évolution des recherches dans le
domaine de la modélisation des pertes de rendement en présence de pathosystèmes.
2 site officiel : http://agrifish.jrc.it/marsstat/Crop_Yield_Forecasting/cgms.htm

6
Au cours de cette introduction, un bref historique des études dans le domaine de la quantification
et de la modélisation des pertes de récoltes montre le cheminement de la recherche, les différentes
voies explorées, leur comparaison et l’intérêt de certaines par rapport à d’autres dans un premier
volet. Ce premier volet nous amène à l’apparition des premières recherches posant les fondements
de l’approche qui a été envisagée. Les quatre volets introductifs suivants exposent l’évolution des
connaissances dans des domaines formant les piliers de l’approche de ce travail. La problématique
ainsi que la structure et la logique qui animent ce travail seront ensuite exposées au travers du
dernier volet introductif.
Historique de la quantification et de la modélisation des pertes de récoltes. Zadoks (1987) définit 3 périodes dans l’histoire de la recherche sur les pertes de récoltes : la
période exploratoire, d’urgence et d’implantation.
La période exploratoire est caractérisée par un intérêt pour les pertes de récoltes initiées par des
épidémies sévères comme celle du mildiou dans les années 1940. Des concepts et des
méthodologies standardisées de recherche ont alors vu le jour.
La période d’urgence couvre les deux guerres mondiales, période qui a vu paraître un grand
nombre de travaux sur les méthodes pour estimer les pertes de récoltes des aliments de base tels
que la pomme de terre et le blé (Large, 1993 ; Luo et al.,1993).
La FAO (Food and Agriculture Organisation) (Chiarappa, 1971 ; Chiarappa, 1981) a ensuite initié
les premiers efforts pour standardiser les concepts, les méthodes et les estimations des pertes de
rendement (potentiel3, accessible4 et réel5), en réponse à des contraintes (maladies, insectes,
environnement physique, physiologie et génétique). Il a également été fait la promotion de
l’utilisation de modèles mathématiques simples et empiriques pour des études quantitatives
(Zadoks et Schein, 1979). James (1974) expose les modèles à « points critiques 6», « points
multiples7 » et les modèles utilisant l’AUDPC8 (Area Under Disease Progress Curve).
3 Rendement dépendant du rayonnement, de la température et du génotype. Sans ravageurs et sans carence hydrique ou nutritive. 4 Rendement tenant compte des ressources hydriques et minérales de la situation de culture. 5 Rendement obtenu avec toutes les contraintes des conditions de culture (ravageurs inclus) 6 Relie la sévérité de la maladie à un stade déterminé aux pertes de rendement. Le stade choisi étant celui pour lequel la sévérité présente la meilleure relation avec les pertes de rendement. 7 Idem points critiques, mais plusieurs stades sont choisis. 8 Relie les pertes de rendement à l’aire sous la courbe de progression de la maladie obtenue à partir de la sévérité observée à plusieurs stades au cours du développement de la culture.

7
Ces modèles permettent de définir des seuils qui correspondent aux premiers dégâts causés par le
pathogène (seuil de dommage), au seuil à partir duquel l’agriculteur doit agir s’il ne veut aucune
perte due au pathogène (seuil d’action), ou au seuil à partir duquel l’agriculteur doit se préparer à
traiter (seuil d’avertissement).
L’apparition d’ordinateurs de bureau puissants dans les années 80 a permis d’explorer ces concepts
avec cependant la nécessité d’une meilleure coopération interdisciplinaire pour comprendre
comment les rendements sont affectés par des facteurs biotiques et abiotiques durant la période de
formation des rendements (Savary et al., 2006).
Ainsi, la théorie des seuils est basée sur des modèles qui ne prennent pas en compte des paramètres
tels que le rayonnement reçu en fonction de la surface verte, la température ou la variété. Pour ces
raisons, ces modèles sont utiles mais en théorie non transposables d’une situation à une autre
(Rossing, 1991).
C’est également au cours des années 80 que la focalisation de la recherche est passée du
pathogène (symptômes), avec la plante hôte comme propriété secondaire, à un intérêt pour la
croissance de la plante et de son développement comme variable prioritaire. L’effet des maladies
sur la morphologie et la physiologie des plantes a alors été pris en compte en fonction de chaque
pathogène (Boote et al., 1983 ; Kropff et al., 1995 ; Rabbinge et Rijsdijk, 1983). Une étude de
Bryson et al. (1995) sur les pertes de rendement causées par la rouille jaune illustre bien cette
nouvelle orientation des recherches. Ce travail montre que le rendement est prédit de façon stable
sur deux années à partir du cumul des radiations interceptées par les tissus verts alors que la
relation entre les symptômes de la rouille jaune et le rendement diffère d’une année à l’autre.
L’identification, la quantification et la traduction en un certain nombre d’équations des effets
potentiels des maladies sur les plantes (par exemple réduction de la surface verte, réduction du
rythme de la photosynthèse) ont ensuite permis d’estimer la baisse de croissance et de rendement
des peuplements malades (par exemple Bastiaans, 1991 ; Bassanezi et al., 2001 ; Robert et al.,
2006).
Les études ayant pour objectif d’identifier et de comprendre au niveau moléculaire les
perturbations physiologiques engendrées par des maladies ne quantifient pas ces perturbations à
l’échelle de la plante ou du couvert. Ceci explique pourquoi elles ne sont pas abordées ici.
L’identification et la quantification des perturbations engendrées par les complexes parasitaires
foliaires à l’échelle de la feuille, de la plante et du couvert concernent de près notre démarche.
Pour cette raison, l’évolution des connaissances dans ces domaines sera plus détaillée au travers
des trois chapitres qui suivent.

8
Comme l’a énoncé Bastiaans et al. (1994), la détermination des effets d’une maladie sur la
physiologie des feuilles prises individuellement est le premier pas vers la compréhension des
dommages causés par les pathogènes dans une culture.
En accord avec cette remarque de Bastiaans, les connaissances accumulées sur les effets causés par
des pathogènes foliaires sur la photosynthèse de la surface encore verte de la feuille sont abordées
dans la partie introductive suivante.
Effets des pathogènes foliaires sur la photosynthèse à l’échelle de la feuille. Afin de déterminer les effets des parasites sur les rendements, il est nécessaire de quantifier les
effets des parasites sur le fonctionnement des feuilles malades (Bastiaans, 1991).
Pour atteindre cet objectif, Bastiaans (1991) propose un modèle pour estimer les perturbations de
la photosynthèse des feuilles atteintes par un parasite. Il met en relation dans son modèle la
photosynthèse d’une feuille saine (PS0) avec celle présentant les symptômes d’une maladie (PSx).
Un paramètre β permet de prendre en compte un effet de la maladie sur la photosynthèse qui est
égal (β=1) ou supérieur (β>1) à la surface couverte par les symptômes (x) :
PSx / PS0 = (1 – x)β
Selon ce modèle, les maladies foliaires ont un effet sur la photosynthèse égal, supérieur ou
inférieur à la surface couverte par les symptômes. Quand l’effet est égal, la photosynthèse de la
surface foliaire non couverte par les symptômes n’est pas affectée alors que la surface couverte par
les symptômes ne photosynthétise plus. Lorsque l’effet est supérieur à la surface couverte par les
symptômes, le parasite affecte aussi la photosynthèse de la surface ne présentant pas de
symptômes. Bastiaans (1991) propose plusieurs hypothèses pour expliquer ce dernier cas.
Le pathogène peut secréter des enzymes ou d’autres composés phytotoxiques qui diffusent ou sont
emportés par le flux transpiratoire dans les parties non colonisées de la feuille. La croissance du
mycélium à l’intérieur des feuilles peut entraîner des perturbations dans les flux d’eau et la
destruction des chloroplastes. Dans certains cas, l’effet du parasite sur la photosynthèse est
inférieur à la surface couverte par les symptômes. Le parasite ne perturbe alors pas complètement
le fonctionnement de la photosynthèse de la surface présentant les symptômes. Ou encore, une
augmentation de la photosynthèse autour des lésions est provoquée par celles-ci.

9
Au Grand-Duché de Luxembourg et dans les régions du nord de la France, c’est principalement la
septoriose et la rouille brune qui affectent la culture du blé d’hiver (El Jarroudi, 2005 ; Bancal et
al., 2007). Concernant la septoriose, le paramètre β de Bastiaans reste globalement proche de un
(Robert et al., 2006), contrairement à certaines valeurs élevées de ce paramètre calculées pour
d’autres pathogènes nécrotrophes (Bassanezi et al., 2001). La septoriose affecterait donc peu la
photosynthèse des feuilles infectées au-delà de la surface couverte par les symptômes.
Cela semble en accord avec les effets physiologiques immédiats de la septoriose sur les cellules
foliaires de l’hôte. La septoriose agit sur les tissus de l’hôte en détruisant les cellules, soit lorsque
le mycélium pénètre dans celles-ci, soit par l’intermédiaire d’enzymes pectolytiques et
cellulolytiques. Elles perdent alors toute capacité à photosynthétiser ou à transpirer (Shtienberg,
1992). La septoriose n’affecte donc pas directement la photosynthèse des surfaces foliaires vertes.
Cependant, selon les études de Magboul et al. (1992) et de Robert et al. (2006), elle accélère la
sénescence des feuilles qu’elle colonise.
Les effets physiologiques de la rouille brune entraînent une baisse de la photosynthèse des tissus
infectés. Le rapport chlorophylle a / chlorophylle b diminue au fur et à mesure que la maladie se
développe (McGrath et Pënnypacker, 1990). L’effet de la rouille brune sur la photosynthèse de
l’orge a montré que l’assimilation nette de la partie infectée est diminuée, mais que la
photosynthèse totale peut être augmentée (Scholes et Farrar, 1986). Le comportement de puits du
champignon et l’intensification de la respiration seraient à l’origine d’une demande très forte en
carbone, ce qui entraînerait une augmentation locale de la photosynthèse. La stimulation locale de
la photosynthèse permettrait à la plante de compenser en partie la perte d’activité photosynthétique
des tissus infectés. Les études sur la quantification des perturbations de la photosynthèse par la
rouille brune sont assez contradictoires. Shtienberg (1992) obtient des diminutions de 20% à 40%
de la photosynthèse dans un couvert atteint par la rouille brune, et calcule un β=3. A l’échelle
foliaire, le β a été évalué à ~ 1,26 (Spitters et al., 1990 ; Bastiaans, 1991). Une autre étude montre
qu’une feuille de blé infectée à 9% par Puccinia recondita a une réduction de 60% de l’activité
photosynthétique, ce qui suggère un β de 10 (Mac Grath et Pennypacker,1990). Ces études
tiennent peu en compte l’effet de compensation possible entre zones infectées et non infectées, au
sein d’une même feuille ou entre étages foliaires.
Enfin, il a été confirmé récemment par une étude de 5 années menée sur des couverts de blé
attaqués par la rouille brune et la septoriose (Bancal et al., 2007) que ces deux maladies n’affectent
pas l’efficacité d’utilisation du rayonnement par les parties vertes du couvert. Leur travail suggère

10
que la valeur de β=1 peut-être utilisée pour la septoriose et la rouille brune, seules ou en complexe,
pour des couverts soumis à différentes pratiques culturales.
D’autre part, Lopes (2001) a également déterminé le paramètre β du formalisme de Bastiaans pour
deux maladies du haricot. Il suggère d’utiliser, au moyen du formalisme de Bastiaans, la portion
exacte de feuille fonctionnant comme une feuille saine pour le calcul du HAD9 et du HAA10, qui
sont des prédicteurs robustes du rendement pour cette culture. La même année, Bassanezi et al.
(2001) ont obtenu, sur la même plante et pour les mêmes maladies, un HAD et un HAA qui tient
compte de l’effet des maladies sur la proportion de surfaces foliaires vertes adjacentes aux surfaces
foliaires couvertes par les symptômes. Ces nouveaux indices qui tiennent compte des effets des
maladies sur le fonctionnement des surfaces foliaires non couvertes par les symptômes sont mieux
corrélés au rendement que les HAD et HAA anciennement calculés pour ces pathosystèmes
(Bergamin Filho et al., 1997 ; Gianasi, 1999 ; Lopes, 2001). Ceci est d’autant plus vrai que le β de
Bastiaans et la sévérité des maladies sont élevés.
Cette brève synthèse nous suggère que la rouille brune et la septoriose n’ont que peu d’impact sur
la photosynthèse des surfaces asymptomatiques des feuilles malades. Sachant cela, l’historique des
études concernant la relation entre la proportion de surface verte foliaire et le rendement peut être
abordé afin de s’informer des connaissances actuelles se rapprochant du sujet traité.
Historique des études concernant la relation entre la surface verte et le rendement De nombreuses études ont déjà illustré le lien existant entre la croissance, ou le rendement, et le
rayonnement intercepté par diverses cultures (Watson, 1947 ; Bergamin Filho et al., 1997 ; Bryson
et al., 1997 ; De Jesus Junior et al., 2001 ; Robert et al., 2004 ).
En 1947, Watson a suggéré que le rendement est une fonction de l’aire foliaire d’une face des
feuilles présentes dans un volume contenu au-dessus d’une unité de surface horizontale au sol
(LAI pour Leaf Area Index) intégrée sur la saison de croissance. Monteith (1977) a ensuite
suggéré que le rendement en matière sèche produit est une fonction du rayonnement intercepté par
la canopée durant une période de croissance. Le concept fut adapté par Waggoner et Berger (1987)
9 Le HAD ou Healthy Leaf Area Duration est la somme de la proportion de tissus foliaires sains journalier pour toute la saison de croissance. 10 Le HAA ou Healthy Leaf Area Absorption est la somme du rayonnement journalier intercepté par la proportion de tissus foliaires sains journalier pour toute la saison de croissance.

11
pour une utilisation sur des couverts malades en prenant en compte la proportion de surface foliaire
saine (HLAI=LAI(1-X) où X est la sévérité de la maladie exprimée en pourcentage de surface
foliaire couverte par les symptômes) pour chaque jour de mesure, et en l’intégrant sur la saison de
culture (HAD pour healthy leaf area duration). Dans cette étude, ces chercheurs calculèrent
également le rayonnement intercepté par la proportion de tissus sains (HRI pour Healthy Radiation
Interception), en tenant compte du HLAI et du rayonnement incident intercepté chaque jour (Ii). La
loi de Beer Lambert a été utilisée pour calculer le rayonnement intercepté. Ils obtinrent alors le
rayonnement intercepté par la proportion de l’aire foliaire saine au cours de la période de
croissance (HAA pour Healthy leaf Area Absorption) en intégrant le HRI sur la saison avec des
données récoltées chaque semaine. La formule se présente comme suit :
HAA= Ii ({ %SVi[1-e(-kLAIi)]+%SVi+1[1-e(-kLAI
i+1)] }/2 )(ti+1-t)
où Ii est le rayonnement reçu par m2 par jour i (MJ.m-2.jour-1) dans la gamme du Rayonnement
Photosynthétiquement Actif (R.P.A., longueur d’onde de 400 à 800 nm). %SVi est le pourcentage
de surface verte foliaire du jour i, k le coefficient d’extinction des strates foliaires considérées et t
le temps en jours. Le HAA est calculé sur toute la saison de croissance, avec i=1 pour le premier
jour.
Ces approches-ci présentent l’avantage de ne pas devoir prendre chaque maladie ou stress
physique individuellement dans l’étude des pertes de rendement. En effet, les variables utilisées
intègrent les différents facteurs jouant sur le rendement via la dégradation de la surface verte.
Relier le rendement à la sévérité de plusieurs maladies avait déjà causé beaucoup de problèmes
auparavant à cause des interactions qu’elles ont entre elles et avec la plante hôte (James et Teng,
1991).
Par la suite un certain nombre de travaux (Aquino et al., 1992 ; Ferrandino et Elmer, 1992 ;
Johnson et al., 1992 ; Rossing et al., 1992 ; Wright et Gaunt, 1992 ; Bastiaans, 1993 ; Bastiaans et
Kropff, 1993 ; Bastiaans et al., 1994 ; Madeira et al., 1994 ; Bryson et al., 1995 ; Nutter et al.,
1996) établirent une relation directe entre le rendement et la proportion de tissus sains plutôt que
indirectement sur base de la proportion de tissus malades. Dans leur étude de 1997 sur la maladie
foliaire Phaeoisariopsis griseola de Phaseolus vulgaris L. , Bergamin Filho et al. comparent
l’approche basée sur le rayonnement intercepté par les tissus foliaires sains durant la saison de
culture à l’approche basée sur la relation qui lie la sévérité de la maladie au rendement. Cette
recherche met en évidence une relation linéaire stable, pour deux années de données obtenues
∑
−
=
1
1
n
i

12
chaque fois sur deux sites, entre le rayonnement intercepté par la surface foliaire saine au cours de
la saison (HRI) et le rendement. La comparaison montre également la supériorité de la corrélation
entre le rendement et le HRI par rapport à celle entre le rendement et la progression de la maladie
au cours de la saison (AUDPC pour Area Under Desease Progress Curve). En outre, le HAD et le
HAA qui ont été calculés montrent également une relation linéaire avec le rendement dans
respectivement cinq et quatre cas sur cinq. La même conclusion a été obtenue par Bryson pour la
rouille jaune sur blé d’hiver (Bryson et al., 1997). De plus, Bryson montre qu’il n’y a pas eu de
différence, entre deux années de mesures, du rapport poids sec en grain sur quantité de
rayonnement incident intercepté par les tissus verts à partir de la floraison (R.U.E.11 de 1,4 g / MJ).
Cela indique la présence d’une relation directe entre la quantité de rayonnement intercepté par la
proportion de surfaces foliaires vertes et le rendement en grains obtenu.
En 2000, Gooding et al. montrent, dans une étude exhaustive menée sur dix-sept années et
comparant différents cultivars de blé traités ou non par des fongicides, que l’effet des fongicides
sur le rendement en grain et le poids moyen est étroitement lié à la durée de la surface verte de la
dernière feuille. Le rayonnement intercepté n’est pas pris en compte, mais l’effet sur les
rendements de la durée pendant laquelle la surface verte foliaire est maintenue, ainsi que
l’augmentation de cette durée par les fongicides sont estimés.
Dimmock et Gooding (2002) montrent également que lorsque la durée de vie de la surface verte de
la dernière feuille augmente, la durée de remplissage des grains et le rendement augmentent
également, et cela pour plusieurs années, cultivars et traitements fongicides.
Ces relations varient cependant en fonction du cultivar. En effet, deux variétés peuvent avoir un
remplissage des grains synchrone mais des cinétiques de sénescence foliaire différentes. Par
ailleurs, une même augmentation de la durée de vie des feuilles (LAD pour Leaf area duration)
pour deux variétés après l’application de fongicides peut se produire à différents stades du
remplissage des grains. Si la LAD survient plus tard, la surface verte restante et le rendement sont
alors moins importants. Il a également été mis en évidence la possibilité de ne pas obtenir
d’augmentation de rendement pour une augmentation de la LAD entre des parcelles témoins et des
parcelles traitées. En effet, certaines variétés atteignent une limite physiologique qui détermine la
taille des grains, liée au contenu en eau des grains au moment des divisions des cellules de
l’endosperme (Schnyder et Baum, 1992 ; Macbeth et al., 1996). Dès lors, une augmentation de la
11 Le R.U.E. ou Radiation Use Efficiency indique le rythme auquel le rayonnement intercepté par le couvert est utilisé pour produire de la biomasse au travers des processus de photosynthèse. Il est variable au cours du temps, mais a été utilisé avec succès sous forme de valeur constante sur la saison (Madden et al., 2007).

13
durée de la surface verte des feuilles ne permet plus d’obtenir de gains supplémentaires lorsque la
culture de ce type de variété est traitée.
Pepler et al. (2005) déterminent une limite potentielle à l’association entre la durée de la surface
verte de la dernière feuille et le rendement en grain. L’analyse de deux années de données sur 6
cultivars et plusieurs traitements fongicides combinés à la ré-analyse des deux années de données
de Dimmock et Gooding (2002a) les amènent à la conclusion que toute augmentation de la durée
de la surface verte au-delà de 700°C jours après la floraison ne se traduit pas par une augmentation
de rendement. La relative constance de ce seuil de 700°C jours suggère que la relation entre la
durée de la surface verte et le rendement en grains dépend du stade de développement du blé.
Ils montrent également sur ces quatre années que la limite de l’association entre durée de vie de la
canopée et gain en rendement est plus étroitement reliée aux stades de développement en °C jours
de la culture que la limite estimée en jours.
Récemment, un modèle permettant de calculer la matière sèche journalière aux stades qui
correspondent à la période de remplissage du grain et la croissance du couvert ainsi que son
rendement a été mis au point pour le blé d’hiver par l’unité Environnement et Grandes Cultures de
l’INRA Paris Grignon. Ce modèle utilise le formalisme de Bastiaans (Robert et al., 2004 ;
Bastiaans, 1991) et l’approche de Monteith (Monteith, 1972) et tient compte de la mobilisation des
sucres accumulés dans les tiges durant la période préfloraison pour le remplissage des grains. Le
rayonnement intercepté par la surface verte pour chaque jour de remplissage des grains, de la
floraison à la maturité complète est également pris en compte (Bancal et al., 2007).
L’étude, qui a été menée sur 5 années de données issues d’expérimentations aux champs, montre
clairement qu’un modèle simple entre croissance et rendement impliquant du blé d’hiver attaqué
par la rouille brune et la septoriose peut être dérivé :
a) d’une estimation du R.U.E. des cultures saines qui prennent en compte d’autres
stress.
b) d’une simple relation entre croissance et rendement, quelle que soit l’épidémie,
pourvu qu’une constante quantité de réserves pré-floraison soit remobilisée aux
grains pour une variété donnée.
c) d’une estimation correcte du aPAR (Rayonnement photosynthétiquement actif
absorbé ou absorb Photosynthetic Active Radiation) par traitement, en utilisant la
somme du rayonnement intercepté par surface verte journalière de chaque strate.
Autrement dit, les résultats de cette étude suggèrent fortement qu’un modèle simple basé sur la
surface verte foliaire par strate et les réserves de préfloraison peuvent prédire à la fois la croissance

14
et les rendements du blé tendre d’hiver. Et ceci pour des cultures atteintes d’épidémies tardives de
maladies foliaires sur une variation de pratiques culturales.
Un certains nombre de facteurs ont un impact sur le maintien de la surface foliaire verte et le lien
qu’elle a avec le rendement. Parmi ces facteurs, les fongicides ont été abordés à plusieurs reprises
dans la littérature.
Afin de mieux comprendre l’influence du facteur « fongicide » sur les surfaces vertes et le
rendement, le volet suivant développe plus particulièrement les connaissances présentes sur le sujet
dans la littérature.
Effets des fongicides sur la surface verte et gains en rendement. En 2000, Gooding et al. publient une étude qui s’étend sur dix-sept années de données. L’étude
porte sur la décroissance de la surface verte du blé d’hiver au cours de la saison, avec ou sans
fongicides. Dans ces travaux, il est montré que l’effet des fongicides sur le rendement en grains et
le poids moyen est étroitement lié à leur effet sur la durée de la surface verte de la dernière feuille.
Les fongicides retardent la diminution de la surface verte plutôt qu’ils en réduisent la vitesse de
dégradation, une fois celle-ci commencée. L’importance de ces effets varie grandement suivant les
expériences et les cultivars.
Les groupes de fongicides qui contrôlent les pathogènes par l’inhibition de la respiration
mitochondriale, comme les strobilurines ont montré des effets de prolongation de la durée de la
surface verte de la dernière feuille et d’augmentation du poids moyen en grain (Gooding et al.
2000). Cette augmentation est significativement plus importante que celle provoquée par leurs
prédécesseurs comme les triazoles et les morpholines (Bayles et Hilton, 2000 ; Bryson et al. 2000).
Cependant, augmenter la durée de la surface verte n’augmente pas toujours la taille des grains
(Davis et al., 1984). Schnyder et Baum (1992) et Macbeth et al. (1996) proposent que le poids
final du grain soit limité par le contenu maximum en eau du grain. Ils firent cette proposition après
avoir constaté que le nombre de cellules de l’albumen est positivement corrélé au poids du grain à
maturité et que durant la division des cellules de l’endosperme il y a une rapide augmentation du
contenu en eau du grain qui cesse à la fin des divisions cellulaires. La taille physique du grain à ce
stade impose, selon eux, une limitation spatiale de développement du grain. Cette limitation
pourrait dès lors imposer une restriction sur le gain de poids final du grain obtenu en retardant la
sénescence des dernières feuilles. Cette hypothèse est confirmée par Dimmock et Gooding (2002a)

15
qui trouvent que le poids final du grain est corrélé au contenu maximum en eau atteint durant le
développement de celui-ci. Dans leur expérience, ils démontrent sur plusieurs années, cultivars et
position de grains sur l’épi, que les traitements fongicides prolongent souvent la durée de
remplissage des grains de blé sans modifier le rythme de croissance des grains.
En effet, le poids maximum en grains peut être substantiellement augmenté en prolongeant la
durée de remplissage des grains par une augmentation de la durée de la surface verte de la dernière
feuille émergée. A l’exception des cultivars pour lesquels la limite maximale de contenu en eau est
déjà atteinte dans les parcelles non traitées. Ceci suggère que les augmentations de LAD après
avoir atteint le contenu maximum en eau du grain sont moins utiles que les augmentations
effectuées avant le contenu maximum en eau du grain, pour augmenter le remplissage du grain ou
le rendement et le poids spécifique.
En conclusion, les travaux de Dimmock et Gooding montrent que les différents types de fongicides
ont des effets qui confirment la relation qui lie la durée de la surface verte et le rendement.
Ils montrent également qu’une augmentation de la durée de la surface verte due à des fongicides
peut ne pas être accompagnée par une augmentation de rendement pour certains cultivars qui
atteignent déjà un maximum physiologique de remplissage du grain dans les parcelles non traitées.
Elaboration des rendements et cycles des pathogènes présents sur les feuilles de blé au Grand-Duché de Luxembourg. La fécondation des fleurs de blé marque le début de la formation et du remplissage des grains.
Pour cette raison, de nombreux travaux considèrent que le remplissage des grains de blé
commence à la floraison. Ces recherches utilisent les degrés jour à partir de ce stade pour calculer
la durée de remplissage des grains (Pepler et al., 2006; Wiegand et Cuellar, 1981; Calderini et al.,
2001). Comme cela a été cité plus haut, la durée de remplissage des grains des céréales a été
associée à la durée de la surface verte de la canopée, et plus particulièrement celle de la dernière
feuille émergée en considérant l’effet de différents traitements phytosanitaires (Dimmock et
Gooding, 2002), différents cultivars (Hanft et Wych, 1982), et la sécheresse (Egli, 1998).
La durée de remplissage des grains se détermine donc en cumulant les degrés jours de la période
s’étendant de la floraison à la maturité, stade auquel est observée la sénescence partielle ou
complète des dernières feuilles émergées. Cette durée de remplissage est calculée en cumulant les
degrés jours jusqu’à la sénescence complète des trois dernières feuilles, dans le cas où celle-ci
survient avant la maturité des grains. Pepler et al. (2005) ont montré que la durée de remplissage

16
des grains atteint une limite située à un cumul de 700°C jours après la floraison. Par conséquent, la
présence de surfaces foliaires vertes au-delà de ce seuil n’est pas suivie d’une augmentation de
rendement correspondante.
La vitesse de sénescence des dernières feuilles à la période post-floraison dépend aussi des
maladies présentes. Les maladies foliaires du blé concernent de nombreuses maladies dont la
présence et l’importance relative peut varier d’une région à l’autre. Dans notre cas, la sénescence
induite est provoquée principalement par la rouille brune et la septoriose (El Jarroudi, 2005). La
rouille brune (Puccinia triticina) provoque une diminution des surfaces foliaires vertes
approximativement égale à celle des surfaces couvertes par les lésions de l’épiderme dues à la
sporulation du pathogène (Bancal et al., 2007). Par contre, la septoriose (causée par
Mycosphaerella graminicola) induit une sénescence accélérée (Magboul et al., 1992).
Le remplissage des grains est dépendant de la cinétique de dégradation de la surface verte des
dernières feuilles émergées. Cette cinétique est en partie dépendante du développement et de
l’apparition des symptômes de la rouille brune et de la septoriose, ainsi que de l’apparition de leurs
symptômes. Par conséquent, une connaissance du cycle de ces pathogènes par rapport à la période
de remplissage des grains est importante pour la compréhension de l’implication de ces deux
maladies sur la cinétique de dégradation de la surface verte des dernières feuilles durant cette
période.
Le cycle de la rouille brune (Puccinia triticina) par rapport à la période de remplissage des
grains.
La rouille brune appartient à la classe des Basidiomycètes, ordre des urédinales, famille des
Pucciniacées. C’est un champignon biotrophe dont seule la forme asexuée est observée dans nos
régions. A la moisson du blé en été, les urédospores de ce parasite obligatoire sont dispersées par
le vent mais ne trouvent plus de blé pour se développer. Elles se développent alors sur les
graminées adventices jusqu’à l’automne, période à laquelle la rouille brune retrouve son hôte
préférentiel. Elle hiverne sous forme de mycélium sur les repousses de céréales ou sur les semis
d’automne. Le mycélium, engagé dans un monocycle (latence - apparition des pustules à la surface
des feuilles –sporulation - dissémination par le vent – germination - pénétration) est stoppé en
hiver et reprend au printemps. Les monocycles s’enchainent alors en une phase épidémique selon
les conditions climatiques (vent, température et humidité)
Les urédospores, issues de la reproduction asexuée, peuvent être disséminées sur de grandes
distances par le vent. Les spores déposées sur le couvert de blé nécessitent une température
optimale de 15°C, et maximale de 35°C, en présence d’eau pour germer (Clifford et Harris, 1981).

17
Une durée minimale d’humectation de 4h est nécessaire à l’infection en conditions optimales, et la
mort du parasite est observée si une période de sècheresse survient entre la germination et la
pénétration (De vallavieille-Pope et al., 1995). Les cycles d’infection au sein du couvert, avec
dissémination verticale et horizontale dépendent de la température et du vent.
Dans les régions où cette maladie est importante, l’agriculteur peut diminuer le développement de
la maladie par l’application d’un semis tardif, le labour après récolte pour supprimer les graminées
et repousses sur le champ à l’automne, l’utilisation de variétés résistantes et l’obtention d’un
couvert peu dense afin d’éviter la présence d’une humidité stagnante. L’agriculteur évitera aussi
l’application d’une fumure azotée non équilibrée qui favorise le développement de la maladie.
A la floraison, début de la période de remplissage des grains, la présence de la maladie et l’état
d’avancement de l’épidémie dépendent donc de nombreux facteurs tels que la présence de
l’inoculum sur place, les choix de l’agriculteur (variété, date de semis, fumure, traitements
phytosanitaires et la densité de semis) et les conditions climatiques.
Une étude de 4 années (2000 à 2003) sur 4 variétés, menée sur le site d’essai utilisé dans ce travail
montre que cette maladie ne s’est développée que les 2 premières années, avec une apparition des
premiers symptômes à l’épiaison (El Jarroudi, 2005). Pour les années pendant lesquelles elle a été
présente, elle intervient dans la dégradation de la surface verte des dernières strates foliaires qui a
lieu durant la période de remplissage des grains car elle est déjà présente à l’épiaison.
Le cycle de la septoriose (Septoria tritici) par rapport à la période de remplissage des grains.
Septoria tritici appartient aux Dothideomycètes. Mycosphaerella graminicola est la forme sexuée
de S. tritici. Ce champignon nécrotrophe présente deux formes de reproduction qui génèrent deux
types de cycles infectieux permettant le développement de la maladie.
A l’automne, les ascospores (forme sexuée) produit par la forme sexuée sont véhiculées dans l’air,
ce qui expliquerait une répartition homogène des symptômes observés dès la fin de l’automne et
pendant l’hiver sur les cultures de blé (Thomas et al., 1989 ; Lovell et al., 2002).
S. tritici ne peut jouer le rôle d’inoculum primaire qu’à partir de pycnidiospores présents sur les
déchets de culture. Ceux-ci sont lourds et ne peuvent être disséminés que par les gouttes de pluie
sur quelques décimètres. Une infection primaire par S. tritici n’est donc possible que sur la culture
d’un deuxième blé ou sur une succession de céréales sensibles. On peut observer les pycnides de S.
tritici dès l’automne et pendant tout l’hiver, ce qui suggère l’accomplissement de premiers cycles
asexués dès l’apparition des premières feuilles. Au printemps, les pycnidiospores sont formés dans
les pycnides édifiés dans les parties nécrosées des feuilles. Avec l’humidité ambiante, les pycnides
se resserrent et les spores sont expulsées au sein d’une gelée (cirrhe). Ces cirrhes sont alors

18
projetés vers les étages foliaires supérieurs par les éclaboussures créées par les gouttes de pluie.
Les spores disséminées germent et un tube germinatif pénètre au sein de la feuille par les stomates.
S’ensuit une période de latence pendant laquelle le mycélium de S. tritici progresse au sein des
espaces intercellulaires du mésophylle. Après 15 à 20 jours, la croissance du mycélium s’amplifie
et détruit les parois des cellules. Les symptômes apparaissent et des pycnides se forment au niveau
des stomates. La mort des cellules de l’hôte devient importante avec l’apparition des pycnides
(Jorgensen et Smedegaard-Peterson, 1999).
Le déroulement des cycles successifs menant à l’infection des étages foliaires supérieurs dépend
des conditions d’humidité et de température. Pour germer, les spores ont besoin d’eau libre sur les
feuilles. Les conditions précises nécessaires à l’infection ont été détaillées par Lemaire et al.
(2003) : Une heure avec au moins 0,1 mm de pluie, suivie d’une heure avec au moins 0,5 mm de
pluie. Ces pluies doivent ensuite être suivies par au moins 16 heures avec au moins 60%
d’humidité relative. La température doit être supérieure à 4°C durant 24 heures à partir de la
première heure de pluie à 0,1 mm.
L’agriculteur peut diminuer le développement de la maladie par l’enfouissement des résidus de
culture après récolte, l’utilisation de variétés résistantes et les traitements phytosanitaires.
Une étude de 4 années (2000 à 2003) sur 4 variétés, menée sur le site d’essai utilisé dans ce travail
montre que cette maladie est présente chaque année bien avant la floraison (El Jarroudi, 2005). La
cinétique de dégradation de la surface verte durant la période de remplissage des grains est
influencée par l’évolution de cette maladie car celle-ci détruit les cellules de l’hôte.

19
Problématique et approche envisagée.
Hypothèses de travail.
L’objectif de ce travail est de déterminer la validité d’un modèle prévisionnel de simulation des
risques de pertes de rendement basé sur l’étude des surfaces photosynthétiquement actives
(surfaces vertes). L’objectif à terme étant d’estimer les pertes de récoltes à venir au cours de la
saison de culture.
Pour atteindre cet objectif, nous devons tout d’abord obtenir un modèle d’estimation des
rendements validé pour le blé d’hiver au Grand-Duché de Luxembourg. La démarche de ce travail
consiste à modéliser la dégradation de la surface verte du couvert et du rayonnement intercepté par
celle-ci au cours de la période de remplissage des grains et d’établir un lien entre celle-ci et le
rendement. L’aspect prévisionnel sera évalué grâce à l’utilisation de données de rayonnement des
années passées et des pourcentages de surface verte estimés à l’aide du modèle Proculture pour ces
mêmes années.
Les hypothèses de travail auxquelles ce travail sera amené à répondre pour atteindre ces objectifs
sont les suivantes :
� La cinétique de dégradation de la surface verte foliaire d’un couvert malade ou sain et
le rayonnement intercepté par cette surface sont-ils des variables suffisantes pour
estimer le rendement ?
� L’estimation du rendement à partir des variables précitées permet-elle d’obtenir un
modèle prenant en compte l’effet variétal, l’année et les traitements ?
� La perte de surface verte potentielle prédite par Proculture, couplée à un modèle simple
d’estimation des rendements, peuvent-ils aider à gérer les fongicides de manière plus
respectueuse de l’environnement ?

20
Approche envisagée. L’approche envisagée est issue de l’étude de la bibliographie concernant notre sujet.
La littérature met en évidence deux principaux modèles d’estimation des rendements basés sur la
cinétique des surfaces vertes et sur l’interception du rayonnement par celles-ci. Ce travail prend en
compte trois échelles (Feuille – plante – couvert) au travers des méthodes d’acquisition des
données et des deux principaux modèles d’estimation des rendements qui seront testés.
La variété du blé, l’ensemble des paramètres climatiques ainsi que les maladies foliaires ayant une
incidence sur la dégradation de la surface verte des trois dernières feuilles sont pris en compte au
travers de la dégradation de celle-ci (1-fig.1). Le rayonnement photosynthétiquement actif
journalier intercepté par la surface verte est comptabilisé pour chaque strate foliaire (2-fig.1). Un
passage à l’échelle du couvert est effectué en tenant compte ou pas, selon le modèle, de la densité
de semis et des brins moyens (3-fig.1). Deux modèles issus de la littérature sont testés. Une
relation est ensuite effectuée entre les sorties des deux modèles et le rendement.
Le modèle le plus adéquat sera finalement utilisé pour tester son aspect prévisionnel.
Pour cette dernière étape, sera utilisée la dégradation de la surface verte foliaire des trois dernières
feuilles estimée à partir de données issues de plusieurs années passées à l’aide du modèle
épidémiologique Proculture. Le rayonnement incident utilisé en prévisionnel sera également issu
de l’utilisation de données historiques.
Figure 1 : Schéma des principaux facteurs intervenant dans les modèles d’estimations des pertes de rendements basé sur l’étude de la surface verte.
Maladies foliaires
Climat
Variété
1
1
Surface Verte
F1,2 et 3
Sénescence
1
Techniques culturales
1
+ Rayonnement
2
Rendement
Interception du couvert.
Interception par strate.
3

21
Structure du travail.
Ce travail est organisé en trois grands Chapitres. Chacun de ces chapitres fournira des données ou
des informations nécessaires à l’élaboration du chapitre suivant.
Ils sont disposés dans un ordre logique qui va de l’acquisition des données (PARTIE 1) à
l’utilisation de celles-ci comme données d’entrée des modèles qui feront l’objet du chapitre sur la
validation, la simplification et l’amélioration d’u n modèle d’estimation des rendements
(PARTIE II). Le modèle estimé comme étant le plus adéquat des deux testés pour l’estimation des
rendements sur nos données sera utilisé pour effectuer une simulation des risques de pertes de
rendement (PARTIE III) à l’aide de données estimées de façon prévisionnelle et issues de
l’exploitation du modèle PROCULTURE. L’utilisation de ce modèle étant possible grâce à sa
validation au Grand-Duché de Luxembourg par Moussa El Jarroudi durant les années de sa thèse et
du projet MACRY. Un schéma détaillé de la structure de ce travail est présenté en Annexe I.

22
PARTIE I
METHODOLOGIE D’ACQUISITION DES SURFACES ET DES SURFACES
FOLIAIRES VERTES.

23
Note préalable
Cette première partie expose en détail et valide les méthodes utilisées pour obtenir les données
d’entrée les plus déterminantes et difficiles à obtenir des modèles qui seront testés dans la
PARTIE II . Ces données sont les surfaces foliaires vertes ou les pourcentages de surfaces foliaires
vertes et le LAI (Leaf Area Index).
La détermination des méthodes les plus précises, exactes et automatisables est présentée dans le
CHAPITRE I pour l’obtention des surfaces foliaires et des surfaces foliaires vertes, et dans le
CHAPITRE II pour l’obtention du LAI.

24
CHAPITRE I : Acquisition des surfaces foliaires L’acquisition des surfaces comprend ici les surfaces foliaires, qui seront utilisées par après pour
l’obtention du LAI, des surfaces vertes ou du pourcentage de surface verte. Deux méthodes ont été
testées ici.
Une première méthode passe par la prise d’images de feuilles et l’analyse de celles-ci (I.1.). Cette
méthode fournit la surface foliaire et la surface verte en cm². Elle a pour caractéristiques de
permettre, avec d’autres données, d’obtenir également le pourcentage de surface verte foliaire et le
LAI. Elle est très lente en raison du temps nécessaire pour prendre et analyser une image, mais elle
fournit des valeurs proches des valeurs réelles. La validation de celle-ci a été effectuée au moyen
d’une autre méthode qui semble encore plus fiable afin de s’assurer de la réelle fiabilité de la
méthode d’analyse d’images.
Une seconde méthode consiste à estimer visuellement le pourcentage de surface verte foliaire.
Cette méthode présente l’avantage d’être plus rapide pour l’obtention du pourcentage de surface
verte foliaire. Elle doit cependant être validée avant d’être utilisée pour l’estimation d’un
pourcentage de surface verte pris en compte dans un modèle d’estimation des rendements. La
validité de ces estimations va être abordée en étudiant à quel point elles sont corrélées aux valeurs
obtenues avec la méthode d’estimation par l’analyse d’images (I.2.).
I.1. Acquisition des surfaces par l’analyse d’images et validation de la méthode.
Matériel et méthodes.
Les expérimentations effectuées en 2006 et 2007 ont eu lieu sur des parcelles expérimentales de
culture mises en place au Grand-Duché de Luxembourg, à Everlange (Latitude N49°47’08.91’’ ;
Longitude E 5°58’01.36’’ ; Altitude 274 m), dans le cadre du projet MACRY coordonné par le
CRP-GL (Centre de Recherche Public Gabriel Lippmann) en collaboration avec le Département
des Sciences et Gestion de l’Environnement de l’Université de Liège. La mise en place de ces
essais a été possible grâce au support technique du Lycée Technique Agricole d’Ettelbrück et de la
Chambre d’Agriculture du Grand-Duché de Luxembourg. Ce site est situé sur un plateau, comme
la plupart des cultures de blé au Grand-Duché de Luxembourg. Everlange est donc représentatif de
la région céréalière Redange-sur-Attert (16 790 ha de surface agricole utilisée). Le sol est de
texture argilo-sableuse (El Jarroudi, 2005. ppI.5 – I.11). Une station météorologique horaire de

25
l’ASTA (Administration des Services Techniques de l’Agriculture du Grand-Duché de
Luxembourg) est située à proximité du site d’essai (Station d’Useldange), condition nécessaire à
l’utilisation des données dans le modèle d’avertissement PROCULTURE.
Les images numériques des trois dernières feuilles ont été prises à l’aide d’un appareil photo
numérique Canon 7MG pixels. Dans un premier temps, il a été utilisé un support de fond blanc
(moitié des images de 2006) sur lequel la feuille était maintenue, de façon non destructive, à l’aide
d’une languette en plexiglas transparent le temps de la prise de photo. Une autre partie des mesures
a été effectuée en prenant des images à l’aide d’un support de fond bleu. La feuille y a été
maintenue temporairement à plat sur la planche, de manière non destructive, à l’aide de papier
collant double face. L’appareil photo est maintenu à une distance de 39 cm du support à fond bleu,
sur une latte en aluminium, de façon à prendre des photographies à la perpendiculaire de la feuille
de blé dans son entièreté (Figure 2). Cette méthode s’est avérée plus rapide. De plus le fond bleu
facilite après l’analyse d’image.
Figure 2 : Image du dispositif de prise de photos numériques de feuilles de blé d’hiver.
Seules les trois dernières feuilles de blé ont été suivies (F1, F2, F3). Le choix de suivre uniquement
les trois dernières feuilles a été fait sur base d’un certain nombre d’études telles que celle de
Lupton (1972) et celle de Gooding et al. (2000). En effet, bien que la matière sèche du grain puisse
être accumulée d’assimilas provenant de la photosynthèse des épis, des tiges et des feuilles basses,
aussi bien que des dernières feuilles, Lupton a montré qu’il pouvait être estimé que 45 % des

26
carbohydrates du grain sont dérivés de la photosynthèse de la dernière feuille. D’autres études
montrent par la suite la forte relation existant entre la dégradation de la surface verte de la dernière
feuille (F1) et le rendement (Gooding et al., 2000 ; Dimmock et Gooding, 2002a), bien qu’il ait
également été montré la participation non négligeable des réserves des tiges, accumulées durant la
préfloraison, au remplissage des grains (Bancal et al., 2007). Il est considéré ici que le suivi des
trois dernières feuilles constitue un bon indicateur de l’état des processus photosynthétiques
opérant pour le remplissage des grains. D’autre part, l’intérêt de prendre en compte les trois
dernières feuilles plutôt que seulement la F1 sera testé dans l’utilisation des modèles d’estimation
des rendements.
Les images des 3 dernières feuilles de 5 plantes marquées de repères par répétition sur 4 répétitions
par variante ont été prises de façon hebdomadaire ou une fois toutes les deux semaines à partir de
l’émergence de la dernière feuille, entre le stade GS 37 et GS 39 (stades phénologiques de
ZADOKS et al. , 1974. voir Annexe II). Chaque répétition (parcelle) mesure 1,5m sur 7,5m et est
séparée des parcelles qui l’entourent par un couloir de 1 m de largeur. Les 5 plantes suivies sont
marquées d’un repère et ont été choisies de manière aléatoire au sein de chaque répétition. La
disposition des répétitions est également aléatoire et a été mise en place dans le cadre du projet
MACRY (Tableau 1 , 2 et 3). Afin de déterminer si le nombre de prise d’images servant à
l’obtention de la surface foliaire et de la surface verte par répétition est suffisant, un test de
comparaison des variances par répétition a été effectué. Les résultats du test montrent des
variances par répétition non significativement différentes, ce qui valide le nombre de mesures
utilisées par répétition.

27
Tableau 1 : Dispositif expérimental 2005-2006 installé pour chaque combinaison variété/précédent cultural.
GS31+
GS59
GS45 GS37 GS31 Triple
traitement
(GS31-37-59)
GS59 Témoin GS39
Témoin GS59 GS39 GS45 GS37 GS31 Triple
traitement
(GS31-37-59)
GS31+GS59
GS31 GS37 Triple
traitement
(GS31-37-59)
Témoin GS39 GS31+
GS59
GS45 GS59
GS39 GS31+
GS59
Témoin GS59 GS45 GS37 GS31 Triple
traitement
(GS31-37-59)
( stades phénologiques de ZADOKS et al. , 1974, signification en Annexe II).
Légende des traitements phytosanitaires effectués aux différents stades :
GS31 : 1,6 l/ha Input pro set + 1l/ ha Bravo
GS37 : 1,6 l/ha Input pro set + 1l/ ha Bravo
GS39 : 1,6 l/ha Input pro set + 1l/ ha Bravo
GS45 : 1,6 l/ha Input pro set + 1l/ ha Bravo
GS59 : 1,6 l/ha Input pro set + 1l/ ha Bravo
GS31+59 : 0,75l/ha Opus team + 1l/ha Bravo ; GS59 =1,6 l/ha Input pro set + 1l/ ha Bravo
GS31-37-59 : GS31=0,7l/ha de Stereo +1l/ha Bravo, GS37=1,6 l/ha Input pro set + 1l/ ha Bravo et
GS 59 = : 0,75l/ha Opus team + 1l/ha Bravo

28
Tableau 3: Plan expérimental de Akteur précédent pois à Everlange en 2006-2007. GS 39 GS31 Témoin GS31 +
GS 59
GS 31 +
GS 59
GS 37 GS 59 GS 45
Témoin GS 31 +
GS 37 +
GS 59
GS 39 GS 37
GS 59 Témoin GS 45 GS 31
GS 45 GS 39 GS 37 GS 31 +
GS 37 +
GS59
GS 37 GS 31 +
GS 59
GS 31 GS59
GS 31 GS 45 GS 31 +
GS 37 +
GS 59
Témoin
GS 31 +
GS 37 +
GS 59
GS 59 GS 31 +
GS 59
GS 39
Légende : Témoin : pas de traitement fongicide GS31 (premier nœud) : 1.6 l/ha Input pro set + 1l/ha Bravo GS37 (apparition dernière feuille) : 1.6 l/ha Input pro set + 1l/ha Bravo GS39 (ligule de l’épis visible) :1.6 l/ha Input pro set + 1l/ha Bravo GS45 (gaine de l’épis éclatée) :1.6 l/ha Input pro set + 1l/ha Bravo GS59 (épis dégagé) :1.6 l/ha Input pro set + 1l/ha Bravo GS31+GS59 :GS31: 0.75l/ha Opus team + 1l/ha Bravo et GS59:1.6 l/ha Input pro set + 1l/ha Bravo GS31+GS37+GS59 : GS31: 0.7l/ha Stereo + 1l/ha Bravo, GS37: 1.6 l/ha Input pro set + 1l/ha Bravo et GS59: 0.75l/ha Opus team + 1l/ha Bravo.
Matières actives :
• Input®pro set : prothioconazole 250g/l + spiroxamine 500 g/l • Bravo : chlorothalonil 500g/l • Opus team : époxiconazole 84g/l + fenpropimorphe 250g/l • Stereo : cyprodinil 250g/l + propiconazole 62.5g/l
Tableau 2: Plan expérimental de Flair et Achat précédent pois ainsi que Achat précédent blé deux années à Everlange en 2006-2007. GS 31 +
GS 37 +
GS 59
Témoin GS 37-39 GS 31 +
GS 59
GS 31 +
GS 59
GS 31 +
GS 37 +
GS 59
Témoin GS 37-39
GS 37-39 GS 31 +
GS 59
GS 31 +
GS 37 +
GS 59
Témoin
Témoin GS 37-39 GS 31 +
GS 59
GS 31 +
GS 37 +
GS 59

29
En 2006, les variantes Achat témoin (non traité) et Achat triple traitement (3 traitements) sur
précédent jachère ont été suivies de façon hebdomadaire. Cette variété a été choisie car elle
présente les caractéristiques des variétés utilisées par les agriculteurs au Grand-Duché de
Luxembourg ainsi que pour sa sensibilité moyenne à la septoriose (Anonymous, 2002), principale
maladie affectant le blé dans la région étudiée. Le choix d’un témoin non traité et d’un témoin
traité à trois reprises avait pour objectif d’évaluer l’effet d’une protection complète contre la
septoriose. En 2007, un plus grand nombre de mesures a pu être effectué, grâce à l’amélioration du
dispositif de prise d’images. Les variantes Achat témoin ainsi que Achat triple traitement ont été
suivies de manière hebdomadaire afin d’obtenir 2 années de données pour les variantes utilisées en
2006. Les variantes Flair témoin et Akteur témoin sur précédent jachère ont été ajoutées, celles-ci
étant utilisées au Grand-Duché de Luxembourg et présentant une résistance à la septoriose
différente de celle d’Achat. La variété Flair est résistante à la septoriose et la variété Akteur est
sensible (Anonymous, 2002). La variante Achat témoin avec un semis précoce (10/10/2006) sur
précédent blé deux années consécutives a également été utilisée afin de tester l’effet d’un
précédent blé. Toutes les variantes ajoutées en 2007 ont été suivies une fois toutes les deux
semaines. Cela a permis de suivre plus de variantes, et le suivi moins détaillé obtenu s’est avéré
suffisant pour l’utilisation qui en a été faite à la suite de ce travail. Enfin, la variante achat témoin
semis tardif (26/10/2006) sur précédent blé deux années consécutives a été suivie pour une seule
date correspondant à l’observation du maximum de surface foliaire verte. Cette mesure unique a pu
être utilisée pour calculer le LAI. Le récapitulatif des caractéristiques de prises d’images est
présenté au sein du Tableau 4. Les produits utilisés pour les traitements phytosanitaires ainsi que
leur stade d’application sont consultables dans la légende des Tableaux 2 et 3. L’ensemble du
dispositif au sein duquel se trouvaient les parcelles suivies est présenté aux Tableaux 1, 2 et 3. Les
autres caractéristiques de l’itinéraire cultural qui a été suivi pour 2006 et 2007 sont présentées au
sein du Tableau 5 pour 2006 et du Tableau 6 pour 2007.
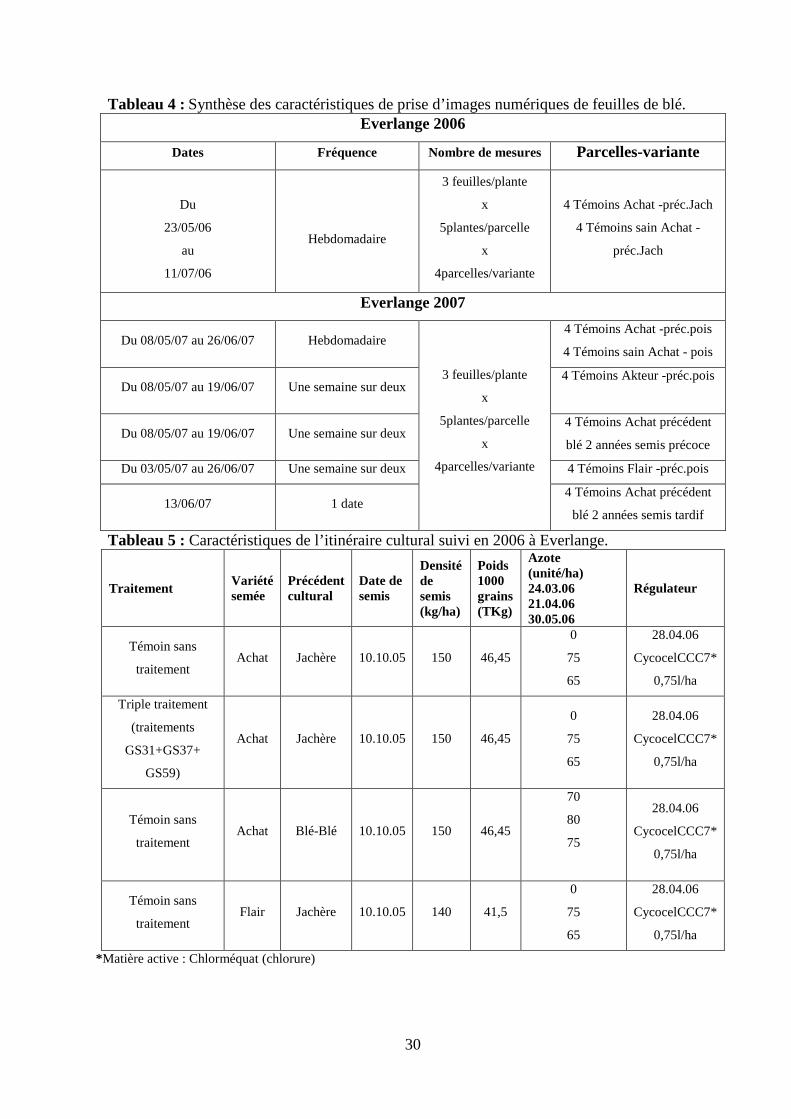
30
Tableau 4 : Synthèse des caractéristiques de prise d’images numériques de feuilles de blé. Everlange 2006
Dates Fréquence Nombre de mesures Parcelles-variante
Du
23/05/06
au
11/07/06
Hebdomadaire
3 feuilles/plante
x
5plantes/parcelle
x
4parcelles/variante
4 Témoins Achat -préc.Jach
4 Témoins sain Achat -
préc.Jach
Everlange 2007
Du 08/05/07 au 26/06/07 Hebdomadaire 4 Témoins Achat -préc.pois
4 Témoins sain Achat - pois
Du 08/05/07 au 19/06/07 Une semaine sur deux 4 Témoins Akteur -préc.pois
Du 08/05/07 au 19/06/07 Une semaine sur deux 4 Témoins Achat précédent
blé 2 années semis précoce
Du 03/05/07 au 26/06/07 Une semaine sur deux 4 Témoins Flair -préc.pois
13/06/07 1 date
3 feuilles/plante
x
5plantes/parcelle
x
4parcelles/variante
4 Témoins Achat précédent
blé 2 années semis tardif
Tableau 5 : Caractéristiques de l’itinéraire cultural suivi en 2006 à Everlange.
Traitement
Variété semée
Précédent cultural
Date de semis
Densité de semis (kg/ha)
Poids 1000 grains (TKg)
Azote (unité/ha) 24.03.06 21.04.06 30.05.06
Régulateur
Témoin sans
traitement Achat Jachère 10.10.05 150 46,45
0
75
65
28.04.06
CycocelCCC7*
0,75l/ha
Triple traitement
(traitements
GS31+GS37+
GS59)
Achat Jachère 10.10.05 150 46,45
0
75
65
28.04.06
CycocelCCC7*
0,75l/ha
Témoin sans
traitement Achat Blé-Blé 10.10.05 150 46,45
70
80
75
28.04.06
CycocelCCC7*
0,75l/ha
Témoin sans
traitement Flair Jachère 10.10.05 140 41,5
0
75
65
28.04.06
CycocelCCC7*
0,75l/ha
*Matière active : Chlorméquat (chlorure)
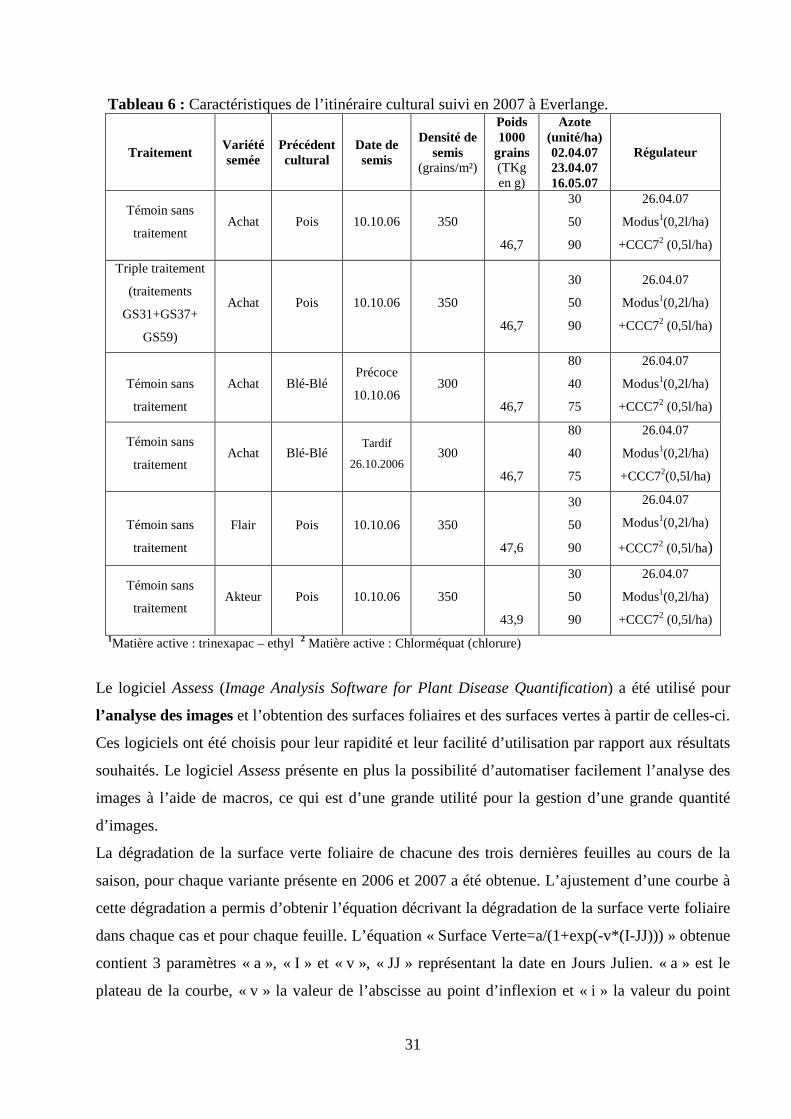
31
Tableau 6 : Caractéristiques de l’itinéraire cultural suivi en 2007 à Everlange.
Traitement
Variété semée
Précédent cultural
Date de semis
Densité de semis
(grains/m²)
Poids 1000
grains (TKg en g)
Azote (unité/ha) 02.04.07 23.04.07 16.05.07
Régulateur
Témoin sans
traitement Achat Pois 10.10.06 350
46,7
30
50
90
26.04.07
Modus1(0,2l/ha)
+CCC72 (0,5l/ha)
Triple traitement
(traitements
GS31+GS37+
GS59)
Achat Pois 10.10.06 350
46,7
30
50
90
26.04.07
Modus1(0,2l/ha)
+CCC72 (0,5l/ha)
Témoin sans
traitement
Achat Blé-Blé Précoce
10.10.06 300
46,7
80
40
75
26.04.07
Modus1(0,2l/ha)
+CCC72 (0,5l/ha)
Témoin sans
traitement Achat Blé-Blé
Tardif
26.10.2006 300
46,7
80
40
75
26.04.07
Modus1(0,2l/ha)
+CCC72(0,5l/ha)
Témoin sans
traitement
Flair Pois 10.10.06 350
47,6
30
50
90
26.04.07
Modus1(0,2l/ha)
+CCC72 (0,5l/ha)
Témoin sans
traitement Akteur Pois 10.10.06 350
43,9
30
50
90
26.04.07
Modus1(0,2l/ha)
+CCC72 (0,5l/ha) 1Matière active : trinexapac – ethyl 2 Matière active : Chlorméquat (chlorure)
Le logiciel Assess (Image Analysis Software for Plant Disease Quantification) a été utilisé pour
l’analyse des images et l’obtention des surfaces foliaires et des surfaces vertes à partir de celles-ci.
Ces logiciels ont été choisis pour leur rapidité et leur facilité d’utilisation par rapport aux résultats
souhaités. Le logiciel Assess présente en plus la possibilité d’automatiser facilement l’analyse des
images à l’aide de macros, ce qui est d’une grande utilité pour la gestion d’une grande quantité
d’images.
La dégradation de la surface verte foliaire de chacune des trois dernières feuilles au cours de la
saison, pour chaque variante présente en 2006 et 2007 a été obtenue. L’ajustement d’une courbe à
cette dégradation a permis d’obtenir l’équation décrivant la dégradation de la surface verte foliaire
dans chaque cas et pour chaque feuille. L’équation « Surface Verte=a/(1+exp(-v*(I-JJ))) » obtenue
contient 3 paramètres « a », « I » et « v », « JJ » représentant la date en Jours Julien. « a » est le
plateau de la courbe, « v » la valeur de l’abscisse au point d’inflexion et « i » la valeur du point

32
d’inflexion. La recherche d’une relation entre les paramètres de ces courbes et le rendement a
ensuite été effectuée.
L’analyse effectuée par Gooding et al. (2000) a également été appliquée. Celle-ci consiste à
adapter une courbe de Gompertz modifiée à la décroissance de la surface verte des trois dernières
strates foliaires à partir du stade de développement complet de la F1 (GS39). Cette courbe a pour
équation,
%SV=100*exp{-exp(-k(t-m))}
où %SV est le pourcentage de la surface des 3 dernières strates foliaires, k est le rythme de
décroissance de la surface verte en jour-1, t est le temps en jours, et m le temps qui s’écoule
jusqu’au point d’inflexion de la courbe en jours également. Dans le cas présent, les degrés jours
ont été utilisés comme variable de temps à la place des jours. Le cumul des degrés jours depuis le
stade GS39 a été utilisé pour la variable t. Les valeurs initiales de k et m qui ont permis d’obtenir
l’ajustement de ce type de courbe sont respectivement -0,02 degrés-1 et 600 degrés jours.
Les valeurs de m obtenues pour chaque répétition de chaque variante suivie en 2006 et 2007 ont
été comparées entre les différents traitements et variétés. La relation linéaire entre m et les
rendements obtenus a également été explorée, celle-ci ayant déjà été mise en évidence avec succès
par Gooding et al. (2000).
La validation de la méthode d’obtention des surfaces foliaires et des surfaces vertes par
l’analyse d’images a été effectuée par la comparaison de surfaces foliaires obtenues à l’aide de
cette méthode à celles obtenues par une méthode consistant à découper et peser le poids de papier
des photocopies des feuilles de blés dont on a fait l’analyse des images. Cette méthode a déjà été
utilisée à plusieurs reprises de manière similaire (ex. : Alem et al., 2002). 60 feuilles vertes de blé
ont été prélevées au hasard. Chacune d’elle a été photographiée avec le dispositif utilisé durant
toute la saison 2007 (dispositif à fond bleu). Les images obtenues ont été analysées au moyen du
logiciel Assess. Les feuilles une fois photographiées ont été photocopiées. Les photocopies de
feuilles ont été découpées et pesées. 10 carrés de 25cm² du même papier ont été pesés.

33
La surface correspondant au poids de papier a ensuite été calculée à partir de la connaissance du
poids de 25cm² comme suit :
Surface foliaire (cm²) = (Poids en papier de la feuille (g) / poids de 25cm² de papier (g) ) * 25 cm² Une régression des moindres carrés a ensuite été effectuée sur les données de surfaces foliaires
issues de l’analyse d’image et de la méthode du poids de papier.
Résultats et discussion.
Une illustration des images de feuilles de blé obtenues et de la classification des pixels de ces
images est présentée en Annexe III . Les seules variantes suivies par la prise et l’analyse d’images
à la fois en 2006 et 2007 sont Achat témoin précédent jachère (ou pois) et Achat triple traitement
précédent jachère (ou pois).
Les conclusions suivantes peuvent être tirées de l’observation de l’évolution dans le temps (1 date
de mesure par semaine ou toute les deux semaines) des moyennes des surfaces vertes des 20 F1,
F2 ou F3 de chaque variante suivie à la fois en 2006 et 2007 (Figure 3) :
- Les trois traitements (Triple traitement) retardent la diminution de la surface verte sur les
trois dernières feuilles, mais la décroissance du triple traitement est plus brutale une fois
qu’elle survient.
Cette observation est en accord avec l’étude de
Gooding (2000), dont l’une des conclusions est que
les fongicides retardent l’arrivée de la décroissance
de la surface verte plutôt qu’ils ne réduisent le
rythme de progression de la décroissance, une fois
celle-ci commencée. Dans ce cas-ci, on peut
observer un gain de rendement associé à un retard
de décroissance de la surface verte (Tableau 7).
- Les courbes de la F1 en 2006 présentent un
plateau situé à plus de 10 cm² au-dessus de celles
de 2007.
Les surfaces vertes de 2006 sont plus importantes dès le début et le restent jusqu’à la fin de la
décroissance. Aucune augmentation de rendement correspondant aux différences de surfaces
foliaires vertes entre 2006 et 2007 n’est observée (Tableau 7). La présence de surfaces vertes
foliaires et de surfaces foliaires des F1 plus importantes en 2006 sans augmentation de
Tableau 7 : Rendements moyens des 4 répétitions pour chaque variante étudiée en 2006 et 2007.
2007 Rdmt net en grain à
15% d’humidité (t/ha) Témoin sain 12,012
Témoin 9,356
Flair Te 9,020
Akteur Te 8,801
Achat bb Te 7,445
2006 Rdmt net en grain à
15% d’humidité (t/ha) Témoin sain 9,339
Témoin 8,356

34
rendement correspondant par rapport à 2007 peut être expliquée, à l’échelle de la parcelle, par
un nombre de brins (tallage) plus important au m² en 2007. Un plus grand nombre de brins au
m² rend le couvert plus dense, ce qui réduit la quantité des ressources disponibles par plante.
Les parcelles présentant un nombre de brins plus important au m² peuvent donc, malgré des
surfaces foliaires réduites, fournir un rendement parcellaire plus important. C’est pourquoi
cette donnée (le nombre de brins par m²) est prise en compte dans le calcul du LAI utilisé pour
les modèles d’estimation des rendements (Chapitre II ). Aucune étude n’a été trouvée,
montrant un effet important de l’année sur la surface foliaire du blé, comme cela est observée
ici. L’étude de Rossi et al. (1997) fait cependant état de surfaces foliaires de blé atteignant 40
cm² pour la F1, alors que les plus grandes surfaces foliaires de F1 observées ici atteignent un
maximum de 32 cm² (Figure 3, SF = SVmax). Cette étude, effectuée en Italie, montre que la
surface foliaire du blé peut variée dans une certaine gamme dont les paramètres (climat,
variété, amendements, traitements phytosanitaires…) restent à être déterminés. Concernant
l’amendement azoté, l’étude de Lovell et al. (1997) fait état de surfaces foliaires plus
importantes sur les parcelles ayant reçu un amendement important.
- La chute plus précoce de la surface verte du témoin en 2006 peut être due à des conditions
très peu favorables pour le maintien de cette surface verte après le stade GS40. La
sécheresse de 2006 pourrait expliquer cette différence. En effet, l’augmentation des
températures hâte l’arrivée de la sénescence (Spiertz, 1977 ; Gooding, 2000). Cette chute
ne peut pas être due aux maladies qui étaient peu présentes au stade GS 40, stade auquel la
chute commence.

35
Tracé de Moyennes - Evolution de la surface verte de la F1 de Achat témoin et Achat témoin sain en 2006 et 2007
stade (Zadoks, 1976)
SV
(cm
²)
30 35 40 45 50 55 60 65 70 75 80 85 90 95 100-5
0
5
10
15
20
25
30
35
SV/Variante: Achat préc. pois 2007 SV/Variante: Achat Témoin Sain 2007 SV/variante Achat témoin 2006
SV/Variante: Achat TeSain2006
Tracé de Moy ennes (F2_SV_Cm²_AchtTe_AchtTeSain_2006ET2007 10v*660c)
stade (Zadoks, 1976)
SV
(cm
²)
30 35 40 45 50 55 60 65 70 75 80 85 90 95 100-5
0
5
10
15
20
25
30
35
SV/Variante: Achat préc. pois2007 SV/Variante: Achat Témoin Sain2007 SV/Variante: Achat Te2006 SV/Variante: Achat TeSain2006
Tracé de Moy ennes (F3_SV_Cm²_AchtTe_AchtTeSain_2006ET2007 9v*651c)
s tade (Zadoks, 1976)
SV
(cm
²)
30 35 40 45 50 55 60 65 70 75 80 85 90 95 100-2
0
2
4
6
8
10
12
14
16
18
20
22
24
26
28
SV/Variante: Achat préc. pois2007
SV/Variante: Achat Témoin Sain2007 SV/Variante: AchatTe2006 SV/Variante: AchatTesain2006
Figure 3 : Cinétique de dégradation de la surface verte foliaire des trois dernières strates de Achat témoin et Achat triple traitement (=Témoin Sain) sur un précédent jachère en 2006 et 2007. A. Cinétique de la F1 B. Cinétique de la F2 C. Cinétique de la F3
L’observation de l’évolution des moyennes des surfaces vertes des trois dernières feuilles des
variantes témoins suivies par la prise et l’analyse d’images uniquement en 2007 (Flair témoin,
Akteur témoin et Achat témoin et Achat témoin précédent blé 2 années) permet de tirer les
conclusions suivantes (Figure 4):
- Les 3 dernières feuilles de Flair témoin suivent la même cinétique de dégradation de la
surface verte que celles de Achat précédent blé-blé témoin de 2007 (pas de distinction
significative (p<0,05) par ANOVA).
- Les 3 dernières feuilles de Akteur témoin suivent une cinétique de dégradation de la
surface verte de plateau plus élevé et de chute plus précoce que Achat témoin et Flair
témoin. Le rendement de Akteur est légèrement inférieur à celui des deux autres variétés
(Tableau 7).
A. B.
C.
36
- Les 3 dernières feuilles de Achat témoin suivent une cinétique de dégradation de la surface
verte de chute plus tardive que celle des autres variantes. Il est notamment constaté une
chute beaucoup plus précoce de la surface verte lorsque cette variété est utilisée sur un
précédent blé deux années. Le rendement de Achat témoin est supérieur au rendement des
autres variantes (Tableau 7).
Tracé de Moyenne (F1_ SV en Cm& - Everlange 2007)
39 48 59 62 69 77 82 90
stade (Zadoks , 1976)
-2
0
2
4
6
8
10
12
14
16
18
20
22
SV
(c
m²)
F1 Achat Te2007F1 Achat bb Te 2007F1 Flair Te 2007F1 Akteur Te 2007
Tracé de Moyennes (F2_SV_Cm²_Everlange 2007)
39 48 59 62 69 77 82 90
stade (Zadoks , 1976)
-5
0
5
10
15
20
25
30
35
SV
(cm
²)
F2 Achat Te 2007 F2 Achat bb Te 2007 F2 Flair Te 2007 F2 Akteur Te 2007
Tracé de Moyennes (F3_SV_Cm²_ Everlange 2007)
39 40 48 55 59 62 69 77 82 90
Stad e (Zadoks , 1976 )
-5
0
5
10
15
20
25
30
SV
(c
m²)
F3 Achat Te 2007 F3 Achat bb Te 2007F3 Flair Te 2007F3 Akteur Te 2007
Figure 4 : Cinétique de dégradation de la surface foliaire verte des trois dernières strates des variétés non traitées présentes en 2007. A. Cinétique de la F1 B. Cinétique de la F2 C. Cinétique de la F3. L’adaptation de la courbe d’équation « Surface Verte=a/(1+exp(-v*(I-JJ))) » à la dégradation de la
surface verte a fourni des valeurs des paramètres « a », « v » et « i » différentes pour chaque
parcelle de chaque variante.
La linéarité de la relation entre le rendement et un des paramètres « a », « i » et « v » a été explorée
par l’établissement d’une régression linéaire multiple. Les résultats montrent qu’aucun de ces
paramètres n’explique la variabilité des rendements observés (Tableau 8).
Le coefficient Bêta du Tableau 8, qui est un coefficient de régression obtenu si on effectue
d'abord un centrage-réduction de toutes les variables afin d'obtenir pour chacune une moyenne
A. B.
C.
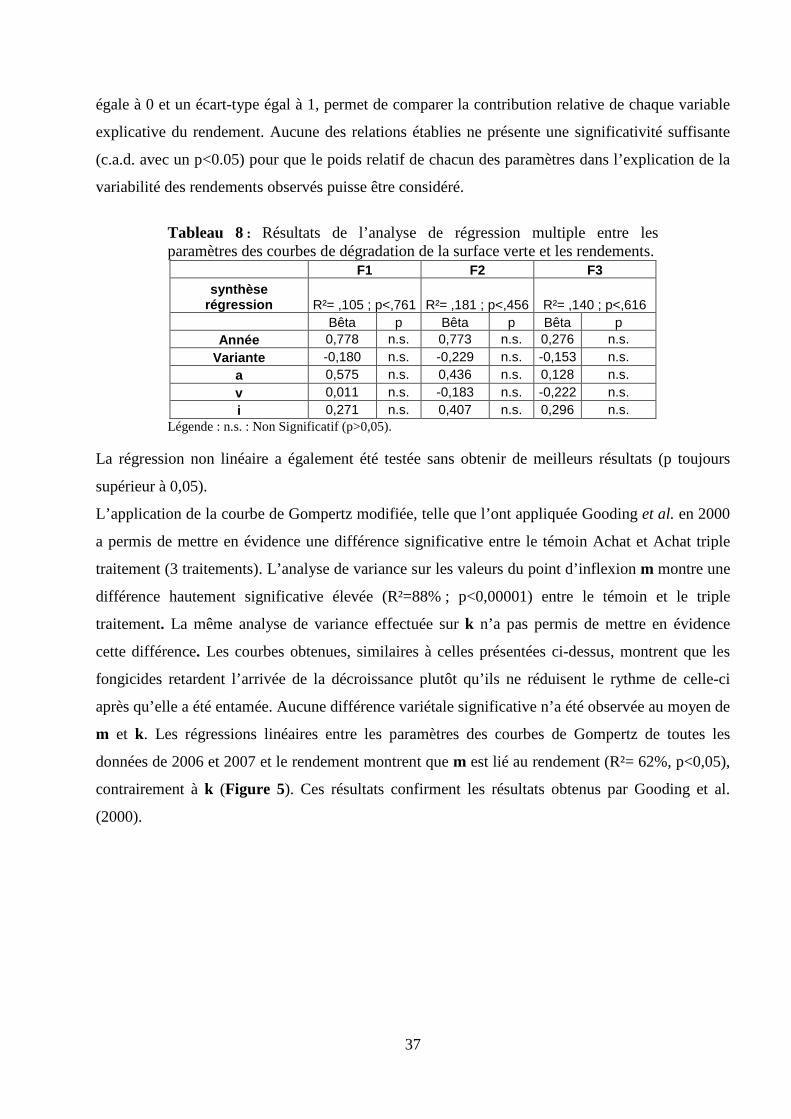
37
égale à 0 et un écart-type égal à 1, permet de comparer la contribution relative de chaque variable
explicative du rendement. Aucune des relations établies ne présente une significativité suffisante
(c.a.d. avec un p<0.05) pour que le poids relatif de chacun des paramètres dans l’explication de la
variabilité des rendements observés puisse être considéré.
Tableau 8 : Résultats de l’analyse de régression multiple entre les paramètres des courbes de dégradation de la surface verte et les rendements.
F1 F2 F3 synthèse
régression R²= ,105 ; p<,761 R²= ,181 ; p<,456 R²= ,140 ; p<,616 Bêta p Bêta p Bêta p
Année 0,778 n.s. 0,773 n.s. 0,276 n.s. Variante -0,180 n.s. -0,229 n.s. -0,153 n.s.
a 0,575 n.s. 0,436 n.s. 0,128 n.s. v 0,011 n.s. -0,183 n.s. -0,222 n.s. i 0,271 n.s. 0,407 n.s. 0,296 n.s.
Légende : n.s. : Non Significatif (p>0,05).
La régression non linéaire a également été testée sans obtenir de meilleurs résultats (p toujours
supérieur à 0,05).
L’application de la courbe de Gompertz modifiée, telle que l’ont appliquée Gooding et al. en 2000
a permis de mettre en évidence une différence significative entre le témoin Achat et Achat triple
traitement (3 traitements). L’analyse de variance sur les valeurs du point d’inflexion m montre une
différence hautement significative élevée (R²=88% ; p<0,00001) entre le témoin et le triple
traitement. La même analyse de variance effectuée sur k n’a pas permis de mettre en évidence
cette différence. Les courbes obtenues, similaires à celles présentées ci-dessus, montrent que les
fongicides retardent l’arrivée de la décroissance plutôt qu’ils ne réduisent le rythme de celle-ci
après qu’elle a été entamée. Aucune différence variétale significative n’a été observée au moyen de
m et k. Les régressions linéaires entre les paramètres des courbes de Gompertz de toutes les
données de 2006 et 2007 et le rendement montrent que m est lié au rendement (R²= 62%, p<0,05),
contrairement à k (Figure 5). Ces résultats confirment les résultats obtenus par Gooding et al.
(2000).
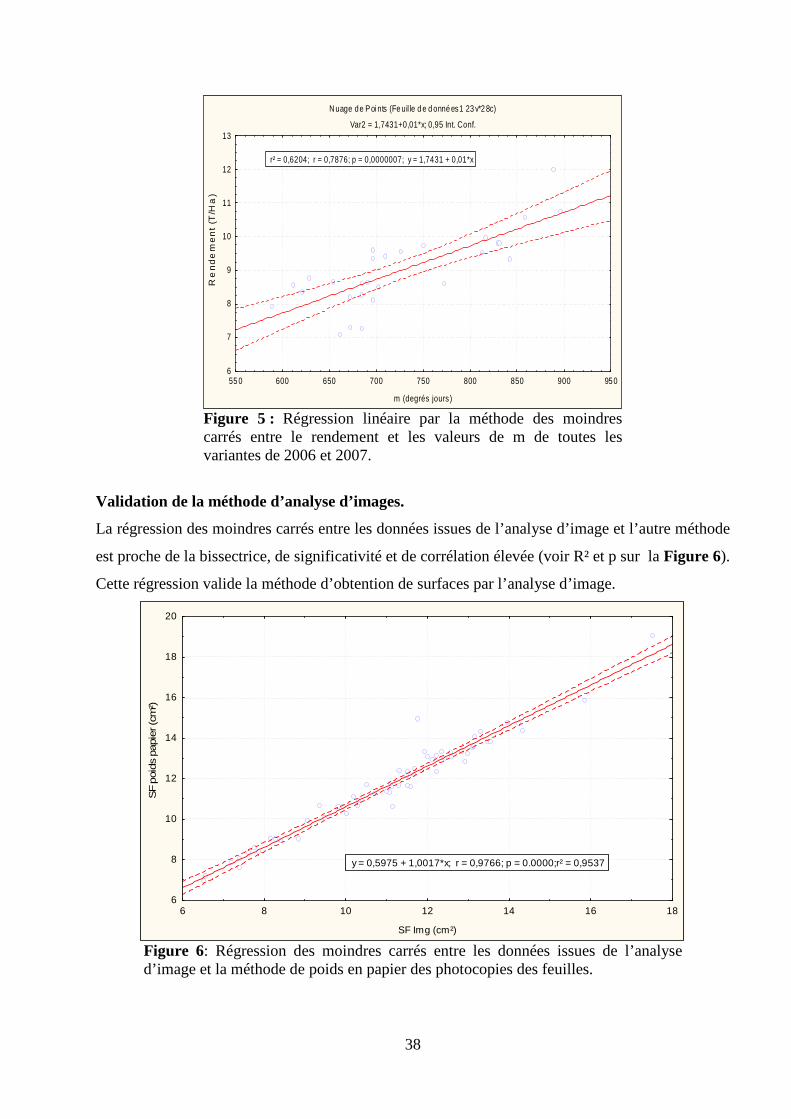
38
N uage de Points (Fe uil le d e d onné es1 23 v*2 8c)
Var2 = 1,7431+0,01*x; 0,95 Int. Conf.
55 0 600 650 700 750 800 850 900 950
m (degrés jours )
6
7
8
9
10
11
12
13
Re
nd
em
en
t (T
/Ha
)
r² = 0,6204; r = 0,7876; p = 0,0000007; y = 1,7431 + 0,01*x
Figure 5 : Régression linéaire par la méthode des moindres carrés entre le rendement et les valeurs de m de toutes les variantes de 2006 et 2007.
Validation de la méthode d’analyse d’images.
La régression des moindres carrés entre les données issues de l’analyse d’image et l’autre méthode
est proche de la bissectrice, de significativité et de corrélation élevée (voir R² et p sur la Figure 6).
Cette régression valide la méthode d’obtention de surfaces par l’analyse d’image.
6 8 10 12 14 16 18
SF Img (cm²)
6
8
10
12
14
16
18
20
SF p
oid
s papie
r (c
m²)
y = 0,5975 + 1,0017*x; r = 0,9766; p = 0.0000;r² = 0,9537
Figure 6: Régression des moindres carrés entre les données issues de l’analyse d’image et la méthode de poids en papier des photocopies des feuilles.

39
I.2. Acquisition du pourcentage de surface verte par la méthode d’estimation visuelle et corrélations avec les données issues de la méthode d’estimation par l’analyse d’images.
Introduction.
Beaucoup de systèmes d’avertissement visant à déterminer le rendement ou le moment optimum
pour l’application d’un traitement phytosanitaire utilisent la proportion de surface foliaire saine, ou
de la surface infectée par une maladie (de Jesus et al., 2001 ; Bassanezi et al., 2001 ; Lopes et
Berger, 2001 ; Bergamin Filho et al., 1997 ; Bryson et al., 1997). La méthode d’estimation visuelle
de la proportion de tissus sains, nécrosés ou malades, est utilisée dans la plupart des
expérimentations car elle est facile d’utilisation, rapide, économique et applicable pour un grand
nombre de conditions. Elle remplirait les critères idéaux d’une bonne méthode d’estimation de la
sévérité d’une maladie énoncés par Nutter et al. (1993a) si elle fournissait des résultats exacts
(moyennes proches de la valeur de référence) et précis (dont la répétition de la mesure mène à des
valeurs proches les unes des autres) pour toutes les études dans lesquelles elle est utilisée.
Ceci n’est pas toujours le cas, comme le montrent plusieurs études qui ont été entreprises afin de
déterminer la validité de la méthode d’estimation visuelle de symptômes de maladies (Parker et al.,
1995 ; Martin et Rybicki, 1998 ; Forrest et al., 1995 ; Moya et al., 2005 ; Price et al., 1993). Il a
souvent été mis en évidence l’existence de biais entre les estimations effectuées par différents
observateurs (Sherwood et al., 1983 ; Moya et al., 2005, Forrest et al., 1995 ; Martin et Rybicki,
1998 ; Parker et al., 1995) et par un même observateur entre deux séances d’estimation sur les
mêmes échantillons (Nutter et al., 1993a). Des logiciels, tels que DISTRAIN (Tomerlin et Howell,
1988) et Disease.Pro (Nutter et Worawitlikit, 1989), d’entraînement des observateurs à la
reconnaissance des surfaces couvertes par les symptômes de certaines maladies ont été mis au
point afin d’améliorer la précision des estimations. En conditions de simulation, des améliorations
ont pu être obtenues dans la capacité des observateurs à estimer les proportions couvertes par des
symptômes grâce à ces logiciels (Forrest et al., 1995 ; Nutter et Worawitlikit, 1989). Les
estimations effectuées par un observateur sur son ordinateur sont cependant loin de celles qu’il
devra effectuer sur des plantes chaque semaine, parce que l’ordinateur ne permet pas de considérer
les dimensions de la feuille de la même manière que sur le terrain. Afin de déterminer la validité
des mesures d’estimations visuelles, il est nécessaire de les comparer et de les valider avec des
mesures reflétant mieux la réalité.

40
L’utilisation de l’ordinateur pour le traitement et l’analyse d’images a été développée en réponse à
cette nécessité (Martin et Rybicki, 1998 ; Niemira et al., 1999 ; Olmstead et al., 2001). Martin et
Rybicki (1998) ont montré que la précision et l’exactitude des mesures obtenues par l’analyse
d’images de symptômes de maladies de plantes étaient significativement plus élevées que celles
obtenues par l’estimation visuelle. Les études menées utilisant l’analyse d’images considèrent ces
mesures comme « véritables » (Sherwood et al., 1983 ; Nutter et al., 1993a ; Parker et al., 1995 ;
Martin et Rybicki, 1998 ; Moya et al., 2005 ; Price et al., 1993). Les images analysées sont issues
dans certains cas de l’image projetée de feuilles et des symptômes de maladies qu’elles
présentaient (Sherwood et al., 1983 ; Nutter et al., 1993a ; Parker et al., 1995). Dans d’autres cas
l’analyse d’images est obtenue par la prise de photos en noir et blanc, en couleur, en numérique,
sous éclairage particulier ou sans éclairage (Martin et Rybicki, 1998 ; Moya et al., 2005 ; Price et
al., 1993).
Les conclusions des études comparant les estimations utilisant l’analyse d’images à la méthode
d’estimation visuelle sont variables. Elles sont fonction de la gamme de pourcentage de surface
couverte par les symptômes estimés, du complexe hôte parasite considéré (Horsf all et Barratt,
1945 ; Lindow et webb, 1983 ; Sherwood et al., 1983 ; Parker et al., 1995), ainsi que de la forme
et du nombre des lésions étudiées (Lindow et webb, 1983 ; Webb et Lindow, 1981). Dans leur
étude, Horsfall et Barratt (1945) suggèrent que l’estimation visuelle est imprécise quand le
pourcentage de surface foliaire affecté est proche de 50 %. Nutter et al. (1995) ont obtenu des
erreurs d’estimation apparaissant systématiquement dans la gamme de 40 % à 60 %. Parker et al.
(1995) constatent une surestimation du pourcentage de surface couverte par les symptômes dans la
gamme 0-10 % et une sous-estimation au-delà. Enfin, Forbes et Jeger (1987) ont rapporté que la
plupart des erreurs apparaissent à 25 % de sévérité pour plusieurs maladies.
L’ensemble de ces études montre que les différents biais introduits sont fonction des observateurs,
de la forme des feuilles et des symptômes étudiés, ou encore de la surface verte étudiée. La
méthode d’estimation visuelle est donc une méthode qui devrait être validée pour chaque équipe la
pratiquant, et en fonction des caractéristiques de la surface verte ou des symptômes qui sont
étudiés.
Dans cette partie, l’estimation visuelle du pourcentage de surface verte des trois dernières feuilles
de parcelles expérimentales de blé d’hiver aux champs a été comparée à l’estimation du
pourcentage de surface verte obtenue à partir de l’analyse d’images numériques.

41
Matériel et méthodes.
La méthode d’estimation visuelle du pourcentage de surface verte (Tomerlin et Howell, 1988) a
été utilisée sur les mêmes plantes, les mêmes feuilles et aux mêmes dates que celles utilisées pour
les estimations par l’analyse d’images. Les estimations visuelles ont été effectuées par l’équipe de
biométéorologie du Département des Sciences et Gestion de l’Environnement de l’Université de
Liège et avec la collaboration du CRP-GL dans le cadre du projet MACRY. Les observateurs
étaient au nombre de 4 à 6 suivant les dates d’observation. Dans cette étude, tous les observateurs
sont considérés comme ayant la même aptitude à estimer le pourcentage de surface verte.
L’objectif étant de déterminer les erreurs typiques effectuées par un observateur. Les estimations
par l’analyse d’images ont été effectuées telles que décrites dans le matériel et méthodes du
chapitre I.1.
Une régression linéaire par la méthode des moindres carrés a été appliquée aux données, afin
d’estimer la relation entre les données issues de l’estimation visuelle et les données issues de
l’estimation par l’analyse d’images. Une représentation graphique mettant en relation le logarithme
népérien de « % surface verte visuelle / % surface verte images » au logarithme népérien de « % de
surface verte images » a été effectuée afin de mettre en évidence les zones de pourcentage pour
lesquelles une surestimation ou une sous-estimation du pourcentage de surface verte est observée
(Parker et al., 1995).
Résultats et discussion
L’analyse de régression des moindres carrés entre les estimations du pourcentage de surface verte
feuille par feuille de façon visuelle et par l’analyse d’images donne de bons résultats. En effet, la
relation est linéaire et hautement significative (p=0.0000), de R² proche de un et de petit intercept
(-4,267) (Figure 7). Cette observation n’est cependant pas valable car la régression est influencée
par une distribution non Normale et bimodale des données (Figure 8). Il n’est donc pas possible de
conclure en l’existence d’une relation linéaire entre les deux types d’estimation. Les méthodes
classiques visant à rétablir la normalité en appliquant l’arc sinus ou l’arc tangente n’ont pas
fonctionné. Une méthode utilisée par Parker et al. (1995) pour comparer deux manières d’estimer
des symptômes de maladies foliaires a permis de tirer quelques conclusions.
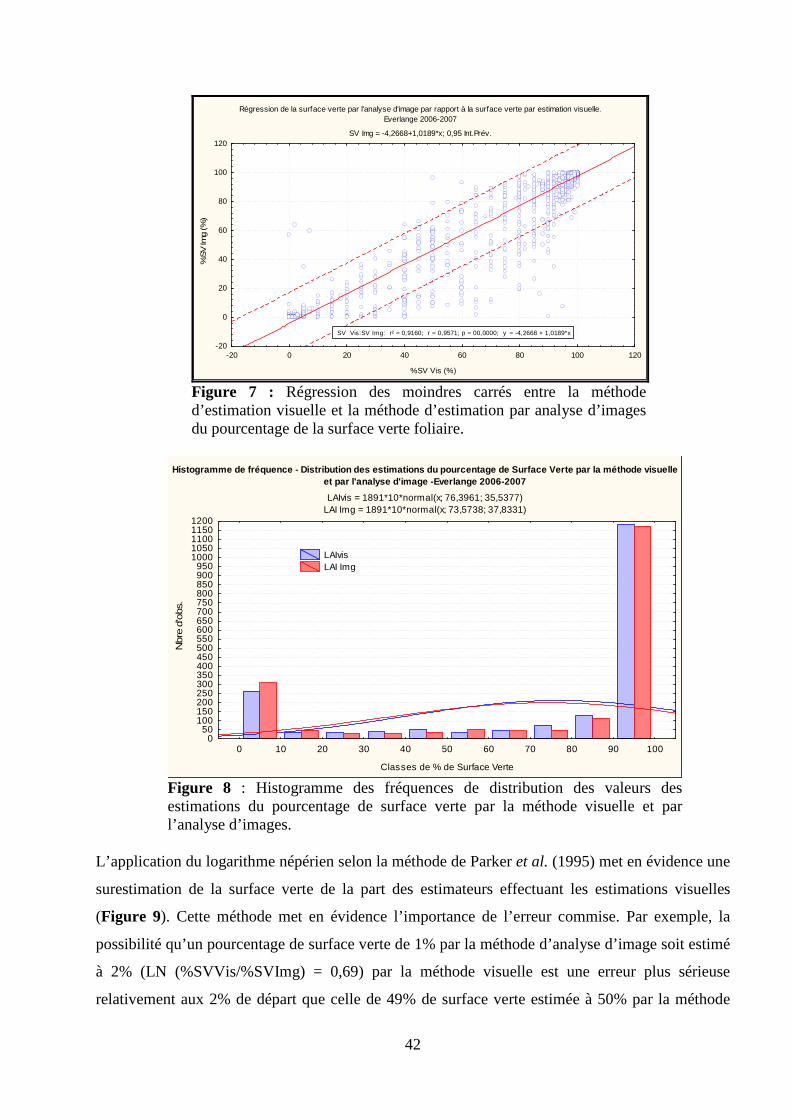
42
Régression de la surface verte par l'analyse d'image par rapport à la surface verte par estimation visuelle.Everlange 2006-2007
SV Img = -4,2668+1,0189*x; 0,95 Int.Prév.
-20 0 20 40 60 80 100 120
%SV Vis (%)
-20
0
20
40
60
80
100
120
%SV Im
g (%
)
SV Vis:SV Img: r² = 0,9160; r = 0,9571; p = 00,0000; y = -4,2668 + 1,0189*x
Figure 7 : Régression des moindres carrés entre la méthode d’estimation visuelle et la méthode d’estimation par analyse d’images du pourcentage de la surface verte foliaire.
Histogramme de fréquence - Distribution des estimat ions du pourcentage de Surface Verte par la méthode visuelle
et par l'analyse d'image -Everlange 2006-2007
LAIvis = 1891*10*normal(x; 76,3961; 35,5377)LAI Img = 1891*10*normal(x; 73,5738; 37,8331)
0 10 20 30 40 50 60 70 80 90 100
Classes de % de Surface Verte
050
100150200250300350400450500550600650700750800850900950
10001050110011501200
Nbre
d'o
bs.
LAIvis LAI Img
Figure 8 : Histogramme des fréquences de distribution des valeurs des estimations du pourcentage de surface verte par la méthode visuelle et par l’analyse d’images.
L’application du logarithme népérien selon la méthode de Parker et al. (1995) met en évidence une
surestimation de la surface verte de la part des estimateurs effectuant les estimations visuelles
(Figure 9). Cette méthode met en évidence l’importance de l’erreur commise. Par exemple, la
possibilité qu’un pourcentage de surface verte de 1% par la méthode d’analyse d’image soit estimé
à 2% (LN (%SVVis/%SVImg) = 0,69) par la méthode visuelle est une erreur plus sérieuse
relativement aux 2% de départ que celle de 49% de surface verte estimée à 50% par la méthode
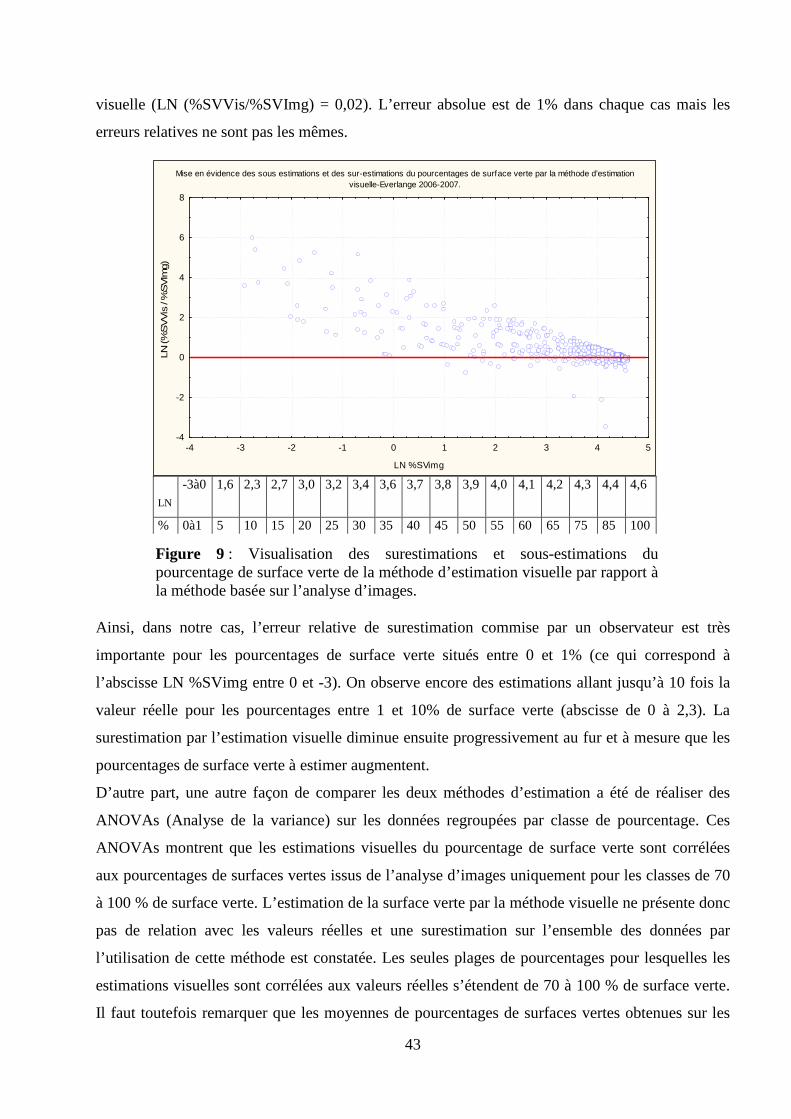
43
visuelle (LN (%SVVis/%SVImg) = 0,02). L’erreur absolue est de 1% dans chaque cas mais les
erreurs relatives ne sont pas les mêmes.
Figure 9 : Visualisation des surestimations et sous-estimations du pourcentage de surface verte de la méthode d’estimation visuelle par rapport à la méthode basée sur l’analyse d’images.
Ainsi, dans notre cas, l’erreur relative de surestimation commise par un observateur est très
importante pour les pourcentages de surface verte situés entre 0 et 1% (ce qui correspond à
l’abscisse LN %SVimg entre 0 et -3). On observe encore des estimations allant jusqu’à 10 fois la
valeur réelle pour les pourcentages entre 1 et 10% de surface verte (abscisse de 0 à 2,3). La
surestimation par l’estimation visuelle diminue ensuite progressivement au fur et à mesure que les
pourcentages de surface verte à estimer augmentent.
D’autre part, une autre façon de comparer les deux méthodes d’estimation a été de réaliser des
ANOVAs (Analyse de la variance) sur les données regroupées par classe de pourcentage. Ces
ANOVAs montrent que les estimations visuelles du pourcentage de surface verte sont corrélées
aux pourcentages de surfaces vertes issus de l’analyse d’images uniquement pour les classes de 70
à 100 % de surface verte. L’estimation de la surface verte par la méthode visuelle ne présente donc
pas de relation avec les valeurs réelles et une surestimation sur l’ensemble des données par
l’utilisation de cette méthode est constatée. Les seules plages de pourcentages pour lesquelles les
estimations visuelles sont corrélées aux valeurs réelles s’étendent de 70 à 100 % de surface verte.
Il faut toutefois remarquer que les moyennes de pourcentages de surfaces vertes obtenues sur les
Mise en évidence des sous estimations et des sur-estimations du pourcentages de surface verte par la méthode d'estimationvisuelle-Everlange 2006-2007.
-4 -3 -2 -1 0 1 2 3 4 5
LN %SVimg
-4
-2
0
2
4
6
8
LN (%
SVVis
/ %
SVIm
g)
LN
-3à0 1,6 2,3 2,7 3,0 3,2 3,4 3,6 3,7 3,8 3,9 4,0 4,1 4,2 4,3 4,4 4,6
% 0à1 5 10 15 20 25 30 35 40 45 50 55 60 65 75 85 100

44
données regroupées par classe de pourcentage de 10 unités ne se différencient pas plus que d’une
marge de 1% maximum entre les deux méthodes. Il faut donc relativiser l’importance de ces
surestimations en fonction de l’utilisation qui est faite des données. L’utilisation de valeurs
moyennes parcellaires issues d’estimations visuelles de 10 plantes par parcelle pourrait fournir des
valeurs presque identiques aux moyennes parcellaires issues d’estimation par l’analyse d’images.
Dans ce travail, les données de %SV sont utilisées dans des modèles d’estimation des rendements.
Les résultats obtenus en utilisant un modèle d’estimation des rendements avec les estimations
visuelles du %SV ont été comparés aux résultats obtenus en utilisant les estimations issues de
l’analyse d’images (PARTIE II, Chapitre II.3. ). La comparaison met en évidence une
surestimation moyenne de 10% des paramètres du modèle lorsqu’on utilise les %SV issus de
l’estimation visuelle. Cette surestimation est probablement une conséquence de la tendance à la
surestimation du %SV par l’utilisation de la méthode d’estimation visuelle. D’autre part,
l’utilisation des estimations du %SV issues de l’analyse d’images permet d’obtenir des modèles
qui expliquent une plus grande proportion de la variabilité des rendements (20% en plus dans 3 cas
sur 4). Ces différences sont non négligeables, alors que les analyses de comparaison des deux
méthodes d’estimation du %SV par classe montrent de faibles différences d’estimation entre les
deux méthodes. Ceci est dû à une utilisation des données par strate foliaire dans le modèle
d’estimation des rendements, alors que la comparaison des deux méthodes d’estimation du %SV
montrant une différence de 1% a été effectuée par classe de %SV.

45
CHAPITRE II : Obtention du LAI. Le vrai LAI ne peut être obtenu que par une méthode permettant d’obtenir directement les surfaces
foliaires (Breda, 2003).
Dans ce Chapitre, le LAI sera calculé en utilisant directement des surfaces foliaires issues de
l’analyse d’images de feuilles. Méthode elle-même validée par une autre méthode d’estimation des
surfaces foliaires dans un chapitre précédent (I.1). Ensuite sera déterminé et validé un modèle
mettant en relation le LAI de référence obtenu à l’aide des surfaces foliaires au PAI (Plant Area
Index) obtenu par une méthode d’obtention à distance, plus rapide et automatisable (II.1. ). La
validation d’un modèle d’obtention du LAI à partir de valeurs de PAI obtenues de façon rapide et
automatisable est d’un grand intérêt pour une utilisation routinière des modèles d’estimation des
rendements et les fonctions de dommages.
Le passage à l’obtention d’un LAI à grande échelle au moyen d’images aériennes a également été
testé par comparaison du LAI obtenu à partir de surfaces foliaires à celui obtenu à l’aide d’images
aériennes prises à quelques mètres du sol dans la dernière partie de ce chapitre (II.2. ).
II.1. Obtention et validation d’un modèle de détermination du LAI à partir de valeurs de PAI obtenues à l’aide d’une méthode indirecte.
Introduction.
Le LAI est défini comme étant la surface foliaire (une face) de toutes les feuilles du couvert
contenues dans un volume situé au-dessus de un mètre carré de surface de sol horizontal (Watson,
1947). Les premières méthodes d’estimation au sol du LAI étaient destructrices et consommatrices
de temps. Les chercheurs coupaient les plantes comprises dans une surface unitaire au sol,
séparaient les feuilles des autres parties de la plante et mesuraient la surface des feuilles
individuelles pour obtenir la surface moyenne par plante. Le produit de la surface foliaire par
plante et le nombre de plantes donnaient le LAI (Wilhelm et al., 2000).
Par la suite, cette méthode a été appliquée de manière non destructrice en déterminant la surface
foliaire à partir de la mesure de la longueur et de la largeur des feuilles (Hopkins, 1939 ; Lal et
Subba Rao, 1950 ; van Arkel, 1978). Une autre méthode directe de mesure consiste à utiliser un
planimètre, instrument qui permet de connaître la surface foliaire à partir de l’obtention du contour
de la feuille. Les méthodes d’estimation directes du LAI fournissent une référence pour la

46
calibration et l’évaluation des méthodes indirectes qui, elles, ne passent pas par une estimation
directe des surfaces foliaires.
Un grand nombre de méthodes indirectes d’obtention du LAI ont été développées. Celles-ci
passent pour la plupart par l’étude de la réflectance du couvert ou par des images de celui-ci.
D’autres se basent sur l’étude du rayonnement transmis et intercepté par le couvert. Campbell et
Norman (1989), ainsi que Welles et Cohen (1996) ont résumé les théories et les équations
existantes pour estimer le LAI à partir de la structure du couvert et de la fraction du rayonnement
direct et/ou indirect intercepté par la culture. Ces relations ont mené au développement de
différents appareils commerciaux (Sunscan, Li-cor, etc…) d’estimation du LAI à partir de la
mesure du rayonnement direct, diffus et transmis au travers du couvert. Beaucoup d’articles
comparent les méthodes d’estimation à distance du LAI de cultures aux méthodes d’estimation du
LAI effectuées directement par prélèvement (par ex. Brenner et al., 1995 ; Levy et Jarvis, 1999).
Bien qu’il ait été publié certains cas de figures pour lesquels la corrélation est élevée entre la
méthode d’estimation directe et une méthode d’estimation à distance du LAI (de Jesus et al.,
2001), la plupart des validations croisées entre ces deux types de méthodes ont mis le doigt sur une
sous-estimation significative du LAI avec les méthodes d’estimation indirectes (Breda, 2003). Plus
particulièrement, il a été montré dans une étude de Wilhelm et al. (2000) comparant trois méthodes
d’estimation du LAI à distance à une méthode d’estimation par prélèvement d’échantillons sur le
maïs que les méthodes d’estimation à distance (AccuPAR, LAI-2000 et Sunscan) sous-estiment
toutes le LAI par rapport à la méthode destructrice.
D’autre part, les instruments qui estiment le LAI à partir de mesures du rayonnement transmis au
travers du couvert estiment en réalité un PAI (Plant Area Index) car ils mesurent aussi
l’interception du rayonnement des tiges, branches et autres éléments non foliaires (White et al.,
2000). Beaucoup d’auteurs ont établi des corrections empiriques pour calculer le LAI à partir des
valeurs de PAI obtenues avec ce genre d’instrument (Chen, 1996 ; Deblonde et al., 1994 ;
Fassnacht et al., 1994 ; Gower et Norman, 1991).
Les conclusions tirées des précédentes études montrent donc que les valeurs de LAI obtenues à
l’aide de tels instruments ne sont pas utilisables telles que l’instrument les fournit. Si une relation
existe toutefois entre les valeurs fournies par ce genre d’instruments et les vraies valeurs de LAI,
celle-ci doit probablement être estimée pour chaque culture à cause des différences de structure du
couvert ainsi que de la participation à l’interception du rayonnement des tiges et des éventuels
organes reproducteurs.
L’objectif de la présente étude est de comparer dans un premier temps l’estimation du PAI de la
partie supérieure d’un couvert de blé d’hiver obtenu par une méthode indirecte utilisant un

47
instrument de mesure du rayonnement transmis par le couvert (Sunscan) disposé sous les trois
dernières feuilles au LAI obtenu par une méthode de référence basée sur l’analyse d’images des
trois dernières feuilles. La méthode de référence est validée par une méthode destructrice de
mesure du LAI. La comparaison mènera à l’obtention d’une régression linéaire par la méthode des
moindres carrés entre la méthode de référence et la méthode d’estimation à distance.
Dans un deuxième temps, la validité d’un modèle permettant d’obtenir le LAI à partir du PAI
fourni par le Sunscan sera estimée en effectuant la validation croisée de la régression des moindres
carrés obtenue entre les deux méthodes auparavant.
Les différences observées entre les LAIs des variantes (années, variétés, précédent) utilisées seront
également discutées.
Matériel et méthodes.
Les expérimentations ont été menées durant deux années sur le site expérimental d’Everlange au
Grand-Duché de Luxembourg. Les valeurs de LAI ont été obtenues à raison de un LAI par parcelle
rectangulaire de +/- 1,5 m sur 7,5 m pour les mêmes parcelles avec les deux méthodes. 4 parcelles
de la variété Achat non traitée et 4 de la même variété traitée trois fois ont été utilisées en 2006. En
2007, la variété Flair non traitée, Achat témoin semis précoce non traité précédent blé deux années
et Achat témoin semis tardif non traité précédent blé deux années en plus des deux variantes de
2006 ont été ajoutées. Les parcelles ont été disposées de façon aléatoire, selon le dispositif
aléatoire présenté au Chapitre I . Les LAI utilisés pour la comparaison sont obtenus à partir de
mesures effectuées à la même date pour les deux méthodes. La date choisie est celle du LAI
maximum, juste après l’apparition de la dernière feuille. 28 valeurs de LAI ont été obtenues avec
chaque méthode.
Le LAI obtenu à l’aide des surfaces foliaires issues de l’analyse d’images a été calculé comme
suit :
∑=
=n
i
NEbSFiLAI1
10000/)/(
Où SFi est la surface foliaire de la Fi, pour i allant de la Feuille1 (strate 1) à la Feuille 3 (strate 3). La surface
foliaire de chaque strate a été obtenue à partir de la moyenne des surfaces foliaires des 5 plantes par parcelle
pour chacune de ces strates.
NE est le nombre d’épis par m²

48
b est un coefficient de conversion qui permet de passer du maître brin au brin moyen, dont les feuilles sont
plus petites.
Le paramètre b a été obtenu pour chaque répétition de chaque variante au moyen de la formule
suivante :
Ng
NMBNBb
))((=
Où NB est le nombre de brins par m² obtenu par comptage manuel.
NMB est le nombre de grains par maître brin obtenu à partir des 20 brins par variante suivis par la prise
d’images.
Ng est le nombre de grains par m² obtenu à partir de la surface parcellaire, le poids de 1000 grains et le poids
des grains récoltés sur chaque parcelle (répétition)
Rem. : Le paramètre b n’a été calculé que pour l’année 2007 car les données nécessaires à son
calcul n’ont pas été collectées en 2006. L’hypothèse est ici émise que le paramètre b ne
varie pas beaucoup en appliquant le paramètre b de Achat témoin et Achat triple traitement
de 2007 aux variantes Achat témoin et Achat triple traitement de 2006. D’autres études
menées sur la variété Soisson ont montré qu’il varie dans un intervalle de 1,2 à 1,5 (M. O.
Bancal, Communication personnelle).
Le LAI obtenu par la méthode indirecte, à l’aide du Sunscan, a été effectué sur les mêmes parcelles
que celles utilisées pour la méthode précédente. Le Sunscan mesure le rayonnement incident
direct, diffus et transmis, dans la gamme du RPA (Rayonnement Photosynthétiquement Actif),
sous la canopée. En plus des différentes mesures de rayonnement, il fournit le LAI qu’il calcule sur
base de la loi de Beer Lambert inversée et adaptée aux cultures.
Le Sunscan est composé de trois éléments (Figure 10) :
� Le premier élément est la sonde avec laquelle le RPA est mesuré sous les trois dernières
feuilles de la culture. Cette sonde est une tige d’un mètre de long contenant soixante-quatre
photodiodes. Une mesure de RPA obtenue est le résultat d’une moyenne des mesures prises par les
soixante-quatre photodiodes. L’extrémité par laquelle est manipulée la sonde est terminée par un
boîtier avec une poignée et un bouton permettant de déclencher la prise de mesures. Ce boîtier est
relié aux deux autres éléments par deux câbles.

49
� Le deuxième élément, appelé BFS (Beam Fraction Sensor) permet de mesurer le RPA reçu
par un sol nu au moment où la mesure du RPA est prise sous la culture. Il est composé de sept
photodiodes disposées en cercle sur un support monté sur trépied. Certaines d’entre elles sont
ombragées par un anneau métallique, quelle que soit l’orientation du BFS. De cette manière le
RPA incident direct et diffus peut être mesuré au même instant que celui de la prise de mesure
sous la culture.
� Le troisième élément permet d’effectuer des réglages paramétriques, de visionner et
d’enregistrer les données issues des mesures obtenues à l’aide des deux premiers éléments. Il se
présente sous la forme d’une grosse calculatrice qui peut également être connectée à un ordinateur
pour le transfert et le traitement des données.
Figure 10 : Les trois éléments du SunScan, appareil de mesure du RPA et de détermination du LAI.
L’instrument a été utilisé à raison de quatre mesures par parcelle sur quatre parcelles par variante.
L’instrument est disposé au sein de la parcelle, de manière sécante aux lignes de semis, de façon à
éviter au maximum de mesurer le rayonnement direct dans l’interligne (Figure 11). Des piquets
ont été disposés comme repères dans chaque parcelle de mesure afin d’effectuer les mesures aux
mêmes endroits à chaque série de mesures.

50
Figure 11 : Prise de mesures sur une parcelle expérimentale. Le SunScan prend en compte un certain nombre de paramètres tels que l’angle des feuilles par
rapport à la tige, la latitude et la longitude, ainsi que l’angle zénithal qui en découle. L’angle des
feuilles par rapport à la tige ainsi que la fréquence et le nombre de mesures par parcelle ont été
choisis sur base des lectures concernant l’utilisation de ce type d’instrument (Wilhelm et al, 2000 ;
Weiss, 2004).
Une régression linéaire par la méthode des moindres carrés a été effectuée, à l’aide du logiciel
Statistica, entre le LAI issu des images et le PAI issu du Sunscan. La distribution des données
suggérant la possibilité d’une relation non linéaire, une régression de type exponentiel par la
méthode des moindres carrés a également été effectuée. Une validation croisée a ensuite été
effectuée afin de tester à quel point de telles régressions peuvent permettre de prédire le LAI de
référence à partir du PAI du Sunscan. La méthode de validation croisée type « Leave-one-out » a
été retenue car celle-ci est la plus adaptée lorsque peu de données sont disponibles. Elle consiste à
obtenir une régression par la méthode des moindres carrés entre le LAI et le PAI en enlevant un
couple « LAI et PAI correspondant » de données à l’ensemble des données utilisées pour obtenir la
régression qui constitue le modèle. L’opération est ensuite répétée autant de fois qu’il y a de
données, en réintroduisant le couple de données enlevées précédemment et en enlevant un autre
couple de données. On obtient alors autant de régressions qu’il y a de données, chaque régression
ayant été obtenue avec l’ensemble des données moins un couple de données. L’étape suivante
consiste à utiliser chaque régression obtenue pour estimer le LAI vrai (LAI image) à partir du PAI
du couple de données enlevé pour obtenir cette même régression. Le LAI ainsi estimé est alors
soustrait au LAI image réellement mesuré. La différence est mise au carré et est nommée carré de
l’erreur pour cette régression. Chaque régression va fournir ainsi un carré de l’erreur. La racine
carrée de la somme des carrés des erreurs divisée par le nombre de données est le RMSEv (Root
Mean Square Error of validation). Le RMSEv en % d’erreurs est le RMSEv divisé par la moyenne
des valeurs mesurées de LAI.
Ligne de semis
7,5 m
1,5 m
Sonde du SunScan

51
Résultats et discussion
Les valeurs de LAI obtenues sont, pour la plupart, dans l’intervalle des valeurs maximales de LAI
observées pour les cultures annuelles. Celles-ci varient entre 2 et 4 pour les cultures annuelles
(Beadles, 1993).
Les résultats issus de la régression linéaire d’une part et exponentielle d’autre part sont présentés
en Figure 12
Nuage de Points (Tab LAI IMG Vs Sunscan2006-2007-2colonnes 6v*56c)
0 1 2 3 4 5
LAI Sunscan
0
1
2
3
4
5
6
7
LA
Iim
g
y = -3,3626 + 1,7389*x; r² = 0,6525; r = 0,8077; p = 0,0000002 y = -0,3101+exp(-1,0747+(0,599524)*x); r² = 0,6869; r = 0,82884
Figure 12 : Régressions des moindres carrés entre le LAI calculé à partir des surfaces foliaires et le PAI issu du Sunscan.
La régression linéaire obtenue est la suivante :
LAIimg = -3,36 + 1.74 * LAIsunscan ,
La régression non linéaire obtenue est la suivante :
LAIimg = -0,31 + e(-1,07+0,60*LAIsunscan) Les coefficients de détermination et de significativité (Figure 12) indiquent une bonne linéarité
ainsi qu’une bonne relation exponentielle. La significativité élevée est la même pour les deux
relations (p=0.0000002), mais la relation exponentielle permet d’expliquer une plus grande
proportion des LAI « image » à partir du LAI issu du Sunscan (R² relation exponentielle = 69% et
R² relation linéaire = 65%).

52
La validation croisée a été menée sur chacune de ces deux régressions. Les détails concernant les
régressions obtenues avec chaque jeu de données « leave-one-out » de la validation croisée sont
présentés en ANNEXE IV. Le RMSEv divisé par la moyenne des LAIs observés peut être
interprété comme le pourcentage d’erreurs dans la prédiction. L’erreur de prédiction peut donc être
estimée à ~34% et la « prédictivité » du modèle à 66 % pour la régression linéaire, contre des
valeurs de, respectivement, ~33% et 67% pour la régression non linéaire. Ces deux relations
montrent donc des résultats très proches.
Contrairement aux observations faites par Wilhelm et al. (2000) sur le maïs avec le Sunscan,
l’appareil surestime globalement le LAI par rapport à la méthode de référence. Cette surestimation
du Sunscan est mise en évidence par les relations obtenues. Sur les 28 points de la relation, les 20
issus des observations de 2007 montrent une surestimation du Sunscan alors que les 8 points issus
des observations de 2006 montrent une sous-estimation du Sunscan (Figure 13).
Trois facteurs peuvent expliquer cette différence :
- L’utilisation du paramètre b obtenu en 2007 dans le calcul du LAI de 2006.
- L’utilisation d’un autre Sunscan en 2007 par rapport à 2006 pour cause de
disfonctionnement de celui utilisé en 2006.
- Les surfaces foliaires des trois dernières feuilles plus importantes en 2006 qu’en 2007
peuvent être à l’origine d’une surestimation du LAI par le Sunscan à cause de la perte de
sensibilité de cet appareil lorsque l’intensité lumineuse est trop faible (phénomène
mentionné par le constructeur).
Les modèles obtenus présentent des coefficients de détermination faibles (R²=0,65 et 0,68), ce qui
n’indique pas une grande proximité entre les valeurs obtenues de LAI et de PAI.
En conclusion, les modèles ne présentent pas une corrélation suffisamment élevée pour qu’on
puisse prendre en considération sa valeur prédictive, qui est de toute manière peu élevée
également. Une amélioration de la relation pourrait sans doute être obtenue à partir de valeurs de
PAI obtenues avec un seul appareil et des valeurs de LAI obtenues avec le paramètre b de la
culture et de l’année en cours.
L’analyse des résultats obtenus en utilisant la méthode de calcul du LAI de référence sur les
différentes variantes (Figure 13, LAI image) nous amène aux interprétations suivantes :
- Les différences entre les LAI observés en 2006 par rapport à ceux observés en 2007, toutes
variantes confondues, sont très significatives (teste F : p = 0,0000001 ; R²=0,80).
- Les valeurs du témoin Achat ne se distinguent pas de celles du Triple traitement Achat
lorsque les deux années sont confondues (Test F non significatifs).
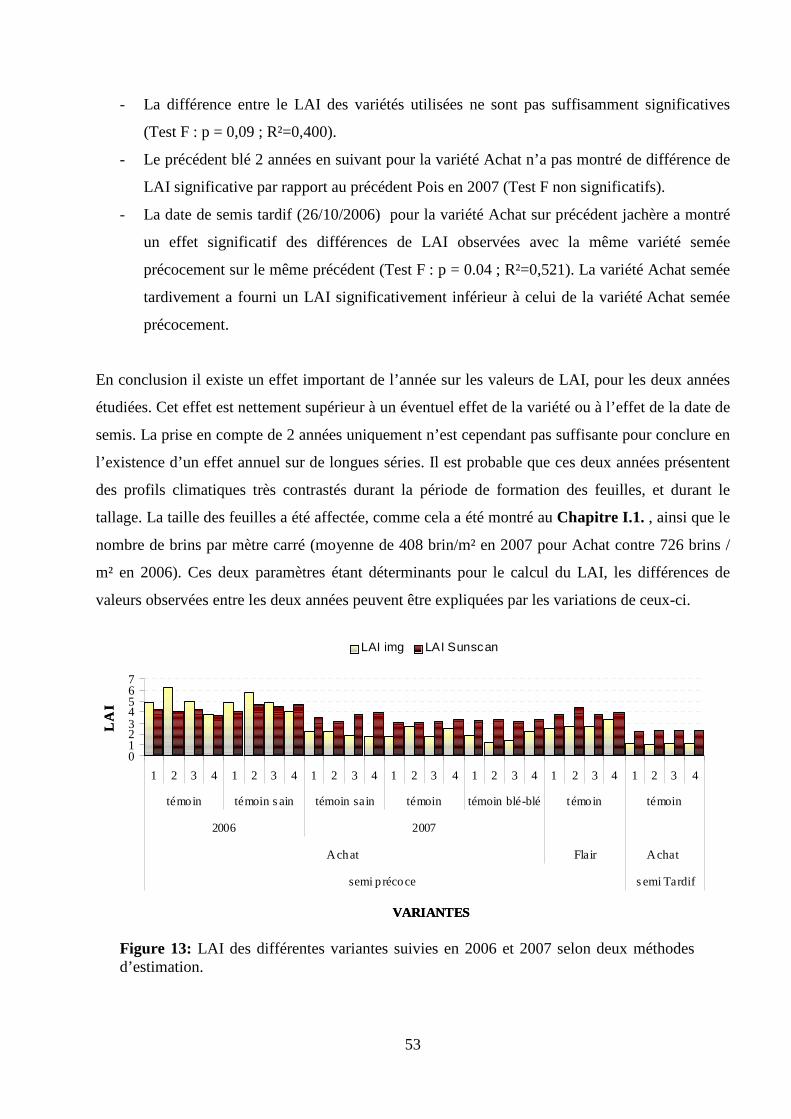
53
- La différence entre le LAI des variétés utilisées ne sont pas suffisamment significatives
(Test F : p = 0,09 ; R²=0,400).
- Le précédent blé 2 années en suivant pour la variété Achat n’a pas montré de différence de
LAI significative par rapport au précédent Pois en 2007 (Test F non significatifs).
- La date de semis tardif (26/10/2006) pour la variété Achat sur précédent jachère a montré
un effet significatif des différences de LAI observées avec la même variété semée
précocement sur le même précédent (Test F : p = 0.04 ; R²=0,521). La variété Achat semée
tardivement a fourni un LAI significativement inférieur à celui de la variété Achat semée
précocement.
En conclusion il existe un effet important de l’année sur les valeurs de LAI, pour les deux années
étudiées. Cet effet est nettement supérieur à un éventuel effet de la variété ou à l’effet de la date de
semis. La prise en compte de 2 années uniquement n’est cependant pas suffisante pour conclure en
l’existence d’un effet annuel sur de longues séries. Il est probable que ces deux années présentent
des profils climatiques très contrastés durant la période de formation des feuilles, et durant le
tallage. La taille des feuilles a été affectée, comme cela a été montré au Chapitre I.1. , ainsi que le
nombre de brins par mètre carré (moyenne de 408 brin/m² en 2007 pour Achat contre 726 brins /
m² en 2006). Ces deux paramètres étant déterminants pour le calcul du LAI, les différences de
valeurs observées entre les deux années peuvent être expliquées par les variations de ceux-ci.
Figure 13: LAI des différentes variantes suivies en 2006 et 2007 selon deux méthodes d’estimation.
01234567
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
témoin témoin s ain témoin sain témoin témoin blé-blé témoin témoin
2006 2007
Achat Flair Achat
semi précoce s emi Tardif
LAI img LAI Sunscan
LA
I
VARIANTES
01234567
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
témoin témoin s ain témoin sain témoin témoin blé-blé témoin témoin
2006 2007
Achat Flair Achat
semi précoce s emi Tardif
LAI img LAI Sunscan
LA
I
VARIANTES

54
II.2. Estimation de la relation existant entre la fraction du sol couverte par la canopée verte, le LAI et la surface verte foliaire des trois dernières feuilles du blé d’hiver.
Introduction.
Le LAI et la proportion de tissus verts sont des paramètres importants des modèles de croissance et
de modélisation des rendements. Ils contribuent directement à la détermination du potentiel
d’absorption des assimilas par la plante. C’est pourquoi de nombreuses tentatives d’estimation du
LAI et du pourcentage de surface verte du couvert à distance ont été mises en place (Blackmer et
White, 1998; Weiss, 2004). La télédétection de la végétation inclut diverses méthodes non
destructives d’analyses spectrales à distance allant des satellites jusqu’aux plates-formes sur le
terrain (Nilsson, 1995). L’intérêt de l’estimation à distance réside dans sa capacité à donner un
aperçu de la répartition spatiale et de l’évolution au cours du temps de la variable étudiée, pour une
région agricole, un territoire ou même à l’échelle globale. Contrairement aux méthodes
d’estimation directe du LAI et de la proportion de surface verte foliaire, l’estimation à distance
n’est pas laborieuse et coûteuse en temps. Malheureusement, et malgré les efforts menés depuis 40
ans, l’estimation du LAI à distance reste un sujet difficile (Guissard, 2005).
L’utilisation de méthodes d’estimation à courte distance mettant en jeu des plates-formes sur le
terrain constitue une étape facile à mettre en œuvre et permet la mise en évidence d’une première
relation entre une méthode d’estimation à distance et une méthode d’estimation de référence du
LAI ou de la surface verte. Dans ce domaine, White (2000) montre que la couverture verte du sol
obtenue à partir de l’analyse d’images aériennes prises à 2,8 m du sol est statistiquement identique
à celle obtenue à partir de l’analyse d’images prises le long d’un transect vertical allant de 0 à 25m
de haut dans un écosystème aride.
L’objectif de cette étude est d’estimer la relation qui peut être établie entre la couverture verte du
sol obtenue à partir d’images aériennes prises à quelques mètres du sol et le LAI ou la surface
verte des trois dernières feuilles d’une culture de blé d’hiver.

55
Matériel et méthodes.
Les expérimentations ont été effectuées en 2007 sur des parcelles expérimentales de cultures mises
en place à Everlange (N49°47’08.91’’- E 5°58’01.36’’- Alt 274 m) au Grand-Duché de
Luxembourg.
Le taux de couverture verte du sol par la canopée a été déterminé à l’aide d’images prises
perpendiculairement au sol, à +/- 3m de haut (Figure 14) entre le 08/05/07 et le 03/07/07. La
surface couverte par les images à la hauteur de la canopée était de 1,10 x 1,40 m. Les images ont
été prises une fois par semaine à l’aide d’un appareil photo numérique Canon 7MG pixels à raison
d’une photo par répétition sur 4 répétitions par variante pour 7 variantes. La prise d’images a été
effectuée à chaque reprise au-dessus du même point.
Figure 14 : Image du dispositif de prise d’images à la verticale et de l’expérimentateur.
Les images obtenues ont ensuite été analysées au moyen de deux logiciels (Assess (APS-Press) et
Ccover-tool (Guissard, 2005)) permettant la classification des pixels de l’image en fonction de
leurs caractéristiques spectrales dans la gamme du visible. La discrimination des pixels a permis
d’obtenir une ségrégation des pixels en 2 groupes bien distincts. L’un comprenant les pixels verts
sous forte, moyenne ou faible luminosité, et l’autre les pixels allant du jaune au brun sous forte,
Appareil photo fixé à l’extrémité d’un axe horizontal.
Double niveau garantissant une prise de vue perpendi- culaire au sol.
Axe vertical.
Déclencheur à distance (système à poire injectant de l’air sous pression dans un fil creux muni d’une tige mobile à son extrémité)

56
moyenne ou faible luminosité. Les résultats obtenus sont en pourcentage de surface couverte par
les pixels verts. Le LAI de référence et le pourcentage de surfaces vertes des trois dernières feuilles
ont été obtenus à partir de l’analyse d’images des trois dernières feuilles prises individuellement.
Les images des feuilles ont été obtenues à l’aide du même appareil photo numérique que celui
utilisé pour la prise d’images verticales de la couverture verte du sol. Les feuilles ont été
photographiées de façon non destructive, en les maintenant temporairement à plat sur un support
de fond bleu à l’aide de papier collant double face. L’appareil photo a été fixé à une distance de 39
cm du support à fond bleu, sur une latte en aluminium, de façon à prendre des photographies à la
perpendiculaire de la feuille de blé dans son entièreté. La surface foliaire a été utilisée pour le
calcul du LAI selon la méthode détaillée dans la section II.1.
Le pourcentage de surface foliaire verte des 3 dernières strates a été calculé par répétition à l’aide
des surfaces foliaires et des surfaces vertes foliaires issues de l’analyse d’images de feuilles
individuelles effectuée avec le logiciel Assess. Ces images ont été prises 2 à 4 fois par mois, à
partir de l’apparition de la dernière feuille et durant 6 à 7 semaines. Une régression des moindres
carrés entre le LAI et la couverture verte du sol a été testée. Seules les valeurs de LAI à la date de
leur maximum et les valeurs de couverture verte du sol aux mêmes dates ont été utilisées car le
LAI des trois dernières feuilles de ces dates-là correspondent au GLAI (Green Leaf Area Index).
La couverture verte a donc plus de chance de correspondre au LAI à ces dates.
Une régression des moindres carrés entre le pourcentage de surface foliaire verte des 3 dernières
feuilles et la couverture verte du sol a également été testée. Dans ce cas, toutes les dates pour
lesquelles on a pu obtenir une surface foliaire verte des 3 dernières strates et une couverture verte
du sol ont été utilisées. Cette dernière analyse a fait l’objet d’une publication (Mackels et Tychon,
2008 ; ANNEXE IV )
Résultats et discussion
L’obtention d’un modèle d’estimation du LAI à partir du pourcentage de couverture verte du sol
par la culture du blé d’hiver a été tentée avec deux logiciels. Chaque logiciel utilisé pour
l’obtention du pourcentage de couverture verte du sol a fourni un modèle.
Le modèle obtenu avec le Logiciel Assess est le suivant :
LAI = 0.045 * GrnCoverAssess– 1.875 ; (p<0,05 ; R²=0,37)
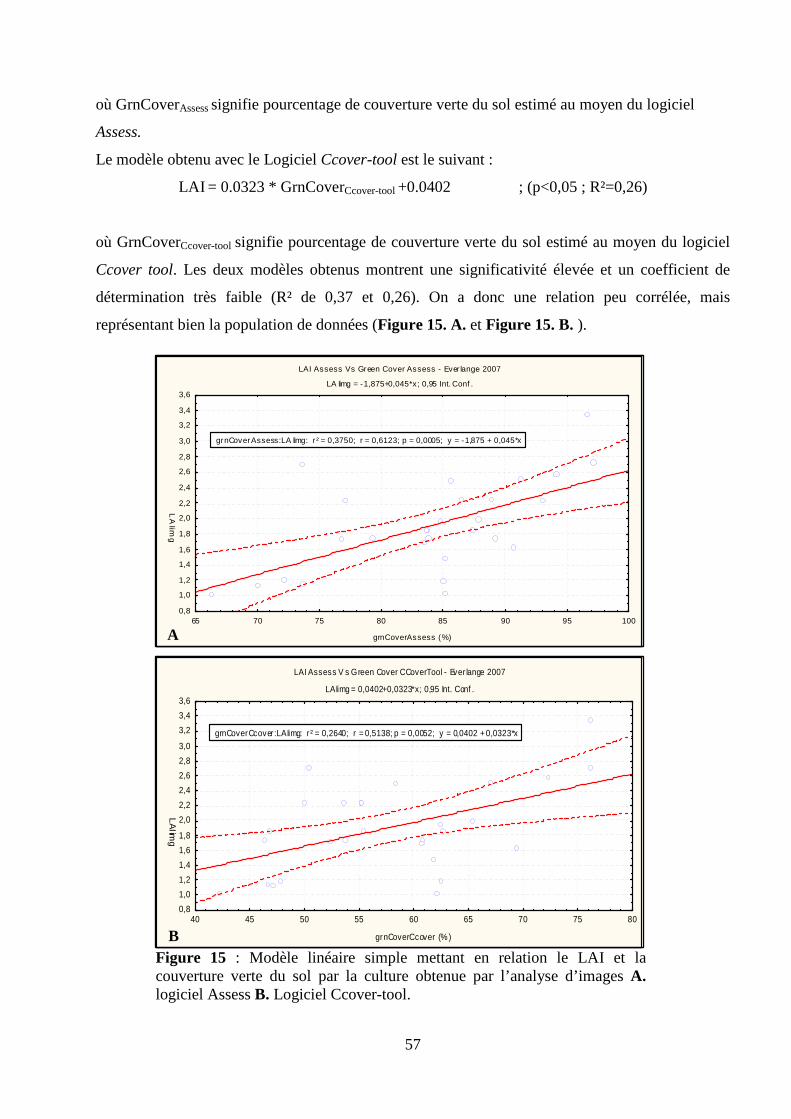
57
où GrnCoverAssess signifie pourcentage de couverture verte du sol estimé au moyen du logiciel
Assess.
Le modèle obtenu avec le Logiciel Ccover-tool est le suivant :
LAI = 0.0323 * GrnCoverCcover-tool +0.0402 ; (p<0,05 ; R²=0,26)
où GrnCoverCcover-tool signifie pourcentage de couverture verte du sol estimé au moyen du logiciel
Ccover tool. Les deux modèles obtenus montrent une significativité élevée et un coefficient de
détermination très faible (R² de 0,37 et 0,26). On a donc une relation peu corrélée, mais
représentant bien la population de données (Figure 15. A. et Figure 15. B. ).
Figure 15 : Modèle linéaire simple mettant en relation le LAI et la couverture verte du sol par la culture obtenue par l’analyse d’images A. logiciel Assess B. Logiciel Ccover-tool.
LAI Assess Vs Green Cover Assess - Ever lange 2007
LA Iimg = -1,875+0,045*x; 0,95 Int. Conf .
65 70 75 80 85 90 95 100
grnCoverAssess (%)
0,8
1,0
1,2
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
3,2
3,4
3,6
LA
Iimg
grnCoverAssess:LA Iimg: r ² = 0,3750; r = 0,6123; p = 0,0005; y = -1,875 + 0,045*x
LAI Assess V s Green Cover CCoverTool - Ever lange 2007
LAIimg = 0,0402+0,0323*x; 0,95 Int. Conf .
40 45 50 55 60 65 70 75 80
grnCoverCcover (% )
0,8
1,0
1,2
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
3,2
3,4
3,6
LA
Iimg
grnCoverCcover :LAIimg: r ² = 0,2640; r = 0,5138; p = 0,0052; y = 0,0402 + 0,0323*x
A
B
LAI Assess Vs Green Cover Assess - Ever lange 2007
LA Iimg = -1,875+0,045*x; 0,95 Int. Conf .
65 70 75 80 85 90 95 100
grnCoverAssess (%)
0,8
1,0
1,2
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
3,2
3,4
3,6
LA
Iimg
grnCoverAssess:LA Iimg: r ² = 0,3750; r = 0,6123; p = 0,0005; y = -1,875 + 0,045*x
LAI Assess V s Green Cover CCoverTool - Ever lange 2007
LAIimg = 0,0402+0,0323*x; 0,95 Int. Conf .
40 45 50 55 60 65 70 75 80
grnCoverCcover (% )
0,8
1,0
1,2
1,4
1,6
1,8
2,0
2,2
2,4
2,6
2,8
3,0
3,2
3,4
3,6
LA
Iimg
grnCoverCcover :LAIimg: r ² = 0,2640; r = 0,5138; p = 0,0052; y = 0,0402 + 0,0323*x
A
B

58
Les données n’étant pas suffisamment corrélées, un tel modèle ne peut être utilisé pour tester sa
valeur prédictive du LAI à partir de valeurs de couverture verte du sol.
Parmi les hypothèses concernant les causes qui peuvent être à l’origine de modèles si peu corrélés,
on peut citer :
- La présence de vent, qui couche les feuilles de blé au moment de la prise d’image. La
couverture verte est alors plus élevée qu’elle ne devrait l’être par rapport aux jours pour
lesquels les mesures ont été effectuées sans vent.
- Une luminosité d’intensité élevée sur certaines images, moyenne et faible sur d’autres. La
classification des pixels d’une image est influencée par cette intensité lumineuse.
- Le LAI ne tient pas compte du recouvrement des feuilles, alors que la couverture verte du
sol en tient compte.
Malgré le recouvrement, on pourrait s’attendre à une bonne relation entre le LAI et la
couverture verte visible en prise de vue verticale.
- L’influence des feuilles encore vertes situées sous les trois strates supérieures du couvert.
L’obtention d’un modèle d’estimation du pourcentage de surface verte des trois dernières feuilles à
partir du pourcentage de couverture verte du sol par la culture du blé d’hiver a également été testée
avec deux logiciels (Mackels et Tychon, 2008 ; ANNEXE IV ). Chaque logiciel utilisé pour
l’obtention du pourcentage de couverture verte du sol a fourni un modèle.
Le modèle obtenu avec le Logiciel Assess est le suivant :
%SVF1,2et3 = 1.6178* GrnCoverAssess– 44.727 ; (p<0,05 ; R²=0,74)
Le modèle obtenu avec le Logiciel Ccover-tool est le suivant :
%SVF1,2et3 = 1.7235* GrnCoverCcover– 15.818 ; (p<0,05 ; R²=0,71)
où %SVF1,2et3 signifie pourcentage de surface verte des trois dernières strates foliaires.
Les deux modèles obtenus montrent une significativité élevée et un coefficient de détermination
aux alentours de 70%. On a donc une relation corrélée à 70 % et représentant bien la population de
données (Figure 16.A. et Figure 16.B.). La régression est cependant influencée par une
distribution des données présentant un plus grand nombre de données dans l’intervalle allant de 90

59
à 100 % de surface verte des trois dernières strates. L’étendue horizontale du plateau formé par
l’accumulation de points dans cette zone montre qu’au grand nombre de valeurs de %SV des trois
dernières strates situées entre 90 et 100% correspondent des taux de couverture verte du sol allant
de 70 à 95%. A cette dispersion s’ajoute celle présente tout le long de la régression. Plus
particulièrement, on constate une part plus importante de dispersion à droite de la droite de
régression. Cette dernière est due, pour la plupart des points, à une surestimation de la couverture
verte du sol engendrée par la prise d’image par jour venteux. Le vent couche les feuilles de blé, ce
qui augmente la surface verte visible sur l’image.
Figure 16 : Modèle linéaire simple mettant en relation le pourcentage de surface verte des trois dernières strates foliaires et la couverture verte du sol par la culture obtenue par l’analyse d’images verticales. A. Logiciel Assess. B. Logiciel Ccover-tool.
Pourcentage de surface verte des 3 dernières strates Vs Couverture verte du sol par la canopée - Everlange 2007
% SV total 3 strates (%) = -44,7271+1,6178*x; 0,95 Int. Conf.
0 10 20 30 40 50 60 70 80 90 100 110
Couverture verte du sol - logiciel Assess (%)
-20
0
20
40
60
80
100
120
% S
V to
tal 3
strate
s (%
)
r² = 0,7363; r = 0,8581; p = 00,0000; y = -44,7271 + 1,6178*x
Pourcentage de surface verte des trois dernières strates Vs pourcentage de couverture verte du sol par la canopéeavec le Ccover-tool - Everlange 2007
% SV total 3 strates (%) = -15,0666+1,7105*x; 0,95 Int. Conf.
-10 0 10 20 30 40 50 60 70 80 90
Couverture verte du sol - logiciel Ccover - tool (%)
-20
0
20
40
60
80
100
120
% S
V to
tal 3
strate
s (%
)
r² = 0,7107; r = 0,8430; p = 00,0000; y = -15,0666 + 1,7105*x
A.
B.
Pourcentage de surface verte des 3 dernières strates Vs Couverture verte du sol par la canopée - Everlange 2007
% SV total 3 strates (%) = -44,7271+1,6178*x; 0,95 Int. Conf.
0 10 20 30 40 50 60 70 80 90 100 110
Couverture verte du sol - logiciel Assess (%)
-20
0
20
40
60
80
100
120
% S
V to
tal 3
strate
s (%
)
r² = 0,7363; r = 0,8581; p = 00,0000; y = -44,7271 + 1,6178*x
Pourcentage de surface verte des trois dernières strates Vs pourcentage de couverture verte du sol par la canopéeavec le Ccover-tool - Everlange 2007
% SV total 3 strates (%) = -15,0666+1,7105*x; 0,95 Int. Conf.
-10 0 10 20 30 40 50 60 70 80 90
Couverture verte du sol - logiciel Ccover - tool (%)
-20
0
20
40
60
80
100
120
% S
V to
tal 3
strate
s (%
)
r² = 0,7107; r = 0,8430; p = 00,0000; y = -15,0666 + 1,7105*x
A.
B.

60
Afin de tenter une diminution de l’effet de cette distribution non équilibrée des données, le nombre
de valeurs situées dans l’intervalle 90 à 100 % de surface verte a été diminué en effectuant une
régression sur les données obtenues à partir de la date du 7 juin (Effectué uniquement avec
l’utilisation du logiciel Assess). La distribution obtenue est Gaussienne et les résultats sont
proches de ceux obtenus en prenant toutes les données (R² = 0,7257 ; p = 00,0000 ; y = -41,2758 +
1,4078*x), mais avec un plateau moins important dans la zone des 90% à 100%SV des trois
dernières strates.
Une validation croisée de type «Leave-one-out » a été effectuée sur les données obtenues.
Le RMSEv (RMSE de validation) entre les valeurs prédites à chaque étape du «Leave-one-out» et
les valeurs observées donnent une indication sur l’aspect prédictif des modèles obtenus. Le
RMSEv du modèle obtenu avec le logiciel Assess est de 19,327, et celui obtenu avec le logiciel
Ccover-tool est de 20,251. Le RMSEv exprimé en pourcentage d’erreurs prédites est de
respectivement 27% et 28%. Ces pourcentages correspondent à un R² de validation de
respectivement 73% et 72%. Ceci indique que ce genre de modèle peut prédire le pourcentage de
surface verte des trois dernières strates avec un taux de réussite de ~ 73 %.
En conclusion, il est difficile d’obtenir un LAImax du blé d’hiver à partir de l’analyse d’images
prises à quelques mètres du sol en utilisant une relation linéaire entre le LAImax et la couverture
verte du sol. Il existe cependant une relation linéaire entre la couverture verte du sol obtenue à
partir de l’analyse de photos aériennes et le pourcentage de surface verte du blé d’hiver de
l’apparition à la sénescence de ces trois dernières feuilles. Cette relation est prédictive à 73 % et
pourrait être testée et améliorée par l’obtention de données supplémentaires. Des améliorations de
la relation peuvent être attendues également si on tient compte des éléments suivants :
- La présence de vent, qui augmente les valeurs de couverture verte du sol.
- L’intensité lumineuse, qui influence la classification des pixels de l’image. En radiométrie
de la canopée, il y a une influence de la variation de l’angle solaire durant le jour sur la
réflectance de la canopée et ses interactions avec l’orientation des lignes de semis (Jackson,
1978 ; Pinter et al., 1983)
- L’influence des feuilles encore vertes situées sous les trois strates supérieures du couvert.
Une meilleure discrimination des pixels très ombragés pourrait permettre d’exclure ces
feuilles.
Si l’observation faite par White (2000), selon laquelle la couverture verte du sol obtenue à partir de
l’analyse d’images aériennes à quelques mètres du sol est identique à celle obtenue à partir de
l’analyse d’images prises à plus haute altitude se confirme pour la culture du blé d’hiver, cela
aurait un intérêt pour les modèles d’estimation des rendements.

61
En effet, ceux-ci utilisent la proportion de tissus sains des trois dernières strates foliaires. Cette
proportion nécessite pour le moment beaucoup de temps à être obtenue à l’échelle des feuilles, des
plantes et des parcelles. Il serait donc intéressant de pouvoir l’obtenir rapidement et pour de plus
grandes surfaces que celles des parcelles expérimentales au moyen d’analyses d’images aériennes.
Dans la littérature, des images aériennes de basse altitude ont déjà été utilisées auparavant pour
étudier la variabilité spatiale de la croissance de certaines cultures et la relation existant avec la
variabilité spatiale du rendement (Buerkert et al., 1996 ; Gérard et Buerkert, 2001). Dans leur
étude de 2001, Gérard et Buerkert ont montré qu’il était possible de suivre la croissance des
cultures de mil à partir du NDVI, mais aussi à partir de la normalisation de la véritable bande verte
d’images en vraies couleurs. La relation entre la surface verte des trois dernières strates foliaires
des couverts de blé et un indice basé sur la normalisation de la bande verte d’images aériennes
dans le visible pourrait également être étudié dans notre cas. Une relation entre un tel indice et le
rendement pourrait également être étudiée. Ce genre de recherche sur le blé pourrait servir d’étape
intermédiaire aux études tentant d’établir un lien entre le rendement observé au sol et l’analyse de
données satellites.

62
Conclusion générale de la partie I Cette première partie concerne l’acquisition des principales données d’entrée des modèles
d’estimation des rendements qui seront testés dans la seconde partie. Ces données d’entrée sont le
LAI et le pourcentage de surface verte. La méthode d’obtention du LAI et du pourcentage de
surface verte par l’analyse d’images ont été choisies comme méthode de référence et ont été
validées avec succès.
L’analyse de la dégradation de la surface verte foliaire des trois dernières strates foliaires de 2006
et 2007 a permis de mettre en évidence la présence d’un effet dû à l’année sur la taille des surfaces
foliaires vertes. Il a été observé que les fongicides retardent la décroissance de la surface verte
plutôt qu’ils ne réduisent son rythme de progression, celle-ci une fois entamée. Aucune différence
variétale, de traitement phytosanitaire ou de précédent cultural n’a été relevée entre les sigmoïdes
ajustées à la dégradation de la surface verte. Aucune des deux relations (régression linéaire et non
linéaire) établies entre le plateau ou le point d’inflexion de ces courbes et le rendement n’a fourni
de résultats significatifs.
L’analyse de la dégradation de la surface verte par l’étude du pourcentage de surface verte des
trois dernières strates a mené à de meilleurs résultats. Une courbe de Gompertz simplifiée, de
forme similaire à la sigmoïde adaptée à la dégradation des surfaces vertes en cm², a été adaptée au
pourcentage de surface verte. Les valeurs du point d’inflexion, m, de cette courbe ont permis de
distinguer la variante traitée de la variété Achat du témoin non traité, sans pour autant mettre de
différence variétale en évidence. Une bonne corrélation a été obtenue entre le rendement et les
valeurs de m pour l’ensemble des variantes testées en 2006 et 2007. Ces conclusions supportent
celles obtenues par Gooding et al.(2000), selon lesquelles les traitements phytosanitaires retardent
l’arrivée de la décroissance de la surface verte, et que ce maintien supplémentaire de la surface
verte est corrélé à l’obtention de rendements plus importants. La corrélation obtenue entre le
paramètre m et les rendements confirme l’intérêt d’explorer les modèles d’estimation des
rendements utilisant celle-ci comme principale variable explicative.
La comparaison de la méthode de référence avec la méthode d’estimation visuelle du pourcentage
de surface verte a été effectuée afin de déterminer les différences entre une méthode de référence
fiable et une méthode plus rapide. Cette comparaison a mis en évidence une tendance à la
surestimation du pourcentage de surface verte de la part des utilisateurs de cette dernière méthode.
Cette surestimation est relativement plus importante pour les pourcentages de surface verte
inférieurs à 15%. La surestimation diminue ensuite progressivement pour les pourcentages
supérieurs à cette valeur.

63
La présence d’une corrélation avec les valeurs de référence uniquement pour les pourcentages
supérieurs à 70 % ne nous permet pas de considérer ces valeurs comme étant proches des valeurs
réelles. Les estimations visuelles peuvent cependant avoir leur utilité si on utilise la moyenne par
classe de pourcentage, puisqu’on a montré que les moyennes par classe de pourcentage (10 classes
de 10%) sont proches des moyennes obtenues avec les valeurs de référence. De plus, l’utilisation
de valeurs moyennes ont déjà montré leur utilité dans plusieurs travaux sans pour autant avoir été
utilisés par classe de pourcentage (El Jarroudi et al., 2002 ; El Jarroudi, 2005).
La méthode de référence d’obtention du pourcentage de surface verte des trois dernières strates a
également été comparée à la méthode plus rapide, automatisable et spatialisable d’obtention de la
couverture verte par l’analyse d’images aériennes à quelques mètres du sol. Le modèle linéaire
simple obtenu entre le pourcentage de surface verte des trois dernières strates foliaires et la
couverture verte du sol présente une bonne corrélation. Cette relation, soumise à une validation
croisée du type « leave-one-out » indique que la surface verte des trois dernières strates peut être
estimée à partir de la couverture verte du sol dans 73 % des cas. L’obtention d’une meilleure
relation semble possible à partir d’un certain nombre d’améliorations des prises de mesures.
L’analyse du LAI, autre principale donnée d’entrée des modèles d’estimation des rendements
utilisés par la suite, et issue de l’utilisation des surfaces foliaires obtenue par l’analyse d’images a
mis en évidence un effet dû à l’année, en plus d’un effet de la date de semis. Cette méthode
d’obtention du LAI est la méthode de référence car elle utilise des mesures proches de la réalité.
Elle a été utilisée afin de tester la validité de la méthode d’obtention du LAI à l’aide d’un
radiomètre (Sunscan), méthode plus rapide et automatisable. Le Sunscan ne peut pas être utilisé
pour les modèles d’estimation du rendement car le LAI qu’il fournit n’est pas suffisamment
corrélé à celui issu de la méthode de référence. Les valeurs fournies par le Sunscan ont été
considérées comme étant des valeurs de PAI (Plant Area Index). Un modèle linéaire simple et un
modèle non linéaire entre le PAI et le LAI de référence n’ont pas été suffisamment corrélés pour
qu’ils puissent être utilisés de manière à fournir un LAI à partir de valeurs de PAI du Sunscan.
Le LAI de référence et la couverture verte du sol ont également été mis en relation. La relation
linéaire a fourni des résultats insuffisants pour qu’on puisse utiliser la méthode d’analyse d’images
aériennes de la couverture verte du sol à des fins d’estimation du LAI sur une grande échelle.

64
PARTIE II
MODELISATION DES PERTES DE RENDEMENT.

65
Note préalable
La bibliographie abordée dans l’introduction générale a montré l’historique des démarches
envisagées afin d’estimer les pertes de rendement à l’aide de modèles simples. Ces modèles,
souvent empiriques, étudient la capacité qu’ont certains paramètres (par exemple les symptômes
des maladies ou la surface verte) à rendre compte des processus physiologiques complexes de
production des rendements et des pertes observées.
Il est clairement mis en évidence, dans l’évolution des démarches, que les approches étudiant les
surfaces foliaires saines ont été envisagées suite à la difficulté de relier les symptômes de maladies
au rendement (Aquino et al., 1992 ; Bastiaans et al., 1994 ; Nutter et Littrell, 1996).
Une approche reliant le rendement au rayonnement journalier intercepté par la surface foliaire
saine de cultures attaquées par des maladies a montré de très bons résultats (Waggoner et Berger,
1987). La supériorité de la relation entre rendement et rayonnement intercepté par la surface
foliaire saine à celle entre rendement et évolution des symptômes a même été mise en évidence
(Bergamin Filho et al., 1997).
Récemment, Bancal et al. (2007) ont montré, pour un couvert de blé tendre d’hiver attaqué par la
rouille et la septoriose, qu’il est possible de prédire à la fois la croissance et le rendement à partir
d’un modèle simple prenant en compte principalement la proportion de surface verte foliaire par
strate, les réserves de préfloraison ainsi que le rayonnement intercepté par chaque strate foliaire.
Dans cette PARTIE II , le LAI de référence ainsi que la proportion journalière de tissus sains
obtenus dans la PARTIE I seront utilisés afin de valider (CHAPITRE I), améliorer et simplifier
(CHAPITRE II) le modèle de Waggoner et Berger (1987) et le modèle de Bancal et al. (2007) pour
différentes variétés, traitements et précédents culturaux du blé tendre d’hiver au Grand Duché de
Luxembourg.
Les résultats obtenus avec ces deux modèles seront comparés et le modèle le plus adéquat sera
ensuite retenu pour effectuer une simulation des risques de pertes de rendement (PARTIE III ).

66
CHAPITRE I : Validation d’un modèle d’estimation des rendements du blé d’hiver au Grand Duché de Luxembourg.
Ce chapitre aborde la validation des modèles d’estimation des rendements de Waggoner et Berger
(1987) et de Bancal et al. (2007), tous deux basés sur l’interception du rayonnement incident
journalier par la surface verte foliaire des trois dernières feuilles, de la floraison à la maturité.
Le modèle de Bancal et al. (2007) se distingue de celui de Waggoner et Berger principalement par
deux aspects.
D’une part, le modèle de Bancal et al. (2007) prend en compte, au travers de l’efficience
d’interception du rayonnement (Ei), l’ombrage des strates supérieures dans le calcul du
rayonnement reçu par les F2 et les F3.
D’autre part, le modèle de Bancal et al. (2007) permet d’estimer la matière sèche journalière
produite grâce à l’utilisation d’un paramètre rendant compte du rythme auquel le rayonnement
intercepté par la culture est converti en biomasse par les processus de la photosynthèse. Ce
paramètre est l’efficience d’utilisation du rayonnement ou R.U.E. (Radiation Use Efficiency) et
s’exprime en grammes de matière sèche par Méga Joule.
I.1. Validation du modèle de calcul du HAA.
Matériel et méthodes.
Dans cette méthode, il est considéré que le rendement peut être exprimé comme une fonction du
rayonnement intercepté par la surface foliaire saine journalière (HRI pour Healthy Radiation
Interception) intégrée sur la période de remplissage des grains (HAA pour Healthy leaf Area
Absorption).
Le calcul de la proportion du rayonnement intercepté par la surface foliaire saine en un jour donné
est basé sur la loi de Beer et s’exprime comme suit :

67
( )[ ]eISVHRI ikLAIii −= −1%
Où %SV i est le pourcentage de surface verte de la ou des strates foliaires considérées au jour i. Il est issu de
l’analyse des images des trois dernières feuilles de 5 plantes par répétition pour 4 répétitions par variante le
jour i (cf. PARTIE I, Chapitre I.1). Les images sont prises de façon hebdomadaire pour chaque feuille des 5
plantes marquées au moyen d’une bague en plastique. Les variantes étudiées sont la variété Achat témoin non
traitée et la variété Achat triple traitement (3 traitements) en 2006, ainsi que les variétés Achat témoin non
traitée, Achat triple traitement (3 traitements), Flair témoin non traitée, Akteur témoin non traitée et Achat
témoin précédent blé deux années non traitée en 2007. Les expérimentations ont été mises en place à
Everlange. La disposition des parcelles, les traitements appliqués, les caractéristiques des prises d’images
effectuées pour l’analyse des surfaces vertes ainsi que les caractéristiques de l’itinéraire cultural sont repris
respectivement dans les Annexes II, III et V.
I i correspond au rayonnement solaire incident du jour i exprimé en MJm-2. Il est obtenu à partir des données
issues du pyranomètre de la station météorologique la plus proche.
k a une valeur de 0,68 et correspond au coefficient d’absorption des feuilles de blé (issu du modèle CERES).
Le LAI est l’indice d’aire foliaire calculé à partir des données de surface foliaire issues de
l’analyse des images, de la même manière et pour les mêmes variantes qu’au Chapitre II.1 de la
PARTIE I . Le LAI est exprimé en m² par m² de sol.
Le HAA a ensuite été calculé, pour chacune des trois dernières feuilles, pour 5 plantes par
répétition pour 4 répétitions par variante sur la période qui s’étend de l’apparition de la F1 jusqu’à
la maturité finale. Les variantes étudiées sont la variété Achat témoin non traitée et la variété Achat
triple traitement (3 traitements) en 2006, ainsi que les variétés Achat témoin non traitée, Achat
triple traitement (3 traitements), Flair témoin non traitée, Akteur témoin non traitée et Achat
témoin précédent blé deux années non traitée en 2007.
Le HAA a été obtenu comme suit :
HAA = I i ({%SVi [1-e(-kLAIi)] + %SVi+1 [1-e(-kLAI
i+1)]} /2) (t i+1-t)
Où %SV i correspond au pourcentage de surface verte issu de l’analyse d’image le jour i
I i correspond au rayonnement solaire incident du jour i exprimé en MJm-². Il est obtenu à partir des données
issues du pyranomètre de la station météorologique la plus proche.
k a une valeur de 0,68 et correspond au coefficient d’interception des feuilles de blé. Cette valeur est issue du
modèle CERES.
LAI est l’indice d’aire foliaire calculé à partir des données de surfaces foliaires issues de l’analyse des images
et exprimé en m² par m² de sol.
∑
−
=
1
1
n
i

68
Les rendements utilisés pour les analyses ont été obtenus par répétition en récoltant chacune
d’elles à l’aide d’une moissonneuse batteuse. D’autre part, pour l’année 2007, chaque plante suivie
individuellement par la prise d’images hebdomadaires a été récoltée préalablement à la récolte des
répétitions. Pour chacune de ces plantes, le nombre et le poids à 15% d’humidité des grains ont été
déterminés.
Deux types d’analyses ont été entrepris :
- Les analyses descriptives du HAA total obtenu de la floraison à la maturité et ses
relations avec les rendements.
- Les analyses des caractéristiques des courbes d’évolution du cumul du HAA de la
floraison à la maturité et les relations entre celles-ci et les rendements.
Analyses du HAA total obtenu de la floraison à la maturité .
1. Une description graphique des différences de HAA total obtenues sur la période visée
entre les différentes répétitions de chaque variante sera exposée.
2. Une régression multiple et des analyses de variance mettent en évidence les éventuelles
différences significatives entre les années, les variétés, les traitements, le précédent et la
date de semis.
3. Une régression multiple, et des régressions simples par la méthode des moindres carrés
entre le HAA de la floraison à la maturité et les rendements à Everlange mettront en
évidence une éventuelle relation avec les rendements obtenus.
L’analyse est effectuée sur les données des deux années, sur une année ou sur la variété
Achat seule à raison d’une valeur moyenne de HAA par répétition pour les 4 répétitions par
variante. Une analyse sur les données de la variété Achat seule peut être entreprise car c’est
la principale variété présente. Il y a donc un nombre de données suffisamment important
pour que des analyses sur cette variété seule puissent être entreprises.
4. La régression par la méthode des moindres carrés entre le HAA de la floraison à la
maturité de chaque plante et les caractéristiques de rendement de chacune des 5 plantes
par répétition suivies en 2007.
Analyses des caractéristiques des courbes d’évolution du cumul du HAA.
5. Une analyse graphique de l’ajustement d’une courbe à l’évolution du cumul du HAA en
fonction du cumul des degrés jours (dj) depuis la floraison jusqu’à la maturité sur
l’ensemble des données de 2006 et 2007, pour chaque répétition de chaque variante suivie.
La courbe obtenue est une sigmoïde d’équation HAA=a/(1+exp(v*(i-dj))).

69
6. Une régression multiple et des analyses de variance mettront en évidence les éventuelles
différences significatives entre les paramètres des différentes courbes obtenues.
7. Une régression multiple, et des régressions linéaires simples par la méthode des
moindres carrés entre les paramètres a (plateau) ou i (point d’inflexion) de chaque courbe
obtenue et le rendement de la répétition correspondante.
Toutes les analyses statistiques ont été effectuées à l’aide des logiciels Excel et Statistica.
Résultats et discussion.
Analyses du HAA total obtenu de la floraison à la maturité .
1. Les valeurs de HAA des variantes suivies à la fois en 2006 et 2007 sont nettement supérieures
en 2006 (Figure 17). Cette figure permet également de visualiser une différence entre le témoin et
le triple traitement de Achat (= Témoin Sain dans la figure).
Figure 17 : Histogramme des HAA obtenus pour chaque répétition de chaque variante en 2006 et 2007 à Everlange (Témoin Sain = triple traitement).
2. La régression multiple effectuée entre le HAA et les différentes modalités d’année, traitement,
variété, et précédent montre un effet significatif de chaque modalité à l’exception du précédent
cultural (Tableau 9). Le paramètre Bêta du Tableau 9 donne l’importance relative de chaque
modalité. Dans un ordre croissant, c’est l’année, le traitement et la variété qui expliquent les
variations de HAA observées.
01000020000300004000050000600007000080000
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère blé-blé Jachère Jachère
Témoin Sain Témoin Témoin Témoin
Achat Akteur Flair
2006 2007
HA
A (
MJ
m-2)
VARIANTES
01000020000300004000050000600007000080000
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère blé-blé Jachère Jachère
Témoin Sain Témoin Témoin Témoin
Achat Akteur Flair
2006 2007
HA
A (
MJ
m-2)
VARIANTES

70
Tableau 9 : Résultats de la régression multiple effectuée entre le HAA et les différentes modalités présentes.
HAA
synthèse régression R²= ,910 ; p<0,000001
Bêta p Année -0,797 ***
Variété 0,178 *
Traitement 0,448 ***
Précédent n.s. n.s. Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Cet effet important de l’année a été mis en évidence par l’analyse de variance entre le témoin
Achat des deux années (p=0.0005 et R²=0,88), et entre la variété Achat traitée des deux années
(p=0.000001 et R²=0,99). Cette différence est principalement due aux valeurs de LAI élevées
obtenues en 2006, en raison des surfaces foliaires supérieures observées. Une analyse de variance
de tous les effets (variété, traitement, précédent et date de semis), à l’exception de l’année, sur
l’ensemble des données montre que seule la variante Achat Triple traitement se distingue des
autres de façon hautement significative (p=0,0002). L’analyse de variance entre Achat Témoin et
triple traitement, pour chaque année prise individuellement, montre un effet significatif élevé du
traitement sur les valeurs de HAA (p=0,00005, R²= 0,95 en 2006 et p=0,03, R²=0,56 en 2007).
3. La régression multiple entre les rendements et le HAA, les différentes modalités d’année,
variété, traitement et précédent n’ont pas fourni de résultats significatifs. Des régressions simples
par la méthode des moindres carrés entre le HAA de la floraison à la maturité et le rendement
(ou le poids de 1000 grains) ont été testées afin de déterminer plus en détail un éventuel effet. Ces
analyses visent à évaluer l’influence de l’année et de la principale variété présente sur la qualité de
la relation linéaire entre le HAA et le rendement. La régression simple entre le rendement et le
HAA, effectuée sur l’ensemble des données de 2006 et 2007 n’est pas significative, que toutes les
variantes pour lesquelles il y a un suivi de la surface verte soient utilisées, ou que seule la variété
Achat soit utilisée (Tableau 10).
Les régressions simples significatives sont celles effectuées par année. En 2006, la relation très
significative et de coefficient de détermination élevé (R²=82%) n’est établie que sur les 8 données
présentes cette année là. De plus, seule la variété Achat est présente en 2006. En 2007, la relation
explique une moins grande proportion des rendements obtenus (R²=46%), mais 20 données sont
utilisées avec 3 variétés et deux traitements (Tableau 10 et Figure 22). La relation portant
uniquement sur la variété Achat en 2007, variété la plus présente, augmente le R². Toutes ces
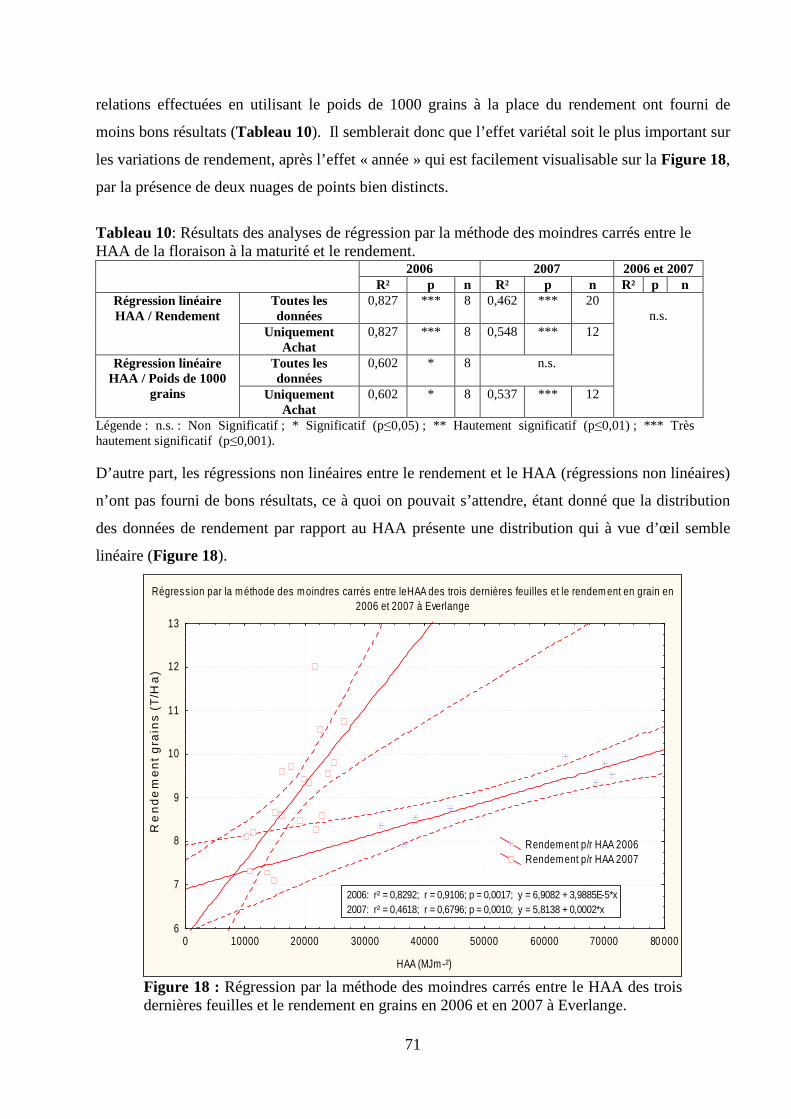
71
relations effectuées en utilisant le poids de 1000 grains à la place du rendement ont fourni de
moins bons résultats (Tableau 10). Il semblerait donc que l’effet variétal soit le plus important sur
les variations de rendement, après l’effet « année » qui est facilement visualisable sur la Figure 18,
par la présence de deux nuages de points bien distincts.
Tableau 10: Résultats des analyses de régression par la méthode des moindres carrés entre le HAA de la floraison à la maturité et le rendement.
2006 2007 2006 et 2007 R² p n R² p n R² p n
Toutes les données
0,827 *** 8 0,462 *** 20 Régression linéaire HAA / Rendement
Uniquement Achat
0,827 *** 8 0,548 *** 12
Toutes les données
0,602 * 8 n.s. Régression linéaire HAA / Poids de 1000
grains Uniquement Achat
0,602 * 8 0,537 *** 12
n.s.
Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001). D’autre part, les régressions non linéaires entre le rendement et le HAA (régressions non linéaires)
n’ont pas fourni de bons résultats, ce à quoi on pouvait s’attendre, étant donné que la distribution
des données de rendement par rapport au HAA présente une distribution qui à vue d’œil semble
linéaire (Figure 18).
Régress ion par la méthode des moindres carrés entre leHAA des trois dernières feuilles et le rendem ent en grain en2006 et 2007 à Everlange
0 10000 20000 30000 40000 50000 60000 70000 80 000
HAA (MJm-²)
6
7
8
9
10
11
12
13
Re
nd
em
en
t g
rain
s (
T/H
a)
2006: r² = 0,8292; r = 0,9106; p = 0,0017; y = 6,9082 + 3,9885E-5*x 2007: r² = 0,4618; r = 0,6796; p = 0,0010; y = 5,8138 + 0,0002*x
Rendement p/r HAA 2006 Rendement p/r HAA 2007
Figure 18 : Régression par la méthode des moindres carrés entre le HAA des trois dernières feuilles et le rendement en grains en 2006 et en 2007 à Everlange.

72
4. Les régressions par la méthode des moindres carrés entre le HAA de la floraison à la maturité
et les caractéristiques du rendement par plante en 2007 ne fournissent aucun bon résultat
(Tableau 11). Ce type de régression a été mené entre le HAA et le nombre de grains par épi, le
poids des grains par épi à 15% d’humidité et le poids de 1000 grains de chaque épi. Quand la
relation est significative, la régression obtenue n’explique qu’une faible proportion de la
distribution des données (R²<33%). La distribution des données montre dans chaque cas une
grande dispersion (exemple Figure 19), ce qui ne suggère pas qu’il soit possible d’adapter une
autre relation que la régression linéaire.
Tableau 11 : Résultats des analyses de régression par la méthode des moindres carrés entre le HAA de la floraison à la maturité et les caractéristiques de rendement par plante en 2007.
2007 R² p n
Toutes les données 0,106 *** 100 Régression linéaire HAA / poids en grains épi Uniquement Achat 0,152 ** 60 Toutes les données 0,213 *** 100 Régression linéaire HAA / Poids de 1000 grains plte Uniquement Achat 0,322 *** 60 Toutes les données n.s. 100 Régression HAA et nbre de grains par épi. Uniquement Achat n.s. 60
Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Nuage de Points (Feuille de données1 10v*100c)
H AA = 205,716 +6,171 1*x; 0,95 In t. Conf.
20 25 30 35 40 45 50 55 60 65
pds 1000 grains (g)
250
300
350
400
450
500
550
600
650
700
750
HA
A (
MJ
/m²)
pds 1000grains :HAA: r² = 0,2131; r = 0,4616; p = 0,000002; y = 205,716 + 6,1711*x
Figure 19 : Régression par la méthode des moindres carrés entre le HAA et le poids de 1000 grains par plante en 2007.
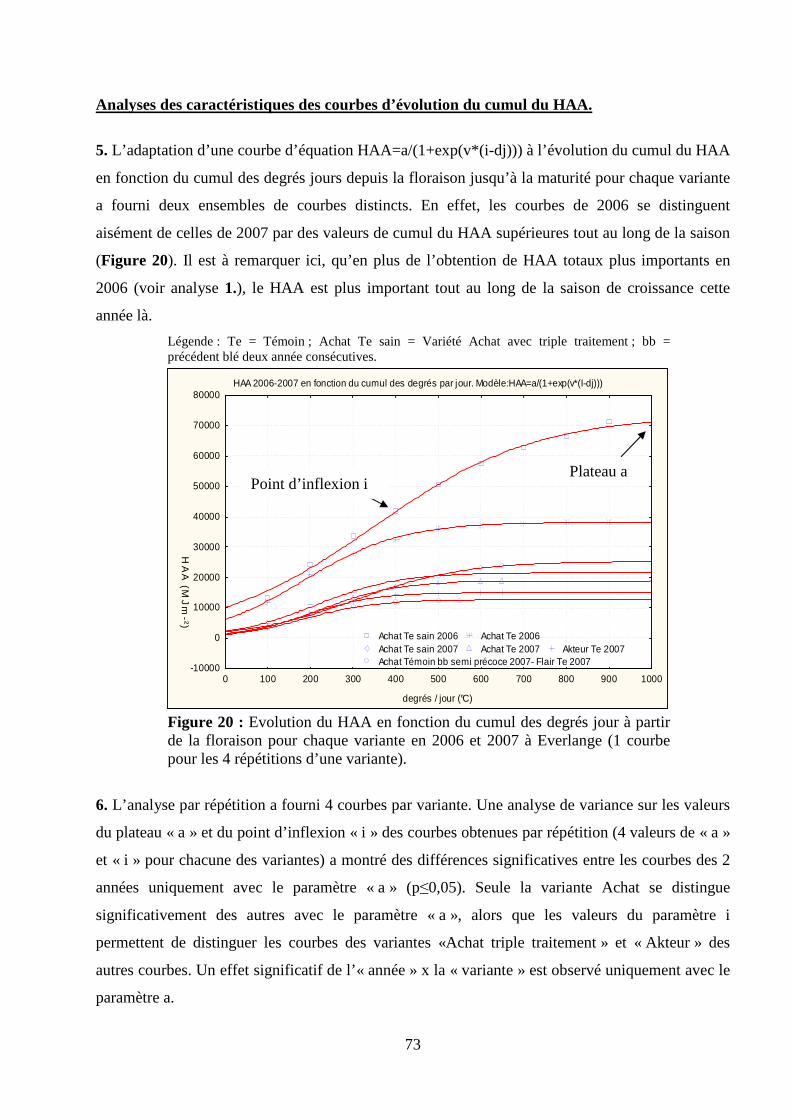
73
Analyses des caractéristiques des courbes d’évolution du cumul du HAA.
5. L’adaptation d’une courbe d’équation HAA=a/(1+exp(v*(i-dj))) à l’évolution du cumul du HAA
en fonction du cumul des degrés jours depuis la floraison jusqu’à la maturité pour chaque variante
a fourni deux ensembles de courbes distincts. En effet, les courbes de 2006 se distinguent
aisément de celles de 2007 par des valeurs de cumul du HAA supérieures tout au long de la saison
(Figure 20). Il est à remarquer ici, qu’en plus de l’obtention de HAA totaux plus importants en
2006 (voir analyse 1.), le HAA est plus important tout au long de la saison de croissance cette
année là.
Légende : Te = Témoin ; Achat Te sain = Variété Achat avec triple traitement ; bb = précédent blé deux année consécutives.
Figure 20 : Evolution du HAA en fonction du cumul des degrés jour à partir de la floraison pour chaque variante en 2006 et 2007 à Everlange (1 courbe pour les 4 répétitions d’une variante).
6. L’analyse par répétition a fourni 4 courbes par variante. Une analyse de variance sur les valeurs
du plateau « a » et du point d’inflexion « i » des courbes obtenues par répétition (4 valeurs de « a »
et « i » pour chacune des variantes) a montré des différences significatives entre les courbes des 2
années uniquement avec le paramètre « a » (p≤0,05). Seule la variante Achat se distingue
significativement des autres avec le paramètre « a », alors que les valeurs du paramètre i
permettent de distinguer les courbes des variantes «Achat triple traitement » et « Akteur » des
autres courbes. Un effet significatif de l’« année » x la « variante » est observé uniquement avec le
paramètre a.
HAA 2006-2007 en fonction du cumul des degrés par jour. Modèle:HAA=a/(1+exp(v*(I-dj)))
0 100 200 300 400 500 600 700 800 900 1000
degrés / jour (°C)
-10000
0
10000
20000
30000
40000
50000
60000
70000
80000
HA
A (M
Jm
-²)
Achat Te sain 2006 Achat Te 2006Achat Te sain 2007 Achat Te 2007 Akteur Te 2007 Achat Témoin bb semi précoce 2007- Flair Te 2007
Point d’inflexion i Plateau a

74
7. Bien qu’il n’y ait pas toujours de différences significatives entre les valeurs des paramètres « a »
et « i » des courbes d’évolution du HAA des différentes variantes, il se peut qu’il y ait une relation
entre les valeurs de ces paramètres et les rendements observés. Une régression multiple entre le
rendement et les deux variables explicatives de la forme des courbes de cumul du HAA montre
que seuls le traitement et le précédent cultural expliquent les variations de rendement. Le
traitement a un poids plus important (Tableau 12).
La régression multiple montre un coefficient de régression très peu élevé (R² de 55 %). Il faut donc
considérer les résultats de celle-ci comme peu explicatifs. Ces résultats sont probablement dus au
grand nombre de variables de cette régression par rapport au nombre de données.
Tableau 12 : Résultats de la régression multiple effectuée entre le rendement et les variables explicatives de la forme des courbes de cumul du HAA.
Rendement
Synthèse régression Test F : R²= ,55 ; p<0,0008 ; n=28
Bêta p Année -0,123 n.s. Variété -0,0364 n.s.
Traitement -0,636 ** Précédent -0,524 ***
a -0,690 n.s. i 0,319 n.s.
Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Des régressions simples entre le rendement et le paramètre « i » ou « a » ont été effectuées afin de
déterminer la corrélation de ces paramètres avec les rendements obtenus sans distinguer les
variétés ou traitements. Les relations obtenues par année sont non significatives lorsque
uniquement les données de l’année 2006 ou uniquement les données de la variété Achat sont
utilisées (Tableau 13). En 2006, peu de données étaient disponibles, raison qui pourrait expliquer
pourquoi les relations ne sont pas significatives.
Le paramètre « i » (point d’inflexion) est corrélé au rendement lorsque toutes les données de 2007
(R² de 84%), ou de 2006 et 2007 (R² de 62%) sont utilisées. Le paramètre « a » (plateau) est
corrélé au rendement uniquement lorsque l’analyse porte sur les données de 2007. Il semblerait
donc que la valeur du point d’inflexion i des courbes d’évolution du HAA de la floraison à la
maturité en fonction des degrés jour soit le seul bon indicateur du rendement.

75
Tableau 13 : Résultats des analyses de régression par la méthode des moindres carrés entre le paramètre « i » ou « a » et le rendement.
2006 2007 2006 et 2007 R² p n R² p n R² p n
Toutes les données 0,843 * 20 0,614 * 28 Régression linéaire « i » / Rendement Uniquement Achat n.s. n.s. Toutes les données 0,841 * 20 n.s. Régression linéaire « a » / Rendement Uniquement Achat
n.s.
n.s. n.s. Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
I.2. Validation du modèle de calcul de la matière sèche.
Matériel et méthodes.
Cette méthode consiste à modéliser la production de matière sèche journalière de la culture ainsi
qu’à établir une relation entre le cumul de la production journalière de matière sèche de la
floraison à la maturité et les rendements obtenus. La mobilisation des réserves de la préfloraison
accumulées dans les tiges n’est pas prise en compte ici, contrairement à l’étude de Bancal et al.
(2007).
La matière sèche journalière produite est calculée à partir du rayonnement global (Rg) journalier
intercepté par la surface verte des trois dernières feuilles. La matière sèche a été calculée, tout
comme le HAA, pour chacune des trois dernières feuilles, pour 5 plantes par répétition pour 4
répétitions par variante. Les variantes étudiées sont la variété Achat témoin non traitée et la variété
Achat triple traitement (3 traitements) en 2006, ainsi que les variétés Achat témoin non traitée,
Achat triple traitement (3 traitements), Flair témoin non traitée, Akteur témoin non traitée et Achat
témoin précédent blé deux années non traitée en 2007. La formule utilisée est la formule de
Monteith adaptée comme suit :
MSJ = Ei(j) Eb Ij
Où MSJ est la matière sèche journalière en g /m² jour.
Eb est l’efficience d’utilisation du rayonnement ou R.U.E. (Radiation Use Efficiency). Pour cette année, une
valeur de 2,8 issue d’une étude de Bryson et al. (1995) a été utilisée. L’ Eb correspond au rapport poids sec en
grain (g) / quantité de rayonnement incident intercepté par les tissus verts dans la gamme du rayonnement
photosynthétiquement actif à partir de la floraison (Mjoules).

76
I j correspond dans cette formule au rayonnement solaire global dans la gamme du rayonnement
photosynthétiquement actif du jour i exprimé en MJm-2. Il est obtenu à partir du rayonnement solaire incident
global issu du pyranomètre de la station météorologique la plus proche des parcelles expérimentales (~50%
du rayonnement incident global).
Ei(j) est l’efficience d’interception du jour j, qui tient compte du LAI et du coefficient d’interception des
feuilles de blé.
Ei(j) est obtenu de la façon suivante,
Ei(j) = %SVF1j (1-e(-k LAIF1
))
+
%SVF2j (e(-k LAI
F1) – e(-k(LAI
F1+ LAI
F2)))
+
%SVF3j e(-k (LAI
F1+ LAI
F2) – exp(-k(LAI
F1+ LAI
F2+ LAI
F3)))
Où Ei(j) est l’efficience d’interception pour chaque feuille i et pour le jour j.
%SVF i(j) correspond au pourcentage de surface verte issu de l’analyse d’image de la feuille i le jour j . Les
images desquelles sont issus les pourcentages de surface verte ont été prises à raison d’une fois par semaine.
Une valeur de pourcentage de surface verte a été obtenue pour chaque jour de la floraison à la maturité, grâce
à l’ajustement d’une régression non linéaire aux valeurs hebdomadaires de surface verte foliaire. Les
variantes utilisées sont les mêmes que celles utilisées pour l’obtention du HAA.
k a une valeur de 0,68 et correspond au coefficient d’interception des feuilles de blé issu du modèle CERES.
LAIFi est l’indice d’aire foliaire calculé à partir des données de surface foliaire issues de l’analyse des
images. Il est exprimé en m² par m² de sol pour chaque Fi du jour j. Ce LAI et la méthode d’obtention de
celui-ci sont les mêmes que ceux utilisés pour l’obtention du HAA.
Les mêmes variantes, les mêmes répétitions et les mêmes rendements que ceux du HAA ont été
utilisés ici.
De même que pour le HAA, deux types d’analyses ont été entrepris:
- Les analyses descriptives de la matière sèche (M.S.) totale obtenue de la floraison à la
maturité et ses relations avec les rendements.
- Les analyses des caractéristiques des courbes d’évolution du cumul de la matière
sèche de la floraison à la maturité et les relations entre celles-ci et les rendements.

77
Analyses de la matière sèche (M.S.) totale obtenue de la floraison à la maturité.
1. Une description graphique des différences de M.S. totales obtenues sur la période visée
entre les différentes répétitions de chaque variante sera exposée.
2. Une régression multiple et des analyses de variance mettent en évidence les éventuelles
différences significatives de M.S. entre les années, les variétés, les traitements, le précédent
et la date de semis.
3. Une régression multiple, et des régressions simples par la méthode des moindres carrés
entre la M.S. de la floraison à la maturité et les rendements, le poids de 1000 grains ou la
matière sèche réellement récoltée à Everlange mettront en évidence une éventuelle relation
avec les rendements obtenus.
L’analyse est effectuée sur les données des deux années, sur une année ou sur la variété
Achat seule à raison d’une valeur moyenne de M.S. par répétition pour les 4 répétitions par
variante. Une analyse sur les données de la variété Achat seule peut être entreprise car c’est
la principale variété présente. Il y a donc un nombre de données suffisamment important
pour que des analyses puissent être faites sur cette variété seule.
4. La régression par la méthode des moindres carrés entre la M.S. de la floraison à la
maturité de chaque plante et les caractéristiques de rendement de chacune des 5 plantes
par répétition suivies en 2007.
Analyses des caractéristiques des courbes d’évolution du cumul de la matière sèche.
5. Une analyse graphique de l’ajustement d’une courbe à l’évolution du cumul de la
Matière Sèche en fonction du cumul des degrés jour (dj) depuis la floraison jusqu’à la
maturité sur l’ensemble des données de 2006 et 2007, pour chaque répétition de chaque
variante suivie. La courbe obtenue est une sigmoïde d’équation MS=a/(1+exp(v*(i-dj))).
6. Une régression multiple et des analyses de variance mettront en évidence les éventuelles
différences significatives entre les paramètres des différentes courbes de cumul de la
matière sèche obtenue.
7. Une régression multiple, et des régressions linéaires simples par la méthode des
moindres carrés entre les paramètres a (plateau) ou i (point d’inflexion) de chaque courbe
obtenue et le rendement de la répétition correspondante.
Toutes les analyses statistiques ont été effectuées à l’aide des logiciels Excel et Statistica.

78
Résultats et discussion.
1.Tout comme pour le HAA, les valeurs de M.S. des variantes Achat témoin et Achat Triple
traitement, suivies à la fois en 2006 et 2007 sont supérieures en 2006 (Figure 21). On observe
également des valeurs supérieures du témoin Achat par rapport au triple traitement.
Figure 21 : Histogramme de la matière sèche totale obtenue pour chaque répétition de chaque variante en 2006 et 2007 à Everlange (Témoin Sain = triple traitement).
2. La régression multiple effectuée entre la M.S. et les différentes modalités d’année, traitement,
variété, et précédent montre un effet significatif de chaque modalité à l’exception de la variété
(Tableau 14). Le paramètre Bêta du Tableau 14 donne l’importance relative de chaque modalité.
Dans un ordre croissant, c’est l’année, le traitement et le précédent qui expliquent les variations de
la M.S. observées. L’année et le traitement sont les deux premiers facteurs expliquant le
rendement, comme pour le HAA. La M.S. se distingue toutefois du HAA, ce dernier ayant la
variété comme troisième facteur explicatif, alors que la variété n’est pas un facteur significatif pour
la M.S. Par contre, le précédent, qui est le troisième facteur explicatif des variations de la M.S.
n’est pas significatif pour le HAA. L’effet important de l’année a également été mis en évidence
par l’analyse de variance sur le témoin Achat entre les deux années (p=0,01 ; R²=69%) et sur le
triple traitement Achat entre les deux années (p=0,000041 ; R²=95%).
L’analyse de variance sur tous les effets montre un effet significatif, par ordre d’importance, de
l’année 2006 par rapport à 2007, du triple traitement Achat par rapport aux témoins, du précédent
blé deux années par rapport au précédent jachère, et de la variété Akteur par rapport aux autres
variétés.
VARIANTES
0100200300400500600700
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère blé-blé Jachère Jachère
Témoin Sain Témoin Témoin Témoin
Achat Akteur Flair
2006 2007
M. s
èche
(g
m-2 )
VARIANTES
0100200300400500600700
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère blé-blé Jachère Jachère
Témoin Sain Témoin Témoin Témoin
Achat Akteur Flair
2006 2007
M. s
èche
(g
m-2 )
79
Tableau 14 : Résultats de la régression multiple effectuée entre la M.S. et les différentes modalités présentes.
M.S.
Synthèse Régression R²= 0,87 ; p<0,000001 ; n= 28
Bêta p Année -0,613 ***
Variété n.s. n.s.
Traitement 0,422 ***
Précédent 0,260 ** Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
3. Une régression multiple entre le rendement et la matière sèche totale de la floraison à la
maturité, l’année, la variété, le traitement et le précédent montre que seuls le précédent et le
traitement expliquent les variations de rendement de façon significative (Tableau 15).
Tableau 15 : Résultats de la régression multiple effectuée entre la M.S. et les différentes modalités présentes.
Rendement
Synthèse Régression R²=0,78; p<0,000001; n= 28
Bêta p Année n.s. n.s.
Variété n.s. n.s.
Traitement -0,758 ***
Précédent 0,574 ***
M.S. n.s. n.s. Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Cette analyse montre que la M.S. n’explique pas les différents rendements obtenus. L’analyse de
régression simple entre le cumul de la M.S. de la floraison à la maturité et le rendement sur
l’ensemble des données de 2006 et 2007 montre également que la M.S. explique peu les variations
de rendement. Cette conclusion est vérifiée, d’une part lorsq’on utilise toutes les variantes pour
lesquelles on disposait d’un suivi de la surface verte (R² =18%) et d’autre part lorsqu’on utilise
uniquement la variété Achat (R² =21 %). Seule la relation entre la M.S. et le rendement par année
explique une plus grande proportion des rendements (R²=84% en 2006, et R²=39% en 2007). Une
utilisation de la variété Achat seule augmente légèrement le R² en 2007. Cependant, les résultats de
2006 sont à relativiser, car seules 8 données sont disponibles.
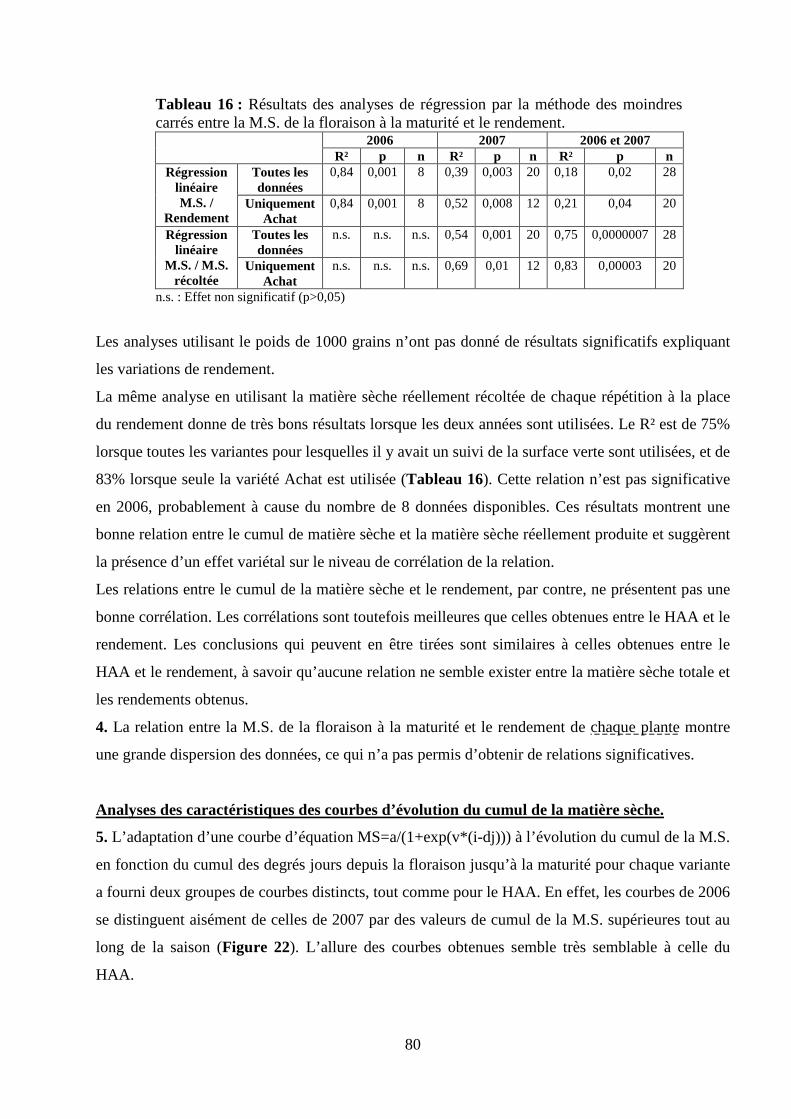
80
Tableau 16 : Résultats des analyses de régression par la méthode des moindres carrés entre la M.S. de la floraison à la maturité et le rendement.
2006 2007 2006 et 2007 R² p n R² p n R² p n
Toutes les données
0,84 0,001 8 0,39 0,003 20 0,18 0,02 28 Régression linéaire M.S. /
Rendement Uniquement
Achat 0,84 0,001 8 0,52 0,008 12 0,21 0,04 20
Toutes les données
n.s. n.s. n.s. 0,54 0,001 20 0,75 0,0000007 28 Régression linéaire
M.S. / M.S. récoltée
Uniquement Achat
n.s. n.s. n.s. 0,69 0,01 12 0,83 0,00003 20
n.s. : Effet non significatif (p>0,05)
Les analyses utilisant le poids de 1000 grains n’ont pas donné de résultats significatifs expliquant
les variations de rendement.
La même analyse en utilisant la matière sèche réellement récoltée de chaque répétition à la place
du rendement donne de très bons résultats lorsque les deux années sont utilisées. Le R² est de 75%
lorsque toutes les variantes pour lesquelles il y avait un suivi de la surface verte sont utilisées, et de
83% lorsque seule la variété Achat est utilisée (Tableau 16). Cette relation n’est pas significative
en 2006, probablement à cause du nombre de 8 données disponibles. Ces résultats montrent une
bonne relation entre le cumul de matière sèche et la matière sèche réellement produite et suggèrent
la présence d’un effet variétal sur le niveau de corrélation de la relation.
Les relations entre le cumul de la matière sèche et le rendement, par contre, ne présentent pas une
bonne corrélation. Les corrélations sont toutefois meilleures que celles obtenues entre le HAA et le
rendement. Les conclusions qui peuvent en être tirées sont similaires à celles obtenues entre le
HAA et le rendement, à savoir qu’aucune relation ne semble exister entre la matière sèche totale et
les rendements obtenus.
4. La relation entre la M.S. de la floraison à la maturité et le rendement de chaque plante montre
une grande dispersion des données, ce qui n’a pas permis d’obtenir de relations significatives.
Analyses des caractéristiques des courbes d’évolution du cumul de la matière sèche.
5. L’adaptation d’une courbe d’équation MS=a/(1+exp(v*(i-dj))) à l’évolution du cumul de la M.S.
en fonction du cumul des degrés jours depuis la floraison jusqu’à la maturité pour chaque variante
a fourni deux groupes de courbes distincts, tout comme pour le HAA. En effet, les courbes de 2006
se distinguent aisément de celles de 2007 par des valeurs de cumul de la M.S. supérieures tout au
long de la saison (Figure 22). L’allure des courbes obtenues semble très semblable à celle du
HAA.
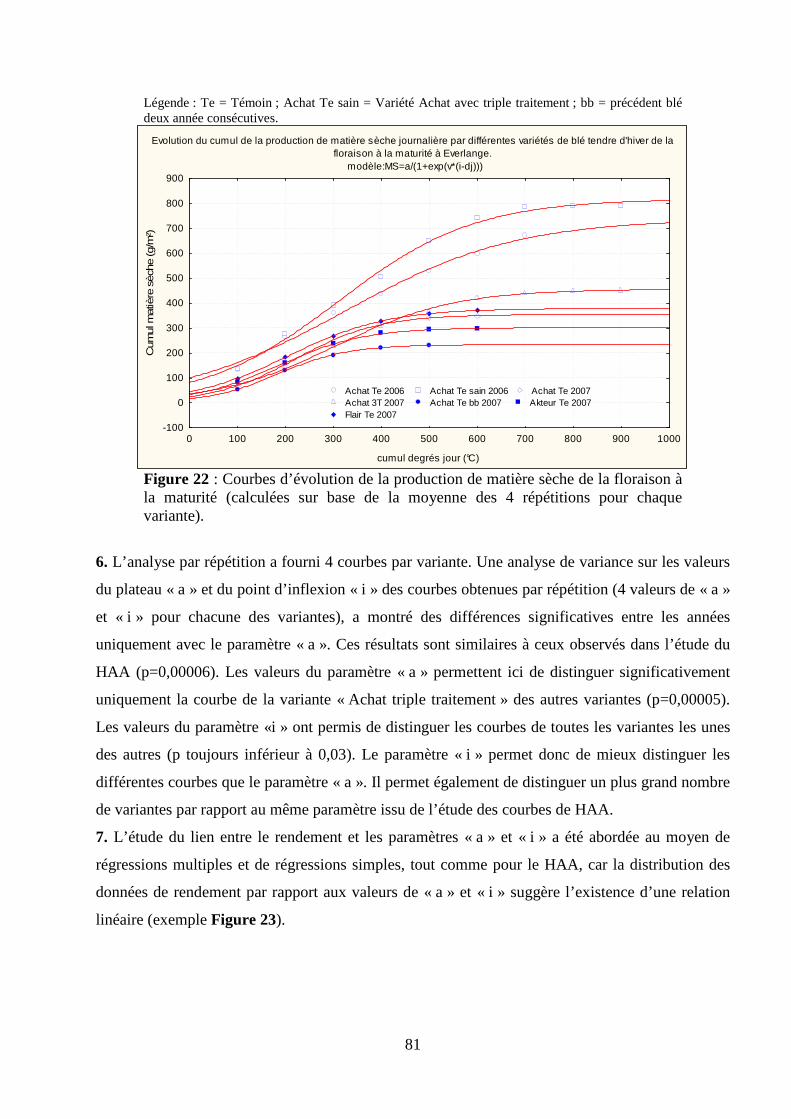
81
Légende : Te = Témoin ; Achat Te sain = Variété Achat avec triple traitement ; bb = précédent blé deux année consécutives.
Evolution du cumul de la production de matière sèche journalière par différentes variétés de blé tendre d'hiver de lafloraison à la maturité à Everlange.
modèle:MS=a/(1+exp(v*(i-dj)))
0 100 200 300 400 500 600 700 800 900 1000
cumul degrés jour (°C)
-100
0
100
200
300
400
500
600
700
800
900
Cum
ul m
atiè
re s
èche
(g/m
²)
Achat Te 2006 Achat Te sain 2006 Achat Te 2007Achat 3T 2007 Achat Te bb 2007 Akteur Te 2007Flair Te 2007
Figure 22 : Courbes d’évolution de la production de matière sèche de la floraison à la maturité (calculées sur base de la moyenne des 4 répétitions pour chaque variante).
6. L’analyse par répétition a fourni 4 courbes par variante. Une analyse de variance sur les valeurs
du plateau « a » et du point d’inflexion « i » des courbes obtenues par répétition (4 valeurs de « a »
et « i » pour chacune des variantes), a montré des différences significatives entre les années
uniquement avec le paramètre « a ». Ces résultats sont similaires à ceux observés dans l’étude du
HAA (p=0,00006). Les valeurs du paramètre « a » permettent ici de distinguer significativement
uniquement la courbe de la variante « Achat triple traitement » des autres variantes (p=0,00005).
Les valeurs du paramètre «i » ont permis de distinguer les courbes de toutes les variantes les unes
des autres (p toujours inférieur à 0,03). Le paramètre « i » permet donc de mieux distinguer les
différentes courbes que le paramètre « a ». Il permet également de distinguer un plus grand nombre
de variantes par rapport au même paramètre issu de l’étude des courbes de HAA.
7. L’étude du lien entre le rendement et les paramètres « a » et « i » a été abordée au moyen de
régressions multiples et de régressions simples, tout comme pour le HAA, car la distribution des
données de rendement par rapport aux valeurs de « a » et « i » suggère l’existence d’une relation
linéaire (exemple Figure 23).

82
Régress ion par la méthode des moindres carrés entre le point d'inflexion i de la courbe d'évolution de la production dematiè re sè che et l e re ndeme nt en 20 06 et 20 07.
16 0 180 200 22 0 240 260 28 0 300 320 34 0
i
6
7
8
9
10
11
12
13R
en
de
me
nt
(T/H
a)
i:Rendement (T/Ha): r² = 0,5700; r = 0,7550; p = 0,000003; y = 5,0965 + 0,0166*x
Figure 23 : Régression par la méthode des moindres carrés entre le point d’inflexion i de la courbe d’évolution de la production de matière sèche et le rendement en 2006 et 2007.
Une régression multiple entre le rendement et les différentes variables explicatives de l’allure des
courbes de cumul de la matière sèche montre que le précédent cultural et les deux paramètres « a »
et « i » expliquent les variations de rendement (Tableau 17). Le paramètre « i » a le poids le plus
important (Bêta le plus élevé). Cette régression met en évidence le caractère explicatif élevé de ces
deux paramètres. Les régressions simples entre chacun des paramètres « a » et « i » et les
rendements devraient donc fournir une relation significative et explicative d’une bonne proportion
des rendements.
La régression entre le paramètre i (point d’inflexion) de la courbe d’évolution de la production de
matière sèche de la floraison à la maturité en fonction des degrés jour ( 0°C jour à la Floraison) et
les rendements sur l’ensemble des données de 2006 et 2007 est significative et explique plus de
50% des rendements observés (Tableau 18). La relation est encore améliorée (R² plus élevé)
lorsque seules les données de la variété Achat sont utilisées. Cela suggère un effet variétal. Une
amélioration est également observée lorsque l’utilisation des données est effectuée par année (2006
ou 2007 seules), ce qui suggère un effet de l’année.
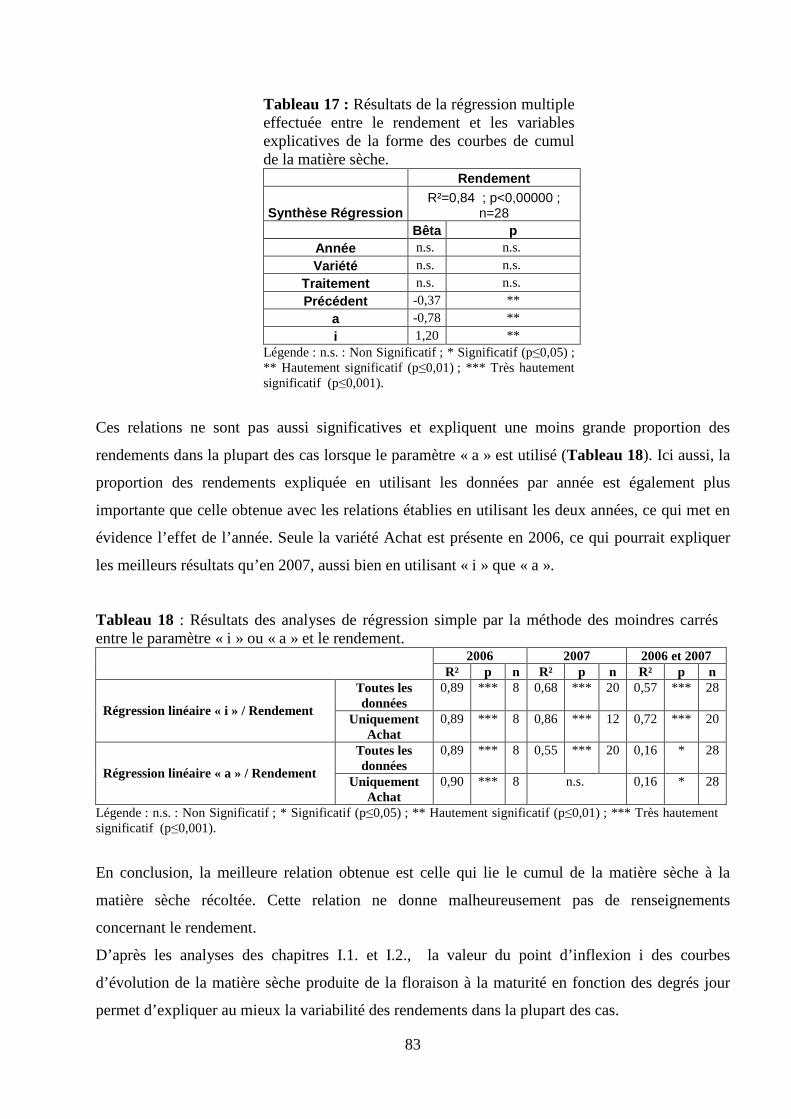
83
Tableau 17 : Résultats de la régression multiple effectuée entre le rendement et les variables explicatives de la forme des courbes de cumul de la matière sèche.
Rendement
Synthèse Régression R²=0,84 ; p<0,00000 ;
n=28 Bêta p
Année n.s. n.s.
Variété n.s. n.s.
Traitement n.s. n.s.
Précédent -0,37 **
a -0,78 **
i 1,20 ** Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Ces relations ne sont pas aussi significatives et expliquent une moins grande proportion des
rendements dans la plupart des cas lorsque le paramètre « a » est utilisé (Tableau 18). Ici aussi, la
proportion des rendements expliquée en utilisant les données par année est également plus
importante que celle obtenue avec les relations établies en utilisant les deux années, ce qui met en
évidence l’effet de l’année. Seule la variété Achat est présente en 2006, ce qui pourrait expliquer
les meilleurs résultats qu’en 2007, aussi bien en utilisant « i » que « a ».
Tableau 18 : Résultats des analyses de régression simple par la méthode des moindres carrés entre le paramètre « i » ou « a » et le rendement.
2006 2007 2006 et 2007 R² p n R² p n R² p n
Toutes les données
0,89 *** 8 0,68 *** 20 0,57 *** 28
Régression linéaire « i » / Rendement Uniquement
Achat 0,89 *** 8 0,86 *** 12 0,72 *** 20
Toutes les données
0,89 *** 8 0,55 *** 20 0,16 * 28
Régression linéaire « a » / Rendement Uniquement
Achat 0,90 *** 8 n.s. 0,16 * 28
Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
En conclusion, la meilleure relation obtenue est celle qui lie le cumul de la matière sèche à la
matière sèche récoltée. Cette relation ne donne malheureusement pas de renseignements
concernant le rendement.
D’après les analyses des chapitres I.1. et I.2., la valeur du point d’inflexion i des courbes
d’évolution de la matière sèche produite de la floraison à la maturité en fonction des degrés jour
permet d’expliquer au mieux la variabilité des rendements dans la plupart des cas.

84
La régression par la méthode des moindres carrés entre ce paramètre i et les rendements (mais
cette fois de la courbe d’évolution du cumul du HAA) est globalement moins bonne, en particulier
lorsque l’ensemble des données obtenues en 2006 et 2007 est utilisé (I.1).
Ces relations peuvent être améliorées et simplifiées de différentes façons. La stabilité de celles-ci
d’une année à l’autre ou d’une variété à l’autre doit également être vérifiée en utilisant plus de
données (cf. Chapitre II de cette partie).

85
CHAPITRE II : Simplification et amélioration d’un modèle d’estimation des rendements du blé d’hiver au Grand Duché de Luxembourg. Ce deuxième chapitre est dédié à la mise en œuvre d’une démarche de simplification de modèles
précédemment testés ainsi que l’amélioration du modèle fournissant les meilleurs résultats.
La simplification consistera à supprimer successivement un ou deux étages foliaires pour la
construction des modèles. Cette simplification est motivée par une étude de Gooding et al. (2000).
Cette étude a mis en évidence une relation entre la dégradation de la surface verte de la dernière
feuille émergée et le rendement. L’utilisation des modèles qui prennent en compte uniquement la
F1, ou uniquement la F1 et la F2, rendra ceux-ci beaucoup plus simples d’utilisation puisque
l’obtention du pourcentage de surface verte et des surfaces foliaires est la donnée nécessitant le
plus de temps à obtenir.
La démarche d’amélioration consistera à ajouter des données issues d’expérimentations antérieures
à cette thèse pour la construction du modèle qui aura fourni les meilleurs résultats aux étapes
précédentes. Ces données n’ont pas été utilisées directement au CHAPITRE I car elles
comportent des lacunes. Ces lacunes nous ont amenés à émettre des hypothèses pour pouvoir
utiliser ces données, ce qui a pu introduire des biais. D’autre part, la méthode d’estimation visuelle
du %SV utilisée pour l’obtention de ces données introduit probablement des biais à son tour. Ces
derniers seront d’ailleurs estimés par la comparaison des résultats obtenus entre l’utilisation du
modèle retenu avec les estimations visuelles ou avec les estimations par l’analyse d’images en
2006 et 2007 au Chapitre II.3.
II.1. Simplification du modèle du calcul du HAA. La simplification du modèle mettant en relation le HAA de la floraison à la maturité et les
rendements en 2006-2007 par suppression successive de la F3 et de la F2 améliore légèrement la
corrélation de la relation dans les deux cas où cette relation montrait déjà des R² supérieurs à 50%
(Tableau 19).
En supprimant la F3, puis la F3 et la F2 dans nos calculs, on passe d’un R² de 83 % à un R² de
84% puis 87% pour la relation entre le HAA et le rendement en 2006, ainsi qu’un passage de 60%
à 62% si le poids de 1000 grains est utilisé à la place du rendement en grains pour cette relation.
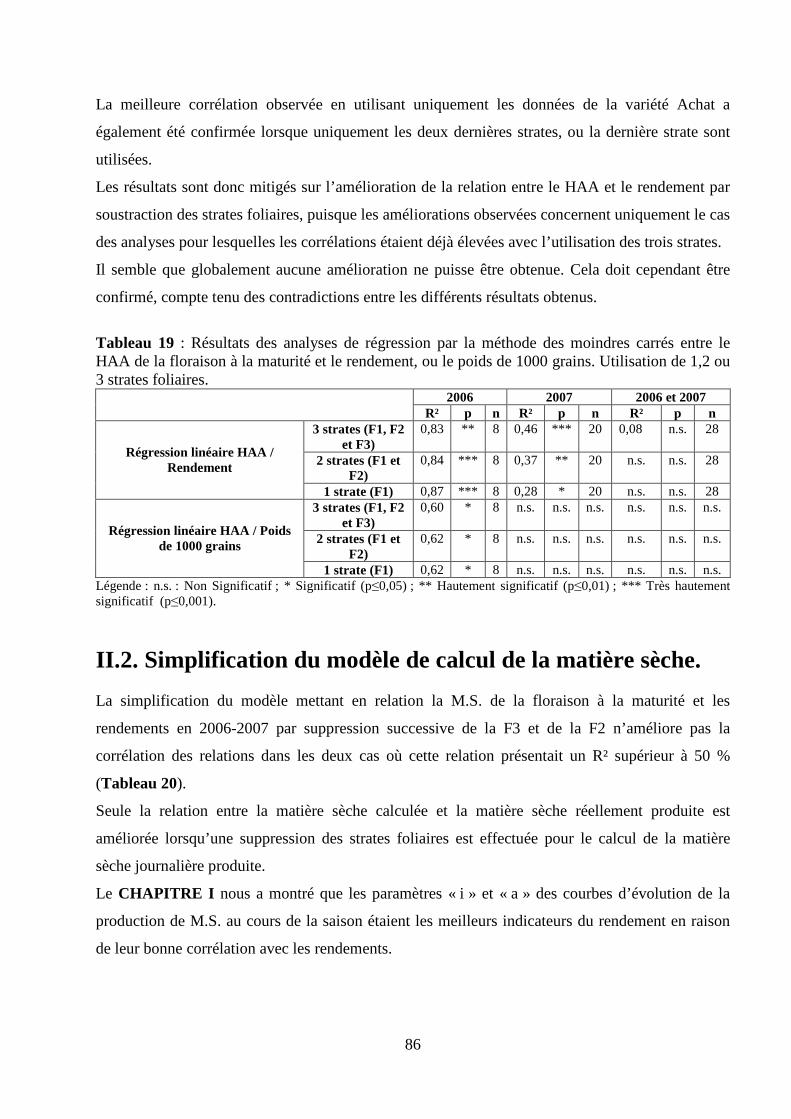
86
La meilleure corrélation observée en utilisant uniquement les données de la variété Achat a
également été confirmée lorsque uniquement les deux dernières strates, ou la dernière strate sont
utilisées.
Les résultats sont donc mitigés sur l’amélioration de la relation entre le HAA et le rendement par
soustraction des strates foliaires, puisque les améliorations observées concernent uniquement le cas
des analyses pour lesquelles les corrélations étaient déjà élevées avec l’utilisation des trois strates.
Il semble que globalement aucune amélioration ne puisse être obtenue. Cela doit cependant être
confirmé, compte tenu des contradictions entre les différents résultats obtenus.
Tableau 19 : Résultats des analyses de régression par la méthode des moindres carrés entre le HAA de la floraison à la maturité et le rendement, ou le poids de 1000 grains. Utilisation de 1,2 ou 3 strates foliaires.
2006 2007 2006 et 2007 R² p n R² p n R² p n
3 strates (F1, F2 et F3)
0,83 ** 8 0,46 *** 20 0,08 n.s. 28
2 strates (F1 et F2)
0,84 *** 8 0,37 ** 20 n.s. n.s. 28 Régression linéaire HAA /
Rendement
1 strate (F1) 0,87 *** 8 0,28 * 20 n.s. n.s. 28 3 strates (F1, F2
et F3) 0,60 * 8 n.s. n.s. n.s. n.s. n.s. n.s.
2 strates (F1 et F2)
0,62 * 8 n.s. n.s. n.s. n.s. n.s. n.s. Régression linéaire HAA / Poids
de 1000 grains
1 strate (F1) 0,62 * 8 n.s. n.s. n.s. n.s. n.s. n.s. Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
II.2. Simplification du modèle de calcul de la matière sèche. La simplification du modèle mettant en relation la M.S. de la floraison à la maturité et les
rendements en 2006-2007 par suppression successive de la F3 et de la F2 n’améliore pas la
corrélation des relations dans les deux cas où cette relation présentait un R² supérieur à 50 %
(Tableau 20).
Seule la relation entre la matière sèche calculée et la matière sèche réellement produite est
améliorée lorsqu’une suppression des strates foliaires est effectuée pour le calcul de la matière
sèche journalière produite.
Le CHAPITRE I nous a montré que les paramètres « i » et « a » des courbes d’évolution de la
production de M.S. au cours de la saison étaient les meilleurs indicateurs du rendement en raison
de leur bonne corrélation avec les rendements.

87
Tableau 20 : Résultats des analyses de régression par la méthode des moindres carrés entre la matière sèche de la floraison à la maturité et le rendement ou la matière sèche récoltée. Utilisation de 1,2 ou 3 strates foliaires.
2006 2007 2006 et 2007 R² p n R² p n R² p n
3 strates (F1, F2 et F3) 0,84 *** 8 0,39 ** 20 0,18 * 28
2 strates (F1 et F2) 0,87 *** 8 0,35 ** 20 n.s. n.s. n.s. Régression linéaire M.S. / Rendement
1 strate (F1) 0,60 * 8 0,28 * 20 n.s. n.s. n.s.
3 strates (F1, F2 et F3) n.s. n.s. n.s. 0,54 *** 20 0,75 *** 28
2 strates (F1 et F2) n.s. n.s. n.s. 0,45 ** 20 0,87 *** 28 Régression linéaire M.S. / M.S. récoltée
1 strate (F1) n.s. n.s. n.s. 0,64 *** 20 0,93 *** 28
Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001). L’amélioration de la relation entre les paramètres « i » ou « a » et le rendement par suppression
des strates foliaires ne sera pas testée car l’effet de la suppression des strates foliaires sur la
production de M.S. journalière et sa relation au rendement montre qu’aucune conclusion
d’amélioration ne peut être déduite. Une meilleure corrélation doit être observée tout d’abord sur la
relation entre la production de matière sèche et le rendement pour que l’amélioration entre le
paramètre i et le rendement soit envisageable. La relation entre le rendement et le cumul de la
production journalière de matière sèche calculée avec la F1 seule pourrait être mieux corrélée si le
pourcentage auquel les F2 et F3 sont affectées par la maladie était pris en compte. Cette
interprétation se base sur l’effet de compensation de la F1, qui varie probablement à mesure que la
photosynthèse de la F2 et la F3 est affectée (Gooding et al., 2000).
II.3. Analyse des biais introduits par l’utilisation des estimations visuelles du pourcentage de surface verte. La proximité entre les estimations visuelles du pourcentage de surface verte et les estimations par
analyse d’images a été abordée au chapitre I.2. de la PARTIE I. A l’issue de ce chapitre, il a été
constaté une surestimation du pourcentage de surface verte foliaire par la méthode d’estimation
visuelle, pour les pourcentages inférieurs à 70%. D’autre part, une analyse des moyennes par
classe de pourcentage a mis en évidence une différence très faible entre la méthode visuelle et la
méthode par l’analyse d’images.
Au cours de ce chapitre-ci, l’impact des surestimations et des biais introduits par l’utilisation de la
méthode d’estimation visuelle sur l’estimation des rendements va être analysé. Pour ce faire, les
résultats issus de l’utilisation du modèle de calcul de la matière sèche avec les pourcentages de
surfaces vertes issus des deux méthodes d’estimation vont être comparés. Le modèle de calcul de

88
la matière sèche a été choisi pour cette comparaison car il est très proche du modèle du HAA et
présente de meilleurs résultats.
Matériel et méthodes.
Les estimations visuelles et les estimations par l’analyse d’images du pourcentage de surface verte
foliaire (%SV) ont été obtenues suivant les méthodes décrites au Chapitre I.1. et I.2 de la
PARTIE I . Les données de %SV par répétition, pour toutes les variantes suivies en 2006 et 2007
par la prise et l’analyse d’images sont utilisées, ainsi que les estimations visuelles correspondantes.
Celles-ci sont présentées au Chapitre I .
La matière sèche journalière produite de la floraison à la maturité (stade GS77) a été calculée
suivant la méthode présentée au Chapitre I.2., avec les %SV issus des deux méthodes
d’estimation. Le calcul a été effectué par répétition, avec 5 plantes par répétition lorsque
l’estimation du %SV est obtenue par l’analyse d’images et 10 plantes par répétition lorsque
l’estimation du %SV est obtenue par la méthode d’estimation visuelle.
Le cumul de la matière sèche journalière de la floraison à la maturité, obtenu en 2006 et 2007 avec
les %SV issus des deux méthodes d’estimation ont été comparés. Les paramètres « a » (plateau) et
« i » (point d’inflexion) des courbes d’évolution du cumul de la matière sèche produite obtenues
avec les %SV issus des deux méthodes d’estimation ont également été comparés, ainsi que la
linéarité de la relation entre ces paramètres et le rendement. Le paramètre « v » n’a pas été utilisé
car il n’explique pas les variations du rendement (Chapitre I.2.).
Résultats et discussion.
Le cumul de la production journalière de matière sèche suit une courbe sigmoïde d’équation
MS=a/(1+exp(v*(i-dj))), où MS est la matière sèche, « a », « v » et « i » sont des paramètres, et
« dj » le cumul des degrés jours. Une illustration de ces courbes obtenues avec l’utilisation de
%SV issus de l’analyse d’images est présentée au Chapitre I.2.
Le cumul de la matière sèche journalière, calculé de la floraison à la maturité avec un %SV obtenu
selon les deux méthodes pour les 4 répétitions de chaque variante est illustré par la Figure 24. Les
valeurs obtenues varient de 204 g/m² à 602 g/m² selon les répétitions, avec des différences entre les
deux méthodes allant jusqu’à 76 g/m². En conséquence, le cumul de la matière sèche obtenu avec
les estimations visuelles du %SV montre des différences de 0 à 23% par rapport au cumul de la
matière sèche obtenu avec les estimations du %SV par l’analyse d’images.
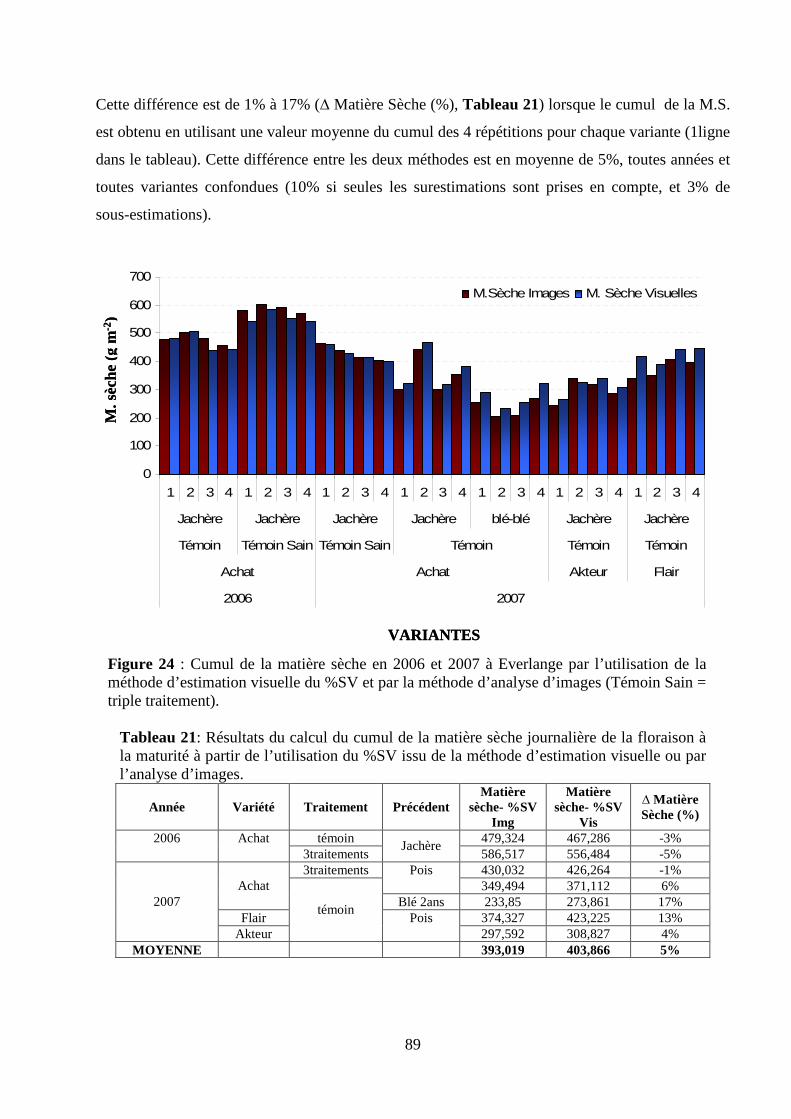
89
Cette différence est de 1% à 17% (∆ Matière Sèche (%), Tableau 21) lorsque le cumul de la M.S.
est obtenu en utilisant une valeur moyenne du cumul des 4 répétitions pour chaque variante (1ligne
dans le tableau). Cette différence entre les deux méthodes est en moyenne de 5%, toutes années et
toutes variantes confondues (10% si seules les surestimations sont prises en compte, et 3% de
sous-estimations).
Figure 24 : Cumul de la matière sèche en 2006 et 2007 à Everlange par l’utilisation de la méthode d’estimation visuelle du %SV et par la méthode d’analyse d’images (Témoin Sain = triple traitement).
Tableau 21: Résultats du calcul du cumul de la matière sèche journalière de la floraison à la maturité à partir de l’utilisation du %SV issu de la méthode d’estimation visuelle ou par l’analyse d’images.
Année Variété Traitement Précédent Matière
sèche- %SV Img
Matière sèche- %SV
Vis
∆ Matière Sèche (%)
témoin 479,324 467,286 -3% 2006
Achat 3traitements
Jachère 586,517 556,484 -5%
3traitements 430,032 426,264 -1% Pois 349,494 371,112 6% Achat
Blé 2ans 233,85 273,861 17% Flair 374,327 423,225 13%
2007
Akteur
témoin Pois
297,592 308,827 4% MOYENNE 393,019 403,866 5%
VARIANTES
M. s
èche
(g
m-2)
0
100
200
300
400
500
600
700
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère Jachère Jachère blé-blé Jachère Jachère
Témoin Témoin Sain Témoin Sain Témoin Témoin Témoin
Achat Achat Akteur Flair
2006 2007
M.Sèche Images M. Sèche Visuelles
VARIANTES
M. s
èche
(g
m-2)
0
100
200
300
400
500
600
700
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère Jachère Jachère blé-blé Jachère Jachère
Témoin Témoin Sain Témoin Sain Témoin Témoin Témoin
Achat Achat Akteur Flair
2006 2007
M.Sèche Images M. Sèche Visuelles

90
Les paramètres « a » (plateau) et « i » (point d’inflexion) des courbes d’évolution du cumul de la
matière sèche varient du simple au quadruple et du simple au double, selon les répétitions, avec
des différences importantes entre les deux méthodes (Figure 25). Le paramètre « a » obtenu avec
les estimations visuelles du %SV montre des différences de 1% à 29% par rapport au paramètre
« a » obtenu avec les estimations du %SV par l’analyse d’images. Le paramètre « i » montre lui
des différences de 1% à 25%. Cette différence est de 2% à 19% pour « a » et de 2% à 18% pour
« i » (Tableau 22) lorsque ces paramètres sont obtenus en utilisant une valeur moyenne du cumul
des 4 répétitions pour chaque variante (1ligne dans le tableau). La moyenne de cette différence est
de 8%, aussi bien pour le paramètre « a » que pour le paramètre « i ».
Figure 25 : Variations des valeurs des paramètres a et i de la courbe de cumul de la matière sèche en 2006 et 2007 par variante à Everlange selon deux méthodes d’estimations du pourcentage de surface verte (Témoin Sain = triple traitement).
0
100
200
300
400
500
600
700
800
900
1000
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère Jachère Jachère blé-blé Jachère Jachère
Témoin Témoin Sain Témoin Sain Témoin Témoin Témoin
Achat Achat Akteur Flair
2006 2007
Plateau "a" images Plateau "a" visuelles
Pt inflexion "i" images Pt inflexion "i" visuelles
VARIANTES
I ou
a
0
100
200
300
400
500
600
700
800
900
1000
1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4 1 2 3 4
Jachère Jachère Jachère Jachère blé-blé Jachère Jachère
Témoin Témoin Sain Témoin Sain Témoin Témoin Témoin
Achat Achat Akteur Flair
2006 2007
Plateau "a" images Plateau "a" visuelles
Pt inflexion "i" images Pt inflexion "i" visuelles
VARIANTES
I ou
a

91
Tableau 22 : Résultats du calcul du cumul de la matière sèche journalière de la floraison à la maturité à partir de l’utilisation du %SV issu de la méthode d’estimations visuelles ou par l’analyse d’images.
La linéarité de la relation entre le rendement de chaque répétition et le paramètre « a » ou le
paramètre « i » sur l’ensemble des données de 2006 et 2007 est meilleure lorsqu’elle est testée
entre le rendement et le paramètre « a » ou « i » issu des courbes de cumul obtenues à partir du
%SV estimé par l’analyse d’images. La régression par la méthode des moindres carrés obtenue
avec les %SV issus des images présente une significativité et un coefficient de détermination plus
élevés, qu’elle soit effectuée entre « i » et le rendement, ou entre « a » et le rendement (Tableau
23). Les résultats nous amènent à la même conclusion lorsque seules les données de l’année 2007
sont analysées. La différence de R² entre les deux méthodes dépasse 20% dans 3 cas sur 4 si on ne
considère pas les régressions établies avec les données de 2006 seules (ces dernières sont établies à
partir de 8 données). Ceci indique que l’utilisation d’un modèle linéaire simple entre le rendement
et un paramètre de la courbe du cumul de la matière sèche journalière pour faire une estimation des
rendements fournira ~20% d’erreurs d’estimation supplémentaires lorsque le modèle aura été
obtenu à partir d’estimations visuelles du %SV.
Tableau 23 : Résultats des analyses de régression simple par la méthode des moindres carrés entre le paramètre « i » ou « a » et le rendement. Comparaison des résultats obtenus selon la méthode d’obtention du %SV par l’analyse d’images à la méthode d’obtention du %SV par l’estimation visuelle.
2006 2007 2006 et 2007 R² p n R² p n R² p n
Images 0,89 *** 8 0,68 *** 20 0,57 *** 28 Régression linéaire « i » / Rendement
Visuelles 0,91 *** 8 0,46 *** 20 0,35 *** 28
Images 0,89 *** 8 0,55 *** 20 0,16 * 28 Régression linéaire « a » / Rendement
Visuelles 0,92 *** 8 0,31 ** 20 n.s. n.s. n.s. Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Année Variété Traitement Précé-dent
a %SV visuel
i %SV visuel
a %SV image
i %SV image
∆a (%)
∆i (%)
Achat témoin Jachère 566,730 242,625 553,441 224,220 2% 8% 2006
Achat 3traitements Jachère 846,496 340,188 818,822 311,71 3% 9% Achat témoin Pois 376,995 236,774 354,723 222,325 6% 6% Achat 3traitements Pois 447,098 296,886 456,662 304,099 -2% -2% Flair témoin Pois 451,762 235,878 379,263 206,940 19% 14%
Akteur témoin Pois 323,391 188,740 302,275 186,528 7% 1% 2007
Achat témoin Blé 2ans 277,266 219,148 233,677 185,729 19% 18%
Moyenne 469,963 251,463 442,695 234,508 8% 8%

92
La comparaison de la matière sèche obtenue avec les deux méthodes différentes, sur une étude de
deux années, 7 variantes en 2007 et 2 en 2006, conduit aux conclusions suivantes :
- En moyenne, une surestimation de l’estimation de la matière sèche produite et des paramètres
«a » et « i » de la courbe d’évolution du cumul de la matière sèche aux alentours de ~ 10%
(moyenne obtenue sans tenir compte des quelques sous-estimations) a été mise en évidence
lorsque les %SV issus de l’estimation visuelle sont utilisés. Cette surestimation est
probablement une conséquence de la tendance à la surestimation du %SV par l’utilisation de
la méthode d’estimation visuelle. Surestimation mise en évidence au Chapitre I.2. de la
PARTIE I .
- L’utilisation, pour les calculs de la matière sèche journalière, de moyennes par strate foliaire
par répétition (1parcelle=1répétition), puis de moyennes de 4 répétitions a montré une
diminution des écarts maximaux. Ces écarts maximaux restent cependant élevés (de ~20%
pour le cumul de la matière sèche et pour les paramètres i et a).
- Un modèle simple d’estimation des rendements de 2006 et 2007, basé sur une régression
linéaire des moindres carrés entre le paramètre i ou a et le rendement explique un plus grand
pourcentage des rendements lorsqu’il se base sur l’utilisation d’un %SV issu des estimations
par l’analyse d’images. En moyenne, l’utilisation du %SV issu de l’analyse d’images permet
d’expliquer ~20% en plus de la variation des rendements obtenus par rapport à l’utilisation
du %SV issu des estimations visuelles.
- Les modèles issus de l’utilisation des estimations du %SV par l’analyse d’images montrent
une significativité plus élevée que ceux issus de l’utilisation des estimations du %SV de
l’estimation visuelle.

93
II.4. Amélioration de l’application du modèle de la matière sèche à la culture du blé d’hiver sur le site expérimental d’Everlange. Dans ce chapitre, la possibilité d’améliorer le modèle ayant fourni les meilleurs résultats avec les
données de 2006 et 2007 sera testée en ajoutant des données issues des années 2000 à 2005. Ce
modèle met en relation le point d’inflexion i des courbes d’évolution de la matière sèche produite
de la floraison à la maturité en fonction des degrés jour et le rendement.
Un plus grand nombre de données disponibles permettra aussi de tester l’influence des variables
année, traitement, variété, précédent, date de semis et de la valeur « a » du plateau de la courbe
d’évolution de la production de matière sèche.
Les données ajoutées n’ont pas été obtenues suivant les mêmes méthodes que celles utilisées en
2006 et 2007. Des adaptations ont été effectuées et certaines hypothèses ont dû être émises pour
que ces données puissent être introduites. Il se peut que les différentes méthodes utilisées et les
hypothèses émises introduisent des biais.
Pour cette raison, l’utilisation de ces données n’a pas été effectuée directement dans l’étape de
validation au CHAPITRE I. L’ajout de ces données doit confirmer tout en améliorant les résultats
obtenus avec le modèle qui a montré les meilleures corrélations avec le rendement. Cependant, il
se pourrait qu’il n’existe pas de confirmation ou d’amélioration des relations précédemment
obtenues si les biais introduits sont trop importants.
Matériel et méthodes.
Les données ajoutées sont issues du projet Sintama et du travail effectué par Moussa El Jarroudi
durant sa thèse (El Jarroudi, 2005). Seul le site expérimental d’Everlange est concerné.
Les hypothèses suivantes ont été émises afin de pouvoir utiliser ces données dans notre modèle :
• Les données des années 2000 à 2005 se composent uniquement des estimations visuelles du
pourcentage de surface foliaire couverte par les symptômes des maladies présentes durant ces
années.
Afin d’obtenir le pourcentage de surface verte, le pourcentage de surface foliaire présentant
les symptômes de la septoriose et de la rouille brune a été soustrait à 100% de surface

94
foliaire. L’hypothèse a été émise que le pourcentage de surface verte obtenu de cette façon
est, en moyenne, similaire à celui obtenu par l’analyse d’images en 2006 et 2007.
La surface sénescente induite par la septoriose a été considérée comme surface présentant des
symptômes dans l’estimation du pourcentage de surface foliaire couverte par ceux-ci.
D’autre part, il a été considéré que le pourcentage maximum de surface foliaire couverte par
les symptômes de la rouille brune ne peut excéder les 66% (Robert et al., 2005 ;
Communication téléphonique M.O.Bancal, 2007). L’estimation visuelle de ce pourcentage a
été effectuée en tenant compte du halo qui peut être observé autour des pustules de rouille
brune et un recalage des estimations obtenues a été effectué sur base d’un maximum de 66%
de surface foliaire couverte par les symptômes de la rouille brune.
• Ne disposant pas de données concernant la surface foliaire et le nombre d’épis par m²,
l’hypothèse a été émise que ceux-ci ne varient pas d’une année à l’autre pour une même
variété. Ces données sont nécessaires au calcul du LAI (voir I.1.).
Un LAI des 3 dernières strates ainsi qu’un pourcentage de participation de chaque strate
constant pour chaque variété ont été utilisés. Ceux-ci ont été calculés sur base d’une
moyenne obtenue en 2006 et 2007 pour chaque variété suivie ces années-là.
• Les données de rayonnement de la station météorologique du réseau ASTA (Administration
des Services Techniques de l’Agriculture du Grand Duché de Luxembourg) la plus proche de
notre site d’essai ne sont pas disponibles pour les années 2000 à 2002. Pour ces années, les
données de rayonnement d’une maille du réseau européen CGMS (Crop Growth Monitoring
System) ont été utilisées. La maille qui a été choisie est celle qui a présenté les données les
mieux corrélées avec les données de la station de L’ASTA la plus proche de Everlange pour
les années 2006 et 2007.
Les données utilisées sont issues des variétés Achat, Flair et Akteur de l’année 2000 à l’année
2007, suivant les modalités indiquées au Tableau 24.
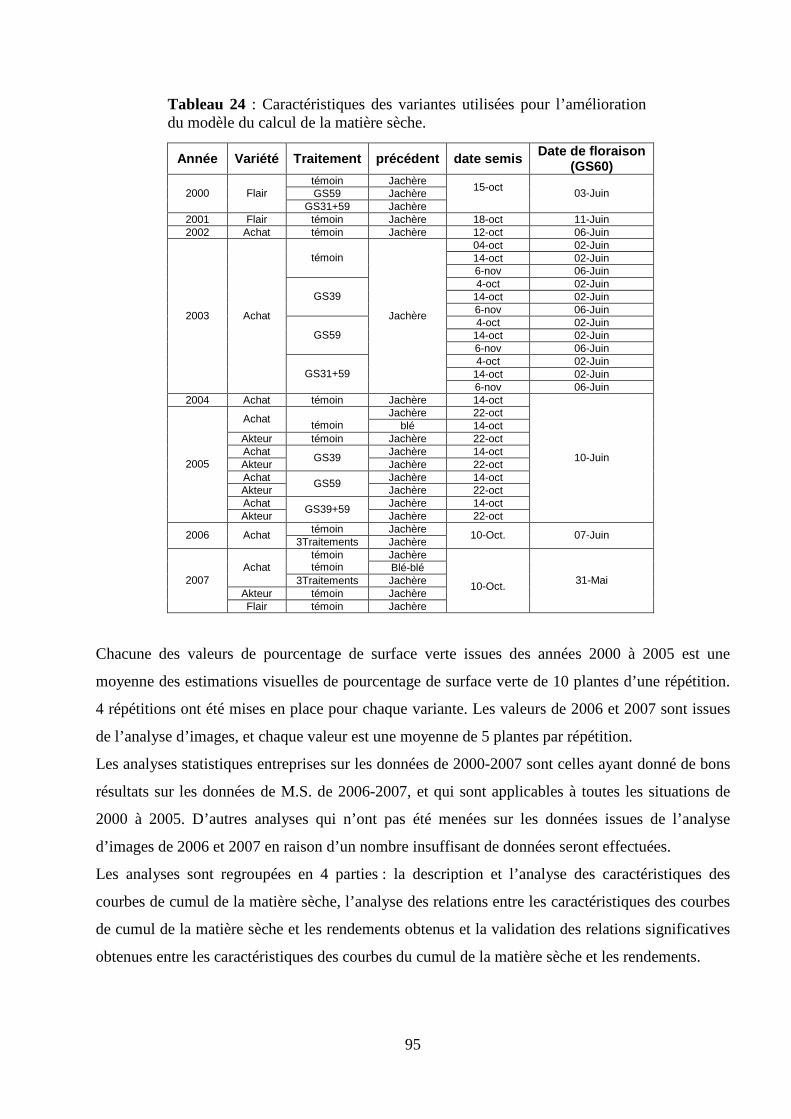
95
Tableau 24 : Caractéristiques des variantes utilisées pour l’amélioration du modèle du calcul de la matière sèche.
Chacune des valeurs de pourcentage de surface verte issues des années 2000 à 2005 est une
moyenne des estimations visuelles de pourcentage de surface verte de 10 plantes d’une répétition.
4 répétitions ont été mises en place pour chaque variante. Les valeurs de 2006 et 2007 sont issues
de l’analyse d’images, et chaque valeur est une moyenne de 5 plantes par répétition.
Les analyses statistiques entreprises sur les données de 2000-2007 sont celles ayant donné de bons
résultats sur les données de M.S. de 2006-2007, et qui sont applicables à toutes les situations de
2000 à 2005. D’autres analyses qui n’ont pas été menées sur les données issues de l’analyse
d’images de 2006 et 2007 en raison d’un nombre insuffisant de données seront effectuées.
Les analyses sont regroupées en 4 parties : la description et l’analyse des caractéristiques des
courbes de cumul de la matière sèche, l’analyse des relations entre les caractéristiques des courbes
de cumul de la matière sèche et les rendements obtenus et la validation des relations significatives
obtenues entre les caractéristiques des courbes du cumul de la matière sèche et les rendements.
Année Variété Traitement précédent date semis Date de floraison (GS60)
témoin Jachère GS59 Jachère 2000 Flair
GS31+59 Jachère
15-oct
03-Juin
2001 Flair témoin Jachère 18-oct 11-Juin 2002 Achat témoin Jachère 12-oct 06-Juin
04-oct 02-Juin 14-oct 02-Juin témoin 6-nov 06-Juin 4-oct 02-Juin
14-oct 02-Juin GS39 6-nov 06-Juin 4-oct 02-Juin
14-oct 02-Juin GS59 6-nov 06-Juin 4-oct 02-Juin
14-oct 02-Juin
2003 Achat
GS31+59
Jachère
6-nov 06-Juin 2004 Achat témoin Jachère 14-oct
Jachère 22-oct Achat témoin blé 14-oct
Akteur témoin Jachère 22-oct Achat Jachère 14-oct Akteur
GS39 Jachère 22-oct
Achat Jachère 14-oct Akteur
GS59 Jachère 22-oct
Achat Jachère 14-oct
2005
Akteur GS39+59
Jachère 22-oct
10-Juin
témoin Jachère 2006 Achat 3Traitements Jachère
10-Oct. 07-Juin
Jachère témoin témoin Blé-blé
Achat
3Traitements Jachère Akteur témoin Jachère
2007
Flair témoin Jachère
10-Oct. 31-Mai

96
Description et analyse des caractéristiques des courbes de cumul de la matière sèche.
1- Une description graphique des valeurs du paramètre « a » ou « i » des courbes d’évolution du
cumul de la matière sèche journalière obtenues pour chaque variante de chaque année de 2000 à
2007, à raison d’une valeur moyenne des 4 répétitions par variante.
2- Des analyses de variance entre les valeurs du paramètre « a » ou « i » afin de déterminer la
significativité des différences entre les différentes modalités d’année, de variété, de traitement, de
précédent et date de semis. Une régression multiple permettra également de déterminer le poids
relatif de chaque modalité dans l’explication des variations des valeurs de « a » ou « i ».
Analyse des relations entre les caractéristiques des courbes de cumul de la matière sèche et
les rendements.
3- Une régression multiple entre le rendement, les paramètres « a » et « i », et les modalités
d’obtention de ceux-ci, afin de déterminer le poids relatif de chaque variable explicative dans les
variations des rendements.
4- Des régressions simples entre la variable dépendante « rendement » et une des deux variables
indépendantes explicatives les plus déterminantes, « a » ou « i », de la régression multiple
précédente. Ses régressions simples sont effectuées avec :
• toutes les données sont utilisées, à l’exception des années 2004 et 2008 qui sont gardées
pour la validation qui fera l’objet de la PARTIE III de ce document.
• toutes les données en omettant les données exceptionnelles du précédent blé deux années.
Cette régression, comparée à la précédente, permettra de déterminer l’influence de
l’utilisation du précédent blé-blé, précédent qu’il est rare de rencontrer chez un agriculteur.
• toutes les données en omettant les données exceptionnelles issues de l’année 2005 qui sont
influencées par une sécheresse survenue pendant le remplissage des grains.
• chaque variété prise individuellement afin de déterminer si un modèle linéaire simple par
variété ne fournit pas de meilleurs résultats.
• toutes les données à l’exception de celles issues de celles de la variété Flair qui montrent un
comportement sensible aux stress hydriques.
Ces différents modèles simples seront comparés afin de déterminer l’influence des différents
facteurs sur un modèle simple mettant en relation « a » ou « i » et le rendement.
5- Une régression multiple entre la variable dépendante « rendement » et les variables
indépendantes « a » et « i » pour chacune des possibilités des analyses précédentes, afin de

97
déterminer si le modèle linéaire tenant compte des deux variables « a » et « i » ne fournit pas de
meilleurs résultats que les modèles simples utilisant uniquement l’un ou l’autre.
Validation des relations significatives obtenues entre les caractéristiques des courbes du
cumul de la matière sèche et les rendements.
6- Une validation croisée type « leave-one-out » a été effectuée sur les modèles significatifs
(p<0 ,05) expliquant plus de 50 % des rendements (R²>0,5) afin de pouvoir comparer leur aspect
prédictif.
Résultats et discussion.
Description et analyse des caractéristiques des courbes de cumul de la matière sèche.
1. L’histogramme représentant les valeurs des paramètres a et i des courbes d’évolution du cumul
de la matière sèche journalière obtenues pour chaque variante de chaque année de 2000 à 2007 met
en évidence l’effet « année » sur les valeurs des paramètres (Figure 26), surtout entre les années
aux conditions favorables à la production de bons rendements (2000, 2006 et 2007) et les années
aux conditions moins favorables. D’autres différences entre variétés, traitements, date de semis et
précédent sont présentes, mais moins visibles au premier coup d’œil.
2. Les résultats des analyses de variance (ANOVA) sur les différences entre les effets « variété »,
« traitement » , « précédent » et « date de semis » ont permis de déterminer si celles-ci sont
significatives.
L’analyse variétale : Une ANOVA effectuée sur l’ensemble des témoins de toutes variétés, en
excluant les différentes dates de semis et précédents autres que la jachère montre une différence
variétale significative uniquement entre le « a » et le « i » de la variété Flair et le « a » et le « i »
des autres variétés (p=0,000024).
L’analyse des traitements : L’effet significatif du traitement avait déjà été mis en évidence pour
le triple traitement de la variété Achat (3 traitements) sur les années 2006 et 2007 au Chapitre I.2.
(p=0,00005). Pour cette même variété, une analyse portant uniquement sur les précédents jachère à
date de semis équivalente, a permis de montrer une différence significative entre les valeurs du
paramètre « a » pour le simple traitement au stade GS 59 (p=0,03) et pour le double traitement au
stade GS39+GS59 (p=0,003) par rapport au témoin. Les différences de valeurs du paramètre « i »
ne sont significatives qu’avec le double traitement GS39+GS59 (p=0,007). Pour la variété Flair et
Akteur, aucun traitement ne montre de différences significatives pour le paramètre « i » ou « a ».
Très peu de données étaient disponibles pour ces deux variétés.

98
Légende : te=témoin ; s2=date de semis 14 Octobre ; s3=date de semis 6 Novembre ; GS=stades zadoks (Annexe II). Figure 26 : Valeurs des paramètres a et i des courbes d’évolution du cumul de la matière sèche journalière pour chaque variante de chaque année de 2000 à 2007.
L’analyse des précédents : Seule la variété Achat présente des précédents différents du précédent
Jachère en 2005 (précédent blé) et en 2007 (précédent blé deux années). Une ANOVA effectuée
sur l’ensemble des données « Achat témoin Jachère » et « Achat témoin précédent blé » ou « blé
deux années » a montré que les différences entre les valeurs de « a » et « i » étaient significatives
(p≤0,01 pour « a » et p≤0,0001 pour « i ») entre la jachère et le précédent blé.
L’analyse de la date de semis : Seule la variété Achat en 2005 a été étudiée avec différentes dates
de semis. Un effet de la date de semis est observé sur les valeurs du paramètre « i » (p≤0,01) du
témoin Achat (p≤0,001), sur les valeurs du paramètre « a » (p≤0,05) et « i » (p≤0,001) de « Achat
GS39, sur les valeurs du paramètre « a » (p≤0,01) et « i » (p≤0,01) de « Achat GS59», et sur les
valeurs du paramètre « a » (p≤0,001) et « i » (p≤0,001) de « Achat GS39+GS59».
L’analyse de variance faisant intervenir les interactions entre les différents facteurs
(multifactorielle du type « année » X « traitement ») n’a pas été possible car elle nécessite un
nombre suffisant et égal de données par variante.
Une régression multiple menée sur l’ensemble des facteurs explicatifs des variations de « a » ou
« i » montre que le principal facteur explicatif est la variété, aussi bien pour « a » que pour « i »
0
100
200
300
400
500
600
700
800
900
Fla
ir te
200
0Fla
ir G
S59
200
0Fla
ir G
S31
+59
200
0Fla
ir te
200
1A
chat
te 2
002
Ach
at te
200
3Ach
at te
s2
2003
Ach
at te
s3
2003
Ach
at G
S39
200
3A
chat
GS39
s2
2003
Ach
at G
S39
s3
2003
Ach
at G
S59
200
3A
chat
GS59
s2
2003
Ach
at G
S59
s3
2003
Ach
at G
S31
+59
200
3A
chat
GS
31+59
s2
2003
Ach
at G
S31
+59
s3
2003
Ach
at te
200
4A
chat
te 2
005
Ach
at te
blé
200
5Akt
eur te
200
5A
chat
GS
39 2
005
Akt
eur G
S39
200
5A
chat
GS
59 2
005
Akt
eur G
S59
200
5Ach
at G
S39
+59
200
5A
kteu
r G
S39
+59
200
5 A
chat
te 2
006
Ach
at te
Sai
n 20
06 A
chat
te 2
007
Ach
at te
Sai
n 20
07Fla
ir te
200
7Akt
eur te
200
7Ach
at b
lé-b
lé 2
007
Variantes
a ou
i i a
i

99
(Tableau 25). Le second facteur explicatif est le traitement pour « a » et la date de semis pour
« i ». Cette observation est valable pour n’expliquer que 49% des variations de « a » et 55% des
variations de « i », comme l’indique le coefficient de détermination.
Tableau 25 : Résultats de la régression multiple entre « a » ou « i » et les facteurs explicatifs des variations observées de 2000 à 2007.
a i
Synthèse Régression R²=0,49; p<0,000001; n= 136 R²=0,55; p<0,00001; n= 136
Bêta p Bêta p Année n.s. n.s. -0,211236 ** Variété -0,632 *** -0,502 ***
Traitement 0,160 ** 0,237 *** Précédent n.s. n.s. n.s. n.s.
Date de semis -0,149 * -0,317 *** Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Analyse des relations entre les caractéristiques des courbes de cumul de la matière sèche et
les rendements.
3. L’utilisation des valeurs des paramètres « a » et « i » pour expliquer la variabilité des
rendements obtenus par répétition des différentes variantes observées entre 2000 et 2007 passe
également par l’analyse des facteurs explicatifs présents, autres que « a » et «i ». L’importance
relative des différentes variables explicatives est évaluée par la régression multiple entre le
rendement et celles-ci (Tableau 26). Cette régression montre que la variable « i » est celle qui
explique le mieux les rendements obtenus (Bêta beaucoup plus élevé que les autres), avec une
significativité très élevée. La variable « a » est la deuxième variable explicative.
Tableau 26 : Résultats de la régression entre le rendement et ses facteurs explicatifs entre 2000 et 2007.
Rendement
Synthèse Régression R²=0,68; p<0,00001; n= 136
Bêta p Année 0,477 ***
Variété 0,188 **
Traitement n.s n.s
Précédent -0,212 ***
Date de semis n.s n.s
a -0,545 ***
i 1,391 *** Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).

100
4. Les relations entre les variables explicatives des rendements les plus importantes, « i » ou « a »,
et le rendement ont été étudiées plus en détail par l’établissement de régressions linéaires simples.
Le Tableau 27 montre les résultats de ces régressions simples, effectuées avec l’ensemble des
données, en supprimant les précédents culturaux blé, l’année ayant subi la sècheresse, la variété
Flair, ou en utilisant uniquement les données d’une variété.
Tableau 27 : Résultats statistiques des régressions simples obtenues entre les paramètres a ou i et les rendements de 2000 à 2007.
Régression linéaire « a »/ Rendement
Régression linéaire « i » / Rendement
R² p R² p n Toutes les données 0,17 *** 0,37 *** 136
Sans précédent blé-blé 0,14 *** 0,31 *** 132 Sans l’année sècheresse 2005 n.s. n.s. n.s. n.s. 100
Uniquement Achat 0,45 *** 0,67 *** 96 Uniquement Akteur 0,94 *** 0,77 *** 20 Uniquement Flair n.s. n.s. n.s. n.s. 20
Toutes les données sans Flair 0,49 *** 0,66 *** 116 Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Les régressions par année ne sont pas présentées car chaque année contient un nombre de données
différent, certaines années n’ont qu’une variété et toutes les années n’ont pas les mêmes variétés
présentes. Les traitements ont un léger effet sur la variabilité des rendements (Tableau 26), ce qui
explique également pourquoi les régressions entre la variable « i » ou « a », et le rendement n’ont
pas été étudiées par traitement.
L’interprétation des résultats des régressions simples (Tableau 27) entre la variable « a » ou la
variable « i » et le rendement sur l’ensemble des données ou sur une partie (suivant les modalités
de précédent, sècheresse, variété et traitements) nous amène aux conclusions suivantes :
Dans tous les cas où la relation est significative, la variable « i » est mieux corrélée au rendement
que la variable « a », à l’exception de la variété Akteur pour laquelle c’est la variable « a » qui est
mieux corrélée. Cette variété semble avoir une relation entre le cumul de la matière sèche totale
qu’elle produit après la floraison et le rendement en grains plus important que les autres variétés.

101
Les seules relations significatives et expliquant plus de 50% (R²>50%) des rendements sont :
- La relation entre la variable « i » et le rendement de la variété Achat.
- La relation entre la variable « i », et le rendement de la variété Akteur.
- La relation entre la variable « a », et le rendement de la variété Akteur.
- La relation entre la variable « i », et le rendement de toutes les données à l’exception de
celles de la variété Flair.
Proximité entre les modèles simples significatifs utilisant « i » : Toutes les régressions
significatives et expliquant au moins 50% des rendements présentent un intercept entre 3,9 et 4,6 et
une pente entre 0,019 et 0,020.
Le modèle global obtenu au chapitre I.2. sur les données de 2006 et 2007 (y = 5,0965 + 0,0166*x)
présente des valeurs d’intercept et de coefficient angulaire également très proches de celles-là. Une
analyse de variance entre ces différents modèles montre que leur pente et intercept ne sont pas
significativement différents. Le modèle global obtenu avec toutes les données de 2006 et 2007
explique cependant une plus grande proportion des rendements que le modèle global utilisant les
données de 2000 à 2007, avec un coefficient de détermination de 57%. Ceci est peut-être dû à
l’utilisation de valeurs plus proches de la réalité pour ces deux années que pour celles de 2000 à
2005, dont l’obtention et l’utilisation a nécessité d’émettre un certain nombre d’hypothèses. La
présence dominante de données issues d’une même variété (Achat) en 2006 et 2007 peut aussi être
mise en cause. L’effet variétal sur la variabilité des rendements est particulièrement important
comme cela sera montré plus loin.
Discussion des effets « précédent » et « variété » sur les résultats des modèles simples
utilisant « i » ou « a » : D’après les résultats (Tableau 27), la suppression des données
« précédent blé », au nombre de 8 données, n’a pas beaucoup amélioré le coefficient de
détermination du modèle. Cette constatation est en accord avec les conclusions de l’analyse des
résultats de la régression multiple indiquant un faible poids du précédent dans l’explication de la
variabilité des rendements obtenus.
L’utilisation des données par variété a amélioré le coefficient de détermination des modèles pour
deux variétés sur les trois utilisées, ce qui suggère un effet variétal important. La régression entre
le rendement et « i » ou « a » montre un coefficient de détermination (R²) supérieur à 50%
uniquement lorsque cette régression est effectuée par variété pour 2 des 3 variétés. Cela signifie
que 50% du rendement ou plus (jusqu’à 94% pour la relation de « a » par rapport au rendement de
Akteur) est expliqué par la variable « i » ou « a » uniquement lorsque la relation est étudiée par
variété.

102
Le modèle mettant en relation la variable « i » et le rendement sur les données de 2006 et 2007 de
la variété Achat seule présentait lui aussi un meilleur coefficient de détermination (R² de 72%) que
celui obtenu en utilisant la variable « i » et les rendements de toutes les variétés. Les deux modèles
obtenus pour la variété Achat sont extrêmement proches (y = 3,8963 + 0,0206*x pour le modèle de
2006-2007 et y = 6,378 + 0,0055*x pour le modèle 2000-2007).
Les mauvais résultats observés avec la variété Flair sont probablement dus au comportement de
cette variété vis-à-vis de l’eau. Il a en effet été observé au cours des années 2006 et 2007 que la
variété Flair est la première affectée par des périodes où le sol est loin de sa capacité au champ
sans pour autant qu’il soit question de sècheresse. Les parcelles de la variété Flair souffraient
visiblement (dessèchement prématuré des feuilles) assez rapidement d’un manque en eau.
Lorsque l’hypothèse est posée que cette variété a un comportement qui n’est pas pris en compte
par le modèle de production de matière sèche duquel les variables « i » et « a » sont issues, et par
conséquent qu’on exclut la variété Flair de nos analyses, les régressions entre les variables « i » ou
« a » et le rendement présentent un coefficient de détermination nettement amélioré (Tableau 27).
Cette exclusion entraîne malheureusement la perte de toutes les données de 2000 et 2001, puisque
seules les données de la variété Flair sont disponibles pour ces deux années.
En conclusion, l’étude de la relation entre la variable « i » ou la variable « a » et le rendement
montre :
- Que globalement la régression simple avec la variable « i » explique mieux la variabilité
des rendements que celle utilisant « a », à l’exception des régressions établies avec les
données de la variété Akteur ;
- Des meilleurs résultats lorsque la relation par variété est établie, par rapport à la relation
utilisant l’ensemble des données ;
- Des résultats moins satisfaisants avec la variété Flair qu’avec les autres variétés, peut-être à
cause de sa sensibilité aux stress hydriques ;
- Des meilleurs résultats pour la régression utilisant toutes les données à l’exception de celles
de la variété Flair que pour la régression utilisant toutes les données ;
- Aucun effet sécheresse, ni précédent sur la relation « i » / rendement.
5. Pour se rapprocher au plus près de la véritable relation qui lie le rendement obtenu à la forme de
la courbe du cumul de la production de matière sèche journalière, il faut caractériser cette relation
avec le plus de variables décrivant cette courbe. C’est pourquoi il a aussi été étudié la relation entre

103
le rendement et les 2 variables (point d’inflexion (i) et plateau (a)) caractérisant la courbe en
utilisant la régression multiple du rendement par rapport à « i » et « a ». Chacune des analyses
précédentes, effectuées avec la variable « a » seule ou la variable « i » seule, a été menée en
utilisant cette fois une régression multiple entre le rendement et les deux variables indépendantes
« a » et « i » (Tableau 28). Chaque relation significative obtenue montre un coefficient de
détermination toujours supérieur à celui obtenu avec une seule de ces deux variables.
L’interprétation des différences observées entre les modèles est la même que celle des modèles
simples.
Tableau 28 : Résultats statistiques des régressions multiples obtenues entre les paramètres « a » et « i », et les rendements de 2000 à 2007.
Régression linéaire « a » et « i » / Rendement
Synthèse régression a i R² p n Bêta p Bêta p
Toutes les données 0,43 *** 136 -0,532 *** 1,07 *** Sans précédent blé-blé 0,36 *** 132 -0,49 *** 0,99 ***
Sans l’année sécheresse 2005 0,13 *** 100 -0,55 ** 0,65 ***
Uniquement Achat 0,67 *** 96 n.s. n.s. 0,82 ***
Uniquement Akteur 0,94 *** 20 0,90 *** n.s. n.s. Uniquement Flair n.s. n.s. 20 n.s.
n.s. n.s. n.s.
Toutes les données sans Flair 0,66 *** 116 n.s. n.s. 0,80 ***
Légende : n.s. : Non Significatif ; * Significatif (p≤0,05) ; ** Hautement significatif (p≤0,01) ; *** Très hautement significatif (p≤0,001).
Ces relations ne peuvent cependant être utilisées de manière prévisionnelle que si la valeur du
plateau « a » peut être déterminée avant la maturité finale.
Une des possibilités qui permettrait d’obtenir la variable « a » avant la maturité finale est d’utiliser
la relation linéaire simple existant entre « i » et « a » des années 2000 à 2007 (Figure 27). La
prévision par l’utilisation de ces régressions multiples peut alors être effectuée dès que la valeur du
point d’inflexion (i) est connue, en cours de saison, tout comme pour la régression simple
impliquant uniquement le i. La régression par la méthode des moindres carrés entre « a » et « i »
montre un R² de 75%, ce qui indique qu’une grosse proportion de « a » peut-être déterminée à
partir du i (Figure 27).
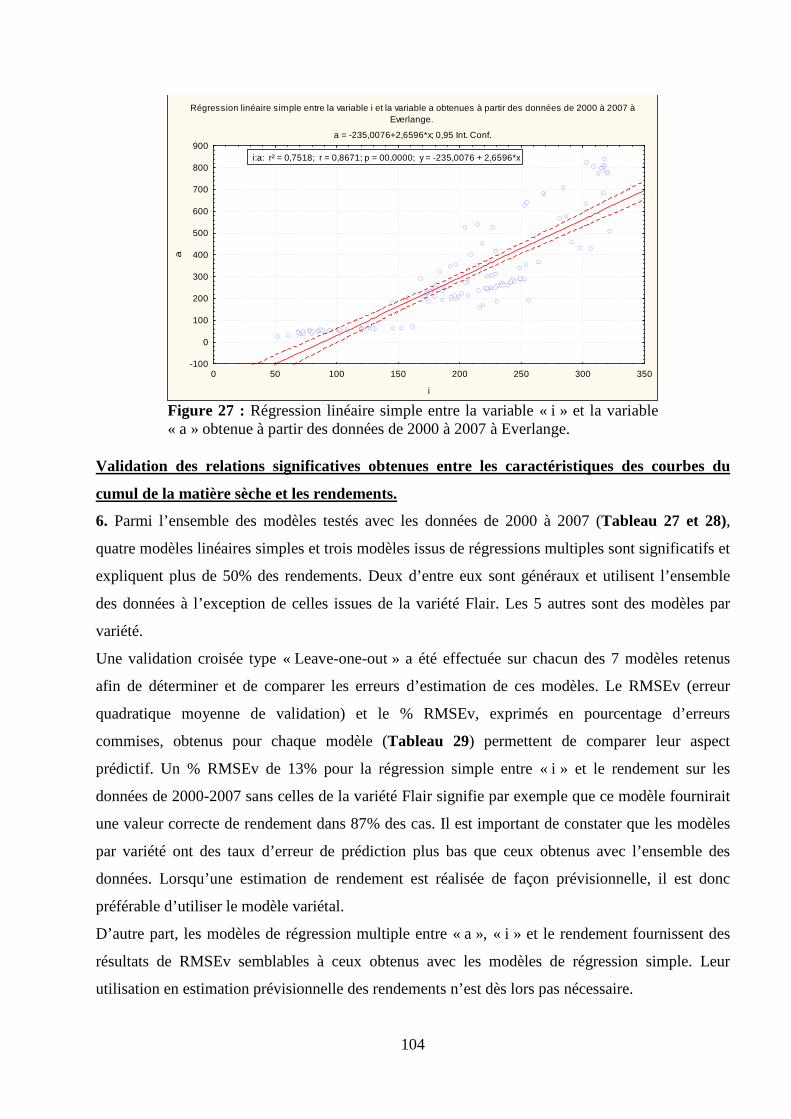
104
Régression linéaire simple entre la variable i et la variable a obtenues à partir des données de 2000 à 2007 àEverlange.
a = -235,0076+2,6596*x; 0,95 Int. Conf.
0 50 100 150 200 250 300 350
i
-100
0
100
200
300
400
500
600
700
800
900
a
i:a: r² = 0,7518; r = 0,8671; p = 00,0000; y = -235,0076 + 2,6596*x
Figure 27 : Régression linéaire simple entre la variable « i » et la variable « a » obtenue à partir des données de 2000 à 2007 à Everlange.
Validation des relations significatives obtenues entre les caractéristiques des courbes du
cumul de la matière sèche et les rendements.
6. Parmi l’ensemble des modèles testés avec les données de 2000 à 2007 (Tableau 27 et 28),
quatre modèles linéaires simples et trois modèles issus de régressions multiples sont significatifs et
expliquent plus de 50% des rendements. Deux d’entre eux sont généraux et utilisent l’ensemble
des données à l’exception de celles issues de la variété Flair. Les 5 autres sont des modèles par
variété.
Une validation croisée type « Leave-one-out » a été effectuée sur chacun des 7 modèles retenus
afin de déterminer et de comparer les erreurs d’estimation de ces modèles. Le RMSEv (erreur
quadratique moyenne de validation) et le % RMSEv, exprimés en pourcentage d’erreurs
commises, obtenus pour chaque modèle (Tableau 29) permettent de comparer leur aspect
prédictif. Un % RMSEv de 13% pour la régression simple entre « i » et le rendement sur les
données de 2000-2007 sans celles de la variété Flair signifie par exemple que ce modèle fournirait
une valeur correcte de rendement dans 87% des cas. Il est important de constater que les modèles
par variété ont des taux d’erreur de prédiction plus bas que ceux obtenus avec l’ensemble des
données. Lorsqu’une estimation de rendement est réalisée de façon prévisionnelle, il est donc
préférable d’utiliser le modèle variétal.
D’autre part, les modèles de régression multiple entre « a », « i » et le rendement fournissent des
résultats de RMSEv semblables à ceux obtenus avec les modèles de régression simple. Leur
utilisation en estimation prévisionnelle des rendements n’est dès lors pas nécessaire.

105
Tableau 29 : Résultats des validations croisées effectuées sur chaque modèle d’estimation des rendements significatifs et présentant un coefficient de détermination supérieur à 50%.
RMSEv
RMSEv
(%)
Sans la variété Flair :
Rendement = 3,950 + 0,0196*i 0,976 13%
Achat :
Rendement = 3,887 + 0,0196*i 0,859 11%
Régression simple i/Rdmt
données 2000-2007
Akteur :
Rendement = 4,633 + 0,0202*i 0,553 8%
Régression simple a/Rdmt
données 2000-2007
Akteur :
Rendement = 5,973 + 0,0094*a 0,295 4%
Sans la variété Flair :
Rendement = 3,977+ 0,0193*i + 0,000163*a 0,979 13%
Achat sans précédent blé :
Rendement = 3,888+ 0,020*i + 0,000007*a 0,864 11%
Régression i et a /Rdmt
données 2000-2007
Akteur :
Rendement = 5,857+ 0,001*i + 0,009*a 0,319 5%
Conclusion
Le modèle du calcul de la matière sèche journalière (Bancal et al., 2007) a été retenu pour ses
meilleurs résultats à l’étape de validation. Les relations entre les paramètres point d’inflexion (i) et
/ou plateau (a) de la courbe d’évolution du cumul de la matière sèche journalière et le rendement
ont été étudiées sur un ensemble de données plus grand, grâce à l’ajout de données issues
d’expérimentations menées entre 2000 et 2005 à celles de 2006 et 2007.
Des différences de valeurs de « i » et « a » sont présentes entre les années pour une même variété.
L’analyse de la variance des valeurs de « i » et « a » n’a pas permis de détacher de différences
variétales, à l’exception de Flair qui se distingue des autres variétés. Les valeurs de « i » ont
permis de distinguer le double traitement du témoin de Achat, et les valeurs de « a » ont permis de
distinguer le double et le simple traitement du témoin de Achat. Les autres variétés n’ont pas
présenté suffisamment de données de traitements pour que l’analyse ait pu être effectuée. Le
précédent blé s’est distingué du précédent jachère, aussi bien avec les valeurs de « i » qu’avec les
valeurs de « a ». La date de semis a également montré des différences significatives de valeurs de
« a » et « i ». La régression multiple a mis en évidence l’ordre d’importance des facteurs
expliquant ces différences. Avec les valeurs de « a », il a été obtenu, par ordre d’importance, la

106
variété, les traitements et la date de semis. L’année n’est pas significative. Avec les valeurs de
« i », il a été obtenu, par ordre d’importance, la variété, la date de semis, l’année et les traitements.
L’obtention de la variété comme effet principal alors que seules les valeurs de « a » et « i » de Flair
se distinguent des autres variétés pourrait bien être due à la faible valeur du coefficient de
détermination de la régression multiple. Celle-ci indique que la régression multiple n’explique que
~50% des valeurs de « a » et « i ». Le poids de l’année dans l’explication des valeurs de « a » et
« i » est également inférieur à celui attendu.
Une régression multiple, établie cette fois entre le rendement et les différentes variables
explicatives de celui-ci a montré l’importance relative de chacune d’entre elles. Par ordre
d’importance, les variables expliquant le rendement sont la variable « i », la variable « a », l’année,
le précédent et finalement la variété. Les résultats de cette régression multiple ont motivé le choix
d’utiliser le paramètre « i » ou/et « a » (qui sont des variables, et non plus des paramètres, dans
cette régression) pour construire notre modèle.
Différents modèles, construits avec « i » ou/et « a » sur des échantillons de données comprenant
toutes les données de 2000 à 2007 (modèle global), en tenant compte du précédent ou pas, de la
sècheresse, ou par variété ont été élaborés. L’effet année n’a pas pu être testé car chaque année
présente un nombre de données très différent. Les modèles les plus significatifs et expliquant le
mieux le rendement sont ceux effectués par variété sur Achat et Akteur. Ce résultat est en accord
avec les résultats de l’analyse précédente ayant montré l’importance de la variété sur les valeurs
« a » ou « i » (régression multiple entre « i » ou « a » et les variables explicatives). La variété Flair
montre de mauvais résultats. L’hypothèse que cette variété présente un comportement non
expliqué par les variables ou paramètres pris en compte dans le modèle du calcul de matière sèche
journalière peut-être avancée. Ce pourrait être le comportement variétal vis-à-vis du contenu en
eau du sol. Celui-ci n’est pas pris en compte par le modèle et il a été observé en 2006 et 2007 que
cette variété présente une grande sensibilité au stress hydrique. L’effet variétal suggère qu’il est
nécessaire d’utiliser un modèle différent par variété. Les modèles globaux, utilisant l’ensemble des
données de 2000 à 2007 ont présenté de mauvais résultats. En tenant compte de l’hypothèse
précitée sur le comportement de la variété Flair, les données issues de celle-ci ont été extraites et le
modèle global a été ré-estimé. Deux modèles globaux expliquant plus de 50 % des rendements ont
été obtenus de cette façon. L’un d’eux entre le rendement et la variable « i » et l’autre entre le
rendement et les variables « i » et « a ». Les modèles par variété restent toutefois bien meilleurs
que les modèles globaux sans Flair. Il a été constaté cependant que les modèles par variété sont
très proches du modèle global. On a obtenu une grande stabilité des coefficients du modèle de

107
régression linéaires simple. Les valeurs des pentes et des intercepts ne varient pas
significativement entre les modèles obtenus par variété, le modèle global et le modèle obtenu avec
les données de 2006 et 2007. Les modèles multiples, prenant en compte à la fois le point
d’inflexion (i) et le plateau (a) pour expliquer les rendements présentent un coefficient de
détermination à chaque fois plus élevé que le coefficient de détermination des modèles simples
utilisant « i » ou « a » correspondants. Ces modèles expliquent donc une plus grande proportion de
la variabilité des rendements utilisés que les modèles simples.
Enfin, le précédent cultural, facteur explicatif des variations de rendement le plus important après
« i », « a » et l’année, a été abordé. Le retrait des données de précédent blé pour l’élaboration des
modèles simples ou multiples entre le rendement et « a » ou « i » a fourni des modèles moins
performants. Les données de rendement des précédents blé sont moins élevées que celles des
précédents jachère, et la matière sèche produite également. La courbe de cumul de la matière sèche
montre des valeurs moins élevées, et les paramètres « a » et « i » de celle-ci sont influencés par le
précédent. L’effet du précédent sur les rendements est également visible sur « a » et « i », ce qui
explique peut-être pourquoi le retrait de ces données diminue la performance des modèles.
Des validations croisées type « Leave-one-out » menées sur chacun des 7 modèles significatifs et
expliquant plus de 50% des rendements ont montré que les modèles variétaux fournissent moins
d’erreurs de prédiction que les modèles globaux. Les résultats restent toutefois assez proches
puisque le modèle le moins bon montre un pourcentage d’erreur de 13% et le meilleur un
pourcentage d’erreur de 4%. Les modèles multiples n’ont pas fourni de meilleurs résultats que les
modèles simples pour la validation croisée et ne sont donc pas de meilleurs « prédicteurs » du
rendement.
Bien que ces modèles montrent de bons résultats, il ne faut pas négliger l’importance de l’effet
année, au jugé de la première analyse de ce chapitre. Il se pourrait bien que certains modèles
fournissent de meilleurs résultats par groupes d’années, comme cela a été montré pour la
régression simple entre le rendement et le cumul de matière sèche de l’étude de Bancal et al.
(2007). Cette étude montre aussi que la contribution aux rendements des réserves pré-floraison
varie significativement en fonction des années, de l’amendement azoté et de la densité du couvert,
mais pas en fonction des fongicides et des génotypes. Les réserves pré-floraison n’ont pas été
prises en compte dans cette étude-ci, mais il se pourrait bien qu’elles jouent également un rôle
important dans la valeur de l’intercept de nos modèles. L’intercept des différents modèles ne varie
pas beaucoup, mais c’est probablement dû à une densité du couvert variant peu et à un

108
amendement azoté semblable pour les différentes expérimentations. Il se pourrait que les valeurs
de celui-ci varient d’une année à l’autre, ou suivant des groupes d’années.

109
Conclusion générale de la partie II
Les données acquises durant la première partie de cette thèse ont été utilisées afin de tester la
validité de deux modèles d’estimation des rendements du blé d’hiver au Grand-Duché de
Luxembourg. Ces modèles, basés sur l’interception du rayonnement journalier par la surface verte
des trois dernières strates foliaires, ont été utilisés suivant différentes modalités de traitements et
variétés avec les données de 2006 et 2007.
Des analyses statistiques classiques ont été menées afin de déterminer quelles sont les sorties des
modèles qui présentent les meilleures corrélations avec les rendements obtenus. Pour chacun des
deux modèles, la meilleure corrélation a été obtenue entre le rendement et un paramètre descriptif
de la courbe d’évolution du cumul des valeurs journalières des sorties du modèle.
C’est l’analyse portant sur le paramètre descriptif «i», point d’inflexion des courbes décrivant
l’évolution du cumul de la production journalière de matière sèche, qui a fourni les meilleurs
résultats. En effet, la corrélation entre le rendement et le point d’inflexion « i » est plus élevée
lorsqu’il s’agit du point d’inflexion des courbes décrivant l’évolution du cumul de la production
journalière de matière sèche que lorsqu’il s’agit du point d’inflexion de la courbe d’évolution du
cumul du HAA journalier. Les résultats suggèrent un effet variétal et un effet « année » qui doivent
être confirmés, étant donné le peu de données utilisées lorsqu’on sépare les variétés ou les années
sur l’ensemble des deux années de données utilisées.
Une simplification a été menée sur les deux modèles, afin de déterminer s’il était intéressant
d’utiliser uniquement la surface verte des deux dernières strates foliaires ou de la dernière strate
foliaire seule. Cette simplification apporterait un gain considérable de temps lors des mesures
hebdomadaires de surfaces foliaires vertes, celles-ci étant les données d’entrée des modèles les
plus coûteuses en temps à obtenir. La simplification des deux modèles par la suppression
successive de la troisième et de la deuxième strate foliaire a fourni, dans la plupart des cas, de
moins bonnes corrélations avec les rendements lorsqu’on utilise uniquement une ou deux strates
foliaires.
L’utilisation d’estimations visuelles du pourcentage de surface foliaire verte à la place des
estimations de ce pourcentage par l’analyse d’images fournirait également un gain de temps dans
l’obtention de ces données. Des données d’estimation visuelle obtenues sur les mêmes plantes que
les estimations par l’analyse d’images en 2006 et 2007 ont été utilisées pour l’obtention des mêmes
modèles de calcul de la matière sèche journalière. La comparaison des résultats montre une
différence moyenne de 5% entre le cumul de la matière sèche obtenue selon les deux méthodes

110
d’estimation du pourcentage de surface verte. Les paramètres « i » et « a » des modèles montrent
une différence moyenne de 8%. Les coefficients de détermination des modèles de régression entre
ces deux paramètres et les rendements ont montré des différences de 20% entre les deux méthodes
dans 3 cas sur 4. De plus, la significativité des régressions entre le « a » ou le « i » et les
rendements est toujours supérieure lorsque le « a » et le « i » sont issus de l’utilisation des
estimations du pourcentage de surface verte par l’analyse d’images. Ces différences sont non
négligeables. Cependant, des données de pourcentage de surface verte issues d’estimations
visuelles des années 2000 à 2005 ont tout de même été utilisées pour valider les modèles
d’estimation des rendements retenus sur un plus grand nombre de données.
Malgré les nombreuses hypothèses qui ont dû être émises pour pouvoir ajouter les données
supplémentaires de 2000 à 2005, les résultats obtenus sont proches de ceux obtenus en utilisant
uniquement les données de 2006 et 2007. De plus, les conclusions concernant l’existence d’un
effet variétal sont confirmées. L’effet important de l’année sur les valeurs de cumul de la matière
sèche indique que les performances de ces modèles pourraient être améliorées s’ils étaient élaborés
par groupe d’années. Cela n’a pas été envisagé car il ne serait plus possible d’utiliser de tels
modèles de manière prédictive.
Des 7 modèles retenus, il est constaté que les modèles par variété mettant en relation le rendement
avec une des deux ou les deux variables descriptives, « i » (point d’inflexion) et « a » (plateau), du
modèle de production journalière de la matière sèche présentent de meilleurs résultats. Ces
meilleurs résultats des modèles par variété sont observés tant pour expliquer la variabilité des
rendements utilisés pour construire le modèle (R² de validation de 77% à 94% pour les modèles
variétaux, contre 66% pour le modèle général « i » / rendement) que en prédiction (%RMSE
validation croisée indiquant de 8% à 11% d’erreurs pour les modèles variétaux contre 13% pour
les modèles généraux).
Ces modèles doivent maintenant être testés sur des échantillons externes, indépendants de ceux
utilisés pour construire ce modèle. Les résultats obtenus avec chaque modèle variétal doivent être
comparés aux résultats obtenus en utilisant les mêmes données avec le modèle général. Les
résultats issus des modèles multiples (avec « i » et « a ») doivent également être comparés aux
résultats issus des modèles simples (avec « i » ou « a »).
Dans la PARTIE III de ce document, il sera montré comment ce genre de modèle peut être utilisé à
des fins prédictives, afin d’estimer les rendements le plus précocement en cours de saison.

111
PARTIE III
APPLICATION DU MODELE EN
CONDITIONS REELLES.

112
Note préalable
Cette dernière PARTIE a pour objectif d’exposer, au travers d’un exemple, de quelle manière un
modèle, tel que ceux obtenus à l’issue de la PARTIE II, peut être utilisé de manière prévisionnelle.
La méthode utilisée consiste à faire une estimation de rendement à la floraison, en utilisant les
valeurs des variables d’entrée du modèle obtenues en effectuant des mesures jusqu’à cette date, et
en déterminant comment vont se comporter ces variables dans les semaines post floraison.
Matériel et méthodes.
Les modèles testés sont le modèle général simple et le modèle général multiple ainsi que le modèle
simple spécifique des variétés utilisées. Ces modèles sont ceux obtenus à l’issue de la PARTIE II
(Tableau 29). Ces modèles seront testés de façon prévisionnelle sur les données du témoin de
Achat de 2004 sur précédent jachère et sur les données du témoin de Rosario précédent jachère et
Tommi précédent jachère de 2008 en suivant les étapes de la Figure 28. La détermination du
rendement par l’utilisation de chacun de ces modèles nécessite la connaissance des valeurs de la
variable « i » seule pour les modèles simples, ou des variables « i » et « a » pour les modèles
multiples (Figure 28-Etape 6.).
L’obtention de la valeur de ces variables le plus tôt possible dans la saison ne peut se faire que
moyennant la connaissance du comportement de la courbe d’évolution du cumul de la production
journalière de matière sèche, car le paramètre « i » représente le point d’inflexion de la courbe et
« a » le plateau (Figure 28-Etape 5.). La formule mathématique de la courbe est obtenue par une
méthode d’estimation non linéaire sur base de quelques points pour lesquels la matière sèche a été
calculée à l’aide du modèle du calcul de la matière sèche. Il est donc nécessaire de connaître les
valeurs des termes de cette formule de calcul de la matière sèche avant la récolte également
(Figure 28-Etape 4.).
Ces termes, présentés dans la PARTIE II de ce document sont :
MSJ = Ei(j) Eb Ij
Les valeurs de chacun de ces termes sont obtenues à partir de l’utilisation des mesures (Figure 28-
Etape 2.) effectuées jusqu’à la date à laquelle une estimation du rendement de l’année en cours est
entreprise (= en pré-estimation). Après la date à laquelle une estimation du rendement (= en post-
estimation) de l’année en cours est effectuée, les valeurs de ces termes doivent également être

113
estimées afin d’avoir un nombre de valeurs de matière sèche journalière suffisant pour que puisse
être adaptée une courbe d’évolution du cumul de la matière sèche à ces valeurs (Figure 28-Etape
1.).
Détermination des termes de l’équation Ei(j) Eb et Ij en pré-estimation et en post-estimation.
Le terme I j, rayonnement solaire incident dans la gamme du rayonnement photosynthétiquement
actif, peut être obtenu facilement à partir des mesures du pyranomètre en pré-estimation (Figure
28-Etape 2.). Les valeurs post-estimation de rayonnement sont nécessaires afin d’établir la courbe
du cumul de la production journalière de la matière sèche. Pour les obtenir, c’est la moyenne
historique (8 années) des valeurs de rayonnement des jours juliens des semaines considérées qui
est prise en compte (Figure 28-Etape 1.). Dans l’exemple présenté, le rayonnement mesuré sera
utilisé jusqu’à la floraison (valeurs pré-estimation), date à partir de laquelle une estimation des
rendements et des risques de perte de récolte est envisageable. Le rayonnement estimé à partir de
la moyenne historique sera ensuite utilisé (valeurs post-estimation). Le stade de la floraison est
choisi comme date d’estimation car c’est celui à partir duquel une estimation du paramètre b,
nécessaire au calcul du LAI, est possible (voir ci-dessous).
Le terme Eb (Figure 28-Etape 3 et 4.), efficience d’utilisation du rayonnement ou R.U.E.
(Radiation Use Efficiency) est considéré comme constant et égal à 2,8 dans notre cas. Il n’est donc
pas nécessaire d’utiliser une méthode de prédiction pour l’obtenir en post-estimation puisque sa
valeur reste constante.
Le terme Ei(j) est l’efficience d’interception du jour j, qui tient compte du LAI et du coefficient
d’interception des feuilles de blé. Pour obtenir les valeurs post-estimation de ce terme-ci, le LAI
ainsi que l’évolution de la dégradation de la surface verte post floraison (Figure 28-Etape 3.)
doivent être connus. Le calcul du LAI est effectué, suivant la formule présentée au chapitre I.1. de
la PARTIE II et demande une connaissance des surfaces foliaires des trois dernières strates, du
nombre d’épis par m² et de la proportion de brins moyens. Les surfaces foliaires peuvent être
déterminées par la prise d’images et leur analyse dès que la F1 est totalement déployée (stade
GS39). Le nombre de brins par m² ainsi que la proportion de brins moyens peuvent être obtenus à
la floraison à partir de prélèvements et du comptage (Figure 28-Etape 2.). Le LAI déterminé à la
floraison ne varie plus, toutes les feuilles étant déployées. Par conséquent, il n’est pas nécessaire
de prédire le LAI puisque celui-ci peut-être déterminé dès la floraison, date d’estimation des
rendments. Dans le cas des années 2004 et 2008, les mesures de surfaces foliaires n’ont pas été
effectuées. Le LAI obtenu en 2007 pour la variété Achat sera utilisé en 2004 pour cette même
variété et le LAI moyen de 2007 sera utilisé en 2008 pour les variétés Rosario et Tommi.
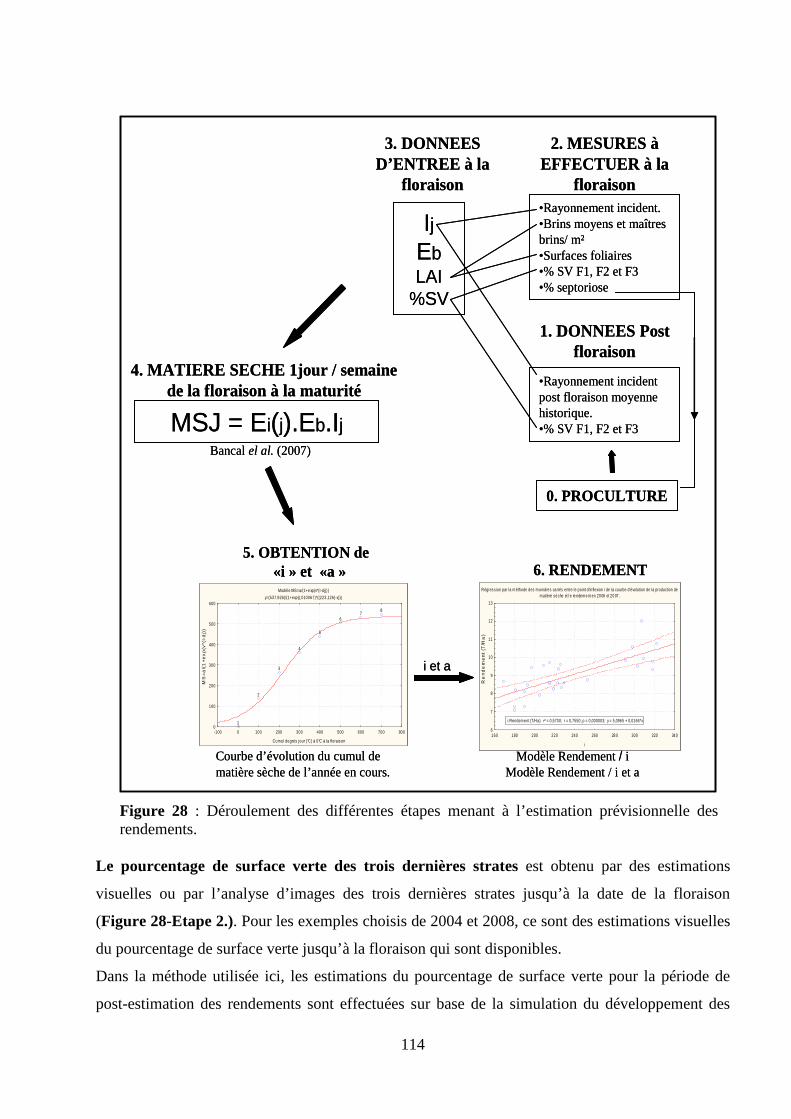
114
Figure 28 : Déroulement des différentes étapes menant à l’estimation prévisionnelle des rendements.
Le pourcentage de surface verte des trois dernières strates est obtenu par des estimations
visuelles ou par l’analyse d’images des trois dernières strates jusqu’à la date de la floraison
(Figure 28-Etape 2.). Pour les exemples choisis de 2004 et 2008, ce sont des estimations visuelles
du pourcentage de surface verte jusqu’à la floraison qui sont disponibles.
Dans la méthode utilisée ici, les estimations du pourcentage de surface verte pour la période de
post-estimation des rendements sont effectuées sur base de la simulation du développement des
Bancal el al. (2007)
Courbe d’évolution du cumul de matière sèche de l’année en cours.
Modèle Rendement / i Modèle Rendement / i et a
6. RENDEMENT
i et a
5. OBTENTION de«i » et «a »
Régres sion par la m éthode des m oindres carrés entre le point d'inflexion i de la courbe d'évolution de la production dem atiè re sèche et l e re ndem ent en 2006 et 2007.
160 180 200 220 240 260 28 0 300 320 340
i
6
7
8
9
10
11
12
13
Re
nd
em
en
t (T
/Ha
)
i:Rendem ent (T/Ha): r² = 0,5700; r = 0,7550; p = 0,000003; y = 5,0965 + 0,0166*x
MSJ = Ei(j).Eb.Ij
4. MATIERE SECHE 1jour / semaine de la floraison à la maturité
Modèle:MS=a/(1+e xp(v*(I-d j)) )
y=(537,926)/(1+exp((,010067)*((223,126)-x)))
1
2
3
4
5
67
8
-100 0 100 200 300 400 500 600 700 800
Cumul degrés jour (°C) à 0°C à la floraison
0
100
200
300
400
500
600
MS
=a
/(1
+e
xp
(v*(
I-d
j))
3. DONNEES D’ENTREE à la
floraison
IjEbLAI
%SV
2. MESURES àEFFECTUER à la
floraison•Rayonnement incident.•Brins moyens et maîtres brins/ m²•Surfaces foliaires•% SV F1, F2 et F3•% septoriose
1. DONNEES Post floraison
•Rayonnement incident post floraison moyenne historique.•% SV F1, F2 et F3
0. PROCULTURE
Bancal el al. (2007)
Courbe d’évolution du cumul de matière sèche de l’année en cours.
Modèle Rendement / i Modèle Rendement / i et a
6. RENDEMENT
i et a
5. OBTENTION de«i » et «a »
Régres sion par la m éthode des m oindres carrés entre le point d'inflexion i de la courbe d'évolution de la production dem atiè re sèche et l e re ndem ent en 2006 et 2007.
160 180 200 220 240 260 28 0 300 320 340
i
6
7
8
9
10
11
12
13
Re
nd
em
en
t (T
/Ha
)
i:Rendem ent (T/Ha): r² = 0,5700; r = 0,7550; p = 0,000003; y = 5,0965 + 0,0166*x
MSJ = Ei(j).Eb.Ij
4. MATIERE SECHE 1jour / semaine de la floraison à la maturité
Modèle:MS=a/(1+e xp(v*(I-d j)) )
y=(537,926)/(1+exp((,010067)*((223,126)-x)))
1
2
3
4
5
67
8
-100 0 100 200 300 400 500 600 700 800
Cumul degrés jour (°C) à 0°C à la floraison
0
100
200
300
400
500
600
MS
=a
/(1
+e
xp
(v*(
I-d
j))
3. DONNEES D’ENTREE à la
floraison
IjEbLAI
%SV
2. MESURES àEFFECTUER à la
floraison•Rayonnement incident.•Brins moyens et maîtres brins/ m²•Surfaces foliaires•% SV F1, F2 et F3•% septoriose
1. DONNEES Post floraison
•Rayonnement incident post floraison moyenne historique.•% SV F1, F2 et F3
0. PROCULTURE

115
symptômes de la septoriose sur les 3 derniers étages foliaires. Seul est pris en compte le
développement des symptômes qui apparaissent suite à la latence des infections qui ont lieu avant
la date d’estimation du rendement. La phase de latence et le développement des symptômes sont
estimés à partir des degrés jours issus d’une base de données historiques. Les nouvelles infections,
survenues après la date d’estimation ne sont pas prises en compte car elles nécessitent la
connaissance des pluies à venir. Le fait qu’il y ait ou non des nouvelles infections n’intervient de
toute manière pas dans l’évolution des symptômes pour les 17 jours qui suivent la date
d’estimation du rendement, puisque une nouvelle infection nécessite ~17 jours de latence (Figure
29 et 30) avant que puisse être observés des symptômes affectant le pourcentage de surface foliaire
verte.
Figure 29 : Schéma temporel du déroulement des estimations de rendement.
Le modèle PROCULTURE est utilisé afin de déterminer l’évolution du pourcentage de surface
verte des trois dernières strates en post estimation (Figure 29 et Figure 28-Etape 1 et 0). Ce
modèle épidémiologique contient une équation, présentée ci-dessous, qui modélise le
développement futur des symptômes de la septoriose, principale maladie responsable de la
dégradation de la surface verte du blé d’hiver au Grand-Duché de Luxembourg. La validation du
module d’estimation de la sévérité de la maladie par ce modèle a été effectuée au Grand-Duché de
Luxembourg sur des cultivars sensibles et semi-sensibles à la septoriose (El Jarroudi et al., 2002).
L’estimation du comportement de la maladie détermine le pourcentage de surface foliaire couverte
par les symptômes de celle-ci et de la sénescence qu’elle induit au cours des semaines qui suivent
la date de simulation. Le pourcentage de surface verte des trois dernières strates est alors déterminé
pour ces semaines par soustraction du pourcentage de la maladie à 100% de surface foliaire.
Période d’estimation visuelle de la surface verte
Période d’estimation en prédiction de la surface verte (PROCULTURE)
Dernière infection prise en compte
Latence dernière infection (14 à 17jours)
temps stades GS39 1er Date d’estimation des
rendements. (floraison en 2004 et 2008)
maturité

116
y = b / (e(abc) e(-abx))
Où y représente le pourcentage de la surface foliaire infectée.
X représente la somme de degrés jour depuis la fin de la période de latence (détermination sur base de
données historiques).
a représente le paramètre du rythme.
b représente le maximum de pourcentage de surface malade.
c est une constante
Les paramètres varient en fonction de l’âge de la feuille et de la variabilité de l’inoculum durant la
période d’infection. Ces variations sont estimées sur base de tables (Lemaire et al., 2003). Si
différentes périodes d’infection apparaissent sous différentes conditions sur une même feuille, les
courbes correspondantes sont ajoutées. Ce modèle est ajustable en cours de saison par des
observations au champ (Lemaire et al., 2003). Les données de températures utilisées pour la
simulation de la période post-estimation sont issues d’une base de données de 10 ans. L’évolution
du pourcentage de surface verte connue, la matière sèche peut alors être calculée pour chaque date
à laquelle le pourcentage de surface verte a été obtenu jusqu’à ce que le nombre de valeurs de
matière sèche suffise à l’ajustement d’une courbe. Les paramètres « a » et « i » de la courbe sont
obtenus et le rendement correspondant peut alors être estimé au moyen des modèles d’estimation
des rendements de la PARTIE II (Figure 28-étape 6). L’estimation du rendement peut-être
entreprise dès l’épiaison et peut-être renouvelée régulièrement, en réajustant le modèle
PROCULTURE. Dans notre cas, le nombre d’épis/m² a été obtenu à la floraison, raison pour
laquelle la première estimation du rendement a lieu à cette date (Figure 29) en 2004 et 2008. Des
nouvelles estimations du rendement ont ensuite été menées chaque semaine, en recalant les
données de %SV de pré-estimation introduites dans Proculture à chaque reprise.
Résultats et discussion.
La date la plus précoce à laquelle une estimation des rendements a été entreprise est la floraison.
Trois autres estimations ont ensuite été testées à trois semaines consécutives uniquement en 2008
(Floraison le 2 Juin, 8, 16 et 29 Juin). Les données considérées nécessaires au calcul de la matière
sèche sont connues avant la date de la floraison pour la première estimation des rendements
(données de pré-estimation). Après la date d’estimation des rendements, les variables
« température journalière », « rayonnement journalier » et « %SV des 3 dernières strates » ont été
obtenues, respectivement, à partir de la base de données historiques de température de Proculture,

117
la moyenne journalière (en jours juliens) des 8 dernières années et les simulations d’un module de
PROCULTURE.
PROCULTURE a effectué une simulation de la progression des symptômes de la septoriose pour
toute la saison de culture à partir de l’introduction dans le modèle du pourcentage de formation de
la F3 et du pourcentage de surface foliaire couverte par les symptômes de la septoriose sur la F5 à
la date d’estimation des rendements. En plus de la progression des symptômes présents, ce modèle
a permis de déterminer les infections des différents étages foliaires en fonction de conditions
météorologiques précises. Ces conditions d’infection se basent sur la connaissance des différentes
étapes du cycle de la septoriose (description au dernier chapitre de l’introduction générale de la
PARTIE I).
La Figure 30 décrit, pour l’année 2008, les conditions météorologiques de précipitation et de
température (A) qui étaient favorables aux nouvelles infections (B) en fonction de la connaissance
de l’inoculum présent et de la simulation du développement de la maladie. Les infections qui ont
pu être prises en compte pour l’estimation de la surface verte nécessaire à l’estimation des
rendements effectuée le 08 Juin et le 16 Juin sont entourées (respectivement le cercle de gauche et
le cercle de droite). A la date de la première estimation des rendements du 02 Juin, aucune de ces
infections n’a pu être prise en compte car elles ont eu lieu après cette date. La période de latence
qui suit chaque infection (graphique C pour la simulation au 2 Juin, D pour la simulation au 8 Juin
et E pour la simulation au 16 Juin) est calculée à partir d’une équation polynomiale qui tient
compte de la température (Shaw, 1990). Elle est représentée en ordonnée de ces graphiques, en
fraction accomplie par jour du délai de latence. Lorsque la somme des fractions journalières est
égale à 1, la latence est terminée (Rapilly, 1991). A chaque fin de latence succède l’apparition de
nouveaux symptômes. Le développement des symptômes est ensuite simulé à partir de la
connaissance des infections présentes à chaque date d’estimation des rendements. Cette simulation
est effectuée à l’aide de l’équation du module de Proculture présentée en matériel et méthodes. Ce
module utilise des données historiques de température. L’évolution simulée des symptômes est
présentée en partie C de la Figure 30 pour la variété Rosario en 2008. Le développement de
chaque feuille, de la F5 à la F1, en fonction des phyllothermes (130 degrés jours) est représenté par
une courbe allant de 0 à 100% pour chaque feuille. Une ligne horizontale marque le début du
développement de la feuille suivante.
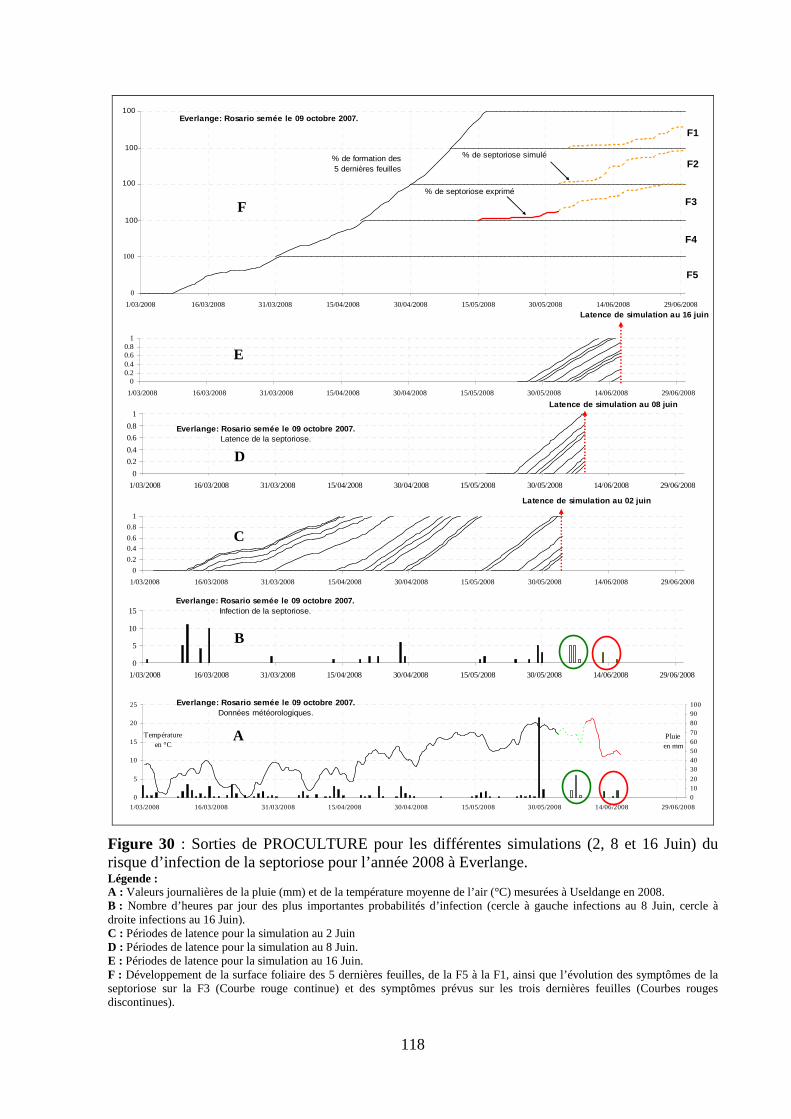
118
0
5
10
15
1/03/2008 16/03/2008 31/03/2008 15/04/2008 30/04/2008 15/05/2008 30/05/2008 14/06/2008 29/06/2008
0
5
10
15
20
25
1/03/2008 16/03/2008 31/03/2008 15/04/2008 30/04/2008 15/05/2008 30/05/2008 14/06/2008 29/06/2008
01020
3040506070
8090100Everlange: Rosario semée le 09 octobre 2007.
Données météorologiques.
Pluie en mm
Températureen °C
0
0.2
0.4
0.6
0.8
1
1/03/2008 16/03/2008 31/03/2008 15/04/2008 30/04/2008 15/05/2008 30/05/2008 14/06/2008 29/06/2008
0
0.2
0.4
0.6
0.8
1
1/03/2008 16/03/2008 31/03/2008 15/04/2008 30/04/2008 15/05/2008 30/05/2008 14/06/2008 29/06/2008
Everlange: Rosario semée le 09 octobre 2007.Latence de la septoriose.
00.20.40.60.8
1
1/03/2008 16/03/2008 31/03/2008 15/04/2008 30/04/2008 15/05/2008 30/05/2008 14/06/2008 29/06/2008
0
100
200
300
400
500
1/03/2008 16/03/2008 31/03/2008 15/04/2008 30/04/2008 15/05/2008 30/05/2008 14/06/2008 29/06/2008
F4
F5
F2
F3
F1
100
100
100
100
Everlange: Rosario semée le 09 octobre 2007.
% de formation des 5 dernières feuilles
% de septoriose exprimé
% de septoriose simulé
Latence de simulation au 02 juin
Latence de simulation au 08 juin
Latence de simulation au 16 juin
D
F
B
A
C
E
Everlange: Rosario semée le 09 octobre 2007.Infection de la septoriose.
Figure 30 : Sorties de PROCULTURE pour les différentes simulations (2, 8 et 16 Juin) du risque d’infection de la septoriose pour l’année 2008 à Everlange. Légende : A : Valeurs journalières de la pluie (mm) et de la température moyenne de l’air (°C) mesurées à Useldange en 2008. B : Nombre d’heures par jour des plus importantes probabilités d’infection (cercle à gauche infections au 8 Juin, cercle à droite infections au 16 Juin). C : Périodes de latence pour la simulation au 2 Juin D : Périodes de latence pour la simulation au 8 Juin. E : Périodes de latence pour la simulation au 16 Juin. F : Développement de la surface foliaire des 5 dernières feuilles, de la F5 à la F1, ainsi que l’évolution des symptômes de la septoriose sur la F3 (Courbe rouge continue) et des symptômes prévus sur les trois dernières feuilles (Courbes rouges discontinues).

119
A partir de la F3, l’évolution des symptômes de la septoriose est représentée, en pourcentage de
surface couverte, par une courbe rouge continue lorsqu’il s’agit de données réelles, et par une
courbe discontinue lorsqu’il s’agit de l’évolution des symptômes obtenue à partir des données
historiques de température. La connaissance de l’évolution des pourcentages de surface foliaire
couverte par la septoriose a permis, aussi bien en 2004 qu’en 2008, de déterminer le %SV des trois
dernières strates en post-estimation des rendements. Le %SV étant obtenu par soustraction du
pourcentage de surface couverte par la septoriose à 100% de surface foliaire. Les taux d’infection
de la septoriose en 2004 étaient très faibles. En conséquence, et étant donné les conditions
climatiques favorables au maintien de la surface verte de cette année-là, seule la surface verte de la
F3 a décru progressivement de la floraison à la maturité. Ces conditions exceptionnelles n’ont pas
rendu possible l’utilisation des F1 et F2 qui ont présenté un %SV constamment à 100%.
L’estimation des rendements à la floraison a tout de même été testée en utilisant uniquement le
%SV de la F3. La courbe de dégradation de la surface verte de la F3, obtenue avec les données
simulées de l’évolution des symptômes par Proculture à partir de la floraison, est très proche de la
courbe obtenue avec les données réelles en 2004 (Figure 31-C). Une proximité entre les courbes
simulées et les courbes obtenues avec les données réelles en 2008 est également observée, pour
chaque date de simulation et chaque strate foliaire des variétés Rosario et Tommi (Figure 31-A et
31-B). Les %SV pour une même date entre la simulation et la courbe réelle ne s’éloignent jamais
de plus de 10%, à l’exception des courbes de la F2 de Rosario. La comparaison des Figures 31-A
et 31-B montre que la variété Rosario présente une dégradation de la surface verte initiée
approximativement une semaine avant la variété Tommi.
Comme cela a été expliqué pour la Figure 30, le %SV a dû être recalé pour chacune des trois
estimations du rendement qui ont suivi celle de la floraison (2 juin). Une nouvelle simulation de
Proculture commençant à chaque nouvelle date d’estimation a permis de recalculer les %SV à
venir en fonction des infections et symptômes connus à la nouvelle date d’estimation des
rendements. La dégradation de la surface verte présentée dans les Figure 31-A et 31-B a été
utilisée dans le calcul de la matière sèche effectué pour la première estimation effectuée à la
floraison (02/06/08 et 10/06/04). La dégradation de la surface verte et de la matière sèche
journalière a ensuite été obtenue pour les deux variétés aux dates d’estimation du 08, 16 et 29 Juin
2008.
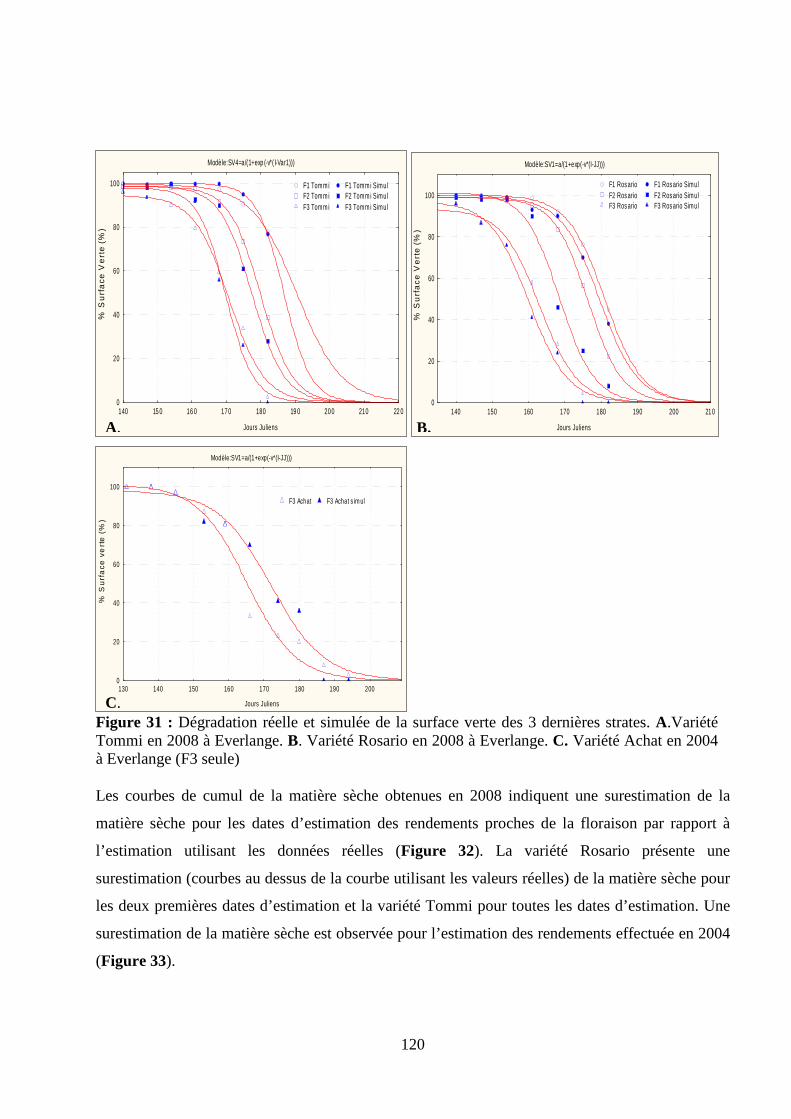
120
Modèle:SV4=a/(1+exp (-v*( I-Var1)))
140 15 0 16 0 170 180 19 0 200 210 22 0
Jours Juliens
0
20
40
60
80
100
% S
urf
ace
Ve
rte
(%
)
F1 Tomm i F1 Tomm i SimulF2 Tomm i F2 Tomm i Simul F3 Tomm i F3 Tomm i Simul
Modèle:SV1=a/(1+exp(-v*(I-JJ)))
140 150 160 170 180 190 200 21 0
Jours Juliens
0
20
40
60
80
100
% S
urf
ac
e V
ert
e (
%)
F1 Rosario F1 Rosario Simul F2 Rosario F2 Rosario SimulF3 Rosario F3 Rosario Simul
Modèle:SV1=a/(1+exp(-v*(I-JJ)))
130 140 150 160 170 180 190 200
Jours Juliens
0
20
40
60
80
100
% S
urf
ace
ve
rte
(%
)
F3 Achat F3 Achat s im ul
Figure 31 : Dégradation réelle et simulée de la surface verte des 3 dernières strates. A.Variété Tommi en 2008 à Everlange. B. Variété Rosario en 2008 à Everlange. C. Variété Achat en 2004 à Everlange (F3 seule) Les courbes de cumul de la matière sèche obtenues en 2008 indiquent une surestimation de la
matière sèche pour les dates d’estimation des rendements proches de la floraison par rapport à
l’estimation utilisant les données réelles (Figure 32). La variété Rosario présente une
surestimation (courbes au dessus de la courbe utilisant les valeurs réelles) de la matière sèche pour
les deux premières dates d’estimation et la variété Tommi pour toutes les dates d’estimation. Une
surestimation de la matière sèche est observée pour l’estimation des rendements effectuée en 2004
(Figure 33).
A. B.
C.

121
Les courbes de cumul de la matière sèche obtenues montrent également des différences entre les
variétés. La variété Rosario a fourni des courbes plus proches de celles obtenues avec les valeurs
réelles que la variété Tommi. Il est également constaté que plus la courbe est estimée tard dans la
saison, plus elle se rapproche de la courbe obtenue avec les données réelles de toute la saison (16
Juin par rapport à 02 Juin).
Les courbes de cumul de la matière sèche de 2004 présentent des valeurs de très loin inférieures à
celles de 2008, car le calcul de la matière sèche n’est effectué que sur une feuille (Figure 33).
Seule la variété Achat est étudiée cette année-là. La courbe de cumul obtenue avec les données
réelles présente des valeurs inférieures à celles de la courbe obtenue avec les données estimées.
Modèle:MS1=a/(1+exp(v*(I-dj)))
0 200 400 600 80 0 1000
degré jours (°C)
0
100
200
300
400
500
600
Cu
mu
l M
ati
ère
Sè
ch
e(g
/m²)
Rosa réelles Rosa 02/06 Rosa 08/06 *Rosa 16/06 Rosa 29/06
Tommi réelles Tommi 02/06 Tommi 08/06 + Tommi 16/06 Tommi 29/06
Figure 32 : Ajustement du cumul de la matière sèche aux valeurs réelles et aux différentes simulations de %SV en 2008.

122
Modèle:Nv_Var6=a/(1+exp(v*(I-Nv_Var5)))
0 200 400 600 800 1000
degré jours (°C)
0
10
20
30
40
50
Cu
mu
l M
ati
ère
Sè
ch
e (
g/m
²)
Simulation à la floraison Achat - 2004Cumul MS Achat données réelles -2004
Figure 33 : Ajustement du cumul de la matière sèche aux valeurs réelles et aux différentes simulations de %SV en 2004.
Il semblerait, au vu des valeurs de rayonnement journalier (Figure 34), que le rayonnement puisse
être à l’origine des surestimations de la matière sèche lorsque la date d’estimation est proche de la
floraison. En effet, le rayonnement journalier réel d’une année (2004, 2008 ou toute autre année)
pris isolément est très variable d’un jour à l’autre. Les pics de rayonnement ne se produisent pas
les mêmes jours juliens d’une année à l’autre. En conséquence, la moyenne de plusieurs années
pour chaque rayonnement moyen journalier fournit des valeurs journalières influencées par les pics
de chaque année. Cet effet est très bien visualisable à la Figure 34, sur laquelle est représenté le
rayonnement journalier observé de 2004 et 2008 par rapport au rayonnement journalier obtenu sur
base de la moyenne des années 2000 à 2007. Lorsque le rayonnement journalier moyen de 8
années est utilisé pour les jours de post-estimation de 2008 ou 2004, le rayonnement journalier est
de 2 à 3 fois supérieur, pour la plupart des jours, au rayonnement journalier réel de l’année en
cours. L’utilisation de ce rayonnement pour chaque jour post-estimation de la matière sèche amène
dans la plupart des cas à une surestimation de la matière sèche journalière produite. Cet effet
explique pourquoi la courbe de cumul de la matière sèche est d’autant plus surestimée qu’elle
utilise un grand nombre de jours avec un rayonnement issu de la moyenne historique. C’est donc
l’estimation faite à la floraison qui fournit des valeurs de matière sèche journalière les plus
surestimées, et celle faite le plus proche de la récolte qui fournit des valeurs de matière sèche
journalière les moins surestimées (Figure 32). Il peut aussi être mentionné que le rayonnement des
années 2004 et 2008 est exceptionnellement bas par rapport à celui des années utilisées pour le
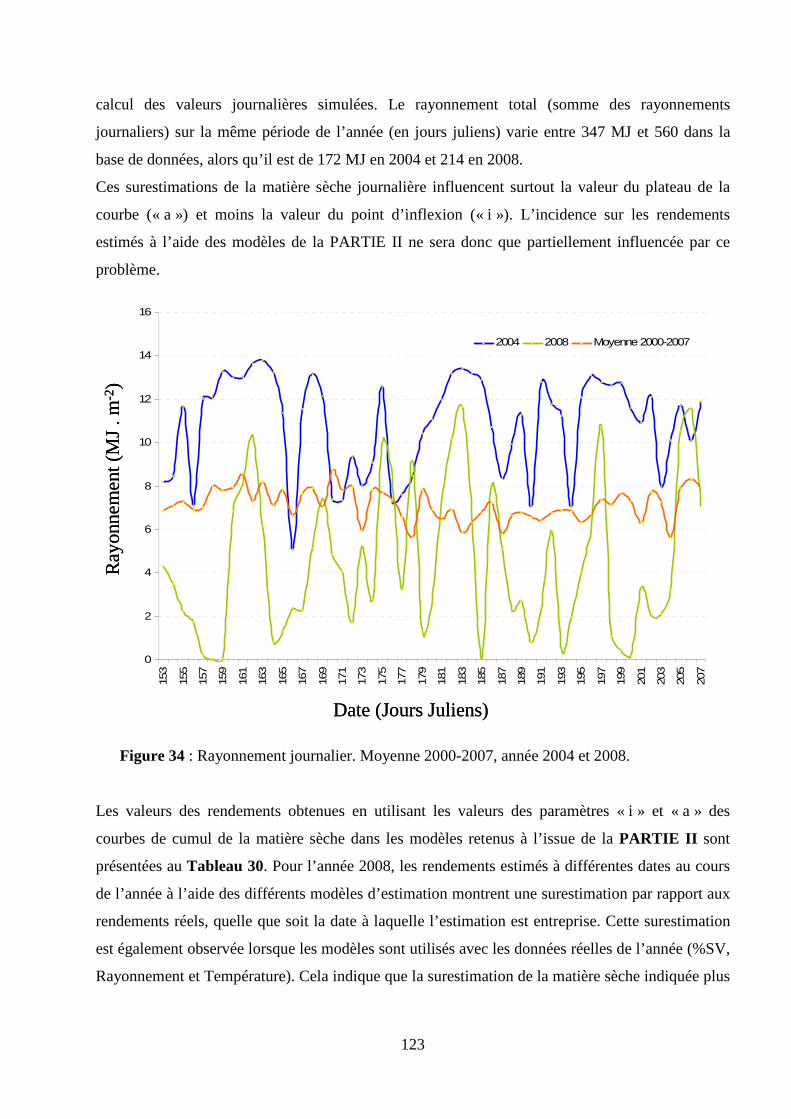
123
calcul des valeurs journalières simulées. Le rayonnement total (somme des rayonnements
journaliers) sur la même période de l’année (en jours juliens) varie entre 347 MJ et 560 dans la
base de données, alors qu’il est de 172 MJ en 2004 et 214 en 2008.
Ces surestimations de la matière sèche journalière influencent surtout la valeur du plateau de la
courbe (« a ») et moins la valeur du point d’inflexion (« i »). L’incidence sur les rendements
estimés à l’aide des modèles de la PARTIE II ne sera donc que partiellement influencée par ce
problème.
Figure 34 : Rayonnement journalier. Moyenne 2000-2007, année 2004 et 2008.
Les valeurs des rendements obtenues en utilisant les valeurs des paramètres « i » et « a » des
courbes de cumul de la matière sèche dans les modèles retenus à l’issue de la PARTIE II sont
présentées au Tableau 30. Pour l’année 2008, les rendements estimés à différentes dates au cours
de l’année à l’aide des différents modèles d’estimation montrent une surestimation par rapport aux
rendements réels, quelle que soit la date à laquelle l’estimation est entreprise. Cette surestimation
est également observée lorsque les modèles sont utilisés avec les données réelles de l’année (%SV,
Rayonnement et Température). Cela indique que la surestimation de la matière sèche indiquée plus
Date (Jours Juliens)
0
2
4
6
8
10
12
14
16
153
155
157
159
161
163
165
167
169
171
173
175
177
179
181
183
185
187
189
191
193
195
197
199
201
203
205
207
2004 2008 Moyenne 2000-2007
Ray
onne
me
nt (
MJ
. m-2 )
Date (Jours Juliens)
0
2
4
6
8
10
12
14
16
153
155
157
159
161
163
165
167
169
171
173
175
177
179
181
183
185
187
189
191
193
195
197
199
201
203
205
207
2004 2008 Moyenne 2000-2007
Ray
onne
me
nt (
MJ
. m-2 )
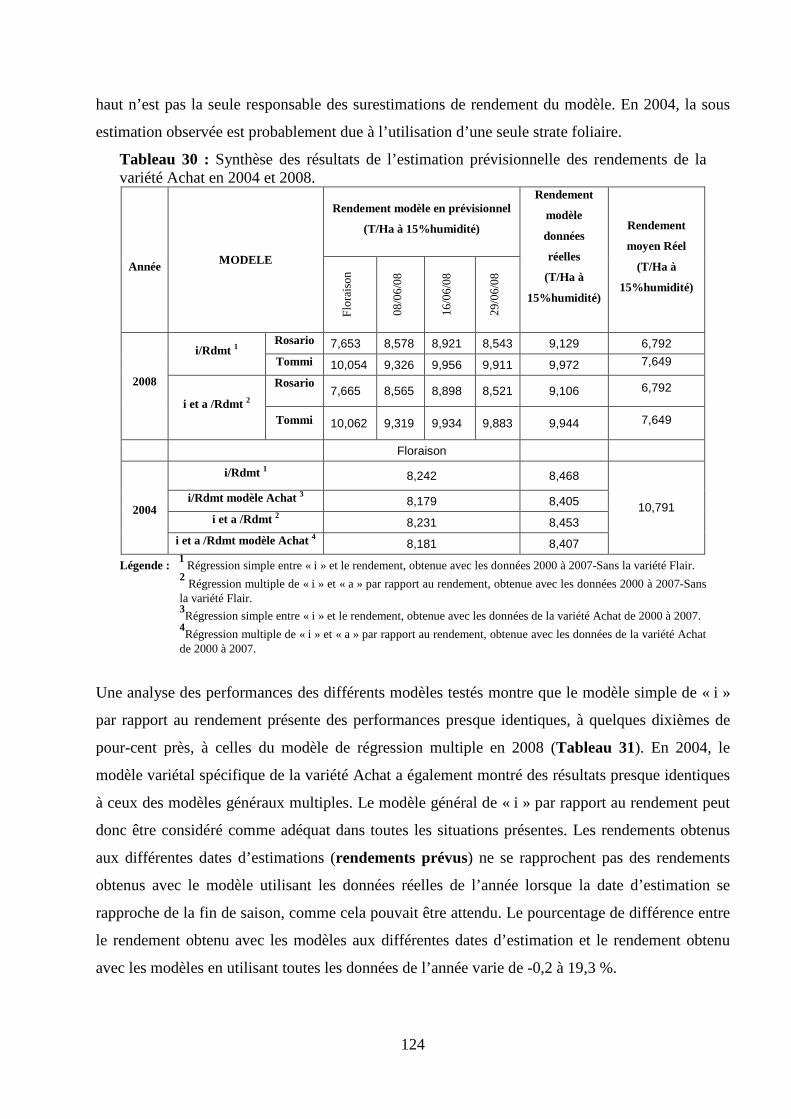
124
haut n’est pas la seule responsable des surestimations de rendement du modèle. En 2004, la sous
estimation observée est probablement due à l’utilisation d’une seule strate foliaire.
Tableau 30 : Synthèse des résultats de l’estimation prévisionnelle des rendements de la variété Achat en 2004 et 2008.
Rendement modèle en prévisionnel
(T/Ha à 15%humidité)
Année MODELE
Flo
rais
on
08
/06
/08
16
/06
/08
29
/06
/08
Rendement
modèle
données
réelles
(T/Ha à
15%humidité)
Rendement
moyen Réel
(T/Ha à
15%humidité)
Rosario 7,653 8,578 8,921 8,543 9,129 6,792 i/Rdmt 1
Tommi 10,054 9,326 9,956 9,911 9,972 7,649
Rosario 7,665 8,565 8,898 8,521 9,106 6,792
2008
i et a /Rdmt 2
Tommi 10,062 9,319 9,934 9,883 9,944 7,649
Floraison
i/Rdmt 1 8,242 8,468
i/Rdmt modèle Achat 3 8,179 8,405
i et a /Rdmt 2 8,231 8,453
2004
i et a /Rdmt modèle Achat 4 8,181 8,407
10,791
Légende : 1
Régression simple entre « i » et le rendement, obtenue avec les données 2000 à 2007-Sans la variété Flair. 2 Régression multiple de « i » et « a » par rapport au rendement, obtenue avec les données 2000 à 2007-Sans
la variété Flair. 3Régression simple entre « i » et le rendement, obtenue avec les données de la variété Achat de 2000 à 2007.
4Régression multiple de « i » et « a » par rapport au rendement, obtenue avec les données de la variété Achat
de 2000 à 2007.
Une analyse des performances des différents modèles testés montre que le modèle simple de « i »
par rapport au rendement présente des performances presque identiques, à quelques dixièmes de
pour-cent près, à celles du modèle de régression multiple en 2008 (Tableau 31). En 2004, le
modèle variétal spécifique de la variété Achat a également montré des résultats presque identiques
à ceux des modèles généraux multiples. Le modèle général de « i » par rapport au rendement peut
donc être considéré comme adéquat dans toutes les situations présentes. Les rendements obtenus
aux différentes dates d’estimations (rendements prévus) ne se rapprochent pas des rendements
obtenus avec le modèle utilisant les données réelles de l’année lorsque la date d’estimation se
rapproche de la fin de saison, comme cela pouvait être attendu. Le pourcentage de différence entre
le rendement obtenu avec les modèles aux différentes dates d’estimation et le rendement obtenu
avec les modèles en utilisant toutes les données de l’année varie de -0,2 à 19,3 %.
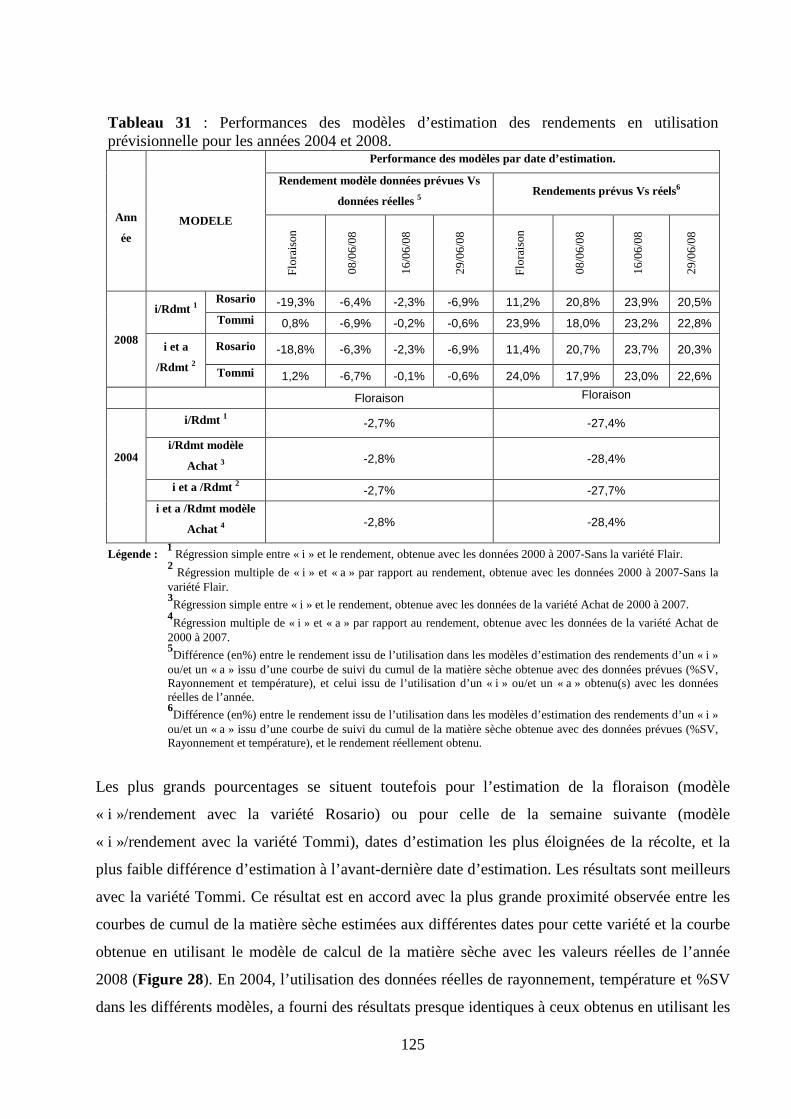
125
Tableau 31 : Performances des modèles d’estimation des rendements en utilisation prévisionnelle pour les années 2004 et 2008.
Performance des modèles par date d’estimation.
Rendement modèle données prévues Vs
données réelles 5 Rendements prévus Vs réels6
Ann
ée MODELE
Flo
rais
on
08
/06
/08
16
/06
/08
29
/06
/08
Flo
rais
on
08
/06
/08
16
/06
/08
29
/06
/08
Rosario -19,3% -6,4% -2,3% -6,9% 11,2% 20,8% 23,9% 20,5% i/Rdmt 1
Tommi 0,8% -6,9% -0,2% -0,6% 23,9% 18,0% 23,2% 22,8%
Rosario -18,8% -6,3% -2,3% -6,9% 11,4% 20,7% 23,7% 20,3%
2008 i et a
/Rdmt 2 Tommi 1,2% -6,7% -0,1% -0,6% 24,0% 17,9% 23,0% 22,6%
Floraison Floraison
i/Rdmt 1 -2,7% -27,4%
i/Rdmt modèle
Achat 3 -2,8% -28,4%
i et a /Rdmt 2 -2,7% -27,7%
2004
i et a /Rdmt modèle
Achat 4 -2,8% -28,4%
Légende : 1
Régression simple entre « i » et le rendement, obtenue avec les données 2000 à 2007-Sans la variété Flair. 2 Régression multiple de « i » et « a » par rapport au rendement, obtenue avec les données 2000 à 2007-Sans la
variété Flair. 3Régression simple entre « i » et le rendement, obtenue avec les données de la variété Achat de 2000 à 2007.
4Régression multiple de « i » et « a » par rapport au rendement, obtenue avec les données de la variété Achat de
2000 à 2007. 5Différence (en%) entre le rendement issu de l’utilisation dans les modèles d’estimation des rendements d’un « i »
ou/et un « a » issu d’une courbe de suivi du cumul de la matière sèche obtenue avec des données prévues (%SV, Rayonnement et température), et celui issu de l’utilisation d’un « i » ou/et un « a » obtenu(s) avec les données réelles de l’année. 6Différence (en%) entre le rendement issu de l’utilisation dans les modèles d’estimation des rendements d’un « i »
ou/et un « a » issu d’une courbe de suivi du cumul de la matière sèche obtenue avec des données prévues (%SV, Rayonnement et température), et le rendement réellement obtenu.
Les plus grands pourcentages se situent toutefois pour l’estimation de la floraison (modèle
« i »/rendement avec la variété Rosario) ou pour celle de la semaine suivante (modèle
« i »/rendement avec la variété Tommi), dates d’estimation les plus éloignées de la récolte, et la
plus faible différence d’estimation à l’avant-dernière date d’estimation. Les résultats sont meilleurs
avec la variété Tommi. Ce résultat est en accord avec la plus grande proximité observée entre les
courbes de cumul de la matière sèche estimées aux différentes dates pour cette variété et la courbe
obtenue en utilisant le modèle de calcul de la matière sèche avec les valeurs réelles de l’année
2008 (Figure 28). En 2004, l’utilisation des données réelles de rayonnement, température et %SV
dans les différents modèles, a fourni des résultats presque identiques à ceux obtenus en utilisant les

126
données estimées de façon prévisionnelle à la floraison avec ces modèles (moins de 3% de
différence).
Les rendements prévus par les modèles et les rendements réels montrent des différences allant
de ~11% à 24%, avec une surestimation des rendements pour les deux variétés utilisées en 2008.
La meilleure performance de 11% est observée pour l’utilisation du modèle « i » /rendement à la
floraison sur la variété Rosario. C’est également à la floraison que l’estimation du modèle en
prévisionnel est la plus éloignée de l’estimation faite avec les données réelles. Il peut donc être
estimé, pour cette raison, qu’elle n’est pas fiable. L’ensemble des autres estimations du rendement
ont des performances très proches des 20% de différences avec les rendements réels, que
l’estimation du rendement soit ou non effectuée proche de la récolte.
En 2004, les différents modèles sont proches entre eux et présentent tous des rendements de 27% à
28% inférieurs aux rendements réels obtenus.
Conclusion.
Les modèles obtenus à l’issue de la PARTIE II ont été testés de manière prévisionnelle sur la
variété Achat en 2004, ainsi que sur la variété Rosario et la variété Tommi en 2008. L’utilisation
de ces modèles de manière prévisionnelle nécessite la connaissance de la température, du
rayonnement et du %SV pour chaque jour s’écoulant entre la date d’estimation des rendements et
la maturité (post-estimation). Le %SV a pu être obtenu, pour les jours suivant la date d’estimation
des rendements (post-estimation) grâce à la connaissance des infections en cours et à la
modélisation de l’évolution des symptômes de la septoriose. Cette modélisation a été effectuée sur
les trois dernières strates à l’aide d’un module du modèle Proculture. Ce module, qui utilise une
base de données historique de température, a permis d’obtenir des courbes de dégradation de la
surface verte très proches des courbes obtenues avec les données réelles pour les 3 strates des deux
variétés présentes en 2008 (écart <10% entre données réelles de %SV et %SV estimés sur 5 des
2x3 strates des variétés Rosario et Tommi). Il a également été constaté en comparant les courbes
de dégradation de la surface verte que la variété Rosario a initié la dégradation de la surface verte
de chaque strate, environ 1 semaine avant Tommi. En 2004, seule la F3 a pu être utilisée car les
deux premières strates présentaient un %SV à 100% durant toute la saison. Pour cette année-là, les
écarts entre la dégradation de la surface verte réelle et simulée étaient également faibles.
L’utilisation des %SV obtenus ainsi que du rayonnement journalier issu de la moyenne journalière,
en jours juliens, de 2000 à 2007 ont permis ensuite de calculer la matière sèche journalière des
jours post-estimation. Les courbes de cumul de la matière sèche obtenues pour chaque variété, en

127
utilisant les données de rayonnement, température et %SV prédites à partir de 4 dates d’estimation
des rendements différentes, et espacées d’une semaine à partir de la floraison, étaient presque
toutes surestimées par rapport aux mêmes courbes obtenues avec les données réelles. Les valeurs
du rayonnement moyen journalier issues d’une moyenne historique sont responsables de ces
surestimations, celles-ci étant presque pour chaque jour supérieures aux valeurs de rayonnement
réel du jour considéré en 2008 et 2004. La présence de surestimations plus importantes de la
matière sèche pour les courbes effectuées aux dates d’estimation des rendements proches de la
floraison est également expliquée par l’utilisation de ce rayonnement, puisque plus l’estimation est
effectuée tôt dans la saison, plus elle utilisera un grand nombre de valeurs de rayonnement issues
de la moyenne historique.
L’utilisation des paramètres « i » et « a » des courbes de cumul de la matière sèche avec les
différents modèles d’estimation des rendements permet de conclure en l’obtention de modèles
fournissant des estimations aux alentours de 20% supérieures aux valeurs de rendement réelles (à
l’exception de l’estimation à la floraison qui est de 11% supérieure) en 2008. En 2004, l’utilisation
de la F3 seule a fourni des valeurs de rendement aux alentours de 28% inférieures aux valeurs
réelles. Cette sous-estimation est expliquée par l’utilisation des trois strates pour la construction
des modèles d’estimation des rendements, alors que ces modèles sont utilisés avec une strate en
2004. Les différents modèles fournissent des résultats très semblables, ce qui était prévisible,
compte tenu de la proximité entre leur valeur de pente et d’intercept. Enfin, la proximité entre les
résultats des modèles simples et multiples incite à l’utilisation de modèles simples.
Avec ces modèles, à chaque valeur de «i » correspond un rendement en considérant que ce « i » est
issu d’une courbe de cumul de la matière sèche obtenue avec trois strates, et non un « i » de valeur
peu élevée obtenu avec la F3 seule. Un modèle construit uniquement avec la F3 des 7 années aurait
probablement fourni en 2004 de meilleurs résultats, plus proches de ceux obtenus en 2008.
Une faible différence entre les erreurs d’estimation des rendements en prévisionnel et en non
prévisionnel des modèles est observée dans la plupart des cas. Cette faible différence d’erreurs
commises entre le prévisionnel et le non prévisionnel montre que l’utilisation en prévisionnel selon
la méthode exposée fonctionne bien. De plus, ceci nous amène à la conclusion qu’une grande
partie des erreurs commises sont issues des étapes précédant la partie prévisionnelle.
L’amélioration des performances de ces modèles passe donc par une estimation correcte des
variables et paramètres qui ne sont pas à prévoir mais qui sont obtenus avant la date d’estimation
des rendements. Une amélioration peut être attendue par une connaissance du nombre de brins
moyens et de maîtres brins de l’année en cours, ainsi que par une estimation à la floraison de la
surface foliaire de la variété considérée pour l’année en cours.

128
D’autres améliorations sont également possibles par l’utilisation de données de meilleure qualité
pour la construction des modèles. En effet, il est probable que de meilleures performances soient
obtenues si la qualité des données utilisées pour construire les modèles ne nous amène pas à
émettre des hypothèses introduisant des biais, comme cela a été le cas pour pouvoir utiliser les
données de 2000 à 2005.
Enfin, une adaptation concernant l’utilisation du modèle PROCULTURE à la simulation de
l’évolution du pourcentage de surface verte des 3 dernières strates de variétés traitées pourrait être
envisagée. Ce modèle se base sur le niveau d’infection de ces 3 strates par la septoriose à la date
d’estimation des rendements afin de simuler l’évolution des symptômes uniquement sur le témoin
non traité. Dans le cas de parcelles traitées, les infections, les symptômes et la réduction de la
surface verte évoluent moins vite. Une adaptation du modèle permettrait de déterminer ces
évolutions en fonction du nombre et du type de traitement. L’évolution du pourcentage de surface
verte, le nombre de brins par mètre carré et la surface foliaire seraient alors pris en compte en
fonction des traitements. Ces données sont indispensables pour effectuer des estimations de
rendement différentes d’un traitement à l’autre.

129
DISCUSSION GENERALE ET PERSPECTIVES

130
Contexte et domaine de l’étude. L’étude présentée dans ce document s’inscrit dans un contexte de préservation de l’environnement
et de maximisation des rendements. La connaissance des rendements pour différents cultivars et
traitements phytosanitaires en cours de saison peut, en complément d’avertissements
phytosanitaires, amener l’agriculteur à raisonner l’application de produits fongicides afin de
maximiser son rendement économique. En outre, c’est par sa contribution à l’amélioration de ce
système d’avertissement que ce travail peut participer à la préservation de l’environnement ainsi
qu’à la préservation de la qualité des eaux de ruissellement et d’infiltration.
L’estimation des rendements est abordée ici par la modélisation suivant une approche mécaniste
qui a émergé d’études issues de la phytopathologie. Ce travail se situe cependant en marge de cette
discipline puisque les maladies des plantes et l’étude de leurs symptômes sont étudiées au travers
des conséquences de leur présence sur la réduction des surfaces foliaires photosynthétiquement
actives. L’interaction entre la plante et l’atmosphère, ainsi que l’impact de cette interaction sur les
rendements sont pris en compte au travers de l’intervention de données météorologiques dans les
modèles d’estimation des rendements considérés. Par ces aspects, il est considéré que ce travail se
situe dans le domaine de l’agrométéorologie.
Approches envisagées aux différentes étapes.
L’objectif de ce travail est de contribuer à la recherche d’un modèle fonctionnel d’estimation des
risques de pertes de récoltes pour différentes variétés et traitements du blé d’hiver au Grand-Duché
de Luxembourg. Cette contribution se veut exploratoire, tant par la recherche des techniques de
mesures à utiliser pour obtenir les données d’entrée des modèles testés que par la recherche du
modèle le plus adapté à l’utilisation de ces données pour l’estimation des rendements.
Afin d’atteindre ces objectifs, cette recherche s’est orientée vers l’étude des effets aisément
quantifiables au champ de la perturbation des processus photosynthétiques dont le produit est
impliqué dans le remplissage des grains. Les principaux organes photosynthétiques dont le produit
de l’activité est responsable du remplissage des grains sont les trois dernières feuilles (Gooding et
al., 2000). Leur activité peut être perturbée par des facteurs biotiques (pestes) ou abiotiques
(sécheresse). Les effets quantifiables des perturbations étudiées sont la réduction de la surface
verte des trois dernières feuilles. Dans le contexte topo climatique de la région étudiée, la cinétique
de dégradation de la surface verte est conditionnée par la sécheresse, ainsi que par le principal
complexe parasitaire Septoriose-Rouille brune.
Il ressort d’études précédentes que certains paramètres décrivant la courbe de décroissance du
pourcentage de surface verte foliaire peuvent expliquer dans certains cas (Bergamin Filho et al.,

131
1997) une grosse proportion des rendements obtenus. Il a également été mis en évidence qu’une
relation avec les rendements plus stables est obtenue, pour différents LAI, lorsque cette relation est
établie avec l’interception du rayonnement par la surface verte présente (Madden et al., 2007).
Lorsque ce travail a commencé, les études effectuées sur la relation entre le rendement et la
cinétique de dégradation de la surface verte du blé d’hiver étaient peu nombreuses (Bryson et al.,
1997 ; Gooding et al., 2000). Ces études ne prenaient pas en compte le rayonnement intercepté par
la surface verte au cours de sa dégradation. Des études avaient été menées dans ce sens sur
d’autres cultures, en présence d’autres complexes parasitaires (Waggoner et Berger, 1987 ;
Bergamin Filho et al., 1997). Les résultats de ces dernières ont motivé la recherche de la relation
entre le rendement et le rayonnement journalier intercepté par la surface verte du blé d’hiver. Cette
étude n’avait jamais été menée auparavant et aucune relation semblable n’avait été étudiée dans le
contexte topo-climatique de la région étudiée au Grand-Duché de Luxembourg. C’est pour ces
raisons que cette recherche a pour objectif de répondre à la question de la validité d’une telle
relation pour le blé d’hiver au Grand-Duché de Luxembourg.
Considérant les études précédentes menées sur la dégradation de la surface verte, il est à noter
qu’une grande diversité d’outils et de méthodes différentes ont été utilisées pour obtenir des
données permettant de caractériser cette dégradation. L’importance et la difficulté d’obtenir des
valeurs de LAI et de surfaces vertes proches de la réalité est à l’origine de certaines études de
comparaison entre différentes méthodes d’obtention de ces données (Brenner et al., 1995 ; Levy
and Jarvis, 1999). Pour le blé d’hiver, peu d’études de ce genre sont disponibles. De plus, dans
notre cas, au critère de qualité des données s’ajoute également la reproductibilité des mesures ainsi
que l’automatisation. Ces critères sont en effet cruciaux s’il est envisagé par la suite d’obtenir ces
données annuellement afin de faire fonctionner un modèle d’estimation des rendements de manière
routinière. Au regard de la bibliographie, aucune étude ne compare et ne met en évidence une
méthode précise, reproductible et automatisable pour l’obtention de la surface verte foliaire et du
LAI (PARTIE I - CHAPITRE II). Une première étape de ce travail a donc consisté à rechercher
une méthode rencontrant les critères recherchés et la possibilité d’obtenir une méthode
d’estimation à l’échelle d’un territoire. Deux méthodes d’estimation de la surface verte ont été
comparées, ainsi que la proximité entre les données issues de ces méthodes et les données issues
d’une méthode de référence, considérée comme indiquant des valeurs très proches de la réalité. La
même démarche a été mise en place pour l’obtention du LAI.
La recherche de la relation entre le rendement et le rayonnement intercepté par la surface verte tout
au long de la dégradation de la surface verte des trois dernières strates a été entreprise à partir,
entre autre, des données de LAI et de surface verte issues des méthodes ayant fourni les meilleurs

132
résultats. La bibliographie parcourue a fait ressortir deux modèles assez proches et adaptés à
l’approche envisagée (PARTIE I). Le plus récent des deux a été étudié sur le blé et prend en
compte l’ombrage des strates inférieures par les strates supérieures ainsi que le calcul de la
production journalière de matière sèche. Ces deux modèles ont été comparés par leurs résultats et
des possibilités de simplification et d’amélioration de leur utilisation ont été testées (PARTIE II).
Les différentes variantes du modèle fournissant les meilleurs résultats ont été retenues et validées.
Enfin, l’utilisation de ceux-ci en mode prévisionnel a été abordée et mise en œuvre sur deux
exemples. L’utilisation en mode prévisionnel implique l’obtention des données de surface verte et
de rayonnement de toute la saison en cours à la date à laquelle l’estimation du rendement est
envisagée. Pour ce faire, la méthode de simulation de la dégradation de la surface verte du modèle
PROCULTURE ainsi que la moyenne, pour les années disponibles, du rayonnement journalier de
chaque jour julien ont été utilisées.
Validité de l’étude.
L’étude a été menée sur des parcelles expérimentales mises en place au Grand-Duché de
Luxembourg, à Everlange. Le site est situé sur un plateau, sans ombrage. Le sol est de texture
argilo-sableuse. Ce travail ne prend donc pas en compte d’autres conditions topo-climatiques ayant
un effet sur la cinétique de dégradation de la surface verte.
Seules certaines variétés utilisées au Grand-Duché de Luxembourg ont été prises en compte. Les 3
variétés utilisées ont des caractéristiques différentes, dont certaines ont été observées dans cette
étude. Par exemple, les variétés Akteur, Achat et Flair n’ont pas un tallage identique et la variété
Flair est moins tolérante au stress hydrique (PARTIE II). Les modèles étudiés prennent en compte
certains paramètres qui varient d’une variété à l’autre. Pour cette raison, l’étude ne doit être
considérée que par rapport à ces variétés et doit être adaptée pour une éventuelle utilisation sur
d’autres variétés.
Seules la rouille brune et la septoriose ont été considérées. Ces deux maladies sont cependant les
plus présentes et les plus nuisibles sur le blé au Grand-Duché de Luxembourg. De plus, l’étude de
la dégradation de la surface verte prend en compte, de façon indirecte, l’ensemble des facteurs
biotiques et abiotiques qui perturbent les processus photosynthétiques de façon proportionnelle à la
surface verte dégradée.
Cette étude ne couvre pas l’ensemble des stress biotiques ou abiotiques ayant un effet sur le
rendement, par exemple par détournement des assimilats de la photosynthèse, sans en avoir sur la
cinétique de dégradation de la surface verte.

133
Les réserves des tiges, accumulées durant la préfloraison et remobilisées pour le remplissage des
grains ne sont pas prises en compte dans cette étude.
Les expérimentations ont été menées dans un cadre de fertilisation homogène, en prenant en
compte toutefois les carences engendrées par le précédent blé.
Les relations établies avec le rendement ne concernent que les aspects quantitatifs. Toute perte
qualitative de récolte n’est pas envisagée dans cette thèse.
Enfin, ce travail a été réalisé suivant différentes modalités de traitements, dont un traitement triple
procurant une protection presque totale contre la septoriose et la rouille brune. Les données
obtenues sont donc issues de couverts atteints à non atteints. Les données utilisées couvrent
l’ensemble des conditions de traitement qui peuvent être rencontrées sur blé d’hiver au Grand-
Duché de Luxembourg.
Synthèse des résultats issus de l’acquisition des données.
L’acquisition des données nécessaires à la construction des modèles d’estimation des rendements
considérés concerne essentiellement la surface verte et le LAI (PARTIE I).
La méthode d’obtention des surfaces foliaires vertes par analyse d’images a été choisie comme
méthode de référence. Elle a été validée avec succès à l’aide de la méthode du poids de papier,
considérée comme une méthode fournissant des données encore plus proches des valeurs réelles.
La méthode de référence a été comparée à la méthode d’estimation visuelle, plus simple et rapide
d’utilisation. Les résultats obtenus montrent une surestimation de la surface foliaire verte de la
méthode d’estimation visuelle par rapport à l’estimation par l’analyse d’images, surtout pour les
faibles pourcentages de surface verte. L’estimation visuelle fournit cependant des résultats
similaires à ceux de l’analyse d’images lorsque les moyennes sont comparées. Ces derniers
résultats justifient l’utilisation des estimations visuelles à partir de moyennes parcellaires (10
plantes par parcelle), dans la suite du travail, pour les années où seules celles-ci étaient
disponibles. En outre, l’étude des surfaces vertes issues de l’analyse d’images des deux années de
mesures effectuées durant ce travail de thèse a montré que les fongicides retardent l’arrivée de la
décroissance de la surface verte plutôt que de réduire son rythme. Cette observation confirme les
résultats obtenus par Gooding et al.(2000). Entre les deux années, la surface verte reste plus élevée
en 2006 qu’en 2007 durant toute la saison pour une même variété, sans engendrer une
augmentation de rendement correspondante. La cinétique de décroissance de la surface verte a été
étudiée de façon différente, par l’ajustement de deux types de courbes, une équation sigmoïde, et
une courbe de Gompertz simplifiée. Aucune différence variétale, de traitement phytosanitaire ou
de précédent cultural n’a été relevée entre les sigmoïdes, et aucune des relations (régression

134
linéaire et non linéaire) établies entre le plateau ou le point d’inflexion des sigmoïdes et le
rendement n’a fourni de résultats significatifs. Les valeurs du point d’inflexion des courbes de
Gompertz ont, par contre, permis de distinguer le témoin de la variante traitée trois fois sur les
données de 2006 et 2007. Une bonne corrélation (R=79%) a été obtenue entre les valeurs du point
d’inflexion des courbes de Gompertz et les rendements de 2006 et 2007. Ces conclusions
concernant l’utilisation de la courbe de Gompertz confirment les conclusions obtenues par
Gooding et al. (2000).
Afin de tester la possibilité d’obtenir des valeurs de surfaces vertes directement à une échelle
supérieure à celle de la feuille, un modèle linéaire simple a été obtenu entre la surface verte foliaire
obtenue par l’analyse d’images des trois dernières feuilles prises individuellement et la couverture
verte du sol obtenue par l’analyse d’images aériennes de l’apparition à la sénescence de ces trois
dernières feuilles. La relation explique 73% des données de surfaces vertes foliaires des trois
dernières strates à partir de données de couverture verte du sol. Il est probable que cette relation
puisse être améliorée et utilisée moyennant une meilleure discrimination des pixels de l’image de
la couverture verte du sol, ainsi qu’une standardisation des conditions (vent et luminosité) dans
lesquelles des images de couverture verte du sol sont prises.
La méthode d’analyse d’images a également permis d’obtenir des valeurs de surfaces foliaires très
proches des valeurs réelles. En effet, la comparaison des valeurs issues de cette méthode avec les
valeurs obtenues sur les mêmes feuilles à l’aide d’une méthode de référence montre une
correspondance à 95%. Ces bons résultats nous ont confortés dans l’utilisation des valeurs de
surfaces foliaires obtenues par l’analyse d’images pour le calcul du LAI.
Pour l’obtention du LAI, trois méthodes ayant chacune leurs avantages et inconvénients ont été
utilisées et comparées. La méthode de référence, utilisant entre autre les surfaces foliaires issues de
l’analyse d’images, a été comparée aux deux autres méthodes, qui permettent d’obtenir un LAI
plus rapidement et à plus grande échelle. Le LAI issu de la mesure du rayonnement intercepté par
le couvert (à l’aide du Sunscan meter) n’explique que 65 % du LAI obtenu avec la méthode de
référence. De plus, une surestimation du LAI a été observée lors de la première année de mesures,
suivie d’une sous-estimation la deuxième année. Les erreurs observées peuvent être expliquées par
différentes causes, en rapport avec la sensibilité de l’appareil utilisé, des dysfonctionnements
observés et son remplacement pour la seconde année de données. Le LAI, obtenu de manière
instantanée avec cet instrument, nécessite une validation supplémentaire avant d’être utilisé de
manière routinière.
Une autre méthode d’estimation du LAI, rapide et permettant de tester la possibilité de passer à la
détermination du LAI à l’échelle du territoire consiste à établir une relation entre le LAI et la

135
couverture verte du sol à la date du maximum de surface verte, obtenue par l’analyse d’images
aériennes prises à quelques mètres du sol. Il a été montré que la relation entre la couverture verte
du sol et le LAI n’était pas assez bonne pour être utilisée à des fins d’estimation du LAI. Parmi les
causes à l’origine de la mauvaise relation obtenue, il y a l’intensité du vent et l’intensité lumineuse
au niveau des strates inférieures, qui ne sont pas les mêmes à chaque prise d’image. D’autre part,
le LAI de référence est obtenu à partir des trois premières strates, alors que la couverture verte du
sol tient compte des strates plus basses.
Synthèse des résultats issus de la validation, la simplification et l’amélioration d’un modèle
d’estimation des rendements.
De tous les modèles d’estimation des rendements testés avec les données de 2006 et 2007, ce sont
les modèles issus de l’établissement de la relation entre le point d’inflexion « i » et/ou le plateau
«a » de la courbe décrivant l’évolution du cumul de la production journalière de matière sèche et le
rendement qui ont fourni les meilleurs résultats. Cette relation, étudiée suivant les différentes
modalités présentes ces deux années, montre qu’un effet variétal et un effet « année » est plausible.
Ceux-ci doivent être confirmés par l’utilisation d’un échantillon de données plus large.
Les simplifications menées sur les modèles étudiés n’ont pas permis de conclure en une possibilité
de réduction du nombre de strates foliaires, la suppression de strates fournissant, dans la plupart
des cas, des résultats moins bons. L’utilisation de la méthode d’estimation visuelle du pourcentage
de surface verte constitue également une simplification. En effet, son utilisation est plus simple et
plus rapide. La comparaison des résultats des modèles obtenus en utilisant les pourcentages de
surface verte issus de l’analyse d’images aux résultats obtenus en utilisant les pourcentages de
surface verte issus de l’estimation visuelle montre une surestimation lorsque sont utilisées les
estimations visuelles. Les paramètres « i » et « a » des modèles montrent une différence moyenne
de 8% entre les deux méthodes. Les coefficients de détermination des modèles de régression entre
ces deux paramètres et les rendements ont montré des différences de 20% dans la plupart des cas,
avec une significativité plus faible lorsque sont utilisées les estimations visuelles.
L’ajout aux données de 2006 et 2007 de données supplémentaires issues d’expérimentations
menées de 2000 à 2005 (à l’exception de 2004) a permis de confirmer l’effet variétal obtenu avec
les données de 2006 et 2007 seules. Une régression multiple a montré que les traitements
phytosanitaires n’expliquent pas les rendements, ce qui semble logique, compte tenu qu’ils
augmentent la durée de maintien de la surface verte et que la vitesse de dégradation de la surface
verte est prise en compte par les modèles. L’effet année est le plus important après « i » et « a »,

136
mais il est difficile dans ce cas-ci de le distinguer de la variété, certaines années ne comportant
qu’une variété.
Des modèles ont été construits par variété, avec toutes les données ou en supprimant certaines
données pour des raisons précises. Les modèles élaborés entre « i » et/ou le plateau «a » et le
rendement à partir des données de toutes les années par variété montrent des meilleurs résultats (R²
de 66 à 94%) que ceux élaborés à partir de l’ensemble des données (modèle général, R² de 37% ;
modèle général sans Flair, R² de 66%). La variété Flair n’a cependant pas fourni un modèle plus
performant que le modèle général. Elle a un comportement sensible au moindre stress hydrique, ce
qui pourrait expliquer les résultats moins performants.
Les validations croisées menées sur les meilleurs modèles (R²>50%) ont fourni des résultats
indiquant que ces modèles peuvent fournir des estimations des valeurs de rendement avec des taux
de réussite situés entre 87% et 96%, suivant le modèle considéré.
Synthèse des résultats issus de la proposition d’un modèle de simulation des risques de perte
de rendement.
L’exemple d’utilisation des modèles obtenus dans un mode prévisionnel a été testé sur deux
années qui n’ont pas été utilisées pour la construction des modèles (2004 et 2008). Les rendements
à venir ont été estimés à la floraison ainsi que durant les trois semaines suivantes à raison d’une
fois par semaine en 2008. Les valeurs de rendement obtenues sont approximativement de 20%
supérieures aux valeurs réelles cette année là, à l’exception de l’estimation à la floraison qui est de
11% supérieure. L’année 2004 était exceptionnelle, ce qui ne nous a pas permis d’utiliser les F1 et
F2 dont la surface foliaire est restée verte jusqu’à la maturité. L’utilisation de la F3 seule a fourni
des valeurs de rendement aux alentours de 28% inférieures aux valeurs réelles. Cette sous-
estimation est expliquée par l’utilisation des trois strates pour la construction des modèles
d’estimation des rendements, alors que ces modèles sont utilisés avec une seule strate cette année-
là.
Les modèles multiples, faisant intervenir à la fois « i » et « a » fournissent des résultats très
proches de ceux obtenus avec les modèles simples. L’analyse des résultats obtenus indique que les
améliorations possibles des performances des modèles en mode prévisionnel passent par
l’amélioration de la qualité des données obtenues avant la date à laquelle une estimation du
rendement à venir (à la floraison par exemple) est entreprise. Une autre amélioration peut
également provenir de l’utilisation de données de meilleure qualité pour la construction des
modèles. Le grand nombre d’hypothèses émises pour pouvoir utiliser les données disponibles entre

137
2000 et 2005 peut en effet avoir introduit un certain nombre d’erreurs diminuant les performances
des modèles obtenus.
Enfin, une adaptation du module d’estimation de l’évolution de la dégradation de la surface verte
des trois dernières strates du modèle PROCULTURE aux différents traitements pour les variétés
utilisées est nécessaire s’il est prévu d’utiliser des modèles tels que ceux construits en mode
prévisionnel dans ces cas là.
Conclusion et perspectives.
Les résultats obtenus permettent de répondre aux objectifs et aux hypothèses de travail présentés
au début de ce document.
- L’objectif principal de ce travail était de déterminer la validité d’un modèle prévisionnel de
simulation des risques de pertes de rendement basé sur l’étude des surfaces vertes.
Cette thèse a permis d’obtenir un modèle d’estimation des risques de pertes de rendement basé sur
l’étude du rayonnement journalier intercepté par la surface verte des trois dernières strates foliaires
du blé d’hiver. Ce modèle a été validé pour le site d’Everlange au Grand-duché de Luxembourg
sur 7 années de données, 3 variétés et 3 modalités de traitements phytosanitaires. Les performances
des modèles d’estimation des rendements obtenus sont assez encourageantes, car certains de ces
modèles expliquent plus de 90% des rendements obtenus en validation. De telles performances
pour un modèle construit sur 7 années de données suggèrent l’existence d’une relation fiable.
L’aspect prévisionnel des modèles a été testé sur deux années non considérées pour la construction
du modèle (2004 et 2008). Les prévisions de rendements ont fourni des valeurs de rendement
approximativement 20% supérieures aux valeurs de rendement réelles. Une estimation du
rendement a pu être entreprise dès la floraison, grâce à l’obtention des données de surface verte de
la floraison à la maturité. Les résultats obtenus ne concernent que le témoin de deux variétés
suivies en 2008, 2004 étant une année exceptionnelle à cause du maintien de sa surface verte des 2
dernières strates de la floraison à la maturité. Les modèles obtenus et plus particulièrement le
modèle mettant en relation le point d’inflexion des courbes de cumul de la matière sèche calculée
et les rendements est donc valide avec une erreur avoisinant les 20%. D’autre part, les bons
résultats obtenus en validation répondent à la première hypothèse de ce travail en montrant que la
cinétique de dégradation de la surface verte d’un couvert malade ou sain, et le rayonnement
intercepté par celle-ci sont des variables suffisantes pour estimer le rendement. Ces variables sont
suffisantes, mais sont-elles nécessaires ? Cette question peut-être discutée au vu des résultats
obtenus en mettant en relation uniquement un paramètre d’une courbe décrivant la dégradation de

138
la surface verte et le rendement (Chapitre I.2. PARTIE I). Ces résultats issus de l’application de
l’approche de Gooding et al. (2000) sont en effet très proches de ceux obtenus en mettant en
relation le rendement avec les paramètres du modèle retenu utilisant l’interception du rayonnement
(R² de 62% avec l’approche de Gooding et R² de 57% avec l’approche du modèle retenu sur les
données de 2006 et 2007. R² de 65% avec l’approche de Gooding et R² de 72% avec le modèle
retenu lorsque le modèle est construit uniquement avec les données de Achat). L’approche de
Gooding et al. (2000) a fourni de très bons résultats dans leur étude menée sur 17 années de
données. Cette étude ne prend pas en compte le rayonnement intercepté par la surface verte. Le
rayonnement mis en relation avec la surface verte présente sur les dernières feuilles émergées est
pourtant responsable en grande partie du remplissage des grains (Bryson et al., 1997). Une étude a
également montré que la relation entre le rendement et le rayonnement intercepté par la surface
verte durant la période de remplissage des grains (HAA) explique une plus grande variation de
rendements que la relation entre le rendement et la durée de la surface verte (HAD) sur la même
période (Madden et al., 2007). Une utilisation des données de Gooding et al. avec notre modèle
devrait donc aboutir à de meilleurs résultats. Une prolongation de ce travail de thèse, comparant les
résultats d’estimation des rendements obtenus en prévisionnel à l’aide du paramètre m de la courbe
de Gompertz aux résultats obtenus en prévisionnel avec le modèle retenu dans ce travail pourrait
également être envisagée. Cette comparaison permettrait également de déterminer si l’approche
utilisant le rayonnement intercepté par la surface verte apporte de meilleures prévisions de
rendement. Cette comparaison serait d’autant plus intéressante que l’aspect prévisionnel du
rayonnement dans notre démarche peut également être discuté, compte tenu de la difficulté à
obtenir le rayonnement de manière prédictive, comme cela a été montré dans ce dernier chapitre.
- La deuxième hypothèse de travail concerne la possibilité de prendre en compte l’effet variétal,
l’année et les traitements dans un modèle d’estimation des rendements basé sur l’utilisation des
surfaces vertes et du rayonnement intercepté par celles-ci.
Un effet variétal important a été mis en évidence, ce qui a conduit à considérer l’adaptation d’un
modèle par variété en plus du modèle général obtenu. Dans la construction du modèle, l’utilisation
des données par variété a mené à des modèles variétaux fournissant de meilleurs résultats que le
modèle général utilisant toutes les données. Il a cependant été montré que les valeurs de pentes et
d’intercepts des modèles linéaires variétaux ne sont pas significativement différents de ceux
obtenus avec le modèle global prenant en compte toutes les données. L’utilisation du modèle
variétal en prévisionnel n’a pas pu être testée étant donné que les variétés présentes les années pour
lesquelles l’utilisation en prévisionnelle a été testée ne sont pas celles utilisées pour la construction
du modèle. Seul le modèle global a été appliqué, ce qui ne permet pas de faire une comparaison

139
entre les performances en prévisionnel du modèle global et du modèle variétal. Dans d’autres
études établissant une relation entre le rendement et des variables descriptives de la décroissance
de la surface verte (Gooding et al., 2000 ; Bryson et al., 1997) ainsi que le rayonnement intercepté
(Bancal et al., 2007), la composante variétale n’a pas été distinguée. Les variétés utilisées par les
agriculteurs changent rapidement, ce qui motive probablement la recherche de relation concernant
l’ensemble des variétés utilisées au cours du temps.
L’effet « année » sur la variation des rendements est important. L’année explique cependant
beaucoup moins les variations de rendement que le paramètre « i » du modèle de calcul de la
matière sèche retenu. Au cours de ce travail, l’objectif était d’obtenir un modèle applicable au plus
grand nombre d’années. Cette démarche s’inscrit à la suite d’autres travaux (Bryson et al., 1997 ;
Gooding et al., 2000 ; Dimmock et Gooding, 2002) qui ont étudié la possibilité d’établir une
relation, commune à plusieurs années, entre le rendement et la dégradation de la surface verte .
Une analyse visant à appliquer des modèles fonctionnant pour des groupes d’années fournissant
des résultats semblables au sein d’une même variété n’a pas été menée. Ce genre d’étude n’est
possible que sur un grand nombre d’années et vise à isoler des comportements de la culture liés à
des conditions climatiques communes à plusieurs années. Ce type d’analyse n’aurait été possible
que sur la variété Achat qui est présente sur 6 années de données. Cette étude montre qu’il est
possible d’obtenir un modèle d’estimation des rendements expliquant une grande proportion des
rendements sur 7 années. Chacune de ces années montre des rendements différents pour une même
variété. C’est la prise en compte du profil de la dégradation de la surface verte foliaire, ainsi que
des données de température et de rayonnement qui permet l’explication des variations de
rendement.
La deuxième hypothèse de travail émise au début de ce travail aborde également la question de la
prise en compte des traitements phytosanitaires dans l’approche d’estimation des rendements
envisagée. Les traitements phytosanitaires ont un effet significatif sur les valeurs des paramètres
du modèle retenu (« i » et « a ») mis en relation avec les rendements. Les traitements
phytosanitaires ont également un effet significatif sur les valeurs de rendement. L’effet des
traitements sur les rendements est donc lié à l’effet sur les valeurs du modèle. Notre approche
prend donc les traitements en compte dans l’estimation des rendements, au travers des variations
des paramètres du modèle.
Dans le modèle retenu, c’est le LAI et la partie tenant compte de la dégradation de la surface verte
qui varient par rapport aux traitements. Il a été constaté que l’effet des fongicides sur le rendement
en grains est étroitement lié à l’effet sur la durée de la surface verte des dernières feuilles, tout
comme Gooding et al. (2000) l’avaient déjà mis en évidence.

140
L’utilisation du modèle d’estimation des rendements tient compte des traitements, uniquement
lorsque l’estimation n’est pas prévisionnelle. L’estimation prévisionnelle, en cours de saison, de
parcelles traitées nécessite l’obtention de la dégradation de la surface verte de toute la période
post-estimation. A l’heure actuelle, cette estimation peut être obtenue grâce à Proculture
uniquement pour le témoin non traité. Une adaptation de Proculture à la modélisation de
l’évolution des symptômes et des infections des principales maladies du blé sur des plantes traitées
permettrait l’utilisation du modèle en prévisionnel également pour les parcelles traitées.
- La troisième hypothèse de travail de cette thèse concerne la possibilité d’améliorer la gestion de
l’application des fongicides de manière à mieux respecter l’environnement grâce à la prise en
compte des informations issues à la fois de Proculture et du modèle d’estimation des rendements.
Cette hypothèse implique une détermination bien ciblée des dates d’application des produits
fongicides, afin d’en diminuer le nombre. Les sorties de Proculture sont déjà utilisées à cette fin,
au travers des avertissements et conseils de traitements élaborés et émis par le CRP, en
collaboration avec l’Université de Liège, chaque semaine en cours de saison à l’intention des
agriculteurs. Ces conseils pourraient être accompagnés d’une estimation des rendements à venir, à
partir d’une certaine date (la floraison) et en fonction des traitements phytosanitaires appliqués. Un
module de rendement économique pourrait également être mis au point à l’aide de modèles tels
que ceux présentés dans ce travail. Ce complément aux avertissements aurait comme intérêt pour
l’agriculteur de situer l’estimation de rendements correspondant à son choix de traitement et à le
situer par rapport aux autres choix possibles. Dans le cas où les estimations de rendement
économique sont meilleures pour un nombre d’applications de produits moindres, l’agriculteur en
serait informé à chaque avertissement hebdomadaire. Cette information pourrait alors motiver son
choix de traiter moins l’année suivante, et donc de mieux respecter l’environnement. Comme cela
a été expliqué plus haut, une adaptation de Proculture à l’estimation du développement des
symptômes et des infections des principales maladies du blé sur des plantes ayant subi différents
traitements sera nécessaire pour atteindre cet objectif.
L’adaptation du modèle d’estimation des rendements étudié dans ce travail à d’autres conditions
topo-climatiques est également envisageable. L’utilisation en mode prévisionnel du modèle
d’estimation des rendements ou des modèles variétaux sur d’autres sites, aux conditions de cumul
des températures journalières et de rayonnement différentes, nécessitera la connaissance des
données historiques de rayonnement et de température pour ces mêmes sites.

141
Une autre perspective d’amélioration de notre modèle concerne la prise en compte de données
prévisionnelles concernant le développement des symptômes de la rouille brune. Pour atteindre cet
objectif, il est nécessaire d’obtenir une modélisation du comportement de la maladie, tel que cela
est réalisé pour la septoriose. Dans cette étude, la région considérée n’a pas montré de
développement important de la rouille brune. Cette maladie n’a donc pas été prise en compte, ce
qui est sans conséquence importante pour l’obtention de la surface verte en prévisionnel. A
l’avenir, il se pourrait qu’un développement plus important de la rouille brune nécessite une
estimation prévisionnelle de son développement afin d’obtenir l’estimation prévisionnelle de la
surface verte. Cette éventualité est à envisager en cas d’adoucissement du climat, étant donné que
le développement de cette maladie nécessite des températures nocturnes élevées par rapport à la
septoriose.
Les méthodes d’acquisition des données de base du fonctionnement des modèles jusqu’à la
floraison (LAI et surface verte) sont diverses et peuvent également être améliorées dans leur
précision ainsi que dans leur automatisation. Plusieurs pistes ont été explorées dans ce travail et
dans de nombreux autres travaux (Bancal et al., 2007 ; de Jesus et al., 2001 ; Bassanezi et al.,
2001 ; Lopes et Berger, 2001 ; Bergamin Filho et al., 1997 ; Bryson et al., 1997) pour l’obtention
de ces données.
Il a été montré au cours de ce travail que la méthode d’estimation visuelle, la plus utilisée dans les
précédents travaux pour l’estimation du pourcentage de surface verte, est plus rapide mais moins
précise que l’analyse d’images. Cette dernière méthode présente en plus l’avantage de fournir la
surface foliaire, de manière très précise, en plus de la surface verte, ce qui permet de l’utiliser
également pour le calcul du LAI. Une amélioration du dispositif de prise d’image pour le rendre
plus rapide d’utilisation rendrait cette méthode définitivement plus intéressante pour les études à
l’échelle de petites parcelles expérimentales.
Un passage de l’estimation des rendements de l’échelle de la parcelle à celle du champ est à
envisager également pour adapter le modèle d’estimations des rendements à l’hétérogénéité des
grandes surfaces cultivées. L’obtention des principales données d’entrée du modèle (LAI et %SV)
à partir d’images aériennes a montré des résultats encourageants. Cependant, cette méthode
nécessite encore des améliorations qui pourraient être explorées par la suite. Une autre méthode
utilisant un radiomètre mesurant la réflectance du couvert en hyperspectrale pourrait également
être envisagée à grande échelle. Celle-ci a déjà fourni des résultats intéressants dans une étude
portant sur le HLAI du riz (Nutter, 1990).

142
Enfin, une amélioration des modèles peut également être attendue par la prise en compte des
réserves accumulées durant la préfloraison dans les tiges et remobilisées pour le remplissage des
grains par la suite (Bancal et al., 2007).

143
Bibliographie
Alem, C., Labhilili, M., Brahmi, K., Jlibene, M., Nasrallah, N. et Maltouf, A. F. 2002. Adaptation hydrique et photosynthétique du blé et du blé tendre au stress salin. Comptes Rendus Biologies 325, 1097-1109. Anonymous. 2002. BSA, Beschreibende Sortenliste, Getreide, Mais, Ölfrüchte, Leguminosen, Hackfrüchte, Deutscher Landwirtschaftsverlag GmbH 10-17. Aquino, V.M., Shokes, F.M., Berger, R.D., Gorbet, D.W. and Kucharek, T.A. 1992. Relationships among late leafspot, healthy leaf area duration, canopy reflectance, and pod yield of peanut. Phytopathology 82, 546-552. Bancal, M.O., Robert, C. and Ney, B. 2007. Modelling wheat growth and yield losses from late epidemics of foliar diseases using loss of green leaf area per layer and pre-anthesis reserves. Annals of Botany, volume 4, 777-789. Bassanezi, R.B., Amorim, L., Bergamin Filho, A., Hau, B. and Berger, R.D. 2001. Accounting for photosynthetic efficiency of bean leaves with rust, angular leaf spot and anthracnose to assess crop damage. Plant Pathology 50, 443-452. Bastiaans, L. 1991. Ration between virtual and visual lesion size as a measure to describe reduction in leaf photosynthesis of rice due to leaf blast. Phytopathology 81, 611-615. Bastiaans, L. 1993. Effects of leaf blast on growth and production of a rice crop. 2. Analysis of the reduction in dry matter production, using two models with different complexity. Netherlands Journal of Plant Pathology 99,19-28. Bastiaans, L. and Kropff, M.J . 1993. Effects of leaf blast on photosynthesis of rice. 2. Canopy photosynthesis. Netherlands Journal of Plant Pathology 99(Suppl.3), 205-217. Bastiaans, L., Rabbinge, R. and Zadoks, J.C. 1994. Understanding and modeling leaf blast effects on crop physiology and yield. Rice Blast Disease eds. C.A.B. International, Wallingford, England, 357-380. Bayles, R.A. and Hilton, G. J. 2000. Variety as a factor in the response of cereals to strobilurins. In Proceeding of the Brighton Crop Protection Conference – Pests and disease, 731-738. Beadle, C.L. 1993. Growth analysis. Photosynthesis and production in a changing environment. Hall, D.O., et al. (Eds), A field and laboratory manual. Chapman & Hall, London, 36-46. Bergamin Filho, A., Carneiro, S.M.T.P.G., Godoy, C.V., Amorim, L., Berger, R. D. and Hau, B. 1997. Angular leaf spot of Phaseolus beans: Relationships between disease, healthy leaf area, and yield. Phytopathology 87, 506-515. Blackmer, A.M. and White, S.E.. 1998. Using precision farming technologies to improve management of soil and fertilizer nitrogen. Australian Journal of Agricultural Research 49, 555-564.

144
Boote, K.J., Jones, J.W., Mishoe, J.W. and Berger, R.D. 1983. Coupling pests to crop growth simulators to predict yield reductions. Phytopathology 73, 1581-1587. Boote, K.J., Jones, J.W. and Pickering, N.B. 1996. Potential uses and limitations of crop models. Agronomy Journal 88, 704-716. Bréda, N. 2003. Ground-based measurements of leaf area index : a review of methods, instruments and current controversies. Journal of Experimental Botany, 54, 2403-2417. Brenner, A.J., Cueto Romero, M., Garcia Haro, J., Gilabert, M.A., Incoll, L.D., Martinez Fernandez, J., Porter, E., Pugnaire, F.I. and Younis, M.T.. 1995. A comparison of direct and indirect methods for measuring leaf and surface areas of individual bushes. Plant, Cell and Environment 18, 1332-1340. Bryson, R.J., Leandro, L. and Jones, D.R. 2000. The physiological effects of kresoxim-methyl on wheat leaf greenness and the implication for crop yield. Proceeding of the Brighton Crop Protection Conference – Pests and diseases 739-747. Bryson, R.J. , Paveley, N.D. , Clark, W.S. , Sylvester-Bradley, R. and Scott, R.K. 1997. Use of in-field measurements of green leaf area and incident radiation to estimate the effects of yellow rust epidemics on the yield of winter wheat. European Journal of Agronomy 7, 53-62. Bryson, R.J., Sylvester-Bradley, R., Scott, R.K. and Paveley, N.D. 1995. Reconciling the effects of yellow rust on yield of winter wheat through measurements of green leaf area and radiation interception. Physiological Responses of Plants to Pathogens volume 42, 9-18. Buerkert, A., Mahler, F. and Marschner, H. 1996. Soil productivity management and plant growth in the Sahel: Potential of an aerial monitoring technique. Plant Soil 180, 29-38. Calderini, D.F., Savin, R., Abeledo, L.G., Reynolds, M.P. and Slafer, G.A. 2001. The importance of the period immediately preceding anthesis for grain weight determination in wheat. Euphytica 119, 199-204. Campbell, G.S. and Norman, J.M. 1989. The description and measurement of plant canopy structure. In G. Russell et al. (eds.). Plant canopies: Their growth, form and function. Cambridge University Press, Cambridge, UK. 1-19. Chen, J.M. 1996. Optically-based methods for measuring seasonal variation of leaf area index in boreal conifer stands. Agricultural and Forest Meteorology 80, 135-163. Chiarappa, L. 1971. Crop Loss Assessment Methods: FAO Manual on the Evaluation and Prevention of Losses by Pests, Diseases and Weeds. Farnham Royal: FAO/CAB. Chiarappa, L. 1981. Crop Loss Assessment Methods. Supplement N◦3. FAO/CAB, Farnham, England. 123 pp. Clifford, B.C. and Harris, R.G. 1981. Controlled environment studies of the epidemic potential of Puccinia recondita f. sp. tritici on wheat in Britain. Transactions of the British Mycological Society 77, 351-358.

145
Davies, W. P., Kettlewell, P. S., Green, H. J., Hocking, T. J. and Jarai, M. J. 1984. Senescence and net photosynthesis of the flag leaf and grain growth of wheat in response to fungicide. Annals of Applied Biology 105, 303-312. Deblonde, G., Penner, M. and Royer, A. 1994. Measuring leaf area index with the LI–COR LAI-2000 in pine stands. Ecology 75, 1507-1511. De Jesus, W.C. Jr., Ribeiro do Vale, X., Coelho, R.R. and Costa, L.C. 2001. Comparison of two methods for estimating leaf area index on common Bean. Agronomy Journal 93, 989-991. De Jesus, W.C. Jr., Ribeiro do Vale, X., Coelho, R.R., Hau, B., Zambolim, L., Costa, L.C. and Bergamin Filho, A. 2001. Effects of angular leaf spot and rust on yield loss of Phaseolus vulgari. Phytopathology 91, 1045-1053. De Vallavieille-Pope, C., Hubert, L., Lecomte, M. and Goyeau, H. 1995. Comparative effects of temperature and interrupted wet periods on germination, penetration and infection of Puccinia recondita f.sp. tritici and P. striiformis urediniospores on wheat seedlings. Phytopathology 85, 409-415. De Winne. 1994. Introduction of the updated system description of the WOFOST crop growth simulation model as implemented in the Crop Growth Monitoring System applied by the European Commission. http://supit.net/main.php?q=aXRlbV9pZD02MQ== Dimmock, J.P.R.E. and Gooding, M.J., 2002a. The effects of fungicides on rate and duration of grain filling in winter wheat in relation to maintenance of flag leaf green area. Journal of Agricultural Science 138, 1-16. Egli, D.B. 1998. Seed biology and the yield of grain crops. CAB International, Wallingford, UK. El Jarroudi, M., Delfosse, P., Maraite, H., Hoffmann, L. and Tychon, B. Validation of a simulation model of Septoria leaf bloch disease progress in winter wheat in Luxembourg. Plant Pathology. Soumis in July 2008. El Jarroudi, M. 2005. Evaluation des paramètres épidémiologiques des principales maladies cryptogamiques affectant les feuilles du blé d’hiver au Grand-Duché de Luxembourg : Calibration et validation d’un modèle de prévision .Thèse de doctorat Université de Liège. Département des Sciences et Gestion de l’Environnement. El Jarroudi, M., Tychon, B. and Maraite, H. 2002. Validation of « proculture » model to predict Septoria tritici development on winter wheat in the Grand-Duché of Luxembourg in 2001. In proceedings of the ESA VII Congress, Cordoba, Spain 275-276. Fassnacht, K.S., Gower, S.T., Norman, J.M. and McMurtie, R.E. 1994. A comparison of optical and direct methods for estimating foliage surface area index in forests. Agricultural and Forest Meteorology 71, 183-207. Ferrandino, F.J. and Elmer, W.H. 1992. Reduction in tomato yield due to Septoria leaf spot. Plant Disease 76, 208-211.

146
Forbes, G.A., and Jeger, M.J. 1987. Factors affecting the estimation of disease intensity in simulated plant structures. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 94, 113-120. Forrest, W., Nutter, Jr. and Schultz, P.M. 1995. Improving the accuracy and precision of disease assessments : selection of methods and use of computer-aided training programs. Canadian Journal of Plant Pathology 17, 174-184. Gérard, B. and Buerkert, A. 2001. Estimation of spatial variability in pearl millet growth using non-destructive methods. Experimental Agriculture 37, 373-389. Gianasi, L. 1999. Antracnose do Feijoeiro: QuantificacËaÄo de Danos e Efeito do Trifenil Acetato de Estanho no Crescimento do Hospedeiro e no Progresso da DoencËa. Piracicaba, Brazil: Universidade de Sao Paulo, PhD thesis. Gooding, M.J, Dimmock, J.P.R.E., France, J. and Jones, S.A. 2000. Green leaf area decline of wheat flag leaves: the influence of fungicides and relationships with mean grain weight and grain yield. Annals of Applied Biology 136, 77-84. Gower, S.T. and Norman, J.M. 1991. Rapid estimation of leaf area index in conifer and broad-leaf plantations. Ecology 72, 1896-1900. Guissard, V. 2005. SPIE Europe Int. Symposium – Remote Sensing, Bruges, Belgium, Proceedings. Hanft, J.M. and Wych, R.D. 1982. Visual indicators of physiological maturity of hard red spring wheat. Crop Science 22, 584-588. Hopkins, J.W. 1939. Estimation of leaf area in wheat from linear dimensions. Canadian Journal of Research 17, 300-304. Horsfall, J.G. and Barratt, R.W. 1945. An improved grading system for measuring plant disease (abstract). Phytopathology 35, 655. Jackson, H.R., Wallen, V.R. and Downer, J.F. 1978. Analysis and electronic area measurement of complex aerial photographic images. Journal of Applied Photographic Engineering 4, 101-106 James, W.C. 1974. Assessment of plant diseases and losses. Annual Review of Phytopathology, 12, 27-48. James, W.C., Teng, P.S. and Nutter, Jr. F.W. 1991. Estimated losses of crops from plant pathogens. CRC Handbook of Pest Management. CRC Press, Boca Raton. 1 and 2, 15-51. Johnson, K.B. 1992. Evaluation of a mechanistic model that describes potato crop losses caused by multiple pests. Phytopathology 82, 363-369. Jorgensen, H.J.L. and Smedegaard-Peterson, V. 1999. Host–pathogen interactions in the Septoria-disease complex. Lucas JA, Bowyer P, Anderson HM, eds. Septoria on cereals: a study of pathosystems. Wallingford, UK: CABI Publishers, 131-160.

147
Kropff, M.J., Teng, P.S. and Rabbinge, R. 1995. The challenge of linking pest and crop models. Agricultural Systems 49, 413-434. Lal, K.N. and Subba Rao, M.S. 1950. A rapid method for estimation of leaf area in growing maize plants. Current Science 19,179-180. Large, E.C. 1966. Measuring plant disease. Annual Review of Phytopathology 4, 9-28. Lemaire, D., Amand, O. and Maraite, H. 2003. Evolution of Proculture, a disease risk simulation model for decision taking in Mycosphaerella graminicola control. 6thInternational Symposium on Septoria/Stagonospora Diseases of cereals/ Tinis-Tunisia, December 8-12. Levy, P.E. and Jarvis, P.G. 1999. Direct and indirect measurements of LAI in millet and fallow vegetation in HAPEX-Sahel. Agricultural and Forest Meteorology 97, 199-212. Lindow, S.E. and Webb, R.R. 1983. Quantification of foliar plant disease symptoms by microcomputer-digitized video image analysis. Phytopathology 73, 520-524. Lopes, D.B. and Berger, R.D. 2001. The effects of rust and anthracnose on the photosynthetic competence of diseased bean leaves. Phytopathology 91, 212-220. Lovell, D.J., Parker, S.R., Hunter, T., Royle, D.J. and Coker, R.R. 1997. Influence of crop growth and structure on the risk of epidemics by Mycosphaerella graminicola (Septoria tritici) in winter wheat. Plant Pathology 46, 126-138. Lovell, D.J., Parker, S.R., Petghem, P.V., Webb, D.A. and Welham, S.J. 2002. Quantification of raindrop kinetic energy for improved prediction of plash-dispersed pathogens. Phytopathology 92, 497-503. Lupton, F.G.H. 1972. Further experiments on photosynthesis and translocation in wheat. Annals of Applied Biology 71, 69-71. Lyman, G.R. 1918. The relation of phytopathologists to plant disease survey work. Phytopathology 8, 219-228. Macbeth, J.E., Kettlewell, P.S. and Sylvester Bradley, R. 1996. Methods for pre-harvest prediction of grain weight in winter wheat. Agri-food Quality: An Interdisciplinary Approach (Eds G. R. Fenwick, C. Hedley, R. L. Richards & S. Khokhar), Cambridge: The Royal Society of Chemistry 83-86. Mackels, C. and Tychon, B. 2008. Green leaf area decline of the last three leaves of wheat. Determination of the relationships with remote sensing green land cover. Italian Journal of Agronomy, 3, supplement, 671-672. Madden, L.V., Hughes, G. and Van den Bosch, F. 2007. The study of plant disease epidemics. APS Press. page 432. Madeira, A.C., Clark, J.A., and Rossall, S. 1994. Growth and light interception in field bean (Vicia faba) infected by Ascochyta fabae. Journal of Agricultural Science 123, 225-232.

148
Magboul, A.M., Geng, S., Gilchrist, D.G. and Jackson, L.F. 1992. Environmental influence on the infection of wheat by Mycosphaerella graminicola. Phytopathology 82, 1407-1413. Marcus, K.G. and Heitkemper, L.J. 1994. Commodity market planning worldwide. Proceedings of the Seminar: Crop Yield Forecasting Methods. Villefranche-sur-Mer, 24 to 27 October 1994. Office for Official Publications of the European Union, Luxembourg 75-84. Martin, D.P. and Rybicki, E.P. 1998. Microcomputer-based quantification of maize streak virus symptoms in Zea mays. Phytopathology 88, 422-427. McGrath, M.T. and Pennypacker, S.P. 1990. Alteration of physiological processes in wheat flag leaves caused by stem rust and leaf rust. Phytopathology 80, 677-686. Mitchell, P.L. 1997. Misuse of regression for empirical validation of models. Agricultural Systems 54, 313-326. Monteith, J.L. 1972. Solar radiation and productivity in tropical ecosystems. Journal of Applied Ecology 9, 747-766. Monteith , J.L. 1977. Climate and the efficiency of crop production in Britain. Philosophical Transactions of the Royal Society of London 281, 277-294. Moya, E.A., Barrales, L.R. and Apablaza, G.E. 2005. Assessment of the disease severity of squash powdery mildew through visual analysis, digital image analysis and validation of these methodologies. Crop Protection 24, 785-789. Niemira, B.A., Kirk, W.W. and Stein, J.M. 1999. Screening for late blight susceptibility in potato tubers by digital analysis of cut tuber surfaces. Plant Disease 83, 469-473. Nilsson, H. 1995. Remote sensing and image analysis in plant pathology. Canadian Journal of Plant Pathology 17, 154-166. Nutter, F.W.Jr , 1990. Remote sensing and image analysis for crop loss assessment. In: ABC. (Eds.), Crop Loss Assessment in Rice 93-105. Nutter, F.WJr, Gleason, M.L., Jenco, J.H. and Christians, N.C. 1993a. Assessing the accuracy, intra-rater repeatability and inter-rater reliability of disease assessment systems. Phytopathology 83, 806-812. Nutter, F.W.Jr. and Littrell, R. H. 1996. Relationships between defoliation, canopy reflectance and pod yield in the peanut-late leafspot pathosystem. Crop Protection. 15, 135-142. Nutter, F.W. and Schutz, P.M. 1995. Improving the accuracy and precision of disease assessments : selection of methods and use of computer-aided training programs. Canadian Journal of Plant Pathology 17, 174-184. Nutter, F.W.Jr ., and Worawitlikit, O. 1989. Disease.Pro : a computerized program for evaluating and improving a persons ability to assess disease proportion. Phytopathology 79, 11-35.

149
Olmstead, J.W., Lang, G.A. and Grove, G.G. 2001. Assessment of severity of powdery mildew infection of sweet cherry leaves by digital images analysis. HortScience 36, 107-111. Parker, S.R., Shaw, M.W. and Royle, D.J. 1995. The reliability of visual estimates of disease severity on cereal leaves. Plant Pathology 44, 856-864. Pepler, S., Gooding, M.J., Ford, K.E. and Ellis, R.H. 2005. A temporal limit to the association between flag leaf life extension by fungicides and wheat yields. European Journal of Agronomy 22, 363-373. Pepler, S., Gooding, M.J. and Ellis, R.H. 2006. Modelling simultaneously water content and dry matter dynamics of wheat grains. Field Crops Research 95, 49-63. Pinter, P.J.Jr, Jackson, R.D., ldso, S.B. and Reginato, R.I. 1983. Diurnal patterns of wheat spectral reflectances. IEEE Transactions on Geoscience & Remote Sensing 21, 156-163. Price, T.V., Gross, R., Ho Wey, J. and Osborne, C.F. 1993. A comparaison of visual and digital image-processing methods in quantifying the severity of coffee leaf rust (Hemileia vastatrix). Australian Journal of Experimental Agriculture 33, 97-101. Rabbinge, R. and Rijsdijk, F.H. 1981. Disease and crop physiology: a modeler’s point of view. Effects of Disease on the Physiology of the Growing Plant, Cambridge University Press ed. PG Ayres. 201-20. Rapilly, F. 1991. Epidémiologie en pathologie végétale: mycoses aériennes. L. T. Doc. Paris. Institut National de la Recherche Agronomique INRA. ISBN 2-7380-0297-8. 317 p. 240-261. Robert, C., Bancal, M.O., Lannou, C. and Ney, B. 2006. Quantification of the effects of Septoria tritici blotch on wheat leaf gas exchange with respect to lesion age, leaf number, and leaf nitrogen status. Journal of Experimental Botany 57, 225-234. Robert, C., Bancal, M.O., Ney, B. and Lannou, C. 2005. Wheat leaf photosynthesis loss due to leaf rust, with respect to lesion development and leaf nitrogen status. New Physiologist 165, 227-241. Robert, C., Bancal, M.O., Nicolas, P., Lannou, C. and Ney, B. 2004. Analysis and modelling of effects of leaf rust and Septoria tritici blotch on wheat growth. Journal of Experimental Botany, 55, 1-16. Rossi, V., Racca, P., Giosue, S., Pancaldi, D. and Alberti, I. 1997. A simulation model for the development of brown rust epidemics in winter wheat. European Journal of Plant Pathology 103, 453-465. Rossing, W.A.H. 1991. Simulation of damage in winter wheat caused by the grain aphid Sitobion avenae. 3. Calculation of damage at various attainable yield levels. Netherlands Journal of Plant Pathology 97, 87-103. Rossing, W.A.H., van Oijen, M., van der Werf, W., Bastiaans, L., and Rabbinge, R. 1992. Modelling the effects of foliar pests and pathogens on light interception, photosynthesis, growth

150
rate and yield of field crops. Pests and Pathogens-Plant Responses to Foliar Attack. P. G. Ayres, ed. Biosience Scientific Publishers, Oxford 161-180 Savary, S., Teng, P.S., Willocquet, L. and Nutter, F.W.Jr. 2006. Quantification and modeling of crop losses: A review of purposes. Annual Review of Phytopathology 44, 89-112. Schnyder, H. and Baum, U. 1992. Growth of the grain of wheat (Triticum aestivum L.). The relationship between water content and dry matter accumulation. European Journal of Agronomy 1, 51-57. Scholes, J.D. 1992. Photosynthesis : cellular and tissue aspects in diseased leaves. Pests and pathogens – plant responses to foliar attacks. E. Lancaster. P.G. Ayres ed., 85-101. Scholes, J.D. and Farrar, J.F. 1986. Increased rates of photosynthesis in localised regions of a barley leaf infected with brown rust. New Phytologist 104, 601-612. Shaw, M.W. 1990. Effects of temperature, leaf wetness and cultivar on the latent period on Mycosphaerella graminicola on winter wheat. Plant Pathology 39, 255-268. Shaw, M.W. and Royle, D.J. 1989. Estimation and validation of function describing the rate at which Mycosphaerella graminicola causes yield loss in winter wheat. Annals of Applied Biology 115, 425-442. Sherwood, R.J., Berg, C.C., Hoover, M.R., and Zeiders, K.E. 1983. Illusions in visual assessment of Stagonospora leafspot of orchard grass. Phytopathology 73, 173-177. Shtienberg, D. 1992. Effects of foliar diseases on gas exchange processes : a comparative study. Phytopathology 82, 760-765. Sofield, I., Wardlaw, I.F., Evans, L.T. and Zee, S.Y. 1977b. Nitrogen, phosphorus and water contents during grain development and maturation in wheat. Australian Journal of Plant Physiology 4, 799-810. Spiertz, J.H.J. 1977. The influence of temperature and light intensity on grain growth in relation to the carbohydrate and nitrogen economy of the wheat plant. Netherlands Journal of Agricultural Science 25, 182-197. Spitters, C.J.T., Roermund, H.J.W.V., Nassau, H.G.M.G.V., Scheppers, J. and Mesdag, J. 1990. Genetic variation in partial resistance to leaf rust in winter wheat : disease progress, foliage senescence and yield reduction. Netherlands Journal of Plant Pathology 96, 3-15. Thomas, M.R., Cook, R.J., and King, J.E. 1989. Factors affecting development of Septoria tritici in winter wheat and its effect on yield. Plant Pathology 38, 246-257. Tomerlin, J.R., and Howell, T.A. 1988. DISTRAIN : a computer program for training people to estimate disease severity on cereal leaves. Plant Disease 72, 455-459. Van Arkel , H. 1978. Leaf area determinations in sorghum and maize by the length-width method. Netherlands Journal of Agricultural Science 26, 170-180.

151
Waggoner, P.E. and Berger, R.D. 1987. Defoliation, disease, and growth. Phytopathology 77, 393-398. Watson, D.J. 1947. Comparative physiological studies in the growth of field crops. I. Variation in net assimilation rate and leaf area between species and varieties, and within and between years. Annals of Botany 11, 41-76. Webb, R., and Lindow, S.E. 1981. Evaluation of Ascochyta pteridium as a potential biological control agent of bracken fern. (Abstr.) Phytopathology 71, 911.
Weiss, M., Baret, F., Smith, G.J., Jonckheere, I., and Coppin, P. 2004. Review of methods for in situ leaf area index (LAI) determination Part II. Estimation of LAI, errors and sampling. Agricultural and Forest Meteorology 121, 37-53. Welles, J.M., and Cohen, S. 1996. Canopy structure measurement of gap fraction analysis using commercial instrumentation. Journal of Experimental Botany 47, 1335-1342. White, M.A., Asner, G.P., Nemani, R.R., Privette, J.L. and Running, S.W. 2000. Measuring fractional cover and leaf area index in arid ecosystems: Digital camera, radiation transmittance, and laser altimetry methods. Remote Sensing of Environment 74, 45-57. Wiegand, C.L., Cuellar, J.A. 1981. Duration of grain filling and kernel weight of wheat as affected by temperature. Crop Science 21, 95-101. Wilhelm, W.W., Ruwe, K. and Schlemmer, M.R. 2000. Comparison of three leaf area index meters in a corn canopy. Crop Science 40, 1179-1183. Wright, A.C., and Gaunt, R.E. 1992. Disease-yield relationships in barley. I. Yield, dry matter accumulation and yield loss models. Plant Pathology 41, 676-687. Zadoks, J.C. 1987. Rationale and concepts of crop loss assessment for improving pest management and crop protection. Crop loss assessment and pest management 123, 1-5. Zadoks, J.C., Chang T.T. and Konzak, B.F. 1974. A decimal code for the growth stages of cereals. Weed Research 14, 415-421. Zadoks, J.C. and Schein, R.D. 1979. Epidemiology and Plant Disease Management. New York: Oxford University Press.427pp.


















