
UIC I Génétique - Cours 4 UIC I Génétique - Cours 4 LEXPRESSION GENIQUE.

L’EXPRESSION DES GENES
Denise Aragnol
Instabilité du génome et cancérogenèse
CNRS/Université de la Méditerranée
?
Chez les eucaryotes

I-DEFINITION D’EXPRESSIONGENIQUE
Expression génique : recouvre l’ensemble des mécanismes quiconduisent à l’apparition d’un produit fonctionnel d’un gène
Produit fonctionnel:-Une protéine� transcription + traduction
-Un ARN � transcription
Exemples de « produit final fonctionnel » de type ARN:ARNt, ARNr, de nombreux (petits) ARN cytoplasmiques et nucléaires dont les fonctions sontencore inconnues
The RNA content ofa cell. This schemeshows the types ofRNA present in allorganisms(eukaryotes,bacteria andarchaea) and thosecategories foundonly in eukaryotic.The non-codingRNAs of archaeahave not yet beenfully characterizedand it is not clearwhich types arepresent in additionto rRNA and tRNA
traduction

II-LA TRANSCRIPTION
LES ARN POLYMERASES EUCARYOTES
-3 ARNpol différentes: ARN pol I, ARN Pol II, ARN Pol III
compositions et fonctions différentes, structures similaires
-multimériques (8 à 12 s. u.); >500kDa
Functions of the three eukaryotic nuclear RNA polymerases
Polymerase Genes transcribed____________________________________________________
RNA polymerase I (nucléole) 28S, 5.8S and 18S ribosomal RNA (rRNA) genes
RNA polymerase II Protein-coding genes; mostsmall nuclear RNA (snRNA)genes
RNA polymerase III Genes for transfer RNAs (tRNA), 5S rRNA, U6-snRNA,small nucleolar (sno) RNAs,small cytoplasmic (sc) RNA
L’ARNpol II eucaryote est constituée de plus de 10 sous-unités
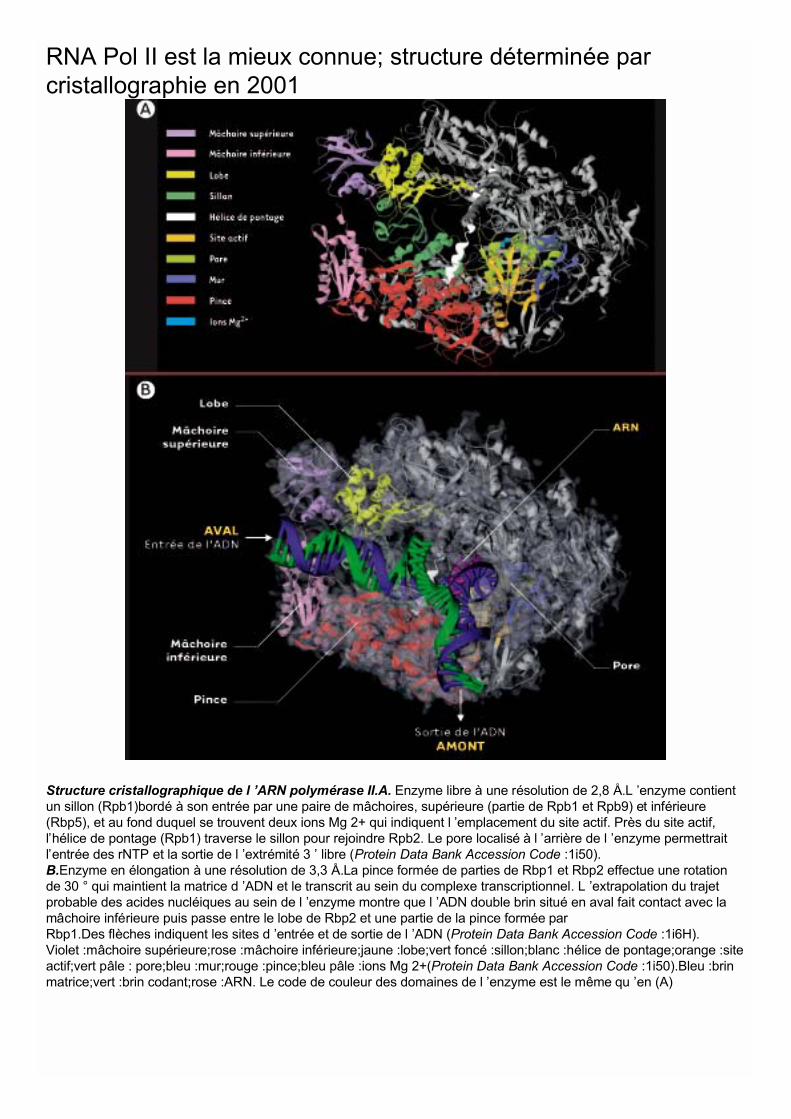
Structure cristallographique de l ’ARN polymérase II.A. Enzyme libre à une résolution de 2,8 Å.L ’enzyme contientun sillon (Rpb1)bordé à son entrée par une paire de mâchoires, supérieure (partie de Rpb1 et Rpb9) et inférieure(Rbp5), et au fond duquel se trouvent deux ions Mg 2+ qui indiquent l ’emplacement du site actif. Près du site actif,l’hélice de pontage (Rpb1) traverse le sillon pour rejoindre Rpb2. Le pore localisé à l ’arrière de l ’enzyme permettraitl’entrée des rNTP et la sortie de l ’extrémité 3 ’ libre (Protein Data Bank Accession Code :1i50).B.Enzyme en élongation à une résolution de 3,3 Å.La pince formée de parties de Rbp1 et Rbp2 effectue une rotationde 30 ° qui maintient la matrice d ’ADN et le transcrit au sein du complexe transcriptionnel. L ’extrapolation du trajetprobable des acides nucléiques au sein de l ’enzyme montre que l ’ADN double brin situé en aval fait contact avec lamâchoire inférieure puis passe entre le lobe de Rbp2 et une partie de la pince formée parRbp1.Des flèches indiquent les sites d ’entrée et de sortie de l ’ADN (Protein Data Bank Accession Code :1i6H).Violet :mâchoire supérieure;rose :mâchoire inférieure;jaune :lobe;vert foncé :sillon;blanc :hélice de pontage;orange :siteactif;vert pâle : pore;bleu :mur;rouge :pince;bleu pâle :ions Mg 2+(Protein Data Bank Accession Code :1i50).Bleu :brinmatrice;vert :brin codant;rose :ARN. Le code de couleur des domaines de l ’enzyme est le même qu ’en (A)
RNA Pol II est la mieux connue; structure déterminée parcristallographie en 2001
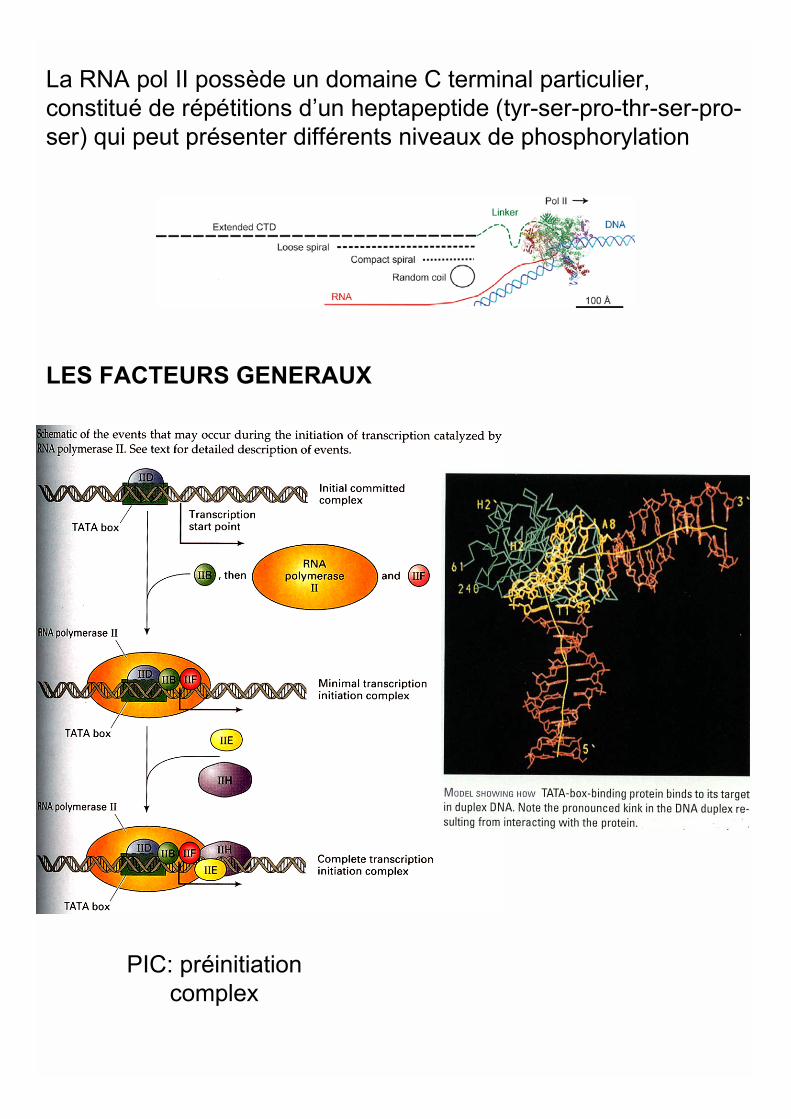
LES FACTEURS GENERAUX
La RNA pol II possède un domaine C terminal particulier,constitué de répétitions d’un heptapeptide (tyr-ser-pro-thr-ser-pro-ser) qui peut présenter différents niveaux de phosphorylation
PIC: préinitiationcomplex

LES FACTEURS EN AMONT
LES PROMOTEURS EUCARYOTES SONT COMPLEXES
EXEMPLE DE PROMOTEURS RECONNUS PAR L’ARN POL II
Chez les eucaryotes le promoteur comprend toutes lesséquences importantes pour l’initiation de la transcription. Il estmodulaire et complexe
Promoteur basal: -TATAbox (-25) TATAWAW avec W=A ou T
-Inr YYA(+1)NWYY avec Y=C ou T
Eléments en amont: CAAT ou GC ou octamère (oct)
En général les promoteurs eucaryotes ne contiennent pasl’ensemble de ces éléments
Les nombres et positions des éléments en amont sont variables

L’ASSEMBLAGE DU COMPLEXE D’INITIATION DE LATRANSCRIPTION DEFINIT LE CHOIX DU BRIN MATRICE
Les interactions entre l’ARN Pol, les différents modules dupromoteur et les différents facteurs protéiques du complexed’initiation de la transcription définissent le « start » detranscription et déterminent le choix du brin matrice
TAACCAAT TATA
ATAT
TAACCAAT TATA
ATAT
5’
5’
5’
5’
3’
3’
3’
3’
5’
5’
L’ELONGATION PEUT COMMENCER
III-LA MATURATION-Addition de la coiffe en 5’ The C-Terminal Domain
(CTD) of RNA PolymeraseII CoordinatesTranscription and Pre-mRNA ProcessingTheCTD consists of 52repeats of the consensusheptapeptide Tyr-Ser-Pro-Thr-Ser-Pro-Ser andserves as a platform forthe ordered assembly ofthe factors responsible fortranscription, pre-mRNA 5_capping, splicing, and 3_processing at differentstages in the synthesis ofthe nascent transcript.

Capping of eukaryotic mRNA. The top part of the diagramshows the capping reaction in outline. A GTP molecule(drawn as Gppp) reacts with the 5 end of the mRNA togive a triphosphate linkage. In the second step of theprocess, the terminal G is methylated at nitrogen number 7.The bottom part of the diagram shows the chemicalstructure of the type 0 cap, with asterisks indicating thepositions where additional methylations might occur toproduce type 1 and type 2 cap structures.
-L’excision des introns et l’épissage des exonscommencent pendant l’élongation
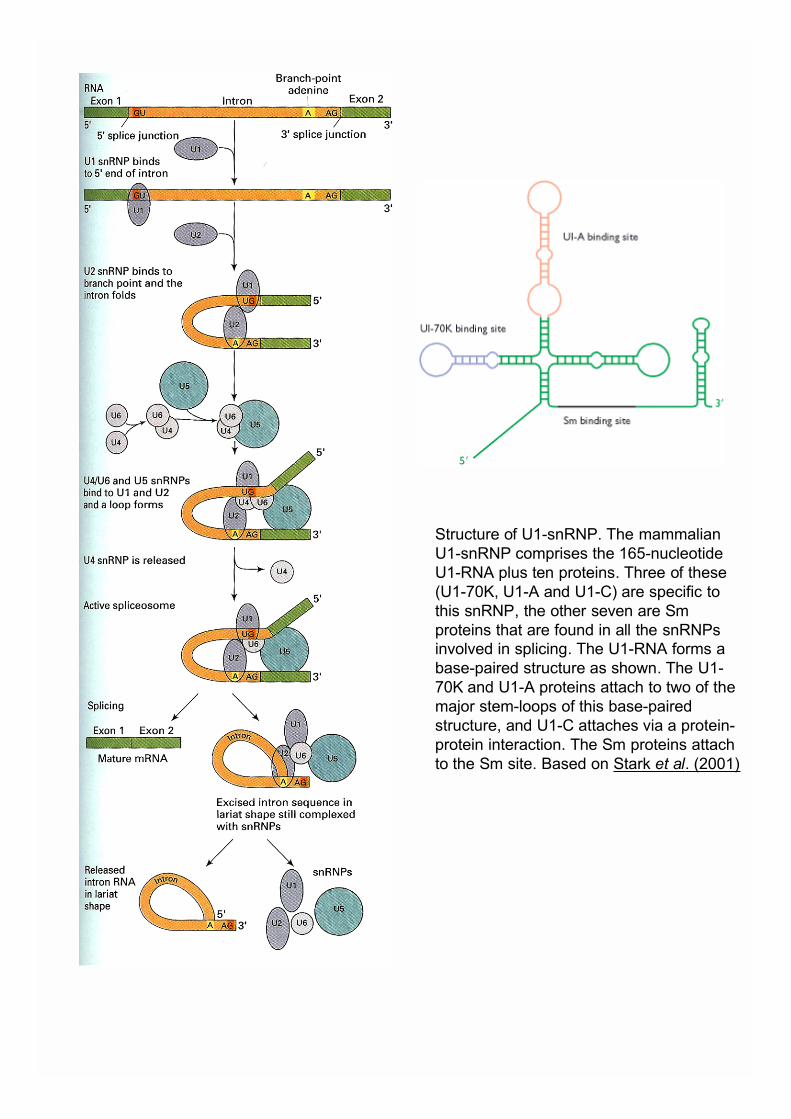
Structure of U1-snRNP. The mammalianU1-snRNP comprises the 165-nucleotideU1-RNA plus ten proteins. Three of these(U1-70K, U1-A and U1-C) are specific tothis snRNP, the other seven are Smproteins that are found in all the snRNPsinvolved in splicing. The U1-RNA forms abase-paired structure as shown. The U1-70K and U1-A proteins attach to two of themajor stem-loops of this base-pairedstructure, and U1-C attaches via a protein-protein interaction. The Sm proteins attachto the Sm site. Based on Stark et al. (2001)
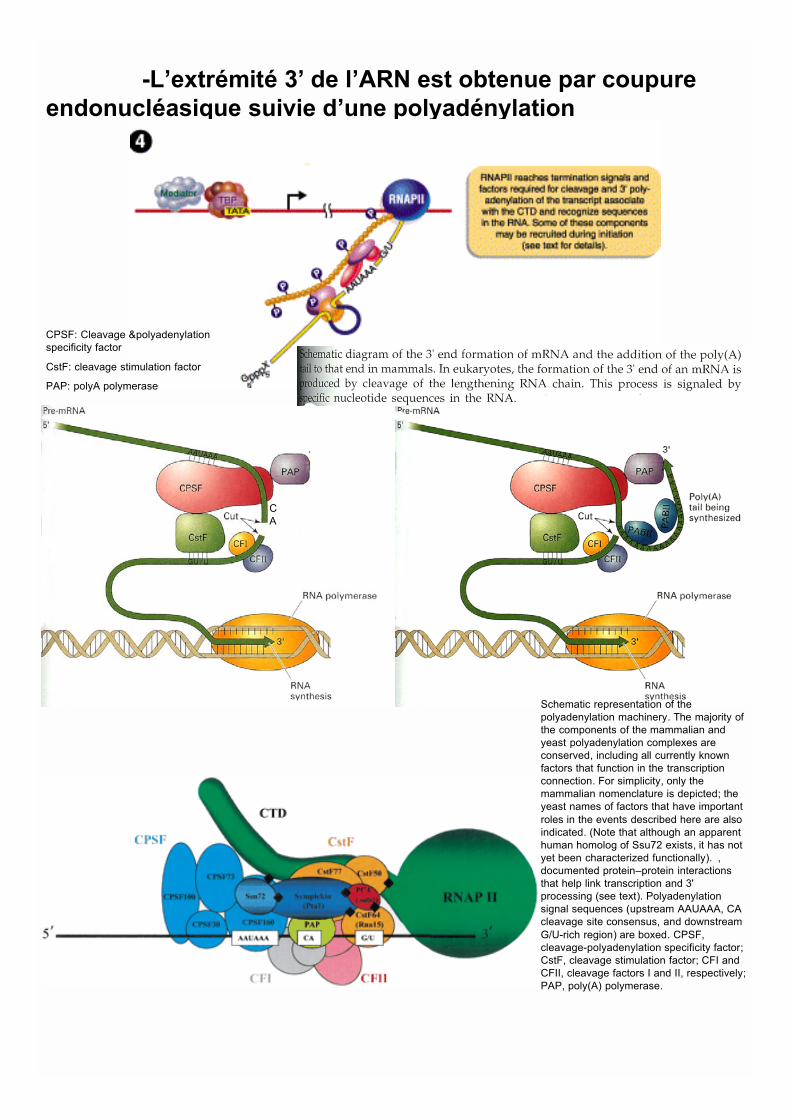
-L’extrémité 3’ de l’ARN est obtenue par coupureendonucléasique suivie d’une polyadénylation
Schematic representation of thepolyadenylation machinery. The majority ofthe components of the mammalian andyeast polyadenylation complexes areconserved, including all currently knownfactors that function in the transcriptionconnection. For simplicity, only themammalian nomenclature is depicted; theyeast names of factors that have importantroles in the events described here are alsoindicated. (Note that although an apparenthuman homolog of Ssu72 exists, it has notyet been characterized functionally). ,documented protein–protein interactionsthat help link transcription and 3'processing (see text). Polyadenylationsignal sequences (upstream AAUAAA, CAcleavage site consensus, and downstreamG/U-rich region) are boxed. CPSF,cleavage-polyadenylation specificity factor;CstF, cleavage stimulation factor; CFI andCFII, cleavage factors I and II, respectively;PAP, poly(A) polymerase.
CA
CPSF: Cleavage &polyadenylationspecificity factor
CstF: cleavage stimulation factor
PAP: polyA polymerase
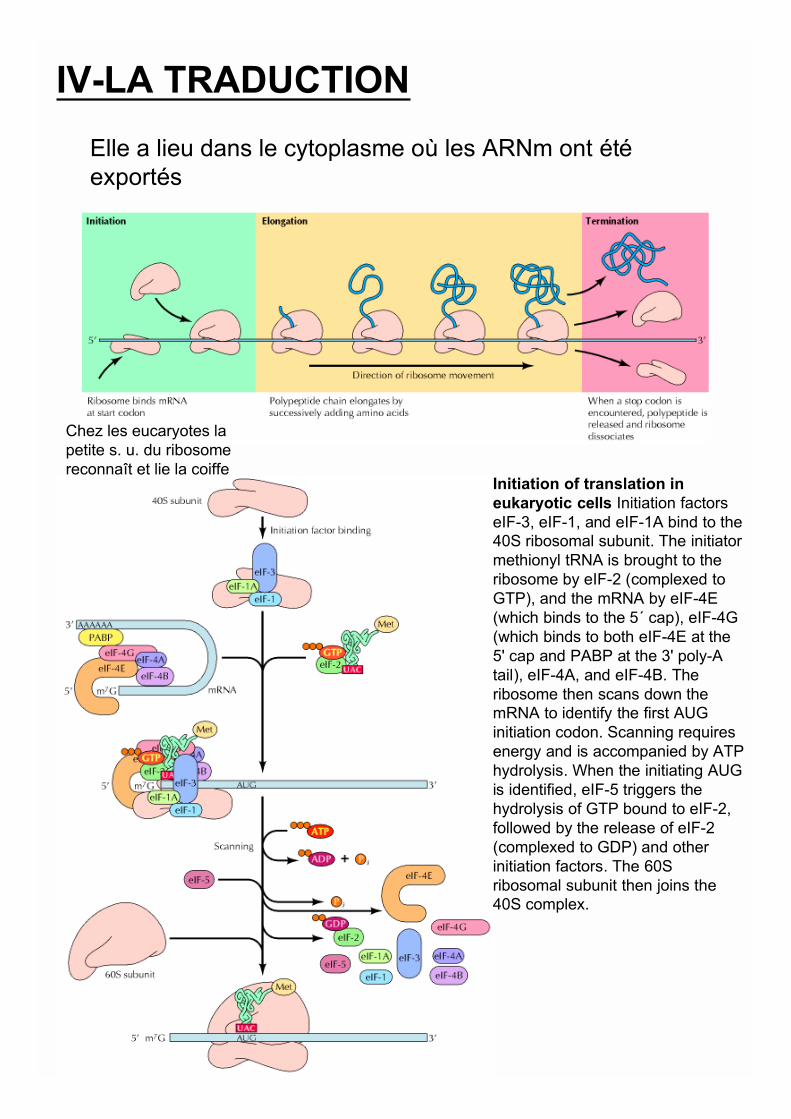
IV-LA TRADUCTION
Elle a lieu dans le cytoplasme où les ARNm ont étéexportés
Chez les eucaryotes lapetite s. u. du ribosomereconnaît et lie la coiffe
Initiation of translation ineukaryotic cells Initiation factorseIF-3, eIF-1, and eIF-1A bind to the40S ribosomal subunit. The initiatormethionyl tRNA is brought to theribosome by eIF-2 (complexed toGTP), and the mRNA by eIF-4E(which binds to the 5´ cap), eIF-4G(which binds to both eIF-4E at the5' cap and PABP at the 3' poly-Atail), eIF-4A, and eIF-4B. Theribosome then scans down themRNA to identify the first AUGinitiation codon. Scanning requiresenergy and is accompanied by ATPhydrolysis. When the initiating AUGis identified, eIF-5 triggers thehydrolysis of GTP bound to eIF-2,followed by the release of eIF-2(complexed to GDP) and otherinitiation factors. The 60Sribosomal subunit then joins the40S complex.
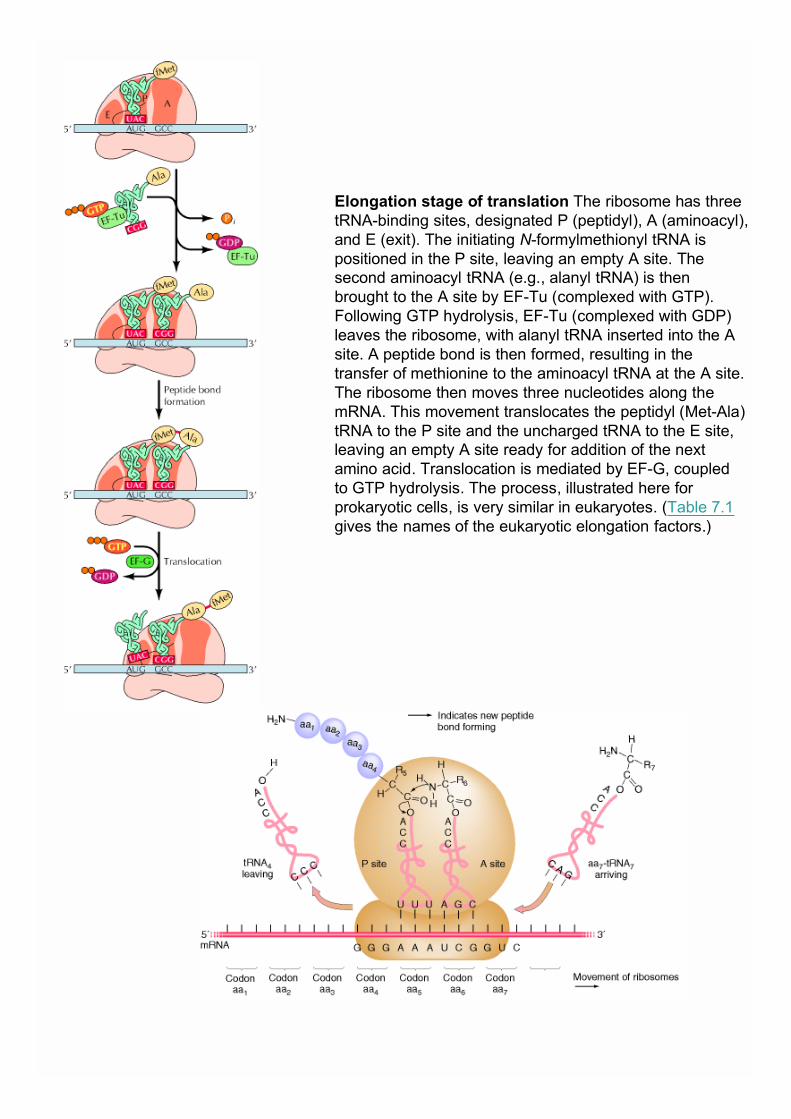
Elongation stage of translation The ribosome has threetRNA-binding sites, designated P (peptidyl), A (aminoacyl),and E (exit). The initiating N-formylmethionyl tRNA ispositioned in the P site, leaving an empty A site. Thesecond aminoacyl tRNA (e.g., alanyl tRNA) is thenbrought to the A site by EF-Tu (complexed with GTP).Following GTP hydrolysis, EF-Tu (complexed with GDP)leaves the ribosome, with alanyl tRNA inserted into the Asite. A peptide bond is then formed, resulting in thetransfer of methionine to the aminoacyl tRNA at the A site.The ribosome then moves three nucleotides along themRNA. This movement translocates the peptidyl (Met-Ala)tRNA to the P site and the uncharged tRNA to the E site,leaving an empty A site ready for addition of the nextamino acid. Translocation is mediated by EF-G, coupledto GTP hydrolysis. The process, illustrated here forprokaryotic cells, is very similar in eukaryotes. (Table 7.1gives the names of the eukaryotic elongation factors.)
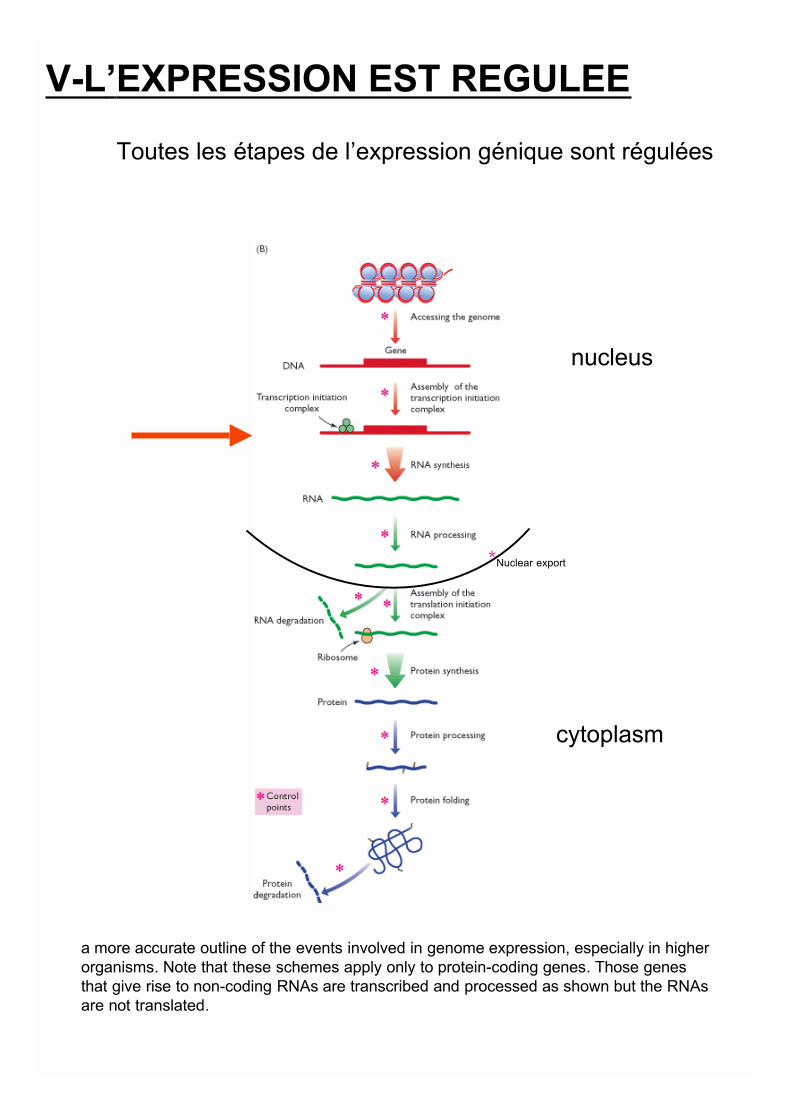
nucleus
cytoplasm
*Nuclear export
a more accurate outline of the events involved in genome expression, especially in higherorganisms. Note that these schemes apply only to protein-coding genes. Those genesthat give rise to non-coding RNAs are transcribed and processed as shown but the RNAsare not translated.
V-L’EXPRESSION EST REGULEE
Toutes les étapes de l’expression génique sont régulées
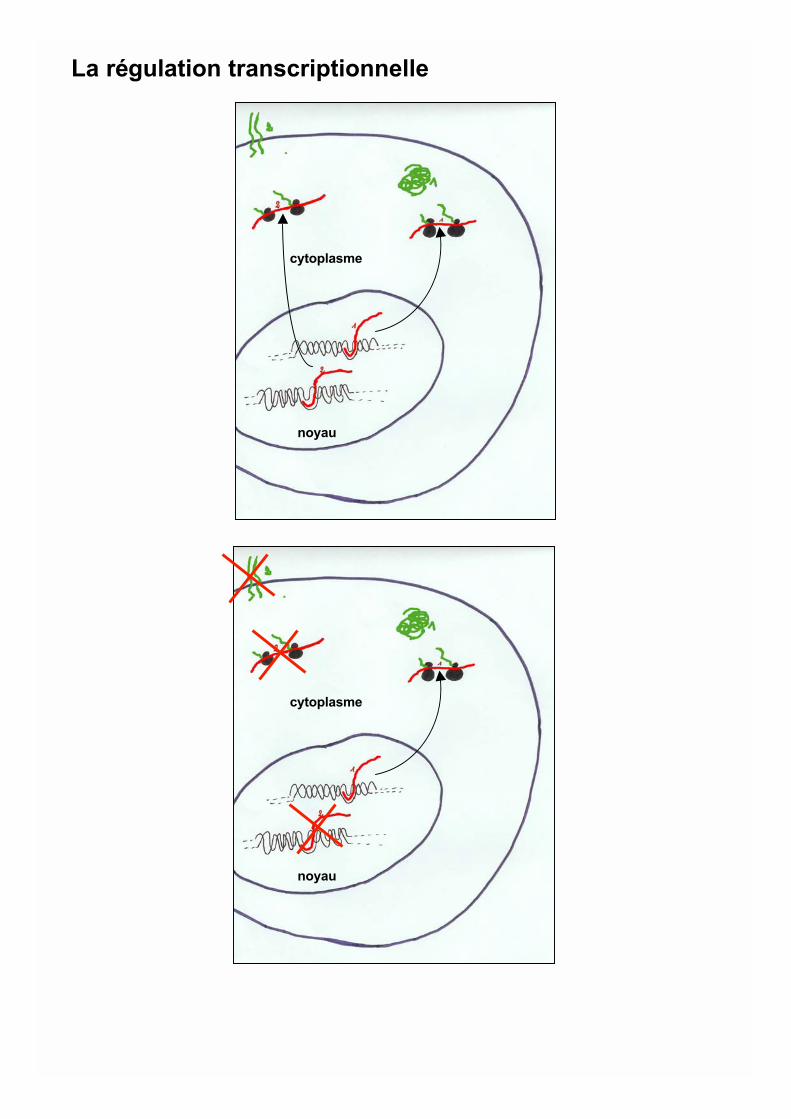
cytoplasme
noyau
cytoplasme
noyau
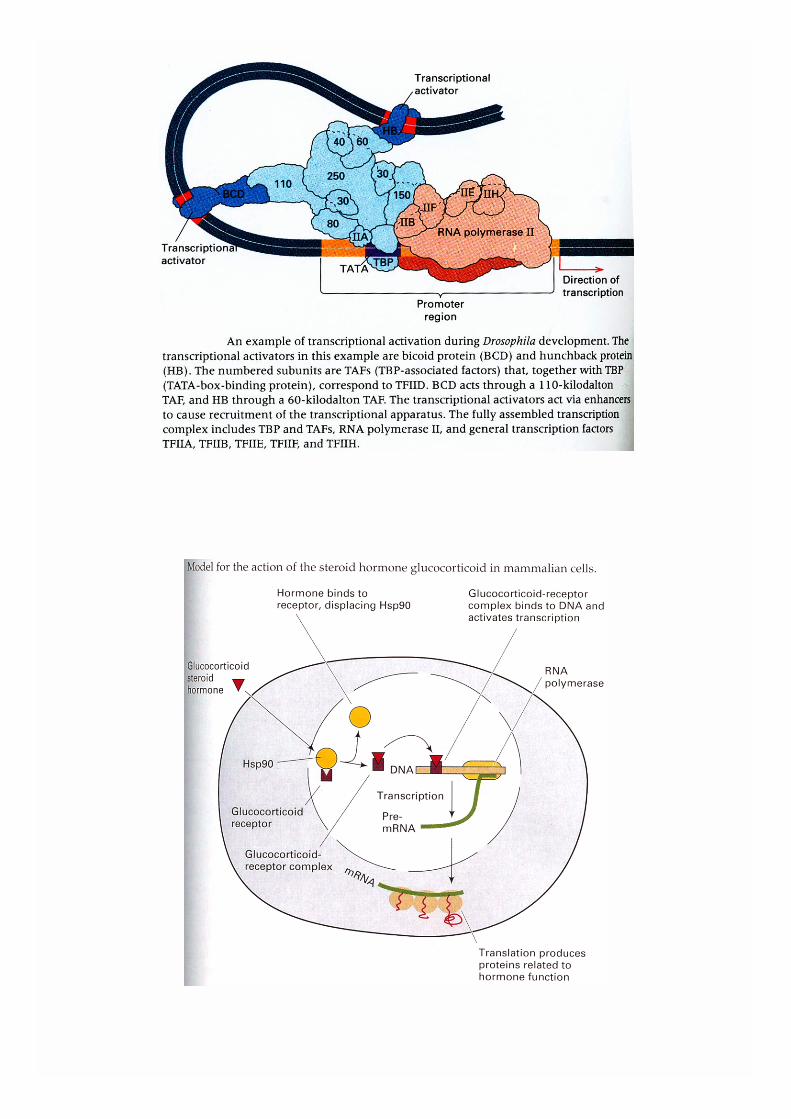
La régulation transcriptionnelle
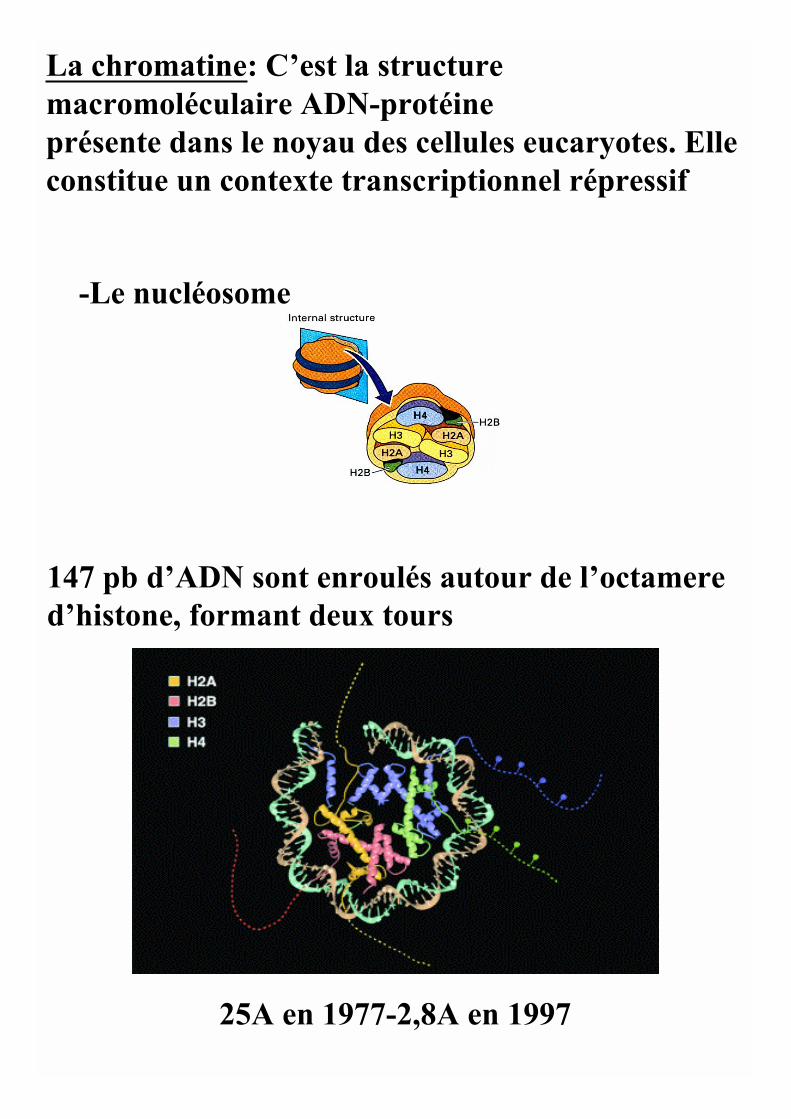
La chromatine: C’est la structuremacromoléculaire ADN-protéineprésente dans le noyau des cellules eucaryotes. Elleconstitue un contexte transcriptionnel répressif
-Le nucléosome
147 pb d’ADN sont enroulés autour de l’octamered’histone, formant deux tours
25A en 1977-2,8A en 1997
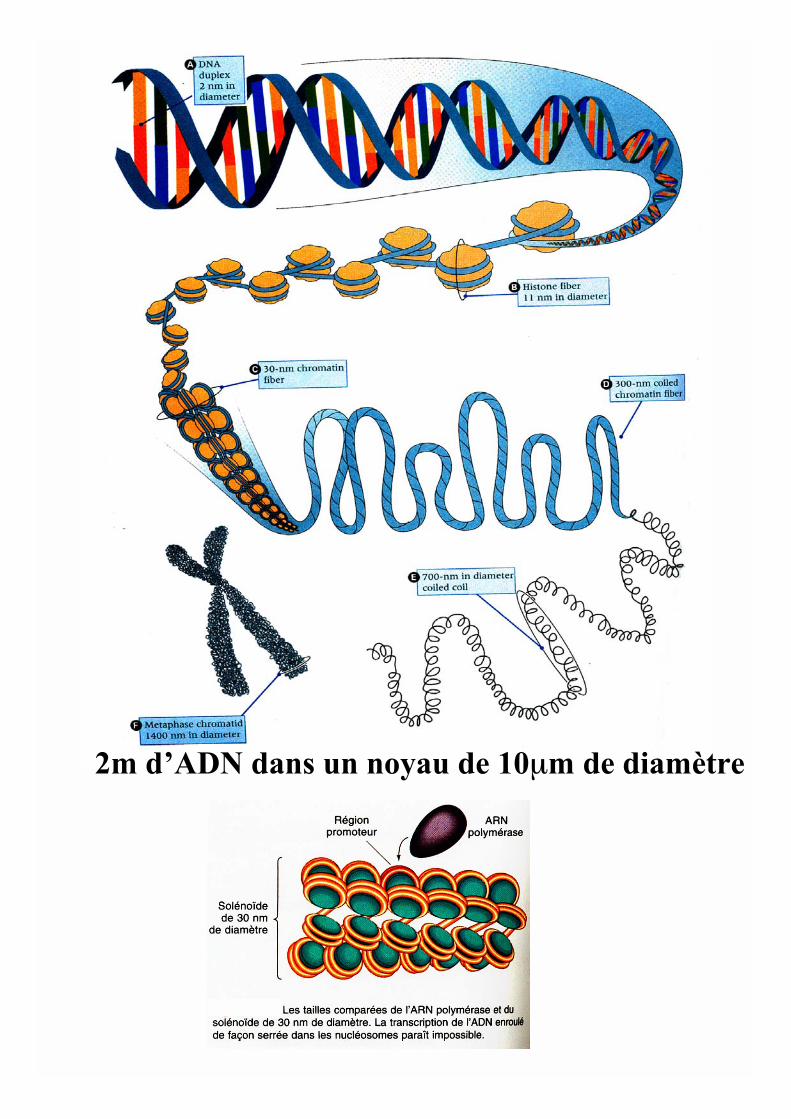
2m d’ADN dans un noyau de 10μm de diamètre

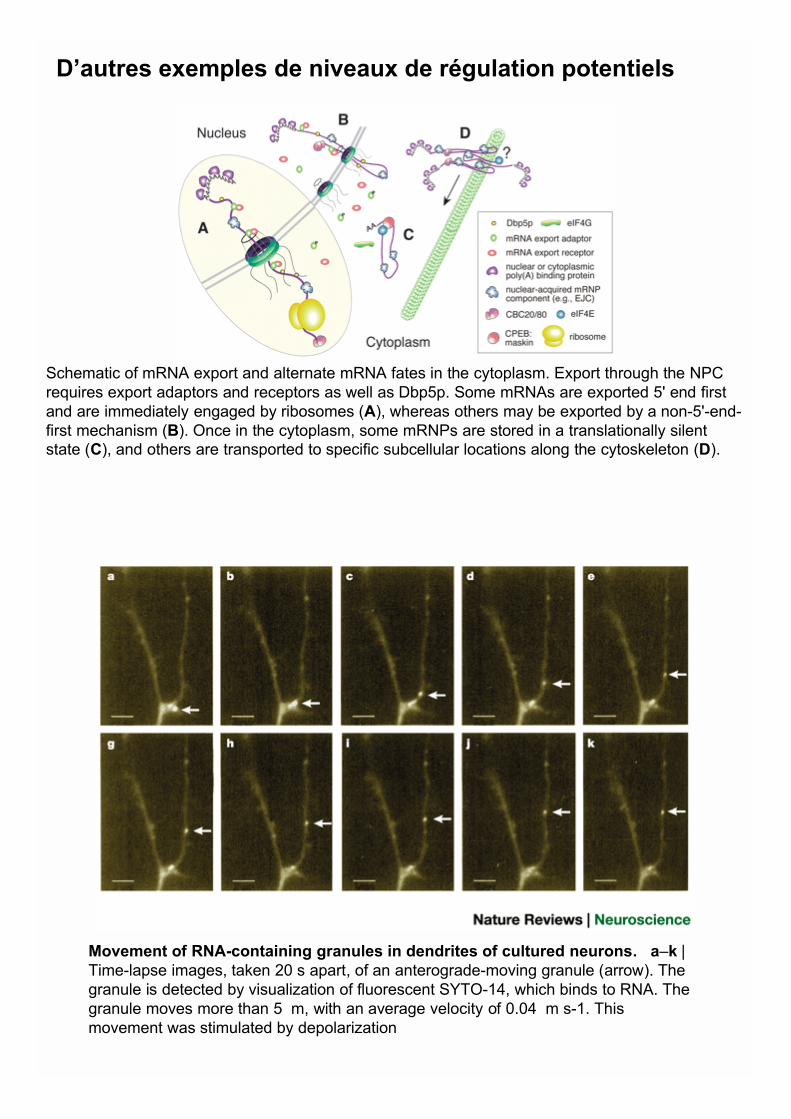
Movement of RNA-containing granules in dendrites of cultured neurons. a–k |Time-lapse images, taken 20 s apart, of an anterograde-moving granule (arrow). Thegranule is detected by visualization of fluorescent SYTO-14, which binds to RNA. Thegranule moves more than 5 m, with an average velocity of 0.04 m s-1. Thismovement was stimulated by depolarization
Schematic of mRNA export and alternate mRNA fates in the cytoplasm. Export through the NPCrequires export adaptors and receptors as well as Dbp5p. Some mRNAs are exported 5' end firstand are immediately engaged by ribosomes (A), whereas others may be exported by a non-5'-end-first mechanism (B). Once in the cytoplasm, some mRNPs are stored in a translationally silentstate (C), and others are transported to specific subcellular locations along the cytoskeleton (D).
D’autres exemples de niveaux de régulation potentiels






![Algorithmes génétiques - old.i2m.univ-amu.frpreaux/PDF/pdf_proteges/Optimisation... · Algorithmes génétiques ! Inventés par Holland dans les années 70 [Holland, 75] pour imiter](https://static.fdocuments.fr/doc/165x107/5b98fbf809d3f22f0a8cf311/algorithmes-genetiques-oldi2muniv-amufr-preauxpdfpdfprotegesoptimisation.jpg)



