
Les polyphénols du thé vert : des molécules à double ...€¦ · polyphénols du thé vert en...
160
LES POLYPHÉNOLS DU THÉ VERT : DES MOLÉCULES À DOUBLE ACTION CONTRE LA MALADIE PARODONTALE Mémoire Marie-Pierre Morin Maîtrise en microbiologie Maître ès sciences (M. Sc.) Québec, Canada © Marie-Pierre Morin, 2015
Transcript of Les polyphénols du thé vert : des molécules à double ...€¦ · polyphénols du thé vert en...

LES POLYPHÉNOLS DU THÉ VERT : DES MOLÉCULES À DOUBLE ACTION CONTRE LA MALADIE PARODONTALE
Mémoire
Marie-Pierre Morin
Maîtrise en microbiologie
Maître ès sciences (M. Sc.)
Québec, Canada
© Marie-Pierre Morin, 2015


iii
RÉSUMÉ
Dans ce projet de recherche, les propriétés antibactériennes et le potentiel anti-destruction tissulaire
des polyphénols du thé vert, plus particulièrement l’épigallocatéchine-gallate (EGCG), ont été évalués.
Dans un premier temps, l’effet antibactérien d’un extrait de thé vert et de l’EGCG a été déterminé sur
trois bactéries parodontopathogènes d’importance, soit Porphyromonas gingivalis, Aggregatibacter
actinomycetemcomitans et Fusobacterium nucleatum. Les concentrations minimales inhibitrices se
sont avérées être entre 1000 et 62,5 µg/ml. De plus, des effets synergiques et additifs des polyphénols
du thé vert ont été observés lorsqu’utilisés en association avec le métronidazole ou la tétracycline, des
antibiotiques couramment utilisés en thérapie parodontale. Dans un deuxième temps, un modèle de
co-culture constitué de fibroblastes gingivaux intégrés dans un gel de collagène et recouvert de
macrophages a été utilisé pour évaluer la capacité des polyphénols du thé vert à inhiber la sécrétion
de métalloprotéinases matricielles (MMPs), notamment les MMP-3, MMP-8 et MMP-9. Il a été
démontré que les polyphénols du thé vert atténuent la sécrétion des MMPs par le modèle de co-culture.
En conclusion, les résultats de cette étude ont apporté des évidences supportant le potentiel des
polyphénols du thé vert en vue d’une utilisation préventive et thérapeutique pour le contrôle des
maladies parodontales.


v
TABLE DES MATIÈRES
RÉSUMÉ ................................................................................................................................................... III
TABLE DES MATIÈRES ............................................................................................................................... V
LISTE DES TABLEAUX .............................................................................................................................. VII
LISTE DES FIGURES .................................................................................................................................. IX
LISTE DES ABRÉVIATIONS ........................................................................................................................ XI
REMERCIEMENTS ....................................................................................................................................XV
1. INTRODUCTION .................................................................................................................................................... 1
1.1 Structure et histologie du parodonte ................................................................................................................. 1
1.1.1 Gencive .................................................................................................................................................. 2
1.1.1.1 Épithélium buccal ......................................................................................................................... 2
1.1.1.2 Épithélium sulculaire .................................................................................................................... 4
1.1.1.3 Épithélium de jonction .................................................................................................................. 4
1.1.1.4 Tissu conjonctif gingival ............................................................................................................... 6
1.1.2 Ligament parodontal ............................................................................................................................... 7
1.1.3 Cément ................................................................................................................................................... 7
1.1.4 Os alvéolaire .......................................................................................................................................... 8
1.2 Écosystème buccal ........................................................................................................................................... 8
1.3 Maladies parodontales ...................................................................................................................................... 9
1.3.1 Généralités ............................................................................................................................................. 9
1.3.1.1 Conséquences systémiques des maladies parodontales .......................................................... 11
1.3.2 Gingivite ............................................................................................................................................... 13
1.3.3 Parodontite ........................................................................................................................................... 14
1.3.3.1 Facteurs étiologiques bactériens ............................................................................................... 15
1.3.3.1.1 Porphyromonas gingivalis ....................................................................................................... 18
1.3.3.1.2 Aggregatibacter actinomycetemcomitans ............................................................................... 28
1.3.3.1.3 Fusobacterium nucleatum ...................................................................................................... 31
1.3.3.2 Facteurs étiologiques immunitaires ........................................................................................... 32
1.3.3.2.1. Métalloprotéinases matricielles ........................................................................................ 35
1.3.4 Traitements des maladies parodontales .............................................................................................. 38
1.4 Polyphénols .................................................................................................................................................... 40
1.4.1 Généralités ........................................................................................................................................... 40
1.4.2 Classification ........................................................................................................................................ 40
1.4.3 Thé ....................................................................................................................................................... 41
1.4.3.3 Thé vert ...................................................................................................................................... 43
1.4.3.4 Composition ............................................................................................................................... 43
1.4.4 Effets bénéfiques du thé vert ................................................................................................................ 44
1.4.4.1 Halitose buccale ........................................................................................................................ 45
1.4.4.2 Carie dentaire ............................................................................................................................ 48
1.4.4.3 Maladies parodontales ............................................................................................................... 51
2. PROBLÉMATIQUE .............................................................................................................................................. 55
2.1 Hypothèse de recherche ................................................................................................................................. 56
2.2 Objectifs spécifiques ....................................................................................................................................... 56
3. MATÉRIEL ET MÉTHODES ................................................................................................................................. 57
3.1 Molécules à l’étude ......................................................................................................................................... 57
3.2 Espèces bactériennes à l’étude et conditions de culture ................................................................................ 58

3.3 Activité antibactérienne des composés .......................................................................................................... 58
3.3.1 Détermination de la concentration minimale inhibitrice ....................................................................... 58
3.3.2 Détermination de la concentration minimale bactéricide ..................................................................... 60
3.4 Évaluation de l’effet synergique ..................................................................................................................... 60
3.4.1 Préparation des microplaques ............................................................................................................. 61
3.5 Culture cellulaire ............................................................................................................................................. 63
3.6 Cytotoxicité des composés à l’étude sur les lignées cellulaires individuelles ................................................ 64
3.7 Mise au point d’un modèle de co-culture macrophages/fibroblastes ............................................................. 65
3.8 Capacité des polyphénols d’intérêt à inhiber la sécrétion de métalloprotéinases matricielles (MMP-3, MMP-8
et MMP-9) par le modèle de co-culture .......................................................................................................... 67
3.9 Analyses statistiques ...................................................................................................................................... 69
4. RÉSULTATS ....................................................................................................................................................... 71
4.1 Activité antibactérienne des composés ...................................................................................................... 71w2
4.1.1 Concentrations minimales inhibitrices et bactéricides ......................................................................... 71
4.1.2 Détermination de l’effet synergique ..................................................................................................... 74
4.2 Inhibition de la sécrétion de métalloprotéinases matricielles (MMP-3, MMP-8 et MMP-9) par le modèle de
co-culture macrophages/fibroblastes ................................................................................................................... 78
4.2.3 Réponse du modèle de co-culture macrophages/fibroblastes stimulé avec le LPS d’A.
actinomycetemcomitans en condition faiblement (ratio 1 :10) et fortement inflammatoire (ratio 10:1) 78
4.2.4 Effet de l'extrait de thé vert et de l’EGCG sur la sécrétion basale des MMPs en conditions faiblement
et fortement inflammatoires .................................................................................................................. 83
4.2.5 Effet du thé vert et de l’EGCG sur la sécrétion des différentes MMPs en conditions faiblement et
fortement inflammatoires à des fins préventives .................................................................................. 88
4.2.6 Effet du thé vert et de l’EGCG sur la sécrétion des différentes MMPs en conditions faiblement et
fortement inflammatoires à des fins thérapeutiques ............................................................................. 93
5. DISCUSSION ...................................................................................................................................................... 99
5.1 Propriétés antibactériennes .......................................................................................................................... 100
5.1.1 Concentrations minimales inhibitrices et bactéricides ....................................................................... 100
5.1.2 Détermination de l’effet synergique ................................................................................................... 103
5.2 Propriétés anti-destruction tissulaire ............................................................................................................ 104
6. CONCLUSION ................................................................................................................................................... 111
7. PERTINENCE DU PROJET .............................................................................................................................. 113
8. BIBLIOGRAPHIE ............................................................................................................................................... 115
9. ANNEXES ......................................................................................................................................................... 141

vii
LISTE DES TABLEAUX
TABLEAU 1. Les différentes classes de composés phénoliques (Tableau adapté de [336]). ........................................... 41
TABLEAU 2. Composition de l’extrait de thé vert GT#9 .................................................................................................... 57
TABLEAU 3. Combinaisons de molécules d’intérêt utilisées lors des essais d’effets synergiques ................................... 61
TABLEAU 4. Détermination des concentrations minimales inhibitrices et bactéricides des polyphénols du thé vert vis-à-vis trois espèces bactériennes parodontopathogènes ......................................................................... 73
TABLEAU 5. Détermination de l’effet synergique des polyphénols du thé vert en association avec le métronidazole ou la tétracycline sur trois espèces bactériennes parodontopathogènes ..................................................... 76
TABLEAU 6. Détermination de l’effet synergique des polyphénols du thé vert en association avec le métronidazole ou la tétracycline sur P. gingivalis ................................................................................................................. 77
TABLEAU 7. Sécrétion des MMP-3, MMP-8 ET MMP-9 par le modèle de co-culture macrophages/fibroblastes à un ratio 1:10 et 10:1 ........................................................................................................................................... 79


ix
LISTE DES FIGURES
FIGURE 1. Représentation schématique des tissus de soutien de la dent (Figure traduite et adaptée de Color Atlas of Histology, fifth edition, 2009 [6]). ...................................................................................................................... 1
FIGURE 2. Évaluation clinique d'un parodonte sain, affecté d'une gingivite ou d'une parodontite (Figure tirée de [55]) .... 11
FIGURE 3. Schéma représentant les interactions bactériennes des 6 complexes de colonisation (Figure tirée de [108]). 17
FIGURE 4. Schématisation du processus de fabrication du thé et composition en polyphénols des divers thés (FIGURE adaptée de [339]. ........................................................................................................................................... 42
FIGURE 5. Structure moléculaire de l’épigallocatéchine-3-gallate. ..................................................................................... 44
FIGURE 6. Disposition des dilutions des composantes à l’étude et des contrôles dans la plaque 96 puits pour les essais de synergie.. ................................................................................................................................................... 62
FIGURE 7. Représentation schématique du modèle de co-culture macrophages/fibroblastes ........................................... 67
FIGURE 8. Sécrétion de la MMP-3 par le modèle de co-culture stimulé avec le LPS d’A. actinomycetemcomitans en condition faiblement inflammatoires (ratio 1 macrophage : 10 fibroblastes) .................................................. 80
FIGURE 9. Sécrétion de la MMP-3 par le modèle de co-culture stimulé avec le LPS d’A. actinomycetemcomitans en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) ..................................................... 80
FIGURE 10. Sécrétion de la MMP-8 par le modèle de co-culture stimulé avec le LPS d’A. actinomycetemcomitans en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) .................................................... 81
FIGURE 11. Sécrétion de la MMP-8 par le modèle de co-culture stimulé avec le LPS d’A. actinomycetemcomitans en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) ..................................................... 81
FIGURE 12. Sécrétion de la MMP-9 par le modèle de co-culture stimulé avec le LPS d’A. actinomycetemcomitans en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) .................................................... 82
FIGURE 13. Sécrétion de la MMP-9 par le modèle de co-culture stimulé avec le LPS d’A. actinomycetemcomitans en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) ..................................................... 82
FIGURE 14. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-3 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) ..................................................................................................................... 85
FIGURE 15. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-3 en condition faiblement inflammatoire (ratio 10 macrophages : 1 fibroblaste) ..................................................................................................................... 85
FIGURE 16. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-8 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) ..................................................................................................................... 86
FIGURE 17. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-8 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) ..................................................................................................................... 86
FIGURE 18. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-9 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) ..................................................................................................................... 87
FIGURE 19. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-9 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) ..................................................................................................................... 87
FIGURE 20. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins préventives .................................................................................... 90
FIGURE 21. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins préventives ...................................................................................... 90
FIGURE 22. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins préventives .................................................................................... 91

FIGURE 23. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins préventives ..................................................................................... 91
FIGURE 24. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins préventives ................................................................................... 92
FIGURE 25. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins préventives ..................................................................................... 92
FIGURE 26. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins thérapeutiques .............................................................................. 95
FIGURE 27. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins thérapeutiques ................................................................................ 95
FIGURE 28. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins thérapeutiques .............................................................................. 96
FIGURE 29. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins thérapeutiques ................................................................................ 96
FIGURE 30. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition faiblement inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins thérapeutiques .............................................................................. 97
FIGURE 31. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition fortement inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins thérapeutiques ................................................................................ 97

xi
LISTE DES ABRÉVIATIONS
spp. ATCC P. gingivalis P. intermedia F. nucleatum A. actinomycetemcomitans T. denticola T. forsythia S. mutans S. sobrinus S. moorei PMN LPS IL IL-1Ra TNF-α TGF-β EGF PDGF PGE2 MMP TIMPS RANK RANKL OPG NF-κB MAPK GT9 EGCG EC ECG EGC ELISA ̊C % h CO2
NaOH RPMI DMEM FBS PMA mM l, ml, µl g, mg, µg
Espèce "American Type Culture Collection" Porphyromonas gingivalis Prevotella intermedia Fusobacterium nucleatum Aggregatibacter actinomycetemcomitans Treponema denticola Tannerella forsythia Streptococcus mutans Streptococcus sobrinus Solobacterium moorei Polymorphonucléaire neutrophile Lipopolysaccharide Interleukine Récepteur antagoniste de l'IL-1 "Tumor necrosis factor"-α "Transforming growth factor"-β "Epidermal growth factor" "Platelet-derived growth factor" Prostaglandine E2 Métalloprotéinases matricielles Inhibiteurs tissulaires de MMPs "Receptor Activator of Nuclear Factor-κ B" Ligand de RANK Ostéoprotégérine "Nuclear factor-kappa B" "Mitogen-activated protein kinase" "Green tea" 9 Épigallocatéchine gallate Épicatéchine Épicatéchine gallate Épigallocatéchine "Enzyme-linked immunosorbent assay" Degré Celcius Pourcentage Heure Dioxyde de carbone Hydroxyde de sodium "Roswell Park Memorial Institute medium" "Dulbecco's modified Eagle's medium" "Foetal bovine serum" "Phorbol myristic acid" Millimolaire Litres, Millilitre, Microlitre Gramme, Milligramme, Microgramme


xiii
« Que ton alimentation soit ton seul médicament »
Hippocrate (460 – 370 av. J.-C.)


xv
REMERCIEMENTS
En préambule à ce mémoire, je souhaite adresser mes remerciements les plus sincères aux personnes
qui m’ont apporté leur aide et qui ont contribué à l’élaboration de ce mémoire ainsi qu’à la réussite de
ma maîtrise. Je témoigne ainsi toute ma reconnaissance aux personnes suivantes, pour l’expérience
enrichissante et pleine d’intérêts qu’elles m’ont fait vivre durant ces dernières années au sein du
Groupe de Recherche en Écologie Buccale (GREB).
On ne peut voir le potentiel et le travail d’une personne avant de lui offrir une première chance. C’est
pour cette raison que je désire d’abord remercier Dr Daniel Grenier, de m’avoir accueillie au sein de
son équipe de laboratoire, et d’avoir pu bénéficier de sa confiance et de son support constant.
J’aimerais remercier les membres de mon comité d’encadrement, Dre Fatiha Chandad ainsi que Dre
Juliana N. Santos. Leur disponibilité de même que leurs critiques constructives ont contribué
grandement à la réalisation de ce projet.
Je tiens également à remercier tous mes collègues du GREB et plus spécifiquement l’équipe du Dr
Daniel Grenier. Mes remerciements s’adressent à Jabrane Azelmat, Katy Vaillancourt et Geneviève
LeBel, professionnels de recherche, Telma Bedran, Amel Ben Lagha ainsi que Bruno Haas (étudiants
au doctorat), Jade Fournier Larente (étudiante à la maîtrise), pour leur appui, leur patience sans limites
et la grande générosité dont ils ont fait preuve, et cela, malgré vos charges de travail respectives, tant
professionnelles qu’académiques.
Merci à mes parents ainsi qu’à mes sœurs qui ont toujours été présents pour moi, qui m’ont soutenue
et encouragée tout au long de mes études. Je tiens également à remercier mon copain pour sa
compréhension, son écoute et ses encouragements. C’est entre autres grâce à eux que j’y suis arrivée!
Enfin, je désire remercier mes amies pour leur présence. Leurs encouragements et leur appui tout au
long de ma maîtrise m’ont permis de persévérer afin d’accomplir avec succès ce projet.


1
1. INTRODUCTION
1.1 Structure et histologie du parodonte
Par définition, le parodonte est un ensemble de tissus entourant la dent et participant au soutien de
celle-ci [1]. Sa principale fonction est de maintenir les dents au sein de l’os alvéolaire tout en conservant
l’intégrité de la surface muqueuse de la cavité buccale [2-4]. Le parodonte comporte plusieurs
structures, dont la gencive, l’os alvéolaire, le cément ainsi que le ligament alvéolodentaire [5] (Figure
1).
Constituant une structure dynamique [2], le parodonte est formé tout au long de la vie par un
arrangement vasculaire unique, un système lymphatique et réseau nerveux hautement spécialisé [2].
Ces quatre structures, devant agir en unité, ont chacune leur importance particulière et leur(s) rôle(s)
spécifique(s) à jouer dans le maintien des organes dentaires au sein de la cavité buccale.
Figure 1. Représentation schématique des tissus de soutien de la dent (figure traduite et adaptée de
Color Atlas of Histology, Fifth Edition, 2009 [6]).

2
Les connaissances actuelles sur l’histologie du parodonte permettent de mieux comprendre les
différents types cellulaires retrouvés au sein des tissus parodontaux, les implications/interactions
possibles entre ces derniers et ce qui se produit d’un point de vue cellulaire et moléculaire lors des
différentes étapes entourant l’initiation et la progression des maladies parodontales.
1.1.1 Gencive
La gencive représente la portion épithéliale du parodonte. Cette dernière recouvre l’os alvéolaire,
protège contre les agressions extérieures [7], et son apparence permet de déterminer l’état de santé
du parodonte [2].
La gencive est divisée en trois composantes majeures. Dans un premier temps, la gencive marginale
(libre) qui est fixée par simple adhérence à la dent sans y être attachée. La gencive libre forme un repli
sur la surface de la dent, communément appelé le sillon gingival. La deuxième composante de la
gencive, la gencive attachée, adhère fermement au cément de la dent et à l’os alvéolaire. Cette portion
de la gencive est affectée lors d’une maladie parodontale et résultera en la formation d’une poche
parodontale et d’une perte d’attache de la dent et la formation d’une poche parodontale. La dernière
composante de la gencive est la muqueuse alvéolaire, caractérisée par une vascularisation plus
visible.
La jonction dento-gingivale est une adaptation de la muqueuse buccale comprenant une composante
épithéliale ainsi qu'une composante conjonctive. Le revêtement épithélial de la gencive est divisé en
trois compartiments fonctionnels ; l’épithélium gingival, sulculaire et jonctionnel.
1.1.1.1 Épithélium buccal
L’épithélium buccal recouvre la gencive libre ainsi que la gencive attachée [2, 8, 9]. Il s’agit d’un
épithélium pavimenteux stratifié kératinisé, composé de quatre couches cellulaires ; la couche basale

3
(stratum germinativum), la couche épineuse (stratum spinosum), la couche granuleuse (stratum
granulosum), ainsi que la couche cornée (stratum corneum) et présentant des digitations épithéliales
dans le chorion gingival [7].
La couche basale (stratum germinativum) représente la couche germinative, puisqu’elle est constituée
de cellules germinales à grand potentiel prolifératif. Cette couche est formée par divers types cellulaires
majoritairement cubiques dont les kératinocytes (90%), qui sont fixés entre eux ainsi qu’à la membrane
basale sous-jacente au moyen des ‘gap’ jonctions et des hémidesmosomes [2]. Ces cellules issues de
la couche basale vont migrer afin de traverser l’ensemble de l'épithélium buccal et, au bout de quelques
jours, atteindre la couche cornée [10]. En plus des kératinocytes, la couche basale est caractérisée
par la présence de mélanocytes (cellules pigmentaires) [11], ainsi que de cellules de Langerhans
(macrophages, cellules immunitaires) qui joueront un rôle dans les mécanismes de défense de la
muqueuse buccale [2, 4, 12, 13]. Les cellules de la couche basale servent à protéger l’intégrité de
l’épithélium ainsi que des structures sous-jacentes et à produire de nouvelles cellules épithéliales. De
plus, ces cellules synthétisent et sécrètent les macromolécules constituant la lame basale et
permettant de séparer l’épithélium du tissu conjonctif, la lamina propria [14].
La strate épineuse (stratum spinosum), supra-adjacente à la couche basale, est composée de
kératinocytes cuboïdes. Ces cellules auront tendance, au cours de leur migration vers les couches
épithéliales apicales, soit la strate granuleuse (stratum granulosum) et la strate cornée (stratum
corneum), à s’aplatir et à suivre un programme de différenciation en cellules mortes remplies de
kératine et entourées d’une couche cornée de protéines et de lipides, constituant ainsi une des
première ligne de défense qui est la barrière épithéliale. Les cellules mortes sont éliminées dans le
sillon gingival par l’intermédiaire du fluide créviculaire [15].
En plus de former une barrière physique, les kératinocytes sécrètent des molécules impliquées dans
l’adhésion cellules-cellules comme l’acide hyaluronique, de même que dans l’adhésion cellules-matrice
comme les intégrines [16]. Ils sécrètent également des collagénases [17], des cytokines telles que le

4
«Tumor Necrosis Factor α» (TNF-α), l’interleukine-1 (IL-1α, IL-1ß), ainsi que des chimiokines dont
l’interleukine-8 (IL-8). Les cellules de l’épithélium gingival buccal libèrent à un niveau basal des
peptides antimicrobiens comme les ß-défensines (hBD) [18].
1.1.1.2 Épithélium sulculaire
L’épithélium sulculaire est une extension de l’épithélium buccal au sein du sillon gingival. Il s’agit d’un
épithélium pavimenteux stratifié, qui, contrairement à l’épithélium buccal, est non kératinisé [2].
Puisqu’il est localisé dans le sillon gingival, ce dernier est constamment en contact avec le biofilm
dentaire sous-gingival. Dans la littérature, il est soupçonné que l’absence de kératine pourrait expliquer
la susceptibilité à l’infiltration bactérienne ainsi qu’à leurs produits potentiellement nocifs [19, 20].
1.1.1.3 Épithélium de jonction
L’épithélium de jonction, la troisième et dernière composante épithéliale du parodonte, est située à une
interface d'importance stratégique entre le sillon gingival, colonisé par des bactéries, et les tissus
conjonctifs parodontaux mous et minéralisés qui doivent être protégés de l’exposition aux bactéries
ainsi qu’à leurs produits. Par conséquent, son adaptation, tant structurelle que fonctionnelle permet à
l'épithélium de jonction de protéger contre la provocation microbiologique constante tout en fournissant
le mécanisme d’attachement de l’épithélium à la dent [2].
Il s’agit d’une mince couche épithéliale située sous le fond du sillon gingival, contre la surface de la
dent (émail), et qui s’étend jusqu’à la jonction amélo-cémentaire [21]. Fondamentalement, il s’agit d’un
épithélium pavimenteux, pluristratifié (malpighien) et indifférencié qui constitue une entité histologique
particulière, puisque ce dernier est caractérisé par un taux très élevé de renouvellement des cellules
[2]. Il est plus large au niveau du fond du sulcus gingival, et se rétrécit jusqu'à une épaisseur de
quelques cellules qui descend le long de l'apex de la dent.

5
La structure permettant l’attachement de l’épithélium à la dent se nomme l’attache épithéliale [22]. D’un
point de vue fonctionnel, l’attache épithéliale permet aux cellules épithéliales spécialisées d’adhérer
aux tissus dentaires calcifiés, protégeant par le fait même les structures sous-jacentes, de l'invasion
par des substances étrangères. D’un point de vue anatomique, l’attache épithéliale consiste en une
lame basale (interne, externe) et des hémidesmosomes, les deux produits par l’épithélium
longitudinalement orienté qu’est l’épithélium de jonction [21]; il est en contact intime avec la surface
extérieure de la dent, soit le cément ou l’émail. Tel un mécanisme dynamique, l'épithélium de jonction
se renouvelle continuellement. De nouvelles cellules ayant l’apparence des cellules de la couche
basale (aplaties et allongées ; orientation longitudinalement et parallèlement à la dent) sont produites
à la base de ce dernier, et les cellules migrent (polarisation de la migration [23] jusqu’à sa surface, tout
en maintenant la fixation biologique de la dent. Ce mécanisme complexe d’attachement consiste en
une séquence d’événements qui se produit dès l’éruption de la dent à travers la muqueuse buccale
[2].
À l’intérieur de l’épithélium jonctionnel sain, il est possible de noter la présence constante d’un infiltrat
de cellules inflammatoires, telles que les monocytes, les macrophages ainsi que les
polymorphonucléaires neutrophiles (PMNs). Ces cellules, occupant de 1 à 3% des espaces
intercellulaires, répondent aux sollicitations continues auxquelles sont soumis les tissus parodontaux
[2, 24, 25]. Les cellules de l’épithélium jonctionnel expriment à leur surface du N-acétyl-lactosamine,
de l’EGF (Epidermal Growth Factor), du LAF-3 (Lymphocyte Activating Factor), de l’IL-8 (interleukine-
8) et de l’ICAM-1 (Intercellular Adhesion Molecule-1) qui leur permet de communiquer avec les cellules
immunitaires [24].
Dans les étapes entourant le déclenchement et la progression de la maladie parodontale, le sillon
gingivo-dentaire se creuse à la suite à la désintégration des cellules superficielles de l’épithélium de
jonction. L’environnement inflammatoire qui accompagne l’éruption avec la présence de nombreux
PMNs au niveau de cet épithélium favorise la désintégration cellulaire [26].

6
1.1.1.4 Tissu conjonctif gingival
Le tissu conjonctif gingival, ou lamina propria, est le tissu prédominant de la gencive [2] et celui-ci
assure, d’un point de vue fonctionnel, l’herméticité ainsi que la protection du parodonte profond [13].
La composante épithéliale de la gencive est séparée de la composante conjonctive par la lame basale,
qui est composée de la lamina densa et de la lamina lucida, assurant la cohésion entre les différentes
composantes de la gencive ainsi que les échanges gazeux et moléculaires entre celles-ci. Le tissu
conjonctif gingival se compose, dans un état sain, d’approximativement 60 à 65% de collagène
organisé en fibres, fibrilles, puis faisceaux de fibres (attache conjonctive ; insertion de fibres dans le
cément) [7, 27, 28], de cellules (8%), représentées majoritairement par des fibroblastes (5%) ainsi que
des cellules immunitaires (3%). Le volume restant, soit près de 35%, est occupé par le compartiment
vasculaire ainsi que les éléments nerveux, incorporés dans une substance fondamentale [2, 27],
composée principalement de diverses glycoprotéines et protéoglycanes [29, 30].
Les fibroblastes sont les principales cellules du tissu conjonctif, puisque ces dernières sont présentes
dans des proportions pouvant aller jusqu’à 65% [27]. Les fibroblastes sont des acteurs importants dans
la physiologie des tissus conjonctifs par des processus successifs de synthèse et de destruction [31-
35] des fibres (collagène I, II, III, IV, réticuline, oxtalalnes, élastiques)[36] ainsi que de la matrice
extracellulaire (glycoprotéines et glycosaminoglycanes ; milieu dans lequel se retrouvent les cellules
et permettant le transport de l’eau, des électrolytes, des nutriments, des métabolites) [2, 37]. Par
conséquent, le capital de tissu conjonctif demeure constant et suffisant pour remplir sa fonction [7].
Les fibroblastes gingivaux participent également à la réponse immunitaire inflammatoire en répondant
à des stimuli provenant de molécules synthétisées et sécrétées par les cellules immunitaires de l’hôte
ou provenant des bactéries parodontopathogènes présentes dans le biofilm sous-gingival [2].
Le tissu conjonctif contient également des leucocytes (neutrophiles, monocytes, macrophages,
lymphocytes, mastocytes, plasmocytes) qui interagissent parfois avec les fibroblastes gingivaux [29,
38]. Les PMNs sont observés en plus grande proportion lorsque le tissu conjonctif est sain ou
faiblement inflammé [26]. En plus de les retrouver dans le tissu conjonctif, ces derniers peuvent être
présents dans l’épithélium de jonction, puis dans le sulcus gingival afin de répondre à la présence
bactérienne de la plaque dentaire sous-gingivale ainsi qu’à leurs métabolites potentiellement nocifs.

7
Au sein du tissu conjonctif gingival, il est également possible de retrouver des populations de
lymphocytes CD4+ et CD8+ [26].
1.1.2 Ligament parodontal
Le ligament parodontal, également appelé desmodonte, est un tissu conjonctif dense d’origine
ectomésenchymateuse. Ce dernier, richement vascularisé et innervé [15, 21], est situé autour de la
racine de la dent et s'étend de la partie interne de l’alvéole dentaire jusqu'au cément [7]. Les cellules
ligamentaires les plus abondantes sont les fibroblastes (65%) qui sont responsables du métabolisme
des composantes de la matrice extracellulaire, dont le collagène [39]. D’un point de vue fonctionnel,
en plus de la synthèse du réseau de fibres de collagène dont il est constitué, le ligament parodontal
ancre la dent au niveau de son alvéole [2] par l'intermédiaire des fibres de Sharpey et absorbe les
forces de mastication ainsi que les forces occasionnées par d'autres mouvements buccaux.
1.1.3 Cément
Le cément est un tissu conjonctif minéralisé d’origine ectomésenchymateuse qui se caractérise par un
dépôt continu au cours de la vie. Cette mince couche de tissu minéralisé (50 à 200 µm) entoure la
racine en recouvrant la dentine radiculaire et permet l’ancrage des fibres du ligament parodontal à la
racine de la dent [2, 7, 15, 24, 39]. En ce qui concerne sa composition, ce tissu possède des similitudes
avec l'os alvéolaire, à l'exception que celui-ci n'est ni vascularisé, ni innervé [21]. Par conséquent, ce
dernier est entièrement dépendant du desmodonte quant à l’apport de ses éléments nutritifs [7]. Le
cément est formé d’une matrice organique qui se compose principalement de collagène (type I et type
IIl), d'eau ainsi que de sels minéraux tels que des cristaux d'hydroxyapatite [40].
D'un point de vue histologique, le cément se divise en deux types distincts; un cément acellulaire et un
cément cellulaire, caractérisés par la présence ou non de cémentocytes à l’intérieur de celui-ci ainsi
que par l’origine des fibres de collagène (fibres de Sharpey) de la matrice (cémentoblastes ou
fibroblastes) [15, 41, 42].

8
1.1.4 Os alvéolaire
L'os alvéolaire est la partie de l’os maxillaire et mandibulaire contenant les alvéoles dentaires [21].
Reconnu comme le principal élément de soutien du parodonte, ce dernier apporte rigidité à cette
structure et assure la fixation des fibres parodontales [2]. Ce tissu conjonctif calcifié comprend une
corticale externe, un os spongieux médian ainsi qu’une corticale externe (lame cribriforme) [2, 21]. Il
possède également de nombreuses perforations à travers lesquelles il est possible de retrouver les
vaisseaux sanguins ainsi que les fibres nerveuses [39]. Le tissu osseux possède la capacité de se
remodeler grâce à l'action des ostéoblastes et des ostéoclastes. Ces derniers sont respectivement
responsables de la formation et de la résorption du tissu osseux selon la demande fonctionnelle et
métabolique [7, 39].
1.2 Écosystème buccal
L’environnement buccal, lieu d’échange direct entre l’organisme et son environnement [43], est un
milieu complexe dans lequel cohabitent plus de 700 espèces de microorganismes [44, 45]. La
microflore buccale résidente est amenée à varier en terme d’abondance et de composition en fonction
des divers sites de l’individu, et selon divers facteurs tels que l’alimentation et le niveau d’hygiène
buccale de celui-ci [46]. Malgré la présence de mycètes, de protozoaires, ainsi que de virus, les
bactéries demeurent toutefois la classe de microorganismes prédominante à l’intérieur de la cavité
buccale [47].
La salive, les dents (plaque dentaire supra-gingivale et sous-gingivale), la muqueuse buccale ainsi que
la langue sont toutes des composantes de la cavité buccale où l’écosystème bactérien diffère, et donc,
où différentes communautés microbiennes distinctes sont retrouvées. Celles-ci sont constituées de
plusieurs genres bactériens comprenant à la fois des bactéries Gram positives et négatives et pouvant
appartenir aux catégories de bactéries anaérobies, strictes ou facultatives, ainsi qu’aux bactéries
aérobies. La salive renferme principalement des espèces appartenant aux genres Streptococcus,
Veillonella et Actinomyces [48]. Sur la surface des dents, Streptococcus spp., Prevotella spp. et
Actinomyces spp. sont principalement retrouvés [45]. Des bacilles Gram positifs de même que des
bactéries du genre Veillonella font partie de la flore bactérienne au niveau de la langue [45], tandis que

9
les Streptococcus sont retrouvés au niveau des muqueuses buccales [49]. Quant au sillon gingival,
une personne montrant une bonne santé buccodentaire voit ce site colonisé majoritairement par une
faible charge bactérienne composée entre autres de bactéries Gram positives et anaérobies
facultatives [50] appartenant en grande partie aux genres Streptococcus et Actinomyces
(Streptococcus sanguis, Streptococcus mitis, Actinomyces naeslundii, Actinomyces viscosus) [21, 51].
Une modification de la flore microbienne en faveur des bactéries Gram négatives anaérobies strictes
(voir section 1.3.3.1) dans le sillon gingival est fortement associée au développement des maladies
parodontales et à la destruction éventuelle des tissus de support de la dent [50]. Ces bactéries
colonisent le sillon gingival sous la forme de plaque dentaire, également appelée biofilm dentaire, qui
peut être supra-gingivale ou sous-gingivale, cette dernière étant un facteur étiologique des maladies
parodontales [21]. Cette structure est constituée essentiellement de bactéries qui adhèrent à la fois à
la surface de la dent et entre elles [21, 52]. Son organisation rigoureuse, et hiérarchisée permet de
créer un environnement stable dans lequel il est possible de retrouver des interactions bactériennes
importantes d’un point de vue adhésif et nutritionnel [52].
1.3 Maladies parodontales
1.3.1 Généralités
Outre la carie dentaire, les maladies des gencives, plus communément appelées maladies
parodontales, constituent le principal problème buccodentaire chez la population adulte. Il est possible
de distinguer deux grands types d’atteinte des tissus de soutien de la dent, soit la gingivite et la
parodontite, celles-ci comportant de nombreux facteurs modifiants ou aggravants. Il est estimé à ce
jour qu’environ 80 % de la population de l’Occident souffrira un jour de l’un ou l’autre de ce type
d’affection [43].
Depuis plusieurs années maintenant, microbiologistes, immunologistes et dentistes travaillent de
concert, avec des méthodes de plus en plus sophistiquées, sur l’étiologie des maladies parodontales.
Récemment, de nouvelles connaissances et de nouveaux concepts en relation avec les pathologies
du parodonte, ont permis d’améliorer le système de classification de celles-ci et ainsi de décrire la
parodontite comme une pathologie inflammatoire d’origine infectieuse et multifactorielle [43].

10
Représentée par un équilibre fragile entre l’agression bactérienne et la réponse de l’hôte, la parodontite
est influencée par divers facteurs innés environnementaux tels que l’âge, le sexe, l’hygiène buccale,
les prédispositions génétiques et hormonales, la situation socio-économique, le stress, l’usage du
tabac ainsi que diverses conditions systémiques telles que le diabète et le sida [53]. Pouvant mener à
la perte éventuelle de la dent, il est primordial de comprendre l’étiologie des maladies parodontales.
L’état sain d’un individu se caractérise par une cohabitation harmonieuse des membres de la flore
microbienne avec les cellules eucaryotes constituant les tissus parodontaux [50]. Cet état d’équilibre
fragile entre l’agressivité bactérienne et la réponse de l’hôte est essentiel au maintien d’une bonne
santé buccodentaire et celui-ci est régulé par trois facteurs, soit les conditions environnementales, les
proportions de bactéries pathogènes/commensales ainsi que le système immunitaire de l’individu [54].
La microflore buccale étant constamment perturbée, dans la mesure où celle-ci ne regagne pas son
état d’origine, elle traversera différents complexes de colonisation suivant une succession écologique
ordonnée. En effet, des modifications quantitatives et qualitatives de cette flore en faveur des bactéries
Gram négatives anaérobies strictes peuvent causer une rupture de l’équilibre et favorise l’apparition
d’une réaction inflammatoire exagérée de l’hôte. Ce type de transition peut ainsi engendrer
l’émergence de manifestations cliniques infectieuses et inflammatoires telles que les maladies
parodontales [50]. Les étapes de progression qui caractérisent la maladie parodontale sont
schématisées dans la figure 2 et seront décrites dans les sections qui suivent.

11
Figure 2. Évaluation clinique d'un parodonte sain, affecté d'une gingivite ou d'une parodontite (figure
tirée de [55])
1.3.1.1 Conséquences systémiques des maladies parodontales
Les maladies parodontales sont des maladies inflammatoires d’origine infectieuse qui affectent, à long
terme, le système d’attache parodontal. Leurs signes et leurs symptômes rencontrent les signes
classiques de l’inflammation, une perte d’attache, une destruction des tissus de soutien de la dent et,
éventuellement, la perte de cette dernière. Bien que les maladies parodontales entraînent des
conséquences au niveau local, il est maintenant connu que cette pathologie pourrait significativement
augmenter le risque de certaines affections systémiques [56].
Au cours des dernières décennies, la relation entre la médecine dentaire et la médecine interne, en
particulier en ce qui concerne le concept de la théorie de l’infection focale, a longtemps été un sujet de
débats. Autrefois, la pathogénie des maladies focales a classiquement été attribuée à des pathologies
de la pulpe dentaire ainsi qu’aux infections périapicales. Par contre, au cours des dernières années,
leur rôle a été délaissé et confronté à l’intérêt croissant de la communauté scientifique quant aux
associations possibles entre l’infection parodontale et les maladies systémiques. En effet, les agents

12
parodontopathogènes et leurs produits, ainsi que les médiateurs de l’inflammation engendrés par cette
présence bactérienne au sein des tissus parodontaux, peuvent pénétrer dans le sang (épithélium
compromis ou ulcéré à l’intérieur des poches parodontales), ce qui provoque des effets systémiques
et/ou contribuant à différentes affections systémiques [57, 58].
Les infections focales d’origine buccodentaire peuvent être définies comme des infections survenant
à distance, de façon généralisées ou localisées à divers appareils ou organes, et qui sont causées par
des microorganismes, directement, ou leurs produits, indirectement, provenant de la cavité buccale
[59]. Bien que ce concept demeure controversé, puisqu’il est difficile de prouver hors de tout doute
l’origine buccale des microorganismes responsables d’infections extrabuccales, ce concept a été
davantage pris en considération récemment par les communautés dentaires et médicales. Des études
cliniques, de même que des expérimentations animales ont permis de suggérer trois principaux
mécanismes physiopathologiques [60-62] :
1. Infection métastatique causée par la translocation de bactéries Gram négatives de la poche
parodontale jusque dans la circulation sanguine (ex. : endocardites bactériennes) [54, 60, 63] :
Les procédures dentaires, comprenant entre autres l’extraction des dents, les traitements
endodontiques, la chirurgie parodontale ainsi que le surfaçage radiculaire, peuvent provoquer
l’introduction de microorganismes buccaux dans le sang ou le système lymphatique [59, 64-
67]. Plus important encore, la présence d’infections chroniques au sein de la cavité buccale,
telle que la parodontite, est une condition importante qui favorise l’entrée des bactéries dans
les tissus conjonctifs sous-muqueux et la circulation sanguine [68, 69]. La mastication, de
même que les procédures d’hygiène buccale de routine peuvent également induire une
bactériémie spontanée, plus particulièrement si les sujets présentent une mauvaise santé
buccodentaire [68, 70]. Les bactériémies spontanées sont plus fréquentes et plus importantes
sur une période cumulée que lors d’un acte isolé [59, 60, 71].
2. Lésion métastatique, telle que les lésions vasculaires, résultant de la circulation de toxines
microbiennes ainsi que des médiateurs pro-inflammatoires [54] : Dans d’autres cas, les agents
infectieux restent au site de la lésion initiale, mais les toxines bactériennes sont libérées et

13
peuvent atteindre un organe ou tissu quelconque via la circulation sanguine et provoquer des
blessures métastatiques. Plus particulièrement, les endotoxines ou le lipopolysaccharide
(LPS), libérés après la mort des bactéries Gram négatives [59], stimulent la production et la
sécrétion de médiateurs de l’inflammation tels que l’IL-1, l’IL-6, l’IL-8, le facteur de nécrose
tumorale (TNF), des lipides bioactifs (prostaglandines E2, thromboxane A2, facteur d’activation
plaquettaire), ainsi que des radicaux libres (oxygène actif, peroxyde d’hydrogène (H2O2),
monoxyde d’azote (NO)) [60]. Ces derniers jouent un rôle important dans le développement
de certaines maladies focales.
3. Inflammation métastatique en raison de la réponse immunitaire face aux
parodontopathogènes et leurs toxines [54, 72] : Des bactéries ou leurs antigènes solubles
entrent dans la circulation sanguine et ces derniers réagissent avec les anticorps circulant pour
former des complexes immuns [59, 60]. Ces complexes immuns peuvent être responsables
d’un certain nombre de réactions inflammatoires aiguës ou chroniques dans les sites de dépôts
[66]. Dans certains cas, certains anticorps contre des bactéries buccales peuvent également
réagir avec les récepteurs tissulaires de l’hôte, résultant en de potentielles lésions
immunitaires. Un tel phénomène de mimétisme microbien a été démontré pour la bactérie S.
mutans dans le tissu cardiaque humain [61, 73].
En se basant sur des études épidémiologiques, de nombreuses associations entre la parodontite
chronique et les maladies cardiovasculaires [59, 74, 75], les maladies respiratoires [59, 76], le diabète
[60], l’ostéoporose [54, 77], les enfants prématurés de faible poids à la naissance (hypotrophie) [54,
59, 60, 78, 79] et, plus récemment, le cancer du pancréas [54, 80], le syndrome métabolique [54, 81],
l’insuffisance rénale chronique [54, 82], la polyarthrite rhumatoïde [54, 59, 83-85] ainsi que les maladies
neurodégénératives telles que la maladie d’Alzheimer [54, 86] ont été proposés et recensés dans la
littérature.
1.3.2 Gingivite
Parmi les deux principaux types de maladie parodontale, la gingivite est l’affection buccale la plus
rencontrée au sein de la population, tous les âges confondus. Une étude épidémiologique réalisée aux

14
États-Unis dénombre que jusqu’à 85 % des enfants seraient atteints [87], et que 80 % à 90 % des
adultes américains souffriraient de maladies parodontales, bien que seulement 5 % à 20 % soient
atteints d’une forme sévère et généralisée de la maladie [88, 89]. La gingivite est définie comme une
inflammation réversible, non spécifique et localisée, qui affecte la gencive libre [90]. C’est la forme la
plus précoce et la moins sévère de la maladie parodontale qui n’entraîne aucune atteinte et destruction
des tissus de support sous-jacents à la dent [55].
L’apparition de la gingivite est intimement liée à une hygiène buccale déficiente et à l’accumulation
conséquente de différentes communautés bactériennes au niveau des sites sous-gingivaux [43, 91].
D’un point de vue clinique, la gingivite se manifeste par la présence d’un ou plusieurs des symptômes
typiques de l’inflammation, soit la rougeur, un œdème ainsi que la présence occasionnelle de
saignements gingivaux lors du brossage/sondage ou de façon spontanée [55]. Dans le but de prévenir
l’apparition de maladies parodontales, plus précisément la gingivite, une bonne hygiène buccale et des
visites régulières chez le dentiste sont nécessaires. Dans une optique thérapeutique, une élimination
mécanique de la plaque sous-gingivale (détartrage) est réalisée par le dentiste. Bénigne dans sa forme
simple, elle nécessite toutefois un suivi régulier. En effet, dans la mesure où aucun traitement n’est
effectué ou que celui-ci n’est pas effectif, la gingivite peut dégénérer et conduire, par la colonisation
des tissus plus profonds du parodonte, à la forme de maladie parodontale la plus sévère et agressive,
la parodontite [54, 92].
1.3.3 Parodontite
Les parodontites sont des infections polymicrobiennes aux étiologies et pathogénies variées [93].
Aujourd’hui considérée comme une maladie inflammatoire progressive comportant plusieurs degrés
de sévérité, elle est davantage retrouvée au sein de la population adulte et chez les personnes âgées
[93].
Étiologiquement, les maladies parodontales, dont la parodontite, sont des infections polymicrobiennes
causées par un groupe spécifique de bactéries opportunistes à Gram négatif [94], et significativement
liées à la présence de plaque dentaire (biofilm) et de tartre. En conséquence à une mauvaise hygiène

15
buccale, le développement de ce type d’affection démontre un manque de maîtrise du facteur bactérien
et résulte en une accumulation et une prolifération de communautés bactériennes regroupées sous la
forme de complexes au niveau des sites sous-gingivaux. Les diverses formes de parodontites sont
associées à une flore bactérienne différente et les sites d’un même sujet peuvent différer en terme de
composition bactérienne [95-97]. Caractérisée par la formation de poches parodontales [98], la
parodontite affecte, de façon irréversible, toutes les structures de soutien de la dent, soit le cément, le
ligament parodontal et l’os alvéolaire [99]. La colonisation des tissus parodontaux par les bactéries
parodontopathogènes entraîne plusieurs dommages tissulaires, de façon directe, en stimulant la
réponse immuno-inflammatoire [21]. D’un point de vue clinique, les parodontites se manifestent par la
présence de douleurs gingivales, de récession gingivale, d’altérations du système de mastication et la
mobilité dentaire [43]. Cette condition pathologique progresse de façon épisodique et oscille entre des
phases cycliques d’exacerbation, de rémission et de latence [100-102].
Il existe différents types de parodontite, tels que la parodontite chronique et la parodontite agressive.
Les microorganismes qui sont associés à ces deux catégories peuvent différer. La résultante demeure
toutefois la même, soit la présence d’inflammation gingivale, la formation de poches parodontales et
la destruction des tissus contribuant au soutien de la dent [43, 92]. La parodontite chronique est la
forme la plus commune et montre un taux de progression de lent à modéré. Elle affecte toutes les
catégories d’âge, mais la prévalence et l’étendue des destructions parodontales augmentent en
fonction du vieillissement et d’une hygiène buccale inadéquate [103-105]. Porphyromonas gingivalis
est reconnu comme l’un des principaux agents étiologiques responsables de cette forme de
parodontite [54, 94], et joue également un rôle dans les formes agressives généralisées [106]. La
parodontite agressive, quant à elle, affecte une faible proportion de la population et montre une
progression très rapide en ce qui a trait à la perte d’attache des tissus parodontaux et la résorption de
l’os alvéolaire. Ce type d’atteinte possède une composante génétique, puisque le sujet atteint peut
présenter une bonne santé générale [105]. Aggregatibacter actinomycetemcomitans est une bactérie
pathogène fortement associée à certaines formes de parodontites agressives [98], localisées ou
généralisées, réfractaires, et plus rarement à la parodontite chronique [106].
1.3.3.1 Facteurs étiologiques bactériens

16
Bien que les bactéries puissent se retrouver à l’état planctonique à l’intérieur de la cavité buccale [43],
la majorité d’entre elles s’organisent en un biofilm bactérien, plus connu dans le domaine clinique sous
le nom de plaque dentaire [52]. Il s’agit de communautés bactériennes qui adhèrent à la fois entre elles
ainsi qu’à la surface des dents pour former une organisation complexe et structurée [43].
Il est possible de distinguer deux types de plaque dentaire selon leur localisation, soit la plaque supra-
gingivale et la plaque sous-gingivale. La plaque supra-gingivale se retrouve sur la surface de l’émail
des dents et est baignée par la salive, constituant un environnement aérobie. Quant à la plaque sous-
gingivale, elle se retrouve au niveau du sillon gingival et par conséquent, est entièrement submergée
par le fluide créviculaire, qui crée un milieu anaérobie [107]. Continuité apicale de la plaque supra-
gingivale, la plaque sous-gingivale est principalement à l’origine des maladies parodontales [21].
La pellicule exogène acquise, constituée de glycoprotéines salivaires qui se déposent peu après le
brossage sur la surface des dents, représente la première phase de formation de la plaque supra-
gingivale et permet alors l’adhésion de microorganismes [52]. La colonisation subséquente, se faisant
de façon ordonnée et organisée, est répartie, comme le démontre la Figure 3, sous la forme de 6
complexes bactériens regroupant les colonisateurs primaires et secondaires.

17
Figure 3. Schéma représentant les interactions bactériennes des 6 complexes de colonisation (figure
tirée de [108]).
Les espèces pionnières, colonisatrices primaires, s’attachent de façon réversible à cette pellicule
exogène à l’aide de leurs adhésines de surface. Cette catégorie comprend majoritairement des
bactéries Gram positives telles que les espèces du genre Streptococcus et les Actinomyces
(complexes jaune, vert, violet et bleu). Ces complexes favorisent le développement de conditions
d’anaérobiose et par le fait même, la prolifération de colonisateurs secondaires, des bactéries Gram
négatives anaérobies strictes incluant les espèces du genre Fusobacterium, Prevotella, Streptococcus,
Peptostreptococcus, Eubacterium et Campylobacter [109]. Ces bactéries, appartenant au complexe
orange, viennent soit remplacer ou joindre les espèces bactériennes déjà présentes au sein du biofilm
et agissent à titre d’intermédiaires entre les colonisateurs primaires et les bactéries du complexe rouge,
bactéries colonisatrices secondaires [110]. Les bactéries qui se développent au sein de la plaque sous-
gingivale telles que P. gingivalis, T. denticola et T. forsythia sont les bactéries constituant le complexe
rouge et qui sont le plus souvent associées au développement des maladies parodontales [108, 111].
Leur colonisation provoque l’approfondissement du sillon gingival pour former une poche parodontale,

18
à l’intérieur de laquelle se développe un écosystème bactérien difficile à éliminer [43]. Les colonisateurs
secondaires synthétisent une matrice d’exopolysaccharides conférant au biofilm un environnement
difficile d’accès aux cellules du système immunitaire ainsi qu’aux agents antimicrobiens. De plus, la
virulence de cette structure stable est accentuée, puisque les structures antigéniques sont
continuellement libérées dans le milieu et contribuent à la destruction des structures du parodonte [52,
112].
Les bactéries du complexe rouge possèdent plusieurs facteurs de virulence leur permettant à la fois
une colonisation efficace des sites sous-gingivaux, d’échapper aux défenses de l’hôte afin de créer
des dommages au niveau des tissus constituant le parodonte [113]. Les fimbriae, les adhésines de
surfaces de même que certaines protéines de la membrane externe permettent l’adhésion des
bactéries parodontopathogènes aux cellules de l’hôte [113]. La prolifération bactérienne et la
colonisation efficace des sites de l’hôte sont assurées par des protéines de la membrane externe, des
vésicules membranaires, le lipopolysaccharide (LPS) ainsi que des exopolysaccharides [113]. De plus,
les bactéries pathogènes peuvent endommager les tissus de façon directe, en élaborant des enzymes
protéolytiques qui causent la dégradation des protéines de l’hôte telles que la fibronectine et le
collagène. De façon indirecte, la destruction des tissus du parodonte peut être la conséquence de
l’activation des mécanismes inflammatoires et immunitaires de l’hôte par le LPS menant à la sécrétion
de médiateurs inflammatoires tels que les cytokines et les métalloprotéinases matricielles (MMPs)
[114].
1.3.3.1.1 Porphyromonas gingivalis
P. gingivalis est une espèce bactérienne dominante de la flore cultivable de la poche parodontale en
phase active de destruction [7]. En effet, de nombreuses études ont démontré que ce membre du
complexe rouge est une espèce d’importance, tant en prévalence qu’en nombre, dans la plupart des
lésions des parodontites chez l’adulte, et plus spécifiquement la forme chronique, tandis que ce dernier
ne se retrouve pas, ou en très faibles proportions chez un individu sain [115].

19
P. gingivalis est un coccobacille non-motile à Gram négatif, asaccharolytique et anaérobie strict,
mesurant approximativement 1 µm de diamètre par 2 µm de long [7, 113]. Lorsque cultivée sur gélose
sang enrichie d’hémine et de vitamine K, cette bactérie se caractérise par la formation de colonies à
pigmentation noire. Cette particularité résulte de l’accumulation au niveau de la membrane
bactérienne, d’un pigment identifié comme le fer (III) protoporphyrine IX sous la forme µ-oxo dimère
[Fe(III)PPIX]2O dérivant de l’hémine (fer) contenue dans le milieu de culture [116-119].
Ce parodontopathogène est le microorganisme le plus intensément étudié d’un point de vue
moléculaire et sa pathogénicité est attribuée à un ensemble de facteurs de virulence, dont la capsule,
les fimbriae, les enzymes protéolytiques (cystéines protéinases; gingipaïnes), les hémagglutinines, le
lipopolysaccharide (LPS) ainsi que les vésicules membranaires [7, 113, 116, 120-122]. Bien que la
coopération bactérienne soit importante dans le déclenchement et la progression de la parodontite
[123], les facteurs de virulence propres à P. gingivalis lui permettent de coloniser les sites sous-
gingivaux, de contourner les mécanismes de défense immunitaire et d’engendrer la destruction
tissulaire et osseuse caractéristique de la parodontite.
En plus d’être un contributeur prédominant dans le processus infectieux des maladies parodontales,
P. gingivalis est également impliqué dans diverses conditions systémiques, notamment les maladies
cardiaques (athérosclérose, ACV), le diabète, les pneumonies d’aspiration, les accouchements
prématurés d’enfants de faible poids à la naissance ainsi que l’arthrite rhumatoïde [72, 76, 120, 124-
133].
1.3.3.1.1.1 Facteurs de virulence de Porphyromonas gingivalis
1.3.3.1.1.1.1 Capsule
Lorsqu’un agent pathogène s’introduit dans l’organisme, le système immunitaire répond à cette
nouvelle infection de deux manières; soit par la détection et la reconnaissance de cet organisme
invasif, suivie de la destruction de celui-ci par une réponse immunitaire très complexe et élaborée.

20
La surface des microorganismes pathogènes constitue l’interface centrale entre l’hôte et la bactérie
pathogène, et la reconnaissance de diverses composantes de surface (LPS, peptidoglycane, ADN
bactérien [134]) par le système immunitaire est essentielle pour initier les processus d’élimination de
cette dernière [135]. La stratégie la plus efficace employée par les bactéries parodontopathogènes afin
d’éviter leur reconnaissance par les cellules de l’hôte [136], tout en exposant des molécules clés dans
le processus d’invasion (adhésines, invasines) est d’empêcher la reconnaissance des motifs
moléculaires associés aux pathogènes (PAMPs) se retrouvant à leur surface [135, 137, 138]. Pour ce
faire, de nombreuses bactéries buccales Gram négatives telle que P. gingivalis possède une capsule
de polysaccharides chargée négativement et hydrophile [139] recouvrant sa membrane externe, et
dont la synthèse est modulée par divers facteurs environnementaux [140-142]. La capsule de ce
microorganisme a été démontrée comme un important facteur dans l’établissement d’infections
expérimentales, plus particulièrement dans un modèle murin [139, 142, 143]. Dans la littérature
actuelle, six sérotypes capsulaires distincts (K1- K6) du point de vue de la composition de la capsule
et de la virulence sont actuellement décrits [7, 139, 142, 144]. La capsule est considérée comme un
facteur de virulence important puisque celle-ci permet à la bactérie de résister à l’action de diverses
substances présentes dans la salive (lysozyme, ions métalliques toxiques) [145], tout en prévenant le
dépôt des anticorps et des protéines du complément sur leur surface, leur permettant ainsi d’échapper
à l’opsonisation et à la phagocytose qui en résulte (activation innée des défenses adaptatives) [135,
136, 142]. Réduisant la capacité des cellules effectrices de l’hôte à avoir accès aux bactéries invasives,
les mécanismes de défenses contre ces bactéries encapsulées doivent être accrus.
1.3.3.1.1.1.2 Fimbriae
De nombreuses études épidémiologiques ont suggéré que P. gingivalis est un acteur important dans
le déclenchement et la progression des maladies parodontales [108, 146, 147]. Bien que les
mécanismes pathogéniques impliqués ne soient pas encore entièrement élucidés, l’adhésion
bactérienne et la colonisation subséquente des tissus de l’hôte sont des étapes cruciales afin que la
bactérie puisse exercer son pouvoir pathogène. Les fimbriae, des filaments longs et fins organisés de
façon péritriche à la surface de la bactérie, ont été reconnus pour exercer ces fonctions [121, 148-150].

21
À ce jour, les fimbriae sont considérés comme des facteurs essentiels intervenant dans les interactions
bactériennes avec les cellules de l’hôte et l’invasion des tissus.
Des examens ultrastructuraux des différentes souches de P. gingivalis ont montré que ces structures
varient entre 0,3 à 3 µm de long et 5 nm de diamètre [121, 149-152]. Dans la littérature, il a été
démontré que P. gingivalis exprime à sa surface deux types de fimbriae distincts; les fimbriae majeurs
de type 1 (FimA; longs), ainsi que les fimbriae mineurs (Mfa; courts). Tous deux semblent impliqués
dans le développement des maladies parodontales [147, 149, 153, 154]. Les fimbriae majeurs de
type 1 (FimA) sont constitués d’une juxtaposition de sous-unités d’une protéine, la fimbrilline, cette
dernière étant sous le contrôle du gène fimA [7, 149, 155], présent qu’en un seul exemplaire sur le
chromosome de la bactérie [149, 156-158]. En se basant sur la variation de la séquence nucléotidique,
le gène a été classé en six types (I, Ib, II, III, IV, V) [147, 149, 156, 159-162]. Il a été suggéré que les
fimbriae majeurs, dus à leur longueur, sont les premières composantes bactériennes à interagir autant
avec les autres espèces bactériennes que les cellules de l’hôte dans les sites sous-gingivaux [163].
Ces interactions de coagrégation/coadhésion sont possibles par l’intermédiaire de différents
récepteurs présents sur la surface bactérienne, dont la glycéraldéhyde -3-phosphate déshydrogénase
(GAPDH) chez certains streptocoques ainsi que la dentilisine de T. denticola [149, 155].
Les fimbriae mineurs sont des homopolymères provenant du gène mfa1 codant pour la sous-unité
protéique Mfa [7, 149, 155]. Ce type de fimbriae permet également la coadhésion aux bactéries déjà
présentes [149, 164-167].
Les fimbriae, en plus d’être responsables des propriétés adhésives de ce microorganisme et de médier
l’adhérence de celui-ci à une grande variété de molécules et substrats au niveau de la cavité buccale
[168], stimulent diverses cellules de l’hôte (cellules épithéliales, monocytes) via l’activation de la voie
NF-κB (intermédiaire TLR-2, CD14) résultant en la sécrétion de diverses cytokines pro-inflammatoires
(IL- 1β , IL-6 et TNF-α) [149].

22
1.3.3.1.1.1.3 Hémagglutinines
Les protéines membranaires, telles que les hémagglutinines, jouent un rôle crucial dans l’adhésion
de P. gingivalis dans les sites sous-gingivaux [169-171]. Bien que cinq hémagglutinines différentes
(HagA à HagE) soient exprimées à la surface cellulaire, ces dernières sont toutes impliquées dans la
liaison de ce microorganisme aux récepteurs des cellules humaines, essentiellement des
oligosaccharides [93, 121, 171].
De plus, P. gingivalis possède une activité hémolytique par l’intermédiaire des hémagglutinines. Ces
protéines membranaires permettent de lier les érythrocytes puis de les lyser, libérant ainsi de
l’hémoglobine, une source de fer essentielle à la croissance de ce microorganisme [121, 172-174].
1.3.3.1.1.1.4 Gingipaïnes
Certaines bactéries impliquées dans l’étiologie de la maladies parodontale possèdent un fort pouvoir
protéolytique, qui, en plus de permettre la dégradation des protéines de l'hôte dans un contexte
nutritionnel, interfère avec les mécanismes de défense mis en place par l'hôte lors de la réponse
immunitaire [175], en plus de causer la destruction tissulaire et osseuse [116, 176]. Ces protéinases
sont considérées comme un facteur de virulence important, puisqu'elles permettent la survie et la
prolifération de la bactérie à l'intérieur de l'hôte, ce qui représente un aspect fondamental de la
pathogénèse bactérienne [138].
P. gingivalis est une bactérie qui possède à sa surface et sécrète sous forme soluble une multitude de
protéinases ayant différentes spécificités (cystéine endoprotéinase ; gingipaïnes, collagénases,
dipeptidyl aminopeptidase IV), dont les gingipaïnes [175, 177, 178]. L'activité protéolytique de P.
gingivalis la mieux caractérisée est reliée à trois cystéines protéinases possédant une activité
amidolytique pour des acides aminés spécifiques, appelées gingipaïnes R (RgpA et RgpB ; cystéines
protéinases spécifiques à l'arginine) et K (Kgp ; cystéine protéinase spécifique à la lysine), qui sont,
selon plusieurs auteurs, fortement associées à la virulence de cette bactérie [116, 122, 177, 179, 180].

23
Les Arg-gingipaïnes A et B, de même que la Lys-gingipaïne clivent la région carboxy-terminale de leur
résidu respectif, soit l’arginine ainsi que la lysine; cette activité catalytique étant liée à la présence d’un
groupement thiol au site actif de l’enzyme [181].
Trois gènes codent pour les cystéines protéinases de P. gingivalis:
rgpA code pour la protéinase spécifique à Arg-X RgpA (95 kDa), qui possède un domaine
catalytique ainsi qu'un domaine adhésine/hémagglutinine (hag sequence domain)(HA);
rgpB code pour la protéinase spécifique à Arg-X RgpB (50 kDa) qui ne possède pas le domaine
adhésine/hémagglutinine à l'exception d'une petite extrémité C-terminale codant pour le
domaine catalytique; et
kgp code pour la protéinase spécifique à Lys-X Kgp (≈ 110 kDa), qui possède un domaine
catalytique ainsi qu'un domaine adhésine/hémagglutinine (hag sequence domain)(HA).
Ainsi donc, RgpA ainsi que Kgp possèdent des domaines hémagglutinines qui contribuent à la
colonisation de la cavité buccale par P. gingivalis [177, 182-186].
P. gingivalis est une bactérie asacharolytique qui nécessite de courts peptides ainsi que des acides
aminés afin de subvenir à ses besoins énergétiques [121, 187]. Cette acquisition de nutriments est
possible suivant la dégradation des protéines environnantes par les protéinases, plus spécifiquement
les gingipaïnes. Ces dernières peuvent dégrader un certain nombre de substrats potentiellement
importants au milieu du sillon gingival, incluant le collagène, la fibronectine, le fibrinogène et la laminine
[121, 188].
Le fer est également un facteur de croissance indispensable à P. gingivalis, qui utilise l’hémine (fer
protoporphyrine IX) afin de satisfaire ses besoins nutritionnels [121, 188-191]. Un certain nombre de
composés contenant du fer ou des dérivés de l’hémine, tels que l’hémoglobine, la transferrine [192],
l’haptoglobine, la myoglobine, l’hémopexine, la méthémoglobine, l’oxyhémoglobine, l’albumine, la
lactoperoxydase, la catalase ainsi que le cytochrome c peuvent être dégradés par les protéases de P.

24
gingivalis à des fins nutritionnelles [121, 188-191]. De plus, grâce à l’hémolysine, ce
parodontopathogène s’approprie du fer en libérant l’hémoglobine des érythrocytes [121, 193].
Bien que la fonction primaire des protéases sécrétées par les bactéries asaccharolytiques telle que P.
gingivalis est de fournir de courts peptides pour subvenir à ses besoins énergétiques (croissance), les
protéases sont également impliquées directement dans l’adhérence et la colonisation des tissus [121,
194]. En effet, la colonisation des sites sous-gingivaux par P. gingivalis est facilitée par sa capacité à
adhérer à de nombreuses surfaces biotiques, dont les cellules épithéliales, la matrice extracellulaire,
de même que les globules rouges et les autres bactéries. Bien que cette adhérence puisse être médiée
par les fimbriae, les gingipaïnes de P. gingivalis peuvent également être impliquées, par des
mécanismes directs ou indirects [195]. De plus, le fort potentiel protéolytique de P. gingivalis peut
favoriser l’adhérence des parodontopathogènes aux tissus gingivaux. Ainsi, la dégradation de la
fibronectine (matrice extracellulaire) permet la libération et l’exposition des cryptitopes à la surface des
cellules eucaryotes, des sites d’adhésion masqués [196] permettant l’adhérence des bactéries [197-
200].
Puisque la bactérie P. gingivalis est considérée comme un intrus pour le corps humain, cette dernière
deviendra une cible naturelle pour le système immunitaire de l'hôte, qui tentera de l’éliminer en
déployant divers mécanismes impliqués dans la réponse immunitaire innée, non-spécifique
(complément, peptides antimicrobiens, neutrophiles et macrophages résidents), de même que dans la
réponse immunitaire adaptative, plus spécifique (anticorps, macrophages activés, lymphocytes T)
[116, 176]. Ces deux composantes de la réponse immunitaire sont en constante interaction et sont
synchronisées par diverses cytokines [195]. Afin de survivre aux attaques du système immunitaire, P.
gingivalis possède la capacité de manipuler ces deux branches de la réponse immunitaire, et il est
soupçonné que les gingipaïnes constituent des acteurs clés dans ce processus.
L’invasion bactérienne induit, dans la cavité buccale, l’expression de peptides antimicrobiens
cationiques tels que les β-défensines, la cathélicidine LL-37 ainsi que les α-défensines dérivées des

25
neutrophiles [201]. Il a été démontré que les gingipaïnes de P. gingivalis dégradent ces protéines,
perturbant par le fait même l’une des premières ligne de défense de la réponse immunitaire innée et
permettant la survie de cette bactérie au sein des lésions parodontales [195, 202-205].
Le système du complément joue un rôle significatif dans la réponse immunitaire innée selon 3 voies :
la voie classique, la voie des lectines ainsi que la voie alternative. Ce système en cascade implique
plusieurs glycoprotéines aboutissant à la formation d’un complexe d’attaque membranaire (MAC),
composés des facteurs C5b, C6, C7, C8, C9. Ce dernier causera la mort des microorganismes
pathogènes [206]. Les gingipaïnes interviennent dans les étapes initiales de la cascade du complément
en dégradant les composantes C3, C4 et C5, empêchant l’activation de la composante C5b et la
formation subséquente du MAC [7, 176, 195, 207-211]. De plus, la dégradation du récepteur C5a
résulte en une suppression de la migration des neutrophiles dans les sites infectés [116, 212, 213].
Bien que le système du complément permette une élimination efficace des microorganismes
pathogènes, une mauvaise régulation de ce dernier peut engendrer la destruction tissulaire. Afin de
prévenir cette dégradation, les cellules de l’hôte expriment un certain nombre de protéines régulatrices
liées à la membrane, dont le CD46, un cofacteur permettant l’inactivation des facteurs C3b et C4b du
complément [207, 214, 215]. Les protéases de P. gingivalis possèdent la capacité de cliver le CD46,
provoquant son relargage et favorisant l’activation du complément et par conséquent, la progression
de la maladie parodontale [195, 207, 216, 217].
Le système immunitaire inné comprend de nombreux types cellulaires, et le rôle d'élimination des
agents pathogènes ayant envahi l'organisme est accompli majoritairement par les neutrophiles ainsi
que les macrophages résidents dans les tissus. Les gingipaïnes de P. gingivalis ont la capacité
d'entraîner une dysfonction des cellules phagocytaires en clivant le récepteur FMLP (formyl-Met-Leu-
Phe) [116]. Cette dégradation résulte en une inactivation des neutrophiles et une incapacité de ceux-
ci à reconnaître les agents pathogènes envahisseurs [116, 176, 218].

26
Il est également connu que P. gingivalis puisse déréguler les mécanismes de la défense immunitaire
de l’hôte par le biais des cytokines [116, 219, 220], incluant les protéines de signalisation ainsi que
leurs récepteurs. Il a été démontré que les gingipaïnes dégradent et inactivent plusieurs composantes
de ce réseau, dont l’IL-1β, l’IL-4, l’IL-6, l’IL-8, l’IL-12, le récepteur de l’IL-6, l’interféron-α, le TNF-α, le
CD4, le CD8, le CD14 et le CD54 [194, 195, 221-223], résultant en une interruption de communication
entre les deux branches de la réponse immunitaire. La dégradation des cytokines résulte en une forte
perturbation des réactions inflammatoires locales et une rupture du gradient de chimiokines à proximité
de la plaque sous-gingivale et, par conséquent, une inhibition de la convergence des neutrophiles au
site de l’infection [177, 223]. En déréglant le réseau des cytokines de l’hôte, la bactérie est capable
d’échapper à son élimination par les cellules immunitaires.
La capacité de P. gingivalis à dégrader les immunoglobulines, plus spécifiquement les IgA, IgG, ainsi
que les IgM, est un autre rôle assumé par les protéases et lui assure, de même qu’aux espèces
bactériennes environnantes, une certaine protection [207, 208, 224-226]. Les immunoglobulines
exercent de nombreux effets protecteurs, notamment en prévenant l’adhérence des bactéries aux
cellules de l’hôte, en causant l’agglutination et l’opsonisation pré-phagocytaire de celles-ci, en
neutralisant les différentes toxines et enzymes qu’elles sécrètent dans l’environnement et en
permettant l’activation de la lyse médiée par le complément [134].
Les protéases de P. gingivalis sont également impliquées dans la destruction tissulaire observée au
cours de la progression des parodontites. P. gingivalis possède une activité collagénase responsable
de la dégradation de certaines protéines structurales telles que le collagène de type II, IV, V et plus
spécifiquement le collagène de type I, le constituant majeur du tissu conjonctif gingival. Les gingipaïnes
de P. gingivalis sont responsables de cette activité de dégradation du collagène puisqu'une inactivation
de ces dernières entraîne une incapacité pour cette bactérie à dégrader le collagène de type I [175,
227]. Les gingipaïnes de P. gingivalis dégradent également des composantes de la membrane basale
et de la matrice extracellulaire, incluant la laminine et la fibronectine [88, 121, 175, 177, 194, 207].

27
Les inhibiteurs plasmatiques des protéases contribuent également aux mécanismes de défense
immunitaire en inhibant les activités protéolytiques dérivées à la fois de l’hôte et des bactéries. Par
ailleurs, il a été démontré que P. gingivalis dégrade de nombreux inhibiteurs de protéases tels que
l’α1-antitrypsine, l’α2-macroglobuline, l’antithrombine III, l’α2-antiplasmine et le C1-inhibiteur, qui ont
pour fonction de réguler l'activité de nombreuses enzymes protéolytiques de la réponse inflammatoire
[121, 194, 207, 209]. De plus, l’activation du plasminogène en plasmine par les protéases de
P. gingivalis permet de moduler la dégradation de protéines de la matrice extracellulaire (fibronectine
et laminine) ainsi que l'activation des formes latentes de métalloprotéinases matricielles (MMPs) [121,
194, 209]. Ces mécanismes, soit la dégradation des inhibiteurs de protéases de même que l'activation
des MMPs résultent en une destruction incontrôlée des tissus parodontaux.
1.3.3.1.1.1.5 Lipopolysaccharide (LPS)
Comme toutes les bactéries Gram négatives, P. gingivalis possède à sa surface du lipopolysaccharide
(LPS). La structure de ce dernier comprend 3 parties ; l’antigène O, le polysaccharide central ainsi que
le lipide A. Le lipide A, la structure biologiquement active du LPS, joue un rôle important lors de la
maladie parodontale, puisque celui-ci stimule indirectement la réponse inflammatoire en déclenchant
la production de TNF-α, d’IL-1β, d’IL-6 ainsi que d’IL-8 par les cellules de l’hôte. Il agit également de
façon directe sur les cellules endothéliales, en inhibant l’expression de l’E-sélectine, cette dernière
participant habituellement aux mécanismes de l’inflammation [7, 43, 228].
Il est connu que le LPS de P. gingivalis est biochimiquement différent de celui des autres bactéries
Gram négatives ; il est atypique par sa faible teneur en heptose et en 2-kéto-3-déoxyoctonate [113].
Par conséquent, ce dernier se caractérise par un faible pouvoir endotoxique lorsque celui-ci est
comparé au LPS d’autres espèces bactériennes (bactéries entériques) [7].

28
1.3.3.1.1.1.6 Vésicules membranaires
Les vésicules membranaires sont des excroissances de la membrane externe qui possèdent des
propriétés antigéniques. Ces dernières sont produites par la bactérie P. gingivalis [229] et relarguées
dans le milieu environnant. Elles contiennent les principaux facteurs de virulence de ce
parodontopathogène. Les vésicules membranaires possèdent une faible dimension (entre 10 et 50
nm), leur permettant une diffusion facilitée à travers les tissus parodontaux en véhiculant, dans les
tissus sous-jacents, les différents facteurs de virulence qu’elles contiennent [229, 230]. Dans un
contexte in vivo, les vésicules fixent une partie des anticorps initialement dirigés contre la bactérie, lui
permettant d’échapper à l’opsonisation et à la bactériolyse [7].
1.3.3.1.2 Aggregatibacter actinomycetemcomitans
A. actinornyceterncornitans est un agent pathogène important impliqué dans les formes sévères et
récurrentes de la parodontite. Effectivement, il s'agit du parodontopathogène majeur dans le cas de la
parodontite agressive localisée, avec une fréquence de détection élevée, soit chez plus de 90% des
patients, ainsi qu’une présence en forte proportion dans les lésions de cette classe de parodontopathie
[7]. La forme agressive de la maladie parodontale est une forme rare (0,1 à 18% des adolescents)
[231], caractérisée par une perte d’attache rapide [232] et la présence d’une faible quantité de plaque
sous-gingivale [29, 233]. Les études ont démontré qu’un facteur génétique pouvait être impliqué dans
le développement de ce type d’affection (composante familiale) [234, 235]. A. actinomycetemcomitans
est également fréquent dans les lésions de certaines parodontites de l'adulte, où il est isolé dans 30 à
50% des cas [236-239]. Cette bactérie parodontopathogène est également impliquée dans les
parodontites réfractaires, sans doute grâce à sa capacité d'invasion des tissus parodontaux [115, 240,
241].
A. actinomycetemcomitans est un coccobacille à Gram négatif, non motile et mesurant 0,7 µm par 1,0
μm [237, 239]. Ce microorganisme est dit capnophile, puisque sa croissance est stimulée dans un
environnement enrichi de 5 à 10% de gaz carbonique (CO2) et sous des conditions d’anaérobiose
[239]. La diversité antigénique présente à la surface de cette bactérie a permis de mettre en évidence

29
6 sérotypes (a à f) [235, 242, 243]. Le stéréotype b, étant le plus virulent, est retrouvé en proportion
élevée chez les patients atteints de parodontite agressive [237, 239].
1.3.3.1.2.1 Facteurs de virulence d’A. actinomycetemcomitans
La bactérie A. actinomycetemcomitans possède de nombreux facteurs de virulence, plus
spécifiquement une leucotoxine, une cytotoxine, le LPS et les vésicules membranaires [244]. La
leucotoxine d’A. actinomycetemcomitans est le facteur de virulence le plus étudié chez cette bactérie
[240, 245, 246]. Il s’agit une protéine de 116 kDa appartenant à la famille des Repeats-in-ToXin (RTX)
[247]. Cette dernière agit par la formation de pores dans la membrane des neutrophiles
polymorphonucléaires (PMNs) [248], des monocytes circulants [249] ainsi que de certaines sous-
populations de lymphocytes, perturbant par le fait même les mécanismes de défense primaires au sein
du sulcus gingival [237, 250, 251]. Son tropisme pour les cellules de la lignée myéloïde repose sur une
interaction avec la B2-intégrine-LFA-1 (lymphocyte function associated molecule) à la surface des
cellules cibles [7, 252]. Le gène codant pour la leucotoxine (ltxA) réside dans un opéron constitué de
quatre gènes, C, A, B et D [237, 253]. Les gènes ltxB et ltxD codent pour des protéines qui sont
impliquées dans le transport de la toxine à la surface de la cellule, tandis que ltxC active la toxine de
façon post-traductionnelle. L'opéron de la toxine semble être présent chez toutes les souches étudiées
[237, 254, 255], bien que plus ou moins exprimé selon les souches (régions promotrices) [237, 256,
257]. De plus, l’expression de la leucotoxine est dépendante de l'environnement (oxygène et fer) [255,
258].
Les fibroblastes sont les cellules les plus abondantes et les plus importantes au sein du tissu conjonctif
gingival. Effectivement, ces derniers, constituant une source majeure de collagène, assure l’intégrité
structurale de la gencive. Plusieurs bactéries buccales, incluant A. actinomycetemcomitans, expriment
une toxine inhibant la prolifération des fibroblastes [237, 259-262]. Considérant son effet sur les
fibroblastes, la cytotoxine de A. actinomycetemcomitans appartient à la famille des « cytolethal-
distensding toxin » (Cdt) [237, 263, 264] et est considérée comme un facteur de virulence [237, 265].
Cette cytotoxine de 50 kDa bloque la synthèse d'ADN chez les fibroblastes, inhibant ainsi leur
prolifération [237, 266].

30
Le LPS d’A. actinomycetemcomitans possède un fort potentiel en ce qui concerne la destruction
tissulaire caractéristique de la parodontite. C’est la raison pour laquelle ce dernier a été largement
étudié et caractérisé au fil du temps [240, 267]. En plus de causer la nécrose de la peau, la résorption
osseuse ainsi que l’agrégation plaquettaire, ce dernier peut activer les macrophages [237].
Effectivement, le LPS d’A. actinomycetemcomitans, biologiquement plus actif que celui des autres
bactéries parodontopathogènes, stimule à faible concentration la production d'IL-1β ainsi que de TNF-
α par les macrophages [268-271]. Ces cytokines sont reconnues pour leur effet pro-inflammatoire et
leur implication dans les processus de résorption osseuse [237, 272]. À une concentration plus élevée,
le LPS stimule la production de cytokines pro-inflammatoires et anti-inflammatoires. Il a été proposé
que le ratio de production de ces deux types de cytokines affecte la balance entre l’évolution ou non
de la maladie parodontale [175].
Certaines bactéries à Gram négatif, dont A. actinomycetemcomitans, libèrent dans le milieu
environnant des vésicules membranaires. Ces structures résultent d’une excroissance de la membrane
externe et se retrouvent en continuité avec cette dernière, ou libérées dans l’environnement [237, 273].
Cette libération de vésicules est directement proportionnelle au degré de virulence de la souche.
Effectivement, les souches exprimant fortement la leucotoxine libèrent davantage de vésicules,
contrairement aux souches l’exprimant peu [274]. Les vésicules membranaires contiennent de
nombreux facteurs de virulence solubles (leucotoxine, cytotoxine, protéine avec activité de résorption
osseuse) de la bactérie, en plus de composantes intrinsèques (LPS [275], adhésines) de cette
dernière. De plus, les vésicules de ce parodontopathogène stimulent la résorption osseuse grâce à
une protéine, localisée dans les vésicules relâchées, activant les ostéoclastes [275].
En plus de la leucotoxine, de la cytotoxine, du LPS et des vésicules membranaires, d’autres facteurs
de virulence ont également été identifiés et partiellement caractérisés incluant des adhésines, des
invasines, un antigène de la capsule polysaccharidique (CPA), de même qu’une collagénase [237].

31
En plus de son implication au sein de la maladie parodontale, la bactérie A. actinomycetemcomitans
serait un agent causal de nombreuses infections extra-buccales, notamment l'endocardite, la
péricardite, la méningite, l'ostéomyélite et les abcès sous-cutanés [7, 237, 244, 276].
1.3.3.1.3 Fusobacterium nucleatum
Fusobacterium nucleatum est une espèce bactérienne prédominante au sein de la cavité buccale, tant
dans un contexte sain qu’un contexte pathologique [277-281]. Il s’agit d’un acteur important dans
certaines maladies infectieuses, dont les maladies parodontales, dans sa forme moins sévère et
réversible de cette affection, soit la gingivite, jusqu’à sa forme la plus sévère, la parodontite (chronique,
agressive). F. nucleatum est l’espèce la plus commune prélevée dans les sites sous-gingivaux, toutes
conditions cliniques confondues [279, 282].
La prévalence de cette bactérie parodontopathogène augmente en fonction de la gravité de la maladie
et, par conséquent avec la progression de l’inflammation ainsi que la profondeur des poches
parodontales [277, 279, 283, 284]. Parmi les cinq sous-espèces, qui ont été identifiées, F. nucleatum
ss nucleatum est fortement associée à la maladie [111, 285].
F. nucleatum est une espèce anaérobie à Gram négatif dont les cellules ont une forme caractéristique
en fuseau aux extrémités pointues (0,4 à 0,7 µm par 3 à 10 µm). De nombreuses études ont suggéré
un rôle important pour cette bactérie dans l’écologie microbienne de la cavité buccale, et plus
particulièrement en ce qui a trait à la formation du biofilm [286]. Effectivement, il a été proposé que F.
nucleatum, en raison des nombreuses adhésines reconnaissant des récepteurs sur plus de 10 espèces
bactériennes de la flore buccale, joue un rôle clé dans la formation du biofilm. Il s’agit d’une des
premières espèces à Gram négatif à s’établir au sein de cette structure, et contribue à créer les
conditions préalables à l'émergence de bactéries anaérobies. Par conséquent, elle contribue à
l’évolution vers plus de complexité de la plaque dentaire, en partant des colonisateurs primaires jusqu’à
l’acquisition des bactéries appartenant au complexe rouge de colonisation soit les bactéries anaérobies
strictes P. gingivalis, T. denticola et T. forsythia [286-288]. C’est entre autre pour cette raison que F.

32
nucleatum ss. nucleatum est incluse dans le complexe de colonisation orange des bactéries à potentiel
parodontopathique [111].
En plus d’être un microorganisme important dans le déclenchement et la progression des maladies
parodontales, F. nucleatum est l'une des espèces buccales les plus communes isolées dans des
infections extra-orales [277], plus précisément dans le sang, dans les articulations, dans les os, dans
l’abdomen, dans les poumons, dans le cerveau, dans les infections de la tête et du cou ainsi que dans
les infections gynécologiques [277]. De nombreuses recherches évaluent l’implication de F. nucleatum
dans diverses affections telles que les résultats défavorables de la grossesse (chorioamniotite,
naissances prématurées, septicémie néonatale, pré-éclampsie), des troubles gastro-intestinaux
(cancer colorectal, maladie inflammatoire de l'intestin, appendicite), ainsi que les maladies
cardiovasculaires, l'arthrite rhumatoïde, les infections des voies respiratoires, le syndrome de Lemierre
et la maladie d'Alzheimer [277, 286].
1.3.3.2 Facteurs étiologiques immunitaires
Bien que la présence bactérienne dans les sites sous-gingivaux soit une composante essentielle à
l'initiation et à la progression de la maladie, elle ne suffit pas à elle seule à expliquer la dégénérescence
tissulaire caractéristique de la parodontite. Un rôle clé dans la progression de celle-ci a été associé
étroitement aux réactions inflammatoires déclenchées par l'hôte, en réponse à une agression
constante de la cavité buccale par les communautés bactériennes [88, 111, 289-292]. En effet,
l'efficacité de cette réponse est observée par la cyclicité des différentes phases de la maladie
parodontale et permet ainsi de confirmer l'étroite relation entre le système immunitaire et la progression
de cette condition pathologique [293].
Afin de conserver un parodonte intact, une réaction inflammatoire quasi constante, dite physiologique,
est mise en place par l'organisme afin de garder l'équilibre (homéostasie) entre le milieu extérieur, soit
les bactéries formant la plaque/biofilm dentaire, et les tissus parodontaux. Si les mécanismes de la
réaction inflammatoire sont opérationnels et efficaces, un parodonte sain est observé et, ce, malgré la

33
présence d'un grand nombre de microorganismes, majoritairement des bactéries, dont certaines
peuvent être pathogènes pour l'individu [294].
En premier lieu, la protection des tissus parodontaux est assurée par les barrières naturelles de
défense de l'hôte, qu'elles soient mécaniques ou immunitaires, spécifiques ou non (épithélium gingival,
salive et fluide créviculaire) [292]. Les barrières épithéliales constituent la première ligne de défense.
Celles-ci s'opposent mécaniquement à la pénétration des bactéries pathogènes dans les tissus
parodontaux. En effet, la capacité des épithéliums à desquamer permet une élimination constante des
bactéries adhérant à la surface des muqueuses. Sur la surface dentaire, c'est le flux salivaire qui
permet une évacuation continuelle des bactéries potentiellement agressives pour le parodonte. En plus
de l'élimination mécanique, les différents composants antibactériens de la salive, tels que les
immunoglobulines à pouvoir agglutinant (IgA sécrétoires), le lysozyme, la lactoferrine, les peroxydases,
etc. permettent, en agissant en synergie, une élimination des microorganismes pathogènes [94, 295,
296]. Les β-défensines 1 produites par les glandes salivaires possèdent également un rôle
antibactérien important. En plus des mécanismes de protection salivaires, un afflux constant de fluide
créviculaire dans les sites sous gingivaux assure également une certaine protection.
Le système immunitaire est capable, dans une certaine mesure, de faire face aux métabolites
potentiellement dommageables des microorganismes qui colonisent le sillon gingival afin de les
empêcher d'infiltrer les tissus parodontaux et prévenir les attaques microbiennes locales [292]. Celui-
ci répond localement par les leucocytes et les macrophages présents dans l'épithélium de jonction et
par la migration des lymphocytes dans la zone tissulaire sous-jacente [294].
Lors d'un changement quantitatif et qualitatif de la flore buccale, indiquant une transition vers des
espèces bactériennes plus virulentes, l'hôte répondra en augmentant ses capacités de défense locales
et régénératrices des tissus parodontaux, via le recrutement et la migration des neutrophiles (cellules
leucocytaires) au site de l’infection. En raison de la chronicité infectieuse, une amplification de la
réponse inflammatoire de l'hôte sera observée d'une part, et les facteurs de virulence exprimés par les
communautés bactériennes adjacentes neutraliseront la réponse de l'hôte d'autre part, conduisant à

34
la destruction des matrices tissulaires avoisinantes. Les bactéries parodontopathogènes peuvent
contrer la réponse immunitaire de l'hôte en freinant la réponse inflammatoire aiguë, en résistant à
l'attaque des neutrophiles et en produisant des enzymes hydrolytiques lui permettant de détruire
différentes molécules impliquées dans la défense de l'organisme, soit les immunoglobulines ainsi que
les composantes du système du complément [88]. La destruction parodontale est alors initiée [297].
Une stimulation des cellules de l'hôte par les parodontopathogènes eux-mêmes ou par certaines de
leurs composantes [292], dont les LPS, puissant antigène de la membrane externe des bactéries Gram
négatives, induit une réponse en diffusant à travers l'épithélium de jonction [294] et en stimulant la
sécrétion d'enzymes hydrolytiques de même que de nombreux médiateurs de l'inflammation [88, 298]
par les monocytes, les neutrophiles, les lymphocytes, les fibroblastes et les cellules épithéliales [88].
Certaines voies de dégradation tissulaire sont par le fait même activées (ostéoclasie) [43]. Le LPS
peut également activer une voie indépendante des médiateurs inflammatoires qui se nomme le
système RANK/RANKL [43].
En ce qui concerne les cytokines, telles que l'IL-1β, l'IL-6, l'IL-8, ainsi que le TNF-α, elles ont
grandement été associées à la progression de la maladie [292]. Présente en quantité importante dans
le fluide créviculaire et les tissus parodontaux, leur sécrétion par les cellules impliquées dans la
réponse inflammatoire est en lien avec le recrutement des cellules immunitaires, plus particulièrement
les leucocytes, au site de l'inflammation et la destruction subséquente des tissus de soutien de la dent
[228].
L'IL-1β est impliquée dans la réponse inflammatoire et la résorption osseuse en favorisant la production
de récepteurs cellulaires, de cytokines, de MMPs [299], et de prostaglandines (PGE2) par les
monocytes et les macrophages [228]. Le TNF-α agit en étroite collaboration avec l'IL-1. Une relation
synergique a été mise en évidence entre ces deux cytokines en ce qui a trait à la résorption osseuse
[228]. En effet, ces deux cytokines possèdent des propriétés biologiques semblables au niveau pro-
inflammatoire, par l'augmentation de la production de médiateurs de l'inflammation, la stimulation de
la synthèse des MMPs, la production d'éicosanoïdes et la résorption osseuse [88, 228]. C'est
principalement le LPS qui induit sa sécrétion par les monocytes et les fibroblastes. L'IL-6 est produite

35
entre autres par les lymphocytes, les monocytes/macrophages et les fibroblastes. Elle stimule la
croissance et la différenciation des lymphocytes B en plasmocytes (cellules sécrétrices d'anticorps) et
stimule la sécrétion des protéines de l'inflammation comme le fibrinogène et la protéine C réactive.
L'IL-6 stimule la formation des ostéoclastes et joue, par conséquent, un rôle lors de la résorption
osseuse [228, 298]. En réponse au LPS bactérien, à l'IL-1 ainsi qu'au TNF-α, les macrophages, les
cellules épithéliales ainsi que les fibroblastes produisent et sécrète une chimiokine, l'IL-8. Comme son
rôle l'indique, cette dernière influence la migration des leucocytes et augmente, par chimiotactisme,
l'activité des neutrophiles et diminue leur adhésion. L'IL-8 contribue à la destruction du collagène en
stimulant la libération de MMPs par les neutrophiles [228]. La PGE2 est un médiateur de l'inflammation
jouant un rôle primordial dans la destruction osseuse et est fortement associée à la parodontite [88,
228]. Synthétisée par les monocytes et les fibroblastes, elle est reconnue également pour ses
propriétés vaso-actives [228].
1.3.3.2.1. Métalloprotéinases matricielles
La sévérité de la parodontite est associée avec une augmentation de divers médiateurs, dans le fluide
créviculaire, soit l'IL-1β, le TNF-α, les PGE2, ainsi que les MMPs [58, 300-303]. En effet, les MMPs
occupent un rôle central lorsqu'il est question du renouvellement tissulaire, tant dans un contexte sain
que dans un contexte pathologique et il a été suggéré que la présence de ces dernières dans le fluide
créviculaire reflète l'état de santé du parodonte.
Les MMPs sont des protéinases, enzymes protéolytiques (endopeptidases) qui ont la particularité de
posséder un ion Zn++ au niveau du site actif [88, 304]. Ces dernières sont capables de dégrader le
collagène, la fibronectine et la laminine, composantes majoritaires de la matrice extracellulaire [305] et
sont classifiées en collagénases, gélatinases, stromélysines et MMP membranaires en fonction de leur
spécificité de substrat [306]. La majorité des types cellulaires constituants le parodonte, plus
spécifiquement les cellules épithéliales, les fibroblastes, les macrophages ainsi que les neutrophiles
ont la capacité de synthétiser des MMPs [307]. La production ainsi que la sécrétion des MMPs est
sous le contrôle de divers médiateurs, ces derniers modulant l'expression des gènes codant pour les
MMPs [308], tels que les cytokines (IL-1, TNF-α), les métabolites de l’acide arachidonique et les

36
facteurs de croissance («Epidermal growth factor » ; EGF, « Platelet-derived growth factor » ; PDGF)
[307, 308]. Synthétisées sous la forme de pro-enzyme (latente) et nécessitant un clivage protéolytique
pour l’activation de ses fonctions (plasmine, enzymes, agents oxydants) [98], ce n'est qu'une fois
activées qu'elles sont impliquées dans la destruction des composantes de la matrice cellulaire ainsi
que la résorption osseuse [304, 307]. Les MMPs sont sous le contrôle d’inhibiteurs, mieux connus sous
le nom de «Tissue Inhibitor of Metallo Proteinases » (TIMPs) [307, 309]. Dans un état non
pathologique, les MMPs ont un rôle de remodelage de la matrice extracellulaire et de remaniement
tissulaire (homéostasie tissu conjonctif gingival). En revanche, dans un contexte pathologique, la
sécrétion des cytokines et des médiateurs de l'inflammation favorisent la production excessive et la
suractivité des MMPs (MMP-2, MMP-3, MMP-8, MMP-9), provoquant par le fait même la destruction
tissulaire et osseuse caractéristique de la maladie parodontale [88, 228, 304, 307, 308]. Au cours du
processus inflammatoire, des produits bactériens, des cytokines, ainsi que divers facteurs de
croissance agissent sur les cellules inflammatoires du parodonte. Celles-ci produiront des quantités
croissantes de MMPs au détriment de la production des inhibiteurs tissulaires des MMPs [310, 311].
Ce déséquilibre initie la destruction tissulaire des différents constituants du parodonte chez les patients
atteints de cette affection.
De nombreuses études ont démontré une forte corrélation entre la sévérité de la maladie parodontale
et le taux de MMPs, plus spécifiquement les MMP-1, MMP-2, MMP-3, MMP-8 et MMP-9, présent dans
la salive, le fluide créviculaire et les tissus parodontaux [312-314]. De ce fait, les MMPs pourraient être
reconnus comme des biomarqueurs dans un contexte d'affection parodontale.
Par définition, un biomarqueur ou un marqueur biologique [315], est une substance qui est mesurée et
évaluée de manière objective comme un indicateur de processus biologiques normaux, de processus
pathogènes ou de réponses pharmacologiques à une intervention thérapeutique. Puisque la salive et
fluide créviculaire sont facilement récoltés et contiennent des marqueurs tant au niveau local et
systémique découlant de la maladie parodontale, ces derniers peuvent être analysés pour une
évaluation de biomarqueurs spécifiques chez le patient atteint de parodontite ou de d'autres affections
systémiques [316, 317]. En raison de la nature non invasive et simple de leur collecte, l'analyse de la

37
salive et du fluide créviculaire peut représenter une alternative particulièrement bénéfique dans la
détermination de l'état de santé parodontal et dans les procédures de rémission suite à l'application
d'une stratégie thérapeutique [318, 319]. De nombreuses études ont montré que la détermination des
niveaux de médiateurs inflammatoires dans les fluides biologiques est un bon indicateur de l'activité
inflammatoire.
La collagénase MMP-8, synthétisée majoritairement par les neutrophiles, participe activement dans la
destruction du parodonte, puisqu'elle intervient dans la dégradation du collagène fibrillaire, incluant les
collagènes de type I, II, III et VI. Le clivage protéolytique du collagène par cette dernière entraîne la
formation de gélatine (collagène dénaturé), qui sera par la suite dégradé par les gélatinases [306].
Dans les maladies de nature inflammatoire, la MMP-8 peut également être produite par les fibroblastes,
les cellules épithéliales (kératinocytes), les odontoblastes, ainsi que les monocytes/macrophages
[320]. Le rôle de la MMP-8 dans la destruction tissulaire observée au cours de la parodontite est très
bien établi. En effet, des niveaux plus élevés de cette protéinase ont été observés dans la salive et le
fluide créviculaire des patients atteints [321].
La MMP-3, également connue sous le nom de stromélysine, favorise l'activation des collagénases et
de la MMP-9 (gélatinase) en plus d'être impliquée dans la dégradation de la fibronectine et des
protéoglycanes de la matrice extracellulaire [88, 98, 320]. La MMP-3 est principalement produite par
les cellules épithéliales (kératinocytes) ainsi que les fibroblastes [322]. Une étude a démontré que la
MMP-3 et la MMP-8 ne sont pas détectables dans les sites cliniquement sains. Par contre, ces MMPs
sont dosées en grande quantités dans les sites démontrant des signes cliniques de parodontite [313,
323].
Les gélatinases, dont la MMP-9, qui ont également été associées à la progression de la parodontite,
possèdent une activité protéolytique dirigée contre la gélatine (collagène interstitiel dégradé par les
collagénases) et le collagène de type IV et V de la membrane basale. L'expression de la MMP-9 est
faible ou absente dans les tissus cliniquement sains et est associée à certains types cellulaires tels

38
que les kératinocytes, les ostéoclastes, les éosinophiles, les neutrophiles ainsi que les
monocytes/macrophages [320].
Contrairement aux molécules pro-inflammatoires, de nombreuses cytokines ont été reconnues pour
leur rôle protecteur, par leur capacité à inhiber le phénomène inflammatoire de même que l'action des
ostéoclastes sur l'os alvéolaire, prévenant ainsi sa résorption. En effet l'IL-1Ra est un antagoniste du
récepteur de l'IL-1. Étant un ligand au même titre que l'IL-1 pour son récepteur, l'IL-1Ra (antagoniste
du récepteur de l'IL-1) empêche la cascade de signalisation cellulaire subséquente à la liaison de se
produire, d'où le potentiel anti-inflammatoire qui lui est attribué. L'IL-4 et l'IL-13, qui partagent de
nombreux effets biologiques [299], sont également reconnues dans la protection de l'os alvéolaire, en
inhibant la formation des ostéoclastes. D'une part, ceux-ci agissent en ciblant directement les
progéniteurs d’ostéoclastes et d'autre part, elles peuvent agir en diminuant l'activité
d'ostéoclastogénèse des ostéoblastes. L'IL-10 inhibe la formation des ostéoclastes en ciblant
directement les progéniteurs des ostéoclastes et indirectement le ratio RANKL/OPG, favorisant ainsi
la conservation de la matrice osseuse [299]. Quant au TGF-β, son rôle protecteur résulte de sa capacité
d’augmenter la synthèse d'inhibiteurs tissulaires de MMPs (TIMPs) et à inhiber la synthèse des MMPs
telles que les collagénases [228].
1.3.4 Traitements des maladies parodontales
Pour tenter d'éradiquer la parodontite, le traitement à administrer est dépendant de l'individu affecté,
mais, dans la majorité des cas, une intervention mécanique seule peut suffire. Il y a deux étapes
essentielles découlant de la thérapie initiale qui permettent d'éliminer le dépôt bactérien sur les
surfaces dentaires et dans les sites sous-gingivaux. La première étape consiste en un détartrage,
permettant d'enlever la plaque et le tartre de la poche parodontale. Cette étape est suivie d'un
surfaçage radiculaire, qui lisse la racine de la dent, enlevant le tartre résiduel et une fine couche de
cément, afin de permettre un rattachement de l'épithélium et ainsi entraîner la guérison [21]. Pour
éliminer les dépôts bactériens sous-gingivaux et ainsi permettre une régénération tissulaire (gencive)
et osseuse, le curetage parodontal est une technique chirurgicale qui suit parfois le surfaçage

39
radiculaire, et qui s'avère utile dans le cas des abcès parodontaux (élimination du tissu gingival infiltré
au niveau de certains sites où le tissu de granulation abonde) [21].
Des agents antimicrobiens complémentaires aux traitements mécaniques peuvent également être
administrés, tant au niveau local (traitement des lésions circonscrites par des antimicrobiens appliqués
de façon locale) que systémique (sites multiples à traiter) [324]. Procurant des résultats équivalents à
ceux obtenus par le détartrage ainsi que le surfaçage radiculaire, ils ne sont cependant pas priorisés
et sont administrés seulement dans les sites parodontaux non-répondants à la première ligne
d'interventions mécaniques (traitements conventionnels) [88]. Par contre, une antibiothérapie
systémique peut s'avérer utile en ce qui concerne les parodontites réfractaires ou à progression rapide.
Des agents chimiothérapeutiques peuvent également être utilisés pour une réduction quantitative et
qualitative de la flore microbienne buccale [88].
De plus, la convergence entre recherche fondamentale et clinique a permis l'acquisition de nouvelles
connaissances et l'élaboration subséquente de nouvelles options préventives et thérapeutiques. Parmi
celles-ci, de nouvelles approches tendent, en plus de contrôler la composante bactérienne, à moduler
la réponse inflammatoire de l'hôte par des inhibiteurs des protéinases, des agents anti-inflammatoires
non-stéroïdiens, des inhibiteurs de l'oxyde nitrique, des cytokines exogènes anti-inflammatoires, des
antagonistes de l'IL-1 et le TNF-α, l'interruption des voies de signalisation NF-κB et MAPK, une action
sur le ratio RANKL/ostéoprotégérine (OPG) de même que la régulation du métabolisme osseux [88].
L'utilisation de composés issus de plantes, dont les polyphénols, pourraient par conséquent
représenter une alternative favorable au traitement des maladies parodontales.

40
1.4 Polyphénols
1.4.1 Généralités
Il existe une grande variété de molécules organiques dans le règne végétal. Parmi celles-ci, on retrouve
les polyphénols, qui se définissent comme des métabolites secondaires des végétaux et qui sont
essentiels à la croissance, à la nutrition de même qu'à la survie de ces derniers. En effet, en plus d'être
responsables des propriétés organoleptiques [325, 326], les polyphénols sont impliqués dans divers
rôles fonctionnels chez la plante, incluant la résistance contre les microorganismes pathogènes ainsi
que les prédateurs herbivores, en plus d'assurer une protection contre les radiations solaires (rayons
ultraviolets ; UVB) [327-331]. La nature des polyphénols, de même que la quantité et la qualité de
ceux-ci dépendent de la plante en question et du stade physiologique de cette dernière [332].
Puisqu’ils sont omniprésents chez les plantes, les polyphénols font partie intégrante de notre
alimentation et ces derniers possèdent de nombreux bienfaits reconnus pour la santé, d’où l’intérêt de
les consommer.
1.4.2 Classification
La structure moléculaire des polyphénols est caractérisée par la présence de un ou plusieurs
groupements phénoliques (aromatiques), auxquels sont attachés un/des groupement(s) hydroxyle(s).
À l'heure actuelle, il existe plus de 8000 polyphénols dont les caractéristiques et la structure ont été
répertoriées [333, 334]. La classification de ces derniers est basée sur de nombreux critères
comprenant leur provenance, leur fonction biologique ainsi que leur squelette carboné [334], plus
spécifiquement le nombre de cycles phénoliques, de même que les éléments structurels liants ceux-ci
[334, 335].
Le Tableau 1 présente la classification des polyphénols telle que proposée dans la littérature [325,
335]. Les premières distinctions sont établies en fonction de l'appartenance des polyphénols à la classe
des non-flavonoïdes, comportant les acides phénoliques, les stilbènes et les lignanes, ainsi qu'à la
classe des flavonoïdes [325, 335]. Les flavonoïdes, qui partagent une structure commune composés
de 2 anneaux aromatiques, sont liés ensemble par 3 atomes de carbone, formant ainsi un hétérocycle

41
oxygéné. Selon le type d'hétérocycle impliqué, ces derniers peuvent être divisés en sous-classes; les
flavanones, les flavones, les isoflavonoïdes, les flavonols, les anthocyanines, de même que les
chalcones.
Tableau 1. Les différentes classes de composés phénoliques (Tableau adapté de [336]).
1.4.3 Thé
Le thé, provenant de la plante Camellia sinensis, est reconnu pour contenir des polyphénols en grande
quantité. Cette boisson d'origine chinoise prend racine dans les plus vieilles traditions de l'Orient, et
ce, depuis l'Antiquité. Le thé représente maintenant, après l'eau, la boisson la plus consommée au
Classes Sous-classes Exemple de composés
Flavonoïdes
Flavanones Naringénine
Flavones Lutéoline
Isoflavonoïdes Glabridine
Flavonols Quercétine, Myricétine
Flavanols
Mononères Constitué de catéchine
Polymères (Tannins)
Condensés Constitué de sous-unités catéchines ou épicathéchines
Hydrolysables Constitué d'acide gallique ou éllagique
Anthocyanines Cyanidine
Chalcones Butéine
Non-Flavonoïdes
Acides phénoliques Acide gallique, Eugénol
Stilbènes Trans resvératrol
Lignanes Magnolol, Honokiol
Coumarins Collinine, Lacinartine

42
monde, s'adaptant aux coutumes des différentes populations [337, 338]. Autrefois et encore
aujourd'hui, le thé est avant tout consommé pour les vertus médicinales qu'il possède.
Tous les types de thés, qu’ils s’agissent du thé blanc, du thé vert, du thé oolong ou du thé noir,
proviennent d’une seule espèce de plante, Camellia sinensis. La principale différence entre les
différents thés réside dans le mode de préparation de ceux-ci, qui aura une influence sur le contenu
qualitatif et quantitatif en polyphénols. Selon le procédé de fabrication, les thés sont classés en 3
types : non-fermentés (vert), partiellement fermentés (oolong) ou fermentés (noir) [339-342].
Contrairement au thé noir, qui est fermenté, le thé vert est produit à travers un processus dans lequel
il ne subit aucune fermentation. La fermentation résulte dans la formation de composés phénoliques
condensés de haut poids moléculaire [339]. Les différentes méthodes de fabrication de chacun des
thés ainsi que les polyphénols qu’ils contiennent sont présentés dans la Figure 4.
Figure 4. Schématisation du processus de fabrication du thé et composition en polyphénols des divers
thés (Figure adaptée de [339].

43
1.4.3.3 Thé vert
Le thé vert est considéré comme un thé non-fermenté puisqu’une fois récoltées, les feuilles matures
subissent un traitement à la vapeur afin de prévenir la fermentation par l’enzyme polyphénol oxydase
avant le séchage des feuilles. Cette enzyme est responsable de la conversion des flavanols en
composés polyphénoliques foncés qui colorent le thé noir [339, 340, 343, 344]. Chaque année, environ
2.5 millions de tonnes de thé sont produites [345]. Il y a approximativement 76 à 78% de cette
production ainsi que de la consommation mondiale de thé qui est consacrée au thé noir, tandis que le
thé vert représente 20 à 22%. Ce dernier est majoritairement consommé en Chine, au Japon et en
Inde [338, 339, 346].
1.4.3.4 Composition
Les pratiques horticoles, le climat, les saisons, de même que le degré de maturation des feuilles de
thé sont des facteurs qui influencent la composition du thé [347]. La composition chimique du thé vert
est complexe et unique ; il contient des protéines (15 à 20% du poids sec) dont les enzymes qui
occupent une fraction importante ; des acides aminés (1-4% en poids sec) dont la théanine, dérivée
de la glutamine et propre au thé, l'acide glutamique, le tryptophane, la glycine, la sérine, l'acide
aspartique, la tyrosine, la valine, la leucine, la thréonine, l'arginine, la lysine ; des hydrates de carbone
(7.5% en poids sec) tels que la cellulose, les pectines, le glucose, le fructose, le saccharose; des lipides
tels que les acides linoléiques ; des stérols comme le stigmastérol ; des vitamines (B, C, E) ; les bases
xanthiques comme la caféine (3%) et la théophylline ; des pigments (caroténoïdes et chlorophylle) ;
des composés volatils tels que des aldéhydes, des alcools, des esters, des lactones, des
hydrocarbures; et finalement, des minéraux ainsi que divers oligo-éléments (5% de poids sec) tels que
Ca, Mg, Cr, Mn, Fe, Cu, Zn, Mo, Se, Na, P, Co, Sr, Ni, K, F et A [340, 348-350].
Les composantes les plus importantes du thé vert sont les polyphénols appartenant à la classe des
flavonoïdes, et plus spécifiquement, les catéchines [348]. Les catéchines sont des composés incolores,
solubles dans l’eau qui confèrent l’astringence au thé. Les quatre principales catéchines (flavan-3-ols)
qui sont retrouvées dans le thé sont l’épigallocatéchine-3-gallate (EGCG), constituant 59% des
catéchines totales, l’épigallocatéchine (EGC ; 19%), l’épicatéchine-3-gallate (ECG ; 13,6%), ainsi que

44
l’épicatéchine, correspondant à environ 6,4% du contenu total [340, 341, 350-352]. L’EGCG, présente
dans des proportions plus élevées, est considéré comme la molécule du thé vert possédant le plus
grand pouvoir antioxydant. Par conséquent, il a été recensé dans la littérature que cette dernière,
présentée dans la figure 5, serait la molécule biologiquement active du thé vert [346, 353, 354].
Figure 5. Structure moléculaire de l’épigallocatéchine-3-gallate.
1.4.4 Effets bénéfiques du thé vert
Les feuilles de thé contiennent 3 composantes principales pouvant agir dans la promotion de la santé
humaine : les bases xanthiques telles que la caféine et la théophylline, les huiles essentielles ainsi que
les polyphénols. Malgré les effets intéressants observés chez ces deux premières composantes [340,
355], l’intérêt de la communauté scientifique quant aux effets bénéfiques du thé se dirige davantage
vers les polyphénols, des molécules antioxydantes auxquelles ont été attribuées de nombreuses
propriétés biologiques. Plus précisément, ces effets favorables ont été associés à leurs propriétés anti-
inflammatoires, anti-mutagéniques, antioxydantes et antimicrobiennes [339, 340, 356].
Suite à la réalisation d’études épidémiologiques et cliniques, de nombreuses publications scientifiques
démontrent et reconnaissent maintenant les bienfaits du thé vert et de ses composantes sur la santé

45
générale. Effectivement, la consommation de thé vert pourrait être liée à une diminution du risque et
de la gravité de nombreuses conditions pathologiques et maladies systémiques [357], incluant les
cancers (buccal, peau, poumon, œsophage, estomac, foie, pancréas, intestin grêle, colon, vessie,
prostate, ovaires et sein) [339-341, 356, 358-361]. Le potentiel antioxydant du thé vert peut également
contribuer à la prévention des maladies cardiovasculaires (athérosclérose, AVC [354, 362]) en
diminuant la tension artérielle, en permettant un meilleur fonctionnement des cellules endothéliales
ainsi qu’en interférant avec la modification oxydative du cholestérol en lipoprotéine de basse densité
(LDL) [339, 340, 356, 363-366]. De plus, les polyphénols du thé vert pourraient constituer de nouvelles
molécules permettant de prévenir et de traiter certaines maladies de nature inflammatoire telle que
l’arthrite rhumatoïde [367-371]. Les polyphénols du thé vert peuvent également permettre un contrôle
de l’obésité [340, 372-375], du diabète [339, 340, 369, 376, 377], de certaines maladies
neurodégénératives telles que la maladie d’Alzheimer et le Parkinson [86, 369, 378-380], en plus
d’augmenter la densité minérale osseuse [381]. Malgré de nombreuses expérimentations in vitro et in
vivo réalisées chez des modèles expérimentaux, les effets bénéfiques du thé vert demeurent
controversés. Dans ce contexte, des études subséquentes de nature clinique devront être réalisées
afin de démontrer hors de tout doute le potentiel du thé en tant qu’agent préventif et thérapeutique de
certaines affections.
Plus récemment, différentes études ont démontré que le thé vert et ses principaux polyphénols peuvent
également exercer un effet positif sur la prévention et le traitement de diverses affections buccales,
plus particulièrement en ce qui a trait à l’halitose buccale, à la carie dentaire ainsi qu’à la maladie
parodontale.
1.4.4.1 Halitose buccale
L'halitose ou, de façon plus commune, la mauvaise haleine, est un terme général utilisé dans le
domaine dentaire afin de décrire une mauvaise odeur émanant de la cavité buccale. Affectant une
grande proportion de la population, et possédant de nombreuses implications sociales, l’halitose
demeure une condition à considérer dans la promotion de la santé et la prévention de certaines
maladies. En effet, bien que les facteurs responsables de l’halitose soient d’origine buccale dans 80%

46
à 90% des cas [382, 383], l'halitose peut être la manifestation de différents désordres systémiques
importantes. Cette affection est causée par la production, dans la cavité buccale, de composés sulfurés
volatils (CSVs), principalement le sulfure d'hydrogène (H2S), le méthyl mercaptan (CH3SH) et le
diméthylsulfate [384-388].
Les composés sulfurés volatils sont majoritairement produits par les espèces bactériennes Gram
négatives anaérobies strictes présentes sur la surface dorsale de la langue, dont F. nucleatum, P.
intermedia, P. gingivalis, T. denticola et Solobacterium moorei [389, 390]. Ces bactéries dégradent les
substrats protéiques provenant de la salive, du fluide créviculaire, des cellules épithéliales buccales
desquamées, des cellules sanguines, des débris alimentaires, avant de transformer les acides aminés
contenant du soufre (cystéine et méthionine) en composés sulfurés volatils.
Les traitements actuels de l'halitose visent à contrôler le facteur microbien, et plus particulièrement à
diminuer le nombre de microorganismes potentiellement nocifs au sein de la cavité buccale. Par
conséquent, l'élimination mécanique par le brossage quotidien des dents et de la langue permet de
diminuer la sévérité de l'halitose [391-394]. Conjointement, des produits de soins bucco-dentaires
supplémentés avec des agents antimicrobiens (chlorhexidine, chlorure de cétylpyridinium, triclosan,
huiles essentielles) ou des substances permettant la neutralisation des CSVs en composés non-
odorants (dioxyde de chlore, sels de zinc) peuvent être utilisés [387, 391, 395-397]. Des recherches
ont récemment démontré que les polyphénols du thé vert pourraient exercer de tels effets [397].
De nombreuses études ont démontré que le thé vert et, par conséquent, l'EGCG, pourrait représenter
une alternative thérapeutique naturelle aux agents antimicrobiens chimiques couramment utilisés pour
lutter contre la mauvaise haleine. Considérant le rôle des bactéries anaérobies parodontopathogènes
dans la production des composés sulfurés volatils, les recherches tendent à démontrer que les
polyphénols du thé vert peuvent réduire la mauvaise haleine en inhibant la croissance de ces dernières.
De plus, les polyphénols du thé vert, dont l'EGCG, inhibent de nombreux facteurs de virulence des
bactéries parodontopathogènes, également impliqués dans la production des CSVs [338, 397-399].

47
Effectivement, à titre d’exemple, l'enzyme L-cystéine désulfhydrase (CD), présente chez certaines
bactéries, agit sur la L-cystéine afin de produire du pyruvate, de l'ammoniac, ainsi que du H2S, tandis
que la L-méthionine-α-désamino-γ-mercaptométhane-lyase (METase), mène à la production de l'α-
cétobutyrate, de l'ammoniac, ainsi que du méthyl mercaptan à partir de la L- méthionine [387, 400].
La bactérie P. gingivalis est connue pour produire une grande quantité de CH3SH à partir de la
METase, codée par le gène mgl [387, 400]. L'EGCG possède la capacité d’inhiber, à des
concentrations sous-inhibitrices, l'expression du gène mgl, et par conséquent, de diminuer la
production du méthyl mercaptan (CH3SH) par P. gingivalis [387].
Il a récemment été démontré qu’un extrait de thé vert ainsi que l'EGCG inhibent la croissance de S.
moorei, avec des valeurs de CMI respectives de 500 et 250 µg/ml [401]. Des analyses en microscopie
électronique à transmission, de même qu’un test de perméabilisation ont révélé que la membrane de
la cellule bactérienne était la cible principale des polyphénols du thé vert. En ce qui concerne les effets
de ces polyphénols sur les propriétés de colonisation de S. moorei, il a été constaté que la formation
du biofilm sur des surfaces prétraitées avec de l’EGCG est affectée de façon significative, et que ces
derniers peuvent entraîner une désorption du biofilm préalablement formé par cette bactérie. En outre,
il a également été démontré que l'extrait de thé vert et l'EGCG réduisent l'adhérence de S. moorei aux
cellules épithéliales buccales. L'activité β-galactosidase de S. moorei, laquelle joue un rôle clé dans la
production des CSVs, est de plus atténuée de façon dose-dépendante par les polyphénols du thé vert.
Enfin, l’expression du gène de cette enzyme est inhibée par une concentration d’EGCG correspondant
à ½ CMI.
L’utilisation du thé vert sous la forme d’un rince-bouche a montré un effet sur la quantité des CSVs
présents [402]. Également, les effets des catéchines du thé vert sur le méthyl mercaptan, une source
majeure dans l'halitose, ont été étudiés. Plus spécifiquement, une gomme à mâcher contenant des
catéchines du thé diminue significativement la production de méthyl mercaptan dans la salive et par
conséquent, cette dernière s’est avéré utile dans la réduction de la mauvaise haleine. L'activité
déodorante diminue selon l'ordre suivante : EGCG > EGC > ECG > EC. L'EGCG serait, à cause de sa

48
structure chimique, un agent permettant la neutralisation efficace des mauvaises odeurs via une
réaction chimique avec le groupement thiol des CSVs [338, 387, 397-399].
Des essais in vitro ont également démontré qu'à différents pH, un extrait de thé vert a la capacité de
diminuer significativement les quantités respectives de H2S et de CH3SH au niveau de la cavité
buccale. Par conséquent, certains produits permettant de réduire la mauvaise haleine, tels que les
gommes à mâcher et les aérosols buccaux contiennent des polyphénols du thé [403].
1.4.4.2 Carie dentaire
La carie dentaire est la maladie la plus répandue des tissus calcifiés des dents dans la population
[404]. Il s'agit d'une maladie infectieuse d'origine multifactorielle, qui résulte d’un processus dynamique
nécessitant un hôte sensible, une microflore cariogène ainsi qu'un substrat approprié qui doit être
présent pour une durée de temps suffisante au sein de la cavité buccale [405-407]. L'étiologie de la
carie dentaire a été associée à une dissolution acide des composants dentaires minéraux, l'acide étant
produit par les bactéries buccales utilisant des glucides alimentaires en tant que substrat [408, 409].
Les principaux agents étiologiques incriminés dans cette pathologie sont principalement les
streptocoques α-hémolytiques Streptococcus mutans et Streptococcus sobrinus. Les lactobacilles
ainsi que les actinomycètes peuvent également être impliqués, mais à moindre degrés [354, 410-412].
Bien que certains produits alimentaires contribuent au développement et à la progression de la carie
dentaire, d’autres aliments, dont le thé vert, sont considérés comme des agents anti-cariogènes. [412,
413]. Par conséquent, le thé a été étudié en détail pour son rôle protecteur dans les caries dentaires
et sa capacité à influencer le processus de formation et l’évolution de ces dernières [404]. Les feuilles
de thé sont riches en fluorure, ce dernier étant reconnu pour améliorer la santé dentaire et prévenir la
formation des caries [414-417]. Cependant, les possibles avantages attribués au thé sur la santé
dentaire ne sont pas limités au fluorure, mais impliquent également d'autres composantes du thé, dont
les catéchines [338, 385, 414, 418-422]. Effectivement, les thés et plus spécifiquement le thé vert,
contiennent une quantité importante de catéchines. Celles-ci représentent un sous-groupe de

49
flavonoïdes et sont soupçonnées d’être responsables des bienfaits pour la santé généralement
attribués au thé [340, 404, 414, 420].
Il existe de nombreuses évidences permettant d’affirmer que les composants bioactifs présents dans
le thé vert, et plus particulièrement les polyphénols, sont en mesure d'influencer le processus de
formation de la carie dentaire à plusieurs niveaux. En effet, ces derniers peuvent inhiber la croissance
de S. mutans et S. sobrinus, de même que la production d’acide par ces derniers. De plus, les
polyphénols du thé peuvent interférer avec le processus d'adhésion des bactéries cariogènes à l'émail
des dents et agir comme inhibiteurs de la glucosyltransférase, limitant ainsi la biosynthèse du glucane,
et de l'amylase salivaire et bactérienne (métabolisme et activité enzymatique) [354, 414, 423-431]. Des
études utilisant des modèles animaux ont démontré que les effets in vitro observés sont reproductibles
dans un contexte in vivo et que le thé vert pourrait être utilisé dans la prévention de la carie dentaire
[432, 433]
1.4.4.2.1. Action antibactérienne
Dans un premier temps, une étude utilisant un rince-bouche à base d'une infusion thé vert a pu établir
un lien entre la diminution du nombre de bactéries cariogènes dans l’environnement buccal
(Streptococcus spp. et Lactobacillus spp.) et l’utilisation de ce rince-bouche [411]. Plus spécifiquement,
de nombreuses équipes de chercheurs ont démontré que le thé vert, et plus particulièrement l'EGC et
l'EGCG, possèdent une forte activité antibactérienne et bactéricide contre les bactéries impliquées
dans l'étiologie de la carie dentaire, S. mutans ainsi que S. sobrinus [404, 405, 426, 432, 434-436].
Il a été rapporté que l’activité antibactérienne de l'EGCG serait liée à une action sur la membrane
cytoplasmique des bactéries [437]. D'autres études sont allées plus loin en affirmant que les altérations
de la membrane cytoplasmique résultent de la génération de peroxyde d'hydrogène par l'EGCG à
l'intérieur de la bicouche phospholipidique [438].

50
1.4.4.2.2. Inhibition de la production acide par les bactéries cariogènes
Certains microorganismes, dont la bactérie S. mutans, produisent des acides, majoritairement l’acide
lactique comme sous-produits de la fermentation des sucres présents dans l'alimentation, et plus
spécifiquement le saccharose. Cette production d'acide entraîne la déminéralisation de l'émail menant
aux lésions carieuses. Il a été démontré que le rinçage de la cavité buccale avec de l’EGCG, permet
de réduire la quantité d’acide produite par les bactéries [424].
1.4.4.2.3. Inhibition du gène codant pour la glucosyl-transférase
Tel que mentionné précédemment, la formation de la plaque dentaire joue un rôle important dans le
développement des caries. Les bactéries cariogènes, et plus particulièrement S. mutans, utilise le
saccharose alimentaire afin de produire des polymères de glucanes (solubles ou insolubles ;
monomères de glucose) à l’aide de l’enzyme glucosyltransférase (GTase) [354, 405, 414]. Ces
polysaccharides insolubles (polymères insolubles de glucanes) contribuent à l'adhérence de bactéries
aux surfaces dentaires [410], à la formation du biofilm ainsi qu’à la production d’énergie lorsque la
disponibilité de glucides fermentescibles exogènes dans la cavité buccale est réduite [414]. L’EGCG à
des concentrations sous-inhibitrices cause une inhibition chez la bactérie S. mutans de l’expression du
gène gtf, le gène codant pour la glucosyltransférase, inhibant ainsi l’adhérence de la bactérie à la
surface dentaire et la formation subséquente du biofilm [412, 414, 426-428, 431].
1.4.4.2.4. Inhibition de l'amylase salivaire et bactérienne
Les substrats utilisés par les bactéries cariogènes peuvent être disponibles directement, via le sucre
ingéré à travers l'alimentation, ou être dérivé de l'amidon alimentaire suite à l’action d'amylases
bactériennes et/ou salivaires [422]. Il a été rapporté dans la littérature que l’EGCG inhibe l’activité de
l’amylase salivaire et bactérienne impliqués dans le métabolisme des glucides [405]. Par conséquent,
l’EGCG réduit la libération du glucose et abaisse ainsi le potentiel cariogène de la nourriture contenant
de l’amidon [341, 439, 440]. L’ajout de polyphénols du thé dans des friandises et dans des gommes à
mâcher a démontré un effet inhibiteur sur la formation de plaque dentaire ainsi que sur le
développement de la carie dentaire [385, 441].

51
1.4.4.3 Maladies parodontales
La maladie parodontale est une maladie inflammatoire d’origine infectieuse affectant, à long terme, les
tissus entourant la dent et participant au soutien de cette dernière. Étant une maladie multifactorielle,
les facteurs microbiologiques, immuno-inflammatoires et environnementaux participent à l’initiation et
à la progression de cette condition pathologique. En effet, la présence de certaines bactéries
anaérobies à Gram négatif au sein du biofilm dentaire sous gingival est considérée comme le facteur
étiologique primaire de cette affection. Subséquemment, une réponse immunitaire exagérée de l’hôte
vis-à-vis les bactéries pathogènes mène à la production de médiateurs inflammatoires (cytokines)
activant diverses voies de dégradation tissulaire et osseuse (MMPs) par les cellules de l’hôte. Afin de
contrôler efficacement l’initiation et la progression des maladies parodontales, les deux composantes
étiologiques doivent être contrôlées [442].
Une vaste étude épidémiologique a révélé une association inverse entre la consommation de thé vert
et les divers paramètres cliniques indiquant une affection parodontale (profondeur des poches
parodontales, niveau d'attachement clinique, saignements lors du sondage) [443]. Les recherches
actuelles visent à identifier les mécanismes d’action par lesquels les polyphénols présents dans le thé
exercent leurs effets bénéfiques contre la maladie parodontale.
D’une part, il est essentiel de limiter la colonisation de la cavité buccale par les bactéries
parodontopathogènes. Effectivement, ces dernières sont fortement impliquées directement ou non,
dans la destruction des tissus parodontaux et leur élimination permettrait d’interférer avec l’initiation
des maladies parodontales et éventuellement, conduire à une amélioration des paramètres cliniques
dans les sites atteints [88, 385, 444]. L'adhésion des bactéries aux cellules de l’hôte constitue la
première étape de l'infection bactérienne. Cette adhésion permet la colonisation des tissus
parodontaux ainsi que l’exposition, dans le milieu environnant, de différents facteurs de virulence ayant
un effet direct sur les cellules de l’hôte [445]. Il a été démontré que le thé vert, et plus spécifiquement
l’EGCG, inhibe la croissance et l’adhérence de P. gingivalis aux cellules épithéliales buccales [354,

52
385, 445, 446]. Une autre étude a également démontré le potentiel bactéricide des catéchines
présentes dans le thé vert. En effet, grâce à l’utilisation d’un système de délivrance à libération lente
saturé en thé (bandes d’hydroxypropylcellulose) appliqué directement dans les poches parodontales,
il est possible, en combinaison avec le traitement mécanique, de réduire significativement les
proportions de Prevotella spp. (intermedia et nigrescens) et P. gingivalis, en plus d’entraîner une
diminution de la profondeur des poches parodontales [447]. Il a récemment été rapporté que certains
facteurs de virulence associés à ces agents étiologiques bactériens, notamment les protéases, sont
neutralisés par l’EGCG [340, 354, 446, 448, 449]. Une autre étude a également démontré les effets
anti-protéases du thé vert, en plus de propriétés antibactérienne, anti-adhérence et anti-inflammatoire
de ce dernier sur la bactérie P. gingivalis [450].
D’autre part, suite à l’infection bactérienne, la réponse immunitaire est perpétuée, ce qui cause la
destruction des tissus parodontaux. Effectivement, les facteurs de virulence des bactéries stimulent
les cellules inflammatoires à libérer des médiateurs de l’inflammation et activent les voies de
dégradation tissulaire qui perturbent l'équilibre avec leurs inhibiteurs, entraînant la destruction [88].
L’inhibition et/ou la modulation de la réponse immunitaire permettrait de réduire l’incidence des
maladies parodontales. En plus d’agir sur la composante bactérienne, il a été rapporté dans la
littérature que le thé vert, et plus particulièrement l’EGCG, ont le potentiel de prévenir l’inflammation
gingivale ainsi que la résorption tissulaire et osseuse caractéristique de la parodontite.
L'épithélium gingival est un tissu qui agit comme une interface entre l'environnement extérieur et le
tissu conjonctif sous-jacent. Il joue un rôle clé dans la résistance contre les infections parodontales en
agissant comme une barrière physique contre les parodontopathogènes. De plus, les cellules
épithéliales buccales élaborent une réponse immunitaire innée par la sécrétion de peptides
antimicrobiens, comprenant les β-défensines humaines (hBDs). Ces peptides antimicrobiens
cationiques interagissent avec la membrane de la cellule bactérienne, entraînant à la formation de
pores et ultimement, la lyse des bactéries parodontopathogènes [451-455]. Il a été démontré que le
thé vert ainsi que l’EGCG augmentent l’expression des gènes codant pour les β-défensines et, par

53
conséquent, la production de ces dernières par les cellules épithéliales buccales [451]. De plus, le thé
vert et l’EGCG préviendraient la dégradation de celles-ci par les protéases de P. gingivalis [451].
Le « Nuclear Factor kappa B » (NF-κB) est un facteur de transcription [456] qui coordonne l'expression
d'une grande variété de gènes responsables de l’inflammation [457]. Ce dernier est également impliqué
dans la maladie parodontale, puisque celui-ci régule la réaction chronique inflammatoire et la
production de cytokines pro-inflammatoires qui engendrent la destruction tissulaire et osseuse [458,
459]. Les ostéoclastes activés détruisent l'os alvéolaire par la voie de destruction osseuse impliquant
le « receptor activator of NF-κB ligand » (RANKL), RANK, ainsi que l’ostéoprotégérine (OPG). Il en
résulte en un cercle vicieux, puisque l'accumulation du biofilm dans les poches parodontales stimule
la réponse immunitaire de l'hôte et conséquemment, la destruction des tissus engendré par cette voie
de signalisation [405, 460]. L’EGCG bloque directement ou non la destruction tissulaire induite par NF-
kB en prévenant la production de certains médiateurs de l’inflammation, dont l’IL-1β, le TNF-α ainsi
que les protéases destructrices, les MMPs [405, 461, 462]. Plus spécifiquement, des études in vitro
et in vivo ont démontré que les catéchines du thé vert diminuent la résorption osseuse en inhibant la
production de cytokines pro-inflammatoires (IL-1β) au niveau de la muqueuse gingivale dont
l’inflammation est induite par le LPS (ou directement l’ostéoclastogénèse) [463, 464]. L’expression du
TNF-α dans les lésions parodontales induites chez le rat a été diminuée lors de l’ajout de catéchines
du thé vert à l’intérieur d’un dentifrice [442, 463, 465]. Enfin, certaines études ont évalué la capacité
de l’EGCG de prévenir la destruction tissulaire et osseuse médiée par les MMPs. Par ailleurs, une de
ces études a démontré que l’EGCG inhibe l’expression de l’ARNm de la MMP-9, en plus d’inhiber la
formation des ostéoclastes [443, 466-468].
La cathélicidine LL-37 est un peptide antimicrobien exprimé par divers types cellulaires, tels que les
monocytes, les neutrophiles ainsi que les cellules épithéliales [469]. Elle est reconnue pour posséder
des propriétés anti-inflammatoires en inhibant la sécrétion de diverses cytokines contribuant à
l’inflammation associée à la parodontite chronique [470-472]. Il a récemment été démontré, en utilisant
un modèle de co-culture 3D comprenant des cellules épithéliales gingivales et des fibroblastes, que le
LL-37, en combinaison avec l’EGCG, agissent en synergie afin de réduire la sécrétion de GRO-α (CXC-

54
chemokine ligand 1), de G-CSF (granulocyte colony-stimulating factor), d’IL-6, d’IL-8, et de l’IP-10
(interferon-γ inducible protein 10), en plus d’exercer un effet additif sur la sécrétion de MCP-1
(monocyte chemoattractant protein-1)[473]. Par conséquent, cette combinaison LL-37 et EGCG
démontre des résultats prometteurs en vue d’une thérapie d’appoint pour le traitement des maladies
parodontales de nature inflammatoire.

55
2. PROBLÉMATIQUE
Outre la carie dentaire, les maladies parodontales représentent le principal problème de santé
buccodentaire au sein de la population et ces dernières engendrent un impact socio-économique
important. Depuis plusieurs années, la convergence entre la recherche fondamentale et clinique a
permis d’en apprendre davantage sur les différentes composantes étiologiques impliquées dans cette
pathologie. Or, beaucoup reste à découvrir. Étant une maladie multifactorielle, les facteurs
microbiologiques, immuno-inflammatoires et environnementaux contribuent à l’initiation et à la
progression de cette condition pathologique. La présence de certaines bactéries anaérobies à Gram
négatif au sein du biofilm dentaire est considérée comme le facteur étiologique primaire de cette
affection. Subséquemment, une réponse immunitaire exagérée de l’hôte vis-à-vis les bactéries
pathogènes mène à la production de médiateurs inflammatoires activant diverses voies de dégradation
tissulaire et osseuse par les cellules de l’hôte.
L’utilisation de polyphénols de plante, dont les catéchines retrouvées dans le thé vert, pourrait modifier
l’interaction bactérienne avec son environnement (mécanismes de pathogénicité), agir comme
modulateur de la réponse immuno-inflammatoire destructrice de l’hôte, et ainsi éviter les
conséquences cliniques qu’entraîne la parodontite, tant au niveau local que systémique. En effet, par
des mécanismes directs et indirects, les parodontopathogènes impliqués dans la maladie parodontale
peuvent avoir une incidence sur le développement de diverses affections systémiques, notamment les
maladies cardiovasculaires, d’où l’intérêt de comprendre les divers mécanismes par lesquels cette
pathologie s’impose à l’hôte.
Puisque les agents microbiens démontrent de plus en plus de résistances aux traitements actuels
impliquant des antibiotiques, les polyphénols du thé vert pourraient représenter de nouvelles molécules
à portée préventive et thérapeutique en vue de leur utilisation pour le contrôle des maladies
parodontales de nature inflammatoire.

56
2.1 Hypothèse de recherche
Les polyphénols du thé vert possèdent des propriétés antibactériennes vis-à-vis les principales
bactéries parodontopathogènes. De plus, ces derniers agissent en synergie avec des antibiotiques
couramment utilisés en thérapie complémentaire.
L’utilisation d’un modèle de co-culture macrophages/fibroblastes permet de mieux évaluer les
propriétés anti-destruction tissulaire des polyphénols du thé vert, par l’inhibition de la sécrétion de
métalloprotéinases matricielles (MMPs) par les cellules de l’hôte.
2.2 Objectifs spécifiques
Afin de vérifier nos hypothèses, trois objectifs ont été définis :
1. Évaluer les propriétés antibactériennes d’un extrait de thé vert et de l’EGCG sur les principales
bactéries parodontopathogènes;
2. Évaluer la capacité d’un extrait de thé vert et de l’EGCG à agir en synergie avec des
antibiotiques utilisés couramment en thérapie complémentaire;
3. Évaluer la capacité d’un extrait de thé vert et de l’EGCG à inhiber la sécrétion de
métalloprotéinases matricielles (MMPs) dans un modèle de co-culture
macrophages/fibroblastes.

57
3. MATÉRIEL ET MÉTHODES
3.1 Molécules à l’étude
L’extrait de thé vert (GT#9; BCY-CT002) a été obtenu de Gosun Biotechnologies Co., Ltd. (Hangzhou,
Chine), et la solution stock issue de cet extrait a été préparée à une concentration 10 mg/ml dans de
l’eau distillée. La préparation a été stérilisée par filtration (filtre 0.2 µm) et conservée pour une durée
maximale d’une semaine à une température de 4 °C à l’abri de la lumière. La composition de l'extrait
selon les analyses de la compagnie est présentée dans le Tableau 2.
Tableau 2. Composition de l’extrait de thé vert GT#9
Composé Pourcentage
Polyphénols totaux 98,42%
Catéchines totales 82,60%
Épigallocatéchine gallate (EGCG) 47,92%
Épigallocatéchine (EGC) 7,56%
DL-Catéchines 2,16%
Épicatéchine (EC) 6,19%
Gallocatéchine gallate (GCG) 4,54%
Épicatéchine gallate (ECG) 14,23%
Caféine Maximum 1,0%
L’épigallocatéchine gallate (EGCG), le principal flavonoïde composant le thé vert, a été obtenu de
Sigma-Aldrich Corp. (St-Louis, MO, États-Unis). Tout comme l'extrait de thé vert, une solution stock a
été préparée à 10 mg/ml dans de l’eau distillée, pour ensuite être stérilisée par filtration (filtre 0.2 µm).
Cette solution a été entreposée à l’abri de la lumière à une température de -20 °C, afin de permettre
sa conservation à plus long terme.

58
Puisque certains agents antibiotiques tels que la tétracycline et le métronidazole peuvent parfois être
administrés en complémentarité aux traitements mécaniques dans la thérapie parodontale, ceux-ci ont
été testés en combinaison avec l'extrait de thé vert et l'EGCG. À cet effet, des solutions de ces deux
antibiotiques ont été préparées à une concentration de 1 mg/ml dans de l’eau distillée, stérilisées par
filtration (filtre 0.2 µm) et entreposées à -20 °C, à l’abri de la lumière.
3.2 Espèces bactériennes à l’étude et conditions de culture
Plusieurs espèces bactériennes ont été utilisées au cours de cette recherche. Chacune d’entre elles
est associée à l’étiologie des maladies parodontales et contribue, de près ou de loin, au développement
de celles-ci. Les bactéries étudiées ont été P. gingivalis American Type Culture Collection (ATCC)
33277, ATCC 49417, W83 et HW24D1, F. nucleatum ATCC 25586 ainsi qu’A. actinomycetemcomitans
ATCC 29522. Ces dernières ont toutes été cultivées en condition anaérobie (N2/H2/CO2 75:10:15) dans
des milieux qui diffèrent d’une espèce à l’autre, à une température de 37 °C pour une période de 24
heures.
P. gingivalis et F. nucleatum ont été cultivées dans le milieu « Todd Hewith broth » (Becton, Dickinson
and Compagny, Sparks, MD, USA) supplémenté de 0,001% d’hémine (10 µg/ml) et de 0,0001% de
vitamine K (1 µg/ml) (THB-HK). A. actinomycetemcomitans a été cultivée dans un milieu « Todd
Hewith broth » supplémenté d’extrait de levure à concentration de 1%.
3.3 Activité antibactérienne des composés
3.3.1 Détermination de la concentration minimale inhibitrice
La capacité du thé vert et de sa principale composante, l’EGCG, à inhiber la croissance des
microorganismes à l’étude a été vérifiée en microplaque 96 puits à fond plat (Sarstedt, St-Léonard,
QC, Canada). À partir d’une culture fraîche (24 h) des microorganismes, ensemencés la veille dans
leur milieu respectif, une dilution a été effectuée afin d’obtenir une densité optique à 655 nm de 0,2

59
(DO655). Par la suite, 100 µl de la suspension bactérienne a été ajoutée à chacun des puits, en
présence de 100 µl d’une concentration décroissante (concentration finale de 8 mg/ml à 15,625 µg/ml)
de l'extrait de thé vert et d’EGCG, dont les dilutions ont préalablement été préparées dans du milieu
THB-HK stérile (dilution sérielle 1:2). D’un puits à l’autre, la concentration de la molécule d’intérêt est,
par conséquent, diminuée de moitié.
Deux types de contrôles ont été utilisés afin d'assurer une standardisation de la méthode
expérimentale. Tout d’abord, certains puits contenaient la culture bactérienne seule (100 µl de la
suspension bactérienne à une DO655 de 0,2 et 100 µl de milieu de culture). Un contrôle de la molécule
d’intérêt seule (100 µl de l’extrait à la dilution appropriée et 100 µl de milieu de culture) a également
été effectué.
Après une incubation de 24 heures à 37 °C sous des conditions d’anaérobiose, la croissance
bactérienne a été évaluée par une observation visuelle et validée en mesurant la DO655 à l’aide d’un
lecteur de microplaque (BioRad Laboratories, Minississauga, ON, Canada). Les valeurs (µg/ml) de la
concentration minimale inhibitrice (CMI) sont définies comme la plus faible concentration de la
molécule d’intérêt pour laquelle, visuellement ainsi qu’à l’aide du lecteur de microplaque, aucune
croissance n’est notée.
Pour assurer la reproductibilité des résultats obtenus ainsi que la validité de la méthode, les différents
essais ont été effectués en triplicata et pour un minimum de trois fois. De plus, afin d’établir une
comparaison entre la molécule étudiée et les traitements thérapeutiques utilisés actuellement dans le
domaine clinique, l’activité antibactérienne de la tétracycline ainsi que celle du métronidazole ont
également été vérifiées en utilisant la méthode expérimentale décrite ci-haut (concentration finale de
200 à 0 µg/ml).

60
3.3.2 Détermination de la concentration minimale bactéricide
Dans les puits où la croissance bactérienne a été inhibée par les substances à l’étude, la concentration
minimale bactéricide (CMB), qui correspond à la plus faible concentration ayant permis de tuer tous
les microorganismes provenant de l’inoculum de départ, a été établie. Pour déterminer cette valeur, un
aliquote de 5 µl des puits de la microplaque qui ne révèlent pas de croissance microbienne visible a
été prélevé et déposé sur leur milieu respectif de culture gélosé pour ensuite être incubé à 37 °C, pour
une durée de 5 jours, en respectant les conditions de culture des microorganismes étudiés. La CMB a
été définie par la plus petite concentration à laquelle aucune croissance sur le milieu gélosé n’est
observée. Pour assurer la reproductibilité des résultats obtenus ainsi que la validité de la méthode, les
différents essais ont été effectués en triplicata.
3.4 Évaluation de l’effet synergique
L’évaluation des effets synergiques potentiels entre les polyphénols du thé vert en combinaison avec
un antibiotique classique (tétracycline, métronidazole) a été réalisée par la méthode de microdilution
en damier décrite précédemment par Hall, Middleton et Westmacott [474], Betts et coll. [475] ainsi que
Hwang et coll. [476]. Ces analyses ont été effectuées sur les bactéries P. gingivalis ATCC 33277,
ATCC 49417, W83 et HW24D1, F. nucleatum ATCC 25586 ainsi que A. actinomycetemcomitans
ATCC 29522. Les concentrations minimales inhibitrices des composés seuls ont tout d’abord été
déterminées par la méthode décrite précédemment (voir 3.3.1) et les différentes combinaisons de
molécules pour lesquelles les effets synergiques ont été évalués sont présentées dans le Tableau 3.

61
Tableau 3. Combinaisons de molécules d’intérêt utilisées lors des essais d’effets synergiques
3.4.1 Préparation des microplaques
Les différentes solutions des composantes à l’étude ont été préparées en étant quatre fois plus
concentrées, afin que les dilutions subséquentes n’altèrent en rien la concentration de départ désirée.
Dilution sérielle en plaque pour le composé A (Thé vert et EGCG; axe de l’abscisse)
Pour le composé A, une dilution sérielle (1:2) en microplaque a été effectuée. À cet effet, un volume
de 50 µl de la molécule à l’étude se retrouve en concentration décroissante (concentration finale de
2 mg/ml à 7,8125 µg/ml) dans les puits des colonnes 3 à 11 de la Figure 6.
Dilution sérielle en tube pour le composé B (Tétracycline et Métronidazole; axe de l’ordonnée)
Les dilutions du composé B ont été effectuées en tubes afin de ne pas affecter la concentration finale
du composé A, ajouté préalablement dans les puits des colonnes 3 à 11. Un volume correspondant de
50 µl a été déposé dans chacun des puits des rangées A à F.
Combinaisons Composé A Composé B
1
Extrait de thé vert
Métronidazole
2 Tétracycline
3
EGCG
Métronidazole
4 Tétracycline

62
Ajout de l’inoculum bactérien
Suite à la conception de la microplaque, un inoculum de 100 µl de la suspension bactérienne de P.
gingivalis, F. nucleatum ou A. actinomycetemcomitans a été ajouté dans chacun des puits, à partir
d’une culture fraîche de 18 à 24 heures dont la DO660 a été ajustée à 0,2 à l’aide de milieu de culture
frais. Des puits sans bactéries ou sans composés ont été ajoutés à titre de contrôles dans cet essai.
Par la suite, les microplaques ont été incubées à 37 °C, en condition anaérobie pour une période de
24 h. La représentation schématique de la microplaque se retrouve dans la Figure 6.
Figure 6. Disposition des dilutions des composantes à l’étude et des contrôles dans la plaque 96 puits
pour les essais de synergie. Dans cette représentation schématique, les contrôles utilisés sont illustrés
en couleur. En bleu on observe le composé B seul et en vert le composé B seul avec la suspension
bactérienne. La couleur orange correspond au composé A seul et la suspension bactérienne en présence
du composé A est représenté par le jaune. Le contrôle bactérien sans composé est illustré par la couleur
grise et le milieu de culture seul est représenté en noir.

63
Après la période d’incubation, la croissance bactérienne a été évaluée par une lecture de la DO655 à
l’aide d’un lecteur de microplaque (BioRad Laboratories) ainsi qu’une observation visuelle. Ces
observations permettent de confirmer la présence ou non d’une croissance bactérienne en fonction de
la turbidité du milieu. À l’aide des données de CMI recueillies, étant définie comme la plus faible
concentration à laquelle aucune croissance bactérienne n’est observée, il est possible de calculer le
« Fractional inhibitory concentration index » qui est basé sur la méthode décrite par Hall, Middleton et
Westmacott [474], et qui est défini comme la somme des FICs de chaque agent antimicrobien testé :
CMI AB : Plus petite dilution du composé A qui inhibe la croissance bactérienne à la concentration
sous-inhibitrice de B
CMI BA : Plus petite dilution du composé B qui inhibe la croissance bactérienne à la concentration
sous-inhibitrice de A
Selon Hwang et coll. [476] et Mazumdar et coll. [477], une valeur de FICI de < 0.5 indique un effet
synergique, 0.5 ≤ FICI < 1 indique une synergie partielle, un FICI = 1 révèle un effet additif, 2 ≤ FICI <
4 montre un effet d’indifférence entre les composés et un effet antagoniste est obtenu dans la mesure
si l’indice FICI > 4.
3.5 Culture cellulaire
Les monocytes U937 ainsi que les fibroblastes gingivaux humains HGF-1 sont les deux lignées
cellulaires utilisées dans cette étude. D’une part, les monocytes U937, une lignée cellulaire
immortalisée de la leucémie monoblastique humaine obtenue de l’American Type Culture Collection
(ATCC CRL-1593.2), ont été cultivés dans le milieu Roswell Park Memorial Institute 1640 (RPMI;
Mediatech Inc., A Corning Subsidiary, Manassas, VA 20109, États-Unis) supplémenté de L-glutamine,
de 10% de sérum fœtal bovin (FBS) inactivé par la chaleur (56 °C; 30 minutes) ainsi que de pénicilline-
streptomycine (100 μg/ml) (RPMI-FBS) à une température de 37 °C et dans une atmosphère contenant
5% de CO2.
FICindex= FICA+FICB FICA= CMI AB /CMI A FICB= CMI BA/CMI B

64
Afin d’induire la différenciation des monocytes en macrophages adhérents, ceux-ci ont été incubés
(6,25 x 106 cellules/ml) dans du milieu RPMI-FBS contenant 10 ng/ml de « phorbol-12-myristate-13-
acetate » (PMA; Sigma Chemical. Co., St-Louis, MO, USA) pour une durée de 24 h. Par la suite, le
milieu a été remplacé par du milieu frais et les cellules ont été incubées dans les conditions de culture
décrites précédemment pour 24 h supplémentaires. Les macrophages, maintenant adhérés à la paroi
du flasque, ont été récupérés en râtelant le fond de celui-ci et suspendus à la concentration désirée
(Objectif 3.7).
D’autre part, les fibroblastes gingivaux humains (HGF-1) obtenus de l’ATCC (CRL-2014) ont été
cultivés dans le milieu « Dulbecco's modified Eagle's medium » (DMEM), supplémenté de 4,5 g/l de
glucose, L-glutamine et sodium pyruvate (Mediatech Inc.), ainsi que de 15% de FBS (Sigma-Aldrich
Corp.) inactivé par la chaleur, de pénicilline et de streptomycine (100 µg/ml) ainsi que d’amphotéricine
B (0,5 μg/ml). Les cellules ont été incubées à 37 °C dans un environnement contrôlé contenant 5% de
CO2. Le milieu de culture est renouvelé tous les 2 à 3 jours, et ce, jusqu’à la confluence des cellules.
Pour la réalisation des différents objectifs du projet, ces cellules ont été récoltées suite à l’ajout d’une
solution de trypsine-acide éthylènediaminetétraacétique (trypsine-EDTA à une concentration de
0.05%; Gibco Life Technologies Grand Island, NY, États-Unis). Les cellules encore attachées au fond
du flasque ont été récupérées en râtelant le fond de celui-ci et suspendues à la concentration désirée
(Objectif 3.7).
3.6 Cytotoxicité des composés à l’étude sur les lignées cellulaires
individuelles
Avant d’aborder la suite de ce volet expérimental, la cytotoxicité du thé vert et de l’EGCG à l’égard des
monocytes U937 différenciés en macrophages ainsi que des fibroblastes gingivaux humains a été
déterminée à l’aide d’un essai colorimétrique MTT (Bromure de 3-[4, 5-diméthylthiazol-2-yl]-2, 5-
diphényltétrazolium), en suivant le protocole établi par le manufacturier (Roche Diagnostics, Manheim,
Allemagne). Ce test est basé sur le changement de coloration du sel de tétrazolium MTT, du jaune au
violet (formazan), lors de sa réduction par une enzyme mitochondriale, la succinate déshydrogénase;

65
ainsi, la viabilité cellulaire est estimée par une mesure colorimétrique proportionnelle à la quantité de
cellules vivantes et à l’activité métabolique de ces dernières [478].
Les deux lignées cellulaires, cultivées en microplaque, ont été traitées avec des concentrations
croissantes de thé vert et d’EGCG (entre 500 et 0,78125 µg/ml) pour une durée de 24 h, durée ayant
été prédéterminée pour les expérimentations subséquentes, avant l’ajout de 10 μl de réactif MTT. Par
la suite, une incubation de 4 h a permis l’observation d’un précipité violet dans les puits. Avant la
mesure de l’absorbance à 550 nm (A550), 100 μl de la solution de solubilisation est ajouté à chacun des
puits suivis d’une incubation sur la nuit.
Pour tous les volets subséquents de cette étude, des concentrations de polyphénols déterminées par
ces tests de viabilité ont été utilisées et sont présentées dans le Tableau 2 de l’annexe. Les
concentrations maximales non cytotoxiques pour les deux types cellulaires se situent autour de
62.5 µg/ml en ce qui concerne l'extrait de thé vert et l’EGCG.
3.7 Mise au point d’un modèle de co-culture macrophages/fibroblastes
Le second objectif de l’étude était d’évaluer les propriétés anti-destruction tissulaire du thé vert et de
son principal polyphénol, soit l’EGCG. Pour ce faire, un modèle de co-culture macrophages-
fibroblastes a été utilisé, puisque celui-ci permet de prendre en considération les interactions entre les
fibroblastes gingivaux humains, cellules constitutives les plus importantes de la gencive, ainsi que les
macrophages, cellules prédominantes lors du processus inflammatoire caractéristique de la
parodontite. Les effets de polyphénols du thé vert sur l’inhibition de la sécrétion de MMPs ont été
évalués.
Comme la littérature ne mentionne pas les proportions in vivo de ces deux types cellulaires lors d’un
état faiblement/fortement inflammatoire, les deux ratios suivants ont été définis arbitrairement et ont
été testés, le ratio 1:10 (macrophages : fibroblastes) étant utilisé pour représenter un état faiblement

66
inflammatoire de la poche parodontale (santé parodontale) et le ratio 10:1 (macrophages : fibroblastes)
pour refléter un état fortement inflammatoire du parodonte [479, 480].
Dans le modèle de co-culture utilisé, les macrophages ont été déposés sur une matrice de collagène
contenant les fibroblastes (Figure 7). Tout d’abord, la solution de collagène a été préparée en
mélangeant, à la température ambiante, 125 µl de milieu DMEM (10X) dont le pH a été préalablement
ajusté à 7.34 (1 ml de milieu DMEM [10X] auquel ont été ajoutés 95 µl de NaOH 1N), ainsi que 1 ml
d’une solution de collagène. Le « Purified Bovine Collagen Solution » (1ml), possédant une
concentration initiale de 3 mg/ml (pH ~ 2.0), a été alcalinisé à l’aide de 250 µl de NaOH 0.1N. L’ajout
de NaOH favorise une alcalinisation du milieu près de la neutralité, permettant ainsi sa solidification
[481, 482]. Suite à la trypsinisation des fibroblastes, ceux-ci ont été resuspendus dans ce milieu à une
concentration de 1,5 x 106 cellules/ml pour le ratio 1:10, ainsi qu’à une concentration de 0,15 x 106
cellules/ml pour le ratio 10:1. Un ml de la solution de collagène contenant les fibroblastes a été disposé
à l’intérieur d’une plaque de 12 puits (Sarstedt) et celle-ci a été incubée à 37 °C pour une durée de 2 h
dans une atmosphère où la concentration de CO2 n’est pas contrôlée. Cette incubation permet au
milieu de se gélifier. Après l’incubation, 1 ml de milieu RPMI-1% FBS (avec antibiotiques) a été ajouté
à la surface du gel afin d’éviter la déshydratation et la plaque a été incubée à 37 °C dans un
environnement contenant 5% de CO2 jusqu’à la fin de la journée. À la fin de la journée, 1 ml des
macrophages suspendus selon les deux rations cellulaires préalablement établis sont ajoutés à la
surface de la matrice de collagène dans le milieu RPMI-1% FBS (sans antibiotiques). Par la suite, la
plaque est incubée toute la nuit (37 °C dans une atmosphère contenant 5% CO2) afin de permettre
l’adhésion des macrophages. Le lendemain, le milieu dans lequel baignent les macrophages a été
aspiré et remplacé par 1,5 ml de milieu RPMI-1% FBS (sans antibiotiques) frais. La stimulation a été
effectuée subséquemment selon les différents protocoles (Objectifs 3.8 et 3.9).

67
Figure 7. Représentation schématique du modèle de co-culture macrophages/fibroblastes
3.8 Capacité des polyphénols d’intérêt à inhiber la sécrétion de
métalloprotéinases matricielles (MMP-3, MMP-8 et MMP-9) par le modèle de
co-culture
Cet objectif a permis d’évaluer l’effet des polyphénols à l’étude sur l’inhibition de la sécrétion de
diverses MMPs par le modèle de co-culture, lorsque celui-ci est stimulé, préalablement ou non, avec
le LPS de la bactérie A. actinomycetemcomitans ATCC 29522.
Dans un premier temps, l’effet des polyphénols sur la sécrétion basale du modèle de co-culture (sans
LPS) a été vérifié. L’extrait de thé vert ainsi que l’EGCG ont été ajoutés à quatre concentrations non-
cytotoxiques (62,5 µg/ml, 31,25 µg/ml, 15,25 µg/ml, 7,8125 µg/ml) au modèle de co-culture
macrophages/fibroblastes (ratios 1 :10 et 10 :1). Une incubation à une température de 37 °C dans un
environnement contenant 5% de CO2 pour une durée de 24 h a été réalisée. Suite à l’incubation, le
surnageant a été recueilli, centrifugé pour éliminer les cellules restantes et conservé à -20 °C jusqu’à
la détermination des concentrations de MMP-3, MMP-8 et MMP -9. La détection a été effectuée par un
Milieu RPMI – 1% FBS 1,5 ml
Matrice de collagène 1,5 ml

68
dosage immuno-enzymatique ELISA en utilisant des trousses commerciales (R&D systems). La
présente expérimentation a été effectuée en duplicata. Les résultats de cet objectif ont permis de
mettre en lumière l’effet des polyphénols du thé vert sur la sécrétion des MMPs lors de l’absence
d’inflammation gingivale.
Afin de vérifier l’intérêt des composés naturels à des fins préventives, ceux-ci ont été ajoutés à quatre
concentrations non cytotoxiques (62,5 µg/ml, 31,25 µg/ml, 15,25 µg/ml, 7,8125 µg/ml) au modèle de
co-culture macrophages/fibroblastes (ratios 1 :10 et 10 :1). Une incubation à une température de 37 °C
dans un environnement contenant 5% de CO2 pour une durée de 2 h a été réalisée avant de stimuler
le modèle avec le LPS d’A. actinomycetemcomitans à une concentration finale de 1 µg/ml. Après 22 h
d’incubation, le surnageant a été recueilli, centrifugé pour éliminer les cellules restantes et conservé à
-20 °C jusqu’à la détermination des concentrations de MMP-3, MMP-8 et MMP -9. La détection a été
effectuée par un dosage immuno-enzymatique ELISA en utilisant des trousses commerciales (R&D
systems). La présente expérimentation a été effectuée en duplicata. Les résultats de cet objectif ont
permis de mettre en évidence la capacité des polyphénols du thé vert à réduire la sécrétion des MMPs
et ainsi permettre leur utilisation potentielle à titre d’agents préventifs de la maladie parodontale.
De plus, afin de vérifier l’intérêt thérapeutique des divers composés naturels à l’étude, ceux-ci ont été
ajoutés à différentes concentrations suivant une stimulation du modèle de co-culture
macrophages/fibroblastes (ratios 1 :10 et 10 :1) avec le LPS d’A. actinomycetemcomitans à une
concentration finale de 1 µg/ml. Une incubation à une température de 37 °C dans un environnement
contenant 5% de CO2 pour une durée de 2 h s’est effectuée avec le LPS avant de traiter le modèle à
l’aide des polyphénols d’intérêt à quatre concentrations non cytotoxiques (62,5 µg/ml, 31,25 µg/ml,
15,25 µg/ml, 7,8125 µg/ml). Après 22 h d’incubation, le surnageant a été recueilli, centrifugé pour
éliminer les cellules restantes et conservé à -20 °C jusqu’à la détermination des concentrations de
MMP-3, MMP-8 et MMP -9. La détection a été effectuée par un dosage immuno-enzymatique ELISA
en utilisant des trousses commerciales (R&D systems). La présente expérimentation a été effectuée
en duplicata. Les résultats de cet objectif ont permis vérifier la capacité des polyphénols d’intérêt à
réduire la réponse inflammatoire déjà prédominante dans certains cas clinique en inhibant la sécrétion
métalloprotéinases matricielles.

69
3.9 Analyses statistiques
La moyenne et l’écart-type des deux réplicas a été calculée et le test t de Student a été utilisé pour les
analyses statistiques avec une valeur de p ≤ 0.05 considérée comme statistiquement significative.

70

71
4. RÉSULTATS
4.1 Activité antibactérienne des composés
4.1.1 Concentrations minimales inhibitrices et bactéricides
Au cours de ce projet, l’activité antibactérienne d'un extrait de thé vert et de l'EGCG a été évaluée sur
trois espèces bactériennes associées à l'étiologie des maladies parodontales, soit P. gingivalis (ATCC
33277, ATCC 49417, W83, HW24D1), F. nucleatum ATCC 25586, et A. actinomycetemcomitans
ATCC 29522. Pour la réalisation de cette analyse, les différents microorganismes ont été cultivés pour
une période de 24 heures en condition d’anaérobiose, en présence de concentrations décroissantes
(8000 à 7,8125 µg/ml) d'un extrait de thé vert ou d’EGCG. Les concentrations minimales inhibitrices
et bactéricides de l’extrait de thé vert, de l'EGCG ainsi que de deux antibiotiques (métronidazole et
tétracycline) utilisés actuellement en thérapie parodontale sont présentées dans le Tableau 4.
Dans un premier temps, les résultats ont démontré qu’à une concentration de 250 µg/ml, l’extrait de
thé vert inhibe totalement la croissance de P. gingivalis ATCC 33277 en milieu THB-HK, et tue
l’inoculum de départ à une concentration de 500 µg/ml, qui correspond ainsi à la CMB. Les valeurs de
CMI et de CMB des trois autres souches de P. gingivalis (ATCC 49417, HW24D1, W83) se situaient à
500 µg/ml et 1000 µg/ml, respectivement.
De façon générale, il est possible de constater qu’à l’exception de P. gingivalis ATCC 49417, qui
démontre une sensibilité semblable au thé vert et à l’EGCG (CMI = 500 µg/ml, CMB = 1000 µg/ml),
les autres souches de P. gingivalis s'avèrent plus sensibles à l’action de l’EGCG. Effectivement, une
concentration d’EGCG de 125 µg/ml (CMB =250 µg/ml) permet d’inhiber la croissance de P. gingivalis
ATCC 33277, tandis que P. gingivalis W83 et HW24D1, quant à elles, montrent une CMI de 250 µg/ml
et une CMB de 500 µg/ml.

72
En ce qui concerne le métronidazole, une concentration de 6,25 µg/ml (CMB = 12,5 µg/ml) s'est avérée
nécessaire pour inhiber la croissance de P. gingivalis ATCC 33277. L’effet inhibiteur du métronidazole
a été observé à une concentration de 50 µg/ml (CMB = 100 µg/ml) pour la bactérie P. gingivalis ATCC
49417, et à 1,5625 µg/ml (CMB = 6,25 µg/ml) pour P. gingivalis W83 et HW24D1. La CMI pour la
tétracycline se situait entre 3,125 µg/ml et 1,5625μg/ml, alors que la CMB était entre 6,25 et 50 µg/ml.
Dans le cas de F. nucleatum, la CMI pour l’extrait de thé vert se situait à 1000 µg/ml, alors que la CMB
correspondait à 2000 µg/ml. Une inhibition totale de la croissance a été observée lors de l’ajout
d’EGCG à une concentration de 500 µg/ml, tandis que 1000 µg/ml (CMB) étaient nécessaires afin de
tuer complètement l'inoculum de départ. D'une part, le métronidazole a démontré une CMI de
1,5625 µg/ml et une CMB de 3,125 µg/ml. D'autre part, la CMI et la CMB de la tétracycline
correspondait à 0,1953125 µg/ml et 0,78125 µg/ml, respectivement.
Finalement, en ce qui concerne l'espèce A. actinomycetemcomitans, la CMI de l'extrait de thé vert se
situait à une valeur de 1000 µg/ml (CMB= 4000 µg/ml), tandis qu’une concentration d’EGCG de
500 µg/ml était nécessaire pour inhiber la croissance de cette dernière. La CMB de l'EGCG contre A.
actinomycetemcomitans était supérieure à la plus forte concentration testée, soit 4000 μg/ml. Le
métronidazole inhibait totalement la croissance d’A. actinomycetemcomitans à une concentration de
50 µg/ml (CMB = 100 µg/ml), tandis que la tétracycline nécessitait des concentrations plus faibles pour
inhiber la croissance bactérienne (CMI = 12,5 µg/ml), et causer la mort de l'inoculum (CMB = 200
µg/ml). A. actinomycetemcomitans était l'espèce bactérienne qui démontrait en général une sensibilité
moindre à l’action de l’extrait de thé vert et de l’EGCG.

73
Tableau 4. Détermination des concentrations minimales inhibitrices et bactéricides vis-à-vis trois espèces
bactériennes parodontopathogènes des polyphénols du thé vert

74
4.1.2 Détermination de l’effet synergique
Tel que présenté dans le Tableau 4, les espèces bactériennes à l'étude se sont avérées relativement
sensibles aux effets du thé vert et de EGCG, de même qu’aux antibiotiques utilisés à titre de contrôle.
Dans l'optique d'un contrôle préventif et thérapeutique des maladies parodontales, l’activité synergique
de l'extrait de thé vert et du EGCG en combinaison avec différents antibiotiques actuellement utilisés
en thérapie, plus spécifiquement le métronidazole et la tétracycline, a été évaluée. À l'aide de la
méthode de microdilution en damier, les différentes combinaisons polyphénols/antibiotiques ont été
testées sur P. gingivalis, F. nucleatum et A. actinomycetemcomitans (Tableaux 5 et 6).
D'une part, dans leur association avec le métronidazole, l’extrait de thé vert et l’EGCG ont démontré
un effet synergique, avec des FICIs respectifs de 0,28125 et 0,3125 envers P. gingivalis ATCC 33277
(Tableau 5). D'autre part, un effet de synergie partielle a été obtenu en combinant la tétracycline avec
les polyphénols à l’étude (FICI=0,75).
Trois souches additionnelles de P. gingivalis (W83, HW24D1 et ATCC 49417) ont été utilisées pour
confirmer ces résultats de synergie. P. gingivalis W83 s'est avéré la souche la plus sensible aux
différentes combinaisons qui ont été testées (Tableau 6). Effectivement, l’extrait de thé vert, en
association avec le métronidazole et la tétracycline, ont démontré un effet synergique, avec des FICIs
respectifs de 0,375 et 0,3125. L’EGCG, lors de son association avec le métronidazole, a également
résulté en un effet synergique (FICI = 0,3125), tandis qu’une synergie partielle a été obtenue avec la
combinaison EGCG et tétracycline (FICI = 0,5). Tant l'extrait de thé vert que l'EGCG, en combinaison
avec le métronidazole, ont démontré un effet de synergie partielle, avec un FICI de 0,5 sur P. gingivalis
HW24D1. Un effet additif a été observé lors de la combinaison de la tétracycline avec l’extrait de thé
vert et l’EGCG sur cette même bactérie. En général, P. gingivalis ATCC 49417 s'est avéré la bactérie
la moins sensible aux combinaisons étudiées. L’EGCG et le métronidazole ont démontré une synergie
partielle, avec un FICI de 0,75. L’EGCG avec la tétracycline, de même que la combinaison de l’extrait
de thé vert et du métronidazole ont démontré un effet additif (FICI = 1). La combinaison de l’extrait de
thé vert et de la tétracycline s’est avérée moins efficace, puisqu’un effet d’indifférence a été observé
(FICI = 1,5).

75
En ce qui concerne F. nucleatum, la presque totalité des combinaisons qui ont été testées, soit l’extrait
de thé vert avec les deux antibiotiques testés, de même que l’EGCG avec le métronidazole ont
démontré des effets synergiques (Tableau 5). Seule la combinaison de l’EGCG et de la tétracycline a
démontré un effet de synergie partielle, avec un FICI de 0,5. Dans le cas d'A. actinomycetemcomitans,
des effets de synergie partielle ont été observés lors de la combinaison du métronidazole avec l’extrait
de thé vert et de l’EGCG, avec des FICI de 0,5, tandis que la tétracycline en combinaison avec les
polyphénols à l’étude résultait en un effet additif (FICI = 1) (Tableau 5).
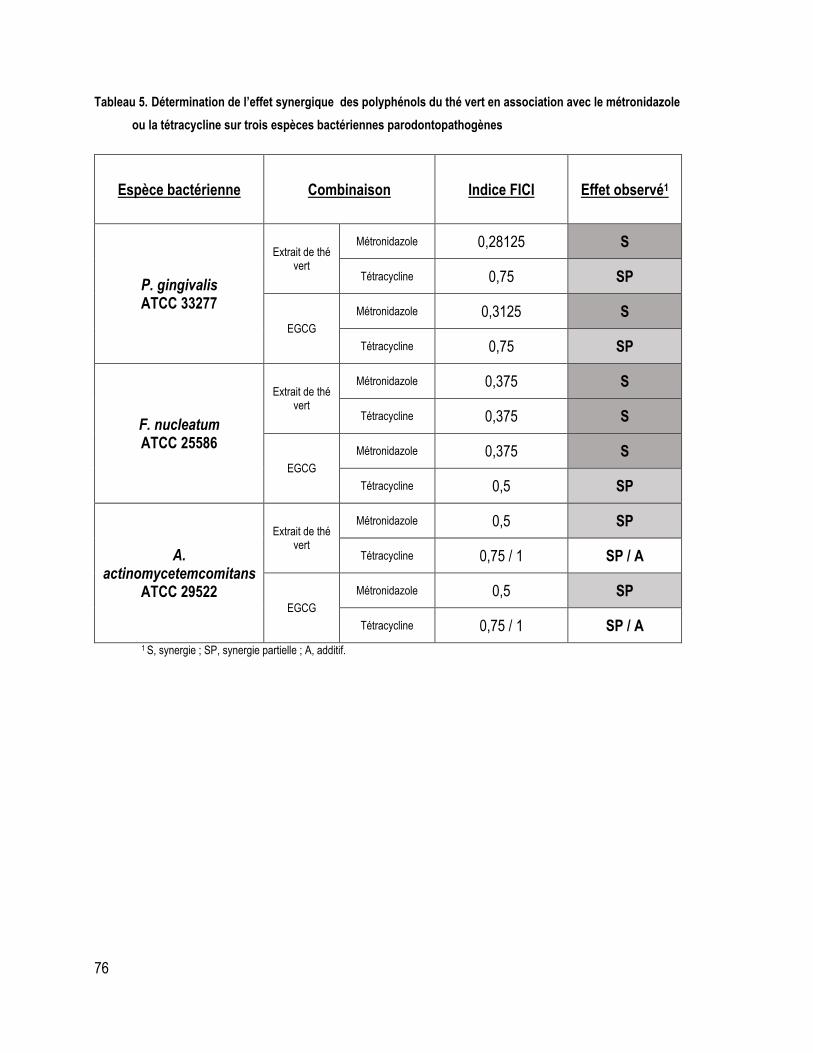
76
Tableau 5. Détermination de l’effet synergique des polyphénols du thé vert en association avec le métronidazole
ou la tétracycline sur trois espèces bactériennes parodontopathogènes
1 S, synergie ; SP, synergie partielle ; A, additif.
Espèce bactérienne Combinaison Indice FICI Effet observé1
P. gingivalis ATCC 33277
Extrait de thé vert
Métronidazole 0,28125 S
Tétracycline 0,75 SP
EGCG
Métronidazole 0,3125 S
Tétracycline 0,75 SP
F. nucleatum ATCC 25586
Extrait de thé vert
Métronidazole 0,375 S
Tétracycline 0,375 S
EGCG
Métronidazole 0,375 S
Tétracycline 0,5 SP
A. actinomycetemcomitans
ATCC 29522
Extrait de thé vert
Métronidazole 0,5 SP
Tétracycline 0,75 / 1 SP / A
EGCG
Métronidazole 0,5 SP
Tétracycline 0,75 / 1 SP / A

77
Tableau 6. Détermination de l’effet synergique des polyphénols du thé vert en association avec le métronidazole
ou la tétracycline sur P. gingivalis
1 S, synergie ; SP, synergie partielle ; A, additif ; I, indifférence
Souches Combinaison Indice FICI Effet observé1
P. gingivalis W83
Extrait de thé vert
Métronidazole 0,375 S
Tétracycline 0,3125 S
EGCG
Métronidazole 0,3125 S
Tétracycline 0,5 SP
P. gingivalis HW24D1
Extrait de thé vert
Métronidazole 0,5 SP
Tétracycline 1 A
EGCG
Métronidazole 0,5 SP
Tétracycline 1 A
P. gingivalis ATCC 49417
Extrait de thé vert
Métronidazole 1 A
Tétracycline 1,5 I
EGCG
Métronidazole 0,75 SP
Tétracycline 1 A

78
4.2 Inhibition de la sécrétion de métalloprotéinases matricielles (MMP-3, MMP-
8 et MMP-9) par le modèle de co-culture macrophages/fibroblastes
Cet objectif a permis d’évaluer l’effet des polyphénols du thé vert sur la sécrétion de diverses MMPs
par le modèle de co-culture macrophages/fibroblastes, lorsque celui-ci est stimulé, préalablement ou
non, avec le LPS d’A. actinomycetemcomitans (ATCC 29522).
Tout d'abord, les effets de l’extrait de thé vert et de l’EGCG sur la viabilité des macrophages et des
fibroblastes ont préalablement été déterminés. Un traitement de 24 h des deux lignées cellulaires avec
les molécules à l’étude à des concentrations jusqu'à 62,5 μg/ml en présence de 1 µg/ml du LPS d’A.
actinomycetemcomitans n'a démontré aucun effet cytotoxique basé sur les résultats de l'essai MTT
(voir Annexe). Pour toutes les concentrations testées de l'extrait de thé vert et d'EGCG, la viabilité
cellulaire est demeurée supérieure à 80%, lorsque comparée aux cellules non traitées.
4.2.3 Réponse du modèle de co-culture macrophages/fibroblastes stimulé avec le
LPS d’A. actinomycetemcomitans en condition faiblement (ratio 1 :10) et fortement
inflammatoire (ratio 10:1)
Le modèle de co-culture macrophages/fibroblastes a été utilisé afin de déterminer la capacité d’un
extrait de thé vert et d’EGCG à diminuer la sécrétion des MMPs, plus spécifiquement la MMP-3, la
MMP-8 et la MMP-9, des enzymes hydrolytiques contribuant à la destruction tissulaire caractéristique
de la parodontite. Tout d’abord, la réponse du modèle de co-culture à la suite d’une stimulation ou non
avec le LPS d’A. actinomycetemcomitans a été évaluée, tant en condition mimant une faible
inflammation, représentée par le ratio 1 :10, qu’en condition mimant une forte inflammation,
représentée par le ratio 10 :1. La méthode de dosage immuno-enzymatique ELISA a été employée
pour la détection des MMPs dans le surnageant de culture. Tel que démontré dans le Tableau 7, en
absence de stimulation avec le LPS, le modèle de co-culture à un ratio 1 macrophage / 10 fibroblastes
(faible inflammation) sécrétait des niveaux de MMP-3, MMP-8 et MMP-9 significativement plus faibles
en comparaison avec le ratio 10 macrophages / 1 fibroblaste (forte inflammation)(Figures 8 à 13).

79
Tableau 7. Sécrétion des MMP-3, MMP-8 et MMP-9 par le modèle de co-culture macrophages/fibroblastes à un
ratio 1:10 et 10:1
MMP Quantité de MMP sécrétées (pg/ml)
Ratio 1:10 Ratio 10:1
MMP-3 7401 21462
MMP-8 32 465
MMP-9 1522 128477
La sécrétion de la MMP-3 en réponse à une stimulation avec le LPS s'est avérée significativement
augmentée dans les deux ratios cellulaires, celle-ci étant doublée (7401 pg/ml à 12669 pg/ml) en
condition faiblement inflammatoire et est augmentée d’un facteur 1,5 (21462 pg/ml à 30365 pg/ml)
lors d’une condition fortement inflammatoire (Figures 8 et 9).
La sécrétion de la MMP-8 par le modèle de co-culture a également été significativement augmentée
suite à la stimulation de ce dernier avec le LPS d’A. actinomycetemcomitans. Effectivement, lors de la
comparaison avec le surnageant non-stimulé, la concentration de MMP-8 est doublée (32 pg/ml à 55
pg/ml) dans une situation faiblement inflammatoire et est augmentée par un facteur de 1,5 (465 pg/ml
à 648 pg/ml) lors d’un état fortement inflammatoire (Figures 10 et 11).
Finalement, la sécrétion de la MMP-9 par le modèle de co-culture a été augmentée d'une manière
comparable aux autres MMPs. Plus spécifiquement, la sécrétion de la MMP-9 a été significativement
augmentée pour les deux ratios cellulaires, celle-ci étant doublée (1522 pg/ml à 2637 pg/ml) en
condition faiblement inflammatoire et augmentée d’un facteur 1,5 (128477 pg/ml à 183473 pg/ml) lors
d’une condition fortement inflammatoire (Figures 12 et 13).

80
Figure 8. Sécrétion de la MMP-3 par le modèle de co-culture stimulé avec le LPS d’A.
actinomycetemcomitans en condition faiblement inflammatoires (ratio 1 macrophage : 10
fibroblastes)
Figure 9. Sécrétion de la MMP-3 par le modèle de co-culture stimulé avec le LPS d’A.
actinomycetemcomitans en condition fortement inflammatoire (ratio 10 macrophages : 1
fibroblaste)
*
0
5000
10000
15000
20000
25000
30000
35000
Contrôle milieu (-) Contrôle LPS (+)
MM
P-3
(p
g/m
l)
Ratio 10:1
*
0
2000
4000
6000
8000
10000
12000
14000
16000
Contrôle milieu (-) Contrôle LPS (+)
MM
P-3
(p
g/m
l)
Ratio 1:10

81
Figure 10. Sécrétion de la MMP-8 par le modèle de co-culture stimulé avec le LPS d’A.
actinomycetemcomitans en condition faiblement inflammatoire (ratio 1 macrophage : 10
fibroblastes)
Figure 11. Sécrétion de la MMP-8 par le modèle de co-culture stimulé avec le LPS d’A.
actinomycetemcomitans en condition fortement inflammatoire (ratio 10 macrophages : 1
fibroblaste)
*
0
10
20
30
40
50
60
70
Contrôle milieu (-) Contrôle LPS (+)
MM
P-8
(p
g/m
l)
Ratio 1:10
*
0
100
200
300
400
500
600
700
800
Contrôle milieu (-) Contrôle LPS (+)
MM
P-8
(p
g/m
l)
Ratio 10:1

82
Figure 12. Sécrétion de la MMP-9 par le modèle de co-culture stimulé avec le LPS d’A.
actinomycetemcomitans en condition faiblement inflammatoire (ratio 1 macrophage : 10
fibroblastes)
Figure 13. Sécrétion de la MMP-9 par le modèle de co-culture stimulé avec le LPS d’A.
actinomycetemcomitans en condition fortement inflammatoire (ratio 10 macrophages : 1
fibroblaste)
0
20000
40000
60000
80000
100000
120000
140000
160000
180000
200000
Contrôle milieu (-) Contrôle LPS (+)
MM
P-9
(p
g/m
l)
Ratio 1:10
*
0
500
1000
1500
2000
2500
3000
3500
Contrôle milieu (-) Contrôle LPS (+)
MM
P-9
(p
g/m
l)
Ratio 1:10

83
4.2.4 Effet de l'extrait de thé vert et de l’EGCG sur la sécrétion basale des MMPs en
conditions faiblement et fortement inflammatoires
Avant de vérifier le potentiel des polyphénols du thé vert à des fins préventives et thérapeutiques, l’effet
de l’extrait de thé vert et de l’EGCG sur la sécrétion basale de MMPs par le modèle de co-culture
macrophages/fibroblastes a préalablement été déterminé. Après un traitement d'une durée de 24 h
avec des concentrations croissantes des molécules à l’étude, les surnageants ont été recueillis et les
concentrations de MMP-3, MMP-8 et MMP-9 ont été déterminées par ELISA.
Pour les deux ratios cellulaires utilisés, correspondant à des conditions faiblement et fortement
inflammatoires, l’extrait de thé vert a diminué de façon significative et dose-dépendante la sécrétion de
la MMP-3 (Figures 14 et 15). Plus spécifiquement, à une concentration de 62,5 µg/ml, l’extrait de thé
vert a inhibé la sécrétion basale de la MMP-3 de 36,3% (ratio 1 :10) et de 38,7% (ratio 10 :1). Les effets
inhibiteurs de l’EGCG ont ensuite été vérifiés selon les mêmes conditions. Dans une condition
faiblement inflammatoire, seule la concentration d’EGCG de 62,5 µg/ml a démontré une diminution
significative de la sécrétion de la MMP-3. Par contre, pour ce qui est du ratio représentant une condition
fortement inflammatoire, l’EGCG inhibait, de façon significative et dose dépendante la sécrétion de la
MMP-3 par le modèle de co-culture à partir de 31,25 µg/ml. Une inhibition de 28,9% a été observée à
une concentration de 31,25 µg/ml.
En ce qui concerne la MMP-8 (Figures 16 et 17), il a été démontré qu’en condition faiblement
inflammatoire, l’extrait de thé vert n’a aucun effet sur la sécrétion basale de cette MMP, tandis que
l’EGCG diminue de manière significative la sécrétion de la MMP-8 aux concentrations de 15,625 µg/ml
et 62,5 µg/ml. À la concentration la plus élevée, soit 62,5 µg/ml, une inhibition de 56,6% a été observée
pour l’EGCG. Lors de l’utilisation d’un ratio illustrant une condition fortement inflammatoire, une
diminution a été observée avec l’EGCG à 62,5 µg/ml, quoique non-significative.
Finalement, tant dans un ratio faiblement que fortement inflammatoire, l’extrait de thé vert et l’EGCG
ont été en mesure de diminuer significativement et de façon dose-dépendante la sécrétion de la MMP-

84
9 par le modèle de co-culture (Figures 18 et 19). Pour l’extrait de thé vert, la diminution s’est avérée
significative à 62,5 µg/ml, correspondant à un pourcentage d’inhibition de 36,1% par rapport aux
cellules non-stimulées pour le ratio 1 :10, tandis que cette diminution s’est avérée significative à partir
de 31,25 µg/ml (diminution de 72,9%) en ce qui concerne le ratio 10 :1. Pour l’EGCG, la diminution
s’est avérée significative à partir d’une concentration de 31,25 µg/ml, tant pour le ratio 1:10 (diminution
de 33,4%) que pour le ratio 10 :1 (diminution de 34,6%).
Les résultats de cet objectif ont permis de mettre en évidence la capacité des polyphénols du thé vert
à réduire la sécrétion basale des MMPs, plus spécifiquement les MMP-3 et MMP-9, ce qui supporte
leur potentiel à atténuer la destruction tissulaire engendrée par ces dernières.

85
Figure 14. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-3 en condition faiblement
inflammatoire (ratio 1 macrophage : 10 fibroblastes)
Figure 15. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-3 en condition faiblement
inflammatoire (ratio 10 macrophages : 1 fibroblaste)
* *
**
* *
0
5000
10000
15000
20000
25000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
Ratio 10:1 SANS LPS
MM
P-3
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
**
0
2000
4000
6000
8000
10000
12000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
Ratio 1:10 SANS LPS
MM
P-3
(p
g/m
l)
Concentration de l'extrait thé vert et d'EGCG (μg/ml)

86
Figure 16. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-8 en condition faiblement
inflammatoire (ratio 1 macrophage : 10 fibroblastes)
Figure 17. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-8 en condition fortement
inflammatoire (ratio 10 macrophages : 1 fibroblaste)
0
100
200
300
400
500
600
700
800
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
Ratio 10:1 SANS LPS
MM
P-8
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
*
*
0
10
20
30
40
50
60
70
80
90
100
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
Ratio 1:10 SANS LPS
MM
P-8
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)

87
Figure 18. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-9 en condition faiblement
inflammatoire (ratio 1 macrophage : 10 fibroblastes)
Figure 19. Effet du thé vert et de l’EGCG sur la sécrétion basale de MMP-9 en condition fortement
inflammatoire (ratio 10 macrophages : 1 fibroblaste)
****
*
**
0
50000
100000
150000
200000
250000
300000
350000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
Ratio 10:1 SANS LPS
MM
P-9
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
*
**
0
500
1000
1500
2000
2500
3000
3500
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
Ratio 1:10 SANS LPS
MM
P-9
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)

88
4.2.5 Effet du thé vert et de l’EGCG sur la sécrétion des différentes MMPs en
conditions faiblement et fortement inflammatoires à des fins préventives
Afin de vérifier l’intérêt des composés naturels à des fins préventives, le modèle de co-culture
macrophages/fibroblastes a préalablement été traité pour une durée de 2 h avec des concentrations
croissantes des molécules à l’étude, soit l’extrait de thé vert et l’EGCG, avant l’ajout du LPS d’A.
actinomycetemcomitans, utilisé comme agent stimulateur. Après une incubation subséquente de 24h,
les surnageants ont été recueillis et à l’aide d’une méthode de dosage immuno-enzymatique ELISA,
les concentrations de MMP-3, MMP-8 et MMP -9 sécrétées en réponse à cette stimulation ont été
déterminées.
Dans les deux ratios cellulaires utilisés, correspondant à des conditions faiblement et fortement
inflammatoires, l’extrait de thé vert a diminué de façon significative et dose-dépendante la sécrétion de
la MMP-3 à partir d’une concentration 15,625 µg/ml (Figures 20 et 21). Plus spécifiquement, à une
concentration de 62,5 µg/ml, l’extrait de thé vert a inhibé la sécrétion de la MMP-3 de 43,7% (ratio
1 :10) et de 34,4% (ratio 10 :1). Les effets inhibiteurs de l’EGCG ont ensuite été vérifiés selon les
mêmes conditions. Dans une condition faiblement inflammatoire, seule la concentration d’EGCG de
31,25 µg/ml démontre une diminution significative de la sécrétion de la MMP-3. Par contre, pour ce
qui est du ratio représentant une condition fortement inflammatoire, l’EGCG inhibait, de façon
significative et dose dépendante la sécrétion de la MMP-3 par le modèle de co-culture à partir de 31,25
µg/ml. Une inhibition de 34% a été observée à une concentration de 31,25 µg/ml.
Des effets relativement similaires ont été observés en ce concerne la MMP-8 (Figures 22 et 23).
Effectivement, il a été démontré qu’en condition faiblement inflammatoire, l’extrait de thé vert et l’EGCG
diminuent significativement et de manière dose-dépendante la sécrétion de la MMP-8 dans le
surnageant de culture, à partir d’une concentration 31,25 µg/ml et 15,625 µg/ml respectivement. À la
concentration la plus élevée, soit 62,5 µg/ml, une inhibition de 48,6% a été observée pour l’extrait de
thé vert, tandis que l’EGCG inhibait la sécrétion de la MMP-8 de 32,2% de. Lors de l’utilisation d’un
ratio illustrant une condition fortement inflammatoire, une diminution dose-dépendante quoique

89
seulement significative à des concentrations de 15,625 µg/ml et 62,5 µg/ml a été observée pour
l’extrait de thé vert, et 15,625 µg/ml et 31,25 µg/ml pour l’EGCG.
Finalement, tant dans un ratio faiblement que fortement inflammatoire, l’extrait de thé vert et l’EGCG
ont été en mesure de diminuer significativement et de façon dose-dépendante la sécrétion de la MMP-
9 par le modèle de co-culture (Figures 24 et 25). Pour l’extrait de thé vert, la diminution s’est avérée
significative à partir de 31,25 µg/ml, jusqu’à l’atteinte d’un pourcentage d’inhibition de 59,8% à 62,5
µg/ml pour le ratio 1 :10, tandis que cette diminution s’est avérée significative à partir de 15,625 µg/ml
(20,1 %) en ce qui concerne le ratio 10 :1. Pour l’EGCG, la diminution s’est avérée significative à partir
d’une concentration de 15,625 µg/ml pour le ratio 1 :10, tandis que cette diminution a été significative
à partir de 31,25 µg/ml en ce qui concerne le ratio 10 :1.
Les résultats de cet objectif ont permis de mettre en lumière la capacité des polyphénols du thé vert à
atténuer la sécrétion des MMPs par un modèle de co-culture macrophage/fibroblaste stimulé avec le
LPS. Ces résultats supportent leur utilisation potentielle à titre d’agents préventifs de la maladie
parodontale.

90
Figure 20. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition faiblement inflammatoire
(ratio 1 macrophage : 10 fibroblastes) à des fins préventives
Figure 21. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition fortement inflammatoire
(ratio 10 macrophages : 1 fibroblaste) à des fins préventives
*
**
*
* *
0
5000
10000
15000
20000
25000
30000
35000
40000
45000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 10:1 AVEC LPS
MM
P-3
(p
g/m
l)
Concentration de l'extrait thé vert et d'EGCG (μg/ml)
*
**
*
0
2000
4000
6000
8000
10000
12000
14000
16000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 1:10 AVEC LPS
MM
P-3
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
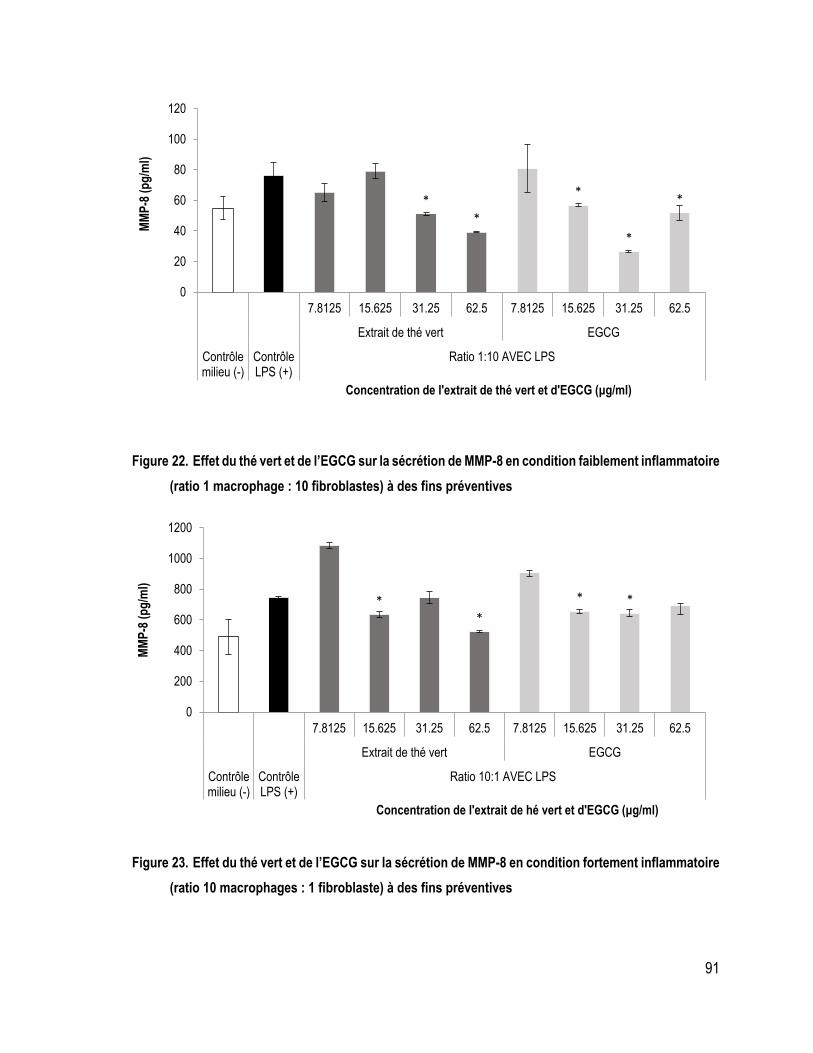
91
Figure 22. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition faiblement inflammatoire
(ratio 1 macrophage : 10 fibroblastes) à des fins préventives
Figure 23. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition fortement inflammatoire
(ratio 10 macrophages : 1 fibroblaste) à des fins préventives
*
*
* *
0
200
400
600
800
1000
1200
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 10:1 AVEC LPS
MM
P-8
(p
g/m
l)
Concentration de l'extrait de hé vert et d'EGCG (μg/ml)
*
*
*
*
*
0
20
40
60
80
100
120
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 1:10 AVEC LPS
MM
P-8
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)

92
Figure 24. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition faiblement
inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins préventives
Figure 25. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition fortement
inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins préventives
*
* ** **
0
50000
100000
150000
200000
250000
300000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 10:1 AVEC LPS
MM
P-9
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
*
*
*
*
*
*
0
500
1000
1500
2000
2500
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 1:10 AVEC LPS
MM
P-9
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)

93
4.2.6 Effet du thé vert et de l’EGCG sur la sécrétion des différentes MMPs en
conditions faiblement et fortement inflammatoires à des fins thérapeutiques
Afin de vérifier l’intérêt des composés naturels à des fins thérapeutiques, le modèle de co-culture
macrophages/fibroblastes a préalablement été traité pour une durée de 2 h avec le LPS d’A.
actinomycetemcomitans, utilisé comme agent stimulateur, avant l’ajout de concentrations croissantes
des molécules à l’étude, soit l’extrait de thé vert et l’EGCG. Après une incubation subséquente de 24h,
les surnageants ont été recueillis et à l’aide d’une méthode de dosage immuno-enzymatique ELISA,
les concentrations de MMP-3, MMP-8 et MMP -9 sécrétées en réponse à cette stimulation ont été
déterminées.
Dans les deux ratios cellulaires utilisés, correspondant à des conditions faiblement et fortement
inflammatoires, l’extrait de thé vert s’est avéré une substance intéressante, puisque ce dernier a
diminué de façon significative et dose-dépendante la sécrétion de la MMP-3 à partir d’une
concentration 7,8125 µg/ml (Figures 26 et 27). Plus spécifiquement, à une concentration de 62,5
µg/ml, l’extrait de thé vert inhibe la sécrétion de la MMP-3 de 47,2% (ratio 1 :10) et de 43,9% (ratio
10 :1). Les effets inhibiteurs de l’EGCG ont ensuite été vérifiés selon les mêmes conditions. Dans une
condition faiblement inflammatoire, l’EGCG a diminué de façon significative et dose-dépendante la
sécrétion de la MMP-3 à partir d’une concentration 7,8125 µg/ml. En ce qui concerne le ratio
représentant une condition fortement inflammatoire, l’EGCG a inhibé, de façon significative et dose
dépendante la sécrétion de la MMP-3 par le modèle de co-culture à partir de 15,625 µg/ml. Une
inhibition de 18,9% a été observée à cette concentration. Comme il a été observé précédemment,
l’extrait de thé vert s’est avéré plus efficace que l’EGCG à inhiber la sécrétion de la MMP-3. Par
conséquent, il est permis de supposer que l’extrait contient d’autres molécules capables d’en réduire
la sécrétion.
Les mêmes effets n’ont pas été observés en ce concerne la MMP-8 (Figures 28 et 29). Il a été démontré
qu’en condition faiblement inflammatoire, l’extrait de thé vert et l’EGCG n’influencent pas
significativement la sécrétion de la MMP-8. Cependant, lors de l’utilisation d’un ratio illustrant une
condition fortement inflammatoire, une diminution dose-dépendante quoique seulement significative à

94
une concentration de 31,25 µg/ml a été observée pour l’extrait de thé vert, et à partir de 15,625 µg/ml
pour l’EGCG.
Finalement, tant dans un ratio faiblement que fortement inflammatoire, l’extrait de thé vert et l’EGCG
ont été en mesure de diminuer significativement et de façon dose-dépendante la sécrétion de la MMP-
9 par le modèle de co-culture (Figures 30 et 31). Pour l’extrait de thé vert, la diminution s’est avérée
significative à partir de 7,8125 µg/ml, jusqu’à l’atteinte d’un pourcentage d’inhibition de 72% à 62,5
µg/ml pour le ratio 1 :10, tandis que cette diminution s’est avérée significative à partir de 31,25 µg/ml
(26,4%) en ce qui concerne le ratio 10 :1. Pour l’EGCG, la diminution s’est avérée significative à partir
d’une concentration de 7,8125 µg/ml pour le ratio 1 :10 (67% à 62,5 µg/ml), tandis que cette diminution
a été significative à partir de 15,625 µg/ml (32,9%) en ce qui concerne le ratio 10 :1.
Les résultats de cet objectif ont permis de mettre en évidence la capacité des polyphénols du thé vert
à réduire la sécrétion des MMPs par un modèle de co-culture préalablement stimulé par le LPS. Ces
résultats supportent leur utilisation potentielle à titre d’agents thérapeutiques de la maladie parodontale
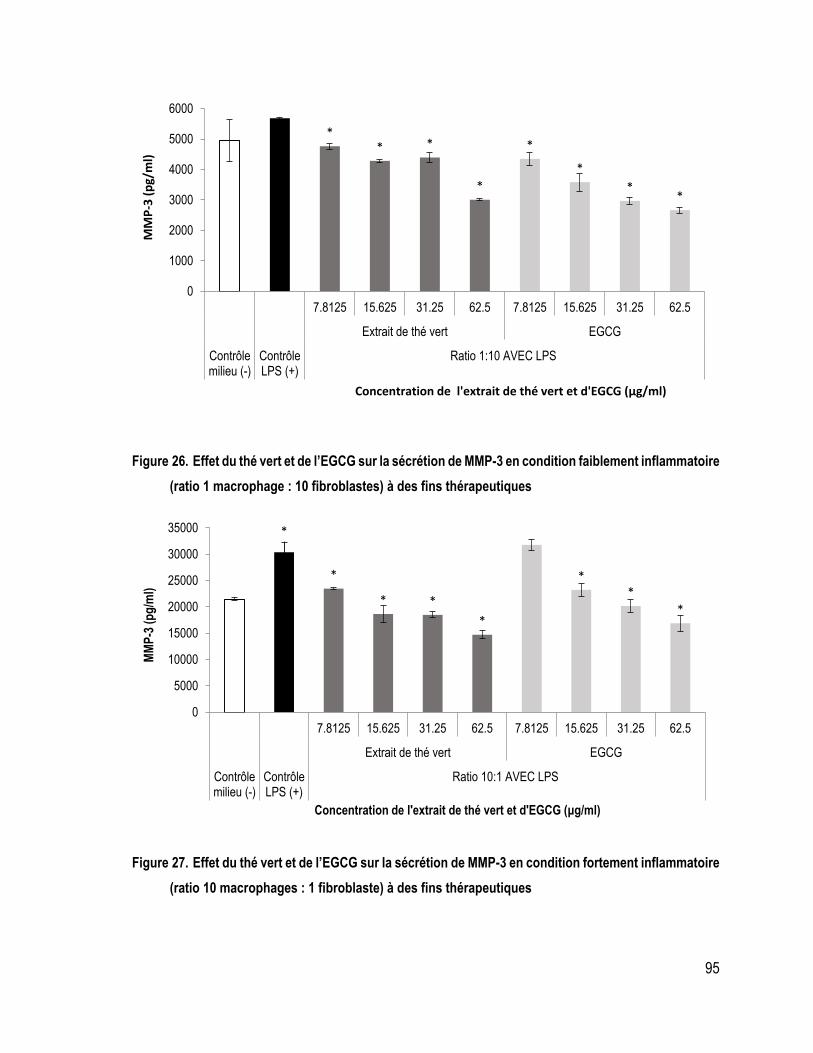
95
Figure 26. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition faiblement inflammatoire
(ratio 1 macrophage : 10 fibroblastes) à des fins thérapeutiques
Figure 27. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-3 en condition fortement inflammatoire
(ratio 10 macrophages : 1 fibroblaste) à des fins thérapeutiques
*
*
* *
*
**
*
0
5000
10000
15000
20000
25000
30000
35000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 10:1 AVEC LPS
MM
P-3
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
** *
*
*
*
**
0
1000
2000
3000
4000
5000
6000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 1:10 AVEC LPS
MM
P-3
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)

96
Figure 28. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition faiblement inflammatoire
(ratio 1 macrophage : 10 fibroblastes) à des fins thérapeutiques
Figure 29. Effet du thé vert et de l’EGCG sur la sécrétion de MMP-8 en condition fortement inflammatoire
(ratio 10 macrophages : 1 fibroblaste) à des fins thérapeutiques
*
*
*
*
0
100
200
300
400
500
600
700
800
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 10:1 AVEC LPS
MM
P-8
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
0
10
20
30
40
50
60
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 1:10 AVEC LPS
MM
P-8
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)

97
Figure 30. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition faiblement
inflammatoire (ratio 1 macrophage : 10 fibroblastes) à des fins thérapeutiques
Figure 31. Effet du thé vert et de l’EGCG sur la sécrétion de la MMP-9 en condition fortement
inflammatoire (ratio 10 macrophages : 1 fibroblaste) à des fins thérapeutiques
*
*
*
* *
0
50000
100000
150000
200000
250000
300000
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 10:1 AVEC LPS (1μg/mL)
MM
P-9
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)
*
**
*
*
* **
*
0
500
1000
1500
2000
2500
3000
3500
7.8125 15.625 31.25 62.5 7.8125 15.625 31.25 62.5
Extrait de thé vert EGCG
Contrôlemilieu (-)
ContrôleLPS (+)
Ratio 1:10 AVEC LPS
MM
P-9
(p
g/m
l)
Concentration de l'extrait de thé vert et d'EGCG (μg/ml)

98

99
5. DISCUSSION
Outre la carie dentaire, les maladies des gencives, plus communément appelées maladies
parodontales, constituent le principal problème buccodentaire chez la population adulte. Il s’agit d’une
infection polymicrobienne considérée comme une maladie chronique inflammatoire [21]. Ces maladies
ont pour conséquence d’entraîner la destruction de l’ensemble des tissus de soutien des dents
constituant le parodonte et peuvent éventuellement conduire à la perte des dents atteintes. En fait, il
est reconnu que ces pathologies représentent la cause principale de la perte des dents chez l’adulte
[483]. La cavité buccale abrite un écosystème microbien complexe, composé de quelque 700 espèces
microbiennes différentes, dont une modification de l’équilibre peut mener à des pathologies buccales
telles que les maladies parodontales. La présence de bactéries spécifiques au sein du biofilm sous-
gingival, notamment P. gingivalis, F. nucleatum et A. actinomycetemcomitans, est considérée comme
le facteur étiologique primaire de cette maladie, tant dans sa forme chronique qu’agressive [45, 110].
Subséquemment, l’expression des facteurs de virulence bactériens engendre une réponse immunitaire
exagérée de l’hôte, menant à la production d’enzymes hydrolytiques (MMPs) et de médiateurs
inflammatoires qui activent diverses voies de dégradation tissulaire et osseuse par les cellules de l’hôte
et sont responsables de la progression et de la sévérité de la parodontite.
Depuis une dizaine d'années, l’intérêt de plusieurs groupes de recherche s'est dirigé vers les composés
naturels, tels que les polyphénols, qui ont démontré de nombreux effets bénéfiques en ce qui a trait à
la prévention de diverses pathologies telles que les malades cardiovasculaires, le cancer, le diabète
et l’arthrite rhumatoïde [339, 340, 450, 484].
Au cours de ce projet de recherche, le potentiel préventif et thérapeutique des polyphénols du thé vert
et de sa principale composante, l'EGCG, a été évalué, notamment par la détermination des propriétés
antibactériennes et anti-destruction tissulaire de ces molécules. Quoique des études antérieures aient
pu démontrer certaines propriétés bénéfiques de ces dernières au niveau systémique, plus
spécifiquement en ce qui concerne leurs propriétés antioxydantes, antibactériennes, anti-
inflammatoires et anti-cancers, peu d’études ont été réalisées dans un contexte d’affections bucco-

100
dentaires. Plus spécifiquement, quelques études ont démontré que les polyphénols du thé vert
neutralisent les composés sulfurés volatils (CSVs) causant l’halitose buccale [395, 397]. Les
polyphénols du thé vert permettent également d’'influencer le processus de formation de la carie
dentaire à plusieurs niveaux, notamment en inhibant la prolifération des streptocoques cariogènes et
la production acide par ces derniers, d’interférer avec le mécanisme d'adhésion des bactéries
cariogènes à l'émail des dents ou d’agir comme inhibiteurs de la glucosyltransférase et de l'amylase
salivaire et bactérienne. En ce qui concerne les maladies parodontales, des études de nature
épidémiologiques ont indiqué une association inverse entre la consommation de thé vert et cette
affection, plus spécifiquement la diminution des divers paramètres cliniques indiquant une affection
parodontale (profondeur des poches parodontales, niveau d'attachement clinique, saignements lors du
sondage) [443]. Toutefois, les mécanismes par lesquels le thé vert et l’EGCG peuvent contribuer à la
prévention des maladies parodontales ne sont pas clairement élucidés et demeurent hypothétiques.
5.1 Propriétés antibactériennes
5.1.1 Concentrations minimales inhibitrices et bactéricides
L’évaluation des propriétés antibactériennes des molécules à l’étude vis-à-vis P. gingivalis, F.
nucleatum ainsi qu'A. actinomycetemcomitans a permis de démontrer qu’une concentration de thé vert
de 250 µg/ml (CMB = 500 µg/ml) est en mesure de réduire significativement la croissance P. gingivalis
ATCC 33277, et qu’une concentration de 500 µg/ml (CMB = 1000 µg/ml) permet l’inhibition de la
croissance des 3 autres souches de P. gingivalis (ATCC 49417, W83 et HW24D1). Une CMI (200
µg/ml) ainsi qu’une CMB (500 µg/ml) comparables ont été retrouvées lors d’une autre étude sur la
bactérie P. gingivalis ATCC 33277 utilisant le même extrait de thé vert. Des extraits de thé blanc,
oolong et noir ont également démontré, au cours de cette même étude, des CMI et des CMB
équivalentes [450]. En ce qui concerne F. nucleatum et A. actinomycetemcomitans, bien que les CMI
soient équivalentes (1000 µg/ml), les CMB se sont avérées plus élevées, avec des valeurs respectives
de 2000 et 4000 µg/ml. En ce qui concerne l’EGCG, une concentration de 125 µg/ml (CMB = 250
µg/ml) a permis l’inhibition de la croissance de P. gingivalis ATCC 33277, une concentration de 250
µg/ml (CMB = 500 µg/ml) a inhibé la croissance de P. gingivalis HW24D1 et W83, tandis qu’une
concentration de 500 µg/ml (CMB = 1000 µg/ml) a permis l’inhibition de la croissance de P. gingivalis

101
ATCC 49417. En ce qui concerne F. nucleatum et A. actinomycetemcomitans, bien que les CMI se
sont avérées équivalentes (500 µg/ml), les CMB se sont avérées plus élevées, avec des valeurs
respectives de 1000 et >4000 µg/ml.
Il est estimé qu’une tasse de thé (2,5 g de feuilles de thé / 200 ml d’eau) contient 90 mg d’EGCG sont
retrouvés, représentant une concentration finale de 450 µg/ml [338]. Différentes études ont démontré
des valeurs de CMI et de CMB comparables en ce qui concerne P. gingivalis. Effectivement, l’une
d’entre elles indique que les polyphénols retrouvés dans le thé, dont l’EGCG inhibe la croissance de
cette bactérie parodontopathogène à des concentrations allant de 250 à 1000 µg/ml [445, 450].
Considérant ces données, il est raisonnable de penser que les effets antibactériens du thé vert et de
l’EGCG, bien que les CMI varient entre les différentes souches et espèces bactériennes, sont
envisageables dans un contexte in vivo. De plus, une étude a démontré que grâce à l’utilisation d’un
système de délivrance à libération lente saturé en Sunphenon, un extrait de feuilles de thé vert ayant
une teneur très élevée en catéchines, et appliqué directement dans les poches parodontales (bandes
d’hydroxypropylcellulose), il est possible, en combinaison avec le traitement mécanique, de réduire
significativement les proportions de Prevotella spp. ainsi que de P. gingivalis. Par conséquent, cette
étude illustre les possibles applications des présents résultats dans une optique de traitement des
maladies parodontales [447].
Bien que des concentrations relativement élevées des molécules à l’étude soient parfois nécessaires
pour inhiber la croissance des différents microorganismes étudiés, des concentrations plus faibles de
ces dernières peuvent suffire cependant à modifier certaines fonctions biologiques des bactéries
parodontopathogènes, notamment l’adhérence aux cellules épithéliales, de même que l’expression
des gènes codant pour certains de leurs facteurs de virulence, tels que les fimbriae, les
hémagglutinines ainsi que les gingipaïnes chez la bactérie P. gingivalis (données en révision pour
publication), diminuant par ailleurs le potentiel pathogène de ce dernier [398, 445, 446, 448, 449].

102
En plus d’inhiber la croissance d'importantes bactéries impliquées dans l’étiologie des maladies
parodontales [445, 447], il a également été démontré que l’EGCG possède un effet sur la croissance
de diverses bactéries pathogènes incriminées dans la carie dentaire ainsi que l’halitose buccale. Plus
spécifiquement, la croissance de S. mutans et S. sobrinus, deux bactéries cariogènes, de même que
S. moorei, impliquée dans l’halitose buccale, sont inhibées par les polyphénols du thé [401, 434]. De
plus, la croissance de bactéries pathogènes impliquées dans diverses infections extra-orales est
également inhibée par l’EGCG, soit Staphylococcus spp. [485-487], Streptococcus pyogenes [488],
Bacillus spp., Clostridium spp. [489], Salmonella typhi [490], ainsi que E. coli [491].
Diverses hypothèses ont été émises afin d’expliquer les mécanismes impliqués dans l’inhibition de la
croissance bactérienne. D’une part, les composantes du thé, soit les catéchines du thé vert et les
théaflavines du thé noir, endommagent de façon irréversible la membrane cytoplasmique bactérienne
[437, 438, 492]. Une analyse en microscopie électronique a permis de mettre en lumière ce mécanisme
à l’aide de la bactérie Pseudomonas aeruginosa. En effet, lors d’essais sur milieu gélosé, il a été
démontré que l’EGCG inhibe la croissance de ce microorganisme. Les analyses subséquentes ont
démontré que l'inhibition de la croissance était la conséquence de la perte de l’intégrité de la membrane
externe de la bactérie [493]. Plus spécifiquement, l'EGCG génère du peroxyde d'hydrogène dans la
bicouche lipidique de la membrane cytoplasmique bactérienne, entraînant la fuite du contenu
intracellulaire [437]. D’autre part, une seconde étude a démontré que l'action antibactérienne des
catéchines du thé vert contre Stenotrophomonas maltophila, un pathogène opportuniste à Gram
négatif, est due à sa capacité à inhiber la dihydrofolate réductase cytoplasmique. Cette enzyme réduit
l'acide dihydrofolique en acide tétrahydrofolique, qui est requis par les bactéries afin de synthétiser les
purines, la thymidylate, ainsi que les précurseurs d'acides nucléiques, qui sont très importants pour la
prolifération cellulaire et la croissance [494]. L'inhibition de cette enzyme par les catéchines du thé vert
a également été observée chez la bactérie E. coli [495].

103
5.1.2 Détermination de l’effet synergique
Par définition, le terme synergie signifie « l’interaction de deux ou plusieurs composés utilisés dans le
traitement de la même pathologie, dont l'effet thérapeutique est supérieur aux effets additionnés de
chacun d'eux pris isolément » [496, 497]. Les plantes, incluant le thé, diffèrent entre eux par la nature
des composés bioactifs qui les composent et par les propriétés antioxydantes de ces derniers. Depuis
la découverte des effets synergiques et leur importance dans le domaine thérapeutique, les
interactions entre les agents biologiquement actifs tels que le thé vert et les divers antibiotiques sont
devenues un sujet important pour la recherche scientifique.
Dans leur association avec le métronidazole et la tétracycline, l’extrait de thé vert et l’EGCG ont
démontré un effet synergique ou partiellement synergique envers P. gingivalis. En ce qui concerne F.
nucleatum, cette bactérie s’est montrée la plus sensible à la presque totalité des combinaisons qui ont
été testées, à l’exception de l’EGCG et de la tétracycline, qui a démontré un effet de synergie partielle.
Enfin, pour A. actinomycetemcomitans, des effets de synergie partielle ont été observés lors de la
combinaison du métronidazole avec les l’extrait de thé vert et de l’EGCG, tandis que la tétracycline en
combinaison avec les polyphénols à l’étude résultait en un effet additif.
Les cibles d’action complémentaires des molécules utilisées en combinaison au cours de cette étude
pourraient expliquer les effets observés. Tel que mentionné précédemment, l’EGCG exerce un effet
antibactérien par la formation de pores dans les membranes bactériennes [437]. Le métronidazole est
un antibiotique appartenant à la famille des nitro-imidazolés; les métabolites du métronidazole
interagissent avec l’ADN des microorganismes, inhibant sa synthèse et menant à la lyse des cellules
[498]. Cet antibiotique s’avère plus approprié pour le traitement des parodontites chroniques, puisque
ce dernier permet l’éradication des bactéries anaérobies strictes, dont P. gingivalis [88]. La tétracycline,
quant à elle, s’attache à la sous-unité ribosomale 30S et interfère avec l’ARNm, inhibant ainsi la
réplication bactérienne et/ou la synthèse des protéines nécessaires à ses fonctions [7, 499]. Retrouvés
en grande concentration dans le fluide créviculaire suite à son administration, les tétracyclines sont
surtout utilisées dans le traitement des parodontites agressives causées par A.
actinomycetemcomitans [500].

104
Dans un contexte extra-buccal, différentes études reconnaissent maintenant les effets antibactériens
du thé vert et ont repoussé les limites de ce concept par la réalisation d’études ayant pour but de
vérifier si les catéchines du thé vert pouvaient agir en synergie avec des agents antimicrobiens connus.
Plusieurs de ces recherches portent sur des bactéries pathogènes telles que Staphylococcus aureus
résistant à la méthycilline (SARM), Staphylococcus epidermidis et Escherichia coli, entre autres. Les
résultats de ces études ont démontré que les catéchines du thé vert agissent en synergie avec la
tétracycline contre S. aureus et S. epidermidis [486, 501], avec la pénicilline contre S. epidermidis
[502], avec la pénicilline, l’oxacilline, l’ampicilline/le sulbactam, le carbapenem et l’imipenem contre
SARM [503-506] et finalement, avec le chloramphénicol, la ciprofloxacine, et la cefotaxime contre la
bactérie pathogène E. coli présentant divers degrés de résistance [501, 507].
5.2 Propriétés anti-destruction tissulaire
Les maladies parodontales sont des maladies infectieuses conduisant à la destruction des tissus de
support de la dent. D’une part, les maladies parodontales sont initiées par la présence d’un sous-
groupe particulier de bactéries Gram négatives incluant entre autre P. gingivalis, F. nucleatum et A.
actinomycetemcomitans [94], colonisant les sites sous-gingivaux sous la forme d'un biofilm. Ceci
constitue le facteur étiologique primaire de cette affection. D’autre part, la réponse de l’hôte à ces
bactéries et à leurs métabolites toxiques constitue également un facteur clé modulant la progression
et la sévérité de cette condition pathologique. Plus précisément, le LPS des bactéries Gram-négatives
est un puissant inducteur de médiateurs pro-inflammatoires et peut ainsi initier de nombreux processus
destructeurs médiés par l’hôte [508]. Les fibroblastes, cellules constitutives des tissus parodontaux, de
même que les macrophages, cellules prédominantes dans les lésions parodontales actives [292], en
réponse aux bactéries parodontopathogènes, produisent et sécrètent plusieurs cytokines et
médiateurs de l’inflammation et de la réponse immunitaire, plus spécifiquement l’IL-1, le TNF-α, l’IL-6,
ainsi que l’IL-8 [509]. Ces cytokines jouent un rôle direct dans la destruction parodontale, puisqu’elles
stimulent les fibroblastes et les macrophages, entre autre à augmenter la production, l’expression et
l'activation des MMPs, entraînant la destruction progressive du parodonte [443]. Les MMPs, dont la
stromélysine MMP-3, la collagénase MMP-8 et la gélatinase MMP-9, dégradent le collagène et la
gélatine constituant la matrice extracellulaire des tissus parodontaux. Par conséquent, l'activité des
MMPs joue un rôle important dans la pathogenèse et la progression de la maladie parodontale.

105
D'ailleurs, leur dosage dans le fluide créviculaire est reconnu pour refléter l’état de santé du parodonte
[300, 320, 321, 323, 510]. Depuis une dizaine d'années, les composés bioactifs dotés d'une capacité
de moduler la réponse inflammatoire de l'hôte reçoivent une attention considérable, car ces derniers
pourraient représenter de nouveaux agents thérapeutiques pour le traitement des maladies
parodontales [270, 291].
Dans notre étude, un modèle de co-culture macrophages/fibroblastes a été utilisé afin d'évaluer la
capacité d’un extrait de thé vert et d’EGCG à diminuer la sécrétion des MMPs, plus spécifiquement la
MMP-3, la MMP-8 et la MMP-9, enzymes hydrolytiques grandement impliquées dans la destruction
tissulaire caractéristique de la parodontite. Les effets des différents composés à l’étude ont été évalués
lorsque le modèle de co-culture a été stimulé ou non, avec le LPS d’A. actinomycetemcomitans en
situation mimant une condition faiblement inflammatoire (ratio 1 :10) et une condition fortement
inflammatoire (ratio 10 :1). Bien que les deux ratios cellulaires utilisés au cours de cette étude aient
été déterminés de façon arbitraire à prime abord, les résultats obtenus supportent que le ratio 10:1
puisse représenter une condition fortement inflammatoire par rapport au ratio 1:10. En effet, des
concentrations significativement plus élevées pour les MMP-3,-8 et -9 sont obtenus pour le ratio 10 :1,
lorsque ce dernier est comparé à la concentration dosée lors d’une situation faiblement inflammatoire,
représentée par le ratio 1 :10.
Avant de vérifier le potentiel de l'extrait de thé vert et de l'EGCG à des fins préventives et
thérapeutiques, leurs effets sur la sécrétion basale de MMPs par le modèle de co-culture
macrophages/fibroblastes ont préalablement été déterminés. Les résultats de cette analyse ont permis
de mettre en lumière la capacité des polyphénols du thé vert à réduire les niveaux basaux des MMPs
sécrétées, plus spécifiquement les MMP-3 et MMP-9. Puisque dans un contexte sain, les MMPs
demeurent importantes dans le remodelage tissulaire, l’utilisation d’une concentration de 62,5 µg/ml
pourrait être à privilégier puisqu'elles n'amène pas une réduction significative de leur sécrétion.

106
D’une part, l’intérêt de l’utilisation des composés naturels à l’étude à des fins préventives a été évalué
dans le modèle de co-culture macrophages/fibroblastes. Les résultats de cet objectif ont permis de
mettre en lumière la capacité des polyphénols du thé vert à prévenir la sécrétion des MMPs (MMP-3,
MMP-8 et MMP-9) par le modèle de co-culture préalablement traité avant d’être stimulé avec le LPS.
D’autre part, le potentiel d'utilisation des composés naturels à l’étude à des fins thérapeutiques a
également été évalué. Les résultats de cet objectif ont démontré la capacité des polyphénols du thé
vert à réduire la sécrétion des MMPs, plus spécifiquement les MMP-3 et -9 dans le modèle de co-
culture préalablement stimulé avec le LPS. L'ensemble des résultats obtenus supportent le potentiel
d'utilisation à titre d’agents préventifs ou thérapeutiques de la maladie parodontale.
Dans des situations reflétant un état faiblement (ratio 1 macrophage : 10 fibroblastes) ou fortement
inflammatoires (ratio 10 macrophages : 1 fibroblaste), un prétraitement avec l’extrait de thé vert a réduit
significativement la sécrétion de la MMP-3 par le modèle de co-culture stimulé avec le LPS, à partir
d’une concentration de 15,625 µg/ml. Les effets inhibiteurs de l’EGCG ont ensuite été vérifiés selon
les mêmes conditions, et ces derniers se sont avérés moins efficaces, ce qui laisse supposer la
présence d’autres molécules bioactives dans l’extrait de thé vert. Pour les deux ratios cellulaires testés,
l’EGCG, a nécessité une concentration d'au moins 31,25 µg/ml pour entraîner une diminution
significative de la sécrétion de la MMP-3.
Dans un contexte thérapeutique de la maladie parodontale, l’extrait de thé vert pourrait représenter
une substance intéressante puisque ce dernier a entraîné une réduction significative de la sécrétion
de la MMP-3, à partir d’une concentration 7,8125 µg/ml pour les deux ratios cellulaires testés. Les
effets inhibiteurs de l’EGCG ont ensuite été vérifiés selon les mêmes conditions. Dans une situation
faiblement inflammatoire, l’EGCG a diminué de façon significative et dose-dépendante la sécrétion de
la MMP-3 à partir d’une concentration 7,8125 µg/ml. En ce qui concerne le ratio représentant une
situation fortement inflammatoire, l’EGCG inhibait, de façon significative et dose dépendante la
sécrétion de la MMP-3 par le modèle de co-culture à partir de 15,625 µg/ml.

107
Dans une alternative préventive (traitement du modèle de co-culture avec les polyphénols avant la
stimulation avec le LPS), il a été démontré qu’en situation faiblement inflammatoire, l’extrait de thé vert
et l’EGCG diminuent significativement la sécrétion de la MMP-8 à partir de 31,25 µg/ml et 15,625
µg/ml, respectivement. Lors de l’utilisation d’un ratio illustrant une situation fortement inflammatoire,
une diminution dose-dépendante quoique seulement significative à des concentrations de 15,625
µg/ml et 62,5 µg/ml a été observée pour l’extrait de thé vert, et 15,625 µg/ml et 31,25 µg/ml pour
l’EGCG.
Dans une optique de traitement de la maladie parodontale (stimulation du modèle de co-culture avec
le LPS avant l'ajout des polyphénols), il a été démontré qu’en situation faiblement inflammatoire,
l’extrait de thé vert et l’EGCG n’influencent pas significativement la sécrétion de la MMP-8. Lors de
l’utilisation d’un ratio illustrant une situation fortement inflammatoire, une diminution dose-dépendante,
quoique seulement significative à une concentration de 31,25 µg/ml, a été observée pour l’extrait de
thé vert, et à partir de 15,625 µg/ml pour l’EGCG.
Dans les deux ratios cellulaires qui ont été utilisés, illustrant un état faiblement ou fortement
inflammatoire, un prétraitement avec l’extrait de thé vert ou l’EGCG a été en mesure de prévenir la
sécrétion de la MMP-9 par le modèle de co-culture stimulé avec le LPS. Pour l’extrait de thé vert, la
diminution s’est avérée significative à partir de 31,25 µg/ml pour le ratio 1 :10, tandis que cette
diminution s’est avérée significative à partir de 15,625 µg/ml en ce qui concerne le ratio 10 :1. Pour
l’EGCG, la diminution s’est avérée significative à partir d’une concentration de 15,625 µg/ml pour le
ratio 1 :10, tandis que cette diminution a été significative à partir de 31,25 µg/ml en ce qui concerne le
ratio 10 :1.
Dans une alternative thérapeutique, l'ajout de l’extrait de thé vert au modèle de co-culture pré-stimulé
avec le LPS a permis, à une concentration de 7,8125 µg/ml, de diminuer significativement la sécrétion
de la MMP-dans la condition faiblement inflammatoire (ratio 1 :10). Cette diminution s’est avérée
significative à partir de 31,25 µg/ml en ce qui concerne le ratio 10 :1. Pour l’EGCG, 7,8125 µg/ml ont

108
été nécessaire afin de réduire significativement la sécrétion de la MMP-9 pour le ratio 1 :10, tandis que
cette diminution a été significative à partir de 15,625 µg/ml en ce qui concerne le ratio 10 :1.
L’inhibition de la sécrétion des MMPs dans un contexte préventif et thérapeutique des affections
parodontales constitue un aspect qui a été peu étudiée jusqu’à présent. Tel qu’il a été mentionné
précédemment, une tasse de thé contient jusqu’à 450 µg/ml d'EGCG, ce qui supporte que l’inhibition
des MMPs dans une situation in vivo pourrait être réaliste. Les études portant sur l’EGCG, plus
spécifiquement sur sa capacité à inhiber la sécrétion des MMPs, ont utilisés d’autres types cellulaires,
tels que les cellules cancéreuses (gastriques, etc.) ainsi que les ostéoblastes [511-514]. Il a été
démontré que l'EGCG contrôle l’expression des MMPs chez ces types cellulaires par la suppression
de l’activation des MAPK ainsi que de la protéine activatrice 1 (AP-1) .
Yun et coll. ont démontré que l'EGCG inhibe la formation d'ostéoclastes dans un modèle de co-culture
impliquant des cellules ostéoblastiques et des cellules primaires de la moelle osseuse, et induit la mort
cellulaire par apoptose d’ostéoclastes multinucléés de manière dose-dépendante [466, 468].
Récemment, il a été démontré que l'EGCG inhibe l'activité des MMPs en inhibant la collagénase MMP-
2 et la gélatinase MMP-9 par le mécanisme mentionné ci-haut chez des ostéoblastes [466]. Plusieurs
études ont fourni des preuves suggérant que l'EGCG inhibe la voie des MAPK, résultant dans la
régulation négative des MMPs.
En plus des catéchines du thé vert, le potentiel anti-destruction tissulaire associé à d'autres
polyphénols a déjà été démontré. Il a été rapporté que les théaflavines, composantes présentes
majoritairement dans le thé noir, ont la capacité d’inhiber la sécrétion et l’expression (ARNm) des MMP-
1 et -2 chez les fibroblastes gingivaux humains lors de leur stimulation par P. gingivalis [515]. Bodet et
coll. ont démontré qu'une fraction de canneberge enrichie en proanthocyanidines inhibe la sécrétion
de MMP-3 et MMP-9 par les fibroblastes gingivaux stimulés avec le LPS d’A. actinomycetemcomitans
[516, 517]. Cette fraction de la canneberge semblait exercer son pouvoir via une inhibition de la
cascade de phosphorylation (protéines de signalisation cellulaire) et une réduction de l’expression de

109
la protéine activatrice chez les fibroblastes (AP-1), impliquée dans la régulation transcriptionnelle de
nombreux médiateurs pro-inflammatoires et de MMPs [270, 516, 518]. Les proanthocyanidines de type
A de la canneberge sont également reconnues pour inhiber la sécrétion des MMPs par les
macrophages stimulés avec le LPS d’A. actinomycetemcomitans, ainsi que l’activité catalytique des
MMP-1 et MMP-9 [518].
Une étude récente réalisée par notre équipe a également mis en lumière la capacité d’un extrait de
bleuet sauvage à réduire la sécrétion des MMP-8 et MMP-9 par les macrophages stimulés par la
bactérie F. nucleatum [519, 520]. Il a été démontré qu'en plus d’inhiber la production de ces MMPs,
l’extrait de bleuet inhibe l’activité catalytique de la MMP-9 (gélatinase-2). Par conséquent, il a été
suggéré que le bleuet pourrait contribuer à réduire les dommages tissulaires caractéristiques de la
parodontite [519].
Il a également été démontré que l'auraptène et le lacinartin, des polyphénols appartenant à la classe
des coumarins, inhibent de manière dose-dépendante la sécrétion de la MMP-8 et MMP-9 chez les
macrophages humains de la lignée U937 stimulés avec le LPS d’A. actinomycetemcomitans [521]. De
plus, la licoricidine ainsi que le licorisoflavan A, des isoflavonoïdes dérivés de la réglisse, inhibent la
sécrétion des MMP-7, -8 et -9 par les macrophages stimulés avec le LPS d’A. actinomycetemcomitans
en diminuant l'activation de NF-kB p65 [522].
Tout comme il a été observé dans cette étude pour le thé vert, il a récemment été démontré que la
myricétine peut agir sur les deux composantes étiologiques impliquées dans le développement de la
parodontite. En plus de posséder des propriétés antibactériennes, la myricétine prévient, de manière
dose-dépendante, l’activation du facteur de transcription NF-κB chez les monocytes. De plus, la
myricétine inhibe la sécrétion d'IL-6, d’IL-8 ainsi que de MMP-3 par les fibroblastes stimulés avec la
bactérie parodontopathogène P. gingivalis [523].

110

111
6. CONCLUSION
En conclusion, cette étude a apporté des évidences supportant le potentiel des polyphénols du thé vert
et de sa composante principale, l’EGCG, en vue d’une utilisation préventive et thérapeutique pour le
contrôle des maladies parodontales. En effet, les résultats obtenus ont démontré les propriétés
antibactériennes de ces composantes via une inhibition de la croissance de trois importantes espèces
parodontopathogènes (P. gingivalis, F. nucleatum et A. actinomycetemcomitans). De plus, l’extrait de
thé vert ainsi que l’EGCG ont démontré, par une utilisation de la méthode de microdilution en damier,
des effets synergiques ou additifs lorsqu’utilisés en association avec le métronidazole ou la
tétracycline, des antibiotiques couramment utilisés en thérapie parodontale. Puisque ces bactéries sont
considérées comme des agents étiologiques majeurs des parodontites chroniques et agressives, leur
élimination s'avère critique en vue d'un traitement efficace de cette affection. De plus, un modèle de
co-culture constitué de fibroblastes gingivaux intégrés dans un gel de collagène et recouvert de
macrophages a été utilisé pour évaluer la capacité des polyphénols du thé vert à inhiber la sécrétion
de MMPs, notamment les MMP-3, MMP-8 et MMP-9, qui sont bien connus pour participer à la
destruction tissulaire et osseuse lors de la parodontite. L'extrait de thé vert et l’EGCG ont inhibé
significativement et de manière dose-dépendante la sécrétion des MMPs par le modèle de co-culture,
plus spécifiquement la MMP-3 et la MMP-9.
Les propriétés antibactériennes, de même les propriétés immuno-modulatrices de ces composés
naturels en font une alternative intéressante à la thérapie conventionnelle, tout en permettant de
diminuer considérablement le risque à l’apparition de résistance aux antibiotiques conventionnels chez
les microorganismes. Les résultats obtenus dans ce projet ouvrent la voie de la recherche clinique afin
de vérifier le réel potentiel des polyphénols du thé vert pour une utilisation à long terme dans la thérapie
des affections parodontales. En effet, ces derniers pourraient être employés à des fins thérapeutiques
en application topique sous forme de gel ou de fibre au niveau des sites sous-gingivaux atteints ou
simplement être utilisés à titre d'additifs à l'intérieur de produits d’hygiène buccodentaire (dentifrices,
rince-bouches ou gommes à mâcher).
En perspective à ce projet de recherche, bien que les propriétés antibactériennes et le potentiel anti-
destruction de l’extrait de thé vert et de l’EGCG aient été mises en évidence au cours de cette étude,

112
il n’en demeure pas moins que beaucoup reste à étudier. En premier lieu, dans une optique
thérapeutique de la maladie parodontale, une évaluation du temps minimal nécessaire pour causer la
mort de l’inoculum bactérien pourrait s’avérer justifiée. De plus, bien que P. gingivalis, F. nucleatum et
A. actinomycetemcomitans sont des bactéries parodontopathogènes importantes dans l’initiation et la
progression de la maladie parodontale, ces dernières n’agissent pas seules. Par conséquent, une
analyse de l’effet des polyphénols du thé vert sur d’autres bactéries parodontopathogènes utilisées en
association pourrait s’avérer intéressant afin de bien comprendre les différentes interactions
bactériennes in vivo. Il pourrait également s’avérer pertinent d’étudier le potentiel anti-inflammatoire
des polyphénols à l’étude, tant sur l’inhibition de la sécrétion de cytokines pro-inflammatoires que
l’augmentation de la sécrétion de cytokines anti-inflammatoires. Finalement, une évaluation des effets
d’autres classes de polyphénols du thé vert pourrait être intéressante dans la mesure où les propriétés
bioactives de l’extrait de thé vert ne soient pas exclusivement attribuables à l’EGCG.

113
7. PERTINENCE DU PROJET
Les maladies parodontales représentent un problème de santé important et leur développement est la
conséquence de facteurs microbiologiques, immuno-inflammatoires et environnementaux. En plus de
causer des dommages au niveau buccal, la parodontite peut entraîner des répercussions systémiques
telles que le diabète, les cardiopathies, les affections pulmonaires ainsi que des naissances
prématurées de bébés de faible poids, d’où l’importance de les traiter et de les prévenir.
Considérant que les maladies parodontales touchent une proportion importante de la population et
qu’elles ont un impact considérable sur la qualité de vie des individus affectés au quotidien, la
réalisation des différents objectifs de cette étude a permis d’élargir les connaissances actuelles quant
au potentiel de certains polyphénols pour la santé buccodentaire. Ayant déjà démontré des effets
bénéfiques au niveau systémique, les polyphénols tels que ceux contenus dans le thé vert (EGCG)
pourraient agir à titre préventif et thérapeutique en élargissant les options de traitements pour la
maladie parodontale.
En raison de leur contenu complexe, l’utilisation d’extraits de plantes permet d’agir sur plus d’une cible
thérapeutique, offrant une alternative intéressante aux traitements traditionnels en visant à la fois la
composante microbiologique et la réponse inflammatoire destructrice de l’hôte, tout en
empêchant/diminuant l’apparition de résistances chez les pathogènes.

114

115
8. BIBLIOGRAPHIE
1. Dorland, W.A.N., Dorland's illustrated medical dictionary. 1957, Saunders: Philadelphia. 2. Hassell, T.M., Tissues and cells of the periodontium. Periodontol 2000, 1993. 3: p. 9-38. 3. Listgarten, M.A., Dental plaque: its structure and prevention. ASDC J Dent Child, 1972. 39(5):
p. 347-52. 4. Lindhe, J., N.P. Lang, and T. Karring, Clinical periodontology and implant dentistry. 2008,
Blackwell Munksgaard: Oxford, UK ; Ames, Iowa. 5. Wolf, H.F., E.M. Rateitschak, and K.H. Rateitschak, Parodontologie. 3e éd. entièrement
remaniée et complétée. ed. 2005, Issy-les-Moulineaux: Masson. xi, 532 p. 6. Gartner, L.P. and J.L. Hiatt, Color atlas of histology. 5th ed. 2009, Philadelphia: Wolters
Kluwer/Lippincott William & Wilkins. xv, 459 p. 7. Charon, J.A. and M.-È. Bezzina-Moulierac, Parodontie médicale : innovations cliniques. 2e éd.
ed. Collection JPIO,. 2010, Rueil-Malmaison: CdP :. xix, 471 p. 8. Muller-Glauser, W. and H.E. Schroeder, Exfoliative cytology and ultrastructure of superficial
epithelial cells from the normal human oral mucosa. J Biol Buccale, 1983. 11(4): p. 317-26. 9. Schroeder, H.E. and J. Theilade, Electron microscopy of normal human gingival epithelium. J
Periodontal Res, 1966. 1(2): p. 95-119. 10. Skougaard, M., Turnover of the gingival epithelium in marmosets. Acta Odontol Scand, 1965.
23(6): p. 623-43. 11. Schroeder, H.E., Melanin containing organelles in cells of the human gingiva. 3. Connective
tissue cells. Helv Odontol Acta, 1969. 13(1): p. 46-55. 12. Newcomb, G.M., G.J. Seymour, and R.N. Powell, Association between plaque accumulation
and Langerhans cell numbers in the oral epithelium of attached gingiva. J Clin Periodontol, 1982. 9(4): p. 297-304.
13. Schroeder, H.E. and M.A. Listgarten, The gingival tissues: the architecture of periodontal protection. Periodontol 2000, 1997. 13: p. 91-120.
14. Karring, T. and H. Loe, The three-dimensional concept of the epithelium-connective tissue boundary of gingiva. Acta Odontol Scand, 1970. 28(6): p. 917-33.
15. Mattout, P. and C. Mattout, Les thérapeutiques parodontales et implantaires. 2003, Paris: Quintessence International. 496 p.
16. Winning, T.A. and G.C. Townsend, Oral mucosal embryology and histology. Clin Dermatol, 2000. 18(5): p. 499-511.
17. Reynolds, J.J., R.M. Hembry, and M.C. Meikle, Connective tissue degradation in health and periodontal disease and the roles of matrix metalloproteinases and their natural inhibitors. Adv Dent Res, 1994. 8(2): p. 312-9.
18. Dale, B.A., Periodontal epithelium: a newly recognized role in health and disease. Periodontol 2000, 2002. 30: p. 70-8.
19. Caffesse, R.G., et al., The effect of mechanical stimulation on the keratinization of sulcular epithelium. J Periodontol, 1982. 53(2): p. 89-92.
20. Fry, H.R. and G.R. App, Histologic evaluation of the effects of intrasulcular toothbrushing on human sulcular epithelium. J Periodontol, 1978. 49(4): p. 163-73.
21. Bercy, P. and H. Tenenbaum, Parodontologie : du diagnostic à la pratique. 1996, Paris ; Bruxelles: De Boeck Université. 289 p.
22. Schroeder, H.E. and M.A. Listgarten, Fine structure of the developing epithelial attachment of human teeth. Monogr Dev Biol, 1971. 2: p. 1-134.

116
23. Frank, D.E. and W.G. Carter, Laminin 5 deposition regulates keratinocyte polarization and persistent migration. J Cell Sci, 2004. 117(Pt 8): p. 1351-63.
24. Nanci, A. and D.D. Bosshardt, Structure of periodontal tissues in health and disease. Periodontol 2000, 2006. 40: p. 11-28.
25. Brill, N. and B.O. Krasse, The passage of tissue fluid into the clinically healthy gingival pocket. Acta Odontol Scand, 1958. 16(3): p. 233-245.
26. Page, R.C. and H.E. Schroeder, Pathogenesis of inflammatory periodontal disease. A summary of current work. Lab Invest, 1976. 34(3): p. 235-49.
27. Schroeder, H.E., S. Munzel-Pedrazzoli, and R. Page, Correlated morphometric and biochemical analysis of gingival tissue in early chronic gingivitis in man. Arch Oral Biol, 1973. 18(7): p. 899-923.
28. Arnim, S.S. and D.A. Hagerman, The connective tissue fibers of the marginal gingiva. J Am Dent Assoc, 1953. 47(3): p. 271-81.
29. Page, R.C. and H.E. Schroeder, Periodontitis in man and other animals : a comparative review. 1982, Basel: Karger. viii, 330.
30. Narayanan, A.S. and R.C. Page, Connective tissues of the periodontium: a summary of current work. Coll Relat Res, 1983. 3(1): p. 33-64.
31. Roodman, G.D., Osteoclast differentiation. Crit Rev Oral Biol Med, 1991. 2(3): p. 389-409. 32. Birkedal-Hansen, H., et al., Synthesis and release of procollagenase by cultured fibroblasts. J
Biol Chem, 1976. 251(10): p. 3162-8. 33. Birkedal-Hansen, H., et al., Cleavage of bovine skin type III collagen by proteolytic enzymes.
Relative resistance of the fibrillar form. J Biol Chem, 1985. 260(30): p. 16411-7. 34. Birkedal-Hansen, H., From tadpole collagenase to a family of matrix metalloproteinases. J Oral
Pathol, 1988. 17(9-10): p. 445-51. 35. Pettigrew, D.W., et al., Effect of oxygen tension and indomethacin on production of collagenase
and neutral proteinase enzymes and their latent forms by porcine gingival explants in culture. Arch Oral Biol, 1978. 23(9): p. 767-77.
36. Birkedal-Hansen, H., Proteolytic remodeling of extracellular matrix. Curr Opin Cell Biol, 1995. 7(5): p. 728-35.
37. Mariotti, A., The extracellular matrix of the periodontium: dynamic and interactive tissues. Periodontol 2000, 1993. 3: p. 39-63.
38. Listgarten, M.A., J. Lindhe, and L. Hellden, Effect of tetracycline and/or scaling on human periodontal disease. Clinical, microbiological, and histological observations. J Clin Periodontol, 1978. 5(4): p. 246-71.
39. Charon, J.A. and C. Mouton, Parodontie médicale. Collection JPIO. 2003, Paris: Editions CdP. xix, 434 p.
40. Bosshardt, D.D. and K.A. Selvig, Dental cementum: the dynamic tissue covering of the root. Periodontol 2000, 1997. 13(1): p. 41-75.
41. Schroeder, H.E., The periodontium. Handbook of microscopic anatomy ; v. V, no. 5. 1986, Berlin ; New York: Springer-Verlag. vi, 418.
42. Cho, M.I. and P.R. Garant, Development and general structure of the periodontium. Periodontol 2000, 2000. 24: p. 9-27.
43. Doucet, P. and M. Lowenstein, Osteoclasts activation by bacterial endotoxins during periodontal diseases. Med Sci (Paris), 2006. 22(6-7): p. 614-20.
44. Tanabe, S. and D. Grenier, Characterization of volatile sulfur compound production by Solobacterium moorei. Arch Oral Biol, 2012. 57(12): p. 1639-43.

117
45. Aas, J.A., et al., Defining the normal bacterial flora of the oral cavity. J Clin Microbiol, 2005. 43(11): p. 5721-32.
46. Al-Ahmad, A., et al., Change in diet and oral hygiene over an 8-week period: effects on oral health and oral biofilm. Clin Oral Investig, 2010. 14(4): p. 391-6.
47. Bhatia, R., Ichhpujani, RL., Microbiology for Dental Students. 3rd edition, 2003. New Delhi : Jaypee Brothers Publishers, 346 p.
48. Bagg, J., Essentials of microbiology for dental students. 2006, Oxford University Press: Oxford ; New York. 317 p.
49. Piette, E. and M. Goldberg, La dent normale et pathologique. 2001, Bruxelles: De Boeck Université. 392 p.
50. Feng, Z. and A. Weinberg, Role of bacteria in health and disease of periodontal tissues. Periodontol 2000, 2006. 40: p. 50-76.
51. Slots, J., Microflora in the healthy gingival sulcus in man. Scand J Dent Res, 1977. 85(4): p. 247-54.
52. Marsh, P.D., Dental plaque as a microbial biofilm. Caries Res, 2004. 38(3): p. 204-11. 53. Burt, B., Position paper: epidemiology of periodontal diseases. J Periodontol, 2005. 76(8): p.
1406-19. 54. Pizzo, G., et al., Dentistry and internal medicine: from the focal infection theory to the
periodontal medicine concept. Eur J Intern Med, 2010. 21(6): p. 496-502. 55. Armitage, G.C., Clinical evaluation of periodontal diseases. Periodontol 2000, 1995. 7: p. 39-
53. 56. Mealey, B.L., Influence of periodontal infections on systemic health. Periodontol 2000, 1999.
21: p. 197-209. 57. Scannapieco, F.A., Position paper of The American Academy of Periodontology: periodontal
disease as a potential risk factor for systemic diseases. J Periodontol, 1998. 69(7): p. 841-50. 58. Offenbacher, S., Periodontal diseases: pathogenesis. Ann Periodontol, 1996. 1(1): p. 821-78. 59. Gendron, R., D. Grenier, and L.-F. Maheu-Robert, The oral cavity as a reservoir of bacterial
pathogens for focal infections. Microb Infect, 2000. 2(8): p. 897-906. 60. Persac, S., et al., An update on focal infection of oral origin. Rev Stomatol Chir Maxillofac,
2011. 112(6): p. 353-9. 61. Thoden van Velzen, S.K., L. Abraham-Inpijn, and W.R. Moorer, Plaque and systemic disease:
a reappraisal of the focal infection concept. J Clin Periodontol, 1984. 11(4): p. 209-20. 62. Miller, W.D., The human mouth as a focus of infection. The Lancet. 138(3546): p. 340-2. 63. Wilson, W., et al., Prevention of infective endocarditis: guidelines from the American Heart
Association: a guideline from the American Heart Association Rheumatic Fever, Endocarditis and Kawasaki Disease Committee, Council on Cardiovascular Disease in the Young, and the Council on Clinical Cardiology, Council on Cardiovascular Surgery and Anesthesia, and the Quality of Care and Outcomes Research Interdisciplinary Working Group. J Am Dent Assoc, 2007. 138(6): p. 739-45, 747-60.
64. Baumgartner, J.C., J.P. Heggers, and J.W. Harrison, Incidence of bacteremias related to endodontic procedures. II. Surgical endodontics. J Endod, 1977. 3(10): p. 399-402.
65. Lofthus, J.E., et al., Bacteremia following subgingival irrigation and scaling and root planing. J Periodontol, 1991. 62(10): p. 602-7.
66. Debelian, G.J., I. Olsen, and L. Tronstad, Systemic diseases caused by oral microorganisms. Endod Dent Traumatol, 1994. 10(2): p. 57-65.
67. Guntheroth, W.G., How important are dental procedures as a cause of infective endocarditis? American Journal of Cardiology. 54(7): p. 797-801.

118
68. Sconyers, J.R., J.J. Crawford, and J.D. Moriarty, Relationship of bacteremia to toothbrushing in patients with periodontitis. J Am Dent Assoc. 87(3): p. 616-622.
69. Wahl, M.J., Myths of dental-induced endocarditis. Arch Interl Med, 1994. 154(2): p. 137-144. 70. Barco, C.T., Prevention of infective endocarditis: A review of the medical and dental literature.
J Periodontol, 1991. 62(8): p. 510-23. 71. Roberts, G.J., Dentists are innocent! "Everyday" bacteremia is the real culprit: a review and
assessment of the evidence that dental surgical procedures are a principal cause of bacterial endocarditis in children. Pediatr Cardiol, 1999. 20(5): p. 317-25.
72. Li, X., et al., Systemic diseases caused by oral infection. Clin Microbiol Rev, 2000. 13(4): p. 547-58.
73. Ferretti, J.J., C. Shea, and M.W. Humphrey, Cross-reactivity of Streptococcus mutans antigens and human heart tissue. Infect Immun, 1980. 30(1): p. 69-73.
74. DeStefano, F., et al., Dental disease and risk of coronary heart disease and mortality. BMJ, 1993. 306(6879): p. 688-91.
75. Mattila, K.J., et al., Association between dental health and acute myocardial infarction. BMJ : BMJ, 1989. 298(6676): p. 779-81.
76. Scannapieco, F.A., G.D. Papandonatos, and R.G. Dunford, Associations Between Oral Conditions and Respiratory Disease in a National Sample Survey Population. Ann Periodontol, 1998. 3(1): p. 251-6.
77. Kuo, L.-C., A.M. Polson, and T. Kang, Associations between periodontal diseases and systemic diseases: A review of the inter-relationships and interactions with diabetes, respiratory diseases, cardiovascular diseases and osteoporosis. Public Health. 122(4): p. 417-33.
78. Offenbacher, S., et al., Periodontal Infection as a Possible Risk Factor for Preterm Low Birth Weight. J Periodontol, 1996. 67(10s): p. 1103-113.
79. Hill, G.B., Preterm Birth: Associations With Genital and Possibly Oral Microflora. Ann Periodontol, 1998. 3(1): p. 222-32.
80. Michaud, D.S., et al., A prospective study of periodontal disease and pancreatic cancer in US male health professionals. J Natl Cancer Inst, 2007. 99(2): p. 171-5.
81. Li, P., et al., Relationship of metabolic syndrome to chronic periodontitis. J Periodontol, 2009. 80(4): p. 541-9.
82. Craig, R.G., Interactions between chronic renal disease and periodontal disease. Oral Dis, 2008. 14(1): p. 1-7.
83. Bartold, P.M., R.I. Marshall, and D.R. Haynes, Periodontitis and rheumatoid arthritis: a review. J Periodontol, 2005. 76(11 Suppl): p. 2066-74.
84. Lens, J.W. and W. Beertsen, Injection of an antigen into the gingiva and its effect on an experimentally induced inflammation in the knee joint of the mouse. J Periodonl Res, 1988. 23(1): p. 1-6.
85. Tolo, K. and L. Jorkjend, Serum antibodies and loss of periodontal bone in patients with rheumatoid arthritis. J Clin Periodontol, 1990. 17(5): p. 288-91.
86. Kamer, A.R., et al., Alzheimer's disease and peripheral infections: the possible contribution from periodontal infections, model and hypothesis. J Alzheimers Dis, 2008. 13(4): p. 437-49.
87. Page, R.C., Oral health status in the United States: prevalence of inflammatory periodontal diseases. J Dent Educ, 1985. 49(6): p. 354-67.
88. Houle, M.A., Grenier D., Currents concepts in periodontal diseases. Médicine et Maladies Infectieuses, 2003. 33(7): p. 331-40.

119
89. Papapanou, P.N., Epidemiology of periodontal diseases: an update. J Int Acad Periodontol, 1999. 1(4): p. 110-6.
90. Armitage, G.C., Periodontal diagnoses and classification of periodontal diseases. Periodontol 2000, 2004. 34: p. 9-21.
91. Loe, H., E. Theilade, and S.B. Jensen, Experimental gingivitis in man. J Periodontol, 1965. 36: p. 177-87.
92. Listgarten, M.A., Pathogenesis of periodontitis. J Clin Periodontol, 1986. 13(5): p. 418-30. 93. Bodet, C., F. Chandad, and D. Grenier, Pathogenic potential of Porphyromonas gingivalis,
Treponema denticola and Tannerella forsythia, the red bacterial complex associated with periodontitis. Pathol Biol (Paris), 2007. 55(3-4): p. 154-62.
94. Haffajee, A.D. and S.S. Socransky, Microbial etiological agents of destructive periodontal diseases. Periodontol 2000, 1994. 5: p. 78-111.
95. Moore, W.E., et al., Variation in periodontal floras. Infect Immun, 1984. 46(3): p. 720-6. 96. Mandell, R.L., J.L. Ebersole, and S.S. Socransky, Clinical immunologic and microbiologic
features of active disease sites in juvenile periodontitis. J Clin Periodontol, 1987. 14(9): p. 534-40.
97. Evian, C.I., E.S. Rosenberg, and M.A. Listgarten, Bacterial variability within diseases periodontal sites. J Periodontol, 1982. 53(10): p. 595-8.
98. Feghali, K., et al., Cranberry Proanthocyanidins: Natural Weapons against Periodontal Diseases. J Agric Food Chem, 2012, 60 (23), pp 5728–35
99. Wilson, T.G. and K.S. Kornman, Fundamentals of periodontics. 1996, Chicago: Quintessence Pub. Co. xii, 564 p.
100. Goodson, J.M., et al., Patterns of progression and regression of advanced destructive periodontal disease. J Clin Periodontol, 1982. 9(6): p. 472-81.
101. Lindhe, J., A.D. Haffajee, and S.S. Socransky, Progression of periodontal disease in adult subjects in the absence of periodontal therapy. J Clin Periodontol, 1983. 10(4): p. 433-42.
102. Socransky, S.S., et al., New concepts of destructive periodontal disease. J Clin Periodontol, 1984. 11(1): p. 21-32.
103. Armitage, G.C., Classifying periodontal diseases--a long-standing dilemma. Periodontol 2000, 2002. 30: p. 9-23.
104. Highfield, J., Diagnosis and classification of periodontal disease. Aust Dent J, 2009. 54 Suppl 1: p. S11-26.
105. T. Duyninh, O.J., P. Bousquet, P. Gibert, V. Orti, Classification des maladies parodontales. EMC - Odontologie, March 2005. Volume 1( Issue 1): p. 58–66.
106. Slots, J. and M.A. Listgarten, Bacteroides gingivalis, Bacteroides intermedius and Actinobacillus actinomycetemcomitans in human periodontal diseases. J Clin Periodontol, 1988. 15(2): p. 85-93.
107. Darveau, R.P., A. Tanner, and R.C. Page, The microbial challenge in periodontitis. Periodontol 2000, 1997. 14: p. 12-32.
108. Socransky, S.S. and A.D. Haffajee, Dental biofilms: difficult therapeutic targets. Periodontol 2000, 2002. 28: p. 12-55.
109. Listgarten, M.A., The structure of dental plaque. Periodontol 2000, 1994. 5: p. 52-65. 110. Holt, S.C. and J.L. Ebersole, Porphyromonas gingivalis, Treponema denticola, and Tannerella
forsythia: the "red complex", a prototype polybacterial pathogenic consortium in periodontitis. Periodontol 2000, 2005. 38: p. 72-122.
111. Socransky, S.S., et al., Microbial complexes in subgingival plaque. J Clin Periodontol, 1998. 25(2): p. 134-44.

120
112. Marsh, P.D., Dental plaque: biological significance of a biofilm and community life-style. J Clin Periodontol, 2005. 32 Suppl 6: p. 7-15.
113. Holt, S.C., et al., Virulence factors of Porphyromonas gingivalis. Periodontol 2000, 1999. 20: p. 168-238.
114. Dumitrescu, A.L. and M. Kawamura, Involvement of psychosocial factors in the association of obesity with periodontitis. J Oral Sci, 2010. 52(1): p. 115-24.
115. Slots, J., Update on Actinobacillus Actinomycetemcomitans and Porphyromonas gingivalis in human periodontal disease. J Int Acad Periodontol, 1999. 1(4): p. 121-6.
116. Li, N. and C. Collyer, Gingipains from Porphyromonas gingivalis — complex domain structures confer diverse functions. Eur J Microbiol and Immunol, 2011. 1(1): p. 41-58.
117. Kusaba, A., et al., Cloning and expression of a Porphyromonas gingivalis gene for protoporphyrinogen oxidase by complementation of a hemG mutant of Escherichia coli. Oral Microbiol Immunol, 2002. 17(5): p. 290-5.
118. Roper, J.M., et al., The enigma of cobalamin (Vitamin B12) biosynthesis in Porphyromonas gingivalis. Identification and characterization of a functional corrin pathway. J Biol Chem, 2000. 275(51): p. 40316-23.
119. Smalley, J.W., et al., The HA2 haemagglutinin domain of the lysine-specific gingipain (Kgp) of Porphyromonas gingivalis promotes micro-oxo bishaem formation from monomeric iron(III) protoporphyrin IX. Microbiol, 2006. 152(Pt 6): p. 1839-45.
120. Hajishengallis, G., Immune evasion strategies of Porphyromonas gingivalis. J Oral Biosci, 2011. 53(3): p. 233-240.
121. Lamont, R.J. and H.F. Jenkinson, Life below the gum line: pathogenic mechanisms of Porphyromonas gingivalis. Microbiol Mol Biol Rev, 1998. 62(4): p. 1244-63.
122. Potempa, J., et al., Gingipains, the major cysteine proteinases and virulence factors of Porphyromonas gingivalis: structure, function and assembly of multidomain protein complexes. Curr Protein Pept Sci, 2003. 4(6): p. 397-407.
123. Mayrand, D. and S.C. Holt, Biology of asaccharolytic black-pigmented Bacteroides species. Microbiol Rev, 1988. 52(1): p. 134-52.
124. Pihlstrom, B.L., B.S. Michalowicz, and N.W. Johnson, Periodontal diseases. Lancet, 2005. 366(9499): p. 1809-20.
125. Kebschull, M., R.T. Demmer, and P.N. Papapanou, "Gum bug, leave my heart alone!"--epidemiologic and mechanistic evidence linking periodontal infections and atherosclerosis. J Dent Res, 2010. 89(9): p. 879-902.
126. Meurman, J.H., M. Sanz, and S.J. Janket, Oral health, atherosclerosis, and cardiovascular disease. Crit Rev Oral Biol Med, 2004. 15(6): p. 403-13.
127. Offenbacher, S., et al., Maternal periodontitis and prematurity. Part I: Obstetric outcome of prematurity and growth restriction. Ann Periodontol, 2001. 6(1): p. 164-74.
128. Scannapieco, F.A., Systemic effects of periodontal diseases. Dent Clin North Am, 2005. 49(3): p. 533-50.
129. Chun, Y.-H.P., et al., Biological foundation for periodontitis as a potential risk factor for atherosclerosis. J Periodont Res, 2005. 40(1): p. 87-95.
130. Demmer, R.T. and M. Desvarieux, Periodontal infections and cardiovascular disease: the heart of the matter. J Am Dent Assoc, 2006. 137 Suppl: p. 14S-20S.
131. Leishman, S.J., H.L. Do, and P.J. Ford, Cardiovascular disease and the role of oral bacteria. J Oral Microbiol, 2010. 2.
132. Iida, Y., et al., Brain abscess in which Porphyromonas gingivalis was detected in cerebrospinal fluid. British Journal of Oral and Maxillofacial Surgery, 2004. 42(2): p. 180.

121
133. van Winkelhoff, A.J. and J. Slots, Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis in nonoral infections. Periodontology 2000, 1999. 20(1): p. 122-35.
134. Kuby, J., et al., Immunologie: Le cours de Janis Kuby. 7th edition 2008: Paris :Dunod. 832 p. 135. Gottschalk, M. and S. Laurent-Lewandowski, [Vaccine development: strategies for coping with
the antigenic diversity of bacteria]. Rev Sci Tech, 2007. 26(1): p. 91-103. 136. Singh, A., et al., The capsule of Porphyromonas gingivalis leads to a reduction in the host
inflammatory response, evasion of phagocytosis, and increase in virulence. Infect Immun, 2011. 79(11): p. 4533-42.
137. Johannessen, M., et al., Bacterial interference with canonical NFkappaB signalling. Microbiol, 2013. 159(Pt 10): p. 2001-13.
138. Hornef, M.W., et al., Bacterial strategies for overcoming host innate and adaptive immune responses. Nat Immunol, 2002. 3(11): p. 1033-40.
139. Laine, M.L., B.J. Appelmelk, and A.J. van Winkelhoff, Prevalence and distribution of six capsular serotypes of Porphyromonas gingivalis in periodontitis patients. J Dent Res, 1997. 76(12): p. 1840-4.
140. Krinos, C.M., et al., Extensive surface diversity of a commensal microorganism by multiple DNA inversions. Nature, 2001. 414(6863): p. 555-8.
141. Tran, Q.T., et al., The Salmonella enterica serotype Typhi Vi capsular antigen is expressed after the bacterium enters the ileal mucosa. Infect Immun, 2010. 78(1): p. 527-35.
142. Brunner, J., et al., The capsule of Porphyromonas gingivalis reduces the immune response of human gingival fibroblasts. BMC Microbiol, 2010. 10: p. 5.
143. Grenier, D. and D. Mayrand, Selected characteristics of pathogenic and nonpathogenic strains of Bacteroides gingivalis. J Clin Microbiol, 1987. 25(4): p. 738-40.
144. Menard, C. and C. Mouton, Clonal diversity of the taxon Porphyromonas gingivalis assessed by random amplified polymorphic DNA fingerprinting. Infect Immun, 1995. 63(7): p. 2522-31.
145. Dufour, T. and J.M. Svoboda, Pathogénie bactérienne des parodontolyses. EMC-Odontologie, 2005. 1(1): p. 46-57.
146. Darveau, R.P., Periodontitis: a polymicrobial disruption of host homeostasis. Nat Rev Microbiol, 2010. 8(7): p. 481-90.
147. Nagano, K., et al., Porphyromonas gingivalis FimA fimbriae: fimbrial assembly by fimA alone in the fim gene cluster and differential antigenicity among fimA genotypes. PLoS One, 2012. 7(9): p. e43722.
148. Listgarten, M.A. and C.H. Lai, Comparative ultrastructure of Bacteroides melaninogenicus subspecies. J Periodontal Res, 1979. 14(4): p. 332-40.
149. Enersen, M., K. Nakano, and A. Amano, Porphyromonas gingivalis fimbriae. J Oral Microbiol, 2013. 5.
150. Okuda, K., J. Slots, and R. Genco, Bacteroides gingivalis, Bacteroides asaccharolyticus, andBacteroides melaninogenicus subspecies: Cell surface morphology and adherence to erythrocytes and human buccal epithelial cells. Curr Microbiol, 1981. 6(1): p. 7-12.
151. Handley, P.S. and L.S. Tipler, An electron microscope survey of the surface structures and hydrophobicity of oral and non-oral species of the bacterial genus Bacteroides. Arch Oral Biol, 1986. 31(5): p. 325-35.
152. Slots, J. and R.J. Gibbons, Attachment of Bacteroides melaninogenicus subsp. asaccharolyticus to oral surfaces and its possible role in colonization of the mouth and of periodontal pockets. Infect Immun, 1978. 19(1): p. 254-64.
153. Yoshimura, F., et al., Surface components of Porphyromonas gingivalis. J Periodontal Res, 2009. 44(1): p. 1-12.

122
154. Amano, A., et al., Variations of Porphyromonas gingivalis fimbriae in relation to microbial pathogenesis. J Periodontal Res, 2004. 39(2): p. 136-42.
155. Amano, A., Bacterial adhesins to host components in periodontitis. Periodontol 2000, 2010. 52(1): p. 12-37.
156. Fujiwara, T., et al., Molecular cloning and sequencing of the fimbrilin gene of Porphyromonas gingivalis strains and characterization of recombinant proteins. Biochem Biophys Res Commun, 1993. 197(1): p. 241-7.
157. Nelson, K.E., et al., Complete genome sequence of the oral pathogenic bacterium Porphyromonas gingivalis strain W83. J Bacteriol, 2003. 185(18): p. 5591-601.
158. Dickinson, D.P., et al., Molecular cloning and sequencing of the gene encoding the fimbrial subunit protein of Bacteroides gingivalis. J Bacteriol, 1988. 170(4): p. 1658-65.
159. Kuboniwa, M., H. Inaba, and A. Amano, Genotyping to distinguish microbial pathogenicity in periodontitis. Periodontol 2000, 2010. 54(1): p. 136-59.
160. Nakagawa, I., et al., Distribution and molecular characterization of Porphyromonas gingivalis carrying a new type of fimA gene. J Clin Microbiol, 2000. 38(5): p. 1909-14.
161. Nakagawa, I., et al., Identification of a new variant of fimA gene of Porphyromonas gingivalis and its distribution in adults and disabled populations with periodontitis. J Periodontal Res, 2002. 37(6): p. 425-32.
162. NM, O.B.-S., et al., Porphyromonas gingivalis gingipains: the molecular teeth of a microbial vampire. Curr Protein Pept Sci, 2003. 4(6): p. 409-26.
163. Whittaker, C.J., C.M. Klier, and P.E. Kolenbrander, Mechanisms of adhesion by oral bacteria. Annu Rev Microbiol, 1996. 50: p. 513-52.
164. Kuboniwa, M. and R.J. Lamont, Subgingival biofilm formation. Periodontol 2000, 2010. 52(1): p. 38-52.
165. Lamont, R.J. and O. Yilmaz, In or out: the invasiveness of oral bacteria. Periodontol 2000, 2002. 30: p. 61-9.
166. Hamada, S., et al., The importance of fimbriae in the virulence and ecology of some oral bacteria. Oral Microbiol Immunol, 1998. 13(3): p. 129-38.
167. Andrian, E., D. Grenier, and M. Rouabhia, Porphyromonas gingivalis-epithelial cell interactions in periodontitis. J Dent Res, 2006. 85(5): p. 392-403.
168. Amano, A., et al., Structural domains of Porphyromonas gingivalis recombinant fimbrillin that mediate binding to salivary proline-rich protein and statherin. Infect Immun, 1996. 64(5): p. 1631-7.
169. Song, H., et al., Hemagglutinin B is involved in the adherence of Porphyromonas gingivalis to human coronary artery endothelial cells. Infect Immun, 2005. 73(11): p. 7267-73.
170. Amano, A., Molecular interaction of Porphyromonas gingivalis with host cells: implication for the microbial pathogenesis of periodontal disease. J Periodontol, 2003. 74(1): p. 90-6.
171. Du, L., et al., Fimbriae and the hemagglutinating adhesin HA-Ag2 mediate adhesion of Porphyromonas gingivalis to epithelial cells. Infect Immun, 1997. 65(9): p. 3875-81.
172. Saito, T., et al., Cloning, expression, and sequencing of a protease gene from Bacteroides forsythus ATCC 43037 in Escherichia coli. Infect Immun, 1997. 65(11): p. 4888-91.
173. Grenier, D., Characteristics of hemolytic and hemagglutinating activities of Treponema denticola. Oral Microbiol Immunol, 1991. 6(4): p. 246-9.
174. Chu, L., et al., Hemolytic activity in the periodontopathogen Porphyromonas gingivalis: kinetics of enzyme release and localization. Infect Immun, 1991. 59(6): p. 1932-40.
175. Lamont, R.J., Oral microbiology and immunology. 2006, Washington, D.C.: ASM Press. xxiv, 458 p.

123
176. Potempa, J., A. Banbula, and J. Travis, Role of bacterial proteinases in matrix destruction and modulation of host responses. Periodontol 2000, 2000. 24: p. 153-92.
177. Imamura, T., The role of gingipains in the pathogenesis of periodontal disease. J Periodontol, 2003. 74(1): p. 111-8.
178. Kadowaki, T., et al., Porphyromonas gingivalis proteinases as virulence determinants in progression of periodontal diseases. J Biochem, 2000. 128(2): p. 153-59.
179. Curtis, M.A., et al., Molecular genetics and nomenclature of proteases of Porphyromonas gingivalis. J Periodontal Res, 1999. 34(8): p. 464-72.
180. Potempa, J., R. Pike, and J. Travis, The multiple forms of trypsin-like activity present in various strains of Porphyromonas gingivalis are due to the presence of either Arg-gingipain or Lys-gingipain. Infect Immun, 1995. 63(4): p. 1176-82.
181. Kadowaki, T., et al., Purification and characterization of a novel arginine-specific cysteine proteinase (argingipain) involved in the pathogenesis of periodontal disease from the culture supernatant of Porphyromonas gingivalis. J Biol Chem, 1994. 269(33): p. 21371-8.
182. Okamoto, K., et al., Cloning and sequencing of the gene encoding a novel lysine-specific cysteine proteinase (Lys-gingipain) in Porphyromonas gingivalis: structural relationship with the arginine-specific cysteine proteinase (Arg-gingipain). J Biochem, 1996. 120(2): p. 398-406.
183. Pavloff, N., et al., Molecular cloning and characterization of Porphyromonas gingivalis lysine-specific gingipain. A new member of an emerging family of pathogenic bacterial cysteine proteinases. J Biol Chem, 1997. 272(3): p. 1595-600.
184. Pavloff, N., et al., Molecular cloning and structural characterization of the Arg-gingipain proteinase of Porphyromonas gingivalis. Biosynthesis as a proteinase-adhesin polyprotein. J Biol Chem, 1995. 270(3): p. 1007-10.
185. Barkocy-Gallagher, G.A., et al., Analysis of the prtP gene encoding porphypain, a cysteine proteinase of Porphyromonas gingivalis. J Bacteriol, 1996. 178(10): p. 2734-41.
186. Okamoto, K., et al., Involvement of a lysine-specific cysteine proteinase in hemoglobin adsorption and heme accumulation by Porphyromonas gingivalis. J Bio Chem, 1998. 273(33): p. 21225-31.
187. Shah, H.N. and S.E. Gharbia, Ecological events in subgingival dental plaque with reference to Bacteroides and Fusobacterium species. Infection, 1989. 17(4): p. 264-8.
188. Holt, S.C. and T.E. Bramanti, Factors in virulence expression and their role in periodontal disease pathogenesis. Crit Rev Oral Biol Med, 1991. 2(2): p. 177-281.
189. Smalley, J.W., et al., The periodontopathogen Porphyromonas gingivalis binds iron protoporphyrin IX in the mu-oxo dimeric form: an oxidative buffer and possible pathogenic mechanism. Biochem J, 1998. 331 ( Pt 3): p. 681-5.
190. Barua, P.K., D.W. Dyer, and M.E. Neiders, Effect of iron limitation on Bacteroides gingivalis. Oral Microbiol Immunol, 1990. 5(5): p. 263-8.
191. Bramanti, T.E. and S.C. Holt, Roles of porphyrins and host iron transport proteins in regulation of growth of Porphyromonas gingivalis W50. J Bacteriol, 1991. 173(22): p. 7330-9.
192. Brochu, V., et al., Acquisition of iron from human transferrin by Porphyromonas gingivalis: a role for Arg- and Lys-gingipain activities. Oral Microbiol Immunol, 2001. 16(2): p. 79-87.
193. Griffen, A.L., et al., Prevalence of Porphyromonas gingivalis and periodontal health status. J Clin Microbiol, 1998. 36(11): p. 3239-42.
194. Travis, J., J. Potempa, and H. Maeda, Are bacterial proteinases pathogenic factors? Trends Microbiol, 1995. 3(10): p. 405-7.

124
195. Guo, Y., K.A. Nguyen, and J. Potempa, Dichotomy of gingipains action as virulence factors: from cleaving substrates with the precision of a surgeon's knife to a meat chopper-like brutal degradation of proteins. Periodontol 2000, 2010. 54(1): p. 15-44.
196. Gibbons, R.J., Bacterial adhesion to oral tissues: a model for infectious diseases. J Dent Res, 1989. 68(5): p. 750-60.
197. Gibbons, R.J., et al., Role of cryptic receptors (cryptitopes) in bacterial adhesion to oral surfaces. Arch Oral Biol, 1990. 35 Suppl: p. 107S-14S.
198. Lamont, R.J. and H.F. Jenkinson, Subgingival colonization by Porphyromonas gingivalis. Oral Microbiol Immunol, 2000. 15(6): p. 341-9.
199. Kontani, M., et al., Cysteine protease of Porphyromonas gingivalis 381 enhances binding of fimbriae to cultured human fibroblasts and matrix proteins. Infect Immun, 1996. 64(3): p. 756-62.
200. Pike, R., et al., Lysine- and arginine-specific proteinases from Porphyromonas gingivalis. Isolation, characterization, and evidence for the existence of complexes with hemagglutinins. J Bio Chem, 1994. 269(1): p. 406-11.
201. Diamond, G., N. Beckloff, and L.K. Ryan, Host defense peptides in the oral cavity and the lung: similarities and differences. J Dent Res, 2008. 87(10): p. 915-27.
202. Carlisle, M.D., R.N. Srikantha, and K.A. Brogden, Degradation of human alpha- and beta-defensins by culture supernatants of Porphyromonas gingivalis strain 381. J Innate Immun, 2009. 1(2): p. 118-22.
203. Gutner, M., et al., Saliva enables the antimicrobial activity of LL-37 in the presence of proteases of Porphyromonas gingivalis. Infect Immun, 2009. 77(12): p. 5558-63.
204. Kuula, H., et al., Human β-defensin-1 and -2 and matrix metalloproteinase-25 and -26 expression in chronic and aggressive periodontitis and in peri-implantitis. Arch Oral Biol. 53(2): p. 175-86.
205. Odell, E.W. and P.J. Wu, Susceptibility of Porphyromonas gingivalis and P. asaccharolytica to the non-oxidative killing mechanisms of human neutrophils. Arch Oral Biol. 37(8): p. 597-601.
206. Peitsch, M.C. and J. Tschopp, Assembly of macromolecular pores by immune defense systems. Curr Opin Cell Biol, 1991. 3(4): p. 710-6.
207. Grenier, D. and V.D. La, Proteases of Porphyromonas gingivalis as important virulence factors in periodontal disease and potential targets for plant-derived compounds: a review article. Curr Drug Targets, 2011. 12(3): p. 322-31.
208. Sundqvist, G., et al., Degradation of Human Immunoglobulins G and M and Complement Factors C3 and C5 by Black-Pigmented Bacteroides. J Med Microbiol, 1985. 19(1): p. 85-94.
209. Grenier, D., Degradation of host protease inhibitors and activation of plasminogen by proteolytic enzymes from Porphyromonas gingivalis and Treponema denticola. Microbiol, 1996. 142 ( Pt 4): p. 955-61.
210. Slaney, J.M. and M.A. Curtis, Mechanisms of evasion of complement by Porphyromonas gingivalis. Front Biosci, 2008. 13: p. 188-96.
211. Grenier, D., et al., Effect of Inactivation of the Arg- and/or Lys-Gingipain Gene on Selected Virulence and Physiological Properties of Porphyromonas gingivalis. Infection and Immunity, 2003. 71(8): p. 4742-8.
212. Jagels, M.A., et al., Proteolytic inactivation of the leukocyte C5a receptor by proteinases derived from Porphyromonas gingivalis. Infect Immun, 1996. 64(6): p. 1984-91.
213. Jagels, M.A., et al., Cleavage of the human C5A receptor by proteinases derived from Porphyromonas gingivalis: cleavage of leukocyte C5a receptor. Adv Exp Med Biol, 1996. 389: p. 155-64.

125
214. Miwa, T. and W.-C. Song, Membrane complement regulatory proteins: insight from animal studies and relevance to human diseases. Int Immunopharmacol, 2001. 1(3): p. 445-59.
215. Riley-Vargas, R.C., et al., CD46: expanding beyond complement regulation. Trends Immunol. 25(9): p. 496-503.
216. Mahtout, H., et al., Porphyromonas gingivalis mediates the shedding and proteolysis of complement regulatory protein CD46 expressed by oral epithelial cells. Oral Microbiol Immunol, 2009. 24(5): p. 396-400.
217. Lovkvist, L., et al., CD46 Contributes to the severity of group A streptococcal infection. Infect Immun, 2008. 76(9): p. 3951-8.
218. Lala, A., et al., Porphyromonas gingivalis trypsin-like protease: A possible natural ligand for the neutrophil formyl peptide receptor. Biochem Biophys Res Comm, 1994. 199(3): p. 1489-96.
219. Kjeldsen, M., P. Holmstrup, and K. Bendtzen, Marginal periodontitis and cytokines: a review of the literature. J Periodontol, 1993. 64(11): p. 1013-22.
220. Wilson, M., R. Seymour, and B. Henderson, Bacterial perturbation of cytokine networks. Infect Immun, 1998. 66(6): p. 2401-9.
221. Sugawara, S., et al., Proteolysis of human monocyte CD14 by cysteine proteinases (gingipains) from Porphyromonas gingivalis leading to lipopolysaccharide hyporesponsiveness. J Immunol, 2000. 165(1): p. 411-8.
222. Oleksy, A., et al., Proteolysis of interleukin-6 receptor (IL-6R) by Porphyromonas gingivalis cysteine proteinases (gingipains) inhibits interleukin-6-mediated cell activation. Microb Pathog, 2002. 32(4): p. 173-81.
223. Duncan, L., et al., Loss of lipopolysaccharide receptor CD14 from the surface of human macrophage-like cells mediated by Porphyromonas gingivalis outer membrane vesicles. Microb Pathog, 2004. 36(6): p. 319-25.
224. Kilian, M., Degradation of immunoglobulins A2, A2, and G by suspected principal periodontal pathogens. Infect Immun, 1981. 34(3): p. 757-65.
225. Birkedal-Hansen, H., et al., Characterization of collagenolytic activity from strains of Bacteroides gingivalis. J Periodontal Res, 1988. 23(4): p. 258-64.
226. Abe, N., et al., Biochemical and functional properties of lysine-specific cysteine proteinase (Lys-Gingipain) as a virulence factor of Porphyromonas gingivalis in periodontal disease. J Biochem, 1998. 123(2): p. 305-12.
227. Houle, M.A., et al., The collagenase activity of Porphyromonas gingivalis is due to Arg-gingipain. FEMS Microbiol Lett, 2003. 221(2): p. 181-5.
228. Okada, H. and S. Murakami, Cytokine expression in periodontal health and disease. Crit Rev Oral Biol Med, 1998. 9(3): p. 248-66.
229. Grenier, D. and D. Mayrand, Functional characterization of extracellular vesicles produced by Bacteroides gingivalis. Infect Immun, 1987. 55(1): p. 111-7.
230. Mayrand, D. and D. Grenier, Biological activities of outer membrane vesicles. Can J Microbiol, 1989. 35(6): p. 607-13.
231. Saxen, L., Heredity of juvenile periodontitis. J Clin Periodontol, 1980. 7(4): p. 276-88. 232. Haffajee, A.D. and S.S. Socransky, Attachment level changes in destructive periodontal
diseases. Journal of Clinical Periodontology, 1986. 13(5): p. 461-72. 233. Moore, W.E., et al., Bacteriology of experimental gingivitis in young adult humans. Infect
Immun, 1982. 38(2): p. 651-67. 234. Tonetti, M.S. and A. Mombelli, Early-onset periodontitis. Ann Periodontol, 1999. 4(1): p. 39-53.

126
235. Henderson, B., J.M. Ward, and D. Ready, Aggregatibacter (Actinobacillus) actinomycetemcomitans: a triple A* periodontopathogen? Periodontol 2000, 2010. 54(1): p. 78-105.
236. Asikainen, S. and S. Alaluusua, Bacteriology of dental infections. Eur Heart J, 1993. 14 Suppl K: p. 43-50.
237. Fives-Taylor, P.M., et al., Virulence factors of Actinobacillus actinomycetemcomitans. Periodontol 2000, 1999. 20(1): p. 136-67.
238. Slots, J., et al., The occurrence of Actinobacillus actinomycetemcomitans, Bacteroides gingivalis and Bacteroides intermedius in destructive periodontal disease in adults. J Clin Periodontol, 1986. 13(6): p. 570-7.
239. Zambon, J.J., Actinobacillus actinomycetemcomitans in human periodontal disease. J Clin Periodontol, 1985. 12(1): p. 1-20.
240. Wang, P.L., et al., Effect of Actinobacillus actinomycetemcomitans protease on the proliferation of gingival epithelial cells. Oral Dis, 2001. 7(4): p. 233-7.
241. Slots, J., et al., Actinobacillus actinomycetemcomitans in human periodontal disease. Association, serology, leukotoxicity, and treatment. J Periodontal Res, 1982. 17(5): p. 447-8.
242. Zambon, J.J., L.A. Christersson, and J. Slots, Actinobacillus actinomycetemcomitans in human periodontal disease. Prevalence in patient groups and distribution of biotypes and serotypes within families. J Periodontol, 1983. 54(12): p. 707-11.
243. Kaplan, J.B., et al., Structural and genetic analyses of O polysaccharide from Actinobacillus actinomycetemcomitans serotype f. Infect Immun, 2001. 69(9): p. 5375-84.
244. Wilson, M. and B. Henderson, Virulence factors of Actinobacillus actinomycetemcomitans relevant to the pathogenesis of inflammatory periodontal diseases. FEMS Microbiol Rev, 1995. 17(4): p. 365-79.
245. Tsai, C.C., et al., Extraction and partial characterization of a leukotoxin from a plaque-derived Gram-negative microorganism. Infect Immun, 1979. 25(1): p. 427-39.
246. Tsai, C.C., et al., Extraction and isolation of a leukotoxin from Actinobacillus actinomycetemcomitans with polymyxin B. Infect Immun, 1984. 43(2): p. 700-5.
247. Baehni, P., et al., Interaction of inflammatory cells and oral microorganisms. VIII. Detection of leukotoxic activity of a plaque-derived gram-negative microorganism. Infect Immun, 1979. 24(1): p. 233-43.
248. Baehni, P.C., et al., Leukotoxic activity in different strains of the bacterium Actinobacillus actinomycetemcomitans isolated from juvenile periodontitis in man. Arch Oral Biol, 1981. 26(8): p. 671-6.
249. Research, Science, and Therapy Committee of The American Academy of Periodontology The pathogenesis of periodontal diseases. J Periodontol, 1999. 70(4): p. 457-70.
250. Taichman, N.S., et al., Comparative studies on the biology of Actinobacillus actinomycetemcomitans leukotoxin in primates. Oral Microbiol Immunol, 1987. 2(3): p. 97-104.
251. Iwase, M., et al., Effects of cations and osmotic protectants on cytolytic activity of Actinobacillus actinomycetemcomitans leukotoxin. Infect Immun, 1990. 58(6): p. 1782-8.
252. Kachlany, S.C., Aggregatibacter actinomycetemcomitans leukotoxin: from threat to therapy. J Dent Res, 2010. 89(6): p. 561-70.
253. DiRienzo, J.M. and T.L. McKay, Identification and characterization of genetic cluster groups of Actinobacillus actinomycetemcomitans isolated from the human oral cavity. J Clin Microbiol, 1994. 32(1): p. 75-81.

127
254. Guthmiller, J.M., D. Kolodrubetz, and E. Kraig, Mutational analysis of the putative leukotoxin transport genes in Actinobacillus actinomycetemcomitans. Microb Pathog, 1995. 18(5): p. 307-21.
255. Spitznagel, J., Jr., E. Kraig, and D. Kolodrubetz, Regulation of leukotoxin in leukotoxic and nonleukotoxic strains of Actinobacillus actinomycetemcomitans. Infect Immun, 1991. 59(4): p. 1394-401.
256. Kolodrubetz, D., et al., cis Elements and trans factors are both important in strain-specific regulation of the leukotoxin gene in Actinobacillus actinomycetemcomitans. Infect Immun, 1996. 64(9): p. 3451-460.
257. Brogan, J.M., et al., Regulation of Actinobacillus actinomycetemcomitans leukotoxin expression: analysis of the promoter regions of leukotoxic and minimally leukotoxic strains. Infect Immun, 1994. 62(2): p. 501-8.
258. Hritz, M., E. Fisher, and D.R. Demuth, Differential regulation of the leukotoxin operon in highly leukotoxic and minimally leukotoxic strains of Actinobacillus actinomycetemcomitans. Infect Immun, 1996. 64(7): p. 2724-9.
259. Kamin, S., et al., Inhibition of fibroblast proliferation and collagen synthesis by capsular material from Actinobacillus actinomycetemcomitans. J Med Microbiol, 1986. 22(3): p. 245-9.
260. Letzelter, C., et al., Supernatant cytotoxicity and proteolytic activity of selected oral bacteria against human gingival fibroblasts in vitro. Arch Oral Biol, 1998. 43(1): p. 15-23.
261. Shenker, B.J., M.E. Kushner, and C.C. Tsai, Inhibition of fibroblast proliferation by Actinobacillus actinomycetemcomitans. Infect Immun, 1982. 38(3): p. 986-92.
262. Stevens, R.H., C. Gatewood, and B.F. Hammond, Cytotoxicity of the bacterium Actinobacillus actinomycetemcomitans extracts in human gingival fibroblasts. Arch Oral Biol, 1983. 28(11): p. 981-7.
263. Mayer, M.P., et al., Identification of a cytolethal distending toxin gene locus and features of a virulence-associated region in Actinobacillus actinomycetemcomitans. Infect Immun, 1999. 67(3): p. 1227-37.
264. Sugai, M., et al., The cell cycle-specific growth-inhibitory factor produced by Actinobacillus actinomycetemcomitans is a cytolethal distending toxin. Infect Immun, 1998. 66(10): p. 5008-19.
265. Moore, W.E., Microbiology of periodontal disease. J Periodontal Res, 1987. 22(5): p. 335-41. 266. Helgeland, K. and O. Nordby, Cell cycle-specific growth inhibitory effect on human gingival
fibroblasts of a toxin isolated from the culture medium of Actinobacillus actinomycetemcomitans. J Periodontal Res, 1993. 28(3): p. 161-5.
267. Kiley, P. and S.C. Holt, Characterization of the lipopolysaccharide from Actinobacillus actinomycetemcomitans Y4 and N27. Infect Immun, 1980. 30(3): p. 862-73.
268. Wilson, M., K. Reddi, and B. Henderson, Cytokine-inducing components of periodontopathogenic bacteria. J Periodontal Res, 1996. 31(6): p. 393-407.
269. Nishida, E., et al., Bone resorption and local interleukin-1alpha and interleukin-1beta synthesis induced by Actinobacillus actinomycetemcomitans and Porphyromonas gingivalis lipopolysaccharide. J Periodontal Res, 2001. 36(1): p. 1-8.
270. Bodet, C., F. Chandad, and D. Grenier, Anti-inflammatory activity of a high-molecular-weight cranberry fraction on macrophages stimulated by lipopolysaccharides from periodontopathogens. J Dent Res, 2006. 85(3): p. 235-9.
271. Noguchi, K., et al., Prostaglandin production via induction of cyclooxygenase-2 by human gingival fibroblasts stimulated with lipopolysaccharides. Inflammation, 1996. 20(5): p. 555-68.

128
272. Saglie, F.R., et al., Lipopolysaccharide from Actinobacillus actinomycetemcomitans stimulates macrophages to produce interleukin-1 and tumor necrosis factor mRNA and protein. Oral Microbiol Immunol, 1990. 5(5): p. 256-62.
273. Holt, S.C., A.C. Tanner, and S.S. Socransky, Morphology and ultrastructure of oral strains of Actinobacillus actinomycetemcomitans and Haemophilus aphrophilus. Infect Immun, 1980. 30(2): p. 588-600.
274. Lai, C.H., M.A. Listgarten, and B.F. Hammond, Comparative ultrastructure of leukotoxic and non-leukotoxic strains of Actinobacillus actinomycetemcomitans. J Periodontal Res, 1981. 16(4): p. 379-89.
275. Nowotny, A., et al., Release of toxic microvesicles by Actinobacillus actinomycetemcomitans. Infect Immun, 1982. 37(1): p. 151-4.
276. Kaplan, A.H., et al., Infection due to Actinobacillus actinomycetemcomitans: 15 cases and review. Rev Infect Dis, 1989. 11(1): p. 46-63.
277. Han, Y.W., Fusobacterium nucleatum: a commensal-turned pathogen. Curr Opin Microbiol, 2015. 23: p. 141-147.
278. Field, C.A., et al., Investigation and quantification of key periodontal pathogens in patients with type 2 diabetes. J Periodontal Res, 2012. 47(4): p. 470-8.
279. Moore, W.E.C. and L.V.H. Moore, The bacteria of periodontal diseases. Periodontol 2000, 1994. 5(1): p. 66-77.
280. Griffen, A.L., et al., Distinct and complex bacterial profiles in human periodontitis and health revealed by 16S pyrosequencing. ISME J, 2012. 6(6): p. 1176-85.
281. Loozen, G., et al., Inter-bacterial correlations in subgingival biofilms: a large-scale survey. J Clin Periodontol, 2014. 41(1): p. 1-10.
282. Moore, W.E., et al., Comparative bacteriology of juvenile periodontitis. Infect Immun, 1985. 48(2): p. 507-19.
283. Yang, N.-Y., et al., Progression of periodontal inflammation in adolescents is associated with increased number of Porphyromonas gingivalis, Prevotella intermedia, Tannerella forsythensis, and Fusobacterium nucleatum. Int J Paediatric Dent, 2014. 24(3): p. 226-33.
284. Riep, B., et al., Are putative periodontal pathogens reliable diagnostic markers? J Clin Microbiol, 2009. 47(6): p. 1705-11.
285. Gharbia, S.E., et al., The distribution and frequency of Fusobacterium nucleatum subspecies in the human oral cavity. Oral Microbiol Immunol, 1990. 5(6): p. 324-7.
286. Signat, B., et al., Fusobacterium nucleatum in periodontal health and disease. Curr Issues Mol Biol, 2011. 13(2): p. 25-36.
287. Kolenbrander, P.E., Oral microbial communities: biofilms, interactions, and genetic systems. Annu Rev Microbiol, 2000. 54: p. 413-37.
288. Kolenbrander, P.E., et al., Communication among oral bacteria. Microbiol Mol Biol Re, 2002. 66(3): p. 486-505.
289. Njoroge, T., et al., A role for fimbriae in Porphyromonas gingivalis invasion of oral epithelial cells. Infect Immun, 1997. 65(5): p. 1980-4.
290. Quirynen, M. and C.M. Bollen, The influence of surface roughness and surface-free energy on supra- and subgingival plaque formation in man. A review of the literature. J Clin Periodontol, 1995. 22(1): p. 1-14.
291. Paquette, D.W. and R.C. Williams, Modulation of host inflammatory mediators as a treatment strategy for periodontal diseases. Periodontol 2000, 2000. 24: p. 239-52.
292. Kornman, K.S., R.C. Page, and M.S. Tonetti, The host response to the microbial challenge in periodontitis: assembling the players. Periodontol 2000, 1997. 14: p. 33-53.

129
293. Bidault, P., F. Chandad, and D. Grenier, Systemic antibiotic therapy in the treatment of periodontitis. J Can Dent Assoc, 2007. 73(6): p. 515-20.
294. Miller, N., et al., Mécanismes immunopathologiques dans la maladie parodontale. Encyclopédie Médico-Chirurgicale (Editions Scientifiques et Médicales Elsevier SAS, Paris), Odontologie, 23-435-B-10, , 2002: 9 p.
295. Kaufman, E. and I.B. Lamster, Analysis of saliva for periodontal diagnosis. J Clinl Periodontol, 2000. 27(7): p. 453-65.
296. Bonass, W.A., et al., Expression of β-defensin genes by human salivary glands. Oral Microbiol Immunol, 1999. 14(6): p. 371-4.
297. Socransky, S.S., Relationship of bacteria to the etiology of periodontal disease. J Dent Res, 1970. 49(2): p. 203-22.
298. Bodet, C., F. Chandad, and D. Grenier, Porphyromonas gingivalis-induced inflammatory mediator profile in an ex vivo human whole blood model. Clin Exp Immunol, 2006. 143(1): p. 50-7.
299. Liu, Y.C., U.H. Lerner, and Y.T. Teng, Cytokine responses against periodontal infection: protective and destructive roles. Periodontol 2000, 2010. 52(1): p. 163-206.
300. Taba, M., Jr., et al., Diagnostic biomarkers for oral and periodontal diseases. Dent Clin North Am, 2005. 49(3): p. 551-71.
301. Gapski, R., et al., Effect of systemic matrix metalloproteinase inhibition on periodontal wound repair: a proof of concept trial. J Periodontol, 2004. 75(3): p. 441-52.
302. Ryan, M., S. Ramamurthy, and L. Golub, Matrix metalloproteinases and their inhibition in periodontal treatment. Curr opin periodontol, 1995. 3: p. 85-96.
303. Makela, M., et al., Matrix metalloproteinases (MMP-2 and MMP-9) of the oral cavity: cellular origin and relationship to periodontal status. J Dent Res, 1994. 73(8): p. 1397-406.
304. Birkedal-Hansen, H., et al., Matrix metalloproteinases: a review. Crit Rev Oral Biol Med, 1993. 4(2): p. 197-250.
305. Page, R.C., The role of inflammatory mediators in the pathogenesis of periodontal disease. J Periodontal Res, 1991. 26(3 Pt 2): p. 230-42.
306. Lindner, D., et al., Differential expression of matrix metalloproteases in human fibroblasts with different origins. Biochem Res Int, 2012. 2012: p. 875742.
307. Reynolds, J.J. and M.C. Meikle, Mechanisms of connective tissue matrix destruction in periodontitis. Periodontol 2000, 1997. 14: p. 144-57.
308. Birkedal-Hansen, H., Role of matrix metalloproteinases in human periodontal diseases. J Periodontol, 1993. 64(5 Suppl): p. 474-84.
309. Murphy, G., et al., The role of plasminogen activators in the regulation of connective tissue metalloproteinases. Ann N Y Acad Sci, 1992. 667: p. 1-12.
310. Ingman, T., et al., Matrix metalloproteinases and their inhibitors in gingival crevicular fluid and saliva of periodontitis patients. J Clin Periodontol, 1996. 23(12): p. 1127-1132.
311. Heikkinen, A.M., et al., Smoking affects diagnostic salivary periodontal disease biomarker levels in adolescents. J Periodontol, 2010. 81(9): p. 1299-307.
312. Dahan, M., et al., Expression of matrix metalloproteinases in healthy and diseased human gingiva. J Clini Periodontol, 2001. 28(2): p. 128-36.
313. Tonetti, M., et al., Detection of interleukin‐8 and matrix metalloproteinases transcripts in healthy and diseased gingival biopsies by RNA/PCR. J Periodontal Res, 1993. 28(6): p. 511-513.
314. Aiba, T., et al., Matrix metalloproteinases‐1 and‐8 and TIMP‐1 mRNA levels in normal and diseased human gingivae. Eur J Oral Sci, 1996. 104(5‐6): p. 562-9.

130
315. Biomarkers Definitions Working, G., Biomarkers and surrogate endpoints: Preferred definitions and conceptual framework. Clin Pharm Ther, 2001. 69(3): p. 89-95.
316. Ozmeric, N., Advances in periodontal disease markers. Clin Chim Acta, 2004. 343(1-2): p. 1-16.
317. Kaufman, E. and I.B. Lamster, Analysis of saliva for periodontal diagnosis--a review. J Clin Periodontol, 2000. 27(7): p. 453-65.
318. Zambon, J.J., M. Nakamura, and J. Slots, Effect of periodontal therapy on salivary enzymatic activity. J Periodontal Res, 1985. 20(6): p. 652-9.
319. Hayakawa, H., et al., Collagenase activity and tissue inhibitor of metalloproteinases-1 (TIMP-1) content in human whole saliva from clinically healthy and periodontally diseased subjects. J Periodontal Res, 1994. 29(5): p. 305-8.
320. Sorsa, T., et al., Matrix metalloproteinases: Contribution to pathogenesis, diagnosis and treatment of periodontal inflammation. Annals of Medicine, 2006. 38(5): p. 306-21.
321. Salminen, A., et al., Salivary biomarkers of bacterial burden, inflammatory response, and tissue destruction in periodontitis. J Clin Periodontol, 2014. 41(5): p. 442-50.
322. Domeij, H., T. Yucel-Lindberg, and T. Modeer, Signal pathways involved in the production of MMP-1 and MMP-3 in human gingival fibroblasts. Eur J Oral Sci, 2002. 110(4): p. 302-6.
323. Gupta, N., et al., Role of salivary matrix metalloproteinase-8 (MMP-8) in chronic periodontitis diagnosis. Front Med, 2015. 9(1): p. 72-6.
324. Guidelines for periodontal therapy. The American Academy of Periodontology. J Periodontol, 1998. 69(3): p. 405-8.
325. Manach, C., et al., Polyphenols: food sources and bioavailability. Am J Clin Nutr, 2004. 79(5): p. 727-47.
326. Cheynier, V., Polyphenols in foods are more complex than often thought. Am J Clin Nutr, 2005. 81(1): p. 223S-9S.
327. Dixon, R.A. and N.L. Paiva, Stress-induced phenylpropanoid metabolism. Plant Cell, 1995. 7(7): p. 1085-97.
328. Koes, R.E., F. Quattrocchio, and J.N.M. Mol, The flavonoid biosynthetic pathway in plants: Function and evolution. BioEssays, 1994. 16(2): p. 123-32.
329. Quideau, S., et al., Plant polyphenols: chemical properties, biological activities, and synthesis. Angew Chem Int Ed Engl, 2011. 50(3): p. 586-621.
330. Beckman, C.H., Phenolic-storing cells: keys to programmed cell death and periderm formation in wilt disease resistance and in general defence responses in plants? Physiol Mol Plant Path, 2000. 57(3): p. 101-10.
331. Pandey, K.B. and S.I. Rizvi, Plant polyphenols as dietary antioxidants in human health and disease. Oxid Med Cell Longev, 2009. 2(5): p. 270-8.
332. Schijlen, E.G., et al., Modification of flavonoid biosynthesis in crop plants. Phytochem, 2004. 65(19): p. 2631-48.
333. Bravo, L., Polyphenols: chemistry, dietary sources, metabolism, and nutritional significance. Nutr Rev, 1998. 56(11): p. 317-33.
334. König, H., G. Unden, and J. Fröhlich, Biology of Microorganisms on Grapes, in Must and in Wine. 2009: Springer.
335. Tsao, R., Chemistry and biochemistry of dietary polyphenols. Nutrients, 2010. 2(12): p. 1231-46.
336. Kluth, D., et al., Modulation of pregnane X receptor- and electrophile responsive element-mediated gene expression by dietary polyphenolic compounds. Free Radic Biol Med, 2007. 42(3): p. 315-25.

131
337. Gardner, E.J., C.H. Ruxton, and A.R. Leeds, Black tea--helpful or harmful? A review of the evidence. Eur J Clin Nutr, 2007. 61(1): p. 3-18.
338. Wu, C.D. and G.X. Wei, Tea as a functional food for oral health. Nutrition, 2002. 18(5): p. 443-4.
339. Cooper, R., Green tea and theanine: health benefits. Int J Food Sci Nutr, 2012. 63 Suppl 1: p. 90-7.
340. Cabrera, C., R. Artacho, and R. Gimenez, Beneficial effects of green tea--a review. J Am Coll Nutr, 2006. 25(2): p. 79-99.
341. McKay, D.L. and J.B. Blumberg, The role of tea in human health: an update. J Am Coll Nutr, 2002. 21(1): p. 1-13.
342. Willson, K.C., Coffee, cocoa and tea. 1999, New York: CABI Pub. 343. Crespy, V. and G. Williamson, A review of the health effects of green tea catechins in in vivo
animal models. J Nutr, 2004. 134(12 Suppl): p. 3431S-40S. 344. Chacko, S.M., et al., Beneficial effects of green tea: a literature review. Chin Med, 2010. 5: p.
13. 345. Pastore, R.L. and P. Fratellone, Potential health benefits of green tea (Camellia sinensis): a
narrative review. Explore (NY), 2006. 2(6): p. 531-9. 346. Wierzejska, R., Tea and health--a review of the current state of knowledge. Przegl Epidemiol,
2014. 68(3): p. 501-6, 595-9. 347. Lin, Y.S., et al., Factors affecting the levels of tea polyphenols and caffeine in tea leaves. J
Agric Food Chem, 2003. 51(7): p. 1864-73. 348. Graham, H.N., Green tea composition, consumption, and polyphenol chemistry. Prev Med,
1992. 21(3): p. 334-50. 349. Astill, C., et al., Factors affecting the caffeine and polyphenol contents of black and green tea
infusions. J Agric Food Chem, 2001. 49(11): p. 5340-7. 350. Narotzki, B., et al., Green tea: a promising natural product in oral health. Arch Oral Biol, 2012.
57(5): p. 429-35. 351. Fernández, P.L., et al., Study of Catechin and Xanthine Tea Profiles as Geographical Tracers.
J Agr Food Chem, 2002. 50(7): p. 1833-9. 352. Reygaert, W.C., The antimicrobial possibilities of green tea. Front Microbiol, 2014. 5: p. 434. 353. Hara, Y., Green Tea: Health Benefits and Applications. 2001: CRC Press. 354. Taylor, P.W., J.M. Hamilton-Miller, and P.D. Stapleton, Antimicrobial properties of green tea
catechins. Food Sci Technol Bull, 2005. 2: p. 71-81. 355. Varnam, A. and J.M. Sutherland, Beverages: technology, chemistry and microbiology. 2012:
Springer US. 356. da Silva Pinto, M., Tea: A new perspective on health benefits. Food Res Int, 2013. 53(2): p.
558-67. 357. Cooper, R., D.J. Morre, and D.M. Morre, Medicinal benefits of green tea: Part I. Review of
noncancer health benefits. J Altern Complement Med, 2005. 11(3): p. 521-8. 358. Cooper, R., D.J. Morre, and D.M. Morre, Medicinal benefits of green tea: part II. review of
anticancer properties. J Altern Complement Med, 2005. 11(4): p. 639-52. 359. Yang, C.S., et al., Tea and tea polyphenols in cancer prevention. J Nutr, 2000. 130(2S Suppl):
p. 472S-8S. 360. Yang, C.S. and Z.-Y. Wang, Tea and cancer. J Natl Cancer Inst, 1993. 85(13): p. 1038-49. 361. Katiyar, S. and H. Mukhtar, Tea in chemoprevention of cancer. J Onc, 1996. 8(2): p. 221-238. 362. Keli, S.O., et al., Dietary flavonoids, antioxidant vitamins, and incidence of stroke: The zutphen
study. Arch Intern Med, 1996. 156(6): p. 637-42.

132
363. Humphrey, L.L., et al., Periodontal disease and coronary heart disease incidence: a systematic review and meta-analysis. J Gen Intern Med, 2008. 23(12): p. 2079-86.
364. Hertog, M.G., et al., Dietary antioxidant flavonoids and risk of coronary heart disease: the Zutphen Elderly Study. Lancet, 1993. 342(8878): p. 1007-11.
365. Hollman, P.C., E.J. Feskens, and M.B. Katan, Tea flavonols in cardiovascular disease and cancer epidemiology. Proc Soc Exp Biol Med, 1999. 220(4): p. 198-202.
366. Tijburg, L., et al., Tea flavonoids and cardiovascular diseases: a review. Crit Rev Food Sci Nutr, 1997. 37(8): p. 771-85.
367. Haqqi, T.M., et al., Prevention of collagen-induced arthritis in mice by a polyphenolic fraction from green tea. Proc Natl Acad Sci U S A, 1999. 96(8): p. 4524-9.
368. Akhtar, N. and T.M. Haqqi, Epigallocatechin-3-gallate suppresses the global interleukin-1beta-induced inflammatory response in human chondrocytes. Arthritis Res Ther, 2011. 13(3): p. R93.
369. Chatterjee, A., et al., Green tea: A boon for periodontal and general health. J Indian Soc Periodontol, 2012. 16(2): p. 161-7.
370. de Pablo, P., et al., Periodontitis in systemic rheumatic diseases. Nat Rev Rheumatol, 2009. 5(4): p. 218-24.
371. Smolik, I., D. Robinson, and H.S. El-Gabalawy, Periodontitis and rheumatoid arthritis: epidemiologic, clinical, and immunologic associations. Compend Contin Educ Dent, 2009. 30(4): p. 188-90.
372. Bell, S.J. and G.K. Goodrick, A functional food product for the management of weight. Crit Rev Food Sci Nutr, 2002. 42(2): p. 163-78.
373. Klaus, S., et al., Epigallocatechin gallate attenuates diet-induced obesity in mice by decreasing energy absorption and increasing fat oxidation. Int J Obes (Lond), 2005. 29(6): p. 615-23.
374. Furuyashiki, T., et al., Tea catechin suppresses adipocyte differentiation accompanied by down-regulation of PPARgamma2 and C/EBPalpha in 3T3-L1 cells. Biosci Biotechnol Biochem, 2004. 68(11): p. 2353-9.
375. Han, L.K., et al., Anti-obesity action of oolong tea. Int J Obes Relat Metab Disord, 1999. 23(1): p. 98-105.
376. Oliver, R.C. and T. Tervonen, Periodontitis and tooth loss: comparing diabetics with the general population. J Am Dent Assoc, 1993. 124(12): p. 71-6.
377. Katz, P.P., et al., Epidemiology and prevention of periodontal disease in individuals with diabetes. Diabetes Care, 1991. 14(5): p. 375-85.
378. Kamer, A.R., et al., Inflammation and Alzheimer's disease: possible role of periodontal diseases. Alzheimers Dement, 2008. 4(4): p. 242-50.
379. Kamer, A.R., et al., TNF-α and antibodies to periodontal bacteria discriminate between Alzheimer’s disease patients and normal subjects. J Neuroimmunol, 2009. 216(1-2): p. 92-7.
380. Guo, S., et al., Protective effects of green tea polyphenols in the 6-OHDA rat model of parkinson’s disease through inhibition of ROS-NO pathway. Biol Psychiatry. 62(12): p. 1353-62.
381. Hegarty, V.M., H.M. May, and K.T. Khaw, Tea drinking and bone mineral density in older women. Am J Clin Nutr, 2000. 71(4): p. 1003-7.
382. Attia, E.L. and K.G. Marshall, Halitosis. Can Med Assoc J, 1982. 126(11): p. 1281-5. 383. Rosenberg, M., Bad breath, diagnosis and treatment. Univ Tor Dent J, 1990. 3(2): p. 7-11. 384. Tonzetich, J. and V.J. Richter, Evaluation of volatile odoriferous components of saliva. Arch
Oral Biol, 1964. 9: p. 39-46.

133
385. Arab, H., et al., Review of The therapeutic effects of Camellia sinensis (green tea) on oral and periodontal health. J Med Plant Res, 2011. 5(23): p. 5465-9.
386. Nakano, Y., M. Yoshimura, and T. Koga, Methyl mercaptan production by periodontal bacteria. Int Dent J, 2002. 52 Suppl 3: p. 217-20.
387. Xu, X., X.D. Zhou, and C.D. Wu, Tea catechin EGCg suppresses the mgl gene associated with halitosis. J Dent Res, 2010. 89(11): p. 1304-8.
388. Nakano, Y., M. Yoshimura, and T. Koga, Correlation between oral malodor and periodontal bacteria. Microb Infect, 2002. 4(6): p. 679-83.
389. Loesche, W.J. and C. Kazor, Microbiology and treatment of halitosis. Periodontol 2000, 2002. 28: p. 256-79.
390. Persson, S., et al., The formation of hydrogen sulfide and methyl mercaptan by oral bacteria. Oral Microbiol Immunol, 1990. 5(4): p. 195-201.
391. Armstrong, B.L., M.L. Sensat, and J.L. Stoltenberg, Halitosis: a review of current literature. J Dent Hyg, 2010. 84(2): p. 65-74.
392. Rosenberg, M., Clinical assessment of bad breath: current concepts. J Am Dent Assoc, 1996. 127(4): p. 475-82.
393. Greenstein, R.B.-N., et al., Reduction of oral malodor by oxidizing lozenges. J Periodontol, 1997. 68(12): p. 1176-81.
394. Tangerman, A., Halitosis in medicine: a review. Int Dent J, 2002. 52 Suppl 3: p. 201-6. 395. Farina, V.H., et al., Effects of the medicinal plants Curcuma zedoaria and Camellia sinensis
on halitosis control. Braz Oral Res, 2012. 26(6): p. 523-9. 396. van den Broek, A.M., L. Feenstra, and C. de Baat, A review of the current literature on aetiology
and measurement methods of halitosis. J Dent, 2007. 35(8): p. 627-35. 397. Lodhia, P., et al., Effect of green tea on volatile sulfur compounds in mouth air. J Nutr Sci
Vitaminol (Tokyo), 2008. 54(1): p. 89-94. 398. Sakanaka, S. and Y. Okada, Inhibitory effects of green tea polyphenols on the production of a
virulence factor of the periodontal-disease-causing anaerobic bacterium Porphyromonas gingivalis. J Agr Food Chem, 2004. 52(6): p. 1688-92.
399. Yasuda, H. and T. Arakawa, Deodorizing mechanism of (–)-epigallocatechin gallate against methyl mercaptan. Biosci, Biotech Biochem, 1995. 59(7): p. 1232-6.
400. Yoshimura, M., et al., Formation of methyl mercaptan from L-methionine by Porphyromonas gingivalis. Infect Immun, 2000. 68(12): p. 6912-6.
401. Morin, M.P., et al., Green tea extract and its major constituent epigallocatechin-3-gallate inhibit growth and halitosis-related properties of Solobacterium moorei. BMC Complement Altern Med, 2015. 15: p. 48.
402. Rassameemasmaung, S., P. Phusudsawang, and V. Sangalungkarn, Effect of green tea mouthwash on oral malodor. ISRN Prev Med, 2013. 2013: p. 975148.
403. Zeng, Q.C., A.Z. Wu, and J. Pika, The effect of green tea extract on the removal of sulfur-containing oral malodor volatiles in vitro and its potential application in chewing gum. J Breath Res, 2010. 4(3): p. 036005.
404. Goenka, P., et al., Camellia sinensis (Tea): Implications and role in preventing dental decay. Pharmacogn Rev, 2013. 7(14): p. 152-6.
405. Gaur, S. and R. Agnihotri, Green tea: a novel functional food for the oral health of older adults. Geriatr Gerontol Int, 2014. 14(2): p. 238-50.
406. van Houte, J., Role of micro-organisms in caries etiology. J Dent Res, 1994. 73(3): p. 672-81. 407. Keyes, P.H., The infectious and transmissible nature of experimental dental caries. Findings
and implications. Arch Oral Biol, 1960. 1: p. 304-20.

134
408. Bonifait, L., F. Chandad, and D. Grenier, Probiotics for oral health: myth or reality? J Can Dent Assoc, 2009. 75(8): p. 585-90.
409. Selwitz, R.H., A.I. Ismail, and N.B. Pitts, Dental caries. Lancet, 2007. 369(9555): p. 51-9. 410. Hamada, S. and H.D. Slade, Biology, immunology, and cariogenicity of Streptococcus mutans.
Microbiol Rev, 1980. 44(2): p. 331-84. 411. Ferrazzano, G.F., et al., Antimicrobial properties of green tea extract against cariogenic
microflora: an in vivo study. J Med Food, 2011. 14(9): p. 907-11. 412. Wang, H., G.J. Provan, and K. Helliwell, Tea flavonoids: their functions, utilisation and analysis.
Trends Food Sci Tech, 2000. 11(4–5): p. 152-160. 413. Moynihan, P., Foods and factors that protect against dental caries. Nutrition Bulletin, 2000.
25(4): p. 281-286. 414. Xu, X., X.D. Zhou, and C.D. Wu, The tea catechin epigallocatechin gallate suppresses
cariogenic virulence factors of Streptococcus mutans. Antimicrob Agents Chemother, 2011. 55(3): p. 1229-36.
415. Newbrun, E., Effectiveness of water fluoridation. J Public Health Dent, 1989. 49(5 Spec No): p. 279-89.
416. Featherstone, J.D., Prevention and reversal of dental caries: role of low level fluoride. Community Dent Oral Epidemiol, 1999. 27(1): p. 31-40.
417. Featherstone, J.D., Remineralization, the natural caries repair process--the need for new approaches. Adv Dent Res, 2009. 21(1): p. 4-7.
418. Kashket, S., et al., In-vitro inhibition of glucosyltransferase from the dental plaque bacterium Streptococcus mutans by common beverages and food extracts. Arch Oral Biol, 1985. 30(11-12): p. 821-6.
419. Rosen, S., et al., Anticariogenic effects of tea in rats. J Dent Res, 1984. 63(5): p. 658-60. 420. Yu, H., T. Oho, and L.X. Xu, Effects of several tea components on acid resistance of human
tooth enamel. J Dent. 23(2): p. 101-5. 421. Onisi, M., et al., An experimental evidence of caries preventive activity of non-fluoride
component in tea. Koku Eisei Gakkai Zasshi, 1981. 31(2): p. 158-62. 422. Ferrazzano, G.F., et al., Anti-cariogenic effects of polyphenols from plant stimulant beverages
(cocoa, coffee, tea). Fitoterapia, 2009. 80(5): p. 255-62. 423. Xu, X., X.D. Zhou, and C.D. Wu, Tea catechin epigallocatechin gallate inhibits Streptococcus
mutans biofilm formation by suppressing gtf genes. Arch Oral Biol, 2012. 57(6): p. 678-83. 424. Hirasawa, M., K. Takada, and S. Otake, Inhibition of acid production in dental plaque bacteria
by green tea catechins. Caries Res, 2006. 40(3): p. 265-70. 425. Sakanaka, S., et al., Preventive effect of green tea polyphenols against dental caries in
conventional rats. Biosci, Biotech, Biochem, 1992. 56(4): p. 592-4. 426. Sakanaka, S., et al., Antibacterial Substances in Japanese Green Tea Extract against
Streptococcus mutans, a Cariogenic Bacterium. Agric Biol Chem, 1989. 53(9): p. 2307-11. 427. Otake, S., et al., Anticaries effects of polyphenolic compounds from Japanese green tea.
Caries Res, 1991. 25(6): p. 438-43. 428. Hattori, M., et al., Effect of tea polyphenols on glucan synthesis by glucosyltransferase from
Streptococcus mutans. Chem Pharm Bull (Tokyo), 1990. 38(3): p. 717-20. 429. Nakahara, K., et al., Inhibitory effect of oolong tea polyphenols on glycosyltransferases of
mutans Streptococci. Appl Environ Microbiol, 1993. 59(4): p. 968-73. 430. Saito, N., Anti-caries effects of polyphenol compound from Camellia sinensis. Nichidai Koko
Kagaku, 1990. 16(2): p. 154-63.

135
431. Sakanaka, S., et al., Inhibitory effects of green tea polyphenols on glucan synthesis and cellular adherence of cariogenic streptococci. Agric Biol Chem, 1990. 54(11): p. 2925-2929.
432. Hamilton-Miller, J.M., Anti-cariogenic properties of tea (Camellia sinensis). J Med Microbiol, 2001. 50(4): p. 299-302.
433. Bodet, C., et al., Potential oral health benefits of cranberry. Crit Rev Food Sci Nutr, 2008. 48(7): p. 672-80.
434. Rasheed, A. and M. Haider, Antibacterial activity of Camellia sinensis extracts against dental caries. Arch Pharm Res, 1998. 21(3): p. 348-52.
435. Wilson, M., Food constituents and oral health: current status and future prospects. 2009: Elsevier.
436. Wu, C., G. Wei, and M. Wilson, Tea as a functional food for oral health. Food constituents and oral health: current status and future prospects, 2009: p. 396-417.
437. Ikigai, H., et al., Bactericidal catechins damage the lipid bilayer. Biochim Biophys Acta, 1993. 1147(1): p. 132-6.
438. Arakawa, H., et al., Role of hydrogen peroxide in bactericidal action of catechin. Biol Pharm Bull, 2004. 27(3): p. 277-81.
439. Zhang, J. and S. Kashket, Inhibition of salivary amylase by black and green teas and their effects on the intraoral hydrolysis of starch. Caries Res, 1998. 32(3): p. 233-8.
440. Kashket, S. and V.J. Paolino, Inhibition of salivary amylase by water-soluble extracts of tea. Arch Oral Biol. 33(11): p. 845-6.
441. Sakanaka, S., Green tea polyphenols for prevention of dental caries. Chemistry and Application of Green Tea, 1997: p. 87-101.
442. Maruyama, T., et al., Supplementation of green tea catechins in dentifrices suppresses gingival oxidative stress and periodontal inflammation. Arch Oral Biol, 2011. 56(1): p. 48-53.
443. Kushiyama, M., et al., Relationship between intake of green tea and periodontal disease. J Periodontol, 2009. 80(3): p. 372-7.
444. Magnusson, I., et al., Recolonization of a subgingival microbiota following scaling in deep pockets. J Clin Periodontol, 1984. 11(3): p. 193-207.
445. Sakanaka, S., et al., Inhibitory effects of green tea polyphenols on growth and cellular adherence of an oral bacterium, Porphyromonas gingivalis. Biosci Biotechnol Biochem, 1996. 60(5): p. 745-9.
446. Makimura, M., et al., Inhibitory effect of tea catechins on collagenase activity. J Periodontol, 1993. 64(7): p. 630-6.
447. Hirasawa, M., et al., Improvement of periodontal status by green tea catechin using a local delivery system: A clinical pilot study. J Periodontal Res, 2002. 37(6): p. 433-8.
448. Okamoto, M., et al., Inhibitory effects of green tea catechins on protein tyrosine phosphatase in Prevotella intermedia. Oral Microbiol Immunol, 2003. 18(3): p. 192-5.
449. Okamoto, M., et al., Inhibitory effect of green tea catechins on cysteine proteinases in Porphyromonas gingivalis. Oral Microbiol Immunol, 2004. 19(2): p. 118-20.
450. Zhao, L., V.D. La, and D. Grenier, Antibacterial, antiadherence, antiprotease, and anti-inflammatory activities of various tea extracts: Potential benefits for periodontal diseases. J Med Food, 2013. 16(5): p. 428-36.
451. Lombardo Bedran, T.B., et al., Green tea extract and its major constituent, epigallocatechin-3-gallate, induce epithelial beta-defensin secretion and prevent beta-defensin degradation by Porphyromonas gingivalis. J Periodontal Res, 2014. 49(5): p. 615-23.
452. Chung, W.O., et al., Expression of defensins in gingiva and their role in periodontal health and disease. Curr Pharml Des, 2007. 13(30): p. 3073-83.

136
453. Komatsuzawa, H., et al., Susceptibility of periodontopathogenic and cariogenic bacteria to defensins and potential therapeutic use of defensins in oral diseases. Curr Pharm Des, 2007. 13(30): p. 3084-95.
454. Joly, S., et al., Human β-defensins 2 and 3 demonstrate strain-selective activity against oral microorganisms. J Clin Microbiol, 2004. 42(3): p. 1024-9.
455. Gursoy, U.K. and E. Könönen, Understanding the roles of gingival beta-defensins. J Oral Microbiol, 2012. 4: p. 10.3402/jom.v4i0.15127.
456. Sen, R. and D. Baltimore, Multiple nuclear factors interact with the immunoglobulin enhancer sequences. Cell, 1986. 46(5): p. 705-16.
457. Blackwell, T.S. and J.W. Christman, The role of nuclear factor-κB in cytokine gene regulation. Am J Respir Cell Mol Biol, 1997. 17(1): p. 3-9.
458. Arabaci, T., et al., Immunohistochemical and stereologic analysis of NF-κB activation in chronic periodontitis. Eur J Dent, 2010. 4(4): p. 454-61.
459. Nichols, T.C., et al., Role of nuclear factor-kappa B (NF-kappa B) in inflammation, periodontitis, and atherogenesis. Ann Periodontol, 2001. 6(1): p. 20-9.
460. Kirkwood, K.L., et al., Novel host response therapeutic approaches to treat periodontal diseases. Periodontol 2000, 2007. 43: p. 294-315.
461. Syed, D.N., et al., Green tea polyphenol EGCG suppresses cigarette smoke condensate-induced NF-kappaB activation in normal human bronchial epithelial cells. Oncogene, 2007. 26(5): p. 673-82.
462. Vasto, S., et al., Inflammatory networks in ageing, age-related diseases and longevity. Mech Ageing Dev, 2007. 128(1): p. 83-91.
463. Nakamura, H., et al., Green tea catechin inhibits lipopolysaccharide-induced bone resorption in vivo. J Periodontal Res, 2010. 45(1): p. 23-30.
464. Rogers, J., et al., Epigallocatechin gallate modulates cytokine production by bone marrow-derived dendritic cells stimulated with lipopolysaccharide or muramyldipeptide, or infected with Legionella pneumophila. Exp Biol Med (Maywood), 2005. 230(9): p. 645-51.
465. Yang, F., et al., Green tea polyphenols block endotoxin-induced tumor necrosis factor-production and lethality in a murine model. Journal Nutr, 1998. 128(12): p. 2334-40.
466. Yun, J.H., et al., Inhibitory effects of green tea polyphenol (-)-epigallocatechin gallate on the expression of matrix metalloproteinase-9 and on the formation of osteoclasts. J Periodontal Res, 2004. 39(5): p. 300-7.
467. Yun, J.H., et al., (-)-Epigallocatechin gallate induces apoptosis, via caspase activation, in osteoclasts differentiated from RAW 264.7 cells. J Periodontal Res, 2007. 42(3): p. 212-8.
468. Nakagawa, H., et al., Fenton reaction Is primarily involved in a mechanism of (−)-epigallocatechin-3-gallate to induce osteoclastic cell death. Biochem Biophys Rese Comm, 2002. 292(1): p. 94-101.
469. Dürr, U.H.N., U.S. Sudheendra, and A. Ramamoorthy, LL-37, the only human member of the cathelicidin family of antimicrobial peptides. Biochimica et Biophysica Acta (BBA) - Biomembranes, 2006. 1758(9): p. 1408-25.
470. Inomata, M., T. Into, and Y. Murakami, Suppressive effect of the antimicrobial peptide LL-37 on expression of IL-6, IL-8 and CXCL10 induced by Porphyromonas gingivalis cells and extracts in human gingival fibroblasts. Eur J Oral Sci, 2010. 118(6): p. 574-81.
471. Rosenfeld, Y., N. Papo, and Y. Shai, Endotoxin (lipopolysaccharide) neutralization by innate immunity host-defense peptides: peptide properties and plausible modes of action. J Biol Chem, 2006. 281(3): p. 1636-43.

137
472. Gorr, S.U., Antimicrobial peptides in periodontal innate defense. Front Oral Biol, 2012. 15: p. 84-98.
473. Lombardo Bedran, T.B., D. Palomari Spolidorio, and D. Grenier, Green tea polyphenol epigallocatechin-3-gallate and cranberry proanthocyanidins act in synergy with cathelicidin (LL-37) to reduce the LPS-induced inflammatory response in a three-dimensional co-culture model of gingival epithelial cells and fibroblasts. Arch Oral Biol. 60(6): p. 845-53.
474. Hall, M.J., R.F. Middleton, and D. Westmacott, The fractional inhibitory concentration (FIC) index as a measure of synergy. J Antimicrob Chemother, 1983. 11(5): p. 427-33.
475. Jonathan W. Betts , C.M., Stephen M. Kelly , Stephen J. Haswell, Synergistic antibacterial effects of theaflavin in combination with ampicillin against hospital isolates of Strenotrophomonas maltophilia. 2012.. J Microbiol Biotech Food Sci, p. 1068-78.
476. Hwang, I.S., et al., Synergistic effect and antibiofilm activity between the antimicrobial peptide coprisin and conventional antibiotics against opportunistic bacteria. Curr Microbiol, 2013. 66(1): p. 56-60.
477. Mazumdar, K., et al., In vitro and in vivo synergism between tetracycline and the cardiovascular agent oxyfedrine HCl against common bacterial strains. Biol Pharm Bull, 2005. 28(4): p. 713-7.
478. Mosmann, T., Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. J Immunol Methods, 1983. 65(1-2): p. 55-63.
479. Bodet, C., F. Chandad, and D. Grenier, Modulation of cytokine production by Porphyromonas gingivalis in a macrophage and epithelial cell co-culture model. Microbes Infect, 2005. 7(3): p. 448-56.
480. Bodet, C., F. Chandad, and D. Grenier, Inflammatory responses of a macrophage/epithelial cell co-culture model to mono and mixed infections with Porphyromonas gingivalis, Treponema denticola, and Tannerella forsythia. Microbes Infect, 2006. 8(1): p. 27-35.
481. Gursoy, U.K., et al., Biofilm formation enhances the oxygen tolerance and invasiveness of Fusobacterium nucleatum in an oral mucosa culture model. J Periodontol, 2010. 81(7): p. 1084-91.
482. Gursoy, U.K., et al., A novel organotypic dento-epithelial culture model: effect of Fusobacterium nucleatum biofilm on B-defensin-2, -3, and LL-37 expression. J Periodontol, 2012. 83(2): p. 242-7.
483. Pihlstrom, B.L., B.S. Michalowicz, and N.W. Johnson, Periodontal diseases. The Lancet. 366(9499): p. 1809-20.
484. Hilal, Y. and U. Engelhardt, Characterisation of white tea – Comparison to green and black tea. Journal für Verbraucherschutz und Lebensmittelsicherheit, 2007. 2(4): p. 414-21.
485. Cui, Y., et al., AFM study of the differential inhibitory effects of the green tea polyphenol (-)-epigallocatechin-3-gallate (EGCG) against Gram-positive and Gram-negative bacteria. Food Microbiol, 2012. 29(1): p. 80-7.
486. Sudano Roccaro, A., et al., Epigallocatechin-gallate enhances the activity of tetracycline in staphylococci by inhibiting its efflux from bacterial cells. Antimicrob Agents Chemother, 2004. 48(6): p. 1968-73.
487. Hisano, M., et al., Inhibitory effect of catechin against the superantigen staphylococcal enterotoxin B (SEB). Arch Dermatol Res, 2003. 295(5): p. 183-9.
488. Hull Vance, S., M. Tucci, and H. Benghuzzi, Evaluation of the antimicrobial efficacy of green tea extract (egcg) against streptococcus pyogenes in vitro. Biomed Sci Instrum, 2011. 47: p. 177-82.

138
489. Sakanaka, S., L.R. Juneja, and M. Taniguchi, Antimicrobial effects of green tea polyphenols on thermophilic spore-forming bacteria. J Biosci Bioeng, 2000. 90(1): p. 81-5.
490. Yoda, Y., et al., Different susceptibilities of Staphylococcus and Gram-negative rods to epigallocatechin gallate. J Infect Chemother, 2004. 10(1): p. 55-8.
491. Lee, K.M., et al., Antipathogenic properties of green tea polyphenol epigallocatechin gallate at concentrations below the MIC against enterohemorrhagic Escherichia coli O157:H7. J Food Prot, 2009. 72(2): p. 325-31.
492. Sirk, T.W., M. Friedman, and E.F. Brown, Molecular binding of black tea theaflavins to biological membranes: relationship to bioactivities. J Agric Food Chem, 2011. 59(8): p. 3780-7.
493. Yi, S.-m., et al., Tea polyphenols inhibit Pseudomonas aeruginosa through damage to the cell membrane. International J Food Microbiol, 2010. 144(1): p. 111-7.
494. Navarro-Martínez, M.D., et al., Antifolate activity of epigallocatechin gallate against Stenotrophomonas maltophilia. antimicrobial agents and chemotherapy, 2005. 49(7): p. 2914-20.
495. Spina, M., et al., Mechanism of inhibition of wt-dihydrofolate reductase from E. coli by tea epigallocatechin-gallate. Proteins: Struct Funct Bioinf, 2008. 72(1): p. 240-251.
496. Denes, É. and N. Hidri, Synergie et antagonisme en antibiothérapie. Antibiotiques, 2009. 11(2): p. 106-115.
497. Page, C.P., Pharmacologie intégrée. 1999, Paris ; Bruxelles: De Boeck Université. x, 606 p. 498. Muller, M., Mode of action of metronidazole on anaerobic bacteria and protozoa. Surgery,
1983. 93(1 Pt 2): p. 165-71. 499. Chopra, I. and M. Roberts, Tetracycline antibiotics: mode of action, applications, molecular
biology, and epidemiology of bacterial resistance. Microbiol Mol Biol Rev, 2001. 65(2): p. 232-60.
500. Seymour, R.A. and P.A. Heasman, Tetracyclines in the management of periodontal diseases. A review. J Clin Periodontol, 1995. 22(1): p. 22-35.
501. Fanaki, N.H., et al., Influence of aqueous green tea extract on the antimicrobial activity of some antibiotics against multiresistant clinical isolates. Egypt. J Med Microbiol, 2008. 17: p. 449-60.
502. Haghjoo, B., et al., The synergistic effects of green tea polyphenols and antibiotics against potential pathogens. Adv Biosci Biotechnol, 2013. 4(11): p. 959.
503. Hu, Z.-Q., et al., Epigallocatechin gallate synergistically enhances the activity of carbapenems against methicillin-resistant Staphylococcus aureus. Antimicrob agents Chemother, 2002. 46(2): p. 558-60.
504. Hu, Z.-Q., et al., Epigallocatechin gallate synergy with ampicillin/sulbactam against 28 clinical isolates of methicillin-resistant Staphylococcus aureus. J Antimicrob Chemother, 2001. 48(3): p. 361-4.
505. Zhao, W.-H., et al., Mechanism of synergy between epigallocatechin gallate and β-lactams against methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother, 2001. 45(6): p. 1737-42.
506. Stapleton, P.D., et al., Potentiation of catechin gallate-mediated sensitization of Staphylococcus aureus to oxacillin by nongalloylated catechins. Antimicrob Agents Chemother, 2006. 50(2): p. 752-5.
507. Cui, Y., et al., AFM Probing the mechanism of synergistic effects of the green tea polyphenol (−)-epigallocatechin-3-gallate (EGCG) with cefotaxime against extended-spectrum beta-lactamase (ESBL)-producing Escherichia coli. PLoS ONE, 2012. 7(11): p. e48880.

139
508. Henderson, B., S. Poole, and M. Wilson, Bacterial modulins: a novel class of virulence factors which cause host tissue pathology by inducing cytokine synthesis. Microbiol Rev, 1996. 60(2): p. 316-41.
509. Schwartz, Z., et al., Mechanisms of alveolar bone destruction in periodontitis. Periodontol 2000, 1997. 14: p. 158-72.
510. AlRowis, R., et al., Oral fluid-based biomarkers in periodontal disease – Part 2. Gingival crevicular fluid. J Inter Oral Health : JIOH, 2014. 6(5): p. 126-35.
511. Yang, G.Y., et al., Inhibition of growth and induction of apoptosis in human cancer cell lines by tea polyphenols. Carcinogenesis, 1998. 19(4): p. 611-6.
512. Ahmad, N., et al., Green tea constituent epigallocatechin-3-gallate and induction of apoptosis and cell cycle arrest in human carcinoma cells. J Natl Cancer Inst, 1997. 89(24): p. 1881-6.
513. Paschka, A.G., R. Butler, and C.Y. Young, Induction of apoptosis in prostate cancer cell lines by the green tea component, (-)-epigallocatechin-3-gallate. Cancer Lett, 1998. 130(1-2): p. 1-7.
514. Taniguchi, S., et al., Effect of (-)-epigallocatechin gallate, the main constituent of green tea, on lung metastasis with mouse B16 melanoma cell lines. Cancer Lett, 1992. 65(1): p. 51-4.
515. Kong, L., et al., Theaflavins inhibit pathogenic properties of P. gingivalis and MMPs production in P. gingivalis-stimulated human gingival fibroblasts. Arch Oral Biol, 2015. 60(1): p. 12-22.
516. Bodet, C., F. Chandad, and D. Grenier, Inhibition of host extracellular matrix destructive enzyme production and activity by a high-molecular-weight cranberry fraction. J Periodontal Res, 2007. 42(2): p. 159-68.
517. Bodet, C., F. Chandad, and D. Grenier, Cranberry components inhibit interleukin-6, interleukin-8, and prostaglandin E production by lipopolysaccharide-activated gingival fibroblasts. Eur J Oral Sci, 2007. 115(1): p. 64-70.
518. La, V.D., A.B. Howell, and D. Grenier, Cranberry proanthocyanidins Inhibit MMP production and activity. J Dent Res, 2009. 88(7): p. 627-32.
519. Ben Lagha, A., et al., Wild Blueberry (Vaccinium angustifolium Ait.) polyphenols target Fusobacterium nucleatum and the host inflammatory response: Potential innovative molecules for treating periodontal diseases. J Agric Food Chem, 2015. 63(31): p. 6999-7008.
520. Matchett, M.D., et al., Inhibition of matrix metalloproteinase activity in DU145 human prostate cancer cells by flavonoids from lowbush blueberry (Vaccinium angustifolium): possible roles for protein kinase C and mitogen-activated protein-kinase-mediated events. J Nutr Biochem. 17(2): p. 117-25.
521. Marquis, A., et al., The plant coumarins auraptene and lacinartin as potential multifunctional therapeutic agents for treating periodontal disease. BMC Complement Altern Med, 2012. 12: p. 80.
522. La, V.D., et al., Modulation of matrix metalloproteinase and cytokine production by licorice isolates licoricidin and licorisoflavan A: potential therapeutic approach for periodontitis. J Periodontol, 2011. 82(1): p. 122-8.
523. Grenier, D., et al., Dual Action of Myricetin on Porphyromonas gingivalis and the inflammatory response of host cells: A Promising Therapeutic Molecule for periodontal diseases. PLoS One, 2015. 10(6): p. e0131758.

140

141
9. ANNEXES
Tableau 1. Polyphénols à l'étude et méthode de préparation
Polyphénols Préparation de la solution mère
Thé vert
Extrait de thé vert "GT9" 10 mg/ml dans H2O distillée et filtration (0,2 µm)
EGCG 10 mg/ml dans H2O distillée et filtration (0,2 µm)
Tableau 2. Synthèse des tests de viabilité MTT
Composantes
Monocytes de la Lignée U937 différenciés en Macrophages
Concentrations maximales (µg/ml) montrant une baisse de viabilité de ≤ 20%
Test 1 Test 2 Test 3 Test 4
Thé vert 62,5 62,5 100 62,5
EGCG 62,5 62,5 125 62,5
Composantes
Fibroblastes gingivaux humains
Concentrations maximales (µg/ml) montrant une baisse de viabilité de ≤ 20%
Test 1 Test 2 Test 3 Test 4
Thé vert 62,5 62,5 62,5 -
EGCG ≥ 125 62,5 62,5

142
Au cours de mes études de deuxième cycle, j’ai été amené à collaborer à deux projets qui ont mené à
des publications scientifiques. Mon implication dans ces projets portait sur la détermination de CMIs,
CMBs, ainsi que des analyses de synergie
Publication scientifique 1:
Marie-Pierre Morin, Telma Bedran, Jade Fournier-Larente, Bruno Haas, Jabrane Azelmat and Daniel Grenier,
Green tea extract and its major constituent epigallocatechin-3-gallate inhibit growth and halitosis-related
properties of Solobacterium moorei. BMC Complementary and Alternative Medicine. 2015
ABSTRACT:
BACKGROUND
Solobacterium moorei is a volatile sulfide compound (VSC)-producing Gram-positive anaerobic bacterium that
has been associated with halitosis. The aim of this study was to investigate the effects of green tea extract and
its major constituent epigallocatechin-3-gallate (EGCG) on growth and several halitosis-related properties of S.
moorei.
METHODS
A microplate dilution assay was used to determine the antibacterial activity of green tea extract and EGCG
against S. moorei. Their effects on bacterial cell membrane integrity were investigated by transmission electron
microscopy and a fluorescence-based permeability assay. Biofilm formation was quantified by crystal violet
staining. Adhesion of FITC-labeled S. moorei to oral epithelial cells was monitored by fluorometry. The
modulation of β-galactosidase gene expression in S. moorei was evaluated by quantitative RT-PCR.
RESULTS
The green tea extract as well as EGCG inhibited the growth of S. moorei, with MIC values of 500 and 250 μg/ml,
respectively. Transmission electron microscopy analysis and a permeabilization assay brought evidence that
the bacterial cell membrane was the target of green tea polyphenols. Regarding the effects of green tea
polyphenols on the S. moorei colonization properties, it was found that biofilm formation on EGCG-treated
surfaces was significantly affected, and that green tea extract and EGCG can cause the eradication of pre-
formed S. moorei biofilms. Moreover, both the green tea extract and EGCG were found to reduce the adherence
of S. moorei to oral epithelial cells. The β-galactosidase activity of S. moorei, which plays a key role in VSC
production, was dose-dependently inhibited by green tea polyphenols. In addition, EGCG at ½ MIC significantly
decreased the β-galactosidase gene expression.
CONCLUSION
Our study brought evidence to support that green tea polyphenols possess a number of properties that may
contribute to reduce S. moorei-related halitosis. Therefore, these natural compounds may be of interest to be
used to supplement oral healthcare products.

143
Publication scientifique 2 :
Daniel Grenier, Marie-Pierre Morin, Jade Fournier-Larente, Huangqin Chen, Vitamin D inhibits the growth of
and virulence factor gene expression by Porphyromonas gingivalis and blocks activation of the nuclear factor
kappa B transcription factor in monocytes. Journal of Periodontal Research 2015; doi: 10.1111/jre.12315.
ABSTRACT:
BACKGROUND AND OBJECTIVE
Increasing evidence suggests that 1,25-dihydroxyvitamin D3(1,25(OH)2 D3 ), a fat-soluble secosteroid
hormone, has a positive impact on periodontal health through diverse mechanisms. The present study was
aimed at investigating the effect of 1,25(OH)2D3 on the growth of and virulence factor gene expression by the
periodontopathogenic bacterium Porphyromonas gingivalis. The effect of 1,25(OH)2 D3 on P. gingivalis-
mediated activation of nuclear factor kappa B (NF-κB) transcription factor in monocytes was also assessed.
MATERIAL AND METHODS
A broth microdilution assay was used to determine the antibacterial activity of 1,25(OH)2 D3 . The modulation of
virulence factor gene expression in P. gingivalis was assessed by quantitative reverse transcription-polymerase
chain reaction. NF-κB activation was assessed using a human monocytic cell line stably transfected with a
luciferase reporter containing NF-κB binding sites.
RESULTS
Minimal inhibitory concentrations of 1,25(OH)2 D3 against P. gingivalis ranged from 3.125 to 6.25 μg/ml.
Moreover, a partial synergistic effect was observed when 1,25(OH)2 D3 was used in association with
metronidazole. 1,25(OH)2 D3 attenuated the virulence of P. gingivalis by reducing the expression of genes
coding for important virulence factors, including adhesins (fimA, hagA and hagB) and proteinases (rgpA, rgpB
and kgp). 1,25(OH)2 D3 dose-dependently prevented P. gingivalis-induced NF-κB activation in a monocyte
model.
CONCLUSION
Our study suggested that 1,25(OH)2 D3 selectively inhibits the growth of and virulence factor gene expression
by P. gingivalis, in addition to attenuating NF-κB activation by this periodontopathogen. This dual action on
P. gingivalis and the inflammatory response of host cells may be of particular interest with a view to developing
a novel and inexpensive preventive/therapeutic strategy.

144















![Thé Vert [en français] (por: mujtabaalirazmi / carlitosrangel)](https://static.fdocuments.fr/doc/165x107/5582f509d8b42a2c168b4cdd/the-vert-en-francais-por-mujtabaalirazmi-carlitosrangel.jpg)

