LA PROCRÉATION MÉDICALEMENT ASSISTÉE (PMA) - BioDj...
14
40 La procréation médicalement assistée est l’ensemble des méthodes permettant d’induire une grossesse en dehors de l’union naturelle de l’homme et de la femme, en particulier l’insémination artificielle, la fécondation in vitro avec transfert d’embryons (FIVETE) et la fécondation par micro injection. Louise Brown, de nationalité britannique, née le 25 juillet 1978 est le premier « bébé éprouvette » au monde, c'est-à-dire conçu par fécondation in vitro (FIV). La naissance de Louise a ouvert un nouveau chapitre de la médecine relatif au traitement de la stérilité. Depuis cette naissance, les techniques de procréation médicalement assistée se sont banalisées. L'infertilité peut être définie comme «l'incapacité pour un couple de procréer ou de mener une grossesse à terme après un an ou plus de rapports sexuels réguliers non protégés». Dans le monde, on estime qu'un couple sur six est stérile. La prévalence de l'infertilité est pratiquement la même chez l'homme et chez la femme. Les troubles de l'ovulation sont la cause la plus courante d'infertilité féminine. Des anomalies anatomiques (trompes de Fallope bouchées, infectées…..) comptent également pour une grande part des cas d'infertilité. Chez l'homme, ce sont les troubles de la production de spermatozoïdes qui constituent la cause la plus fréquente d'infertilité. Des obstructions anatomiques et des problèmes immunitaires sont également à l'origine de cas d'infertilité masculine. Pour la femme comme pour l'homme, les options de traitement varient en fonction du type d'infertilité diagnostiqué. Elles comprennent la thérapie médicamenteuse, la chirurgie et les techniques de procréation médicalement assistée (PMA). Les taux de succès des traitements sont fonction de divers facteurs essentiels, notamment, l'âge de la femme, la durée de l'infertilité du couple et l'existence de problèmes d'infertilité chez les deux partenaires. Parmi les femmes ayant suivi un traitement d'induction de l'ovulation (thérapie médicamenteuse), 80% conçoivent un enfant après plusieurs cycles de traitement. Les taux de réussite des techniques de PMA sont en constante progression depuis dix ans. Actuellement, on peut s'attendre à un taux de réussite d'environ 25% par cycle pour une fécondation in vitro pratiquée dans des conditions optimales ce qui correspond au taux naturel de réussite. 5.1 Les techniques 5.1.1 L’insémination artificielle Cette technique (figure 24) consiste à injecter, à l’aide d’une pipette, les spermatozoïdes, recueillis après masturbation, directement dans l’utérus, le jour de l’ovulation. Dans les cas de stérilité masculine importante, l’insémination peut se faire avec le sperme d’un donneur. 5.1.2 La Fécondation In Vitro Et Transfert d’Embryon (FIVETE) La fécondation in vitro et transfert d’embryon (figure 25), comme son nom l’indique, va permettre de mettre en présence, 5 LA PROCRÉATION MÉDICALEMENT ASSISTÉE (PMA) Figure 24 : L’insémination artificielle
-
Upload
truongcong -
Category
Documents
-
view
216 -
download
0
Transcript of LA PROCRÉATION MÉDICALEMENT ASSISTÉE (PMA) - BioDj...

40
La procréation médicalement assistée est l’ensemble des méthodes permettant d’induire une grossesse en dehors de l’union naturelle de l’homme et de la femme, en particulier l’insémination artificielle, la fécondation in vitro avec transfert d’embryons (FIVETE) et la fécondation par micro injection.
Louise Brown, de nationalité britannique, née le 25 juillet 1978 est le premier « bébé éprouvette » au monde, c'est-à-dire conçu par fécondation in vitro (FIV). La naissance de Louise a ouvert un nouveau chapitre de la médecine relatif au traitement de la stérilité. Depuis cette naissance, les techniques de procréation médicalement assistée se sont banalisées.
L'infertilité peut être définie comme «l'incapacité pour un couple de procréer ou de mener une grossesse à terme après un an ou plus de rapports sexuels réguliers non protégés». Dans le monde, on estime qu'un couple sur six est stérile. La prévalence de l'infertilité est pratiquement la même chez l'homme et chez la femme.
Les troubles de l'ovulation sont la cause la plus courante d'infertilité féminine. Des anomalies anatomiques (trompes de Fallope bouchées, infectées…..) comptent également pour une grande part des cas d'infertilité.
Chez l'homme, ce sont les troubles de la production de spermatozoïdes qui constituent la cause la plus fréquente d'infertilité. Des obstructions anatomiques et des problèmes immunitaires sont également à l'origine de cas d'infertilité masculine.
Pour la femme comme pour l'homme, les options de traitement varient en fonction du type d'infertilité diagnostiqué. Elles comprennent la thérapie médicamenteuse, la chirurgie et les techniques de procréation médicalement assistée (PMA).
Les taux de succès des traitements sont fonction de divers facteurs essentiels, notamment, l'âge de la femme, la durée de l'infertilité du couple et l'existence de problèmes d'infertilité chez les deux partenaires. Parmi les femmes ayant suivi un traitement d'induction de l'ovulation (thérapie médicamenteuse), 80% conçoivent un enfant après plusieurs cycles de traitement. Les taux de réussite des techniques de PMA sont en constante progression depuis dix ans. Actuellement, on peut s'attendre à un taux de réussite d'environ 25% par cycle pour une fécondation in vitro pratiquée dans des conditions optimales ce qui correspond au taux naturel de réussite.
5.1 Les techniques 5.1.1 L’insémination artificielle
Cette technique (figure 24) consiste à injecter, à l’aide d’une pipette, les spermatozoïdes, recueillis après masturbation, directement dans l’utérus, le jour de l’ovulation. Dans les cas de stérilité masculine importante, l’insémination peut se faire avec le sperme d’un donneur.
5.1.2 La Fécondation In Vitro Et
Transfert d’Embryon (FIVETE)
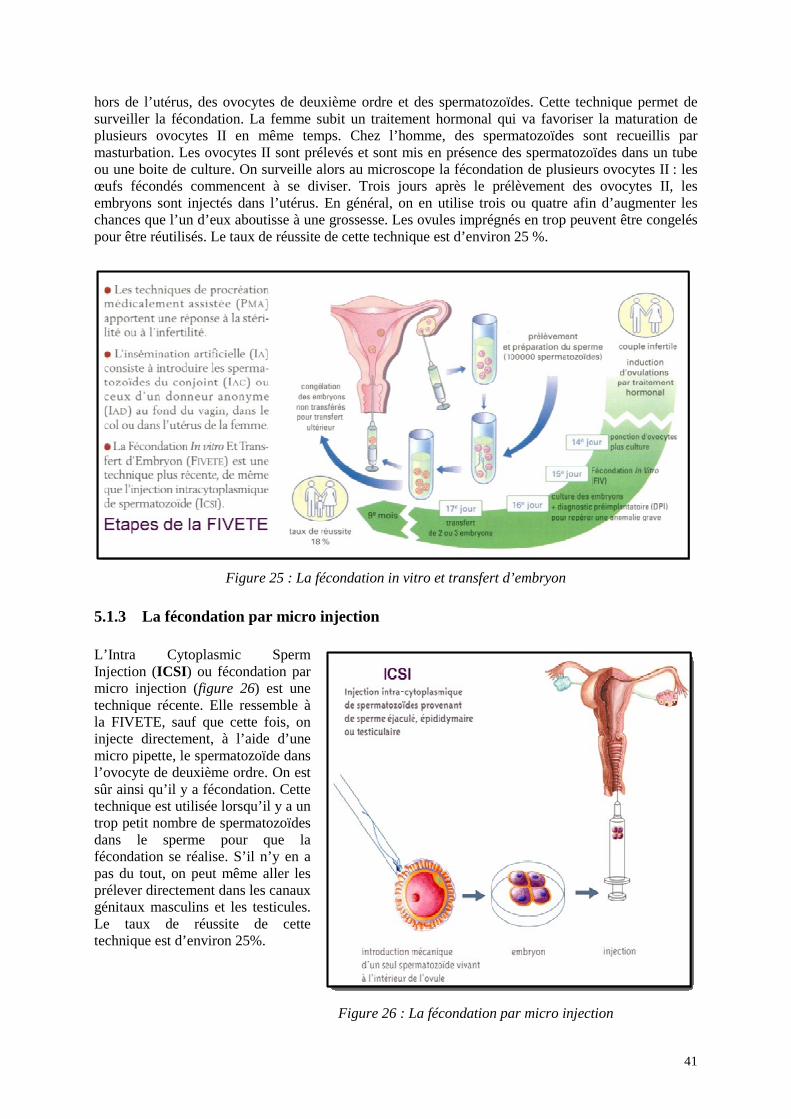
La fécondation in vitro et transfert d’embryon (figure 25), comme son nom l’indique, va permettre de mettre en présence,
5 LA PROCRÉATION MÉDICALEMENT ASSISTÉE (PMA)
Figure 24 : L’insémination artificielle
41
hors de l’utérus, des ovocytes de deuxième ordre et des spermatozoïdes. Cette technique permet de surveiller la fécondation. La femme subit un traitement hormonal qui va favoriser la maturation de plusieurs ovocytes II en même temps. Chez l’homme, des spermatozoïdes sont recueillis par masturbation. Les ovocytes II sont prélevés et sont mis en présence des spermatozoïdes dans un tube ou une boite de culture. On surveille alors au microscope la fécondation de plusieurs ovocytes II : les œufs fécondés commencent à se diviser. Trois jours après le prélèvement des ovocytes II, les embryons sont injectés dans l’utérus. En général, on en utilise trois ou quatre afin d’augmenter les chances que l’un d’eux aboutisse à une grossesse. Les ovules imprégnés en trop peuvent être congelés pour être réutilisés. Le taux de réussite de cette technique est d’environ 25 %.
Figure 25 : La fécondation in vitro et transfert d’embryon
5.1.3 La fécondation par micro injection L’Intra Cytoplasmic Sperm Injection (ICSI ) ou fécondation par micro injection (figure 26) est une technique récente. Elle ressemble à la FIVETE, sauf que cette fois, on injecte directement, à l’aide d’une micro pipette, le spermatozoïde dans l’ovocyte de deuxième ordre. On est sûr ainsi qu’il y a fécondation. Cette technique est utilisée lorsqu’il y a un trop petit nombre de spermatozoïdes dans le sperme pour que la fécondation se réalise. S’il n’y en a pas du tout, on peut même aller les prélever directement dans les canaux génitaux masculins et les testicules. Le taux de réussite de cette technique est d’environ 25%.
Figure 26 : La fécondation par micro injection

42
La PMA pose des problèmes beaucoup plus généraux tels que le diagnostic préimplantatoire , les embryons surnuméraires, le clonage humain. Actuellement, les techniques de PMA ne sont pas optimales et c'est pourquoi il existe des taux d'échecs non négligeables.
La recherche sur les embryons surnuméraires permettrait :
- une optimisation du diagnostic préimplantatoire. Cette technique est autorisée alors qu'elle n'est pas optimale. Des recherches permettraient donc de rendre vraiment fiable cette méthode afin d'implanter in utero des embryons viables.
- une amélioration de la réimplantation des embryons. En effet cette technique ne connaît pas un bon taux de réussite. Les chercheurs souhaiteraient donc faire des travaux sur différents milieux pour l'améliorer. Cela permettrait de réimplanter moins d'embryons in utero et de réduire ainsi les grossesses multiples.
La loi CH sur la PMA se trouve sur le lien suivant : www.admin.ch/ch/f/as/2000/3055.pdf

43
Tout a débuté avec la culture de tissus au début du XXe siècle. Puis après avoir perfection cette technique, la culture de cellules est apparue 50 ans plus tard. Il aura fallu la trypsinisation des tissus qui consiste à digérer ceux-ci à l’aide de la trypsine pour obtenir des cellules isolées ou des amas de cellules capables de se diviser in vitro.
La culture de tissus est le terme général englobant le prélèvement de cellules, tissus ou organes d’un animal ou d’une plante et leur placement ultérieur dans un environnement artificiel conduisant à leur croissance.
6.1 Culture cellulaire La culture cellulaire est devenue un des outils majeurs utilisés aujourd’hui dans les sciences de la vie. La culture cellulaire est le maintien en dehors de l'organisme, des cellules non organisées en tissu mais capable de se diviser in vitro et d'exprimer des métabolismes et des fonctions spécifiques. On distingue 2 types de cellules:
� Les cellules libres et circulantes comme les cellules du sang.
� Les cellules en cohésion les unes avec les autres, constituant un tissu.
Les techniques d'obtention de ces 2 classes de cellules sont différentes.
• Les cellules circulantes sont obtenues par prélèvement et centrifugation.
• Les cellules organisées en tissus nécessitent la mise en œuvre de techniques plus originales qui peuvent être divisées en 2 groupes:
� La méthode des explants
Cette méthode est la plus ancienne. Elle a permis aux précurseurs de la culture de tissu d'obtenir les premières cellules in vitro.
De petits morceaux de tissus sont fixés sur un récipient de culture en verre ou en plastique traité et baignés dans du milieu de culture. Après quelques jours, des cellules individuelles se déplacent de l’explant de tissus vers la surface du récipient de culture ou le substrat où elles commencent à se diviser et proliférer.
� La méthode par digestion enzymatique.
C’est la méthode la plus généralement utilisée. Les enzymes utilisées sont des enzymes protéolytiques qui digèrent la trame protéique qui entoure les cellules dans le tissu. On utilise souvent la trypsine.
Nous obtenons une suspension de cellules individuelles qui sont placées dans des récipients de culture contenant un milieu de culture pour les laisser pousser et se diviser.
Les avantages et les inconvénients de ces méthodes sont :
6 CULTURE DE CELLULES ET TISSUS

44
- La méthode des explants est souvent utilisée quand le tissu à mettre en culture est très petit. L'obtention nécessaire pour avoir des couches cellulaires confluentes est relativement long (environ 30 jours).
- La méthode enzymatique est beaucoup plus rapide avec un bon rendement mais certaines cellules à membrane fragile peuvent être lésées par cette méthode.
Les méthodes de culture:
� La culture stationnaire ou monocouche:
Le principe général de cette méthode est lié à l'affinité des cellules au support car la plupart appartient à des tissus solides organisés. Le support peut être du verre ou du plastique traité pour la culture. Le plastique peut être recouvert de support physiologique: collagène, fibronectine ou d'une membrane basale reconstituée ou encore l'utilisation de gel d'agarose, de gel de collagène pour une orientation. Les cellules sont placées dans des boites micropuits. Le rendement de cette technique est important. Cela étant dû à une surface d'adhérence plus grande.
� La culture en suspension:
Comme la plupart des cellules ont besoin d'un support pour croître, il faut donc trouver un artifice pour maintenir la plupart de celles-ci en suspension. En 1933, on arrive à faire croître dans des tubes en verres, des cellules en suspension tournant à grande vitesse.
Il faut contrôler la fonctionnalité des cellules en culture ; pour cela, on doit considérer deux caractéristiques :
• La prolifération des cellules.
• La préservation des fonctions spécialisées. L'évolution des cellules en culture: Les cellules in vitro présentent 2 propriétés fondamentales qui sont: la capacité proliférative et leur fonction différentiée. Ces propriétés ont tendance à évoluer de manières très différentes quelle que soit la méthode de culture employée. Les cellules conservent la plupart du temps leur potentiel de division, pouvant être stimulé au début de la culture par des facteurs de croissances, mais celui-ci peut noter un ralentissement au cours du temps. A l'inverse, les cellules en culture voient souvent leur fonction différenciée se modifier et même disparaître. Ce phénomène de dédifférenciation peut être visible morphologiquement.

45
6.2 Culture de tissus végétaux Les cultures in vitro végétales sont des cultures d’explants de plantes, sur un milieu synthétique, dans des conditions stériles, dans un environnement contrôlé et dans un espace réduit. On parle de culture de tissus lorsque les cellules restent liées entre elles après les multiples divisions. La culture in vitro permet d'obtenir une quantité importante d'individus tous génétiquement identiques à l'aide, en théorie, d'une seule cellule végétale. La multiplication végétative in vitro consiste à prélever un fragment d'organe de la plante, à le placer dans un milieu approprié de façon à ce que ce fragment puisse régénérer une plante entière. Le fragment peut être un morceau de feuille, de tige, un bourgeon... La multiplication végétative est effectuée en éprouvette ou dans un récipient analogue. Ce type de multiplication est basé sur une propriété importante des cellules végétales, inexistante chez les cellules animales. Une cellule végétale différenciée peut redevenir indifférenciée, se diviser à nouveau activement et puis se différencier à nouveau en une cellule, peut-être tout à fait différente de ce qu'elle était avant. Cette propriété est nommée la totipotence de la cellule végétale. Toutes les cellules d'un végétal sont donc capables d'exprimer toutes les informations du code génétique contenu dans leur ADN. C'est un phénomène qui n'est en aucun cas valable chez les Animaux; en effet lorsque la zygote se développe, seules les toutes premières cellules sont totipotentes mais bien vite ce potentiel de la cellule diminue pour engendrer les cellules spécialisées au bout de quelques stades et pourtant toutes les cellules possèdent l'ensemble du code génétique. La Technique :
1. Des cellules isolées ou des tissus sont prélevés dans une feuille ou un autre organe et maintenus en vie sur un substrat nutritif.
2. On stérilise la surface du tissu prélevé, et l'on dépose ce fragment de tissu vivant sur un milieu
nutritif solidifié avec de l'agar-agar.
3. Les tissus dépourvus de chloroplastes sont hétérotrophes; par conséquent le substrat doit contenir, en plus des sels minéraux indispensables, une certaine quantité de sucre sous forme de saccharose ou de glucose.
4. Souvent il faut ajouter des hormones végétales, des phytohormones, qui favorisent la
croissance et la division cellulaire.
5. La masse cellulaire forme à la surface un cal constitué d'un amas de cellules indifférenciées. Si l'on attend trop longtemps, ce cal présentera des processus de différenciation. Mais on peut prélever sur ce cal des cellules encore non différenciées pour les cultiver séparément. La dissociation des cellules calleuses est réalisée par suspension dans un milieu liquide.
6. On récupère les cellules isolées par filtration à travers une gaze. Des cals formés à partir d'une
seule cellule peuvent être cultivés à l'obscurité ou à la lumière. En faisant varier les concentrations et les combinaisons des phytohormones, on peut induire la formation d'organes.
7. Quand les plantules présentent des radicelles et de petites feuilles, elles sont prêtes à être
transplantées. Elles forment un clone.

46
Figure 27 : Culture de tissus végétaux "in vitro"
Cette technique de clonage est remarquable pour ses rendements (production de plantes homogènes en grand nombre). Un simple bourgeon de rosier permet de produire jusqu'à 400'000 nouveaux rosiers génétiquement identiques. Le clonage est une stratégie de multiplication d'abord naturelle. La reproduction du fraisier, des pommes de terre, des jonquilles (par bulbes), n'est rien d'autre que du clonage. La présence de la paroi pectocellulosique des cellules est une des barrières aux échanges d'information génétique. On peut séparer les cellules d'un tissu végétal grâce à l'action d'enzymes généralement extraites de champignons, qui dégradent la cellulose de la paroi. Des agents stabilisants sont ajoutés au milieu pour empêcher l'éclatement de la cellule. On obtient ainsi des cellules sans paroi qui deviennent sphériques : les protoplastes. Ceux-ci peuvent ensuite être utilisés expérimentalement comme s'il s'agissait de cellules animales. Les protoplastes peuvent être obtenus à partir de n'importe quel tissu végétal, mais ce sont généralement les jeunes feuilles qui sont utilisés pour leur préparation. Si les conditions de milieu sont favorables, la paroi végétale se reconstitue. Les organites cellulaires se réarrangent et les cellules entrent en division. Elles donnent ainsi naissance à des microcolonies, puis des cals, amas de cellules indifférenciées. Transférés sur un milieu de régénération, les cals se développent en embryons qui donneront des plantules. S'il est apparemment possible d'obtenir des protoplastes chez toutes les espèces végétales, leur culture puis la régénération d'une plante entière à partir des protoplastes posent encore de nombreuses difficultés et constituent une limite de cette technique. Il semble notamment que les monocotylédones (céréales) soient plus récalcitrantes vis-à-vis de la culture des protoplastes que les dicotylédones. Les protoplastes sont donc un matériel privilégié pour le transfert direct de gènes. Débarrassées de leur paroi pectocellulosique, ces cellules ne présentent plus d'obstacle à l'intégration de l'ADN. Quelques utilisations des protoplastes :
A. L'hybridation somatique.
La propriété la plus importante des protoplastes est leur capacité à fusionner entre eux lorsqu'ils sont placés dans un milieu approprié. Cette technique permet de surmonter les

47
barrières liées à la reproduction sexuée et de créer de nouvelles combinaisons entre noyau et cytoplasme. De telles hybridations somatiques ont été réalisées entre espèces différentes (à l’intérieur d’un genre) et incompatibles sur le plan de la reproduction, par exemple Petunia, Daucus (carotte) et Solanum (pomme de terre, tomate, aubergine). Le but de telles expériences est de créer des plantes à très grande productivité, combinant plusieurs qualités: résistances supplémentaires, et à l'avenir fixation d'azote atmosphérique.
Figure 28 : L'hybridation somatique
Cette technique d'hybridation est illustrée par le tabac (ci-dessus). Des cellules des feuilles (riches en chloroplastes) de deux sortes de tabac sont traitées par des enzymes qui dissolvent la paroi cellulaire. Les protoplastes qui en résultent sont placés dans une solution de polyéthèneglycol qui diminue la tension superficielle, ce qui facilite la fusion. Après centrifugation, les cellules sont placées sur un milieu de culture spécial sur lequel seules les cellules hybrides vont se multiplier. Plus tard, elles se différencieront en tige et feuilles. On les greffe alors sur un des plants parents. Les graines de ces hybrides germent, se développent en jeunes pousses puis en plantes adultes. Dans ces dernières, les génomes des deux plantes de départ sont présents. La première démonstration de fusion entre des protoplastes différents remonte aux travaux de Melchers en 1978. Il recherchait des tomates cultivables à basse température et réalisa, à cette fin, des hybrides entre la tomate et la pomme de terre par fusion de protoplastes : la pomate. Cette nouvelle espèce est malheureusement un exemple théorique, car elle est stérile. La pomme de terre cultivée (Solanum tuberosum) est une espèce chez laquelle l'introduction de caractères par fusion de protoplastes est facilement réalisable. Ainsi, on a pu introduire des gènes de résistance à certains virus, au mildiou et à la pourriture bactérienne, à partir des espèces sauvages d'Amérique du Sud, notamment Solanum brevidens. De nouvelles lignées mâles stériles de colza résistantes à l'atrazine (herbicide) ont pu également être obtenues par cette technique.

48
Au cours de la reproduction sexuée, les informations génétiques contenues dans le cytoplasme (mitochondrie) sont transmises par la mère. En revanche, la fusion de protoplastes conduit à une hybridation des noyaux, mais aussi à celle des cytoplasmes. Ceci est très intéressant pour le transfert et l'amélioration de caractères à hérédité cytoplasmique, comme la stérilité mâle. On parle alors d'hybridation somatique (car issue de cellules non reproductrices de la plante). Les protoplastes sont des cellules chargées négativement et la fusion spontanée n'est que très rarement observée. La fusion peut être obtenue de deux manières :
� Grâce à divers agents chimiques (cations Ca2+ et pH élevé) on peut neutraliser la charge électrique des protoplastes. On utilise ensuite le polyéthylène glycol (PEG) qui provoque une forte agrégation des cellules et déstabilise la membrane plasmique. Après retour aux conditions initiales, les protoplastes fusionnent.
� L'électrofusion, plus récente, utilise des champs électriques intenses et de courte durée, qui en
déstabilisant les membranes entraînent la fusion des protoplastes. Ce système semble être plus efficace.
Figure 29 : Techniques de fusion des protoplastes
Lors de la fusion, tous les échanges sont possibles entre deux protoplastes. On peut ainsi obtenir des degrés de fusion très variables :
• Fusion des noyaux et des cytoplasmes : lorsque la fusion des noyaux a lieu, il peut y avoir une recombinaison plus ou moins importante entre les chromosomes des deux parents. Ce phénomène peut être utilisé pour transférer des gènes nucléaires. On cherchera notamment à obtenir des hybrides somatiques asymétriques, où seuls quelques fragments d'ADN du parent donneur seront introduits dans l'espèce receveuse. En effet, les cas de fusion importante de génomes entre espèces conduisent à des plantes souvent stériles comme la pomate. Pour favoriser ce transfert partiel, l'ADN du parent donneur est déstabilisé par irradiation ménagée des protoplastes avant la fusion.
• Fusion unique des cytoplasmes : les cybrides : très souvent, la fusion des noyaux n'a pas lieu
et au cours des divisions successives, il ne subsistera que l'un des noyaux parentaux. Celui-ci sera associé à un cytoplasme composite ou recombiné. Il contient les organites cytoplasmiques de l'un ou l'autre parent. On constate souvent une recombinaison des mitochondries. En revanche, les chloroplastes de l'un des deux parents sont souvent éliminés. Il y a alors modification des relations nucléo-cytoplasmiques. L'obtention de ces cybrides peut être également provoquée. On utilise dans ce cas des doses létales d'irradiation pour les cellules du

49
parent donneur, afin d'inactiver complètement le noyau. Seuls seront transférés ses mitochondries et ses chloroplastes. Le parent receveur peut en plus être traité à l'iodo-acétate, entraînant le blocage de ses organites. Ainsi, les cybrides issus de la fusion seront constitués du noyau du parent receveur et des organites du parent donneur.
Les caractères sous la dépendance de l'ADN mitochondrial ou chloroplastique ne sont pas à négliger. La résistance aux herbicides est codée par exemple par l'ADN chloroplastique.
Les hybrides retenus :
Figure 30 : Les différents hybrides obtenus
B. La transformation génétique. Du fait de l'absence de la paroi pectocellulosique, l'introduction directe de l'ADN dans les cellules est facilitée.

50
6.3 Culture de tissus animaux Il est possible de réaliser des cultures de cellules et de tissus prélevés sur des animaux ou des hommes. On isole les cellules par des moyens mécaniques ou chimiques, puis on les place dans un milieu nutritif contenant du sérum animal. Le sérum contient certains polypeptides jouant le rôle de facteurs de croissance. Ils permettent l'adhérence des cellules sur un support solide. En effet, la plupart des cellules animales en culture ne peuvent croître et se diviser que si elles sont placées sur une surface solide, telle que le verre ou le plastique du fond des boîtes de culture. La culture de cellules provenant directement d'un tissu ou d'un organe est appelée culture primaire . Quand la culture atteint une certaine densité, on peut en prélever une partie et l'incuber, on obtient ainsi une culture secondaire. De cette façon, on peut multiplier le nombre de cultures. La plupart du temps, les cellules conservent les propriétés qu'elles présentaient sur l'organisme. Ainsi, des cellules embryonnaires de muscle squelettique fusionnent en grandes fibres musculaires capables de se contracter; les cellules épithéliales s'assemblent en surfaces cellulaires comme dans un organisme intact; les cellules nerveuses forment même des fibres excitables. Souvent, après avoir servi dans plusieurs cultures secondaires successives, les cellules présentent des signes de vieillissement similaires à ceux rencontrés dans l'organisme. Elles meurent après un certain nombre de divisions cellulaires qui leur est spécifique. Elles reflètent probablement la limite de durée de vie de l'organisme. Pourtant dans certaines cultures, on trouve des cellules qui survivent durant des années et se reproduisent même à l'infini: elles forment des lignées cellulaires permanentes, à disposition des chercheurs. On peut les conserver dans de l'azote liquide à -196 oC, sans avoir besoin de les cultiver. Il existe aujourd'hui des banques de cellules qui envoient du matériel dans le monde entier, afin que les chercheurs travaillent sur les mêmes lignées.
Figure 31 L'obtention de cellules hybrides

51
Par adjonction de certaines substances dans le milieu de culture, on parvient à faire fusionner des cellules de types différents, ce qui permet d'obtenir des cellules à deux noyaux, appelées hétérocaryons. Si les deux noyaux fusionnent, il se forme une cellule hybride. De telles cellules permettent d'étudier les interactions entre les constituants cellulaires provenant de cellules différentes. Quand on fait fusionner des cellules humaines et animales, il arrive fréquemment que l'un ou l'autre chromosome humain se perde. On a ainsi la possibilité d'attribuer un gène à un chromosome déterminé, par l'observation des perturbations. La fusion cellulaire (entre une cellule cancéreuse et un lymphocyte) a par exemple permis, entre autre, la production d’anticorps monoclonaux. Cette hybridation (qui donne naissance à des hybridomes) est très courante dans la recherche et permet d’obtenir des lignées rendues comme « immortelles » grâce à la fusion avec des cellules cancéreuses. Les anticorps monoclonaux sont des anticorps issus d’un seul clone de plasmocytes, reconnaissant qu'un seul type d'épitope sur un antigène donné, donc très spécifique. Désormais, on n’obtient plus des populations d’anticorps différents (en prenant le sérum d’un animal immunisé par exemple) dirigés contre une multitude d’éléments caractéristiques de la protéine (de l’antigène), mais on peut, par cette technique, isoler des anticorps dirigés contre un seul épitope et pas un autre ; comme si, au lieu de reconnaître l’ensemble d’un visage, un système de détection reconnaissait le coin d’un œil, une ride de la joue ou la forme d’une oreille. Produire des anticorps monoclonaux in vitro a longtemps été difficile, en raison de la faible durée de vie des cellules sécrétrices d'anticorps, les plasmocytes. Les anticorps étaient alors obtenus in vivo en injectant chez l'animal un antigène donné et en recueillant les anticorps dans son sang. Cette méthode coûteuse ne donnait qu'une faible quantité d'anticorps, pollués par de nombreuses impuretés. A la fin des années 1970, César Milstein et Georges Köhler ont développé la technique des hybridomes. L'antigène est injecté chez l'animal, et des cellules de rate en sont prélevées après quelques semaines. Dans ces cellules se trouvent des plasmocytes sécrétant des anticorps dirigés spécifiquement contre l'antigène choisi. Ces plasmocytes sont alors fusionnés in vitro avec des myélomes, qui sont des cellules tumorales ayant la propriété de se multiplier indéfiniment. Les cellules hybrides obtenues (dites « hybridomes » sont sélectionnées.
Figure 32 : Les hybridomes

52
La sélection des hybridomes : Après la fusion cellulaire, il y a trois catégories de cellules :
1) Les cellules de myélome non fusionnées ou fusionnées entre elles.
2) Les plasmocytes non fusionnés ou fusionnés entre eux.
3) Les hybridomes Il faut donc sélectionner les hybridomes. Cette sélection se fait sur un milieu de culture spécifique, milieu HAT (Hypoxanthine Aminoptérine Thymidine) car les cellules cancéreuses (cellules de myélome) utilisées sont des cellules qui, par sélection génétique, ont perdu la faculté de produire une enzyme impliquée dans la synthèse de nucléotide. Sans cette enzyme, la cellule ne peut pas vivre dans ce milieu. Après quelques jours de culture :
- Les cellules de myélome qui n’ont pas fusionné meurent.
- Les plasmocytes qui n’ont pas fusionné meurent car leur espérance de vie est limitée.
- Les hybridomes sont les seules cellules qui peuvent survivre dans ce milieu. Une fois les hybridomes sélectionnés, il faut sélectionner les anticorps monoclonaux. Pour cela :
1. On isole au hasard quelques hybridomes que l’on met en culture séparément sur une plaque contenant des puits (un hybridome par puit).
2. On laisse les cellules se diviser pendant quelques jours. On obtient un clone d’hybridomes
dans chacune des puits et chaque clone produit un type d’anticorps (anticorps monoclonaux).
3. Les anticorps monoclonaux se trouvent dans le surnageant de chaque puit.
4. Il faut ensuite vérifier la compatibilité des anticorps avec l’antigène. Le génie génétique permet aujourd'hui de produire des anticorps monoclonaux utilisables en pratique clinique humaine. Mais la plupart des anticorps étant produits dans des cellules de rongeurs (souris, rat, hamster, lapin), ils déclenchent une réaction immunitaire lors de leur injection à un patient. Cette immunité inactive progressivement l'action bénéfique de l'anticorps. Pour éviter cela, on cherche à produire des anticorps chimériques "humanisés", modifiés par génie génétique pour remplacer au maximum les fragments constants de l'espèce d'origine par des fragments humains. Les applications des anticorps monoclonaux sont nombreuses. Ils sont très largement utilisés à la fois comme outils de diagnostic et dans des buts thérapeutiques. Dans le domaine du diagnostic, ils permettent, par exemple, de détecter la présence de virus (hépatite B, herpès ou SIDA), de bactéries ou de cellules tumorales. Couplés à des toxines puissantes, ils permettent de détruire sélectivement des cellules malades. On peut les cultiver pour occuper la place d’un agent infectieux, sur son récepteur cellulaire, et lui fermer ainsi la porte d’entrée des cellules. On estime qu’un tiers des entreprises de biotechnologie dans le monde commercialise des produits à base d’anticorps monoclonaux.

53
6.4 Les cellules souches embryonnaires (cellule ES) Tout d’abord, une cellule souche est une cellule précurseur non différenciée capable de se renouveler et de se différencier. Il existe différents types de cellules souches : Les cellules souches totipotentes : Cellules embryonnaires jusqu’au stade 8 cellules au
maximum. Ces cellules sont capables de se développer jusqu’au stade complet d’un individu.
Les cellules souches pluripotentes : Cellules embryonnaires à partir du stade 16 cellules jusqu’au
stade blastocyste (100 cellules environ). Ces cellules sont capables de se développer en tous les types de cellules de l’organisme.
Les cellules souches multipotentes : Cellules adultes qui ne peuvent générer qu’un éventail limité
de cellules. Les cellules souches de moelle osseuses, par exemple, à partir desquelles se forment les cellules sanguines.
Peu de branches de la recherche au sein des biosciences se développent avec autant de dynamisme que la recherche sur les cellules souches. Le 6 novembre1998 une équipe américaine révèle qu'elle a réussi à isoler et à mettre en culture des "cellules souches embryonnaires" humaines. Ces cellules se multiplient à l'infini in vitro. La mise en culture des cellules ES humaines laisse envisager la création d’embryons-clone humains destinés à produire des tissus spécialisés sains, qui remplaceraient leurs homologues chez un adulte malade. Fin 2007, il a fallu définitivement abandonner un dogme biologique, à savoir l’impossibilité de reprogrammer des cellules. Deux équipes de chercheurs, au Japon et aux Etats-Unis, ont en effet réussi, grâce à l’adjonction de quatre gènes, à ramener un des cellules de la peau mature à un stade quasi embryonnaire. Ces cellules souches pluripotentes induites (dites cellules iPS), très similaires aux cellules embryonnaires, ont suscité de nombreuses vagues bien au-delà du monde scientifique. La fascination exercée par les cellules souches embryonnaires réside dans leur aptitude à se transformer en chacun des quelques 200 types de cellulaires de l’organisme humain. Or, ces précisément cette aptitude qui intéresse les chercheurs. Ils étudient quelles sont les conditions nécessaires pour que les cellules souches se développent en cellules cardiaques, nerveuses, musculaires ou productrice d’insuline. La compréhension de ces processus biologiques fondamentaux ouvre de nouvelles possibilités à la médecine : grâce à des thérapies dites de « remplacement cellulaire », les chercheurs veulent à l’avenir traiter des maladies comme le diabète, dans lesquelles certains types cellulaires sont absents ou ne fonctionnent pas. Production de cellules souches pluripotentes : Insérer la figure de gènes et dialogue juin 2008 Le chemin menant de la recherche fondamentale sur les cellules souches embryonnaires à des thérapies efficaces est semé d’embûches. Les chercheurs sont conscients qu’un long travail de laboratoire sera nécessaire avant que nous puissions compter sur des applications cliniques. Et la culture des cellules iPS n’a pas non plus rendu plus simple les questions éthiques qui se posent. Ce serait un tort de penser que la science pourrait désormais renoncer aux cellules souches embryonnaires que certains contestent. Seules des analyses détaillées et des comparaisons à long terme avec des cellules souches embryonnaires montreront s’il est effectivement possible de les remplacer par des cellules iPS.