
L. GAUVRY - Espace pédagogique de l'Université du...
26
Année : 2016-2017 L. GAUVRY M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Septembre – Novembre 2016
Transcript of L. GAUVRY - Espace pédagogique de l'Université du...

Année : 2016-2017
L. GAUVRY
M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Septembre – Novembre 2016

Les reproductions d'oeuvres protégées contenues dans ce document sont réalisées avec l'autorisation du CFC(20, rue des grands Augustins. 75006 Paris). CFC : Centre Français d'exploitation du droit de Copie.
M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Septembre Octobre – 2016
ENSEIGNEMENT DE BIOLOGIE CELLULAIRE
COURS (4H)
RAPPELS SUR L'EMBRYOLOGIE pages 1 - 4 DES CELLULES SOUCHES A LA THERAPIE CELLULAIRE
pages 5 - 8 LA MATRICE EXTRACELLULAIRE pages 9 - 10 LES COMPLEXES JONCTIONNELS ET LES PROTEINES
D'ADHERENCE pages 11 - 18
TRAVAUX DIRIGES (3H)
TD1 ET TD2 pages 20 -24
L. GAUVRY

CM1 : Rappels sur l'embryologie
I. Définitions et rappels sur l'embryogenèse. Embryologie : C'est la science qui étudie l’embryon.
L'embryogenèse : L'étude des modifications du futur individu de la fécondation (œuf ou zygote) jusqu'au moment où cet individu peut mener une existence autonome ; celui-ci peut être une larve ou un jeune (poussin ; nouveau-né) qui possède tous les organes de l'adulte mais dont certains ne sont pas encore fonctionnels (appareil génital). C'est la succession des étapes (fécondation, segmentation, gastrulation, organogenèse, mise en place des cellules germinales) qui suivent la fécondation jusqu'à la naissance (espèces vivipares) ou l'éclosion (espèces ovipares). L'embryogenèse comprend la multiplication cellulaire, la croissance, l'organisation des cellules en tissus et l'édification des organes aptes à remplir des fonctions spécifiques.
On nomme embryon l'organisme en voie de développement à partir de la fécondation jusqu'à la naissance ou l'éclosion.
(Remarque : chez les vertébrés on parle d'embryon à partir de la fécondation. Chez l'homme, le terme d'embryon est
utilisé jusqu'au 3ème mois (phase pendant laquelle les principaux organes ont été formés), ensuite on parle de fœtus (phase de croissance).
Ontogenèse : comprend l' ensemble des processus du développement qui, chez un animal ou un végétal, conduisent du zygote à l'adulte. On a les développements embryonnaire et post-embryonnaire. Développement embryonnaire : désigne les transformations qu'il y a à l'intérieur des enveloppes de l'embryon ou de l'œuf.
Développement post-embryonnaire : désigne les transformations externes qui vont de l'éclosion à la forme adulte. Ce dernier peut s'effectuer : soit de façon directe (ou condensée) : le jeune acquiert progressivement les caractéristiques de l'état adulte par des
processus de croissance ( Oiseaux, mammifères...). L'individu présente à la naissance une forme semblable à l'adulte. Cela est possible lorsque les oeufs ont des réserves vitellines importantes (poulet) ou quand l'embryon utilise des réserves externes (maternelles) grâce à des annexes embryonnaires (organes supplémentaires qui disparaissent à la naissance. (mammifères placentaires)
Soit de façon indirecte (ou dilatée) : le jeune est différent de l’adulte, on parle de larves et il peut subir des transformations (mues) (L1, L2…) avant la métamorphose (passage d'une forme larvaire à une forme adulte). (têtard ---> grenouille).
II. Historique
► 18ième : Wölff (1759) introduisit la notion d'épigenèse avec l'apparition progressive des formes à partir d'un ovule relativement informe.
► 19ième : Von Baer (1828) découverte des feuillets embryonnaires
- l'ontogenèse récapitule la phylogenèse
(le développement de l'individu reproduit l'évolution généalogique de l'espèce (exple : branchie, doigt palmé ---> espèce animal aquatique)(Remarque : Phylogenèse : science qui étudie le mode de formation des espèces et leur évolution).
► 20ième : Lewis (1978) découvre les gènes du développement (gènes homéotiques)
: Gearhart et Thomson (1998) isolent des cellules souches humaines.
III. Les stades de l'embryogénèse
Ils regroupent la fécondation, la segmentation, la gastrulation et l'organogénèse
1
M 32B02 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Octobre 2014M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Octobre 2016

.LA Fecondation : La fécondation c'est la fusion entre deux gamètes (haploides) mâle et femelle pour former un zygote (diploïde).
LA SEGMENTATION : LA SEGMENTATION : …......................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................
Les différents types d’oeufs et de segmentation
Les œufs sont de tailles variables avec la présence de réserves nutritives (le vitellus) ce qui rend possible le développement de l’embryon en l’absence de ressources nutritives externes.
Les types d'oeuf et la segmentation
Le mode de segmentation dépend : de la quantité et de la distribution des réserves en vitellus accumulées par l’oeuf pendant l’ovogenèse. des facteurs cytoplasmiques qui influencent l’orientation du fuseau mitotique et le moment de sa formation. Ces facteurs seront à l’origine des segmentations radiaire (divisions parallèles et perpendiculaires à l’axe PA-PV), spirale (plans de divisions obliques), bilatérale et rotationnelle.
Ainsi, un zygote se divise d’autant plus vite que ses réserves vitellines sont faibles. La quantité de vitellus peut ralentir ou inhiber la segmentation de l’œuf.
On observe ainsi :
Des segmentations totales (le sillon de division s’étend à la totalité de l’oeuf qui est le plus souvent pauvre en vitellus). Des segmentations partielles ou méroblastiques (une partie seulement du cytoplasme est segmentée). Ceci concerne des oeufs riches en vitellus.
Les oeufs alécithes : Ce sont des oeufs très pauvres en vitellus. On peut citer les oeufs d’amphioxus dont la segmentation est totale et égale car tous les blastomères ont la même taille. Le premier plan de segmentation coupe l’oeuf en deux blastomères dans le plan méridien (axe pôle animal-pôle végétatif) puis le second plan de segmentation est perpendiculaire au premier. Ensuite, il y a une division avec un plan de segmentation équatorial. Ensuite, les divisions méridiennes alternent avec les divisions latidudinales. On obtient alors une blastula avec un blastocèle régulier.
Les oeufs hétérolécithes ou mésolécithes : Ce sont des oeufs assez riches en vitellus (exemple : oeuf de grenouille). Celui-ci est localisé selon un gradient du PA au PV avec une augmentation importante au-dessous du niveau de l’équateur avec une augmentation importante de la taille des plaquettes vitellines. La segmentation est totale et inégale.
Les oeufs télolécithes : Ce sont des oeufs très riches en vitellus. Ce vitellus occupe presque la totalité de la cellule qui est de taille géante (exemple : l’oeuf de poule). Cet oeuf de quelques centimètres renferme une zone sans vitellus de 3 mm de diamètre, appelée la cicatricule. Cette zone contient le noyau et le cytoplasme.
2

LA Gastrulation :
Définition :
1.
3
c) Immigration
e) Prolifération polaire
d) Délamination
b) Epibolie
…...................................................................................................................................................................................................................................................................................................................................................................................
…...................................................................................................................................................................................................................................................................................................................................................................................
…...................................................................................................................................................................................................................................................................................................................................................................................
…..............................................................................................................................................................................................................................................................................................................................................................................................................................................................
…...................................................................................................................................................................................................................................................................................................................................................................................
a) Embolie
…....................................................................................................................................................................................................................................................................................................................................................................................................................................................................

DESTINEE DES FEUILLETS EMBRYONNAIRES CHEZ LES VERTEBRES
L'organogénèse : ...................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................................
épiderme
stomodeum, proctodeum EPIDERME cristallin, vésicules
olfactives et otiques, hypophyse
ECTOBLASTE ECTODERME
tube nerveux encéphale, moëlle épinièreNEURODERME
crêtes neurales ganglions nerveux (crâniens, spinaux, sympathiques)
MESODERME AXIAL corde dorsale
sclérotome squelette axialMESOBLASTE somites myotome muscles striés
dermatome derme et mésen MESODERME LATERAL pièces intermédiaires appareil urogénital
splanchnopleure muscles lisses , myocarde, endocarde lames coelome cavité générale (péritonéale,
péricardiaque) latérales somatopleure musculature viscérale de la tête
péricardemuscles et squelettes des membres
épithélium du tube Digestif et des gl. AnnexesENDOBLASTE ENDODERME épithélium respiratoire (branchies, poumons)
thyroïde
Exemple de modèle l'amphibien
La fin blastula la gastrula organogénèse
la neurula
Organogénèse
formation des somites
espèce
Amphibien
4

CM 1 : Des cellules souches … à la thérapie cellulaire
I . Introduction
II . Définitions des cellules souches et iPS
III . Comment fabrique t-on des cellules souches embryonnaires et iPS ?
IV. Applications et limite
V . La plasticité cellulaire
5
M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Octobre 2016
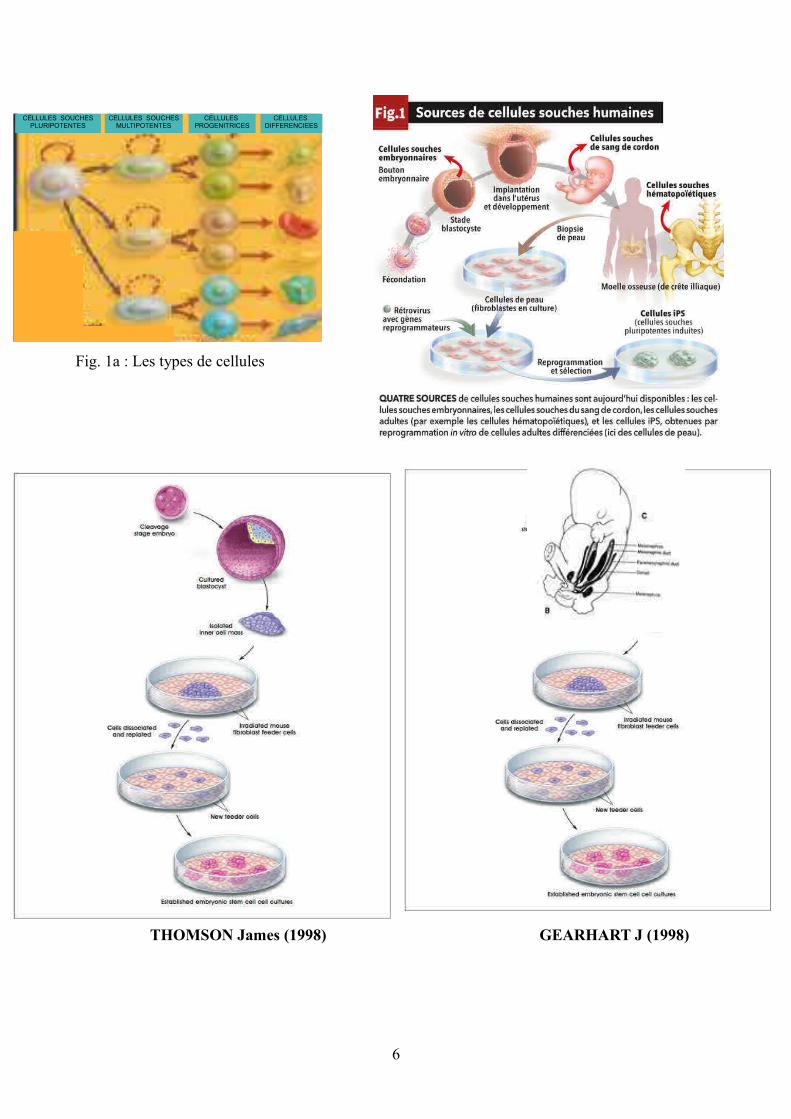
Fig. 1a : Les types de cellules
6
THOMSON James (1998) GEARHART J (1998)
CELLULES SOUCHES PLURIPOTENTES
CELLULES SOUCHES MULTIPOTENTES
CELLULES PROGENITRICES
CELLULES DIFFERENCIEES
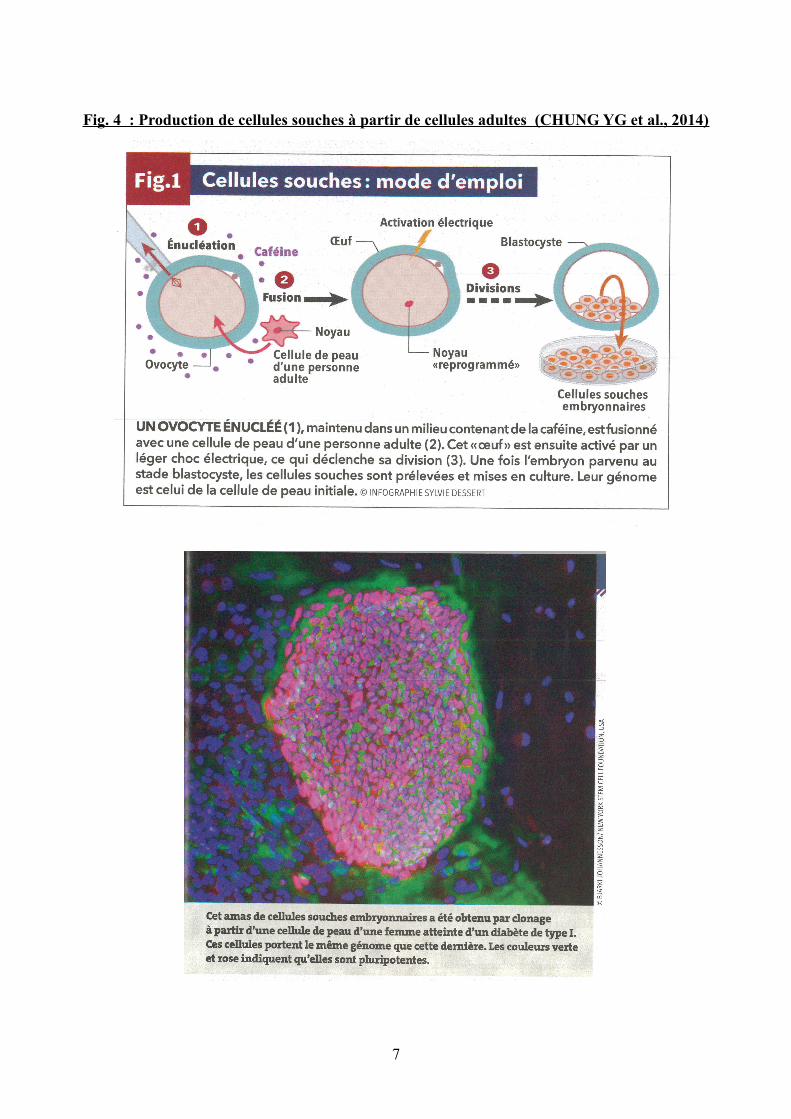
Fig. 4 : Production de cellules souches à partir de cellules adultes (CHUNG YG et al., 2014)
7
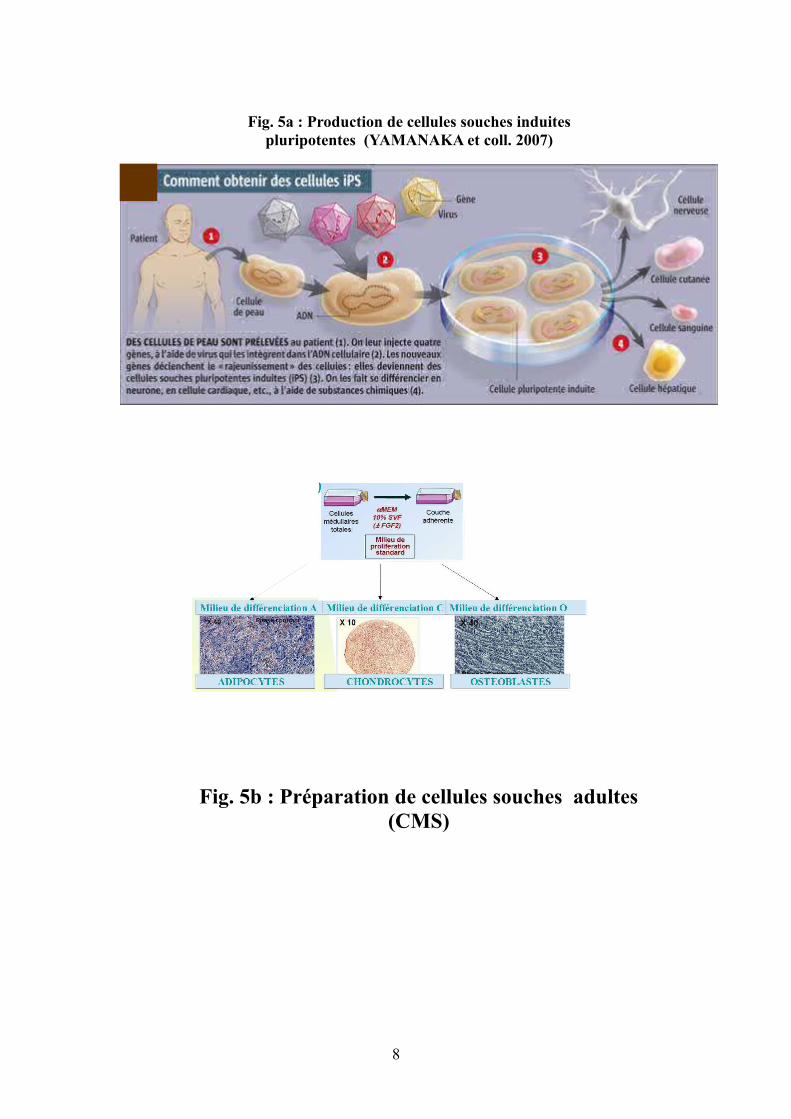
8
Fig. 5a : Production de cellules souches induites pluripotentes (YAMANAKA et coll. 2007)
Fig. 5b : Préparation de cellules souches adultes (CMS)

La matrice extracellulaire (MEC)
I. Définition
La MEC correspond à un enchevêtrement complexe de macromolécules interdépendantes
(glycoprotéines et polysaccharides).
Elle sera en contact avec les cellules des organismes pluricellulaires (elle remplit l’espace
entre les cellules).
Les principaux rôles sont dans :
Le soutien structural
La reconnaissance cellulaire, le guidage et la mobilité.
La séquestration de facteurs de croissance.
Parmi les cellules qui élaborent les éléments de la MEC, on trouve les fibroblastes ou encore les chondroblastes ou les ostéoblastes.
Les éléments constitutifs de la MEC sont classés en 5 catégories :
les collagènes
Ce sont des protéines très abondantes (numérotées de I, II ,III, etc ), résistantes. Elles forment des collagènes fibrillaires au niveau de la MEC, laminaires au niveau de la LB, collagène de jonctions. Rôles dans le maintien et dans la capture de l'eau.
Les fibres élastiques
C'est un matériau composite formé de microfibrille de Fibrilline (avec 2 isoformes) enfouie dans un réseau d’élastine réticulée (qui représente 90 %). L’élastine est une protéine de 70 kDa riche en proline et en glycine et qui possède peu d’hydroxyproline et pas d’hydroxylysine.
◦ Rôles dans la formation d'un réseau de fibres élastiques entrelacées avec celui des fibres de collagène, limitant ainsi l’ampleur de l’étirement et évitant le déchirement du tissu. Rôle dans le fonctionnement de structure comme la peau, les vaisseaux et les poumons est lié à leur élasticité en plus de leur résistance à la tension.
Les glycosaminoglycanes (ou GAG) Ce sont des longs polysaccharides constitués d’unités répétitives de disaccharide qui sont généralement du
type :
Acide hexuronique (acide glucuronique , acide iduronique)
Hexosamine (N acétylglucosamine, N acétylgalactosamine) Ils sont au nombre de cinq :
Le hyaluronane (ou acide hyaluronique) ; la chondroïtine sulfate (cartilage élastique, cartilage hyalin, os, derme, cornée) ; l’héparane sulfate (foie, poumon, aorte) ; le kératane sulfate (cartilage, cornée) ; le dermatane sulfate (derme, tendon, ligament, cartilage fibreux).
Les protéoglycanes
Les protéoglycanes correspondent à l’association de GAGs avec des protéines centrales.
Les rôles sont de remplissage et de permettre de résister aux forces de compression, de participer
9
M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Octobre 2015

à la rétention d’eau, de limiter la recirculation d’eau et la diffusion, rôle de lubrifiant (au niveau des articulations avec le hyaluronane et les protéoglycanes), dans l’expression séquentielle de protéoglycanes au cours du développement et de la cicatrisation.
les glycoprotéines adhérentes
Ces molécules constituent une colle moléculaire mais aussi des signaux nécessaires au développement ou à la réparation tissulaire.
Elles renferment de nombreux domaines (du complément, des domaines riche en cystéine, EGF, fibronectine I, II et III (FN-I), (FN-II) et (FN-III), fibrinogène, d’immunoglobulines et de lectines), aux RGD (qui se fixent aux intégrines). Exemple: la fibronectine forme des structures en V qui se composent de deux sous-unités de 235 kDa environ reliées par des ponts disulfures situés près de l’extrémité COOH terminale.
Les rôles de la fibronectine sont dans la formation de la MEC au cours du développement embryonnaire avant d’être remplacée par une matrice à base de collagène et dans le guidage où elle procurerait des sites d’adhérence lors des déplacements. L’adhérence entre les cellules et la MEC pourraient participer au pontage de molécule matricielle.
II. La lame basale
La lame basale est une matrice qui possède principalement qu’un type de collagène de type IV qui forme des structures lamillaires et de la laminine (Fig.2.)
Rôles :
Soutien mécanique
Barrière cellulaire sélective
Dans la régénération tissulaire après lésion
La laminine (Fig. 2) est une glycoprotéine constituée de 3 sous unités protéiques reliées entre elles par des ponts disulfures. Elle forme une croix asymétrique de 850 kDa. L’autoassemblage du réseau de la laminine, puis du collagène se produit au contact des cellules qui les sécrètent (où elles sont en fortes concentrations) mais aussi grâce aux récepteurs membranaires de la laminine (intégrines) et du collagène IV (le dystroglycane).
10
Fig. 2 : La structure de la laminine. La schématisation de la molécule est présentée sur la figure (a), et des photographies électroniques de molécule de laminine marquée au platine (b). Cette glycoprotéine à domaine multiple est composée de trois polypeptides (α, β et χ) qui sont reliés par des ponts disulfures dans la structure asymétrique.
Fig. 1: Des pontages entre les deux réseaux sont assurés par l’entactine (ou nidogène) (glycoprotéine d’adhérence en bâtonnet) et le perlécane (protéoglycanes à héparane sulfate). Ils se lient aux 4 types de molécules (laminine, collagène, entactine, perlécane).

Les complexes jonctionnels et les protéines d'adhérence
I. Introduction
II. Quelques principes généraux de l’adhérence cellulaire
III. Les principales familles des récepteurs d’adhésion
III.1. Les cadhérines
III.2. Les Molécules d’Adhérence Cellulaire de la famille des immunoglobulines
(Ig-CAM)
III.3. Les intégrines
III.4. Les sélectines
III.5. Les mucines
III.6. Les autres
11
M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Octobre 2016
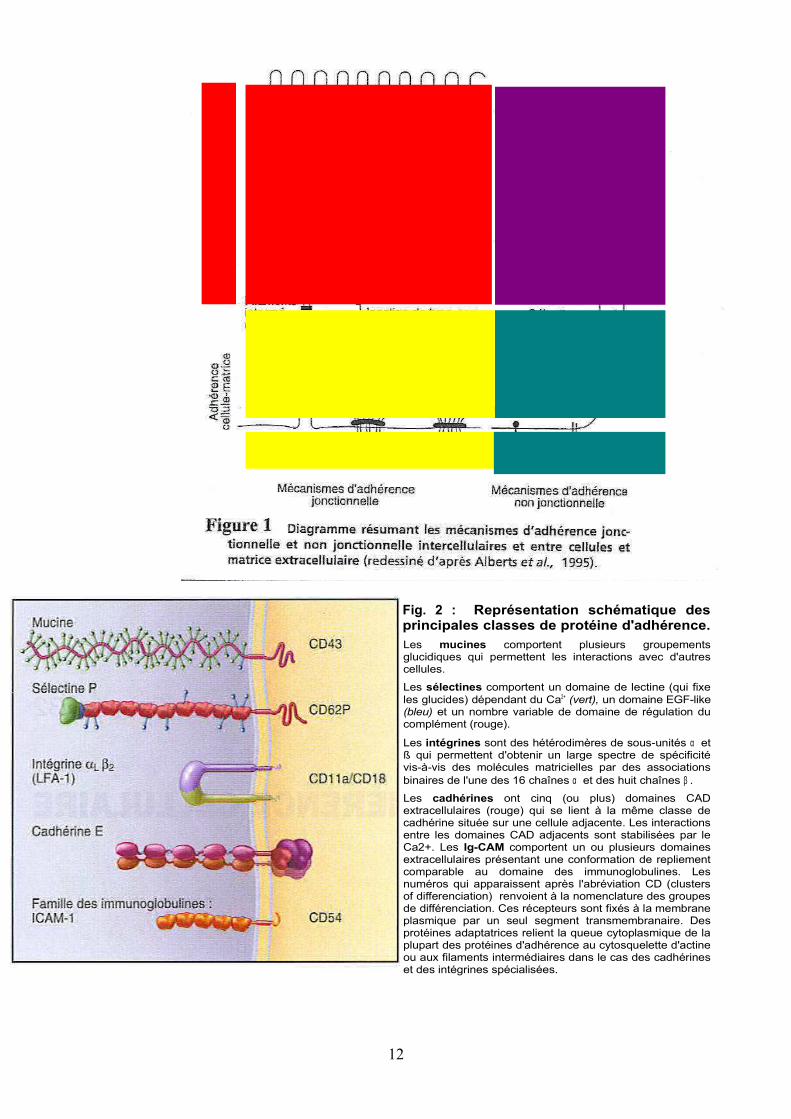
12
Fig. 2 : Représentation schématique des principales classes de protéine d'adhérence. Les mucines comportent plusieurs groupements glucidiques qui permettent les interactions avec d'autres cellules.
Les sélectines comportent un domaine de lectine (qui fixe les glucides) dépendant du Ca2+ (vert), un domaine EGF-like (bleu) et un nombre variable de domaine de régulation du complément (rouge).
Les intégrines sont des hétérodimères de sous-unités α et ß qui permettent d'obtenir un large spectre de spécificité vis-à-vis des molécules matricielles par des associations binaires de l'une des 16 chaînes α et des huit chaînes β .
Les cadhérines ont cinq (ou plus) domaines CAD extracellulaires (rouge) qui se lient à la même classe de cadhérine située sur une cellule adjacente. Les interactions entre les domaines CAD adjacents sont stabilisées par le Ca2+. Les Ig-CAM comportent un ou plusieurs domaines extracellulaires présentant une conformation de repliement comparable au domaine des immunoglobulines. Les numéros qui apparaissent après l'abréviation CD (clusters of differenciation) renvoient à la nomenclature des groupes de différenciation. Ces récepteurs sont fixés à la membrane plasmique par un seul segment transmembranaire. Des protéines adaptatrices relient la queue cytoplasmique de la plupart des protéines d'adhérence au cytosquelette d'actine ou aux filaments intermédiaires dans le cas des cadhérines et des intégrines spécialisées.

13
14
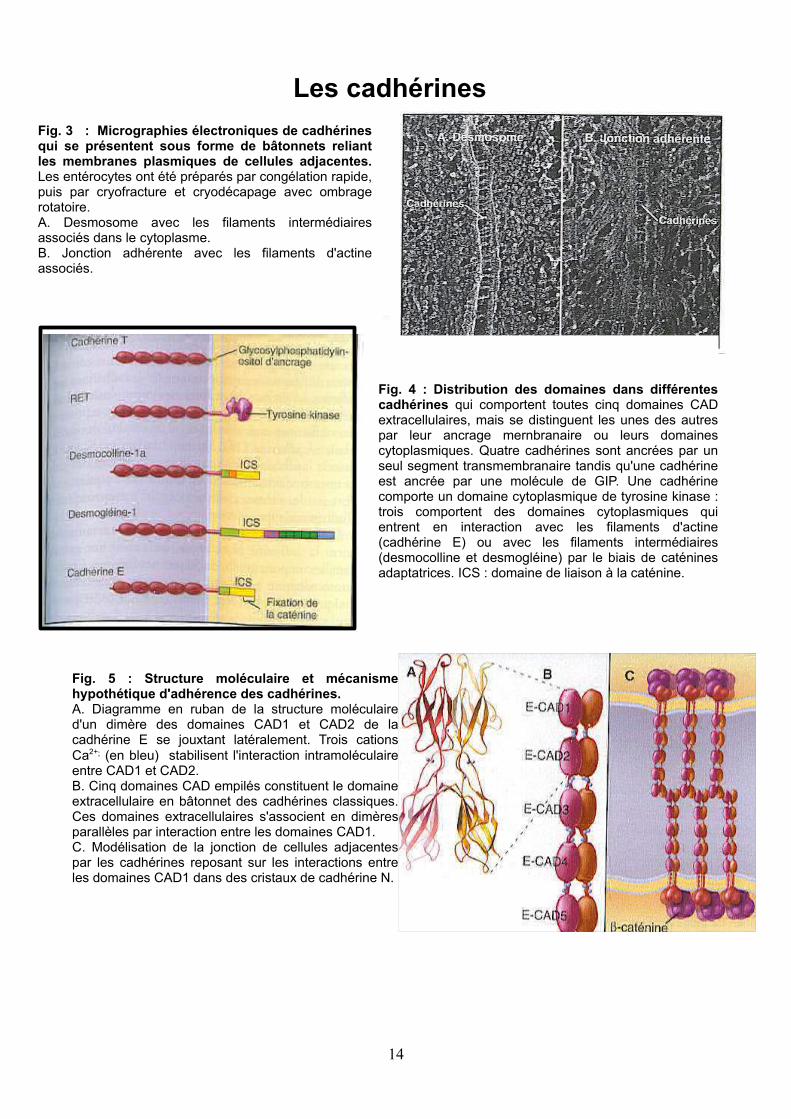
Fig. 5 : Structure moléculaire et mécanisme hypothétique d'adhérence des cadhérines. A. Diagramme en ruban de la structure moléculaire d'un dimère des domaines CAD1 et CAD2 de la cadhérine E se jouxtant latéralement. Trois cations Ca2+; (en bleu) stabilisent l'interaction intramoléculaire entre CAD1 et CAD2. B. Cinq domaines CAD empilés constituent le domaine extracellulaire en bâtonnet des cadhérines classiques. Ces domaines extracellulaires s'associent en dimères parallèles par interaction entre les domaines CAD1. C. Modélisation de la jonction de cellules adjacentes par les cadhérines reposant sur les interactions entre les domaines CAD1 dans des cristaux de cadhérine N.
Les cadhérines
Fig. 3 : Micrographies électroniques de cadhérines qui se présentent sous forme de bâtonnets reliant les membranes plasmiques de cellules adjacentes. Les entérocytes ont été préparés par congélation rapide, puis par cryofracture et cryodécapage avec ombrage rotatoire.A. Desmosome avec les filaments intermédiaires associés dans le cytoplasme. B. Jonction adhérente avec les filaments d'actine associés.
Fig. 4 : Distribution des domaines dans différentes cadhérines qui comportent toutes cinq domaines CAD extracellulaires, mais se distinguent les unes des autres par leur ancrage mernbranaire ou leurs domaines cytoplasmiques. Quatre cadhérines sont ancrées par un seul segment transmembranaire tandis qu'une cadhérine est ancrée par une molécule de GIP. Une cadhérine comporte un domaine cytoplasmique de tyrosine kinase : trois comportent des domaines cytoplasmiques qui entrent en interaction avec les filaments d'actine (cadhérine E) ou avec les filaments intermédiaires (desmocolline et desmogléine) par le biais de caténines adaptatrices. ICS : domaine de liaison à la caténine.
15
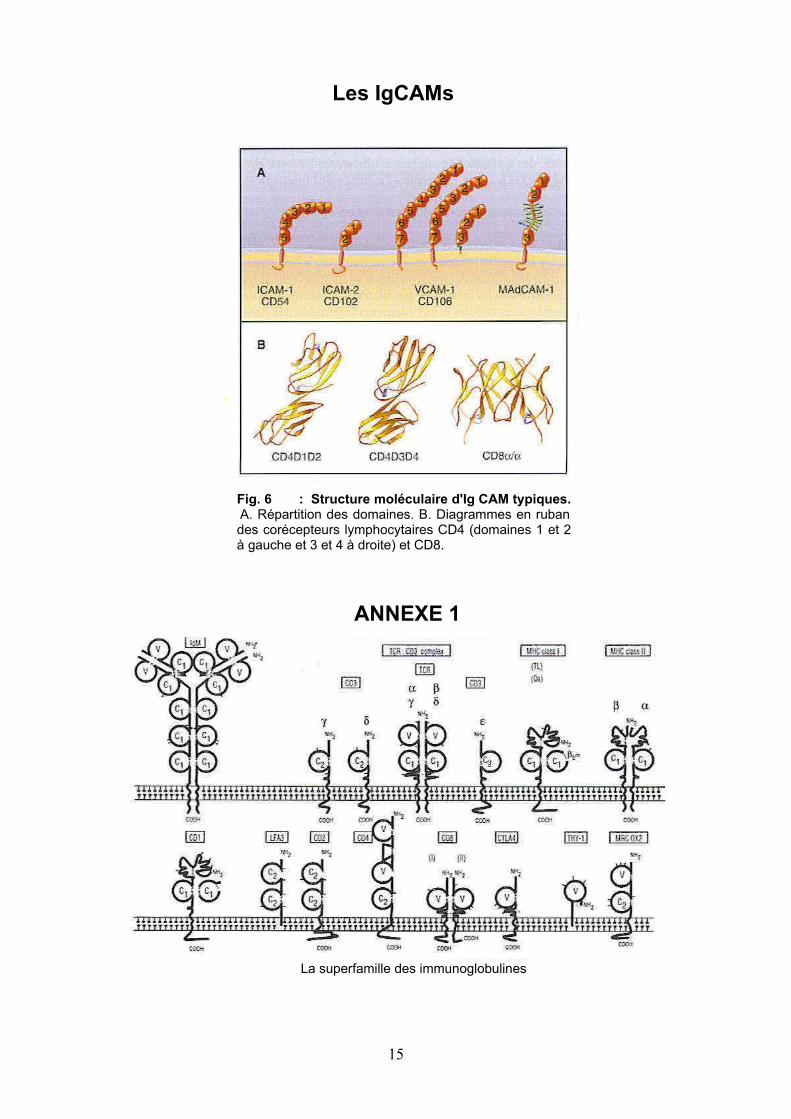
Fig. 6 : Structure moléculaire d'Ig CAM typiques. A. Répartition des domaines. B. Diagrammes en ruban des corécepteurs lymphocytaires CD4 (domaines 1 et 2 à gauche et 3 et 4 à droite) et CD8.
ANNEXE 1
La superfamille des immunoglobulines
Les IgCAMs
16
Les intégrines
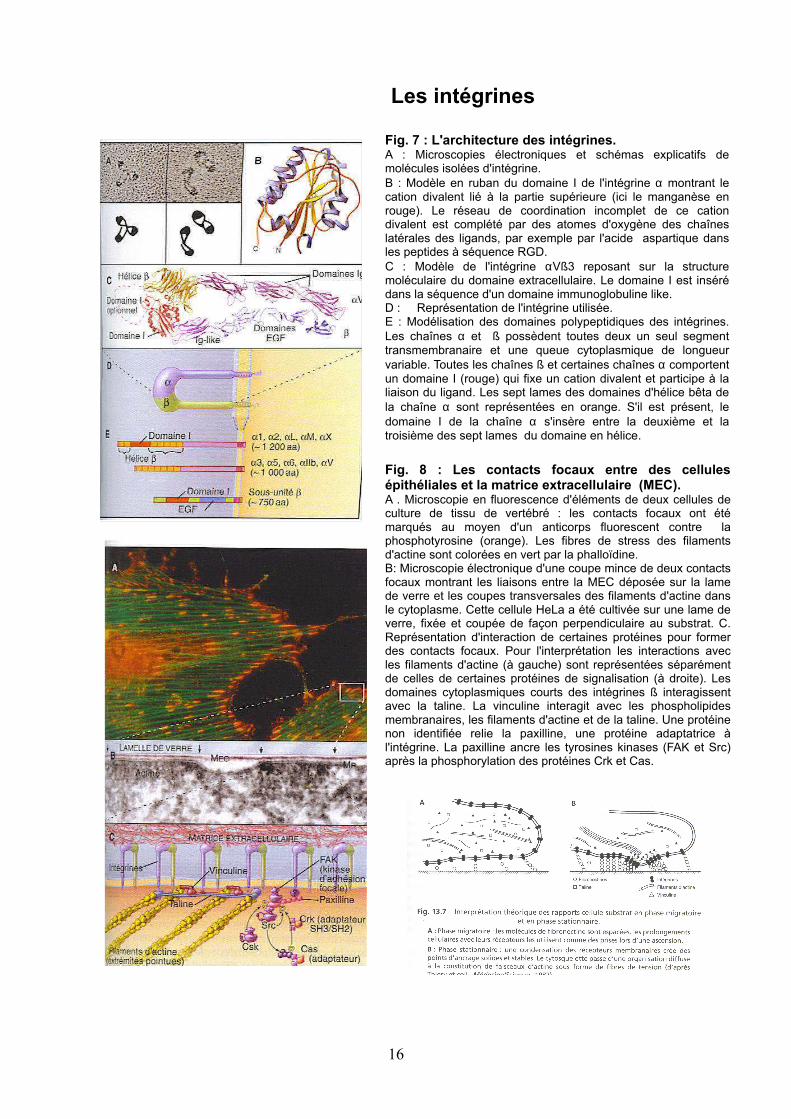
Fig. 7 : L'architecture des intégrines.A : Microscopies électroniques et schémas explicatifs de molécules isolées d'intégrine.B : Modèle en ruban du domaine I de l'intégrine α montrant le cation divalent lié à la partie supérieure (ici le manganèse en rouge). Le réseau de coordination incomplet de ce cation divalent est complété par des atomes d'oxygène des chaînes latérales des ligands, par exemple par l'acide aspartique dans les peptides à séquence RGD.C : Modèle de l'intégrine αVß3 reposant sur la structure moléculaire du domaine extracellulaire. Le domaine I est inséré dans la séquence d'un domaine immunoglobuline like.D : Représentation de l'intégrine utilisée.E : Modélisation des domaines polypeptidiques des intégrines. Les chaînes α et ß possèdent toutes deux un seul segment transmembranaire et une queue cytoplasmique de longueur variable. Toutes les chaînes ß et certaines chaînes α comportent un domaine I (rouge) qui fixe un cation divalent et participe à la liaison du ligand. Les sept lames des domaines d'hélice bêta de la chaîne α sont représentées en orange. S'il est présent, le domaine I de la chaîne α s'insère entre la deuxième et la troisième des sept lames du domaine en hélice.
Fig. 8 : Les contacts focaux entre des cellules épithéliales et la matrice extracellulaire (MEC).A . Microscopie en fluorescence d'éléments de deux cellules de culture de tissu de vertébré : les contacts focaux ont été marqués au moyen d'un anticorps fluorescent contre la phosphotyrosine (orange). Les fibres de stress des filaments d'actine sont colorées en vert par la phalloïdine.B: Microscopie électronique d'une coupe mince de deux contacts focaux montrant les liaisons entre la MEC déposée sur la lame de verre et les coupes transversales des filaments d'actine dans le cytoplasme. Cette cellule HeLa a été cultivée sur une lame de verre, fixée et coupée de façon perpendiculaire au substrat. C. Représentation d'interaction de certaines protéines pour former des contacts focaux. Pour l'interprétation les interactions avec les filaments d'actine (à gauche) sont représentées séparément de celles de certaines protéines de signalisation (à droite). Les domaines cytoplasmiques courts des intégrines ß interagissent avec la taline. La vinculine interagit avec les phospholipides membranaires, les filaments d'actine et de la taline. Une protéine non identifiée relie la paxilline, une protéine adaptatrice à l'intégrine. La paxilline ancre les tyrosines kinases (FAK et Src) après la phosphorylation des protéines Crk et Cas.
17
Les sélectines et les mucines
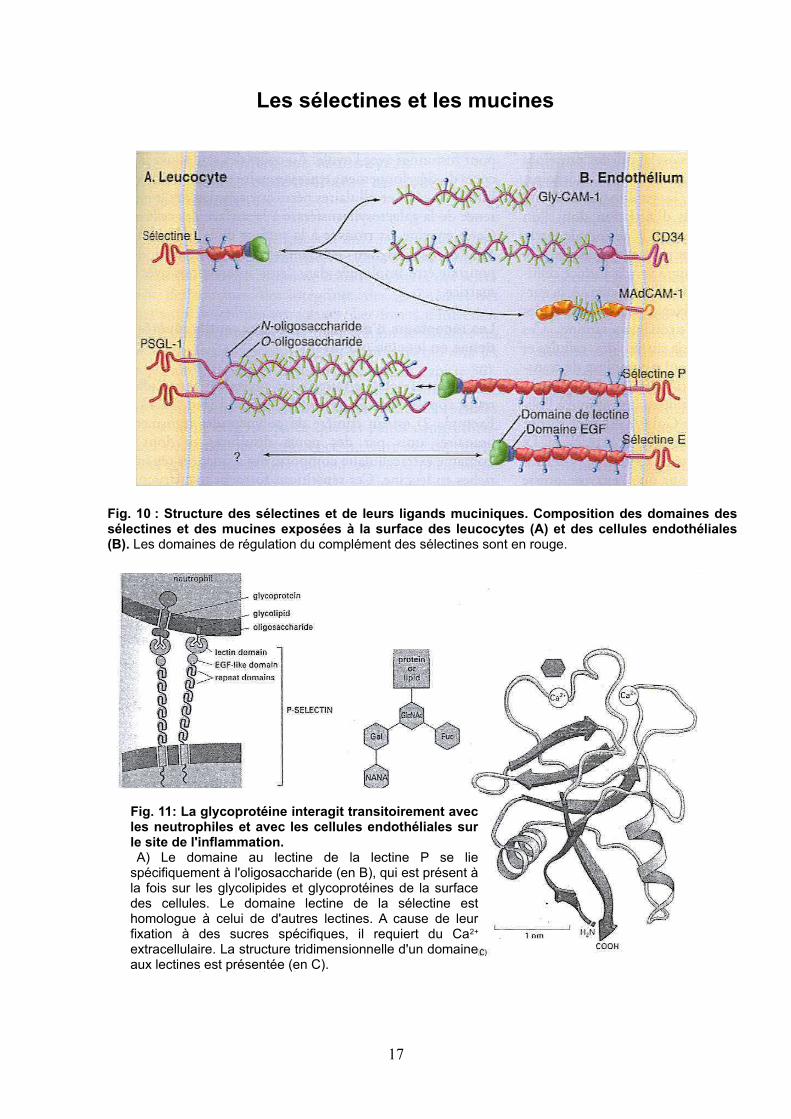
Fig. 10 : Structure des sélectines et de leurs ligands muciniques. Composition des domaines des sélectines et des mucines exposées à la surface des leucocytes (A) et des cellules endothéliales (B). Les domaines de régulation du complément des sélectines sont en rouge.
Fig. 11: La glycoprotéine interagit transitoirement avec les neutrophiles et avec les cellules endothéliales sur le site de l'inflammation.
A) Le domaine au lectine de la lectine P se lie spécifiquement à l'oligosaccharide (en B), qui est présent à la fois sur les glycolipides et glycoprotéines de la surface des cellules. Le domaine lectine de la sélectine est homologue à celui de d'autres lectines. A cause de leur fixation à des sucres spécifiques, il requiert du Ca2+ extracellulaire. La structure tridimensionnelle d'un domaine aux lectines est présentée (en C).
Fig. 11: La glycoprotéine interagit transitoirement avec les neutrophiles et avec les cellules endothéliales sur le site de l'inflammation.
A) Le domaine au lectine de la lectine P se lie spécifiquement à l'oligosaccharide (en B), qui est présent à la fois sur les glycolipides et glycoprotéines de la surface des cellules. Le domaine lectine de la sélectine est homologue à celui de d'autres lectines. A cause de leur fixation à des sucres spécifiques, il requiert du Ca2+ extracellulaire. La structure tridimensionnelle d'un domaine aux lectines est présentée (en C).
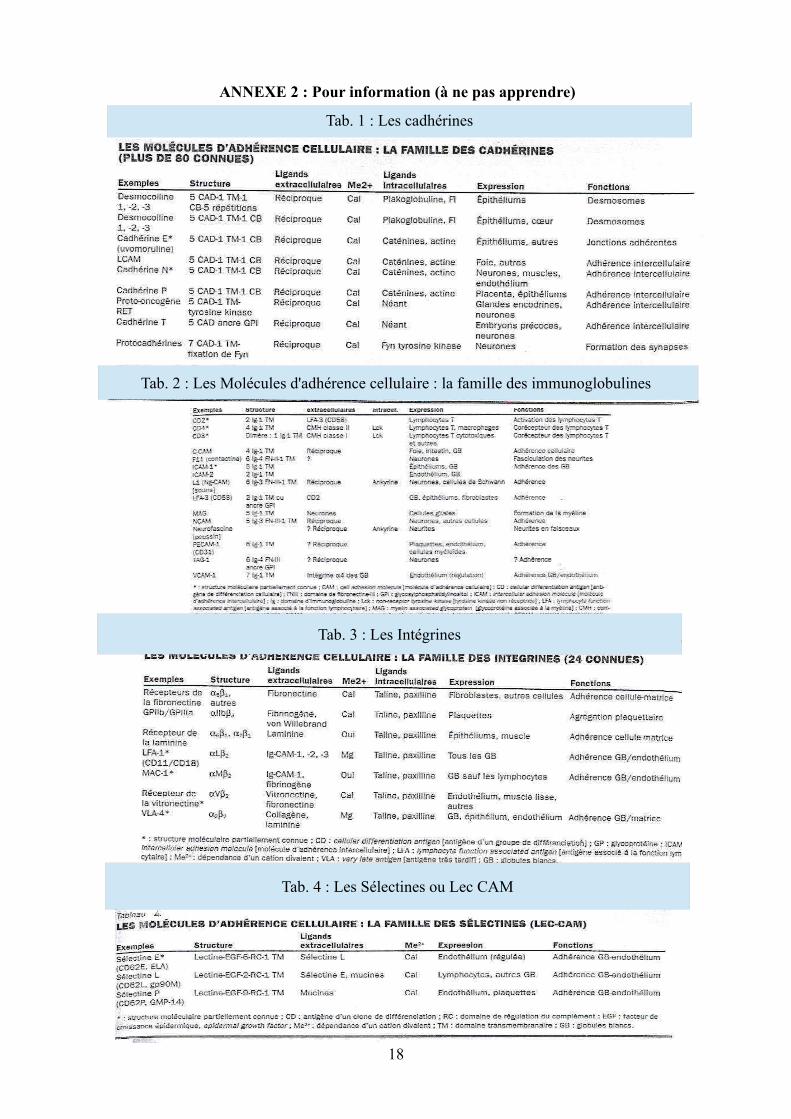
18
Tab. 3 : Les Intégrines
Tab. 1 : Les cadhérines
Tab. 2 : Les Molécules d'adhérence cellulaire : la famille des immunoglobulines
Tab. 4 : Les Sélectines ou Lec CAM
ANNEXE 2 : Pour information (à ne pas apprendre)

TRAVAUX DIRIGES
19

TD1 et 2 : Rappels sur
LES CELLULES ET LES LIGNEES.
3 parties :
Information sur les TP
Rappels sur les cellules
Exercices sur les cellules en culture
1. Définir en une phrase les mots et les sigles suivants :
Culture primaire :
Lignée cellulaire :
Un milieu de culture :
A partir de l'annexe 3 de votre poly TP, vous classerez ces éléments :
Pouvez-vous préciser les informations concernant les cellules Hela ?Origine :niveau de sécurité :milieu de culture utilisé :
Un incubateur à CO2 :
20
Tests de connaissance
M 3202 – BIOLOGIE CELLULAIRE ET METHODES ALTERNATIVES A L'EXPERIMENTATION ANIMALE Septembre -octobre 2016

2. Exercice sur les cellules
Au niveau du laboratoire des tests sont réalisés pour optimiser des cultures animales de reins et plus précisement des cellules épithéliales rénales. A partir d'explants rénaux préalablement congelés dans du MEM supplémenté en DMSO 10%, deux tests de dissociation des cellules sont réalisés sur ces organes.Le premier à partir de collagénase pendant 1h à 37°C.Le second à partir d'un automate le Gentle Macs (Miltenyi Biotec) qui permet en plus de digérer les échantillons de les broyer mécaniquement. Ici les échantillons sont incubées en présence de collagénase pendant 10 min à 37°C.Pour chaque test les cellules sont lavées puis comptées avant d'être mises en culture dans des flasques T25.Les résultats sont présentés sur la cellule de Malassez ci-dessous :
Résultat des comptages 1) Cellules dissociées pendant une heure sans le Gentle Macs
Photos de cellules comptées
2) Cellules dissociées pendant 10 min avec le Gentle Macs
Concentrations à calculer à partir de 8 rectangles ?
…................................................ …................................................
Figure 1
21

1/ Pourquoi utilise -t-on de la collagénase ?
2/ Après avoir déterminé les concentrations cellulaires à partir des résultats de la figure 1, vous conclurez sur l'intérêt de la méthode ?
Les échantillons sont analysées par cytométrie en flux en utilisant des anticorps phénotypiques des cellules épithéliales rénales et les résultats sont présentés dans un tableau ci-dessous.
Analyse par Cytométrie en flux 1) Cellules dissociées pendant une heure sans le Gentle Macs
2) Cellules dissociées pendant 10 min avec le Gentle Macs
Pourcentage de cellules épithéliales rénales
80,00% 60,00%
Autres phénotypes 20,00% 40,00%
3/ Interprétez et concluez sur l'intérêt de la méthode ?
3. Entretien et suivi d'une culture de cellules L 929.
En procédant comme pour l'étude d'une croissance bactérienne, construire la courbe de croissance de cette culture à partir des données suivantes (Tableau 1) : la croissance est suivie par l'évolution de la densité surfacique des cellules L 929.
N.B. - Densité surfacique = nombre de cellules par unité de surface.
Tableau 1 : Résultats d'une croissance de cellules L 929.
Durée d’incubation (h) 0 8 12 22 30 34 48 52 56 60 70 79 94 100Densité surfacique (X 104 cellules.cm-2)
3,0 3,1 3,35 4,5 5,95 6,7 10,2 11,5 12,9 14,8 20,0 26,5 38,2 40,5
Ln
2/ Déterminer le taux de croissance népérien (ou vitesse de croissance maximale spécifique) et le temps de génération de cette culture.
Une culture de cellules L 929, au plateau de densité, dans un flacon de 25 cm 2 de surface utile (sur laquelle peuvent se multiplier les cellules), est remise en suspension selon le protocole décrit dans le document 1. 9 mL de suspension cellulaire (S1) sont alors recueillis. Le résultat de l'observation X 190 de S1 à l'hématimètre de Malassez (volume total 1 mm3 (ou 1µL)) est fourni par le document 1.
3/ Déterminer la concentration exacte de la suspension S1.
22

4/ Quelle était la densité surfacique de cellules dans le flacon avant trypsinisation ?
On veut ensemencer un flacon de 175 cm2 avec 3,5.105 cellules et une hauteur de milieu de 5 mm.
5/ Quel volume total doit contenir le flacon ?
6/ Quelle sera la concentration cellulaire réalisée ?
7/ Quelle dilution finale aura-t-on effectuée ?
8/ Quelle sera la densité surfacique de cellules dans le flacon ainsi repiqué ?
9/ Combien de générations seront nécessaires pour atteindre la densité surfacique de cellules maximale ?
Sachant qu'une phase de latence de 18 heures est régulièrement observée après un repiquage, quelle est la durée d'incubation prévisible avant d'obtenir cette densité ?
Document 1 : Numération de la suspension S1, hématimètre de Malassez (X 190)
N.B. - Caractéristiques de la cellule de Malassez : Hauteur................................................................................................................. 0,2 mmSurface quadrillée (grand rectangle)......………………………….…………………. 5 mm2
Nombre de rectangles dans le quadrillage ....………………………………….….... 100Distance entre deux lignes.........…............………………………………………..…. 50 µm
23

ANNEXE 1 : Comptage avec une cellule de Malassez
Le comptage de la concentration d’une culture grâce à la cellule de Malassez (haemacytomètre). Cette méthode est la plus simple pour dénombrer une culture de cellules animales, micro-algues, etc .. Le principe peut être appliqué à d’autres types de cellules. Il faut alors reprendre le calcul pour l’adapter à la profondeur et à la surface de la cellule utilisée.
Coupe de la cellule de Malassez
Le gravage vertical de la cellule de Malassez est composé de 5 groupes de 6 lignes, plus une ligne seule. Les groupes de lignes sont espacés de 0,25 mm. Les lignes des groupes sont espacées de 0,05 mm. Le gravage horizontal est constitué de 5 groupes de 5 lignes, plus une ligne seule. Les groupes de lignes sont espacés de 0,2 mm. Les lignes des groupes sont espacés de 0,05 mm. La profondeur de la cellule est de 0,200 mm.
Vue du gravage de la cellule de Malassez
On utilise donc pour compter :
25 rectangles de 0,2 x 0,25 mm divisés en 20 carrés de 0,05 mm de côté (ces rectangles sont appelés "périmètres de comptage" -en bleu sur le schéma-).
Chaque périmètre a un volume de 0,2x0,2x0,25=0,01 mm3
soit 10-5 mL. La totalité de la cellule, soit 100 rectangles de 0,2x0,25 mm, dont certains sont divisés horizontalement, verticalement, les deux ou pas du tout. Le volume de la cellule est de 0,2x2x2,5 = 1 mm3 = 1 µL = 10-3 mL.
Détail de la cellule de Malassez
Exemple : on compte une culture de cellules. Ce genre de culture compte environ 5 à 10.106
cellules par mL choisit de compter 3 périmètres de comptage.
Sur ces trois périmètres, on dénombre 251 cellules. 251/3=83,6 cellules par périmètres, soit 8,36.106
cellules par mL
Discussion sur la méthode :
La valeur du résultat dépend en partie du soin que l’on aura apporté à la préparation de l’échantillon et de la cellule. On prendra donc bien soin durant cette étape a ne pas trop faire déborder l’échantillon dans les rainures. Si on utilise du formol, les cellules mortes décantent très vite, il faut donc bien homogénéiser l’échantillon avant de le placer sur la lame.
Une autre source d’erreur est le choix du nombre de périmètres que l’on compte, surtout pour de faibles concentrations. Un nombre de 150 cellules comptées semble être un minimum. Pour ne pas oublier de compter un périmètre, on compte méthodiquement ! Le plus simple lorsqu’on ne compte que 4 ou 5 périmètres est de suivre une diagonale.
24
Utilisation :On commence tout d’abord par humecter les bords de la lamelle avec de l'eau, puis elle est déposée sur les supports. Celle-ci doit rester collée à la lame. Ensuite l’échantillon est introduit à l’aide d’une pipette pasteur dans l’espace crée entre la lame et la lamelle par capillarité.
La lame est placée sous un microscope, un grossissement de 400 est pratique. Les cellules peuvent alors être comptées. Les cellules à cheval sur un bord ne sont comptées que pour une moitié. Selon la concentration et la taille des cellules, on choisit la surface sur laquelle on va dénombrer : 3, 4 ou 5 périmètres de comptage pour des cultures très denses (de l’ordre de 2 à 20 millions de cellules par ml). Ensuite, on rapporte le nombre que l’on trouve à un seul périmètre de comptage. Il suffit ensuite
de multiplier ce résultat par 105 et l’on obtient un résultat en nombre de cellules par mL. 25 périmètres, ou la totalité des 100 rectangles pour des cultures beaucoup moins denses (de l’ordre de 500 000 cellules par ml). Si on ne compte que 25 périmètres, on multiplie le résultat par 4 pour rapporter le nombre à la surface totale de la cellule. Il suffit ensuite de multiplier le chiffre que l’on
obtient par 103 et l’on obtient un résultat en nombre de cellules par mL.
Zone de dépôt par capillarité et de comptage


















![PARCOURS D'OEUVRES D'ART EN VILLEblogacabdx.ac-bordeaux.fr/artsculturecitoyennete40/wp... · 2019. 10. 15. · Microsoft PowerPoint - MDM SCULPTURE DIAPO version Prof.odp [Réparé]](https://static.fdocuments.fr/doc/165x107/60d1ef2dc53aa6199801e046/parcours-doeuvres-dart-en-2019-10-15-microsoft-powerpoint-mdm-sculpture.jpg)