
Involvement of heparin affin regulatory peptide in human prostate cancer
11
Involvement of Heparin Affin Regulatory Peptide in Human Prostate Cancer Francis Vacherot, 1 Danie `le Caruelle, 1 Dominique Chopin, 2 Sixtina Gil-Diez, 2 Denis Barritault, 1 * Jean Pierre Caruelle, 1 and Jose ´ Courty 1 1 Laboratoire de Recherche sur la Croissance Cellulaire, la Re ´paration et la Rege ´ne ´ration Tissulaires, Universite ´ Paris XII-Val de Marne, Cre ´teil, France 2 Laboratoire d’Urologie, Ho ˆpital Henri-Mondor, Cre ´teil, France BACKGROUND. Heparin affin regulatory peptide (HARP) composes, together with midkine (MK), a new family of heparin-binding growth/differentiation factors. Recently, HARP was incriminated in cancer progression, as an angiogenic factor and as a tumor growth factor. In this study, we analyzed the possible involvement of HARP in human prostate cancer (Pca). METHODS. The localization of HARP protein and its mRNAs in normal prostate (n = 5), benign prostate hyperplasia (BPH) (n = 7), and prostate cancer (Pca) (n = 9) was analyzed by immunohistochemistry and in situ hybridization. The mitogenic activity of this growth factor for prostate epithelial cells was determined with a thymidine incorporation assay. HARP cDNA was transfected into normal prostate epithelial (PNT-1A) cells, and their growth was evaluated by soft-agar growth assay. RESULTS. We found HARP protein associated with epithelial cells in PCa but not in normal prostate or BPH, while the corresponding mRNAs were located in the stromal compartment. Furthermore, HARP is mitogenic for PNT-1A, LNCaP, and DU-145 cells. Overexpression of the human HARP in PNT-1A transfected cells induced both anchorage-independent growth and growth at low serum concentrations. CONCLUSIONS. Our results suggest that HARP may act in a paracrine manner from mes- enchymal to tumoral epithelial cells, and may play a role in the molecular mechanisms that regulate prostate tumor cell growth. Prostate 38:126–136, 1999. © 1999 Wiley-Liss, Inc. KEY WORDS: prostate; growth factor; HARP; tumor Abbreviations: PCa, prostate cancer; EGF, epidermal growth factor; FGF, fibroblast growth factor; IGF, insulin-like growth factor; TGF- b, transforming growth factor beta; HARP, heparin affin regulatory peptide; MK, midkine; HB-GAM, heparin-binding growth- associated molecule; PTN, pleiotrophin; BPH, benign prostate hy- perplasia; FCS, fetal calf serum; G 418, geneticin. Grant sponsor: Association pour la Recherche sur le Cancer; Grant number: 6595; Grant sponsor: Ministe `re de l’Education Nationale (DRED); Grant sponsor: Ligue Nationale contre le Cancer; Grant sponsor: Naturalia et Biologia; Grant sponsor: Association pour la Recherche sur les Tumeurs de Prostate. *Correspondence to: Denis Barritault, Laboratoire de Recherche sur la Croissance Cellulaire, la Re ´paration et la Rege ´ne ´ration Tissu- laires, Universite ´ Paris XII-Val de Marne, Avenue du Ge ´ne ´ral de Gaulle, 94010 Cre ´teil, France. E-mail: [email protected] Received 23 February 1998; Accepted 19 June 1998 The Prostate 38:126–136 (1999) © 1999 Wiley-Liss, Inc.
Transcript of Involvement of heparin affin regulatory peptide in human prostate cancer

Involvement of Heparin Affin Regulatory Peptidein Human Prostate Cancer
Francis Vacherot,1 Daniele Caruelle,1 Dominique Chopin,2 Sixtina Gil-Diez,2
Denis Barritault,1* Jean Pierre Caruelle,1 and Jose Courty1
1Laboratoire de Recherche sur la Croissance Cellulaire, la Reparation et la RegenerationTissulaires, Universite Paris XII-Val de Marne, Creteil, France
2Laboratoire d’Urologie, Hopital Henri-Mondor, Creteil, France
BACKGROUND. Heparin affin regulatory peptide (HARP) composes, together with midkine(MK), a new family of heparin-binding growth/differentiation factors. Recently, HARP wasincriminated in cancer progression, as an angiogenic factor and as a tumor growth factor. Inthis study, we analyzed the possible involvement of HARP in human prostate cancer (Pca).METHODS. The localization of HARP protein and its mRNAs in normal prostate (n = 5),benign prostate hyperplasia (BPH) (n = 7), and prostate cancer (Pca) (n = 9) was analyzed byimmunohistochemistry and in situ hybridization. The mitogenic activity of this growth factorfor prostate epithelial cells was determined with a thymidine incorporation assay. HARPcDNA was transfected into normal prostate epithelial (PNT-1A) cells, and their growth wasevaluated by soft-agar growth assay.RESULTS. We found HARP protein associated with epithelial cells in PCa but not in normalprostate or BPH, while the corresponding mRNAs were located in the stromal compartment.Furthermore, HARP is mitogenic for PNT-1A, LNCaP, and DU-145 cells. Overexpression ofthe human HARP in PNT-1A transfected cells induced both anchorage-independent growthand growth at low serum concentrations.CONCLUSIONS. Our results suggest that HARP may act in a paracrine manner from mes-enchymal to tumoral epithelial cells, and may play a role in the molecular mechanisms thatregulate prostate tumor cell growth. Prostate 38:126–136, 1999. © 1999 Wiley-Liss, Inc.
KEY WORDS: prostate; growth factor; HARP; tumor
Abbreviations: PCa, prostate cancer; EGF, epidermal growth factor;FGF, fibroblast growth factor; IGF, insulin-like growth factor; TGF-b, transforming growth factor beta; HARP, heparin affin regulatorypeptide; MK, midkine; HB-GAM, heparin-binding growth-associated molecule; PTN, pleiotrophin; BPH, benign prostate hy-perplasia; FCS, fetal calf serum; G 418, geneticin.Grant sponsor: Association pour la Recherche sur le Cancer; Grantnumber: 6595; Grant sponsor: Ministere de l’Education Nationale(DRED); Grant sponsor: Ligue Nationale contre le Cancer; Grantsponsor: Naturalia et Biologia; Grant sponsor: Association pour laRecherche sur les Tumeurs de Prostate.*Correspondence to: Denis Barritault, Laboratoire de Recherche surla Croissance Cellulaire, la Reparation et la Regeneration Tissu-laires, Universite Paris XII-Val de Marne, Avenue du General deGaulle, 94010 Creteil, France. E-mail: [email protected] 23 February 1998; Accepted 19 June 1998
The Prostate 38:126–136 (1999)
© 1999 Wiley-Liss, Inc.

INTRODUCTION
Prostate cancer (PCa) is the second most commoncause of death from cancer in men and the leadingtype of cancer in men over 75. The prostate showsmild growth during childhood, accelerates in puberty,and reaches adult size during the third decade [1].Inductive interactions between mesenchymal and ep-ithelial tissues are necessary for prostate developmentand function. Mesenchymal cells induce specific sig-nals that regulate the growth and differentiation ofprostate epithelial cells during development of thegland [2]. Cell communications also have an impor-tant influence on the morphogenesis of several otherorgans, such as lung [3] and breast [4]. In adulthood,the mesenchyme contributes to the homeostatic equi-librium and repair of epithelial layers [5,6]. Althoughthe mediators that regulate mesenchymal-epithelialinteractions are poorly understood, it is known thatdisturbances may result in disease [7]. Numerous re-ports suggest that growth factors have a key role aseffectors of mesenchymal-epithelial interactions.Among these growth factors, the epidermal growthfactor (EGF), fibroblast growth factor (FGF), insulin-like growth factor (IGF), and transforming growth fac-tor beta (TGF-b) families have been implicated as po-tential cellular regulators in prostate homeostasis,through autocrine and/or paracrine mechanisms [8,9].
Heparin affin regulatory peptide (HARP) is an 18-kDa protein that belongs to a novel family of heparin-binding molecules, including the structurally relatedprotein, midkine (MK) [10], also called retinoic acid-induced heparin-binding factor [11]. This growth fac-tor has been independently identified by different ap-proaches, leading to several names including heparin-binding growth-associated molecule (HB-GAM) [12]and pleiotrophin (PTN) [13]. The amino-acid sequenceof HARP is highly conserved among human, rat, bo-vine, and mouse species, with 98% similarity [13,14].First isolated from neonatal rat brain as a moleculethat induced neurite outgrowth of embryonic neurons[15], more recent studies have shown that this poly-peptide is also present in non neuronal tissue includ-ing heart, uterus, cartilage, and bone extracts [16]. Thebiological activities of this molecule have been inves-tigated by several laboratories; despite controversialresults, it is now clear that HARP induces the prolif-eration of endothelial, epithelial, and fibroblastic cells[17,18]. In addition, recent studies have shown thatexogenous heparin can modulate the mitogenic activ-ity of HARP in vitro [19].
Given its angiogenic and oncogenic activity [18,20–23], HARP has been suspected of being involved intumor progression. HARP mRNA has been detectedin several tumor cell lines (PC-3, T47Dco, MDA-
MB231, MDA-MB361, Hs-578T, A1827, and PA-1) andin tumor tissues, suggesting a direct role in cell trans-formation [18,20].
In this study, we investigated the tissular locationof HARP mRNA and its translation product in normalprostate, BPH, and PCa. We also studied the mitogen-ic properties of HARP and its function as a tumorgrowth factor by using prostate-derived in vitro mod-els.
MATERIALS AND METHODS
Materials
Prostate tissue samples were provided by the De-partment of Urology (Hopital Henri Mondor, Creteil,France). Normal prostates (n = 5), benign prostate hy-perplasia (n = 7), and prostate cancer (n = 9) wereobtained by prostatectomy, transurethral resection, ornormal donors. All specimens were examined at theDepartment of Pathology (University Paris XII, Cre-teil, France). Tumors were staged according to theTNM classification [24], and histologic grade wasevaluated according to the grading system of Gleason[25]. Fresh prostatic tissue samples were snap-frozenin isopropanol and stored at −80°C. The human epi-thelial cell line derived from normal prostatic secre-tory luminal cells (PNT 1A) and the tumoral prostatecell line (DU 145) were a generous gift from O. Cus-senot (Hospital St. Louis, Paris, France). Recombinanthuman HARP was purified as previously described[21], and human recombinant FGF-2 was a kind giftfrom M. Mazue (Famitalia, Italy).
Cell Culture
PNT 1A, DU 145, and LNCaP cells were cultured inRPMI-1640 medium (Gibco BRL, France) containing,respectively, 5%, 5%, and 10% (v/v) fetal calf serum(FCS) (Eurobio, France) and antibiotics (100 U/mlpenicillin, 100 mg/ml streptomycin, and 0.5 mg/mlfungizone; Gibco BRL). The PNT 1A and LNCaPgrowth medium was supplemented with 1 nM dihy-drotestosterone (DHT) (Fluka, France). PC 3 cells werecultured in DMEM/HAM F12 medium (1:1; GibcoBRL) supplemented with 5% FCS and antibiotics. Bo-vine epithelial lens (BEL) cells were cultured in pres-ence of 1 ng/ml of FGF-2 as previously described [19].All cells were cultured at 37°C in an atmosphere con-taining 7.5% CO2.
Production of Antibodies to Recombinant HARP
Rabbit antibodies were produced by injecting 100mg of human recombinant HARP in Freund’s com-
HARP in Human Prostate Tumors 127

plete adjuvant in several subcutaneous fractions.Booster injections were given 4 and 8 weeks after pri-mary immunization. Immunoglobulins were first pu-rified by affinity on protein A-Sepharose, and specificanti-HARP immunoglobulins were then purified onHARP-Hitrap (Pharmacia Biotech, France) as recom-mended by the manufacturer. The specificity of anti-bodies was tested by Western blot analysis with hu-man recombinant HARP and MK, which shares 50%sequence homology with HARP.
Immunohistochemistry
Frozen sections of prostatic tissue (5-mm thickness)were fixed in acetone at 4°C for 10 min. After threewashes in phosphate-buffered saline (PBS), sectionswere incubated with 1.5% normal goat serum in PBSfor 30 min. The sections were then incubated for 90min at 37°C with anti-HARP (1 mg/ml) antibodies di-luted in PBS containing 0.2% Tween-20 and 1.5% nor-mal goat serum. Binding was revealed by using theABC alkaline phosphatase kit (Vectastin ABC-AP Kit,Vector Laboratories, Biosys, France). Briefly, the sec-tions were washed, and incubated with biotinylatedgoat anti-rabbit IgG (1/200) for 30 min at room tem-perature and then with avidin-biotin-linked alkalinephosphatase. The sections were rinsed and then incu-bated in alkaline phosphatase substrate (AlkalinePhosphatase Substrate Kit I, Vector Laboratories, Bio-sys) supplemented with 1 mM levamisol. The sectionswere washed and lightly counterstained with hema-toxylin.
Experiments using available monoclonal antibodieswere performed as described above, except that theantibodies were diluted 1:50 for anti-human Von Wil-lebrand factor (factor VIII-related antigen, Dako,France) and 1:200 for anti-human cytokeratin 18(Sigma, France). The alkaline phosphatase-conjugatedrabbit anti-mouse immunoglobulins were used at 1:50dilution (Dako, France).
Western Blot Analysis of HARP
Proteins were separated on SDS-15% polyacryl-amide gel [26] and were transferred to Immobilon-Pmembranes (Millipore, France) in 50 mM CAPS buffer,pH 11, containing 10% methanol for 20 min at 260 mA.Membranes were blocked with 3% gelatin in PBS-0.2%Tween-20, and then incubated with 1 mg/ml of puri-fied anti-HARP antibodies. The presence of antibodiesassociated with Immobilon-P was assayed usinghorseradish peroxidase-conjugated goat anti-rabbitIgG (Diagnostics Pasteur, France) and ECL chemilu-minescence (Amersham, France) according to themanufacturer’s recommendations.
In Situ Hybridization
Sections of 10-mm thickness were cut and thaw-mounted on Super Frost Plus slides. The sections werefixed with 4% paraformaldehyde in PBS for 30 min atroom temperature and treated with 10 mg/ml of pro-teinase K in PBS (Boehringer, France) for 30 min at37°C. In situ hybridization studies were done withdigoxygenin-labeled probes obtained by unidirec-tional PCR using a sense primer (58 GAAAATTTG-CAGCTGCCT 38; 300–319) and an antisense primer(58TTCTCCTGTTTCTTGCCT 38; 745–762) as de-scribed by Ledoux et al. [27].
Transfection Experiments
Approximately 5 × 104 PNT 1A cells/cm2 wereseeded and incubated overnight in DMEM/F12 (1/1)medium containing 5% FCS. The cells were then trans-fected with 10 mg of pJK12 [21] and 1 mg of pAG60[28], using a Transfectam kit (Promega, France) as rec-ommended by the manufacturer. Control cells weretransfected with only 10 mg of pAG60. Cells were se-lected in geneticin (G 418) (200 mg/ml) for 15 days.Resistant colonies were isolated and allowed to growas monoclonal populations. Clones expressing thehighest amounts of HARP, as determined by Westernblot analysis, were selected for further studies.
Growth Factor Assays
Thymidine incorporation assays were performed aspreviously described [19]. Briefly, cells (PC 3, DU 145,PNTIA, LNCaP, or BEL) were seeded at 1 × 104 cells/well in 48-well culture plates (Costar, France) ingrowth medium. The medium was discarded 72 hrlater and cells were maintained for 24 hr in FCS-freemedium. HARP was added for a further 24 hr. DNAsynthesis was determined by labeling the cells with[methyl 3H]thymidine (0.5 mCi/well) between 18–24hr after addition of growth factors.
Cell Proliferation Assay
Transfected cells were seeded at 3,500 cells/well in12-well culture plates (Costar) in DMEM/F12 (1/1)medium (1 ml/well) containing 5% FCS supple-mented with 200 mg/ml G 418. Medium was removed12 hr later and replaced by fresh medium containing1.25% FCS. Each day, cells were trypsinized andcounted in a Coulter counter. The samples were as-sayed in triplicate.
128 Vacherot et al.

Soft-Agar Growth Assays
The soft-agar assay was performed essentially asdescribed in Fang et al. [18]. Briefly, 40,000 cells in0.35% agar (Sigma) were plated on top of 1 ml of so-lidified 0.6% agar in a 35-mm dish. Growth mediumwas composed of DMEM/F12 (1/1) containing 5%FCS supplemented with 200 mg/ml of G 418 and wasincluded in both agar layers. Colonies more than 60mm in diameter were counted after 2–3 weeks of in-cubation.
RESULTS
Location of HARP Transcripts in Normal, BenignHyperplastic, and Adenocarcinoma Human
Prostate Tissue
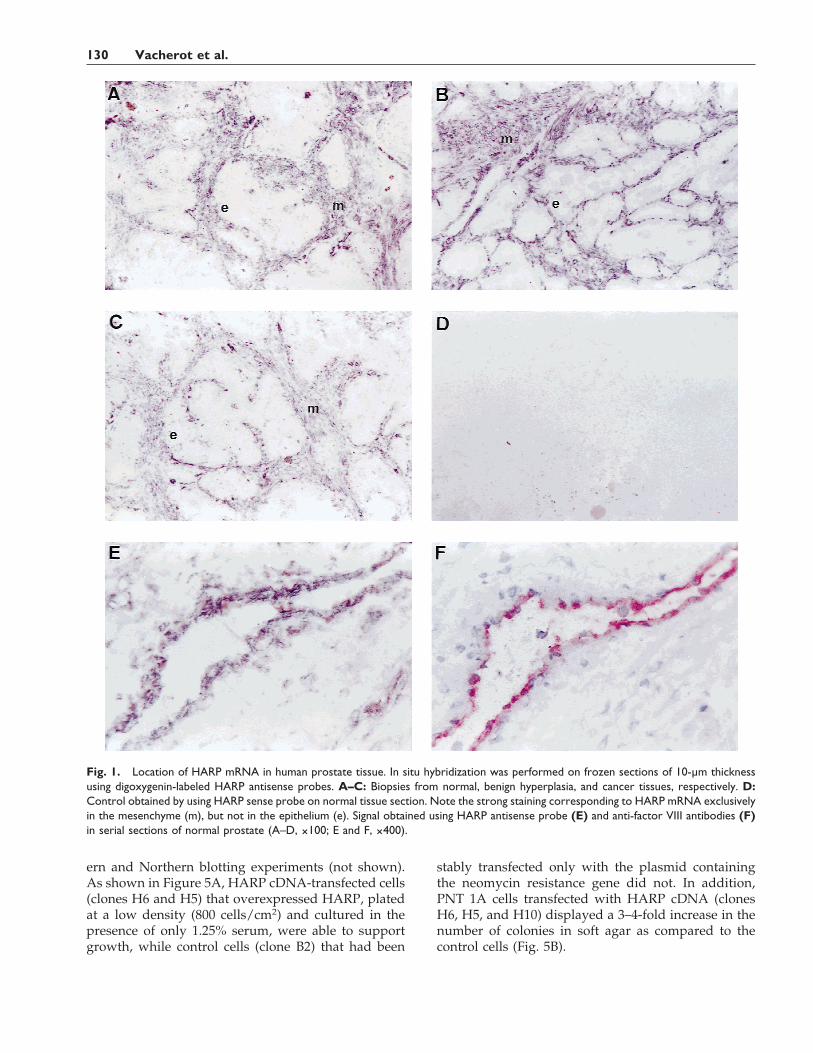
To study the functional role of HARP in the pros-tate, we investigated the cellular location of HARPmRNA by in situ hybridization on frozen sections ofhuman prostate tissue. In all the studied sections (5cases of normal prostate; 7 cases of BPH, and 9 cases ofPCa), HARP mRNA was localized to the fibromuscu-lar stroma in normal tissue (Fig. 1A), BPH (Fig. 1B),and PCa (Fig. 1C). As shown in Figure 1D, no signalwas detected using the HARP sense probe, attesting tothe specificity of staining. In all sections, studied in-cluding normal prostate, BPH, and PCa, no signal wasdetected in epithelial cells (Fig. 1A–C), whose pres-ence was confirmed by hematoxylin counterstaining.Endothelial cells were positive for HARP mRNA (Fig.1E, normal prostate section). We also observed a co-localization of mRNA staining with the one obtainedwith anti-factor VIII antibodies (Fig. 1F, normal pros-tate section) performed in serial sections of normalprostate.
Specificity of Anti-HARP Immunoglobulins
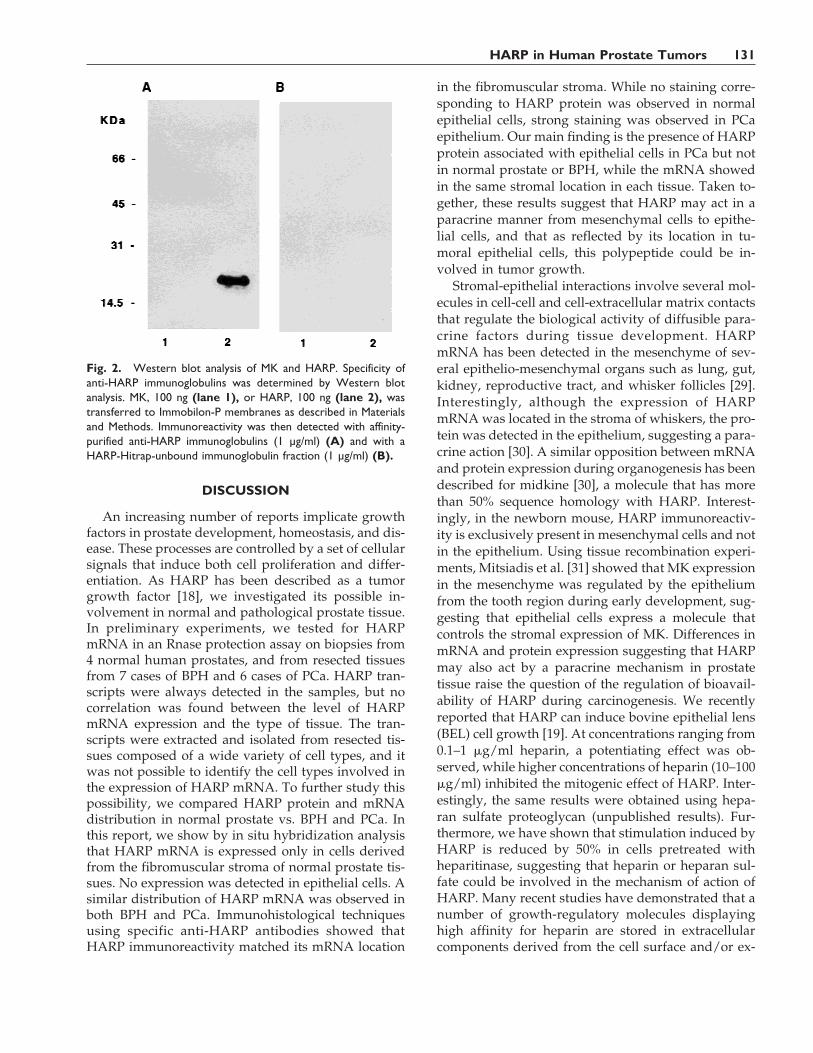
The specificity of affinity-purified anti-HARP im-munoglobulins was tested by Western blot analysis,using human recombinant HARP and also MK, whichshares 50% sequence homology with HARP. Asshown in Figure 2A, affinity-purified anti-HARPyielded a single immunoreactive signal correspondingto HARP protein (Fig. 2A, lane 2). In contrast, no sig-nal was observed with MK protein (Fig. 2A, lane 1),demonstrating a lack of cross-reactivity. No signal wasobserved using nonspecific immunoglobulins, show-ing the specificity of the anti-HARP immunoglobulinsused in this study (Fig. 2B, lanes 1 and 2).
Immunolocation of HARP Protein in Normal,Benign Hyperplastic, and Adenocarcinoma Human
Prostate Tissue
To locate HARP in prostate tissues and to confirmthat stromal cells expressed HARP, we studied immu-
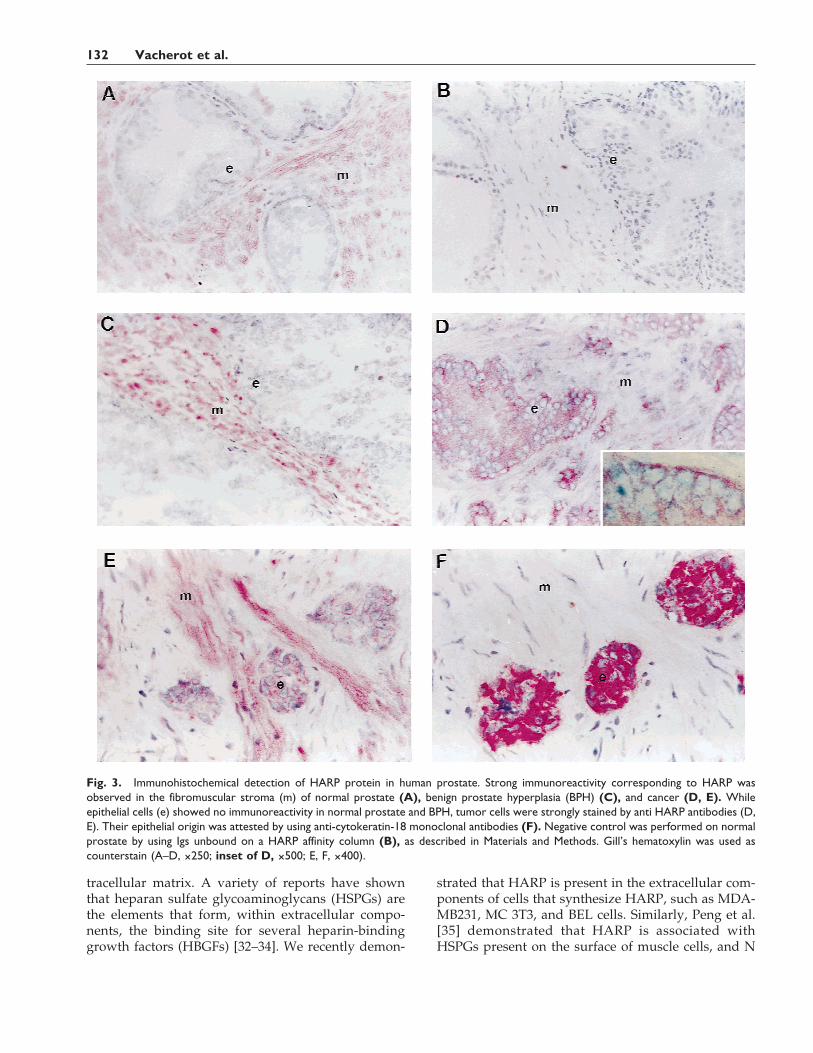
noreactivity in human prostate tissues with specificrabbit polyclonal anti-HARP immunoglobulins. Asshown in Figure 3, staining corresponding to HARPwas observed in the fibromuscular stroma of normalprostate (Fig. 3A), BPH (Fig. 3C), and PCa (Fig. 3D)tissue sections. While epithelial cells from normalprostate (5 cases) and BPH (7 cases) had no detectableimmunoreactivity (Fig. 3A,C), strong HARP staininglocalized to epithelial cells in all the histological slicesstudied from all 9 PCa samples investigated, as illus-trated in Figure 3D. In a preliminary attempt to betterlocate HARP on the epithelial cells, an enlarged mag-nification is presented (inset of Fig. 3D). However, atthis stage, it is unclear whether HARP binds exclu-sively at the cell surface. Interestingly, in biopsies ofinvasive carcinoma, clusters of epithelial cells werealso positive with anti-HARP antibodies (Fig. 3E).These tumoral cells were stained positively with anti-cytokeratin-18 monoclonal antibodies (Fig. 3F), dem-onstrating their epithelial origin. As shown in Figure3B, no immunostaining corresponding to HARP wasobserved with nonspecific antibodies used as negativecontrol.
Growth Properties of HARP for Human ProstateEpithelial Cells
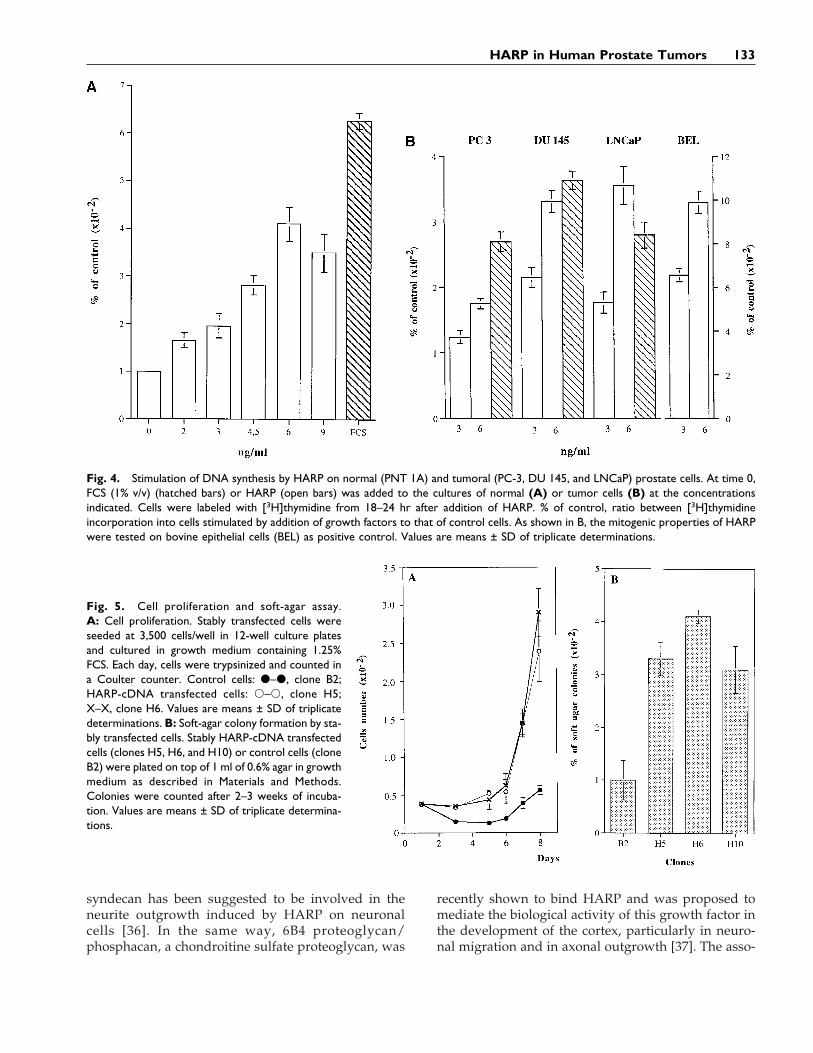
As previously reported, HARP displayed in vitromitogenic activity for a wide variety of cells includingepithelial cells derived from various sources. To testfor possible involvement of HARP in prostate physi-ology or pathophysiology, we tested the in vitro mi-togenic activity of HARP for normal (PNT 1A) andtumor (DU 145, LNCaP, and PC 3) epithelial cells de-rived from prostate tissues. As illustrated in Figure 4,addition to quiescent cells of various amounts of hu-man recombinant HARP induced the incorporation of[3H]thymidine in a concentration-dependent mannerin PNT 1A cells (Fig. 4A) and in DU 145 and LNCaPcell lines. A weak but significant response was ob-served with PC 3 cells (Fig. 4B). Contrary to DU 145and LNCaP cell lines, PC 3 cells expressed HARP [18](unpublished data), suggesting an autocrine stimula-tion of HARP on PC 3 cells.
In a second set of experiments, the mitogenic effectof HARP was investigated by using stably HARP-transfected PNT-1A cells and measuring clonalgrowth. Stably transfected PNT-1A cells were ob-tained by cotransfecting cells with pJK12 (containingfull-length human HARP cDNA) and pAG60 (contain-ing the resistance gene to neomycin). Neomycin-resistant cells were selected by incubation with 200mg/ml of G 418 and the resulting stable neomycin-resistant clones were selected for further studies.Overexpression of HARP was investigated by West-
HARP in Human Prostate Tumors 129
ern and Northern blotting experiments (not shown).As shown in Figure 5A, HARP cDNA-transfected cells(clones H6 and H5) that overexpressed HARP, platedat a low density (800 cells/cm2) and cultured in thepresence of only 1.25% serum, were able to supportgrowth, while control cells (clone B2) that had been
stably transfected only with the plasmid containingthe neomycin resistance gene did not. In addition,PNT 1A cells transfected with HARP cDNA (clonesH6, H5, and H10) displayed a 3–4-fold increase in thenumber of colonies in soft agar as compared to thecontrol cells (Fig. 5B).
Fig. 1. Location of HARP mRNA in human prostate tissue. In situ hybridization was performed on frozen sections of 10-µm thicknessusing digoxygenin-labeled HARP antisense probes. A–C: Biopsies from normal, benign hyperplasia, and cancer tissues, respectively. D:Control obtained by using HARP sense probe on normal tissue section. Note the strong staining corresponding to HARP mRNA exclusivelyin the mesenchyme (m), but not in the epithelium (e). Signal obtained using HARP antisense probe (E) and anti-factor VIII antibodies (F)in serial sections of normal prostate (A–D, ×100; E and F, ×400).
130 Vacherot et al.
DISCUSSION
An increasing number of reports implicate growthfactors in prostate development, homeostasis, and dis-ease. These processes are controlled by a set of cellularsignals that induce both cell proliferation and differ-entiation. As HARP has been described as a tumorgrowth factor [18], we investigated its possible in-volvement in normal and pathological prostate tissue.In preliminary experiments, we tested for HARPmRNA in an Rnase protection assay on biopsies from4 normal human prostates, and from resected tissuesfrom 7 cases of BPH and 6 cases of PCa. HARP tran-scripts were always detected in the samples, but nocorrelation was found between the level of HARPmRNA expression and the type of tissue. The tran-scripts were extracted and isolated from resected tis-sues composed of a wide variety of cell types, and itwas not possible to identify the cell types involved inthe expression of HARP mRNA. To further study thispossibility, we compared HARP protein and mRNAdistribution in normal prostate vs. BPH and PCa. Inthis report, we show by in situ hybridization analysisthat HARP mRNA is expressed only in cells derivedfrom the fibromuscular stroma of normal prostate tis-sues. No expression was detected in epithelial cells. Asimilar distribution of HARP mRNA was observed inboth BPH and PCa. Immunohistological techniquesusing specific anti-HARP antibodies showed thatHARP immunoreactivity matched its mRNA location
in the fibromuscular stroma. While no staining corre-sponding to HARP protein was observed in normalepithelial cells, strong staining was observed in PCaepithelium. Our main finding is the presence of HARPprotein associated with epithelial cells in PCa but notin normal prostate or BPH, while the mRNA showedin the same stromal location in each tissue. Taken to-gether, these results suggest that HARP may act in aparacrine manner from mesenchymal cells to epithe-lial cells, and that as reflected by its location in tu-moral epithelial cells, this polypeptide could be in-volved in tumor growth.
Stromal-epithelial interactions involve several mol-ecules in cell-cell and cell-extracellular matrix contactsthat regulate the biological activity of diffusible para-crine factors during tissue development. HARPmRNA has been detected in the mesenchyme of sev-eral epithelio-mesenchymal organs such as lung, gut,kidney, reproductive tract, and whisker follicles [29].Interestingly, although the expression of HARPmRNA was located in the stroma of whiskers, the pro-tein was detected in the epithelium, suggesting a para-crine action [30]. A similar opposition between mRNAand protein expression during organogenesis has beendescribed for midkine [30], a molecule that has morethan 50% sequence homology with HARP. Interest-ingly, in the newborn mouse, HARP immunoreactiv-ity is exclusively present in mesenchymal cells and notin the epithelium. Using tissue recombination experi-ments, Mitsiadis et al. [31] showed that MK expressionin the mesenchyme was regulated by the epitheliumfrom the tooth region during early development, sug-gesting that epithelial cells express a molecule thatcontrols the stromal expression of MK. Differences inmRNA and protein expression suggesting that HARPmay also act by a paracrine mechanism in prostatetissue raise the question of the regulation of bioavail-ability of HARP during carcinogenesis. We recentlyreported that HARP can induce bovine epithelial lens(BEL) cell growth [19]. At concentrations ranging from0.1–1 mg/ml heparin, a potentiating effect was ob-served, while higher concentrations of heparin (10–100mg/ml) inhibited the mitogenic effect of HARP. Inter-estingly, the same results were obtained using hepa-ran sulfate proteoglycan (unpublished results). Fur-thermore, we have shown that stimulation induced byHARP is reduced by 50% in cells pretreated withheparitinase, suggesting that heparin or heparan sul-fate could be involved in the mechanism of action ofHARP. Many recent studies have demonstrated that anumber of growth-regulatory molecules displayinghigh affinity for heparin are stored in extracellularcomponents derived from the cell surface and/or ex-
Fig. 2. Western blot analysis of MK and HARP. Specificity ofanti-HARP immunoglobulins was determined by Western blotanalysis. MK, 100 ng (lane 1), or HARP, 100 ng (lane 2), wastransferred to Immobilon-P membranes as described in Materialsand Methods. Immunoreactivity was then detected with affinity-purified anti-HARP immunoglobulins (1 µg/ml) (A) and with aHARP-Hitrap-unbound immunoglobulin fraction (1 µg/ml) (B).
HARP in Human Prostate Tumors 131
tracellular matrix. A variety of reports have shownthat heparan sulfate glycoaminoglycans (HSPGs) arethe elements that form, within extracellular compo-nents, the binding site for several heparin-bindinggrowth factors (HBGFs) [32–34]. We recently demon-
strated that HARP is present in the extracellular com-ponents of cells that synthesize HARP, such as MDA-MB231, MC 3T3, and BEL cells. Similarly, Peng et al.[35] demonstrated that HARP is associated withHSPGs present on the surface of muscle cells, and N
Fig. 3. Immunohistochemical detection of HARP protein in human prostate. Strong immunoreactivity corresponding to HARP wasobserved in the fibromuscular stroma (m) of normal prostate (A), benign prostate hyperplasia (BPH) (C), and cancer (D, E). Whileepithelial cells (e) showed no immunoreactivity in normal prostate and BPH, tumor cells were strongly stained by anti HARP antibodies (D,E). Their epithelial origin was attested by using anti-cytokeratin-18 monoclonal antibodies (F). Negative control was performed on normalprostate by using Igs unbound on a HARP affinity column (B), as described in Materials and Methods. Gill’s hematoxylin was used ascounterstain (A–D, ×250; inset of D, ×500; E, F, ×400).
132 Vacherot et al.
syndecan has been suggested to be involved in theneurite outgrowth induced by HARP on neuronalcells [36]. In the same way, 6B4 proteoglycan/phosphacan, a chondroitine sulfate proteoglycan, was
recently shown to bind HARP and was proposed tomediate the biological activity of this growth factor inthe development of the cortex, particularly in neuro-nal migration and in axonal outgrowth [37]. The asso-
Fig. 4. Stimulation of DNA synthesis by HARP on normal (PNT 1A) and tumoral (PC-3, DU 145, and LNCaP) prostate cells. At time 0,FCS (1% v/v) (hatched bars) or HARP (open bars) was added to the cultures of normal (A) or tumor cells (B) at the concentrationsindicated. Cells were labeled with [3H]thymidine from 18–24 hr after addition of HARP. % of control, ratio between [3H]thymidineincorporation into cells stimulated by addition of growth factors to that of control cells. As shown in B, the mitogenic properties of HARPwere tested on bovine epithelial cells (BEL) as positive control. Values are means ± SD of triplicate determinations.
Fig. 5. Cell proliferation and soft-agar assay.A: Cell proliferation. Stably transfected cells wereseeded at 3,500 cells/well in 12-well culture platesand cultured in growth medium containing 1.25%FCS. Each day, cells were trypsinized and counted ina Coulter counter. Control cells: d–d, clone B2;HARP-cDNA transfected cells: s–s, clone H5;X–X, clone H6. Values are means ± SD of triplicatedeterminations. B: Soft-agar colony formation by sta-bly transfected cells. Stably HARP-cDNA transfectedcells (clones H5, H6, and H10) or control cells (cloneB2) were plated on top of 1 ml of 0.6% agar in growthmedium as described in Materials and Methods.Colonies were counted after 2–3 weeks of incuba-tion. Values are means ± SD of triplicate determina-tions.
HARP in Human Prostate Tumors 133

ciation of HARP with proteoglycans could be one invitro mechanism that controls both the bioavailabilityand the biological activity of HARP, and it is temptingto speculate that this mechanism could occur in vivoin prostate tissue.
We also present evidence that endothelial cells ex-press HARP mRNA. In addition, we detected HARPmRNA in capillaries derived from adult rat brain,uterus [38], and human breast [27]. Several studies,including our own, have demonstrated that purifiedhuman recombinant HARP is mitogenic for endothe-lial cells [17,18,20] and is also able to exert angiogenicactivity in vitro and in vivo [21,23]. Recent reportshave demonstrated that HARP, and more preciselythe C-terminal part of the molecule, enhance plasmin-ogen activator levels and downregulate PAI-1 expres-sion in ABAE cells, supporting the functional role ofHARP in angiogenesis [39]. Similar results have beenreported with MK [40]. Recently, it was shown thatHARP belongs to the family of heparin-releasable pro-teins [41]. This family, that includes lipoprotein lipase,tissue factor pathway inhibitor, extracellular superox-ide dismutase, histaminase, platelet factor 4, and tu-mor necrosis factor-binding protein-1, contributes tovascular homeostasis. Although the precise physi-ological role of HARP located in blood vessels remainsto be clarified, it is tempting to speculate that HARPprotein may play an important role in angiogenesismechanisms during physiological or pathophysiologi-cal processes, particularly tumor progression. It isnoteworthy that the physiological function of HARPin normal or tumoral epithelial cells remains un-known. Interestingly, although a growth-promotingeffect of HARP has been observed in vitro on normalepithelial cells, no HARP immunoreactivity was ob-served in situ in normal and benign hyperplasic tis-sues, leading to speculation that no (or no available)HARP binding sites were present in situ in normalepithelial cells. Taken together, these observationssuggest that stroma components or extracellular ma-trix components could regulate the presence or avail-ability of HARP-binding sites on the surface of normalepithelial cells that display a low cellular proliferationindex. It has been extensively reported that cells pro-liferating in tissue culture in response to variousgrowth factors and expressing the correspondinggrowth factor receptor do not respond to the samegrowth factor when these cells are in the original tis-sue. The absence of HARP protein immunostaining intissue sections containing prostatic epithelial cells istherefore not in contradiction with the fact that in tis-sue culture, normal epithelial prostatic cells such asPNT 1A cells respond to HARP. Further in vitro stud-ies using coculture systems are required to character-ize more precisely these regulatory components.
The location of HARP polypeptide in tumor epithe-lial cells points to the presence of an unidentified spe-cific receptor. The presence of high-affinity bindingsites for HARP in NIH 3T3 cells has been reported[42]. These high-affinity binding sites were also foundin other cell types, including normal rat kidney cells(NRK), human mammary adenocarcinoma cells (SK-BR-3), human epidermoid carcinoma cells (A431), hu-man hepatocarcinoma cells (HepG2), mouse neuro-blastoma cells (NB41A3), and pheochromocytomacells (PC12). However, it is noteworthy that all thesestudies were performed with HARP that had little orno mitogenic activity for the cells used. The character-ization and regulation of the expression of this recep-tor during cell transformation are extremely promis-ing in the investigation of HARP involvement in PCa.
It is noteworthy that several heparin bindinggrowth factors were suggested to be involved in pros-tatic carcinoma progression. As previously reported,the basic fibroblast growth factor (FGF 2) is the maingrowth factor produced by human prostatic stromalcells [43]. In BPH, immunoreactive FGF 2 was found instromal cells, and no immunostaining was seen in theprostatic epithelium. In prostatic carcinomas, FGF 2was present both in mesenchymal and tumor cells.Immunostaining in tumor cells ranged from weakly tostrongly positive [44,45]. In prostatic cell cultures, FGF2 is a potent mitogen for mesenchymal cells and, to alesser extent, for epithelial cells [43,46]. FGF 2 tran-scripts were detected in DU-145 and PC-3 cells but notin LNCaP cells [45]. In this context, the normal pros-tatic epithelial cell line (PNT 1A), transfected with FGF2 cDNA, acquired a transforming phenotype [47]. Thekeratinocyte growth factor (KGF), another member ofthe FGF family, has been reported to have a key role asparacrine factor in the human prostate [48]. KGFmRNA is produced exclusively by stromal cells, andthe protein stimulates only the growth of epithelialcells [49,50]. The gene product of fibroblast growthfactor receptor 2 (FGFR 2) was found in the humanprostate, and the FGFR 2 IIIb isoform was only de-tected in the primary prostate epithelial cell cultures.During the development of prostate cancer in a ratmodel, the FGF binding characteristics of prostate epi-thelium changed to a higher affinity for FGF 2 andFGF 1 but not for KGF, resulting from a switch ofFGFR expression from the IIIb to the IIIc isoform [51].Structural modification and an increased level ofgrowth factor receptors could be associated with pros-tate cancer growth and progression. In this way, tu-mor cells have been reported to overexpress c-METprotein, the hepatocyte growth factor receptor. c-METprotein was slightly detected by immunohistochemis-try in normal and hyperplastic glands and wasstrongly observed in adenocarcinoma [52,53].
134 Vacherot et al.

The hormonal regulation of HARP mRNA has beeninvestigated using prostate-derived epithelial cells.HARP mRNA synthesis is upregulated by dihydrotes-tosterone and inhibited by anandron, a specific andro-gen inhibitor [54]. Our data raise questions about theputative hormonal regulation of HARP mRNA duringhormone-dependent pathophysiological processessuch as prostate tumor progression.
In conclusion, our results point to a crucial role ofHARP in the pathophysiology of prostate tissue.HARP could act as a tumor growth factor for epithelialcells in PCa, and its inhibition in vivo is an excitingnew research area for prostate cancer therapy.
ACKNOWLEDGMENTS
F.V. is the recipient of a grant from the Associationpour la Recherche sur les Tumeurs de Prostate.
REFERENCES
1. Brandes D. Prostate gland embryology, anatomy and histology.In: Mill GS, editor. Uropathology. New York: Churchill Living-stone; 1989. p 1165–1180.
2. Cunha GR, Donjacour AA, Cooke PS, Mee S, Bigsby RM, Hig-gins SJ. The endocrinology and the developmental biology ofthe prostate. Endocr Rev 1987;8:338–363.
3. Bissel MJ, Hall HG, Parry GJ. How does the extracellular matrixdirect gene expression? Theor Biol 1982;99:31–68.
4. Streuli CH, Bailey N, Bissel MJ. Control of mammary epithelialdifferentiation: basement membrane induces tissue-specificgene expression in the absence of cell-cell interaction and mor-phological polarity. J Cell Biol 1991;115:1383–1395.
5. Cunha GR, Alarid ET, Donjacour AA, Boutin EL, Foster BA.Normal and abnormal development of the male urogenital tract:role of androgens, mesenchymal-epithelial interactions andgrowth factors. J Androl 1992;13:85–105.
6. Chang SM, Chung LWK. Interactions between prostatic fibro-blasts and epithelial cells in culture. Endocrinology 1989;125:2719–2727.
7. Sakakura T, Nishizuka Y, Dawe CJ. Mesenchyme-dependentmorphogenesis and epithelium-specific cytodifferentiation inmouse mammary gland. Science 1976;194:1439–1441.
8. Story MT. Regulation of prostate growth by fibroblast growthfactors. World J Urol 1995;13:297–305.
9. Cunha GR, Foster B, Thomson A, Sugimura Y, Tanji N, Tsuji M,Terada N, Finch PW, Donjacour AA. Growth factors as media-tors of androgen action during the development of the maleurogenital tract. World J Urol 1995;13:264–276.
10. Kadomatsu K, Tomomura M, Muramatsu T. cDNA cloning andsequence of a new gene intensely expressed in early differen-tiation stages of embryonal carcinoma cells and in mid-gestationperiod of mouse embryogenesis. Biochem Biophys Res Com-mun 1988;151:1312–1318.
11. Raulais D, Lagente-Chevallier O, Guettet C, Duprez D, CourtoisY, Vigny M. A new heparin binding protein regulated by reti-noic acid from chick embryo. Biochem Biophys Res Commun1991;174:708–715.
12. Merenmies J, Rauvala H. Molecular cloning of the 18-kDagrowth associated protein of developing brain. J Biol Chem1990;265:16721–16724.
13. Li YS, Milner PG, Chauhan AK, Watson MA, Hoffman RM,Kodner CM, Milbrandt J, Deuel TF. Cloning and expression of adevelopmentally regulated protein that induces mitogenic andneurite outgrowth activity. Science 1990;250:1690–1694.
14. Tezuka K, Takeshita S, Haneda Y, Kumegawa M, Kikuno R,Hashimoto-Gotoh T. Isolation of mouse and human cDNAclones encoding a protein expressed specifically in osteoblastsand brain tissues. Biochem Biophys Res Commun 1990;173:246–251.
15. Rauvala H. An 18-kd heparin-binding protein of developingbrain that is distinct from fibroblast growth factors. EMBO J1989;8:2933–2941.
16. Laaroubi K, Vacherot F, Delbe J, Caruelle D, Barritault D,Courty J. Biochemical and mitogenic properties of the heparin-binding growth factor HARP. Prog Growth Factor Res 1995;6:25–34.
17. Courty J, Dauchel MC, Caruelle D, Perderiset M, Barritault D.Mitogenic properties of a new endothelial cell growth factorrelated to pleiotrophin. Biochem Biophys Res Commun 1991;180:145–151.
18. Fang W, Hartmann N, Chow DT, Riegel AT, Wellstein A. Pleio-trophin stimulates fibroblasts and endothelial and epithelialcells and is expressed in human cancer. J Biol Chem 1992;267:25889–25897.
19. Delbe J, Vacherot F, Laaroubi K, Barritault D, Courty J. Effect ofheparin on bovine epithelial lens cells proliferation induced byheparin affin regulatory peptide. J Cell Physiol 1995;164:47–54.
20. Wellstein A, Fang W, Khatri A, Lu Y, Swain SS, Dickson RB,Sasse J, Riegel AT, Lippman ME. A heparin-binding growthfactor secreted from breast cancer cells homologous to a devel-opmentally regulated cytokine. J Biol Chem 1992;267:2582–2587.
21. Laaroubi K, Delbe J, Vacherot F, Desgranges P, Tardieu M, JayeM, Barritault D, Courty J. Mitogenic and in vitro angiogenicactivity of human recombinant heparin affin regulatory peptide.Growth Factors 1994;10:89–98.
22. Chauhan AK, Li YS, Deuel TF. Pleiotrophin transforms NIH 3T3cells and induces tumors in nude mice. Proc Natl Acad Sci USA1993;90:679–682.
23. Choudhuri R, Zhang HT, Donnini S, Ziche M, Bicknell R. Anangiogenic role for the neurokines midkine and pleiotrophin intumorigenesis. Cancer Res 1997;57:1814–1819.
24. Schroder FH, Hermanek P, Denis L, Fair WR, GospodarowiczMK, Pavone-Maculaso M. The TNM classification of prostatecancer. Prostate 1992;4:129–138.
25. Gleason DF. Histologic grading of prostate cancer. Hum Pathol1992;23:273–279.
26. Laemmli UK. Cleavage of structural proteins during the assem-bly of the head of bacteriophage T4. Nature 1970;227:680–685.
27. Ledoux D, Caruelle D, Liu J, Crepin M, Barritault D, Courty J.Cellular distribution of the angiogenic factor heparin affin regu-latory peptide (HARP) mRNA and protein in the human mam-mary gland. J Histochem Cytochem 1997;45:1239–1245.
28. Colbere-Garapin F, Horodniceanu F, Kourilsky P, Garapin AC.A new dominant hybrid selective marker for higher eucaryoticcells. J Mol Biol 1981;150:1–14.
29. Vanderwinden JM, Mailleux P, Schiffmann SN, VanderhaeghenJJ. Cellular distribution of the new growth factor pleiotrophin(HB-GAM) mRNA in developing and adult rat tissues. AnatEmbryol (Berl) 1992;186:387–406.
30. Mitsiadis TA, Salmivirta M, Muramatsu T, Muramatsu H, Rau-vala H, Lehtonen E, Jalkanen M, Thesleff I. Expression of theheparin-binding cytokines, midkine (MK) and HB-GAM (pleio-trophin), is associated with epithelial-mesenchymal interactionsduring fetal development and organogenesis. Development1995;121:37–51.
HARP in Human Prostate Tumors 135

31. Mitsiadis TA, Muramatsu T, Muramatsu H, Thesleff I. Midkine(MK), a heparin-binding growth/differentiation factor, is regu-lated by retinoic acid and epithelial-mesenchymal interactionsin the developing mouse tooth, and affects cell proliferation andmorphogenesis. J Cell Biol 1995;129:267–281.
32. Vigny M, Ollier-Hartmann MP, Lavigne M, Fayein N, JeannyJC, Laurent M, Courtois Y. Specific binding of basic fibroblastgrowth factor to basement membrane-like structures and to pu-rified heparin sulphate proteoglycan of the EHS tumor. J CellPhysiol 1988;137:321–328.
33. Gitay-Goren H, Soker S, Vlodavsky I, Neufeld G. The binding ofvascular endothelial growth factor to its receptor is dependenton cell surface-associated heparin-like molecules. J Biol Chem1992;267:6093–6098.
34. Chernousov MA, Carey DJ. N-syndecan (syndecan 3) from neo-natal rat brain binds basic fibroblast growth factor. J Biol Chem1993;268:16810–16814.
35. Peng HB, Ali AA, Dai Z, Daggett DF, Raulo E, Rauvala H. Therole of heparin-binding growth-associated molecule (HB-GAM)in the postsynaptic induction in cultured muscle cells. J Neuro-sci 1995;15:3027–3038.
36. Raulo E, Chernousov MA, Carey DJ, Nolo R, Rauvala H. Isola-tion of a neuronal cell surface receptor of heparin bindinggrowth-associated molecule (HB-GAM). J Biol Chem. 1994;269:12999–13004.
37. Maeda N, Nishiwaki T, Shintani T, Hamanaka H, Noda M. 6B4:proteoglycan/phosphacan, an extracellular variant of receptor-like protein-tyrosine phosphatase j/RPTPb, binds pleiotro-phin/heparin-binding growth-associated molecule (HB-GAM).J Biol Chem 1996;271:21446–21452.
38. Milhiet PE, Vacherot F, Caruelle, JP, Barritault D, Caruelle D,Courty J. Upregulation of the angiogenic factor heparin affinregulatory peptide by progesterone in rat uterus. J Endocrinol1998;158:389–399.
39. Kojima S, Inui T, Kimura T, Sakakibara S, Muramatsu H, Ama-numa H, Maruta H, Muramatsu. Synthetic peptides derivedfrom midkine enhance plasminogen activator activity in bovineaortic endothelial cells. Biochem Biophys Res Commun 1995;206:468–473.
40. Kojima S, Muramatsu H, Amanuma H, Muramatsu T. Midkineenhances fibrinolytic activity of bovine endothelial cells. J BiolChem 1995;270:9590–9596.
41. Novotny WF, Maffi T, Mehta RL, Milner PG. Identification ofnovel heparin-releasable proteins, as well as the cytokines mid-kine and pleiotrophin, in human postheparin plasma. Arterio-scler Thromb 1993;13:1798–1805.
42. Kuo MD, Huang SS, Huang JS. Characterization of heparin-binding growth-associated factor receptor on NIH 3T3 cells. Bio-chem Biophys Res Commun 1992;182:188–194.
43. Story MT, Livingston B, Baeten L, Swartz SJ, Jacobs SC, BegunFP, Lawson RK. Cultured human prostate-derived fibroblastsproduce a factor that stimulates their growth with propertiesindistinguishable from basic fibroblast growth factor. Prostate1989;15:355–365.
44. Sinowatz F, Amselgruber W, Lincoln D, Sasse J, Kolle S, PlendlJ, Kayser K. Role of basic fibroblast growth factor in prostatictumors. Nutrition 1995;11:619–621.
45. Cronauer MV, Hittmair A, Eder IE, Hobisch A, Culig Z,Ramoner R, Zhang J, Bartsch G, Reissigl A, Radmayr C, Thurn-her M, Klocker H. Basic fibroblast growth factor levels in cancercells and in sera of patients suffering from proliferative disor-ders of the prostate. Prostate 1997;31:223–233.
46. McKeehan WL, Adams PS, Fast D. Different hormonal require-ments for androgen-independent growth of normal and tumorepithelial cells from rat prostate. In Vitro Cell Dev Biol 1987;23:147–152.
47. Ropiquet F, Berthon P, Villette JM, Le Brun G, Maitland NJ,Cussenot O, Fiet J. Constitutive expression of FGF2/bFGF innon-tumorigenic human prostatic epithelial cells results in theacquisition of a partial neoplastic phenotype. Int J Cancer 1997;72:543–547.
48. Yan G, Fukabori Y, Nikolaropoulos S, Wang F, McKeehan WL.Heparin-binding keratinocyte growth factor is a candidate stro-mal-to-epithelial-cell andromedin. Mol Endocrinol 1992;6:2123–2128.
49. Story MT, Hopp KA, Molter M, Meier DA. Characteristics ofFGF-receptors expressed by stromal and epithelial cells culturedfrom normal and hyperplastic prostates. Growth Factors 1994;10:269–280.
50. Ittman M, Mansukhani A. Expression of fibroblast growth fac-tors (FGFs) and FGF receptors in human prostate. J Urol 1997;157:351–356.
51. Yan G, Fukabori Y, McBride G, Nikolaropolous S, McKeehanWL. Exon switching and activation of stromal and embryonicfibroblast growth factor (FGF)-FGF receptor genes in prostateepithelial cells accompany stromal independence and malig-nancy. Mol Cell Biol 1993;13:4513–4522.
52. Pisters LL, Troncoso P, Zhau HE, Li W, von Eschenbach AC,Chung LW. c-met proto-oncogene expression in benign and ma-lignant human prostate tissues. J Urol 1995;154:293–298.
53. Humphrey PA, Zhu X, Zarnegar R, Swanson PE, Ratliff TL,Vollmer RT, Day ML. Hepatocyte growth factor and its receptor(c-MET) in prostatic carcinoma. Am J Pathol 1995;147:386–396.
54. Vacherot F, Laaroubi K, Caruelle D, Delbe J, Barritault D, Ca-ruelle JP, Courty J. Upregulation of heparin-affin regulatorypeptide by androgen. In Vitro Cell Dev Biol 1995;31:647–648.
136 Vacherot et al.


















