G. Cozon Février 2010allergo.lyon.inserm.fr/M1_2009-2010/Cozon_11-02-2010.pdfintestinaux,...
90
Transcript of G. Cozon Février 2010allergo.lyon.inserm.fr/M1_2009-2010/Cozon_11-02-2010.pdfintestinaux,...

Introduction Généralités
• Équilibre entre les 2 partenaires• Co-évolution progressive de l'espèce hôte
et de l'espèce parasitaire.• But de chaque être vivant : survivre, se
multiplier pour assurer sa descendance.• Un pathogène trop agressif qui tue son
hôte s'expose à ne plus trouver de terrain de développement. Ex la myxomatose en Australie

Co-sélection de l'hôte et de son parasite
• Développement de mécanismes de résistances aux parasites et de mécanismes d'échappement à cette résistance.
• Impératifs, buts de chaque espèce :– de l'agent infectieux : survivre et se multiplier pour
assurer sa descendance– de l'hôte : prévenir ou limiter la colonisation de ces
tissus
• Équilibre entre le pouvoir pathogène de l'agent infectieux et les mécanismes de défense de l'hôte.

Équilibre entre le pouvoir pathogène de l'agent infectieux et les mécanismes de
défense de l'hôte • Équilibre progressif au cours de l'évolution par
des sélections successives de l'agent infectieux et de son hôte.
• Sélection d’hôtes résistants
• Sélection de parasites capables d’échapper aux moyens de défense.
• Adaptation réciproque– Adaptation au Système immunitaire– Adaptation au traitement (ex chloroquine et palu)

Exemple d’équilibre hôte-parasite
• Paludisme = maladie grave protozoaire• Drépanocytose homozygote maladie grave du GR• Protection des hétérozygotes vis-à-vis du
paludisme.

Accidents de cette co-évolution
• Myxomatose du lapin• Vaccination antivariolique

Prévalence des parasitoses (1)

Prévalence des parasitoses (2)

Prévalence des parasitoses (3)

Mécanismes de la pathogénicité des parasites
• Pathogénicité par spoliation• Pathogénicité par traumatisme et point de
départ de surinfections.• Pathogénicité mécanique• Pathogénicité par toxicité• Pathogénicité par action immunodépressive
du parasite sur son hôte• Pathogénicité par mécanisme allergique• Pathogénicité par mécanisme
immunopathologique

Pathogénicité par spoliation
• Le plus souvent la spoliation induite par les parasites est insignifiante, parfois importante.
• Anémie des ankylostomes par spoliation en GR.
• Anémie de type Biermérienne de la Bothriocéphalose par carence en Vit B12 captée par ce cestode

Ankylostome
L'ankylostome adulte est un petit (1 cm) ver rond qui vit 4 à 7 ans dans le
jéjunum ou le duodénum.L'infestation massive peut causer une anémie grave

Pathogénicité par traumatisme et point de départ de surinfections
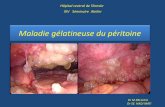
• Amibe du colon � une inflammation puis une érosion et enfin une ulcération muqueuse siège de surinfection bactérienne et d'abcès amibien en bouton de chemise. Les complications mécaniques seront la perforation de la paroi intestinale et la possibilité de péritonite.
• La dracunculose ou filaire de Médine • Migration anarchique grave d'anguillule
(cerveau)• Parasites vecteurs de maladies infectieuses :
tiques et borréliose ou encéphalite à tiques

Pathogénicité mécanique
• Rupture de cellules infectées : GR et paludisme anémie
• Occlusion intestinale par un amas des vers (ascaris)
• Occlusion des canaux biliaires (douves + réaction inflammatoire, angiocholite)
• Obstruction lymphatique (filaire lymphatique éléphantiasis)
• Envahissement d'un tissu sain (échinococcose alvéolaire)

Amas d’ascaris dans un intestin de chevalhttp://www.cvm.okstate.edu/~users/jcfox/htdocs/clinpara/lecture.htm
Éléphantiasis modéré http://www.tigr.org/tdb/e2k1/bma1/elephantiasis.shtml

Pathogénicité par toxicité
• Toxicité locale : amibes histolytiques abcès
• Toxicité neurotrope (ascaris, ixode)• Toxicité générale à type de fièvre prurit à
distance, urticaire

Pathogénicité par action immunodépressive du parasite
sur son hôte
• Paludisme• Bilharziose• Leishmaniose

Pathogénicité par mécanisme allergique
• Ascaridiose• Onchocercose• Distomatose urticaire à distance• Filariose avec la loase : encéphalite
allergique fatale si lyse massive de parasite par un traitement filaricide trop agressif.

Pathogénicité par mécanisme immunopathologique
• Hémolyse par des anticorps dans le paludisme
• CIC glomérulonéphrite• Pathogénicité par la formation d'un
granulome inflammatoire sérofibrineux dont le but est de contenir le parasite : sclérose fibrose hépatique ou vésicale lors des bilharzioses.

Mécanismes de défense de l'hôte contre le parasite
• Les mécanismes spécifiques– Les anticorps
– Les lymphocytes T et B
• Les mécanismes non spécifiques– Mécanismes solubles ou humoraux– Mécanismes cellulaires

Anticorps et parasites
• Activation du complément (sporozoïtes du plasmodium, Trypano, vers intestinaux)
• Inhibition de fixation du parasite (Ac anti-mérozoïtes du plasmodium, Ac anti-chondroïtine-4-sulfate et primipare, cryptosporidium)
• Activation de la phagocytose (Plasmodium, Trypanosoma brucei)
• ADCC éosino, neutro, plaquettes, macrophages dans les filarioses, Trypanosoma cruzi, Trichinellaspiralis, Schistosoma mansoni).
• Activation mastocytaire (IgE) pour les parasites intestinaux, hypersécrétion de mucus et augmentation du péristaltisme

Lymphocytes T et parasites
• Fonction auxiliaire Th1, Th2, Th3 variable selon les parasites soit protectrice soit délétère. Ex Leishmania
• Fonction cytotoxique (perforine, granzyme, FAS-Ligand) Ex Toxoplasma gondii

Les mécanismes non spécifiques
• Mécanismes solubles ou humoraux– complément voie classique ou alterne (Trichinella
spiralis, les schistosomules de Schistosoma mansoni)– Protéine C réactive (opsonisation, activation du C.)�
protection (poisson) ou facilitation (leishmania)– L'haptoglobuline + protéine proche de
l'haptoglobuline +hémoglobine = complexe à activité peroxydasique (résistance spontanée de l'homme à Trypanosoma brucei brucei.)

Mécanismes non spécifiques cellulaires
• Monocyte/macrophage, polynucléaires neutrophiles, basophiles et éosinophiles, mastocytes et plaquettes,
• Récepteurs de surface FcR IgG, IgE IgA• Stimulation des cellules par des cytokines

Monocyte/macrophage
• Rôle central• Cellule hôte• Cellule présentatrice d'antigène• Cellule effectrice de l'immunité anti-parasitaire• Cellule régulatrice (monokines de l’inflammation,
IL-10, PgE2, TGFß)• Activation de fibroblastes (cicatrisation granulome)

Phagocytose
• FcR, Récepteurs du complément, récepteurs du mannose et fucose, «Toll-like receptors»
• Importance dans les trypanosomiases (T brucei, ou T. rhodesiense) diminution rapide de la parasitémie.
• La phagocytose médiée par des molécules d'opsonine permet parfois l'entrée du parasite (L. donovani )

Phagolysosome
earlyendosomes
lateendosomes
lysosomes

Moyens de destruction des MΦ
• Radicaux libres de l'oxygène (O2-, H2O2, OH•) des peroxysomes.
• Enzymes lysosomales pH acide• Monoxyde d'azote.
• Nécessité d’un signal d’activation (IFNγet/ou TNFα)
• Libération de monokines de l’inflammation (IL-1, TNFα) parfois délétères (neuropalu)

Le polynucléaire neutrophile
• Importance dans certaines parasitoses– Trypanosoma cruzi– larves de S. mansoni
– nématodes comme Trichinella spiralis
Trichinose avec œdème périorbitaire, dleursmusculaires, diarrhée, éosino 28%

Les polynucléaires éosinophiles.
• augmentés dans les infections par les helminthes
• sous la dépendance d'IL-5 : la cytokine facteur de croissance pour les éosinophiles (eosino-CSF)
• dégranulation au contact du parasite

Contenu des granules des éosinophiles.
• Des arylsulfatases, bêta-glucuronidase, collagénase
• protéine basique majeure (neutralisant l'héparine)
• Neurotoxine ou EDN/EPX, • Peroxydase ou EPO, • Protéine Cationique de l'Éosinophile ou ECP

L'éosinophile = cellule inflammatoire ,
• médiateurs lipidiques (leucotriènes, prostaglandines, thromboxane B2, facteur d'activation plaquettaire ou PAF-acether...),
• des chemokines (RANTES, MIP-1a , IL-8) • des cytokines pro-inflammatoires (IL-1, IL-6,
TNFα).

L'éosinophile et parasite
• Cytolytique pour les parasites multicellulaires comme les helminthes :– Schistosomules, ou T. spiralis.

MASTOCYTE
• Histamine,• Héparine• Protéase (tryptase) • Dérivés de l’acide arachidonique :
prostaglandines et leucotriènes : les E-CFA et N-CFA (eosinophilic et neutrophilic chemotacticfactor of anaphylaxis)

Les plaquettes
• Granules lysosomiaux: contenant des phosphatases acides, aryl sulfatases, collagénases,
• Action cytotoxique vis à vis des parasites comme les schistosomules, T. gondii et T cruzi.

Réponse anti-parasitaire dans les muqueuses

Réponse anti-parasitaire anti-helminthes
systémiques

Variations des réponses anti-helminthes

Mécanismes d'échappement parasitaire aux défenses de l'organisme
• Adaptation fruit de la pression de sélection• Résistance à la destruction par la cascade
du complément• Dissimulation par masquage ou
recouvrement antigénique• Diversion par leurre antigénique• Commutation antigénique • Modification des réponses immunitaires

Résistance à la destruction par la cascade du complément
• Tachyzoïtes de Toxoplasma gondii non activateurs de la voie alterne par pauvreté en protéines et glycoprotéines de surface.
• Trypanosoma cruzi possède à sa surface une glycoprotéine de 87-93 kDa qui présente 50% d'homologie avec la protéine membranaire DAF (facteur accélérateur de dissociation)
• Schistosomules de S. mansoni captent les protéines DAF
• Promastigotes de Leishmania major

Le cycle parasitaire de Leischmania
Stade intracellulaireHôte les mammifères
Stade extracellulaire flagelléHôte le phlébotome

Métacyclogénèse du promastigote
stade extracellulaire • Promastigote procyclique non infestant• Promastigote métacyclique
– transformation des lipophosphoglycanes (LPG) de surface qui s’allongent
– migration vers le pharynx de l'insecte

Lipophosphoglycanes (LPG)de surface
• Glycocalix dense qui recouvre entièrement le parasite.
• 4 constituants :– Structure terminale disaccharidique de
mannose,– un polymère de disaccharides phosphorylés
répétitif,– un noyau de phosphosaccharides et– un ancrage lipidique phosphatidylinositol.

Promastigote
• Forme procyclique : LPG courts (7 nm) avec des résidus terminaux de galactose qui permettent l'ancrage du parasite sur les molécules lectines-like des cellules épithéliales du phlébotome.
• Forme métacyclique : allongement des séquences répétées (17nm).
• Résistance à la lyse par le système du complément de son hôte vertébré.


Inhibition du complexe d'attaque C5b-C9 = complexe membranaire d'attaque (CMA)
Par mutagenèse la diminution de la longueur des LPG diminue la virulence de L. Major car le CMA peut alors lyser le parasite
Protéine kinase extracellulaire gp63 permet la phosphorylation du composant C3 du complément et donc son inactivation en C3bi favorisant l’opsonisation du parasite et la fixation sur le CR3.

Dissimulation par masquage ou recouvrement antigénique
• les schistosomules peuvent se travestir en se recouvrant de molécules de l'hôte : glycolipides de groupes sanguins A, B, O, glycoprotéines diverses comme les molécules du complexe majeur d'histocompatibilité de classe I et II. Le ver n'est alors plus reconnu comme étranger à l'organisme mais comme étant un constituant toléré de l'organisme hôte.
• Expression d’une protéine proche d'une protéine inhibitrice de l'organisme : T. cruzi exprime une protéine de type DAF capable de bloquer l'activation de la cascade du complément.

Diversion par leurre antigénique
• Épitopes de surface répétitifs de Plasmodium réponse non efficace.
• Variation antigénique et génétique des souches (protéine d'adhérence de surface PfEMP1 de P. Falciparum codée par les gènes var dont il existe environ 50 copies par génome) pas d'immunité protectrice croisée entre variants

Commutation antigénique.
• Trypanosomes africains qui expriment à leur surface un seul type de glycoprotéine variante (Variable Surface coat Gp : VSG). Il existe plus de 1000 gènes et pseudogènes codant pour cette glycoprotéine.
• Succession de gènes différents

Commutation antigénique
http://diverge.hunter.cuny.edu/~weigang/Images/22-15_trypanosomes_1.jpg

Variations antigéniques des Trypanosomes
http://www.gla.ac.uk/centres/wcmp/research/barry/research.html

Variations antigéniques des Trypanosomes
http://home.inje.ac.kr/~lecture/immunobiotech/ch2/E$$00010.JPG

Commutation chez Schistosoma
• Schistosomules expriment une gp majeure de surface (gp 38) réponse Ac
• Les adultes n'expriment plus cette gp 38 à la surface mais libèrent une glycoprotéine gp 115 responsable d'une immunité croisée vis à vis de gp 38, immunité qui protège l'hôte d'une surinfection par de nouveaux schistosomules.

Modification des réponses immunitaires
• Modification de la présentation de l'antigène
• Modification des fonctions des phagocytes mononucléés
• Modification de la réponse T

Modification de la présentation de l'antigène
• Capacités de phagocytose et de présentation de l'antigène sont diminuées (plasmodium, trypanoafricaine).
• Leishmaniose viscérale il existe un blocage de l'expression des antigènes de classe II
• Diminution de l'expression de co-molécules d'activation ou de la production de cytokines comme l'IL-1 �présentation incomplète.

Modification des fonctions des phagocytes mononucléés
• Blocage des fonctions des monocytes macrophages (GR avec plasmodies ou leishmanies intra-C)
• "Désactivation" des phagocytes mononucléés par la production de molécules inhibitrices comme la prostaglandine E2 ou le TGFß,

Modification de la réponse T
• Inhibition des réponses prolifératives des lymphocytes (T. cruzi ) par défaut d'expression des molécules α et ß du IL-2R, induit par un facteur soluble parasitaire immunosuppresseur.
• T. brucei induit un blocage du cycle cellulaire des lymphocytes T
• Les schistosomes adultes produisent aussi un facteur inhibiteur de faible poids moléculaire qui inhibe la prolifération lymphocytaire
• L. major et L. donovani produisent une protéine majeure de surface capable de cliver la molécule CD4 des lymphocytes humains

Induction de réponse lymphocytaire spécifique à activité suppressive
• Blocage de l'interféron γ par TGFß ou l'IL-10 (T. cruzi, Leishmania chagasi, Schistosoma mansoni )

Variations de réponse selon l’hôte
• Exemple de l’infection par Leishmania

La réponse du système immunitaire de l'hôte
• Rôle limiter la prolifération du parasite voire l’éliminer
• Souches souris résistantes et sensibles• Parallélisme imparfait entre souris et
homme quant à la sensibilité ou la résistance.
• Nature variable de la réponse.

Chez l’homme
• F. localisées– Réponse humorale (Ac anti-lsh)– Réponse cellulaire (HSR leishmanine)
• F. diffuses– Réponse humorale (Ac anti-lsh) et activation
polyclonale– Déficit de la réponse cellulaire spécifique :
• HSR négative à la leishmanine• Lymphopénie inconstante• Inhibition des réponses prolifératives

Chez la souris
• Souche sensible (Balb/c) :– Inoculum (104-107) dans le coussinet plantaire
– Essaimage dans les ganglions périphériques– Puis les organes lymphoïdes en 3 semaines
– Mort en 10 à 12 semaines selon l’inoculum.
• Souches résistante (C57BL/6) :– Oedème puis régression

Génes de suceptibilité chez la souris
•Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002;2 : 845-58.

Relation susceptibilité et
réponse Th1/Th2
http://www.wehi.edu.au/research/divisions/inf/labs/handman/genetic_basis.html

Anomalies immunitaires précoces
• Activation réduite des MΦ infectés par les promastigotes
• Capacité réduite à activer les mécanismes de cytotoxicité intracellulaire
• Diminution de production d’IFN-γ• Déséquilibre rapport Th1/Th2• Diminution de l’activité des cellules NK• Production de TGFβ

Anomalies précoces
• Absence de transcription de gènes d’IL-12, MIP-1α, TNFα, IL-10 et du gène de la NO synthase inductible dans les macrophages du derme de souris Balb/c.
• Production retardée d’IL-12 (7-12 jours)• Défaut de présentation par le CMH de
classe II (souris CMH cl.II -/- infection fatale) rôle des lymphocytes T CD4+.

Activation des macrophages
• Indispensable pour le contrôle de l'infection intracellulaire (souris SCID ou souris athymiques nude)
• Rôle majeur de l’interféron-γ (IFN-γ). La délétion du gène de l'IFN-γ ou du récepteur d'IFN-γ chez les souches résistantes de souris les rend sensibles à l'infection par L. Major

Mécanismes de l’IFN-γ• Production d'oxyde nitrique (NO) par la NO
synthase inductible (iNOS)• Blocage de la production de NO in vitro ou in
vivo rend les cultures ou les animaux sensibles à l'infection par L. major
• Anomalie de production de NO chez les animaux KO IFN-γ -/- ou KO récepteur d'IFN-γ -/-et chez le hamster infecté par L. donovani
• L'IFN-γ est la seule cytokine capable de stimuler la transcription de iNOS et la libération de NO

Action synergique d’autres cytokines
• Le TNFα. Les anticorps anti-TNF neutralisants peuvent rendre sensibles à L. major les souris résistantes CBA ou C3H.
• l'IL-2, l'IL-7 et même l'IL-4 • Autres cytokines aussi régulatrices de la
réponse anti-parasitaire et peuvent la bloquer : l'IL-3, L'IL-5 et le GM-CSF

Développement des lymphocytes T CD4 effecteurs
• Description des lymphocytes CD4+ Th1 et Th2 par Mossman et Coffman en 1986
• Leishmanioses murines bon modèle de cette théorie
• BALB/c expriment des transcrits d'IL-4 alors que les souches résistantes expriment des transcrits d'IFN-γ
• Transfert de lignées cellulaires Th1 résistance à des souris Balb/c.

Rôle des ly T, de l’IL-4 et IL-12
• Immuno-intervention (irradiation ou cyclophosphamide, d'anti-CD4, d'anti-IL-4, d'anti-IL12 ou de blocage de l'interaction CD28-B7) très précoce après l'inoculation du parasite.
• Sécrétion initiale d’IL-4 : chez les souris résistantes la production d'IL-4 décroît rapidement alors qu'elle ne décroît que peu chez les souris sensibles BALB/c.
• L’IL-4 inhibe la production d’IFN-γ

Rôle des cellules NK et CD8+
• Incertain– l'apport de lymphocytes CD4 seuls permet la
protection de souris SCID;– les souris beige (NK-/-) sont résistante à l'infection
par l. Major alors que la production initiale d’IFN-γ est fonction de la présence de cellules NK;
– les souris ß2 microglobuline -/- de souches résistantes gardent leur résistance.
– Rôle possible pour le contrôle initial de l’infection de la peau.

Rôle de l'IL-4
• BALB/c, cinétique de production d'IL-4 bimodale après injection dans le coussinet plantaire de Leishmanies : – une première production d'IL-4 maximum à 16 heures
puis à J5. – Après injection voie intraveineuse de Leishmanies
chez la souris BALB/c on retrouve une production précoce d'IL-4 par les splénocytes murins à 90 minutes.
– un anti-IL-4 injecté précocement permet de rendre résistantes des souris sensibles

Rôle de l'IL-12
• Le blocage de l'IL-12 chez des souris résistantes les rend sensibles à l'infection
• L'IL-12 injectée au tout début après l'inoculation peut rendre résistante des souris BALB/c.
• L'action non par l'induction d'IFN-γ mais par la capacité de l'IL-12 à bloquer la production d'IL-4.

Modèle du développement Th1/Th2
•Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002;2 : 845-58.

Rôle variable des cytokines TH2
The relative disease exacerbatory activity of Th2 and regulatory cytokines in BALB/c mice
+++?+TGF-β
++++++IL-10
−*+*+IL-13
−+++++IL-4
L. donovaniL. mexicanaL. major
(+++) major mediator; (++) significant activity; (+) moderate activity; (−) no activity.
Alexander J, Bryson K. T helper (h)1/Th2 and Leishmania: paradox rather than paradigm.Immunol Lett. 2005 15;99:17-23.

Nature des cellules produisant précocement de l’IL-4
• Cellules T Vα8/Vβ4• Déplétion en T Vα8/Vβ4 avec un virus
MMT SIM (spécifique de Vβ4) absence de production précoce d’IL-4 d’où résistance. (Launois Immunity 1997 6: 541-9).
• Pas d’effet avec un virus MMT contrôle spécifique de Vβ6.

Spécificité des cellules Vα8/Vß4
• Les cellules Vα8/Vß4 reconnaissent une protéine LACK de 36 Kd (Leishmania homologof receptor for activated C kinase) et produisent de l'IL-4
• La stimulation de souris résistantes C57BL/6 par l'antigène LACK ne produit pas de sécrétion d'IL-4.
• Des souris tolérantes à l’antigène LACK par insertion du gène dans le génome murin ne produisent plus d’IL-4 précocement et sont résistantes à l’infection à L. major.

Modulation des cellules Vα8/Vß4
• L’immunisation par la protéine LACK confère une protection si l’immunisation se fait en présence d’IL-12
• l’immunisation par de l’ADN codant pour la protéine LACK induit une protection identique à celle induite par la protéine avec l’IL-12
• Rôle de la cellule présentatrice d’antigène.

Évènement précoce : production d’IL-4
•Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002;2 : 845-58.

Évènement tardif chez Balb/c
•Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002;2 : 845-58.

Évènement tardif s. résistante
•Sacks D, Noben-Trauth N. The immunology of susceptibility and resistance to Leishmania major in mice. Nat Rev Immunol. 2002;2 : 845-58.

Modulation de la réponse Th1/Th2
• Rôle de l’APC (cellules dendritiques DC1 ou DC2)
• Rôle du TGF-β : L’injection de TGF-β peut exacerber l’infection parasitaires chez des souris de susceptibilité intermédiaire alors que le traitement par un anti-TGF-βpermet une guérison de ces mêmes souris.

Modulation de la réponse Th1/Th2
• Rôle des hormones stéroïdiennes : Au cours de la gestation il existe un risque accrue de leishmaniose grave même chez les souris sensibles (J. Immunol. 1996 ;156 :644-52)
• Rôle des co-molécules d'activation CD80 et CD86 : rôle variable selon les modèles – Le blocage de la molécule CD86 par un anticorps
anti-CD86 augmente la production d'IFN-γ et favorise la guérison dans un modèle souris BALB/c leishmania donovani (J. Immunol 1997; 159: 4460-6).

Modulation de la réponse Th1/Th2
• Rôle de la quantité d'antigène : A très faible dose de parasite une souris sensible peut faire une réponse protectrice et devenir résistante (Eur. J. Immunol. 1998 ; 28 : 4020-8)
• Rôle de modulateurs de la réponse Th1/Th2 : ex Imiquimod (J Inf Dis 1999 ; 179 : 1485-94)

Rôle de cellules T régulatrices
• C57BL/6 inoculation 103 promastigotes guérison spontanée en 8-10 semaines.
• Persistance 102-104 parasites vivants• Rôle local de cellules T CD4+ CD25+ à
fonction régulatrice.• L’injection de ces cellules rend susceptible
des C57BL/6.• → Survie du parasite

Réponse équilibrée ?
Alexander J, Bryson K. T helper (h)1/Th2 and Leishmania: paradox rather than paradigm. Immunol Lett. 2005 15;99:17-23.

• Le modèle d’infection par Leishmania a permis de mieux comprendre l’équilibre Th1/Th2 (limites)
• Il existe un équilibre entre l'hôte et le parasite qui permet dans la plupart des cas aux 2 partenaires de survivre sauf dans certaines situations (immunodépression; gestation etc...)

http://www.biology.leeds.ac.uk/staff/awtr/
Cinétique de la RI à Plasmodium chabaudichez la souris

Équilibre hôte/parasite
• Équilibre entre survie de l’hôte et du parasite
• Équilibre entre effet protecteur et effet délétère de la réponse immune.
• Formes cliniques différentes en fonction de la souche infectieuse et de la réponse de l’hôte.