
Fc récepteur et polynucléaire neutrophile
8
LE POLYNUCLÉAIRE DANS TOUS SES ÉTATS ! REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462 // 39 article reçu le 20 décembre 2013, accepté le 21 janvier 2014. © 2014 – Elsevier Masson SAS – Tous droits réservés. RÉSUMÉ Les récepteurs Fc (FcRs) sont des molécules qui reconnaissent spécifique- ment les fragments Fc des différentes classes d’immunoglobulines. Quatre classes de FcRs (pour les IgG, IgA, IgE et IgM) ont été décrites à ce jour. Ils sont exprimés sur les leucocytes, essentiellement sur les phagocytes et sur les lymphocytes B. Les polynucléaires neutrophiles au repos expriment fortement deux types de FcγRs, le FcγRIIA et le FcγRIIIB, ainsi que le FcαRI (récepteur aux IgA). Par contre, ils n’expriment pas le FcεRI ou le FcμRI. Les fonctions de ces récepteurs sur le neutrophile sont multiples et essentielles dans la défense de l’hôte contre les microorganismes. Les FcRs peuvent activer ou inhiber les réponses immunitaires. Les FcRs activateurs jouent un rôle clé dans la phagocytose et la sécrétion de facteurs solubles comme des cytokines ou des substances vasoactives. L’activation des FcRs induit des voies de signalisation qui modulent la mise en route des réponses immunitaires, par exemple la production de cytokines. Certains FcRs comme le FcαRI, peuvent avoir une double fonctionnalité en utilisant leur unique motif de transduction (à ITAM) soit pour inhiber soit pour activer les réponses immunitaires, selon la structure du ligand reconnu (par exemple des IgA monomériques ou poly- mériques). Finalement, des polymorphismes dans les gènes codant pour les FcRs peuvent être impliqués dans l’initiation et/ou aggravation de certaines maladies infectieuses ou autoimmunes. Sur le polynucléaire neutrophile, ces polymorphismes peuvent générer un changement de liaison vis-à-vis de certains ligands et une réponse immunitaire altérée. Anticorps – immunoglobuline – régulation de la réponse immunitaire – phagocytose – neutrophile – récepteur Fc. Renato C. Monteiro a,b,c,d, *, Sanae Ben Mkaddem b,c,d , Margarita Hurtado-Nédelec a,b,c,d Fc récepteur et polynucléaire neutrophile til l 20 dé b 2013 té l 21 j i 2014 a UF « Dysfonctionnements immunitaires » Département d’immuno-hématologie Hôpital Bichat-Claude Bernard (AP-HP) – Université Paris 7 16, rue Henri-Huchard 75877 Paris cedex 18 b Inserm U1149 – Centre de recherche sur l’inflammation c Université Paris Diderot – Université Sorbonne Paris Cité Laboratoire d’excellence Inflamex – Paris d ERL CNRS 8252 – Paris * Correspondance [email protected] SUMMARY Fc receptor and neutrophils Fc receptors ( FcRs ) are molecules with specificity for the Fc fragments of different antibody types. There are currently four classes of FcRs for IgG, IgA, IgM and IgE expressed on various leukocytes, mainly on pha- gocytes and B cells. The neutrophil strongly express two forms of FcγRs, the FcγRIIA and FcγRIIIB, as well as the FcαRI but not FcμRI or FcεRs. The functions of these receptors on neutrophils are multiple and essen- tial for host defense against microorganisms. FcRs can activate or inhibit immune responses. Activating neutrophil FcRs play a key role in phagocytosis and the secretion of soluble factors such as cytokines or vasoactive substances. Following the activation of FcRs, this mechanism will play various roles such as, for example, modulation of the immune response into play with cytokines. Some FcRs as FcαRI behaves like Janus using their unique transduction motif (ITAM) to either activate or inhibit responses depending on the type of engagement by the ligand, either monome- ric or polymeric IgA for example. Finally, the various FcRs neutrophils can be involved in the initiation and/ or aggravation of infectious or autoimmune diseases depending on the type of allelic variants expressed which usually generates changes in ligand binding leading to altered host defense. Antibody – immunoglobulin – immuneregulation – Fc receptor – phagocytosis – neutrophil. Il existe quatre classes de récepteurs bien définis, les FcγR pour les IgG, les FcεR pour les IgE, les FcαR pour les IgA et les FcμR pour les IgM, le FcδR restant le seul FcR non encore identifié au niveau moléculaire. La lettre grecque désigne la spécificité pour l’isotype de chaîne lourde, caractéristique de la classe des immunoglobulines (Igs), et le chiffre romain la classe de récepteur [1]. Trois sous-classes de récepteurs ont été identifiées pour les IgG chez l’homme et dénommées FcγRI, FcγRII et FcγRIII [2]. Des récepteurs pour les autres classes d’anticorps ont été identifiés depuis les années 90, notamment le récepteur pour les IgA 1 et 2 sur les monocytes et neutrophiles, appelé FcaRI [3-5], et le récepteur pour les IgM sur les lymphocytes B, le FcμRI [6, 7]. Récemment, un nouveau type de FcR a été identifié, le tripartite motif-containing protein (TRIM) 21. Ce FcR possède des caractéristiques particulières, à savoir : une expression intracellulaire, la capacité de reconnaître plusieurs classes d’Igs et un rôle particulier dans l’immunité anti-infectieuse [8]. Dans la numérota- tion des classes de différenciation (CD), les FcγRI, II et III 1. Introduction Les récepteurs pour les fragments Fc des immunoglobulines (FcRs) sont des glycoprotéines membranaires de la super- famille des immunoglobulines, dont la portion extracellulaire comprend deux à trois domaines immunoglobuliniques. Ces récepteurs partagent une grande homologie structu- relle, en lien avec leur fonction commune de liaison aux différentes classes d’anticorps (IgG, IgE, IgA, IgM et IgD).
Transcript of Fc récepteur et polynucléaire neutrophile

LE POLYNUCLÉAIRE DANS TOUS SES ÉTATS !
REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462 // 39
article reçu le 20 décembre 2013, accepté le 21 janvier 2014.
© 2014 – Elsevier Masson SAS – Tous droits réservés.
RÉSUMÉ
Les récepteurs Fc (FcRs) sont des molécules qui reconnaissent spécifique-ment les fragments Fc des différentes classes d’immunoglobulines. Quatre classes de FcRs (pour les IgG, IgA, IgE et IgM) ont été décrites à ce jour. Ils sont exprimés sur les leucocytes, essentiellement sur les phagocytes et sur les lymphocytes B. Les polynucléaires neutrophiles au repos expriment fortement deux types de FcγRs, le FcγRIIA et le FcγRIIIB, ainsi que le FcαRI (récepteur aux IgA). Par contre, ils n’expriment pas le FcεRI ou le FcμRI. Les fonctions de ces récepteurs sur le neutrophile sont multiples et essentielles dans la défense de l’hôte contre les microorganismes. Les FcRs peuvent activer ou inhiber les réponses immunitaires. Les FcRs activateurs jouent un rôle clé dans la phagocytose et la sécrétion de facteurs solubles comme des cytokines ou des substances vasoactives. L’activation des FcRs induit des voies de signalisation qui modulent la mise en route des réponses immunitaires, par exemple la production de cytokines. Certains FcRs comme le FcαRI, peuvent avoir une double fonctionnalité en utilisant leur unique motif de transduction (à ITAM) soit pour inhiber soit pour activer les réponses immunitaires, selon la structure du ligand reconnu (par exemple des IgA monomériques ou poly-mériques). Finalement, des polymorphismes dans les gènes codant pour les FcRs peuvent être impliqués dans l’initiation et/ou aggravation de certaines maladies infectieuses ou autoimmunes. Sur le polynucléaire neutrophile, ces polymorphismes peuvent générer un changement de liaison vis-à-vis de certains ligands et une réponse immunitaire altérée.
Anticorps – immunoglobuline – régulation de la réponse immunitaire – phagocytose – neutrophile – récepteur Fc.
Renato C. Monteiroa,b,c,d,*, Sanae Ben Mkaddemb,c,d, Margarita Hurtado-Nédeleca,b,c,d
Fc récepteur et polynucléaire neutrophile
ti l l 20 dé b 2013 té l 21 j i 2014
a UF « Dysfonctionnements immunitaires »Département d’immuno-hématologieHôpital Bichat-Claude Bernard (AP-HP) – Université Paris 716, rue Henri-Huchard75877 Paris cedex 18 b Inserm U1149 – Centre de recherche sur l’inflammation c Université Paris Diderot – Université Sorbonne Paris CitéLaboratoire d’excellence Inflamex – Paris d ERL CNRS 8252 – Paris
SUMMARYFc receptor and neutrophils
Fc receptors ( FcRs ) are molecules with specificity for the Fc fragments of different antibody types. There are currently four classes of FcRs for IgG, IgA, IgM and IgE expressed on various leukocytes, mainly on pha-gocytes and B cells. The neutrophil strongly express two forms of FcγRs, the FcγRIIA and FcγRIIIB, as well as the FcαRI but not FcμRI or FcεRs. The functions of these receptors on neutrophils are multiple and essen-tial for host defense against microorganisms. FcRs can activate or inhibit immune responses. Activating neutrophil FcRs play a key role in phagocytosis and the secretion of soluble factors such as cytokines or vasoactive substances. Following the activation of FcRs, this mechanism will play various roles such as, for example, modulation of the immune response into play with cytokines. Some FcRs as FcαRI behaves like Janus using their unique transduction motif (ITAM) to either activate or inhibit responses depending on the type of engagement by the ligand, either monome-ric or polymeric IgA for example. Finally, the various FcRs neutrophils can be involved in the initiation and/or aggravation of infectious or autoimmune diseases depending on the type of allelic variants expressed which usually generates changes in ligand binding leading to altered host defense.
Antibody – immunoglobulin – immuneregulation – Fc receptor – phagocytosis – neutrophil.
Il existe quatre classes de récepteurs bien définis, les FcγR pour les IgG, les FcεR pour les IgE, les FcαR pour les IgA et les FcμR pour les IgM, le FcδR restant le seul FcR non encore identifié au niveau moléculaire. La lettre grecque désigne la spécificité pour l’isotype de chaîne lourde, caractéristique de la classe des immunoglobulines (Igs), et le chiffre romain la classe de récepteur [1]. Trois sous-classes de récepteurs ont été identifiées pour les IgG chez l’homme et dénommées FcγRI, FcγRII et FcγRIII [2]. Des récepteurs pour les autres classes d’anticorps ont été identifiés depuis les années 90, notamment le récepteur pour les IgA 1 et 2 sur les monocytes et neutrophiles, appelé FcaRI [3-5], et le récepteur pour les IgM sur les lymphocytes B, le FcμRI [6, 7]. Récemment, un nouveau type de FcR a été identifié, le tripartite motif-containing protein (TRIM) 21. Ce FcR possède des caractéristiques particulières, à savoir : une expression intracellulaire, la capacité de reconnaître plusieurs classes d’Igs et un rôle particulier dans l’immunité anti-infectieuse [8]. Dans la numérota-tion des classes de différenciation (CD), les FcγRI, II et III
1. Introduction
Les récepteurs pour les fragments Fc des immunoglobulines (FcRs) sont des glycoprotéines membranaires de la super-famille des immunoglobulines, dont la portion extracellulaire comprend deux à trois domaines immunoglobuliniques. Ces récepteurs partagent une grande homologie structu-relle, en lien avec leur fonction commune de liaison aux différentes classes d’anticorps (IgG, IgE, IgA, IgM et IgD).

40 // REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462
Les gènes codant pour les récepteurs, FCGR2/3, sont, quant à eux, localisés au sein d’un cluster en position 1q23. De nombreux polymorphismes affectent ces gènes FCGR2/3, le plus connu d’entre eux étant le polymorphisme V158F du gène FCGR3A. Une substitution du nucléotide à la position 559 de l’ADNc de FCGR3A conduit au codage d’une valine (V) ou d’une phénylalanine (F) au niveau du domaine proximal (acide aminé 158) impliqué dans le site de liaison des IgG. Il n’est, dès lors, pas surprenant que ce polymorphisme influence la liaison de ces dernières. Le gène FCGR3A présente également un polymorphisme tri-allélique en position 48 (domaine distal), entraînant la présence d’une leucine (L), d’une arginine (R) ou d’une histidine (H). Un polymorphisme de FCGR2A se traduit par la présence en position 131 du domaine immuno-globulinique distal d’une R (alloforme 131R) ou d’une H (alloforme 131H). Cette substitution, connue depuis 1991, affecte fortement l’affinité du récepteur FcγRIIA pour les IgG1 de souris. Les deux variants alléliques FCGR2A-131H (appelé LR pour low-responder) et FCGR2A-131R (appelé HR pour high-responder) lient les IgG2a et IgG2b de souris, mais seul l’allèle FCGR2A-131R lie fortement les IgG1 de souris.
2. Expression cellulaire des FcRs
et leur capacité de liaison aux Igs
Les FcRs sont exprimés sur la majorité des cellules myé-loïdes et sur certaines cellules lymphoïdes. Chaque récep-teur présente un profil d’expression particulier, tandis que chaque type cellulaire exprime le plus souvent plusieurs classes de FcRs. Le polynucléaire neutrophile exprime le FcγRIIA, le FcγRIIIB et le FcαRI, mais pas le FcγRI, le FcεRI ou le FcμR en conditions d’homéostasie, c’est-à-dire au repos (figure 1). Le FcγRI (CD64) peut-être induit sur le polynucléaire neutrophile au cours d’une réaction inflam-matoire notamment par l’IFN-γ probablement pour assurer une bonne phagocytose des microorganismes. En effet, une hyperexpression de ce récepteur par le neutrophile est associée aux infections systémiques sévères [11, 12]. Les FcRs sont donc engagés ensemble par des complexes immuns, les amenant à coopérer positivement, comme par exemple FcγRIIA et FcγRIIIB sur les neutrophiles,
Abréviations
ADCC : antibody-dependent cytotoxicity
ANCA : anti-neutrophil cytoplasmic antibody
AP-1 : adaptor protein 1
CD : cluster différentiation
CMH : complexe majeur d’histocomptabilité
ERK : extracellular-regulated kinase
FcR : récepteur Fc
HR : high responder
Ig : immunoglobuline
ITAM : immunoreceptor tyrosine-based activation motif
ITAMi : inhibitory ITAM
ITIM : immunoreceptor tyrosine-based inhibition motif
IRF : interferon-regulatory factor
LR : low responderNF : nuclear factorNK : natural killerMPO : myélopéroxidasePPR : pattern recognition receptorRas : rat sarcoma protein, membre de la famille des petites protéines GTPases.SHP-1 : Src homology 2-containing tyrosine phosphataseSHIP : Src homology 2-containing inositol phosphataseSNP : single nucleotide polymorphismSyk : spleen tyrosine kinaseTLR : toll-like receptorsTRAIL : tumor necrosis factor-related apoptosis-inducing ligandTRIM : tripartite motif-containing protein
correspondent respectivement aux CD64, CD32 et CD16 et le FcαRI au CD89.Le clonage moléculaire des FcRs a permis d’identifier diffé-rentes formes de récepteurs. Les FcγRII (FcγRIIA, FcγRIIB et FcαRIIC) et FcγRIII (FcγRIIIA et FcγRIIIB), se distinguent par leur ancrage membranaire et/ou leur motif de signa-lisation [9]. Les récepteurs activateurs FcγRI, FcγRIIIA et FcaRI comportent une région transmembranaire possédant un résidu chargé et une courte région intracytoplasmique. Le résidu chargé permet au récepteur de s’associer à des dimères d’une chaîne transductrice, la chaine FcR-γ. Cette chaîne possède une très courte région extracellulaire, une région transmembranaire et une région intracytoplasmique comportant un motif de transduction ITAM (immunoreceptor tyrosine-based activation motif) impliqué dans l’activation cellulaire, qui se produit secondairement à l’agrégation des FcRs par des complexes immuns formés par des IgG ou des IgA associées à leurs antigènes spécifiques. Les récepteurs FcγRIIA et FcγRIIC sont également des récep-teurs activateurs se distinguant des précédents par le fait qu’ils possèdent leur propre motif de transduction ITAM dans leur partie intracytoplasmique et qu’ils n’ont donc pas besoin de chaînes accessoires pour entraîner l’acti-vation cellulaire. De la même façon, le récepteur FcγRIIB contient son propre motif de transduction de type ITIM (immunoreceptor tyrosine-based inhibition motif), ce qui fait de lui l’unique FcγR inhibiteur [10]. Enfin, le récepteur FcγRIIIB est un récepteur atypique du fait qu’il s’agit d’une protéine glypiée, c’est-à-dire reliée au feuillet externe de la membrane par une liaison glycosylphosphatidylinositol (GPI) qui la localise principalement dans les radeaux lipidiques. Sa structure ne lui confère aucune capacité intrinsèque de transduction du signal, mais il peut coopérer avec d’autres récepteurs, comme le FcγRIIA, amenant à proximité de ces derniers des éléments de signalisation associés aux radeaux lipidiques.Les gènes codant pour les récepteurs FcγRII et FcγRIII sont localisés sur le chromosome 1. Par contre, le gène codant pour le FcαRI est localisé sur le chromosome 19. Les gènes FCGR1 (FCGR1A, FCGR1B et FCGR1C) sont localisés de part et d’autre du centromère (1p23 et 1q21), et seul FCGR1A semble coder pour le récepteur de forte affi-nité, les deux autres gènes présentant des codons-stop au niveau de l’exon codant le troisième domaine extracellulaire.
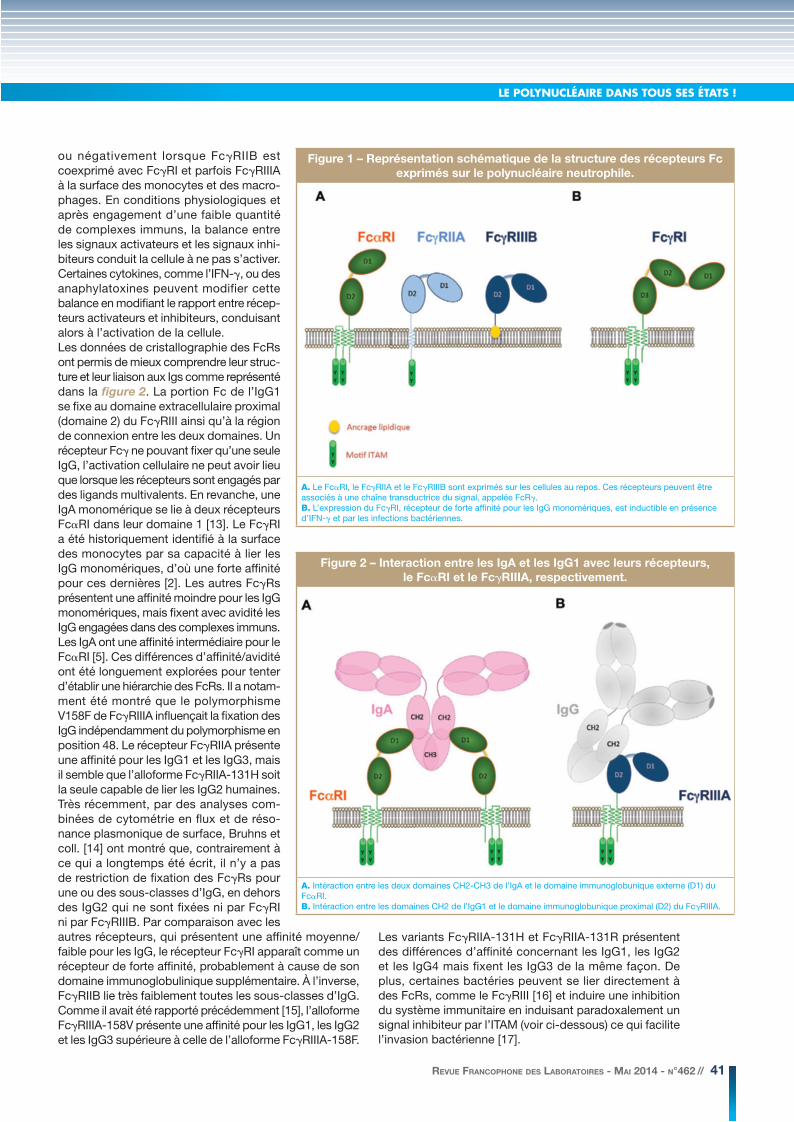
LE POLYNUCLÉAIRE DANS TOUS SES ÉTATS !
REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462 // 41
ou négativement lorsque FcγRIIB est coexprimé avec FcγRI et parfois FcγRIIIA à la surface des monocytes et des macro-phages. En conditions physiologiques et après engagement d’une faible quantité de complexes immuns, la balance entre les signaux activateurs et les signaux inhi-biteurs conduit la cellule à ne pas s’activer. Certaines cytokines, comme l’IFN-γ, ou des anaphylatoxines peuvent modifier cette balance en modifiant le rapport entre récep-teurs activateurs et inhibiteurs, conduisant alors à l’activation de la cellule.Les données de cristallographie des FcRs ont permis de mieux comprendre leur struc-ture et leur liaison aux Igs comme représenté dans la figure 2. La portion Fc de l’IgG1 se fixe au domaine extracellulaire proximal (domaine 2) du FcγRIII ainsi qu’à la région de connexion entre les deux domaines. Un récepteur Fcγ ne pouvant fixer qu’une seule IgG, l’activation cellulaire ne peut avoir lieu que lorsque les récepteurs sont engagés par des ligands multivalents. En revanche, une IgA monomérique se lie à deux récepteurs FcαRI dans leur domaine 1 [13]. Le FcγRI a été historiquement identifié à la surface des monocytes par sa capacité à lier les IgG monomériques, d’où une forte affinité pour ces dernières [2]. Les autres FcγRs présentent une affinité moindre pour les IgG monomériques, mais fixent avec avidité les IgG engagées dans des complexes immuns. Les IgA ont une affinité intermédiaire pour le FcαRI [5]. Ces différences d’affinité/avidité ont été longuement explorées pour tenter d’établir une hiérarchie des FcRs. Il a notam-ment été montré que le polymorphisme V158F de FcγRIIIA influençait la fixation des IgG indépendamment du polymorphisme en position 48. Le récepteur FcγRIIA présente une affinité pour les IgG1 et les IgG3, mais il semble que l’alloforme FcγRIIA-131H soit la seule capable de lier les IgG2 humaines. Très récemment, par des analyses com-binées de cytométrie en flux et de réso-nance plasmonique de surface, Bruhns et coll. [14] ont montré que, contrairement à ce qui a longtemps été écrit, il n’y a pas de restriction de fixation des FcγRs pour une ou des sous-classes d’IgG, en dehors des IgG2 qui ne sont fixées ni par FcγRI ni par FcγRIIIB. Par comparaison avec les autres récepteurs, qui présentent une affinité moyenne/faible pour les IgG, le récepteur FcγRI apparaît comme un récepteur de forte affinité, probablement à cause de son domaine immunoglobulinique supplémentaire. À l’inverse, FcγRIIB lie très faiblement toutes les sous-classes d’IgG. Comme il avait été rapporté précédemment [15], l’alloforme FcγRIIIA-158V présente une affinité pour les IgG1, les IgG2 et les IgG3 supérieure à celle de l’alloforme FcγRIIIA-158F.
Figure 1 – Représentation schématique de la structure des récepteurs Fc
exprimés sur le polynucléaire neutrophile.
A. Le FcαRI, le FcγRIIA et le FcγRIIIB sont exprimés sur les cellules au repos. Ces récepteurs peuvent être associés à une chaîne transductrice du signal, appelée FcRγ.B. L’expression du FcγRI, récepteur de forte affinité pour les IgG monomériques, est inductible en présence d’IFN-γ et par les infections bactériennes.
Les variants FcγRIIA-131H et FcγRIIA-131R présentent des différences d’affinité concernant les IgG1, les IgG2 et les IgG4 mais fixent les IgG3 de la même façon. De plus, certaines bactéries peuvent se lier directement à des FcRs, comme le FcγRIII [16] et induire une inhibition du système immunitaire en induisant paradoxalement un signal inhibiteur par l’ITAM (voir ci-dessous) ce qui facilite l’invasion bactérienne [17].
Figure 2 – Interaction entre les IgA et les IgG1 avec leurs récepteurs,
le FcαRI et le FcγRIIIA, respectivement.
A. Intéraction entre les deux domaines CH2-CH3 de l’IgA et le domaine immunoglobunique externe (D1) du FcαRI.B. Intéraction entre les domaines CH2 de l’IgG1 et le domaine immunoglobunique proximal (D2) du FcγRIIIA.
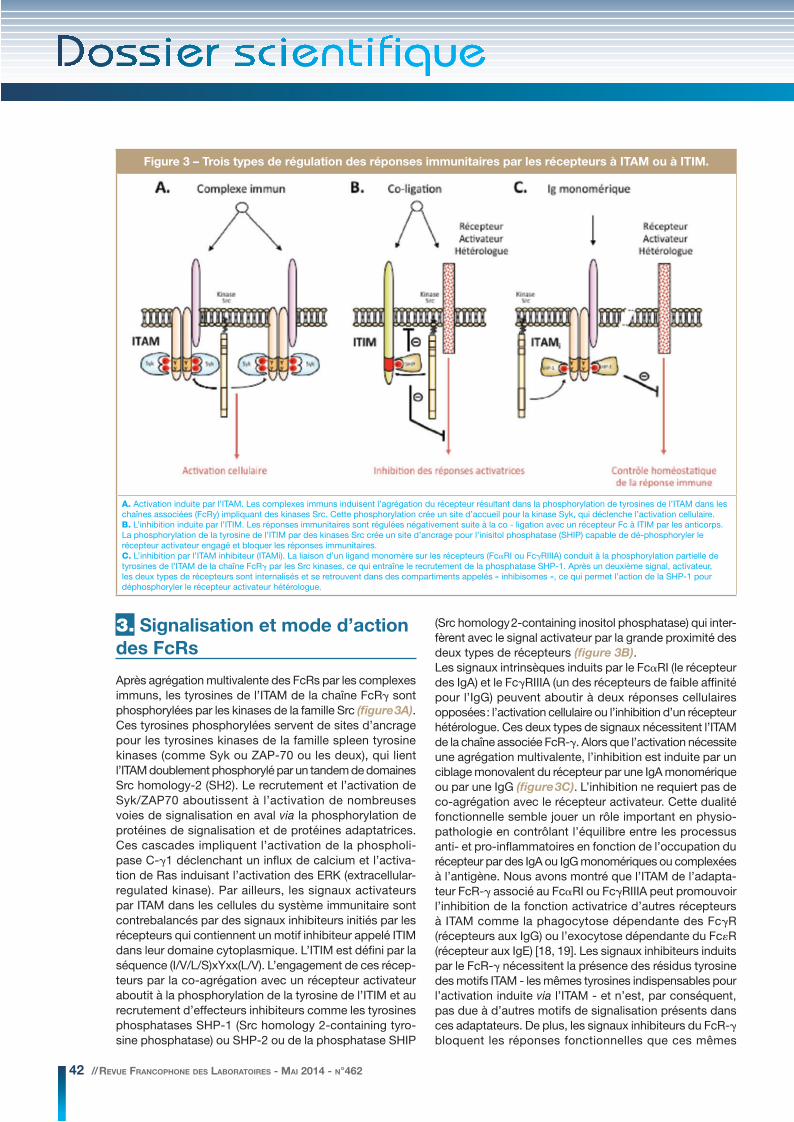
42 // REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462
3. Signalisation et mode d’action
des FcRs
Après agrégation multivalente des FcRs par les complexes immuns, les tyrosines de l’ITAM de la chaîne FcRγ sont phosphorylées par les kinases de la famille Src (figure 3A). Ces tyrosines phosphorylées servent de sites d’ancrage pour les tyrosines kinases de la famille spleen tyrosine kinases (comme Syk ou ZAP-70 ou les deux), qui lient l’ITAM doublement phosphorylé par un tandem de domaines Src homology-2 (SH2). Le recrutement et l’activation de Syk/ZAP70 aboutissent à l’activation de nombreuses voies de signalisation en aval via la phosphorylation de protéines de signalisation et de protéines adaptatrices. Ces cascades impliquent l’activation de la phospholi-pase C-γ1 déclenchant un influx de calcium et l’activa-tion de Ras induisant l’activation des ERK (extracellular-regulated kinase). Par ailleurs, les signaux activateurs par ITAM dans les cellules du système immunitaire sont contrebalancés par des signaux inhibiteurs initiés par les récepteurs qui contiennent un motif inhibiteur appelé ITIM dans leur domaine cytoplasmique. L’ITIM est défini par la séquence (I/V/L/S)xYxx(L/V). L’engagement de ces récep-teurs par la co-agrégation avec un récepteur activateur aboutit à la phosphorylation de la tyrosine de l’ITIM et au recrutement d’effecteurs inhibiteurs comme les tyrosines phosphatases SHP-1 (Src homology 2-containing tyro-sine phosphatase) ou SHP-2 ou de la phosphatase SHIP
(Src homology 2-containing inositol phosphatase) qui inter-fèrent avec le signal activateur par la grande proximité des deux types de récepteurs (figure 3B).Les signaux intrinsèques induits par le FcαRI (le récepteur des IgA) et le FcγRIIIA (un des récepteurs de faible affinité pour l’IgG) peuvent aboutir à deux réponses cellulaires opposées : l’activation cellulaire ou l’inhibition d’un récepteur hétérologue. Ces deux types de signaux nécessitent l’ITAM de la chaîne associée FcR-γ. Alors que l’activation nécessite une agrégation multivalente, l’inhibition est induite par un ciblage monovalent du récepteur par une IgA monomérique ou par une IgG (figure 3C). L’inhibition ne requiert pas de co-agrégation avec le récepteur activateur. Cette dualité fonctionnelle semble jouer un rôle important en physio-pathologie en contrôlant l’équilibre entre les processus anti- et pro-inflammatoires en fonction de l’occupation du récepteur par des IgA ou IgG monomériques ou complexées à l’antigène. Nous avons montré que l’ITAM de l’adapta-teur FcR-γ associé au FcαRI ou FcγRIIIA peut promouvoir l’inhibition de la fonction activatrice d’autres récepteurs à ITAM comme la phagocytose dépendante des FcγR (récepteurs aux IgG) ou l’exocytose dépendante du FcεR (récepteur aux IgE) [18, 19]. Les signaux inhibiteurs induits par le FcR-γ nécessitent la présence des résidus tyrosine des motifs ITAM - les mêmes tyrosines indispensables pour l’activation induite via l’ITAM - et n’est, par conséquent, pas due à d’autres motifs de signalisation présents dans ces adaptateurs. De plus, les signaux inhibiteurs du FcR-γ bloquent les réponses fonctionnelles que ces mêmes
Figure 3 – Trois types de régulation des réponses immunitaires par les récepteurs à ITAM ou à ITIM.
A. Activation induite par l’ITAM. Les complexes immuns induisent l’agrégation du récepteur résultant dans la phosphorylation de tyrosines de l’ITAM dans les chaînes associées (FcRy) impliquant des kinases Src. Cette phosphorylation crée un site d’accueil pour la kinase Syk, qui déclenche l’activation cellulaire.B. L’inhibition induite par l’ITIM. Les réponses immunitaires sont régulées négativement suite à la co - ligation avec un récepteur Fc à ITIM par les anticorps. La phosphorylation de la tyrosine de l’ITIM par des kinases Src crée un site d’ancrage pour l’inisitol phosphatase (SHIP) capable de dé-phosphoryler le récepteur activateur engagé et bloquer les réponses immunitaires.C. L’inhibition par l’ITAM inhibiteur (ITAMi). La liaison d’un ligand monomère sur les récepteurs (FcαRI ou FcγRIIIA) conduit à la phosphorylation partielle de tyrosines de l’ITAM de la chaîne FcRγ par les Src kinases, ce qui entraîne le recrutement de la phosphatase SHP-1. Après un deuxième signal, activateur, les deux types de récepteurs sont internalisés et se retrouvent dans des compartiments appelés « inhibisomes », ce qui permet l’action de la SHP-1 pour déphosphoryler le récepteur activateur hétérologue.

LE POLYNUCLÉAIRE DANS TOUS SES ÉTATS !
REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462 // 43
adaptateurs peuvent déclencher quand ils se trouvent au sein de récepteurs agrégés : dégranulation mastocytaire ou phagocytose IgG-dépendante et production de cytokines. Cette inhibition nécessite la tyrosine phosphatase SHP-1 dont l’effet aboutit in vitro à la désensibilisation du récep-teur cible et in vivo à la prévention du développement de l’asthme ainsi que la prévention du développement d’une glomérulonéphrite [19, 20].Ainsi, les récepteurs associés à des adaptateurs contenant un motif ITAM peuvent, après un engagement transitoire, fonctionner comme inhibiteurs de multiples réponses comme les réactions inflammatoires dues aux FcR ou les signaux induits par les pathogènes via les TLR (toll-like receptors). Au contraire de l’inhibition induite par les ITIM, l’inhibition induite par l’ITAM, dorénavant appelé ITAMi pour ITAM inhibiteur, ne nécessite pas de co-agrégation avec les récepteurs activateurs. Notre hypothèse est que la fonction de ces deux types d’ITAM, (ITAM ou ITAMi) est associée à de nouveaux mécanismes intrinsèques qui permettent une orientation vers les états activateurs ou inhibiteurs. L’inhibition par l’ITAMi d’un récepteur activateur hétéro-logue est effectuée par la phosphatase SHP-1 (et non par SHIP ou SHP-2) recrutée par l’ITAMi du FcR engagé qui se fait grâce à la formation de nouvelles structures appelées « inhibisomes » qui sont induites après endocytose des récepteurs regroupés au niveau des radeaux lipidiques membranaires [21] (figure 3C). L’inhibition par l’adapta-teur FcR-γ associé au FcαRI, par exemple, fournit une explication à un phénomène observé depuis longtemps dans lequel les IgA monomériques, majoritaires dans le sérum, peuvent inhiber l’activation IgG-dépendante des cellules myéloïdes, affectant ainsi un large panel de fonc-tions telles que la phagocytose, la réponse oxydative et la production de cytokines [5]. Conformément à nos données, nous proposons que l’absence de régulation négative par le FcαRI chez les patients déficients en IgA peut expliquer la majoration de l’incidence des maladies auto-immunes et des allergies observée chez ces patients [22].
4. Fonctions des FcRs
sur les phagocytes
Le déclenchement d’une réponse cellulaire via les FcRs va être conditionné, pour les récepteurs de faible affinité, par l’agrégation de ces récepteurs grâce à l’interaction avec des complexes immuns solubles ou avec des cel-lules opsonisées/sensibilisées par des Igs. L’agrégation de ces récepteurs conduira à leur migration au sein de radeaux lipidiques. Cette étape est indispensable pour la transduction des signaux. En fonction du type cellulaire et de son équipement en récepteur FcγRs, on distingue cinq grands types de fonctions cellulaires pouvant découler de ces voies d’activation.La première de ces fonctions est la phagocytose, exercée par les phagocytes « professionnels » que sont les macro-phages et les neutrophiles [23]. En présence d’agents pathogènes, de cellules ou de débris cellulaires opsoni-sés par des IgG, les phagocytes vont émettre des pseu-dopodes autour des complexes immuns pour former un phagosome. Celui-ci va fusionner avec des lysosomes ou
différentes granulations pour former un phagolysosome contenant diverses enzymes hydrolytiques, des protéines cationiques, des peptides anti-microbiens, des cytokines…. Ce processus s’accompagne aussi d’une explosion oxy-dative par activation de la NADPH oxydase, produisant l’anion superoxyde à l’origine des autres formes réactives de l’oxygène (voir l’article de M. Hurtado-Nedelec et al. dans ce numéro).La présentation antigénique constitue une deuxième voie de réponse enclenchée par les FcRs et permettra d’activer une réponse immunitaire adaptative. Ainsi, les cellules den-dritiques vont pouvoir endocyter les particules antigéniques via leurs FcγRs ou FcaRI, les dégrader et les présenter sous forme de peptides apprêtés dans des molécules du CMH [24]. En règle générale, ces peptides issus de protéines exogènes sont présentés par des molécules du CMH de classe II, induisant l’activation de lymphocytes T auxiliaires CD4+, mais la capture par les FcγRs semble également favori-ser la présentation de ces peptides exogènes via le CMH de classe I (cross-présentation), permettant l’initiation de réponses T CD8+ cytotoxiques [24, 25].Le troisième type de réponse enclenché par l’activation des FcγRs est la cytotoxicité à médiation cellulaire dépendante d’anticorps ou ADCC. Il s’agit de la plus connue dans le contexte d’une action antitumorale ; les cellules NK et les monocytes/macrophages en sont les principaux acteurs. Après la liaison à la cellule cible par les FcγRs, les granu-lations contenues dans le cytoplasme des cellules NK sont acheminées jusqu’à la membrane plasmique et déversées dans l’espace intercellulaire séparant l’effecteur et la cible. En se polymérisant à la surface des cibles, la perforine va créer des pores conduisant à une déstabilisation de la bicouche lipidique. La perméabilité ainsi obtenue va favoriser l’entrée de granzymes, autres constituants cytotoxiques des granulations NK. Parmi eux, le granzyme B va agir sur la cascade des caspases et entraîner l’apoptose des cibles ; d’autres granzymes vont coopérer et provoquer la mort des cellules cibles. En ce qui concerne les macrophages, cette cytotoxicité cellulaire va prendre la forme d’une « phagocy-tose extériorisée » : les granulations normalement déversées dans les phagosomes sont exocytées au niveau de l’espace intercellulaire, permettant ainsi la lyse de la cellule cible grâce à leur contenu particulièrement riche en enzymes protéo-lytiques, en formes réactives de l’oxygène et en NO (chez la souris). Il est vraisemblable qu’un tel phénomène existe aussi avec les polynucléaires neutrophiles [26].Le quatrième mécanisme activé par les FcγRs est la sécrétion de facteurs solubles comme des cytokines ou des subs-tances vasoactives. Les cytokines sécrétées jouent un rôle déterminant dans la régulation des réponses immunitaires. L’engagement des FcγRs joue également un rôle dans la cytotoxicité des cellules NK (natural killer) avec l’intervention de cytokines membranaires que sont les ligands de récep-teurs de mort, comme le ligand de FAS ou TRAIL (tumor necrosis factor-related apoptosis-inducing ligand) [27].Le cinquième et dernier mécanisme montre qu’au cours de l’infection par des agents pathogènes, les anticorps peuvent être effecteurs dans la cellule infectée, où ils sont détectés au niveau intracellulaire par le récepteur d’anti-corps exprimé de façon ubiquitaire, le TRIM21 cytosolique. La reconnaissance d’anticorps intracellulaires par TRIM21

44 // REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462
active une cascade de signalisation où TRIM21 catalyse la formation de chaînes d’ubiquitine et stimule les voies du facteur de transcription NF (nuclear factor) -kB, AP-1 (adaptor protein 1), IRF (interferon-regulatory factor) 3, IRF5 et IRF7. Cette activation aboutit également à la produc-tion de cytokines pro-inflammatoires, à la modulation des ligands naturels tueurs liés au stress et à l’induction d’un état antiviral. Ainsi, le système de détection intracellulaire des anticorps par le récepteur TRIM21 fournit une puissante activation du système immunitaire inné indépendamment des PPRs (pattern recognition receptors) [28].L’étude des souris dont les gènes des récepteurs Fc ont été invalidés permet une évaluation de l’implication de ces récepteurs dans le mécanisme d’action des anticorps thérapeutiques. Malgré la relative proximité phylogénétique entre les deux espèces, il existe de nombreuses différences entre les FcγRs humains et murins. La souris, contrairement à l’homme, ne possède que trois récepteurs activateurs, le FcγRI, le FcγRIII et le FcγRIV plus récemment décrit, et un seul récepteur inhibiteur (FcγRII), avec des profils d’expression différents des récepteurs FcγRs humains [10]. Bien qu’il existe un risque d’immunogénicité chez les souris immunocompétentes quand une IgG1 humaine est administrée, l’utilisation d’anticorps thérapeutiques chez la souris est possible et présente une certaine pertinence du fait que les IgG1 humaines présentent de l’affinité pour les FcγRs murins, alors que l’inverse n’est pas vrai. En revanche, l’antigène cible humain est rarement partagé avec la souris, ce qui oblige à développer, pour l’étude des effets thérapeutiques d’anticorps humanisé, des modèles de greffe de cellules tumorales humaines exprimant l’anti-gène cible humain (modèles xénogéniques reposant sur des souris immunodéficientes), ou des modèles où l’antigène cible humain est exprimé par les cellules murines (modèles syngéniques chez des souris immunocompétentes).
5. Polymorphisme du gène
du FcγRIIA et propension
à l’infection et l’autoimmunité
Trois isoformes de FcgRII ont été identifiés chez l’homme : le FcγRIIA, le FcγRIIB et le FcγRIIC. Le FcRγIIA est un récepteur activateur puissant de l’inflammation et est le plus répandu ; FcRγIIA est exprimé par toutes les cellules myéloïdes, fortement sur les neutrophiles mais pas sur les plaquettes. Les FcγRIIA et IIC possèdent un motif ITAM-like dans leur domaine intracytoplasmique. Contrairement aux FcRI et RIIIA, FcγRIIA n’est pas associé à la sous-unité chaine γ [29]. C’est un récepteur à faible affinité qui interagit avec les quatre sous-classes d’IgG humaines [14], ainsi que les sous-classes IgG1, IgG2a, IgG2b de souris [30]. Il n’existe pas d’homologie de FcgRIIA chez la souris. Après agrégation multivalente du FcγRIIA, les tyrosines de l’ITAM sont phosphorylées par les kinases de la famille Src. Ces tyrosines phosphorylées servent de sites d’ancrage aux tyrosines kinases de la famille Syk comme Syk (spleen tyrosine kinase), qui se lie par un tandem de domaines Src homology-2 (SH2) à l’ITAM doublement phosphorylé. Le recrutement et l’activation de Syk aboutissent à l’in-duction de nombreuses voies de signalisation en aval via
la phosphorylation de protéines effectrices de signalisation et de protéines adaptatrices. Ces cascades impliquent l’acti-vation de la phospholipase C-γ1 (PLC-γ1) déclenchant un influx calcique et l’activation de Ras induisant l’activation de la MAP kinase Erk. Plusieurs fonctions cellulaires sont dépen-dantes de l’intégrité du motif ITAM comme la phagocytose, la production des cytokines, la présentation antigénique, la cytotoxicité à médiation cellulaire dépendante des anticorps (ADCC), et la production des anions superoxydes. Parmi les trois classes de récepteurs aux immunoglobulines, seul FcγRII est capable de fixer les IgG2. Il existe 2 allotypes de FcγRIIA (FcγRIIA-R131 et FcγRIIA-H131) en fonction de la présence d’une arginine (R) ou d’une histidine (H) en position 131 dans le domaine extracellulaire du récepteur. Cette variabilité, qui ne concerne qu’un acide aminé, a des conséquences majeures sur la fonctionnalité du récepteur. In vitro, les neutrophiles possédant l’allotype IIA-H/H131 peuvent phagocyter normalement les bactéries opsonisées par l’IgG2 [31]. Au contraire les neutrophiles qui expriment à leur surface le récepteur IIA-R/R131, présent chez 25 % de la population caucasienne, se révèlent incapables de phagocyter ces bactéries. In vivo, il a été montré que les patients âgés de plus de 5 ans et porteurs de l’allotype IIA-R/R131 étaient plus susceptibles aux infections méningococ-ciques que les patients IIA-H/H131 [32]. Deux autres études ont clairement montré que le récepteur FcγRIIA R/R131 conférait un risque important de développer des pneu-monies mortelles à pneumocoques [33] et des méningo-coccies létales [34]. De plus, plusieurs études suggèrent que l’allotype IIA-R/R131 est associé à un risque élevé dans le développement d’une polyarthrite rhumatoïde [35]. Des études récentes montrent que la souris transgénique pour le récepteur FcγRIIA développe spontanément des maladies auto-immunes avec un spectre complexe de symptômes semblables à ceux trouvés dans la polyarthrite rhumatoïde humaine et le lupus érythémateux systémique (LES) (anticorps antinucléaires, glomérulonéphrite avec un dépôt de complexes immuns dans la membrane basale) [36]. De plus, il a été montré que le FcγRIIA soluble est capable d’éliminer complètement les complexes immuns et par conséquent prévient l’activation des cellules inflammatoires [37]. Ces résultats montrent clairement que le FcγRIIA humain joue un rôle majeur dans l’initiation de l’inflammation induite par des complexes immuns (figure 4A).
6. Activation des FcRs
du polynucléaire neutrophile
et ANCA
Les polynucléaires neutrophiles humains expriment prin-cipalement trois types de FcRs, le FcγRIIA, le FcγRIIIB et le FcαRI. Il est intéressant de noter que FcγRIIA, contient un motif ITAM dans sa région cytoplasmique. Il s’agit d’un récepteur d’activation unique qui est dépourvu d’homologie chez la souris. En outre, le FcγRIIIB exprimé par les neu-trophiles est lié à la membrane plasmique par des lipides (ancrage GPI), ne possède pas de domaine transmem-branaire et ne peut donc pas activer seul la signalisation intracellulaire. Enfin, le FcαRI représente le seul récepteur associé à la chaine FcR-γ sur les neutrophiles au repos.
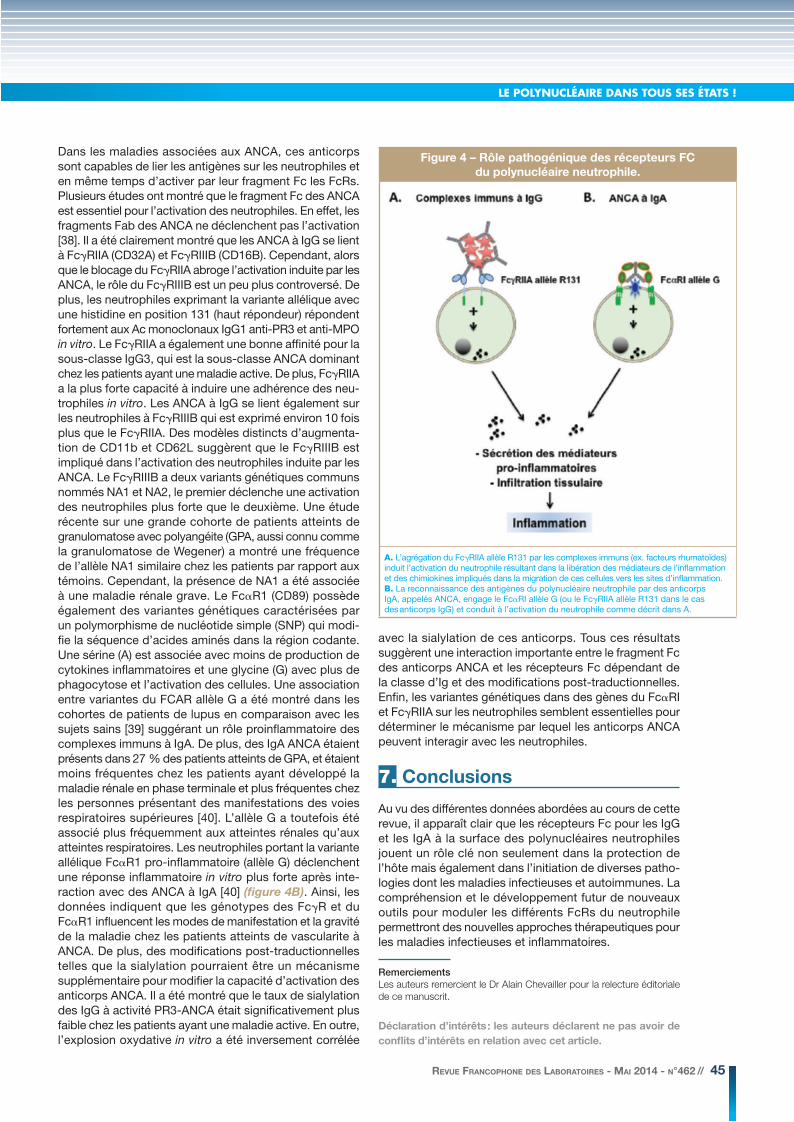
LE POLYNUCLÉAIRE DANS TOUS SES ÉTATS !
REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462 // 45
Dans les maladies associées aux ANCA, ces anticorps sont capables de lier les antigènes sur les neutrophiles et en même temps d’activer par leur fragment Fc les FcRs. Plusieurs études ont montré que le fragment Fc des ANCA est essentiel pour l’activation des neutrophiles. En effet, les fragments Fab des ANCA ne déclenchent pas l’activation [38]. Il a été clairement montré que les ANCA à IgG se lient à FcγRIIA (CD32A) et FcγRIIIB (CD16B). Cependant, alors que le blocage du FcγRIIA abroge l’activation induite par les ANCA, le rôle du FcγRIIIB est un peu plus controversé. De plus, les neutrophiles exprimant la variante allélique avec une histidine en position 131 (haut répondeur) répondent fortement aux Ac monoclonaux IgG1 anti-PR3 et anti-MPO in vitro. Le FcγRIIA a également une bonne affinité pour la sous-classe IgG3, qui est la sous-classe ANCA dominant chez les patients ayant une maladie active. De plus, FcγRIIA a la plus forte capacité à induire une adhérence des neu-trophiles in vitro. Les ANCA à IgG se lient également sur les neutrophiles à FcγRIIIB qui est exprimé environ 10 fois plus que le FcγRIIA. Des modèles distincts d’augmenta-tion de CD11b et CD62L suggèrent que le FcγRIIIB est impliqué dans l’activation des neutrophiles induite par les ANCA. Le FcγRIIIB a deux variants génétiques communs nommés NA1 et NA2, le premier déclenche une activation des neutrophiles plus forte que le deuxième. Une étude récente sur une grande cohorte de patients atteints de granulomatose avec polyangéite (GPA, aussi connu comme la granulomatose de Wegener) a montré une fréquence de l’allèle NA1 similaire chez les patients par rapport aux témoins. Cependant, la présence de NA1 a été associée à une maladie rénale grave. Le FcαR1 (CD89) possède également des variantes génétiques caractérisées par un polymorphisme de nucléotide simple (SNP) qui modi-fie la séquence d’acides aminés dans la région codante. Une sérine (A) est associée avec moins de production de cytokines inflammatoires et une glycine (G) avec plus de phagocytose et l’activation des cellules. Une association entre variantes du FCAR allèle G a été montré dans les cohortes de patients de lupus en comparaison avec les sujets sains [39] suggérant un rôle proinflammatoire des complexes immuns à IgA. De plus, des IgA ANCA étaient présents dans 27 % des patients atteints de GPA, et étaient moins fréquentes chez les patients ayant développé la maladie rénale en phase terminale et plus fréquentes chez les personnes présentant des manifestations des voies respiratoires supérieures [40]. L’allèle G a toutefois été associé plus fréquemment aux atteintes rénales qu’aux atteintes respiratoires. Les neutrophiles portant la variante allélique FcαR1 pro-inflammatoire (allèle G) déclenchent une réponse inflammatoire in vitro plus forte après inte-raction avec des ANCA à IgA [40] (figure 4B). Ainsi, les données indiquent que les génotypes des FcγR et du FcαR1 influencent les modes de manifestation et la gravité de la maladie chez les patients atteints de vascularite à ANCA. De plus, des modifications post-traductionnelles telles que la sialylation pourraient être un mécanisme supplémentaire pour modifier la capacité d’activation des anticorps ANCA. Il a été montré que le taux de sialylation des IgG à activité PR3-ANCA était significativement plus faible chez les patients ayant une maladie active. En outre, l’explosion oxydative in vitro a été inversement corrélée
avec la sialylation de ces anticorps. Tous ces résultats suggèrent une interaction importante entre le fragment Fc des anticorps ANCA et les récepteurs Fc dépendant de la classe d’Ig et des modifications post-traductionnelles. Enfin, les variantes génétiques dans des gènes du FcαRI et FcγRIIA sur les neutrophiles semblent essentielles pour déterminer le mécanisme par lequel les anticorps ANCA peuvent interagir avec les neutrophiles.
7. Conclusions
Au vu des différentes données abordées au cours de cette revue, il apparaît clair que les récepteurs Fc pour les IgG et les IgA à la surface des polynucléaires neutrophiles jouent un rôle clé non seulement dans la protection de l’hôte mais également dans l’initiation de diverses patho-logies dont les maladies infectieuses et autoimmunes. La compréhension et le développement futur de nouveaux outils pour moduler les différents FcRs du neutrophile permettront des nouvelles approches thérapeutiques pour les maladies infectieuses et inflammatoires.
RemerciementsLes auteurs remercient le Dr Alain Chevailler pour la relecture éditoriale de ce manuscrit.
Déclaration d’intérêts : les auteurs déclarent ne pas avoir de
conflits d’intérêts en relation avec cet article.
Figure 4 – Rôle pathogénique des récepteurs FC
du polynucléaire neutrophile.
A. L’agrégation du FcγRIIA allèle R131 par les complexes immuns (ex. facteurs rhumatoïdes) induit l’activation du neutrophile résultant dans la libération des médiateurs de l’inflammation et des chimiokines impliqués dans la migration de ces cellules vers les sites d’inflammation.B. La reconnaissance des antigènes du polynucléaire neutrophile par des anticorps IgA, appelés ANCA, engage le FcαRI allèle G (ou le FcγRIIA allèle R131 dans le cas des anticorps IgG) et conduit à l’activation du neutrophile comme décrit dans A.

46 // REVUE FRANCOPHONE DES LABORATOIRES - MAI 2014 - N°462
Références[1] Nomenclature of Fc receptors. IUIS/WHO Subcommittee on Nomenclature of Fc receptors. Bull World Health Organ 1994;72(5): 809-10.[2] Unkeless JC, Scigliano E, Freedman VH. Structure and function of human and murine receptors for IgG. Annu Rev Immunol 1988;6: 251-81.[3] Monteiro RC, Kubagawa H, Cooper MD. Cellular distribution, regu-lation, and biochemical nature of an Fc alpha receptor in humans. J Exp Med 1990;171(3):597-613.[4] Chevailler A, Monteiro RC, Kubagawa H, et al. Immunofluorescence analysis of IgA binding by human mononuclear cells in blood and lym-phoid tissue. J Immunol 1989;142(7):2244-9.[5] Monteiro RC, Van De Winkel JG. IgA Fc receptors. Annu Rev Immunol 2003;21:177-204.[6] Ohno T, Kubagawa H, Sanders SK, et al. Biochemical nature of an Fc mu receptor on human B-lineage cells. J Exp Med 1990;172(4):1165-75.[7] Kubagawa H, Oka S, Kubagawa Y, et al. Identity of the elusive IgM Fc receptor (FcmuR) in humans. J Exp Med 2009;206(12):2779-93.[8] McEwan WA, Tam JC, Watkinson RE, et al. Intracellular antibo-dy-bound pathogens stimulate immune signaling via the Fc receptor TRIM21. Nat Immunol 2013;14(4):327-36.[9] Fridman WH, Bonnerot C, Daeron M, et al. Structural bases of Fc gamma receptor functions. Immunol Rev 1992;125:49-76.[10] Nimmerjahn F, Ravetch JV. Fcgamma receptors as regulators of immune responses. Nat Rev Immunol 2008;8(1):34-47.[11] Hirsh M, Mahamid E, Bashenko Y, et al. Overexpression of the high-affinity Fcgamma receptor (CD64) is associated with leukocyte dysfunc-tion in sepsis. Shock 2001;16(2):102-8.[12] Qureshi SS, Lewis SM, Gant VA, et al. Increased distribution and expression of CD64 on blood polymorphonuclear cells from patients with the systemic inflammatory response syndrome (SIRS). Clin Exp Immunol 2001;125(2):258-65.[13] Woof JM, Burton DR. Human antibody-Fc receptor interactions illu-minated by crystal structures. Nat Rev Immunol 2004;4(2):89-99.[14] Bruhns P, Iannascoli B, England P, et al. Specificity and affinity of human Fcgamma receptors and their polymorphic variants for human IgG subclasses. Blood 2009;113(16):3716-25.[15] Koene HR, Kleijer M, Algra J, et al. Fc gammaRIIIa-158V/F poly-morphism influences the binding of IgG by natural killer cell Fc gamma-RIIIa, independently of the Fc gammaRIIIa-48L/R/H phenotype. Blood 1997;90(3):1109-14.[16] Salmon JE, Kapur S, Kimberly RP. Opsonin-independent ligation of Fc gamma receptors. The 3G8-bearing receptors on neutrophils mediate the phagocytosis of concanavalin A-treated erythrocytes and nonopsonized Escherichia coli. J Exp Med 1987;166(6):1798-813.[17] Pinheiro da Silva F, Aloulou M, Skurnik D, et al. CD16 promotes Escherichia coli sepsis through an FcR gamma inhibitory pathway that prevents phagocytosis and facilitates inflammation. Nat Med 2007;13(11):1368-74.[18] Aloulou M, Ben Mkaddem S, Biarnes-Pelicot M, et al. IgG1 and IVIg induce inhibitory ITAM signaling through FcgammaRIII controlling inflammatory responses. Blood 2012;119(13):3084-96.[19] Pasquier B, Launay P, Kanamaru Y, et al. Identification of FcalphaRI as an inhibitory receptor that controls inflammation : dual role of FcRgamma ITAM. Immunity 2005;22(1):31-42.[20] Kanamaru Y, Pfirsch S, Aloulou M, et al. Inhibitory ITAM signaling by Fc alpha RI-FcR gamma chain controls multiple activating responses and prevents renal inflammation. J Immunol 2008;180(4):2669-78.
[21] Pfirsch-Maisonnas S, Aloulou M, Xu T, et al. Inhibitory ITAM signa-ling traps activating receptors with the phosphatase SHP-1 to form polarized “inhibisome” clusters. Sci Signal 2011;4(169):ra24.[22] Jacob CM, Pastorino AC, Fahl K, et al. Autoimmunity in IgA defi-ciency : revisiting the role of IgA as a silent housekeeper. J Clin Immunol 2008;28 Suppl 1 :S56-61.[23] Indik ZK, Park JG, Hunter S, Schreiber AD. The molecular dis-section of Fc gamma receptor mediated phagocytosis. Blood 1995;86(12):4389-99.[24] Geissmann F, Launay P, Pasquier B, et al. A subset of human den-dritic cells expresses IgA Fc receptor (CD89), which mediates internali-zation and activation upon cross-linking by IgA complexes. J Immunol 2001;166(1):346-52.[25] Regnault A, Lankar D, Lacabanne V, et al. Fcgamma receptor-mediated induction of dendritic cell maturation and major histocom-patibility complex class I-restricted antigen presentation after immune complex internalization. J Exp Med 1999;189(2):371-80.[26] Valerius T, Repp R, de Wit TP, et al. Involvement of the high-affinity receptor for IgG (Fc gamma RI; CD64) in enhanced tumor cell cytotoxi-city of neutrophils during granulocyte colony-stimulating factor therapy. Blood 1993;82(3):931-9.[27] Oshimi Y, Oda S, Honda Y, et al. Involvement of Fas ligand and Fas-mediated pathway in the cytotoxicity of human natural killer cells. J Immunol 1996;157(7):2909-15.[28] Randow F, MacMicking JD, James LC. Cellular self-defense : how cell-autonomous immunity protects against pathogens. Science 2013;340(6133):701-6.[29] Daeron M. Fc receptor biology. Annu Rev Immunol 1997;15:203-34.[30] Jonsson F, Mancardi DA, Kita Y, et al. Mouse and human neutro-phils induce anaphylaxis. J Clin Invest 2011;121(4):1484-96.[31] Parren PW, Warmerdam PA, Boeije LC, et al. On the interaction of IgG subclasses with the low affinity Fc gamma RIIa (CD32) on human monocytes, neutrophils, and platelets. Analysis of a functional polymor-phism to human IgG2. J Clin Invest 1992;90(4):1537-46.[32] Yee AM, Phan HM, Zuniga R, et al. Association between FcgammaRIIa-R131 allotype and bacteremic pneumococcal pneumo-nia. Clin Infect Dis 2000;30(1):25-8.[33] Platonov AE, Shipulin GA, Vershinina IV, et al. Association of human Fc gamma RIIa (CD32) polymorphism with susceptibility to and severity of meningococcal disease. Clin Infect Dis 1998;27(4):746-50.[34] Bredius RG, Derkx BH, Fijen CA, et al. Fc gamma receptor IIa (CD32) polymorphism in fulminant meningococcal septic shock in child-ren. J Infect Dis 1994;170(4):848-53.[35] Morgan AW, Griffiths B, Ponchel F, et al. Fcgamma receptor type IIIA is associated with rheumatoid arthritis in two distinct ethnic groups. Arthritis Rheum 2000;43(10):2328-34.[36] Tan Sardjono C, Mottram PL, van de Velde NC, et al. Development of spontaneous multisystem autoimmune disease and hypersensitivity to antibody-induced inflammation in Fcgamma receptor IIa-transgenic mice. Arthritis Rheum 2005;52(10):3220-9.[37] Ierino FL, Hulett MD, McKenzie IF, et al. Mapping epitopes of human Fc gamma RII (CDw32) with monoclonal antibodies and recom-binant receptors. J Immunol 1993;150(5):1794-803.[38] Kettritz R. How anti-neutrophil cytoplasmic autoantibodies activate neutrophils. Clin Exp Immunol 2012;169(3):220-8.[39] Wu J, Ji C, Xie F, et al. FcalphaRI (CD89) alleles determine the proin-flammatory potential of serum IgA. J Immunol 2007;178(6):3973-82.[40] Kelley JM, Monach PA, Ji C, et al. IgA and IgG antineutrophil cytoplas-mic antibody engagement of Fc receptor genetic variants influences granu-lomatosis with polyangiitis. Proc Natl Acad Sci USA 2011;108(51):20736-41.


















