
Electrophysiologie: mécanismes et méthodes Activité...
18
biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM Electrophysiologie: mécanismes et méthodes Activité électrique du cœur I. Cellule cardiaque: excitabilité, conduction, automatisme et contractilité 1.1 Anatomie et histologie cardiaque De par leur nature histologique et leur fonction physiologique, on distingue deux types de tissus cardiaques : Le tissu nodal : il est constitué du nœud sinusal (1), du nœud auriculoventriculaire (2), du tronc et des branches du faisceau de His (3) et du réseau de Purkinje (4). Son rôle essentiel est l’élaboration et la conduction de l’influx. Le tissu myocardique : dont la fonction principale est la contraction. Dans les conditions physiologiques l’influx nait périodiquement du nœud sinusal de façon automatique, sans qu’aucune intervention cérébrale ne soit nécessaire. Cet influx est conduit à travers les oreillettes dont il provoque la contraction par des voies anatomiquement mal individualisées. Cet influx passe ensuite au niveau du nœud auriculoventriculaire ( ou nœud de Keith et Flack )situé à la base des oreillettes où il subit un retard d’environ 150 ms , nécessaire à la séparation dans le temps des contractions auriculaires et ventriculaires. Il traverse alors la jonction auriculoventriculaire et chemine à l’intérieur du septum interventriculaire, puis dans les parois ventriculaires en suivant le tronc, puis le faisceau de His. Il est rapidement conduit aux cellules myocardiques ventriculaires par le réseau de Purkinje. Dans les conditions physiologiques, l’influx ne peut passer des oreillettes aux ventricules que par le nœud auriculoventriculaire et le tronc du faisceau de His : il existe en effet, entre les oreillettes et les ventricules, un anneau fibreux ( 5 ) qui n’est traversé par aucune cellule myocardique ou nodale autre que celle du tronc du faisceau de His et qui réalise ainsi l’isolation électrique nécessaire entre oreillettes et ventricules pour que leur contraction respective puisse être décalée dans le temps. Le rôle de conduction du tissu nodal découle d’une vitesse de conduction élevée ( 4 m/s ) par rapport au tissu myocardique ( 0,4 m/s ), sauf au niveau du nœud auriculoventriculaire où la vitesse de conduction, beaucoup plus lente que celle du tissu myocardique, permet la génération du retard d’environ 150 ms Tissu myocardique Tissu nodal 5 3 1 2 3 3 4 3
Transcript of Electrophysiologie: mécanismes et méthodes Activité...

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
Electrophysiologie: mécanismes et méthodes
Activité électrique du cœur
I. Cellule cardiaque: excitabilité, conduction, automatisme et contractilité
1.1 Anatomie et histologie cardiaque
De par leur nature histologique et leur fonction physiologique, on distingue deux types de tissus cardiaques :
Le tissu nodal : il est constitué du nœud sinusal (1), du nœud auriculoventriculaire (2), du tronc et des branches du faisceau de His (3) et du réseau de Purkinje (4). Son rôle essentiel est l’élaboration et la conduction de l’influx.
Le tissu myocardique : dont la fonction principale est la contraction. Dans les conditions physiologiques l’influx nait périodiquement du nœud sinusal de façon automatique, sans qu’aucune intervention cérébrale ne soit nécessaire. Cet influx est conduit à travers les oreillettes dont il provoque la contraction par des voies anatomiquement mal individualisées. Cet influx passe ensuite au niveau du nœud auriculoventriculaire ( ou nœud de Keith et Flack )situé à la base des oreillettes où il subit un retard d’environ 150 ms , nécessaire à la séparation dans le temps des contractions auriculaires et ventriculaires. Il traverse alors la jonction auriculoventriculaire et chemine à l’intérieur du septum interventriculaire, puis dans les parois ventriculaires en suivant le tronc, puis le faisceau de His. Il est rapidement conduit aux cellules myocardiques ventriculaires par le réseau de Purkinje. Dans les conditions physiologiques, l’influx ne peut passer des oreillettes aux ventricules que par le nœud auriculoventriculaire et le tronc du faisceau de His : il existe en effet, entre les oreillettes et les ventricules, un anneau fibreux ( 5 ) qui n’est traversé par aucune cellule myocardique ou nodale autre que celle du tronc du faisceau de His et qui réalise ainsi l’isolation électrique nécessaire entre oreillettes et ventricules pour que leur contraction respective puisse être décalée dans le temps. Le rôle de conduction du tissu nodal découle d’une vitesse de conduction élevée ( 4 m/s ) par rapport au tissu myocardique ( 0,4 m/s ), sauf au niveau du nœud auriculoventriculaire où la vitesse de conduction, beaucoup plus lente que celle du tissu myocardique, permet la génération du retard d’environ 150 ms
Tissu myocardique Tissu nodal
5
3
1
2
3
3
4
3

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
1.2 électrophysiologie cardiaque
Comme dans le cas des cellules nerveuses, la membrane de la cellule cardiaque est polarisée et le caractère excitable de cette cellule se traduit par l’apparition d’un potentiel d’action. Néanmoins les cellules cardiaques possèdent leur propre spécificité.
1.2.1 potentiel de membrane des cellules cardiaques
* Les cellules myocardiques ont un potentiel de repos de – 90 mV. Le potentiel d’action d’une cellule cardiaque possède un plateau, ce qui lui confère une durée dans le temps.
* Les cellules du faisceau de His et du réseau de Purkinje ont également un potentiel d’action en plateau, mais elle présente la particularité d’avoir un potentiel de repos qui n’est pas stable dans le temps : la polarisation membranaire diminue avec le temps.
Il existe donc une dépolarisation spontanée de la cellule, caractérisée par la pente de dépolarisation diastolique ( diastole = durée pendant laquelle le cœur est en relâchement musculaire ). Lorsqu‘elle atteint le seuil d’excitation, cette dépolarisation engendre un potentiel d’action. L’existence de cette pente a pour conséquences :
Cette pente est responsable de l’automatisme de la cellule
Des drogues et des stimulations nerveuses pouvant augmenter cette pente augmentent la fréquence de l’automatisme ; inversement des drogues et des stimulations nerveuses pouvant diminuer cette pente diminuent la fréquence de l’automatisme
* les cellules du nœud sinusal et du nœud auriculoventriculaire ont une polarisation moins importante et présente une montée lente du potentiel d’action, rendant compte d’une vitesse de conduction très diminuée, ce qui retarde l’influx.
La pente de dépolarisation diastolique est importante, rendant compte d’un automatisme à fréquence élevée. Cette pente est maximale pour les cellules du nœud sinusal, ce qui explique que ces cellules ont normalement le contrôle du rythme cardiaque.

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
La figure ci-dessous recense les potentiels d’action des différentes cellules cardiaques :
1.2.2 phénomènes ioniques
Comme pour les autres cellules excitables, le potentiel d’action des cellules cardiaques est dû à une variation de la perméabilité membranaire aux différents ions.
Phase 1 : phase de dépolarisation rapide due à l’activation du canal sodique, comme dans le
cas de la fibre nerveuse
Phase 2 : phase en plateau due à l’activation plus lente d’une protéine canal laissant entrer par diffusion les ions sodium et calcium. Le maintien d’une perméabilité membranaire élevée au sodium explique le maintien du potentiel à une valeur proche du potentiel d’équilibre du sodium L’entrée du calcium par diffusion dans la cellule qui n’en contient quasiment pas est responsable de la contraction musculaire
Phase 3 : phase de repolarisation. Cette phase correspond à une augmentation de la perméabilité membranaire du potassium
Phase 4 : pour récupérer son état initial, la cellule doit faire sortir le sodium et le calcium entrés pendant la phase de dépolarisation et faire entrer le potassium sorti pendant la phase de repolarisation. Cela nécessite un transport actif de sodium, de potassium, mais aussi de calcium.
La phase de dépolarisation diastolique des cellules du tissus nodal semble être due à une diminution progressive de la perméabilité membranaire au potassium pendant la diastole. Le potentiel membranaire s’adapte donc à cette variation de perméabilité. Durant toutes les phases du potentiel d’action, la relation de Goldman est vérifiée

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
1.3 physiopathologie
* troubles de l’automatisme : Il peut s’agir
D’un défaut d’automatisme ( pause sinusale )
De l’apparition d’un foyer automatique ectopique ( qui n’est pas situé à l’endroit normal, c’est-à-dire non situé au niveau du nœud sinusal ) rentrant alors en concurrence avec le nœud sinusal et responsable « d’extrasystoles » ou de crises de « tachycardie ».
* troubles de la conduction : Il peut s’agir :
D’un défaut de conduction des voies normales de conduction ( par exemple : du tronc ou des branches du faisceau de His ), responsable de blocs auriculoventriculaires ( BAV )
De l’existence d’une voie anormale de conduction ( par exemple : le faisceau de Kent, constitué de cellules myocardiques qui relient l’oreillette et le ventricule, court-circuite le nœud auriculoventriculaire sans provoquer le retard physiologique de 0,15 s [syndrome de Wolff-Parkinson-White] ). Cette voie anormale est responsable d’une dépolarisation prématurée du ventricule.
* troubles de l’excitabilité : De façon physiologique, la cellule devient de moins en moins excitable lors de la phase 4 de dépolarisation diastolique. * phénomène de réentrée : Entre deux points, existent deux voies de conduction, non nécessairement anatomiquement distinctes, mais fonctionnellement distinguables par la durée de leur période réfractaire. Un influx arrive précocément, par rapport à l’influx précédent, à l’origine de ces deux voies en période réfractaire pour l’une voie, et non pour l’autre. Le démarrage d’un cycle de tachycardie par réentrée est alors possible. B A B A B A L’influx descendant arrive à l’instant où la voie B est réfractaire et non la voie A. l’influx est transmis aux ventricules uniquement par la voie A.
La voie B sort de sa période réfractaire lorsque l’influx venant de A l’atteint par voie rétrograde
L’influx ayant traversé B se retransmet à nouveau aux deux ventricules par la voie A.

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
1.4 pharmacologie et traitement
Les troubles de l’automatisme peuvent être prévenus par des drogues agissant sur la pente de dépolarisation diastolique. Les troubles de la conduction peuvent être modifiés par des médicaments agissant sur la vitesse de la phase de dépolarisation rapide ( phase 0 ) ou sur la durée de période réfractaire.
Ainsi, en augmentant la période réfractaire de l’une des deux voies d’un circuit de réentrée, on peut bloquer le cycle responsable de la tachycardie. Les médicaments arythmiques, répartis en différentes classes, agissent plus ou moins sélectivement sur l’une des phases membranaires :
L’inhibition du canal sodique rapide ( phase 0 ) par les médicaments « quinidine-like » entraînant un ralentissement de la conduction, en agissant sur la pente de dépolarisation rapide.
L’inhibition du courant lent calcicosodique ( phase 2 ) par les médicaments appelés « inhibiteurs calciques » déprime l’excitabilité et la conduction des fibres à réponse lente. La diminution intramyocardique du calcium entraîne également une dépression de la contractibilité, expliquant le fait que ce médicament est contre indiqué dans l’insuffisance cardiaque
Les béta bloquants diminuent la pente de dépolarisation diastolique Lorsque les drogues sont inactives, l’implantation d’un stimulateur cardiaque intracorporel ( pacemaker ) peut devenir nécessaire. Celui-ci est un boitier relié au cœur par une sonde et délivrant une impulsion de stimulation lorsqu’il ne détecte aucune stimulation spontanée.

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
II. électrocardiogramme L’électrocardiogramme consiste en l’enregistrement en fonction du temps de l’activité électrique du coeur au moyen d’électrodes placées à la surface du corps Ainsi recueille t_on des potentiels en milieu conducteur L’ECG reconstitue à partir de l’enregistrement du potentiel en certains points du milieu extérieur, l’état d’activation du coeur dans son ensemble On considère que l’organisme est un corps conducteur homogène constitué de solutions contenant des ions
2.1 évolution du potentiel en fonction de l’état d’excitation d’une fibre myocardique
Une fibre ou un groupe de fibres au repos impose entre les compartiments intra et extra cellulaires une différence de potentiel définissant le potentiel de repos. Tous les points du compartiment extracellulaires sont au même potentiel et deux électrodes placées sur la surface du corps n’enregistreront par conséquent aucune différence.
Il en est de même pour une fibre ou un groupement de fibres complètement dépolarisés :
En revanche, une fibre ou un groupement de fibres partiellement dépolarisées créent un gradient de potentiel dans le compartiment extracellulaire du fait de l’existence de courants locaux.

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
2.2 potentiel créé par un dipôle
Entre 2 charges électriques placées à distance, une force électrique de coulomb apparaît.
Chaque charge crée un champ électrique E
qui dérive du potentiel V : E
= - grad V
Une charge en un point P crée un gradient de potentiel. Ce potentiel V évolue inversement avec la distance qui sépare la charge de ce point P :
V = r
q
4
1
avec = o.r où o est la permittivité du vide telle que o4
1= 9.109 ou o =
36
1.10-9 et r est la
permittivité relative du milieu ou la constante diélectrique du milieu. Dans l’eau r est de l’ordre de 80. Un dipôle électrostatique est constitué de deux charges opposées +q et –q distantes de d, distance très petite devant la distance r d’un point P au barycentre de ce dipôle. Les deux charges du dipôle créent donc en un point P le potentiel V donné par :
En milieu liquide, le maintien de ce dipôle nécessite de l’énergie, ce présentant sous la forme d’un courant électrique.
V =
214
1
r
q
r
q
On définit le moment dipolaire de ce dipôle par la
relation M
= q. d
Le potentiel s’écrit sous la forme :
V = ²
.
4
1
r
uM p
pu
étant un vecteur unitaire

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
2.3 potentiel créé par une fibre isolée au repos ou complètement dépolarisée
L’élément de potentiel électrique créé en P par dS s’écrit sous la forme : dV = ²
.
4
1
r
uMd p
Ce qui donne en calculant le produit scalaire : dV = ²
cos...
4
1
r
dS
On appelle puisse de l’élément de feuillet la quantité µ =
On obtient donc : dV = ²
cos..
4
1
r
dSµ
On appelle d l’élément d’angle solide sous lequel le point P observe l’élément de surface dS. On a
d = ²
cos.
r
dS .
L’élément de potentiel dV s’écrit donc : dV = dµ4
1
Si la densité surfacique de charge et l’épaisseur membranaire sont uniformes, la puissance µ du feuillet est aussi uniforme. On peut donc intégrer l’ensemble des éléments de potentiels créés par le feuillet.
Une face de la membrane ( face présentée à P ) génère le potentiel V+ = 4
.µ
P
L’autre face de la membrane, de même répartition surfacique, de même épaisseur dipolaire, mais
présentant à P non pas les charges + mais les charges -, génère le potentiel V- = -4
.µ
La somme des deux potentiels créés par ces deux feuillets donne V = V+ + V- = 0 Au repos, ou complètement dépolarisée, le potentiel créé par une fibre est nul.
On appelle feuillet électrique un ensemble de deux distributions de charges de signes contraires réparties sur les faces d’une lame très mince ( dont l’épaisseur est très faible par rapport à ses dimensions de surface ). Une membrane cellulaire est assimilable à un feuillet pour les mêmes raisons.
On considère un feuillet d’épaisseur et de densité de
charge surfacique = dq / dS. Le point P, distant de r de cet élément de surface, voit cet élément de surface comme un élément de dipôle
électrostatique de moment dipolaire dM = dq = dS.

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
2.4 potentiel créé par une fibre isolée partiellement dépolarisée
En voie de dépolarisation ou de repolarisation, Ω1 et Ω3 ne contribuent pas au potentiel : V = 0
Par contre, sous 2, l’angle solide contient deux portions de feuillet présentant à P leur face négative. Le potentiel créé en P par ces deux
portions de feuillet est : V = 221 )(4
1 µ
Si A est la surface de dépolarisation de la
membrane, et r suffisamment grand, 2 = ²
cos.
r
A
Dans ce cas, V = ²
cos.)(
4
121
r
Aµ
.
Cette relation montre qu’une fibre partiellement dépolarisée est assimilable à un dipôle dont le
moment M
est perpendiculaire au front d’activation, orienté de la zone dépolarisée vers la zone au repos et de valeur (µ1 + µ2 ) A. Ce dipôle se déplace avec le front d’excitation. La propagation de ce moment dipolaire crée un potentiel en P évoluant au cours du temps : V t
2.5 cas d’un groupement de fibres
Si on considère un groupe de fibres partiellement dépolarisées, on peut remplacer chaque fibre par son dipôle équivalent. L’ensemble des dipôles forme un feuillet dont la frontière est entre la zone active et la zone encore au repos. Ce feuillet se déplace avec le front de dépolarisation (activation). Donc cette propagation peut-être représentée par la progression d’un dipôle avec un pôle positif vers le milieu non encore dépolarisé. Donc dans le cas d’une dépolarisation :
• si le front de dépolarisation va vers P, P voit l’extrémité + du dipôle s’approcher et donc son potentiel devenir de plus en plus positif.
• A l’opposé si le front de dépolarisation s’éloigne de P, son potentiel est de plus en plus négatif.
• Dans le cas d’une repolarisation, quand le front s’approche de P, P voit son extrémité négative s’approcher et donc son potentiel de plus en plus négatif. A l’inverse quand ce front s’éloigne de P son potentiel est de plus en plus positif
Dans cet exemple, le front d’excitation s’approche de P puis s’en éloigne

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
Le potentiel recueilli par une électrode placée en un point fixe varie si la valeur de projection du moment du dipôle sur l’axe de la dérivation varie
– en amplitude – en direction – en sens
Le potentiel varie aussi si le moment (même d’amplitude, de direction et de sens constants ) se déplace : s’approche ou s’éloigne de l’électrode
2.6 cas d’un organe
Pour un organe, Il en résulte un dipôle global variable M
(t), somme vectorielle de tous les dipôles cellulaires instantanés μ(t). La mesure du signal électrophysiologique traduit les variations du potentiel aux bornes d’une
dérivation (par projection de M
(t) sur l’axe de la dérivation).
Vague de dépolarisation
biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
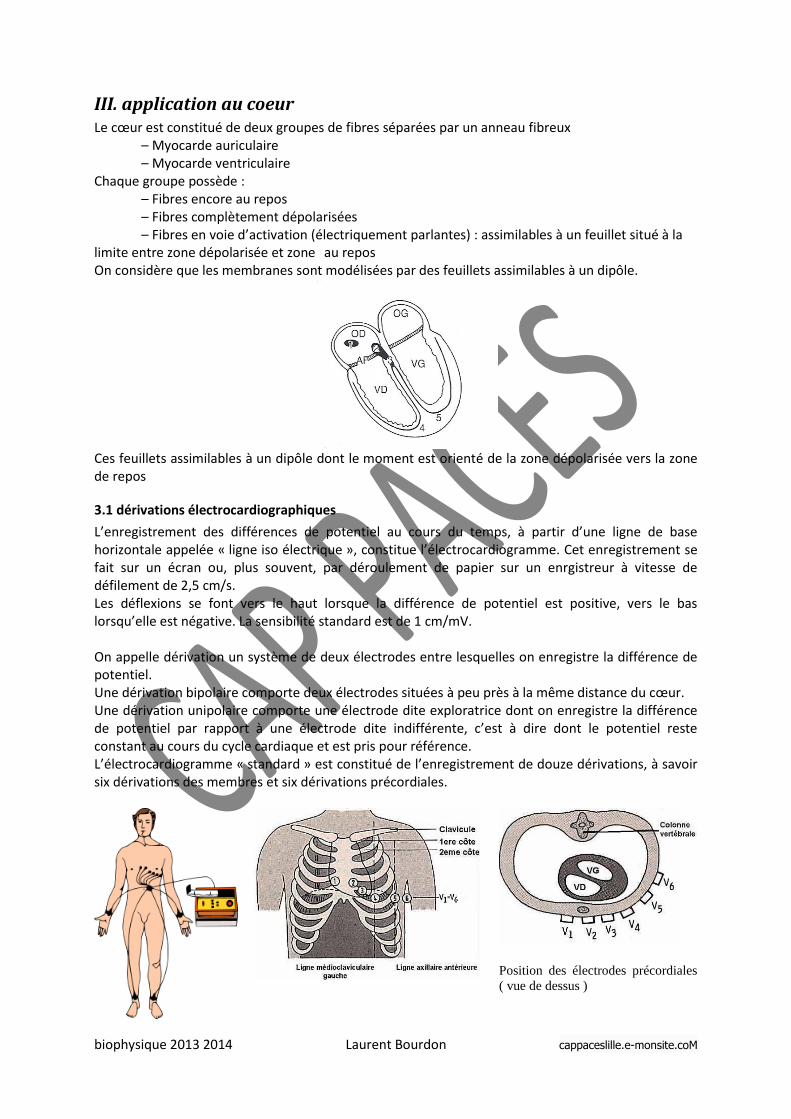
III. application au coeur Le cœur est constitué de deux groupes de fibres séparées par un anneau fibreux
– Myocarde auriculaire – Myocarde ventriculaire
Chaque groupe possède : – Fibres encore au repos – Fibres complètement dépolarisées – Fibres en voie d’activation (électriquement parlantes) : assimilables à un feuillet situé à la
limite entre zone dépolarisée et zone au repos On considère que les membranes sont modélisées par des feuillets assimilables à un dipôle.
Ces feuillets assimilables à un dipôle dont le moment est orienté de la zone dépolarisée vers la zone de repos
3.1 dérivations électrocardiographiques
L’enregistrement des différences de potentiel au cours du temps, à partir d’une ligne de base horizontale appelée « ligne iso électrique », constitue l’électrocardiogramme. Cet enregistrement se fait sur un écran ou, plus souvent, par déroulement de papier sur un enrgistreur à vitesse de défilement de 2,5 cm/s. Les déflexions se font vers le haut lorsque la différence de potentiel est positive, vers le bas lorsqu’elle est négative. La sensibilité standard est de 1 cm/mV. On appelle dérivation un système de deux électrodes entre lesquelles on enregistre la différence de potentiel. Une dérivation bipolaire comporte deux électrodes situées à peu près à la même distance du cœur. Une dérivation unipolaire comporte une électrode dite exploratrice dont on enregistre la différence de potentiel par rapport à une électrode dite indifférente, c’est à dire dont le potentiel reste constant au cours du cycle cardiaque et est pris pour référence. L’électrocardiogramme « standard » est constitué de l’enregistrement de douze dérivations, à savoir six dérivations des membres et six dérivations précordiales.
Position des électrodes précordiales
( vue de dessus )

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
3.1.1 dérivations des membres
Cette dénomination provient de ce que les électrodes sont placées sur les membres : les poignets droit et gauche et une jambe ( conventionnellement la gauche ). En théorie, ces électrodes devraient être placées sur les épaules droite ( R ) et gauche ( L ) et sur le bas ventre ( F ). On considère que lors du recueil de ces potentiels, les membres jouent simplement le rôle de conducteur. VR : potentiel de l’électrode placée sur le membre supérieur droit ( Right ) VL : potentiel de l’électrode placée sur le membre supérieur gauche ( Left ) VF : potentiel de l’électrode placée sur l’un ou l’autre membre inférieur ( Foot )
Les dérivations des membres comportent : 3 dérivations bipolaires:
– DI = VL –VR ; – DII = VF –VR ; – DIII = VF –VL
3 dérivations unipolaires : – VR = VR –VW ; – VL = VL –VW ; – VF = VF –VW
VW étant un potentiel d’électrode constant au cours du temps 3.1.2 dérivations précordiales
Les dérivations précordiales comportent six dérivations unipolaires V1 à V6, constituées de l’enregistrement de référence VW du potentiel V1 à V6 de six électrodes placées sur le thorax en des endroits précis et universellement adoptés. Elles explorent le cœur dans un plan horizontal et sont placées relativement près de lui, ce qui ne permet plus d’assimiler le cœur à un dipôle : la théorie du feuillet doit donc y être appliquée

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
3.2 théorie d’Einthoven
Cette théorie simplificatrice permet d’expliquer les tracés observés dans les dérivations périphériques (enregistrement à grande distance du coeur). * Première hypothèse : A chaque instant, le potentiel créé par le cœur en voie de dépolarisation ou de repolarisation peut être assimilé à celui créé par un dipôle unique dans un milieu conducteur homogène. Le vecteur moment varie en origine, module, direction et sens au cours du cycle cardiaque.
D’où Vp = ²
.
4
1
r
uM P
avec P = R, L ou F.
* Deuxième hypothèse : L’origine du vecteur moment peut être considérée comme fixe. On l’appelle centre électrique du cœur. Il se situe au point O, dans le ventricule gauche.
Centre électrique du cœur
vectocardiogramme
La courbe décrite par l’extrémité du vecteur moment s’appelle vectocardiogramme. * Troisième hypothèse : Les 3 points R, L, et F sont assimilés aux sommets d’un triangle équilatéral dont le centre de gravité est le centre électrique du cœur : les sommets R L F sont à égale distance du centre électrique du cœur.
Si on pose k = ²4
1
r
VR = k M
. Ru
VL = k M
. Lu
VF = k M
. Fu
De par l’inclinaison des trois vecteurs Ru
, Lu
et Fu
, on
aboutit à : VR + VL + VF = 0 Pour obtenir le potentiel de référence nécessaire à l’enregistrement des dérivations unipolaires ( dérivations précordiales et dérivations unipolaires des membres ), il suffit donc d’enregistrer le potentiel VR + VL + VF. Ce potentiel est obtenu à l’aide d’un circuit additionneur équivalent à trois résistances égales et de valeur R suffisamment élevée pour ne pas modifier les valeurs des potentiels VR, VL et VF.
On construit alors la borne centrale de Wilson dont le potentiel VW est bien nul. La loi des nœuds de Kirschoff permet d’écrire :
R
VV WR = R
VV WL =R
VV WF = 0
Soit VW = 3
1( VR + VL + VF )

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
En ce qui concerne les dérivations bipolaires des membres, si l’on prend pour exemple l’enregistrement de DI, on a :
DI = VL – VR = ²4
1
rM
. ( Lu
- Ru
) = k M
. ( Lu
- Ru
)
Or, Lu
- Ru
est de même direction et de même sens que R L et donc que le vecteur unitaire DIu
de la
direction DI.
Néanmoins, Lu
- Ru = 2 Ru
sin 30 = 3 = 1,7.
On en déduit que : DI = VL – VR = ²4
1
rM
. ( Lu
- Ru
) 3 = 3 k M
DIu
= k’ M
DIu
Ainsi, les différences de potentiel DI, DII et DIII sont elles aussi, proportionnelles à la projection
du moment cardiaque sur l’axe DIu
, DIIu
, DIIIu
de la dérivation correspondante. Cependant, le coefficient de proportionnalité intervenant dans les dérivations bipolaires est supérieur ( d’un facteur 1,7 ) à celui intervenant dans les dérivations unipolaires. On a pris l’habitude d’enregistrer les dérivations augmentées :
aVR = 1,5 VR aVL = 1,5 VL aVF = 1,5 VF Ces dérivations augmentées fournissent un tracé identique à celui des dérivations unipolaires classiques correspondantes. Elles présentent l’avantage de présenter un coefficient de
proportionnalité ( 1,5 ) proche de 3 = 1,7.
3.3 tracé électrocardiogramme
Quelle que soit la dérivation considérée, on enregistre toujours la même séquence au cours du cycle cardiaque.
L’onde P correspond à la dépolarisation auriculaire. Elle est normalement positive en D1, D2 et D3.
Le complexe qRs correspond à la dépolarisation ventriculaire. Il comprend dans sa forme complète une déflexion négative q, une déflexion positive R et une déflexion négative s
L’onde T correspond à la repolarisation ventriculaire. L’onde de repolarisation auriculaire n’est pas vue, car elle est masquée par le complexe qRs. L’intervalle PR compris entre le début de l’onde P et le début du complexe qRs est normalement iso électrique et traduit le délai entre l’activation des oreillettes et celle des ventricules. Ce délai est obtenu grâce au retard pris par l’influx dans le nœud auriculoventriculaire. Le segment ST entre la fin du complexe qRs et le début de l’onde T est normalement iso électrique.
biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
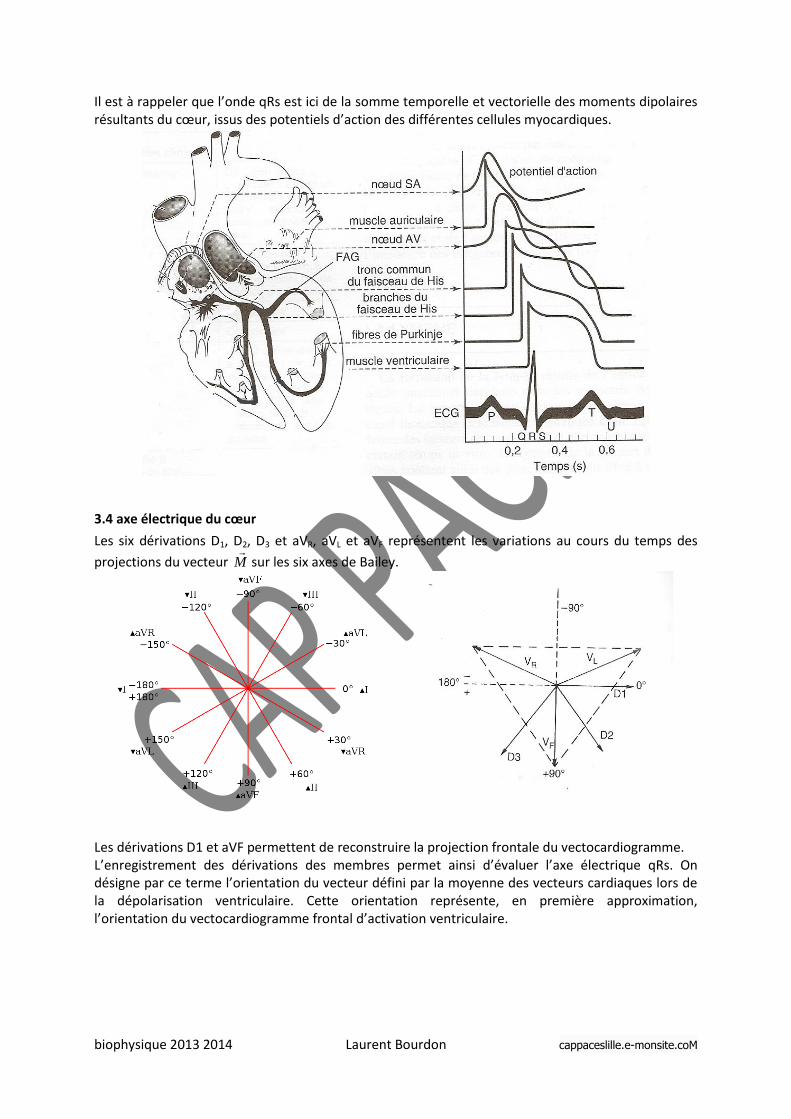
Il est à rappeler que l’onde qRs est ici de la somme temporelle et vectorielle des moments dipolaires résultants du cœur, issus des potentiels d’action des différentes cellules myocardiques.
3.4 axe électrique du cœur
Les six dérivations D1, D2, D3 et aVR, aVL et aVF représentent les variations au cours du temps des
projections du vecteur M
sur les six axes de Bailey.
Les dérivations D1 et aVF permettent de reconstruire la projection frontale du vectocardiogramme. L’enregistrement des dérivations des membres permet ainsi d’évaluer l’axe électrique qRs. On désigne par ce terme l’orientation du vecteur défini par la moyenne des vecteurs cardiaques lors de la dépolarisation ventriculaire. Cette orientation représente, en première approximation, l’orientation du vectocardiogramme frontal d’activation ventriculaire.

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
Ci-dessous sont représentées les projections du complexe qRs en D1, D2, VF et VL.
Ci-dessous sont représentées les déviations de l’axe électrique de qRs.
3.5 rythme cardiaque
Le rythme régulier sinusal est de l’ordre de 70 pulsations par minute. La fréquence cardiaque est déterminée grâce au laps de temps existant entre deux qRs stables consécutifs. Chaque onde P est suivie d’un complexe qRs L’ECG permet de distinguer :
Bradycardies :
Tachycardies :
Arythmies :
pauses :
extrasystoles :

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
3.6 troubles de la conduction
* L’intervalle PR est normalement compris entre 0,12 et 0,20 s. Un intervalle PR supérieur à 0,20 s traduit un bloc auriculoventriculaire. * Les troubles de la conduction ( blocs ) peuvent siéger sur l’une des branches du faisceau de His ( blocs de branche droit ou gauche ). Dans ce cas, le cycle qRs est supérieur à 0,12 s, s’il est complet. * il peut s’agir d’un bloc auriculoventriculaire, dont le trouble de la conduction se situe soit sur les deux branches, soit en amont ( nœud sinusal, nœud auriculoventriculaire, tronc du faisceu de His ). Ce bloc possède trois degrés de gravités croissante : * bloc du premier degré : l’excitation passe toujours, mais avec un retard, entraînant un allongement de l’intervalle PR, devenant supérieur à 0,2 seconde * bloc du troisième degré : l’excitation ne passe jamais et l’onde P n’est pas suivie du complexe qRs : l’onde P est dite bloquée. La contraction est assurée par les cellules en aval du bloc qui peuvent alors exprimer leur faculté d’automatisme, néanmoins moins fiable et de fréquence plus lente que l’automatisme sinusal. Ce bloc justifie la pause d’un pacemaker * le bloc de deuxième degré : il est de gravité intermédiaire et se traduit par la présence d’ondes P bloquées survenant plus ou moins périodiquement. * Le syndrome de Wolff-Parkinson-White est un trouble de la conduction dû à la présence d’une voie accessoire de conduction traversant l’anneau fibreux et court-circuite le nœud auriculoventriculaire. Il entraîne une diminution de l’intervalle PR car une partie du ventricule se trouve excitée en avance par l’influx provenant des oreillettes qui est transmis directement par la voie accessoire, sans subir le retard physiologique d’environ 0,15 s au niveau du nœud auriculoventriculaire.
3.7 hypertrophie et troubles ischémiques
L’hypertrophie de l’un des ventricules entraîne une déviation de l’axe qRs vers le ventricule hypertrophié. L’axe sera compris entre +90° et + 180° se traduisant par un complexe qRs anormalement négatif en D1 en cas d’hypertrophie ventriculaire droit.
L’axe sera compris entre 0° et - 90° se traduisant par un complexe qRs anormalement négatif en VF en cas d’hypertrophie ventriculaire gauche. L’ischémie des cellules myocardiques ( angine de poitrine ) entraîne des modifications de la repolarisation. On observe un sous décallage ou sus décallage du segment ST La nécrose correspond à des cellules myocardiques ( infarctus du myocarde ) remplacées par un tissu fibreux non contractile et dépourvu d’activité électrique

biophysique 2013 2014 Laurent Bourdon cappaceslille.e-monsite.coM
L’ECG permet en outre un diagnostic topologique si l’on remarque que pour le ventricule gauche : V2, V3 regardent la face antérieure D1, VL et V4, V5, V6 regardent la face latérale D2, D3, aVF regardent la face postérodiaphragmique Quant au ventricule droit, il est au mieux examiné par les dérivations D3 et V1, V2, V3.


















