
Rapport final Incertitude & Flexibilité des Traitements de Substitution

Effet de la réduction et de la substitution du NaCl dans le Camembert sur la croissance de la microflore
fongique d'affinage
Mémoire
Marilyne Touchette
Maîtrise en microbiologie agroalimentaire Maître ès sciences (M. Sc.)
Québec, Canada
© Marilyne Touchette, 2016


iii
Résumé
Santé Canada encourage les transformateurs à réduire le contenu en NaCl dans plusieurs aliments, dont les
fromages, puisque sa surconsommation est associée à plusieurs problèmes de santé. L'effet causé par la
réduction du NaCl dans le Camembert, ou de sa substitution partielle par du KCl, sur la qualité globale de ce
fromage n'est pas connu. Le but du projet visait à étudier la cinétique d'incorporation du sodium et du
potassium lors du saumurage d'un Camembert commercial afin de produire des fromages réduits en NaCl et
substitués par du KCl. Ces nouvelles conditions ont ensuite permis d'étudier le comportement des ferments
fongiques d'affinage Penicillium camemberti et Geotrichum candidum. Les résultats ont démontré une
modification du profil de croissance des deux espèces, plus accentuée chez G. candidum, en fonction des
modifications salines apportées. L'impact causé par ces changements de comportement doit maintenant être
évalué au niveau de l'activité spécifique des deux espèces.


v
Table des matières
Résumé ............................................................................................................................................................. iii
Table des matières ............................................................................................................................................ v
Liste des tableaux ............................................................................................................................................ ix
Liste des figures ............................................................................................................................................... xi
Liste des annexes ........................................................................................................................................... xv
Liste des abréviations et des sigles ............................................................................................................ xvii
Remerciements .............................................................................................................................................. xix
Avant-propos .................................................................................................................................................. xxi
Introduction ....................................................................................................................................................... 1
1. Revue de littérature .................................................................................................................................. 3
1.1. Le chlorure de sodium dans l'alimentation et son importance en fromagerie .................................... 5
1.1.1. La consommation du sodium chez les Canadiens .................................................................... 5
1.1.1.1. L'apport en sodium ............................................................................................................... 5
1.1.1.2. Les sources de sodium ......................................................................................................... 6
1.1.2. L'effet du sodium sur la santé humaine ..................................................................................... 7
1.1.3. La réduction du sodium dans l'alimentation .............................................................................. 8
1.1.3.1. La stratégie de réduction du sodium au Canada .................................................................. 9
1.1.4. Les rôles du sel en fromagerie ................................................................................................ 10
1.1.4.1. Le complément d'égouttage................................................................................................ 11
1.1.4.2. La réduction de l'activité de l'eau ........................................................................................ 12
1.1.4.3. Le développement des propriétés sensorielles ................................................................... 16
1.2. Le fromage Camembert ................................................................................................................... 18
1.2.1. La fabrication des fromages de type Camembert ................................................................... 18
1.2.2. Les techniques de salage en fromagerie ................................................................................ 20
1.2.2.1. Le salage à sec................................................................................................................... 21
1.2.2.2. Le saumurage ..................................................................................................................... 21
1.2.3. Le dosage du sel dans les fromages ...................................................................................... 26
1.3. L'affinage du camembert et sa microflore fongique de surface ....................................................... 28
1.3.1. Les phénomènes biochimiques ............................................................................................... 28
1.3.2. L'évolution microbienne .......................................................................................................... 30
1.3.3. Penicillium camemberti ........................................................................................................... 31
1.3.3.1. Les rôles de P. camemberti lors de l'affinage ..................................................................... 31
1.3.4. Geotrichum candidum ............................................................................................................. 33
1.3.4.1. Les rôles de G. candidum lors de l'affinage ........................................................................ 33
1.3.5. L'osmotolérance chez les mycètes ......................................................................................... 34

vi
1.3.5.1. Comportement de P. camemberti en fonction du sel .......................................................... 35
1.3.5.2. Comportement de G. candidum en fonction du sel ............................................................. 35
1.3.6. Le suivi de croissance des mycètes pendant l'affinage ........................................................... 36
1.4. La réduction du sodium dans les fromages ..................................................................................... 39
1.4.1. Les cibles de réduction ............................................................................................................ 39
1.4.2. Les stratégies de réduction du sodium dans les fromages ..................................................... 39
1.4.2.1. Utilisation du KCl à titre de sel de remplacement ............................................................... 41
1.5. Problématique, Hypothèse et objectifs ............................................................................................. 43
2. Étude de la cinétique d'incorporation du sodium et du potassium dans un fromage Camembert industriel saumuré ...................................................................................................................... 45
2.1. Résumé ............................................................................................................................................ 47
2.2. Introduction ...................................................................................................................................... 48
2.3. Matériel et méthodes ....................................................................................................................... 50
2.3.1. Matériel à l'étude ................................................................................................................. 50
2.3.2. Préparation des saumures .................................................................................................. 50
2.3.3. Saumurage et affinage des fromages Camemberts ........................................................... 51
2.3.4. Analyses de composition des fromages .............................................................................. 52
2.4. Résultats et discussion .................................................................................................................... 53
2.4.1. Production d'un Camembert industriel de composition saline standard .................................. 53
2.4.2. Production d'un Camembert industriel réduit en sodium ......................................................... 54
2.4.3. Production d'un Camembert industriel réduit en sodium et enrichie en potassium ................. 56
2.4.3.1. Premier essai de saumurage .............................................................................................. 56
2.4.3.2. Optimisation des paramètres de saumurage (2e essai de saumurage) .............................. 59
2.5. Conclusions et perspectives ............................................................................................................ 62
3. Effet de la réduction du chlorure de sodium dans un Camembert industriel et de sa substitution partielle par du chlorure de potassium sur la croissance de Penicillium camemberti et Geotrichum candidum ................................................................................................................................ 63
3.1. Résumé ............................................................................................................................................ 65
3.2. Introduction ...................................................................................................................................... 66
3.3. Matériel et méthodes ....................................................................................................................... 68
3.3.1. Variation intra-espèce de la résistance au NaCl et KCl chez P. camemberti .......................... 68
3.3.1.1. Matériel biologique à l'étude ............................................................................................... 68
3.3.1.2. Fabrication du fromage Camembert déshydraté ................................................................. 68
3.3.1.3. Préparation des CMC et des milieux de culture synthétiques ............................................. 68
3.3.1.4. Suivi de croissance en fonction du NaCl et du KCl ............................................................. 69
3.3.2. Suivi de la croissance de la microflore fongique d'affinage dans un fromage Camembert industriel ................................................................................................................................ 70
3.3.2.1. Matériel à l'étude ................................................................................................................. 70

vii
3.3.2.2. Préparation des saumures et saumurage ........................................................................... 70
3.3.2.3. Affinage et échantillonnage ................................................................................................ 71
3.3.2.4. Analyses de composition des Camemberts ........................................................................ 72
3.3.2.5. Suivi de croissance de P. camemberti et G. candidum au cours de l'affinage .................... 73
3.3.3. Analyses statistiques............................................................................................................... 74
3.4. Résultats et discussion .................................................................................................................... 76
3.4.1. Variation intra-espèce de la résistance au NaCl et KCl chez P. camemberti .......................... 76
3.4.1.1. Croissance sur milieu de culture synthétique ..................................................................... 76
3.4.1.2. Croissance sur caillé modèle de type Camembert (CMC) .................................................. 79
3.4.2. Suivi de la croissance de la microflore fongique d'affinage dans un fromage Camembert industriel ............................................................................................................................... 82
3.4.2.1. Analyses de composition .................................................................................................... 82
3.4.2.2. Suivi du pH ......................................................................................................................... 87
3.4.2.3. Suivi de croissance des ferments fongiques d'affinage ...................................................... 89
3.5. Conclusion et perspectives .............................................................................................................. 95
Conclusion générale et perspectives ............................................................................................................ 97
Cinétique d'incorporation du sodium et du potassium dans un Camembert industriel saumuré .......... 97
Variabilité intra-spécifique de la résistance au NaCl et au KCl chez P. camemberti ............................ 98
Étude de la réduction du NaCl dans un Camembert industriel et de sa substitution partielle par du KCl sur la croissance des ferments fongiques ................................................................................ 99
Annexes ......................................................................................................................................................... 101
Bibliographie ................................................................................................................................................. 105


ix
Liste des tableaux
Tableau 1.1 – Teneur en NaCl des fromages selon différentes variétés (Adapté de Sutherland, 2002 et Guinee et Fox, 2004)......................................................................................................................................... 11
Tableau 1.2 – Activité de l'eau minimale pour la croissance de différents microorganismes (Adapté de Gillis, 2004; Guinee et Fox, 2004 et Guinee et O'Kennedy, 2007). ................................................................... 13
Tableau 1.3 – Activité de l'eau de différentes variétés fromagères (Adapté de Guinee et Fox, 2004 et Guinee et O'Kennedy, 2007). ............................................................................................................................ 13
Tableau 2.1 – Composition saline des saumures ............................................................................................. 51
Tableau 2.2 – Teneur en humidité et en cendres des fromages au 2e et 4e jour d'affinage .............................. 54
Tableau 2.3 – Teneurs en humidité et en minéraux des fromages au 4e jour d'affinage .................................. 56
Tableau 2.4 – Teneurs en sodium et en potassium des fromages au 4e jour d'affinage et proportion de réduction en sodium correspondante ................................................................................................................ 58
Tableau 2.5 – Teneurs en humidité et en cendres des fromages au 4e jour d'affinage .................................... 59
Tableau 2.6 – Teneurs en sodium et en potassium des fromages au 4e jour d'affinage et proportion de réduction en sodium correspondante ................................................................................................................ 61
Tableau 2.7 – Teneurs en humidité et en cendres des fromages au 4e jour d'affinage .................................... 62
Tableau 3.1 – Composition saline des saumures et temps de saumurage ...................................................... 71
Tableau 3.2 – Journées d’échantillonnage des Camemberts, concordance avec les étapes d’affinage du fromage dans l’industrie participant au projet. .............................................................................................. 72
Tableau 3.3 – Gènes ciblés et séquences nucléotidiques des amorces et des sondes utilisées pour la quantification par PCR en temps réel de P. camemberti et de G. candidum. ................................................... 73
Tableau 3.4 – Table d'ANOVA associée aux analyses statistiques sur la mesure du diamètre des colonies sur milieux PDA acidifié ...................................................................................................................... 78
Tableau 3.5 – Comparaisons multiples sur l'interaction triple souches*types de sel*concentrations en conditionnant sur la concentration ..................................................................................................................... 79
Tableau 3.6 – Table d'ANOVA associée aux analyses statistiques sur la mesure du diamètre des colonies sur caillé modèle ................................................................................................................................. 80
Tableau 3.7 - Comparaisons multiples sur l'interaction double souches*types de sel en conditionnant sur le type de sel ............................................................................................................................................... 81
Tableau 3.8 – Table d'ANOVA associée aux analyses statistiques sur la teneur en humidité, en cendres et en sodium des fromages ................................................................................................................. 84
Tableau 3.9 – Table d'ANOVA associée aux analyses statistiques sur la teneur en potassium des fromages ........................................................................................................................................................... 85
Tableau 3.10 – Table d'ANOVA associée aux analyses statistiques sur le ratio S/H des fromages ................ 85

x
Tableau 3.11 – Analyse de la composition minérale et de la teneur en humidité des Camemberts en fonction des conditions de saumurage et du temps d'affinage. ......................................................................... 86
Tableau 3.12 – Table d'ANOVA associée aux analyses statistiques du pH ..................................................... 88
Tableau 3.13 – Table d'ANOVA associée aux analyses statistiques sur la quantification qPCR des gènes btub et cgl ............................................................................................................................................... 89

xi
Liste des figures
Figure 1.1 - Apport moyen en sodium par catégorie d'âge chez la population canadienne (Tiré intégralement de Santé Canada, 2010) .............................................................................................................. 6
Figure 1.2 - Sources de sodium selon les groupes d'aliments transformés au Canada (Santé Canada, 2010) ................................................................................................................................................................... 7
Figure 1.3 – Influence de la teneur en NaCl sur la production d'acide lactique par des ferments lactiques du genre Lactococcus (Adapté par Ardö et coll., 2014). .................................................................... 14
Figure 1.4 – Cinétique d'incorporation du sel dans un Camembert immergé dans une saumure saturée statique (○) et dans une saumure agitée (●). Le pH des saumures a été ajusté à 4,6 à la température à 14 C (Tirée de Hardy, 2009). ........................................................................................................................... 22
Figure 1.5 – Évolution de la teneur saline au centre d'un fromage Camembert pendant l'affinage. Le symbole (●) correspond aux données prises en surface et le symbole ( ○) correspond aux données prises au centre du fromage (Tirée de Hardy, 2009). ........................................................................................ 26
Figure 1.6 - Voies métaboliques responsables de la génération de composés aromatiques pendant l'affinage des fromages (Tirée de Marilley et Casey, 2004). ............................................................................. 29
Figure 1.7 – Changements physico-chimiques pendant l'affinage du Camembert attribués au développement de P. camemberti (Adaptée de McSweeney, 2004). ............................................................... 32
Figure 1.8 – Quantification de P. camemberti ( ▲), G. candidum (□), K. lactis (●) pendant l'affinage de CMC par dénombrement sur gélose (gauche) et par qPCR (droite). Le symbole (×) correspond à la biomasse (poids sec) du mycélium total (Figure adaptée de Lessard et coll., 2012). ....................................... 37
Figure 2.1 – Teneur en sodium dosée après 2 jours (expérience 1) et 4 jours (expérience 2) d'affinage dans des Camemberts saumurés pendant 70 minutes dans une saumure expérimentale statique (26 % NaCl (p/p)) en comparaison avec une saumure agitée (saumure industrielle). Les résultats sont représentés par la moyenne ± l'erreur-type. ..................................................................................................... 53
Figure 2.2 – Incorporation du sodium dans des fromages Camemberts en fonction du temps de saumurage (vert) et proportions de réduction en sodium obtenues comparativement aux fromages salés pendant 70 minutes (violet). Les fromages ont été salés dans des saumures statiques saturées à 26 % de NaCl (p/p). Puisque les temps de saumurage 0, 30 et 70 minutes ont été expérimentés à deux reprises, les résultats représentés pour ces conditions proviennent de la moyenne obtenue pour ces deux essais. Pour tous les autres temps, les résultats proviennent de seulement un essai de saumurage. Les résultats sont représentés par la moyenne ± l'erreur type. .................................................... 55
Figure 2.3 – Teneur en sodium et en potassium des fromages Camemberts saumurés 70 minutes dans des saumures statiques fabriquées selon divers ratios de NaCl et de KCl. La saumure sans KCl (100NaCl :0KCl) correspond à saumure standard saturée à 26 % de NaCl (p/p). Le dosage des minéraux a été fait au 4e jour d'affinage. Les résultats sont représentés par la moyenne ± l'erreur type. Les pourcentages correspondent au ratio molaire des éléments sodium et potassium dans chacune des conditions, 100 % correspond au contenu total en sodium et en potassium. ............................................. 57
Figure 2.4 – Incorporation du sodium (en violet) et du potassium (en vert) dans des fromages Camemberts salés pendant 50, 60 et 70 minutes dans des saumures mixtes de ratio 4NaCl:1KCl

xii
comparativement au sodium incorporé dans des fromages salés dans une saumure statique saturée à 26 % de NaCl (p/p) pendant 70 minutes. Le dosage des minéraux a été réalisé au 4e jour d'affinage. Les résultats sont représentés par la moyenne ± l'erreur type. Les pourcentages correspondent au ratio molaire des éléments sodium et potassium dans chacune des conditions, 100 % correspond au contenu total en sodium et en potassium. ......................................................................................................... 60
Figure 3.1 – Influence du NaCl et du KCl sur la croissance de quatre souches de P. camemberti sur milieu de culture synthétique PDA acidifié (pH 3,5) après 10 jours d'incubation à 25 C. Les souches sont représentées par quatre différentes couleurs; LMA-17 (bleu), LMA-30 (orange), LMA-435 (violet), LMA-1029 (vert). Les bandes hachurées représentent la croissance en absence de sel, les bandes foncées représentent la croissance en fonction du NaCl et les bandes pâles représentent la croissance en présence de KCl. Les données sont représentées par la moyenne ± l'erreur type. L'axe des abscisses indique le % en NaCl. Les teneurs en KCl ont été calculées sur une base équimolaire avec le NaCl. ................................................................................................................................................................. 78
Figure 3.2 – Influence du NaCl et du KCl sur la croissance (diamètre des colonies) de quatre souches de P. camemberti sur caillé modèle Camembert après 10 jours d'incubation à 25 C. Les souches sont représentées par quatre différentes couleurs; LMA-17 (bleu), LMA-30 (orange), LMA-435 (violet), LMA-1029 (vert). Les bandes hachurées représentent la croissance en absence de sel, les bandes foncées représentent la croissance en fonction du NaCl et les bandes pâles représentent la croissance en présence de KCl. Les données sont représentées par la moyenne ± l'erreur type. L'axe des abscisses indique le % en NaCl. Les teneurs en KCl ont été calculées sur une base équimolaire avec le NaCl. L'interaction double (types de sel*concentrations) est représentée sur la figure. Des lettres différentes pour une même concentration signifient un effet significatif (p < 0,05). ............................................................. 81
Figure 3.3 – Évolution du pH à la surface (gauche) et au centre (droite) des fromages industriels en fonction de différentes teneurs salines. Chaque couleur représente une condition saline différente; standard (noir), réduit en NaCl (gris) et réduit en NaCl et substitué par du KCl (bleu). L'affinage a été réalisé à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. L'interaction double (conditions*jours d'échantillonnage) est représentée sur chacun des graphiques. Des lettres identiques signifient qu'il n'y a aucune différence significative entre les pH en fonction des conditions salines, pour un jour précis (p > 0,01). .................................................................... 88
Figure 3.4 – Quantification de P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel en fonction de teneurs salines variées à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. L'interaction double (conditions*jours d'échantillonnage) est représentée sur le graphique. Les lettres minuscules permettent de comparer les résultats obtenus entre les trois conditions pour chacun des jours. Les lettres majuscules comparent les résultats obtenus entre les jours d'échantillonnage pour chaque condition. Des lettres identiques indiquent qu'il n'y a aucune différence significative (p > 0,01). ...................... 89
Figure 3.5 – Quantification de G. candidum (LMA-1028) à la surface d'un fromage Camembert industriel en fonction de teneurs salines variées à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. L'interaction double (conditions*jours d'échantillonnage) est représentée sur le graphique. Les lettres minuscules

xiii
permettent de comparer les résultats obtenus entre les trois conditions pour chacun des jours. Les lettres majuscules comparent les résultats obtenus entre les jours d'échantillonnage pour chaque condition. Des lettres identiques indiquent qu'il n'y a aucune différence significative (p > 0,01). ...................... 90


xv
Liste des annexes
Annexe 1 – Quantification de G. candidum (LMA-1028) et P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel de teneur saline standard à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. ........................................................ 101
Annexe 2 – Quantification de G. candidum (LMA-1028) et P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel réduit en NaCl à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. ........................................................ 102
Annexe 3 – Quantification de G. candidum (LMA-1028) et P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel réduit en NaCl et substitué par du KCl à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. .......................... 103


xvii
Liste des abréviations et des sigles
AAC Agriculture et agroalimentaire Canada ADN Acide désoxyribonucléique AMT Apport maximal tolérable ANOVA Analyse de la variance (analysis of variance) AOC Appellation d'origine contrôlée AS Apport suffisant AVC Accidents vasculaires cérébraux aw Activité de l'eau btub -tubuline (gène) CaCl2 Chlorure de calcium cgl Cystathionine-ɣ-lyase (gène) CMC Caillé modèle de type Camembert CMP Caséinomacropeptide DL Degré de liberté ESCC Enquête sur la santé dans les collectivités canadiennes GTS Groupe de travail sur le sodium HR Humidité relative ICP-OES Spectrométrie d'émission à plasma induit (Inductively coupled plasma –
optical emission spectrometry) IOM Institute of Medicine KCl Chlorure de potassium LMA Laboratoire de mycologie alimentaire NaCl Chlorure de sodium OMS Organisation mondiale de la santé PCR Réaction en chaîne de la polymérase (polymerase chain reaction) PDA Potato dextrose agar qPCR Réaction en chaîne de polymérase quantitative ou PCR en temps réel S/H Ratio sel/humidité STD Standard TCA Acide trichloroacétique (trichloroacetic acid)


xix
Remerciements
En premier lieu, je tiens à remercier mon directeur de recherche, Monsieur Steve Labrie, afin de m'avoir fait
confiance dans le cadre de ce projet. Merci pour ton soutien, tes conseils et les encouragements apportés tout
au long de mon cheminement. Je tiens à préciser que je suis grandement reconnaissante envers la patience
dont tu as fait part pendant le long processus de rédaction de ce mémoire. Je suis très fière d'avoir fait partie
de ton équipe à l’intérieur de laquelle j'ai pu relever de grands défis et apprendre énormément! J'aimerais
également remercier Monsieur Daniel St-Gelais et Madame Sylvie Turgeon pour leur participation dans
l'évaluation de ce mémoire.
Ensuite, je remercie tous les membres de l'équipe de recherche du Laboratoire de Mycologie Alimentaire.
Merci pour vos idées et vos conseils ainsi que pour tous les bons moments partagés. Je tiens à remercier
spécialement Catherine et Marie-Hélène pour tous les conseils, votre disponibilité et votre aide si précieuse.
Merci également à Francis pour l'aide apportée en début de parcours et à Marie-Ève pour sa contribution en
laboratoire.
J'aimerais exprimer ma reconnaissance envers les partenaires financiers, soit Agriculture et agroalimentaire
Canada, les Producteurs laitiers du Canada et la Commission canadienne du lait pour les fonds apportés à ce
projet. Merci à l'industrie laitière ayant collaboré au projet et spécialement à Jimmy pour l'aide apportée en
usine.
Merci à mon amie Élisabeth pour ton support et ta présence tout au long de mes études universitaires. Merci à
mes amies de longue date; Fanny, Jessica, Josiane et Marie-Michèle, sur lesquelles j'ai toujours pu compter.
Votre amitié, votre écoute et tous ces moments précieux ont su me divertir et me soutenir durant cette période
de ma vie. Je remercie aussi ma belle-famille pour tous les encouragements apportés.
J'aimerais également remercier mes chers parents, Chantal et Laurent, ainsi que mon frère, Philippe, pour leur
soutien au quotidien, malgré la distance. Grâce à votre support, vos encouragements et à tout ce que vous
m'avez donné et transmis, j'ai pu accomplir de grands défis. Merci pour la confiance, le temps et tous les
merveilleux moments que vous m'avez accordés. Je vous attribue une partie de ma réussite.
Finalement, un merci bien spécial à mon amoureux, Jérôme, qui a su m'épauler et m'encourager tout au long
de mes études universitaires et qui a fait preuve d'une grande patience pendant les périodes les plus difficiles.
Ton positivisme, ton humour et tous les bons moments partagés au quotidien ont contribué à la réussite de
mes études.


xxi
Avant-propos
Ce mémoire de maîtrise est consacré à l’étude de la réduction du chlorure de sodium dans les fromages de
type Camembert et, plus particulièrement, de son effet au niveau de la croissance de la microflore fongique
d'affinage. Le développement de ce projet ainsi que son encadrement ont été réalisés par mon directeur de
recherche, Monsieur Steve Labrie. Ce travail est divisé en trois chapitres, le premier correspondant à une
synthèse de la littérature en lien avec le sujet du projet de recherche. Ensuite, les deuxième et troisième
chapitres correspondent à l'ensemble des travaux effectués ainsi que les résultats et conclusions qui en
découlent.
La revue de littérature se divise en quatre principales sections dont je suis l'entière rédactrice. Dans un
premier temps, elle traite du rôle du sel dans l'alimentation humaine ainsi que de son importance en
fromagerie. Ensuite, la fabrication du fromage Camembert et les techniques de salage y sont abordées.
Subséquemment, l'affinage du Camembert et l’évolution de sa microflore fongique sont traités de façon plus
précise. En dernier lieu, la fin de cette section traite de la réduction du sodium dans les fromages ainsi que
des stratégies utilisées.
Le deuxième chapitre de ce mémoire comprend une étude de la cinétique d'incorporation du sel lors du
saumurage d'un Camembert industriel et correspond au premier objectif de ce projet de recherche. Un volet
de recherche industrielle y est associé puisque les expériences de saumurage ont été entièrement réalisées
dans une industrie productrice du fromage. Il a permis de comprendre davantage les mécanismes de diffusion
du NaCl et du KCl lors du saumurage du Camembert dans des conditions réelles de production et d'identifier
les différents paramètres nécessaires à l’optimisation de la production de fromages réduits en NaCl et/ou
partiellement substitués par du KCl. J'ai été entièrement responsable de la planification des expériences
réalisées dans le cadre cet objectif ainsi que de la réalisation de celles-ci et des analyses qui en ont découlé.
Les travaux ont également été supervisés par mon directeur de recherche.
Finalement, le troisième chapitre de ce mémoire traite de l'effet de la réduction du NaCl dans le Camembert et
de son remplacement partiel par du KCl sur la croissance des ferments fongiques d'affinage, soit
Penicillium camemberti et Geotrichum candidum. Ce chapitre correspond aux deuxième et troisième objectifs
de ce projet de recherche. Les résultats ont permis d'identifier les teneurs en NaCl et KCl optimales à la
croissance de P. camemberti et de comparer l'effet de ces deux sels sur son développement. Les paramètres
optimisés lors du premier objectif ont été utilisés au troisième objectif et ont permis de reproduire des
fromages correspondant aux conditions salines ciblées et de déterminer l'effet de ces modifications sur la
croissance des ferments d’affinage d'intérêt. J'ai été responsable de la planification de ces expériences, de

xxii
leur réalisation et des analyses y étant associées. Marie-Hélène Lessard et Marie-Ève Dubuis ont contribué à
la réalisation d'une partie des expériences associées au troisième objectif de ce projet.

1
Introduction
Le chlorure de sodium (NaCl) ou, le sel, fait partie de l'alimentation de l'humain depuis des milliers d'années.
Son utilisation à titre d'agent de conservation daterait des temps préhistoriques (Guinee et Fox, 2004). Dans
l'alimentation ancestrale, la consommation de ce composé était très faible, soit de 1 à 10 mmol/jour (0,058 à
0,58 g/jour), alors qu'aujourd'hui, elle peut s'élever jusqu'à 205 mmol/jour (12 g/jour) dans certains pays
occidentaux (He et MacGregor, 2008). Au Canada, la consommation du sodium est près de deux fois
supérieure à la teneur recommandée au quotidien (Santé Canada, 2010). Cette augmentation est entre autres
due à la modernisation de l'agriculture et à l'arrivée sur le marché des aliments transformés et des mets
préparés, puisqu'ils contiennent beaucoup de sel ajouté (He et MacGregor, 2008). Or, il est connu depuis
plusieurs décennies que le sel peut engendrer des effets indésirables sur la santé humaine, lorsqu'il est
consommé en quantité excessive. L'ion sodium en est d'ailleurs majoritairement responsable (Doyle et Glass,
2010). Maintenant, il est recommandé d'éviter la consommation d'aliments riches en sodium et beaucoup
d'efforts sont déployés au niveau gouvernemental et industriel afin de réduire sa présence dans les aliments
offerts sur le marché (Webster et al., 2011).
L'initiative du Gouvernement du Canada en ce qui concerne la réduction du sodium dans l'alimentation a été
amorcée en 2007 par la création du Groupe de travail sur le sodium (GTS). Le mandat attribué à ce groupe de
multi-intervenants était d'élaborer et de surveiller la mise en place d'une stratégie visant à améliorer la santé
des Canadiens via la réduction du sodium consommé. Puisque 77 % de l'apport quotidien en sodium des
Canadiens proviennent des aliments transformés, une partie des recommandations découlant de cette
stratégie visait à encourager les industries de transformation alimentaire à réduire le contenu en sodium dans
leurs aliments (Santé Canada, 2010). Les fromages font partie intégrante de l'alimentation des Canadiens et
participent de façon non négligeable à l'apport en sodium au quotidien (Fisher et al., 2009, Santé Canada,
2010). De ce fait, la stratégie de Santé Canada encourage les fromagers à réduire le contenu en sodium dans
leurs aliments et des cibles de réduction volontaires ont été proposées pour la plupart des catégories de
fromages, incluant le fromage de type Camembert (Santé Canada, 2012).
L'inquiétude associée à l'impact causé par la réduction du sel sur la qualité globale des fromages est justifiable
puisque ce composé y joue de nombreux rôles de grande importance. Entre autres, le sel permet de contrôler
la croissance et l'activité de la microflore pathogène, d'altération et d'affinage (Guinee et Fox, 2004, Doyle et
Glass, 2010, Ardö et al., 2014, Labrie et al., 2014). Les fromages de type Camembert possèdent des
caractéristiques intrinsèques propices au développement microbien, soit leur contenu en eau élevé, la
neutralité du pH se développant pendant l’affinage et une diversité microbienne très variée (Spinnler et Gripon,
2004). Puisque les ferments fongiques d'affinage Penicillium camemberti et Geotrichum candidum contribuent

2
grandement au développement des propriétés sensorielles de cette variété pendant l'affinage (Molimard et
Spinnler, 1996, Molimard et al., 1997, Sousa, 2003, Spinnler et Gripon, 2004, Boutrou et Gueguen, 2005,
Boutrou et al., 2006), l'effet associé à la réduction du NaCl ou de son remplacement sur ces microorganismes
doit être déterminé.
À ce jour, peu d'études permettent de comprendre les effets possibles associés à la réduction du sel sur les
ferments fongiques. G. candidum est reconnue pour sa sensibilité au sel, mais son comportement en fonction
de celui-ci est décrit comme étant souche-dépendant (Sansonetti, 1930, Marcellino et al., 2001, Boutrou et
Gueguen, 2005). Pour ce qui est de P. camemberti, sa tolérance face au sel s'avère plus élevée et elle semble
nécessiter une teneur en NaCl minimale pour un développement optimal (O'Nulain, 1986). Or, les informations
actuellement disponibles dans la littérature scientifique s'avèrent insuffisantes afin d'évaluer l'effet qu'aura une
modification saline sur la croissance et l'activité de ces deux ferments d’affinage dans l'écosystème du
Camembert.
La substitution du NaCl par le KCl s'avère être l'alternative de choix afin de réduire le contenu en sodium dans
les fromages (Reddy et Marth, 1991, Hoffmann, 2014). Plusieurs études ont démontré la possibilité d'obtenir
des fromages de qualité acceptable en substituant entre 25 et 50 % du NaCl par du KCl (Fitzgerald et Buckley,
1985, Aly, 1995, Katsiari et al., 1997, 1998, Gomes et al., 2011, Grummer et al., 2012, Thibaudeau et al.,
2015). Cependant, l'impact engendré par ces substitutions sur la qualité globale du fromage Camembert est
peu documenté. De plus, le comportement de la microflore fongique d'affinage en présence de KCl n'est pas
connu.
Ce projet de recherche avait pour but d'évaluer l'effet d'une réduction du NaCl dans le Camembert et d'une
substitution par du KCl sur la croissance des ferments d'affinage P. camemberti et G. candidum. Le premier
objectif de ce projet de maîtrise visait à étudier la cinétique d'incorporation du sodium et du potassium dans un
Camembert industriel saumuré afin d'identifier les paramètres de saumurage permettant d'obtenir des
fromages réduits en NaCl et partiellement substitués avec du KCl. Ensuite, le deuxième objectif consistait à
évaluer la variabilité intraspécifique de la résistance au NaCl et au KCl chez quatre isolats de P. camemberti.
Les paramètres de saumurage identifiés ainsi que la compréhension du comportement des microorganismes
ont par la suite servi à la réalisation du troisième objectif qui consistait à évaluer l'effet de ces modifications
salines sur la croissance de P. camemberti et G. candidum à divers temps clés pendant l'affinage.

3
1. Revue de littérature


5
1.1. LE CHLORURE DE SODIUM DANS L'ALIMENTATION ET SON IMPORTANCE EN FROMAGERIE
1.1.1. La consommation du sodium chez les Canadiens
1.1.1.1. L'apport en sodium
Une consommation excessive en sodium chez l'humain est observée au niveau planétaire (Brown et al.,
2009). De ce fait, l'Organisation mondiale de la santé (OMS) propose que la consommation de cet élément ne
dépasse pas 2 g Na/jour (ou 5 g de NaCl) (OMS, 2013a). Au Canada, la quantité de sodium consommée est
trop élevée chez tous les groupes d'âge (Figure 1.1). Selon l'Enquête sur la santé dans les collectivités
canadiennes (ESCC) réalisée en 2004 (cycle 2,2) et en prenant en considération le sel ajouté par les
consommateurs, l'apport moyen en sodium des Canadiens serait d'environ 3 400 mg/jour (Fisher et al., 2009,
Santé Canada, 2010). Selon le guide de référence des apports nutritionnels de l'Institute of Medicine (IOM),
l'apport suffisant (AS) en sodium, correspondant à l'apport nécessaire pour satisfaire et excéder les besoins
de personnes en bonne santé, se situe à 1 500 mg/jour pour les gens âgés de 9 à 50 ans et d'une
concentration moindre pour les autres groupes d'âge (Figure 1.1) (IOM, 2005). Les Canadiens consomment
donc plus du double de la quantité de sodium dont ils ont réellement besoin. De plus, l'apport maximal
tolérable (AMT) établit par l’IOM est dépassé par la majorité des individus de chaque groupe d’âge, celui-ci
fixé à 2 300 mg de sodium/jour pour la population âgée de 14 ans et plus (Figure 1.1) (IOM, 2005). L'AMT
représente la teneur maximale en sodium pouvant être consommée sans causer d'effets indésirables sur la
santé. La majorité de la population canadienne, en dépassant cette limite supérieure, est donc
particulièrement à risque de souffrir de problèmes de santé reliés à la surconsommation du sodium. D'ailleurs,
environ 19 % des Canadiens âgés entre 20 et 79 ans seraient hypertensifs (Santé Canada, 2012).

6
Figure 1.1 - Apport moyen en sodium par catégorie d'âge chez la population canadienne (Tiré intégralement de Santé Canada, 2010)
1.1.1.2. Les sources de sodium
Le sodium est omniprésent dans l'environnement (Hardy, 2004). Il n'est donc pas étonnant de le retrouver
naturellement dans les aliments. D'ailleurs, 12 % de l'apport quotidien en sodium des Canadiens provient de la
présence naturelle de cet élément dans les aliments. Ainsi, par corollaire, 88 % du sodium consommé
quotidiennement est ajouté de façon volontaire, en majeure partie sous forme de NaCl, pendant la
transformation, la préparation ou lors de la consommation des aliments. Plus précisément, 77 % de l’apport
quotidien provient des aliments transformés, 5 % est ajouté pendant la préparation et 6 % au moment du
repas (Santé Canada, 2010). Parmi les aliments transformés, les données de l'ESCC réalisée en 2004
confirment que les principaux groupes responsables de cette contribution élevée sont les pains et ses dérivés
(14 %), les viandes transformées et les plats à base de légumes (9 % chacun) ainsi que les soupes (7 %)
(Figure 1.2). Selon cette étude, les fromages se retrouvent au 6e rang et représentent 5 % de l'apport
quotidien en sodium des Canadiens (Fisher et al., 2009, Santé Canada, 2010).
Le NaCl ne constitue pas le seul composé contribuant à l'apport alimentaire en sodium. Plusieurs autres
additifs ou ingrédients alimentaires contenant du sodium contribuent à son apport au quotidien (Doyle et
Glass, 2010). Chez les Canadiens, il s'agit principalement du glutamate monosodique, du benzoate de
sodium, du citrate sodium, du bicarbonate de sodium, du nitrite de sodium et du pyrophosphate disodique
(Santé Canada, 2010). Cependant, les ingrédients et les additifs alimentaires contribuent moindrement à
l'apport en sodium, le NaCl y demeure la principale source (Doyle et Glass, 2010).

7
Figure 1.2 - Sources de sodium selon les groupes d'aliments transformés au Canada (Santé Canada, 2010)
1.1.2. L'effet du sodium sur la santé humaine
Une fois consommé, la majorité du sodium est absorbée via l'intestin tandis que l'excès est éliminé par les
reins ou la transpiration (Doyle et Glass, 2010). Dans l'organisme, une quantité importante du sodium se loge
au niveau des fluides extracellulaires (3,3 g/L) (Hardy, 2004). Cet élément s'avère indispensable au bon
fonctionnement du corps humain. Plusieurs rôles essentiels sont réalisés par le sodium dans l'organisme tels
que la transmission de l'influx nerveux, la régulation du volume plasmatique et des fluides extracellulaires, la
détermination du potentiel membranaire des cellules et le transport intermembranaire de certaines molécules.
Il joue également un rôle dans certaines voies métaboliques, notamment au niveau de la dégradation des
hydrates de carbone et des protéines. Un manque de cet élément chez l'humain peut donc lui être fatal
(Hardy, 2004, Doyle et Glass, 2010, Cruz et al., 2011).
Bien qu'il soit fort utile dans le secteur alimentaire et indispensable au fonctionnement du corps humain, le
sodium s'avère néfaste pour la santé lorsqu'il est consommé de façon excessive. Le lien entre la
consommation de sodium et une tension artérielle élevée a été démontré depuis plusieurs années (Elliott et
al., 1988, Poulter et al., 1990). Malgré tout, aujourd'hui, le mécanisme par lequel le sodium affecte la tension

8
artérielle est encore mal compris. Certaines personnes seraient plus sensibles au sel tandis que d'autres y
seraient plus résistants. L'excrétion du sodium par l'urine après une diète riche en sodium ne s’effectuerait pas
aussi bien chez les personnes sensibles comparativement aux autres. Ainsi, une teneur importante en sodium
est retrouvée au niveau du plasma sanguin, ce qui provoque une expansion de celui-ci et une élévation du
rythme cardiaque (Doyle et Glass, 2010). Le sodium est ainsi associé au développement de l'hypertension.
L'hypertension représenterait un facteur de risque pour le développement de plusieurs maladies chroniques
dont les accidents vasculaires cérébraux (AVC) et les maladies cardiovasculaires (Strazzullo et al., 2009,
Whelton et He, 2014). D'ailleurs, selon Statistiques Canada, les maladies du cœur et les ACV représentaient
les deuxième et troisième causes de mortalité au Canada en 2011 (Statistique Canada, 2014). D’autre part,
les enfants sont particulièrement affectés lors d'une exposition fréquente aux aliments riches en sodium. En
effet, plus la consommation de sel est importante, plus les récepteurs du goût salé s'affaibliraient ce qui
diminue sa perception (He et MacGregor, 2006). La consommation d'aliments salés dès le jeune âge les
prédisposerait donc à souffrir d'hypertension en vieillissant puisqu'ils auront tendance à consommer ce type
d'aliment. Par ailleurs, des associations entre l'excès de sodium et d'autres problèmes de santé ont été
proposées. Il a été observé que l'excrétion du calcium et du sodium par le corps humain était plus élevée lors
d'une diète riche en sodium, ce qui favoriserait la déminéralisation des os et le développement d'ostéoporose
(Cappuccio et al., 2000). Enfin, des associations entre l'excès de sodium et le cancer gastrique, l'obésité, la
formation de pierres aux reins et l'asthme ont également été établis (He et al., 2008, Doyle et Glass, 2010).
1.1.3. La réduction du sodium dans l'alimentation
Puisque la majorité du sodium ajouté aux aliments provient du NaCl, réduire ce composé dans les aliments
transformés s'avère être la méthode la plus efficace afin de minimiser sa consommation. De plus, il a été
démontré qu'une réduction modérée de la quantité de sel consommée pendant une période d'au moins un
mois contribuait à abaisser la tension artérielle de personnes en bonne santé et d'individus souffrant
d'hypertension. Conséquemment, les risques de développer des maladies du cœur seront également réduits
(He et al., 2013). Aussi, il a été estimé qu'une réduction de l'apport en sodium d’une quantité de 1 840 mg
Na/jour ( 53 % de réduction) chez les Canadiens permettrait de diminuer la prévalence de l'hypertension de
30 % et de réaliser des économies allant jusqu'à 430 millions de dollars par année dans le système de santé
(Joffres et al., 2007).
Bien que plusieurs évidences aient démontré l'intérêt de réduire le sodium dans la consommation humaine, il
existe une contradiction dans la littérature. Une étude a, entre autres, démontré l'effet du sodium sur
l'hypertension et la pression sanguine via l'analyse de son taux d'excrétion dans l'urine (Stolarz-Skrzypek et
al., 2011). Les résultats observés n'ont pas confirmé l'association normalement connue entre l'excrétion du

9
sodium et les risques d'hypertension ou de maladies cardiovasculaires. Dans une seconde étude, les auteurs
mentionnent que les éléments permettant de relier le taux de morbidité associé aux maladies
cardiovasculaires et le taux de mortalité de toutes causes avec une diète pauvre en sodium seraient
insuffisants (Taylor et al., 2011). Ces études ont cependant été critiquées à plusieurs niveaux, notamment en
ce qui a trait à l'échantillonnage, la collecte et l'interprétation des données ainsi que le pouvoir statistique
(Campbell et al., 2011a, Campbell et al., 2011b). Il n’en demeure pas moins que le débat sur l’importance de
la consommation de sodium et de ses effets sur la santé demeure un sujet de recherche d’actualité.
Aujourd'hui, plusieurs pays ont entrepris leur propre plan de réduction du sodium dans l’alimentation pour leur
population respective. D'ailleurs, l'OMS a publié un document résumant les différentes initiatives sur la
réduction de la consommation du sodium au niveau de plusieurs pays européens (OMS, 2013b). À titre
d'exemple, la Finlande a pu abaisser l'apport moyen en sodium de sa population d'environ 40 % sur une
période de 30 ans. La stratégie du Canada est étroitement basée sur celle adoptée par le Royaume-Uni. La
stratégie de ce dernier comporte trois volets; la sensibilisation de la population sur les risques reliés au
sodium, l'étiquetage des emballages concernant les teneurs en sel (système de «feux de circulation») et la
demande aux entreprises alimentaires de réduire le sodium dans plusieurs aliments transformés en se basant
sur des cibles de réduction préétablies (Santé Canada, 2010).
1.1.3.1. La stratégie de réduction du sodium au Canada
En 2007, le gouvernement canadien a mis sur pied le Groupe de travail sur le sodium (GTS). Supervisé par
Santé Canada, le GTS a été formé afin de créer et d’émettre des recommandations et de proposer une
stratégie de réduction du sodium dans l'alimentation des Canadiens. Le but final est d'améliorer la santé de la
population en diminuant la prévalence des maladies chroniques associées à la surconsommation de sodium.
La Stratégie de réduction du sodium pour le Canada, publiée en 2010, comprend diverses recommandations
réparties en quatre volets; l'approvisionnement alimentaire, la recherche, la sensibilisation et l'éducation ainsi
que la surveillance des progrès accomplis. L'objectif ultime de réduction du sodium fixé par le GTS est que,
chez plus de 95 % de la population, l'apport moyen en sodium se situe entre l'AS et l'AMT. Cependant, un
objectif intérimaire proposé pour 2016 a été établi par Santé Canada où l’apport quotidien ciblé est de 2 300
mg Na/jour (Santé Canada, 2010).
Le volet de l'approvisionnement alimentaire touche principalement les aliments transformés et les mets
préparés qui sont vendus dans divers établissements. Santé Canada encourage les transformateurs, les
détaillants et les restaurateurs à réduire volontairement la quantité de sodium dans leurs aliments (Santé
Canada, 2010). Afin de les guider dans leur démarche, Santé Canada a publié un Document d'orientation
destiné à l'industrie alimentaire sur la réduction du sodium dans les aliments transformés en juin 2012. Ce

10
document contient des cibles graduelles de réduction du sodium par catégories d'aliments transformés. Si la
cible finale est atteinte, elle représenterait une réduction de 25 à 30 % du sodium selon la catégorie
alimentaire en question (Santé Canada, 2012).
La réduction du sodium dans l'alimentation est une démarche multifacette nécessitant la collaboration de toute
la population, en surplus de la contribution industrielle. Ainsi, un volet de la stratégie a été consacré
entièrement à la sensibilisation et à l'éducation de la population au moyen de publicités et d'outils éducatifs. Le
but est d'encourager les consommateurs à s'alimenter plus sainement, à faciliter leurs choix alimentaires et à
comprendre les répercussions sur la santé associées à une surconsommation du sodium. La démarche de
réduction du sodium doit également tenir compte de la qualité nutritionnelle, sensorielle et de l'innocuité du
produit final. Finalement, Santé Canada s'engage à évaluer les progrès accomplis au niveau des
comportements alimentaires des consommateurs et de l'atteinte des cibles de réduction. D'ailleurs, l'ESCC
prévu en 2015 permettra d'évaluer s'il y a eu des changements au niveau des habitudes alimentaires chez la
population canadienne depuis 2004 (CTAC, 2009, Santé Canada, 2010, 2012).
Aujourd'hui, plusieurs fabricants et détaillants ont amorcé des efforts de réduction du sodium dans leurs
aliments. Cependant, la formulation de tels aliments s'avère être une épreuve difficilement réalisable pour
plusieurs catégories de produits. Les enjeux les plus communs concernent l'aspect réglementaire, le temps
nécessaire à l’établissement de nouvelles formulations, les coûts associés aux projets de développement ainsi
que les défis d'ordre technique aboutissant à des changements au niveau de la qualité globale des produits
finis et de leur durée de vie (Agriculture et Agroalimentaire Canada, 2013). Les impacts de la réduction de
sodium sont variables d'un aliment à l'autre et les défis augmentent avec la complexité de l’aliment en lui-
même. Les fromages en sont un bon exemple puisque le sel y joue plusieurs rôles parmi lesquels on retrouve
le contrôle des activités microbiennes lors de l'affinage, ce qui a un impact sur la texture et le développement
du profil aromatique (Guinee et Fox, 2004).
1.1.4. Les rôles du sel en fromagerie
Le sel en fromagerie est indispensable puisqu'il joue plusieurs rôles majeurs lors de la confection des
fromages. Il complète l'égouttage, il réduit l'activité de l'eau et il contribue au développement des propriétés
sensorielles de plusieurs façons (Gillis, 2004, Guinee et Fox, 2004, Guinee et O'Kennedy, 2007, Hardy, 2009).
La quantité de sel dans les fromages est grandement variable, mais très spécifique à une variété de fromage
en particulier. À titre d'exemple, elle peut s'étendre entre 0,6 % dans un fromage Cottage à 4,5 % dans un
fromage Bleu (Tableau 1.1). Ces différences sont attribuables à une combinaison de facteurs, variables selon
les régions, telles que le type de lait utilisé, les méthodes de fabrication et de salage et les paramètres
d'affinage. Ces facteurs amènent également une variation au niveau de la composition globale de l'aliment ce

11
qui permet d'observer une si grande diversité fromagère (Guinee et al., 2014a). Comme le tableau 1.1 le
démontre, la variation du contenu en sel est également visible au sein d'une même catégorie fromagère. Il
existe plusieurs divergences au niveau des recettes utilisées par les différentes fromageries lors de la
fabrication d'un type de fromage en particulier. Parmi celles-ci, les fromagers peuvent choisir les types de lait,
de ferments et de présure utilisés ainsi que la quantité de cette dernière. De plus, les pH au moment de
l'emprésurage, du découpage et du soutirage peuvent varier. D'autres facteurs non-intentionnels doivent aussi
être considérés dont la composition du lait, celle-ci variable en fonction des saisons. Au bout du compte, ces
éléments influenceront l'affinage du fromage et sa composition, dont la teneur en humidité et en sel.
Néanmoins, le contenu en sel, pour une catégorie fromagère en particulier, se doit d'être contrôlé et respecté
afin de générer un produit de qualité dont les caractéristiques sont constantes d'un lot de production à l'autre.
Cela devient possible grâce à l'utilisation de méthodes de fabrication standardisées, contrôlées et validées
(Guinee et al., 2014b).
Tableau 1.1 – Teneur en NaCl des fromages selon différentes variétés (Adapté de Sutherland, 2002 et Guinee et Fox, 2004)
Variétés fromagères Teneur en NaCl (g/100g) Cottage 0,6 – 1,0 Camembert 1,5 – 2,5 Emmental 0,7 – 1,0 Bleu 1,7 – 4,5 Edam/Gouda 1,2 – 2,0 Cheddar 1,5 – 1,9 Feta 2,8 – 4,0 Parmesan 2,0 – 2,6
1.1.4.1. Le complément d'égouttage
Lors du salage, l'absorption du sel dans la matrice fromagère engendre une migration des molécules d'eau
vers la surface afin d'équilibrer la pression osmotique entre la phase aqueuse de l'aliment et le milieu salé.
Une perte d'eau est alors observée et permet d'assurer un complément à l'égouttage du fromage. À titre
d'exemple, dans un fromage saumuré, il existe une relation entre la teneur en eau perdue et le gain de sel. De
façon générale, la perte d'eau est de 2 à 2,5 fois plus grande que la quantité de sel absorbée (p/p) et le poids
du fromage après saumurage représente 96 à 98 % de son poids original (Geurts et al., 1974, Hardy, 1985,
Guinee et Fox, 2004, Mietton et al., 2004, Everett et al., 2014, Guinee et al., 2014a).

12
1.1.4.2. La réduction de l'activité de l'eau
Dans les aliments, l'eau interagit avec les différents constituants, soit les protéines, les lipides, les glucides et
les sels minéraux ce qui fait en sorte qu'une certaine partie de cette eau y est liée, tandis qu'une autre partie
est libre et peut être utilisée comme solvant. Cette dernière représente l'activité de l'eau (aw), c'est-à-dire, l'eau
disponible permettant la croissance microbienne et la réalisation de certaines réactions enzymatiques
nécessitant un milieu aqueux (Hardy, 2009). Théoriquement, l'aw d'une solution est défini par la loi de Raoult
selon la relation suivante :
PPo x aw
où : P = pression de vapeur de la solution ou de l'aliment Po = pression de vapeur de l'eau pure à la même température x = fraction molaire du solvant γ = coefficient d'activité de l'eau dans la solution
Dans les fromages, l'aw dépend de la teneur en humidité et de la concentration en molécules de faibles poids
moléculaires, telles que les petits peptides, les acides aminés, les acides et le phosphate de calcium présents
dans la phase aqueuse. Or, le sel s'avère être le meilleur contributeur à l'abaissement de l'aw des fromages.
Cette capacité s'explique par le fait que les sels, tels que le NaCl et le KCl, possèdent une meilleure solubilité
que de plus grosses molécules comme les lipides ou les protéines. L'aw des fromages est cependant une
valeur non constante qui évolue au cours de la maturation fromagère due à la dégradation des composés
organiques qui contribueront à leur tour à la réduction de ce paramètre (Hardy, 1979, Schlesser et al., 1992,
Guinee et Fox, 2004, Guinee et O'Kennedy, 2007, Hardy, 2009). Cette évolution s'explique également par les
changements de contenu en eau observés pendant l'affinage dus à l'évaporation superficielle, ce qui a
d'ailleurs été observé dans le fromage Camembert (Schlesser et al., 1992).
1.1.4.2.1. L'aw et le développement microbien
Dans les aliments, l'effet préservateur du sel est en grande partie assuré par sa capacité à réduire l'aw. En fait,
chaque microorganisme possède une valeur d'aw minimale où la croissance leur est possible (Tableau 1.2).
Généralement, les bactéries nécessitent une teneur en eau libre plus élevée tandis que les levures et les
moisissures tolèrent des environnements où la disponibilité en eau est plus faible. La réduction de l'aw dans
les aliments a plusieurs conséquences au niveau du développement microbien. Entre autres, la phase de
latence des microorganismes est allongée et une réduction de la vitesse de croissance est alors observée

13
(Gillis, 2004, Hardy, 2009). Cependant, comme le démontre le tableau 1.3, l'aw typiquement retrouvé dans
plusieurs variétés fromagères n'est pas suffisante pour empêcher la croissance des microorganismes. En fait,
le développement microbien est contrôlé par une combinaison de divers paramètres dont la température, la
disponibilité en oxygène, le pH et l'ajout d'additifs alimentaires qui, une fois combinés, exerceront un effet
synergique (Doyle et Glass, 2010). Ainsi, les ferments, les microflores d’affinage ainsi que la microflore
naturelle des fromages sont en mesure de se développer et d’être actifs pendant la durée de vie du produit.
Cependant, chaque espèce réagit différemment, certaines capables de supporter les conditions présentes
dans le fromage alors que d’autres en sont incapables.
Tableau 1.2 – Activité de l'eau minimale pour la croissance de différents microorganismes (Adapté de Gillis, 2004; Guinee et Fox, 2004 et Guinee et O'Kennedy, 2007).
Espèces microbiennes aw minimale typique Streptococcus thermophilus 0,98 Lactococcus lactis 0,96 Lactobacillus helveticus 0,96 Geotrichum candidum 0,95 – 0,97 Escherichia coli 0,95 Salmonella sp. 0,94 Listeria monocytogenes 0,92 Staphylococcus aureus 0,86 Penicillium camemberti 0,86 Penicillium roqueforti 0,75
Tableau 1.3 – Activité de l'eau de différentes variétés fromagères (Adapté de Guinee et Fox, 2004 et Guinee et O'Kennedy, 2007).
Fromages aw typique Ricotta 1,00 Quark 0,99 Cottage 0,99 Camembert 0,98 Emmental 0,97 Edam 0,96 Cheddar 0,95 Gouda 0,95 Parmesan 0,92 Bleu 0,91 – 0,95
Il existe différents mécanismes qui peuvent expliquer l'effet préservateur du sel. Par exemple, il y joue un rôle
important en augmentant la pression osmotique dans la phase aqueuse de l'aliment. En réponse à un ajout de
sel important dans leur environnement, les cellules microbiennes expulsent l'eau de leur cytoplasme et se
retrouvent alors en état de plasmolyse. Cette déshydratation cellulaire aura pour effet d'inhiber leur croissance

14
ou de tuer les cellules à forte concentration en sel (Guinee et Fox, 2004, Doyle et Glass, 2010). L'effet
préservateur du sel peut aussi s'expliquer par sa capacité à diminuer la solubilité de l'oxygène en milieu
aqueux, par une augmentation de la sensibilité au CO2 ainsi que par la toxicité de l'ion chlorure, en trop
grande quantité, sur la cellule microbienne (Hardy, 2004).
Les teneurs salines présentes dans les fromages influencent le comportement de leur population microbienne.
La microflore bactérienne primaire des fromages, soit les bactéries lactiques du genre Lactococcus, est
sensible aux conditions salines ce qui a une incidence au niveau de l'utilisation du lactose et de la production
d'acide lactique à partir de l’étape de salage. À titre d'exemple et comme le démontre la figure 1.3, la
production d'acide lactique par certaines souches de lactocoques semble optimale autour de 1 % de NaCl et
fortement réduite à 2,5 % (Irvine et Price, 1961). Au contraire, une concentration en sel trop faible favoriserait
leur développement et leur activité, ce qui peut causer l'apparition d'amertume dans les fromages (Guinee et
O'Kennedy, 2007). Les bactéries lactiques de la microflore secondaire sont généralement plus tolérantes aux
conditions salines. Pour plusieurs souches de Lactobacillus casei, L. curvatus et L. plantarum, la tolérance au
NaCl a été démontrée à 4, 6 et 8 % sur milieu gélosé et même jusqu'à 10 % pour certaines de ces souches
(Jordan et Cogan, 1993). Dans les fromages, ces bactéries interviennent au niveau de la fermentation du
lactose résiduel. Elles contribuent également à l'affinage par leur action au niveau du catabolisme des
protéines, des lipides et des acides aminés (McSweeney, 2004).
Figure 1.3 – Influence de la teneur en NaCl sur la production d'acide lactique par des ferments lactiques du genre Lactococcus (Adapté par Ardö et coll., 2014).

15
La teneur en sel des fromages influence également la croissance de la microflore fongique d'affinage. En fait,
l'ajout de sel permet la sélection d'une microflore spécifique, celle-ci composée de microorganismes utiles à la
maturation fromagère, afin d'éviter l'implantation de certains microorganismes indésirables (Gillis, 2004). Dans
le fromage bleu, les fortes teneurs en sel permettent de sélectionner la croissance de Penicillium roqueforti,
celle-ci étant relativement tolérante aux concentrations salines élevées, et de limiter la croissance de
microorganismes d'altération (Johnson et al., 2009). La germination des spores de cette espèce est stimulée
entre 1 et 3 % de NaCl tandis que des concentrations supérieures sont inhibitrices (Cantor et al., 2004). Étant
donné l'importance de ce sujet dans le cadre de ce mémoire, la tolérance au sel chez G. candidum et
P. camemberti est traitée de façon distincte à la section 1.3.5.
La présence plus ou moins importante de sel influence également la croissance ou non des bactéries
pathogènes. Cependant, le sel ne peut servir à lui seul de barrière à la croissance de ces microorganismes
puisque plusieurs espèces possèdent une tolérance élevée face au contenu en NaCl (Doyle et Glass, 2010). À
titre d'exemple, S. aureus peut croître jusqu'à 20 % de NaCl. Salmonella sp. et E. coli O157:H7 sont des
espèces moindrement tolérantes, mais leur croissance est possible jusqu'à 6 % et 6,5 %, respectivement.
L. monocytogenes possède aussi une grande tolérance au sel, soit jusqu'à 10 % (Labrie et al., 2014).
D'ailleurs, une étude a démontré que le contenu en sel dans un fromage Cheddar constituait une barrière
mineure à la croissance de cette espèce puisqu'elle s'est avérée similaire en présence de 0,7 % ou 1,8 %
(standard) de NaCl (Shrestha et al., 2011).
1.1.4.2.2. L'aw et les réactions enzymatiques
Les enzymes sont sensibles aux conditions environnementales dans lesquelles elles sont présentes, telles la
température et le pH. De plus, certaines enzymes nécessitent un milieu aqueux afin de pouvoir réaliser leur
activité. Ainsi, par son action au niveau de la réduction de l'aw, le sel contribue à la modulation de l'activité de
plusieurs enzymes. Ces réactions peuvent ainsi être retardées ou inhibées (Ardö et al., 2014). Dans les
fromages, les réactions enzymatiques sont nombreuses et variées; elles contribuent à la formation du caillé
ainsi qu'au développement des propriétés organoleptiques pendant l'affinage grâce au catabolisme des
sucres, des lipides et des protéines (Figure 1.6) (McSweeney, 2004). L'effet du sel est visible au niveau de
l'activité des enzymes natives du lait, des enzymes coagulantes et des enzymes microbiennes (Ardö et al.,
2014).
La plasmine est une enzyme naturellement retrouvée dans le lait. Elle résiste à la pasteurisation et présente
une activité optimale sur les caséines à pH neutre. Peu d'études ont porté sur l'effet du NaCl sur l'activité de la
plasmine. Elle présenterait une activité optimale à 2 % de NaCl et serait inhibée à partir de 4 %. Une légère
activité serait toujours visible à 8 % de NaCl (Guinee et Fox, 2004, Guinee et O'Kennedy, 2007). Dans des

16
fromages Cheddar, la concentration en plasmine active en début d'affinage s'est révélée légèrement plus
élevée à 0,8 % de NaCl comparativement aux fromages contenant entre 1,3 et 2,4 % de NaCl tandis que
l'effet contraire a été observé plus tard durant la maturation. Cependant, ces observations seraient plutôt
associées aux différences de température de cuisson et de pH qu'à la teneur en sel (Møller et al., 2012).
L'activité enzymatique de la présure, essentiellement la chymosine, a été démontrée pendant l'affinage.
Jusqu'à 30 % de la quantité ajoutée au lait peut demeurer active pendant cette période, en fonction du
procédé de fabrication (McSweeney, 2004). La chymosine permet la dégradation des caséines en peptides,
ceux-ci servant de précurseurs pour d'autres réactions enzymatiques (Guinee et O’Kennedy, 2007). En
solution, l'hydrolyse des caséines-s1 par les enzymes coagulantes (chymosine, pepsine, présure d'origine
fongique) serait stimulée par une augmentation du taux de NaCl et serait optimale à 6 % (Guinee et Fox,
2004). Cependant, l'activité de la chymosine sur cette fraction protéique s'est avérée inversement
proportionnelle à la teneur en sel dans un fromage Cheddar (Møller et al., 2012). Au contraire, d'autres ont
observé un retard dans la dégradation de la caséine-s1 dans des fromages pauvres en sel (Guinee et Fox,
2004). La teneur en sel influence également l'hydrolyse de la caséine- par la chymosine. Son activité sur
cette protéine est optimale en absence de sel et inhibée à partir de 5 %. En fait, la présence de sel induirait
des changements de conformation de la caséine- affectant l'activité de la chymosine sur celle-ci (Guinee et
Fox, 2004, Hardy, 2004).
Dans les fromages, l'effet du NaCl sur l'activité enzymatique d'origine microbienne est peu connu (Guinee et
Fox, 2004, Ardö et al., 2014). La plupart des études évoquent des liens indirects entre le contenu en sel et
l'activité enzymatique microbienne en se basant, par exemple, sur le degré de protéolyse ou d'amertume.
Entre autres, la protéolyse dans un Camembert s'est avérée fortement avantagée à de faibles conditions en
NaCl (O'Nulain, 1986) ce qui peut être attribué au métabolisme de P. camemberti. .
1.1.4.3. Le développement des propriétés sensorielles
Le sel affecte le goût, la production de composés aromatiques et la texture des fromages de plusieurs façons.
De manière directe, il contribue au goût par sa saveur salée. En fait, dans la molécule de NaCl, la perception
du goût salé est due à l'ion sodium. Pour être perceptible, la molécule de NaCl doit être solubilisée afin de
permettre au sodium d'interagir avec les récepteurs gustatifs (Doyle et Glass, 2010). Le goût salé des
fromages dépend donc de leur rapport sel/humidité (S/H). De plus, le NaCl a la capacité de rehausser le goût
umami et de masquer l'amertume (Gillis, 2004).
Comme mentionné précédemment, le sel affecte la croissance des microorganismes et l'activité enzymatique
dans les fromages. Généralement, la réduction du sel dans les fromages est associée à une augmentation de

17
la protéolyse (O'Nulain, 1986, Guinee et Fox, 2004). Conséquemment, la fermeté de la pâte fromagère sera
moins importante et la perception de saveurs non désirées, telles l'acidité et l'amertume, augmentera. Outre le
lien entre le NaCl et la protéolyse, ce composé affecte la texture des fromages par d'autres moyens. Par
exemple, il contribue à augmenter l'hydratation des caséines. Ce phénomène est attribué à l'échange entre le
calcium des micelles de caséines et le sodium (Guinee et Fox, 2004, Guinee et O'Kennedy, 2007). Ce calcium
se retrouve alors solubilisé ce qui cause la dissociation des protéines et une modification du pouvoir tampon
du fromage (Sousa, 2003, Gillis, 2004). Lors d'une augmentation de l'hydratation des caséines, un
changement au niveau de la texture serait aussi observé, soit une augmentation de la viscosité et une perte
d'élasticité (Guinee et Fox, 2004). Enfin, dans plusieurs variétés de fromage, le sel est majoritairement présent
à la surface des fromages, à la suite du salage en saumure ou à sec, et contribue à la formation de la croûte
(Mietton et al., 2004). Cette croûte assure une protection au fromage puisqu'elle y constitue une barrière
physique et contribue à l'apparence de l'aliment (Gillis, 2004).

18
1.2. LE FROMAGE CAMEMBERT
Le Camembert est originaire de la région de Normandie, en France. Le célèbre fromage français
« Camembert de Normandie » possède d'ailleurs une appellation d'origine contrôlée (AOC) signifiant que le
lait utilisé ainsi que la fabrication, l'affinage et le conditionnement de l'aliment doivent être réalisés dans une
région géographique précise et selon un cahier de charges défini (INAO, 2013). L'appellation « Camembert »
n'est cependant pas protégée, ce qui fait en sorte que cette variété de fromage est produite partout dans le
monde à partir de lait cru, thermisé ou pasteurisé (Spinnler et Gripon, 2004).
Le Camembert est un fromage à pâte molle, affiné en surface et possédant une croûte fleurie. La moisissure
Penicillium camemberti et/ou la levure Geotrichum candidum colonisent majoritairement la surface du fromage
lui conférant son apparence spécifique (Leclercq-Perlat, 2011). Il se distingue des autres types de fromage par
la présence de son arôme typique de champignon, son onctuosité et son feutrage blanc, celui-ci étant en
majeure partie dû à la présence dominante de P. camemberti (Spinnler et Gripon, 2004, Leclercq-Perlat,
2011). Il est généralement composé de 18 à 30 % de matière grasse, de 50 à 55 % d'humidité et de 17 à 21 %
de protéines (Sutherland, 2002). L'aw de ce fromage est élevée, soit environ 0,98 ce qui fait en sorte que la
durée de vie est relativement courte (Shaw, 1981, Sutherland, 2002), s'étalant jusqu'à 6 semaines pour un
Camembert traditionnel (Poças et Pintado, 2009).
1.2.1. La fabrication des fromages de type Camembert
La transformation fromagère repose sur un processus de concentration sélective de certaines composantes
du lait et est basée sur quatre principales étapes; la coagulation, l'égouttage, le salage et l'affinage (Leclercq-
Perlat, 2011). Dans certains cas, la coagulation est précédée d'une étape de préparation du lait, c'est-à-dire, la
standardisation du contenu en protéine et en matière grasse, le traitement thermique s'il y a lieu, et une étape
de maturation (Mietton et al., 2004). Le fromage Camembert peut être confectionné selon un procédé de
fabrication traditionnel ou industriel, ce dernier étant plus mécanisé afin d'en augmenter la productivité. En
général, les Camemberts traditionnels sont fabriqués à partir de lait cru et peuvent être moulés à la louche,
tandis que la technologie industrielle utilise du lait cru, thermisé ou pasteurisé et emploie des machines pour
mouler les meules (Shaw, 1981, Spinnler et Gripon, 2004).
La coagulation est un processus physico-chimique qui consiste à déstabiliser les micelles de caséine du lait
afin de provoquer leur agglomération. Cette agglomération aboutit à la formation d'un réseau protéique,
appelé gel, où une partie de la matière grasse, des sels minéraux, des protéines solubles, de l'eau et du
lactose y sont retenus. Dans le cas du Camembert, ce changement d'état physique est de type mixte puisqu'il
est provoqué par l'acidification du lait par les bactéries lactiques et par réaction enzymatique de la chymosine
(Shaw, 1981, St-Gelais et Tirard-Collet, 2002). L'acidification du lait est possible grâce au métabolisme des

19
bactéries lactiques, les plus souvent utilisées étant Lactococcus lactis ssp. lactis et L. lactis ssp. cremoris
(Sousa, 2003, Spinnler et Gripon, 2004). Ces bactéries mésophiles et homofermentaires métabolisent le
lactose en acide lactique entraînant ainsi une baisse du pH. L'acidification engendre une migration du calcium
et du phosphate colloïdal de la micelle vers la phase aqueuse ainsi qu'une réduction du nombre de charges
négatives associées aux micelles de caséines. Conséquemment, les caséines s'aggloméreront. La
coagulation par voie enzymatique est réalisée par l'ajout d'une protéase coagulante, la chymosine, qui initie
l'hydrolyse de la caséine- en caséinomacropeptide (CMP) et en paracaséine-. La paracaséine-,
hydrophobe, reste liée aux micelles tandis que le CMP, soluble, se retrouve dans le lactosérum. Puisque le
CMP est responsable de la stabilité des micelles grâce à son caractère hydrophile et à sa charge nette
négative, sa perte dans le lactosérum permet la coagulation des caséines. À la suite de cette coagulation
mixte, il en résulte un coagulum partiellement fragilisé dû à sa forte déminéralisation (St-Gelais et Tirard-
Collet, 2002).
L'égouttage consiste à l'extraction du lactosérum, essentiellement composé d'eau et de matières solubles
telles que des composés azotés non coagulés (dont les protéines du lactosérum), des minéraux et du lactose.
Il permet donc la concentration des caséines et de la matière grasse. Dans le cas du Camembert, cette étape
débute au moment du soutirage, soit au moment où le caillé est mis en moule. Puisque le coagulum du
Camembert est en partie déminéralisé et fragilisé, les seuls traitements mécaniques pouvant être utilisés pour
favoriser l'expulsion du lactosérum sont le découpage, le brassage et le moulage. Suite au moulage, le caillé
s'égoutte de façon naturelle sous l'effet de son propre poids. Plusieurs retournements des moules sont
effectués afin d'accélérer l'expulsion du lactosérum. Tout au long de la période d'égouttage, l'acidification se
poursuit et le pH final du caillé varie normalement entre 4,6 et 4,7 dans le cas d'une fabrication traditionnelle
(Shaw, 1981, St-Gelais et Tirard-Collet, 2002, Sousa, 2003, Leclercq-Perlat, 2011).
En production industrielle, pour des raisons de rentabilité et afin de répondre à la demande de certains
consommateurs, il est possible de produire des fromages Camembert de type stabilisé (Spinnler et Gripon,
2004). Les pâtes de type stabilisées sont obtenues en limitant l'acidification du lait afin que le pH final ne
descende pas sous une valeur de 5,2. Pour ce faire, une partie des ferments mésophiles, ou sa totalité, est
remplacée par des streptocoques thermophiles. La baisse de la température de coagulation limite la
croissance et le métabolisme des ferments thermophiles ce qui ralentit l'acidification. Ainsi, le caillé est moins
acide, moins déminéralisé et moins coulant. Étant moins déminéralisés, les fromages ont un pouvoir tampon
plus élevé que les pâtes traditionnelles ce qui ralentit l'évolution du pH en cours d'affinage. Dans certains cas,
une étape de délactosage peut être ajoutée à cette technique limitant également l'acidification. Par contre, elle
a pour conséquence de réduire l'activité enzymatique pendant l'affinage, générant des fromages moins
goûteux. D'ailleurs, l'activité protéolytique des moisissures d'affinage serait moins importante à pH 5,2, qu'à

20
pH 4,6. La technologie des fromages de type stabilisé est avantageuse pour les industries qui l'utilisent
puisqu'elle prolonge la durée de vie de l'aliment (Lawrence et al., 1987, St-Gelais et Tirard-Collet, 2002).
À la suite de l'égouttage, le salage des fromages de type Camembert est réalisé sur les jeunes caillés au
lendemain de leur production (Leclercq-Perlat, 2011). Il s'agit d'une étape essentielle en technologie
fromagère; le sel (NaCl) y joue de multiples rôles, notamment au niveau du contrôle de la croissance
microbienne et des activités enzymatiques pendant l'affinage (section 1.1.4) (Guinee et Fox, 2004). De plus, il
contribue à la synérèse du caillé ce qui permet de complémenter l'égouttage (Fox et McSweeney, 2004). La
teneur en sel des fromages de type Camembert est variable et s'étend entre 1,5 et 2,5 % (Sousa, 2003). De
même, le ratio sel sur humidité (S/H) peut se situer entre 2,8 et 4,8 (Sutherland, 2002, Guinee et Fox, 2004).
Étant donné l'importance du salage dans le cadre de ce projet, ce sujet est traité de façon distincte à la section
1.2.2.
L'affinage correspond à la période d’entreposage des fromages dans des conditions contrôlées (température
et humidité) au cours de laquelle une série de réactions biochimiques se produisent (section 1.3.1). Ces
changements biochimiques sont dus à l'action catalytique de divers agents, soit les enzymes endogènes du
lait, les agents coagulants et les enzymes microbiennes. Ces agents permettent de métaboliser la matière
grasse, les protéines et le lactose résiduel afin de générer les caractéristiques uniques à chaque variété de
fromage, soit leurs arômes, leurs saveurs et leur texture. Le temps d'affinage est généralement inversement
proportionnel à la teneur en humidité du fromage (Fox et McSweeney, 2004). La période d'affinage des
Camemberts de production traditionnelle est réalisée entre 11 et 13 C et 90 à 95 % d'HR pendant trois
semaines à un mois. Dans le cas des fromages de type Camembert de production industrielle, les paramètres
d'affinage sont sensiblement les mêmes, mais sont de plus courte durée, soit environ deux semaines. Les
fromages sont ensuite emballés et placés entre 4 et 8 C (Shaw, 1981, Spinnler et Gripon, 2004), où le
processus d’affinage se poursuit, mais plus lentement (Leclercq-Perlat, 2011).
1.2.2. Les techniques de salage en fromagerie
En transformation fromagère, trois techniques sont principalement utilisées afin de saler les fromages; le
salage dans la masse, le salage à sec en surface et le saumurage (Sutherland, 2002, Gillis, 2004, Guinee et
Fox, 2004). Le salage à sec en surface et le saumurage sont utilisés dans le cas du Camembert. En
fabrication de type traditionnelle, le salage des Camemberts se fait à sec en surface par saupoudrage manuel,
tandis qu'en technologie industrielle, le saumurage et le salage à sec automatisé sont utilisés (Shaw, 1981).
Aujourd'hui, le saumurage constitue la technique la plus courante pour saler les fromages de type Camembert,
puisqu'il permet une meilleure uniformité du contenu en sel et réduit la manutention (Hardy, 1985, Sousa,
2003).

21
1.2.2.1. Le salage à sec
Le salage à sec est en grande partie utilisé lors de la fabrication des fromages à pâte molle, tels que le
Camembert et le fromage Bleu. Comme son nom l'indique, il consiste à l'ajout de sel sec à la surface des
fromages. Plusieurs techniques peuvent être utilisées afin d'appliquer la quantité de sel désirée; le roulage sur
tapis de sel, le frottage manuel ou le saupoudrage. Une mécanisation de cette dernière version est aujourd'hui
possible et permet l'obtention d'une meilleure régularité au niveau de l'application du sel. Elle fait appel à
l'utilisation de lits fluidisés ou d'appareils à rideau de sel et à l'ajout de sel de granulométrie spécifique
contenant des anti-agglomérants (Gillis, 2004, Mietton et al., 2004).
Lors de l'application du sel, l'humidité superficielle du fromage provoque une dissolution des cristaux ce qui
permet ensuite leur absorption dans la pâte (Gillis, 2004). La pression osmotique engendrée occasionne alors
un mouvement des molécules d'eau du centre du fromage vers la surface et la formation d'une mince couche
de saumure très concentrée localisée immédiatement à la surface et présente pendant plusieurs jours. La
présence prolongée de cette saumure est responsable de la contraction des caséines à la surface du fromage
et d'une importante perte d'eau à cet endroit. Conséquemment, la migration du sel dans la pâte est ralentie et
la prise en sel se fait moins rapidement que celle observée lors du saumurage. Au cours de l'affinage, le ratio
S/H à travers la pâte tend à s'équilibrer, mais ce processus est relativement long (Guinee et Fox, 2004). Les
mécanismes de prise en sel lors du salage à sec diffèrent donc de ceux observés au moment du saumurage
et sont dépendants de l'humidité à la surface du fromage, de la granulométrie du sel et de la quantité ajoutée
(Hardy, 1985).
1.2.2.2. Le saumurage
Le saumurage consiste à submerger les fromages démoulés dans une solution concentrée, la plupart du
temps saturée, en NaCl. La différence de concentration en sel entre la phase aqueuse du fromage et la
saumure provoque une diffusion du sel vers l'intérieur du fromage et la migration du lactosérum dans le sens
opposé afin d'équilibrer la pression osmotique entre ces deux phases. Ainsi, une partie de l'eau, des minéraux
solubles, du lactose, de la matière azotée non coagulée, de la matière grasse et des microorganismes
provenant du fromage se retrouve dans la saumure, ce qui contribue à son enrichissement (Hardy, 1985,
Guinee et Fox, 2004, Mietton et al., 2004). On observe éventuellement un état d’équilibre dans la saumure
pour ces divers composants, après un certain temps d’utilisation. Dans la majorité des entreprises, la saumure
n’est jamais complètement remplacée, mais plutôt complétée au fil du temps par une portion de nouvelle
saumure. Ceci est effectué afin d’éviter de déséquilibrer la saumure de ses divers éléments. Le temps de
saumurage appliqué dépend de la teneur saline visée et varie en fonction des différentes variétés de fromages
(Sutherland, 2002). Il peut s'étendre de quelques minutes à plusieurs jours. Dans le cas du Camembert, cette
opération s'étale généralement entre 30 et 60 minutes (Shaw, 1981).

22
1.2.2.2.1. La prise en sel lors du saumurage
Les phénomènes intervenant au niveau de la prise en sel lors du saumurage ont fait l'objet de nombreuses
études (Geurts et al., 1974, Hardy, 1976, Geurts et al., 1980, Guinee et Fox, 1986b, a, Chamba, 1988, De
Piante et al., 1990, Melilli et al., 2003a, Melilli et al., 2003b, Melilli et al., 2005, Melilli et al., 2006). La figure 1.4
représente le profil associé à la cinétique d'incorporation du sel lors du saumurage d'un fromage de type
Camembert. Tel que démontré, la quantité de sel absorbée augmente en fonction du temps de saumurage
tandis que sa vitesse d'absorption ralentit. Des profils similaires ont été observés chez plusieurs autres
variétés fromagères, dont le Gouda (Geurts et al., 1980), le Romano (Guinee et Fox, 1986b, a), l'Emmental
(Chamba, 1988), le Cuartirolo (De Piante et al., 1990) et le Rugasano (Melilli et al., 2003a, Melilli et al., 2003b,
Melilli et al., 2006). Le ralentissement du taux d'absorption du sel en fonction du temps de saumurage a été
associé à la réduction du gradient de concentration en sel entre la saumure et la phase aqueuse du fromage
(Guinee et Fox, 2004). Bien que semblant simple d'exécution, les multiples facteurs intervenants pendant le
saumurage et susceptibles d'affecter la teneur en sel incorporée demandent un contrôle rigoureux de cette
opération. Les paramètres régissant la prise en sel sont reliés aux caractéristiques intrinsèques des fromages,
à la composition de la saumure et aux paramètres de saumurage (Guinee et Fox, 2004).
Figure 1.4 – Cinétique d'incorporation du sel dans un Camembert immergé dans une saumure saturée statique (○) et dans une saumure agitée (●). Le pH des saumures a été ajusté à 4,6 à la température à 14 C (Tirée de Hardy, 2009).

23
Paramètresreliésauxfromages
La géométrie de l'aliment ainsi que sa composition sont deux facteurs ayant une influence sur l'absorption du
sel pendant le saumurage (Guinee et Fox, 2004). La géométrie des fromages fait référence à leur taille et leur
forme. De façon générale, la vitesse d'absorption du sel sera d'autant plus élevée que la taille du fromage sera
petite. Plus précisément, ce phénomène est dû au ratio entre la surface du fromage et le volume de la
saumure. Pour des fromages ayant le même volume, plus la surface de contact avec la saumure sera élevée,
plus rapide sera la prise en sel. L'effet de la forme des fromages sur la prise en sel s'explique par le nombre
de directions disponibles pour la pénétration des solutés, celles-ci étant plus élevées dans un fromage de
forme rectangulaire que cylindrique. L'incorporation du sel serait avantagée au niveau des surfaces planes
comparativement aux surfaces courbées (Geurts et al., 1980, Guinee et Fox, 1986b, a). Pour ce qui est de la
composition de l'aliment, le contenu en eau avant le saumurage joue un rôle important au niveau de
l'absorption du sel. Plus l'humidité est élevée, plus la porosité dans la matrice fromagère sera élevée. Le sel
diffuse à travers la phase aqueuse du fromage et ne peut passer à travers la matrice protéique ou la matière
grasse (Melilli et al., 2003b). Ainsi, la vitesse d'absorption du sel augmente en fonction de l'importance de la
teneur en humidité (Geurts et al., 1974, Melilli et al., 2003b). Le pH du fromage influencerait également la prise
en sel. Geurts et coll. (1980) ont observé une absorption de sel plus importante dans un Gouda à pH 4,7,
comparativement à un pH de 5,7. La présence du lactate dans la phase aqueuse du fromage provoquerait une
diffusion de celui-ci vers la saumure et un gain d'eau dans la matrice fromagère, ce qui favorise la prise en sel
(Guinee et Fox, 2004).
Paramètresreliésàlacompositiondessaumures
L'absorption du sel est également dépendante de la composition saline des saumures. De façon générale,
plus la teneur saline de celles-ci est importante, plus la pénétration du sel sera élevée (Hardy, 1976, Geurts et
al., 1980, Guinee et Fox, 1986a, Chamba, 1988). Cependant, certains auteurs ont observé un ralentissement
de l'absorption du sel après salage en saumures très concentrées, c'est-à-dire près du taux de saturation, ce
qui s'explique par une plus grande perte d'eau à la surface du fromage (Chamba, 1988, Melilli et al., 2003b,
Melilli et al., 2005, Melilli et al., 2006). Un haut taux de déshydratation superficielle cause un rétrécissement de
la porosité dans la matrice fromagère ce qui crée une barrière à la pénétration du sel (Melilli et al., 2003b,
Melilli et al., 2005). Ensuite, puisque les saumures sont utilisées à répétitions pour plusieurs lots de fromages,
elles s'enrichissent en divers solutés provenant de la phase aqueuse des fromages, ce qui correspond au
phénomène de vieillissement chimique. L'évolution observée dans des saumures destinées au salage de
fromages de type Camembert démontre que leur usage répété permet une augmentation du contenu en acide
lactique, en lactose, en matière azotée et en matière minérale telle que le calcium, le potassium et le
phosphore (Lacrampe et al., 1971, Terré et al., 1985). Ces composés défavorisent les échanges entre la

24
saumure et le fromage, ce qui a été observé dans le Camembert (Terré et al., 1985) et l'Emmental (Chamba,
1988). De plus, la présence de minéraux, autres que le sodium, entrave la solubilisation du NaCl (Mietton et
al., 2004). Parallèlement à cette évolution chimique, la concentration microbienne dans la saumure augmente.
Ces saumures peuvent donc devenir un vecteur de contamination (Hardy, 1985). C’est pourquoi plusieurs
fromageries utilisent des filtres pour réduire la présence de particules fines dans la saumure, mais également
de bactéries (ex. filtration Padovan). La teneur en calcium des saumures doit également être ajustée par
l'ajout de CaCl2. Une trop faible teneur en calcium dans la saumure provoquera une diffusion du calcium
colloïdal vers la saumure. Ainsi, l'hydratation des caséines sera plus importante ce qui a pour effet
d'augmenter la teneur en eau à la surface du fromage et de voir apparaître certains défauts de fromagerie
(croûte poisseuse, croûte pourrie) (Guinee et Fox, 2004).
Paramètresreliésauxconditionsdesaumurage
Comme démontré à la figure 1.4, l'agitation des saumures favorise la prise en sel. Cette opération permet
d'assurer une homogénéité de la saumure, notamment au niveau de la teneur en sel, de la température et du
pH. La perte de lactosérum pendant le saumurage provoque une dilution de la saumure en périphérie des
fromages et réduit donc la teneur saline à laquelle ils sont exposés (Hardy, 1976). Ainsi, l'absorption du sel se
fait moins rapidement dans une saumure statique, tandis que l'agitation permet d'éviter cette dilution locale. La
température de saumurage varie normalement entre 10 et 15 C (Gillis, 2004). L'augmentation de la
température est corrélée à une plus grande teneur en sel incorporée ce qui a été observé dans le Gouda,
(Geurts et al., 1974), le Romano (Guinee et Fox, 1986a), le Camembert (Hardy, 1976), l'Emmental (Chamba,
1988) et le Rugasano (Melilli et al., 2003a). Selon Melilli et al. (2003a), ces observations s'expliqueraient par
une diminution de la viscosité de la phase aqueuse. De plus, à une température inférieure à 10 C, la
cristallisation de la matière grasse est favorisée ce qui freine la prise en sel. Par contre, des températures trop
élevées favorisent la prolifération microbienne (Gaüzère, 2013). Le pH des saumures étant normalement
ajusté à une valeur se rapprochant de celle du caillé, il aurait peu d'effet sur la prise en sel, ce qui a été
démontré dans le cas du Camembert (Hardy, 1976) et de l'Emmental (Chamba, 1988). Par contre, un pH trop
acide (< 4,6) provoquera une précipitation des caséines et une importante perte d'humidité (Hardy, 1976,
Guinee et Fox, 2004). Le volume de fromage par rapport au volume de saumure doit également être pris en
considération puisqu'une trop grande quantité de fromage défavoriserait l'incorporation du sel pendant le
saumurage. Le ratio du volume de saumure devrait normalement être cinq fois supérieur, ou plus, au volume
de fromage afin qu'il n'influence pas l'absorption du sel (tel que cité dans Bintsis, 2006).

25
Estimationdelapriseensel
La prise en sel lors du saumurage peut être exprimée selon une relation mathématique basée sur la loi de
diffusion de Fick, originalement adaptée au fromage Gouda (Geurts et al., 1974). Elle a ensuite été adaptée au
fromage Camembert (Hardy, 1976). Elle permet d'estimer la quantité de sel absorbée lors du saumurage pour
un temps donné et suit l'équation mathématique suivante :
S 2C ∙ ∙
où : S = g de sel/100 g d'eau de fromage ; C = concentration en sel de la saumure (g/100 g d'eau) ; A = surface du fromage (cm2) ; V = volume du fromage (cm3) ; D = coefficient de diffusion du sel, spécifique à chaque fromage (environ 0,20 cm2/jour pour le
Camembert); t = temps de saumurage (jours).
L'application de cette loi nécessite que le saumurage soit effectué dans des conditions précises; le volume de
saumure devrait être dix fois plus élevé que le volume de fromages et la saumure doit être agitée
correctement afin que la température et la concentration en sel soient homogènes en tout temps autour du
fromage (Hardy, 1985, 2009). De plus, le coefficient de diffusion du sel (D), spécifique à chaque fromage, doit
être connu ce qui limite l'utilisation de cette relation mathématique en industrie.
1.2.2.2.2. La diffusion du sel pendant l'affinage
À l'instar du salage à sec en surface, la majorité du sel absorbé à la suite du saumurage se retrouve à la
surface du fromage; l'aw à cet endroit est donc particulièrement faible, soit d'environ 0,75. Puisqu'il existe un
gradient de concentration entre la quantité de sel présente dans les couches superficielles et la teneur en eau
dans le centre de la pâte, le sel continue à diffuser vers l'intérieur du fromage pendant l'affinage (Figure 1.5)
jusqu'à ce qu'une répartition quasi-homogène soit atteinte (Hardy, 2009). Il s'agit d'un processus relativement
long qui peut s'étaler, par exemple, entre 7 à 10 jours dans le cas d'un Camembert de 0,25 kg et de 55 %
d'humidité (Sutherland, 2002). Selon Hardy (2009), cet équilibre pourrait même ne jamais être atteint dû à
l'assèchement de la surface des fromages. Les facteurs influençant l'atteinte de l'équilibre salin dans le
fromage sont semblables à ceux de la prise en sel lors du salage et sont liés à sa géométrie, la température,
le gradient de concentration entre les différentes parties de l'aliment et sa composition. L'humidité favorise la
diffusion du sel tandis que la matière grasse et les protéines obstruent le passage des ions Na+ et Cl-. Aussi,
les pores du réseau protéique ainsi que l'eau liée aux protéines effectuent un effet de tamisage et de friction

26
affectant la mobilité des solutés. De plus, la viscosité de la phase aqueuse influence négativement la diffusion
du sel, celle-ci étant dépendante de la température (Guinee et Fox, 2004, Guinee et al., 2014b).
Figure 1.5 – Évolution de la teneur saline au centre d'un fromage Camembert pendant l'affinage. Le symbole (●) correspond aux données prises en surface et le symbole ( ○) correspond aux données prises au centre du fromage (Tirée de Hardy, 2009).
1.2.3. Le dosage du sel dans les fromages
Plusieurs méthodes peuvent être utilisées afin de doser la quantité de sel dans les fromages. Ainsi, pour un
mélange des sels NaCl et KCl, cette quantification peut être réalisée par le dosage des ions chlorures ou des
ions sodium et potassium. Une fois le dosage effectué, la masse molaire de chaque ion est ensuite utilisée
afin de convertir, par calcul, les résultats générés sous forme de concentration de sels, soit en NaCl ou en KCl
(Gillis, 2004). La titration potentiométrique s'avère être une méthode analytique normalisée (ISO/FIL)
permettant de doser les ions chlorures dans les fromages (ISO 5943, FIL-88) (CCIL, 2015). Le chloruremètre
est un appareil simple d'utilisation et utilisé en fromagerie afin de mesurer les chlorures. Le principe de cet
appareil est étroitement lié à la technique de titration potentiométrique (Gillis, 2004). Cependant, le dosage
des chlorures ne peut discriminer la nature des sels (NaCl, KCl et CaCl2) si les ions tels que le sodium, le
potassium ou le calcium ne sont pas dosés en parallèle. Cette technique de dosage des ions chlorure a
tendance à légèrement surestimer le contenu en NaCl réel des fromages ce qui peut s'expliquer, à titre
d'exemple, par l'ajout de CaCl2 au lait lors de la fabrication fromagère, ce qui est également dosé par l'appareil
(Gillis, 2004). Ceci a d'ailleurs été confirmé par dosage du chlorure et du sodium à partir de mêmes
échantillons de fromages expérimentaux de type Gouda (Geurts et al., 1974). Dans ce cas, il devient donc
pertinent de doser l'ensemble des cations d'intérêt afin d'obtenir un résultat plus précis.

27
Le dosage du sodium et du potassium dans les fromages peut être réalisé par spectrométrie atomique. Parmi
les variantes possibles, l'absorption atomique ou l'émission atomique peuvent être utilisées (Gillis, 2004,
Mermet, 2004). Ces méthodes débutent d'abord par la minéralisation de l'aliment par calcination et/ou par
digestion en milieu acide (Gaucheron, 2004). Les éléments à quantifier sont ensuite atomisés. Dans le cas de
l'absorption atomique, l'atomisation peut être réalisée à l'aide d'un mélange gazeux (air/acétylène ou
N2O/acétylène) sous forme de flamme tandis que pour l'émission atomique, elle est réalisée par exposition de
l'échantillon à un plasma, généralement composé d'argon (inductively coupled plasma ou ICP). Les atomes
excités absorbent et émettent de l'énergie à des longueurs d'ondes spécifiques à chaque élément, ce qui
permet de les identifier. Ces méthodes deviennent difficiles à appliquer de façon routinière dans un cadre
industriel compte tenu de la manutention associée à la préparation des échantillons et l’expertise nécessaire
pour utiliser les appareils. Cependant, elles ont l'avantage d'avoir une grande sensibilité. À titre d'exemple, la
limite de détection du sodium et du potassium par spectrométrie d'émission à plasma induit est de 0,5 µg/L et
de 1 µg/L, respectivement (Perkin-Elmer, 2004).

28
1.3. L'AFFINAGE DU CAMEMBERT ET SA MICROFLORE FONGIQUE DE SURFACE
Parmi les différentes variétés fromagères, le Camembert est reconnu pour la grande complexité de son
affinage et sa rapidité. En fait, cette complexité est attribuée à l'importance de la diversité de la microflore
d'affinage. La microflore d'affinage du Camembert est composée de différentes espèces bactériennes, de
levures et de moisissures. La diversité de cette microflore est variable et est influencée par les procédés de
fabrication tels que la pasteurisation. La microflore secondaire sera d'autant plus complexe si le fromage est
produit à partir de lait cru. La teneur en humidité élevée et le développement des mycètes en surface sont les
principaux facteurs responsables de la rapidité de la vitesse d'affinage. Au cours de cette période de
maturation, les multiples réactions cataboliques issues de l'activité des enzymes coagulantes, de la plasmine
ou des enzymes d'origine microbienne contribuent aux changements chimiques de l'aliment, générant l'aspect
et les propriétés sensorielles typiques à celui-ci. De plus, les modifications de cet environnement laissent
place à l'évolution d'une microflore se développant en fonction des conditions générées (Sousa, 2003,
Spinnler et Gripon, 2004).
1.3.1. Les phénomènes biochimiques
Les trois principaux évènements biochimiques ayant lieu lors de l'affinage des fromages étant responsables
de la formation de composés aromatiques et du développement de la texture sont le métabolisme du lactose
et de l'acide lactique, la protéolyse et la lipolyse (Figure 1.6) (Sousa, 2003, Marilley et Casey, 2004).

29
Figure 1.6 - Voies métaboliques responsables de la génération de composés aromatiques pendant l'affinage des fromages (Tirée de Marilley et Casey, 2004).
Le métabolisme du lactose débute par sa fermentation en acide lactique via l'activité des bactéries lactiques.
Pendant l'affinage du Camembert, le catabolisme du lactate en dioxyde de carbone et en eau par les ferments
fongiques engendre l'implantation d'un gradient de pH entre la surface et le centre de l'aliment ce qui a un
impact sur l'évolution microbienne et le développement de la texture (sections 1.3.2 et 1.3.3) (Sousa, 2003,
McSweeney, 2004, Spinnler et Gripon, 2004).
Dans le Camembert, la protéolyse s'avère être le phénomène biochimique le plus important et le plus
complexe. Elle a un impact direct sur le développement de la texture du Camembert, soit le ramollissement de
la pâte, dû à la dégradation des caséines (McSweeney, 2004). De plus, elle contribue à la réduction de l'aw de
l'aliment puisque les peptides générés ont la capacité de lier une partie de la phase aqueuse (Schlesser et al.,
1992, McSweeney, 2004). Les peptides et acides aminés formés contribuent ensuite au développement des
saveurs ainsi qu'aux défauts d'arômes, tels que l'amertume. Les acides aminés libres servent de précurseurs
à diverses réactions cataboliques qui mènent à la formation de composés aromatiques tels que des amines,
des aldéhydes, des composés soufrés ainsi que divers acides et alcools (Molimard et Spinnler, 1996, Sousa,
2003, Marilley et Casey, 2004, Spinnler et Gripon, 2004). La protéolyse du Camembert débute d'abord par la
dégradation des caséines en différents peptides via l'action de la présure, de la plasmine et autres enzymes
naturellement retrouvées dans le lait. Ces peptides sont par la suite métabolisés à l'aide de protéases et

30
peptidases produites par les bactéries lactiques, coagulantes ou non, la microflore secondaire et les ferments
d'affinage. La protéolyse s'avère être plus intense en surface qu'au centre du Camembert puisque les mycètes
colonisant la croûte de ces fromages y sont les principaux contributeurs (Molimard et Spinnler, 1996, Sousa,
2003, Spinnler et Gripon, 2004). Dans le Camembert, l’expression des gènes liés à la formation de protéases
et de peptidases d'origine fongique se produirait majoritairement durant les 21 premiers jours d'affinage
(Lessard et al., 2014).
Le catabolisme des lipides du Camembert consiste en la dégradation des triglycérides en diglycérides, en
monoglycérides et en acides gras libres. Ces derniers sont d'une grande importance puisqu'ils contribuent à la
texture de l'aliment, au goût et ils servent de précurseurs à la formation de plusieurs molécules aromatiques
telles que les esters, les alcools, les lactones et les méthyl-cétones. D'ailleurs, les méthyl-cétones s'avèrent
être les composés aromatiques les plus abondants dans le Camembert et sont associés aux arômes fruités,
de moisi et de bleu (Molimard et Spinnler, 1996, Spinnler et Gripon, 2004). Tel qu'observé au niveau de la
protéolyse, les principaux contributeurs à la lipolyse du Camembert sont les enzymes d'origines fongiques, ce
qui fait en sorte que la dégradation des lipides est plus intense en surface qu'au centre de l'aliment. L'activité
lipolytique associée aux lipases d'origine fongique serait exprimée dès les premiers jours de l'affinage et se
produirait tout au long de celle-ci (Lessard et al., 2014).
1.3.2. L'évolution microbienne
L'établissement de la microflore du Camembert est enclenché par l'acidification de la pâte due au métabolisme
des bactéries lactiques. Tolérant des environnements acides et salés, les levures sont les premières espèces
à coloniser la surface du Camembert. Elles sont visibles dès les trois premiers jours de l'affinage (Leclercq-
Perlat et al., 2004). La diversité associée à ces espèces est variable. Dans les fromages Camemberts faits de
lait cru, les levures majoritairement retrouvées sont Kluyveromyces marxianus ssp. lactis, Kluyveromyces
marxianus ssp. marxianus, Saccharomyces cerevisiae et Debaromyces hansenii (Schmidt et al., 1978). Elles
ont été identifiées à la surface et au centre de l'aliment et leur répartition varie entre les différents fromages et
le stade d’affinage (Schmidt et al., 1980). Geotrichum candidum est également retrouvée dans les fromages
faits de lait non pasteurisé et, par ces multiples attributs technologiques (section 1.3.4.1), est la levure la plus
importante dans la fabrication des fromages Camemberts (Molimard et Spinnler, 1996, Boutrou et Gueguen,
2005). Dans les fromages faits de lait pasteurisé, K. marxianus, K. lactis, D. hanseniii et G. candidum sont
ensemencées lors de la fabrication fromagère à titre de ferments d'affinage. Penicillium camemberti est la
moisissure la plus importante des fromages Camemberts due à ses multiples rôles (section 1.3.3.1). Ses
spores sont ajoutées volontairement au lait pendant la fabrication fromagère ou vaporisées à la surface du
caillé démoulé. P. camemberti apparaît en surface un peu plus tard que les levures, soit après 6 à 7 jours
d'affinage. Grâce à leurs capacités à métaboliser le lactate et à générer de l'ammoniac, les levures et

31
P. camemberti contribuent à la neutralisation de la pâte du fromage. Cette neutralisation laisse place au
développement d'une microflore bactérienne aérobie, halophile et acido-sensible à la surface du fromage
(Molimard et Spinnler, 1996, Guizani et al., 2002, Sousa, 2003, Leclercq-Perlat et al., 2004, Spinnler et
Gripon, 2004, Leclercq-Perlat, 2011). Cette dernière est responsable de la production de composés
aromatiques et est généralement composée de staphylocoques, de coliformes et de plusieurs espèces
appartenant au groupe des bactéries corynéformes. À l’intérieur de ce groupe, Brevibacterium linens s'avère
être l'espèce la plus abondante et contribue activement au développement de flaveurs par son implication
dans la production de composés volatils soufrés (Richard et Zadi, 1983, Spinnler et Gripon, 2004,
Bockelmann, 2010).
1.3.3. Penicillium camemberti
Les moisissures du genre Penicillium font partie du groupe des ascomycètes (Bockelmann, 2010). Penicillium
camemberti serait une espèce domestiquée de la moisissure d'altération Penicillium commune (Pitt et al.,
1986). La classification des moisissures étant complexe, P. camemberti était originalement connue sous le
nom de Penicillium album et se distinguait par ses conidies de teintes verdâtres. Penicillium caseicolum,
originalement appelée Penicillium candidum, correspondait plutôt à la moisissure formant des thalles de
couleur blanche. Aujourd'hui, ces espèces sont regroupées sous le même nom, soit P. camemberti, et seule la
forme blanche est utilisée dans la fabrication des fromages de type Camembert. Le choix des souches
utilisées à titre de ferment d'affinage est principalement basé sur la densité et la hauteur du mycélium formé
ainsi que sur les propriétés protéolytiques et lipolytiques des souches (Pitt et al., 1986, Cerning et al., 1987,
Spinnler et Gripon, 2004, Bockelmann, 2010). Cependant, peu de variations intra-espèce au niveau de
l'activité protéolytique ont été observées, contrairement à de plus grandes différences pour l'activité lipolytique
(Lenoir et al., 1971, Spinnler et Gripon, 2004).
1.3.3.1. Les rôles de P. camemberti lors de l'affinage
P. camemberti s'avère être l'espèce fongique la plus abondante à la surface des fromages de type Camembert
(Lessard et al., 2012, Lessard et al., 2014). Elle intervient au niveau de l'apparence du fromage, lui conférant
son feutrage blanc typique. De plus, l'implantation de cette couche offre un rôle de protection contre certains
microorganismes indésirables (Bockelmann, 2010). À titre d'exemple, elle possèderait des propriétés
inhibitrices contre le développement des moisissures d'altération Cladosporium herbarum, Penicillum
caseifulvum, P. roqueforti et P. commune. Ces observations sont cependant souche-dépendante et l'inhibition
observée est affectée par la présence de G. candidum (Decker et Nielsen, 2005).
P. camemberti intervient activement au niveau du développement des propriétés sensorielles de l'aliment.
Dans les fromages à pâte molle et à croûte fleurie, elle est reconnue comme étant l'espèce la plus

32
protéolytique (Sousa, 2003). Elle synthétise diverses enzymes protéolytiques telles que des
carboxypeptidases, des métalloprotéases, une aspartyl-protéase et une aminopeptidase et permet la
génération de plusieurs acides aminés libres (Cerning et al., 1987, Sousa, 2003, Spinnler et Gripon, 2004).
Elle est également impliquée au niveau du catabolisme de certains acides aminés, dont la méthionine, celle-ci
étant précurseur à la formation de plusieurs composés aromatiques (Molimard et Spinnler, 1996). Dans les
fromages de type Camembert, P. camemberti est reconnue comme étant l'espèce la plus lipolytique (Sousa,
2003, Lessard et al., 2014). Elle produit entre autres une grande quantité de lipases alcalines extracellulaires
(Spinnler et Gripon, 2004). Son implication au niveau du catabolisme des acides gras permet la production de
méthyl-cétones, il s'agit d'importants contributeurs à l'arôme typique du Camembert. P. camemberti est
notamment responsable de la production de l'alcool oct-1-en-3-ol, celui-ci identifié comme étant le composé
clé responsable de la note de champignon du Camembert (Karahadian et al., 1985, Molimard et Spinnler,
1996). Lors d'une croissance excessive, son métabolisme est également associé à la génération de peptides
hydrophobes reconnus pour leur caractère amérisant (Vassal et Gripon, 1984, Molimard et al., 1994).
Par sa capacité à consommer le lactose, l'acide lactique et à produire de l'ammoniac issu de son métabolisme
protéolytique, P. camemberti est indirectement responsable de l'évolution de la texture du Camembert
(Figure 1.7). En plus de permettre l'implantation de la microflore bactérienne acido-sensible en surface et de
favoriser l'activité de certaines enzymes (plasmine), la neutralisation de la pâte contribue à l'établissement
d'un gradient de pH ainsi qu'un gradient minéral, ce qui permet son ramollissement (McSweeney, 2004).
Figure 1.7 – Changements physico-chimiques pendant l'affinage du Camembert attribués au développement de P. camemberti (Adaptée de McSweeney, 2004).

33
1.3.4. Geotrichum candidum
Geotrichum candidum (téléomorphe: Galactomyces geotrichum) est une levure dimorphique
phylogénétiquement associée au groupe des ascomycètes. Elle peut se présenter sous plusieurs
morphotypes soit sous une forme unicellulaire typiquement observée chez les levures ou former un mycélium,
caractéristique aux moisissures. Il s'agit d'une levure ubiquitaire étant parfois retrouvée dans le lait cru. Elle
colonise la surface de plusieurs variétés fromagères, dont les pâtes molles telles que le Camembert, les
fromages au lait de chèvre et de brebis et les pâtes semi-fermes comme le Saint-Nectaire et le Reblochon.
Cependant, elle peut se retrouver parmi la microflore contaminante de certains fromages tels que le Cottage et
le Bleu (van den Tempel et Nielsen, 2000, Boutrou et Gueguen, 2005). Cette espèce présente une grande
variabilité génétique, ce qui a d'ailleurs été observé entre plusieurs souches isolées de divers environnements
laitiers (Marcellino et al., 2001, Alper et al., 2013). Il n'est donc pas étonnant de constater les variations
observées au niveau des capacités technologiques des souches utilisées à titre de ferment d'affinage dont
leur pouvoir désamérisant et leurs habiletés à former des composés aromatiques (Bockelmann, 2010).
1.3.4.1. Les rôles de G. candidum lors de l'affinage
G. candidum apparaît à la surface et au centre des fromages de type Camembert durant les premiers jours
d'affinage (Leclercq-Perlat et al., 2004, Lessard et al., 2012, Lessard et al., 2014). Tout comme
P. camemberti, son développement en surface contribue à l'établissement du feutrage blanc (Bockelmann,
2010). Sa présence joue également un rôle au niveau de la texture, de l'épaisseur et du maintien de la croûte
(Marcellino et al., 2001). Cependant, une croissance trop prononcée peut mener à l'apparition de défauts de
fromagerie, dont la peau de crapaud (Spinnler et Gripon, 2004). Plusieurs interactions microbiennes
impliquant son développement ont préalablement été observées. Par exemple, sa présence limiterait le
développement de P. camemberti à la surface de fromages à pâte molle et à croûte fleurie (Vassal et Gripon,
1984). Par contre, un effet synergique entre les deux espèces a plutôt été observé en milieu de culture à base
de lactosérum (Aziza et al., 2005). De plus, G. candidum joue un rôle protecteur à la surface du Camembert
par sa capacité à inhiber la croissance de plusieurs moisissures contaminantes du genre Penicillium
(P. commune, P. caseifulvum, P. verrucosum, P. discolor, P. solitum et P. coprophilum) ainsi qu'Aspergillus
versicolor (Nielsen et al., 1998) et Mucor (Boutrou et Gueguen, 2005). Certaines souches produiraient
également des composés capables d'inhiber la croissance de L. monocytogenes (Dieuleveux et al., 1998).
G. candidum contribue largement au développement de composés aromatiques dans les fromages de type
Camembert. Bien que son activité protéolytique dans ces fromages soit plus faible que celle observée chez
P. camemberti, elle synthétise plusieurs protéases et peptidases impliquées dans la formation de composés
aromatiques, celles-ci variant en fonction de la souche utilisée (Boutrou et Gueguen, 2005, Boutrou et al.,

34
2006). Entre autres, elle contribue à l'amélioration de l'aspect sensoriel des fromages par sa production
d'aminopeptidases et de carboxypeptidases capables de dégrader les peptides amers générés par
P. camemberti (Vassal et Gripon, 1984, Molimard et al., 1994, Boutrou et Gueguen, 2005). De plus, sa
présence permet d'atténuer la perception des goûts de carton et de moisi causés par le métabolisme de
P. camemberti (Molimard et al., 1997). Elle participe activement au catabolisme de la méthionine générant des
composés soufrés, ceux-ci étant reconnus pour leur caractère aromatique (Molimard et Spinnler, 1996, Gente
et al., 2007, Lessard et al., 2014). G. candidum contribue également à la lipolyse des fromages de type
Camembert, mais son activité lipolytique s'avère moins prononcée que celle de P. camemberti (Lessard et al.,
2014). Elle contribue à la formation d'importants composés aromatiques, dont les méthyl-cétones et les esters
(Molimard et Spinnler, 1996, Leclercq-Perlat et al., 2007). Son activité lipolytique est cependant souche-
dépendante (Marcellino et al., 2001, Boutrou et Gueguen, 2005).
Tel qu'observé chez P. camemberti, G. candidum contribue à la neutralisation de la pâte des fromages de type
Camembert par sa capacité à produire de l'ammoniac et à consommer l'acide lactique (Boutrou et Gueguen,
2005). Ainsi, elle est également responsable du développement de la texture et de l'implantation de la
microflore bactérienne de surface (Figure 1.7) (McSweeney, 2004).
1.3.5. L'osmotolérance chez les mycètes
De façon générale, les levures et les moisissures sont reconnues pour leur capacité à tolérer et à croître dans
des environnements où les concentrations en ions ou en carbohydrates sont élevées, contrairement aux
bactéries. L'osmotolérance parmi ce groupe microbien serait prévalent chez les ascomycètes. Les
mécanismes moléculaires et physiologiques intervenant au niveau de l'osmotolérance des mycètes ont
principalement été évalués chez les levures Saccharomyces cerevisiae et Debaromyces hansenii, cette
dernière étant reconnue pour sa forte tolérance face au sel (Larsson et al., 1990, Hooley et al., 2003).
L'osmorégulation chez les mycètes est basée sur leur capacité à maintenir leur potentiel osmotique
intracellulaire en fonction de l'environnement dans lequel ils sont présents par augmentation ou diminution de
la concentration en solutés dans leur cytoplasme. En milieu hypertonique, les cellules ont la capacité
d'accumuler des molécules capables de compenser la différence de concentration en solutés avec l'extérieur.
Cependant, l'accumulation de fortes teneurs en ions solubles est reconnue pour son potentiel cytotoxique sur
le métabolisme cellulaire, notamment au niveau de l'activité de certaines enzymes. Ainsi, pour éviter
l'exposition à ces ions toxiques, les mycètes accumulent ou synthétisent des solutés compatibles, aussi
connus sous le nom d'osmoprotecteurs. L'effet toxique de ces composés sur le métabolisme cellulaire serait
faible ou inexistant. Chez les mycètes, ces osmoprotecteurs sont des composés organiques, principalement

35
des polyols ainsi que des acides aminés ou des dérivés de ceux-ci. Le soluté compatible le plus commun chez
ces microorganismes est le glycérol (Blomberg et Adler, 1992, Hooley et al., 2003).
1.3.5.1. Comportement de P. camemberti en fonction du sel
Il existe peu d'informations traitant du comportement de P. camemberti en fonction du stress causé par le sel.
De plus, les mécanismes physiologiques et moléculaires y étant reliés n'ont pas été étudiés chez cette
espèce. Les études se basent principalement sur des analyses du taux de croissance sur milieux de culture
synthétique, sur caillé modèle et sur fromage Camembert soumis à différentes teneurs en NaCl.
Sur milieu de culture Sabouraud, P. camemberti possède une tolérance élevée face au NaCl; sa croissance
est visible après trois jours d'incubation à 8, 10 et 12 % de NaCl. Elle s'avère considérablement retardée à
20 % de NaCl, apparaissant après un mois d'incubation. La sensibilité face au sel chez cette moisissure varie
en fonction du milieu de culture utilisé. Sur caillé modèle salé à 8 % de NaCl, quelques colonies ont été
observées après plus de 11 jours (Sansonetti, 1930). Son optimum de croissance a été observé à une
concentration variant entre 1 et 2 % de NaCl sur milieu solide et en absence de sel lorsque cultivée en milieu
liquide (Choisy et al., 2009). La croissance de P. camemberti sur fromages Camemberts salés entre 0,20 et
2,49 % de NaCl a montré un développement faible et inégal du mycélium après quatre semaines d'affinage
pour des concentrations inférieures à 0,70 %. Parallèlement, après cinq semaines, la protéolyse s'est avérée
plus importante entre 0,20 et 1,14 % de NaCl et moins prononcée entre 1,73 et 2,49 % (O'Nulain, 1986).
1.3.5.2. Comportement de G. candidum en fonction du sel
G. candidum est reconnue pour sa sensibilité relative face au stress salin. Cependant, le comportement de
cette espèce en présence de NaCl montre une grande variabilité en fonction des souches étudiées. Selon
différentes études, les observations faites sur milieu de culture synthétique montrent une croissance optimale
variable se situant à 0 et 1 % de NaCl et une réduction importante du taux de croissance à 3 et 4 %
(Sansonetti, 1930, Marcellino et al., 2001). D'autres ne démontrent aucune croissance à 4 % de NaCl (van
den Tempel et Nielsen, 2000). Dans des fromages expérimentaux, 8 % de NaCl ont permis d'empêcher la
croissance de G. candidum tandis que P. camemberti s'est développée à des concentrations largement
supérieures, même à 20 % de NaCl (Sansonetti, 1930). À l'inverse, un salage insuffisant avantage la
croissance de G. candidum au détriment de P. camemberti ce qui peut causer des défauts de fromagerie dont
le défaut nommé « peau de crapaud » (Spinnler et Gripon, 2004). La tolérance au sel chez G. candidum varie
en fonction de l'origine de la souche. Il semblerait que certaines souches isolées à partir de fromages ont été
sélectionnées pour résister à des concentrations salines plus élevées. À titre d'exemple, la croissance de 60
isolats provenant de fromages Armada a été possible en présence de 5 % de NaCl (Tornadijo et al., 1998).

36
1.3.6. Le suivi de croissance des mycètes pendant l'affinage
Le suivi de croissance de la microflore fongique des fromages a principalement été réalisé par méthode de
microbiologie classique, soit par dénombrement sur milieux de culture synthétique (Guizani et al., 2002,
Leclercq-Perlat et al., 2004, Leclercq-Perlat et al., 2006). Il s'agit de méthodes simples d'exécution, mais
comportant toutefois plusieurs désavantages. Entre autres, elles sont fastidieuses compte tenu de leur
manutention élevée et du temps y étant alloué. Non seulement, certaines espèces microbiennes sont sous-
estimées ou ne peuvent tout simplement pas être détectées dû à leur incapacité de croître en condition in vitro
ou à la dominance et la capacité d'envahissement des autres espèces présentes, dont les moisissures (Jany
et Barbier, 2008). Ces méthodes de culture traditionnelle sont d'autant plus problématiques chez les
moisissures puisque seul les spores et certains amas d’hyphes sont dénombrés (Leclercq-Perlat et al., 2004,
Leclercq-Perlat et al., 2006). Ainsi, la population réelle de ces mycètes s'avère sous-estimée et l'évolution
réelle de ces microorganismes pendant l'affinage ne correspond pas au profil observé sur milieu gélosé (Le
Drean et al., 2010, Lessard et al., 2012, Leclercq-Perlat et al., 2013). Il devient donc difficile d'établir une
corrélation adéquate entre le développement de ces microorganismes pendant l'affinage et les changements
biochimiques observés dans l'aliment. Depuis quelques années, les méthodes de biologie moléculaire sont de
plus en plus exploitées afin d'étudier les communautés microbiennes de divers types fromagers (Jany et
Barbier, 2008). Le PCR en temps réel (qPCR) a, entre autres, été utilisé afin d'étudier l'écosystème fongique
de fromages à pâte molle et à croûte fleurie (Le Drean et al., 2010, Lessard et al., 2012, Leclercq-Perlat et al.,
2013). La quantification par qPCR est possible grâce à l'amplification d'un gène spécifique présent dans le
génome de l’espèce d'intérêt. Plusieurs réactions de qPCR ayant des cibles différentes peuvent être réalisées
sur la même extraction d’ADN d’un écosystème afin de quantifier plusieurs espèces microbiennes. Le choix du
gène d'intérêt permet une quantification spécifique et donc, la discrimination de chaque espèce.
L'utilisation du qPCR comporte plusieurs avantages. Il est, entre autres, reconnu pour sa rapidité, sa
sensibilité, sa spécificité et son degré d'automatisation (Wolffs et al., 2005). Pour ce qui est de l'écosystème
des fromages à pâte molle et à croûte fleurie, il a permis de comprendre davantage la dynamique de
croissance des ferments fongiques, surtout dans les débuts de l'affinage, ou d'y confirmer leur rôle. À titre
d'exemple, Le Dréan et coll. (2010), ont suivi la croissance de P. camemberti dans des fromages industriels de
type Camembert (Coulommiers et Carré). Ils ont pu confirmer le développement actif de cette espèce dans les
tout premiers jours d'affinage, contrairement à ce qui a été observé par méthode de microbiologie classique
(Leclercq-Perlat et al., 2004). La croissance du mycélium a été constatée à partir du deuxième jour d'affinage
avec un taux maximal entre les jours 2 et 4 alors que Leclercq-Perlat et coll. (2004) n'avait observé aucune
variation de croissance durant les 14 premiers jours d'affinage sur milieu synthétique. L'utilisation de cette

37
technique permet donc une meilleure estimation de la biomasse fongique réelle puisque l'ADN présent dans
les spores et les hyphes est quantifié.
Lessard et coll. (2012) ont déterminé le profil de croissance de plusieurs ferments fongiques, soit
P. camemberti, G. candidum, K. lactis et D. hansenii, dans des caillés modèles de type Camembert. Tout
comme Le Dréan et coll. (2010), ils ont observé une phase active de croissance chez P. camemberti en début
d'affinage, soit durant les six premiers jours. Ils ont de plus constaté que P. camemberti et G. candidum
devenaient rapidement les espèces majoritaires parmi cet écosystème, que l'ajout de la levure D. hansenii
provoquait une réduction des comptes chez les trois autres espèces étudiées et que la biomasse déterminée
par qPCR était beaucoup plus importante que les comptes déterminés sur milieu gélosé. De plus, comme le
démontre la figure 1.8, l'utilisation du qPCR a permis de détecter la croissance de K. lactis tout au long de
l'affinage tandis qu'aucune croissance n'a été possible sur milieu gélosé après 14 jours, ce qui est
probablement dû à la présence dominante de P. camemberti et G. candidum qui empêche une quantification
exacte.
Figure 1.8 – Quantification de P. camemberti ( ▲), G. candidum (□), K. lactis (●) pendant l'affinage de CMC par dénombrement sur gélose (gauche) et par qPCR (droite). Le symbole (×) correspond à la biomasse (poids sec) du mycélium total (Figure adaptée de Lessard et coll., 2012).
Leclercq-Perlat et coll (2013) ont également étudié la dynamique de croissance de P. camemberti dans des
fromages de type Camembert par qPCR et par méthode de culture classique. Comme il a été déterminé dans
les études connexes, le qPCR a permis d'observer une phase active de croissance dans les débuts de
l'affinage, débutant après seulement un jour. Ensuite, une légère réduction du taux de croissance est
observée, ce qui a été corrélé avec l'épuisement du lactose dans la croûte du fromage et le début de la
sporulation. Le suivi de croissance par méthode de culture classique a permis de confirmer le moment où la
sporulation a débuté.

38
Ainsi, l'utilisation du qPCR a permis d'éclaircir plusieurs aspects en lien avec la dynamique de croissance des
ferments fongiques et certains phénomènes biochimiques pendant l'affinage du fromage. Dans un contexte
industriel et une fois mise au point, cette technique pourrait permettre un meilleur contrôle des populations
microbiennes et être utile, par exemple, à la détection de microorganismes d'altération et ainsi éviter certains
problèmes lors de l'affinage ou simplement s'assurer du bon déroulement de celui-ci.
Bien que le qPCR possède plusieurs attributs, certains désavantages y sont toutefois associés. Entre autres, il
s'agit d'une méthode complexe qui demande un certain niveau expertise lors de sa mise au point, de son
exécution ainsi que de l'analyse des résultats générés. De plus, certains biais peuvent être causés par la
présence d'inhibiteurs de réactions PCR provenant de la matrice fromagère ou par un mauvais rendement
d'extraction d'ADN, parfois associé à une lyse cellulaire non complète (Jany et Barbier, 2008, Le Dréan et al.,
2010). Finalement, une surestimation de la population microbienne peut être causée par la détection d'ADN
provenant de cellules mortes (Wolffs et al., 2005).

39
1.4. LA RÉDUCTION DU SODIUM DANS LES FROMAGES
1.4.1. Les cibles de réduction
Au Canada, les cibles de réduction du sodium suggérées par Santé Canada (section 1.1.3.1) ont été établies
dans la plupart des aliments en réduisant de 25 à 30 % le contenu en sodium au sein d'une catégorie
alimentaire, celui-ci établie en fonction d'une moyenne pondérée du volume de ventes (Santé Canada, 2012).
Dans le cas du Camembert et du Brie, la cible proposée pour 2016 est de 670 mg de Na/100 g alors que le
contenu moyen des fromages de cette catégorie a été estimé à 643 mg de Na/100 g en fonction du volume
des ventes de 2009 et de 2010 (Santé Canada, 2012). Le problème est que, les cibles de réduction établies
pour les fromages regroupent plusieurs variétés dont le contenu en sodium est grandement variable. Ainsi, en
se basant sur les repères établis par Santé Canada, la cible de réduction est déjà atteinte pour plusieurs
variétés de fromages Camembert et Brie. Cependant, les efforts de réduction du sodium dans les fromages à
pâte molle et à croûte fleurie sont tout de même justifiables puisque les fromages de spécialité, comme le
Camembert, sont réputés comme étant très salés et sont peu recommandés par les nutritionnistes. Par
ailleurs, pour bénéficier d'une allégation « teneur réduite en sodium » selon la législation canadienne, le
fromage doit montrer une réduction d'au moins 25 % de son contenu en sodium par rapport au même fromage
ayant une teneur régulière (CTAC, 2009).
1.4.2. Les stratégies de réduction du sodium dans les fromages
À ce jour, plusieurs stratégies permettant de réduire le contenu en sodium des fromages ont été explorées
(Reddy et Marth, 1991, Guinee et Fox, 2004, Guinee et O’Kennedy, 2007, Doyle et Glass, 2010, Cruz et al.,
2011). Puisque la majorité du sodium présent dans les fromages provient de l'ajout de NaCl pendant le
procédé de transformation (Johnson et al., 2009), les stratégies de réduction étudiées sont principalement
basées sur la réduction ou la substitution de ce composé. Les principales approches sont les suivantes :
Une réduction par palier du contenu en NaCl.
Une réduction partielle ou complète du contenu en NaCl et son remplacement par d'autres sels (KCl,
MgCl2 ou CaCl2).
Une réduction du contenu en NaCl et un remplacement par des exhausteurs de saveurs (ex: extraits
de levures).
Des modifications des procédés de transformations, par exemple :
- Altération des teneurs en minéraux (Ca et P) par augmentation du contenu protéique du lait
de fromagerie via l'ajout de rétentat de lait ultrafiltré.

40
- Centrifugation du lait, stérilisation, réincorporation de microorganismes et réduction du
temps de saumurage.
- Fabrication de fromage sans sel ajouté par délactosage et cuisson du caillé moulé.
Néanmoins, ces différentes stratégies ne s'avèrent pas toutes fructueuses étant donné les multiples rôles
joués par le NaCl. Ainsi, il devient difficile de générer un produit identique à celui d'origine. Par exemple, une
réduction nette du contenu en NaCl sans substitution doit être faite par paliers afin que le consommateur
s'habitue au goût moins salé du produit. Ce processus doit donc s'étaler sur une longue période de temps
(Dötsch et al., 2009) et nécessite donc l’optimisation de la formulation du fromage pour chacun de ces paliers.
De plus, une étude portant sur le Cheddar a révélé que la perception du goût salé s'avérait être l'attribut le
plus important pour le consommateur tandis qu'un fromage non salé a été considéré insipide (Schroeder et al.,
1988). En fait, le NaCl est reconnu pour son goût salé, mais aussi car il possède la propriété de masquer
certaines saveurs indésirables, dont l'amertume et le goût métallique. Pour l'instant, aucun composé ne peut
remplacer de façon équivalente la saveur du NaCl (Reddy et Marth, 1991, Doyle et Glass, 2010).
Bien que la saveur soit un attribut important, les autres rôles fonctionnels joués par le NaCl, soit le contrôle de
la texture, de la croissance et l'activité microbienne, ainsi que de l'innocuité, doivent également être satisfaits.
Pour l'instant, l'approche la plus étudiée dans les fromages, et qui semble la plus réussie, est la réduction
partielle du contenu en NaCl combiné à un remplacement par du chlorure de potassium (KCl) (Reddy et
Marth, 1991, Hoffmann, 2014). Malheureusement, la législation canadienne ne permet pas encore son
utilisation à titre d'additif alimentaire dans le but de remplacer le NaCl (Santé Canada, 2006). Cependant, les
autorités de Santé Canada semblent prêtes à accepter de nouveaux ingrédients ou de nouvelles applications
d’ingrédients, si ceux-ci permettent de réduire l’apport quotidien en sodium dans la population et sont
démontrés comme sécuritaires.
Pour ce qui est des autres sels de remplacement tels que le CaCl2 et le MgCl2, leur utilisation seul ou par
remplacement partiel s'est avérée affecter la texture et/ou le goût de l'aliment (Fitzgerald et Buckley, 1985,
Grummer et al., 2012). Dans l'étude de Fitzgerald et Buckley (1985), des fromages Cheddar salés avec du
MgCl2 ou du CaCl2 seulement étaient totalement inacceptables dû à une amertume beaucoup trop prononcée.
De plus, la lipolyse et la protéolyse s'est avérée plus importante. Dans cette même étude, ainsi que celle de
Grummer et coll. (2012), des fromages Cheddar salés avec un mélange de NaCl/MgCl2 ou NaCl/CaCl2 étaient
moins appréciés que les contrôles (NaCl seulement) dû à la détection de saveurs indésirables dont
l'amertume et la perception d'un goût métallique. Dans ces deux études, le pH s'est avéré plus bas dans les
fromages contenant un mélange de NaCl/MgCl2 et de NaCl/CaCl2 suggérant que ces sels de remplacement
possèdent une activité inhibitrice inférieure sur les ferments lactiques comparativement au NaCl. De plus, il

41
semblerait que le calcium et le magnésium augmentent plus fortement l'hydratation des caséines que le
sodium, ce qui est d'avantage vrai pour le magnésium. De ce fait, la synérèse est désavantagée ce qui peut
expliquer la teneur en humidité plus élevée dans les fromages salés en présence de MgCl2.
1.4.2.1. Utilisation du KCl à titre de sel de remplacement
1.4.2.1.1. Le KCl et la santé humaine
Le potassium est un élément essentiel au corps humain. Il s'agit du cation intracellulaire le plus abondant; il
contribue aux diverses fonctionnalités des cellules. Les hommes canadiens consomment en moyenne entre
3,2 et 3,4 g de potassium/jour alors que les femmes en consomment entre 2,4 et 2,6 g/jour. L'apport en
potassium chez les Canadiens est donc insuffisant puisque l'AS a été fixé à 4,7 g/jour (IOM, 2005).
Le potassium est reconnu pour ses bienfaits sur la santé humaine. Une consommation élevée de cet élément
est associée à une réduction de la tension artérielle, ce qui est principalement visible chez les individus ayant
une diète riche en sodium. De plus, il existe une relation inverse entre la consommation en potassium et les
maladies cardiovasculaires. Sa consommation est également liée à la santé des os. En fait, il existe une
corrélation positive entre l'apport en potassium et la rétention du calcium dans les os, ce qui préviendrait le
risque de développement d'ostéoporose (He et MacGregor, 2008, Weaver, 2013). Bien que le potassium soit
bénéfique pour la santé humaine, certaines personnes souffrant de problèmes rénaux doivent surveiller leur
apport en cet élément. Lorsque l'excrétion du potassium par les reins ne se fait pas adéquatement, celui-ci est
accumulé dans l'organisme ce qui peut provoquer l'hyperkaliémie et, par la suite, l'arythmie cardiaque (He et
MacGregor, 2008).
1.4.2.1.2. Le KCl dans les fromages
Plusieurs études ont démontré la possibilité de générer des fromages de qualité acceptable, c'est-à-dire au
niveau de la texture et du goût, en substituant une portion du NaCl par du KCl. Ces études concernaient les
variétés de fromages Cheddar (Fitzgerald et Buckley, 1985, Grummer et al., 2012), le Feta (Aly, 1995, Katsiari
et al., 1997), le Kefalograviera (Katsiari et al., 1998), un fromage frais (Mina's fresh cheese) (Gomes et al.,
2011) et la Mozzarella (Thibaudeau et al., 2015). Selon la variété fromagère, ces substitutions variaient entre
25 et 50 %. Néanmoins, bien que les fromages contenant un mélange NaCl/KCl soient jugés acceptables, les
fromages de contenu en NaCl standards sont généralement préférés. En fait, le potassium est connu pour
apporter un goût métallique et une amertume, ce qui fait en sorte qu'il ne peut remplacer complètement le
NaCl (Guinee et Fox, 2004, Doyle et Glass, 2010, Hoffmann, 2014). Dans certaines variétés de fromages,
seulement 1 % p/p de KCl sont nécessaires pour que l'amertume y étant associée soit perceptible (Guinee et
Fox, 2004).

42
LeKCletlecomportementmicrobien
L'effet du KCl sur la croissance des ferments fongiques, tels que P. camemberti et G. candidum, n'est pas
documenté. Néanmoins, une étude portant sur la croissance des moisissures P. roqueforti et Aspergillus niger
sur milieu de culture PDA (Potato Dextrose Agar) révèle qu'à la même molalité, le NaCl et le KCl modulent le
taux de croissance et la durée de la phase de latence de façon équivalente (Samapundo et al., 2010). En fait,
l’effet du NaCl et du KCl sur l'aw, à condition de les comparer à la même molalité, serait équivalent dans le
milieu de culture PDA. Puisque la capacité du sel à réduire l'aw est une caractéristique majeure responsable
du contrôle de la croissance microbienne, ces observations pourraient expliquer le comportement similaire de
ces moisissures en fonction des deux sels. Ces mêmes auteurs ont également démontré que le remplacement
de 30 % du NaCl dans un pain blanc par un mélange de KCl et de Sub4Salt (NaCl, KCl et gluconate de
sodium) n'affectait pas le taux de croissance ni la phase de latence de P. roqueforti.
Au niveau bactérien, des observations similaires ont pu être réalisées. En milieu de culture liquide, le
comportement d'espèces pathogènes, dont L. monocytogenes et S. aureus, s'est avéré similaire que ce soit
pour le NaCl ou son équivalent molaire en KCl (Boziaris et al., 2007, Bidlas et Lambert, 2008). Dans des
fromages Cheddar contenant différents ratios de NaCl et de KCl (2:1, 1:1, 1:2 et 3:4), les comptes de bactéries
aérobies totales, de ferments lactiques, de bactéries lactiques de la microflore secondaire, de coliformes, de
levures et moisissures et de spores aérobies se sont avérés similaires aux fromages contrôles (NaCl
seulement) (Reddy et Marth, 1995).
LeKCletlesaumurage
La plupart des études traitant des mécanismes de diffusion du sel lors du saumurage ont été réalisées en
présence de NaCl seulement et non d'un mélange de NaCl et de KCl. Dans le cas du Camembert, les
mécanismes de prise en sel lors du saumurage en présence de ces deux composés n'ont pas été publiés. Or,
les études disponibles pour d'autres variétés de fromages démontrent que le potassium pénètre dans la
matrice fromagère par un processus de diffusion, tout comme le sodium (Zorrilla et Rubiolo, 1994ab, Bona et
al., 2007). Plus la teneur en KCl des saumures est élevée, plus la quantité de potassium dosée dans les
fromages l'est également, ce qui a été observé dans la Mozzarella (Thibaudeau et al., 2015), le Kefalograviera
(Katsiari et al., 1998), le Halloumi (Ayyash et Shah, 2010) et le Nabulsi (Ayyash et Shah, 2011). Même si le
sodium et le potassium sont similaires au point de vue de leur propriété et de leur taille (Zorrilla et Rubiolo,
1994a), ils pourraient avoir une vitesse de diffusion différente. À titre d'exemple, le coefficient de diffusion du
KCl dans un fromage à pâte semi-ferme (fromage Prato) serait plus élevé que celui du NaCl (Bona et al.,
2007).

43
1.5. PROBLÉMATIQUE, HYPOTHÈSE ET OBJECTIFS
Problématique
Chez l'humain, un apport en sodium élevé au quotidien augmente les risques d’hypertension artérielle et, par
conséquent, de plusieurs autres problèmes de santé associés. Ainsi, la réduction du chlorure de sodium dans
les fromages, dont le Camembert, est recommandée par Santé Canada. Or, le sel exerce plusieurs rôles
majeurs dans les fromages, dont la modulation de la croissance et de l'activité microbienne. La microflore
fongique de surface des Camemberts contribue largement au développement des propriétés sensorielles
pendant l'affinage. Modifier la concentration saline à laquelle ces mycètes sont normalement exposés pourrait
entraîner des défauts de texture et de saveurs. Les impacts pouvant résulter de ces modifications doivent
donc être déterminés.
Hypothèse
La réduction du NaCl dans le Camembert modulera la croissance de Penicillium camemberti et Geotrichum
candidum et sa substitution partielle par le KCl rétablira le profil de croissance initial des microorganismes
dans un Camembert.
Objectif général
Déterminer l’effet de la réduction nette du NaCl et de sa substitution partielle par KCl sur la croissance de la
microflore fongique d’affinage du Camembert.
Objectifs spécifiques
1. Évaluer la cinétique d'incorporation du sodium et du potassium dans un Camembert industriel saumuré.
1.1. Déterminer les paramètres de saumurage permettant d'obtenir un fromage Camembert de
composition saline standard.
1.2. Déterminer les paramètres de saumurage permettant d'obtenir un fromage Camembert dont le
contenu en sodium est réduit de 30 %.
1.3. Déterminer les paramètres de saumurage permettant d'obtenir un Camembert dont 30 % du contenu
en sodium est substitué sur une base équimolaire par du potassium.
2. Déterminer la variabilité intraspécifique de la résistance au NaCl et au KCl chez P. camemberti.
2.1. Évaluer l'effet du stress salin sur P. camemberti en milieu de culture synthétique.
2.2. Évaluer l'effet du stress salin sur P. camemberti en caillé modèle de type Camembert.

44
3. Évaluer la croissance de P. camemberti et G. candidum à la surface d'un Camembert industriel en
fonction de différentes teneurs salines.

45
2. Étude de la cinétique d'incorporation du
sodium et du potassium dans un fromage
Camembert industriel saumuré


47
2.1. RÉSUMÉ
L'association entre une consommation excessive en sodium et l'hypertension a mené à la réduction du sel
dans plusieurs aliments, dont les fromages. L'utilisation de KCl à titre de sel substitut s'avère être la stratégie
la plus commune et la plus réussie permettant de réduire partiellement la teneur en sodium dans les fromages.
La cinétique d'incorporation du sodium et du potassium a été étudiée dans un fromage Camembert industriel
de type stabilisé salé en saumure saturée en NaCl (26 % (p/p)) ou en utilisant différents ratios de NaCl et de
KCl (4:1, 7:3, 3:2, 1:1). Le but de cette étude était d'identifier les paramètres de saumurage (temps et
composition saline des saumures) permettant d'obtenir un fromage de teneur en sodium standard (A), réduit
de 30 % en sodium (B) et réduit de 30 % en sodium avec une substitution équimolaire par du potassium (C).
Les teneurs en sodium et en potassium présentes dans les fromages Camemberts, à la suite du saumurage,
ont été déterminées par spectrométrie d'émission à plasma induit (ICP-OES) après quatre jours d'affinage.
Les résultats ont démontré qu’il était possible de produire un fromage ayant une teneur en sodium équivalente
au standard de l’industrie après salage en saumure sans modification du temps de saumurage (70 minutes)
en évitant de surcharger le bac de saumure. Un fromage réduit de 34,8 % (g Na/g de fromage) de sodium a
été obtenu par salage en saumure saturée après 20 minutes de saumurage. Le salage en saumure mixte
(ratio 4NaCl:1KCl) a permis de produire un fromage dont le sodium a été partiellement substitué par du
potassium (22,3 % de réduction en sodium) après 50 minutes de saumurage. Ainsi, les paramètres permettant
d'obtenir les trois cibles salines visées ont pu être identifiés, mais la réduction obtenue dans les fromages
substitués par du potassium était plus faible que désirée.

48
2.2. INTRODUCTION
L'apport quotidien en sodium est trop élevé pour la quasi-totalité de la population mondiale (Brown et al.,
2009). Son excès dans l'alimentation humaine a été associé à l'hypertension artérielle, celle-ci étant un facteur
de risque du développement de maladies cardiovasculaires (Strazzullo et al., 2009, Whelton et He, 2014). De
nombreux pays ont aujourd'hui entrepris des efforts afin de réduire l'apport quotidien en sodium au niveau de
leur population respective (Webster et al., 2011). Leur stratégie vise particulièrement les aliments transformés
qui contribuent pour 77 % de l’apport quotidien en sodium au Canada. En ce qui concerne les fromages, ceux-
ci s'avèrent être contributeurs de 5 % de l'apport quotidien en sodium au Canada. Étant donné que la majorité
du sodium retrouvé dans les fromages provient de l'ajout de sel pendant le procédé de fabrication (Johnson et
al., 2009), sa réduction y est envisagée. Au Canada, la stratégie de réduction du sodium encourage les
industries alimentaires à réduire la teneur en sel dans la plupart des catégories fromagères, incluant les
fromages à pâte molle de type Camembert (Santé Canada, 2012).
Le salage des fromages à pâte molle et à croûte fleurie est généralement réalisé par saumurage (Sousa,
2003). Les mécanismes intervenant au niveau de la diffusion du sel et de la cinétique de son absorption ont
été étudiés dans plusieurs variétés fromagères telles que le Gouda (Geurts et al., 1974), le Camembert
(Hardy, 1976), l'Emmental (Chamba, 1988), le Cuartirolo (De Piante et al., 1990), le Romano (Guinee et Fox,
1986a) et le Rugasano (Melilli et al., 2003a, Melilli et al., 2003b, Melilli et al., 2005, Melilli et al., 2006). Lorsque
les saumures sont agitées et homogénéisées constamment, le sel diffuse dans les fromages selon un
mécanisme dérivant de la loi de diffusion de la matière (loi de Fick), ce qui a préalablement été validé dans le
cas du fromage Camembert (Hardy, 1976). La prise en sel lors du saumurage est spécifique à chaque
fromage puisqu'elle est dépendante des caractéristiques intrinsèques de chacun. Ces caractéristiques
correspondent, entre autres, à la géométrie de l'aliment, sa teneur en eau initiale ainsi qu'à la structure de la
pâte (porosité) (Geurts et al., 1974, 1980, Guinee et Fox, 1986b, Chamba, 1988, Melilli et al., 2003b, Guinee
et Fox, 2004). Outre les éléments reliés aux fromages, la prise en sel est également influencée par les
conditions de saumurage telles que la concentration en sel des saumures, l'agitation et la température (Geurts
et al., 1974, Hardy, 1976, Geurts et al., 1980, Chamba, 1988, Melilli et al., 2003a, Melilli et al., 2003b, Guinee
et Fox, 2004, Melilli et al., 2005, Melilli et al., 2006). Les études réalisées dans le fromage Camembert
démontrent que l'incorporation du sel en fonction du temps de saumurage n'est pas linéaire (Hardy, 1976). En
fait, sa vitesse d'incorporation ralentit en fonction du temps de saumurage en conséquence de la réduction du
gradient en sel formé entre la saumure et le fromage, au cours du temps. Ainsi, dans l'optique où une
réduction du contenu en sodium des fromages doit être réalisée, il devient difficile d'estimer le temps de
saumurage requis afin d'obtenir la cible visée.

49
Autre que la réduction nette du contenu en sel, plusieurs stratégies permettant de réduire la teneur en sodium
dans les aliments ont été proposées telles que l'utilisation de sels substituts (KCl, MgCl2, CaCl2) (Guinee et
Fox, 2004). La substitution partielle du contenu en NaCl par du KCl s'avère être la méthode la plus utilisée et
la plus réussie (Reddy et Marth, 1991, Hoffmann, 2014). De plus, le potassium est reconnu pour ses bienfaits
au niveau de la santé humaine, notamment au niveau de la tension artérielle, de la santé osseuse et du
métabolisme du calcium (Weaver, 2013). Par ailleurs, le potassium est habituellement consommé en trop
faible quantité sur une base journalière (IOM, 2005). Le goût désagréable (amertume et métallique) associé au
potassium limite cependant son utilisation; une substitution d'au plus 30 % générerait des fromages de qualité
acceptable dans plusieurs variétés (Guinee et Fox, 2004, Hoffmann, 2014). Quelques études ont démontré
qu'il est possible de produire des fromages contenant un mélange de NaCl et de KCl en substituant une
portion du NaCl de la saumure par du KCl (Aly, 1995, Katsiari et al., 1997, 1998, Ayyash et Shah, 2010,
Ayyash et Shah, 2011, Thibaudeau et al., 2015). La diffusion du sel dans des fromages à pâte semi-ferme lors
du salage en saumures composées de mélange de NaCl et de KCl a préalablement été étudiée (Zorrilla et
Rubiolo, 1994b, Bona et al., 2007, Bockelmann, 2010). Or, peu d'études portant sur la diffusion de ces deux
sels pendant le saumurage du Camembert ont été réalisées.
L'objectif de l'étude était d'identifier les paramètres de saumurage permettant de produire un fromage
Camembert de composition en sodium correspondant au standard industriel utilisé (STD), réduit de 30 % en
sodium (RED) ou réduit de 30 % en sodium, mais substitué de façon équimolaire par du potassium (KCL).

50
2.3. MATÉRIEL ET MÉTHODES
2.3.1. Matériel à l'étude
Les Camemberts utilisés provenaient d'une entreprise canadienne de production de fromages de spécialité qui
désire demeurer anonyme. Les fromages étaient de forme cylindrique (diamètre: 20 cm, hauteur: 3 cm) et
avaient un poids de 870 g. Ils ont été produits à partir de lait pasteurisé selon un procédé de fabrication de
type industriel dont les étapes demeurent confidentielles. La coagulation a été faite par ajout d'une
combinaison de ferments lactiques thermophiles et mésophiles afin d'obtenir une pâte de type stabilisée. Les
expériences de saumurage ont été réalisées sur place, en industrie. Quatre conditions salines dans les
Camemberts étaient ciblées dans cette étude; une condition en sodium standard (STD) équivalente à celle
normalement retrouvée dans ce fromage après le saumurage en industrie réalisé dans les mêmes
équipements que les autres fromages expérimentaux, une réduction nette de 30 % du contenu en sodium
(RED), une réduction de 30 % du contenu en sodium substitué de façon équimolaire par du potassium (KCL)
et des fromages saumurés dans le bassin de saumure de l’entreprise (IND). Le but de cette étude était de
déterminer les paramètres de saumurage (temps et compositions salines des saumures) permettant d'obtenir
les cibles salines (STD), (RED) et (KCL).
2.3.2. Préparation des saumures
Chaque saumure a été préparée à partir d'eau potable, leur composition en NaCl (Hi-Grade Granulated Salt,
Windsor Salt, Pointe-Claire, Canada) et KCl (Potassium chloride USP/FCC, Thermo Fisher Scientific,
Waltham, États-Unis) sont présentées au tableau 2.1. Le CaCl2 a été ajouté à une concentration finale de 1 %
à l'aide d'une solution de CaCl2 32 % (p/v) (FDC Industries) et le pH a été ajusté à 5,1 ± 0,1 par ajout d'acide
lactique 80 % (FCC solution, Les Produits Chimiques Amplex Ltée, Pointe-Claire, Canada) 24 heures après
leur confection. Elles ont été entreposées à 12 °C ± 2 °C et brassées à tous les jours afin de dissoudre les
sels. Les saumures ont été fabriquées au moins 48 heures avant le saumurage afin de laisser les sels se
solubiliser. Contrairement à la saumure industrielle, les saumures expérimentales ont été confectionnées à
petite échelle dans des bassins statiques (non agités) dont le poids total variait entre 30 et 150 kg et chacune
des saumures n'a été utilisée qu'à une seule reprise.
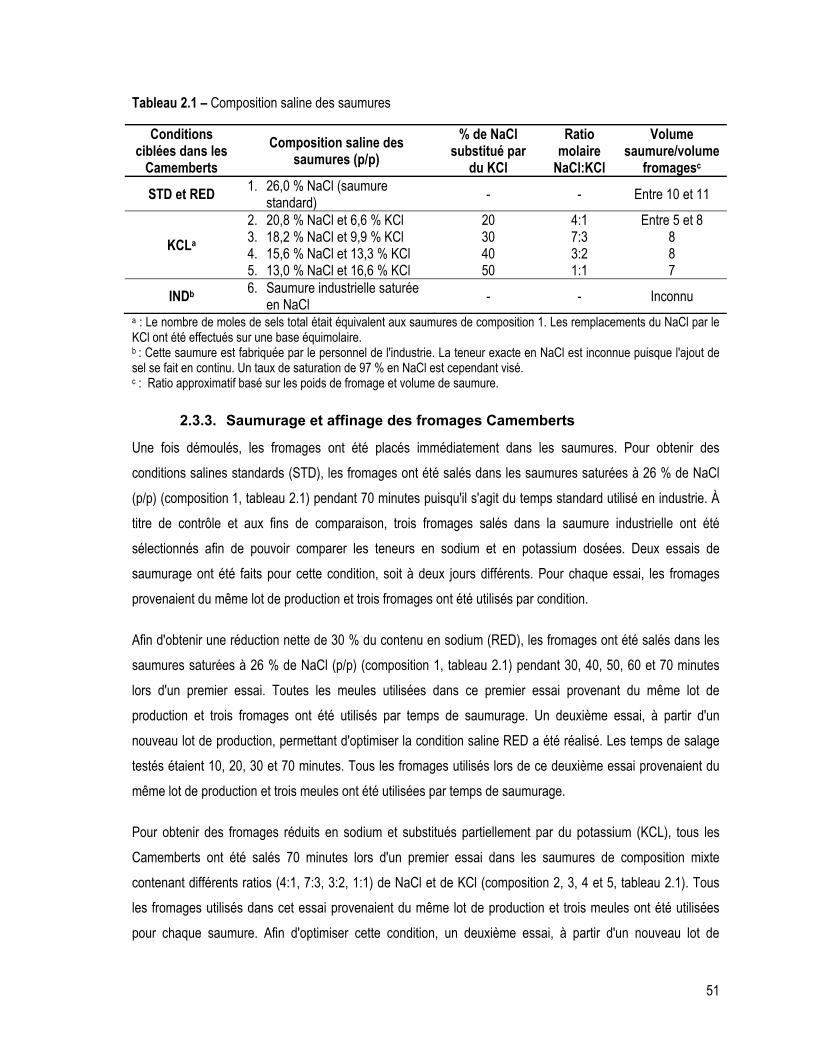
51
Tableau 2.1 – Composition saline des saumures
Conditions ciblées dans les
Camemberts
Composition saline des saumures (p/p)
% de NaCl substitué par
du KCl
Ratio molaire
NaCl:KCl
Volume saumure/volume
fromagesc
STD et RED 1. 26,0 % NaCl (saumure
standard) - - Entre 10 et 11
KCLa
2. 20,8 % NaCl et 6,6 % KCl 20 4:1 Entre 5 et 8 3. 18,2 % NaCl et 9,9 % KCl 30 7:3 8 4. 15,6 % NaCl et 13,3 % KCl 40 3:2 8 5. 13,0 % NaCl et 16,6 % KCl 50 1:1 7
INDb 6. Saumure industrielle saturée en NaCl
- - Inconnu
a : Le nombre de moles de sels total était équivalent aux saumures de composition 1. Les remplacements du NaCl par le KCl ont été effectués sur une base équimolaire. b : Cette saumure est fabriquée par le personnel de l'industrie. La teneur exacte en NaCl est inconnue puisque l'ajout de sel se fait en continu. Un taux de saturation de 97 % en NaCl est cependant visé. c : Ratio approximatif basé sur les poids de fromage et volume de saumure.
2.3.3. Saumurage et affinage des fromages Camemberts
Une fois démoulés, les fromages ont été placés immédiatement dans les saumures. Pour obtenir des
conditions salines standards (STD), les fromages ont été salés dans les saumures saturées à 26 % de NaCl
(p/p) (composition 1, tableau 2.1) pendant 70 minutes puisqu'il s'agit du temps standard utilisé en industrie. À
titre de contrôle et aux fins de comparaison, trois fromages salés dans la saumure industrielle ont été
sélectionnés afin de pouvoir comparer les teneurs en sodium et en potassium dosées. Deux essais de
saumurage ont été faits pour cette condition, soit à deux jours différents. Pour chaque essai, les fromages
provenaient du même lot de production et trois fromages ont été utilisés par condition.
Afin d'obtenir une réduction nette de 30 % du contenu en sodium (RED), les fromages ont été salés dans les
saumures saturées à 26 % de NaCl (p/p) (composition 1, tableau 2.1) pendant 30, 40, 50, 60 et 70 minutes
lors d'un premier essai. Toutes les meules utilisées dans ce premier essai provenant du même lot de
production et trois fromages ont été utilisés par temps de saumurage. Un deuxième essai, à partir d'un
nouveau lot de production, permettant d'optimiser la condition saline RED a été réalisé. Les temps de salage
testés étaient 10, 20, 30 et 70 minutes. Tous les fromages utilisés lors de ce deuxième essai provenaient du
même lot de production et trois meules ont été utilisées par temps de saumurage.
Pour obtenir des fromages réduits en sodium et substitués partiellement par du potassium (KCL), tous les
Camemberts ont été salés 70 minutes lors d'un premier essai dans les saumures de composition mixte
contenant différents ratios (4:1, 7:3, 3:2, 1:1) de NaCl et de KCl (composition 2, 3, 4 et 5, tableau 2.1). Tous
les fromages utilisés dans cet essai provenaient du même lot de production et trois meules ont été utilisées
pour chaque saumure. Afin d'optimiser cette condition, un deuxième essai, à partir d'un nouveau lot de

52
production, dans des saumures de ratio 4NaCl:1KCl a été fait. Les temps 50, 60 et 70 minutes ont été
expérimentés. Tous les fromages utilisés dans ce deuxième essai provenaient du même lot de production et
trois meules ont été utilisées par temps de saumurage.
Tous les fromages ont été retournés à la moitié du temps de saumurage puisque l'équipement utilisé ne
permettait pas de les submerger totalement. Suite au saumurage, les fromages saumurés et les contrôles ont
été affinés sur des claies pendant 4 jours à 12,5 °C et 96,5 % d'humidité relative. Ils ont ensuite été emballés
et transportés à l'Université Laval où ils ont été emballés sous vide et entreposés à – 20 ºC jusqu'au moment
de leurs analyses. Pour chaque essai, trois fromages étaient utilisés par condition de saumurage.
Étant donné le contexte industriel de ce projet, le nombre d'essais possible était limité. Ainsi dans quelques
expériences présentées dans ce chapitre, le nombre de répétitions est faible (essais préliminaires). Ces
résultats sont tout de même indiqués dans ce mémoire afin de présenter la démarche expérimentale et la
tendance de la cinétique de saumurage, sans toutefois en déterminer les valeurs avec une très grande
précision. Cette étape de criblage de paramètres de saumurage a tout de même permis d'établir les conditions
nécessaires, à la suite de l'optimisation des paramètres, afin de produire les fromages ayant différentes
concentrations de sodium et de potassium.
2.3.4. Analyses de composition des fromages
Les fromages emballés sous vide ont été dégelés dans un bac d'eau froide le matin de leurs analyses. Un
quart de chaque fromage coupé en forme de pointe a été homogénéisé à l'aide d'un mélangeur (type blender).
Pour chaque fromage, la teneur en humidité a été déterminée en triplicata par séchage de 1 g de fromage à
100 °C dans un four à vide (Isotemp 280A, Thermo Fisher Scientific, Waltham, États-Unis) pendant 4 heures
(AOAC, 2008). Les cendres ont été obtenues, en triplicata, après calcination (Thermolyne, Thermo Fisher
Scientific, Waltham, États-Unis) de 2 g de fromage à 550 °C pendant 18 à 24 h ou jusqu'à ce qu'elles soient
de couleur grise-blanche (AOAC, 2005). Elles ont ensuite été solubilisées dans du TCA 20 % (p/p)
(Trichloroacetic acid crystals, ACS, J.T. Baker, Center Valley, États-Unis), dissoutes dans de l'eau
déminéralisée 18 m et filtrées sur membrane d'acétate de cellulose (VWR syringe filter 0,45 µm, VWR
International, Montréal, Canada). Pour chaque fromage, le dosage du sodium et du potassium a été fait en
duplicata par spectrométrie d'émission à plasma induit (Optima 4300TM DV ICP-OES, PerkinElmer, Waltham,
États-Unis) au Laboratoire de pédologie forestière de la Faculté de Foresterie, de Géographie et de
Géomatique de l'Université Laval (Thibaudeau et al., 2015). Exceptionnellement, seulement deux fromages
salés 20 minutes, utilisés lors de l’optimisation de la condition saline B ont pu être analysés dû à la perte d'un
des trois échantillons lors de sa décongélation.

53
2.4. RÉSULTATS ET DISCUSSION
2.4.1. Production d'un Camembert industriel de composition saline standard
Les conditions de saumurage utilisées ont été reproduites à petite échelle afin d'obtenir une teneur en sodium
équivalente au standard fixé par l'industrie. Malgré la reproductibilité des conditions de saumurage
industrielles, certains paramètres n'ont pu être respectés tels que l'agitation des saumures et la submersion
totale des fromages. Puisque ces paramètres étaient susceptibles d'influencer négativement la prise en sel
pendant le saumurage, l'incorporation du sodium dans les fromages salés dans les saumures statiques
expérimentales (saumure standard, 26 % NaCl (p/p)) et dans la saumure industrielle a été comparée (figure
2.1). Les résultats présentés à la figure 2.1 démontrent que les concentrations en sodium dosées dans les
fromages salés dans les saumures expérimentales (STD) et dans la saumure industrielle (IND) étaient
similaires, celles-ci étant respectivement de 4,42 ± 0,14 et 4,62 ± 0,20 mg Na/g de fromage à l'expérience 1 et
de 4,60 ± 0,28 mg Na/g et 4,85 ± 0,16 mg Na/g de fromage à l'expérience 2. Au niveau de la teneur en
humidité et de la teneur minérale, les résultats présentés au tableau 2.2 démontrent également une similitude
entre les traitements. Ainsi, cette partie de l’étude a permis de confirmer qu'aucune modification au niveau du
temps de saumurage sélectionné par l'industrie n'était nécessaire afin d'obtenir la teneur en sodium standard
dans les fromages expérimentaux, salés en saumure statique.
Figure 2.1 – Teneur en sodium dosée après 2 jours (expérience 1) et 4 jours (expérience 2) d'affinage dans des Camemberts saumurés pendant 70 minutes dans une saumure expérimentale statique (26 % NaCl (p/p)) en comparaison avec une saumure agitée (saumure industrielle). Les résultats sont représentés par la moyenne ± l'erreur-type.
0
1
2
3
4
5
6
Saumure expérimentale Saumure industrielle
Sodi
um (m
g/g
de fr
omag
e)
Expérience 1 Expérience 2

54
Tableau 2.2 – Teneur en humidité et en cendres des fromages au 2e et 4e jour d'affinage
Saumurage Humidité (%) Cendres (%)
2e jour 4e jour 2e jour 4e jour
Expérimental (statique) 47,4 ± 0,2 45,6 ± 0,6 2,94 ± 0,04 2,89 ± 0,07 Industriel (agité) 47,3 ± 0,1 45,8 ± 0,2 3,03 ± 0,06 2,94 ± 0,07
Les résultats sont présentés par la moyenne ± l'erreur-type. Tous les fromages ont été saumurés pendant 70 minutes.
Sachant que l'agitation des saumures favorise normalement la prise en sel (Hardy, 1976, Chamba, 1988), une
teneur en sodium significativement plus élevée dans les fromages salés dans la saumure industrielle était
attendue. Dans des conditions de saumurage statique, l'absorption du sel et la perte de lactosérum provenant
du fromage produisent une dilution locale de la saumure en périphérie de l'aliment (Hardy, 1976). Ainsi, les
fromages sont théoriquement exposés à de plus faibles teneurs en sodium, ce qui désavantage la prise en sel.
Bien que les teneurs en sodium dosées après 70 minutes de saumurage étaient similaires, il est impossible
d'affirmer que la cinétique d'incorporation du sodium ait été équivalente au niveau des deux conditions. La
détermination du contenu en sodium aux temps de saumurage inférieurs à 70 minutes n'a été réalisée que
dans les fromages salés dans les saumures statiques et il est prudent d'affirmer que la cinétique aurait pu être
différente pendant la même période dans des fromages en saumure industrielle agitée. De plus, il est possible
que la répartition des sels au sein du fromage (centre vs surface) soit différente, ce qui n'a pas été
déterminée.
Les saumures expérimentales ont été fabriquées de façon à respecter les conditions rencontrées dans la
saumure industrielle. Cependant, la démarche expérimentale utilisée ne permettait pas de tenir compte du
phénomène de vieillissement chimique de la saumure industrielle. L'usage répété de cette saumure contribue
à son enrichissement en minéraux, en matières azotées et en lactose et à l'augmentation de son acidité
(Lacrampe et al., 1971) ce qui entrave la solubilisation du NaCl (Mietton et al., 2004) et réduit le transfert des
solutés (Terré et al., 1985). Ainsi, tel qu'observé lors du saumurage d'un fromage Emmental (Chamba, 1988)
et Camembert (Terré et al., 1985), il est possible que le vieillissement de la saumure industrielle ait défavorisé
la prise en sel. De plus, le ratio de volume de fromage par rapport au volume de saumure entre les deux
conditions n'était pas équivalent. D'ailleurs, il a été observé qu'un volume de saumure cinq fois supérieur au
volume total de fromages devrait être respecté afin d'éviter que ce ratio influence la prise en sel (Bintsis,
2006). Or, en industrie, la quantité de fromage salé dans une même saumure peut varier.
2.4.2. Production d'un Camembert industriel réduit en sodium
Afin d'identifier les paramètres de saumurage permettant d'obtenir un Camembert réduit de 30 % en sodium,
la cinétique d'incorporation de cet élément en fonction du temps de salage a été évaluée (figure 2.2). Les

55
résultats représentent la moyenne des teneurs en sodium quantifiée dans les fromages après quatre jours
d'affinage (vert) ainsi que les proportions de réduction en sodium par rapport au fromage standard saumuré
pendant 70 minutes (violet). Telle qu'attendue, une augmentation de la teneur en sodium des fromages en
fonction du temps de saumurage a pu être observée. La vitesse d'absorption du sodium a ralenti en fonction
du temps de saumurage, ce qui correspond au profil normalement observé dans d'autres études (Hardy, 1976,
Geurts et al., 1980, Guinee et Fox, 1986b, a, Chamba, 1988, De Piante et al., 1990, Melilli et al., 2003a, Melilli
et al., 2003b, Melilli et al., 2006). Ce ralentissement s'explique par la diminution du gradient de concentration
en sel entre la phase aqueuse du fromage et la saumure en fonction du temps (Guinee et Fox, 2004).
Figure 2.2 – Incorporation du sodium dans des fromages Camemberts en fonction du temps de saumurage (vert) et proportions de réduction en sodium obtenues comparativement aux fromages salés pendant 70 minutes (violet). Les fromages ont été salés dans des saumures statiques saturées à 26 % de NaCl (p/p). Puisque les temps de saumurage 0, 30 et 70 minutes ont été expérimentés à deux reprises, les résultats représentés pour ces conditions proviennent de la moyenne obtenue pour ces deux essais. Pour tous les autres temps, les résultats proviennent de seulement un essai de saumurage. Les résultats sont représentés par la moyenne ± l'erreur type.
Les résultats obtenus démontrent que le sodium est incorporé rapidement pendant les 10 premières minutes
de saumurage; le contenu en sodium au 4e jour d'affinage était de 0,61 ± 0,06 mg/g de fromage dans les
échantillons non saumurés et de 2,68 ± 0,24 mg/g de fromage au temps 10 minutes. Ainsi, après 10 minutes,
la teneur en sodium correspondait déjà à 56 % du contenu quantifié en fin de saumurage, soit à 70 minutes.
L'absorption du sodium a ensuite été ralentie entre 10 et 50 minutes pour finalement demeurer relativement
stable jusqu'à 70 minutes où 4,76 ± 0,16 mg/g de fromage ont été quantifiés. Parallèlement, un profil similaire
a pu être observé au niveau de l'évolution de la teneur minérale totale (tableau 2.3), ce qui s'explique par la
prise en sel lors du salage. L'objectif de réduction en sodium de 30 % a pu être atteint après seulement
43,7%
34,8%
23,6%
13,5%
7,0%5,1%
0%
5%
10%
15%
20%
25%
30%
35%
40%
45%
50%
0
1
2
3
4
5
6
0 10 20 30 40 50 60 70
Proportion de réduction en sodium (%)
Sodium (mg/g de fromage)
Temps (minutes)
Sodium (mg/g)
Proportion réduite (%)

56
20 minutes de saumurage correspondant à une réduction équivalente à 34,8 % Na (p/p) (figure 2.2). La
portion industrielle associée à ce projet limitait le nombre d'essais pouvant être faits, ce qui a fait en sorte que
cette condition n'a été testée qu'une seule fois. Elle a tout de même été sélectionnée pour la suite du projet.
Les facteurs générant des variations au niveau du contenu en sel incorporé dans les fromages lors du
saumurage sont connus (Guinee et al., 2014b). Puisque ces variations peuvent être associées à des facteurs
qui n'étaient pas contrôlables, tels que la composition des fromages avant saumurage, il est donc possible que
la cinétique d'incorporation du sel pour la portion subséquente du projet diffère. Les résultats obtenus sont tout
de même utiles à la suite du projet, même si les valeurs obtenues sont approximatives. Ceci justifie donc que
le contenu en sodium doit être déterminé à nouveau pour chaque fromage produit ainsi.
Au niveau de la teneur en humidité des fromages, les résultats obtenus ne correspondaient pas aux résultats
attendus. La prise en sel lors du saumurage est accompagnée d'une perte du contenu en eau du fromage. Le
ratio d'eau perdue et de sel absorbé lors du saumurage est généralement équivalent à 2,5 (Geurts et al.,
1974, Hardy, 1985). Or, les teneurs en humidité sont demeurées relativement stables en fonction du temps de
saumurage (tableau 2.3). Il est possible que le sel incorporé ait créé une barrière en surface empêchant l'eau
de sortir de l'aliment.
Tableau 2.3 – Teneurs en humidité et en minéraux des fromages au 4e jour d'affinage
Temps de saumurage (minutes)
0 10 20 30 40 50 60 70
Hum
idité
(%
) Essai 1 48,1 ±
0,1 - -
49,3 ± 0,3
48,6 ± 0,2
49,0 ± 0,6
48,5 ± 0,5
49,0 ± 0,4
Essai 2 46,8 ±
0,8 46,6 ±
0,3 47,0 ±
0,5 47,0 ±
0,4 - - -
46,2 ± 0,1
Cen
dres
(%
) Essai 1 1,79 ± 0,01
- - 2,66 ± 0,01
2,75 ± 0,03
2,88 ± 0,07
2,93 ± 0,02
3,02 ± 0,06
Essai 2 1,84 ± 0,03
2,33 ± 0,05
2,44 ± 0,03
2,54 ± 0,01
- - - 2,89 ± 0,07
Les résultats sont présentés par la moyenne ± l'erreur-type.
2.4.3. Production d'un Camembert industriel réduit en sodium et enrichie en potassium
2.4.3.1. Premier essai de saumurage
Afin de déterminer les conditions de saumurage permettant d'obtenir un fromage réduit de 30 % de sodium et
substitué par du potassium, l'incorporation de ces deux éléments dans un Camembert salé en saumure mixte
(NaCl et KCl) a été étudiée. La concentration de chaque élément quantifié pour chacune des conditions de
saumurage est présentée au niveau moléculaire (mol/100 g) à la figure 2.3 et sur une base pondérale (mg/g)

57
au tableau 2.4. La présentation des résultats sous forme molaire permet de pouvoir comparer les teneurs en
sodium et en potassium puisque le poids moléculaire de ces deux éléments est différent. Les résultats sont
ensuite présentés sur une base pondérale puisqu'il s'agit de la forme communément utilisée en fromagerie et
par Santé Canada quant aux cibles de réduction saline établies (% de sel). Ensuite, les teneurs en cendres et
en humidité sont présentées au tableau 2.5. Toutes les analyses ont été faites sur des fromages saumurés 70
minutes et affinés pendant quatre jours. Comme le démontre la figure 2.3 et le tableau 2.4, les teneurs en
sodium étaient moins importantes après salage en saumures mixtes comparativement au salage en condition
standard pour une même durée de saumurage. Au contraire, des concentrations plus élevées en potassium
ont été quantifiées dans les fromages salés en saumures mixtes comparativement à la condition standard.
Plus les teneurs en KCl des saumures étaient importantes, plus la concentration en potassium absorbée par
les fromages était élevée. Ces résultats sont en lien avec d'autres études similaires pour les variétés de
fromage Mozzarella (Thibaudeau et al., 2015), Kefalograviera (Katsiari et al., 1998), Halloumi (Ayyash et
Shah, 2010) et Nabulsi (Ayyash et Shah, 2011).
Figure 2.3 – Teneur en sodium et en potassium des fromages Camemberts saumurés 70 minutes dans des saumures statiques fabriquées selon divers ratios de NaCl et de KCl. La saumure sans KCl (100NaCl :0KCl) correspond à saumure standard saturée à 26 % de NaCl (p/p). Le dosage des minéraux a été fait au 4e jour d'affinage. Les résultats sont représentés par la moyenne ± l'erreur type. Les pourcentages correspondent au ratio molaire des éléments sodium et potassium dans chacune des conditions, 100 % correspond au contenu total en sodium et en potassium.
0
0,01
0,02
0,03
4NaCl:1KCl 7NaCl:3KCl 3NaCl:2KCl 1NaCl:1KCl 100NaCl:0KCl Non saumuré
Sodi
um e
t pot
assi
um (m
ol/1
00 g
de
from
age)
Proportion molaire en NaCl et KCl des saumures
K Na
31 %
69 %
35 %
65 %
43 %
57 %
48 %
52 % 91 %
9 %
66 %
44 %

58
Tableau 2.4 – Teneurs en sodium et en potassium des fromages au 4e jour d'affinage et proportion de réduction en sodium correspondante
Saumures (NaCl:KCl) Non
saumuré 4:1 7:3 3:2 1:1
100:0 (standard)
Teneur en sodium (mg/g de fromage)
0,65 ± 0,05 3,83 ± 0,06
3,94 ± 0,09
3,17 ± 0,13
3,19 ± 0,07
4,93 ± 0,14
Teneur en potassium (mg/g de fromage)
0,56 ± 0,02 2,97 ± 0,10
3,58 ± 0,09
4,14 ± 0,19
5,01 ± 0,05
0,88 ± 0,02
Réduction en sodiuma (%)
- 22,3 20,1 35,7 35,3 -
Les résultats correspondent à la moyenne ± l'erreur type. Tous les fromages ont été saumurés pendant 70 minutes. a: Comparativement à la teneur en sodium obtenue dans les fromages saumurés pendant 70 minutes dans la saumure 100NaCl:0KCl.
Les résultats présentés à la figure 2.3 démontrent que les ratios de sodium et de potassium des fromages
différaient des proportions correspondantes dans les saumures, à l'exception de la saumure 1:1 (NaCl:KCl).
Généralement, le taux de potassium incorporé était plus élevé que le ratio présent dans la saumure. De plus,
la quantité totale des deux sels (nombre total de moles de sodium et de potassium dosées) était plus élevée
dans les fromages salés en saumure mixte que celle des fromages salés en présence de NaCl seulement, ce
qui était particulièrement visible au niveau des échantillons salés dans les saumures de ratios 7:3 et 1:1
(NaCl:KCl). Ces résultats suggèrent une pénétration plus rapide du potassium dans la matrice fromagère
comparativement au sodium. Certaines observations en lien avec la diffusion du KCl suggèrent qu'il diffuserait
plus rapidement que le NaCl. À titre d'exemple, Bona et al. (2007) ont déterminé que le coefficient de diffusion
du KCl était plus élevé que celui du NaCl dans un fromage Prato. Aussi, le KCl possèderait une meilleure
mobilité que le NaCl en solution aqueuse (Schwartzberg et Chao, 1982).
Au niveau moléculaire, la proportion de sodium et de potassium obtenue dans les fromages salés dans la
saumure mixte de ratio 4:1 (NaCl:KCl) était le plus proche de l'objectif visé; 69 % du total de ces deux
éléments correspondaient à du sodium tandis que 31 %, à du potassium (figure 2.3). Cependant, la réduction
en sodium correspondante était de seulement 22,3 % (p/p) (tableau 2.4) comparativement aux fromages
standards, ce qui était inférieur à la cible expérimentale visée. Ainsi, des temps de saumurage raccourcis (50
et 60 minutes) ont été expérimentés (figure 2.4 et tableau 2.6) dans le but d'obtenir une réduction en sodium
plus importante. Pour les autres saumures mixtes (ratios 7:3, 3:2 et 1:1), les proportions en potassium
quantifiées étaient trop élevées. Par crainte d'obtenir des fromages dont les qualités organoleptiques étaient
altérées, elles ont été éliminées du processus de sélection des paramètres de saumurage.
Au niveau de la teneur minérale (cendres), les résultats présentés au tableau 2.5 montrent des concentrations
plus élevées en minéraux dans les fromages salés en présence de KCl. Ces résultats concordent avec les

59
observations faites par Ayyash et Shah (2010), où une substitution du NaCl par du KCl dans un fromage de
type Halloumi a été réalisée par salage en saumure mixte. Ces résultats pourraient s'expliquer par une
pénétration plus rapide du KCl dans la matrice fromagère comparativement au NaCl. Aucune différence n'a
été observée au niveau de la teneur en humidité à l'exception des fromages salés dans la saumure mixte de
ratio 3:2 (NaCl:KCl). Ces observations concordent aussi avec des études similaires chez les variétés de
fromages saumurés, dont le Kefalograviera (Katsiari et al., 1998), le Feta (Aly, 1995, Katsiari et al., 1997),
l'Halloumi (Ayyash et Shah, 2010) et le Nabulsi (Ayyash et Shah, 2011).
Thibaudeau et coll. (2015) ont observé une teneur en humidité significativement plus élevée à la surface de
fromages Mozzarella salés dans une saumure de ratio 1NaCl:1KCl comparativement à des fromages salés
dans un ratio 3NaCl:1KCl et dans une saumure contrôle (100NaCl:0KCl). Les auteurs expliquent leurs
résultats par le fait que l'exposition à une plus faible teneur en sodium et plus forte teneur en potassium
pourrait avoir réduit la migration de l'eau du fromage vers la saumure. Une teneur en humidité plus élevée a
aussi été observée dans des fromages Fynbo salés dans un mixte de NaCl et de KCl (100g/L de chaque sel)
comparativement au salage en présence de NaCl seulement. L'hypothèse émise est en lien avec l'aw des
saumures, celle-ci étant plus élevée dans la saumure mixte. Conséquemment, la perte d'eau des fromages
salés dans cette dernière pourrait être moins importante (tel que cité dans Zorrilla et Rubiolo, 1994a).
Tableau 2.5 – Teneurs en humidité et en cendres des fromages au 4e jour d'affinage
Saumures (NaCl:KCl)
Non saumuré
4:1 7:3 3:2 1:1 100:0
(standard) Humidité (%) 48,1 ± 0,1 48,9 ± 0,1 49,1 ± 0,2 47,9 ± 0,2 49,0 ± 0,3 49,0 ± 0,4
Cendres (%) 1,79 ± 0,01 3,30 ± 0,16
3,19 ± 0,02
3,15 ± 0,06
3,32 ± 0,02
3,02 ± 0,06
Les résultats sont présentés par la moyenne ± l'erreur type. Tous les fromages ont été saumurés pendant 70 minutes.
2.4.3.2. Optimisation des paramètres de saumurage (2e essai de saumurage)
Les concentrations en sodium et en potassium quantifiées sont présentées à la figure 2.4 (mol/100g) et au
tableau 2.6 (mg/g). Comme il est mentionné à la section 2.4.3.1, les résultats sont dans un premier temps
présentés sous forme molaire afin de pouvoir comparer les teneurs des deux éléments de façon équivalente.
Les résultats sont ensuite présentés sur une base pondérale puisqu'il s'agit de la forme communément utilisée
en fromagerie et par Santé Canada (% de sel). Tel qu'observé lors du premier essai de saumurage (section
2.4.3.1), les résultats ont démontré qu'une substitution partielle du contenu en NaCl de la saumure par du KCl
a provoqué une réduction au niveau de la teneur en sodium incorporé dans les fromages et une augmentation
de la teneur en potassium. La somme des deux éléments (moles) quantifiés était plus élevée dans les
fromages salés en saumures mixtes pendant 60 et 70 minutes, comparativement aux fromages salés en

60
présence de NaCl seulement (70 minutes), mais cette augmentation s'est avérée relativement faible. Une
absorption plus rapide du potassium pourrait également expliquer ces résultats. Dans les fromages salés en
saumure mixte, le ratio de chacun des éléments s'est avéré différent de celui obtenu lors du premier essai de
saumurage. Pour chaque temps de saumurage, un ratio molaire de 3Na:1K (75 % Na, 25 % K) a été
déterminé dans les fromages du 2e essai de saumurage, contrairement à 7Na:3K (69 % Na, 31 % K) au
premier essai. De façon identique à ce qui a été observé au premier essai de saumurage, le ratio des deux
sels utilisés dans la saumure (4NaCl:1KCl) n'était pas équivalent au ratio obtenu dans les fromages (3Na:1K),
ce qui suggère également que le potassium diffuse plus rapidement dans la matrice fromagère que le sodium.
Tel qu'observé lors du premier essai de saumurage (section 2.3.4.1), les teneurs en humidité (tableau 2.7)
sont encore demeurées stables en fonction des différents traitements de saumurage.
Figure 2.4 – Incorporation du sodium (en violet) et du potassium (en vert) dans des fromages Camemberts salés pendant 50, 60 et 70 minutes dans des saumures mixtes de ratio 4NaCl:1KCl comparativement au sodium incorporé dans des fromages salés dans une saumure statique saturée à 26 % de NaCl (p/p) pendant 70 minutes. Le dosage des minéraux a été réalisé au 4e jour d'affinage. Les résultats sont représentés par la moyenne ± l'erreur type. Les pourcentages correspondent au ratio molaire des éléments sodium et potassium dans chacune des conditions, 100 % correspond au contenu total en sodium et en potassium.
0
0,01
0,02
0,03
Non salé 50 min 60 min 70 min 70 min
Sodi
um e
t pot
assi
um (m
ol/1
00 g
de
from
age)
Conditions de saumurage
Na K
75 %
25 %
75 %
25 %
76 %
24 %
92 %
8 %
4NaCl:1KCl 100NaCl:0KCl
54 %
48 %

61
La diffusion des sels s'est avérée différente du profil observé lors du premier essai de saumurage
(section 2.3.4.1). Comparativement aux standards, la réduction en sodium obtenue dans les fromages salés
70 minutes en saumure mixte était de seulement 9,3 % (tableau 2.6) au lieu de 22,3 % comme il a été observé
préalablement. Lors de l'optimisation des paramètres, de plus gros volumes de fromage ont été saumurés,
sans augmenter le volume de saumure ce qui peut expliquer les résultats obtenus. Les teneurs en sodium et
en potassium incorporées après 60 et 70 minutes de salage en saumure mixte étaient similaires ce qui
s'explique par la plus faible vitesse de diffusion du sel vers la fin du saumurage et une stabilisation du contenu
en sel. Un profil équivalent a également été observé au niveau de la teneur minérale (tableau 2.7). Après 50
minutes, il y avait significativement moins de sodium que dans les fromages standards, mais la teneur en
cendres était équivalente. La proportion de réduction en sodium calculée correspondait à 22,2 %, ce qui était
équivalent aux résultats obtenus après 70 minutes de saumurage lors du premier essai (section 2.3.4.1). Ainsi,
les résultats démontrent une grande variabilité au niveau de la prise en sel entre les essais effectués, ce qui
peut s'expliquer par le faible nombre de répétitions effectuées. Certains facteurs non contrôlés peuvent aussi
expliquer l'inconsistance des résultats obtenus entre les lots; soit les différences au niveau de la composition
du fromage avant saumurage, notamment la teneur en humidité initiale et une variabilité au niveau du volume
de fromage par rapport au volume de saumure. Il s'agit de facteurs bien connus en fromagerie qui expliquent
les écarts en sodium souvent importants observés entre les lots de fromage industriels. C'est pourquoi il est
difficile de reproduire des conditions de saumurage et d'obtenir des teneurs en ions sodium et potassium
rigoureusement identiques. Par conséquent, il est impératif de bien doser les proportions d'ions dans les
fromages étudiés avant d'en évaluer l'effet sur la microflore (Chapitre 3).
Tableau 2.6 – Teneurs en sodium et en potassium des fromages au 4e jour d'affinage et proportion de réduction en sodium correspondante
Saumures Non
saumuré
4NaCl:1KCl 100NaCl:0KCl (saumure standard)
70 minutes 50
minutes 60
minutes 70
minutes Teneur en sodium (mg/g de fromage)
0,57 ± 0,13 3,58 ± 0,12
4,10 ± 0,06
4,17 ± 0,11
4,60 ± 0,28
Teneur en potassium (mg/g de fromage)
0,82 ± 0,02 2,05 ± 0,08
2,26 ± 0,21
2,22 ± 0,03
0,65 ± 0,05
Réduction en sodiuma (%) - 22,2 10,9 9,3 - Les résultats sont présentés par la moyenne ± l'erreur type. a: Comparativement à la teneur en sodium obtenue dans les fromages saumurés pendant 70 minutes dans la saumure 100NaCl:0KCl.

62
Tableau 2.7 – Teneurs en humidité et en cendres des fromages au 4e jour d'affinage
Saumures Non saumuré 4NaCl:1KCl 100NaCl:0KCl
(saumure standard) 70 minutes
50 minutes
60 minutes
70 minutes
Humidité (%) 46,8 ± 0,8 46,6 ± 0,1 46,5 ± 0,2 46,3 ± 0,2 46,2 ± 0,1 Cendres (%) 1,84 ± 0,03 2,84 ± 0,02 2,93 ± 0,04 2,97 ± 0,01 2,89 ± 0,07
Les résultats sont présentés par la moyenne ± l'erreur type.
2.5. CONCLUSIONS ET PERSPECTIVES
Les paramètres permettant d'obtenir des fromages de composition saline standard ou réduits en sodium ont
pu être déterminés; une réduction nette sans substitution équivalente à 34,8 % a été obtenue. Cependant,
puisque les résultats ont démontré une grande variabilité, il est possible d'observer une différence lors d'une
répétition subséquente de ces paramètres de saumurage. La cible saline visée dans les fromages substitués
par du potassium n'a pu qu'être partiellement obtenue. Or, les paramètres permettant d'obtenir une réduction
en sodium allant jusqu'à 22 % avec une substitution par du potassium ont pu être atteints en conservant
l’équilibre en sel total du fromage et en tenant compte de la molalité. Les études traitant de l'absorption du sel
dans le Camembert ont généralement été réalisées à partir de saumure standard composée uniquement de
NaCl. Ainsi, cette étude a permis de mieux comprendre les phénomènes liés à l'absorption du sel après
salage en saumure mixte (NaCl et KCl). Les paramètres de saumurage sélectionnés pourront être utilisés afin
d'obtenir des fromages réduits en sodium et ainsi déterminer les effets de ces modifications salines sur la
texture des fromages et le comportement de la microflore fongique d'affinage (Chapitre 3).

63
3. Effet de la réduction du chlorure de sodium
dans un Camembert industriel et de sa
substitution partielle par du chlorure de
potassium sur la croissance de
Penicillium camemberti et
Geotrichum candidum


65
3.1. RÉSUMÉ
Les fromages contribuent de façon importante à l'apport quotidien en sodium au Canada. Puisque ce dernier a
des effets néfastes sur la santé, la réduction du sodium dans les fromages est souhaitable. Or, dans les
fromages de type Camembert, le sel joue plusieurs rôles fondamentaux dont l’un est le contrôle de la
croissance et de l'activité de la microflore fongique. Le but de cette étude était de déterminer l'impact de la
réduction du chlorure de sodium dans un Camembert industriel et de son remplacement partiel par du chlorure
de potassium sur la croissance des ferments d’affinage Penicillium camemberti et Geotrichum candidum.
La résistance intra-espèce au NaCl et KCl chez quatre souches de P. camemberti a été étudiée sur milieu de
culture synthétique (0 – 20 % sel) et sur caillé modèle de type Camembert (0 – 8 % sel). Ensuite, des
fromages Camemberts industriels ont été saumurés afin d'obtenir une teneur en sodium standard, une
réduction d’environ 30 % de sodium et une réduction de 20 à 30 % de sodium substitué par du potassium. La
dynamique de croissance de P. camemberti et G. candidum à la surface des fromages a été évaluée pour
chaque condition saline par PCR en temps réel à différents temps clés de l'affinage. La teneur en sodium, en
potassium, l'humidité, les cendres et le pH des fromages ont aussi été déterminés.
Les résultats ont démontré que le profil de croissance des quatre isolats de P. camemberti était similaire en
fonction des différentes teneurs salines. Une croissance optimale a été atteinte entre 1 et 3 % de sel sur milieu
de culture synthétique et à 1% sur caillé modèle de type Camembert (CMC). Le comportement de chacune
des souches en fonction de la nature des sels (NaCl vs KCl) était similaire aux teneurs salines avoisinant les
teneurs standards du Camembert (2 % sur milieu synthétique et 1 – 3 % sur CMC). Les analyses de
compositions des Camemberts industriels ont démontré que les conditions de saumurage ont eu peu d'impact
sur l'humidité ainsi que le pH des fromages. La quantification par qPCR a démontré que les modifications
salines avaient un léger effet sur la croissance de P. camemberti. Cependant, la croissance de G. candidum a
été accentuée dans les fromages réduits en sodium ou partiellement substitués avec du potassium,
principalement en début d'affinage. De plus, G. candidum devenait l'espèce dominante tout au long de
l'affinage des fromages réduits en sodium contrairement à ce qui est observé dans des conditions standards.
Globalement, les résultats ont démontré que la réduction du sel ou sa substitution affecte l’équilibre et la
croissance des ferments d'affinage. Cependant, il serait intéressant de mesurer l'impact de ces changements
salins sur l'activité métabolique de ces deux espèces.

66
3.2. INTRODUCTION
L'apport alimentaire quotidien en sodium chez l'humain est excessif et ce problème est présent au niveau
mondial (Brown et al., 2009). Dans plusieurs pays, l'apport quotidien en sodium varie entre 3,4 et 3,5 g
(Paquin, 2014) alors que la cible quotidienne recommandée par l'OMS est de seulement 2 g (OMS, 2007).
Une corrélation positive entre le sodium et la tension artérielle a été démontrée depuis plusieurs années
(Elliott et al., 1988, Poulter et al, 1990). De plus, l'hypertension représente un facteur de risque lié au
développement de plusieurs problèmes de santé tels que les maladies cardiovasculaires (Strazzullo et al.,
2009, Whelton et He, 2014), celles-ci étant une importante cause de mortalité au niveau mondial (OMS, 2007).
Plusieurs études ont démontré les effets bénéfiques reliés à une réduction de la consommation du sodium sur
la santé humaine (Sacks et al., 2001, He et MacGregor, 2006, Strazzullo et al., 2009, He et MacGregor, 2011,
He et al., 2013). Ainsi, des stratégies permettant de réduire la consommation en sodium ont été développées
au sein de plusieurs pays (Webster et al., 2011, OMS, 2013b).
Les fromages contribuent de façon importante à l'apport en sodium. Selon Fisher et al., (2009), ils
représentent 5 % de l'apport quotidien des Canadiens. Puisque la majorité de leur contenu en sodium provient
de l'ajout du chlorure de sodium (NaCl) pendant le procédé de transformation (Johnson et al., 2009), la
réduction de ce composé y est envisagée. Or, le sel (ou NaCl) joue plusieurs rôles essentiels dans cet aliment
tels que le contrôle de la croissance des microorganismes et des activités enzymatiques, ce qui contribue au
développement des propriétés organoleptiques recherchées pendant l'affinage (Guinee et Fox, 2004, Ardö et
al., 2014, Everett et al., 2014, Labrie et al., 2014). La réduction du NaCl dans les fromages à pâte molle et à
croûte fleurie comme le Camembert représente un enjeu important puisque cette variété possède des
caractéristiques intrinsèques favorables au développement des microorganismes. Leur contenu en eau et
l'activité de l'eau (aw) sont élevés et le pH en surface atteint rapidement la neutralité en cours d'affinage
(Guinee et Fox, 2004, Leclercq-Perlat et al., 2004, Guinee et O’Kennedy, 2007). Ils possèdent également une
microflore complexe composée de plusieurs espèces fongiques contrairement à d'autres types de fromages
où la population microbienne est beaucoup moins diversifiée (Spinnler et Gripon, 2004).
Penicillium camemberti et Geotrichum candidum sont utilisés comme ferments d'affinage et sont présents en
quantité importante à la surface des fromages de type Camembert (Lessard et al., 2014). Ces deux espèces
participent activement au développement des propriétés sensorielles par leur contribution au niveau de la
lipolyse, de la protéolyse et de l'établissement du gradient de pH (Spinnler et Gripon, 2004). Le comportement
de P. camemberti dans un Camembert industriel réduit en sodium est peu connu. Un développement faible et
inégal du mycélium à la surface d'un Camembert contenant 0,20 et 0,70 % de NaCl (p/p) a préalablement été
observé (O'Nulain, 1986), ce qui laisse croire qu'une quantité minimale en sel est nécessaire afin de permettre

67
une croissance optimale. G. candidum est reconnu pour sa sensibilité au sel, mais son comportement en
fonction de ce composé a déjà été décrit comme étant souche-dépendant (Boutrou et Gueguen, 2005).
L'ajout de sels substituts (MgCl2, CaCl2 et KCl) afin de compenser la réduction en NaCl dans les fromages a
été suggéré, mais à ce jour, aucun sel ne peut remplacer efficacement le NaCl. Plusieurs études ont démontré
la possibilité de générer un fromage de qualité acceptable en substituant une portion du NaCl par du KCl
(Fitzgerald et Buckley, 1985, Aly, 1995, Katsiari et al., 1997, 1998, Gomes et al., 2011, Grummer et al., 2012,
Thibaudeau et al., 2015). L'amertume et le goût métallique associés au KCl limite cependant son utilisation,
une substitution d'au plus 30 % a donc été suggérée dans plusieurs variétés fromagères (Hoffmann, 2014).
L'effet du KCl sur le comportement des ferments fongiques utilisés dans la fabrication fromagère est peu
connu. La croissance de Penicillium roqueforti sur pain blanc où 30 % du contenu en NaCl a été remplacé par
un mélange de sel [KCl et Sub4Salt (NaCl, KCl et gluconate de sodium)] n'a pas été affectée (Samapundo et
al., 2010). Cependant, l'influence du KCl sur la croissance de P. camemberti et G. candidum dans un
écosystème tel que le Camembert n'est pas connu.
Le but de l'étude était d'évaluer l'effet de la réduction du NaCl dans un Camembert industriel ou de sa
substitution partielle par du KCl sur la croissance de l'écosystème fongique d'affinage. Puisque l'effet du sel
sur la croissance de P. camemberti est peu connu, la caractérisation de différentes souches en fonction de la
concentration en NaCl et en KCl a préalablement été étudiée sur milieux de culture synthétique et sur caillés
modèles de type Camembert (CMC). La quantification de P. camemberti et G. candidum à la surface de
fromages Camembert industriels réduits en sodium ou partiellement substitués par du KCl a ensuite été
réalisée par PCR en temps réel à plusieurs temps clés pendant l'affinage.

68
3.3. MATÉRIEL ET MÉTHODES
3.3.1. Variation intra-espèce de la résistance au NaCl et KCl chez P. camemberti
3.3.1.1. Matériel biologique à l'étude
La croissance de quatre isolats de P. camemberti en fonction de différentes teneurs en NaCl et KCl a été
évaluée sur milieux de culture synthétiques et sur CMC. Quatre souches de P. camemberti ont été
sélectionnées dans la banque de souche du Laboratoire de mycologie alimentaire (LMA). Les souches
LMA-17 et LMA-30 ont été isolées à partir de deux fromages à pâte molle du terroir québécois et les souches
LMA-435 et LMA-1029 ont été fournies par deux industries productrices de ferments différentes.
3.3.1.2. Fabrication du fromage Camembert déshydraté
Deux lots de caillés modèles ont été fabriqués, chacun à partir de 120 L de lait. Les deux lots de lait cru
provenaient du Centre de recherche de Deschambault (Deschambault, Québec, Canada). Une fois pasteurisé
(75 C, 16 secondes) et refroidi à 32 °C, le lait a été ensemencé avec 1 % (v/v) de ferments lactiques (Flora
Danica, Chr Hansen, HØrsholm, Danemark) et 0,25 ml/l de CaCl2 (Cal-Sol, 45 % p/v, Fromagex, Rimouski,
Canada). Lorsque le pH du lait a atteint 6,4, l'emprésurage a été réalisé en ajoutant 0,1 ml/l de présure
contenant 520 mg/l de chymosine (CHY-MAX, Chr Hansen, HØrsholm, Danemark). Le temps total de
coagulation (temps de prise et de coagulation) a été de 72 minutes pour le premier lot de production et de 138
minutes pour le deuxième. Le caillé a ensuite été coupé et moulé dans des moules d'un diamètre de 103 mm
et d'une hauteur de 109 mm. Trois retournements ont été faits, soit après 30, 60 et 180 minutes suivant le
moulage. Les caillés ont été placés 16 h à 22 °C, démoulés et défaits en morceaux à la main. Ils ont été
lyophilisés (Virtis-Repp, SP industries, Warminster, États-Unis), moulus au moulin à disque (Glenn Mills Inc.,
Clifton, États-Unis), emballés sous-vide (Spirovac 350, Spiromac, St-Germain-de-Grantham, Canada) et
entreposés à – 30 C jusqu'au moment de leur utilisation. Pendant la fabrication du caillé, aucun ferment
fongique d'affinage n'a été ajouté et aucune étape de salage n'a été effectuée. Le pH final des lots de caillé au
jour 1 était de 4,79 pour le premier lot et de 4,61 pour le deuxième.
3.3.1.3. Préparation des CMC et des milieux de culture synthétiques
Le caillé modèle déshydraté a d'abord été irradié par rayonnement gamma de source cobalt 60 (Gammacell
220 Research Irradiator, Nordion, Ottawa, Canada) à une dose de 5 kGy. Pour chacune des conditions
salines, tous les caillés ont été préparés afin qu'ils soient composés de 44 % (p/p) de masse sèche (caillé et
sel) et 56 % (p/p) d'humidité (eau distillée stérile). Les concentrations en NaCl testées étaient de 0, 1, 2, 3, 4,
5, 6, 7 et 8 % (p/p) ainsi que leur équivalent molaire en KCl. Le sel et l'eau distillée ont été homogénéisés et

69
ajoutés au caillé déshydraté. Le caillé hydraté a ensuite été mélangé et réparti en portion de 30 g dans des
boîtes de Pétri. Les caillés ont été hydratés un maximum de 24 heures avant l'expérience réalisée et ont été
conservés à 4 °C jusqu'à leur utilisation.
Le PDA (DifcoTM Potato Dextrose Agar, BD, Mississauga, Canada) déshydraté (39 g/L) et le sel ont été
homogénéisés dans de l'eau distillée jusqu'à dissolution complète. Ensuite, le volume d'eau final a été ajusté à
l'aide d'un cylindre gradué. Les milieux de culture ont été préparés afin d'obtenir les teneurs en NaCl
suivantes; 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15 et 20 % (p/v). Une deuxième série de milieux de culture a été
effectuée avec du KCl. La quantité de KCl ajoutée aux milieux a été calculée selon l'équimolaire de chaque
concentration en NaCl. Les milieux de culture ont été autoclavés 15 minutes à 121 C. Ils ont été acidifiés (pH
3,5) après la stérilisation en ajoutant 14 ml/l d'acide tartrique stérile 10 % (p/v) (D-Tartaric acid, 99 %, Sigma-
Aldrich, Saint-Louis, États-Unis) et entreposés à 4 °C jusqu'à leur utilisation.
3.3.1.4. Suivi de croissance en fonction du NaCl et du KCl
Afin de préparer les suspensions de spores utiles pour les tests de croissance, chacune des souches de
P. camemberti a été cultivée sur milieu de culture PDA acidifié pendant 7 jours à 25 °C. À partir d'une colonie
isolée, elles ont été transférées sur une nouvelle gélose PDA acidifié de façon à obtenir un mycélium couvrant
toute la surface. Les spores ont été récoltées à l'aide d'écouvillons stériles imbibés de Tween 80 0,5 % (v/v)
stérile après 7 jours de croissance à 25 °C. Elles ont été suspendues dans 1 ml de Tween 80 0,5 % stérile,
dénombrées à l'hématimètre et diluées avec de l'eau peptonée 0,1 % (p/v) (BactoTM peptone, BD,
Mississauga, Canada) stérile à une concentration finale de 100 spores/µl. Pour chaque souche, 1 µl de la
suspension de spores a été inoculé en un point au centre de chaque milieu de culture et CMC. Les cultures
ont été incubées, sans les inverser, à 25 °C. La croissance a été évaluée en mesurant le diamètre des
colonies (mm) à toutes les 24 heures sur une période de 10 jours (López-Dίaz et al., 1996, Cuppers et al.,
1997). Ces expériences ont été réalisées trois fois et chaque répétition a été faite en triplicata. Pour chaque
expérience, de nouveaux CMC et milieux de culture PDA ont été fabriqués.

70
3.3.2. Suivi de la croissance de la microflore fongique d'affinage dans un fromage Camembert industriel
3.3.2.1. Matériel à l'étude
Les fromages Camemberts utilisés provenaient d'une entreprise canadienne de production de fromages de
spécialité. La forme des fromages était cylindrique (diamètre: 20 cm, hauteur: 3 cm) et leur poids était de
870 g. Ils ont été produits à partir de lait pasteurisé selon un procédé de fabrication de type industriel dont les
étapes demeurent confidentielles. La coagulation a été faite par ajout d'une combinaison de ferments lactiques
thermophiles et mésophiles afin d'obtenir une pâte de type stabilisée. Les seuls ferments fongiques inoculés
au lait lors du procédé de fabrication de ce fromage sont P. camemberti LMA-1029 et G. candidum LMA-1028.
3.3.2.2. Préparation des saumures et saumurage
Les saumures ont été confectionnées et entreposées en usine dans une salle tempérée à 12 °C ± 2 °C, 48h
avant leur utilisation. Elles ont été produites dans des bacs de différentes dimensions. Ainsi, leur poids final
variait entre 45 et 154 kg. Chaque saumure a été préparée avec de l'eau potable dans lesquelles le NaCl (Hi-
Grade Granulated Salt (> 99,8 % NaCl), Windsor Salt, Pointe-Claire, Canada) et le KCl (Potassium chloride
USP/FCC, Thermo Fisher Scientific, Waltham, États-Unis) ont été ajoutés. La composition en sel de chaque
saumure est mentionnée au tableau 3.1. Une fois le NaCl et le KCl ajoutés, une solution de CaCl2 32 % (p/v)
(FDC Industries) a été utilisée afin d'obtenir une teneur finale de CaCl2 de 1 %. Vingt-quatre heures après la
confection des saumures, leur pH a été mesuré et ajusté 5,1 ± 0,1 par ajout d'une solution d'acide lactique
80 % (FCC solution, Les Produits Chimiques Amplex Ltée, Pointe-Claire, Canada). Elles ont été brassées à
tous les jours afin de favoriser la dissolution des sels et l'atteinte d'une stabilité, jusqu'à leur utilisation.
Les paramètres de saumurage permettant d'obtenir les conditions salines ciblées ((STD) standard; (RED)
réduction nette de 30 % du contenu en sodium et (KCL) réduction de 22 % du contenu en sodium avec une
substitution par du potassium) ont préalablement été optimisés (Chapitre 2) et ont été utilisés afin de
reproduire des fromages ayant ces compositions. Une fois démoulés, un total de 63 fromages (21/condition
saline), provenant tous du même lot de production, ont été placés immédiatement dans les saumures. Ainsi,
pour chaque condition saline et jours d'échantillonnage, trois fromages ont été saumurés afin d'obtenir un
triplicata (3 fromages/conditions x 3 conditions x 7 jours = 63 fromages). Les paramètres de saumurage
sélectionnés pour chaque condition saline ciblée sont présentés dans le tableau 3.1. Compte tenu du contexte
expérimental du projet, le ratio du volume de saumure par rapport au volume de fromages par saumure se
situait entre 6 et 12. Puisque l'équipement utilisé ne permettait pas de submerger les fromages dans la
saumure, ils ont été tournés à la mi-temps de saumurage. À titre de contrôle, trois fromages saumurés 70

71
minutes dans le bassin de saumure industriel (Condition IND) ainsi que trois fromages non saumurés (NSAL)
ont été sélectionnés.
Tableau 3.1 – Composition saline des saumures et temps de saumurage
Cibles salines des Camemberts
Composition saline des saumures Temps de
saumurage (minutes) Teneur en sels (p/p)
Substitution NaCl par
KCl
Ratio molaire NaCl:KCl
(STD) Standard 26,0 % NaCl - - 70 (RED) Réduction nette en sodium
26,0 % NaCl - - 20
(KCL) Substitution par potassium
20,8 % NaCl et 6,6 % KCla 20 % 4:1 50
(IND) Contrôle industriel Saturée en NaClb-c - - 70 a: Les remplacements du NaCl par le KCl ont été faits sur une base équimolaire et la teneur totale en sel est équivalente à la saumure saturée à 26 % (p/p) de NaCl. b: Il s'agit de la saumure industrielle fabriquée par le personnel de l'industrie qui est normalement utilisée pour saler tous les fromages saumurés de l’usine. c: La teneur exacte en NaCl est inconnue puisque l'ajout de sel dans le bassin se fait en continu. Le taux de saturation en sel atteint cependant 97 %.
3.3.2.3. Affinage et échantillonnage
L'affinage des fromages a été fait selon les paramètres optimisés par l'entreprise. Une fois saumurés, tous les
fromages expérimentaux, ainsi que les fromages utilisés à titre de contrôle, ont été placés sur des claies et
transférés dans la salle d'affinage à 12,5 °C et 96,5 % d'humidité relative. Deux retournements ont été faits,
soit aux jours 5 et 7. Au jour 11, les fromages ont été séchés (12 °C, 2 h), emballés et transportés à
l'Université Laval où ils ont été placés dans une chambre d'affinage à 4 C jusqu'au moment de leur
échantillonnage respectif. Cette procédure correspond au conditionnement normalement appliqué au fromage
dans cette industrie avant son entreposage et sa livraison.
L'échantillonnage des fromages a été fait à plusieurs étapes clés de l'affinage, chacune étant représentée au
tableau 3.2. L'analyse de composition des fromages, c'est-à-dire; la teneur en humidité, en cendres, en
sodium, en potassium et le pH, a été effectuée aux jours 5, 11, 14, 21 et 35. Des prises de pH ont aussi été
effectuées aux jours 6 et 7. À chaque jour d'échantillonnage, trois fromages/condition ont été sélectionnés au
hasard dans la salle d'affinage. Le jour des prélèvements, deux portions d'un quart (pointe), précisément
mesurées, de chaque fromage ont été coupées. Ces portions ont été emballées sous-vide et congelées à
– 30 °C afin de pouvoir effectuer les analyses de composition. Parmi les deux quarts prélevées, un a été
sélectionné pour la prise de pH et l'autre pour la détermination du taux d'humidité et des cendres et le dosage
du sodium et du potassium. La portion restante des fromages a été utilisée pour prélever les échantillons de
mycélium. À chaque jour d'échantillonnage, une surface, précisément mesurée et équivalente à 50 cm2 (25

72
cm2 de chaque côté) de mycélium, a été extraite des fromages à l'aide d'une lame de scalpel stérile en
prenant soin de ne pas prélever la matrice fromagère. Le mycélium a ensuite été broyé à l'azote liquide à
l'aide d'un mortier et d'un pilon stériles. Le mycélium broyé a été conservé à – 80 °C jusqu'à son utilisation. Le
mycélium à jour 5 n'a pu être retiré puisqu'il n'était pas suffisamment développé, le premier jour
d'échantillonnage de la croûte a donc été fait au jour 6. Les autres prélèvements de mycélium ont été faits aux
jours 11, 14, 21 et 35.
Tableau 3.2 – Journées d’échantillonnage des Camemberts, concordance avec les étapes d’affinage du fromage dans l’industrie participant au projet.
Jours Étapes de l'affinage
5 Apparition du mycélium 6 Jour où il y a suffisamment de mycélium pour l’échantillonnage 7 Suivi du pH seulement, jour de contrôle en industrie
11 Emballage, séchage et transfert à 4 °C 14 Distribution et prêt à la consommation 21 Période de consommation 35 Période de consommation
3.3.2.4. Analyses de composition des Camemberts
Le taux d'humidité, le contenu en cendres, en sodium et en potassium ont été déterminés sur les échantillons
prélevés aux jours 5, 11, 14, 21 et 35 (STD, RED, KCL) ainsi qu'au jour 5 seulement pour les fromages
contrôles (IND et NSAL). Le ratio S/H a également été calculé à partir des teneurs en sodium, en potassium et
en humidité pour chacun de ces jours d'échantillonnage. Les échantillons de fromages emballés sous-vide ont
été décongelés dans un bac d'eau froide le matin de leurs analyses. Chaque échantillon a été broyé dans un
robot culinaire (type blender) afin de le rendre homogène. La détermination du taux d'humidité a été réalisée
par séchage au four à vide (Isotemp 280A, Thermo Fisher Scientific, Waltham, États-Unis) à 100 C pendant 4
heures (AOAC, 2008). Les cendres ont été obtenues après calcination (Thermolyne, Thermo Fisher Scientific,
Waltham, États-Unis) à 550 C pendant une période de 18 à 24 h ou jusqu'à ce qu'elles soient de couleur
grise-blanche (AOAC, 2005). Les cendres ont été solubilisées dans du TCA 20 % (p/p) (Trichloroacetic acid
crystals, ACS, J.T. Baker, Center Valley, États-Unis) dissoutes dans de l'eau déminéralisée 18 m et filtrées
sur membrane d'acétate de cellulose (VWR syringe filter 0,45 µm, VWR International, Montréal, Canada). Le
dosage du sodium et du potassium a été réalisé par spectrométrie d'émission à plasma induit (Optima 4300TM
DV ICP-OES, PerkinElmer, Waltham, États-Unis) au Laboratoire de pédologie forestière de la Faculté de
Foresterie, de Géographie et de Géomatique de l'Université Laval (Thibaudeau et al., 2015). Pour chaque
échantillon de fromages, les analyses ont été effectuées en triplicata.

73
Le suivi du pH a été fait au centre et à la surface de chaque échantillon prélevé aux jours 5, 6, 7, 11, 14, 21 et
35. Les échantillons de fromages emballés sous-vide ont été décongelés le matin de leur analyse dans un bac
d'eau froide. Le mycélium à la surface des fromages a été retiré à l'aide d'une lame de scalpel. Les surfaces
des fromages, d'une épaisseur de 5 mm, ont été séparées du centre à l'aide d'une lame de scalpel (Back et
al., 1993). Le centre et la surface ont été homogénéisés séparément à la main. Les prises de pH (Orion 3-star,
Thermo Scientific, Waltham, États-Unis) ont été effectuées une fois à l'aide d'une électrode de pH combinée
en verre (Inlab Cool, Mettler Toledo, Columbus, États-Unis). Pour chaque fromage, trois mesures ont été
prises.
3.3.2.5. Suivi de croissance de P. camemberti et G. candidum au cours de l'affinage
L'ADN génomique totale (ADNg) fongique, prélevé à la surface des fromages, a été extrait selon la méthode
d'Al-Samarrai et Schmid à partir d’une quantité de mycélium broyé à l'azote liquide connue variant entre 17 à
26 mg (Al-Samarrai et Schmid, 2000). Une extraction d'ADNg a été réalisée par échantillon de mycélium, donc
une par fromage. Ainsi, pour chaque condition, trois extractions d'ADN ont été réalisées (1 extraction/fromage
x 3 fromages/condition).
La quantification de P. camemberti LMA-1029 et G. candidum LMA-1028 aux jours 6, 11, 14, 21 et 35 a été
réalisée en triplicata par qPCR en temps réel (ABI 7500 Fast system) selon la méthode optimisée par Lessard
et coll. (2012). La détection des gènes a été réalisée en utilisant une approche qPCR TaqMan. Les gènes
ciblés et la séquence des amorces et des sondes utilisées sont présentés dans le tableau 3.3 (Lessard et al.,
2012). Leur quantification a été réalisée de façon absolue par comparaison des cycles seuils (threshold cycle)
à une courbe standard obtenue à partir de plasmides recombinants contenant les gènes d'intérêt. Seuls les
essais de quantification qPCR ayant un pourcentage d'efficacité se situant entre 90 et 110 % ont été
considérés.
Tableau 3.3 – Gènes ciblés et séquences nucléotidiques des amorces et des sondes utilisées pour la quantification par PCR en temps réel de P. camemberti et de G. candidum.
Espèces Gène ciblé Séquence des amorces et sondes (5' vers 3')
P. camemberti (LMA-1029)
-tubuline (btub)
Amorce F : CCG TTC TCG TCG ATT TGG A Amorce R : CTT TCG GCA AGC TTT TCC G Sonde : Fam– TGG TAC CAT GGA CGC TGT CCG CT –Tamra
G. candidum (LMA-1028)
Cystathionine-ɣ-lyase (cgl)
Amorce F : CGC TTT CAC CAA CTC CAT TGA Amorce R : CTT GTC TGG ATC GAG TCT CCC T Sonde : Fam– CCG AGC TCC GTG ACA TCA TCA CTG A –Tamra

74
3.3.3. Analyses statistiques
Pour l’analyse des données sur les diamètres moyens des colonies de P. camemberti, des modèles d’Anova à
blocs aléatoires complets ont été utilisés. Toutefois, la structure des traitements dans chaque bloc est
différente selon le milieu de culture utilisé, à savoir le milieu PDA acidifié ou caillé modèle Camembert (CMC).
Cette structure est composée de trois facteurs, soit le facteur « souches » (4 niveaux : LMA-1029, LMA-17,
LMA-30, LMA-435), le facteur « type de sel » (3 niveaux : KCl, NaCl, sans sel) et le facteur « concentrations
salines ». Les niveaux utilisés à ce dernier facteur changent selon le milieu d’application. Pour le milieu PDA
acidifié, 12 concentrations salines ont été étudiées, soit les concentrations 1-10 %, 15 % et 20 %. Il est à noter
que les données de la concentration 20 % ont été exclues des analyses, puisque les mesures de diamètres
étaient toutes nulles à cette concentration. En contrepartie, pour le milieu CMC, seulement huit concentrations
salines ont été expérimentées, soit les concentrations 1-8 %.
Puisque la concentration saline a un sens uniquement en présence d’un type de sel, les facteurs « types de
sel » et « concentration saline » ont été regroupés ensemble en un seul facteur, nommé « conditions
salines ». Un total de 23 modalités pour ce facteur ont été considérées dans le milieu PDA, soit les 11
concentrations salines (1-10 %, 15 %) de KCl, avec le NaCl (les mêmes 11 concentrations) et l’absence de
sel. Pour le milieu CMC, il y a 17 modalités considérées pour ce facteur, soit les huit concentrations salines (1-
8 %) avec le KCl, ces mêmes concentrations salines avec le NaCl et l’absence de sel. Au final, la structure
des traitements dans chaque bloc est une structure factorielle 4 x 23 dans le milieu PDA et une structure
factorielle 4 x 17 dans le milieu CMC. Des contrastes ont été construits sur ces sources de variation pour
évaluer l’effet propre des souches, des types de sel et des concentrations salines, de même que leurs
interactions.
Pour tenir compte de l’hétéroscédasticité dans les données, les modèles d’Anova ont été ajustés avec une
variance résiduelle propre à chaque niveau de concentration saline. De plus, pour rencontrer le postulat de
normalité sur les modèles ajustés en milieu PDA, les mesures de diamètres des colonies ont été transformées
par la racine carrée, soit la meilleure transformation identifiée dans la famille de Box-Cox. Aucune
transformation n’a été appliquée en milieu CMC. Pour chacune des analyses effectuées, un seuil de
signification de 5 % a été considéré pour déclarer un effet significatif entre les données.
Pour l’analyse des données sur la teneur en humidité, en cendres, en sodium, en potassium, le ratio S/H, le
pH et la population fongique (gènes btub et cgl) dans les fromages, des modèles d’Anova complètement
aléatoires ont été utilisés. Toutefois, la structure des traitements est différente d’un type de mesure à l’autre.
Pour les mesures de teneur en humidité, en cendres et en sodium des fromages, la structure des traitements
comprend trois conditions salines croisées chacune à cinq jours d’échantillonnage, en plus de deux contrôles,

75
pour un total de 17 combinaisons de traitement. Pour les mesures de teneur en potassium des fromages, la
structure des traitements comprend deux conditions salines croisées chacune avec cinq jours
d’échantillonnage, en plus de trois contrôles, pour un total de 13 combinaisons de traitement. Pour les ratios
S/H des fromages, la structure des traitements comprend trois conditions salines croisées chacune avec cinq
jours d’échantillonnage, en plus d’un contrôle, pour un total de 16 combinaisons de traitement. Des contrastes
ont été construits sur les combinaisons de traitement pour évaluer l’effet propre des conditions salines, des
jours d’échantillonnage et de leur interaction.
Pour les analyses du pH des fromages, la structure des traitements est une structure factorielle comprenant
trois conditions salines (3 niveaux) croisées chacune à sept jours d'échantillonnage, pour un total de
21 combinaisons au centre et à la surface du fromage. Finalement, la structure des traitements dans la
quantification fongique (gènes btub et cgl) comprenait deux facteurs, soit les conditions salines (3 niveaux) et
les jours d'échantillonnage (5 niveaux), pour un total de 15 combinaisons pour chacun des gènes.
Pour chacune des analyses effectuées, la normalité des résidus a été vérifiée par le test de Shapiro-Wilk. Afin
de satisfaire les postulats de normalité, aucune transformation des données n'a été nécessaire pour les
analyses sur l'humidité, les cendres, le sodium, le potassium, le ratio S/H et le pH. Un modèle avec variances
hétérogènes, selon la condition, a été ajusté pour le sodium et le potassium. Dans le cas de la quantification
fongique, afin de satisfaire les hypothèses de normalité et d’homoscédasticité des résidus sous-jacents, les
variables ont été utilisées avec leur logarithme. Pour chacune des analyses effectuées dans les fromages, un
seuil de signification de 1 % a été considéré pour déclarer un effet significatif entre les données.
Tous les modèles ont été ajustés aux données à l'aide de la procédure « MIXED » du logiciel SAS (version 9.4
SAS inc., NC).

76
3.4. RÉSULTATS ET DISCUSSION
3.4.1. Variation intra-espèce de la résistance au NaCl et KCl chez P. camemberti
Puisque la tolérance au sel chez P. camemberti est peu connue, l'étude de son comportement en fonction de
différentes teneurs en NaCl et KCl a été effectuée. Ainsi, son optimum de croissance face au sel ainsi que son
niveau de tolérance ont pu être déterminés. Étant donné que le comportement de G. candidum face au sel a
été mieux documenté (Sansonetti, 1930, Tornadijo et al., 1998, van den Tempel et Nielsen, 2000, Marcellino
et al., 2001), cette espèce n'a pas été incluse dans cette partie de l'étude.
3.4.1.1. Croissance sur milieu de culture synthétique
La croissance de quatre souches de P. camemberti en fonction du NaCl et du KCl a été évaluée sur PDA
acidifié (Figure 3.1). L'évolution de la croissance en fonction des deux sels était similaire pour chaque isolat;
c'est-à-dire, plus les teneurs salines étaient élevées, plus la croissance a été ralentie. Les analyses
statistiques ont démontré un effet significatif pour l'interaction triple souches*types de sel*concentrations
(p < 0,0001, tableau 3.4). Selon cette interaction et dépendamment de la teneur saline, la sensibilité face au
NaCl ou au KCl entre les souches s'est avérée différente pour une même concentration (tableau 3.5). Entre
autres, la sensibilité de la souche LMA-1029 a semblée plus prononcée face aux deux sels comparativement
aux autres souches (figure 3.1).
Selon les résultats présentés à la figure 3.1, l'optimum de croissance en fonction du NaCl pour les quatre
souches se situait à 1 %. Au-dessus de ces teneurs salines, un léger ralentissement de croissance a été
observé. De plus, les résultats suggèrent que les quatre souches ont été stimulées par une teneur en sel
minimale pour une croissance optimale sur ce milieu de culture puisque la croissance a été considérablement
réduite à 0 % de sel. Les quatre souches ont pu se développer jusqu'à 10 % de NaCl, mais aucune croissance
n'a été visible aux concentrations supérieures après 10 jours d'incubation à 25 C. Des résultats semblables
ont été obtenus en présence de KCl. Chaque souche a démontré une préférence pour de faibles teneurs en
KCl. L'optimum de croissance en fonction du KCl chez LMA-17, LMA-30 et LMA-435 se situait à 0,51 M
(équivalent molaire à 3 % de NaCl) et à 0,34 M (équivalent molaire à 2 % NaCl) pour LMA-1029. La
croissance a été visible jusqu'à 2,56 M (équivalent molaire à 15 % de NaCl) pour les quatre souches après
10 jours d'incubation, mais a été considérablement ralentie. Aucune croissance n'a été observée à 3,42 M de
KCl (équivalent à 20 % de NaCl) au 10e jour d'incubation à 25 C.
Les résultats obtenus dans cette étude sont en lien avec ce qui a été observé dans la littérature sur milieux de
culture gélosés; de faibles teneurs salines, soit entre 1 et 2 % de NaCl, stimuleraient le développement de

77
P. camemberti tandis que des concentrations plus élevées retarderaient sa croissance (Sansonetti, 1930,
Choisy et al., 2009). Sa tolérance face au NaCl est élevée puisqu'entre 20 et 30 % seraient nécessaires pour
une inhibition complète de sa croissance (Sansonetti, 1930). Parallèlement aux observations faites à la figure
3.1, Lopez-Diaz et coll. (1996) ont également observé un effet stimulant sur la croissance de P. roqueforti en
présence de faibles concentrations en NaCl lorsque comparées à la croissance en absence de sel.
La figure 3.1 montre une différence au niveau du comportement distinct de chaque isolat en fonction de la
nature des sels. D'ailleurs les analyses statistiques présentées au tableau 3.4 ont démontré un effet important
accordé aux types de sel (p < 0,0001). De plus, l'interaction triple souches*types de sel*concentrations permet
d'observer les effets significatifs selon la nature du sel pour chacune des souches à une concentration précise
(tableau 3.5). À 1 % de sel, seule la souche LMA-435 possède un comportement similaire en fonction du type
de sel, les autres souches préférant croître en fonction du NaCl. À 2 % de sel, toutes les souches ont un
comportement similaire selon la nature du sel. À partir de 3 %, et ce jusqu'à 10 %, la nature du sel affecte
différemment la croissance des souches LMA-17, LMA-30 et LMA-435, le NaCl démontrant un effet plus
inhibiteur. Pour la souche LMA-1029, l'effet plus inhibiteur du NaCl est visible entre 5 et 10 % de sel. Pour
toutes les souches, cet effet devient de plus en plus important avec l’augmentation de la concentration saline.
Les sels influencent le comportement des microorganismes par leur capacité à réduire l'activité de l'eau (aw)
(Guinee et Fox, 2004, Guinee et O’Kennedy, 2007, Choisy et al., 2009, Doyle et Glass, 2010), mais leur
potentiel répresseur dépendrait de la nature du composé en question. Cependant, à la même molalité,
Samapundo et coll. (2010) ont observé que le NaCl et le KCl exerçaient un effet équivalent au niveau de la
répression de l'aw du milieu de culture PDA, ce qui n'a pas été évalué dans notre étude. Dans cette même
étude, les auteurs ont démontré que le NaCl et le KCl, à la même molalité, avaient un effet similaire sur le taux
de croissance et la durée de la phase de latence de Penicillium roqueforti et d’Aspergillus niger. Ils soulignent
que leurs résultats sont en lien avec la capacité de chacun des sels à réduire l'aw. Or, considérant les résultats
obtenus, il se pourrait que l'effet répresseur (ou stimulant) du NaCl et du KCl sur P. camemberti soit causé par
d'autres facteurs que la disponibilité de l'eau et/ou la toxicité de l'ion Cl- puisque ce dernier était présent à la
même concentration.

78
Tableau 3.4 – Table d'ANOVA associée aux analyses statistiques sur la mesure du diamètre des colonies sur milieux PDA acidifié
Sources de variation Diamètre - PDA acidifié
DL F-Value p-Value Souches 3 167,06 <0,0001
Types de sel 2 285,73 <0,0001 Concentrations 10 2621,93 <0,0001
Types de sel*concentrations 10 110,26 <0,0001 Souches*types de sel 6 2,31 0,0354
Souches*concentrations 30 10,18 <0,0001 Souches*types de sel*concentrations 30 2,63 <0,0001
Erreur 180
Total corrigé 271
Figure 3.1 – Influence du NaCl et du KCl sur la croissance de quatre souches de P. camemberti sur milieu de culture synthétique PDA acidifié (pH 3,5) après 10 jours d'incubation à 25 C. Les souches sont représentées par quatre différentes couleurs; LMA-17 (bleu), LMA-30 (orange), LMA-435 (violet), LMA-1029 (vert). Les bandes hachurées représentent la croissance en absence de sel, les bandes foncées représentent la croissance en fonction du NaCl et les bandes pâles représentent la croissance en présence de KCl. Les données sont représentées par la moyenne ± l'erreur type. L'axe des abscisses indique le % en NaCl. Les teneurs en KCl ont été calculées sur une base équimolaire avec le NaCl.
0
10
20
30
40
50
60
0 1 2 3 4 5 6 7 8 9 10 15 20
Diamètre des colonies (m
m)
Concentration en NaCl (%) ou équivalent molaire en KCl
LMA‐17 NaCl LMA‐30 NaCl LMA‐435 NaCl LMA‐1029 NaCl
LMA‐17 KCl LMA‐30 KCl LMA‐435 KCl LMA‐1029 KCl
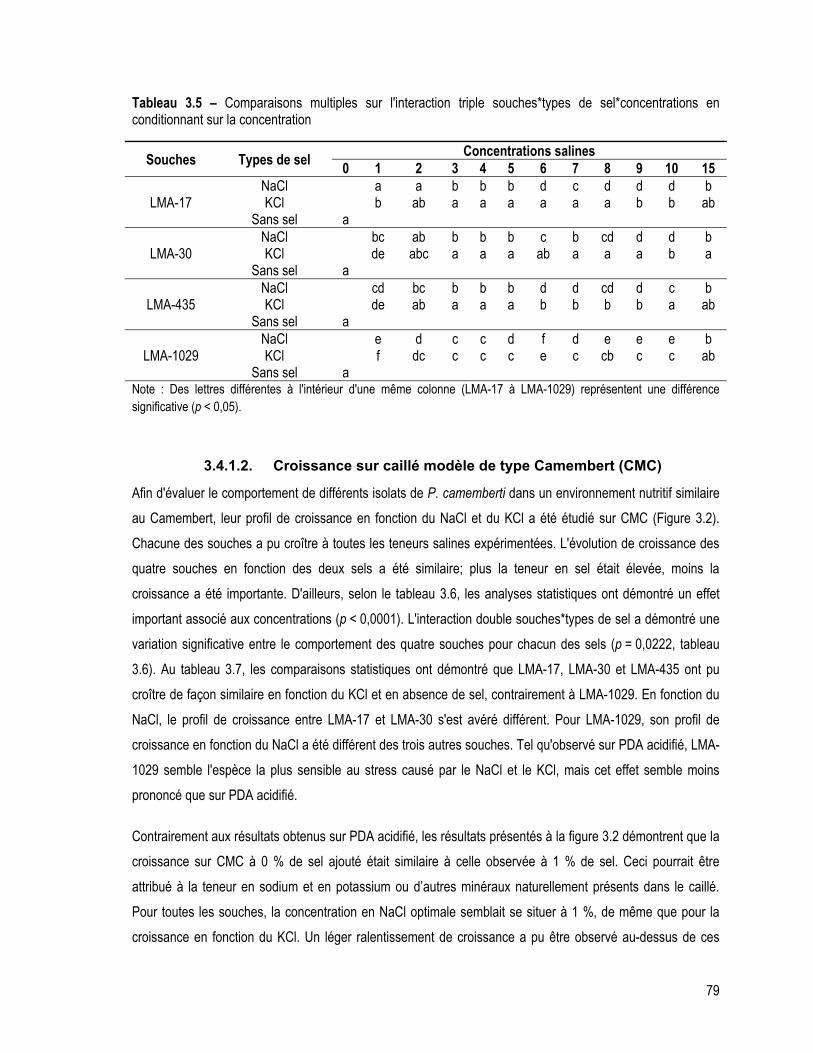
79
Tableau 3.5 – Comparaisons multiples sur l'interaction triple souches*types de sel*concentrations en conditionnant sur la concentration
Souches Types de sel Concentrations salines
0 1 2 3 4 5 6 7 8 9 10 15
LMA-17 NaCl a a b b b d c d d d b KCl b ab a a a a a a b b ab
Sans sel a
LMA-30 NaCl bc ab b b b c b cd d d b KCl de abc a a a ab a a a b a
Sans sel a
LMA-435 NaCl cd bc b b b d d cd d c b KCl de ab a a a b b b b a ab
Sans sel a
LMA-1029 NaCl e d c c d f d e e e b KCl f dc c c c e c cb c c ab
Sans sel a Note : Des lettres différentes à l'intérieur d'une même colonne (LMA-17 à LMA-1029) représentent une différence significative (p < 0,05).
3.4.1.2. Croissance sur caillé modèle de type Camembert (CMC)
Afin d'évaluer le comportement de différents isolats de P. camemberti dans un environnement nutritif similaire
au Camembert, leur profil de croissance en fonction du NaCl et du KCl a été étudié sur CMC (Figure 3.2).
Chacune des souches a pu croître à toutes les teneurs salines expérimentées. L'évolution de croissance des
quatre souches en fonction des deux sels a été similaire; plus la teneur en sel était élevée, moins la
croissance a été importante. D'ailleurs, selon le tableau 3.6, les analyses statistiques ont démontré un effet
important associé aux concentrations (p < 0,0001). L'interaction double souches*types de sel a démontré une
variation significative entre le comportement des quatre souches pour chacun des sels (p = 0,0222, tableau
3.6). Au tableau 3.7, les comparaisons statistiques ont démontré que LMA-17, LMA-30 et LMA-435 ont pu
croître de façon similaire en fonction du KCl et en absence de sel, contrairement à LMA-1029. En fonction du
NaCl, le profil de croissance entre LMA-17 et LMA-30 s'est avéré différent. Pour LMA-1029, son profil de
croissance en fonction du NaCl a été différent des trois autres souches. Tel qu'observé sur PDA acidifié, LMA-
1029 semble l'espèce la plus sensible au stress causé par le NaCl et le KCl, mais cet effet semble moins
prononcé que sur PDA acidifié.
Contrairement aux résultats obtenus sur PDA acidifié, les résultats présentés à la figure 3.2 démontrent que la
croissance sur CMC à 0 % de sel ajouté était similaire à celle observée à 1 % de sel. Ceci pourrait être
attribué à la teneur en sodium et en potassium ou d’autres minéraux naturellement présents dans le caillé.
Pour toutes les souches, la concentration en NaCl optimale semblait se situer à 1 %, de même que pour la
croissance en fonction du KCl. Un léger ralentissement de croissance a pu être observé au-dessus de ces

80
teneurs salines. Tel qu'observé sur PDA acidifié, l'interaction double types de sel*concentrations a démontré
une variation significative du comportement de l'ensemble des souches en fonction de la nature des sels
(p < 0,0001). Pour une même teneur en NaCl et KCl, le comportement des quatre souches s'est avéré
différent à des concentrations supérieures à 4 % (figure 3.2), le NaCl ayant un effet inhibiteur plus prononcé.
Sachant que la teneur saline du Camembert est variable et se situe entre 1,5 et 2,5 % (Sousa, 2003), les
résultats obtenus suggèrent qu'une réduction nette du contenu en NaCl ou sa substitution partielle par du KCl
dans ce type de fromage risque d'affecter légèrement la croissance des quatre isolats étudiés. Bien que les
analyses statistiques aient démontré des effets significatifs reliés aux concentrations salines, la croissance en
demeure tout de même similaire pour des teneurs salines avoisinant celles du Camembert. Ces expériences
ont cependant été réalisées en culture pure, ce qui ne permet pas d'évaluer l'effet du sel au sein de
l'écosystème réel. À titre d'exemple, la teneur en NaCl influence indirectement les interactions entre les
microorganismes, ce qui a été démontré entre G. candidum et P. roqueforti sur milieux gélosés (cheese agar)
(van den Tempel et Nielsen, 2000). Puisque G. candidum se développe bien en absence de sel, sa présence
aurait diminué et retardé la sporulation de P. roqueforti, contrairement à 4 % de NaCl où G. candidum n'a pu
se développer. Des essais en fromagerie ont donc été réalisés pour caractériser le comportement de
G. candidum et P. camemberti en mélange sur des fromages industriels.
Tableau 3.6 – Table d'ANOVA associée aux analyses statistiques sur la mesure du diamètre des colonies sur caillé modèle
Sources de variation Diamètre - Caillé modèle
DL F-Value p-Value Souches 3 29,77 <0,0001
Types de sel 2 1257,57 <0,0001 Concentrations 7 1774,46 <0,0001
Types de sel*concentrations 7 9,84 <0,0001 Souches*types de sel 6 2,56 0,0222
Souches*concentrations 21 1,14 0,3151 Souches*types de sel*concentrations 21 0,58 0,9277
Erreur 134
Total corrigé 201

81
Figure 3.2 – Influence du NaCl et du KCl sur la croissance (diamètre des colonies) de quatre souches de P. camemberti sur caillé modèle Camembert après 10 jours d'incubation à 25 C. Les souches sont représentées par quatre différentes couleurs; LMA-17 (bleu), LMA-30 (orange), LMA-435 (violet), LMA-1029 (vert). Les bandes hachurées représentent la croissance en absence de sel, les bandes foncées représentent la croissance en fonction du NaCl et les bandes pâles représentent la croissance en présence de KCl. Les données sont représentées par la moyenne ± l'erreur type. L'axe des abscisses indique le % en NaCl. Les teneurs en KCl ont été calculées sur une base équimolaire avec le NaCl. L'interaction double (types de sel*concentrations) est représentée sur la figure. Des lettres différentes pour une même concentration signifient un effet significatif (p < 0,05).
Tableau 3.7 - Comparaisons multiples sur l'interaction double souches*types de sel en conditionnant sur le type de sel
Souches Comparaisons statistiques
NaCl KCl Sans sel LMA-17 b a a LMA-30 a a a
LMA-435 ab a a LMA-1029 c b b
0
10
20
30
40
50
60
0 1 2 3 4 5 6 7 8
Diamètre des colonies (m
m)
Concentration en NaCl (%) ou équivalent molaire en KCl
LMA‐17 NaCl LMA‐30 NaCl LMA‐435 NaCl LMA‐1029 NaCl
LMA‐17 KCl LMA‐30 KCl LMA‐435 KCl LMA‐1029 KCl
a a
a a
a a
a b
a b
a b
a ba b

82
3.4.2. Suivi de la croissance de la microflore fongique d'affinage dans un fromage Camembert industriel
3.4.2.1. Analyses de composition
Le tableau 3.11 présente la composition en humidité, en cendres, en sodium et en potassium ainsi que le ratio
S/H des fromages en fonction des différents traitements salins. Pour l'ensemble des conditions (STD, RED et
KCL), les résultats ont montré une variation significative de la teneur en humidité en fonction du temps
d'affinage (p < 0,0001, tableau 3.8). Il y a une réduction de la teneur en humidité particulièrement importante
en début d'affinage pour les fromages standards et pour les fromages substitués par du potassium tandis
qu'une stabilité est atteinte pendant le reste de l'affinage. La réduction du taux d'humidité dans les fromages
réduits en sodium a cependant été visible jusqu'au jour 21. D'autres études ont observé une réduction de la
teneur en humidité dans des fromages de type Camembert en fonction du temps d'affinage (Schlesser et al.,
1992, Guizani et al., 2002, Leclercq-Perlat et al., 2004). De plus, les résultats démontrent que les traitements
salins (STD, RED et KCL) ont eu peu d'influence sur la teneur en humidité des fromages puisqu'aucune
différence significative n'y a été observée (p = 0,6280, tableau 3.8). Ces résultats sont en accord avec d'autres
études où aucune différence au niveau de la teneur en humidité n'a été observée entre des fromages
contrôles et des fromages comportant différents ratios d'un mélange de NaCl et de KCl, soit dans le Cheddar
(Fitzgerald et Buckley, 1985, Reddy et Marth, 1995), le Halloumi (Ayyash et Shah, 2010), le Nabulsi (Ayyash
et Shah, 2011), le Feta (Aly, 1995, Katsiari et al., 1997) et le Kefalograviera (Katsiari et al., 1998). Dans le
Camembert, la teneur en humidité s'est avérée inversement proportionnelle à la concentration en NaCl après
un jour de fabrication. Cependant, après 35 jours, la teneur en humidité de ces fromages était similaire, ce qui
a été attribué à l'évaporation superficielle de la phase aqueuse au cours de l'affinage (O'Nulain, 1986).
Pour l'ensemble des jours d'échantillonnage, une variation significative de la teneur minérale a été observée
entre les conditions STD, RED et KCL (p < 0,0001, tableau 3.8). La teneur en cendres dans les fromages de
conditions salines standards et substitués par du potassium était comparable tandis que celle des fromages
réduits en sodium s'y distinguait. La teneur minérale dans les fromages réduits en sodium était plus faible que
dans les deux autres conditions (STD et KCL) ce qui s'explique par la réduction du contenu en sel dans ceux-
ci. Une variation significative a également été observée au jour 5 entre le contenu en cendres dans les
fromages non salés (NSAL) ainsi que les fromages des conditions STD, RED et KCL (p < 0,0001, tableau
3.8), la teneur minéral étant plus faible dans ces contrôles puisqu'ils n'ont pas été saumurés. Une teneur
minérale plus élevée avait été observée dans les fromages enrichis en potassium comparativement au
standard lors des expériences d'optimisation (Chapitre 2), ce qui avait été attribué à une incorporation plus
rapide du potassium comparativement au sodium. Cependant, ces résultats n'ont pu être observés dans cette
étude. Ayyash et Shah (2010) ont également observé une corrélation positive entre la quantité de cendres

83
dans un fromage de type Halloumi et l'importance de la teneur en KCl. Les analyses statistiques ont
également démontré une variation significative de la teneur minérale, pour l'ensemble des conditions salines
(STD, RED, KCL) en fonction du jour d'échantillonnage (p = 0,0025, tableau 3.8). Les fluctuations étaient
variables d'un jour à l'autre, il était parfois possible d'y observer une augmentation ou une diminution en
fonction du temps d'affinage.
Pour l'ensemble des conditions STD, RED et KCL, aucune variation significative n'a été observée au niveau
des teneurs en sodium et en potassium en fonction du temps d'affinage (p = 0,0928, tableau 3.8 et p = 0,3040,
tableau 3.9, respectivement). Les résultats présentés au tableau 3.11 démontrent que le contenu en sodium et
en potassium est demeuré relativement stable en fonction du temps d'affinage. Pour l'ensemble des jours
d'échantillonnage, le contenu en potassium dans les fromages substitués avec du potassium était
significativement différent des fromages standards (p < 0,0001, tableau 3.9). Selon les résultats présentés au
tableau 3.11, le contenu en potassium était plus élevé dans les fromages substitués par du potassium que
dans les fromages standards, ce qui s'explique par l'absence de KCl dans la saumure utilisée pour produire
les fromages de condition standard. Pour l'ensemble des jours d'échantillonnage, aucune différence
significative entre le contenu en sodium des fromages réduits en sodium et substitués par du potassium n'a
été observée contrairement aux fromages de condition standard (p < 0,0001, tableau 3.8). La teneur en
sodium dans les fromages de condition saline standard était plus élevée que dans les deux autres conditions
(RED et KCL), ce qui s'explique par le temps de saumurage plus court dans les fromages réduits en sodium et
le contenu en NaCl plus faible dans les saumures utilisées pour produire les fromages substitués par du
potassium. Ensuite, l'analyse statistique comparant la teneur en sodium au jour 5 dans les fromages de
condition standard et celle des fromages contrôles industriels (IND) n'a démontré aucune différence
significative (p = 0,3926, non représenté dans la table d'ANOVA). De même, l'analyse statistique comparant
les teneurs en humidité de ces deux conditions (STD et IND) n'a démontré aucune différence significative (p =
0,3043, non représenté dans la table d'ANOVA). Ces résultats indiquent donc que la condition standard se
comparait aux critères industriels au niveau de la teneur en sodium et en humidité. En prenant en
considération que tout le sodium dosé provenait du NaCl, la teneur approximative des fromages standards
serait d'environ 1,38 % à 1,46 % (p/p) de NaCl, selon le jour d'échantillonnage. Ces concentrations se situent
donc légèrement en dessous des teneurs habituellement rencontrées dans le Camembert, soit entre 1,5 et
2,5 % (p/p) (Sousa, 2003). Ainsi, les ratios S/H calculés pour chaque jour d'échantillonnage sont également
faibles, mais les analyses statistiques effectuées au jour 5 n'ont démontré aucune différence significative entre
les fromages standards et les fromages contrôles industriels (p = 0,5483, non représenté dans la table
d'ANOVA). De plus, les ratios S/H des fromages standards et substitués par du potassium se situaient parmi
les valeurs moyennes rencontrées dans ce type de fromage, celles-ci oscillant entre 2,7 et 3,6 (Sutherland,
2002). Pour tous les jours d'échantillonnage, les ratios S/H calculés étaient plus élevés dans les fromages

84
substitués par du potassium que les standards; ils variaient entre 3,10 et 3,32 selon les jours
d'échantillonnage. Cependant, les comparaisons multiples présentées au tableau 3.11 entre ces deux
conditions ont démontré une différence significative au jour 14 seulement. Ces résultats peuvent s'expliquer
par la masse molaire plus élevée du KCl comparativement au NaCl puisque ce ratio est calculé sur une base
pondérale.
Les proportions de réduction en sodium dans les fromages réduits en sodium comparativement aux standards
variaient entre 22 % (jour 14) et 35 % (jour 21) ce qui se rapprochait de la cible établie de 30 %.
Parallèlement, les ratios S/H calculés étaient plus bas que dans les fromages standards et variaient entre 1,88
et 2,28 selon le jour d'échantillonnage. D'ailleurs les comparaisons multiples présentées au tableau 3.11 entre
les conditions à chacun des jours d'échantillonnage ont montré une différence significative entre les ratios S/H
des fromages standards et réduits en sodium. Les proportions de réduction en sodium dans les fromages
substitués par du potassium par rapport aux standards variaient entre 19 % (jours 14 et 35) et 27 % (jour 11)
et les ratios molaires en sodium et en potassium avoisinaient 3:1 (Na:K) pour chaque jour d'échantillonnage.
Pour certains jours (jours 14 et 35), la réduction en sodium dans les fromages réduits en sodium et dans les
fromages substitués par du potassium s'est avérée relativement plus faible que l'objectif visé de 30 %.
Plusieurs paramètres peuvent influencer la prise en sel des fromages, tel que les variations au niveau de la
composition des fromages avant saumurage, ainsi que les conditions de saumurage (Guinee et Fox, 2004,
Guinee et al., 2014b). Cela peut expliquer la variabilité obtenue au niveau des réductions en sodium puisque
des fromages différents étaient utilisés pour chacun des jours d'échantillonnage. Néanmoins, dans le contexte
de ce projet, les résultats obtenus étaient acceptables et se rapprochaient des cibles visées pour les
conditions RED et KCL.
Tableau 3.8 – Table d'ANOVA associée aux analyses statistiques sur la teneur en humidité, en cendres et en sodium des fromages
Sources de variation
DL
Humidité Cendres Sodium F-Value p-Value F-Value p-Value F-Value p-Value
Conditions 2 0,47 0,6280 131,08 <0,0001 90,85 <0,0001 Jours 4 11,20 <0,0001 5,10 0,0025 2,18 0,0928
Conditions*jours 8 0,69 0,6952 1,01 0,4460 0,82 0,5892 Conditions vs contrôle industriel | jour=5 1 0,8 0,3761 0,00 0,9840 8,17 0,0072 Conditions vs contrôle non salé | jour=5 1 0,42 0,5199 450,18 <0,0001 1508,63 <0,0001
Erreur 34
Total corrigé 50
Conditions = STD, RED et KCL
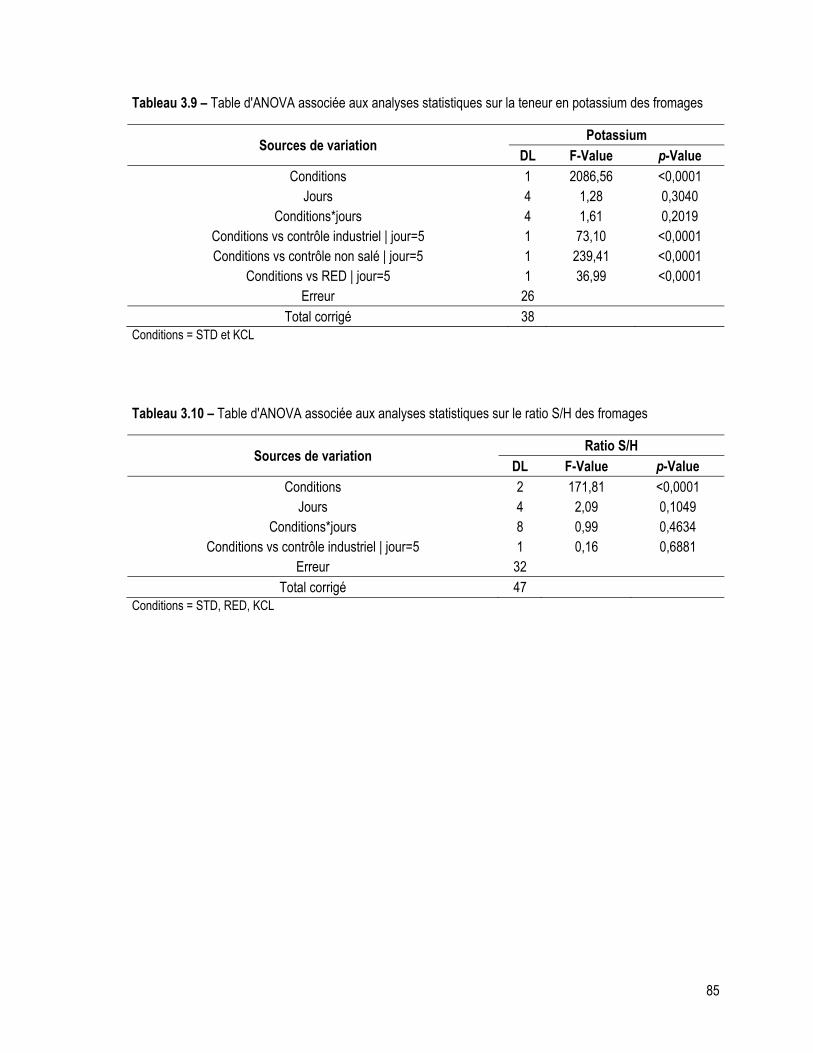
85
Tableau 3.9 – Table d'ANOVA associée aux analyses statistiques sur la teneur en potassium des fromages
Sources de variation Potassium
DL F-Value p-Value Conditions 1 2086,56 <0,0001
Jours 4 1,28 0,3040 Conditions*jours 4 1,61 0,2019
Conditions vs contrôle industriel | jour=5 1 73,10 <0,0001 Conditions vs contrôle non salé | jour=5 1 239,41 <0,0001
Conditions vs RED | jour=5 1 36,99 <0,0001 Erreur 26
Total corrigé 38
Conditions = STD et KCL
Tableau 3.10 – Table d'ANOVA associée aux analyses statistiques sur le ratio S/H des fromages
Sources de variation Ratio S/H
DL F-Value p-Value Conditions 2 171,81 <0,0001
Jours 4 2,09 0,1049 Conditions*jours 8 0,99 0,4634
Conditions vs contrôle industriel | jour=5 1 0,16 0,6881 Erreur 32
Total corrigé 47
Conditions = STD, RED, KCL

86
Tableau 3.11 – Analyse de la composition minérale et de la teneur en humidité des Camemberts en fonction des conditions de saumurage et du temps d'affinage.
Conditions salines
Cendres (%)
Humidité (%)
Na
(mg/g) K
(mg/g) Réduction
en sodium* (%) Ratio S/H**
Jour
5
STD 2,97b 49,53a 5,56b 0,88a 2,85b
RED 2,61a 49,36a 4,22a 0,93 24 2,17a
KCL 2,92b 49,14a 4,31a 2,37b 22 3,15b
IND 2,83 48,91 5,33 0,86 2,77
NSAL 1,79 49,03 0,27 1,04
Jour
11 STD 3,05b 47,71a 5,74b 0,72a 3,06b
RED 2,67a 48,46a 4,11a ND 28 2,15a
KCL 3,00b 48,34a 4,20a 2,31b 27 3,12b
Jour
14 STD 3,04b 47,73a 5,51b 0,79a 2,93b
RED 2,75a 47,97a 4,31a ND 22 2,28a
KCL 3,10b 47,87a 4,46a 2,38b 19 3,32c
Jour
21 STD 3,00b 47,30a 5,43b 0,77a 2,92b
RED 2,53a 47,63a 3,53a ND 35 1,88a
KCL 2,99b 47,71a 4,03a 2,36b 26 3,10b
Jour
35 STD 3,04b 47,28a 5,59b 0,85a 3,00b
RED 2,65a 46,66a 3,83a ND 31 2,08a
KCL 3,07b 47,77a 4,50a 2,23b 19 3,28b
Légende STD : Fromages salés selon les paramètres standards de l'industrie dans les saumures fabriquées dans des conditions standards. RED : Fromages réduits en sodium. KCL : Fromages ayant un contenu en sodium partiellement substitué par du potassium. IND : Contrôles; fromages salés dans la saumure industrielle selon les paramètres standards de l'industrie participante. NSAL : Fromages non saumurés. ND : Non dosé. * : En comparaison avec la condition STD. ** : Sel/humidité x 100; la teneur en sel (chlorure) a été calculée à partir de la quantité de Na et K dosée (Lindsay et al., 1982). Pour les conditions STD, RED et IND, seul le Na a été utilisé pour calculer ce ratio. Les lettres a, b et c font référence aux comparaisons multiples entre les conditions salines à chacun des jours d'échantillonnage. Des lettres identiques indiquent qu'il n'y a aucune différence significative entre les traitements salins pour un jour précis (p > 0,01).

87
3.4.2.2. Suivi du pH
La microflore fongique de surface des fromages à pâte molle est responsable de la désacidification de la pâte
sous la croûte et de l'établissement d'un gradient de pH de la surface, vers le centre. Leurs habiletés à
consommer l'acide lactique et à produire de l'ammoniac suite au catabolisme des acides aminés sont
responsables de cette neutralisation (Le Graët et al., 1983, Leclercq-Perlat et al., 2004, McSweeney, 2004,
Boutrou et Gueguen, 2005). Le suivi du pH au centre et à la surface des Camemberts a été évalué afin de
déterminer l'impact des conditions salines expérimentées sur l'alcalinisation de la pâte (Figure 3.3), ce qui
permet d'évaluer indirectement l'effet de ces trois conditions sur l'activité des ferments fongiques.
Pour les trois conditions salines, les résultats obtenus ont démontré une augmentation globale du pH du jour 5
au jour 35 ce qui correspond au profil normalement observé dans un fromage à pâte molle et à croûte fleurie
de type stabilisée (Champagne et al., 2003, Lessard et al., 2014). Pendant le procédé de fabrication,
l'utilisation de ferments lactiques thermophiles combinée à une réduction de la température lors de la
coagulation permet de limiter l'acidification du caillé et d'obtenir un pH post-production plus élevé que dans
une pâte traditionnelle, soit autour de 5,2 (Lawrence et al., 1987). Les pH initiaux après production (jour 1,
avant saumurage) étaient de 5,07 ± 0,01 au centre et de 5,11 ± 0,03 en surface ce qui se rapprochait de la
valeur ciblée lors de la production de ce type de fromage. Ainsi, pour chaque condition saline, le pH en surface
est demeuré stable jusqu'au jour 5, tandis qu'une légère post-acidification a été visible au centre du fromage.
Les résultats obtenus ont démontré que, pour l'ensemble des jours d'échantillonnage, l'évolution du pH à la
surface, ou au centre des fromages, variait peu en fonction des conditions salines expérimentées (p = 0,3212
et p = 0,1750, tableau 3.12). Cependant, les analyses statistiques présentées à la figure 3.3 ont démontré une
différence significative entre le pH au centre des fromages réduits en sodium et les deux autres conditions aux
jours 7 et 14. Sachant que la teneur en sel module la capacité des bactéries lactiques (genre Lactococcus) à
fermenter le lactose en acide lactique (Irvine et Price, 1961), cet écart pourrait y être relié. Un écart significatif
du pH en surface a été observé au jour 21 entre les fromages substitués par du potassium et les fromages
standards, celui-ci étant plus faible dans cette dernière. Les causes exactes de cet écart demeurent
inconnues, mais pourraient être reliées au métabolisme des ferments fongiques. Parallèlement à cette
observation, la densité cellulaire quantifiée chez G. candidum était significativement plus élevée dans les
fromages substitués par du potassium (voir 3.4.2.3). Cependant, de façon générale, les modifications salines
effectuées dans cette étude semblent avoir eu peu d'impact sur l'aptitude de G. candidum et P. camemberti à
alcaliniser la pâte du fromage. Ces résultats sont en lien avec les conclusions issues d'une autre étude où peu
de variations du pH ont été observées au centre et à la surface de Camemberts affinés pendant 35 jours et
comportant différentes teneurs salines comprises entre 0 et 2,49 % de NaCl (O'Nulain, 1986).

88
Tableau 3.12 – Table d'ANOVA associée aux analyses statistiques du pH
Sources de variation
DL
pH surface pH centre F-Value p-Value F-Value p-Value
Conditions 2 1,17 0,3212 1,82 0,1750 Jours 6 243,92 <0,0001 52,13 <0,0001
Conditions*jours 12 4,3 0,0002 4,85 <0,0001 Erreur 42
Total corrigé 62
Figure 3.3 – Évolution du pH à la surface (gauche) et au centre (droite) des fromages industriels en fonction de différentes teneurs salines. Chaque couleur représente une condition saline différente; standard (noir), réduit en NaCl (gris) et réduit en NaCl et substitué par du KCl (bleu). L'affinage a été réalisé à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. L'interaction double (conditions*jours d'échantillonnage) est représentée sur chacun des graphiques. Des lettres identiques signifient qu'il n'y a aucune différence significative entre les pH en fonction des conditions salines, pour un jour précis (p > 0,01).
4,5
5
5,5
6
6,5
5 15 25 35
pH
a,a,aa,a,a
a,a,a
a,a,a
a,a,a
b,ab,a
a,a,a
96,5 % HR
12,5 C4 C
4,5
5
5,5
6
6,5
5 15 25 35
a,a,aa,b,a
a,a,a
a,b,a
a,a,a
a,a,a
96,5 % HR
12,5 C4 C
a,a,a
Affinage (jours)
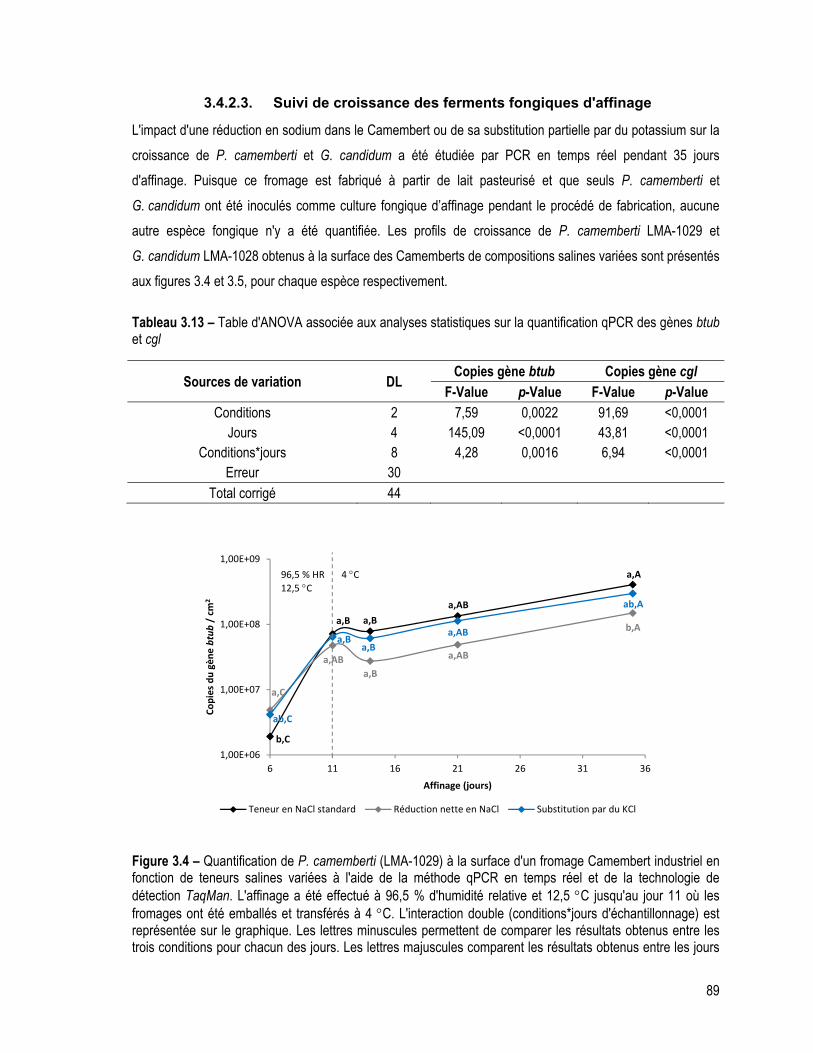
89
3.4.2.3. Suivi de croissance des ferments fongiques d'affinage
L'impact d'une réduction en sodium dans le Camembert ou de sa substitution partielle par du potassium sur la
croissance de P. camemberti et G. candidum a été étudiée par PCR en temps réel pendant 35 jours
d'affinage. Puisque ce fromage est fabriqué à partir de lait pasteurisé et que seuls P. camemberti et
G. candidum ont été inoculés comme culture fongique d’affinage pendant le procédé de fabrication, aucune
autre espèce fongique n'y a été quantifiée. Les profils de croissance de P. camemberti LMA-1029 et
G. candidum LMA-1028 obtenus à la surface des Camemberts de compositions salines variées sont présentés
aux figures 3.4 et 3.5, pour chaque espèce respectivement.
Tableau 3.13 – Table d'ANOVA associée aux analyses statistiques sur la quantification qPCR des gènes btub et cgl
Sources de variation DL Copies gène btub Copies gène cgl
F-Value p-Value F-Value p-Value Conditions 2 7,59 0,0022 91,69 <0,0001
Jours 4 145,09 <0,0001 43,81 <0,0001 Conditions*jours 8 4,28 0,0016 6,94 <0,0001
Erreur 30
Total corrigé 44
Figure 3.4 – Quantification de P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel en fonction de teneurs salines variées à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. L'interaction double (conditions*jours d'échantillonnage) est représentée sur le graphique. Les lettres minuscules permettent de comparer les résultats obtenus entre les trois conditions pour chacun des jours. Les lettres majuscules comparent les résultats obtenus entre les jours
1,00E+06
1,00E+07
1,00E+08
1,00E+09
6 11 16 21 26 31 36
Copies du gène btub/ cm
2
Affinage (jours)
Teneur en NaCl standard Réduction nette en NaCl Substitution par du KCl
b,C
a,C
ab,C
a,B
a,B
a,AB
a,B
a,B
a,B
a,AB
a,AB
a,AB
a,A
ab,A
b,A
96,5 % HR 4 C12,5 C

90
d'échantillonnage pour chaque condition. Des lettres identiques indiquent qu'il n'y a aucune différence significative (p > 0,01).
Figure 3.5 – Quantification de G. candidum (LMA-1028) à la surface d'un fromage Camembert industriel en fonction de teneurs salines variées à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C. L'interaction double (conditions*jours d'échantillonnage) est représentée sur le graphique. Les lettres minuscules permettent de comparer les résultats obtenus entre les trois conditions pour chacun des jours. Les lettres majuscules comparent les résultats obtenus entre les jours d'échantillonnage pour chaque condition. Des lettres identiques indiquent qu'il n'y a aucune différence significative (p > 0,01).
3.4.2.3.1. Croissance dans les Camemberts standards
Une phase active de croissance a été observée en début d'affinage chez les deux espèces due à l'importance
de la biomasse cellulaire quantifiée au jour 6. Une légère réduction de croissance, celle-ci n'étant toutefois pas
significative, est ensuite observée entre les jours 11 à 14 ce qui pourrait être attribué au changement de
métabolisme de P. camemberti après épuisement du lactose (Leclercq-Perlat et al., 2013). En fait, selon
Leclercq-Perlat et coll. (2013), la croissance du mycélium de P. camemberti continue tant qu'il y a du lactose
et une réduction du taux de croissance est ensuite observée, ce qui serait attribué à la mort cellulaire et au
début de la sporulation. Selon la figure 3.4, l'évolution de croissance de P. camemberti s'est ensuite poursuivie
lentement jusqu'au jour 35 pour atteindre un maximum de 4,06 x 108 copies de gènes/cm2. Chez G. candidum,
une densité cellulaire maximale de 9,41 x 107 copies de gène/cm2 a été atteinte au jour 11 puis une légère
diminution a été observée jusqu'au jour 35 (Figure 3.5). Toutefois, les analyses statistiques présentées à la
1,00E+06
1,00E+07
1,00E+08
1,00E+09
6 11 16 21 26 31 36
Copies du gènecgl / cm
2
Affinage (jours)
Teneur en NaCl standard Réduction nette en NaCl Substitution par du KCl
96,5 % HR 4 C12,5 C
c,B
a,A
b,B
a,A
a,A
a,A a,A
ab,A
b,Ab,A
a,A
a,A
b,A
ab,A
a,A

91
figure 3.5 n'ont démontré aucune variation significative entre les jours 11 et 35 chez cette espèce. La
proportion des deux espèces était similaire aux jours 6, 11 et 14 tandis que P. camemberti devenait l'espèce
majoritaire pour les jours suivants (Annexe 1). Au jour 35, une différence de plus d'un log a été observée entre
les dénombrements cellulaires des deux espèces. Le profil de croissance obtenu chez ces deux espèces
correspond à ce qui a été observé à la surface d'un Camembert industriel identique à celui utilisé dans cette
étude (Lessard et al., 2014). Lessard et coll. (2014) ont observé une phase active de croissance chez les deux
espèces dans les cinq premiers jours d'affinage, puis une évolution lente de celles-ci jusqu'en fin d'affinage. La
dominance au niveau du dénombrement cellulaire de P. camemberti a été visible à partir du jour 15.
3.4.2.3.2. Croissance dans les Camemberts réduits en sodium
Le comportement des deux espèces a été étudié dans des conditions réduites en sodium. Comparativement
aux fromages standards, les proportions de réduction en sodium variaient de 22 à 35 % (p/p), selon le jour
d'échantillonnage (Tableau 3.11). Les teneurs en NaCl approximatives calculées à partir du contenu en
sodium dosé variaient donc entre 0,90 et 1,09 % (p/p).
Le profil de croissance de P. camemberti observé dans les fromages réduits en sodium s'est avéré similaire à
celui obtenu dans les fromages standards. Une phase active de croissance a été visible en début d'affinage.
Tel qu'observé à la condition saline standard, une diminution du nombre de copies de gènes a été observée
entre les jours 11 et 14, mais celle-ci n'étant toutefois pas significativement différente. L'augmentation de la
biomasse cellulaire s'est ensuite poursuivie plus lentement au cours de l'affinage pour atteindre un maximum
de 1,49 x 108 copies de gène/cm2 au jour 35. D'ailleurs, les analyses statistiques présentées à la figure 3.4 ont
démontré une variation significative entre les jours 14 et 35. Comparativement au profil de croissance obtenu
dans la condition standard, la densité cellulaire de P. camemberti s'est avérée significativement plus élevée en
début d'affinage (jour 6) suggérant une phase active de croissance plus rapide à ce moment dans les
fromages réduits en sodium. Or, il s'agit du seul temps où la densité cellulaire fût plus importante. En général,
le dénombrement cellulaire a été moins important en condition réduite en sodium. Pour les jours 11 à 35, seul
le jour 35 a démontré une différence significative. Cependant, les différences observées aux jours 6 et 35
étaient relativement faibles. Le dénombrement moléculaire, comparativement à la condition standard,
divergeait de moins d'un log, ce qui est relativement faible d'un point de vue biologique.
Le profil de croissance observé chez G. candidum dans la condition réduite en sodium a été différent de celui
obtenu dans la condition standard (Figure 3.5). En fait, les analyses statistiques visibles à la figure 3,5 ont
démontré que la population de G. candidum est demeurée stable entre les jours 6 à 35. Pendant tous les jours
d'affinage étudiés, la croissance de G. candidum a été favorisée dans les fromages réduits en sodium, lorsque
comparée à la condition standard. Cet effet a été particulièrement important en début d'affinage suggérant une

92
phase active de croissance plus rapide. Une différence significative a d'ailleurs été observée au jour 6, le
dénombrement cellulaire dans les fromages standards était de 1,86 x 106 copies de gène/cm2 et de 1,32 x 108
copies de gènes/cm2 dans les fromages réduits en sodium, ce qui représentait un écart de près de deux logs.
Cependant, comme observé chez P. camemberti, les différences obtenues aux jours 14, 21 et 35,
comparativement au dénombrement moléculaire à la surface des fromages standards, étaient près ou
inférieures à un log.
Ainsi, les résultats ont démontré que l'effet du sel sur la croissance des mycètes a été particulièrement
important dans les débuts de la période d'affinage. Dans les fromages salés par saumurage ou en surface, la
majorité du sel (ou du sodium) en début d'affinage se retrouve concentrée en surface pour ensuite migrer à
travers la pâte jusqu'à l'atteinte d'une quasi-homogénéité (Le Graet et Brulé, 1988, Guizani et al., 2002, Hardy,
2009). Ainsi, une exposition à des teneurs en sodium plus élevées au jour 6 pourrait expliquer les différences
au niveau du dénombrement cellulaire chez P. camemberti et G. candidum. L'effet étant beaucoup plus
marqué chez G. candidum à ce moment, cette observation peut être expliquée par la sensibilité plus
importante de cette espèce face aux conditions salines. Sachant que cette espèce s'implante à la surface du
Camembert dans les trois premiers jours d'affinage (Leclercq-Perlat et al., 2004), sa croissance aurait été
nettement avantagée dans les fromages réduits en sodium pendant cette période.
Dans des conditions standards, la microflore fongique de surface est habituellement dominée par
P. camemberti, ce qui a d'ailleurs été observé dans les études connexes où la quantification de l'écosystème
fongique de surface a été réalisée par PCR en temps réel à partir d'un CMC (Lessard et al., 2012) et d'un
Camembert industriel (Lessard et al., 2014). Les résultats présentés aux figures 3.4 et 3.5 ainsi qu'aux
annexes 1 et 2 ont permis d'observer un débalancement au niveau de la microflore fongique quantifiée. Dans
les fromages réduits en sodium, G. candidum représente l'espèce majoritaire, à l'exception du jour 35 où la
présence des deux espèces était équivalente (Annexe 2). Le sel est reconnu pour son action sélective au
niveau du développement de la microflore fongique de surface des fromages à pâte molle. Préférant de faibles
teneurs salines, la croissance de G. candidum dans les fromages réduits en sodium pourrait avoir été
avantagée au détriment de P. camemberti, causant ainsi ce débalancement. De plus, la capacité de
G. candidum à limiter le développement de P. camemberti à la surface de fromage à pâte molle et à croûte
fleurie a préalablement été démontrée (Vassal et Gripon, 1984) ce qui pourrait également expliquer la
diminution du dénombrement de cette moisissure dans la condition réduite en sodium (jours 14 à 35). Une
prolifération plus importante de G. candidum peut aussi être responsable de l'apparition de défauts de
fromagerie. Cependant, aucun défaut n'a été détecté visuellement dans les fromages, tel que le défaut de
peau de crapaud pouvant être causé par une prolifération rapide de G. candidum entravant ainsi l'implantation
de P. camemberti (Spinnler et Gripon, 2004).

93
3.4.2.3.3. Croissance dans les Camemberts partiellement substitués par du potassium
Le comportement des deux espèces a été étudié dans un Camembert industriel où une portion du sodium a
été remplacée par du potassium. Les réductions en sodium variaient entre 19 et 27 % selon le jour
d'échantillonnage (tableau 3.11). Selon le contenu en sodium et potassium dosé à chaque jour
d'échantillonnage, les teneurs en NaCl calculées variaient entre 1,02 et 1,14 % (p/p), tandis que les teneurs en
KCl s'étendaient entre 0,43 et 0,45 % (p/p). Le ratio molaire de sodium et de potassium avoisinait 3:1 (Na:K)
pour chaque jour d'échantillonnage.
Le profil de croissance de P. camemberti a été similaire à celui observé en condition standard et en condition
réduite en sodium. Une phase active de croissance a été visible en début d'affinage dû à l'importance de la
biomasse cellulaire quantifiée au jour 6. Ensuite, une légère réduction a été visible entre les jours 11 et 14.
Cependant, les analyses statistiques ne démontrent aucune différence significative au niveau du
dénombrement cellulaire à ces deux jours. Pour les jours suivants, la croissance s'est poursuivie plus
lentement jusqu'au jour 35 où une densité cellulaire maximale de 2,97x 108 copies de gène/cm2 a été atteinte.
Pour chacun des jours d'échantillonnage, les analyses statistiques présentées à la figure 3.4 n'ont démontré
aucune différence significative entre la quantité de gènes quantifiés dans les fromages substitués par du
potassium et les fromages standards. Ainsi, malgré le fait que la croissance de P. camemberti dans les
fromages réduits en sodium ne soit que légèrement différente, il semble que la substitution du sodium par du
potassium ait permis de rétablir le profil de croissance initial de P. camemberti. De plus, les analyses
statistiques n'ont démontré aucune différence significative entre le dénombrement cellulaire obtenu à la
surface des fromages réduits en sodium et celui des fromages dont le sodium a été substitué par du
potassium.
Le profil de croissance de G. candidum dans les fromages substitués par du potassium, montre un profil
intermédiaire en qPCR lorsque comparé aux deux autres conditions (Figure 3.5). L'effet de la substitution du
sel a été surtout marqué au début de l'affinage. Le dénombrement moléculaire au jour 6 était équivalent à
1,97 x 107 copies de gène/cm2. Ainsi, le nombre de copie du gène cgl détecté était d’un log supérieur à la
condition standard et un log inférieur à la condition réduite en sodium sans remplacement. D'ailleurs les
analyses statistiques présentées ont démontré une différence significative entre les trois conditions pour ce
jour d'échantillonnage. La croissance de G. candidum a donc été plus rapide dans les débuts d'affinage
comparativement à la condition standard, ce qui peut s'expliquer par l'exposition à une teneur en sodium
moins élevée. Cependant, l'effet du potassium sur cette espèce n'est pas documenté. Les résultats semblaient
toutefois démontrer que l'effet inhibiteur de ce dernier est moins important que celui du sodium. Par contre,

94
pour les jours suivants, peu de variations significatives ont été observées comparativement à la condition STD,
soit seulement au jour 21 avec une différence de 0,54 log.
Au niveau du balancement entre les deux espèces fongiques, un profil similaire à celui observé dans les
conditions standards a pu être observé, mais la dominance de P. camemberti apparaît beaucoup plus tard
dans l'affinage des fromages substitués, soit au jour 35 (Annexe 3). Pour les jours précédents, le
dénombrement moléculaire de G. candidum est légèrement plus élevé entre les jours 6 et 14. Ensuite, une
biomasse équivalente est observée au jour 21. P. camemberti est majoritairement responsable de la formation
de peptides amers pendant l'affinage dû à son activité protéolytique, tandis que G. candidum est reconnu pour
son pouvoir désamérisant (Vassal et Gripon, 1984, Molimard et al., 1994, Boutrou et Gueguen, 2005). Tout
comme dans les fromages réduits en sodium, il serait donc pertinent de vérifier l'impact de ces
débalancements au niveau de la microflore fongique sur le développement des propriétés sensorielles, soit la
formation des composés aromatiques.

95
3.5. CONCLUSION ET PERSPECTIVES
La sensibilité intra-espèce des quatre souches de P. camemberti étudiées était similaire face au NaCl et au
KCl sur milieu de culture synthétique et sur CMC. Chacune des souches montre une meilleure croissance à de
faibles teneurs salines, soit entre 0 et 3 % de sel selon la souche et le milieu de culture. À des concentrations
avoisinant les teneurs salines standards du Camembert, le NaCl et le KCl ont eu un effet similaire sur la
croissance de chacune des souches tandis que le potentiel inhibiteur du NaCl était plus important à des
concentrations supérieures. La réduction du sodium ou son remplacement par du potassium dans un
Camembert industriel a démontré peu d'effet sur la teneur en humidité et l'alcalinisation de la pâte des
fromages au cours de l'affinage. Ces conditions ont légèrement influencé le profil de croissance de
P. camemberti au début et à la fin de l'affinage tandis que celui de G. candidum a été affecté plus fortement en
début d'affinage. Or, l'impact des modifications salines sur l'activité spécifique (expression des gènes,
synthèse d’enzymes, activités des enzymes, etc.) des deux espèces n'est pas connu. Des observations
préalablement réalisées dans un fromage Camembert ont démontré une corrélation positive entre la réduction
de la protéolyse et l'augmentation de la concentration en NaCl (O'Nulain, 1986). P. camemberti et
G. candidum participent activement au catabolisme des protéines pendant l'affinage, ce qui contribue au
développement de la texture et des propriétés organoleptiques. Sachant que la protéolyse associée à ces
deux espèces se produit dans les premières semaines d'affinage (Lessard et al., 2014) et que l'effet du sel sur
leur croissance a été plus prononcé à ce moment, il serait intéressant d'en évaluer l’impact.


97
Conclusion générale et perspectives
Le but de ce projet de maîtrise était de déterminer l'impact d’une réduction du NaCl dans le Camembert et de
sa substitution partielle par du KCl sur la croissance des ferments fongiques P. camemberti et G. candidum.
Dans un premier temps, une étude de la cinétique d'incorporation des ions sodium et potassium pendant le
saumurage d'un Camembert industriel a été réalisée. Ainsi, les paramètres de saumurage nécessaires à la
production de fromages de conditions salines standards, réduits en NaCl et partiellement substitués par du
KCl ont été identifiés en industrie. L’utilisation de fromages provenant d’une fromagerie produisant à grande
échelle a permis de vérifier la faisabilité de la réduction du sodium dans des conditions proches de la réalité
industrielle. L'effet causé par ces modifications salines sur la croissance des deux microorganismes formant le
ferment d’affinage d'intérêt a donc pu être déterminé au sein de leur écosystème. De plus, étant peu
documentée, la variabilité intra-espèce de la résistance au sel chez P. camemberti a été déterminée ce qui a
permis de comprendre davantage le comportement de cette espèce en présence de NaCl et de KCl lorsque
cultivée en culture pure.
Cinétique d'incorporation du sodium et du potassium dans un Camembert industriel saumuré
L'étude de l'incorporation du sodium et du potassium lors du saumurage du Camembert a permis de
démontrer la complexité associée à cette technique et d'identifier quelques-uns des nombreux facteurs
pouvant affecter la prise en sel. En effet, une variabilité importante au niveau de l'incorporation du sel a été
observée entre les différents essais de saumurage ce qui peut être attribuable à la composition de l'aliment
entre les lots de production, aux variations dans la constitution des saumures et des conditions de saumurage,
ainsi qu'au faible nombre de répétitions effectuées dû aux contraintes associées à la portion industrielle du
projet. Toutefois, les conditions ciblées ont pu être obtenues, avec quelques déviations, surtout visibles au
niveau des fromages salés en saumure mixte (mélange de NaCl et KCl). De plus, il a été possible de
comprendre davantage les phénomènes liés à l'incorporation du potassium dans la matrice fromagère lors du
saumurage du Camembert, ceux-ci n'étant pas documentés pour cette variété de fromage.
Sachant que le contenu en sel des fromages influence la synérèse, il serait pertinent d'évaluer l'impact de ces
nouveaux paramètres de saumurage sur la composition globale de l'aliment, par exemple sur le contenu en
protéine et en matière grasse. Cette portion du projet est en cours de réalisation par Mme Christine Vallières,
ce qui permettra d’obtenir ces importantes informations. De plus, bien que le présent projet ait permis de
comprendre davantage les phénomènes liés à l'absorption du sel lors du saumurage, il serait intéressant de
déterminer le profil de migration de ces sels pendant la phase d'affinage. En fait, les sels incorporés dans la
pâte suite au salage se déplacent dans la matrice, de la surface vers le centre du fromage, tandis qu'une

98
migration des molécules d'eau vers la surface est observée pendant la même période. Puisqu'une absorption
plus rapide du potassium dans la matrice fromagère a été suspectée lors du saumurage, il se pourrait que le
profil de migration de cet élément dans la pâte ne soit pas le même que le profil de migration du sodium.
Puisque les ions interagissent avec les constituants de la matrice fromagère, tels que les caséines, les
enzymes et les microorganismes, il serait pertinent d'en évaluer l'effet au niveau de la texture, de l'activité
enzymatique et du développement microbien au centre de l'aliment.
Variabilité intra-spécifique de la résistance au NaCl et au KCl chez P. camemberti
Le deuxième objectif de ce projet de recherche a permis de comprendre davantage le comportement
spécifique de P. camemberti en fonction de la concentration en sel et de comparer le profil de croissance
selon la nature des composés, c'est-à-dire le NaCl et le KCl. La tolérance au NaCl moyennement élevée chez
P. camemberti avait préalablement été documentée. Cependant, les études disponibles sont peu nombreuses;
elles ont essentiellement été réalisées sur milieux de culture synthétiques tandis que la présente étude a
permis de comprendre le comportement de cette espèce dans un environnement plus près du Camembert par
l'utilisation de caillés modèles. De plus, la variabilité intra-espèce de la résistance au sel de cette moisissure
était peu connue et aucune information concernant l'effet du KCl n'était disponible.
Les résultats obtenus lors du deuxième objectif étaient en accord avec les informations disponibles dans la
littérature; l'optimum de croissance des quatre isolats utilisés se situait à de faibles concentrations et elles ont
chacune démontré une tolérance moyennement élevée face aux deux sels. Seulement une des quatre
souches utilisées a démontré une sensibilité plus prononcée au NaCl et au KCl sur PDA, tandis que cette
observation a été moins marquée sur caillé modèle. Ainsi, à la lumière des résultats obtenus, il semblerait y
avoir peu de variabilité intra-espèce au niveau de la résistance au sel entre les isolats utilisés de
P. camemberti. Finalement, les résultats générés ont démontré que le NaCl possédait un effet inhibiteur plus
important sur le développement des quatre souches à des concentrations salines dépassant 3 à 5 % NaCl.
Bien que cette observation ait été faite à des concentrations supérieures à celles normalement retrouvées
dans un fromage de type Camembert, il serait intéressant de comprendre la cause de ce phénomène dans un
contexte plus fondamental, c'est-à-dire, au niveau du fonctionnement, du métabolisme ou de l'intégrité de la
cellule fongique.
Afin de poursuivre la recherche à partir des données obtenues pour cet objectif, il serait intéressant de valider
la capacité de chacun des sels à réduire l'aw dans chacun des milieux utilisés puisqu'en réalité, c'est ce
paramètre qui gouverne majoritairement le comportement microbien dans les aliments. De plus, la variabilité
intra-espèce de la résistance à ces deux sels pourrait être étudiée plus en profondeur chez G. candidum. Plus
précisément, la croissance de cette levure en fonction du NaCl est assez bien documentée, mais le profil de

99
croissance de cette espèce en fonction du KCl n'a pas été étudié. De plus, la tolérance au NaCl et au KCl
pourrait être déterminée chez d'autres ferments fongiques participant à l'affinage des fromages à pâte molle et
à croûte fleurie, tels que Kluyveromyces lactis, Debaryomyces hansenii, Yarrowia lipolitica et plusieurs autres.
Les producteurs fromagers qui désirent réduire le contenu en sel de leur fromage ou le substituer par du KCl
pourraient bénéficier de cette information et s'en servir à titre de critère de sélection.
Étude de la réduction du NaCl dans un Camembert industriel et de sa substitution partielle par du KCl sur la croissance des ferments fongiques
Le troisième objectif de ce projet a permis de déterminer l'effet causé par une réduction du NaCl dans un
Camembert industriel et de son remplacement par du KCl sur la croissance de P. camemberti et de
G. candidum. Les informations disponibles à ce sujet étaient peu nombreuses et plutôt corrélées avec le
développement de P. camemberti et non de l'écosystème fongique dans son ensemble. De plus, selon les
informations disponibles dans la littérature, aucune étude de ce genre n'avait été réalisée dans un Camembert
industriel de type stabilisé.
À la lumière des résultats obtenus, l'hypothèse émise au début du projet de recherche est en partie réfutée.
En effet, les modifications salines effectuées ont eu un impact mineur sur la croissance de P. camemberti,
contrairement à ce qui a été observé chez G. candidum. Plus précisément, la réduction du contenu en NaCl a
provoqué une légère baisse du taux de croissance chez P. camemberti, tandis que la croissance de
G. candidum a été nettement avantagée en début d'affinage. La substitution du NaCl par le KCl a également
causé un effet spécifique à chacune des espèces étudiées. Elle a permis de rétablir le profil de croissance
chez P. camemberti, mais la vitesse de croissance de G. candidum, surtout en début d'affinage, a été
augmentée dans cette condition. Un autre fait intéressant a été remarqué lors de cette étude, soit le
débalancement au niveau de la microflore des fromages réduits en NaCl, ce qui fait en sorte que G. candidum
est présente de façon majoritaire tout au long de l'affinage, à l'exception du jour 35 où sa présence est
équivalente à P. camemberti. Au contraire, la microflore fongique dans cette variété de fromage est
habituellement dominée par P. camemberti. Considérant les rôles joués par chacune des espèces lors de
l'affinage, il serait pertinent de vérifier l'impact de ce débalancement au niveau de la qualité globale de
l'aliment.
Les résultats obtenus dans cet objectif du projet laissent place à de nombreuses perspectives de recherche.
En premier lieu, l'effet engendré par ces modifications salines sur l'activité des deux ferments d'intérêt devrait
être évalué plus en détails. Sachant que ces espèces participent activement à la lipolyse et à la protéolyse, il
serait pertinent d'évaluer les changements induits par une modification des conditions salines sur la production
de lipases et de protéases. Parallèlement, une évaluation sensorielle pourrait être réalisée par un panel de

100
dégustateur entraîné. Ainsi, il serait possible de corréler les changements observés au niveau de l'activité de
chacune des espèces avec les propriétés organoleptiques de l'aliment.
Ensuite, puisque cette partie du projet s'est attardée spécifiquement à l'étude de P. camemberti et
G. candidum, il serait pertinent d'évaluer les conséquences engendrées par ces modifications salines sur la
croissance et l'activité des autres ferments d'affinage, sur la microflore lactique, ainsi que sur la microflore
d’affinage secondaire. À titre d'exemple, l'osmotolérance connue chez la levure D. hansenii pourrait également
causer un débalancement au niveau de la microflore d'affinage lors d'une modification des conditions salines
de l'aliment. Puisque cette espèce participe également à l'affinage du fromage, étant ajoutée dans certains
ferments d’affinage plus complexes, elle pourrait ainsi provoquer une modification des propriétés sensorielles
générées.

101
Annexes
L'annexe 1 représente l'évolution de P. camemberti et G. candidum pendant l'affinage d'un Camembert
industriel de teneur saline standard. L'évolution des deux espèces est équivalente en début d'affinage tandis
que P. camemberti devient l'espèce majoritaire après le jour 14. Ce profil de croissance correspond à ce qui
est normalement observé dans ce type de fromage (Lessard et al., 2012 et 2014).
Annexe 1 – Quantification de G. candidum (LMA-1028) et P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel de teneur saline standard à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C.
1,00E+06
1,00E+07
1,00E+08
1,00E+09
6 11 16 21 26 31 36
Copies de gène/cm
2
Affinage (jours)
P. camemberti
G. candidum

102
L'annexe 2 représente le profil de croissance de P. camemberti et G. candidum dans un Camembert industriel
dont le contenu en sodium est réduit de 24 à 35 % (tableau 3.11), selon le jour d'échantillonnage. Il est
possible d'observer un débalancement au niveau de la microflore fongique puisque, dans ce cas, G. candidum
constitue l'espèce majoritaire alors que P. camemberti domine normalement l'écosystème fongique de ce type
de fromage.
Annexe 2 – Quantification de G. candidum (LMA-1028) et P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel réduit en NaCl à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C.
1,00E+06
1,00E+07
1,00E+08
1,00E+09
6 11 16 21 26 31 36
Copies de gène/cm
2
Affinage (jours)
P. camemberti
G. candidum

103
L'annexe 3 représente le profil de croissance de P. camemberti et G. candidum dans un Camembert industriel
réduit de 22 à 27 % de NaCl (tableau 3.11) et substitué par du KCl. Un débalancement au niveau de la
microflore fongique est observable puisque la dominance de l'espèce G. candidum est visible jusqu'au jour 21.
Le profil habituel, dominé par P. camemberti, semble se rétablir pour les jours suivants, soit beaucoup plus
tard que dans un fromage de condition saline standard (Annexe 1).
Annexe 3 – Quantification de G. candidum (LMA-1028) et P. camemberti (LMA-1029) à la surface d'un fromage Camembert industriel réduit en NaCl et substitué par du KCl à l'aide de la méthode qPCR en temps réel et de la technologie de détection TaqMan. L'affinage a été effectué à 96,5 % d'humidité relative et 12,5 C jusqu'au jour 11 où les fromages ont été emballés et transférés à 4 C.
1,00E+06
1,00E+07
1,00E+08
1,00E+09
6 11 16 21 26 31 36
Copies de gène/cm
2
Affinage (jours)
P. camemberti
G. candidum


105
Bibliographie
Agriculture et Agroalimentaire Canada (2013). Les efforts de réduction du sodium entrepris par l'industrie agroalimentaire canadienne. [En ligne]: http://www.agr.gc.ca/fra/industrie-marches-et-commerce/reglements-sur-les-aliments/enjeux-strategiques-et-reglementaires-en-alimentation/rapports-et-ressources/les-efforts-de-reduction-du-sodium-entrepris-par-lindustrie-alimentaire-canadienne/?id=1363026410711. Consulté le 15 janvier 2014.
Al-Samarrai, T.H. et J. Schmid (2000). A simple method for extraction of fungal genomic DNA, Letters in applied microbiology, 30(1):53-56.
Alper, I., M. Frenette et S. Labrie (2013). Genetic diversity of dairy Geotrichum candidum strains revealed by multilocus sequence typing, Applied microbiology and biotechnology, 97(13):5907-5920.
Aly, M. E. (1995). An attempt for producing low-sodium Feta-type cheese, Food Chemistry, 52(3):295-299.
AOAC (2005). AOAC official method 935.42: Ash of cheese. Association of Official Analytical Chemists.
AOAC (2008). AOAC official method 926.08: Loss on drying (moisture) in cheese. Association of Official Analytical Chemists.
Ardö, Y., S. Skeie et T. Guinee (2014). Salt in cheese ripening, dans The importance of salt in the manufacture and ripening of cheese, P. Paquin et S. Labrie (eds.), International Dairy Federation, pages 21-29.
Ayyash, M.M. et N.P. Shah (2010). Effect of partial substitution of NaCl with KCl on Halloumi cheese during storage: Chemical composition, lactic bacterial count, and organic acids production, Journal of food science, 75(6):C525-C529.
Ayyash, M.M. et N.P. Shah (2011). The effect of substituting NaCl with KCl on Nabulsi cheese: Chemical composition, total viable count, and texture profile, Journal of dairy science, 94(6):2741-2751.
Aziza, M., C. Couriol, A. Amrane et R. Boutrou (2005). Evidences for synergistic effects of Geotrichum candidum on Penicillium camembertii growing on cheese juice, Enzyme and microbial technology, 37(2):218-224.
Back, J.P., S.A. Langford et R.G. Kroll (1993). Growth of Listeria monocytogenes in Camembert and other soft cheeses at refrigeration temperatures, Journal of dairy research, 60(3):421-429.
Bidlas, E. et R.J.W. Lambert (2008). Comparing the antimicrobial effectiveness of NaCl and KCl with a view to salt/sodium replacement, International journal of food microbiology, 124(1):98-102.
Bintsis, T. (2006). Quality of the brine, dans Brined Cheeses, A.Y. Tamime (ed.), Blackwell Publishing Ltd, pages 264-301.

106
Blomberg, A. et L. Adler (1992). Physiology of osmotolerance in fungi, Advances in microbial physiology, 33:145-212.
Bockelmann, W. (2010). Secondary cheese starter cultures, dans Technology of cheesemaking, Law B.A. et A.Y. Tamime (eds.), Wiley-Blackwell, pages 193-230.
Bona, E., R.L. Carneiro, D. Borsato, R.S.S.F. Silva, D.A.S. Fidelis et L.H.M.e. Silva (2007). Simulation of NaCl and KCl mass transfer during salting of Prato cheese in brine with agitation: a numerical solution, Brazilian journal of chemical engineering, 24:337-349.
Boutrou, R. et M. Gueguen (2005). Interests in Geotrichum candidum for cheese technology, International journal of food microbiology, 102(1):1-20.
Boutrou, R., L. Kerriou et J.Y. Gassi (2006). Contribution of Geotrichum candidum to the proteolysis of soft cheese, International dairy journal, 16(7):775-783.
Boziaris, I.S., P.N. Skandamis, M. Anastasiadi et G.J.E. Nychas (2007). Effect of NaCl and KCl on fate and growth/no growth interfaces of Listeria monocytogenes Scott A at different pH and nisin concentrations, Journal of applied microbiology, 102(3):796-805.
Brown, I.J., I. Tzoulaki, V. Candeias et P. Elliott (2009). Salt intakes around the world: implications for public health, International journal of epidemiology, 38(3):791-813.
Campbell, N., R. Correa-Rotter, B. Neal et F.P. Cappuccio (2011a). New evidence relating to the health impact of reducing salt intake, Nutrition, metabolism, and cardiovascular diseases, 21(9):617-619.
Campbell, N., F.P. Cappuccio et S.W. Tobe (2011b). Unnecessary controversy regarding dietary sodium: a lot about a little, Canadian journal of cardiology, 27(4):404-406.
Cantor, M.D., T. van den Tempel, T.K. Hansen et Y. Ardö (2004). Blue cheeses, dans Cheese: chemistry, physics and microbiology, Vol 2, Major cheese groups (3e edn), P.F. Fox, P.L.H. McSweeney, T.M Cogan et T.P. Guinee (eds.), Elsevier, London, pages 175-198.
Cappuccio, F.P., R. Kalaitzidis, S. Duneclift et J.B. Eastwood (2000). Unravelling the links between calcium excretion, salt intake, hypertension, kidney stones and bone metabolism, Journal of nephrology, 13(3):169-177.
CCIL (2015). Normes FIL / ISO. [En ligne]: http://dairyinfo.gc.ca/index_f.php?s1=fil-idf&s2=pub&s3=iso. Consulté le 16 mai 2015.
Cerning, J., J. Gripon, C., G. Lamberet et J. Lenoir (1987). Les activités biochimiques des Penicillium utilisés en fromagerie, Lait, 67(1):3-39.
Chamba, J.,F. (1988). Salage de l'Emmental. I. Influence des conditions de saumurage sur l'absorption du sel et sa cinétique, Lait, 68(2):121-142.

107
Champagne, C.P., L. Soulignac, M. Marcotte et J.P. Innocent (2003). Texture et évolution du pH de fromages de type Brie entreposés en atmosphère contrôlée, Lait, 83(2):145-151.
Choisy, C., M. Desmazeaud, M. Gueguen, J. Lenoir, J.L. Schmidt et C. Tourneur (2009). Les phénomènes microbiens, dans Le fromage : de la science à l'assurance qualité, A. Eck et J.C. Gillis (eds.), Tec & Doc - Lavoisier, Paris, pages 347-440.
Cruz, A.G., J.A.F. Faria, M.A.R. Pollonio, H.M.A. Bolini, R.M.S. Celeghini, D. Granato et N.P. Shah (2011). Cheeses with reduced sodium content: effects on functionality, public health benefits and sensory properties, Trends in food science and technology, 22(6):276-291.
CTAC (2009). Reformulation des produits pour réduire le sodium : Guide de réduction du sodium pour l'industrie alimentaire. [En ligne]: http://www.foodtechcanada.ca/siteimages/Guide%20r%C3%A9duction%20du%20sodium%20pour%20l'industrie%20alimentaire.pdf. Consulté le 14 février 2014.
Cuppers, H.G., S. Oomes et S. Brul (1997). A model for the combined effects of temperature and salt concentration on growth rate of food spoilage molds, Applied and environmental microbiology, 63(10):3764-3769.
De Piante, D., E. Castelao et A. Rubiolo de Reinick (1990). Diffusion coefficient of salt in salting and ripening of soft cheese, Engineering and food, 1(1):493-502.
Decker, M. et P.V. Nielsen (2005). The inhibitory effect of Penicillium camemberti and Geotruchum candidum on the associated funga of white mould cheese, International journal of food microbiology, 104(1):51-60.
Dieuleveux, V., D. Van Der Pyl, J. Chataud et M. Gueguen (1998). Purification and characterization of anti-Listeria compounds produced by Geotrichum candidum, Applied and environmental microbiology, 64(2):800-803.
Dötsch, M., J. Busch, M. Batenburg, G. Liem, E. Tareilus, R. Mueller et G. Meijer (2009). Strategies to reduce sodium consumption: a food industry perspective, Critical reviews in food science and nutrition, 49(10):841-851.
Doyle, M.E. et K.A. Glass (2010). Sodium reduction and its effect on food safety, food quality, and human health, Comprehensive reviews in food science and food safety, 9(1):44-56.
Elliott, P., R.D. Forrest, C.A. Jackson et J.S. Yudkin (1988). Sodium and blood pressure: positive associations in a north London population with consideration of the methodological problems of within-population surveys, Journal of human hypertension, 2(2):89-95.
Everett, D.W., T.P. Guinee et M.E. Johnson (2014). Cheese structure and functionality, dans The importance of salt in the manufacture and ripening of cheese, P. Paquin et S. Labrie (eds.), International Dairy Federation, pages 30-41.

108
Fisher, P.W.F., M. Vigneault, R. Huang, K. Arvaniti et P. Roach (2009). Sodium food sources in the Canadian diet, Applied physiology, nutrition, and metabolism, 34:884-892.
Fitzgerald, E. et J. Buckley (1985). Effect of total and partial substitution of sodium chloride on the quality of Cheddar cheese, Journal of dairy science, 68(12):3127-3134.
Fox, P.F. et P.L.H. McSweeney (2004). Cheese: an overview, dans Cheese: chemistry, physics and microbiology, Vol. 1, P.F. Fox, P.L.H. McSweeney, T.M. Cogan et T.P. Guinee (eds.), Academic Press, pages 1-18.
Gaucheron, F. (2004). Dosage des principaux minéraux du lait et des produits laitiers, dans Minéraux et produits laitiers, F. Gaucheron (ed.), Tec & Doc - Lavoisier, Paris, France, pages 565-583.
Gaüzère, Y. (2013). Technologie fromagère appliquée aux pâtes pressées cuites et non cuites. Centre d'expertise fromagère du Québec, Guide de formation, Saint-Hyacinthe, Canada.
Gente, S., S. La Carbona et M. Guéguen (2007). Levels of cystathionine-γ-lyase production by Geotrichum candidum in synthetic media and correlation with the presence of sulphur flavours in cheese, International journal of food microbiology, 114(2):136-142.
Geurts, T.J., P. Walstra et H. Mulder (1974). Transport of salt and water during salting of cheese. 1. Analysis of the process involved, Netherlands milk and dairy journal, 28:102-129.
Geurts, T.J., P. Walstra et H. Mulder (1980). Transport of salt and water during salting of cheese. 2. Quantities of salt taken up and and moisture lost, Netherlands milk and dairy journal, 34:229-254.
Gillis, J.C. (2004). Manuel du salage en fromagerie - Théorie & pratiques. Arilait recherche, Paris.
Gomes, A.P., A.G. Cruz, R.S. Cadena, R.M.S. Celeghini, J.A.F. Faria, H.M.A. Bolini, M.A.R. Pollonio et D. Granato (2011). Manufacture of low-sodium Minas fresh cheese: Effect of the partial replacement of sodium chloride with potassium chloride, Journal of dairy science, 94(6):2701-2706.
Grummer, J., M. Karalus, K. Zhang, Z. Vickers et T.C. Schoenfuss (2012). Manufacture of reduced-sodium Cheddar-style cheese with mineral salt replacers, Journal of dairy science, 95(6):2830-2839.
Guinee, T.P. et P.F. Fox (1986a). Influence of cheese geometry on the movement of sodium chloride and water during brining, Irish journal of agricultural and food research, 10(2):73-96.
Guinee, T.P. et P.F. Fox (1986b). Transport of sodium chloride and water in Romano cheese slices during brining, Food Chemistry, 19(1):49-64.
Guinee, T.P. et P.F. Fox (2004). Salt in cheese: physical, chemical and biological aspects, dans Cheese: chemistry, physics and microbiology, Vol. 1, P.F. Fox, P.L.H. McSweeney, T.M. Cogan et T.P. Guinee (eds.), Academic Press, pages 207-259.

109
Guinee, T.P., M.E. Johnson et S. Skeie (2014a). Characteristic levels of salt in different cheese varieties, dans The importance of salt in the manufacture and ripening of cheese, P. Paquin et S. Labrie (eds.), International Dairy Federation, pages 46-60.
Guinee, T.P., M.E. Johnson et S. Skeie (2014b). Good manufacturing practices for achieving target salt levels in cheese, dans The importance of salt in the manufacture and ripening of cheese, P. Paquin et S. Labrie (eds.), International Dairy Federation, pages 61-73.
Guinee, T.P. et B.T. O'Kennedy (2007). Reducing salt in cheese and dairy spread, dans Reducing salt in food: pratical strategies, D. Kilcast et F. Angus (eds.), Woodhead Publishing Limited, Cambridge, pages 316-357.
Guizani, N., S. Kasapis, Z.H. Al-Attabi et M.H. Al-Ruzeiki (2002). Microbiological, physicochemical, and biochemical changes during ripening of Camembert cheese made of pasteurized cow's milk, International journal of food properties, 5(3):483-494.
Hardy, J. (1976). Étude de la diffusion du sel dans les fromages à pâte molle de type Camembert. Comparaison du salage à sec et du salage en saumure, Thèse de doctorat, Université de Nancy I, Nancy, France.
Hardy, J. (1979). L'activité de l'eau, le sel et les moisissures des fromages, Revue de laiterie française, 377:19-25.
Hardy, J. (1985). Le chlorure de sodium: propriétés et utilisation en fromagerie, dans Laits et produits laitiers: vache - brebis – chèvre, F.M. Luquet (ed.), Tec & Doc – Lavoisier, Paris, pages 603-615.
Hardy, J. (2004). Le chlorure de sodium dans le lait et les produits fromagers, dans Minéraux et produits laitiers, F. Gaucheron (ed.), Tec & Doc – Lavoisier Paris, France, pages 619-643.
Hardy, J. (2009). L'activité de l'eau et l'affinage des fromages, dans Le fromage: de la science à l'assurance qualité, A. Eck et J.C. Gillis (eds.), Tec & Doc – Lavoisier Paris, France, pages 62-84.
He, F.J., J. Li et G.A. MacGregor (2013). Effect of longer-term modest salt reduction on blood pressure, The Cochrane database of systematic reviews, 4:CD004937.
He, F.J. et G.A. MacGregor (2006). Importance of salt in determining blood pressure in children: meta-analysis of controlled trials, Hypertension, 48(5):861-869.
He, F.J. et G.A. MacGregor (2008). Beneficial effects of potassium on human health, Physiologia plantarum, 133(4):725-735.
He, F.J. et G.A. MacGregor (2011). Salt reduction lowers cardiovascular risk: meta-analysis of outcome trials, Lancet, 378(9789):380-382.

110
He, F.J., N.M. Marrero et G.A. MacGregor (2008). Salt intake is related to soft drink consumption in children and adolescents: a link to obesity?, Hypertension, 51(3):629-634.
Hoffmann, W. (2014). Partial substitution of sodium chloride by potassium chloride in natural cheeses, dans The importance of salt in the manufacture and ripening of cheese, P. Paquin et S. Labrie (eds.), International Dairy Journal, pages 74-83.
Hooley, P., D.A. Fincham, M.P. Whitehead et N.J. Clipson (2003). Fungal osmotolerance, Advances in applied microbiology, 53:177-211.
INAO (2013). Cahier des charges de l'appellation d'origine «Camembert de Normandie» [En ligne]: http://agriculture.gouv.fr/IMG/pdf/CDCCamembertdeNormandie_BO_cle46ec84.pdf. Consulté le 16 mai 2015.
IOM (2005). Dietary reference intakes for water, potassium, sodium, chloride and sulfate [En ligne]: http://www.nap.edu/openbook.php?record_id=10925. Consulté le 16 mai 2015.
Irvine, D.M. et W.V. Price (1961). Influence of salt on the development of acid by lactic starters in skimmilk and in curd submerged in brine, Journal of dairy science, 44(2):243-248.
Jany, J.L. et G. Barbier (2008). Culture-independent methods for identifying microbial communities in cheese, Food microbiology, 25(7):839-848.
Joffres, M.R., N.R. Campbell, B. Manns et K. Tu (2007). Estimate of the benefits of a population-based reduction in dietary sodium additives on hypertension and its related health care costs in Canada, Canadian journal of cardiology, 23(6):437-443.
Johnson, M.E., R. Kapoor, D.J. McMahon, D.R. McCoy et R.G. Narasimmon (2009). Reduction of sodium and fat levels in natural and processed cheeses: scientific and technological aspects, Comprehensive reviews in food science and food safety, 8(3):252-268.
Jordan, K.N. et T.M. Cogan (1993). Identification and growth of non-starter lactic acid bacteria in Irish Cheddar cheese, Irish journal of agricultural and food research:47-55.
Karahadian, C., D.B. Josephson et R.C. Lindsay (1985). Volatile compounds from Penicillium sp. contributing musty-earthy notes to Brie and Camembert cheese flavors, Journal of agricultural and food chemistry, 33(3):339-343.
Katsiari, M.C., L.P. Voutsinas, E. Alichanidis et I.G. Roussis (1997). Reduction of sodium content in Feta cheese by partial substitution of NaCl by KCl, International dairy journal, 7(6–7):465-472.
Katsiari, M.C., L.P. Voutsinas, E. Alichanidis et I.G. Roussis (1998). Manufacture of Kefalograviera cheese with less sodium by partial replacement of NaCl with KCl, Food chemistry, 61(1–2):63-70.

111
Labrie, S., W. Bisig et K. Jordan (2014). Importance of NaCl in cheese manufacture and ripening, dans The importance of salt in the manufacture and ripening of cheese, P. Paquin et S. Labrie (eds.), International Dairy Federation, pages 8-20.
Lacrampe, J.L., J. Hardy, J. Ramet, P. et F. Weber (1971). Contribution à l'étude de l'évolution chimique et du traitement des saumures de fromagerie, Lait, 51(503-504):158-175.
Larsson, C., C. Morales, L. Gustafsson et L. Adler (1990). Osmoregulation of the salt-tolerant yeast Debaryomyces hansenii grown in a chemostat at different salinities, Journal of bacteriology, 172(4):1769-1774.
Lawrence, R.C., L.K. Creamer et J. Gilles (1987). Texture development during cheese ripening, Journal of dairy science, 70(8):1748-1760.
Le Drean, G., J. Mounier, V. Vasseur, D. Arzur, O. Habrylo et G. Barbier (2010). Quantification of Penicillium camemberti and P. roqueforti mycelium by real-time PCR to assess their growth dynamics during ripening cheese, International journal of food microbiology, 138(1-2):100-107.
Le Graet, Y. et G. Brulé (1988). Migration des macro et oligo-éléments dans un fromage à pâte molle de type Camembert, Lait, 68(2):219-234.
Le Graët, Y., A. Lepienne, G. Brulé et P. Ducruet (1983). Migration du calcium et des phosphates inorganiques dans les fromages à pâte molle de type Camembert au cours de l'affinage, Lait, 63(629-630):317-332.
Leclercq-Perlat, M.N. (2011). Cheese | Camembert, brie, and related varieties, dans Encyclopedia of dairy sciences (Second edition), J.W. Fuquay (ed.), Academic Press, San Diego, pages 773-782.
Leclercq-Perlat, M.N., F. Buono, D. Lambert, E. Latrille, H.E. Spinnler et G. Corrieu (2004). Controlled production of Camembert-type cheeses. Part I: Microbiological and physicochemical evolutions, The Journal of dairy research, 71(3):346-354.
Leclercq-Perlat, M.N., G. Corrieu et H.E. Spinnler (2007). Controlled production of Camembert-type cheeses. Part III: role of the ripening microflora on free fatty acid concentrations, The Journal of dairy research, 74(2):218-225.
Leclercq-Perlat, M.N., D. Picque, S.T. Martin Del Campo Barba et C. Monnet (2013). Dynamics of Penicillium camemberti growth quantified by real-time PCR on Camembert-type cheeses under different conditions of temperature and relative humidity, Journal of dairy science, 96(6):4031-4040.
Leclercq-Perlat, M.N., D. Picque, H. Riahi et G. Corrieu (2006). Microbiological and biochemical aspects of Camembert-type cheeses depend on atmospheric composition in the ripening chamber, Journal of dairy science, 89(8):3260-3273.
Lenoir, J., C. Choisy, B. Auberger, J. Marrasse et M. Schmidt (1971). Aptitude de l'espèce Penicillium caseicolum à la production d'enzymes protéolytiques, Lait, 51(503-504):138-157.

112
Lessard, M.H., G. Belanger, D. St-Gelais et S. Labrie (2012). The composition of Camembert cheese-ripening cultures modulates both mycelial growth and appearance, Applied and environmental microbiology, 78(6):1813-1819.
Lessard, M.H., C. Viel, B. Boyle, D. St-Gelais et S. Labrie (2014). Metatranscriptome analysis of fungal strains Penicillium camemberti and Geotrichum candidum reveal cheese matrix breakdown and potential development of sensory properties of ripened Camembert-type cheese, BMC genomics, 15:235.
Lindsay, R.C., S.M. Hargett et C.S. Bush (1982). Effect of sodium/potassium (1:1) chloride and low sodium chloride concentrations on quality of Cheddar cheese, Journal of dairy science, 65(3):360-370.
López-Dίaz, T.M., J. Santos, A. Otero, M.L. Garcίa et B. Moreno (1996). Some technological properties of Penicillium roqueforti strains isolated from a home-made blue cheese, Letters in applied microbiology, 23:5-8.
Marcellino, N., E. Beuvier, R. Grappin, M. Gueguen et D.R. Benson (2001). Diversity of Geotrichum candidum strains isolated from traditional cheesemaking fabrications in France, Applied and environmental microbiology, 67(10):4752-4759.
Marilley, L. et M.G. Casey (2004). Flavours of cheese products: metabolic pathways, analytical tools and identification of producing strains, International journal of food microbiology, 90(2):139-159.
McSweeney, P.L.H. (2004). Biochemistry of cheese ripening, International journal of dairy technology, 57(2-3):127-144.
Melilli, C., D.M. Barbano, M. Caccamo, L. Tuminello, S. Carpino et G. Licitra (2006). Interaction of brine concentration, brine temperature, and presalting on salt penetration in Ragusano cheese, Journal of dairy science, 89(5):1420-1438.
Melilli, C., D.M. Barbano, G. Licitra, G. Portelli, G. Di Rosa et S. Carpino (2003a). Influence of the temperature of salt brine on salt uptake by Ragusano cheese, Journal of dairy science, 86(9):2799-2812.
Melilli, C., D.M. Barbano, G. Licitra, G. Tumino, G. Farina et S. Carpino (2003b). Influence of presalting and brine concentration on salt uptake by Ragusano cheese, Journal of dairy science, 86(4):1083-1100.
Melilli, C., D. Carco, D.M. Barbano, G. Tumino, S. Carpino et G. Licitra (2005). Composition, microstructure, and surface barrier layer development during brine salting, Journal of dairy science, 88(7):2329-2340.
Mermet, J.M. (2004). Intérêt et principes de la spectroscopie atomique dans le dosage des minéraux, dans Minéraux et produits laitiers, F. Gaucheron (ed.), Tec & Doc – Lavoisier Paris, France, pages 815-846.
Mietton, B., F. Gaucheron et F. Salün-Michel (2004). Minéraux et transformation fromagère, dans Minéraux et produits laitiers, F. Gaucheron (ed.), Tec & Doc – Lavoisier Paris, France, pages 471-563.

113
Molimard, P., Lesschaeve, S. Issanchou, M. Brousse et H.E. Spinnler (1997). Effect of the association of surface flora on the sensory properties of mould-ripened cheese, Lait, 77(1):181-187.
Molimard, P., I. Lesschaeve, I. Bouvier, L. Vassal, P. Schlich, S. Issanchou et H.E. Spinnler (1994). Bitterness and nitrogen fractions of mold ripened cheese of Camembert-type: impact of the association of Penicillium camemberti with Geotrichum candidum, Lait, 74(5):361-374.
Molimard, P. et H.E. Spinnler (1996). Review: compounds involved in the flavor of surface mold-ripened cheeses: origins and properties, Journal of dairy science, 79(2):169-184.
Møller, K., F. Rattray, E. Høier et Y. Ardö (2012). Manufacture and biochemical characteristics during ripening of Cheddar cheese with variable NaCl and equal moisture content, Dairy science and technology, 92(5):515-540.
Nielsen, M.S., J.C. Frisvad et P.V. Nielsen (1998). Protection by fungal starters against growth and secondary metabolite production of fungal spoilers of cheese, International journal of food microbiology, 42(1–2):91-99.
O'Nulain, M. (1986). Manufacture of modified Camembert and Cheddar cheeses, Mémoire de maîtrise, Department of dairy and food chemistry, National University of Ireland, Cork.
OMS (2007). Réduire les apports en sel au niveau des populations - Rapport du forum et de la réunion technique OMS. Service de production des documents de l'OMS, Genève, Suisse.
OMS (2013a). Directives: Sur l'apport en sodium chez l'adulte et chez l'enfant - Résumé d'orientation. Service de production de documents de l'OMS, Genève, Suisse.
OMS (2013b). Mapping salt reduction initiatives in the WHO european region. WHO Regional Office for Europe, Copenhague, Danemark.
Paquin, P. (2014). Introduction: Dairy products and salt, dans The importance of salt in the manufacture and ripening of cheese, P. Paquin et S. Labrie (eds.), International Dairy Federation, pages 3-7.
Perkin-Elmer (2004). A guide to inorganic analysis from the leaders in AA, ICP-OES, and ICP-MS. [En ligne]: http://www.d.umn.edu/~jevans1/chem%204242%20s15/Problem%20Sets/for%20ps%203/GDE_InorganicAnalysis%20from%20pe.pdf. Consulté le 16 mai 2015.
Pitt, J.I., R.H. Cruickshank et L. Leistner (1986). Penicillium commune, P. camembertii, the origin of white cheese moulds, and the production of cyclopiazonic acid, Food microbiology, 3(4):363-371.
Poças, M.F. et M. Pintado (2009). Packaging and the shelf life of cheese, dans Food packaging and shelf life: a practicle guide, G.L. Robertson (ed.), Taylor & Francis Group, Boca Raton, États-Unis, pages 103 - 122.

114
Poulter, N.R., K.T. Khaw, B.E. Hopwood, M. Mugambi, W.S. Peart, G. Rose et P.S. Sever (1990). The Kenyan Luo migration study: observations on the initiation of a rise in blood pressure, British medical journal, 300(6730):967-972.
Reddy, K.A. et E.H. Marth (1991). Reducing the sodium content of foods: a review, Journal of food protection, 54(2):132-150.
Reddy, K.A. et E.H. Marth (1995). Microflora of Cheddar cheese made with sodium chloride, potassium chloride, or mixtures of sodium and potassium chloride, Journal of food protection, 58(1):54-61.
Richard, J. et H. Zadi (1983). Inventaire de la flore bactérienne dominante des Camemberts fabriqués avec du lait cru, Lait, 63(623-624):25-42.
Sacks, F.M., L.P. Svetkey, W.M. Vollmer, L.J. Appel, G.A. Bray, D. Harsha, E. Obarzanek, P.R. Conlin, E.R. Miller III, D.G. Simons-Morton, N. Karanja et P.H. Lin (2001). Effects on blood pressure of reduced dietary sodium and the dietary approaches to stop hypertension (DASH) diet. DASH-Sodium collaborative research group, The New England journal of medicine, 344(1):3-10.
Samapundo, S., N. Deschuyffeleer, D. Van Laere, I. De Leyn et F. Devlieghere (2010). Effect of NaCl reduction and replacement on the growth of fungi important to the spoilage of bread, Food microbiology, 27(6):749-756.
Sansonetti, F. (1930). Sur les flores bactérienne et fongique des caillés de lait. Rôle des symbioses dans la maturation du Camembert (Suite), Lait, 10(98):858-874.
Santé Canada (2006). Liste des additifs alimentaires autorisés. [En ligne]: http://www.hc-sc.gc.ca/fn-an/securit/addit/diction/dict_food-alim_add-fra.php. Consulté le 18 mai 2015.
Santé Canada (2010). Stratégie de réduction du sodium pour le Canada - Recommandations du Groupe de travail sur le sodium. [En ligne]: http://www.hc-sc.gc.ca/fn-an/nutrition/sodium/related-info-connexe/strateg/ reduct-strat-fra.php. Consulté le 15 janvier 2014.
Santé Canada (2012). Document d'orientation destiné à l'insdustrie alimentaire sur la réduction du sodium dans les aliments transformés. [En ligne]: http://www.hc-sc.gc.ca/fn-an/alt_formats/pdf/legislation/guide-ld/2012-sodium-reduction-indust-fra.pdf. Consulté le 16 mai 2015.
Schlesser, J.E., S.J. Schmidt et R. Speckman (1992). Characterization of chemical and physical changes in Camembert cheese during ripening, Journal of dairy science, 75(7):1753-1760.
Schmidt, J.,L., J. Lenoir et M. Schmidt (1980). Contribution à l'étude de la flore levure du fromage de Camembert (II), Lait, 60(595-596):272-282.
Schmidt, J.,L., J. Lenoir, M. Schmidt, T. Luu et C. Graffard (1978). Contribution à l'étude de la flore levure du fromage de Camembert. Son évolution au cours de la maturation, Lait, 58(577):355-370.

115
Schroeder, C.L., F.W. Bodyfelt, C.J. Wyatt et M.R. McDaniel (1988). Reduction of sodium chloride in Cheddar cheese: effect on sensory, microbiological, and chemical properties, Journal of dairy science, 71(8):2010-2020.
Schwartzberg, H.G. et R.Y. Chao (1982). Solute diffusivities in leaching process, Food technology, 36(2):73-86.
Shaw, M.B. (1981). The manufacture of soft, surface mould, ripened cheese in France with particular reference to Camembert, International journal of dairy technology, 34(4):131-138.
Shrestha, S., J.A. Grieder, D.J. McMahon et B.A. Nummer (2011). Survival of Listeria monocytogenes introduced as a post-aging contaminant during storage of low-salt Cheddar cheese at 4, 10, and 21 degrees C, Journal of dairy science, 94(9):4329-4335.
Sousa, M.J. (2003). Cheeses | Surface mold-ripened cheese varieties, dans Encyclopedia of food sciences and nutrition (Second edition), B. Caballero (ed.), Academic Press, Oxford, pages 1122-1129.
Spinnler, H.E. et J.C. Gripon (2004). Surface mould-ripened cheeses, dans Cheese: chemistry, physics and microbiology, Vol, 2, P.F. Fox, P.L.H. McSweeney, T.M. Cogan et T.P. Guinee (eds.), Academic Press, pages 157-174.
St-Gelais, D. et P. Tirard-Collet (2002). Fromage, dans Science et technologie du lait - Transformation du lait, Vignola, C.L. (ed.), Presses internationales Polytechnique, Montréal, Canada, pages 349-416.
Statistique Canada (2014). Les dix principales causes de décès, 2011. [En ligne]: http://www.statcan.gc.ca/ pub/ 82-625-x/2014001/article/11896-fra.htm. Consulté le 12 mai 2015.
Stolarz-Skrzypek, K., T. Kuznetsova, L. Thijs, V. Tikhonoff, J. Seidlerova, T. Richart, Y. Jin, A. Olszanecka, S. Malyutina, E. Casiglia, J. Filipovsky, K. Kawecka-Jaszcz, Y. Nikitin, J.A. Staessen et Investigators of European Project on Genes in Hypertension (2011). Fatal and nonfatal outcomes, incidence of hypertension, and blood pressure changes in relation to urinary sodium excretion, Journal of the American medical association, 305(17):1777-1785.
Strazzullo, P., L. D'Elia, N.B. Kandala et F.P. Cappuccio (2009). Salt intake, stroke, and cardiovascular disease: meta-analysis of prospective studies, British medical journal, 339:b4567.
Sutherland, B.J. (2002). Cheeses | Salting of cheese, dans Encyclopedia of dairy sciences, H. Roginski (ed.), Elsevier, Oxford, pages 293-300.
Taylor, R.S., K.E. Ashton, T. Moxham, L. Hooper et S. Ebrahim (2011). Reduced dietary salt for the prevention of cardiovascular disease: a meta-analysis of randomized controlled trials (Cochrane review), American journal of hypertension, 24(8):843-853.
Terré, E., Y. Le Graët, G. Brulé et J.L. Maubois (1985). Étude de transfert des solutés des fromages à pâte molle dans les saumures. Intérêt du traitement par ultrafiltration sur membrane., Technique laitière, 997:39-47.

116
Thibaudeau, E., D. Roy et D. St-Gelais (2015). Production of brine-salted Mozzarella cheese with different ratios of NaCl/KCl, International dairy journal, 40(0):54-61.
Tornadijo, M.E., J.M. Fresno, R. Martin Sarmiento et J. Carballo (1998). Study of the yeasts during the ripening process of Armada cheeses from raw goat's milk, Lait, 78(6):647-659.
van den Tempel, T. et M.S. Nielsen (2000). Effects of atmospheric conditions, NaCl and pH on growth and interactions between moulds and yeasts related to blue cheese production, International journal of food microbiology, 57(3):193-199.
Vassal, L. et J.C. Gripon (1984). L'amertume des fromages à pâte molle de type Camembert: rôle de la présure et de Penicillium caseicolum, moyens de la contrôler, Lait, 64(643-644):397-417.
Weaver, C.M. (2013). Potassium and health, Advances in nutrition, 4(3):368S-377S.
Webster, J.L., E.K. Dunford, C. Hawkes et B.C. Neal (2011). Salt reduction initiatives around the world, Journal of hypertension, 29(6):1043-1050.
Whelton, P.K. et J. He (2014). Health effects of sodium and potassium in humans, Current opinion in lipidology, 25(00):1-5.
Wolffs, P., B. Norling et P. Rådström (2005). Risk assessment of false-positive quantitative real-time PCR results in food, due to detection of DNA originating from dead cells, Journal of microbiological methods, 60(3):315-323.
Zorrilla, S.E. et A.C. Rubiolo (1994a). Fynbo cheese NaCl and KCl changes during ripening, Journal of food science, 59(5):972-975.
Zorrilla, S.E. et A.C. Rubiolo (1994b). Modeling NaCl and KCl movement in Fynbo cheese during salting, Journal of food science, 59(5):976-980.


















