
Science et nature Les probiotiques issus du lait maternel ...

الجمهورية الجزائرية الديمقراطية الشعبية
وزارة التعليم العالي و البحث العلمي
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTAIRE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE
UNIVERSITE D’ORAN
FACULTE DES SCIENCES
DEPARTEMENT DE BIOLOGIE
LABORATOIRE DE GENETIQUE MICROBIENNE
Mémoire de fin d’étude
En vue de l’obtention du Diplôme de Magister
En Biologie
OPTION : Biologie moléculaire et génétique des microorganismes
Thème
Mer AOUES Abdelkader Président Professeur Université d’ORAN
Mme Mesli-Taleb Farida Examinateur Professeur Université d’ORAN
Mer BEKKADA Ahmed Examinateur MCA Centre universitaire Ghelizane
er
2012-2013
Contribution à l’étude des probiotiques et prébiotiques comme alternatives aux antibiotiques
en aviculture
Présenté par : LAROUCI Saliha
M Bensalah Farid Encadreur Professeur Université d’ORAN Soutenue le 20 octobre 2013

Remerciement
Louanges à dieu de nous avoir illuminé le chemin du savoir et nous avoir aidé.
Au terme de ce travail, je tiens à exprimer mes remerciements les plus sincères aux personnes
qui ont contribué de loin ou de prés à l’élaboration de ce mémoire.
J’adresse tout d’abord mes sincères remerciement à Pr BENSALAH Farid, directeur du
Laboratoire de Génétique Microbienne, pour m’avoir encadré et permis de réalisé ce travail
au sein de son laboratoire, je tiens a lui exprimer ma reconnaissance pour la qualité de son
savoir faire, de ses compétences, ses conseils et orientations scientifiques tout le long de ce
travail, je lui exprime ma profonde gratitude pour tout les efforts et le temps qu’il m’a
consacré pour réaliser ce travail.
J’adresse mes vifs remerciements à Pr TALEB- MESLI Farida pour sa gentillesse d’avoir
accepter d’examiner mon travail.
Je remercie aussi les membres de jury :
Pr AOUES Abdelkader de m’avoir honoré en acceptant de présider le jury.
Dr BEKKADA Ahmed d’avoir accepté d’examiner mon travail.
Je témoigne toute ma reconnaissance à mes chers parents, pour leur amour et leur soutien
durant toutes mes années d’études.
Enfin, je remercie tous mes amis, mes proches et toutes les personnes qui me sont chères à
moi pour leur soutien, leur patience, leur compréhension…

Liste des abréviations
Liste des tableaux
Liste des figures
Résumé
Abstract
ملخص
Introduction générale…………………………………………………………………………….
Synthèse bibliographique
Chapitre 1 : Généralités
1. Le microbiote intestinal chez la volaille………………………………………………………
3. Description de la flore digestive du poulet et localisation dans le tractus………………….
2. Description du tube digestif chez la volaille……………………………………………... …..
4. Les pathogènes entériques de la volaille digestif………………………………………..........
4.1. Maladies entériques chez la volaille ……………………………………………………
4.1.1. Escherichia coli ……………………………………………………………………
4.1.2. Salmonella…………………………………………………………………………
5. Contrôle des infections entériques chez la volaille……………………...................................
Chapitre 2 : les probiotiques en aviculture
1. histoire d’utilisation des probiotiques en alimentation animale…………………………….
2. Définition des probiotiques…………………………………………………………………….
3. Les micro-organismes probiotiques autorisés aujourd’hui en alimentation avicole……...
4. Critères de sélection des probiotiques………………………………………………………..
5. Les différents microorganismes probiotiques en aviculture et leurs caractéristiques…….
5.1. Le genre Lactobacillus ………………………………………………………………..
5.2. Le genre Streptococcus………………………………………………………...............
5.3. Le genre Enterococcus………………………………………………………................
5.4. Les genres Leuconostoc, Oenococcus et Weissella …………………………………...
5.5. Les genres Pediococcus ………………………………………………………………..
5.6. Le genre Bifidobacterium……………………………………………………………...
5.7. Les levures……………………………………………………………………………
6. Production de substances antimicrobiennes…………………………………………………
7. Efficacités des probiotique en aviculture……………………………………………………..
7.1 Efficacité sanitaire des probiotiques …………………………………………………..
7.2 Efficacité zootechnique des probiotiques ……………………………………………..
1
3
3
5
7
7
7
8
9
10
11
12
14
18
19
19
20
20
21
21
21
24
26
26
27

Chapitre 3 : les prébiotiques
1. Définition Prébiotiques ………………………………………………………………………..
2. Différentes classes des prébiotiques …………………………………………………………
2.1. Les hexoses……………………………………………………………………………….
2.2. Les disaccharides naturels ……………………………………………………………….
2.3. Les oligosaccharides ……………………………………………………………………..
3. Mode d’action des prébiotiques ………………………………………………………………
4. Caractérisation des exopolysaccharides (prébiotique)…………………………………….
4.1 Classification des EPS des bactéries lactiques…………………………………………..
4.2 Production et détection des EPS …………………………………………………….
4.2.1 Détection de polysaccharides exo-cellulaires……………………………………
4.2.2 Examen visuel …………………………………………………………………….
4.2.3 Les colorations…………………………………………………………………….
4.3 Extraction et purification ……………………………………………………………...
4.4 Dosage colorimétrique des EPS purifiés………………………………………………
4.5 Caractérisation des EPS à l’échelle chromatographique…………………………….
5. Effet des prébiotiques en aviculture ………………………………………………………….
Chapitre 4 : Approche moléculaire
1. l’ADNr 16S comme sémantide bactérienne ………………………………………………….
2. Les méthodes d’identification…………………………………………………………………
2.1. Identification des bactéries par l’ADNr-16S………………………………………………
2.2. Identification bactérienne avec amorces oligonucléotidiques (Amplification génique)…
3. Amplification par PCR………………………………………………………………………..
4. Séquençage de l’ADN………………………………………………………………………….
5. Taxonomie des bactéries lactiques ……………………………………………………………
Matériels et méthodes
1.Échantillonnage ………………………………………………………………………………..
2. Isolement et purification des souches…………………………………………………………
3. Conservation des souches……………………………………………………………………..
4. Souche de référence utilisé ……………………………………………………………………
5. Caractérisation phénotypique………………………………………………………………...
5.1. Examen macroscopique………………………………………………………………….
5.2. Examen microscopique…………………………………………………………………..
5.3. Test de la catalase ………………………………………………………………………..
6. recherche des bactéries mucoides……………………………………………………………..
29
29
29
29
29
30
31
31
32
32
32
32
33
33
34
35
37
38
38
39
39
41
42
44
44
44
45
45
45
45
45
45

6.1. Production et caractérisation des exopolysaccharides (EPS)…………………………
6.1.1. formation des exopolysaccharides ………………………………………………
6.1.2. Mise en évidence des EPS Mise en évidence des EPS en milieu solide au rouge
de ruthénium……………………………………………………………………………….
6.1. 3 Mise en évidence des EPS par microscopie………………….………………….
6.1.4. Extraction des EPS...………….…………………………………………………..
6.1.5 Lyophilisation des EPS……………………...……………………………………..
6.1.6 Dosage colorimétrique des EPS ………………………………………………......
6.1 .7. Activité rhéologique………………………………………………………………
6.2. Analyse de la composition des EPS…………………………………………………... ..
6.2.1 L’hydrolyse des EPS………………………………………………………………
6.2.2 Chromatographie sur couche mince des oses constitutifs des polysaccharides ..
7. Etude de l’activité antimicrobienne des souches probiotiques isolées du tractus digestif vis-à-
vis de souches pathogènes………………………………………………………………….
7.1. Nature de la substance inhibitrice ...................................................................................
8. Caractérisation moléculaire des probiotiques ……………………………………………….
8.1. Extraction d’ADN………………………………………………………………………..
8.1.1. Extraction d’ADN par utilisation de MOBIO KIT……………………………...
8.1.2. Extraction d’ADNr 16S dans sa totalité (1500pb)……………………………….
8.1.3. Vérification qualitatif de l’ADN extrait………………………………………….
8.2. Amplification……………………………………………………………………………..
8.2.1. Amplification de la région interne de l’ADNr 16S (480 pb) du genre
Lactobacillus sp……………………………………………………………………………….......
8.2.2. Séquençage et taxonomie des souches lactobacilles de la flore intestinale ……
8.2.3. Amplification du gène ADNr 16S (1500 pb) dans sa totalité…………………..
8.3. Electrophorèse sur gel d’agarose………………………………………………………..
Résultats et discussion
1. Caractérisation phénotypique ………………………………………………………………..
2. Mise en évidence et caractérisation des EPS…………………………………………………
2.1. Production des exopolysaccharides sur milieux solide…………………………………
2.2. Production des EPS en milieux liquide…………………………………………………
2.3. Mise en évidence des colonies mucoïdes blanchâtres en présence de colorant rouge de
ruthénium sur milieu solide ………………………………………………………………….
2.4. Mise en évidence des EPS par microscopie ……………………………………………..
2.5 Extraction des EPS ……………………………………………………………………….
2.6. Lyophilisation des EPS …………………………………………………………………..
45
45
46
46
46
47
47
48
48
48
49
50
51
51
52
52
53
54
54
54
55
55
56
59
59
59
60
72
72
73
73

2.7. Propriétés rhéologiques des EPS………………………………………………………..
2.8. Dosage colorimétrique …………………………………………………………………..
2.9. Chromatographie sur couche mince des oses constitutifs des différents
polysaccharides……………………………………………………………………………………
3. Etude de l’activité antimicrobienne des souches probiotique de la collection LGM………
4. Caractérisation moléculaire des souches probiotiques ……………………………………...
4.1. Identification du genre Lactobacillus spp. par amplification d’une région interne du gène
ADNr 16S ……………………………………………………………………………….
4.2 Séquençage et taxonomie des souches de la flore intestinale…………………………..
4.3. Amplification du gène de l’ADNr 16S à base de PCR de quelques isolats…………...
Conclusion et perspective………………………………………………………………………...
Références bibliographiques
Annexes
74
75
79
83
91
91
91
92
94

Liste des Abréviation
Noms de genres bactériens
E.: Escherichia
Lb.: Lactobacillus
Lc. : Lactococcus
S. : Staphylococcus
St. : Streptococcus
sp. : Espèce non précisée
ssp. : Sous espèce
Unités de mesures
C° : Degré Celsius
cm, mm, nm : Centimètre, millimètre, nanomètre
g, mg : Gramme, milligramme
h, min, s : heure, minute, seconde
l, ml, μl : Litre, millilitre, microlitre
M, mM : Molaire, millimolaire
N : Normalité
rpm : Rotation par minute
U : Unité
UI : Unité Internationale
V/V : Volume par volume
Autres abréviations
ADN : Acide Désoxyribonucléique
AFCA-CIAL : Association des Fabricants de Compléments pour l'Alimentation animale
ATCC: American Type Culture Collection
BET: Bromure d’Ethidium
dNTP :desoxynucléotides
DO : Densité Optique
EDTA : Acide Ethylene Diamine Tetra acétique
EPS: Exopolysaccharides
FAO: Food and Agriculture Organization

FEMS: Federation of European Microbiological Societies
G+C: Guanine + Cytosine
GRAS: Generally Regarded As Safe
INRA: Institut National de la Recherche Agronomique
LAB : bactéries lactiques
LGM: Laboratoire De Génétique Microbienne
M : marqueur de taille
MRS : de Man-Rogosa et Sharp
OMS : Organisation Mondiale de la Santé
Pb : paire de base
PCR : polymerase chain réaction
pH : Potentiel d’Hydrogène
qsp : Quantité suffisante pour
RFLP : Restriction Fragment Length Polymorphism
TBE : tris-borate-EDTA
TES : Tris-EDTA-saccharose
Tris : trishydroxyméthylaminométhane
UFC : Unité Formant Colonie
UV: Ultra Violet
WHO: World Health Organization
CCM: chromatographie sur couche mince
TLC: thin layer chromatography

Liste des tableaux
Tableau 1: Nombre de bactéries viables (log10 / g de contenu) des groupes majoritaires dan
le tube digestif du poulet (d’après Smith, 1965)
Tableau 2 : Micro-organismes probiotiques autorisés en Europe pour la volaille (liste publiée
par l’AFCA-CIAL, dernière mise à jour Mars 2009).
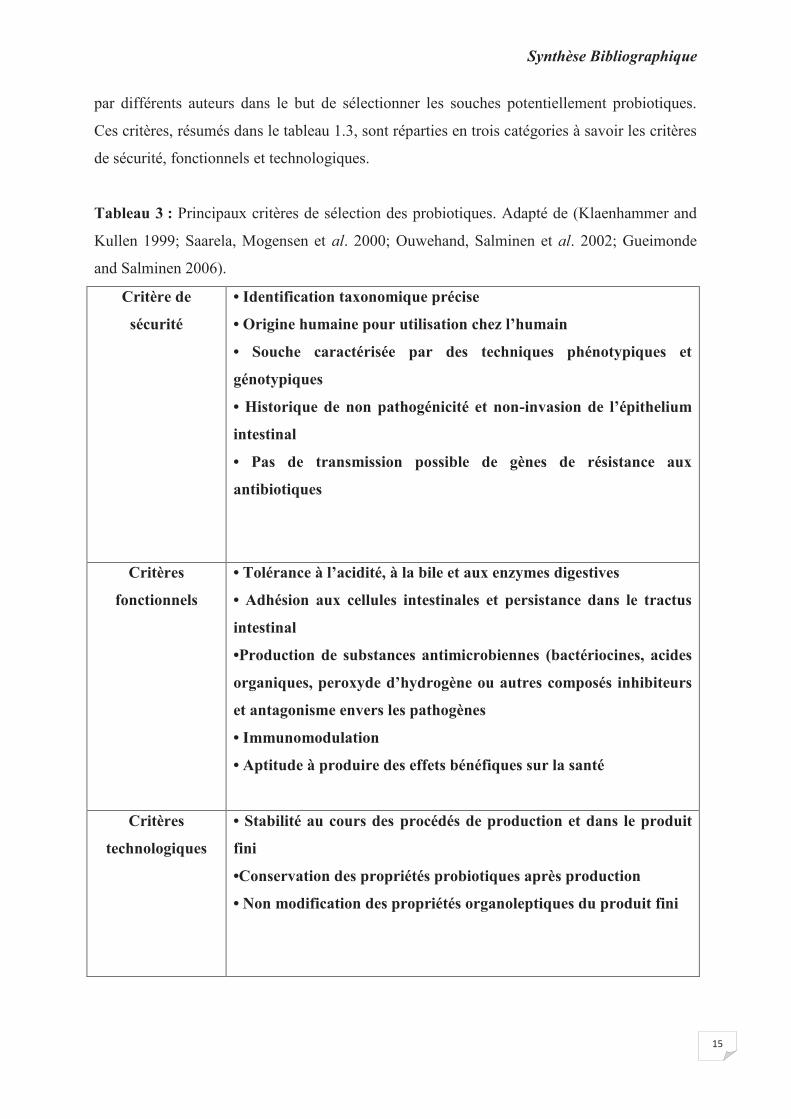
Tableau 3 : Principaux critères de sélection des probiotiques. Adapté de (Klaenhammer and
Kullen 1999; Saarela, Mogensen et al. 2000; Ouwehand, Salminen et al. 2002; Gueimonde
and Salminen 2006).
Tableau 4 : Exemples d’effets probiotiques récemment démontrés en élevage avicole (adapté
de Bernardeau et al., 2006).
Tableau 5 : valeurs obtenues permettant de tracer la courbe d’étalonnage de glucose (0-120
mg eq glucose/l).
Tableau 6: Résultats des différentes concentrations des EPS exprimés en mg eq. glucose/l
Tableau 7 : les différent RF obtenus sur le système 1 (acide acétique /chloroforme/ eau).
Tableau 8 : activité antibactérienne des différents isolats LGM contre des souches
pathogènes.
Tableau 9 : effet inhibiteur des surnageants natifs

Liste des figures
Figure 1: Schéma du tractus digestif des volailles et valeurs des pH des contenus digestifs
(Farner 1942).
Figure 2 : Représentation des différents genres microbiens autorisés en tant qu’additifs en
alimentation avicole en Europe (adapté de AFCA-CIAL, Mars 2009).
Figure 3 : les étapes de déroulement de la PCR.
Figure 4 : Arbre phylogénétique des principaux genres de bactéries lactiques et des genres
associés, obtenu par analyse des ARNr 16S (Stiles et Holzapfel, 1997).
Figure 5 : photo d’un tube digestif réalisée au laboratoire
Figure 6: dépôt des échantillons ADN sur gel d’agarose au laboratoire LGM
Figure 7 : montage de l’électrophorèse du laboratoire LGM
Figure 8: photos des observations microscopiques des isolats identifiés au genre
Lactobacillus sp. thermophiles.
Figure 9 : photos des observations microscopiques des lactobacilles mésophiles.
Figure 10 :photos des observations microscopiques des isolats pré-identifiés au Streptococcus
themophilus
Figure 11 : photos des observations microscopiques des isolats pré-identifiés au genre genre
Enteroccus sp.
Figure 12 : photos des observations microscopiques des isolats pré-identifiés à des
pédiocoques.
Figure 13: photos des observations microscopiques des isolats pré-identifiés à des
Leuconostocs.
Figure 14a : photos des observations microscopiques de Levures REF
Figure 14b: photos des observations microscopiques des isolats de levures.
Figure 15 : photos des observations microscopiques des isolats pré-identifiés au groupe
Bacillus.
Figure 16: photos des observations macroscopiques des souches productrices des EPS sur
milieux gélosé hypersaccharosé.
Figure 17 : observation macroscopiques montrant un trouble avec précipité blanc des EPS en
milieu liquide HJL.
Figure 18 : mise en évidence des EPS produits sur milieu rouge de ruthénium.
Figure 19: localisation des EPS par coloration vitale au bleu de toluidine sous microscope
optique GX1000.

Figure 20a : précipitation des EPS à l’éthanol .
Figure 20b : récupération du culot EPS.
Figure 21 : exopolysaccharides lyophilisés.
Figure 22 : activité rhéologique des EPS sur différents milieux.
Figure 23 : dosage colorimétrique de la gamme glucose et des différents échantillons
Figure 24 : courbe montrant les différentes concentration du glucose (20-120mg/l) en
fonction de la densité optique DO (490nm).
Figure 25 : dosage colorimétrique des deux dilution 1/10 et 1/100 des six EPS.
Figure 26 : exemple de résultats des dosages des dilutions finales d’un échantillon EPS.
Figure 27 : histogramme représentatif des concentrations des échantillons EPS des différents
isolats du tractus digestif
Figure 28: profil de la composition en oses des différents EPS par analyse sur ccm d’un
premier essai dans le système 1 (acide acétique : chloroforme : eau).
Figure 29 : profil de la composition en oses des différents EPS par analyse sur ccm d’un
deuxième essai dans le système 1 (acide acétique : chloroforme : eau).
Figure 30: profile de la composition en oses des différents EPS par analyse sur ccm dans le
système 2 (Acétone: Eau).
Figure 31 : activité antagoniste des différents isolats LGM vis-à-vis de S. aureus ATCC25923
par la méthode des disques.
Figure 32 : activité antagoniste des différents isolats LGM vis-à-vis de E. coli ATCC25922
par la méthode des disques.
Figure 33 : activité antagoniste de différents isolats vis-à-vis S. aureus ATCC 43300 par la
méthode des disques.
Figure 34 : activité antagoniste des différents surnageants natifs de différents isolats LGM
vis-à-vis d’E. coli ATCC2592 par la méthode des disques.
Figure 35 : activité antagoniste des différents surnageants natifs de différents isolats LGM
vis-à-vis de S .aureus ATCC25923 par la méthode des disques.
Figure 36 : activité antagoniste des surnageant neutralisé de différents isolats LGM vis-à-vis
d’E. coli ATCC2592 par la méthode des disques.
Figure 37 : activité antagoniste des surnageant neutralisé de différents isolats LGM vis-à-vis
de S .aureus ATCC25923 par la méthode des disques.
Figure 38 : activité antagoniste des différents Lactobacillus spp. vis-à-vis d’Enterococcus
faecalis JH2-2 par la méthode double couche.

Figure 39:activité antagoniste des différents Lactobacillus spp. vis-à-vis d’Enterococcus
faecalis V583 par méthode double couche.
Figure 40 : Amplification du fragment ADN 480 pb du gène de l’ADNr 16S en
électrophorèse sur gel d'agarose 0.8 %.
Figure 41 : Amplification du fragment ADN 1500 pb du gène de l’ADNr 16S en
électrophorèse sur gel d'agarose 1.3 %.

Contribution à l’étude des probiotiques et prébiotiques comme alternatives aux
antibiotiques en aviculture
Résumé
Le recours aux antibiotiques a connu ses limites en raison de l’émergence de nouvelles
souches multi résistantes causées par l’utilisation abusive de ces composés dans le secteur
avicole, malgré que l’antibioprévention reste actuellement le seul moyen utilisé pour contrôler
les problèmes sanitaires et économiques liés aux pathogènes aviaires, l’utilisation des
probiotiques s’avère nécessaire pour favoriser une bonne microflore antagoniste vis-à-vis des
pathogènes et peut s’inscrire comme stratégie alternative envisagée pour protéger les volailles
des agents pathogènes et pour remplacer les antibiotiques comme facteur de croissance.
Ce travail s’intéresse à l’identification par les méthodes ADN des souches de volaille
productrices de substances inhibitrices vis-à-vis de certains pathogènes Staphylococcus
aureus, Escherichia coli, Salmonella typhi. 11 de ces souches ont été retenues suite à leur
forte action inhibitrice. Au total 48 souches ont été isolées à partir de différents
compartiments du tractus digestif de deux coqs de race locale, dont 6 ont été pré-identifiés à
des leuconostocs, 2 à des pédiocoques, 8 à des entérocoques, 22 à des lactobacilles, 5
apparentés au genre Bacillus et enfin 4 autres appartenant à des levures.
Par ailleurs, l’utilisation des amorces d’une région interne de 480 pb du gène l’ADNr 16S
spécifiques au genre Lactobacillus sp. et le séquençage du fragment d’ADN amplifié ont
permis d’identifier les isolats appartenant à la flore lactobacillaire au nombre de 22.
Lactobacillus acidophilus, Lactobacillus gallinarum, Lactobacillus crispatus et Lactobacillus
johonossi ont été les souches les plus diagnostiquées par la méthode PCR colonie. Une autre
approche moléculaire a été entamée afin d’amplifier le gène ADNr 16S dans sa totalité 1500
pb chez certaines souches, celui-ci reste un taxon de choix en matière de systématique en
bactériologie. D’autres travaux menés sur la détection et l’extraction des prébiotiques tels que
les polysaccharides exogènes (EPS) ainsi que leur caractérisation par chromatographie sur
couche mince qui a montré une similitude des profils avec le dextrane commercial et leur
dosage colorimétrique qui a été déterminé à 1254 mg/l pour la production d’EPS la plus
élevée chez la souche LGM-IL2.
Mot clés : Lactobacillus spp, ADN, PCR, probiotiques, pathogènes, prébiotiques, CCM.

Contribution to the study of probiotics and prebiotics as alternatives to antibiotics in
poultry
Abstract:
The use of antibiotics had its limitations due to the emergence of new multi -resistant strains
caused by the misuse of these compounds in the poultry sector, despite the antibio-prevention
currently remains the only means used to monitor health problems and economic issues
related to avian pathogens , the use of probiotics is necessary to promote healthy microflora
antagonist towards the pathogen and can register as an alternative strategy intended to protect
poultry pathogens and to replace antibiotics as growth factor.
This work focuses on the identification by DNA methods of producing poultry strains of
inhibitory substances against pathogens such as Staphylococcus aureus, Escherichia coli, and
Salmonella typhi. 11 of these strains were selected due to their strong inhibitory action. A
total of 48 strains were isolated from different compartments of the digestive tract of two local
cocks , including pre-identified to leuconostocs was 6 , 2 to pediococci , 8 to enterococci , 22
lactobacilli, 5 related to genus Bacillus and yeasts belonging to 4 .
Furthermore, the use of primers a 480 bp internal region of the gene 16S rDNA specific to
the genus Lactobacillus sp. and sequencing of the amplified DNA fragment were identified
isolates belonging to the lactobacilli flora to 22. Lactobacillus acidophilus, Lactobacillus
gallinarum, Lactobacillus crispatus and Lactobacillus johonossi were the most diagnosed
strains by PCR method colony. Another molecular approach has been initiated in order to
amplify the 16S rDNA gene in its entirety in 1500 bp for some strains, which remains a taxon
choice of systematic bacteriology. Other work on the detection and extraction of prebiotics
such as exogenous polysaccharides (EPS) and their characterization by thin layer
chromatography which showed a similarity of profils with commercial dextrane and
colorimetric assay was determined to 1254 mg / l for the highest production of EPS by strain
LGM-IL2.
Key words: Lactobacillus spp, DNA, PCR, probiotics, pathogens, prebiotics, TLC.


كبدائل للمضادات الحيوية في الدواجن دراسة البروبيوتيك والبريبايوتك المساهمة في
ملخص
لقد عرف استخدام المضادات الحيوية حدوده بنسبة كبيرة نظرا لظهور سلالات بكتيرية مقاومة لهذه المضادات و السبب الاستعمال المفرط لها و التي لا تزال الوسيلة الوحيدة المستخدمة لرصد المشاكل الصحية المتعلقة بمسببات إلىراجع
استخدام البروبيوتيك ضروري لتعزيز الميكلوفلور الصحية المضادة للبكتيريا الممرضة أصبح الدواجن.عند الإمراض .الأمراضتسريع عملية نمو الدواجن و حمايتها من مسببات إلىبديلة تحل مكان المضادات تهدف إستراتيجيةباعتباره
النافعة من الجهاز الهضمي لديوك برية و تشخيصها بالوسائل الحديثة كالبيولوجيا السلالاتبعزل هذه الدراسة و لقد اهتمت . E. coli, S. aureusل تثبيط عمل البكتيريا الممرضة مث على و التي لديها القدرة ADNالجزئية المبنية على كشف ال
(probiotique)ذات منفعة لة سلا 48 عزلت بالإجمال .من هذه السلالات تم اختيارها بسبب قدرتها المثبطة القوية 11 , leuconostocs, 2 pédiocoques,8 entérocoques6 ,:منها من مختلف مناطق الجهاز الهضمي
5 Bacillus ,22 lactobacilles4 levures, .
خاصة بالنوع ADNr 16Sلمنطقة داخلية لمورثة الحمض النووي 480pb ذات تباستعمال قطع من النكليوتيدا
lactobacillus sp و تسلسل مورثة الحمض النووي المضخم (séquençage) عزلة من نوع 22سمح لنا بتحديد lactobacillus sp. .
Lactobacillus acidophilus, Lactobacillus gallinarum, Lactobacillus crispatus , Lactobacillus johonossi في مجملها ADNr 16Sكما استعملنا منهج اخر لتضخيم جينات . PCR ت الأكثر تشخيصا عن طريقالسلالا.كانت
لبعض السلالات . 1500pbذات
وكذلك تحديد خصائصها (EPS) ريبيوتيك يمتعددة الخارجية اللكشف و استخلاص السكريات ال أخرىدراسات أجريتالتي اظهرت تشابها ملحوظا في التركيبة الجزئية مع ديكسترات CCMعن طريق تقنية كروماتوغرافيا الطبقة الرقيقة
(Dextrane) كما تم تحديد الجرعة الخاصة بكل.EPS ب إنتاجكمية أعلىعن طريق التغير اللوني الذي حدد ل./مغ1264
مسببات الإمراض ,بروبيوتيك ,البريبيوتيك ,كرومتوغرافيا ., PCR, ,حمض نوويLactobacillus sp :ة كلمات مفتاحي

1
Introduction
En 2006, l’union Européenne a interdit définitivement l’emploi des antibiotiques comme
promoteurs de croissance en production avicole. L’antibiothérapie a connu ses limites en
raison de l’émergence de nouvelles souches pathogènes multi-résistantes causée par
l’utilisation abusive de ces composés dans le secteur avicole. Récemment de nouvelles
stratégies de prévention ont été proposées comme alternatives aux antibiotiques pour réduire
l’incidence des pathogènes entériques chez la volaille. Parmi ces stratégies le recours aux
probiotiques notamment les lactobacilles (Gionchetti et al., 2000 ; Fritts et al., 2000 ; Jin et
al., 1997) et les prébiotiques (Gibson et Roberfroid, 1995) semblent offrir les résultats les plus
prometteurs. Les microorganismes autorisés aujourd’hui en Europe en alimentation animale
appartiennent aux bactéries du genre Bacillus, Enterococcus, Lactobacillus, Lactococcus,
Streptococcus, Bifidobacterium, Pediococcus et aux levures, ces additifs probiotiques sont
principalement utilisés en mono-souches ou des multi-souches (Bernardeau et Vernoux,
2009). Les probiotiques sont définis par l’OMS comme des « microorganismes vivants, qui
lorsqu’ils sont administrés en quantité adéquate, confèrent un effet bénéfique pour la santé de
l’hôte au-delà de l’effet nutritionnel premier ». Certains probiotiques comme Lb. johsonii
réduit le potage de salmonella (Casey et al., 2004), bifidobacterium , enterococcus et
pediococcus augmentent les paramètres de performances zootechniques et modulent la
composition de la microflore du caecum (Mountzouris et al.,2007), d’autres parmi eux,
comme pediococuus acidilactici ou Saccharomyces boulardii améliorent la résistance aux
coccidioses (Eimeria acervulina , Eimeria tenella) (Lee et al., 2007) .Par ailleurs, les auteurs
ont rapporté que les avantages dus aux lactobacilles en qualité de probiotiques sont mulitples
tels l’inhibition des campylobacters (Messaoudi et al., 2011) et l’inhibition d’Eimeria tenella
in vitro (Tierney et al., 2004). ainsi que l’action de Lactobacillus johnsonii FI9785 dans le
contrôle des entérites nécrotiques endémiques dues à Clostridium perfringens, réduisant les
pertes économiques et l’utilisation d’antibiotiques (La Ragione et al., 2004) .
Au titre de l’usage des probiotiques, un cadre législatif a été établi sur leur utilisation en 2002
par l’EFSA (European Food Safety Agency) d’où ressort trois pricipaux chapitres; l’identité,
la sécurité et l'efficacité de la souche. L’identification des microorganismes par les méthodes
ADN est devenue une méthode incontournable (Gevers et al., 2001, O’Mahony et al., 2000),
elle est plus résolutive et plus discriminante par rapport aux méthodes d’identification et
classification traditionnelles.

2
L’objectif de ce travail consiste à isoler à partir du tube digestif de la volaille fermière des
souches du genre Lactobacillus sp. à fort potentiel probiotique et de les identifier par les
méthodes d’ADN en utilisant des amorces spécifiques au genre Lactobacillus sp. d’une région
interne du gène l’ADNr 16S de 480 pb par l’utilisation des amorces spécifiques (V3/V4). Les
autres souches de la flore totale ont été au même titre isolées, pré-identifiées par des méthodes
phénotypiques et certaines aptitudes ont été caractérisées, notamment chez les leuconostocs
telles que la mise en évidence des polysaccharides produits par microscopie en utilisant le
colorant bleu de toluidine et la sélection des souches mucoïdes productrices d’EPS par
l’utilisation d’un milieu spécifique au rouge au ruthénium .
En outre, une méthode d’extraction de ces exopolysaccharides au nombre de 8 sera effectué
afin de pouvoir les caractériser au niveau monosaccharidique par la chromatographie sur
couche mince ainsi qu’un dosage colorimétrique sera réalisé.
Dans un autre chapitre, une activité à caractère antagonisme sera recherchée in vitro vis-à-vis
de certains pathogènes, en l’occurrence Staphylococcus aureus (ATCC 25923, ATCC 43300),
Escherichia coli ATCC25922, Enterococcus faecalis V583 Enterococcus faecalis JH2-2 et
Salmonella typhi.
Une amplification du gène ADNr 16S de 1500 pb par l’utilisation des amorces universelles de
quelques souches sera effectuée.

Synthèse Bibliographique
3
Chapitre 1 : Généralités
1. Le microbiote intestinal chez la volaille
Selon la définition d'Isolauri et ses collaborateurs, le microbiote intestinal normal est un
consortium complexe et en équilibre de microorganismes qui habitent normalement le tractus
gastro-intestinal et qui remplissent un rôle dans la nutrition, la physiologie et le
fonctionnement du système immunitaire de l'hôte (Isolauri, Sutas et al. 2001). La composition
de ce microbiote intestinal est en équilibre relativement stable dans le tube digestif. Cet
équilibre peut être rompu avec l'âge, les conditions d'hygiène, le stress ou à la suite d'une
agression extérieure comme lors de l'utilisation d'antibiotiques, de facteurs de croissance
(Gabriel, Mallet et al. 2003). Ainsi, on note des populations microbiennes plus élevées chez
des animaux élevés au sol sur litière propre ou litière contaminée par une bande précédente
par rapport à des animaux élevés en cage individuelle (Gabriel, Mallet et al. 2003). Selon les
conditions d'élevage, l'augmentation de la densité d'élevage ou les stress thermiques semblent
globalement augmenter les bactéries néfastes au détriment des bactéries bénéfiques (Gabriel,
Mallet et al. 2005). Outre ces conditions, la présence de parasites intestinaux comme les
coccidies, peut entraîner la dégradation de la muqueuse intestinale et la production de
nouveaux substrats pour la microflore, modifiant ainsi sa composition (Kimura, Shiosaka et
al. 1976). La flore est modifiée aussi par l'alimentation. Ainsi, le type de céréales en
particulier la présence de polysaccharides non amylacés hydrosolubles (Mathlouthi, Mallet et
al. 2002) ou leur mode de présentation (Gabriel, Mallet et al. 2003) entraînent des
changements de la flore. De même, les matières grasses, ou le type d'amidon peuvent avoir un
effet sur la composition du microbiote (Weurding, Enting et al. 2003).
2. Description du tube digestif chez la volaille
Le tractus gastro-intestinal présente quelques particularités anatomiques (Figure 1). On
distingue différents compartiments ; la cavité buccale ne comprend ni lèvres ni dents, mais un
bec corné qui permet la préhension et une certaine fragmentation des aliments. Les glandes
salivaires sont peu développées. Il n'y a ni voile de palais, ni épiglotte, si bien que la
déglutition est un phénomène uniquement mécanique par redressement de la tête. Dans la
bouche, les aliments sont peu fragmentés et grossièrement insalivés (M. Larbier and
Leclercq1992). L'œsophage contient un renflement dont l'épithélium est riche en glandes à
mucus : le jabot. Cet organe de pH variant entre 4,47 et 4,54 (Farner 1942) peut entreposer

Synthèse Bibliographique
4
des aliments qui s'y humectent et s'y ramollissent, il fonctionne chez le poulet alimenté à
volonté. Il est le lieu d'une digestion microbienne et comporte essentiellement des
Lactobacilles, d'une partie de l'amidon (hydrolyse avec formation d'acides lactique) et de
formation d'acide gras volatiles (M. Larbier and Leclercq 1992). Le proventricule est riche en
glandes sécrétoires (acide chlorhydrique et pepsinogène précurseur de la pepsine) et
permettant la digestion chimique : c'est l'estomac chimique. La protéolyse y débute à pH de 3
à 4,5. Dans le gésier et le proventricule, le faible pH fait chuter la population bactérienne
(Farner 1942). Le gésier, estomac mécanique est caractérisé par une couche superficielle très
dure entourée de muscles puissants. Il y règne un pH très bas (2 à 3,5) et il peut contenir de
petits graviers qui sont nécessaires aux animaux consommant des grains intacts. C'est donc au
niveau du gésier que se produit véritablement la protéolyse sous l'action de la pepsine
(Gabriel, Mallet et al. 2005). Dans l'intestin, l'environnement devient plus favorable à la
croissance bactérienne en raison de la plus faible pression d'oxygène et de la faible
concentration en enzyme et en sels biliaires et d'un pH variant dans le duodénum entre 5,68 et
6,07, dans le jéjunum entre 5,72 et 6, dans le caecum entre 5,6 et 5,83, l'iléon entre 6,18 et
6,50 et dans le colon entre 6,08 et 6,58 (Farner 1942).
À la naissance les poussins sont axéniques. Le microbiote spécifique commence à se
développer dès les deux premiers jours pour donner lieu à une flore microbienne
environnementale spécifique après trois à six semaines (Methner, Barrow et al. 1997). En
effet, après l'éclosion, la flore augmente rapidement. Ainsi dès le premier jour, l'iléon et le
caeum hébergent 108
et 1010
bactéries par gramme de contenu digestif. Leur nombre atteint
109
et 1011 bactéries par gramme en 3 jours et reste relativement stable jusqu'à l'âge de 30
jours (Gabriel, Mallet et al. 2005). La flore est composée essentiellement de bactéries à Gram
positif anaérobies facultatives du jabot à l'iléon terminal, alors que le caecum contient en plus
des anaérobies stricts, ces dernières étant dominantes (Gabriel, Mallet et al. 2005).
D'un point de vue qualitatif, dès le premier jour, les coliformes, les streptocoques et les
clostridies colonisent rapidement le tube digestif, du jabot au caecum, alors que les
lactobacilles et les Bactéroïdes ne sont mis en évidence dans le caecum qu'après 3 jours et 5
jours respectivement (Gabriel, Mallet et al. 2005). Les populations bactériennes présentes
dans le tractus digestif représentent une large gamme de types métaboliques et
morphologiques. Leur nombre total est plus important que le nombre de cellules eucaryotes
constituant le corps de l'hôte. On distingue les bactéries dominantes (>106
UFC /g contenu),
sous-dominantes (105
à 103
UFC / g contenu), et résiduelles (<103
UFC / g contenu). Chez le

Synthèse Bibliographique
5
poulet, les sites principaux d'activité bactérienne sont le jabot, le caecum et, dans une moindre
mesure, l'intestin grêle (Cole and Fuller 1984). Ainsi, dans le caecum et l'iléon, on trouve
respectivement 1011
et 109
bactéries par g de contenu (Apajalahti, Kettunen et al. 2004). Les
études effectuées sur le microbiote des oiseaux ont concerné principalement le caecum
(Gabriel, Mallet et al. 2005).
3. Description de la flore digestive du poulet et localisation dans le tractus digestif :
La flore digestive peut se trouver dans la lumière intestinale ou adhérer à la muqueuse
digestive. La flore luminale dépend des nutriments disponibles, de la vitesse de transit et de la
présence ou non de substances antimicrobiennes. La flore des muqueuses dépend de
l’expression par l’hôte de sites d’adhésion spécifiques sur les membranes des entérocytes, de
la vitesse de production de mucus, de la production d’anticorps (Ig) sécrétoires, et de
l’extrusion de matériel cellulaire de la membrane.
La flore digestive comprend des bactéries et des champignons. Chez le poulet, 29 genres
bactériens ont été identifiés (Fuller, 1984). Chaque genre serait représenté par 3 à 4 espèces,
et chaque espèce par 3 à 4 types métaboliques différents, ce qui ferait plus de 200 types
différents, sachant que seulement 25% des souches seraient identifiées. Le tube digestif
contient donc une large population bactérienne de différents types métaboliques et
morphologiques. Ainsi, le nombre total de cellules bactériennes est plus important que le
nombre de cellules eucaryotes constituant le corps de l’hôte.
Tableau 1: Nombre de bactéries viables (log10 / g de contenu) des groupes majoritaires dans
le tube digestif du poulet (d’après Smith, 1965)
log10 Jabot Gésier Duodénum Iléon Caeca
Lactobacilles 8.7 7.3 8.0 8.6 8.7
Entérocoques 4.0 3.7 4.0 4.2 6.7
Coliformes 1.7 - 2.0 2.7 5.6
Levures 2.7 - 1.7 - 2.0
Clostridies - - (-) (-) 9.0
Anaérobies
obligatoire
non sporulant
- - - - 10.0
Streptocoques
anaérobies
- - - - 10.0

Synthèse Bibliographique
6
Figure 1: Schéma du tractus digestif des volailles et valeurs des pH des contenus digestifs
(Farner 1942).
Chez le poulet, les deux sites principaux d’activité bactérienne sont le jabot et les caeca.
Globalement, la flore du jabot à l’iléon terminal est composée principalement d’anaérobies
facultatifs alors que les caeca contiennent en plus des anaérobies stricts, ces derniers étant
dominants (Fuller, 1984). Dans le jabot, on trouve principalement des lactobacilles qui
peuvent être attachés à l’épithélium en formant presque une couche complète. On trouve aussi
des entérocoques, des coliformes, et des levures. Dans le gésier et le proventricule, le faible
pH fait chuter la population bactérienne. Dans le duodénum, le nombre important d’enzymes,
la forte pression en oxygène et la présence de fortes concentrations de composés
antimicrobiens tels que les sels biliaires limitent la croissance bactérienne. On trouve
principalement des lactobacilles ainsi que des entérocoques et des coliformes. Plus loin dans
l’intestin, l’environnement devient plus favorable à la croissance bactérienne à cause de la
plus faible pression d’oxygène, la faible concentration en enzymes et sels biliaires
(réabsorption et dégradation en partie par la microflore). Si les aliments sont bien digestibles,
par manque de substrat, la flore est limitée. Dans l’iléon, on trouve principalement des
lactobacilles attachés aux entérocytes, des entérocoques et des coliformes. Dans le caeca, on

Synthèse Bibliographique
7
trouve une large population de types morphologiques variés, enfouie dans la couche de mucus
et attachée à l’épithélium. En effet, le contenu de cet organe étant rarement renouvelé (1 à 2
fois/jour), cela le rend favorable au développement des bactéries. On trouve en majorité des
anaérobies stricts comme les Eubacterium, des bifidobactéries ou des clostridies. On trouve
aussi des anaérobies facultatifs comme des lactobacilles, des entérocoques, et des coliformes.
4. Les pathogènes entériques de la volaille
4.1. Maladies entériques chez la volaille
Les volailles sont prédisposées à de nombreuses infections parasitaires, bactériennes,
mycoplasmiques et virales en particulier celles des systèmes respiratoire et gastro-intestinal.
Les maladies entériques à forte prévalence comme la cryptosporidiose ou la paratuberculose,
conduisent à des pertes économiques élevées dans les élevages. De nombreuses espèces de
Salmonella peuvent attaquer les oiseaux adultes et causer des taux élevés de mortalité et de
morbidité. Une autre problématique liée à la présence de ces pathogènes dans le tube digestif
est la contamination des denrées alimentaires provenant de la volaille. Les infections par le
Welchia sont responsables de l'entérite nécrosante. Les infections infracliniques résultent dans
des performances altérées et des lésions du foie. La maladie clinique aiguë entraîne une
mortalité accrue (Chafai 2006).
Le traitement de choix suppose l'utilisation d'antibiotiques, mais ne prévient pas la récurrence
de la maladie après arrêt de la médication. Les mesures de contrôle consistent en une pratique
d'hygiène adéquate dans la chaîne de production et au niveau du consommateur.
4.1.1. Escherichia coli :
Les Escherichia coli aviaires, bien que considérés par beaucoup comme pathogènes
secondaires, représentent à l'heure actuelle l'une des plus importantes causes de pertes
économiques dans le secteur avicole. Les Escherichia coli sont des hôtes commensaux du
tractus digestif de la volaille et la plupart des souches ne sont pas pathogènes. Cependant, un
certain nombre de celles-ci appelées « Avian Pathogenic E. coli » ou APEC et appartenant à
des sérotypes bien particuliers sont associées au syndrome de la colibacillose (Stordeur and
Mainil 2002). La voie d'entrée principale de l'agent pathogène est le tractus respiratoire, via
l'inhalation de particules de poussières contaminées par les E. coli excrétées du tractus
digestif d'animaux sains. Les intestins sont, en effet, le réservoir le plus important des E. coli
pathogènes aviaires ou APEC. Après une première multiplication au niveau du tractus
respiratoire supérieur, les bactéries colonisent les voies respiratoires profondes à savoir les

Synthèse Bibliographique
8
sacs aériens et les poumons. Dans une troisième étape, la bactérie atteint le sang et colonise
les organes internes comme le coeur, le foie et la rate (Jordan and Pattison 1996).
Les colibacilloses sont sans doute les infections bactériennes les plus fréquentes et les plus
importantes en pathologie aviaire. Elles peuvent entrainer de la mortalité, des baisses de
performances et des saisies à l'abattoir (Stordeur and Mainil 2002).
Contrairement aux infections des mammifères, les colibacilloses aviaires prennent des formes
générales, avec une voie d'entrée respiratoire ou génitale (Stordeur and Mainil 2002). L'agent
étiologique de la colibacillose est la bactérie Escherichia coli.
4.1.2. Salmonella
Plus de 60 % des toxi-infections dans le monde sont dues à Salmonella. Les salmonelloses
sont de ce fait, devenues un phénomène de santé publique ce qui justifie l'implication de
l'Organisation Mondiale de la Santé (OMS) dans la lutte contre les salmonelloses (Salm-Surv
2005).
Les salmonelles appartiennent à la famille des entérobactéries, ce sont des bacilles mobiles à
Gram négatif, aéro-anaérobies facultatifs, mésophiles qui se cultivent facilement. À nos jours
plus de 2300 serotypes différents de salmonelles sont identifiés, sur le plan épidémiologique
et sont classées en fonction de leur potentiel pathogène pour l'homme ou l'animal. La
virulence des salmonelles est une notion complexe, résultant de nombreux facteurs encore
largement étudiés, tant au niveau biochimique que génétique. Les principaux facteurs de
virulence sont la mobilité (reposant sur les flagelles), l'adhésion par les pili et les fimbriae
(phénomène actif de reconnaissance spécifique entre une adhésine bactérienne et un ligand
présent à la surface d'une cellule hôte), l'invasion (par endocytose pour les entérocytes, par
phagocytose pour les macrophages), la formation de phagosomes spacieux et la fusion avec
les lysosomes (Feuillet 2007).
Actuellement plus de 200 sérotypes de salmonelles sont connus chez la volaille. Salmonella
enteritidis qui infecte les organes profonds (foie, rate, ovaire) est à l'origine d'infection
durable au niveau des troupeaux alors que Salmonella typhimurium est actuellement le
sérotype le plus incriminé dans les salmonelloses aviaires provoquant les formes cliniques les
plus graves, surtout chez les jeunes poussins prenant une allure septicémique, avec une
mortalité brutale dans les jours qui suivent l'éclosion. On observe également de nombreuses
mortalités en coquille. La première étape de colonisation par les salmonelles est une étape de
colonisation intestinale. La bactérie arrive dans l'intestin grêle, où elle se multiplie en
adhérant à l'épithélium, elle pénètre par un phénomène d'endocytose dans les cellules

Synthèse Bibliographique
9
épithéliales iléales et caecales, notamment les tissus lymphoïdes incluant les plaques de Peyer,
les amygdales caecales et dans les cellules M. Dans le cas des salmonelles provoquant des
maladies systémiques, le site d'attachement préférentiel se situe au niveau des plaques de
Peyer (Feuillet 2007). L'infection est strictement limitée à la sphère digestive et peut
correspondre à un portage latent avec élimination épisodique des Salmonelles dans les fèces.
5. Contrôle des infections chez la volaille
Le recours aux antibiotiques demeure l'approche la plus préconisée pour lutter contre ces
infections. L'utilisation des antibiotiques en alimentation animale s'est progressivement
développée dès le début des années 50 et a permis d'améliorer les conditions sanitaires des
animaux et d'accroître la productivité des élevages en réduisant les coûts de production.
Cependant, l'utilisation abusive de ces antibiotiques qui possèdent beaucoup de similitudes avec
ceux utilisés en médecine humaine a entraîné une augmentation inquiétante du nombre de souches
pathogènes multi-résistantes.
Cette situation a conduit l'Union européenne à décréter, en 2006, l'interdiction complète des
antibiotiques à titre de facteurs de croissance dans les aliments pour animaux dont les premières
espèces animales touchées par cette interdiction sont les porcs et les volailles, principales
productions à recourir à ce type d'additifs. Au Canada, l'utilisation des antibiotiques comme
facteur de croissance n'est pas formellement interdit, mais il est déconseillé. Cependant, le
Québec jouit d'une réglementation unique au Canada puisque tous les médicaments destinés aux
animaux ne sont disponibles que sur ordonnance (CAAAQ 2008). De surcroît, aucun producteur
ne peut détenir ou administrer un médicament à un animal de consommation à moins qu'il n'ait
été prescrit par un médecin vétérinaire (selon l'Ordre des médecins vétérinaires du Québec).
Il est donc clair que la recherche de solution alternative susceptible d'assurer une succession
satisfaisante aux antibiotiques en termes d'effets zootechniques devient une urgence.

Synthèse Bibliographique
10
Chapitre 2 : Les probiotiques en aviculture
1. Histoire d’utilisation des probiotiques en alimentation animale
Les probiotiques ont été commercialisés et utilisés dans les fermes à partir des années
1960.Leur utilisation a été encouragée (1) par le Comité Swann en 1969 qui recommandait de
restreindre l’usage des antibiotiques en alimentation animale à la seule fin thérapeutique (leur
utilisation « facteurs de croissance » étant associée à l’augmentation des résistances
bactérienne) ; (2) par la nécessité de faire face aux conséquences d’une production animale
toujours plus intense et stressante pour les animaux (économie d’échelle, augmentation de la
taille des élevages, concentration des animaux, sevrage précoce, …). Entre les années 1970 et
1990, les micro-organismes probiotiques revendiquaient des propriétés zootechniques,
amélioration du gain de poids, du coefficient de digestibilité, et également des effets sanitaires
(diminution des diarrhées, de la morbidité, …). Mais cette période est aussi marquée par
l’absence de cadre réglementaire contribuant à réduire la confiance des utilisateurs et dès le
début des années 1990, on observe un déclin de l’utilisation des probiotiques sur le marché
européen. Cette première vague d’utilisation des probiotiques en alimentation animale
jusqu’en 1993 a été définie par Bernardeau et Vernoux (2009) comme « la première
génération de probiotiques », caractérisée par une efficacité supposée et un cadre
réglementaire peu adapté. L’absence d’efficacité (Simon et al., 2001), de compréhension du
mécanisme d’action et le manque de données scientifiques ont amené les professionnels de la
production animale (vétérinaires, nutritionnistes, éleveurs) à considérer le concept probiotique
avec grand scepticisme (Bernardeau et Vernoux, 2009).
C’est le formidable essor de l’utilisation des probiotiques en alimentation humaine et les
avancées scientifiques en matière d’écologie digestive et d’interactions microbiennes
(Caramia, 2004) qui vont relancer l’utilisation des micro-organismes en alimentation animale.
Stimulées également par la nécessité de palier à l’interdiction des antibiotiques facteurs de
croissance décrétée en 2006 en Europe, les études scientifiques sur les probiotiques sont
mieux établies, réalisées en double aveugle, contrôlées, plus fiables et la directive Européenne
de 1993 remet les micro-organismes probiotiques à l’honneur en les incluant dans la
réglementation des additifs pour l’alimentation animale. Parallèlement, les productions
animales connaissent entre 1980 et 2000, une série de crises qui va remodeler complètement
le paysage réglementaire, aussi bien au niveau institutionnel (création de l’AFSSA – Agence

Synthèse Bibliographique
11
Française de Santé et Sécurité Alimentaire en France 1998 et l’EFSA – European Food Safety
Agency au niveau européen en 2002) que législatif (refonte complète de la réglementation de
l’utilisation des additifs en alimentation animale – Dir. 70/524/EC et clarification de
l’utilisation des micro-organismes comme additifs Reg. 1831/2003/EC). Cette nouvelle
réglementation très rigoureuse exige de la part des industriels des données scientifiques et
technologiques incluant la démonstration de l’innocuité des micro-organismes (pour l’animal,
le travailleur, le consommateur et l’environnement) et la preuve de leur efficacité en accord
avec les revendications zootechniques et/ou digestives (Mantovani et al., 2006). Les trois
volets du dossier d’enregistrement européen appliqués aux probiotiques sont (1) identité et
qualité: caractéristiques de la souche (taxonomie, métabolisme, propriétés, …), processus de
fabrication, stabilité du probiotique (seul ou en mélange), méthode d’analyse ; (2) sécurité :
pour l’espèce cible (innocuité à 10 fois la dose recommandée), pour le manipulateur, le
consommateur (absence d’antibiorésistance, génotoxicité et mutagénicité) et pour
l’environnement ; (3) efficacité : à démontrer pour l’espèce cible par un minimum de trois
études significatives dans deux lieux différents. Le volet efficacité décrit l’espèce cible, les
conditions (âge, stade physiologique, type de production), les doses d’utilisation, les
performances revendiquées ainsi que les mécanismes d’action possibles. Les allégations
possibles pour des probiotiques peuvent concerner des effets sur la performance animale, la
production animale, le bien-être animal ou l’environnement.
La difficulté scientifique, la charge financière et la complexité des dossiers d’autorisation ont
définitivement mis un terme à l’utilisation abusive et non fondée des probiotiques en
alimentation animale. Apparaissent alors sur le marché, des probiotiques plus sûrs, plus
efficaces et plus transparents que Bernardeau et Vernoux (2009) caractérisent de «
probiotiques de deuxième génération »
2. Définition des probiotiques
Le terme "probiotique" est un mot relativement nouveau qui signifie "en faveur de la vie". Le
concept probiotique est né de la théorie de la longévité de Metchnikoff en 1907. Il fut le
premier à proposer l'utilisation des Lactobacilles des yaourts pour la restauration du
microbiote dans le tractus gastro-intestinal. Les probiotiques ont d'abord été développés dans
les années 1960 pour les élevages d'animaux afin de prévenir les infections et stimuler le gain
de poids. La première définition officielle a été proposée par Fuller en 1989 qui définit un
probiotique comme étant « un supplément alimentaire microbien vivant qui affecte
positivement la santé de l'animal en améliorant sa balance microbienne intestinale ». Cette

Synthèse Bibliographique
12
définition a été révisée plusieurs fois, notamment par la FAO (Food and Agriculture
Organization of the United Nations) et la WHO (World Health Organization). En 2001, leur
nouvelle définition s'énonce comme suit : « Les probiotiques sont des microorganismes
vivants qui lorsqu'ils sont administrés en quantité adéquate, produisent un effet bénéfique
pour la santé de l'hôte ».
3. Les micro-organismes probiotiques autorisés aujourd’hui en alimentation avicole
De nombreuses espèces microbiennes ont été utilisées en tant qu’agents probiotiques. Ces
micro-organismes appartiennent aux bactéries du genre Bacillus, Bifidobacterium,
Enterococcus, Escherichia coli, Lactobacillus, Lactococcus, Streptococcus et aux levures du
genre Saccharomyces. Les micro-organismes utilisés en alimentation animale diffèrent
sensiblement de ceux utilisés en alimentation humaine. Ces variantes intègrent les différences
rencontrées au niveau des objectifs d’efficacité, des aspects sécuritaires, des fréquences
d’ingestion, des contraintes de fabrication ou encore de stockage ou encore de la
réglementation.
Les genres Lactobacillus et Bifidobacterium sont majoritairement utilisés pour des
applications en nutrition humaine, alors que les genres Bacillus, Enterococcus et
Saccharomyces sont les micro-organismes les plus utilisés dans les élevages (Simon et al.,
2001) (Tableau 3). Les souches de Bacillus, plus stables car sporulées, sont plus à même de
résister aux processus d’incorporation dans l’aliment, aux paramètres de granulation et aux
conditions non exigeantes de stockage « longue durée » des aliments pour animaux (Simon,
2005). Inversement, les cellules végétatives sont beaucoup plus sensibles, ce qui explique que
les lactobacilles ou les bifidobactéries, pourtant bien documentées, ont été moins utilisées au
début en alimentation animale. Mais les techniques de stabilisation et de protection évoluant
(enrobage, encapsulation), cinq souches de lactobacilles sont aujourd’hui autorisées et
plusieurs dossiers actuellement soumis à l’EFSA pour homologation portent sur ces
microorganismes autrefois considérés comme sensibles (Bifidobacteria, Lactobacillus…).
Ainsi les lactobacilles, avec une grande diversité d’espèces (Tableau 3), représentent
aujourd’hui 21% des souches utilisées comme additifs en alimentation porcine et avicole,
3ème groupe microbien après les genres Bacillus et Enterococcus représentant chacun 29%
des utilisations (Figure 2). De nouvelles tendances sont également perçues concernant le
nombre de souches constitutives des produits. La première génération de probiotiques était
multi-souches. En 2007, avec la nouvelle réglementation en vigueur, les additifs probiotiques
sont principalement mono-souche (85%), seuls quelques additifs contiennent deux souches

Synthèse Bibliographique
13
(15%). Cette tendance tient vraisemblablement du fait que la préparation des dossiers
scientifiques et le processus d’homologation sont longs et difficiles. Cependant, là encore, la
situation évolue et plusieurs produits contenant deux ou plusieurs souches sont actuellement
examinés par l’EFSA.
Tableau 2 : Micro-organismes probiotiques autorisés en Europe pour la volaille (liste publiée par
l’AFCA-CIAL, dernière mise à jour Mars 2009).
Espèces animales Souches avec appellation commerciale N° Enr
Poulets
d’engraissement
Bacillus subtilis C-3120 –DSM 15544 – CALSPORIN – Calpis
co.Ltd – ORFFA
4b1720
Bacillus subtilis DSM 17299 – O35 / Chr.Hansen 4b1821
Bacillus amyloliquefaciens CECT 5940- ECOBIAL/ norel SA 4b1822
Sta
bil
isan
te
de
la f
lore
Enterococcus faecium DSM 3530
BIOMIN IMB52 / Biomin GmbH
4b1850
Dindes
engraissement
Bacillus cereus var. toyoi NCIMB 40112/CNCM I-1012
TOYOCERIN/ Rubinum
4b1701
Mic
ro-o
rgan
ism
es
Dindons
engraissement
Bacillus lichenifornis DSM5749 et Bacillus subtilis DSM5750
BIOPLUS2B
E1700
Enterococcus faecium DSM 10663 /NCIMB10415- Oralin E1707
Lactobacillus farciminis CNCM MA 67/4R - BIACTON 12
Poulets
engraissement
Bacillus cereus var. toyoi NCIMB 40112/CNCM I-1012 -
TOYOCERIN
E1701
Enterococcus faecium NCIMB 10415 - CYLACTIN E1705
Enterococcus faecium DSM 10663/NCIMB 10415 - ORALIN E1707
Enterococcus faecium NCIMB 11181 -LACTIFERM 15
Enterococcus faecium ATCC 59519 et Enterococcus faecium
ATCC 55593 - PROPIOS -PIONEER PDFM
E1709
Enterococcus faecium CECT 4515 - FECINOR 18
Pediococcus acidilacti CNCM MA18/5M- Bactocell-
FERMAID
E1712
Lactobacillus farciminis CNCM MA 67/4R - BIACTON 12
Poulets pondeuses Lactobacillus farciminis CNCM MA 67/4R - BIACTON 12
Lactobacillus acidophilus D2/CLS CECT 4529 E1715

Synthèse Bibliographique
14
Figure 2 : Représentation des différents genres microbiens autorisés en tant qu’additifs en
alimentation avicole en Europe (adapté d’AFCA-CIAL, Mars 2009).
Jusqu’à présent, les additifs microbiens selon cette réglementation, peuvent revendiquer des
propriétés zootechniques (relatives aux performances de croissance des animaux), digestives
et stabilisatrices de la microflore intestinale. Cependant force est de constater qu’en élevage,
les propriétés des microorganismes dépassent les seuls effets « croissance », qui doivent
plutôt être considérés comme une résultante de l’amélioration de l’état de santé général de
l’animal. Ces dernières années, des études scientifiquement ont ainsi élargi le potentiel
d’utilisation des souches probiotiques. Des applications préventives de pathologies digestives
ou immunostimulantes ont ainsi été démontrées aussi bien en élevage porcin qu’avicole
(Tableau 3). Dans un contexte d’assainissement des pratiques d’élevage vers une stratégie
plus naturelle et respectueuse de l’environnement et du bien-être animal, les micro-
organismes probiotiques présentent donc un réel potentiel de développement commercial.
4. Critères de sélection des probiotiques
Les micro-organismes doivent posséder diverses propriétés de survie pour répondre à la
définition des probiotiques (Gagnon 2007). Ils doivent présenter une activité positive et
persister durant leur passage dans le tractus digestif. Ces propriétés sont propres à chaque
souche et ne peuvent pas être extrapolables d'une souche à l'autre même au sein d'une même
espèce (Dunne, O'Mahony et al. 2001). Plusieurs critères majeurs de sélection ont été établis
Bacillus 29%
Enterococcus spp.29%
kluyveromyces spp. 4%
Lactobacillus spp. 21%
Pediococcus spp. 4%
Saccharomyces spp. 13%
Synthèse Bibliographique
15
par différents auteurs dans le but de sélectionner les souches potentiellement probiotiques.
Ces critères, résumés dans le tableau 1.3, sont réparties en trois catégories à savoir les critères
de sécurité, fonctionnels et technologiques.
Tableau 3 : Principaux critères de sélection des probiotiques. Adapté de (Klaenhammer and
Kullen 1999; Saarela, Mogensen et al. 2000; Ouwehand, Salminen et al. 2002; Gueimonde
and Salminen 2006).
Critère de
sécurité
• Identification taxonomique précise
• Origine humaine pour utilisation chez l’humain
• Souche caractérisée par des techniques phénotypiques et
génotypiques
• Historique de non pathogénicité et non-invasion de l’épithelium
intestinal
• Pas de transmission possible de gènes de résistance aux
antibiotiques
Critères
fonctionnels
• Tolérance à l’acidité, à la bile et aux enzymes digestives
• Adhésion aux cellules intestinales et persistance dans le tractus
intestinal
•Production de substances antimicrobiennes (bactériocines, acides
organiques, peroxyde d’hydrogène ou autres composés inhibiteurs
et antagonisme envers les pathogènes
• Immunomodulation
• Aptitude à produire des effets bénéfiques sur la santé
Critères
technologiques
• Stabilité au cours des procédés de production et dans le produit
fini
•Conservation des propriétés probiotiques après production
• Non modification des propriétés organoleptiques du produit fini

Synthèse Bibliographique
16
Parmi les critères reliés à la sécurité, l’identification taxonomique de la souche est une étape
importante dans l’établissement de nouvelles souches potentiellement probiotiques (Holzapfel
et al., 2001). Chaque souche doit être identifiée par des techniques moléculaires fiables et
confrontée à une nomenclature actualisée (FAO/WHO, 2002; Gueimonde et Salminen,
2006). Actuellement, l’hybridation ADN-ADN est la méthode moléculaire de référence pour
identifier l’espèce d’une souche, mais cette méthode est longue et requiert une large collection
de souches de référence (FAO/WHO, 2002). Le séquençage de l’ADN codant pour l’ARN
16S ribosomal est considéré aussi pertinent (FAO/WHO, 2002).
Dans ce dernier cas, il est recommandé que la technique soit combinée avec des tests
phénotypiques pour confirmation. L’origine de la souche est également une condition
importante car l’interaction spécifique avec l’hôte est maximisée lorsqu’elle provient du
même habitat (Alvarez-Olmos et Oberhelman, 2001). Les souches probiotiques doivent
également être sans effet négatif et être sécuritaires pour la santé humaine. À ce titre, les
souches potentiellement probiotiques seront évaluées afin qu’aucun des effets secondaires
suivants ne soient détectés: infections systémiques, activité métabolique nuisible, stimulation
immune excessive chez des individus susceptibles et transfert de gènes (par exemple de
résistance aux antibiotiques) (FAO/WHO, 2002). À ce sujet, les souches de lactobacilles et
bifidobactéries associées aux aliments possèdent un historique de sécurité de longue date et
très peu de corrélations existent entre des infections systémiques et la consommation de ces
probiotiques dans la littérature (Borriello et al., 2003; Marteau et al., 2006).
Pour assurer une fonctionnalité optimale, les souches probiotiques doivent conserver leur
viabilité jusqu’à leur site d’action. Cependant, il existe une controverse autour de ce critère
notamment par rapport à l’effet sur le système immunitaire. Même s’il est reconnu que les
cellules bactériennes mortes peuvent apporter des bénéfices physiologiques (Sanders, 2003),
la définition actuelle des probiotiques insiste sur le paramètre de viabilité (FAO/WHO, 2002).
Avant d’atteindre le tractus intestinal, les probiotiques doivent résister principalement à
l’environnement acide de l’estomac (pH compris entre 2,0 et 3,4) et à la bile sécrétée dans le
duodénum (Dunne et al., 2001; Gueimonde et Salminen, 2006). Le degré de tolérance à ces
conditions diffère pour chaque souche (Havenaar et Huis in’t Veld, 1992). Des tests in vitro
sont réalisés pour sélectionner les probiotiques qui maintiendront leur intégrité cellulaire et
leur activité métabolique lors du passage dans le tractus digestif humain.
La capacité d’adhésion à la muqueuse intestinale est également une des propriétés essentielles
que les souches probiotiques se doivent de posséder (Saarela et al., 2000; Tuomola et al.,
2001). L’adhésion permet d’accroître le temps de rétention des probiotiques dans l’intestin et

Synthèse Bibliographique
17
met en contact étroit les bactéries et les cellules épithéliales (Gueimonde et Salminen, 2006).
Ainsi, un probiotique ayant un fort pourcentage d’adhésion pourra éventuellement stimuler le
système immunitaire et prévenir l’implantation de pathogènes sur les cellules épithéliales de
l’intestin par des mécanismes de compétition (Saarela et al., 2000). Les modèles in vitro pour
évaluer l’adhésion des probiotiques font appels à des lignées cellulaires provenant de côlons
humains telles que les Caco-2 et HT-29 et à des techniques conventionnelles de détection de
l’attachement bactérien telles que l’énumération par comptages sur plaques, par coloration
Gram, par marquage radioactif ou par de nouvelles approches comme l’ELISA (Enzyme
Linked ImmunoSorbent Assay) et la quantification par PCR en temps réel (Le Blay et al.,
2004; Servin, 2004; Candela et al., 2005). Grâce à ces techniques, plusieurs études ont montré
le potentiel d’adhésion de nombreuses souches de Lactobacillus et de Bifidobacterium
(Chauvière et al., 1992; Bernet et al., 1993; Crociani et al., 1995; Moroni et al., 2006). La
capacité d’adhésion évaluée à l’aide de ces modèles in vitro est différente pour chaque
souche. Ceci est probablement lié à la physiologie et aux facteurs d’adhésion tels que les
composés protéiques, polysaccharides, charges ioniques et aux acides lipotéichoiques propres
à chaque souche bactérienne (Crociani et al., 1995).
Selon Ouwehand et al. (2002b), l’adhésion aux cellules intestinales humaines en culture n’est
pas un critère suffisant pour décrire les interactions mucosales des probiotiques. Ces auteurs
ont proposé que des pièces de tissus humains seraient plus appropriées pour étudier l’adhésion
des probiotiques. Parmi les souches ayant fait l’objet d’études préalables sur Caco-2, L.
rhamnosus GG adhère fortement au tissu provenant du côlon alors que L. johnsonii La1 et L.
casei Shirota le font à un degré moindre. Ces différences d’adhésion sur le tissu peuvent
s’expliquer par la présence de bactéries du microbiote intestinal. Le microbiote résident
pourrait affecter l’adhésion des souches probiotiques au tissu du côlon et ceci n’est
généralement pas pris en compte dans les modèles d’adhésion (Ouwehand et al., 2002b).
Ainsi, certains auteurs soulignent la limitation de certains critères de sélection et estiment
qu’il faut restreindre la surinterprétation de la signification de telles évaluations (Holzapfel et
Schillinger, 2002; Sanders, 2003). Cependant, les tests in vitro et in vivo sont des moyens
utiles et assez simples à mettre en oeuvre pour évaluer de façon préliminaire le potentiel
probiotique de chaque souche avant de procéder à des essais cliniques chez l’humain.
Parmi l’ensemble des critères présentés au Tableau 3, l’aptitude à produire des effets
bénéfiques sur la santé demeure encore délicat à évaluer dû notamment au fait que les modes
d’action par lesquels les probiotiques exercent un rôle fonctionnel in vivo sont méconnus
(Klaenhammer et Kullen, 1999). Ainsi, comme le soulignent ces auteurs, la compréhension

Synthèse Bibliographique
18
des mécanismes d’action représente un des défis scientifiques majeurs dans le domaine des
probiotiques.
Enfin, du point de vue technologique, les souches probiotiques doivent posséder plusieurs
qualités telles que la facilité à être cultivée à de hautes densités cellulaires tout en conservant
leurs propriétés biologiques et leur stabilité au cours des procédés de production et
d’entreposage (Champagne et al., 2005). À ce titre, de nouvelles technologies permettant de
produire des souches probiotiques à haute viabilité et fonctionnalité sont actuellement
disponibles (Lacroix et Yildirim, 2007).
5. Les différents microorganismes probiotiques en aviculture et leurs caractéristiques
En alimentation animale de nombreux genres bactériens et fongiques incluant Lactobacillus,
Bifidobacterium, Bacillus, Streptococcus, Pediococcus, Enterococcus, Propionibacterium,
Saccharomyces et Aspergillus sont utilisés, comme probiotique (Tannock 1997). En général,
les souches probiotiques sont sélectionnées pour leurs effets bénéfiques sur la santé tout en
assurant leur sécurité d'utilisation (innocuité). L'utilisation commerciale de probiotiques en
élevages industriels des volailles est relativement nouvelle. Comme pour les autres animaux,
l'utilisation des probiotiques s'est développée à la suite des recherches effectuées sur le tractus
gastro-intestinal qui ont permis une meilleure compréhension du rôle de la microflore et de
son importance sur la santé et l'hygiène digestive des animaux (Gaggia, Mattarelli et al.). Les
différentes études réalisées sur des volailles ont montré que les probiotiques exercent des
activités antibactériennes contre diverses bactéries pathogènes notamment celles responsables
d'infection chez les poulets dont Salmonella sp (Van Immerseel, Cauwerts et al. 2002;
Higgins, Higgins et al. 2008), Campylobacter et Escherichia coli (Zacconi, Scolari et al.
1999; Ragione, Narbad et al. 2004). L'administration de Lactobacillus salivarius A23 à des
poussins a permis d'augmenter le taux des Lactobacilles dans le jabot dès le premier jour
d'administration. Par contre, aucune augmentation significative n'a été observée au niveau du
caecum. Ceci signifie que ce probiotique colonise préférentiellement le jabot (Zacconi,
Scolari et al. 1999). L'administration d'une préparation brut de microflore caecale a permis de
protéger les animaux contre des infections à Salmonella typhimunium et à Salmonella
enteritidis (Andreatti, da Silva et al. 2000; Higgins, Higgins et al. 2008). Enterococcus
faecium J96, une souche probiotique isolée de l'intestin d'une poule a permis de réduire le taux
de croissance de Salmonella pullorum, gallinarum, typhimurim et enteritidis in vitro.

Synthèse Bibliographique
19
L'administration orale de 109
UFC de cette souche à des poussins de 30 h d'âge leur a permis
de résister à une infection à Salmonella pullorum (Audisio, Oliver et al. 2000).
Dans le même ordre d'idée, l'administration simultanée de Salmonella enteritidis et
Lactobacillus salivarius CTC2197 par voie orale à des poussins d'un jour a permis une
élimination complète des Salmonelles (Zacconi, Scolari et al. 1999).
5. 1. Le genre Lactobacillus
Lactobacillus est le genre principal de la famille des Lactobacillaceae, il contient de
nombreuses espèces qui sont des agents de fermentation lactique intervenant dans de
nombreuses industries. Il s’agit de bacilles longs et fins (parfois incurvés) souvent groupés en
chaînes, immobiles, asporulés, catalase négative, se développent à un optimum de température
situé entre 30 et 40°C. Les lactobacilles ont des exigences nutritionnelles très complexes en
acides aminés, en vitamines, en acides gras, en nucléotides, en glucides et en minéraux
(Khalid et Marth, 1990 ; Leclerc et al., 1994).
Le genre Lactobacillus a été subdivisé par Orla-Jensen en trois groupes et cette classification
est encore utilisée en milieu industriel (Tamime, 2002 ; Guiraud et Rosec, 2004) :
Groupe I « Thermobacterium » : comprend les lactobacilles homofermentaires thermophiles
qui se développent à 45°C mais pas à 15°C. Les espèces les plus fréquentes dans
l’alimentation (lait, yaourt, fromage) sont Lb. helveticus, Lb. delbrueckii, Lb. acidophilus.
Groupe II « Streptobacterium » : regroupe les lactobacilles homofermentaires mésophiles et
peuvent être occasionnellement hétérofermentaires en fonction du substrat. Les espèces les
plus fréquentes dans l’alimentation sont Lb. casei, Lb. curvatus, Lb. sake et Lb. plantarum.
Groupe III « Betabacterium » : ce sont des lactobacilles hétérofermentaires. Il comporte les
espèces Lb. fermentum, Lb. brevis et Lb. sanfransisco.
5.2. Le genre Streptococcus
Le genre Streptococcus est toujours large et la classification est très mouvementée. Ce genre
est généralement divisé en trois groupes : pyogène (la plus part des espèces pathogènes et
hémolytiques), oral (tel que St. salivarius, St. bovis) et les autres streptocoques (Scheilfer,
1987). La seule espèce de streptocoques qui soit utilisée en technologie alimentaire est
Streptococcus thermophilus qui a été inclue dans le groupe des « autres streptocoques», mais
ensuite transféré au groupe des streptocoques oraux à cause de leur degré d’homologie avec
l’ADN de Streptococcus salivarius (Stiles et Holzapfel, 1997).

Synthèse Bibliographique
20
Streptococcus thermophilus se différencie par son habitat (lait et produits laitiers) et son
caractère non pathogène. La résistance à la température, la capacité de croitre à 52°C et le
nombre limité des hydrates de carbones permettent de distinguer les St. thermophilus de la
plupart des autres streptocoques (Haddie, 1986 ; Pilet et al ., 2005).
5.3. Le genre Enterococcus
Ce genre regroupe les streptocoques fécaux qui représentent une hémolyse de type λ et β et
qui appartiennent au groupe D. Ce sont des commensaux de l’intestin. Les espèces
rencontrées dans l’alimentation sont essentiellement En. faecalis et les espèces proches. Les
entérocoques sont des coques qui peuvent être mobiles, homofermentaires, généralement
différenciés par la fermentation de l’arabinose et le sorbitol, ils croissent entre 10°C et 45°C
(Tamime, 2002 ; Ho et al., 2007).
5.4. Les genres Leuconostoc, Oenococcus et Weissella
Ils ressemblent les coques lenticulaires en paires ou en chainettes mésophiles, qui possèdent
un caractère hétérofermentaire marqué, avec production d’acide lactique (isomère D), de CO2
et d’éthanol. Les caractéristiques telles que l’hydrolyse de l’esculine, la formation de
dextrane, les conditions de croissance, la capacité à croître à différents pH et température,
l’assimilation de citrate et/ou malate permettent la différenciation entre les genres
Leuconostoc et Weissella (Pilet et al., 1998 ; Ho et al., 2007).
Actuellement, le genre Leuconostoc comprend quatorze espèces, ils sont également
anaérobies facultatifs et exigeants au point de vue nutritionnel et leur croissance est toujours
lente. Le développement des leuconostoc entraîne souvent l’apparition d’une viscosité dans le
milieu grâce à la production des exopolysaccharides. Les leuconostoc principalement Ln.
mesenteroides ssp. cremoris et Ln. lactis sont utilisés en association avec les lactocoques dans
l’industrie laitière pour produire en plus de l’acide lactique et le CO2, des substances
aromatiques telles que le diacétyle et l’acétoïne à partir des citrates du lait (Hassan et Frank,
2001 ; Guiraud, 2003 ; Ogier et al., 2008).
L’espèce Leuconostoc oenos isolée de vins a été classée dans un nouveau genre, Oenococcus
oeni et certaines espèces de lactobacilles hétérofermentaires ont été groupées avec
Leuconostoc paramesenteroides dans le nouveau genre Weissella (Stiles et Holzapfel, 1997).

Synthèse Bibliographique
21
5.5. Les genres Pediococcus
Les Pediococcus sont des coques homofermentaires dont la particularité est le regroupement
en tétrade. Ils sont mésophiles, le plus souvent incapable d’utiliser le lactose, et leur
développement nécessite la présence de divers facteurs de croissance. Certaines espèces se
distinguent par leur capacité à se développer à des teneurs en sels très élevées, comme
Pediococcus halophilus, renommé Tetragenococcus halophilus et Tetragenococcus
muriaticus qui tolère jusqu’à 18% de NaCl (Pilet et al., 2005).
Les espèces de Tetragenococcus ont un rôle crucial dans la fabrication des produits
alimentaires à concentration élevée en sel comme les sauces de soja, alors que les
pediocoques sont parfois utilisés comme levains lactiques pour les charcuteries (Guiraud et
Rosec, 2004 ; Tosukhowong et al., 2005).
5.6. Le genre Bifidobacterium
Le genre Bifidobacterium est considéré comme faisant partie du groupe des bactéries
lactiques grâce à la similarité de ses propriétés physiologiques et biochimiques et à sa
présence dans le même habitat écologique, tel que le tube gastro-intestinal. Ces
microorganismes sont phylogénétiquement sans rapport avec ces dernières. Ils sont davantage
liés au phylum Actinobacteria (anciennement Actinomycètes) des bactéries Gram positif dont
l’ADN est à haut pourcentage de G +C. Les bifidobactéries se caractérisent par leur forme très
irrégulière souvent en forme V mais pouvant être coccoïdes, la présence d'une enzyme, la
fructose-6-phosphate phosphocétolase, celle-ci leur permet de fermenter les hexoses en
produisant de l'acide acétique et de l'acide lactique. Leur température de croissance varie de
36°C à 43°C (Axelsson et al., 2004 ; Pilet et al., 2005 ; Ho et al., 2007).
5.7. Les levures :
Les levures sont des champignons chez lesquels la forme unicellulaire est prédominante. Les
cellules végétatives peuvent être sphériques, ovoïdes, allongées, cylindriques, apiculées,
ogivales ou en forme de citron. La taille cellulaire varie de 2-3 μm de long à 20-50 μm. la
largeur des cellules est de 1 à 10 μm. Le mode de reproduction végétative le plus courant chez
les levures est le bourgeonnement.
Depuis de nombreuses années, les levures sont également utilisées en additifs alimentaires
chez les animaux pour améliorer les performances zootechniques et comme régulateur de la
flore intestinale chez l’homme. Ils induisent des effets positifs en termes de performances de

Synthèse Bibliographique
22
productions chez plusieurs espèces des ruminants et monogastriques, mais ne peuvent pas
coloniser le tractus digestif.
Les levures utilisées comme probiotiques sont des souches de Saccharomyces cerevisiae. Une
souche bien déterminée de cette levure est dénommée Saccharomyces boulardii (Rolfe, 2000;
Toma et al, 2005).

Synthèse Bibliographique
23
Tableau 4 : Exemples d’effets probiotiques récemment démontrés en élevage avicole (adapté
de Bernardeau et al., 2006)
Animal Souche probiotique Commentaires Référence
Dindes Lactobacillus sp. Augmente le gain de poids. Diminue
Les couts de production
Torres- Rodriguez
et al,2007
Pou
lets
de
chai
r
2Lb., 1 bifidobacterium
Enterococcus, Pediococcus
Augmente les parameters de performance
zootechniques. Module la composition de la
microflora du caecum
Mountzouris et al.,
2007
Lb-based probiotic
Effets sur l’immunité locale démontré par
(1) une diminution des taux d’invasion
intestinale et du développement d’oocytes
d’Eimeria acervulina (EA), (2) des taux
supérieurs de sécrétion d’IL-2 et
diminution de la production d’oocytes d’EA
Dalloul et al.,2003
Pediococcus acidilactici et
Saccharomyces boulardii
Améliore la résistance aux coccidioses
(Eimeria acervulina, E. tenella) en
augmentant l’immunité Humorale
Lee et al., 2007
Enterococcus faecium
NCIMB10415
B.subtilis, B.Licheniformis
Augmente le gain de poids, le taux de
conversion, la taille des villosités dans
l’ileum.
Samli et al.,2007
Bacillus subtilis & Bacillus
Licheniformis
Pas d’impact sur les performances de
croissance, le poids du tibiotarsi, sa
longueur, sa robustesse et son % de Ca.
Améliore l’épaisseur la paroi du tibia
median et latéral, de l’index tibiotarsal et du
% de cendre
Mutus et al.,2006
Lactobacillus johnsonii
F19785
Contrôle les entérites nécrotiques
endémiques dues à Clostridium perfringens,
réduisant les pertes économiques et
l’utilisation d’antibiotiques
La Ragione et al.,
2004
Lb. espèces Inhibe Eimeria tenella – in vitro Tierney et al.,2004
Poules
pondeu
-ses,
fin de
période
Lb. espèces
Augmente la production d’œufs, diminue la
mortalité, améliore le taux de conversion
mais pas la qualité des œufs.
Yoruk et al.,2004

Synthèse Bibliographique
24
6. Production de substances antimicrobiennes
C’est un mécanisme d’action des probiotiques concernant l’inhibition de la croissance des
pathogènes grâce à des composés antimicrobiens. Les bactéries appartenant aux genres
Lactobacillus et Bifidobacterium d’origine humaine peuvent produire des substances
antimicrobiennes, telles que les acides organiques, qui sont actives in vitro et in vivo contre
les micro-organismes entérovirulents impliqués dans les cas de diarrhées (Servin, 2004).
L’acide lactique et acétique sont produits via la fermentation des hexoses par les lactobacilles
et bifidobactéries. Ces acides organiques peuvent diffuser passivement à travers la membrane
bactérienne sous leur forme non dissociée (Figure 1.6). Ils acidifient le cytoplasme après
dissociation et inhibent l’activité enzymatique cellulaire des pathogènes acido-sensibles
(Deng et al., 1999). Cette diminution du pH peut donc affecter la viabilité des pathogènes
bactériens (Bruno et Shah, 2002; Servin, 2004).
Dans une étude réalisée in vitro, Hütt et al. (2006) ont observé que l’activité anti-Salmonella
produit par trois souches de lactobacilles, L. paracasei 8700:2, L. plantarum 299V et L.
fermentum ME-3, en condition microaérophile de même que l’activité anti-Shigella de deux
bifidobactéries, B. lactis Bb12 et B. longum 46, en condition anaérobie était associée à un
effet du pH. Une corrélation positive entre la production d’acide lactique et l’activité
inhibitrice a été observée. Cependant, la quantité d’acide acétique et l’activité inhibitrice des
lactobacilles et des bifidobactéries cultivées en conditions anaérobiques étaient corrélées de
façon négative.
En utilisant un modèle murin, Asahara et al. (2001) ont observé qu’une infection à
salmonelles induite lors d’un traitement aux antibiotiques pouvait être réduite de façon
importante par la précolonisation de l’intestin avec la souche B. breve Yakult. Selon ces
auteurs, la production d’acides organiques par la souche probiotique et une réduction du pH
fécal seraient importantes pour l’activité anti-infectieuse obtenue contre la souche Salmonella
typhimurium. En utilisant la même souche, ces mêmes auteurs ont observé que la production
d’une forte concentration d’acides organiques peut également jouer un rôle important dans
l’inhibition de la production de toxines par une souche de E. coli O157:H7 dans un modèle
murin (Asahara et al., 2004).
La production d’autres agents antimicrobiens, tels que les bactériocines, ont aussi été
rapportés principalement lors d’études in vitro. Les bactériocines sont définies comme des
composés protéiques ayant une activité inhibitrice contre un large spectre de souches
bactériennes (Klaenhammer, 1993). Elles agissent principalement sur la membrane externe
des bactéries cibles en formant des pores qui mènent à la libération du contenu intracellulaire

Synthèse Bibliographique
25
et à la mort de la bactérie affectée (Klaenhammer, 1993). Les lactobacilles sont souvent
associés à la production de bactériocines (Fooks et Gibson, 2002). La production de
bactériocines par les souches de bifidobactéries est moins documentée. Néanmoins, il a été
rapporté que la souche B. bifidum NCFB 1454 produit une bactériocine active contre Listeria
monocytogenes (Yildirim et Johnson, 1998).
Un composé antimicrobien produit par des bifidobactéries humaines, Bifidobacterium spp
CA1 et F9, a été identifié récemment et il consistait en une ou plusieurs molécules lipophiles
avec une masse moléculaire de moins de 3,500 Da (Liévin et al., 2000). Ces auteurs ont
démontré que ces composés antimicrobiens étaient actifs contre Salmonella typhimurium
SL1334 et qu’une diminution de 4 logarithmes de l’adhésion du pathogène aux cellules Caco-
2 pouvait être obtenue. Ces auteurs ont également évalué cette activité antimicrobienne in
vivo avec des souris axéniques C3/He/Oujco infectées par S. typhimurium C5. Les souris
recevant les bifidobactéries CA1 et F9 étaient protégées de l’infection létale comparativement
aux souris contrôles qui ne vivaient pas plus de 10 jours après l’infection. Cependant, même si
certaines bactériocines présentent une forte activité in vitro et in vivo chez les souris contre
certains pathogènes, la démonstration de la production et l’activité des bactériocines dans le
tube digestif humain est à établir.
Lors d’expérience in vitro, certains auteurs ont proposé une action synergétique entre les
substances protéiques antimicrobiennes et l’acide organique pour expliquer l’action inhibitrice
des bactéries probiotiques. Ainsi, Gopal et al. (2001) ont mené une série d’expériences pour
étudier l’inhibition in vitro d’une souche d’E. coli O157:H7 par L. rhamnosus DR20, L.
acidophilus HN017 et B. lactis DR10. Le pré-traitement d’E. coli O157:H7 avec les
surnageants de culture des bactéries réduit l’association de ce pathogène avec les cellules
Caco-2 et HT-29. Les molécules inhibitrices sécrétées dans les surnageants par ces bactéries
sont partiellement affectées par des traitements avec la lactate dehydrogénase, trypsine et
protéinase K laissant suggérer que les substances protéiques antibactériennes ne sont pas
uniquement responsable de l’inhibition et qu’elles agissent de concert avec l’acide produit.
Selon Servin et al. (2004) ceci s’expliquerait par une diminution du pH qui provoque une
perméabilisation de la membrane externe des bactéries Gram-négatif facilitant ainsi la
pénétration d’autres substances antimicrobiennes telles que les bactériocines.

Synthèse Bibliographique
26
7. Efficacités des probiotique en aviculture
7.1. Efficacité sanitaire des probiotiques
Leur efficacité première se situe au niveau de l’aspect sanitaire. les probiotiques exercent des
activités antibactériennes contre diverses bactéries pathogènes et notamment contre les
microorganismes fréquemment responsable d’infection chez les poulets : Salmonella sp,
Compylobacter, Escherichia coli. (Van immerseel et al, 2002 ; Van immerseel et al, 2005).
De nombreuses expériences confirment les effets des souches probiotiques, notamment les
lactobacilllus contre les souches d’ Escherichia coli et Salmonella :
· L’administration de Lactobacillus salivarius A23 à des poussins nouvellement
éclospermet d’augmenter le poids et de diminuer le taux des pathogènes (coliformes)
et augmenter le taux des lactobacilles dans le jabot dès le premier jour
d’administration. Par contre, aucune diminution significative n’a été observée au
niveau du cæcum. Ceci signifie que le probiotique agit essentiellement au niveau de
jabot (Zacconi et al ,1999).
· L’administration de La microflore cæcale permet de protéger les animaux contre des
infections par des souches de Salmonella Typhimurium et S.Enteritidis (Andreatti
Filho et al, 2000).
· D’autres bactéries que les lactobacilles ont un effet probiotique .Tel est le cas
d’Enterococcus faecium souche J96 isolé de l’intestin d’une poule. Cette souche réduit
le taux de croissance de Salmonella pullorum, gallinarum, typhimurium et enteritidis
in vitro. L’administration de 109 UFC de cette souche à des poussins de 30 h leurs
permet de survivre à un challenge 24 h plus tard avec 105 UFC de Salmonella
Pullorum (Audisio et al, 2000 cité par Van immerseel, 2003).
· Il y a également des rapports concernant l’emploi de mélanges de différentes souches
Lactobacillus Salivarius et Lactobacillus Plantarum inhibent in vitro Escherichia coli
et Salmonella Typhimurium (Murry et al, 2004). Ainsi il a été rapporté récemment que
la croissance de Salmonella Enteritidis était fortement réduite in vitro en présence
d’un mélange des Lactobacillus Crispatus et de Clostridium Lactatifermantans à pH
5.8 (Van Der Wielden et al, 2002). En revanche, l’administration simultanée de
Salmonella Enteritidis et Lactobacillus salivarius souche CTC2197 par voie orale à
des poussins d’un jour a permis une élimination complète des Salmonelles après 21
jours (Pascual et al, 1999).

Synthèse Bibliographique
27
· L.salivarius additionné au suspension fécale affecte positivement le poids des poussins
et l’exclusion compétitive des Salmonelles (Zacconi et al, 1999). De la même façon
une suspension feacale permet de protéger les poussins contre une colonisation par les
souches : Salmonella typhimurium, S. agona, S. infantis, S. enteritidis (Oliveira et al,
2000 ; Denis et al, 2004).
Ces expériences montrent qu’il serait possible de réaliser, dés l’éclosion chez des poussins,
une colonisation dirigée du tube digestif des animaux avec des souches probiotiques à fort
pouvoir inhibiteur plutôt que de laisser s’installer naturellement une flore lactique quelconque
apportée par l’environnement. Il est évident que la microflore complexe du cæcum d’un
adulte exerce une action protectrice contre la colonisation des bactéries pathogènes de type E
coli, Salmonella et Campylobacter. Par contre, chez les poussins l’infection par des bactéries
pathogènes est beaucoup plus fréquente du faite que la flore intestinale n’est pas
complètement établie. De plus, les poussins étant séparés de leur mère dès leur éclosion, ils
n’ont pas la possibilité d’acquérir la microflore protectrice maternelle. Tout ceci met l’accent
sur l’intérêt d’utiliser des probiotiques en aviculture.
7.2. Efficacité zootechnique des probiotiques
Chez l’animal, l’efficacité zootechnique revendiquée des probiotiques est souvent par
l’amélioration de la croissance (GMQ), de l’indice de consommation (IC), et de l’état
sanitaire voire du bien être des animaux établis par la réduction de la fréquence des diarrhées
ou de la mortalité durant certaines phases critiques d’élevage: stress alimentaires (changement
de régime alimentaire, rations riches en concentré), stress sanitaires (densité des animaux...).
En matière de productivité, les données publiées font apparaître une variabilité importante de
la réponse animale pour le GMQ et pour l’IC, la réponse relative étant d’autant plus marquée
que les conditions nutritionnelles et sanitaires sont médiocres (Edens, 2003).
Une telle variabilité en pratique n’est pas surprenante car l’action supposée passe par la
modification de l’écosystème intestinal qui peut largement différer d’un essai à l’autre en
fonction des microorganismes utilisés (souches) ainsi qu’à leur concentration dans l’aliment,
de l’interaction des probiotiques avec certains composants de l’aliment, de l’âge des animaux
(les plus jeunes présentant des flores digestives moins stables que celle des adultes et une
immunité moins établie), et de leur état nutritionnel et sanitaire.
· L’administration d’une souche d’Enterococcus faecium M-74, à des poussins durant
06 semaines améliore les performances zootechniques des animaux par rapport au

Synthèse Bibliographique
28
groupe témoin : le poids final est de 2168.25 g et un IC est de 2.02 pour le lot traité
contre 1956.10 g et 2.16 pour le lot temoin (P<0.01) (Ivanković et al, 1999).
· L’addition d’un probiotique, à base d’Enterococcus faecium M-74, à l’eau de boisson
(3g/100l) des poussins durant 06 semaines améliore la croissance des animaux de
10.8% par rapport au lot temoin (Kralik et al, 2004).
· Yeo et Kim, (1997) ont étudié sur des poussins les effets zootechniques d’une souche
de Lactobacillus casei : gain de poids, indice de consommation, activité d’uréase
intestinal. La ration des poussins est supplémentée avec la souche de Lactobacillus
casei, un antibiotique, extrait de yucca, ou n’est pas de tout supplémentée (lot témoin).
Les résultas montrent que l’addition d’un probiotique favorise l’amélioration de gain
moyen quotidien durant les 3 premières semaines avec diminution de taux d’uréase
intestinales comparativement aux autres lots.
· D’autres paramètres nutritionnels tel que l’activité des enzymes amylases sont
également améliorés en présence des Lactobacillus. (Jin et al, 2000).
· Un essai de supplémentation par la levure Saccharomyces cerevisiae a été conduit sur
des poussins (4.108 UFC), la mortalité a été significativement diminuée dans le lot
traité (Karaoglu et Dardug, 2005).
· Paramètres environnementaux : Dans l’élevage intensif, la principale préoccupation
environnementale concerne les déjections. l’emploi d’additifs probiotiques permet de
réduire la quantité d’azote dans les effluent, ce qui pourrait représenter un gain
d’efficacité alimentaire, à condition toutefois que l’énergie ainsi épargnée soit rendue
disponible à l’animal. Ainsi son importance, d’un point de vue environnemental ;
(Applegate et Angel, 2005 ; Wood et Abuchar, 1998 ; Rotz, 2004; Ferket et al, 2002 ;
Lee et al, 2006). Selon Chang et Chen, (2003) la présence des souches lactobacilli
dans l’aliment réduit l’excrétion d’ammoniac.
· L’addition de jus de rumen lyophilisé augmente le poids des poulets de chair et
améliore la conversion (Kuçukersan et al, 2002).

Synthèse Bibliographique
29
Chapitre 3 : Les prébiotiques
1. Définition Prébiotiques
Par définition les prébiotiques sont des ingrédients des aliments indigestibles, qui ont un effet
bénéfique sur l’animal par le biais d’une stimulation de la croissance et/ou de l’activité d’un
nombre restreint d’espèces bactériennes déjà résidente dans la flore digestive de l’animal ce
qui peut contribuer à l’amélioration de la santé de l’animal (Gibson et Roberfroi, 1995; Piva,
1999; Schrezenmeir et De Vreseal, 2001; Rastall et Gibson, 2004 ; Cummings et Kong, 2004 ;
Fao/Who, 2004). Par conséquent, un produit sera classé comme prébiotique dés qu’il répond
aux trois conditions suivantes : (Suskovic et al, 2001 ; Ferket, 2002 ; Fooks et Gibson, 2002 ;
Gibson et al, 2004).
-être ni hydrolysé, ni absorbé dans le tractus gastro-intestinal.
- être sélectif pour un nombre limité de bactéries endogènes.
-Modifier la microflore intestinale en améliorant sa composition.
-Tout ceci doit nécessairement induire une modification de la composition de la flore,
améliorant ainsi l’état de la santé de l’hôte.
Compte tenu de ces critères particuliers, on constate que la plupart des prébiotiques sont des
hydrates de carbones non digestibles pour l’hôte.
2. Différentes classes des prébiotiques
On distingue différentes classes de prébiotiques, selon la taille de la molécule ou suivant leur
origine, naturelle ou synthétique (Van immerseel al, 2003 ; Gibson et al, 2004).
2.1. Les hexoses, telles que le fructose, glucose, galactose et mannose, et les pentoses tels que
le ribose, xylose et arabinose sont les monosaccharides prébiotiques les plus importants. Le
galactose est disponible sous forme de disaccharides tels que le lactose. Cependant le
monosaccharide le plus couramment utilisé comme prébiotique est certainement le mannose.
2.2. Les disaccharides naturels
Les plus couramment utilisés sont le sucrose, le lactose et le maltose.
2.3. Les oligosaccharides (Conway, 2001)
Sont produits la plupart du temps par synthèse ou par hydrolyse enzymatique, soit à partir des
hexoses monosaccharidiques, soit à partir de la paroi de cellules microbiennes ou par
fermentation de polysaccharides. Parmi les oligosaccharides, les fructo-oligosaccharides
(FOS) occupent certainement une place importante.

Synthèse Bibliographique
30
Les FOS sont produits par hydrolyse d’inuline ou par synthèse à partir de sucrose ou de
lactose. Les FOS réduisent la colonisation de l’intestin par Salmonella. L’administration de
FOS dans les aliments pour volaille semble également réduire la colonisation de l’intestin par
Campylobacter et les salmonelles (Gibson et Fuller, 2000 ; Van immerseel et al, 2003).
De la même façon Oyarzabal et ses collaborateurs (1995) ont mis en évidence l’efficacité
d’une préparation probiotique associant un E.laecium, L. lactis, ar Pediococcus sp, avec FOS
; l’incorporation permet l’exclusion des salmonelles a partir du ceacum.
Les mannane-oligosaccharides (MOS) sont des constituants naturels de la paroi des levures et
des gommes naturelles. Ce produit est constitué d’un lysat centrifugé de Saccharomyces
cerevisiae. L’administration de ces MOS protège la volaille contre plusieurs pathogènes
provoquant des troubles digestifs en stimulant le système immunitaire, modifiant la flore
intestinale et inactivant les aflatoxines (Anonyme, 2002; Revington, 2002).
3. Mode d’action des prébiotiques
Les prébiotiques agissent en amont des probiotiques. Où le probiotique va fournir directement
un micro-organisme aux actions bénéfiques pour l’hôte, le prébiotique se contente d’apporter
une source nutritive sélective d’une flore bénéfique pour l’hôte. Le mode d’action des
prébiotiques est donc à rapprocher de celui des probiotiques.
Les prébiotiques sont généralement des oligosaccharides ou des polysaccharides à courte
chaîne constitués approximativement de deux à vingt unités de sucre. Ils échappent à la
digestion dans l’intestin grêle et sont des substrats potentiels pour l'hydrolyse et la
fermentation par les bactéries intestinales.
Les prébiotiques doivent agir comme substrat sélectif d’une ou d’un nombre restreint de
souches bactériennes bénéfiques qui résident dans le côlon et en stimuler la croissance. Les
bifidobactéries et les lactobacilles sont les micro-organismes du microbiote intestinal les plus
fréquemment ciblés (Marteau et al., 2004).
De plus en plus utilisé en thérapie (Fujimori et al., 2007 ; Larkin et al., 2007), les prébiotiques
sont également grandement utilisés dans les alicaments et dans l’alimentation animale.
Les FOS sont des composants naturels, que nous retrouvons dans divers végétaux (oignon ou
blé par exemple), qui ne sont pas digérés par les Mammifères mais qui peuvent être
métabolisés par certaines bactéries (Willard et al. 1994a). Ils permettraient d’obtenir une
réduction quantitative de la flore intestinale pathogène en étant à l’origine d’un
environnement plus favorable pour les bactéries utiles qui se développent plus vite que les
bactéries pathogènes qui sont incapables d’utiliser les FOS (Lecoindre 2000).

Synthèse Bibliographique
31
4. Caractérisation des exopolysaccharides (prébiotiques)
4.1. Classification des EPS des bactéries lactiques
Les bactéries lactiques produisent deux principales classes d'EPS. Les homopolysaccharides
et les hétéropolysaccharides.
Les homopolysaccharides sont constitués d'un seul type de monosaccharide joint par
différents types de liaisons et d'embranchements. Ils comprennent, entre autres, les a-D-
glucanes, produits par Leuconostoc mesenteroides, Streptococcus mutans et Streptococcus
sobrinus; les D-glucanes, produits par Propionibacterium spp. (Deutsch et al. , 2008),
Pediococcus spp. Et Streptococcus spp., ainsi que le levane produit par Streptococcus
salivarius (De Vuyst et Degeest, 1999).
Pour leur part, les hétéropolysaccharides sont des répétitions de plusieurs oligosaccharides,
chacun contenant de trois à huit résidus et possédant un nombre limité de monosaccharides
différents (Degeest et al., 2001a). Des résidus acétyle, amino ou phosphate peuvent également
être retrouvés dans les unités répétitives. Les hétéropolysaccharides sont produits par des
LAB mésophiles, comme Lactococcus lactis ssp. lactis, L. lactis ssp. cremoris, Lactobacillus
casei et L. rhamnosus, et par des LAB thermophiles, notamment Lactobacillus acidophilus,
Lactobacillus helveticus et S. thermophilus (De Vuyst et Degeest, 1999). La structure des
hétéropolysaccharides peut varier, entre les espèces et entre les souches, selon la composition
en sucres, la présence et la nature des chaînes latérales, la masse moléculaire, etc.
Les différentes structures possibles des EPS affectent le pouvoir texturant de ces
macromolécules dans les produits alimentaires. Étant donné que les EPS varient dans leur
composition, leur arrangement spatial, leur charge, leur rigidité et leurs interactions avec les
protéines, la corrélation entre la concentration des EPS et la viscosité apparente est
habituellement spécifique à chaque EPS. Malgré tout, certaines caractéristiques semblent
communes à plusieurs EPS. Par exemple, les EPS ayant une masse molaire plus élevée
entraîneront habituellement une plus grande viscosité. Aussi, les EPS neutres contribuent
surtout à la viscosité, mais pas à l'élasticité, contrairement aux EPS chargés négativement qui
vont plutôt contribuer à l'élasticité, mais pas à la viscosité (Jolly et al., 2002).

Synthèse Bibliographique
32
4.2. Production et détection des EPS
4.2.1. Détection de polysaccharides exo-cellulaires
II existe plusieurs façons de détecter la présence de polysaccharides. Ces derniers peuvent être
détectés qualitativement par un examen visuel, par coloration ou par microscopie photonique
électronique. La mesure de la viscosité et le dosage des EPS purifiés permet de les détecter
quantitativement.
4.2.2. Examen visuel
La façon la plus simple de détecter la production d'EPS chez une souche consiste à examiner
une colonie sur un milieu gélosé. La colonie d'une souche productrice aura une apparence
brillante et en touchant la colonie avec un cure-dents ou avec une anse à ensemencer (manche
de Koch), il y aura formation de longs filaments visqueux. Cependant, certaines souches de
bactéries lactiques ne produisent pas de polymères sur un milieu gélosé mais en produisent en
milieu liquide sous certaines conditions bien définie.
4.2.3. Les colorations
II est possible de sélectionner des clones bactériens producteurs d’EPS, appelés clones Mut+,
en ajoutant des colorants cationiques dans le milieu de culture. Les clones Muc+ sont révélés
par le masquage de la coloration par le polysaccharide produit.
Les colorants cationiques ont une affinité pour les polysaccharides anioniques et neutres ; ils
développent des interactions ioniques avec les anions organiques ou inorganiques du milieu
environnant. Additionné au milieu de croissance, le colorant cationique se fixe donc sur le
peptidoglycane de la paroi bactérienne, ce qui entraîne une coloration de la colonie d'une
souche non-productrice. Ces colorants ne présentant pas d'affinité pour les polysaccharides
excrétés, les polysaccharides produits masquent alors la coloration et les clones Muc+
apparaissent blancs (Gancel et al, 1988).
Un exemple de colorant cationique est le rouge de ruthénium qui donne une coloration rose
foncé au milieu de culture. Les colonies Muc+ apparaissent donc blanches sur fond rose par le
masquage de la couleur dû aux exopolysaccharides produits. Les clones Muc- apparaissent
rosés. Cette technique a été utilisée par Bouzar et al. (1995) pour la sélection de clones d'une
souche de Lb. delbrueckii ssp. bulgaricus produisant différents niveaux de polysaccharides
extracellulaires. De cette façon, Stingele et al. (1996) ont sélectionné la souche Streptococcus
thermophilus Sfi6 possédant le phénotype de production d'EPS stable.

Synthèse Bibliographique
33
4.3. Extraction et purification
La meilleure façon de doser quantitativement les EPS est de les extraire du milieu de culture
et de les doser. Les polysaccharides bactériens extracellulaires précipitent en présence
d'éthanol. Après les avoir purifiés pour enlever les protéines et les résidus du milieu pouvant
interférer lors du dosage, il est possible de les doser avec une méthode calorimétrique comme
celle de Dubois (1956) à l'acide sulfurique et phénol. Lors de ce dosage, la courbe étalon est
produite avec du glucose et les unités de mesure sont exprimées en mg/l d'équivalents
glucose. Cette méthode a aussi ses limites et un milieu défini est recommandé. En effet,
certains éléments présents dans le MRS par exemple peuvent précipiter avec l'éhano1 et
fausser les résultats de dosage (Garcia-Garibay et Marshall,1991). L'extraction des EPS dans
le lait est plus compliquée ; les constituants du lait peuvent interférer dans la précipitation des
polymères et les EPS bactériens ont tendance à s'associer aux protéines du lait pour former un
complexe glycoprotéique. II faut alors hydrolyser les caséines avec des enzymes spécifiques
et bien purifier les polymères recueillis par une précipitation à l'éthanol (Bouzar et al., 1997).
4.4. Dosage colorimétrique des EPS purifiés
Les exopolysaccharides purifiés et lyophilisés sont resuspendus dans de l'eau dé-ionisée
(Millipore) et dosés selon la méthode de dosage des sucres totaux de Dubois et al. (1956)
modifiée par Gerhardt et al. en 1981.
Le principe de cette méthode repose sur le fait que l'ajout d'acide sulfurique provoque
l'hydrolyse des polysaccharides et la formation de 5- hydroxyméthyl-furfural qui donne, en
présence de phénol, une coloration orangée. L'intensité de la couleur est directement
proportionnelle à la quantité de sucre présent et est directement mesurable par
spectrophotométrie. La réaction est sensible et la couleur est stable. De petites quantités de
Carbohydrates peuvent être détectées et la présence de protéines ou de peptides n'influence
pas le dosage.
Une courbe standard est préparée à partir d'une solution glucose à une concentration donnée.
Chaque échantillon est préparé et pré-dilué si nécessaire. Le blanc est fait avec la
concentration O mg/l.
La lecture de l'absorbance se faite avec un spectrophotomètre à une longueur d'onde de 499
nm. Les quantités de polysaccharides sont exprimées en mg/l d'équivalent glucose, selon la
courbe standard.

Synthèse Bibliographique
34
4.5. Caractérisation des EPS à l’échelle chromatographique La chromatographie sur couche mince (CCM) est la technique la plus simple, utilisée pour
déterminer la composition qualitative en oses polysaccharidiques. Cette technique a été
utilisée par Souly Frag (1978).
Avant d’être analysés, les polysaccharides doivent subir une hydrolyse acide afin de libérer
les différents oses composants de ce polysaccharide.
Une plaque de chromatographie sur couche mince (TLC, CCM en français) se compose d’un
support, en aluminium ou en verre, sur lequel a été étendue une fine couche d’un milieu de
sorption (p. ex. la silice, SiO2) comme phase stationnaire. Ces plaques sont plongées
d’environ un demi-cm dans une phase mobile. Cette dernière est généralement un mélange
binaire ou ternaire de solvants, adapté au type de séparation recherché. Les composés déposés
à environ un cm du bas de la plaque sont alors humectés et dissous par la phase mobile qui
progresse par capillarité le long de la phase stationnaire.
Selon la nature des phases mobile et stationnaire, deux types de mécanismes d’interaction
permettent la séparation de composés présents en mélange : l’adsorption sur la surface de la
phase stationnaire solide et le partage entre un film de phase stationnaire liquide et la phase
mobile (Pachaly, 1999).
Pour les analyses de routine par TLC des chromatographies monodimensionnelles, on utilise
généralement des plaques de Silicagel 60 F254 prêtes à l’emploi à support en aluminium
(Merck) et celles-ci sont développées dans des cuves en verre conventionnelles (Camag), dont
l’atmosphère aura préalablement été saturée en vapeurs de la phase mobile.
Du fait de ses faibles contraintes techniques, de son emploi simple et de son coût relativement
modeste, la TLC est un outil de choix pour l’analyse de routine d’extraits bruts, de fractions,
ainsi que de produits purs isolés. Les quantités déposées sur les plaques sont normalement de
100 μg pour les extraits et de 10 μg pour les produits purs.
Il s’agit également d’un support facilement utilisable pour caractériser ultérieurement des
substances par leurs réactivités chimiques ou leurs activités sur certaines cibles biologiques.
L’incorporation dans la phase stationnaire des plaques du commerce de produits permettant la
visualisation des composée UV-actifs à 254 nm et 366 nm, augmente le seuil de détection des
produits à faible activité spectrale dans le domaine du visible (environ 450-700 nm).

Synthèse Bibliographique
35
5. Effet des prébiotiques en aviculture
Les prébiotiques constituent une toute autre classe d’additifs. Par définition, les prébiotiques
sont tous les produits ajoutés aux aliments qui sont indigestibles pour le poulet, mais qui
peuvent avoir un effet bénéficiaire sur la santé intestinale par la stimulation spécifique de la
croissance ou de l’activité d’un nombre limité d’espèces bactériennes favorables. Presque tous
les prébiotiques sont des oligosaccharides ou des polysaccharides. Par exemple, plusieurs
études ont montré un effet bénéficiaire de l’utilisation de lactose pendant la période juste
avant l’abattage. Les fructo-oligosaccharides par contre, malgré leur effet bénéficiaire bien
documenté sur la santé intestinale chez l’homme et les mammifères, ne semblent avoir qu’un
effet marginal, voir inexistant, sur la colonisation de l’intestin du poulet par Salmonella. Les
mannan-oligosaccharides cependant ont bien un effet de protection contre les Salmonelles, ce
que s’explique probablement par le blocage de l’attachement des Salmonella sur les cellules
épithéliales de l’intestin. Les auteurs ont montré un effet de protection contre la colonisation
et l’excrétion des Salmonelles dans les fientes d’un additif sur base d’arabinoxylo-
oligosaccharides (AXOS), ce qui est d’autant plus surprenant que les polymères dont ces
AXOS sont dérivés, et qui sont largement présents dans les céréales, tels que le froment et le
seigle, semblent augmenter la sensibilité des poulets pour une infection à Salmonella. Aussi
pour le « guar gum » et « partially hydrolysed guar gum », des effets de protection contre
Salmonella ont été décrits. Il existe encore toute une gamme d’autres prébiotiques, pour
lesquels cependant souvent, les effets de protection contre la colonisation et l’excrétion dans
les fientes des agents zoonotiques n’ont pas été clairement documentés chez le poulet. Les
probiotiques constituent une troisième grande classe d’additifs testés pour une utilisation dans
le cadre de la lutte contre la salmonellose et les autres agents zoonotiques chez le poulet. Les
probiotiques par définition sont des microbes vivants ajoutés aux aliments. La plupart des
souches microbiennes utilisées dans les produits probiotiques appartiennent aux genres
Bifidobacterium ou Lactobacillus. Les lactobacilles peuvent produire des métabolites qui
limitent la croissance des Salmonelles. Ils peuvent, dans certaines conditions, aussi moduler
l’immunité et interférer dans l’attachement des Salmonelles sur les cellules épithéliales de
l’intestin. Par ce biais, les lactobacilles peuvent protéger contre la colonisation par
Salmonella. Certaines souches de lactobacilles ont déjà été mélangées dans les aliments à la
ferme avec de bons résultats. Cependant, la réduction des nombres de Salmonelles retrouvées
au niveau de l’intestin après administration orale de lactobacilles est souvent limitée (Van
Coillie et al., 2007).

Synthèse Bibliographique
36
Les oligosaccharides sont produits la plupart du temps par synthèse ou par hydrolyse
enzymatique, soit à partir des hexoses monosaccharidiques, soit à partir de la paroi de cellules
microbiennes, ou par fermentation de polysaccharides (Iji et Tivey, 1998). Parmi les
oligosaccharides, les fructo-oligosaccharides (FOS) occupent certainement une place
importante. Les FOS sont constitués de courts polymères de fructose (liés en β 1-2). Ils sont
produits par hydrolyse d’inuline ou par synthèse à partir de sucrose ou de lactose (Le Blay et
al., 1999). On a démontré que les FOS réduisent la colonisation de l’intestin par Salmonella
(Fukata et al., 1999), d’autant qu’une flore de compétition est donnée en même temps (Bailey
et al., 1991). L’administration de FOS dans les aliments pour volaille semble également
réduire la colonisation de l’intestin par Campylobacter (Schoeni et Wong, 1994).
L’administration de ces MOS à une concentration de 4000 ppm dans l’aliment de poussins de
3 jours a entraîné une réduction de la concentration de Salmonelles dans les caeca après
challenge de Salmonella typhimurium et de Salmonella dublin (Spring et al., 2000). On a
démontré également que le contenu cæcal des poules recevant des MOS dans les aliments,
administré à des poussins, protégeait ces poussins contre un challenge avec Salmonella
enteritidis (Fernandez et al., 2000).
Les polysaccharides, peu de données sont disponibles, sauf pour la gomme de guar obtenue
par traitement enzymatique des fèves de Cyanopsis tetragonolobus. Le traitement à l’endo-β-
D-mannanase permet de cliver la chaîne centrale de mannose de la gomme. On obtient ainsi
un mélange de galactomannanes que l’on appelle gomme de guar partiellement hydrolysée.
Inclus dans l’aliment pour poule pondeuse à une concentration de 250 ppm cette gomme de
guar partiellement hydrolysée a permis de protéger les poules contre un challenge assez
sévère de Salmonella enteritidis, et en plus, la coquille des œufs ainsi que le blanc et le jaune
des œufs étaient significativement moins contaminés (Ishihara et al., 2000).

Synthèse Bibliographique
37
Chapitre 4 : APPROCHE MOLECULAIRE
Au cours des dernières décennies, différentes méthodes d'identification moléculaire des
bactéries lactiques ont été élaborées afin de réduire le temps d'analyse généralement
nécessaire aux tests phénotypiques. En outre, parce qu'elles ne sont pas influencées par des
facteurs physiologiques et environnementaux, les méthodes moléculaires offrent des résultats
d'identification plus fiables.
Les méthodes moléculaires permettent une identification au niveau de l'espèce et de la souche.
De plus, les souches de bactéries peuvent être différenciées entre elles et, en outre, les souches
porteuses de gènes spécifiques peuvent être détectées quelle que soit l'espèce.
1. l’ADNr 16S comme sémantide bactérienne
De toutes les sémantides bactériennes, le gène de l’ADNr 16S reste certainement la plus
usitée selon Fox et al. (1980) et Weisburg et al. (1991) qui historiquement en firent la
première sémantide bactérienne et la première facilement amplifiable « universellement ».
Aujourd’hui encore, le gène de l’ADNr 16S reste la sémantide la mieux renseignée dans les
bases de données de séquences nucléotidiques (cf figure I.15) et de facto la plus employée
(près de 600000 séquences dans la base « RDPII » (Cole et al., 2007) et plus de 200000
séquences supérieures à 1250 Nt dans la base « Greengenes » en août 2008 (De Santis et al.,
2006).
L’ARN 16S est un constituant de la petite sous-unité ribosomique. De fait, la survie de la
cellule dépend de sa conformation. Or cette dernière dépend de sa séquence et donc la
séquence du gène de l’ADNr 16S est indispensable à la survie de la cellule. Il existe donc des
zones conservées du gène qui sont indispensables à la structure tertiaire de l’ARNr.
Ces régions conservées sont au nombre de dix. De ces régions peuvent être extraits des
amorces ou sondes dites « universelles ». Entre ces régions conservées existent des régions
dites « hypervariables » dont la séquence n’est pas « indispensable » à la conformation
ribosomique. Ces régions hypervariables sont au nombre de neuf et sont représentées et
annotées de V1 à V9 sur la figure I.17. Ces régions hypervariables offrent des possibilités
d’amorçages spécifiques d’un taxon donné ou, couplé avec deux régions conservées et une
amplification, permettent une identification phylogénétique de la bactérie. L’ensemble de ces
régions et la taille relativement restreinte du gène de l’ARNr 16S (1541 bases chez E. coli)
assurent de nombreuses possibilités de travail aux bactériologistes.

Synthèse Bibliographique
38
Malgré son usage intensif, l’emploi de l’ADNr 16S en tant que sémantide dans les études
phylogénétiques souffre néanmoins de plusieurs revers. Dans de tels cas, on peut alors
avantageusement remplacer le gène de l’ADNr 16S par d’autres sémantides bactériennes.
2. Les méthodes d’identification
2.1. Identification des bactéries par l’ADNr-16S
L’indentification des bactéries par les méthodes moléculaires représente un complément
indispensable aux méthodes traditionnelles d’analyse des caractéristiques morphologiques et
physiologiques.
Parmi ces nouvelles techniques, celle basée sur le séquençage direct apparaît comme la plus
efficace : elle consiste à séquencer « litre » une région spécifique du chromosome bactérien
(ADNr-16S) et à comparer cette séquence avec les séquences connues et stockées en banques
de données. Ceci permet de déterminer avec certitude le genre et l’espèce de la souche
étudiée. L’analyse de séquences génétiques discriminantes offre une technique
complémentaire pour l’identification bactérienne de toutes les situations. Le gène codant
l’ADN 16S est l’outil principalement utilisé pour l’identification moléculaire des bectéries.
Il s’agit d’un gène chromosomique d’une taille de 1.500 paires de bases, présent chez toutes
les espèces bactériennes (gène universel), dont la séquence est spécifique de chaque espèce et
dont les extrémités 5’ et 3’ (15 premières et 15 dernières bases) sont conservées dans toutes
les espèces bactériennes.
Techniquement, l’ADN bactérien est extrait par selon des protocoles bien standardisés, puis le
gène ADNr 16S est amplifié par PCR, incorporant deux amorces universelles
complémentaires des extrémités 5’ et 3’ du gène.
Le produit d’amplification est ensuite séquencé et la séquence est alors lue à l’aide d’un
automate, Elle est ensuite comparée par des logiciels aux séquences homologue contenues
dans les banques informatiques (GenBank, EMBL) via le réseau internet par le BLAST.
Le logiciel fournit un pourcentage d’homologie entre la séquence étudiée et celles contenues
dans les banques de banques de données. Une homologie de 99-100% permet d’affirmer
l’identification de la souche bactérienne.
A défaut, la souche peut être positionnée parmi l’ensemble des espèces connues sous forme
d’un arbre phylogénétique car le gène ADNr 16S possède aussi des propriétés d’horloge
moléculaire.

Synthèse Bibliographique
39
2.2. Identification bactérienne avec amorces oligonucléotidiques (Amplification génique)
C’est une méthode génotypique permettant l’identification de locus spécifiques d’ADN des
bactéries au niveau du genre et l’espèce (Jackson et al., 2004), à l’exemple des gènes de
ménages.
De nombreuses méthodes génotypiques utilisant l’analyse de produits d’amplification génique
issus de gènes cibles ont été développées pour identifier les bactéries au niveau de l’espèce.
Ces techniques incluent le séquençage de l’ADNr 16S qui est souvent considéré comme
méthode de référence. Dans ce dernier cas, cependant, l’analyse des données de séquence peut
être compliquée par le fait que des espèces proches peuvent posséder des séquences d’ADNr
16S identiques ou, au contraire, que des séquence d’ADNr 16S différentes peuvent coexister
dans une bactérie donnée.
Pour résoudre ce problème, il est possible d’utiliser des séquences cibles monocopies qui
possèdent une divergence évolutive supérieure à celle de l’ADNr 16S.
Le gène sodA des cocci à Gram positif, qui code pour une superoxide dismutase Mn-
dépendante, satisfait à ces critères et les auteurs ont pu montrer que le séquençage d’un
fragment interne, sodAint, obtenu par PCR à l’aide d’une paire d’amorces dégénérées ou une
paire d’amorces spécifiques à l’espèce permettait l’identification fiable au niveau du genre ou
de l’espèce des streptocoques et des entérocoques.
Des chercheurs ont utilisé la même paire d’amorces universelles pour construire une
génothèque de fragments sodAint correspondant à toutes les espèces de Staphylocoques à
coagulase-négative (SCN) (Poyart et al.,2001), et ont montré que son utilisation permettait
l’identification rapide et fiable d’isolats cliniques de SCN.
Par ailleurs, l’amplification PCR et l’analyse des sequences génétiques discriminantes des
gènes d’aminopeptidases (pepN, pepO), par l’utilisation d’amorces spécifiques (170/171,
493/494) ont permis l’identification de l’espèce Lactococcus lactis (Bensalah et al.,2009).
3. Amplification par PCR
La PCR, réaction de polymérisation en chaîne (Polymerase Chain Reaction), est une
technique permettant d'obtenir, à partir d'un échantillon d'ADN, d'importantes quantités d'une
séquence d'ADN spécifique. Cette amplification repose sur la réplication d'une matrice
d'ADN double brin. Elle se décompose en trois phases : une phase de dénaturation, une phase
d'hybridation avec des amorces et une phase d'élongation. Les produits de chaque étape de
synthèse servent de matrice pour les étapes suivantes, ainsi on réalise une amplification
exponentielle.

Synthèse Bibliographique
40
· Les différentes étapes de la PCR (Figure 3)
A. La dénaturation :
C'est la séparation des deux brins d'ADN, obtenue par élévation de la température.
B. L’hybridation
En abaissant la température, les amorces spécifiques s'hybrident sur les molécules simples
brin d'ADN. Les amorces sont constituées de courtes séquences d'ADN complémentaires de
la séquence de l'ADN à amplifier. Il s'agit toujours d'un couple d'amorces, complémentaire
encadrant le fragment d'ADN à amplifier.
C. L’élongation :
C'est la synthèse du brin complémentaire. Une enzyme polymérase, la Taq polymérase, ajoute
à l'extrémité de l'amorce des oligonucléotides présents dans le milieu de réaction.
Figure 3: les étapes de déroulement de la PCR

Synthèse Bibliographique
41
4. Séquençage de l’ADN
La caractérisation fine d’un gène passe par son séquençage, c'est-à-dire par la connaissance du
nombre, de la nature et de l’ordre des nucléotides qui le composent. Ces agencement permet
de connaître par exemple l’emplacement des différents sites de restriction du gène afin de
mieux le manipuler.
Enfin, la traduction de la séquence nucléotidique en séquence d’acides aminés permet
éventuellement de confirmer ou de proposer une fonction de la protéine par le gène.
Le principe est le suivant :
Le fragment d’ADN à séquencer (il peut s’agir par exemple d’un exon du gène d’intérêt) est
obtenu par PCR puis mis en présence d’un milieu réactionnel contenant :
· L’amorce à partir de laquelle la synthèse du néobrin sera réalisée par une ADN
polymérase,
· Les 4 désoxynucléotides (dA, dT, dC, dG),
· Les 4 désoxynucléotides (ddA, ddT, ddC, ddG) marqués chacun par un fluorochrome
distinct
· Une ADN polymérase (en général la taq DNA polymérase).
L’ensemble est soumis à une succession de cycles de polymérisation au cours desquels l’ADN
polymérase peut, au niveau de chaque nucléotide de l’ADN matrice, incorporer un
désoxynucléotide ou un didésoxynucléotide.
Dans le cas où elle incorpore un désoxynucléotide, la synthèse peut continuer, dans le cas
contraire elle s’arrête. Ce choix étant aléatoire, chaque base de l’ADN matrice aura
statistiquement vu un certain nombre de fois l’incorporation d’un didésoxynucléotide, si bien
que le milieu réactionnel contient l’ensemble des molécules néosynthétisées possibles. Ces
molécules sont ensuite dénaturées puis migrées dans un gel d’électrophorèse afin d’être
séparées selon leur taille.
On peut ainsi reconstituer la séquence en analysant la nature du fluorochrome terminant
chacun de ces fragments néosynthétisées, du plus petit (premier nucléotide de la matrice) au
plus grand (dernier nucléotide de la matrice).
Cette analyse est réalisée à l’aide d’un séquenceur automatique : cet appareil est pourvu d’une
source laser ou infra-rouge qui excite les fluorochromes portés par les didésoxynucléotides
après que les fragments aient migrés sur le gel d’électrophorèse. Cette excitation provoque
une émission de lumière à une longueur d’onde dépendant de la nature du fluorochrome.

Synthèse Bibliographique
42
5. Taxonomie des bactéries lactiques
Depuis la description du Bacterium lactis (actuellement Lactococcus lactis), la taxonomie des
bactéries lactiques est en évolution permanente. Le nombre de nouvelles espèces a augmenté
énormément au cours de ces dix dernières années. Les réorganisations effectuées ont
contribué à fusionner des espèces en une seule, ou identifier une espèce comme un nouveau
genre (Pot, 2008).
La classification des bactéries lactiques peut se faire selon des critères phylogénétiques par
l’utilisation des méthodes moléculaires. Cependant, la caractérisation phénotypique/
biochimique classique demeure pratique dans l’identification préliminaire des
microorganismes. Certaines caractéristiques phénotypiques sont utilisées pour identifier les
espèces à l'intérieur des genres comme la capacité à : fermenter les hydrates de carbone,
tolérer différentes concentrations en bile, produire des polysaccharides extracellulaires, exiger
des facteurs de croissance, produire de l’acétoïne et synthétiser certaines enzymes. La
composition en G+C de l’ADN, la composition en acides gras, la mobilité électrophorétique
de la lactate déshydrogénase sont également d'autres critères qui peuvent être étudiés pour
l'identification des espèces lactiques (Vandamme, 1996 ; Stiles et Holzopfel, 1997 ; Ho et al.,
2007).
La morphologie est considérée comme la caractéristique clé pour décrire et classifier les
genres des bactéries lactiques. De ce fait, les bactéries lactiques peuvent être divisées
arbitrairement en bacilles (Lactobacillus et Carnobacterium) et coques (tous les autres
genres). Le genre Weissella, récemment décrit, est le seul genre qui comporte à la fois des
bacilles et des coques (Collins et al., 1993 ; Ho et al., 2007).
A ce groupe de bactéries lactiques, appartient plusieurs genres comme Aerococcus,
Atopobium, Carnobacterium, Enterococcus, Lactobacillus, Lactococcus, Leuconostoc,
Oenococcus, Pediococcus, Streptococcus, Tetragenococcus, Vagococcus et Weissella. (Stiles
et Holzapfel, 1997 ; Pot, 2008). Des genres nouveaux, par exemple Alloiococcus,
Dolosicoccus, Dolosigranulum, Eremococcus, Facklamia, Globicatella, Helococcus,
Ignavigranum et Lactosphaera, ont également été décrits, comportant des souches qui
montrent des liens physiologiques et phylogénétiques avec les groupe des bactéries lactiques
(Broadbent, 2001 ; Axelsson, 2004).
Le genre Bifidobacterium est actuellement considéré par plusieurs auteurs comme genre de
bactéries lactiques, bien qu’il se distingue par un pourcentage en G+C de 55%, largement
supérieur à celui des autres genres et par une voie métabolique de fermentation des sucres
particulière. Les études phylogénétiques basées sur l’analyse des séquences des ARN

Synthèse Bibliographique
43
ribosomiques ont confirmé l’appartenance de ces différents genres à un même groupe qui
inclut également Clostridium, Bacillus et Propionibacterium (Figure 4) (Stiles et Holzopfel,
1997 ; Pilet et al., 2005).
Figure 4 : Arbre phylogénétique des principaux genres de bactéries lactiques et des genres associés, obtenu par analyse des ARNr 16S (Stiles et Holzapfel, 1997).

Matériels et Méthodes
44
1.Échantillonnage
Deux coqs fermiers ont été sacrifiés et leur tube digestif transporté au laboratoire dans des conditions stériles à froid et disséqués en zone aseptique.
Figure 5 : photo d’un tube digestif réalisée au laboratoire.
2. Isolement et purification des souches
Des prélèvements ont été réalisés à partir de différents compartiments des coqs fermiers :
Jabot, gésier, duodénum, jéjunum, colon… dans des conditions stériles, chaque échantillon
prélevé a été ensemencé en milieu lait liquide et mis en incubation à différentes températures
30°C et 45°C. Après 24h d’incubation toutes les cultures ont été diluées et ensemencées sur
milieu gélosé MRS et incubées en anaérobiose dans les mêmes conditions de température. La
purification des souches a été établit par réalisation de subcultures sur milieux liquide et
gélose MRS jusqu'à l’obtention de colonies bien distinctes et homogènes, la pureté des
souches a été vérifiée par observation microscopique.
3. Conservation des souches
A partir des cultures jeunes des souches sur milieu liquide, 1.5 ml de culture est transféré
dans un tube eppendorf stérile. Les cellules sont récupérées par centrifugation à 3000
tours/min pendant 10min, après élimination du surnageant le culot est resuspendu dans le
milieu de conservation MRS additionné de glycérol à 15%. Les échantillons sont portés au
congélation -20°C.

Matériels et Méthodes
45
4. Souches de référence utilisées
Deux souches de référence ont été utilisées et représentent des espèces types appartenant à la
collection INRA : Lactobacillus delbreuckii ATCC 11842 et Streptococcus thermophilus
CNRZ 1066.
Les souches pathogènes de références utilisées sont Staphylococcus aureus (ATCC 25923,
ATCC 43300), Escherichia coli ATCC25922 et Enterococcus faecalis (V583 et JH2-2).
5. Caractérisation phénotypique
5.1. Examen macroscopique
Les colonies obtenues sur les boites pétri sont observées à l’œil nu afin de déterminer la
forme, la taille, l’aspect, le contour des colonies ainsi que la couleur.
5.2. Examen microscopique
Sur des cultures jeunes, des colorations de Gram ont été réalisées, l’observation
microscopique s’est faite au grossissement (Gx1000) ce qui permettra d’observer la
morphologie des cellules bactériennes (la forme, le mode d’association, taille …) et le type
de Gram des différents isolats.
5.3. Test de la catalase
La catalase est une enzyme qui permet la dégradation de peroxydase d’hydrogène (H2O2).
Le test de catalase est réalisé afin de vérifier l’appartenance des souches isolées au groupe des
bactéries lactiques en mettant en contact une colonie avec une goutte de H2O2, un résultat
positif est estimé par effervescence, les bactéries lactiques étant catalase négatif.
6. Recherche des bactéries mucoïdes
6.1. Production et Caractérisation des exopolysaccharides (EPS)
6.1.1. Formation des exopolysaccharides
Il existe plusieurs milieux de culture pour le développement d’une meilleure production des
exopolysaccharides sur gélose qui est le milieu HJL et le milieu hypersaccharosé. Il existe
aussi plusieurs façons de détecter la présence de polysaccharides. Ces derniers peuvent être
détectés qualitativement par un examen visuel, par coloration ou par microscopie. La mesure
de la viscosité et le dosage des EPS purifiés permet de les détecter quantitativement.

Matériels et Méthodes
46
Les souches à tester ont été ensemencées en stries sur milieu gélose hypersaccharosé et
incubées à 37°C pendant 24h, la production des exopolysaccharides se manifeste par
l’apparition de colonies larges et gluantes (Leveau et al., 1991).
6.1.2. Mise en évidence des EPS en milieu solide au rouge de ruthénium
La production des EPS peut être visualisée en milieu gélosé par coloration au rouge de
ruthénium, un colorant cationique qui se fixe aux composants de la paroi bactérienne
(Stingele et al., 1996).
Il est possible de sélectionner des clones bactériens producteurs d’EPS appelés clone Muc+,
en ajoutant des colorant cationiques dans le milieu de culture, les clones Muc+ sont révélés
par masquage de la coloration par le polysaccharide produit.
Le colorant cationique additionné en milieu de croissance se fixe donc sur le peptidoglycane
de la paroi bactérienne, ce qui entraine une coloration de la colonie d’une souche non
productrice. Ces colorants ne présentent pas d’affinité pour les polysaccharides excrétés, les
polysaccharides produits masquent la coloration et les clones Muc+ apparaissent blancs
(Gancel et al ., 1988).
Un exemple de colorant cationique est le rouge de ruthénium qui donne une coloration rose
foncé au milieu de culture, les colonies Muc+ apparaissent donc blanches sur fond rose par le
masquage de la couleur du aux les exopolysaccharides produits. Les clones Muc- apparaissent
rosés.
6.1.3. Mise en évidence des EPS par microscopie
Le colorant bleu de toluidine est utilisé pour une coloration vitale. La réalisation de cette
coloration se fait entre lame et lamelle, on dépose une goutte du colorant bleu de toluidine
préparé en solution a 1% sur laquelle on étale quelques cellules d’une colonie. On procède à
une observation sous microscope optique (Sheehan DC, Hrapchak BB. 1980).
6.1.4. Extraction des EPS
Les exopolysaccharides sont extraits selon la méthode modifiée par Evans LR and Linker M,
(1973) qui consiste à récupérer la totalité des exopolysaccharides produits sur milieu gélosé
et les faire homogénéiser dans 10 ml d’eau distillée stérile. La suspension obtenue est
centrifugée à 3000 tours pendant 1h.

Matériels et Méthodes
47
Apres récupération du surnageant, on additionne trois volumes d’éthanol à 95% et on laisse
précipiter à froid pendant la nuit. On réalise une dernière centrifugation pour la récupération
du culot contenant les EPS extraits.
6.1.5. Lyophilisation des EPS
La lyophilisation consiste en l'élimination progressive de l'eau du produit préalablement
congelé (phase solide) par passage à la phase vapeur, sans passer par la phase liquide. Ce
changement d'état s'appelle la sublimation.
Les extrait d’EPS ont été congelé a -20°C pendant 24h suivi d’une lyophilisation pendant 48h
avec un appareil CHRIST, ALPHA 1-2 MODEL, Germany. Ce travail a été réalisé au
laboratoire de Nutrition du département de Biologie de l’université es-sénia.
6.1.6. Dosage colorimétrique des EPS
Les exopolysaccharides purifiés et lyophilisés sont resuspendus dans 10 ml d'eau dé-ionisée
(Millipore) et dosés selon la méthode de dosage des sucres totaux de (Dubois et al., 1956)
modifiée par Gerhardt et al. (1981).
Le principe de cette méthode repose sur le fait que l'ajout d'acide sulfurique provoque
l'hydrolyse des polysaccharides et la formation de 5- hydroxyméthyl-furfural qui donne, en
présence de phénol, une coloration orangée. L'intensité de la couleur est directement
proportionnelle à la quantité de sucre présent et est directement mesurable par
spectrophotométrie. La réaction est sensible et la couleur est stable. De petites quantités de
carbohydrates peuvent être détectées et la présence de protéines ou de peptides n'influence pas
le dosage.
Une courbe standard de 0 à 120 mg/l est préparée à partir d'une solution stock. Chaque
échantillon est préparé trois fois et pré-dilué de la manière suivante : les échantillons de
polysaccharides ont été dilués dans un premier temps au 1/10, cependant la concentration des
EPS est restée trop forte, ce qui a emmener à réaliser une deuxième dilution au 1/100 qui n’a
pas donnée les résultats souhaités. Une troisième dilution au 1/200 (20 µl Écha/3980 µl eau)
qui a permis de doser convenablement par spectrophotomètre les échantillons préparés. Le
blanc est fait avec la concentration 0 mg/l. A l‘aide d'une micropipette calibrée, 1 ml de
standard (Glucose) ou d'échantillon (EPS) est placé dans un tube à essai en verre propre. En
utilisant la même micropipette, 1 ml de Phénol 5% est ajouté. Finalement, 3 ml d'acide
sulfurique sont ajoutés au mélange. Les tubes sont immédiatement agités au vortex et

Matériels et Méthodes
48
refroidis pendant environ 10 minutes à température ambiante. A noter que l’acide sulfurique
provoque une augmentation de la température.
La lecture de l’absorbance est faite avec un spectrophotomètre Biochrom Libra S6 à une
longueur d'onde de 490 nm. Les quantités de polysaccharides sont exprimées en mg/l
d'équivalent glucose.
Le taux de sucre est déterminé en se référant à une courbe d’étalonnage tracé avec du glucose.
· Préparation d’une gamme étalon glucose
Une série de dilutions a été effectuée à partir d’une solution mère de glucose à concentration
120 mg/l.
6.1.7. Activité rhéologique
Ce test a été réalisé par ensemencement du lait écrémé stérile par 1ml d’EPS natif.
L’incubation a été faite à température ambiante. Le levain lactique possède un pouvoir
épaississant si le gel formé présente une certaine rhéologie visqueuse (Bourgeois et Leveau,
1991).
6.2. Analyse de la composition des EPS
Les exopolysaccharides selon leur propre origine peuvent présenter des chaines linéaires, des
chaines ramifiées ou des chaines complexes. La nature des sucres qui composent ces chaines
diffère d’un EPS à un autre, en effet certains sont riches en un seul ose (glucose pour
dextrane) et d’autres montrent une diversité (Xanthane).
L’hydrolyse permet une fracture de ces bio –polymères en sucres plus simples tel que les oses
ou disaccharide.
La chromatographie sur couche mince permet de déceler la nature de ces sucres en fonction
de leur migration sur plaque et leur rapport frontale (Rf).
6.2.1. L’hydrolyse des EPS
L’hydrolyse des polysaccharides conduisant à l’identification des monosaccharides s’est
poursuivie sur une période de 4h à 100°C. Un volume de 2 ml d’acide sulfurique a été ajouté
à 20 mg de polysaccharide séché et purifié. L’hydrolyse a été neutralisé avec de l’hydroxyde
de baryum (Graber et coll., 1988) puis centrifugé à 3500 rpm pendant 10min. Le surnageant a
été filtré par l’utilisation de filtre millipore de 45µm (Millex®-GS Sterilizing Filter Unit
SLGS025BS).

Matériels et Méthodes
49
6.2.2. Chromatographie sur couche mince des oses constitutifs des polysaccharides
Il s’agit d’une méthode chromatographique qui permet, suivant un procédé simple et rapide,
l’identification et la séparation d’un très grand nombre de substances organiques et minérales.
On l’utilise dans cette étude afin de séparer les composants des exopolysaccharides extraits et
donner une identification aux monosaccharides constituants ces biopolymères.
La chromatographie sur couche mince a été réalisée selon la méthode décrite par Souly Frag
(1978).
Le protocole de migration se présente comme suit :
· Solution à analyser
10 mg de chacun des échantillons obtenus ainsi que les sucres standards sont dissouts dans
1ml d’un mélange méthanol et eau (1 :1).
Les sucres utilisés : Glucose, Raffinose, Arabinose, Saccharose, Galactose, Lactose et
dextrane.
· Préparation de la cuve
1ère technique utilisée : le mélange solvant chloroforme-acide acétique-eau (30 :35 :5)( est
introduit au fond de la cuve bien fermée par l’étanchéité du couvercle (Souly Frag 1978).
1h d’attente est nécessaire pour assurer la saturation de l’atmosphère de la cuve.
2ème technique utilisée : le travail s’est accompli dans les mêmes conditions précédentes à
l’exception du système solvant utilisé qui est Acétone /Eau 15 :85 (Soczewi ński E et al., 1998)
· Dépôt des échantillons
A l’aide d’un micro-tube capillaire on dépose 20 µl de chaque échantillon sur une plaque de
silice (TLC Silica-gel 60 F254 MERCK).
A la fin, chaque dépôt est bien séché.
· Développement du chromatogramme
Les échantillons déposés sur la plaque ont migré dans une cuve saturée selon les deux
systèmes de solvant décrits si dessus.

Matériels et Méthodes
50
· Révélation
Les solutés étant incolores, il faut les faire apparaître par une réaction chimique. On utilise
pour cela un révélateur : 50 ml de permanganate de potassium à 20 g/l mélangé au moment de
l’emploi à 50 ml de carbonate de sodium anhydre à 40 g/l dans un vaporisateur.
Après le dépôt de la plaque sur une table propre, on vaporise le révélateur sur l’ensemble de la
surface. Le séchage de cette dernière est réalisé à l’aide d’un séchoir.
Pour identifier les constituants du mélange des différents échantillons on calcule le rapport
frontal de chaque tâche de migration (Rf= distance parcourue par la substance (centre de la
tache) / distance parcourue par le solvant).
Le rapport frontal Rf est caractéristique d’une substance donnée dans un solvant donné.
7. Etude de l’activité antimicrobienne des souches probiotiques isolées du tractus digestif
vis-à-vis de souches pathogènes
Des interactions bactériennes englobant les isolats du genre Lactobacillus et des souches
productrices des EPS pré-identifiés à des Leuconostoc et Pediococcus ont été utilisées à
l’encontre de souches pathogènes de référence afin de sélectionner des bactéries d’intérêt
productrices de substances inhibitrices vis-à-vis des bactéries non désirables, utilisées
notamment contre Escherichia coli ATCC 25922, Staphylococcus aureus (ATTC25923,
ATCC 43300), Salmonella typhi, et Enterococcus faecalis (V583/ JH2-2).
Deux méthodes ont été utilisées à savoir celle des disques (porte germes) selon Tadesse et al.
(2004) qui consiste à :
· inonder en surface les boites de Pétri contenant le milieu MH par quelques millilitres
des pré-cultures de la souche pathogène (souche indicatrice),
· laisser sécher les boites pendant 30 min à 37°C, après séchage, déposer à la surface de
la gélose des disques en papier buvard stérile préalablement imprégnés par la pré
culture des bactéries lactiques, ensuite sécher encore une fois les boites pendant 30
min.
· Mettre les boites à 4°C pendant 4h pour assurer la diffusion des substances
responsables de l’interaction, enfin incuber les boites à 37°C pendant 24h.
· L’inhibition de la souche indicatrice se traduit par la formation de zones claires autour
des disques dont le diamètre est mesuré à partir du centre du disque en mm.

Matériels et Méthodes
51
En deuxième lieu, une seconde méthode celle de la double couche selon Fleming et al.
(1975) a été utilisée et qui consiste à :
· Les souches à tester (supposées être inhibitrices) sont ensemencées par spot à la
surface d’un milieu MRS solide. Les boites de pétri ainsi ensemencées sont incubées
pendant 18h (pré-incubation) dans les conditions favorables de la souche.
· Les souches indicatrices (souches pathogènes) sont ensemencées en masse, un volume
de 0.5 ml de culture d’une souche pathogène est rajouté à 7 ml de gélose molle. Ce
mélange est versé sur la surface des boites pétri pré-incubées.
· Après solidification du milieu, une deuxième incubation est nécessaire à 30°C pendant
24h.
· La formation des halos claires autour de la souche indique une inhibition vis-à-vis le
pathogène testé.
7.1. Nature de la substance inhibitrice
Pour montrer que la substance inhibitrice n’est pas d’origine acide (acide lactique), le
surnageant doit être traiter. Une culture de 10 ml a été centrifugée à 3000 tour/min pendant 10
min pour éliminer les cellules bactériennes, puis une neutralisation de l’acidité du milieu a été
effectuée par ajustement du pH du surnageant avec du NaOH 2N suivi d’une filtration à
travers un filtre millipore de 0.22µm (Millex®-GS Sterilizing Filter Unit SLGS025BS).
Le surnageant obtenu a été utilisé en interaction avec les pathogènes.
8. Caractérisation moléculaire des souches probiotiques
A ce titre, un cadre législatif a été établi sur les probiotiques en 2002 par l’EFSA (European
Food Safety Agency) d’où ressort trois principaux chapitres : l’identité, la sécurité et
l'efficacité de la souche.
L’identification des microorganismes par les méthodes ADN est devenue une méthode
incontournable, elle est plus résolutive et plus discriminante par rapport aux méthodes de
classification traditionnelle. La taxonomie des probiotiques utilisés comme additif en
aviculture est régie par la loi.

Matériels et Méthodes
52
8.1. Extraction d’ADN
8.1.1. Extraction d’ADN par utilisation de MOBIO KIT
Les examens macroscopiques et microscopiques étant réalisés, les tests Gram positif et
catalase négative des différents isolats nous ont conduit à une identification préliminaire à des
lactobacilles. En matière de diagnostic de ces souches, une approche plus approfondie a été
consacrée à l’amplification de l’ADNr 16S par la technique PCR en utilisant des amorces
spécifiques d’une région interne du gène de l’ADNr 16S préconisant ainsi un fragment de 480
pb.
Afin d’amplifier par PCR des cours fragments, la méthode d’extraction avec MOBIO KIT est
utilisée. Dans ce cas c’est une région interne de 480 pb de l’ADNr 16S spécifique au genre
Lactobacillus. Au total 13 souches ont été testées en utilisant le couple d’amorce V3/V4
(amorces INRA, communication personnelle).
V3 5’-CTC CTA CGG GAG GCA GCAG-3’
V4 3’-GGA CTA CCAGGGTATCTAAT-5’
Le protocole d’extraction appliqué se présente comme suit :
1. Mettre 1.8 ml de culture microbienne dans un tube de capacité de 2ml, le faire centrifuger à
10.000 tours/min pendant 30 secondes à température ambiante. Eliminer le surnageant.
2. Resuspendre le culot dans 300 µl de la solution Microbead et vortexer. Transférer les cellules
resuspendues dans le tube Microbead.
3. Ajouter 50 µl de solution MD1 au tube Microbead. Vortexer à la vitesse maximale pendant 10
minutes.
4. S’assurer que les tubes de Microbead de 2 ml tournent librement dans la centrifugeuse sans
frottage. Laisser centrifuger à 10.000 tours/min pendant 30 secondes à température ambiante.
5. Transférer les surnageants dans de nouveaux tubes propres de 2 ml. Prévoir 300 µl ou 350 µl
de surnagent.
6. Ajouter 100 µl de solution MD2 au surnagent, vortexer pendant 5 secondes et incuber à 4°C
pendant 5 minutes.
7. Centrifuger les tubes à 10.000 tours/min pendant 1 minute.
8. En évitant le culot, transférer le volume entier du surnagent dans des tubes propres de 2 ml.
Prévoir 450 ml en volume.
9. Secouer la solution MD3 avant l’emploi. Ajouter 900 µl de cette solution au surnagent et
vortexer pendant 5 secondes.
10. Charger environ 700 µl dans le spin-filter et centrifuger à 10.000 tours/min pendant 30
secondes à température ambiante. Jeter le surplus, ajouter le surnagent restant au spin-filter,

Matériels et Méthodes
53
centrifugé à 10.000 tours/min pendant 30 secondes. Un total de 2 ou 3 chargements sont
nécessaires pour chaque échantillon pour avoir le maximum d’ADN concentré.
11. Ajouter 300 µl de la solution MD4 et centrifuger à 10.000 pendant 30 secondes, puis jeter le
surplus.
12. Centrifuger à température ambiante à 10.000 tours/min pendant 1minute.
13. Faire attention à ne pas éclabousser le liquide sur le support de spin-filter, puis le placer dans
un nouveau tube de 2 ml.
14. Ajouter 50 µl de la solution MD5 au centre de la membrane blanche du filtre.
15. Centrifuger à température ambiante à 10.000 tours/min pendant 30 secondes.
16. Jeter le filtre de rotation, l’ADN dans le tube est resuspendu dans 50 µl d’eau distillée
stérile et enfin prêt pour n’importe quel autre application.
Les solutions du KIT
-Solution MD1 : contient le détergeant SDS et d’autres agents de perturbation nécessaires
pour la lyse cellulaire.
-Solution MD2 : contient des réactifs pour la précipitation des matériaux organiques et
inorganiques non-ADN y compris les débris cellulaires et les protéines.
-Solution MD3 : est une solution de concentration élevée en sels, utilisée pour lier l’ADN
avec la membrane du Spin-filter tube.
-Solution MD4 : solution de lavage à base d'éthanol, utilisée pour éliminer les résidus de sel et
d’autres contaminants.
-Solution MD5 : tampon d'élution, contient le Tris 10mM à pH8.
8.1.2. Extraction d’ADNr 16S dans sa totalité (1500pb)
Afin d’uniformiser cette méthode à l’ensemble des souches du laboratoire, des amorces
universels ont été appliquées à 5 genres différents, dont l’un d’eux a été pré-identifié à un
pédiocoque LGM Clo1.
La technique de Gevers et al. (2001) est utilisée dans ce cadre. Des amorces universels
8UA/1492R (Sato T et al.,2003) sont utilisées dans la réaction mix PCR.
Le protocole d’extraction ADN se présente comme suit :
-Les cellules à partir d’une colonie sont resuspendu dans 1ml d’eau distillée stérile et
centrifuger à 3000 tours/min pendant 10 minutes et stocker le culot à -20°C pendant 1h.
-Le culot est ensuite lavé dans 1ml de tampon TES (6.7% sucrose, 50 mM Tris-Hcl pH 8,
1mM EDTA) et resuspendu dans 300 µl de tampon STET (8% sucrose, 5% Triton X-100,
50mM Tris-Hcl pH 8, 50mM EDTA).

Matériels et Méthodes
54
- 75 µl de tampon de lyse (TES contenant 1330U/ml mutanolysine et 40 mg/ml lysosyme)
sont ajoutés à la suspension et incubés à 37°C pendant 1h.
-Ajouter par la suite 40 µl de SDS préchauffé (37°C) dans un tampon TE (10 mM Tris-Hcl,
1mM EDTA, pH 8), les cellules sont vortexées pendant 60 secondes et incubées à 37°C
pendant 10 min suivi d’une incubation à 65°C pendant 10 min.
- 100 µl de tampon TE sont ajoutés et le lysat est extrait avec 1volume de
(phénol/chloroforme/isoamylalcool).
- Les phases sont séparées par centrifugation à 3000 tour/min pendant 20 min. La phase aqueuse est mixée avec 70 µl de NaCl 5M et 1 ml d’isopropanol, l’ADN est précipité dans la
glace pendant 15 min.
- L’extrait ADN est collecté par centrifugation 3000 tours/min pendant 20 min et le culot est lavé dans de l’éthanol froid à 70%.
-Le culot ADN est resuspendu dans 50 µl d’eau distillée stérile pour des fins analytique.
8.1.3. Vérification qualitatif de l’ADN extrait
Afin de vérifier la qualité de l’ADN après extraction, la technique la plus commune est la
migration de l’échantillon sur gel d’agarose 1 % suivi d’une visualisation sous lumière UV.
Si les ADN migrent sous la forme d’une trainée (smear) l’échantillon d’ADN a sans doute
subi une dégradation majeure.
8.2. Amplification
8.2.1. Amplification de la région interne de l’ADNr 16S (480) du genre Lactobacillus spp.
L'amplification d'ADN a été effectuée avec un thermo-cycler d’amplification de type PCR
TECHNE TC-312 (Barloworld Scientific Ltd, stone ST 15 OSA, UK) selon le programme
d’amplification suivant :
Pré-dénaturation à 95°C pendant 5min,
- Dénaturation à 94°C pendant 30 secs
- Hybridation à 60°C pendant 30 sec 35 Cycles
- Elongation à 72°C pendant 1 min
Enfin une étape d’élongation 72°C pendant 7 min.

Matériels et Méthodes
55
8.2.2. Séquençage et taxonomie des souches lactobacilles de la flore intestinale
L’ADN total des différents isolats à été extrait selon la méthode appropriée à MOBIO KIT.
L'amplification d'ADN a été effectuée avec un thermo cycler d’amplification de type PCR
TECHNE TC-312 (Barloworld Scientific Ltd, stone ST 15 OSA, UK) suivant un programme
d’amplification spécifique. La présence des produits amplifiés par PCR est révélée en
électrophorèse sur gel d'agarose à concentration 1.3 % en utilisant un marqueur de taille
Smart Ladder 200 pb. Les réactions de séquençage sont effectuées suivant une méthode
dérivée de celle décrite par Sanger et al.(1977) à l’aide du kit « BigDyeTM Terminator Cycle
Sequencing. Un seul brin a été séquencé, et chaque séquence a été comparée aux séquences
disponibles dans les bases de données du NCBI GenBank en utilisant l’outil de recherche
Blast. Cette partie de l’étude a été réalisée et traitée à l’INRA de Jouy-en-Josas.
8.2.3. Amplification du gène ADNr 16S (1500 pb) dans sa totalité
L'amplification d'ADN a été effectuée dans le même thermo-cycler du laboratoire cité ci-
dessus suivant un programme d’amplification :
Pré-dénaturation à 95°C pendant 15min,
- Dénaturation à 94°C pendant 1min,
- Hybridation à 60°C pendant 1min 35 Cycles
- Elongation à 72°C pendant 1.5min
Enfin une étape d’élongation finale a 72°C pendant 10min.
Les couples d’amorces universels utilisés (Sato et al., 2003) :
8UA: 5’AGAGTTTGATCCTGGCTCAG -3’
1492R : 3’-TACGGGTACCTTGTTACGACTT-5’
· Les mélanges réactionnels utilisés pour les deux amplifications PCR, sont réalisés à un
volume final de 25 µl contenant :
- 2.5 µl 10X tampon PCR (Takara),
- 2µl dNTP (Takara),
- 0.5 µl de chaque amorce,
- 0.25 µl de taq ADN polymerase (Takara)
- H2O stérile en qsp.
Le produit de PCR est une bande ADN préconisé à 1500 pb pour le gène ADNr 16S et une
bande de 480 pb pour la région interne du gène ADNr16S du genre Lactobacillus spp.

Matériels et Méthodes
56
8.3. Electrophorèse sur gel d’agarose
5 µl des produits PCR ont été analysés par électrophorèse sur gel d’agarose à 1.3% pour la
visualisation des grands fragments d’ADN à 1500 pb et 0.8% pour les petits fragments de 480
pb à 100 V pendant 60 minutes.
La présence des produits amplifiés par PCR est révélée en électrophorèse sur ce gel en
utilisant un marqueur de taille Smart Ladder 200 pb.
· Préparation du gel
- Peser 1.3 g d’agarose (pour la recherche des grands fragments d’ADN) et 0.8% (le cas
des petits fragments d’ADN) dans un Erlenmeyer de capacité suffisante.
- Ajouter l’agarose à 100 ml de TBE.
- Chauffer avec agitation jusqu’à ébullition et disparition du trouble, laisser refroidir
jusqu'à ce qu’il devienne possible de saisir le flacon à main nue (température environ
45-50 °C).
- Ajouter 1μl de BET (Bromure d’éthidium) au gel TBE (voir composition dans
l’annexe), agiter doucement pour éviter la formation de bulles d’air.
- Préparer le moule à peigne pour couler le gel.
- Placer le moule sur une surface bien horizontale.
- Couler lentement le gel sur 3 à 5 mm d'épaisseur et laisser se solidifier dans la cuve.
- Laisser refroidir 30 minutes environ avant d’enlever délicatement le peigne.
- Les puits sont placés du côté de la cathode.
- Le gel est enfin prêt pour le dépôt des échantillons.
· Dépôt des échantillons (produits d’amplification)
- Mélanger la solution de charge (bleu de bromophénol et glycérol) avec l’ADN sur une
petite plaque à dépôt (5 μl d’ADN et 3 μl de la solution de charge) avec une
micropipette.
- Remplir les puits du gel avec ce mélange en faisant attention à ne pas déchirer le fond
du gel avec la pointe de l’embout (figure 6).

Matériels et Méthodes
57
Figure 6: dépôt des échantillons ADN sur gel d’agarose au laboratoire LGM
· Migration
- Fermer la cuve, brancher les fils et exercez un voltage de 80 v jusqu’à 100 v
- Laisser migrer jusqu’à ce que le colorant de charge arrive à proximité du bord du gel
(environ 40 minutes).
- Une fois le témoin de migration (colorant bleu) atteint l’extrémité du gel, on arrête la
migration.
Figure 7 : montage de l’électrophorèse du laboratoire LGM

Matériels et Méthodes
58
· Visualisation La visualisation de l’ADN sur le gel d’agarose est réalisée grâce à un colorant fluorescent le
bromure d’éthidium (BET). Ce composé possède la propriété de s’intercaler entre les paires
de bases des acides nucléiques. Le gel est visualisé dans l’obscurité à l’aide d’une plaque à
UV (uvitec SXT-F26M, 312nm, power 180W) puis photographié avec un appareil numérique.

Résultats et Discussion
59
1. Caractérisation phénotypique
Au total 48 isolats de différents compartiments du tube digestif des coqs fermiers ont été
recensés et sélectionnées dans des conditions d’ambiance appropriées et de milieux adéquats
pour une durée de 24 à 48h d’incubation.
Toutes les souches sont de Gram positif et catalase négative et ont été identifiées à des
bactéries lactiques, dont 22 identifié d’une façon préliminaire sur la base d’une étude
phénotypique, selon les critères aspect, forme et la couleur des colonies ainsi que
l’observation des cellules bactériennes en forme de bâtonnets fin longs, moyens, épais courts,
incurvés et absence de spores ont orientés la pré-identification des différents isolats au genre
Lactobacillus (figure 8).
On signale la présence de la diversité d’une flore particulièrement assez riche en forme de
coques en petite en grande structure, en chainettes coutes et longues ainsi que de bacilles
justifiant l’existence d’autres espèces à intérêt telles que les bactéries lactiques de différents
genres dont 6 ont été pré-identifiés à des leuconostocs, 2 à des pédiocoques, 8 à des
entérocoques, ainsi qu’à deux autres groupes non lactiques dont 5 apparentés au genre
Bacillus et 4 appartenant à des levures.
Les résultats des observations microscopiques des souches isolées sont regroupés dans les
figures 8, 9, 10, 11, 12, 13,14, 15.
2. Mise en évidence et caractérisation des EPS
2.1. Production des exopolysaccharides en milieu solide
L’ensemencement des échantillons sur les différents milieux de cultures à permis d’isoler un
certain nombre de bactéries productrices d’exopolysaccharides. On a pu obtenir 8 souches
bactériennes (LGM-IL2, LGM-CLO1, LGM-Co1, LGM-CA1, LGM-Ja1, LGM-Il3, LGM-
Pro4, LGM-jb2) capable de se développer sur milieu hypersaccharosé après 24 à 48 h
d’incubation en anaérobiose à 30°C.
Ces isolats se caractérisent par la formation de colonies mucoïdes à aspect gluant sur ce
milieu gélosé témoignant d’une production massive d’un biopolymère épaississant,
exopolysaccharides (EPS) (figure 16).
Ces résultats rejoignent celles de Bouzar et al. (1997) qui avaient constaté que l’espèce Lc.
lactis est capable de produire des EPS, en culture pure comme en association avec St.
thermophilus. Ainsi, Looijesteijn et al. (2001) ont rapporté qu’au sein d’une même espèce de
bactéries lactiques, les résultats peuvent être différents. Ces auteurs ont pu identifier des

Résultats et Discussion
60
souches productrices d’EPS (voire très fortement) et des souches non productrices sans que ce
caractère n’engendre des disparités de croissance.
2.2. Production des EPS en milieu liquide
La production en EPS des bactéries isolées a été observée également sur milieu HJL liquide,
où l’apparition d’un trouble voir un biofilm est constatée au bout de 24 à 48 heures
d’incubation en anaérobiose à température égale à 30°c. Les résultats sont groupés dans la
figure 17.

Résultats et Discussion
61
La souche LGM- jé1 La souche LGM -jé2
La souche LGM –G2 La souche LGM-jé3
La souche LGM-G7 La souche LGM-C1
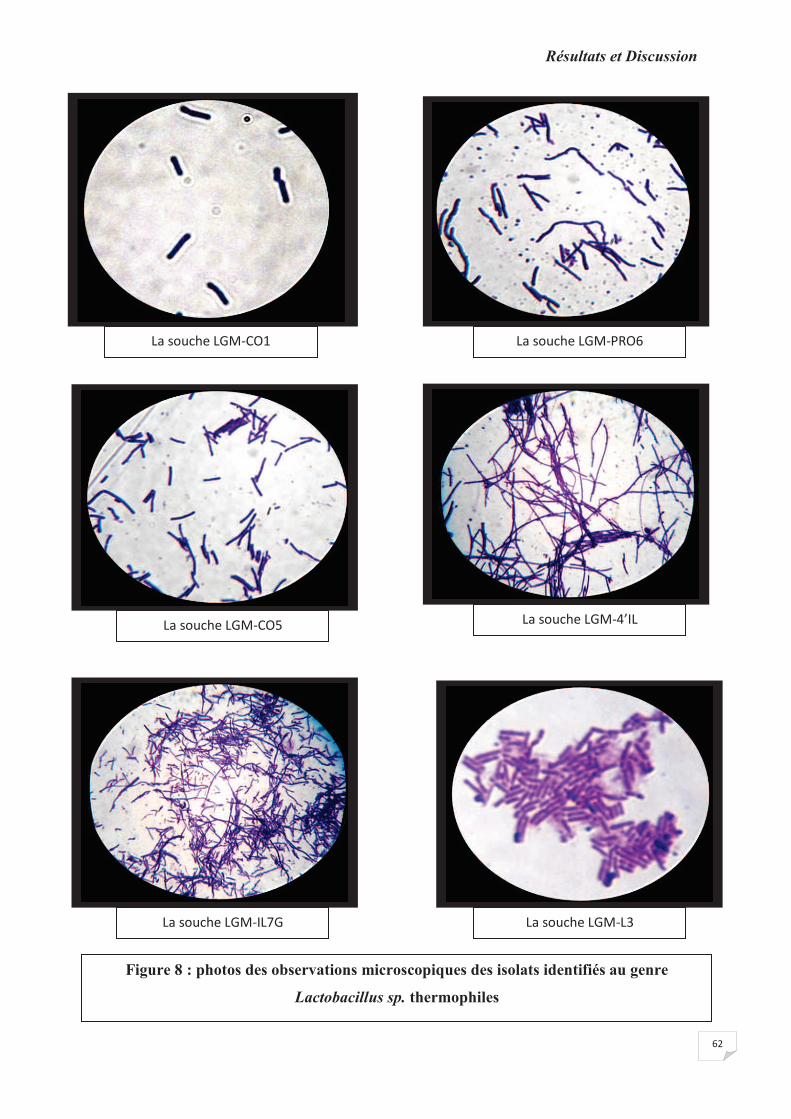
Résultats et Discussion
62
Figure 8 : photos des observations microscopiques des isolats identifiés au genre
Lactobacillus sp. thermophiles
La souche LGM-CO1 La souche LGM-PRO6
La souche LGM-CO5
La souche LGM-4’IL
La souche LGM-IL7G
La souche LGM-L3

Résultats et Discussion
63
Figure 9 : photos des observations microscopiques des lactobacilles mésophiles
La souche LGM-Jé2
La souche LGM-IL2
La souche LGM-CA7

Résultats et Discussion
64
Figure 10 : photos des observations microscopiques des isolats pré-identifiés au
Streptococcus themophilus
La souche LGM-J3
La souche LGM-J1
La souche LGM-Duod
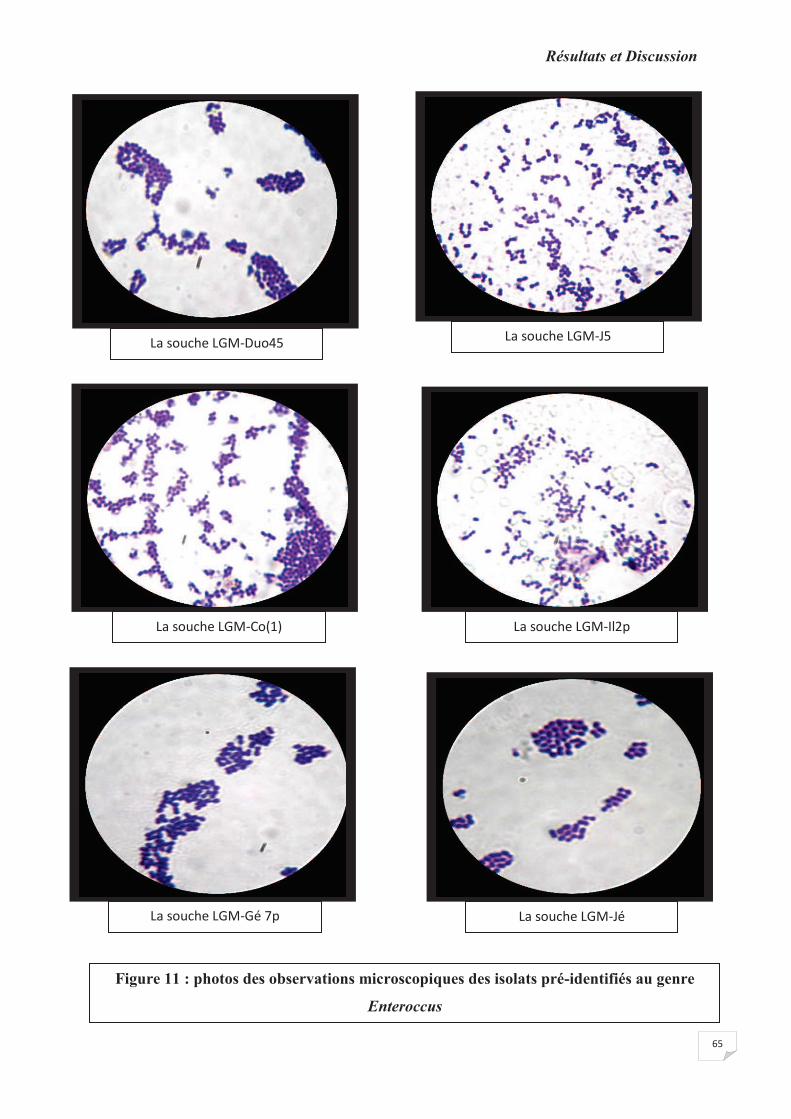
Résultats et Discussion
65
Figure 11 : photos des observations microscopiques des isolats pré-identifiés au genre
Enteroccus
La souche LGM-Gé 7p
La souche LGM-Jé
La souche LGM-Il2p
La souche LGM-Co(1)
La souche LGM-J5
La souche LGM-Duo45

Résultats et Discussion
66
Figure 12 : photos des observations microscopiques des isolats pré-identifiés à des
pédiocoques
La souche LGM-IL2
La souche LGM-CLO1
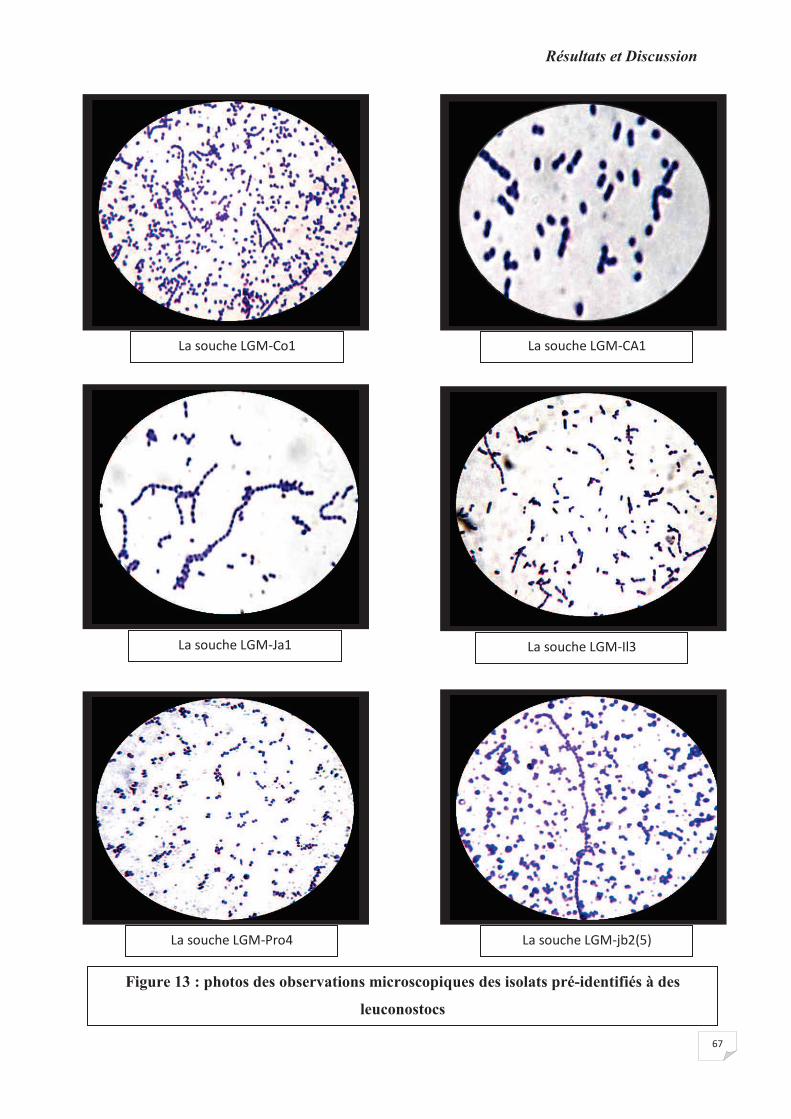
Résultats et Discussion
67
La souche LGM-jb2(5)
La souche LGM-Pro4
La souche LGM-Il3
La souche LGM-Ja1
La souche LGM-CA1
La souche LGM-Co1
Figure 13 : photos des observations microscopiques des isolats pré-identifiés à des
leuconostocs
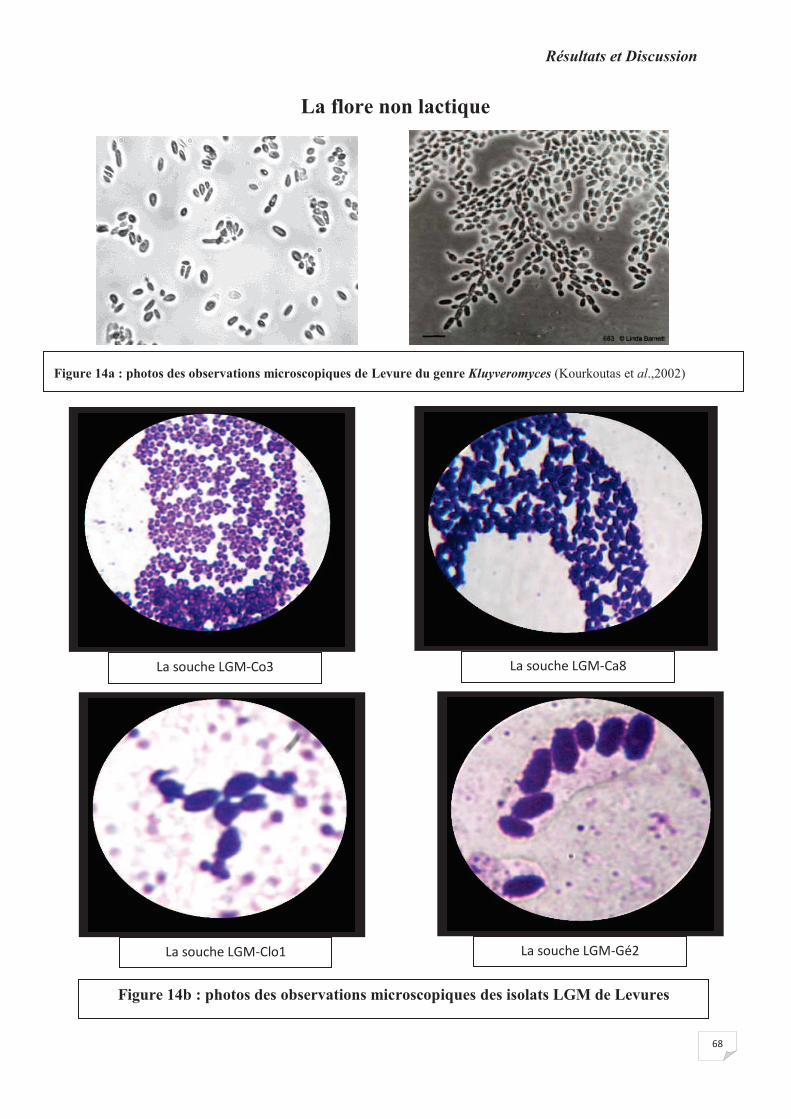
Résultats et Discussion
68
La flore non lactique
Figure 14b : photos des observations microscopiques des isolats LGM de Levures
La souche LGM-Clo1
La souche LGM-Gé2
La souche LGM-Co3
La souche LGM-Ca8
Figure 14a : photos des observations microscopiques de Levure du genre Kluyveromyces (Kourkoutas et al.,2002)
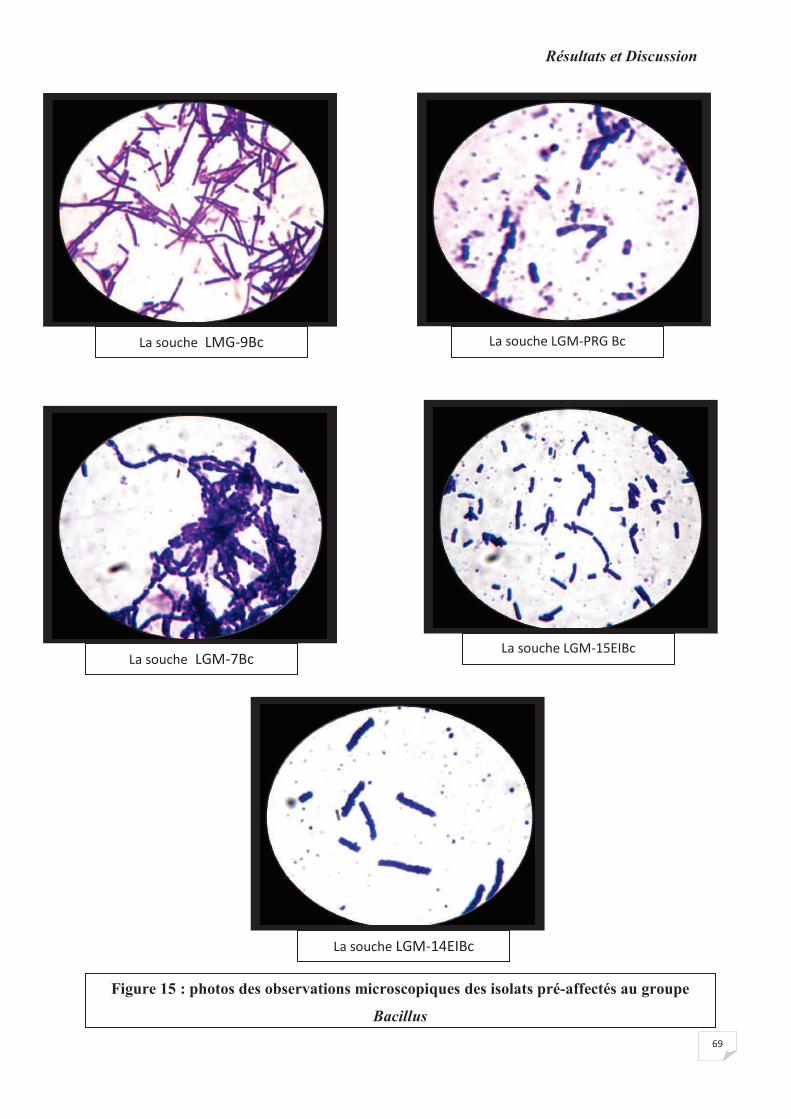
Résultats et Discussion
69
Figure 15 : photos des observations microscopiques des isolats pré-affectés au groupe
Bacillus
La souche LMG-9Bc
La souche LGM-7Bc
La souche LGM-15EIBc
La souche LGM-PRG Bc
La souche LGM-14EIBc
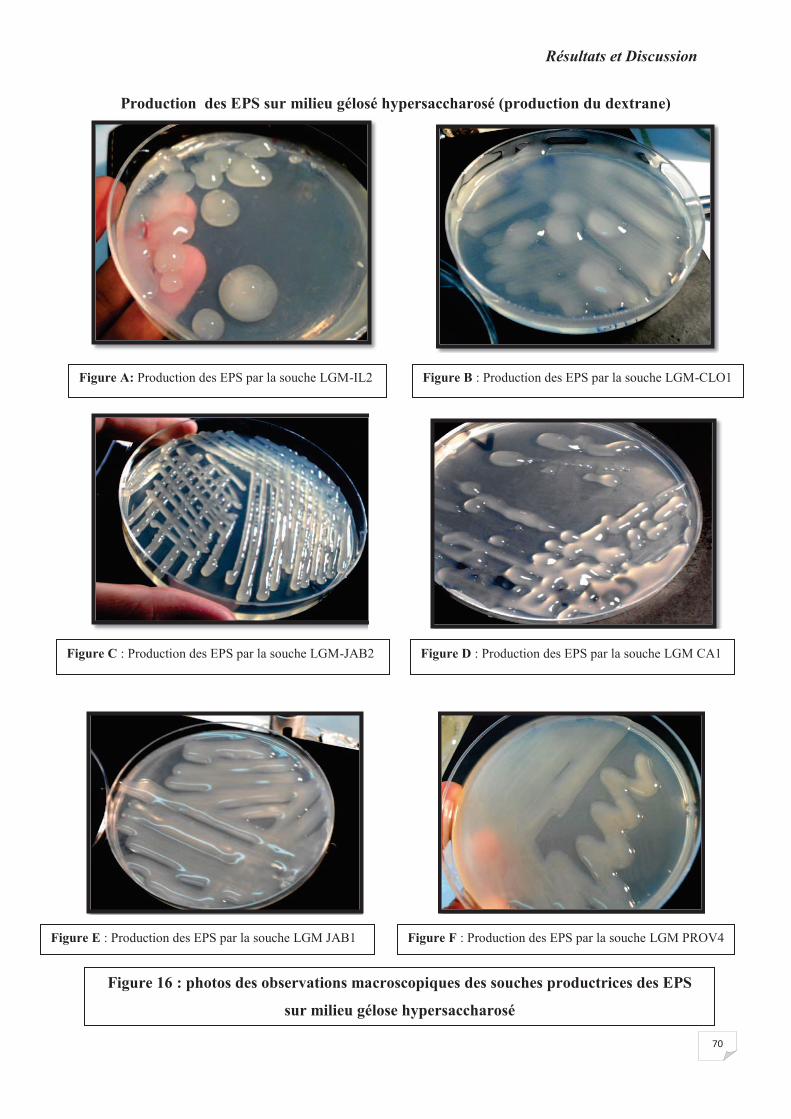
Résultats et Discussion
70
Production des EPS sur milieu gélosé hypersaccharosé (production du dextrane)
Production des EPS sur milieu liquide HJL (production de biofilm)
Figure A: Production des EPS par la souche LGM-IL2 Figure B : Production des EPS par la souche LGM-CLO1
Figure C : Production des EPS par la souche LGM-JAB2
Figure D : Production des EPS par la souche LGM CA1
Figure E : Production des EPS par la souche LGM JAB1
Figure F : Production des EPS par la souche LGM PROV4
Figure 16 : photos des observations macroscopiques des souches productrices des EPS
sur milieu gélose hypersaccharosé

Résultats et Discussion
71
Figure 17 : observations macroscopiques montrant un trouble avec précipité blanc d’EPS
en milieu liquide HJL
TEMOIN
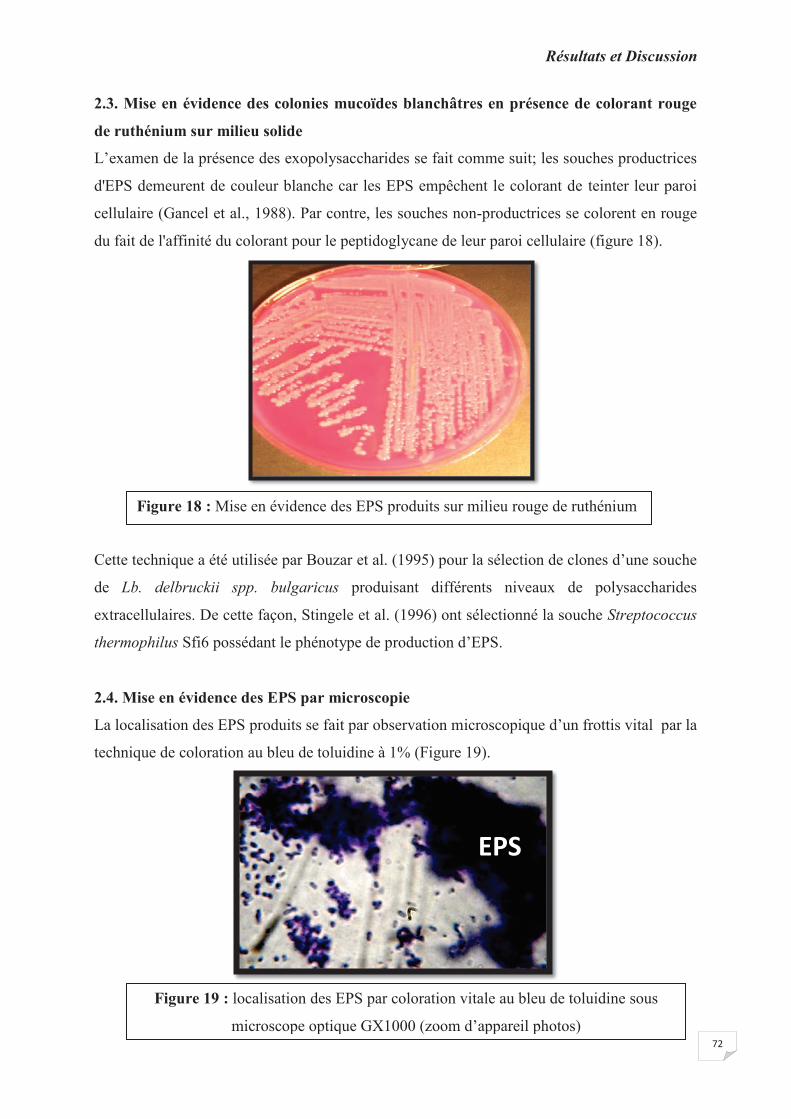
Résultats et Discussion
72
2.3. Mise en évidence des colonies mucoïdes blanchâtres en présence de colorant rouge
de ruthénium sur milieu solide
L’examen de la présence des exopolysaccharides se fait comme suit; les souches productrices
d'EPS demeurent de couleur blanche car les EPS empêchent le colorant de teinter leur paroi
cellulaire (Gancel et al., 1988). Par contre, les souches non-productrices se colorent en rouge
du fait de l'affinité du colorant pour le peptidoglycane de leur paroi cellulaire (figure 18).
Cette technique a été utilisée par Bouzar et al. (1995) pour la sélection de clones d’une souche
de Lb. delbruckii spp. bulgaricus produisant différents niveaux de polysaccharides
extracellulaires. De cette façon, Stingele et al. (1996) ont sélectionné la souche Streptococcus
thermophilus Sfi6 possédant le phénotype de production d’EPS.
2.4. Mise en évidence des EPS par microscopie
La localisation des EPS produits se fait par observation microscopique d’un frottis vital par la
technique de coloration au bleu de toluidine à 1% (Figure 19).
EPS
Figure 18 : Mise en évidence des EPS produits sur milieu rouge de ruthénium
Figure 19 : localisation des EPS par coloration vitale au bleu de toluidine sous
microscope optique GX1000 (zoom d’appareil photos)

Résultats et Discussion
73
2.5. Extraction des exopolysaccharides (EPS)
Après avoir ajouté l’éthanol, la précipitation des exopolysaccharides se fait après 24h à froid
(figure 20.a) et le culot contenant les EPS extraits est récupéré après centrifugation (figure
20.b).
Figure 20a : précipitation des EPS à l’éthanol Figure 20b : récupération du culot EPS
2.6. Lyophilisation des EPS
Obtention du lyophilisat soumis à une action du lyophilisateur pendant une période de 48 h
(figure 21).
Figure 21: exopolysaccharides lyophilisés

Résultats et Discussion
74
2.7. Propriétés rhéologiques des EPS
Le produit lyophilisé a été ajouté à 10 ml du lait liquide stérile et à de l’eau distillée stérile.
Ces derniers sont devenus filants et visqueux, lorsqu’ils coulent sur la paroi interne des tubes
présentent une viscosité élevée d’où le caractère épaississant de l’EPS (figure 22).
Un intérêt grandissant est porté aux exopolysaccharides produits par les bactéries lactiques.
En effet, ceux-ci ont le grade alimentaire GRAS (generally regarded as safe) et ils possèdent
aussi des propriétés rhéologiques particulières permettant d’améliorer la texture des produits
fermentés (Ruas-Madiedo et al., 2002).
D’après Béal et al. (2008), certaines bactéries lactiques sont capables de produire des EPS
dont l’accumulation provoque une viscosité des milieux.
Selon Lin et Chien (2007), la production des EPS dépend des bactéries lactiques et la durée de
la fermentation. Chamba (2008) a rapporté que les lactocoques comme les leuconostocs sont
capables de produire des EPS, ce caractère est généralement porté par un plasmide.
Eau
LAIT EAU
Figure 22 : activité rhéologique des EPS sur différents milieux
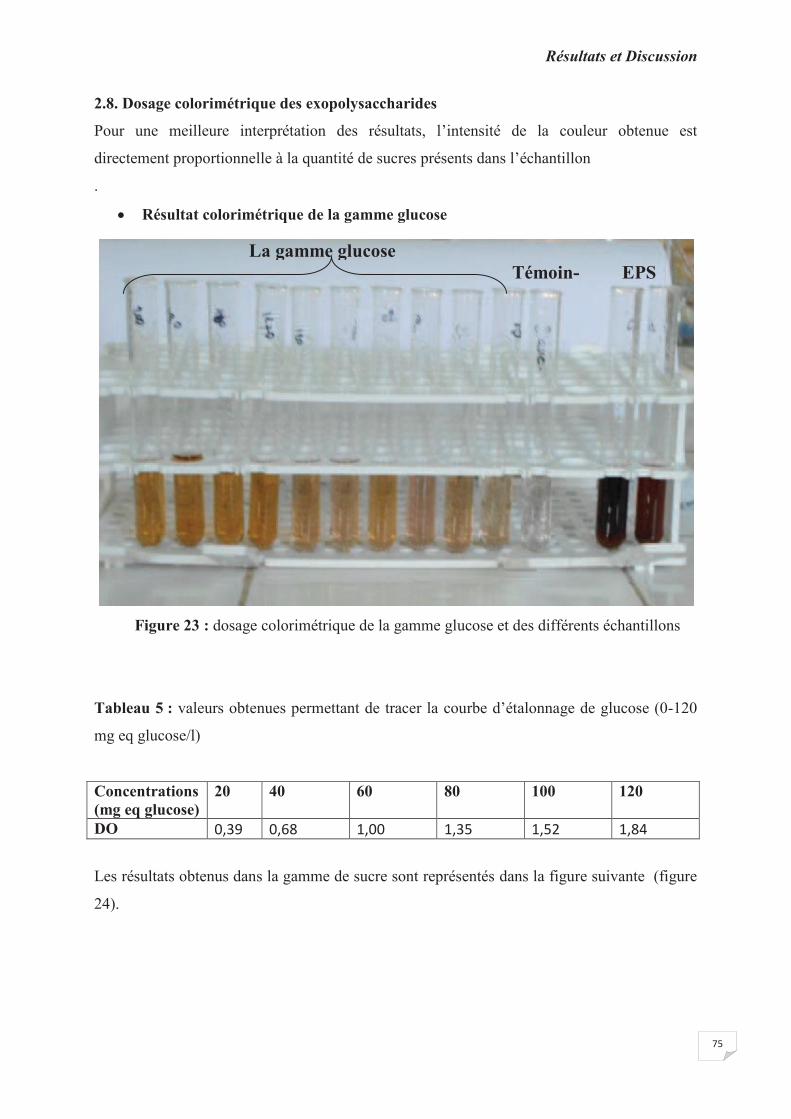
Résultats et Discussion
75
2.8. Dosage colorimétrique des exopolysaccharides
Pour une meilleure interprétation des résultats, l’intensité de la couleur obtenue est
directement proportionnelle à la quantité de sucres présents dans l’échantillon
.
· Résultat colorimétrique de la gamme glucose
Figure 23 : dosage colorimétrique de la gamme glucose et des différents échantillons
Tableau 5 : valeurs obtenues permettant de tracer la courbe d’étalonnage de glucose (0-120
mg eq glucose/l)
Concentrations (mg eq glucose)
20 40 60 80 100 120
DO 0,39 0,68 1,00 1,35 1,52 1,84
Les résultats obtenus dans la gamme de sucre sont représentés dans la figure suivante (figure
24).
EPS Témoin- La gamme glucose

Résultats et Discussion
76
De cette manière, la courbe étalon glucose a été établie.
Figure 24: courbe montrant les différentes concentrations du glucose (0-120mg/l) en fonction
de la densité optique (DO 490nm)
R2 : coefficient de corrélation ≥0.95 (liaison forte entre les 2 séries)
.
· Résultats du dosage colorimétrique des différents EPS
Au total, le dosage a été effectué pour 6 polymères extraits.
Figure 25: dosage colorimétrique des deux dilutions A (1/10) et B (1/100) des 6 EPS (E1, E2,
E3, E4, E5, E6)
y = 0,0788x
R² = 0,9873
0,00
0,20
0,40
0,60
0,80
1,00
1,20
1,40
1,60
1,80
2,00
0 5 10 15 20 25 30
Série1
Linéaire (Série1)
De
nsi
té O
pti
qu
e (
DO
49
0n
m)
A B E1 E2 E3 E4 E5 E6 E1 E2 E3 E4 E5 E6

Résultats et Discussion
77
Figure 26 : exemple de résultats des dosages des dilutions finales d’un échantillon EPS
(A : 1/10, B : 1/100, C : 1/200)
· Résultats des différentes concentrations des EPS
Les résultats obtenus des différents dosages nous ont permis de dresser le tableau suivant.
Le calcul des différentes concentrations a été effectué en se basant sur la formule y = 0.0788x
Tableau 6: Résultats des différentes concentrations des EPS exprimés en mg eq. glucose/l
Les résultats de dosage des différents échantillons d’EPS produits par les six isolats,
notamment LGM-Jb2, LGM-Jb1, LGM-IL3, LGM-Clo1, LGM-IL2 et LGM-PRO4 sont
représentés dans l’histogramme suivant.
Echantillons C équivalent glucose (mg/l) E1 (LGM-Jb2)
E2 (LGM-Jb1)
E3 (LGM-IL3)
E4 (LGM-Clo1)
E5 (LGM-IL2)
E6 (LGM-PRO4)
134 654 716 750 1264 978
A B C

Résultats et Discussion
78
Figure 27 : histogramme représentatif des concentrations des échantillons EPS des différents
isolats du tractus digestif
Les résultats de dosage des différents polymères obtenus montrent que les concentrations des
EPS varient d’un échantillon à un autre, selon la capacité de production de chaque souche.
Par ailleurs, la concentration la plus élevée produite est de 1264 mg/l obtenue avec la souche
LGM-IL2 pré-identifiée au genre Pediococcus sp. et la concentration la plus faible est celle
produite par la souche LGM-Jb2 pré-identifiée au genre Leuconostoc sp. qui est de l’ordre de
134 mg/l.
En général, les résultats obtenus des différentes concentrations des EPS produits à partir de
différents isolats de la collection LGM varient entre 134 mg/l et 1264 mg/l, ce qui concorde
avec celles produites par S. thermophilus d’une gamme de 50 à 350 mg/l (Cerning et al,
1988; Doco et al, 1990), celles produites par Lb bulgaricus de 60 à 150 mg/l (Cerning et al,
1986; Garcia-Garibay and Marshall,1991), et de 80 à 600 mg/l pour celles produites par Lc
lactis subsp cremoris (Cerning et al, 1992).
La quantité des EPS produite par différentes espèces varient considérablement selon la
littérature, d’autres auteurs ont trouvé des concentrations importantes et plus élevées (plus de
1200 mg/l), ce qui concorde avec la production d’EPS produit par la souche LGM-IL2 pré-
identifiée au genre Pediococcus (Van de Berg et al., 1995; De Vuyst and Degeest, 1999).
0
200
400
600
800
1000
1200
1400
E1 E2 E3 E4 E5 E6
E1
E2
E3
E4
E5
E6

Résultats et Discussion
79
2.9. Chromatographie sur couche mince des oses constitutifs des différents
polysaccharides
Les figure 28,29 et 30, laissent apparaitre les résultats d’analyse par chromatographie sur
couche mince des différents extraits de polysaccharides hydrolysés produits par les isolats
LGM pré-identifiés aux genres Leuconostoc et Pediococcus.
Figure 28: Profil de la composition en oses des différents EPS par analyse sur ccm dans le
système 1 (acide acétique : chloroforme : eau)
1 : Raff, 2 : Glu, 3 : Gal, 4 : Sac, 5 : Ara, 6 : Lac, 7: CLO1, 8: CLo1’, 9:IL2, 10:IL2’,
11:JAB1, 12:Co2, 13: IL3, 14:Pro4, 15: Dextrane
Les résultats obtenus montrent une similitude des profils chromatographiques des différents
extraits avec l’échantillon commercial le dextrane, dont on note la présence de 3 spots à
Rf1=0.112, Rf2=0.264 et Rf3=0.5.
Le profil des oses obtenu montre une prédominance du glucose qui est le constituant majeur
du dextrane.
Rf2
Rf3
Rf1
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15

Résultats et Discussion
80
L’analyse des différents oses constitutifs des polysaccharides a montré que les 6 échantillons
ainsi que le dextrane commercial contiennent principalement une tache dont le Rf2=.0264 et
qui correspond au glucose, on note aussi la présence d’autres taches dont le Rf1=0.112 et
Rf3=0.5 de nature non déterminée.
Cela laisse supposer que, soit l’hydrolyse des polysaccharides était partielle et a libéré
certainement différents tri ou disaccharides plus lourd que le glucose, soit ces échantillons
contiennent plusieurs composants de nature glucidique ou autre non déterminés de Rf
différents.
Calcul du facteur de rétention (Rf) des différents sucres et échantillon
Tableau 7 : les différent RF obtenus sur le système 1 (acide acétique /chloroforme/ eau)
Sucres standards
Rf Échantillons EPS Rf
Glu Ara Gal Sac Lac Raf
0.264 0.364 0.225 0.178 0.092 0.059
IL2 0.264 0.112
JAB1 0.264 0.112
CLO1
0.264 0.112
PRO2(4)
0.264 0.112
COL2(1)
0.264 0.112
IL2(3) 0.264 0.112
Dextrane commercial 0.264 0.112

Résultats et Discussion
81
Figure 29 : Profil de la composition en oses des différents EPS par analyse sur ccm dans le
système 1 (acide acétique : chloroforme : eau)
La figure 29 laisse apparaitre le profil des différents oses constitutifs de polysaccharides
analysés par chromatographie sur couche mince.
D’après la figure, on remarque une similitude des profils de différents échantillons (de 8 à 13)
avec le dextrane commercial (14) et notamment une diversité de composants révélés pour
chaque échantillon, on note principalement la présence d’une tache dont le Rf2=0.36 et qui
correspond à celle du glucose (piste 7).
Par ailleurs, on remarque la présence de différentes autres taches dont le Rf1=0.2 qui a migré
un peu moins que celui du glucose, ce qui laisse supposer selon les résultats obtenus par
Boual et al.( 2011) et Guillaume T. et al.(2006) qui montrent une migration d’acide
gluccuronique et d’acide galacturonique à un Rf juste en dessous de celui du glucose, donc
vraisemblablement les échantillons analysés contiennent ce composant. D’autre part, à un
Rf3=0.6 et un Rf4=0.73 qui montre des spots avec une migration plus importante, laisse
suggérer que ce sont des composants plus léger peut être des acides comme le montre le
témoin de la piste 1 (acide ascorbique).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16
Rf 2 Rf 3
Rf 4
Rf 1

Résultats et Discussion
82
Figure 30: Profile de la composition en oses des différents EPS par analyse sur ccm dans le
système 2 (Acétone: Eau)
L’analyse de cette chromatographie, montre une diversité dans les profils obtenus pour
chaque échantillon, dont on note principalement la présence chez touts les échantillons
analysés (6-10) ainsi que le dextrane commercial (11) d’une tache à Rf2=0.5 correspondant à
celle du glucose.
D’autres taches sont présentes chez quelques échantillons (9, 10, 11) dont le Rf1=0.20 et qui
est approximativement semblable à celui du raffinose (2) à Rf=0.24, cela laisse supposer que
ces échantillons n’ont pas subit une hydrolyse totale ce qui a engendré la migration de
composés complexe lourd à ce Rf.
Par ailleurs, la présence de taches à Rf3=0.7 et Rf4 =0.8 est remarquable pour touts les
échantillons analysés, et qui ont migré fortement prés du front de migration se localisent à un
niveau rapproché que celui l’acide ascorbic (1) utilisé dont le Rf=0.6. cela peut traduire que
ces échantillons contiennent d’autres composés de nature non glucidique (acide ou autre
dérivés).
Rf 1
Rf 2
Rf 3
Rf 4
1 2 3 4 5 6 7 8 9 10 11 12 13 14

Résultats et Discussion
83
Les résultats obtenus par les ccm réalisées lors de cette étude, semblent similaires à celles
d’autres auteurs qui ont monté que le glucose et le galactose sont les sucre neutres retrouvés
souvent dans la composition des polysaccharides des LAB (De Vuyst et al., 1998;
Cerning,1990, 1995, Nakajima et al., 1990; Grobben et al., 1995; Gruter et al., 1993).
Les monosaccharides les plus retrouvés dans divers EPS produits des LAB sont le rhamnose
(Cerning et al., 1986, 1988; Nakajima et al., 1990, 1992; Gruter et al., 1993; Grobben et al.,
1995), le mannose (Petit et al., 1991), le fructose (Manca de Nadra et al., 1985), l’arabinose
et le xylose (Cerning et al., 1988, 1992) ou les dérivées des sucres tels que le N-
acetylgalactosamine (Doco et al., 1990; Petit et al., 1991) et le N-acetylglucosamine (Cerning
et al., 1994) sont aussi retrouvés.
3. Etude de l’activité antimicrobienne des souches probiotiques de la collection LGM
L’activité antibactérienne se révèle par la présence d’une zone transparente autour des disques
imprégnés avec les cultures des souches à tester (figure 31). Les souches de la flore totale de
la collection développée lors de cette étude synthétisent une substance inhibitrice de nature
inconnue dirigée contre les pathogènes et qui pourrait vraisemblablement être une
bactériocine. L’activité antagoniste se traduit par la présence d’un halo clair autour du
probiotique notifiant la sensibilité du pathogène suite à l’élaboration de cette substance.
La présence d’une zone d’inhibition peut être due à la production des substances
antimicrobienne comme les bactériocines, des acides organiques, des peroxydes ou dioxyde
de carbone (Piard, Desmazeaud M,J ; 1992).
Au total 20 souches probiotiques ont été utilisées parmi eux des Leuconostoc (LGM-jb2,
LGM-Ca1), des pediocoques (LGM-CLo1, LGM-IL2) et notamment des Lactobacillus spp.
(LGM-CA7, LGM-IL2L, LGM-Jé2L, LGM-7G, LGM-4’L).
D’autres inhibitions ont été réalisées par d’autres bactéries non lactiques à l’exemple de
Bacillus isolé lors de ce travail (LMG-9Bc, LGM-7Bc, LGM-PrGBc, LGM-14EIB), nous
faisons remarqué que ce genre est utilisé comme probiotique en aviculture (Mutus et al.2006).
Ce travail a pu montrer qu’au niveau de la collection de souches élaborée, 11 isolats ont
exercé un effet inhibiteur pour les différents pathogènes utilisés, et cela par la méthode des
disques (Tadesse et al.,2004) et que par ailleurs, leur extrait (surnageant) a au même titre
engendré un effet neutralisant la croissance de ces pathogènes.
Les résultats de ces interactions sont illustrés dans les figures (figure31 au 39) et tableaux
suivants (tableau 8,9).

Résultats et Discussion
84
Tableau 8 : activité antibactérienne des différents isolats LGM contre des souches
pathogènes
pathogènes S.aureus
ATCC
25923
E.coli
ATCC
25922
S.aureus
ATCC
43300
Souches
probiotiques
LGM-CA7
LGM-IL2L
LGM-Jé2L
LGM-7G
LGM-4’L
LGM-Jb1o
LGM-IL2O
LGM-Clo1O
LGM-7Bc
LMG-9Bc
LGM-PrGBc
-
+
-
+
+
+
-
-
-
+
+
-
+
+
-
-
+
+
/
-
-
/
+
+
+
-
-
/
/
/
-
/
+
Selon les résultats montré sur le tableau 1, on note que la souche pathogène S. aureus ATCC
25923 a été inhibée par 6 cultures probiotiques parmi 11 utilisées, et que les souches E. coli
ATCC 25922 et S. aureus ATCC 43300 ont été inhibées seulement par 4 cultures différentes
de nature probiotique.
Ces résultats indiquent que les isolats LGM sont capables de synthétiser des substances
inhibitrices ayant une activité antibactérienne. L’ensemble des souches testées ne présentent
pas le même spectre d’action vis-à-vis des bactéries pathogènes notamment S. aureus ATCC
25923, E. coli ATCC 25922 et S. aureus ATCC 43300.
Par ailleurs, les LAB sont connues par la production d’une multitude de composés
antimicrobiens : les acides organiques, les bactériocines, le diacétyl et le peroxyde
d’hydrogène (Titiek et al.1996 ; Aslam et Qazi, 2010).
Charlier et al. (2009) ont montré que Lactococcus sp. et Leuconostoc sp. présentent une
inhibition à spectre élargie vis-à-vis de Staphylococcus aureus qui est induit par l’effet de
l’acide lactique et des bactériocines.
La capacité inhibitrice in vitro des LAB vis-à-vis des germes pathogènes semble être une
bonne propriété probiotique (Ammor et al., 2006).

Résultats et Discussion
85
Tableau 9 : effet inhibiteur des surnageants natifs
Les résultats du tableau 2 résument la résistance et la sensibilité des différentes souches
pathogènes testées aux composants des surnageants natifs de quelques isolats LGM.
Pathogènes S.aureus
ATCC
25923
E.coli
ATCC
25922
Salmonella
typhi
Souches
probiotiques
LGM-CA7
LGM-IL2L
LGM-Jé2L
LGM-7G
LGM-4’L
LGM-IL2O
LGM-Clo1O
LGM-Jb1o
LMG-9Bc
LGM-7Bc
LGM-PrGBc
LGM-14EIBc
-
-
-
+
+
ND
ND
ND
+
-
+
+
+
+
+
-
-
+
-
+
ND
ND
+
+
+
+
+
-
-
-
-
ND
ND
ND
ND
ND
+ : sensibilité, - : résistance, ND : non déterminée
D’après les résultats obtenues sur le tableau 2, on remarque que la souche Salmonella typhi a
été inhibée par l’effet de 3 surnageants, la souche S. aureus ATCC 25923 a été inhibée par
l’effet de 5 surnageants natifs, et que la souche E. coli ATCC 25922 a été inhibée par l’effet
de 7 autres surnageants différents.
Selon les résultats obtenus, on constate que l’activité antibactérienne des surnageants natifs
des différentes souches n’est pas semblable envers la collection des souches pathogènes. En
effet, les diamètres des zones d’inhibition varient d’une souche à une autre.
Il apparaît que les surnagenants des isolats LGM-CA7, LGM-IL2L, LGM-Jé2L n’arrivent pas
à inhiber la croissance de S. aureus ATCC 25923, alors qu’ils présentent une bonne activité
antimicrobienne contre E. coli ATCC 25922 et Salmonella typhi.
La fraction extracellulaire correspondant au surnageant présente un fort pouvoir antibactérien,
ce qui confirme la production d’agents antimicrobien par les souches lactiques dans le milieu.

Résultats et Discussion
86
Plusieurs études ont montré que la fraction extracellulaire contient des substance responsables
de cette interaction (Metlef et Bouras,2009) .
Les lactocoques sont capables de produire deux substances majeures (acide lactique et
bactériocines) responsables de l’effet antagoniste. Les LAB hétérofermentaires peuvent
produire des quantités notables d’acides organiques autres que l’acide lactique et c’est le cas
des leuconostocs qui produisent autant d’acétate que de lactate. L’acide acétique excerce une
forte action inhibitrice à l’encontre de nombreux micoorganismes (Bourgeois et
Larpents,1996).
Ø Effet du surnageant neutralisé
Après élimination de l’effet des acides organiques, les surangents neutres ont été testés au
nombre de 12 pour leur activité inhibitrice contre deux germes S .aureus ATCC25923 et E.
coli ATCC 25922.
Lés résultats obtenus (figure 36, 37) montrent une perte de l’activité inhibitrice pour les
surnageants utilisés, seulement deux parmi eux (souche LGM-Jé2L et LGM-14EI) sont
arriver a engendré une neutralisation à l’encontre des deux pathogènes testés.
Cela laisse supposer que l’inhibition majeure est due aux acides organiques produits par les
souches LGM. L’apparition de l’activité antagoniste pour les 2 souches LGM-Jé2L et LGM-
14EI, après élimination de l’effet de l’acide, confirme l’existence des autres substances
antibactériennes telles que le diacétyl, le peroxyde d’hydrogène et les bactériocines.
L’intervention du peroxyde d’hydrogène dans le phénomène d’inhibition par les bacteries
lactiques a été établie. Lorsqu’il est libéré à des concentrations suffisantes, il peut provoquer
l’auto-inhibition ou inhiber d’autres contaminants de l’environnement (Bourgeaois et Larpent,
1996).
Par ailleurs, les lactocoques produisent des bactériocines à spectre varié d’inhibition où deux
ont été caractérisées : la nisine produite par Lc. lactis spp. lactis a un spectre d’inhibition large
touchant la plupart des germes à gram positif, et la pedioccine produite par Lc. lactis spp.
cremoris (Bourgeois et Larpent, 1996 ; Campos et al., 2006).
Trias et al. (2008) ont observé un effet antimicrobien exercé par des souches de Leuconostoc
mesenteroides vis-à-vis de Listeria monocytogenes. Les acides organiques, le peroxyde
d’hydrogène et les bactériocines ont été détectés en tant que principaux mécanismes
d’inhibition.

Résultats et Discussion
87
Ø La méthode de la double couche (Fleming et al., 1975)
Cette méthode a été réalisée pour 10 souches appartenant au genre Lactobacillus spp.,
seulement 6 ont montré une inhibition vis-à-vis deux pathogènes d’Enterococcus faecalis
V583 et Enterococcus faecalis JH2-2 (figure 38,39). A ce titre, la méthode de double couche
n’a pas donné satisfaction de la démarche escomptée par rapport aux autres protocoles
réalisés.

Résultats et Discussion
88
Figure 32 : activité antagoniste des
différents isolats LGM vis-à-vis d’E. coli
ATCC25922 par méthode de disques.
Figure 31:activité antagoniste des différents
isolats LGM vis-à-vis S. aureus ATCC25923
par la méthode des disques
Figure 33: activité antagoniste des différentes souches LGM vis-à-vis S. aureus ATCC 43300 par la méthode
des disques

Résultats et Discussion
89
Figure 35:activité antagoniste des différents
surnageant natifs de différents isolats LGM
vis-à-vis S. aureus ATCC25923 par la méthode
des disques
Figure 34 : activité antagoniste des surnageants
natifs de différents isolats LGM vis-à-vis d’E.
coli ATCC2592 par la méthode des disques
Figure 36 : activité antagoniste des surnageants
neutralisés de différents isolats LGM vis-à-vis
d’ E. coli ATCC2592 par la méthode des disques
Figure 37 : activité antagoniste des surnageants
neutralisés de différents isolats LGM vis-à-vis S.
aureus ATCC25923 par la méthode des disques
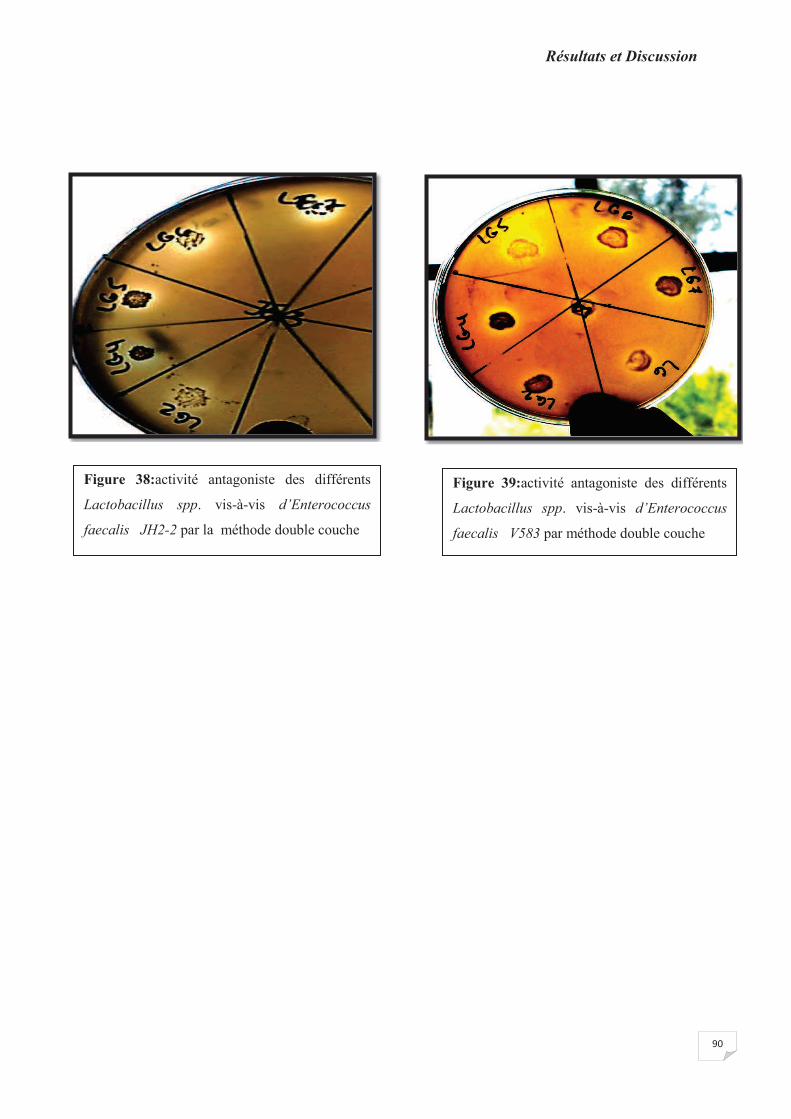
Résultats et Discussion
90
Figure 39:activité antagoniste des différents
Lactobacillus spp. vis-à-vis d’Enterococcus
faecalis V583 par méthode double couche
Figure 38:activité antagoniste des différents
Lactobacillus spp. vis-à-vis d’Enterococcus
faecalis JH2-2 par la méthode double couche

Résultats et Discussion
91
4. Caractérisation moléculaire des souches probiotiques
4.1. Identification du genre Lactobacillus spp. par amplification d’une région interne du
gène ADNr 16S
Une bande d’ADN de 480 pb a été amplifié pour les 13 souches de lactobacilles ainsi que
pour la souche de référence Lactobacillus delbruekii ATTC 11842 utilisée témoin positif,
aucune présence de bande significative n’a été constatée pour la souche Streptococcus
thermophilus CNRZ 1066 de référence utilisée comme témoin négatif (figure 40).
Figure 40 : Amplification du fragment ADN 480 pb du gène de l’ADNr 16S en
électrophorèse sur gel d'agarose 0.8 %.
M: marqueur de taille 200 pb (Smart Ladder), piste 1: témoin positif (Lb. delbruckii ATTC
11842), pistes 2.3.4.5.6.7.8.10.11.12.13.14.15: (isolats lactobacilles), piste 9: témoin négatif
(St. thermophilus CNRZ 1066)
4.2. Séquençage et taxonomie des souches de la flore intestinale
Le séquençage des différents amplifias obtenu a été réalisé et les scores affichés lors de
traitement des séquences par le BLAST via internet ont montré une forte homologie avec les
séquences déposées dans la banque de donnée. Cette forte homologie aux environs de 99% de
similitude entre séquences a notifié la présence de Lb. acidophilus, Lb. gallinarum, Lb.
Crispatus, Lb. Johnossi, Lb. Fermentum et Lb. Helveticus pour la majorité des isolats
examinés. A ce degré d’investigation, les souches sélectionnées ne peuvent être affectées au
niveau de l’espèce et se regroupent dans le genre Lactobacillus spp.
M 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
480 pb
400 pb
200 pb

Résultats et Discussion
92
4.3. Amplification du gène de l’ADNr 16S à base de PCR de quelques isolats
On note la présence de bande d’ADN de poids moléculaire à 1500 pb comparativement aux
bandes du marqueur de taille Smart Ladder 200 pb (figure 41).
L’ADNr 16S reste un taxon de choix en matière de systématique en bactériologie.
Le séquençage de cette bande permettrait de donner une affiliation résolutive de l’espèce une
fois que cette séquence sera traitée au niveau d’une base de données par logiciel BLAST.
Figure 41 : Amplification du fragment ADN 1500 pb du gène de l’ADNr 16S en
électrophorèse sur gel d'agarose 1.3 %.
M : marqueur de taille 200 pb ( Smart Leader), piste 1,2 : S1,S2 (Streptomyces), piste 3 :
N1(lactobacille), piste 4 :R1(Rhodococcus), piste 5 : V1 (leuconostoc) , piste 6 :L2 (tetra
pediococcus ), piste 7 : S3( Streptomyces), piste 8,9 : R2,R3 (Rhodococcus), piste 10 : L3
(leuconostoc), piste 11 : L4 (lactobacile )
Au total 11 souches du laboratoire ont fait l’objet d’une extraction ADN (Gevers et al., 2001)
et une amplification PCR par l’usage d’amorces universelles ADNr 16S (8UA/1492R) et
l’amplifiât est estimé à 1500 pb.
Au total quatre souches appartenant à des genres différents ont montré des amplifias à 1500
pb. La méthode d’extraction ADN peut être optimisée pour les souches n’ayant pas montré un
résultat positif.
1 2 3 4 5 6 7 8 9 10 11 M
M
1500 pb
1000 pb
800 pb
600 pb
400 pb

Résultats et Discussion
93
Dans le contexte de cette étude, le genre Pediococcus produisant des EPS a révélé la présence
de l’amplifiât ADN (piste 6).
L’amplifiât chez les quatre souches a une taille de 1500 pb, ces résultats concordent avec ceux
de la littérature (Sato et al.,2003) .
Il serait intéressant de séquencer la bande ADN afin de confirmer d’une manière résolutive la
taxonomie de ces souches.
La méthode RFLP par digestion de l’ADNr 16S totale avec des enzymes de restrictions
permettait l’obtention de profils de digestion dont l’analyse trancherait quand à la similitude
et l’homologie des séquences ADN des souches du même genre et aiderait à les différencier.

Conclusion et perspectives
94
La microflore présente au niveau du microbiote intestinal montre une diversité enrichissante
de microorganismes dont le profil phénotypique peut être exploité dans le domaine de lutte
contre les pathogènes et aussi par la production de polysaccharides en qualité de prébiotique.
Les souches isolées ont été identifiés au genre Lactobacillus par l’amplification d’un fragment
de 480 pb, ces résultats concordent avec ceux de Dubernet et al. (2002) qui, par
l’amplification d’une région partielle de 250 pb, entre les régions ADNr16S et ADNr 23S ont
démontré l’appartenance de leurs souches au genre Lactobacillus. Les travaux réalisés dans
cette étude sur les inhibitions ont montré que les lactobacilles du tractus digestif se
caractérisent par un large spectre d’action antagoniste à l’égard de pathogènes utilisés en
l’occurrence Staphylococcus aureus et Salmonella typhi. Les études de Makras et al. (2006),
ont déjà confirmé l'activité antagoniste des bactéries lactiques contre les bactéries pathogènes.
Les propriétés antibactériennes des LAB dépendent de plusieurs critères: le pH, le milieu de
croissance et la production de substances antibactériennes comme les bactériocines, les acides
organiques, les acides gras et le peroxyde d'hydrogène (Huttunen et al., 1995). L'acidification
du milieu par l'acide lactique joue un rôle considérable dans la capacité inhibitrice des LAB
contre plusieurs bactéries pathogènes. Hicks et Goepfert (1968) ont bien confirmé que l'acide
lactique et l'acide acétique produits par les bactéries lactiques participent à l'inhibition de
Staphylococcus aureus et les travaux de Sakaridis et al. (2012) ont montré que les LAB se
caractérisent par une forte inhibition vis-à-vis de Salmonella sp. et de Listeria monocytogenes.
Récemment de nouvelles stratégies de prévention ont été proposées comme alternatives aux
antibiotiques pour réduire l’incidence des pathogènes entériques chez la volaille, parmi ces
stratégies, le recours aux probiotiques et prébiotiques semblent offrir les résultats les plus
prometteurs. Notre travail nous a permis de révéler la présence d’une flore digestive riche et
diverse où on retrouve les différents groupes de probiotiques cités dans la littérature
(Bernardeau et Vernoux, 2009 ; Capcarova et al., 2011), à l’exemple des lactobacilles, des
pediocoques, des leuconostoc, des streptocoques, des entérocoques, des Bacillus et
notamment des levures.
Par ailleurs, l’ADNr 16S a permis d’identifier un premier groupe appartenant au genre
Lactobacillus spp., au nombre de 22 dont 13 ont été identifiés par les techniques de biologie
moléculaire, un deuxième groupe dont 06 en forme de coques ovoïdes et longues chainettes ,
pré-identifiée au leuconostoc, 02 en forme de tétracoques pré-identifiés au pédiocoques, 06 en
forme coques en amas pré-identifiés au groupe des entérocoques et 03 en forme de longues
chainettes coques pré-identifiés à Streptococcus thermophilus, et enfin un troisième groupe

Conclusion et perspectives
95
non lactique au nombre de 09 dont 04 levures et 05 Bacillus. Au total 48 souches ont été
répertoriées et conservées.
Les EPS qui ont été repérés chez certaines souches en particulier les leuconostocs et les
pédiocoques ont fait l’objet d’une extraction, purification, et dosage après hydrolyse en
présence d’acide sulfurique, les résultats sont fort intéressants dont le dosage de la production
la plus élevée en EPS à été déterminé par 1264 mg/l, ces travaux concordent avec Van de
Berg et al., 1995 et De Vuyst and Degeest, 1999 .
D’autre part, la chromatographie sur couche mince de différents EPS extrait a conduit à une
similitude des profils avec le dextrane commercial, en montrant une diversité de dérivés dans
le profil ccm. Ces résultats sont en convergence avec ceux de De Vuyst et al., 1998 et
Cerning,1990.
En matière de perspective, en premier plan, un diagnostic moléculaire à base de PCR et
séquençage s’avère plus que nécessaire afin de déterminer l’exactitude de la taxonomie des
bactéries les plus performantes. En second plan, ces souches peuvent faire l’objet d’études
ultérieures dans l’alimentation avicole, afin de valoriser leur potentialité en qualité de
probiotique, en étudiant la prévention de maladies entériques dues à des pathogènes et par la
même occasion, évaluer leurs aptitudes dans le contexte des performances zootechniques et
prévention sanitaire à l’exemple de la souche Pediococcus LGM-IL2 dont les performances
ont été testées par plusieurs auteurs à l’exemple de Pediococcus acidilactici (MA 18/5M)
(Temim et al.,2009).
Les souches productrices d’exopolysaccharides méritent un suivi et doivent être exploitées
pour une éventuelle caractérisation et production de ces prébiotiques. Il a été montré que les
Mannan-oligosaccharides présentent un effet de protection contre les salmonelles chez la
volaille ainsi que les AXOS ( Arabinoxhylo-oligosaccharides) qui ont montré le même effet
de protection de la colonisation de l’épithélium, en plus l’excrétion des salmonelles a été
retrouvée dans les fientes après un aditif de ces prébiotiques (Fukata et al.,1999). Des travaux
peuvent être dirigés contre certaines pathologies, en l’occurrence les coccidioses.
Selon plusieurs chercheurs, les probiotiques et prébiotiques sont bien placés pour prendre la
relève des antibiotiques en raison de leurs aptitudes nutritionnelles et antimicrobiennes fort
intéressantes.

Alvarez-Olmos M. I. and R. A. Oberhelman ,2001. Probiotic agents and infectious diseases: A modem perspective on a traditional therapy.Clinical Infectious Diseases 32(11): 1567-1576.
Ammor S., Tauveron G., Dufor E. et Chevalier I., 2006. Antibacterial activity of lactic acid bacteria against spoilage and pathogenic bacteria isolated from the same meat small-scale facility 1- Screening and characterization of antibacterial compound. Food Control. 17 : 454-461.
Andreatti Filho R. L., Da Silva, E. N., Ribeiro, A. R., Kendo, N., Curi, P. R., 2000. Use of anaerobic cecal microflora, lactose and acetic acid for the protection of broiler chicks against experimental infection with salmonella typhimurium and salmonella enteritidis. Braz. J. Microbiol., 31:107-112.
Apajalahti J., Kettunen A., Graham H., 2004. Characteristics of the gastrointestinal microbial communities, with special reference to the chicken. World’s Poult. Sci. J., 60, 223-232
Applegate T.J., and Angel, R., 2005. Feasibility versus practicality of phosphorus reduction in poultry: progress and future needs. Symposium State of the Science Animal Manure and Waste Management. Annual Report.
Asahara T, Nomoto K, Shimizu K, Watanuki M, Tanaka R. 2001. Increased resistance of mice to Salmonella enterica serovar Typhimurium infection by synbiotic administration of bifidobacteria and transgalactosylated oligosaccharides. J Appl Microbiol. 91: 985-996.
Asahara T, Shimizu K, Nomoto K, Hamabata T, Ozawa A, Takeda Y. 2004. Probiotic bifidobacteria protect mice from lethal infection with shiga toxin-producing Escherichia coli O157:H7. Infect Immun. 72: 2240-2247.
Aslam S. et Qazi J.I., 2010. Isolation of acidophilic lactic acid bacteria antagonistic to microbial contaminants. Pakistan. J. Zool. 42(5) : 567-573.
Audisio M. C, G. Oliver, et al. (2000). "Protective effect of Enterococcus faecium J96, potential probiotic strain, on chicks infected with Salmonella pullorum." Journal of Food Protection 63(10): 1333-1337.
Axelsson L., 2004. Classification and physiology. In : Lactic acid bacteria: Microbiological and functional aspects ((Salminen S., Wright A.V. et Ouwehand A.). 3e Ed., Marcel Dekker,
Inc. New York. 1-66.

Bailey J.S., Blankenship L.C., Cox N.A., 1991. Poultry Sci. 70, 2433-2438.
Béal C., Marin M., Fontaine E., Fonseca F. et Obert J.P., 2008. Production et conservation des ferments lactiques et probiotiques. In : Bactéries lactiques, de la génétique aux ferments (Corrieu G. et Luquet F.M.). Tec & Doc, Lavoisier. Paris. 661-765.
Bensalah F., Delorme C., Renault P., 2009.Characterisation of thermotolerant cocci from indigenous flora of 'leben' in algerian arid area and DNA identification of atypical Lactococcus lactis strains. Curr Microbiol. 59(2):139-46.
Bernardeau M, Vernoux JP Gueguen M., Smith DG, Corona-Barrera E., , 2009. Antagonistic activities of two Lactobacillus strains against Brachyspira. Vet Microbiol. 138(1-2); 184-190.
Bernardeau M, Gueguen M & Vernoux JP (2006) Beneficial lactobacilli in food and feed: long-term use, biodiversity and proposals for specific and realistic safety assessments. FEMS
Microbiol Rev. 2006 Jul;30(4):487-513.
Bernet MF, Brassart D, Neeser JR, Servin AL. 1993. Adhesion of human bifidobacterial strains to cultured human intestinal epithelial cells and inhibition of enteropathogen-cell interactions. Appl Environ Microbiol. 59: 4121-4128
Borriello SP, Hammes WP, Holzapfel W, Marteau P, Schrezenmeir J, Vaara M, Valtonen V. 2003. Safety of probiotics that contain lactobacilli or bifidobacteria. Clin Infect Dis. 36: 775-780.
Boual Z, KEMASSI A., MICHAUD P. et OULD EL HADJ M. D.2011. Caractérisation partielle des polysaccharides hydrosolubles des feuilles d'Asphodelus tenuifolius Cavan (Liliaceae): effet prébiotique des oligosaccharides issus de l'hydrolyse des polysaccharides. Algerian journal of arid environment ,vol. 1: 52-60.
Bourgeois C.M. et Larpent J.P., 1996. Microbiologie alimentaire : Aliments fermentés et fermentations alimentaires. Tec & Doc, Lavoisier. Paris. 432-704.
Bouzar F., Cerning J. et Desmazeaud M., 1997. Exopolysaccharide production and texturepromoting abilities of mixed strain starter cultures in yogurt production. J. Dairy Sci. 80 : 2310- 2317.
Bouzar , Fm, Cerning, J. and Desmazeaud, M. (1 995) Exopolysaccharide production in milk by Lactobacillus delbnreckii ssp. bulgancus CNRZ 1187 and by two colonial variants. Journal of Daity Science 79, 205-21 1.
Bruno FA, and Shah NP. 2002. Inhibition of pathogenic and putrefactive microorganisms by Bifidobacterium sp. Milchwissenschaft. 57: 617-621.

Campos C.A., Rodriguez O., Calo-Mata P., Prado M. et Barros-Velazquez J. 2006. Preliminary characterization of bacteriocins from Lactococcus lactis, Enterococcus faecium
and Enterococcus mundtii strains isolated from turbot (Psetta maxima). Food Res. Int. 39 : 356-364.
Candela M, Seibold G, Vitali B, Lachenmaier S, Eikmanns BJ, Brigidi P. 2005. Real-time PCR quantification of bacterial adhesion to Caco-2 cells: Competition between bifidobacteria and enteropathogens. Res Microbiol. 156: 887-895
Caramia G. 2004. Probiotics: from Metchnikoff to the current preventive and therapeutic possibilities. Pediatr Med Chir. 26(1):19-33.
Casey PG, Casey GD, Gardiner GE, Tangney M, Stanton C, Ross RP, Hill C & Fitzgerald GF (2004) Isolation and characterization of anti-Salmonella lactic acid bacteria from the porcine gastrointestinal tract. Lett ApplMicrobiol 39: 431-438.
Cerning J., Bouillane, C., Desmazeaud, M.J. and Landon, M. (1988) Exocellular polysaccharides production by Streptococcus thennophilus. Biotechnology Lefters I O , 255-260.
Cerning J., Bouillane, Cm, Desmazeaud, M.J. and Landon, M. (1986) Isolation -and characterization of exocellular polysaccharides production by Lacfobacillus bulgaticus. Biotechnology Letfers 8, 625-628.
Cerning J., C. Bouillanne, M. Landon, and M. J. Desmazeaud. 1992. Isolation and characterization of exopolysaccharides from slime-forming mesophilic lactic acid bacteria. J. Dairy Sci. 75:692- 699.
Cerning J., 1990. Exocellular polysaccharide produced by lactic acid bacteria. Microbiol.
Rev. 87: 113-130.
Cerning J (1995). Production of exopolysaccharides by lactic acid bacteria and dairy propionibacteria. Lait 75: 463-472.
Cerning J, Renard CMGC, Thibault JF, Bouillanne C, Landon M, Desmazeaud M, Topisirovic L (1994). Carbon source requirements for exopolysaccharide production by Lactobacillus casei CG11 and partial structure analysis of the polymer. Appl. Environ. Microbiol. 60: 3914-3919.
Chafai S. (2006). Effet de l'addition des probiotiques dans les régimes alimentaires sur les performances zootechnique du poulet de chair.
Chamba F.J., 2008. Application des bactéries lactiques lors des fabrications fromagères. In: Bactéries lactiques de la génétique aux ferments (Corrieu G. et Luquet F.M.). Tec & Doc,
Lavoisier. Paris. 787-813.

Champagne C. P., N. J. Gardner, et al. (2005). "Challenges in the addition of probiotic cultures to foods." Critical Reviews in Food Science and Nutrition 45(1): 61-84.
Chang M. H., and Chen, T.C., 2003. Reduction of broiler house malodor by direct feeding of a lactobacilli containing Probiotics. Poult. Sc., 2 (5): 313-317.
Charlier C., Cretenet M., Even S. et Le Loir Y., 2009. Interactions between Staphylococcus
aureus and lactic acid bacteria: An old story with new perspectives. Int. J. Food Microbiol.
131 : 30-39.
Chauviere G, Coconnier MH, Kerneis S, Fourniat J, Servin AL. 1992. Adhesion of human Lactobacillus acidophilus strain LB to human enterocyte-like Caco-2 cells. J Gen Microbiol. 138: 1689-1696.
Cole C. B. and R. Fuller (1984). "Bile acid deconjugation and attachment of chicken gut bacteria: Their possible role in growth depression." British Poultry Science 25(2): 227-231.
Conway P. L., 2001. Prebiotics and human health: The state-of-the-art and future perspectives. Scand. J. Nutr., 45:13-21.
Crociani J, Grill JP, Huppert M, Ballongue J. 1995. Adhesion of different bifidobacteria strains to human enterocyte-like Caco-2 cells and comparison with in vivo study. Lett Appl Microbiol. 21: 146-148.
Cummings J. H. and Kong, S. C., 2004. Probiotics, prebiotics and antibiotics in in£ammatory bowel disease Infammatory bowel disease_crossroads of microbes, epithelium and immune systems. Wiley, Chichester (NovartisFoundation Symposium263): 99-114.
Dalloul RA, Lillehoj HS, Shellem TA & Doerr JA (2003) Intestinal immunomodulation by vitamin A deficiency and Lactobacillus-based probiotic in Eimeria acervulina-infected broiler chickens. Avian Dis 47: 1313- 1320.
De Oliveira G. H., A. Berchieri, et al. (2000). "Prevention of Salmonella infection by contact using intestinal flora of adult birds and/or a mixture of organic acids." Brazilian Journal of Microbiology 31(2): 116-120.
De Vuyst L, Degeest B (1999). Heteropolysaccharides from lactic acid bacteria. FEMS Microbiol. Rev. 23: 153-177.
De Vuyst L, Vanderveken F, Van De Ven S, Degeest B (1998). Production by and isolation of exopolysaccharides from Streptococcus thermophilus grown in a milk medium and evidence for their growth-associated biosynthesis. J. of Appl. Micobiol. 84: 1059- 1068.
Degeest B, Vaningelgem F, De Vuyst L (2001a) Microbial physiology, fermentation kinetics, and process engineering of heteropolysaccharide production by lactic acid bacteria. International Dairy Journal II: 747-757

Deng Y, Ryu JH, Beuchat LR. 1999. Tolerance of acid adapted and non adapted Escherichia coli O157:H7 cells to reduced pH as affected by type of acidulant. J Appl Microbiol. 88: 203-210.
Denis O. Krause, James D. House, and Nyachoti, C. M., 2004. Alternatives to antibiotics in swine diets: a molecular approach. Department of Animal Science. University of Manitoba. Canada..
Deutsch SM, Falentin H, Dols-Lafargue M, Lapointe G, Roy D (2008) Capsular exopolysaccharide biosynthesis gene of Propionibacterium freudenreichii subsp. shermanii. International Journal of Food Microbiology 125: 252-258
Doco T., J. M. Wieruszelski, B. Fournet, D. Carcano, P. Ramos, and A. Loones. 1990. Structure of an exocellular polysaccharide produced by Streptococcus thermophilus. Carbohydr. Res. 198:313- 321.
Dubois M., K. A. Gilles, J. K. Hamilton, P. A. Rebers, and F. Smith. 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28:350–356.
Dunne C, L. O'Mahony, et al. (2001). "In vitro selection criteria for probiotic bacteria of human origin: correlation with in vivo findings." American Journal of Clinical Nutrition 73(2): 386s-392s.
Edens F.W., 2003. An alternative for antibiotics use in poultry: Probiotics. Rev. Bras. Cienc. Avic.,Vol.5. N°.2
Evans LR and Linker M : Production and characterization of the slime polysaccharides of pseudomonas aeruginosa. J Bact, 116:915-924, 1973.
FAO 2004 Agriculture mondiale : horizon 2015-2030
FAO/WHO (2002). Joint working group report on guidelines for the evaluation of probiotics in food. London, Ontario, Canada.
FAO/WHO 2004. Health and Nutritional Properties of Probiotics in food including powder Milk with Live Lactic acid Bacteria.
Farner D. S. (1942). "The hydrogen ion concentration in avian digestive tracts." Poultry Science 21: 445-450
Ferket P. R., Parks,C. W., and Grimes, J. L., 2002. Benefits of dietary antibiotic and mannanoligosaccharide supplementation for poultry. Department of Poultry Science. North Carolina State University

Fernandez F., Hinton M., Van Gils B., 2000. Avian Pathol. 29, 575-581.
Feuillet L. (2007). Étude comparative des vaccins et des flores bactériennes dans la lutte contre les Salmonelles en élevage de poules pondeuses. Médecine vétérinaire. Alfort, École Nationale vétérinaire d'Alfort: 17-21.
Fleming H.P., Etchells J.L. et Costilow R.N., 1975. Microbial inhibition of isolates of pediococcus from cucumber brine. Appl. Env. Microbiol. 30 : 1040-1042.
Fooks L.J. and Gibson, G. R., 2002. Probiotics as modulators of .the gut flora. Brit. J. Nutr., 88, suppl. I: 39-49.
Fritts C.A., Kersey J.H., Motl M.A., Kroger E.C., Yan F., Si J., Jiang Q., Campos M.M., Waldroup A.L.,Waldroup P.W., 2000. Bacillus subtilis C-3102 (Calsporin) improves live performance and microbiological status of broiler chickens. J. Appl. Poultry Res., 9 : 149-155.
Fujimori S, Tatsuguchi A, Gudis K, Kishida T, Mitsui K, Ehara A, Kobayashi T, Sekita Y, Seo T, Sakamoto C. 2007. High dose probiotic and prebiotic cotherapy for remission induction of active Crohn's disease. J Gastroenterol Hepatol 22: 1199-1204.
Fukata T., Sasai, K.,Amiymoto, T. and Baba, E. 1999 .Inhibitory effects of competitive exclusion and fructo-oligosaccharide, singly and in combination, on Salmonella colonization of chicks. Journal of Food Protection 62: 229-233.
Fuller R., 1989. Probiotics in man and animals. J. Appl. Bacteriol., 66(5):365-378.
Fuller R., 1984. Microbial activity in the alimentary tract of birds. Proc. Nutr. Soc., 43, 55-61
Gabriel I., Mallet, S., Lessire, M., 2003. La microflore digestive : une composante oubliée de la nutrition des volailles. Cinquièmes journées de la recherche avicole. Tours
Gabriel I., Mallet S., Leconte M., Fort G., Naciri M., 2003. Effects of whole wheat feeding on the development of coccidial infection in broiler chickens. Poult. Sci., 82, 1668-1676.
Gabriel I., Mallet, S., Sibille, P., 2005. La microflore digestive des volailles : facteurs de variation et conséquences pour l’animal. INRA. Prod. Anim., 18 (5) : 309-322.
Gaggîa F., P. Mattarelli, et al. "Probiotics and prebiotics in animal feeding for safe food production." International Journal of Food Microbiology In Press, Corrected Proof.
Gagnon M. (2007). Rôle des probiotiques lors d'infection entériques d'origine bactérienne et virale : anlyses in vitre et études in vivo chez des modèles murins. Département des sciences des aliments et de nutrition Québec. Université Laval. Ph.D: 155.
Gancel Fey Novel, G., Carcano, D., Loones, A., Ramos, P. 1988. Procédé de sélection de clones bactériens producteurs d'exopolysaccharides et clones producteurs obtenus. Brevet No: R 2 632 968 - Al. France 88 08009.

Garcia-Garibay M., Marshall, V.M.E. 1991. Polymer production by Lactobacillus delbrueckii ssp. bulgancus. Journal of Applied Bacteriology. 70 : 325-328.
Gevers D, Huys G, Swings J .2001.Applicability of rep-PCR fingerprinting for identification of Lactobacillus species. FEMS Microbiol Lett 205:31-36.
Gibson G. R., Probert, M, H., Loo, V, J., Rastall, A, R., and Roberfroid, B, M., 2004. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutrition Research Reviews., 17: 259–275.
Gibson G. R., and Fuller, R., 2000. Aspects of in vitro and in vivo research approaches directed toward identifying probiotics and prebiotics for human use. J. Nutr., 130: 391–395.
Gibson G.R., Roberfroid, M.B., 1995. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J Nutr., 125(6): 1401 1412.
Gionchetti P., Rizzello F., Venturi A., Campieri M., 2000. Probiotics in effective diarrhea and inflammatory bowel diseases. J. Gastroen Hepatol., 15 : 489-493
Gopal PK, Prasad J, Smart J, Gill HS. 2001. In vitro adherence properties of Lactobacillus rhamnosus DR20 and Bifidobacterium lactis DR10 strains and their antagonistic activity against an enterotoxigenic Escherichia coli. Int J Food Microbiol. 67: 207-216.
Graber M, André MorinCorresponding author contact information, Francis Duchiron, Pierre F. Monsan,Microbial polysaccharides containing 6-deoxysugars, 1988 ,Enzyme and Microbial Technology Volume 10, Issue 4, , Pages 198–206
Grobben GJ , Sikkema J , Smith MR , De Bont JAM (1995). Production of extracellular polysaccharides by Lactobacillus delbrueckii ssp bulgaricus NCFB 2772 grown in a chemically defined medium. J. Appl. Bacteriol 79: 103-107
Gruter M, Leeflang BR, Kuiper J, Kamerling JP, Vliegenthart JFG. (1993). Structural characterization of the exopolysaccharide produced by Lactobacillus delbrueckii subsp bulgaricus rr grown in skimmed milk. Carbohyd. Res. 239: 209-226
Gueimonde M. and S. Salminen (2006). "New methods for selecting and evaluating probiotics." Digestive and Liver Disease(38): S242-S247.
Guillaume T, Catherine M., Jean-Pierre J., Bernard D., Florence C-B. 2006. The eps locus of Streptococcus thermophilus IP6756 is not involved in exopolysaccharide production. International Dairy Journal 16 :467–473.
Guiraud J.P. et Rosec J.P., 2004. Pratique des normes en microbiologie alimentaire. AFNOR. 237-251.
Guiraud J.P., 2003. Microbiologie Alimentaire. Tec & Doc, Dunod. Paris. 90-292.

Haddie J.M., 1986. Other streptococci. In : Bergey's manual of systematic bacteriology (Sneath P.H.A., Mair N.S., Sharpe M.E., Holt J.G.W. et Baltimore W.). 1 : 1070.
Hassan A.N. et Frank J.F., 2001. Starter Cultures and their use. In: Applied Dairy Microbiology (Marth E.H. et Steele J.L.) 2e Ed., Marcel Dekker Inc. New York. 151-205.
Havenaar R, and Huis in’t Veld JHJ. 1992. Selection of strains for probiotic use. In: Fuller R (Ed). Probiotics, the scientific basis. Chapman & Hall, pp.209-224.
Higgins S. E., J. P. Higgins, et al. (2008). "Evaluation of a Lactobacillus-Based Probiotic Culture for the Reduction of Salmonella Enteritidis in Neonatal Broiler Chicks." Poultry Science 87(1): 27-31.
Ho T.N.T., N. Tuan N., Deschamps A. et Caubet R., 2007. Isolation and identification of lactic acid bacteria (LAB) of the Nem Chua fermented meat product of Vietnam. Int.
Workshop on Food Safety and Processing Technology. 134-142.
Holzapfel WH, and Schillinger U. 2002. Introduction to pre- and probiotics. Food Res Int.
35: 109-116.
Hütt P Shchepetova J, Loivukene K, Kullisaar T, Mikelsaar. 2006. Antagonistic activity of probiotic lactobacilli and bifidobacteria against entero- and uropathogens. J Appl Microbiol. 100: 1324-1332
Iji P.A., Tivey D.R., 1998. World’s Poultry Sci. J. 54, 129-143.
Ishihara N., Chu D.C., Akachi S., Jujena L.R., 2000. Poultry Sci. 79, 689-697.
Isolauri E., Sütas, Y., Kankaanpää, P., Arvilommi, H., and salminen, S., 2001. Probiotics: effects on immunity. Am. J. Clin. Nutr., Vol. 73, No. 2, 444-450
Ivanković S., Kralik, G., Milaković, Z., Bogut, I., 1999. Effect of the probiotic vebac. Acta. Agraria. Kaposvariensis., Vol 3. No 2: 353-360.
Jin l. Z., Ho,Y. W., Abdullah, N., and jalaludin, S., 2000. Digestive and bacterial enzyme activities in broilers fed diets supplemented with lactobacillus cultures. Poult. Sci., 79: 886-891.
Jin L.Z., Ho Y.W., Abdullah N., Jalaludin S., 1997. Probiotics in poultry : modes of action. World’ s. Poult. Sci. J., 53 : 351-368.
Jolly L, Vincent SJF, Duboc P, Nesser JR (2002) Exploiting exopolysaccharides from lactic acid bacteria. Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology 82: 367-374

Jordan F. T. W. and M. Pattison (1996). Poultry diseases. S. company. London: 38-43.
Karaoglu M., and Durdag, H., 2005. The Influence of Dietary Probiotic (Saccharomyces cerevisiae) Supplementation and Different Slaughter Age on the Performance, Slaughter and Carcass Properties of Broilers. International Journal of Poult. Sc., 4 (5): 309-316.
Khalid N.M. et Marth E.H., 1990. Lactobacilli, their enzymes and role. In: Ripening and spoilage of cheese. Rev. Dairy Sci. 73 : 158-167.
Klaenhammer TR. 1993. Genetics of bacteriocins produced by lactic acid bacteria. FEMS Microbiol Rev. 12: 39-85.
Klaenhammer T. R. and M. J. Kullen (1999). "Selection and design of probiotics." International Journal of Food Microbiology 50( 1 -2): 45-57.
Kralik G., Milaković, Z., Ivanković, S., 2004. Effect of probiotic supplementation on the performance and the composition of the intestinal microflora in broilers. Acta. Agraria Kaposvariensis ., 8(2): 23-31.
Kuçukersan K., Tuncer,S.D., Sanli, Y., Midilli, M., Goncuoglu, E., Kuçukersan, S., and Tan, H., 2002. The effects of dietary stabilized rumen extract (SRE) and virginiamycine on performance and carcass yield of broilers. Méd. Vét., 153(11) : 723-726.
La Ragione RM, Narbad A, Gasson MJ & Woodward MJ (2004) In vivo characterization of Lactobacillus johnsonii FI9785 for use as a defined competitive exclusion agent against bacterial pathogens in poultry. Lett Appl Microbiol 38: 197-205.
Lacroix C, and Yildirim S. 2007. Fermentation technologies for the production of probiotics with high viability and functionality. Curr Opin Biotechnol. 18: 176-183.
Larbier M, and B. Leclercq (1992). Nutrition et alimentation des volailles. Paris.
Larkin TA, Astheimer LB, Price WE. 2007. Dietary combination of soy with a probiotic or prebiotic food significantly reduces total and LDL cholesterol in mildly hypercholesterolaemic subjects. Eur J Clin Nutr.
Le Blay G., Michel C., Blottière H.M., Cherbut C., 1999. J. Nut. 129, 2231-2235
Le Blay G, Fliss I, Lacroix C. 2004. Comparative detection of bacterial adhesion to Caco-2 cells with ELISA, radioactivity and plate count methods. J Microbiol Meth. 59: 211- 221.
Lecoindre P., (2000) Prolifération bactérienne chronique intestinale, Prat Méd Chir Anim Comp, 35, 511 - 514

Lee S, Lillehoj HS, Park DW, Hong YH, Lin JJ. 2007. Effects of Pediococcus- and Saccharomyces-based probiotic (MitoMax) on coccidiosis in broiler chickens. Comp Immunol Microbiol Infect Dis. 30(4):261-8.
Lee K.W., Lee, S. K. and Lee, B. D.; 2006. Aspergillus oryzae as Probiotic in Poultry. Poult. Sci., 5 (1): 01-03.
Leveau J.Y., Boiux M. et De Roissart H.B., 1991. La flore lactique : technique d’analyse et
de contrôle dans les industries agro alimentaires. 2e Ed., Tec & Doc, Lavoisier. Paris. 3 : 2-40.
Liévin V, Peiffer I, Hudault S, Rochat F, Brassart D, Neeser JR, Servin AL. 2000. Bifidobacterium strains from resident infant human gastrointestinal microflora exert antimicrobial activity. Gut. 47: 646-652.
Lin T.Y. et Chien M.F.C., 2007. Exopolysaccharides production as affected by lactic acid bacteria and fermentation time. Food .Chem. 100 : 1419 1423.
Looijesteijin P.J., Trapel L., de Vries E., Abee T. et Hugenholtz J., 2001. Physiological function of exopolysaccharides produced by Lactococcus lactis. Int. J. Food Microbiol. 64 : 71-80.
Manca De Nadra MC , Strasser De Saad AM , Pesce de Ruiz Holgado AA and Olivier G (1985). Extracellular polysaccharide production by Lactobacillus bulgaricus CRL 420. Milchwissenschaft 40: 409-411.
Mantovani A, Maranghi F, Purificato I, Macrì A. 2006. Assessment of feed additives and contaminants: an essential component of food safety. Ann Ist Super Sanita. 42(4):427-32.
Marteau P, Seksik P, Lepage P, Dore J. 2004. Cellular and physiological effects of probiotics and prebiotics. Mini Rev Med Chem 4: 889-896
Marteau P. 2006. Living drugs for gastrointestinal diseases: the case for probiotics. Dig Dis.
24: 137-147.
Mathlouthi N., Mallet S., Saulnier L., Quemener B., Larbier M., 2002. Effects of xylanase and b-glucanase addition on performance, nutrient digestibility, and physico-chemical conditions in the small intestine contents and caecal microflora of broiler chickens fed a wheat and barley-based diet. Anim. Res., 51, 395-406.

Messaoudi S , , Kergourlay G, Rossero A, Ferchichi M, Prévost H, Drider D, Manai M, Dousset X L. 2011. Identification of lactobacilli residing in chicken ceca with antagonism against Campylobacter. Int. Microbiol. 14 (2):103-10.
Metchnikoff E., 1907. The prolongation of life, Heinemann, London.
Methner U., P. A. Barrow, et al. (1997). "Comparative study of the protective effect against Salmonella colonisation in newly hatched SPF chickens using live, attenuated Salmonella vaccine strains, wild-type Salmonella strains or a competitive exclusion product." International Journal of Food Microbiology 35(3): 223-230.
Metlef S. et Dilmi-Bouras A., 2009. Effet antagoniste de Lactococcus lactis, souches extrêmophiles locales, sur des espèces de la flore intestinale résidente. Rev. Nat. Tec. 1 : 33-44.
Moroni O, Kheadr E, Boutin Y, Lacroix C, Fliss I. 2006. Inactivation of adhesion and
invasion of food-borne Listeria monocytogenes by bacteriocin-producing Bifidobacterium
strains of human origin. Appl Environ Microbiol. 72: 6894-6901.
Mountzouris KC, Tsirtsikos P, Kalamara E, Nitsch S, Schatzmayr G, Fegeros K. 2007. Evaluation of the efficacy of a probiotic containing Lactobacillus, Bifidobacterium,
Enterococcus, and Pediococcus strains in promoting broiler performance and modulating cecal microflora composition and metabolic activities. Poult Sci. 86(2):309-17.
Murry A.C., A Hintonjr, J. R and Morrison, H., 2004. Inhibition of growth of escherichia coli, salmonella typhimurium and clostridia on chicken fee media by lactobacillus salivarius and lactobacillus plantarum perfringens. International journal of poultry science., 3 (9): 603-607.
Mutus R, Kocabagli N, Alp M, Acar N, Eren M, Gezen SS. 2006. The effect of dietary probiotic supplementation on tibial bone characteristics and strength in broilers. Poult Sci.
85(9):1621-5
Nakajima H, Toyoda S, Toba T, Itoh T, Mukai T, Kitazawa H, Adachi S (1990). A novel phosphopolysaccharide from slime-forming Lactococcus lactis ssp.cremoris SBT 0495. J. Dairy Sci. 73: 1472- 1477.
O’Mahony A., O’Sullivan, T., Walsh, Y., Vaughan, A., Maher, M., Fitzgerald, G.F., Van Sinderen, D. 2000.Characterisation of antimicrobial producing lactic acid bacteria from malted barley. J. Inst. Brew. 106, 403–410.

Ogier J.C., Casalta E., Farrokh C. et Saïhi A., 2008. Safety assessment of dairy microorganisms: The Leuconostoc genus. Int. J. Food Microbiol. 126 : 286-290.
Ouwehand AC, Salminen S, Tolkko S, Roberts P, Ovaska J, Salminen E. 2002b. Resected
human colonic tissue: new model for characterizing adhesion of lactic acid bacteria. Clin
Diagn Lab Immunol. 9: 184-186.
Ouwehand A. C, S. Salminen, et al. (2002). "Probiotics: an overview of beneficial effects." Antonie Van Leeuwenhoek International Journal of General and Molecular Microbiology 82(1-4): 279-289.
Oyarzabal O. A., Conner, D. E. and Blevins, W.T., 1995. Fructooligosaccharide utilization by salmonella and potential direct-fed-microbial bacteria for Poultry. J. FoodProt., 58(2): 92-96.
Pachaly P. (1999). Dünnschicht-Chromatographie in der Apotheke. 1. bis 4. Lieferung, Wissenschaftliche Verlagsgesellschaft mbH, Stuttgart
Pascual M., M. Hugas, et al. (1999). "Lactobacillus salivarius CTC2197 prevents Salmonella enteritidis colonization in chickens." Applied and Environmental Microbiology 65(11): 4981-4986.
Petit C, Grill JP, Maazouzi N, Marczak R (1991). Regulation of polysaccharide formation by Streptococcus thermophilus in batch and fedbatch cultures. Appl. Microbiol. Biotechnol. 36: 216-221.
Piard J.C and M. Desmazeaud ,1992. Inhibiting factors produced by lactic acid bacteria. 2. Bacteriocins and other antibacterial substances Lait 72 (1992) 113-142
Pilet M.F., Magras C., Federigh M., 2005. Bactéries lactiques. In : bactériologie alimentaire (Federighi M.). 2e Ed., Economica. Paris. 219-240.
Pilet M.F., Magras C., Federighi M., 1998. Bactéries lactiques. In : Manuel de bactériologie alimentaire (Sutra L., Feederighi M., Jouve J.L.). Polytechnica. Paris. 235-260.
Piva G., Rossi, F., 1999. Possible alternatives to the use of antibiotics as growth promoters .New additives. Ciheam., p. 83-106.
Ragione R. M. L., A. Narbad, et al. (2004). "<i>In vivo</i> characterization of <i>Lactobacillus johnsonii</i> FI9785 for use as a defined competitive exclusion agent against bacterial pathogens in poultry." Letters in Applied Microbiology 38(3): 197-205.

Revington B., 2002. Feeding poultry in the post-antibiotic era.Multi-State Poultry Meeting.http://ag.ansc.purdue.edu/poultry/multistate/multi-state.pdf.
Rolfe R. D., 2000. The Role of Probiotic Cultures in the Control of Gastrointestinal Health. J. Nutr., 130: 396–402.
Rotz , C. A., 2004. Management to reduce nitrogen losses in animal production. J. Anim. Sci., 82: 119–137.
Ruas-Madiedo P., Hugenholtz, J. and Zoon, P. (2002) An overview of the functionality of exopolysaccharides produced by lactic acid bacteria. Inter. Dairy J., 12, 163-171.
Saarela M., G. Mogensen, et al. (2000). "Probiotic bacteria: safety, functional and technological properties." Journal of Biotechnology 84(3): 197-215.
Salm-Surv G. (2005). un reseau de l'OMS pour la surveillance des maladies d'origine alimentaire, OMS.
Samli HE, Senkoylu N, Koc F, Kanter M, Agma A. 2007. Effects of Enterococcus faecium and dried whey on broiler performance, gut histomorphology and intestinal microbiota. Arch
Anim Nutr. 61(1):42-9.
Sanders ME. 2003. Probiotics: considerations for human health. Nutr Rev. 61: 91-99.
Sato T, J.P.Hu, K.Ohki, M.Yamaura, J.Washio, J.Matsuyama, N.Takahashi, Identification of mutans streptococci by restriction fragment length polymorphism analysis of polymerase chain reaction-amplified 16S ribosomal RNA genes. Oral Microbiology and Immunology, Volume 18, Issue 5, pages 323–326, October 2003
Schleifer K.H., 1987. Recent changes in the taxonomy of lactic acid bacteria. FEMS
Microbiol. Letters. 46 : 201-203.
Schrezenmeir J and De Vrese, M., 2001. Probiotics, prebiotics, and synbiotics approaching a definition. Am. J. Clin. Nutr., 73(2): 361-364.
Servin AL. 2004. Antagonistic activities of lactobacilli and bifidobacteria against microbial pathogens. FEMS Microbiol Rev. 28: 405-440.
Sheehan DC, Hrapchak BB. 1980. Theory and practice of histotechnology. St. Louis: C.V.mMosby. p 159– 179.
Simon O., A. Jadamus, et al. (2001). "Probiotic feed additives - effectiveness and expected modes of action." Journal of Animal and Feed Sciences 10: 51-67.

Simon O., 2005. Micro-Organisms as Feed Additives –Probiotics. Advances in Pork Production Volume 16, pg. 161.
Soczewi ński E., Wójciak M., Pachowicz K.: Rapid thin layer chromatographic systems for the analysis of sugars. Chem Anal., 43, 823, 1998.
Souly frag 1978 Separation and Analysis of Some Sugars by Using Thin Layer Chromatography,JOURNAL OF THE A.S.S.B.T.VOL. 20 No. 3 JULY 1978.
Spring P., Wenk C., Dawson K.A., Newman K.E., 2000. Poultry Sci. 79, 205-211.
Stiles M.E. et Holzapfel W.H., 1997. Lactic acid bacteria of foods and their current taxonomy. Int. J. Food Microbiol. 36 : 1-29.
Stingele F., J. R. Neeser, and B. Mollet. 1996. Identification and characterization of the eps (exopolysaccharide) gene cluster from Streptococcus thermophilus Sfi6. J. Bacteriol. 178:1680–1690.
Suskovic J., Kos, B., Goreta, J., and Mato, S., 2001. Role of Lactic Acid Bacteria and Bifidobacteria in Synbiotic Effect. Food. technol. biotechnol., 39 (3): 227-235.
Tadesse G., Ephraim E.et Ashenafi M., 2004. Assessment of the antimicrobial activity of lactic acid bacteria isolated from Borde and Shamita, traditional Ethiopian fermented beverages, on some foodborne pathogens and effect of growth medium on the inhibitory activity. Int. J. Food Safety. 5 : 13-20.
Tamime A.Y., 2002. Microbiology of starter cultures. In: Dairy microbiology handbook (Robinson R.K.). 3e Ed., John Wiley and Sons, Inc., New York. 261-366.
Tannock G. W. (1997). "Probiotic properties of lactic-acid bacteria: Plenty of scope for fundamental R&D." Trends in Biotechnology 15(7): 270-274.
Temim S. , Hammami N. et Bedran L. 2009. Evaluation De l'Efficacité Du Probiotique Pediococcus acidilactici sur Les performances de croissance, la morphométrie et la Flore lactobacillaire de l'intestin du poulet de chair. European Journal of Scientific Research, Vol.38 No.1 (2009), pp.119-128 .
Tierney J, Gowing H, Van Sinderen D, Flynn S, Stanley L, McHardy N, Hallahan S & Mulcahy G .2004. In vitro inhibition of Eimeria tenella invasion by indigenous chicken Lactobacillus species. Vet Parasitol 122:171-182.

Titiek F.D., Endang S.R., Djoko W. et Slamet S., 1996. Antimicrobial substance produced by lactobacillus sp. TGR-2 isoleted from Growol. Indonesian. Food Nutr. Prog. 3(2) : 29-34.
Toma M. M., Raipulis, J., Kalnina, I. and Rutkis, R., 2005. Effect of Probiotic Yeast on Genotoxicity. Food Technol. Biotechnol., 43 (3): 301-305.
Torres-Rodriguez A, Donoghue AM, Donoghue DJ, Barton JT, Tellez G, Hargis BM. 2007. Performance and condemnation rate analysis of commercial turkey flocks treated with a Lactobacillus spp.-based probiotic. Poult Sci. 86(3):444-6.
Tosukhowong A., Nakayama J., Mizunoe Y., Sugimoto S., Fukuda D. et Sonomoto K., 2005. Reconstitution and function of Tetragenococcus halophila chaperonin 60 tetradecamer. J.
Biosci. Bioengin. 99 : 30-37.
Trias R., Badosa E., Montesinosand E. et Bañeras L., 2008. Bioprotective Leuconostoc
strains against Listeria monocytogenes in fresh fruits and vegetables. Int. Dairy J. 18: 1-5.
Tuomola E, Crittenden R, Playne M, Isolauri E, Salminen S. 2001. Quality assurance criteria for probiotic bacteria. Am J Clin Nutr. 73: 393S-398S.
Van Coillie E., Goris J., Cleenwerck I., Grijspeerdt K., Botteldoorn N., Van Immerseel F., De Buck J., Vananneyt M., Swings J., Herman L. et Heyndrickx M. (2007) Identification of Lactobacilli isolated from the cloaca of laying hens and characterization for potential use as probiotics to control Salmonella Enteritidis. Journal of Applied Microbiology 102, 1095- 1106..
Van den Berg DJC, Robijn GW, Janssen AC, Giuseppin MLF, Vreeker R, Kamerling JP, Vliegenthart JFG, Ledeboer AM, Verrips CT (1995). Production of a novel extracellular polysaccharide by Lactobacillus sake 0-1 and characterization of the polysaccharide. Appl. Environ. Microbiol. 61: 2840-2844.
Van Immerseel F., De Buck j., Boyen F., Pasmans F., Bertrand S., Collard J.M., Saegerman C., Hooyberghs J., Haesebrouck F., Ducatelle R., 2005. Salmonella dans la viande de volaille et dans les oeufs : un danger pour le consommateur qui demande la mise en place d’un
programme de lutte efficace. Ann. Méd. Vét., 149: 34-48.
Van Immerseel F., De Buck, J., Pasmans, F., Haesebrouk, F., Ducatelle, R., 2003. Stratégies nutritionnelles pour réduire les agents pathogènes chez la volaille. Cinquièmes journées de la recherche avicole. Tours.
Van Immerseel , F., Cauwerts, K., Devriese, L.A., Haesebrouck, F., and Ducatelle, R., 2002. Feed additives to control Salmonella in poultry. World’s Poultry Science Journal., 58: 501-51.

Weurding R., H. Enting, et al. (2003). "The relation between starch digestion rate and amino acid level for broiler chickens." Poultry Science 82(2): 279-284.
Willard M.D., Simpson R.B., Delles E.K., Cohen N.D., Fossum T.W., Kolp D.L., Reinhard G., (1994a) Effects of dietary supplementation of fructo-oligosaccharides on small intestinal bacterial overgrowth in dogs, Am. J. Vet. Res, 55, 654 - 9
Wood , M. T., 1998. The use of EM in the poultry industry. Suitainable community development, L. L. C
Yildirim Z, and Johnson MG. 1998. Characterization and antimicrobial spectrum of bifidocin B, a bacteriocin produced by Bifidobacterium bifidum NCFB 1454. J Food Prot. 61: 47- 51.
Yoruk MA, Gul M, Hayirli A & Macit M (2004) The effects of supplementation of humate and probiotic on egg production and quality parameters during the late laying period in hens. Poult Sci 83: 84-88.
Zacconi C., Svolari, Fraioli, G.D., Sarra, P.G., 1999. Colonisation of chicken intestinal tract by lactobacillus salivarius A23 strain. Annali di Microbiologia ed Enzimologia., 49:103-115.

Annexe 1 : Composition des milieux de culture et tampon
Milieu MRS (pH 6.5) Peptone ......................................................................................10g
Extrait de viande ....................................................................... 10g
Extrait de levure ..........................................................................5g
Glucose ......................................................................................20g
Tween 80....................................................................................1ml
Phosphate bipotassique .............................................................. 2g
Acétate de sodium .......................................................................5g
Citrate d’ammonium ...................................................................2g
Sulfate de magnesium, 7 H2O ................................................ 0.2g
Sulfate de manganèse, 4 H2O .................................................0.5g
Agar .........................................................................................15g
Eau distillée qsp ................................................................ 1000ml
Gélose hypersaccharosée (pH 6.8)
Extrait de viande ....................................................................10g
Extrait de levure ..................................................................... 3g
Peptone................................................................................ 2.5 g
Saccharose.......................................................................... 150 g
K2HPO4 .................................................................................2 g
NaCl .......................................................................................1 g
MgSO4, 7H2O .....................................................................0.2g
Agar ......................................................................................15g
Eau distillée qsp ............................................................. 1000 ml
Milieu HJL (pH6.5)
Tryptone ………………………………………………….3g
Extrait de bœuf …………………………………………0.2g
Extrait de levure …………………………………………..1g
Lactose ……………………………………………….......1g
KH2PO4 …………………………………………...…...0.5g
Eau distillée qsp……………………………………….100 ml
· Stérilisation des milieux de culture par autoclavage à 120°C pendant 15 min.

Tampon TBE (pH8)
Tris-Hcl 30mM
Acide Borique 89mM
EDTA 2mM
Annexe II : Coloration de Gram
La coloration de Gram a été réalisée selon la technique suivante :
· Sur une lame, fixer à la chaleur une culture bactérienne ;
· Recouvrir la lame avec la solution de violet de gentiane pendant une minute ;
· Ajouter du lugol pendant 30 secondes ;
· Décolorer avec de l’alcool 95°, puis rincer à l’eau ;
· Faire une contre coloration en utilisant la fuschine et laisser agir 20 à 30 secondes ;
· Laver à l’eau ;
· Après séchage, soumettre la lame à une observation microscopique à immersion
(x100).
Les bactéries à Gram positif apparaissent en violet et les bactéries à Gram négatif en rose

RésuméLe recours aux antibiotiques a connu ses limites en raison de l’émergence de nouvelles
souches multi résistantes causées par l’utilisation abusive de ces composés dans le secteur
avicole, malgré que l’antibioprévention reste actuellement le seul moyen utilisé pour contrôler
les problèmes sanitaires et économiques liés aux pathogènes aviaires, l’utilisation des
probiotiques s’avère nécessaire pour favoriser une bonne microflore antagoniste vis-à-vis des
pathogènes et peut s’inscrire comme stratégie alternative envisagée pour protéger les volailles
des agents pathogènes et pour remplacer les antibiotiques comme facteur de croissance.
Ce travail s’intéresse à l’identification par les méthodes ADN des souches de volaille
productrices de substances inhibitrices vis-à-vis de certains pathogènes Staphylococcus
aureus, Escherichia coli, Salmonella typhi. 11 de ces souches ont été retenues suite à leur
forte action inhibitrice. Au total 48 souches ont été isolées à partir de différents
compartiments du tractus digestif de deux coqs de race locale, dont 6 ont été pré-identifiés à
des leuconostocs, 2 à des pédiocoques, 8 à des entérocoques, 22 à des lactobacilles, 5
apparentés au genre Bacillus et enfin 4 autres appartenant à des levures.
Par ailleurs, l’utilisation des amorces d’une région interne de 480 pb du gène l’ADNr 16S
spécifiques au genre Lactobacillus sp. et le séquençage du fragment d’ADN amplifié ont
permis d’identifier les isolats appartenant à la flore lactobacillaire au nombre de 22.
Lactobacillus acidophilus, Lactobacillus gallinarum, Lactobacillus crispatus et Lactobacillus
johonossi ont été les souches les plus diagnostiquées par la méthode PCR colonie. Une autre
approche moléculaire a été entamée afin d’amplifier le gène ADNr 16S dans sa totalité 1500
pb chez certaines souches, celui-ci reste un taxon de choix en matière de systématique en
bactériologie. D’autres travaux menés sur la détection et l’extraction des prébiotiques tels que
les polysaccharides exogènes (EPS) ainsi que leur caractérisation par chromatographie sur
couche mince qui a montré une similitude des profils avec le dextrane commercial et leur
dosage colorimétrique qui a été déterminé à 1254 mg/l pour la production d’EPS la plus
élevée chez la souche LGM-IL2.
Mot clés :Lactobacillus Spp; ADN; PCR; Probiotiques; Aviculture; Pathogènes; Activité Inhibitrice;
Prébiotiques; CCM; Souche LGM-IL2.


















