
ÉCOLOGIE DE LA CÉCIDOMYIE DU SAPIN (PARADIPLOSIS …€¦ · Résumé Les infestations de la...
83
PAYSE MAILHOT ÉCOLOGIE DE LA CÉCIDOMYIE DU SAPIN (PARADIPLOSIS TUMIFEX) : RELATIONS AVEC LA CÉCIDOMYIE INQUILINE DES GALLES (DASINEURA BALSAMICOLA) ET SES PARASITOÏDES Mémoire présenté à la Faculté des études supérieures de l’Université Laval dans le cadre du programme de maîtrise en biologie végétale pour l’obtention du grade de maître ès sciences (M.Sc.) DÉPARTEMENT DE PHYTOLOGIE FACULTÉ DES SCIENCES DE L’AGRICULTURE ET DE L’ALIMENTATION UNIVERSITÉ LAVAL QUÉBEC 2006 © Payse Mailhot, 2006
Transcript of ÉCOLOGIE DE LA CÉCIDOMYIE DU SAPIN (PARADIPLOSIS …€¦ · Résumé Les infestations de la...

PAYSE MAILHOT
ÉCOLOGIE DE LA CÉCIDOMYIE DU SAPIN (PARADIPLOSIS TUMIFEX) : RELATIONS AVEC LA
CÉCIDOMYIE INQUILINE DES GALLES (DASINEURA BALSAMICOLA) ET SES
PARASITOÏDES
Mémoire présenté à la Faculté des études supérieures de l’Université Laval
dans le cadre du programme de maîtrise en biologie végétale pour l’obtention du grade de maître ès sciences (M.Sc.)
DÉPARTEMENT DE PHYTOLOGIE FACULTÉ DES SCIENCES DE L’AGRICULTURE ET DE L’ALIMENTATION
UNIVERSITÉ LAVAL QUÉBEC
2006 © Payse Mailhot, 2006

Résumé Les infestations de la cécidomyie du sapin (Paradiplosis tumifex), un ravageur important du
sapin baumier (Abies balsamea), surviennent périodiquement et ont été observées dans les
plantations du sud du Québec de 2000 à 2002. Divers ennemis naturels causent
probablement les fluctuations cycliques d’abondance, tels que la cécidomyie inquiline des
galles (Dasineura balsamicola) et diverses espèces de parasitoïdes. Cette étude réalisée en
Estrie, visait à documenter leurs interactions, afin d’évaluer leur rôle dans l’effondrement
des populations de P. tumifex survenu en 2003. Cinq espèces de parasitoïdes associés à
P. tumifex ont été identifiées. Des données relatives à la présence de plusieurs parasitoïdes
dans une même galle (multiparasitisme) suggèrent des interactions négatives (interférence)
fréquentes entre les ennemis naturels de P. tumifex. De plus, les forts taux d’inquilinisme et
de parasitisme indiquent que ces ennemis naturels sont probablement responsables de
l’effondrement des populations de ce ravageur, en 2003 dans les plantations du sud-ouest
du Québec.

iii
Remerciements Durant la période où j’étais étudiante à la maîtrise, j’ai accouché trois fois. La première
d’une fille, la seconde d’un garçon et la troisième de ce mémoire. Ces trois «délivrances»
m’ont apporté une grande fierté mais n’auraient pas été possibles sans l’appui de
nombreuses personnes.
Tout d’abord, je voudrais remercier mes directeurs de recherche, messieurs Jacques
Brodeur et Conrad Cloutier pour leur soutien, leurs conseils judicieux, leur compréhension
et surtout pour m’avoir transmis leur passion de l’entomologie et de la recherche du savoir.
Ensuite, j’aimerais remercier le Réseau Biocontrôle du Canada, l’Association des
Producteurs d’Arbres de Noël du Québec et le CDAQ pour leur soutien financier. Merci
aussi aux producteurs de sapins qui ont donné accès à leurs plantations soit messieurs
Francis Côté et Benoît Bédard, ainsi qu’à monsieur André Pettigrew, agronome du Réseau
d’Avertissement Phytosanitaire du MAPAQ, direction générale de l’Estrie et expert en
protection des arbres de Noël, pour son soutien au projet et la collecte de données à la
plantation de Racine.
Merci à monsieur Georges Pelletier du Centre de foresterie des Laurentides, qui m’a
soutenue dans l’identification des parasitoïdes ainsi qu’à messieurs Gary Gibson, John
Huber et Lubomir Masner, taxonomistes au Ministère de Agriculture et de Agroalimentaire
Canada, à Ottawa, pour l’identification des spécimens.
Je voudrais également remercier Christine Jean, Simon Boudreault et Sébastien Jacob,
professionnels de recherche à l’Université Laval, qui ont grandement contribué à la prise de
données et qui m’ont fourni des photos. Merci, à Mylène Blais, Marie-Pierre Migneault et à
tous les étudiants qui ont investi de nombreuses heures de travail à ce projet de recherche.

À Steve, Méliane, Mael et à mes parents qui m’ont fait découvrir la beauté de la nature.

Table des matières
Résumé............................................................................................................................... ii Remerciements.................................................................................................................. iii Table des matières ..............................................................................................................v Liste des tableaux............................................................................................................. vii Liste des figures .............................................................................................................. viii
Chapitre 1. Introduction générale ...........................................................................................1 1.1. Introduction..................................................................................................................2 1.2. Les galles .....................................................................................................................3 1.3. Les cécidomyies...........................................................................................................6
1.3.1. Biologie de la cécidomyie du sapin Paradiplosis tumifex ....................................7 1.3.2. Biologie de la cécidomyie inquiline des galles Dasineura balsamicola ............12 1.3.3. Dynamique des populations................................................................................17
1.4. Hypothèses générales.................................................................................................21 1.5. Objectifs.....................................................................................................................21 1.6. Bibliographie .............................................................................................................22
Chapitre 2. Le parasitisme et l’inquilinisme comme facteurs de régulation des populations de la cécidomyie du sapin Paradiplosis tumifex Gagné. ......................................................26
2.1. Résumé.......................................................................................................................27 2.2. Introduction................................................................................................................28 2.3. Matériels et méthodes ................................................................................................29
2.3.1. Caractéristiques des plantations et parcelles d’échantillonnage .........................30 2.3.2. Mesure de l’activité des insectes adultes ............................................................31 2.3.3. Échantillonnage des pousses...............................................................................32 2.3.4. Élevage des parasitoïdes à partir de la récolte d’aiguilles porteuses de galles ...33 2.3.5. Identification des spécimens...............................................................................35 2.3.6. Analyses des données .........................................................................................35
2.3.6.1. Taux d’infestation ........................................................................................35 2.3.6.2. Inquilinisme .................................................................................................36 2.3.6.3. Effet de la densité de P. tumifex sur D. balsamicola ...................................36 2.3.6.4. Ectoparasitisme............................................................................................37 2.3.6.5. Endoparasitisme...........................................................................................37 2.3.6.6. Relations de densité entre la cécidomyie galligène et ses parasitoïdes .......38
2.4. Résultats.....................................................................................................................39 2.4.1. Taux d’infestation de P. tumifex .........................................................................39 2.4.2. Inquilinisme ........................................................................................................39 2.4.3. Identification des parasitoïdes ............................................................................42 2.4.4. Activité des parasitoïdes adultes.........................................................................43 2.4.5. Ectoparasitisme et endoparasitisme ....................................................................44
2.5. Discussion......................................................................................................................48 2.5.1. Taux d’infestation de P. tumifex .........................................................................49 2.5.2. Inquilinisme ........................................................................................................50

vi
2.5.2.1. Taux d’inquilinisme.....................................................................................50 2.5.2.2. Relation entre D. balsamicola et P. tumifex ................................................50
2.5.3. Parasitisme ..........................................................................................................52 2.5.3.1. Identification des parasitoïdes .....................................................................52 2.5.3.2. Distribution saisonnière des parasitoïdes.....................................................55 2.5.3.3. Ectoparasitisme............................................................................................58 2.5.3.4. Endoparasitisme...........................................................................................60 2.5.3.5. Multiparasitisme ..........................................................................................61
2.5.4. Impacts des ennemis naturels sur P. tumifex ......................................................64 2.6. Bibliographie .............................................................................................................66
Chapitre 3. Conclusions générales........................................................................................70 3.1. Conclusion .................................................................................................................71 3.2. Bibliographie .............................................................................................................74

Liste des tableaux Tableau 1-1. Taxonomie des parasitoïdes hyménoptères associés à la cécidomyie du sapin
Paradiplosis tumifex décrits par MacGown et Osgood (1972).....................................19 Tableau 2-1. Taux saisonnier moyen d’infestation des pousses par Paradiplosis tumifex et
nombre moyen de galles par pousse sur sapin baumier (Abies balsamea) dans des plantations de trois localités du sud du Québec, en 2002. ...........................................39
Tableau 2-2. Identification des parasitoïdes hyménoptères associés à la cécidomyie du sapin Paradiplosis tumifex et obtenus à partir de l’élevage post-dormance en laboratoire. ..42
Tableau 2-3. Régressions linéaires des effets de la densité (n/pousse) des galles de Paradiplosis tumifex sur les pousses du sapin baumier, de leur longueur et de la densité de l’inquiline, Dasineura balsamicola, sur la densité et le pourcentage d’ectoparasitoïdes, à Saint-Fortunat, Québec en 2002. ...............................................47
Tableau 2-4. Régressions linéaires des effets de la densité (n/pousse) des galles de Paradiplosis tumifex sur le sapin baumier et de la longueur des pousses sur la densité des endoparasitoïdes et le taux d’endoparasitisme, dans une plantation à Saint-Fortunat, Québec, en 2002. ..........................................................................................47

Liste des figures Figure 1-1. Femelle de Paradiplosis tumifex pondant ses oeufs sur les aiguilles du sapin
baumier à l’ouverture des bourgeons .............................................................................8 Figure 1-2. Paradiplosis tumifex, adulte mâle (corps 1,8 mm; antennes 1,7 mm). ................9 Figure 1-3. Oeufs de Paradiplosis tumifex ou de Dasineura balsamicola
(longueur = 0,35 mm) sur une jeune aiguille de sapin..................................................10 Figure 1-4. Larve de Paradiplosis tumifex de 3e stade (3,0 mm), à l’intérieur d’une galle
disséquée.......................................................................................................................11 Figure 1-5. Pupe de Paradiplosis tumifex (2,5 mm) dont les extrémités (tête, ailes et pattes)
noircissent après environ cinq jours de pupaison, selon la température. ......................12 Figure 1-6. Larves de deuxième stade de Dasineura balsamicola (gauche) et de
Paradiplosis tumifex (droite) dans la même galle, disséquée en laboratoire. Photo : S. Boudreault. ...............................................................................................................15
Figure 1-7. Dasineura balsamicola adulte mâle (corps 1,8 mm; antennes 1,7 mm). Photo : P. Mailhot. ........................................................................................................16
Figure 2-1. Quatre larves cyclopiformes de premier stade de Platygaster sp., deux à l’intérieur et deux à l’extérieur d’une larve de cécidomyie du sapin, Paradiplosis tumifex brisée et disposée entre lame et lamelle, observée au microscope...................38
Figure 2-2. Taux d’inquilinisme des galles de Paradiplosis tumifex par Dasineura balsamicola à Saint-Fortunat, Québec, en 2002 (moyenne annuelle 16,43 % ; écart-type 5,15 %)..................................................................................................................40
Figure 2-3. Relation entre la densité de l’inquiline Dasineura balsamicola (n/pousse), et celle des galles de son hôte la cécidomyie du sapin, Paradiplosis tumifex (n = 565); plusieurs points sont superposés sur la figure, plus particulièrement dans les régions inférieures (R2 = 0,30381; P = 0,0001; Y = 0,04080+ 0,16700x– 0,00178x2). ............41
Figure 2-4. Distribution et abondance saisonnière, en 2002, des parasitoïdes adultes capturés sur pièges collants jaunes placés au sommet de sapins baumiers à Saint-Fortunat, Québec, dans une plantation infestée de cécidomyie du sapin, Paradiplosis tumifex. .........................................................................................................................43
Figure 2-5. Taux d’ectoparasitisme des larves de Paradiplosis tumifex à l’intérieur des galles sur sapin baumier, Saint-Fortunat, Québec, en 2002. ........................................44
Figure 2-6. Taux d’endoparasitisme des larves de Paradiplosis tumifex évalué à partir de sous-échantillons de larves à l’intérieur des galles sur le sapin baumier, à Saint-Fortunat, Québec, en 2002. ..........................................................................................45
Figure 2-7. Relation entre la densité des ectoparasitoïdes (nombre par pousse) et celle des galles de Paradiplosis tumifex sur le sapin baumier à Saint-Fortunat, Québec, en 2002. . .....................................................................................................................................46
Figure 2-8. Première mention d’un ectoparasitoïde (stade larvaire) sur une larve de Dasineura balsamicola, la cécidomyie inquiline des galles. La photo montre une galle disséquée de P. tumifex, collectée à Saint-Fortunat, le 28 octobre 2002. .....................60

Chapitre 1. Introduction générale

2
1.1. Introduction Les plantations de sapins baumiers (Abies balsamea Mill.) sont destinées à la vente sur le
marché des sapins de Noël. Ces ventes atteignaient 1,4 millions d’arbres sur les marchés
locaux et d’exportation, aux États-Unis et au Mexique en 2002 (APANQ, 2005). Quatre-
vingt pour-cent des arbres de Noël cultivés au Québec sont exportés, ce qui entraîne des
retombées économiques importantes (près de 50 millions de dollars en 2002). Près de
11 000 hectares de terres québécoises sont consacrés à cette culture. Les membres de
l'Association des producteurs d’arbres de Noël du Québec (APANQ) vendent plus de
500 000 arbres aux Québécois, chaque année. Les principales régions productrices et
exportatrices de sapins de Noël sont l’Estrie et la Beauce, où les plantations atteignent une
superficie de 7 000 hectares. D’autres régions telles la Montérégie et les Laurentides sont
également productrices de sapins baumiers mais à une moins grande échelle (APANQ,
2005).
C’est une culture pérenne dont la durée de croissance des arbres avant la coupe, au Québec,
est de 5 à 12 ans (APANQ, 2005). Comme la production de sapins s’échelonne sur une
longue période, de nombreuses étapes de production se suivent et beaucoup d’énergie est
investie pour amener chaque arbre à maturité. Les semences sont d’abord choisies, puis
semées dans une pouponnière, un endroit désigné pour la reproduction des arbres en
pépinières (Bédard, comm. pers., 2002). Les jeunes arbres sont transplantés aux champs au
début de leur deuxième ou troisième année de croissance (Bédard, comm. pers., 2002). Au
cours des années suivantes, certaines activités sylvicoles devront être exécutées telles que la
fertilisation, le désherbage, la taille des arbres, la répression de maladies et d’insectes, la
coupe des arbres et le dessouchage (APANQ, 2005).
Pour produire de beaux arbres, l’entretien général ne suffit pas lorsque des maladies ou des
insectes ravageurs s’attaquent à la plantation. Des connaissances et des outils pour lutter
contre ces problèmes sont essentiels.

3 Les principales maladies associées au sapin baumier sont le balai de sorcière,
(Melampsorella caryophyllacearum), le pourridié-agaric (Armillaria ostoyae), le rouge des
aiguilles (Rhizosphaera spp. et Lirula spp.) et le chancre derméen (Dermea sp.) (APANQ,
2005). Le climat peut également être défavorable pour les arbres en plantation lorsqu’il y a
une dessiccation hivernale soit une déshydratation des aiguilles due aux vents chauds alors
que le sol est gelé (APANQ, 2005).
Les principaux ravageurs du sapin baumier sont le puceron des pousses du sapin (Mindarus
abietinus Koch), la tordeuse des bourgeons de l’épinette (Choristoneura fumiferana
(Clem.)), le diprion du sapin (Neodiprion abietis (Harr.)), le tétranyque de l’épinette
(Oligonychus ununguis (Jac.)), le longicorne noir (Monochamus scutellatus (Say)), le
perce-pousse du sapin (Pleroneura brunneicornis Roh.) et la cécidomyie du sapin
(Paradiplosis tumifex Gagn.) (APANQ, 2005). Cette étude se concentre sur la cécidomyie
du sapin, P. tumifex (Diptera : Cecidomyiidae) un ravageur épisodique à l’origine de la
formation de petites galles sur les aiguilles de sapin.
1.2. Les galles Une galle est une formation végétative résultant d’un développement atypique des cellules,
des tissus ou des organes de la plante (Mani, 1964; Shorthouse et Rohfritsch, 1992).
L’appellation «galle», réfère aux formations produites par l’entremise d’un organisme
parasite, tel que des virus, bactéries, champignons, nématodes, acariens ou insectes (Mani,
1964; Meyer, 1987). De plus, c’est la plante qui doit initier la croissance végétative en
réaction à l’attaque du parasite et cette croissance varie selon la physiologie nutritionnelle
ou le mode d’action utilisé par le parasite pour exploiter son hôte (Mani, 1964; Meyer,
1987). Toutefois, c’est l’organisme parasite qui contrôle le développement de la galle et s’il
meurt avant la maturation de celle-ci, sa croissance sera arrêtée (Stone et Schönrogge,
2003).

4 En général, les galles sont observées sur les plantes (règne végétal) et rarement, sur des
champignons (règne mycètes) (Shorthouse et Rohfritsch, 1992). Dans le règne végétal,
plusieurs types de galles sont rencontrés et, conséquemment, il existe divers types de
classification des galles. Mani (1964) les classe en fonction de la partie de la plante qui est
touchée. Les galles peuvent se retrouver sur tous les organes des plantes-hôtes; en majorité
sur les feuilles (80%) mais également sur les racines, tiges, fleurs, fruits et graines
(Shorthouse et Rohfritsch, 1992). Certains spécialistes les classent plutôt selon qu’il s’agit
d’un organe (galle organoïde) ou d’un tissu (galle histidoïde) qui est affecté (Meyer, 1987;
Shorthouse et Rohfritsch, 1992). Les galles histidoïdes sont divisées en deux catégories :
les galles cataplasmiques dont la différenciation tissulaire est peu élaborée, ce qui donne
des galles à croissance irrégulière de taille et de volume variables; et les galles
prosoplasmiques dont la taille et la forme sont spécifiques grâce à une différenciation
tissulaire bien définie (Meyer, 1987; Shorthouse et Rohfritsch, 1992). La majorité des
zoocecidia, i.e. galles induites par des invertébrés cecidozoaires (nématodes et arthropodes
causant la formation de galles) sont des galles histidoïdes prosoplasmiques (Meyer, 1987).
Les galles sont également classées selon leur forme : étui, rouleau de feuille, déformation
des bourgeons apicaux, rosette, etc. (Felt, 1940; Mani, 1964; Meyer, 1987; Shorthouse et
Rohfritsch, 1992).
Les insectes responsables de la formation de galles sont nommés cécidogènes ou
galligènes. Ils représentent plus de 13 000 espèces et sont largement distribués à travers
divers ordres : diptères (Cecidomyiidae), hémiptères (Tingidae), homoptères (un grand
nombre de familles incluant les pucerons galligènes, super-famille Aphidoidea),
hyménoptères (Eulophidae et Cynipidae), lépidoptères, thysanoptères et coléoptères
(certains charançons) (Meyer, 1987; Shorthouse et Rohfritsch, 1992; Stone et Schönrogge,
2003). La plupart des insectes galligènes sont très spécifiques non seulement à l’espèce de
la plante-hôte, mais également à l’organe de celle-ci (Shorthouse et Rohfritsch, 1992).
Les insectes galligènes sont des herbivores spécialistes qui ont des ancêtres communs avec
d’autres insectes qui n’induisent pas la formation de galles. Ceux-ci ayant évolués
parallèlement, ils demeurent néanmoins très près les uns des autres, d’un point de vue

5 taxonomique (Shorthouse et Rohfritsch, 1992). Selon Stone et Schönrogge (2003), trois
hypothèses sont invoquées pour expliquer l’évolution de la capacité à induire une galle
chez les insectes. La première est que la nourriture dans une galle est abondante, à
proximité, riche en éléments nutritifs et les tissus végétaux y sont moins bien protégés
qu’ailleurs (absence de cire protectrice, diminution ou détournement des réactions de
défense de la plante). La seconde hypothèse présume que l’insecte galligène, vivant à
l’intérieur de la galle, est protégé des conditions environnementales défavorables, par
exemple, de dessiccation ou de fortes précipitations. Troisièmement, la galle permettrait de
protéger l’espèce galligène contre ses ennemis naturels. En principe, la galle protège
physiquement son occupant des prédateurs, parasitoïdes et pathogènes généralistes. Par
contre, les galles confèrent également des inconvénients majeurs car elles sont fixes et donc
plus facilement repérables par les parasitoïdes et prédateurs spécifiques (Murakami, 1999).
L’impact des dommages occasionnés à une plante par un insecte galligène est variable et
difficile à quantifier. On estime cependant que dans la majorité des cas, les dommages ne
sont pas d’une grande importance (Mani, 1964). En effet, les plantes portant un nombre
limité de galles augmenteraient facilement leur photosynthèse, pour compenser les pertes
occasionnées (Bagatto et al., 1996).
L’inconvénient principal pour la plante qui abrite un insecte galligène, est qu’elle doit
investir de l’énergie dans la formation de la galle plutôt que dans sa propre croissance. Le
galligène est en compétition directe avec les organes ou tissus de la plante-hôte. Celui-ci
détourne et consomme les nutriments et produits photosynthétiques des cellules ou du
phloème de sa plante-hôte (Bagatto et al., 1996; Stone et Schönrogge, 2003). Les galles
peuvent également entraîner un effet de dépérissement précoce des organes de la plante-
hôte, par exemple des feuilles ou aiguilles infectées tombent prématurément (West et
Shorthouse, 1982; Williams et Whitham, 1986; Bagatto et al., 1996), des fruits avortent
(Stone et Schönrogge, 2003) et des semences seront infertiles (West et al., 1996; AAC,
2004). Lorsque les fleurs sont attaquées, la production de graines est réduite ce qui entraîne
une baisse de fitness pour la plante ainsi que des pertes de rendement dans le cas de plantes
cultivées (West et al., 1996; Stone et Schönrogge, 2003; AAC, 2004). Par exemple, la

6 cécidomyie orangée du blé Sitodiplosis mosellana (Gehin) (Diptera : Cecidomyiidae) peut
se développer sur le blé, le seigle et l’orge (INRA, 1998). Son attaque précoce sur le blé
réduit la formation des grains et une attaque plus tardive rend les grains fragiles, déformés
ou mous au toucher (INRA, 1998). Une infestation de S. mosellana peut entraîner une
diminution de la qualité des grains, du rendement ainsi que des pertes économiques
importantes (AAC, 2004).
D’autres insectes galligènes peuvent causer des dommages mineurs aux plantes mais
occasionnent des pertes économiques lorsque produites dans un but commercial, surtout
pour des raisons esthétiques comme c’est le cas pour les plantes ornementales. Les galles
de la cécidomyie du rhododendron Clinodiplosis rhododendri (Felt) (Diptera :
Cecidomyiidae) sur Rhododendron catawbiensis Michaux, causent une déformation et une
décoloration des feuilles. La galle a peu d’impact sur la croissance et la fécondité de la
plante-hôte, par contre, elle cause un réel problème dans les pépinières où la production
commerciale de C. rhododendri nécessite des pulvérisations d’insecticides dans la saison
de fort achalandage des pépinières, et augmente l’exposition des clients à des produits
nocifs tout en diminuant les ventes de rhododendrons (Speker et Johnson, 1988; Hanula,
1991).
Toutefois, certaines galles d’insectes ont déjà été utilisées au service de l’homme. Par
exemple, en Europe et en Asie des galles contenant de fortes doses de tanin étaient utilisées
pour la fabrication d’encre et comme combustible dans les lampes. Aux États-Unis, des
galles de feuilles de chêne étaient utilisées comme supplément à l’alimentation du bétail, à
cause de leur riche teneur en protéines (Felt, 1940).
1.3. Les cécidomyies Les cécidomyies sont des diptères de la famille des Cecidomyiidae. La morphologie de
l’adulte s’apparente à celle d’un petit moustique. Certaines espèces sont mycophages
(Gagné, 1981), d’autres sont prédatrices (van Schelt et Mulder, 2000) ou même parasitoïdes
(Poinar, 2003). Certaines larves prédatrices sont utilisées comme agents de lutte biologique

7 contre les pucerons dans les serres, telle l’espèce Aphidoletes aphidimyza (Rond.) (van
Schelt et Mulder, 2000). Toutefois, la majorité des cécidomyies sont galligènes (Gagné,
1981; Shorthouse et Rohfritsch, 1992) et passent un ou plusieurs stades de leur vie à
l’intérieur des tissus modifiés (galles) de la plante-hôte. Les cécidomyies galligènes
produisent, selon l’espèce, différents types et formes de galles, tels que des étuis, des lèvres
recouvrant l’insecte, un enroulement de jeunes feuilles et l’inhibition de croissance du
bourgeon apical qui résulte en une galle en forme d’artichaut (Felt, 1940; Shorthouse et
Rohfritsch, 1992). Les cécidomyies galligènes, habituellement très spécifiques à leur
plante-hôte, sont associées à une grande diversité de plantes. On les retrouve dans les petits
fruits (bleuet, framboise, cassis), les arbres fruitiers (pomme, poire, prune, mangue, figue),
les céréales (blé, riz, orge seigle, avoine), les légumineuses (trèfle, luzerne, lentille, pois,
févier), les conifères (pin, sapin) et un grand nombre de plantes ornementales
(rhododendron, lavande).
Les cécidomyies galligènes sécrètent des substances stimulant la croissance des plantes,
qui, une fois injectées dans les tissus de la plante, provoquent la formation de la galle
(Shorthouse et Rohfritsch, 1992). Par exemple, des gibbérellines ont été trouvées à faibles
taux dans les larves de Janetiella sp. (Diptera : Cecidomyiidae). Ces substances produites
par les glandes salivaires, ou parfois d’autres organes, pourraient augmenter les niveaux
d’auxine dans les portions stimulées de la plante et causer une croissance anormale, en
formant une galle (Shorthouse et Rohfritsch, 1992).
1.3.1. Biologie de la cécidomyie du sapin Paradiplosis tumifex La cécidomyie du sapin (Paradiplosis tumifex Gagné) est un diptère galligène spécifique
aux sapins baumiers (A. balsamea) et frasers (A. fraseri). Cette espèce est univoltine,
indigène de l’Amérique du Nord et son aire de distribution s’étend de la Caroline du Nord,
aux monts Otish dans le Nord québécois près du 53e parallèle, et de Terre-Neuve au
Manitoba (Giese et Benjamin, 1959; Martineau, 1985).

8 Les cécidomyies du sapin adultes émergent du sol au début mai, au Maine (Osgood et al.,
1992) et à la mi-mai au Québec (Jean et al., 2003). Paradiplosis tumifex ressemble à un
petit moustique orangé ou rougeâtre et mesure entre 3,5 et 3,7 mm, en incluant les antennes
(Giese et Benjamin, 1959; Osgood et al., 1992). L’abdomen de la femelle se prolonge en un
ovipositeur lui permettant de pondre ses œufs entre les aiguilles serrées des bourgeons de
sapin (Figure 1-1; Osgood et al., 1992). Lors de l’émergence de la cécidomyie, l’abdomen
de la femelle est rempli d’œufs (moyenne de 120) uniformes et bien développés (Akar,
1987; Osgood et al., 1992). Les appendices du mâle (pattes et antennes) sont plus longs que
ceux de la femelle et une paire de crochets génitaux termine le bout de l’abdomen
(Figure 1-2). Les antennes des mâles et des femelles diffèrent puisqu’il y a alternance de
petits et de gros articles chez les mâles alors qu’ils sont semblables chez les femelles et
moins pubescents (Osgood et Gagné, 1978).
Figure 1-1. Femelle de Paradiplosis tumifex pondant ses oeufs sur les aiguilles du sapin baumier à l’ouverture des bourgeons (Source : Ronald S. Kelley, Vermont Department of Forests, Parks and Recreation, www.forestryimages.org).

9
Figure 1-2. Paradiplosis tumifex, adulte mâle (corps 1,8 mm; antennes 1,7 mm).
Suite à l’accouplement, les femelles pondent leurs œufs dans les bourgeons du sapin
baumier (Abies balsamea Mill.) prêts à débourrer, soit dès que la membrane qui les
recouvre est déchirée. La femelle insère son ovipositeur entre les aiguilles disposées en
faisceau serré et y dépose un ou plusieurs œufs (Figure 1-3). Les œufs sont oblongs
(longueur moyenne de 0,35 mm), parfois blancs translucides (Giese et Benjamin, 1959)
parfois orangés (Osgood et al., 1992) et leur incubation prend de deux à trois jours (Giese
et Benjamin, 1959; Osgood et al., 1992). À l’éclosion, les larves se dirigent sur la surface
adaxiale de l’aiguille, près de la base, où elles commencent à se nourrir (West et
Shorthouse, 1982).

10
Figure 1-3. Oeufs de Paradiplosis tumifex ou de Dasineura balsamicola (longueur = 0,35 mm) sur une jeune aiguille de sapin.
Les larves de premier stade (L-1) se nourrissent en suçant le contenu des cellules de la
jeune aiguille sans causer de blessures ou de nécroses cellulaires (West et Shorthouse,
1982). Selon une hypothèse de Shorthouse et Rohfritsch (1992), certains composés de la
salive des larves stimulent l’induction de la galle. La nature et le mode d’action de ces
composés ne sont cependant pas bien connus à ce jour (Stone et Schönrogge, 2003). Ces
composés provoqueraient la formation d’une galle (Shorthouse et Rohfritsch, 1992) suite à
la prolifération et la différenciation des cellules mésophylliennes autour de la larve (L-1)
(West et Shorthouse, 1982; Shorthouse et Rohfritsch, 1992).
La galle se forme en moins d’une semaine et constitue alors une cavité presque close
recouvrant la larve, à l’exception d’un ostiole sur le dessus (West et Shorthouse, 1982).
C’est par ce même ostiole que les larves quittent la galle l’automne suivant. La galle est
composée d’une seule loge de type prosoplasmique simple, i.e. qu’elle est formée de tissus
différenciés en zones bien définies, de taille et forme déterminées (West et Shorthouse,
1982). Habituellement, une seule galle est formée par aiguille, mais lors de grande
infestation de P. tumifex, plusieurs galles peuvent se juxtaposer, voire se fusionner, sur la
même aiguille (West et Shorthouse, 1982). Les larves se nourriraient en suçant le contenu
des cellules tapissant l’intérieur de la galle, appelées cellules nutritives (Shorthouse et
West, 1986). Ces cellules ne sont pas altérées et rarement endommagées par l’activité
d’alimentation de P. tumifex (Shorthouse et West, 1986). Les cellules nutritives et
11 mésophylliennes constituant en grande partie la galle, contiennent une grande concentration
de granules d’amidon, dont se nourriraient les larves (West et Shorthouse, 1982; Osgood et
al., 1992). La concentration d’amidon augmente jusqu’à maturation complète de la galle,
soit vers la mi-juillet, puis diminuerait lors du vieillissement de la galle (West et
Shorthouse, 1982; Osgood et al., 1992). La galle provoque la sénescence et la chute
prématurée de l’aiguille, l’automne suivant (West et Shorthouse, 1982; Osgood et al.,
1992).
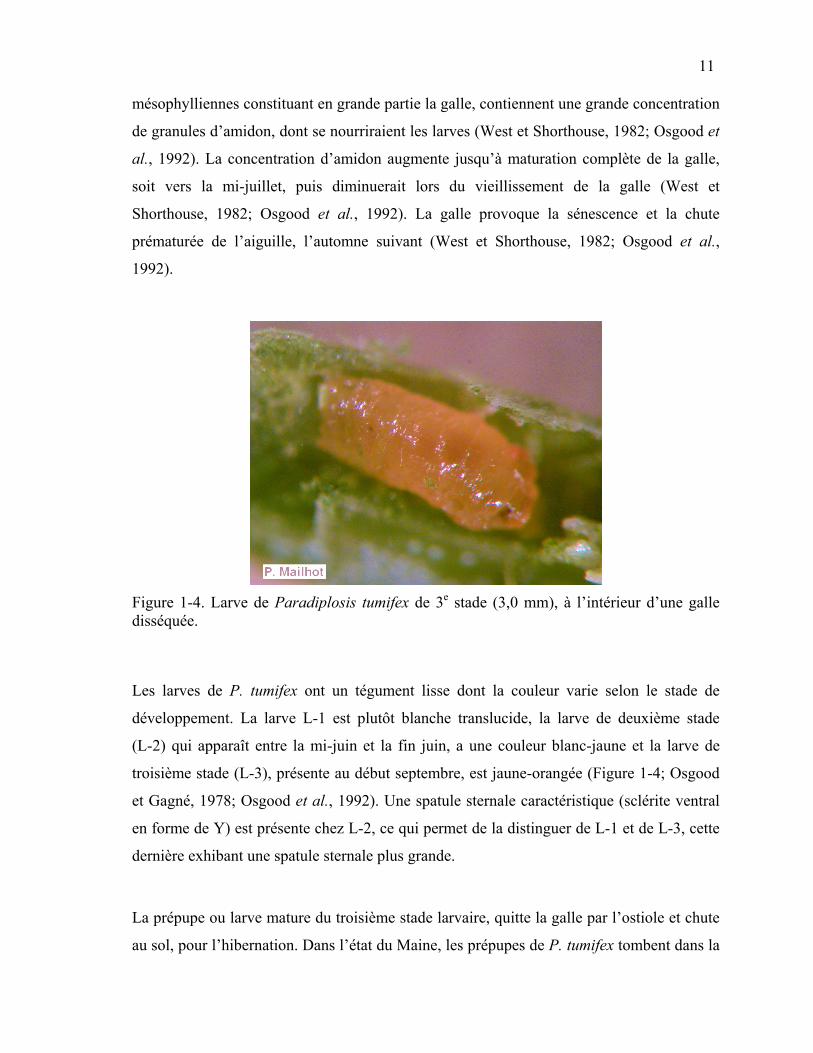
Figure 1-4. Larve de Paradiplosis tumifex de 3e stade (3,0 mm), à l’intérieur d’une galle disséquée.
Les larves de P. tumifex ont un tégument lisse dont la couleur varie selon le stade de
développement. La larve L-1 est plutôt blanche translucide, la larve de deuxième stade
(L-2) qui apparaît entre la mi-juin et la fin juin, a une couleur blanc-jaune et la larve de
troisième stade (L-3), présente au début septembre, est jaune-orangée (Figure 1-4; Osgood
et Gagné, 1978; Osgood et al., 1992). Une spatule sternale caractéristique (sclérite ventral
en forme de Y) est présente chez L-2, ce qui permet de la distinguer de L-1 et de L-3, cette
dernière exhibant une spatule sternale plus grande.
La prépupe ou larve mature du troisième stade larvaire, quitte la galle par l’ostiole et chute
au sol, pour l’hibernation. Dans l’état du Maine, les prépupes de P. tumifex tombent dans la

12 litière de la mi-septembre à la mi-octobre (Osgood et al., 1992). À la fin de la période
d’hibernation dans le sol, P. tumifex s’empupe, soit vers la fin avril ou au début mai, au
Maine (Osgood et al., 1992). La pupe ou nymphe est de type obtecte i.e. que ses appendices
ne sont pas complètement exposés et libres, mais plutôt fusionnés à l’enveloppe
tégumentaire, habituellement bien sclérifiée, qui recouvre l’insecte (Gullan et Cranston,
2000). La pupe est tout d’abord complètement orangée puis peu de temps avant
l’émergence, les appendices (tête, ailes et pattes) noircissent (Figure 1-5; Akar, 1987). Ce
stade est complété en une à deux semaines (P. Mailhot, notes personnelles), puis la
cécidomyie du sapin émerge du sol sous forme de moucheron adulte.
Figure 1-5. Pupe de Paradiplosis tumifex (2,5 mm) dont les extrémités (tête, ailes et pattes) noircissent après environ cinq jours de pupaison, selon la température.
Selon une étude effectuée au Maine, les infestations cycliques de P. tumifex atteignent des
pics de populations tous les 6 à 10 ans (Osgood et al., 1992). En période épidémique, les
populations peuvent infester plus de 80 % des pousses annuelles de sapin et l’infestation
peut durer de une à trois années consécutives. La dynamique de population de P. tumifex
sera discutée et approfondie plus loin.
1.3.2. Biologie de la cécidomyie inquiline des galles Dasineura balsamicola Dans les galles induites par la cécidomyie du sapin, peut s’introduire la larve d’une seconde
espèce de cécidomyie : la cécidomyie inquiline des galles du sapin Dasineura balsamicola
(Lintner).

13 L’inquilinisme est une association entre deux espèces animales où l’espèce inquiline profite
d’un abri que lui procure l’hôte. Par exemple, les éponges abritent une multitude
d’organismes (bactéries et crustacés) inquilins (Harris 1996). Chez les insectes, l’inquiline
a très fréquemment un effet destructeur sur son hôte (Shorthouse, 1980; Torre-Bueno,
1989 ; Ronquist, 1994). Les genres Dasineura et Macrolabis (Cecidomyiidae) comportent
un grand nombre d’espèces inquilines.
Dans certains cas, il est admis que l’espèce inquiline, très proche d’un point de vue
taxonomique de son hôte, aurait évolué à partir de l’insecte galligène auquel il est associé
(Ronquist, 1994). D’ailleurs, un grand nombre d’espèces inquilines ont longtemps été
confondues avec les galligènes auxquels elles sont associées à cause de leurs morphologies
et de leurs cycles de vie analogues (Osgood et Gagné, 1978; Ronquist, 1994). L’origine
évolutive de l’inquilinisme est encore controversée mais Ronquist (1994) a émis deux
hypothèses pour les cas où un lien monophylétique (descendance récente d’une espèce
commune) existe entre les insectes galligènes et inquilins associés. La première suppose
que la compétition pour la ressource végétale, souvent très spécifique en temps et lieu pour
les galligènes, a entraîné l’évolution de l’inquilinisme. La seconde suppose que
l’inquilinisme a évolué d’un comportement de ponte mutualiste où plusieurs femelles
exploitent ensemble un même site. Certains individus ayant perdu la capacité d’induire une
galle auraient alors profité de l’abri de leurs homologues.
Dasineura balsamicola a tout d’abord été confondue avec P. tumifex et présumée être
l’espèce responsable de la formation de galles sur les aiguilles du sapin baumier (Giese et
Benjamin, 1959). Les expériences d’élevage de Osgood et Gagné (1978), ont révélé deux
espèces distinctes de cécidomyies dans la galle, soit D. balsamicola et Paradiplosis sp..
Osgood et Gagné (1978) ont alors clarifié le rôle de chacune, montrant que P. tumifex est
l’espèce galligène et que D. balsamicola est une espèce de cécidomyie inquiline des galles.
Osgood et Gagné (1978) ont déduit que D. balsamicola n’induit pas la formation de la galle
car elle n’est jamais retrouvée seule dans une galle en début de saison. Peu de temps après
la formation de la galle, D. balsamicola s’y trouve toujours en présence d’une larve de

14 P. tumifex, morte ou vivante (Osgood et Gagné, 1978). Les larves inquilines de premier
stade cherchent très activement des larves L-1 galligènes, et dès qu’elles entrent en contact
avec l’une d’elles, elles s’immobilisent, indiquant que D. balsamicola est attirée par
P. tumifex plutôt qu’à une caractéristique structurelle de la surface de l’aiguille (Akar et
Osgood, 1987). Les deux espèces se trouvant côte à côte, lors de la prolifération des
cellules végétales induite par P. tumifex, D. balsamicola est encloisonnée avec celle-ci dans
la galle (Shorthouse et West, 1986). À l’intérieur de la galle, D. balsamicola se nourrirait
indépendamment de P. tumifex, en suçant le contenu des cellules nutritives tapissant la base
de la loge de la galle (Shorthouse et West, 1986). La larve inquiline complète son
développement à l’intérieur des cloisons végétales sans qu’il n’y ait d’interférence
apparente avec P. tumifex. Par contre, avant d’atteindre le troisième stade larvaire,
P. tumifex, en présence de la cécidomyie inquiline, meurt, probablement tuée par cette
dernière (Shorthouse et West, 1986; Osgood et al. 1992). Dasineura balsamicola n’altère
pas la galle de P. tumifex alors que d’autres inquilines entraînent des modifications
importantes dans les structures de la galle même si elles sont incapables de l’induire
(Shorthouse, 1980; Ronquist, 1994). Une différence peut toutefois être notée dans la
concentration d’amidon qui est plus faible dans une galle où seule D. balsamicola est
toujours active (Shorthouse et West, 1986). De plus, comme la larve inquiline n’induit pas
la formation de la galle, Shorthouse et West (1986) suggèrent que si la larve de P. tumifex
meurt avant la maturation de la galle, D. balsamicola pourrait aussi mourir précocement à
cause de l’espace restreint de la galle immature et de carences nutritives.

15
Figure 1-6. Larves de deuxième stade de Dasineura balsamicola (gauche) et de Paradiplosis tumifex (droite) dans la même galle, disséquée en laboratoire. Photo : S. Boudreault.
Dasineura balsamicola a une morphologie et un cycle de vie très semblable à ceux de
P. tumifex mais les deux espèces se distinguent sur quelques points. Tout d’abord,
l’émergence des adultes de D. balsamicola est bien synchronisée avec celle de P. tumifex
mais parfois avec quelques jours de retard (Osgood et Gagné, 1978; Akar, 1987; Jean et al.,
2003). La morphologie des articles antennaires et des ailes des adultes diffère également
(Figures 1-2 et 1-7; Osgood et Gagné, 1978). Les œufs des deux espèces ne sont pas
morphologiquement différentiables, mais la couleur des larves est différente. La larve L-1
de D. balsamicola est jaune alors que celle de P. tumifex est blanc translucide; les larves
L-2 et L-3 de D. balsamicola sont oranges alors que celles de P. tumifex sont plus claires
(Figure 1-6; Osgood et Gagné, 1978). De plus, la larve L-2 de D. balsamicola ne possède
pas de spatule sternale comme celle de P. tumifex.

16
Figure 1-7. Dasineura balsamicola adulte mâle (corps 1,8 mm; antennes 1,7 mm). Photo : P. Mailhot.
Contrairement à P. tumifex, le tégument des larves de D. balsamicola est bosselé ou
«rugueux» avec des soies et spinules (Osgood et Gagné, 1978). Shorthouse et West (1986)
suggèrent que les spinules de la larve de D. balsamicola provoquent l’abrasion du tégument
mince de la larve de P. tumifex et entraîneraient éventuellement, sa mort. Ils suggèrent que
l’espace disponible dans la galle diminue lorsque les larves grossissent et que P. tumifex ne
peut éviter le contact de D. balsamicola. Certaines espèces d’inquilines détruisent leur hôte
très tôt dans leur développement (Shorthouse, 1980) alors que D. balsamicola peut
cohabiter avec son hôte durant tout le premier stade larvaire et une partie du second
(Osgood et Gagné, 1978).
À l’instar de P. tumifex, la prépupe de D. balsamicola dont la croissance est terminée,
quitte la galle et chute au sol, pour l’hibernation. Si P. tumifex gagne la litière aussi tôt qu’à
la mi-septembre (Shorthouse et West, 1986; Osgood et al., 1992), D. balsamicola quitte la
galle plus tard, à partir de la mi-octobre, jusqu’au début décembre (Osgood et al., 1992). À
la fin de la période d’hibernation dans le sol, les deux espèces s’empupent, soit vers la fin
avril ou au début mai, au Maine (Osgood et al., 1992). La nymphe est aussi de type obtecte
chez D. balsamicola mais s’empupe dans un cocon alors que P. tumifex n’en tisse pas
(Osgood et Gagné, 1978). Suite à leur pupaison d’une à deux semaines, selon la
température, elles émergent du sol sous forme adulte (Figure 1-7).

17 L’aire de distribution de D. balsamicola est identique à celle de son hôte P. tumifex, et à
celles des sapins baumier et fraser, soit le nord-est de l’Amérique du Nord.
1.3.3. Dynamique des populations Au Maine, une étude de la dynamique des populations de P. tumifex et de D. balsamicola
échelonnée sur près de 18 années a permis d’observer que les cycles d’infestation de
P. tumifex atteignent des pics de populations environ tous les six à dix ans (Osgood et al.,
1992). Durant les fortes infestations, le taux de défoliation des pousses, suite à la chute des
aiguilles, peut atteindre près de 40% (Martineau, 1985; Osgood et al., 1992). Selon
Martineau (1985) qui discute des niveaux d’infestation observés au Québec depuis les
années 1940, un niveau d’infestation faible est atteint dès que 1 % des rameaux sont
infestés. La période épidémique de la galligène persiste généralement de un à trois ans (et
plus dans certains cas) puis un effondrement des populations survient (Osgood et al., 1992).
C’est ce qui s’est produit récemment dans les plantations de sapins baumiers du sud du
Québec, lorsqu’une importante infestation observée de 2000 à 2002, a été suivie d’une forte
chute des populations en 2003 (Jean et al., 2003; Cloutier et al., 2006).
Les densités de population de l’inquiline semblent suivre celles de P. tumifex et une relation
de densité-dépendance entre les deux espèces a été observée (Osgood, et al., 1992). Les
variations des populations galligènes ont un effet positif direct sur les populations
inquilines, mais avec un décalage d’une année. Cette relation rappelle l’interaction typique
observée dans les modèles de dynamique de population d’un hôte avec son parasitoïde
(Osgood et al., 1992; Ginzburg et Taneyhill, 1994; Berryman, 1996). Comme l’inquiline
cause la mort de la cécidomyie du sapin en association dans la galle, Shorthouse et West
(1986) ont suggéré que l’espèce inquiline régule les populations de la cécidomyie du sapin.
Les populations de cécidomyies ne varient pas uniquement dans le temps mais également
d’un lieu à l’autre, et ce même à l’intérieur d’une région donnée. En effet, des échantillons
de pousses de sapin récoltées dans cinq plantations de localités différentes dans la région

18 centre-est du Maine, ont révélé des fluctuations entre les sites (Osgood et al., 1992). Le
pourcentage d’infestation de P. tumifex pouvait varier de 5 à 45% à l’intérieur d’une même
année. L’étude de Osgood et al. (1992) démontre toutefois un bon synchronisme
d’infestation entre les différents sites avec parfois un léger décalage. Ce synchronisme sur
un territoire donné suggère que les sapins des milieux naturels sont affectés environ au
même moment que les sapins de plantations, mais l’information relative à ce sujet n’est pas
disponible.
En plus de la cécidomyie inquiline D. balsamicola, les facteurs de mortalité naturelle de
P. tumifex incluent d’autres ennemis naturels tels que des oiseaux, des champignons
entomopathogènes et plusieurs espèces de parasitoïdes (Osgood et al., 1992). Au Maine, la
mésange à tête noire est un prédateur des larves arrivant à maturité dans la galle (Osgood et
al., 1992).
Pour ce qui est des champignons, un basidiomycète de l’espèce Uredinopsis mirabilis
Magnus (Uredinales : Pucciniastraceae) serait pathogène de la cécidomyie du sapin lorsque
ses hyphes envahissent les galles (Bergdahl et Mazzola, 1985). Ce champignon cause
également la mort de D. balsamicola ou des parasitoïdes associés, s’il y a lieu (Bergdahl et
Mazzola, 1985). D’autres espèces de champignons découverts sur le sapin baumier
colonisent l’intérieur des galles, telles que Milesia marginalis Faull et Wats. (Osgood et al.,
1992), Aspergillus sp. (MacGown et Osgood, 1972), Hormonema dematioides Lagerberg et
Melin et Phyllosticta sp. (Petrini et al., 1989). Toutefois, la nature de leurs associations
avec les habitants des galles et leurs véritables conséquences sur le développement de
P. tumifex, D. balsamicola ou des parasitoïdes ne sont pas connues.
Aux États-Unis, dix-sept espèces de parasitoïdes ont été associées aux galles de la
cécidomyie du sapin. Pour la majorité d’entre-elles, aucune évidence expérimentale ne
confirme leur relation spécifique avec la cécidomyie du sapin (MacGown et Osgood, 1972).
L’étude de Connor et Osgood (1979), au Maine, a identifié six espèces d’hyménoptères
parasitoïdes (quatre Chalcidoidea et deux Proctotrupoidea), obtenues à partir de l’élevage
des larves de la cécidomyie du sapin (Tableau 1-1). Dans la littérature, il n’y a aucune

19 mention de parasitoïde associé à la cécidomyie inquiline des galles du sapin (Connor et
Osgood, 1979). La distribution saisonnière, au Maine, de ces six parasitoïdes est décrite
sommairement par MacGown et Osgood (1972).
Tableau 1-1. Taxonomie des parasitoïdes hyménoptères associés à la cécidomyie du sapin Paradiplosis tumifex décrits par MacGown et Osgood (1972).
Super-famille Famille Sous-famille Espèce identification 1972
Espèce nom actuel
Proctotrupoidea Platygastridae Platygastrinae Platygaster abicolis
Platygaster abicolis
Platygastrinae Platygaster mainensis
Platygaster mainensis
Chalcidoidea Eulophidae Eulophinae Tetrastichus marcovitchi
Aprostocetus marcovichi
Tetrastichinae Tetrastichus whitmani
Quadrastichus whitmani
Tetrastichinae Tetrastichus solidaginis
Quadrastichus* solidaginis
Tetrastichinae Tetrastichus cecidivorus
Tetrastichus cecidivorus
Encyrtidae Encyrtinae Pseudencyrtus borealis
Pseudencyrtus borealis
* Cette espèce n’a pas été obtenu par élevage, lors de l’étude de Connor et Osgood (1979) mais observé en grand nombre lors de l’étude de MacGown et Osgood (1972).
Platygaster abicollis MacGown et Osgood (Hymenoptera : Platygastridae) et P. mainensis
MacGown et Osgood (Hymenoptera : Platygastridae) sont des endoparasites. La ponte a
lieu au printemps, dans les larves L-1 de P. tumifex peu de temps après leur éclosion. Dans
la plupart des cas, une seule larve de Platygaster sp. par larve hôte est observée, mais
l’incidence de superparasitisme, où deux larves ou plus se côtoient, est fréquent et jusqu’à
cinq larves ont été rapportées par MacGown et Osgood (1972). Le taux de survie des larves
de Platygaster sp. en situation de superparasitisme n’a pas été déterminé.
Aprostocetus marcovichi Crawford (Hymenoptera : Eulophidae) et Quadrastichus
whitmani Girault (Hymenoptera : Eulophidae) sont des ectoparasites de la larve de
cécidomyie, alors que Tetrastichus cecidivorus MacGown et Quadrastichus solidaginis

20 (Burks) sont des endoparasites du stade larvaire. Leurs cycles vitaux sont décrits par
MacGown et Osgood (1972). Pseudencyrtus borealis MacGown (Hymenoptera :
Encyrtidae) est un endoparasite de la larve et l’adulte est actif tout au long de l’été
(MacGown et Osgood, 1972).
En plantations de sapins, certaines pratiques culturales, entre autres le choix de l’espèce
cultivée, minimisent les dommages causés par la cécidomyie du sapin. Les sapins dont les
bourgeons débourrent plus tardivement sont moins attaqués par la galligène, aussi, les
génotypes ou espèces plus tardives, telles que le sapin fraser (Abies fraseri) seraient à
préconiser (Osgood et al., 1992; RAP, 2002). La plupart des producteurs de sapins de Noël
effectuent la taille des arbres à chaque année, dans le but de rencontrer les critères
esthétiques des clients. On peut supposer que cette pratique permettrait de réduire les
populations de cécidomyies car elle élimine des sections de branches infestées.
Lors de cette étude et depuis plusieurs années, l’insecticide recommandé par le Ministère de
l’Agriculture, des Pêcheries et de l’Alimentation du Québec (MAPAQ) contre la
cécidomyie du sapin était le Diazinon (RAP, 2002). C’est un organophosphate très toxique
et à large spectre (EPA, 2000). Selon l’agence de protection environnementale des États-
Unis (EPA), il est particulièrement toxique pour les mammifères, les oiseaux et les insectes
bénéfiques tels que l’abeille domestique (EPA, 2000). Pour que le traitement soit efficace,
il doit être effectué à l’intérieur d’une courte période au printemps, avant que la larve de
P. tumifex ne soit protégée par la galle (RAP, 2002). De plus, le Diazinon, est présentement
sous révision et pourrait bien être retiré des marchés dû à sa grande toxicité (ARLA, 2000;
EPA, 2000). Afin de protéger les plantations de sapins lors des épidémies de la cécidomyie
du sapin, des solutions de rechange devront être développées. Des méthodes alternatives
aux insecticides chimiques sont à préconiser. Pour cela, les connaissances sur la
cécidomyie du sapin et sur son écologie au Québec, devront être enrichies. En effet, jusqu’à
présent les informations publiées proviennent principalement des états du Maine et du
Wisconsin. Aucune évidence ne suggère que celles-ci sont valables aux plantations de
l’Estrie, de la Beauce et des autres régions productrices au Québec, particulièrement quant
à la distribution des parasitoïdes. Cette étude, entreprise en 2002 à l’Université Laval,

21 s’inscrit dans un projet visant à pallier cette lacune, avec l’aide de l’Association des
producteurs d’arbres de Noël du Québec et du Réseau Biocontrôle (Cloutier et al., 2006).
1.4. Hypothèses générales La cécidomyie du sapin P. tumifex a de nombreux ennemis naturels répertoriés au Maine et
au Wisconsin. Par extension, on peut s’attendre à ce qu’une grande diversité biologique de
parasitoïdes ainsi que la cécidomyie inquiline D. balsamicola soient présents dans les
plantations de sapins du sud du Québec. L’inquilinisme et le parasitisme seront importants
dans la régulation des populations et provoquent la fin des cycles épidémiques.
1.5. Objectifs Pour être en mesure de vérifier les hypothèses générales avancées, les objectifs de cette
étude étaient de :
• examiner la densité de P. tumifex dans différentes plantations;
• déterminer le taux d’occupation des galles par les ennemis naturels (inquilines,
endoparasitoïdes et ectoparasitoïdes);
• identifier les parasitoïdes associés à la cécidomyie du sapin au Québec et examiner
la saisonnalité de ces espèces;
• évaluer le rôle de l’inquilinisme et du parasitisme dans l’effondrement des
populations de P. tumifex, survenu en 2002 et documenter les interactions entre ces
derniers.

22
1.6. Bibliographie
AAC. 2004. Agriculture et Agroalimentaire Canada, Prévisions relatives à la cécidomyie du blé dans l’ouest canadien en 1999. http://res2.agr.ca/saskatoon/result/cecidomyieble1999-wheatmidge1999_f.html ; consulté le 7 avril 2006.
Akar, H.Jr. 1987. Some aspects of the biology and rearing of two species of midges,
Paradiplosis tumifex and Dasineura balsamicola, (Diptera : Cecidomyiidae) found in galled needles of balsam fir, Abies balsamea. Thesis. University of Maine. 42 pages.
Akar, H. et E.A. Osgood. 1987. The method by wich larvae of Dasineura balsamicola
(Lintner) (Diptera : Cecidomyiidae) gain access to the interior of galls induced by Paradiplosis tumifex Gagné (Diptera : Cecidomyiidae). The Canadian Entomologist 119 : 395-396.
APANQ. 2005. Mémoire de l’Association des producteurs d'arbres de Noël du Québec à la
Direction des politiques en milieu terrestre Ministère du Développement durable, de l’Environnement et des Parcs le 1er septembre 2005. http://www.apanq.qc.ca/index1.html ; consuté le 9 février 2006.
ARLA. 2000. Agence de réglementation de la lutte antiparasitaire. Mise à jour sur la
réévaluation du diazinon au Canada. www.hc-sc.gc.ca/pmra-arla/ ; consulté le 20 mars 2006.
Bagatto, G., L.C. Paquette et J.D. Shorthouse. 1996. Influence of galls of Phanacis taraxi
on carbon partitioning within common dandelion Taraxacum officinale. Entomologia Experimentalis et Applicata 79 : 111-117.
Bergdahl, D.R. et M. Mazzola. 1985. Some relationships between Uredinopsis mirabilis,
Paradiplosis tumifex and Dasineura balsamicola in the needle of Abies balsamea. Canadian Journal of Forest Research 15: 551-554.
Berryman, A.A. 1996. What causes population cycles of forest Lepidoptera? Trends in
Ecology and Evolution 11 : 28-32. Cloutier, C., P. Mailhot et J. Brodeur. 2006. La cécidomyie du sapin a-t-elle trop d’ennemis
naturels? Le Naturaliste Canadien 130 : 32-36. Connor, J.Y. et E.A Osgood. 1979. Host determination of Chalcidoid and Proctotrupoid
Hymenoptera associated with two species of midges (Diptera : Cecidomyiidae) found in needle galls of balsam fir. The Canadian Entomologist 111 : 945-947.

23 EPA. 2000. U.S. Environmental Protection Agency. Organophosphate pesticides;
Documents for Diazinon. www.epa.gov/pesticides/op/diazinon.htm ; consulté le 20 mars 2006.
Fellt, E.P. 1940. Plant galls and gall makers. Comstock Publishing Company, inc. New
York. 364 pages. Gagné, J.R. 1981. Manual of Nearctic Diptera, vol. 1. Chapter 16 : Cecidomyiidae.
Monograph No.27. Agriculture Canada, Ottawa, pages 257-292. Giese, R.L. et D.M. Benjamin. 1959. The biology and ecology of the balsam gall midge in
Wisconsin. Forest Science 5 : 193-208. Ginzburg, L.R. et D.E. Taneyhill, 1994. Population cycles of forest Lepidoptera : a
maternal effect hypothesis. Journal of Animal Ecology 63 : 79-92. Gullan, P.J. et P.S. Cranston. 2000. The insects, an outline of entomology. Deuxième
édition. Blackwell science. 470 pages. Hanula, J.L. 1991. Seasonal abundance and control of the rhododendron gall midge,
Clinodiplosis rhododendri (Felt), in container grown Rhododendron catawbiense Michaux. Journal of Environmental Horticulture 9: 68-71.
Harris, C.L. 1996. Concepts in zoology. 2e Édition. Harper Collins College Publishers, inc.
485 pages. INRA. 1998. Institut National de la Recherche Agronomique. Cécidomyie orangée du blé.
http ://www.inra.fr.hyppz/ravageur/3sitmos.htm ; consulté le 7 avril 2006. Jean, C., P. Mailhot, J. Brodeur et C. Cloutier. 2003. Outils de lutte intégrée contre la
cécidomyie du sapin dans les plantations d’arbres de Noël. Rapport de recherche CDAQ. 18 pages.
MacGown, M.W. et E.A Osgood. 1972. Taxonomy and biology of Chalcidoid and
Proctotrupoid Hymenoptera associated with Dasineura balsamicola (Diptera : Cecidomyiidae) in Maine. The Canadian Entomologist 104 : 1259-1269.
Mani, M.S. 1964. Ecology of plant galls. Monographiae biologicae. vol.XII. éditeurs W.W.
Weisbach et P. Van Oye. The Hague. 434 pages. Martineau, R. 1985. Insectes nuisibles des forêts de l’est du Canada. Éditions Marcel
Broquet Inc. Ottawa. pages 108-110. Meyer, J. 1987. Plant galls and gall inducers. Éditeur Gebrüder Borntraeger. Berlin.
291 pages.

24 Murakami, M. 1999. Effect of avian predation on survival of leaf-rolling lepidopterous
larvae. Researches on Population Ecology 41: 135-138. Osgood, E.A., R.L. Bradbury et F.A. Drummond. 1992. The balsam gall midge - an
economic pest of balsam fir Christmas trees. Maine Agricultural Experiment Station Technical Bulletin 151 : 1-30.
Osgood, E.A. et R.J. Gagné. 1978. Biology and taxonomy of two gall midges (Diptera :
Cecidomyiidae) found in galls on balsam fir needles with description of a new species of Paradiplosis. Annals of the Entomological Society of America 71 : 85-91.
Petrini, L.E., O. Petrini et G. Laflamme. 1989. Recovery of endophytes of Abies balsamea
from needles and galls of Paradiplosis tumifex. Phytoprotection 70 : 97-103. Poinar, G. Jr. 2003. Cecidomyiidae (Diptera) as insect parasitoids. Proceedings of the
Entomological Society of Washington 105 : 499-501. RAP. 2002. Réseau d’avertissement phytosanitaire: Bulletin d’information. Cécidomyie du
sapin. www.agr.gouv.qc.ca/dgpar/rap ; consulté le 20 mars 2006. Ronquist, F. 1994. Evolution of parasitism among closely related species: phylogenetic
relationships and the origin of inquilinism in gall wasps (Hymenoptera : Cynipidae). Evolution 48 : 241-266.
van Schelt, J. et S. Mulder. 2000. Improved methods of testing and release of Aphidoletes
aphidimyza (Diptera : Cecidomyiidae) for aphid control in glasshouses. European Journal of Entomology 97 : 511-515.
Shorthouse, J.D. 1980. Modification of the galls of Diplolepis polita by the inquiline
Periclistus pirata. Bulletin de la société botanique de France 127 : 79-84. Shorthouse, J.D. et R.J. West. 1986. Role of the inquiline, Dasineura balsamicola
(Diptera : Cecidomyiidae) in the balsam fir needle gall. Proceedings of the Entomological Society of Ontario 117 : 1-7.
Shorthouse, J.D. et O. Rohfritsch. 1992. Biology of insect-induced galls. Oxford University
Press, Oxford, Royaume-Uni, 285 pages. Speker, D.R. et W.T. Johnson. 1988. Biology and immature stages of rhododendron gall
midge Felt (Diptera : Cecidomyiidae). Proceedings of the Entomological Society of Washington 90 : 343-355.
Stone, G.N. et K. Schönrogge. 2003. The adaptive significance of insect gall morphology.
Trends in Ecology and Evolution 18 : 512-522.

25 Torre-Bueno, J.R. 1989. The Torre-Bueno glossary of entomology. Compilé par Stephen
W. Nichols, incluant un supplément de George S. Tulloch. New York Entomological Society, New York, USA. 840 pages.
West, R.J. et J.D. Shorthouse. 1982. Morphology of the balsam fir needle gall induced by
the midge Paradiplosis tumifex (Diptera : Cecidomyiidae). Canadian Journal of Botany 60 : 131-140.
West, S.A., E.A. Herre, D.M. Windsor et P.R.S. Green. 1996. The ecology and evolution
of the new world non-pollinating fig wasp communities. Journal of Biogeography 23 : 447-458.
Williams, A.G. et T.G. Whitham. 1986. Premature leaf abscission: an induced plant defense
against gall aphid. Ecology 67 : 1619-1627.

Chapitre 2. Le parasitisme et l’inquilinisme comme facteurs de régulation des populations de la cécidomyie
du sapin Paradiplosis tumifex Gagné.

27
2.1. Résumé Le marché des sapins de Noël (Abies balsamea Mill.) est en pleine progression au Québec,
tant pour le commerce local que pour l’exportation. Malheureusement, le sapin est la proie
de ravageurs qui endommagent son aspect esthétique et qui diminuent sa valeur sur le
marché. C’est le cas de la cécidomyie du sapin (Paradiplosis tumifex Gagné), un diptère
galligène dont les infestations peuvent indirectement causer la défoliation des arbres de
Noël. Des infestations majeures de P. tumifex surviennent périodiquement, comme ce fut le
cas de 2000 à 2002 au Québec. Les facteurs déterminant le début et la fin des cycles
d’infestation sont mal compris ainsi que le rôle joué par les ennemis naturels, notamment la
cécidomyie inquiline des galles (Dasineura balsamicola Lintner) et diverses espèces
d’ectoparasitoïdes et d’endoparasitoïdes.
L’objectif de cette étude était d’identifier les ennemis naturels et de documenter les
interactions entre ces derniers afin d’évaluer le rôle de l’inquilinisme et du parasitisme dans
l’effondrement des populations de P. tumifex, survenu au Québec, en 2002. À cet effet,
l’évaluation du taux d’infestation des pousses par P. tumifex, la détermination du taux
d’occupation des galles par les ennemis naturels (inquilines, endoparasitoïdes et
ectoparasitoïdes), l’identification partielle des parasitoïdes retrouvés au Québec et la
caractérisation de la distribution saisonnière des parasitoïdes adultes ont été réalisées
Les données ont permis d’estimer des niveaux d’inquilinisme des galles variant de 20 à
60 %, en 2002. Cinq espèces de parasitoïdes associés à P. tumifex appartenant aux familles
des Eulophidae, Encyrtidae et Platygastridae ont été identifiées. Leur période d’activité au
stade adulte a été caractérisée et leur abondance mesurée. Le taux de parasitisme de
P. tumifex diffère selon les taxons des parasitoïdes. Des données relatives à la présence de
plusieurs parasitoïdes dans une même galle (multiparasitisme) suggèrent des interactions
négatives (interférence) fréquentes entre les ennemis naturels de la cécidomyie du sapin. De
plus, les forts taux d’inquilinisme et de parasitisme indiquent que ces ennemis naturels sont
probablement responsables de l’effondrement des populations de la cécidomyie du sapin.

28
2.2. Introduction La cécidomyie du sapin Paradiplosis tumifex Gagné (Diptera : Cecidomyiidae) est un
ravageur épisodique des plantations de sapins baumiers. Selon des données provenant du
Maine, les périodes d’infestation, où les densités de la cécidomyie du sapin sont élevées,
sont relativement courtes soit de deux à quatre ans et il s’écoule de six à dix ans entre deux
épidémies majeures (Osgood et al., 1992). Paradiplosis tumifex est à l’origine de la
formation de petites galles sur les aiguilles des jeunes pousses en croissance au printemps
(West et Shorthouse, 1982).
Les dégâts occasionnés par P. tumifex sont importants en période de forte infestation car les
aiguilles infestées de galles ont tendance à tomber au sol vers la fin de l’automne, ce qui
laisse des rameaux dénudés d’aiguilles juste avant la période des ventes de sapins de Noël
(West et Shorthouse, 1982). Les arbres fortement infestés ont une valeur esthétique
moindre et sont déclassés sur le marché des sapins de Noël. Les producteurs subissent alors
des pertes économiques importantes.
Une cécidomyie inquiline ainsi que plusieurs espèces de parasitoïdes ont été identifiées
comme ayant un rôle dans la régulation des populations de la cécidomyie du sapin (Giese et
Benjamin, 1959; Osgood et Gagné, 1978; Shorthouse et West, 1986; Osgood et al., 1992).
La cécidomyie inquiline des galles Dasineura balsamicola (Lintner) (Diptera :
Cecidomyiidae), est un facteur de mortalité de P. tumifex qui semble efficace pour diminuer
les densités élevées de P. tumifex (Shorthouse et West, 1986; Osgood et al., 1992).
L’inquiline, par définition, est un organisme qui utilise l’abri de son hôte (Torre-Bueno,
1989). Dasineura balsamicola a un cycle de vie semblable et lié à celui de P. tumifex mais
n’a pas la capacité d’initier la production de la galle sur l’aiguille de sapin, tel que
démontré par Osgood et Gagné (1978, voir aussi Shorthouse et West, 1986; Osgood et al.,
1992). À l’éclosion des œufs, la larve néonate adopte un comportement de recherche de
P. tumifex dans le but de se retrouver inclue dans la galle (avec P. tumifex) au moment de sa
formation (Shorthouse et West, 1986). L’inquiline se nourrit des tissus végétaux de la galle

29
mais vers la fin de l’été ou en automne, elle causera la mort de son hôte. Shorthouse et
West (1986) suggèrent que la mort de P. tumifex soit causée par l’abrasion des spinules de
D. balsamicola sur le tégument mince de son hôte, surtout lorsque la croissance des larves,
dans l’espace restreint de la galle, contribue à leur rapprochement. Selon Shorthouse et
West (1986), l’inquiline est un facteur de mortalité majeur car elle peut occuper plus de
50 % des galles. Osgood et al. (1992) suggèrent une relation de densité-dépendance entre
P. tumifex et D. balsamicola, mais avec un décalage d’une année ce qui reflète l’interaction
typique observée dans les modèles de dynamique de population d’un hôte avec son
parasitoïde (Osgood et al., 1992; Ginzburg et Taneyhill, 1994; Berryman, 1996).
Au Maine, six espèces d’hyménoptères parasitoïdes appartenant aux familles Eulophidae,
Encyrtidae et Platygastridae, ont été associées à P. tumifex comme étant des facteurs de
régulation potentiels (Connor et Osgood, 1979; Osgood et al., 1992). Certaines ont été
observées en grand nombre particulièrement Aprostocetus marcovitchi (Hymenoptera:
Eulophidae), mais les données sont imprécises (MacGown et Osgood, 1972).
L’objectif de cette étude était d’identifier au Québec les ennemis naturels de la cécidomyie
du sapin et de documenter les interactions entre ces derniers, afin d’évaluer le rôle de
l’inquilinisme et du parasitisme dans l’effondrement des populations de P. tumifex, survenu
en 2002 (Jean et al., 2003; Cloutier et al., 2006). À cet effet, l’évaluation du taux
d’infestation des pousses par P. tumifex, la détermination du taux d’occupation des galles
par les ennemis naturels (inquilines, endoparasitoïdes et ectoparasitoïdes), l’identification
des parasitoïdes retrouvés au Québec et l’évaluation de la distribution saisonnière des
parasitoïdes adultes ont été réalisées.
2.3. Matériels et méthodes L’échantillonnage a été effectué dans trois plantations commerciales de sapins baumiers du
Québec méridional. La première plantation était située à Saint-Fortunat (45° 58’ N, 71°
36’ O) dans la région Chaudière-Appalaches, à la limite de la région administrative du
Centre du Québec; la deuxième à Inverness (46° 15’ N, 71° 29’ O) dans la région Centre du

30
Québec et la troisième à Racine (45° 30’ N, 72° 15’ O), en Estrie. La collecte de données a
été effectuée de mai à novembre 2002. L’année suivante, étant donné la très faible
abondance de la cécidomyie du sapin en début de saison, l’échantillonnage a été limité du
8 mai au 16 juin 2003.
2.3.1. Caractéristiques des plantations et parcelles d’échantillonnage Le site de Saint-Fortunat est une plantation de sapins baumiers dont les graines provenaient
d’arbres matures sélectionnés en forêt par le producteur. Deux parcelles (A et B) ont été
échantillonnées. Aucun intrant chimique ni pesticide de synthèse n’a été utilisé. Des
traitements phytosanitaires avec des «produits biologiques» (recette maison du producteur)
étaient effectués sauf dans la parcelle A. Pour limiter l’invasion des mauvaises herbes, les
rangs et les entre-rangs ont été fauchés à quelques reprises pendant la saison estivale. En
2002, les arbres avaient en moyenne six ans dans la parcelle A et huit ans dans la parcelle
B. Les arbres des deux parcelles ont été taillés vers le 1er septembre. La parcelle A a été
réservée pour l’échantillonnage à intervalle régulier, dédié à l’évaluation de la densité des
galles et de la diversité des insectes associés. L’aire d’échantillonnage de la parcelle A était
composée de 30 rangées de 50 arbres et subdivisée en 10 blocs. Chaque bloc était composé
de 15 rangées de 10 arbres numérotés à intervalles pour faciliter l’échantillonnage. La
parcelle B était située à environ 30 mètres de la parcelle A, en plus haute altitude (vers
l’ouest) et séparée par une bande d’arbres divers (peupliers faux-trembles, pins, érables).
Cette seconde parcelle a été utilisée pour récolter des galles en grand nombre dans le but
d’en faire l’élevage, en fin de saison. La parcelle B était composée de neuf rangs de 100
arbres chacun et divisée en cinq blocs de neuf rangs et de 20 arbres. Selon le producteur,
l’infestation de la plantation de Saint-Fortunat par la cécidomyie du sapin avait débuté en
2001.
La plantation de sapins baumiers située à Inverness était sous une régie de type
conventionnel avec pesticides. Pour l’année 2002, un traitement herbicide au Round-Up® a
été appliqué sur les rangs entre les arbres alors que les entre-rangs ont été fauchés. L’année
précédente un traitement au Diazinon® avait été appliqué contre la cécidomyie du sapin,

31
dont l’infestation avait débuté en 2000, selon le producteur. Les arbres, d’environ six ans,
ont été taillés vers le 15 juillet. L’aire d’échantillonnage a été définie de la même manière
que celle de la parcelle A de la plantation de Saint-Fortunat : soit 30 rangées de 50 arbres et
divisée en 10 blocs où chaque bloc était composé de 15 rangées de 10 arbres dont certains
étaient numérotés pour repérage rapide.
La plantation de sapins baumiers de Racine est une plantation à régie biologique sans
aucune application de pesticides. Pour restreindre les mauvaises herbes, les rangs et les
entre-rangs ont été fauchés à quelques reprises pendant la saison estivale. Les arbres étaient
âgés de 13 ou 14 ans et la taille a été effectuée à la mi-août. L’aire d’échantillonnage était
identique à celles de la parcelle A de Saint-Fortunat et de la plantation d’Inverness. Les
données relatives à la situation épidémique de P. tumifex l’année précédant
l’échantillonnage, n’étaient pas disponibles.
2.3.2. Mesure de l’activité des insectes adultes Pour déterminer les périodes d’activité (vol) des parasitoïdes et des cécidomyies adultes,
des pièges collants jaunes (10 x 18 cm, recto-verso de surface collante, Seabright
Laboratories) ont été placés dans la parcelle A de la plantation de Saint-Fortunat. Pour une
question de faisabilité, une seule plantation a été suivie durant toute la saison. Le site de
Saint-Fortunat a été sélectionné parce que l’émergence printanière des cécidomyies et des
parasitoïdes y était particulièrement abondante. Dans chaque bloc, un piège collant était
placé au sommet d’un arbre choisi aléatoirement (au total 10 pièges). Les pièges ont été
installés le 6 mai 2002 et étaient remplacés à intervalles de trois ou quatre jours jusqu’au 13
août, puis une fois par semaine jusqu’au 28 octobre. Les pièges relevés étaient conservés
pour identification ultérieure des cécidomyies et parasitoïdes. Les abondances sont
exprimées en valeurs logarithmiques.

32
2.3.3. Échantillonnage des pousses Pour déterminer l’abondance des insectes à l’étude, des pousses annuelles de sapin
baumier, soit les rameaux et leurs aiguilles en croissance à partir de bourgeons débourrés au
printemps 2002, étaient récoltées à intervalles réguliers. La sélection des arbres
échantillonnés était faite de façon aléatoire dans chaque bloc et le même arbre n’était pas
utilisé lors de deux échantillonnages successifs.
Le nombre d’échantillons de pousses variait d’une plantation à l’autre et selon la date de
collecte (voir ci-dessous) pour des raisons de logistique. La plantation de Racine a été
suivie grâce à la collaboration de M. André Pettigrew du MAPAQ en Estrie, qui y a
effectué la collecte. L’effort d’échantillonnage y a été moins intensif, mais les données
ajoutées permettent toutefois d’avoir une meilleure vue d’ensemble de la situation des
plantations du sud-ouest du Québec. À la plantation de Saint-Fortunat, l’abondance et la
diversité des organismes observés durant l’émergence printanière, semblait pouvoir fournir
le plus d’informations sur les différents organismes associés à la cécidomyies du sapin, ce
qui était le but premier de cette étude (Jean et al., 2003). C’est pourquoi un plus grand
effort d’échantillonnage y a été effectué.
En 2002, dans la parcelle A de Saint-Fortunat, une pousse a été récoltée sur cinq arbres de
chacun des blocs, pour un total de 50 pousses par date d’échantillonnage. La récolte des
pousses a débuté le 6 mai, à raison de deux fois par semaine jusqu’au 13 août, puis ensuite
la fréquence d’échantillonnage a été de une fois par semaine jusqu’au 28 octobre 2002.
Dans les plantations d’Inverness et de Racine, un total de 30 pousses ont été
échantillonnées par date, soit trois pour chacun des blocs. À Inverness, l’échantillonnage a
été effectué du 30 mai au 20 août 2002 à raison de deux fois par semaine au début, et une
seule fois à partir du 6 août. À Racine, le prélèvement de 10 pousses par collecte a débuté le
3 juin au rythme d’une fois par semaine et s’est poursuivi jusqu’au 3 octobre. Ensuite, trois
échantillonnages supplémentaires ont été effectués soit les 17 octobre, 7 et 25 novembre
2002.

33
Chaque pousse récoltée était conservée au réfrigérateur pour être examinée dans les deux à
trois jours suivants, au laboratoire. Au début de la saison, alors que la pousse annuelle se
développe, soit avant la formation des galles, les œufs de cécidomyies étaient dénombrés.
La distinction des œufs de P. tumifex et de D. balsamicola n’a pas été effectuée puisque
aucun critère ne permet de les différencier morphologiquement. La longueur de la pousse,
le nombre d’aiguilles présentes, le nombre d’aiguilles sur lesquelles il y avait des galles de
la cécidomyie du sapin, ainsi que les espèces à l’intérieur des galles ont été déterminés. Les
galles étaient disséquées sous une loupe binoculaire (grossissement: 10X à 40X). À l’aide
d’un scalpel, l’ouverture que forme l’ostiole était prolongée puis les tissus végétaux étaient
découpés de façon à ouvrir la galle sans endommager son contenu. Les larves de P. tumifex
et de D. balsamicola étaient identifiées à l’aide de caractères morphologiques décrits dans
la littérature (Osgood et al., 1992) et leur stade larvaire était déterminé selon la présence ou
l’absence de la spatule sternale et de la couleur de la larve (Osgood et Gagné, 1978). La
présence et le nombre des œufs et des larves d’ectoparasitoïdes observés étaient aussi notés.
Pour vérifier la présence d’endoparasitoïdes non visibles à l’œil nu, des sous-échantillons
des larves de P. tumifex et de D. balsamicola étaient utilisés, comprenant en moyenne 35
larves (de 14 à 68 larves selon leur disponibilité). Ces larves étaient placées entre lame et
lamelle, et observées au microscope à lumière transmise (grossissement 40X à 400X),
permettant de voir les œufs ou larves de parasitoïdes dans l’hémolymphe ou les tissus.
2.3.4. Élevage des parasitoïdes à partir de la récolte d’aiguilles porteuses de galles Les connaissances relatives à la taxonomie des larves de parasitoïdes associés à la
cécidomyie du sapin sont incomplètes. Pour identifier à l’espèce les parasitoïdes associés à
P. tumifex ou à D. balsamicola, il faut identifier des spécimens adultes dont l’hôte de
provenance est incontestable. Pour obtenir ces adultes, 4000 aiguilles portant des galles de
la cécidomyie du sapin ont été récoltées, à Saint-Fortunat, en 2002. Pour s’assurer que les
organismes étaient matures et qu’ils avaient emmagasiné toutes les ressources nutritives
essentielles à leur développement jusqu’au stade d’adulte, des aiguilles avec des galles bien
développées ont été récoltées, en fin de saison (Connor et Osgood, 1979; Akar, 1987). Pour
éviter toute interférence avec la mesure du taux d’infestation et de la densité des

34
organismes, ces aiguilles ont été récoltées dans la parcelle B de la plantation de
Saint-Fortunat. L’échantillonnage a été effectué pendant huit semaines, soit du 23
septembre au 11 novembre 2002. Chaque semaine, 25 arbres choisis aléatoirement à raison
de cinq par bloc dans cinq blocs, ont été échantillonnés. Sur chaque arbre, 20 aiguilles ont
été récoltées, soit cinq par pousse sur quatre pousses différentes.
Les aiguilles récoltées ont été placées individuellement dans une éprouvette de type
Ependorf, identifiée selon la provenance et la date. Le couvercle avait été préalablement
percé à l’aide d’une aiguille fine pour permettre l’aération et du coton dentaire humidifié
avec une solution préservative d’eau distillée (0,75 ml) et de méthyl parabenzène (0,05 %),
était ajouté pour favoriser la survie (Akar, 1987). Pour recréer les conditions de
développement normal durant l’hiver, les galles ont été placées dans des conditions de
dormance selon la méthode décrite par Akar (1987). Afin de simuler des conditions de
dormance hivernale prolongée, les cagettes étaient d’abord placées dans une chambre froide
non-éclairée à 4° C pendant deux semaines pour l’acclimatation des larves. Ensuite, elles
étaient déplacées dans une chambre froide non-éclairée dont la température était maintenue
à 2° C, de 80 à 85% d’humidité relative, pendant huit semaines.
Durant la deuxième semaine de la période de dormance, environ le tiers des galles de
chaque échantillon ont été disséquées pour en déterminer le contenu. Les larves de
cécidomyies et leurs stades étaient identifiés, les ectoparasitoïdes étaient notés, de même
que les galles vides ou celles contenant des larves mortes ou des champignons. Les
dissections avaient pour but de valider l’association de parasitoïdes avec l’espèce de
cécidomyie (P. tumifex ou D. balsamicola) et ainsi vérifier si des endoparasitoïdes non
répertoriés à ce jour, étaient associés à l’inquiline. De plus, le multiparasitisme de fin de
saison était observé sauf pour les endoparasitoïdes, car leur méthode d’observation était
destructive.

35
Suite à la période de huit semaines de «dormance» à 2˚ C, les cagettes étaient replacées à
4° C pour deux semaines d’acclimatation, puis dans une chambre de croissance à 15° C,
avec une humidité relative de 80 % et une photopériode de 16L : 8N, permettant le
développement et l’émergence des adultes. Les cécidomyies et parasitoïdes adultes émergés
étaient identifiés. Leur date d’émergence et leur sexe étaient notés. Les galles n’ayant pas
donné d’émergents ont été gardées sous observation au maximum 335 jours dans ces
conditions.
2.3.5. Identification des spécimens Les parasitoïdes émergés au laboratoire ont d’abord été identifiés sur la base de critères
morphologiques selon une clé d’identification des Hyménoptères (Agriculture Canada,
1993) et celle de MacGown et Osgood (1972). Des spécimens de chaque espèce ont aussi
été envoyés à des spécialistes de la Collection nationale canadienne d'insectes, arachnides
et nématodes (Agriculture et Agroalimentaire Canada) à Ottawa, pour fin d’identification.
Les Platygastridae ont été identifiés par Dr Lubomir Masner, les Eulophidae par Dr John
Huber et les Encyrtidae par Dr Gary Gibson. Des spécimens de références ont été déposés à
la Collection nationale canadienne, au Centre de foresterie des Laurentides du Ministère
des ressources naturelles Canada (MRN) à Québec, ainsi qu’à la Collection d’insectes du
Québec au Complexe scientifique situé à Québec (MRN et MAPAQ).
2.3.6. Analyses des données
2.3.6.1. Taux d’infestation
Le taux moyen d’infestation des pousses par la cécidomyie du sapin a été déterminé pour
les trois plantations à partir du 6 juin et durant toute la saison estivale de 2002. Bien que de
nouvelles galles de P. tumifex ne pouvaient se former au cours de la saison, des échantillons
de pousses amassés à d’autres fins ont également contribué à l’obtention d’un plus grand
échantillon pour cette analyse. Dans le calcul du taux d’infestation, une pousse infestée
correspond à une pousse avec au moins une aiguille porteuse d’une galle de la cécidomyie
du sapin P. tumifex. La moyenne saisonnière du taux d’infestation des pousses ainsi que la

36
moyenne du nombre de galles par pousse ont été calculées comme indices du niveau
d’infestation par P. tumifex. Des analyses de variance à une voie (ANOVA) sur l’effet des
blocs (données non-transformées), le taux d’infestation (données non-transformées) et le
nombre moyen de galles par pousse (données non-transformées) ont été réalisés pour
chaque plantation, à l’aide du logiciel SAS system (PROC GLM).
2.3.6.2. Inquilinisme Le taux d’inquilinisme moyen des galles de P. tumifex a été déterminé pour chaque date
d’échantillonnage. Les galles vides ont été exclues de ce calcul, puisqu’ils était impossible
de savoir si P. tumifex y avait été associé à D. balsamicola.
Les échantillons utilisés pour le calcul du taux d’inquilinisme, ont été sélectionnés en
fonction de la date approximative d’établissement des inquilines, soit à partir du 16 juin
2002, d’après l’examen des galles. Les données ultérieures au 9 septembre 2002 ont été
exclues car la larve de P. tumifex avait probablement commencé à quitter la galle pour
hiverner, selon les observations provenant du Maine (Osgood et al., 1992). Dans le cas de
la plantation d’Inverness, où la densité des galles était très faible, l’échantillonnage a été
terminé le 20 août. Les estimés des niveaux d’inquilinisme n’étaient pas possibles pour la
saison 2003 étant donné le déclin important des populations et la très faible abondance de la
cécidomyie du sapin.
2.3.6.3. Effet de la densité de P. tumifex sur D. balsamicola Les données utilisées pour vérifier l’effet de la densité de P. tumifex sur la réponse (nulle,
linéaire ou exponentielle) de la cécidomyie inquiline, proviennent des échantillons de
Saint-Fortunat, où l’échantillonnage était le plus complet, et couvre la période du 16 juin au
9 septembre 2002. Le logiciel SAS system (PROC GLM) a été utilisé pour exécuter les
tests statistiques. Des régressions ont été effectuées pour caractériser les relations (linéaires
ou quadratiques) entre le nombre de galles par pousse et le nombre d’inquilines, ainsi que
l’influence de la longueur des pousses de sapin sur le nombre d’inquilines. Ces analyses ont

37
été réalisées sur les valeurs non-transformées de densité. L’analyse graphique des résidus a
été effectuée pour s’assurer de la normalité et de l’homogénéité des variances.
2.3.6.4. Ectoparasitisme Le taux moyen d’ectoparasitisme a été déterminé à partir des échantillons de pousses
récoltées à Saint-Fortunat, plantation où l’échantillonnage était le plus intensif, en
considérant la période du 10 septembre au 25 novembre 2002. Le taux d’ectoparasitisme a
été déterminé en fonction du nombre de galles disséquées de P. tumifex où au moins un
ectoparasitoïde était présent. Les galles vides ont été exclues de l’analyse.
2.3.6.5. Endoparasitisme Le taux moyen d’endoparasitisme a été mesuré sur les échantillons de pousses provenant de
Saint-Fortunat, plantation où l’échantillonnage était le plus intensif, durant la période du 10
septembre au 25 novembre 2002. Il a été calculé séparément pour les larves du genre
Platygaster et pour l’ensemble des autres endoparasitoïdes. Les larves L-1 de Platygaster
sp. sont très caractéristiques et de type cyclopiforme, soit une «tête» ronde avec de longues
mandibules falciformes et le reste du corps réduit à une «queue» (Figure 2-1). Leurs
mandibules les distinguent clairement des autres larves endoparasitoïdes qui sont de type
vermiforme, mais dont l’identification n’était pas possible avec la documentation
disponible. Les taux d’endoparasitisme ont été déterminés à partir des données prises par
l’observation de sous-échantillons de larves de P. tumifex au microscope, pour chaque date
d’observation. La même méthode a été utilisée pour évaluer l’endoparasitisme chez
D. balsamicola.

38
Figure 2-1. Quatre larves cyclopiformes de premier stade de Platygaster sp., deux à l’intérieur et deux à l’extérieur d’une larve de cécidomyie du sapin, Paradiplosis tumifex brisée et disposée entre lame et lamelle, observée au microscope.
2.3.6.6. Relations de densité entre la cécidomyie galligène et ses parasitoïdes Les données utilisées pour le calcul de ces relations proviennent des échantillons de
pousses récoltées à Saint-Fortunat ainsi que des observations de larves faites au
microscope. Les échantillons du 10 septembre au 25 novembre 2002 ont été utilisés, dates
auxquelles on suppose que la période de ponte de tous les parasitoïdes est terminée. Les
régressions linéaires effectuées avec le logiciel SAS system (PROC GLM), évaluaient
l’effet du nombre de galles par pousse sur : le nombre d’ectoparasitoïdes, le nombre
d’endoparasitoïdes, le taux d’ectoparasitisme et le taux d’endoparasitisme. Ensuite, l’effet
de la présence et du nombre d’inquilines par pousse sur le taux d’ectoparasitoïdes a été
vérifié par régression linéaire (logiciel SAS, PROC GLM) ainsi que l’effet de la longueur
des pousses sur le nombre d’ectoparasitoïdes. Les analyses de résidus ont été effectuées
telles que décrit précédemment.

39
2.4. Résultats
2.4.1. Taux d’infestation de P. tumifex Les pousses de sapin étaient fortement infestées par P. tumifex aux sites de Saint-Fortunat
et de Racine, où plus de 50 % des pousses contenaient une galle ou plus (Tableau 2-1).
L’infestation était plutôt faible à Inverness, avec moins de 10 % de pousses infestées. Il n’y
avait pas de différences significatives entre les blocs du plan expérimental, et ce dans
aucune des plantations échantillonnées (Tableau 2-1).
Tableau 2-1. Taux saisonnier moyen d’infestation des pousses par Paradiplosis tumifex et nombre moyen de galles par pousse sur sapin baumier (Abies balsamea) dans des plantations de trois localités du sud du Québec, en 2002. Des lettres qui sont différentes dans une même colonne indiquent qu’il y a une différence significative entre les sites (P < 0,05).
Plantations Nombre de pousses
échantillonnées
Bloc n = 10
valeur de P
Taux d’infestation ± écart-type
Nb moyen de galles par pousse
± écart-type
Saint-Fortunat 1639 0,2508 56,72 % ± 11,97 a 4,59 ± 1,58 a
Inverness 599 0,5350 9,37 % ± 7,11 b 0,16 ± 0,16 b
Racine 554 0,0801 61,41 % ± 11,18 a 4,03 ± 1,48 a
2.4.2. Inquilinisme Dans la plantation de Saint-Fortunat, en 2002, le taux moyen d’inquilinisme était de plus de
15 % (Figure 2-2). À Inverness, le taux saisonnier moyen d’inquilinisme estimé était de
12,72 % (écart-type = 27,18 %), alors qu’à Racine ce taux était de 66,54 % (écart-type
18,24 %).
À Saint-Fortunat, la majorité des pousses contenaient moins de huit inquilines et même
lorsqu’il y avait de 50 à 80 galles sur la pousse, on n’y comptait pas plus de cinq inquilines.
Exceptionnellement, une pousse portant 46 galles comptait dix-sept inquilines.
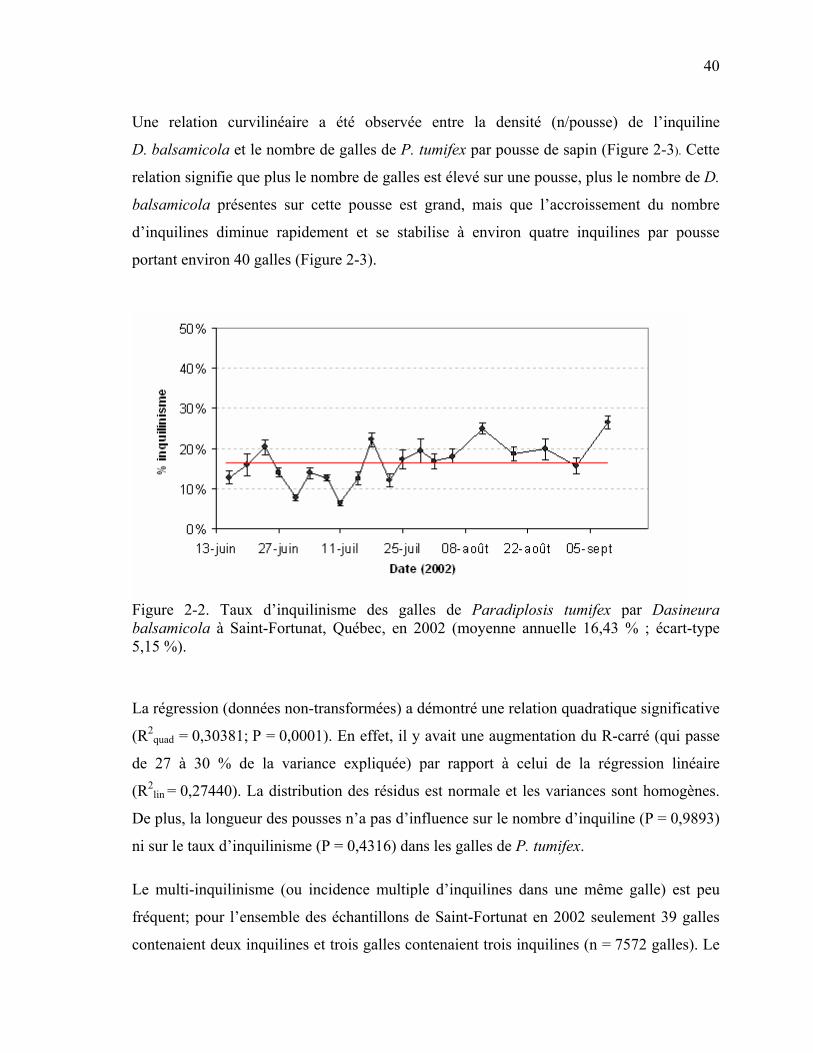
40
Une relation curvilinéaire a été observée entre la densité (n/pousse) de l’inquiline
D. balsamicola et le nombre de galles de P. tumifex par pousse de sapin (Figure 2-3). Cette
relation signifie que plus le nombre de galles est élevé sur une pousse, plus le nombre de D.
balsamicola présentes sur cette pousse est grand, mais que l’accroissement du nombre
d’inquilines diminue rapidement et se stabilise à environ quatre inquilines par pousse
portant environ 40 galles (Figure 2-3).
Figure 2-2. Taux d’inquilinisme des galles de Paradiplosis tumifex par Dasineura balsamicola à Saint-Fortunat, Québec, en 2002 (moyenne annuelle 16,43 % ; écart-type 5,15 %).
La régression (données non-transformées) a démontré une relation quadratique significative
(R2quad = 0,30381; P = 0,0001). En effet, il y avait une augmentation du R-carré (qui passe
de 27 à 30 % de la variance expliquée) par rapport à celui de la régression linéaire
(R2lin = 0,27440). La distribution des résidus est normale et les variances sont homogènes.
De plus, la longueur des pousses n’a pas d’influence sur le nombre d’inquiline (P = 0,9893)
ni sur le taux d’inquilinisme (P = 0,4316) dans les galles de P. tumifex.
Le multi-inquilinisme (ou incidence multiple d’inquilines dans une même galle) est peu
fréquent; pour l’ensemble des échantillons de Saint-Fortunat en 2002 seulement 39 galles
contenaient deux inquilines et trois galles contenaient trois inquilines (n = 7572 galles). Le
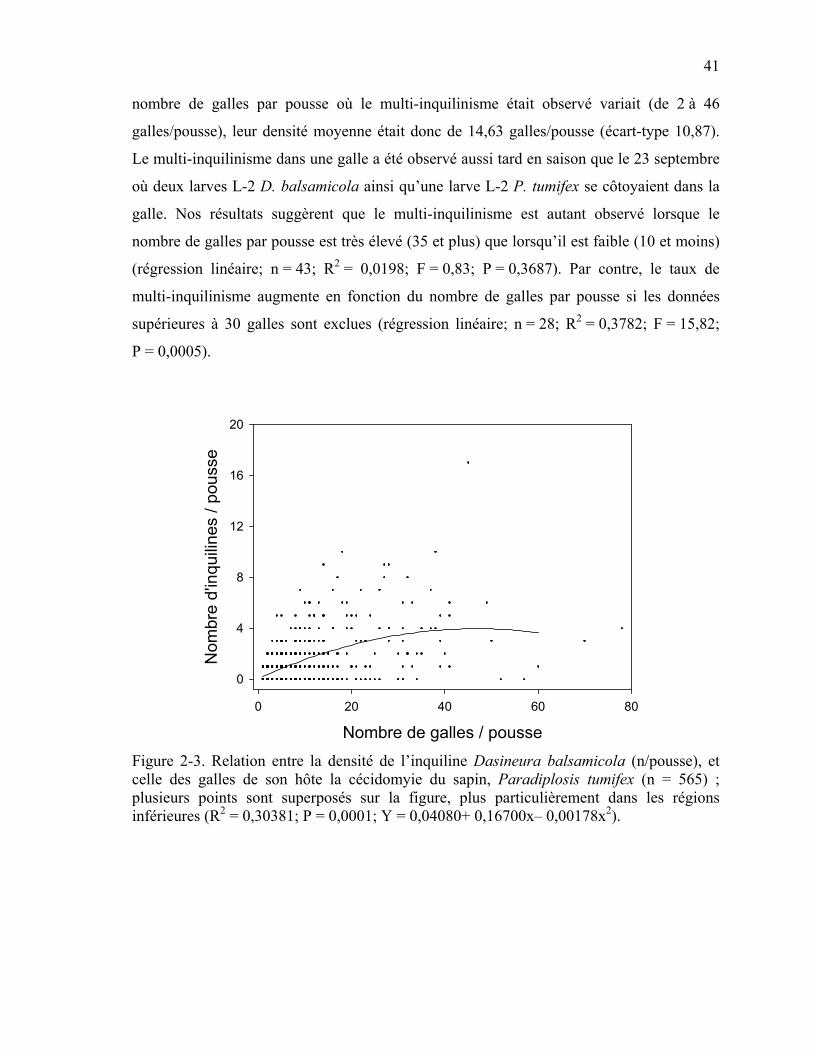
41
nombre de galles par pousse où le multi-inquilinisme était observé variait (de 2 à 46
galles/pousse), leur densité moyenne était donc de 14,63 galles/pousse (écart-type 10,87).
Le multi-inquilinisme dans une galle a été observé aussi tard en saison que le 23 septembre
où deux larves L-2 D. balsamicola ainsi qu’une larve L-2 P. tumifex se côtoyaient dans la
galle. Nos résultats suggèrent que le multi-inquilinisme est autant observé lorsque le
nombre de galles par pousse est très élevé (35 et plus) que lorsqu’il est faible (10 et moins)
(régression linéaire; n = 43; R2 = 0,0198; F = 0,83; P = 0,3687). Par contre, le taux de
multi-inquilinisme augmente en fonction du nombre de galles par pousse si les données
supérieures à 30 galles sont exclues (régression linéaire; n = 28; R2 = 0,3782; F = 15,82;
P = 0,0005).
Nombre de galles / pousse0 20 40 60 80
Nom
bre
d'in
quili
nes
/ pou
sse
0
4
8
12
16
20
Figure 2-3. Relation entre la densité de l’inquiline Dasineura balsamicola (n/pousse), et celle des galles de son hôte la cécidomyie du sapin, Paradiplosis tumifex (n = 565) ; plusieurs points sont superposés sur la figure, plus particulièrement dans les régions inférieures (R2 = 0,30381; P = 0,0001; Y = 0,04080+ 0,16700x– 0,00178x2).

42
2.4.3. Identification des parasitoïdes
Cinq espèces de parasitoïdes appartenant à trois familles d’hyménoptères ont été identifiées
à partir de l’élevage en laboratoire des galles (Tableau 2-2). Un seul adulte par galle a
émergé lors de cette expérience. Le premier adulte parasitoïde émergé de nos échantillons,
34 jours après sa sortie de dormance à basse température, était un spécimen de
Aprostocetus near marcovitchi Crawford. Ce sont les Pseudencyrtus sp. qui ont émergé le
plus rapidement avec un délai moyen de 55 jours, à 15° C. Les Platygaster albicollis
MacGown et Osgood ont pris 187 jours en moyenne pour émerger, Quadrastichus
whitmani Girault 131 et Q. near diarthronomyiae (Gahan) 171 jours. Quelques autres
spécimens (35) appartenant à la famille des Eulophidae ont émergé, mais ceux-ci n’ont pu
être identifiés soit parce que les spécimens étaient abîmés, soit parce que les caractères
taxonomiques permettant de les identifier n’étaient pas visibles. La dernière émergence
observée était un Q. near diarthronomyiae émergé après 315 jours d’incubation post-
dormance, ce qui est très élevé. Les parasitoïdes adultes ont pris en moyenne 148 jours pour
émerger à une température de 15° C, dans des conditions d’humidité relative de 80 % et à
une photopériode de 16L : 8N. Les Eulophidae étaient les plus abondants, plus
particulièrement A. near marcovitchi et Q. near diarthronomyiae avec respectivement 151
et 132 émergences.
Tableau 2-2. Identification des parasitoïdes hyménoptères associés à la cécidomyie du sapin Paradiplosis tumifex et obtenus à partir de l’élevage post-dormance en laboratoire.
Famille
Sous-famille
Espèce Type de
parasitisme
Nombre de spécimens obtenus
n = 4000 galles
Platygastridae Platygastrinae Platygaster albicollis Endoparasitisme 3
Encyrtidae Encyrtinae Pseudencyrtus sp. Endoparasitisme 4
Eulophidae Tetrastichinae Aprostocetus near marcovitchi Ectoparasitisme 151
Eulophidae Tetrastichinae Quadrastichus near diarthronomyiae
inconnu 132
Eulophidae Tetrastichinae Quadrastichus whitmani Ectoparasitisme 36
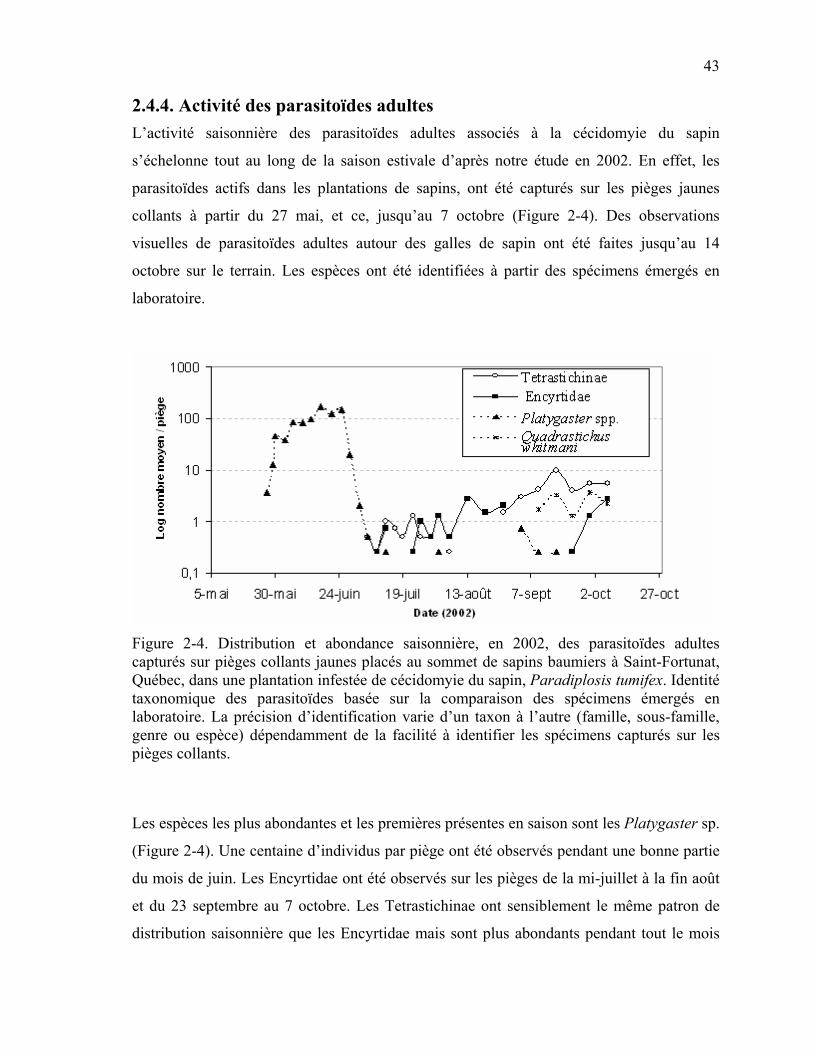
43
2.4.4. Activité des parasitoïdes adultes L’activité saisonnière des parasitoïdes adultes associés à la cécidomyie du sapin
s’échelonne tout au long de la saison estivale d’après notre étude en 2002. En effet, les
parasitoïdes actifs dans les plantations de sapins, ont été capturés sur les pièges jaunes
collants à partir du 27 mai, et ce, jusqu’au 7 octobre (Figure 2-4). Des observations
visuelles de parasitoïdes adultes autour des galles de sapin ont été faites jusqu’au 14
octobre sur le terrain. Les espèces ont été identifiées à partir des spécimens émergés en
laboratoire.
Figure 2-4. Distribution et abondance saisonnière, en 2002, des parasitoïdes adultes capturés sur pièges collants jaunes placés au sommet de sapins baumiers à Saint-Fortunat, Québec, dans une plantation infestée de cécidomyie du sapin, Paradiplosis tumifex. Identité taxonomique des parasitoïdes basée sur la comparaison des spécimens émergés en laboratoire. La précision d’identification varie d’un taxon à l’autre (famille, sous-famille, genre ou espèce) dépendamment de la facilité à identifier les spécimens capturés sur les pièges collants.
Les espèces les plus abondantes et les premières présentes en saison sont les Platygaster sp.
(Figure 2-4). Une centaine d’individus par piège ont été observés pendant une bonne partie
du mois de juin. Les Encyrtidae ont été observés sur les pièges de la mi-juillet à la fin août
et du 23 septembre au 7 octobre. Les Tetrastichinae ont sensiblement le même patron de
distribution saisonnière que les Encyrtidae mais sont plus abondants pendant tout le mois
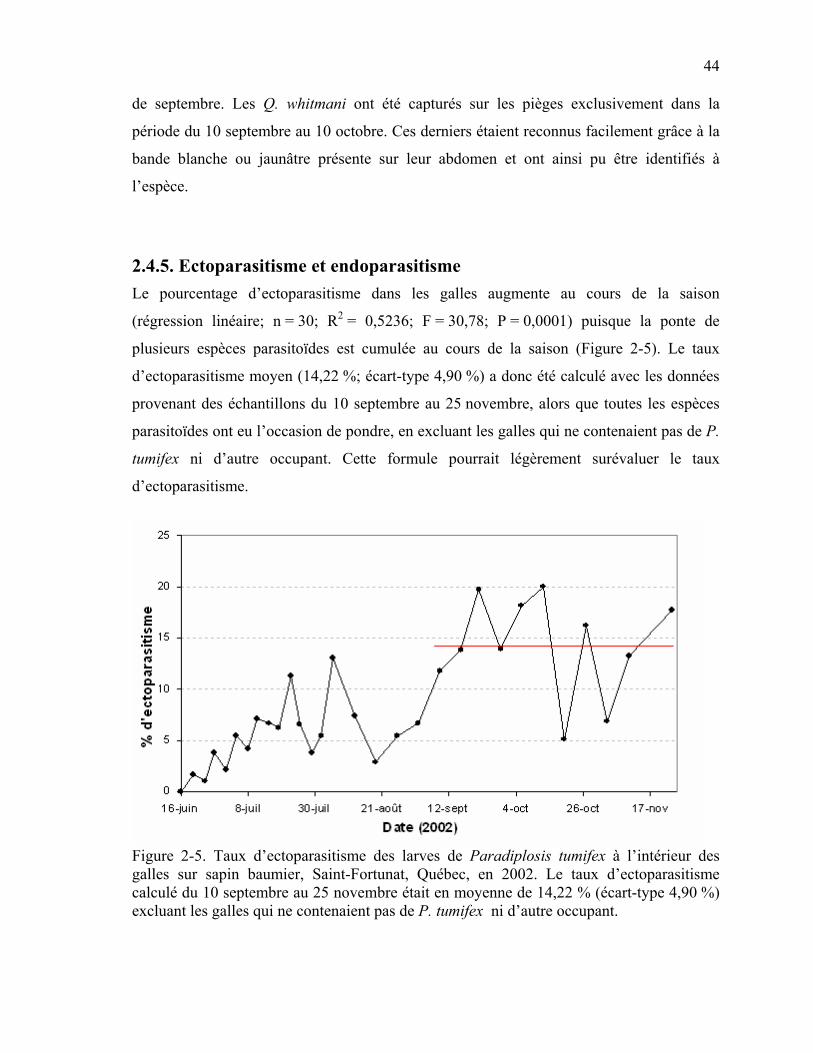
44
de septembre. Les Q. whitmani ont été capturés sur les pièges exclusivement dans la
période du 10 septembre au 10 octobre. Ces derniers étaient reconnus facilement grâce à la
bande blanche ou jaunâtre présente sur leur abdomen et ont ainsi pu être identifiés à
l’espèce.
2.4.5. Ectoparasitisme et endoparasitisme Le pourcentage d’ectoparasitisme dans les galles augmente au cours de la saison
(régression linéaire; n = 30; R2 = 0,5236; F = 30,78; P = 0,0001) puisque la ponte de
plusieurs espèces parasitoïdes est cumulée au cours de la saison (Figure 2-5). Le taux
d’ectoparasitisme moyen (14,22 %; écart-type 4,90 %) a donc été calculé avec les données
provenant des échantillons du 10 septembre au 25 novembre, alors que toutes les espèces
parasitoïdes ont eu l’occasion de pondre, en excluant les galles qui ne contenaient pas de P.
tumifex ni d’autre occupant. Cette formule pourrait légèrement surévaluer le taux
d’ectoparasitisme.
Figure 2-5. Taux d’ectoparasitisme des larves de Paradiplosis tumifex à l’intérieur des galles sur sapin baumier, Saint-Fortunat, Québec, en 2002. Le taux d’ectoparasitisme calculé du 10 septembre au 25 novembre était en moyenne de 14,22 % (écart-type 4,90 %) excluant les galles qui ne contenaient pas de P. tumifex ni d’autre occupant.
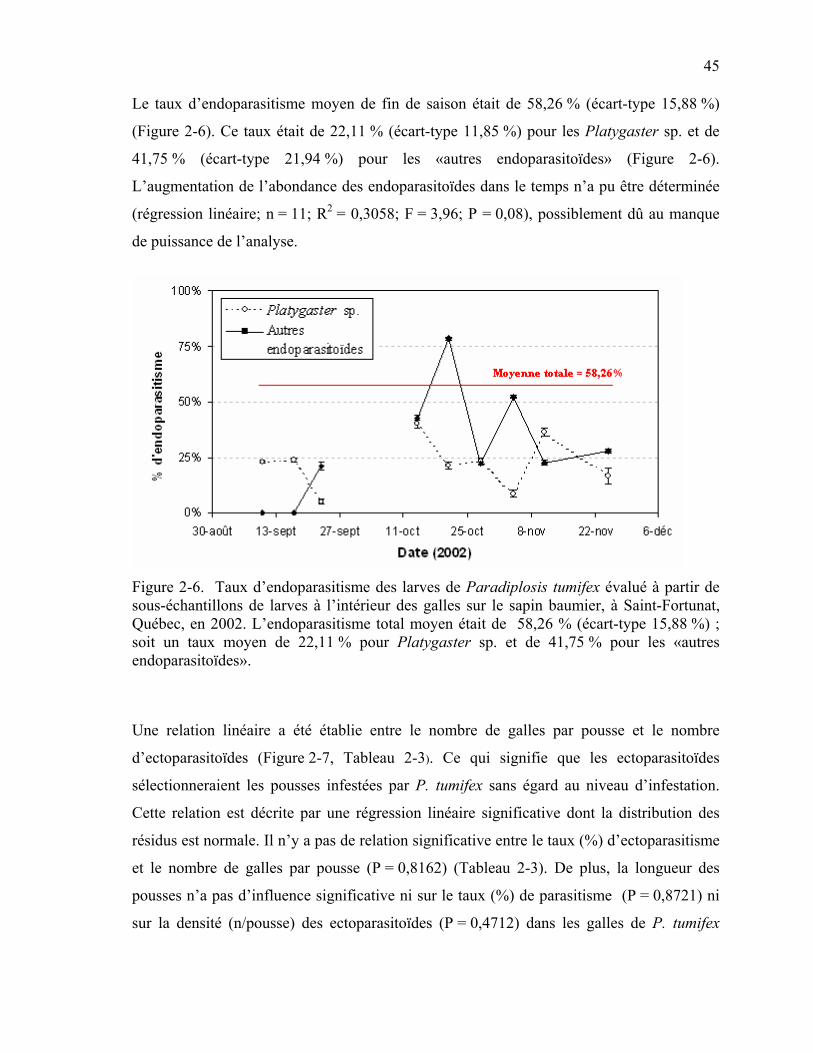
45
Le taux d’endoparasitisme moyen de fin de saison était de 58,26 % (écart-type 15,88 %)
(Figure 2-6). Ce taux était de 22,11 % (écart-type 11,85 %) pour les Platygaster sp. et de
41,75 % (écart-type 21,94 %) pour les «autres endoparasitoïdes» (Figure 2-6).
L’augmentation de l’abondance des endoparasitoïdes dans le temps n’a pu être déterminée
(régression linéaire; n = 11; R2 = 0,3058; F = 3,96; P = 0,08), possiblement dû au manque
de puissance de l’analyse.
Figure 2-6. Taux d’endoparasitisme des larves de Paradiplosis tumifex évalué à partir de sous-échantillons de larves à l’intérieur des galles sur le sapin baumier, à Saint-Fortunat, Québec, en 2002. L’endoparasitisme total moyen était de 58,26 % (écart-type 15,88 %) ; soit un taux moyen de 22,11 % pour Platygaster sp. et de 41,75 % pour les «autres endoparasitoïdes».
Une relation linéaire a été établie entre le nombre de galles par pousse et le nombre
d’ectoparasitoïdes (Figure 2-7, Tableau 2-3). Ce qui signifie que les ectoparasitoïdes
sélectionneraient les pousses infestées par P. tumifex sans égard au niveau d’infestation.
Cette relation est décrite par une régression linéaire significative dont la distribution des
résidus est normale. Il n’y a pas de relation significative entre le taux (%) d’ectoparasitisme
et le nombre de galles par pousse (P = 0,8162) (Tableau 2-3). De plus, la longueur des
pousses n’a pas d’influence significative ni sur le taux (%) de parasitisme (P = 0,8721) ni
sur la densité (n/pousse) des ectoparasitoïdes (P = 0,4712) dans les galles de P. tumifex
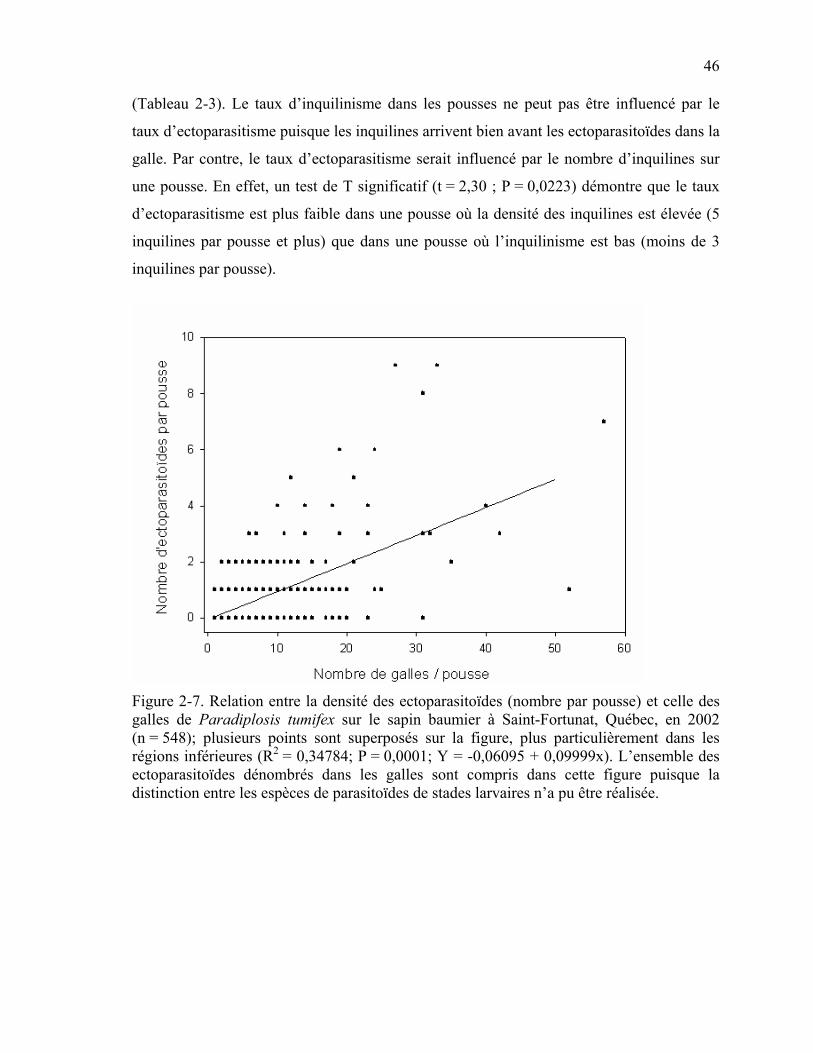
46
(Tableau 2-3). Le taux d’inquilinisme dans les pousses ne peut pas être influencé par le
taux d’ectoparasitisme puisque les inquilines arrivent bien avant les ectoparasitoïdes dans la
galle. Par contre, le taux d’ectoparasitisme serait influencé par le nombre d’inquilines sur
une pousse. En effet, un test de T significatif (t = 2,30 ; P = 0,0223) démontre que le taux
d’ectoparasitisme est plus faible dans une pousse où la densité des inquilines est élevée (5
inquilines par pousse et plus) que dans une pousse où l’inquilinisme est bas (moins de 3
inquilines par pousse).
Figure 2-7. Relation entre la densité des ectoparasitoïdes (nombre par pousse) et celle des galles de Paradiplosis tumifex sur le sapin baumier à Saint-Fortunat, Québec, en 2002 (n = 548); plusieurs points sont superposés sur la figure, plus particulièrement dans les régions inférieures (R2 = 0,34784; P = 0,0001; Y = -0,06095 + 0,09999x). L’ensemble des ectoparasitoïdes dénombrés dans les galles sont compris dans cette figure puisque la distinction entre les espèces de parasitoïdes de stades larvaires n’a pu être réalisée.
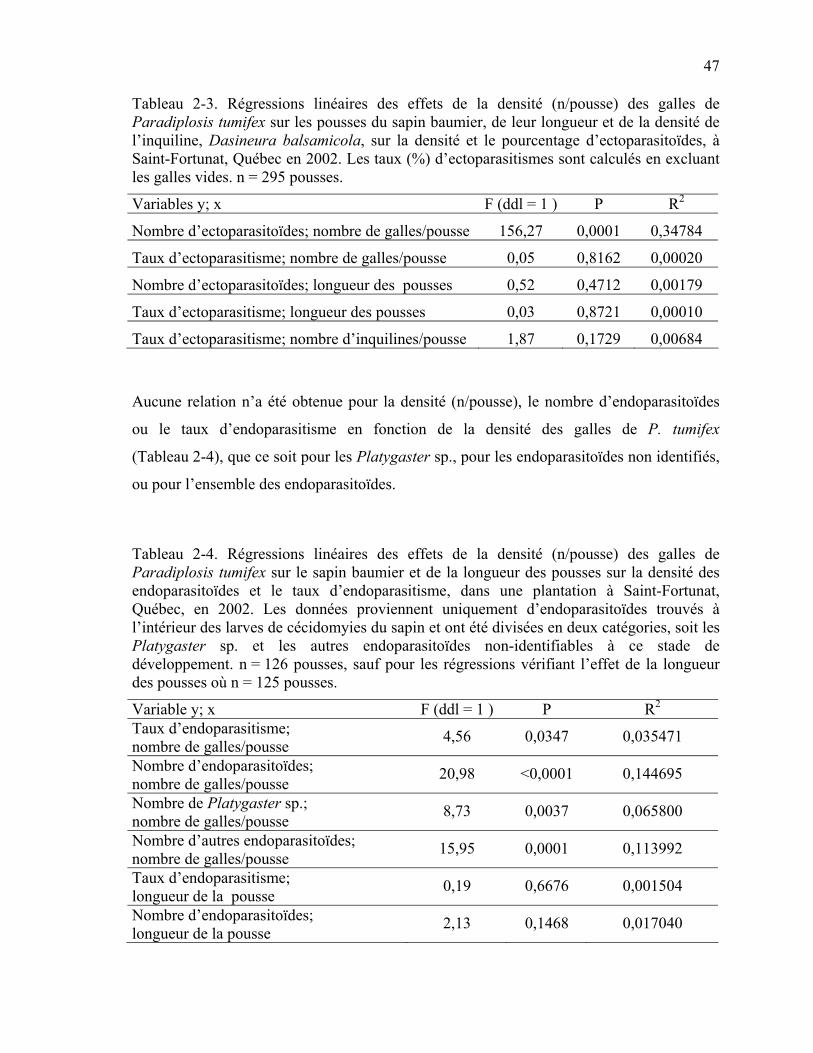
47
Tableau 2-3. Régressions linéaires des effets de la densité (n/pousse) des galles de Paradiplosis tumifex sur les pousses du sapin baumier, de leur longueur et de la densité de l’inquiline, Dasineura balsamicola, sur la densité et le pourcentage d’ectoparasitoïdes, à Saint-Fortunat, Québec en 2002. Les taux (%) d’ectoparasitismes sont calculés en excluant les galles vides. n = 295 pousses.
Variables y; x F (ddl = 1 ) P R2
Nombre d’ectoparasitoïdes; nombre de galles/pousse 156,27 0,0001 0,34784
Taux d’ectoparasitisme; nombre de galles/pousse 0,05 0,8162 0,00020
Nombre d’ectoparasitoïdes; longueur des pousses 0,52 0,4712 0,00179
Taux d’ectoparasitisme; longueur des pousses 0,03 0,8721 0,00010
Taux d’ectoparasitisme; nombre d’inquilines/pousse 1,87 0,1729 0,00684
Aucune relation n’a été obtenue pour la densité (n/pousse), le nombre d’endoparasitoïdes
ou le taux d’endoparasitisme en fonction de la densité des galles de P. tumifex
(Tableau 2-4), que ce soit pour les Platygaster sp., pour les endoparasitoïdes non identifiés,
ou pour l’ensemble des endoparasitoïdes.
Tableau 2-4. Régressions linéaires des effets de la densité (n/pousse) des galles de Paradiplosis tumifex sur le sapin baumier et de la longueur des pousses sur la densité des endoparasitoïdes et le taux d’endoparasitisme, dans une plantation à Saint-Fortunat, Québec, en 2002. Les données proviennent uniquement d’endoparasitoïdes trouvés à l’intérieur des larves de cécidomyies du sapin et ont été divisées en deux catégories, soit les Platygaster sp. et les autres endoparasitoïdes non-identifiables à ce stade de développement. n = 126 pousses, sauf pour les régressions vérifiant l’effet de la longueur des pousses où n = 125 pousses.
Variable y; x F (ddl = 1 ) P R2
Taux d’endoparasitisme; nombre de galles/pousse 4,56 0,0347 0,035471
Nombre d’endoparasitoïdes; nombre de galles/pousse 20,98 <0,0001 0,144695
Nombre de Platygaster sp.; nombre de galles/pousse 8,73 0,0037 0,065800
Nombre d’autres endoparasitoïdes; nombre de galles/pousse 15,95 0,0001 0,113992
Taux d’endoparasitisme; longueur de la pousse 0,19 0,6676 0,001504
Nombre d’endoparasitoïdes; longueur de la pousse 2,13 0,1468 0,017040

48
La grande diversité des parasitoïdes dans les galles de P. tumifex résulte en plusieurs cas de
multiparasitisme. Dans la parcelle A de la plantation de Saint-Fortunat, les résultats de
dissections de galles révèlent 13 galles (sur n = 7572) contenant deux ectoparasitoïdes ou
plus, des larves de P. tumifex contenant des Platygaster sp. en combinaison avec d’autres
espèces endoparasitoïdes (7,21 % des larves observées entre lame et lamelle; n = 323) et
des combinaisons d’endoparasitoïdes et d’ectoparasitoïdes ont été observées (9,37 % des
galles observées; n = 323). De plus, des cas de «superparasitisme» (21,05 % des galles
observées, n = 323, ce pourcentage inclut les cas de multiparasitisme) ont été observés, où
jusqu’à 13 larves de Platygaster sp. ont été retrouvées dans une seule larve de P. tumifex.
Des cas de parasitisme et d’inquilinisme combiné ont également été remarqués : 5,88 % des
galles observées (n = 323) où une inquiline côtoyait P. tumifex endoparasité et 12 galles
(n = 7572) contenant P. tumifex, D. balsamicola ainsi qu’un ectoparasitoïde.
2.5. Discussion La densité de populations de la cécidomyie du sapin oscille entre des niveaux épidémiques
de durée limitée et des niveaux à peine détectables (Martineau, 1985; Osgood et al., 1992).
Nos résultats suggèrent que lors de périodes épidémiques telles que celles survenues au
sud-ouest du Québec en 2001-2002, l’inquilinisme et le parasitisme contribuent grandement
à réduire les populations et sont influencés par la densité des cécidomyies galligènes dans
une pousse. La grande diversité de parasitoïdes associés à P. tumifex, de même que leur
distribution saisonnière, sont similaires à ce qui avait été répertorié lors d’études
précédentes, en particulier au Maine (Osgood et al., 1992). Des cas de multiparasitisme et
de combinaisons de parasitoïdes et d’inquilines ont été observés dans les galles, ce qui
suggère de nombreux types d’interactions compétitives intra- et interspécifiques dans la
galle menant éventuellement à l’élimination de certains co-occupants des galles, au
détriment d’autres.

49
2.5.1. Taux d’infestation de P. tumifex Les infestations des pousses par P. tumifex dans les plantations de sapins de Noël varient
d’une saison à l’autre (Martineau, 1985; Osgood et al., 1992). Les cycles épidémiques de
ce genre sont bien documentés pour les petits mammifères (associés avec leurs prédateurs)
et pour certains insectes herbivores, notamment chez les lépidoptères forestiers (Ginzburg
et Taneyhill, 1994). Chez les insectes, ces variations seraient principalement influencées
par les parasitoïdes, les prédateurs ou les pathogènes, bien que de nombreux facteurs, tels
que la température et la qualité de la nourriture ont un effet direct sur la régulation de ces
populations (Myers, 1990, Ginzburg et Taneyhill, 1994; Berryman, 1996).
En 2002, les taux d’infestation des pousses, par la cécidomyie du sapin, à Saint-Fortunat et
à Racine, étaient caractéristiques d’une période épidémique plus ou moins avancée (Osgood
et al., 1992 ; Jean et al., 2003). En 2001, l’infestation était déjà commencée à Saint-
Fortunat et sûrement à Racine, bien que les données n’étaient pas disponibles.
À Inverness, en 2002, le taux d’infestation était faible et il n’y a pas eu d’infestation
épidémique à ce site, probablement suite à l’usage d’insecticides pour le contrôle de la
cécidomyie. En effet, en 2001, le producteur avait observé un début d’infestation de
P. tumifex et avait traité avec succès (C. Cloutier, comm. pers., 2002).
En 2003, les échantillonnages de cécidomyies sur les pièges jaunes et de galles sur les
pousses pendant le mois de mai et jusqu’à la mi-juin, indiquaient une récession générale de
l’épidémie de P. tumifex qui durait depuis deux ou trois ans au Québec, bien que des
données précises à ce sujet ne soient pas disponibles. Les taux d’infestation par les galles
de P. tumifex étaient très faibles soit entre 0 % et 3,68 % (Saint-Fortunat) et les données du
RAP indiquaient que c’était ainsi dans tous les sites surveillés du sud du Québec.
En comparaison avec les données du Maine, les infestations de Racine et Saint-Fortunat en
2002 étaient très élevées. Alors qu’il y avait autour de 60 % d’infestation au Québec, seule
une des cinq plantations du Maine, et ce durant une seule épidémie (sur un suivi de 16 ans)

50
a atteint ces sommets. L’épidémie au Québec a également chuté drastiquement
comparativement aux chutes de populations plus graduelles observées dans les plantations
du Maine.
2.5.2. Inquilinisme
2.5.2.1. Taux d’inquilinisme Au Maine, le taux d’inquilinisme variait en fonction du site et de l’année où a lieu
l’infestation de P. tumifex; il pouvait être presque nul ou dépasser les 80 % (Osgood et al.,
1992). Dans la plantation de Saint-Fortunat en 2002, l’inquilinisme était relativement faible
(environ 16 %) alors que dans la plantation de Racine, l’inquilinisme a atteint 60 %. Au
Maine, lors d’infestations de P. tumifex comparables à celles observées au Québec en 2002,
l’inquilinisme était également de 60 % (Osgood et al., 1992).
2.5.2.2. Relation entre D. balsamicola et P. tumifex Les périodes d’émergence et d’activité des cécidomyies P. tumifex et D. balsamicola, bien
décrites dans les travaux de Osgood et Gagné (1978) et de Akar (1987) au Maine puis
corroborées par Jean et al. (2003) pour les plantations du sud du Québec, montrent un bon
synchronisme. Ainsi, les deux espèces de cécidomyies sont actives et pondent
simultanément durant le débourrement et la croissance initiale des bourgeons de sapin et ce,
avec un très léger décalage dans le temps, ce qui permet à D. balsamicola d’avoir plus
d’opportunités de rencontrer des œufs de P. tumifex. Il y a donc de nombreuses interactions
possibles entre les deux espèces et nous avons tenté d’en découvrir les principales.
Une relation curvilinéaire entre le nombre de galles par pousse et le nombre d’inquilines
D. balsamicola sur ces pousses a été établie (Figure 2-3). Cette relation montre que
l’abondance de l’inquiline sur une pousse est en partie dépendante de l’abondance des
galles de la cécidomyie galligène, une ressource essentielle pour sa protection et son
développement.

51
La curvilinéarité de cette régression, correspondant à la signification statistique des effets
quadratiques, suggère qu’à une certaine densité de P. tumifex sur une pousse, on observe un
plafonnement de l’inquilinisme, soit à environ quatre inquilines par pousse sur une densité
d’environ 40 galles par pousse (Figure 2-3). Deux hypothèses pourraient expliquer
l’atteinte d’un plateau : soit l’effectif des cécidomyies inquilines D. balsamicola, n’est pas
suffisant pour coloniser toute la pousse; soit les femelles de D. balsamicola ne privilégient
pas ces pousses trop infestées afin d’éviter la compétition.
Premièrement, il est possible que les femelles inquilines D. balsamicola ne soient pas assez
nombreuses ou pas assez fécondes pour répondre aux fortes densités de P. tumifex. Le taux
d’infestation des inquilines était d’ailleurs relativement faible à Saint-Fortunat (16 % des
pousses), en 2002. Il est possible que le nombre insuffisant d’inquilines qui colonisaient les
pousses, puisse résulter de leur mauvaise capacité de vol (Osgood et al., 1992; Briggs et
Latto, 2000). Seules les cécidomyies émergées à proximité de pousses fortement infestées,
ou plus rarement transportées par le vent, peuvent s’y rendre, alors que les cécidomyies des
autres secteurs privilégient des pousses de sapin infestées plus près d’elles (Briggs et Latto,
2000).
En second lieu, l’atteinte d’un plateau dans les taux d’inquilinisme pourrait être due à la
sélection limitée des pousses très infestées par les femelles inquilines. En effet, lorsqu’un
habitat n’est pas convenable parce que la compétition y est trop forte, il est avantageux pour
la femelle de choisir un meilleur site pour sa ponte (van Baaren et Nénon, 1996). Avec
l’augmentation de la densité des pontes de D. balsamicola sur une jeune pousse, il y a une
possibilité accrue que plusieurs inquilines se retrouvent dans la même galle. Nos résultats
suggèrent d’ailleurs que le multi-inquilinisme est observé plus fréquemment dans les
pousses contenant entre 15 et 30 galles. Lorsque le nombre d’inquilines sur une pousse est
élevé, il peut y avoir une augmentation de la compétition directe entre elles et
éventuellement, une seule larve survivra. Les femelles inquilines ont donc avantage à
pondre sur des pousses où la compétition intraspécifique est moins probable. Pour réduire
cette compétition, de nombreuses espèces herbivores et parasitoïdes laissent des traces

52
chimiques (phéromones de marquage) sur leur site de ponte (Roitberg et Prokopy, 1987).
Miller et Borden (1984) ont démontré l’existence d’une phéromone de marquage chez
Contarinia oregonensis Foote (Diptera : Cecidomyiidae), une cécidomyie causant des
dommages aux cônes du sapin Douglas (Pseudotsuga menziessi (Minb.), lui permettant
l’évitement de cônes occupés par les œufs de ses conspécifiques. Une phéromone du même
type pourrait exister chez D. balsamicola puisqu’elle pourrait faire une sélection des
pousses. Aucune mention de phéromone de marquage n’a été répertoriée se rapportant à
une inquiline.
Les courbes de densité des populations présentées par Osgood et al. (1992) sur une période
de 17 ans, suggèrent une relation de densité-dépendance entre la cécidomyie inquiline des
galles et la cécidomyie du sapin. Cette relation évoque la dynamique couplée d’un
prédateur, parasitoïde ou champignon et de sa proie ou son hôte préféré, fortement
influencées l’un par l’autre. Nos données sur la réponse de l’inquiline à la densité de
P. tumifex dans une pousse (Figure 2-3) confirment l’hypothèse selon laquelle l’inquiline
est un facteur de mortalité important pour la cécidomyie du sapin P. tumifex. Cependant,
des données sur plusieurs années seront nécessaires pour vérifier si la relation de densité-
dépendance proposée par Osgood et al. (1992) se vérifie au Québec.
2.5.3. Parasitisme
2.5.3.1. Identification des parasitoïdes L’élevage et la mise en dormance des galles de la cécidomyie du sapin en chambre de
croissance a révélé qu’un minimum de cinq espèces de parasitoïdes, provenant de trois
familles d’hyménoptères, sont associées à la cécidomyie du sapin P. tumifex au Québec.
Ces espèces sont en partie représentatives de la liste établie par Connor et Osgood (1979)
pour le Maine où ils faisaient la mention de Platygaster albicollis, Quadrastichus whitmani
et Aprostocetus near marcovitchi, espèces identifiées dans nos échantillons. Cependant,
trois espèces qui avaient été associées à P. tumifex par MacGown et Osgood (1972), ne se
retrouvent pas dans nos échantillons (Platygaster mainensis, Tetrastichus cecidivorus et
Pseudencyrtus borealis). De plus, l’espèce identifiée au Québec, comme étant très proche

53
taxonomiquement de Quadrastichus diarthronomyiae (synonyme Tetrastichus
diarthronomyiae selon LaSalle, 1993) n’avait jamais été mentionnée et les Pseudencyrtus
sp. ne correspondent pas aux espèces congénériques connues, identifiées par MacGown
(1979).
Notons d’ailleurs que Q. diarthronomyiae est connu comme parasitoïde de la cécidomyie
du chrysanthème, Diarthronomya chrysanthemi Ahlb. (synonymes Diarthronomyia
chrysanthemi et Rhopalomyia chrysanthemi, selon Sundheim et Rafoss, 1999)
(LaSalle, 1993). Ni nos élevages, ni la littérature ne permettent de savoir si
Q. diarthronomyiae est endoparasitoïde ou ectoparasitoïde. D’ailleurs, si Q. near
diarthronomyiae est bel et bien Q. diarthronomyiae, ce parasitoïde n’est pas spécifique et la
cécidomyie du sapin ne serait possiblement qu’un hôte alternatif. D’autres parasitoïdes de
P. tumifex pourraient avoir des hôtes alternatifs, soit : Aprostocetus marcovitchi et
Quadrastichus whitmani (MacGown et Osgood, 1972).
Les parasitoïdes spécialistes de larves d’insectes galligènes ou mineuses et appartenant à la
famille des Eulophidae, possèdent souvent des hôtes alternatifs (Hagvar, et al., 2000, Massa
et al.. 2001). Lorsque l’hôte principal d’une espèce de parasitoïde est absent ou en faible
abondance, ceux-ci se tournent vers ces hôtes alternatifs auxquels ils sont plus ou moins
bien adaptés (Godfray, 1994). Il est même possible, lorsque la variabilité génétique est
suffisante, d’adapter certains parasitoïdes à un hôte cible, par sélection active en
laboratoire, dans l’objectif de produire des agents de lutte biologique (Rathman et al., 1990,
Chow et Heinz, 2006).
La qualité des spécimens de Pseudencyrtus sp. obtenus suite à l’élevage en laboratoire, n’a
pas permis d’identifier l’espèce d’aucun des Encyrtidae émergés des galles de P. tumifex.
Toutefois, il ne s’agit pas de P. cecidomyiae Howard; et selon les caractéristiques des ailes,
ni de P. borealis, que MacGown (1979) avait associé à la cécidomyie du sapin (Gibson,
comm. pers., 2004). Alors, malgré les distributions saisonnières semblables des espèces
observées au Maine et au Québec, il est probable que Pseudencyrtus sp. observé dans cette
étude, soit une espèce nouvelle encore non-décrite. Notons qu’il n’existe présentement que

54
trois espèces décrites de Pseudencyrtus en Amérique du Nord (Gibson comm. pers., 2004).
Pour clarifier cette situation, il faudrait obtenir de nouveaux spécimens québécois et les
comparer taxonomiquement à P. borealis provenant de l’étude du Maine.
Il n’est pas exclu que d’autres espèces que celles identifiées dans cette étude soient
associées à P. tumifex au Québec, vu la durée limitée de notre étude. Par exemple, une seule
espèce de Platygaster a été identifiée suite à l’élevage, mais de quatre à cinq espèces étaient
observées sur les pièges collants jaunes donc, présentes sur le terrain. Il est possible qu’il en
soit de même pour d’autres espèces qui, plus fragiles, n’auraient pas survécu aux
procédures d’élevage et d’hivernement simulé. Ces élevages sont difficiles et entraînent un
fort pourcentage de mortalité comme apparemment ici avec seulement moins de 10 %
(394 sur 4000 galles) des galles élevées au laboratoire ayant produit un insecte adulte, soit
P. tumifex, D. balsamicola ou une guêpe parasitoïde.
L’élevage était en partie basé sur les essais de Akar (1987) qui avaient pour but d’étudier la
diapause des cécidomyies en laboratoire, sous différentes conditions de température et
d’humidité relative. Les conditions identifiées comme optimales par Akar (1987) ont été
très légèrement modifiées ici pour des questions de logistique; soit une acclimatation de
4° C au lieu de 3° C, suivie de huit semaines à 2° C plutôt que 0° C. Les principales
difficultés rencontrées étaient le contrôle et le maintien de l’humidité ainsi que la
contamination par des moisissures. Le contrôle de l’humidité dans les capsules Ependorf
était très difficile, notamment après 90 jours d’incubation. Selon Akar (1987), de plus
grands contenants auraient diminué les risques de dessiccation mais pour nos besoins,
l’espace requis aurait été démesuré, nuisant à l’observation des larves bien cachées. Une
autre recommandation d’Akar (1987) était d’utiliser un substrat stérile en remplacement de
la mousse de tourbe qu’il a utilisée. Malgré l’utilisation de coton dentaire imbibé de méthyl
parabenzène, il y avait toujours une certaine contamination. Akar (1987) mentionnait
l’inefficacité du méthyl parabenzène à 0,25% contrairement à Connor et Osgood (1979) qui
avaient constaté une diminution significative des moisissures avec cet agent de
conservation. La concentration utilisée ici (0,05%) était probablement insuffisante. Les
problèmes reliés à l’humidité et aux moisissures étaient amplifiés par la longue période

55
d’incubation, bien supérieure à celle de Akar (1987). En effet, il s’est écoulé en moyenne
148 jours (34-315) entre la fin de la dormance simulée et l’émergence des adultes, toutes
espèces confondues. L’émergence de P. tumifex s’est produite en moyenne en 35 jours, en
comparaison à 21 jours pour Akar (1987), qui récoltait peu de parasitoïdes, alors que la
grande majorité des émergences ici, était des parasitoïdes (326/394).
Pour référence ultérieure, quelques spécimens de chaque espèce récoltée dans cette étude
ont été déposés comme vouchers à la collection nationale du Canada, Ottawa. Malgré l’aide
précieuse des Dr Gibson, Hubert et Masner, la détermination des espèces n’est pas
complète et mériterait d’être approfondie. La taxonomie des micro-hyménoptères
parasitoïdes est encore peu avancée et a subi beaucoup de révisions au cours des dernières
années, notamment chez les Tetrastichus sp., Quadrastichus sp. et Aprostocetus sp.
(LaSalle, 1993). Les caractères taxonomiques de ces insectes étant difficiles à évaluer, les
identifications proposées pour les spécimens appartenant aux genres Quadrastichus et
Aprostocetus restent en partie incertaines (Q. near diarthronomyiae et A. near marcovitchi)
et doivent être renforcées.
Le problème de l’identification taxonomique spécifique des parasitoïdes n’a pas de
répercussion négative sur cette étude puisque les données recueillies sur leur abondance et
leur impact global sur P. tumifex restent valides et le dépôt de vouchers pourra permettre
éventuellement d’apporter des précisions taxonomiques. D’ailleurs dans un futur rapproché,
il serait possible de procéder à leur caractérisation par analyse et séquençage d’ADN (par
exemple mitochondrial), ce qui permettrait de les identifier plus rapidement comme larves
ou adultes (Sampson et al., 2006). Pour cela, les amorces PCR (réaction de polymérase en
chaîne) spécifiques à chaque espèce de parasitoïde devront être disponibles.
2.5.3.2. Distribution saisonnière des parasitoïdes La distribution saisonnière des parasitoïdes observée au Maine, est similaire à celle
observée lors de cette étude. La période d’activité de certaines espèces de parasitoïdes
associés aux galles de sapin baumier débute, au Maine, dans la seconde moitié du mois de

56
mai (MacGown et Osgood, 1972; Osgood et al., 1992). Au sud-ouest du Québec, en 2002,
les premiers parasitoïdes étaient également observés à la mi-mai, seulement quelques jours
après les premières émergences de la cécidomyie du sapin P. tumifex (Figure 2-4; Jean et
al., 2003). Les dernières observations saisonnières de parasitoïdes, lors de cette étude en
2002, ont été faites à Saint-Fortunat le 14 octobre, alors qu’au Maine, MacGown et Osgood
(1972) mentionnent la présence de Tetrastichus sp. jusqu’à la fin octobre.
Nos données sur l’activité des insectes en vol (collecte au piège jaune sur la cime des
arbres) montrent qu’environ une dizaine d’espèces de parasitoïdes sont actives dans le sapin
baumier et ce, durant toute la saison estivale. Leurs périodes d’activité se chevauchent et
donc le développement des uns à l’intérieur de la galle peut empiéter sur celui des autres.
Les Platygaster sp. sont les premiers parasitoïdes de la saison à être actifs au sud-ouest du
Québec, selon nos observations (Figure 2-4). Les premières émergences des Platygaster sp.
ont été observées le 20 mai au Maine (1969 et 1985) et le 27 mai au Québec (2002) mais ils
étaient capturés plus longtemps lors de cette étude, soit jusqu'au début juillet plutôt que
jusqu’à la mi-juin, au Maine. À Saint-Fortunat, en 2002, l’abondance des Platygaster sp.
était beaucoup plus élevée qu’au Maine (MacGown et Osgood,1972). Il est toutefois
impossible de faire des comparaisons précises de l’abondance des parasitoïdes entre les
données recueillies de cette étude et celles du Maine dû à l’absence de quantification de
l’abondance des parasitoïdes au Maine.
Comme notre étude coïncide avec une culmination de l’infestation de P. tumifex, en 2002
(Jean et al., 2003), il est probable que les populations de Platygaster sp. s’étaient
progressivement établies dans les années précédentes, ce qui expliquerait leur grande
abondance. Les Platygaster sp. sont probablement efficaces pour repérer la cécidomyie du
sapin rapidement, car en général, les endoparasitoïdes seraient mieux adaptés à trouver leur
hôte que les ectoparasitoïdes (Godfray, 1994). Par contre, les fortes populations de
Platygaster sp. pourraient aussi être dues à une surévaluation du nombre de Platygaster sp.
associé à la cécidomyie du sapin. En effet, les identifications ont été limitées au niveau de
la famille sur les pièges collants, ou de façon plus précise lorsque la position et la qualité du

57
spécimen le permettait. Donc, des individus de certaines espèces non associées à la
cécidomyie du sapin, ont pu contribuer à gonfler légèrement le nombre de Platygaster sp.
Pour ce qui est de A. near marcovitchi, il apparaît beaucoup plus hâtivement au Maine
(mi-juin) qu’au Québec (début juillet). Cette espèce était apparemment beaucoup plus
abondante au Maine, que dans les plantations du sud-ouest du Québec.
Au Québec, Q. whitmani a été piégé exclusivement en fin de saison (septembre et octobre),
contrairement au Maine, où leur présence saisonnière s’étend du début juillet jusque vers la
fin de septembre (MacGown et Osgood,1972) et où des spécimens ont été observés pondant
leurs oeufs en septembre (Osgood et Dimond, 1970). Malgré tout, l’abondance de
Q. whitmani y était très faible, et probablement moindre qu’au Québec où de deux à quatre
individus étaient piégés par semaine, durant cinq semaines.
Lors de sa première mention en 1916, Q. whitmani avait été identifié comme parasitoïde
des œufs de Physonota unipunctata Say (Coleoptera : Chrysomelidae), ce qui serait
étonnant. Sauf erreur taxonomique possible, il est probable que la cécidomyie du sapin
P. tumifex, ne soit qu’un hôte alternatif de Q. whitmani. Ainsi, l’abondance de Q. whitmani,
pourrait être relative à celle de son hôte primaire.
L’identification des Encyrtidae collectés sur les plaquettes jaunes collantes n’a pas pu être
précisée jusqu’au genre étant donné leur état inadéquat. Des encyrtides ont été observés dès
le 9 juillet, étant particulièrement abondants de la fin juillet à la fin août et à quelques
reprises du 23 septembre au 7 octobre. Cette distribution saisonnière est sensiblement la
même que celle rapportée par MacGown et Osgood (1972), au Maine, où elle se prolonge
jusqu’à la mi-octobre, bien que les populations les plus abondantes étaient observées à la
mi-juillet.
Selon nos observations, les Tetrastichinae, soit la combinaison A. near marcovitchi,
Q. diarthronomyiae et d’autres espèces non identifiées, ont essentiellement le même patron

58
de distribution saisonnière qu’au Maine (MacGown et Osgood,1972), mais paraissent plus
abondants pendant tout le mois de septembre.
La distribution saisonnière des parasitoïdes de P. tumifex au sud-ouest Québec, en 2002, est
donc très semblable à ce qui a été déterminé au Maine, lors de deux études précédentes
mais l’abondance des Platygaster paraît plus élevée au Québec. Cette étude amène des
précisions sur la diversité et l’abondance relative des parasitoïdes de P. tumifex, au cours de
la saison.
2.5.3.3. Ectoparasitisme Parmi les cinq espèces de parasitoïdes associées à P. tumifex, recensées au cours de cette
étude, au moins deux sont des ectoparasitoïdes, soit Quadrastichus whitmani Girault
(Hymenoptera : Eulophidae) et Aprostocetus near marcovitchi Crawford (Hymenoptera :
Eulophidae). Selon Osgood et Dimond (1970), ces Eulophidae auraient, au Maine, des
hôtes alternatifs en plus de la cécidomyie du sapin. Dans ce cas, l’effectif de leur
population ne serait pas uniquement dépendant des populations de P. tumifex.
Puisque plusieurs espèces de parasitoïdes se succèdent au cours de la saison (Figure 2-4),
leurs pontes ainsi que le nombre de larves d’ectoparasitoïdes dans les galles est croissant de
mai à octobre (Figure 2-5). C’est d’ailleurs pour cette raison que les estimés de
l’ectoparasitisme et la relation entre la densité (nombre par pousse) des ectoparasitoïdes en
fonction de celle des galles de P. tumifex, ont été effectués à partir des données de fin de
saison, au moment où toutes les espèces de parasitoïdes ont eu l’occasion de pondre à
l’intérieur des galles. Comme les ectoparasitoïdes ne sont pas différenciables à l’état
larvaire, ces relations de densité étaient vérifiées pour l’ensemble des ectoparasitoïdes,
alors qu’il y a probablement des différences interspécifiques. Nos résultats livrent donc une
tendance globale de la réponse de l’ensemble des ectoparasitoïdes en fonction du nombre
de galles présentes sur une pousse.

59
Le pourcentage des galles de sapin parasitées par un ectoparasitoïde à Saint-Fortunat, en fin
de saison 2002, est d’environ 10 %. Ce taux semble peu élevé mais combiné aux autres
facteurs de réduction de la population des cécidomyies du sapin, il pourrait s’avérer
important.
Une relation linéaire a été démontrée entre le nombre de galles par pousse et le nombre
d’ectoparasitoïdes (Figure 2-7), ce qui signifie que plus la densité des galles de P. tumifex
sur les pousses est élevée, plus celle des ectoparasitoïdes est élevée. L’ensemble des
ectoparasitoïdes semblent donc avoir une réponse fonctionnelle directe (réponse de type I)
face à la densité de leur hôte P. tumifex.
Il est possible que les ectoparasitoïdes puissent même se développer (ou du moins survivre
temporairement) sur la larve de cécidomyie inquiline. Nous avons observé quatre cas où
une larve de la cécidomyie inquiline des galles, D. balsamicola, sur laquelle était accroché
un ectoparasitoïde (Figure 2-8). Ces observations ont été faites surtout en fin de saison alors
que la population de cécidomyie du sapin P. tumifex, diminuait. Nos données ne permettent
pas de confirmer que les ectoparasitoïdes observés sur D. balsamicola, complètent leur
développement et produisent des pupes pouvant entrer en dormance et émerger au
printemps. Il s’agit d’une première mention de parasitoïdes associés à D. balsamicola.
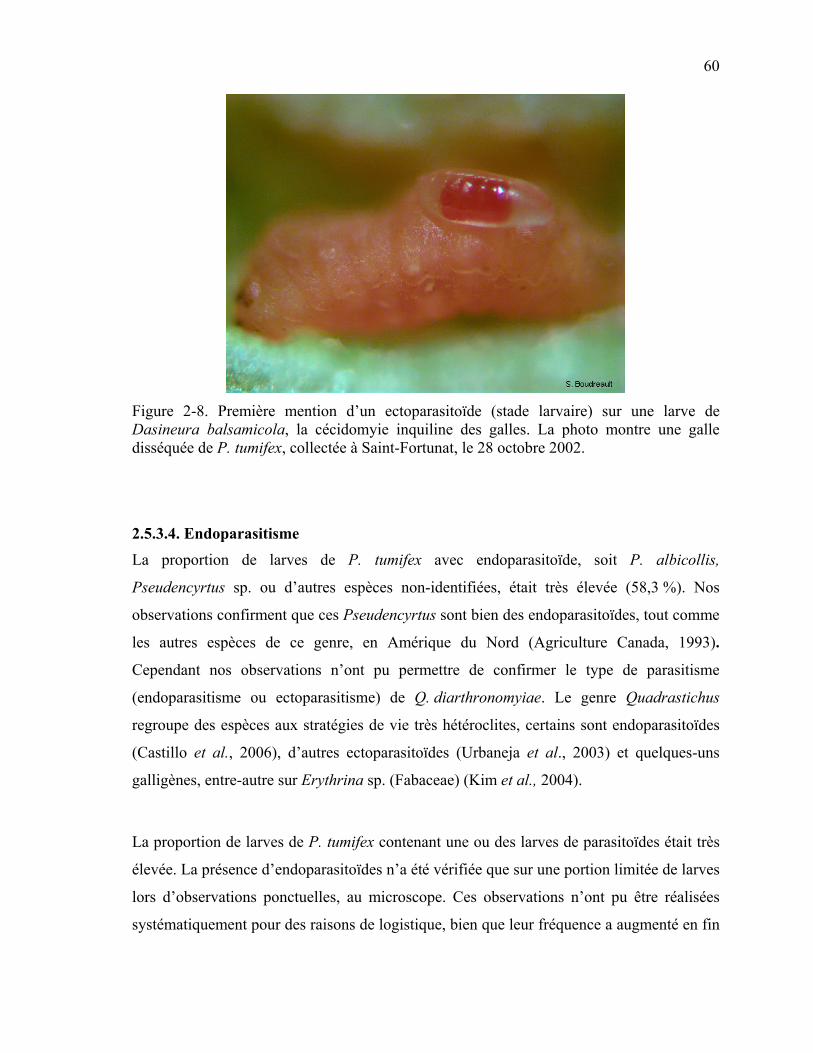
60
Figure 2-8. Première mention d’un ectoparasitoïde (stade larvaire) sur une larve de Dasineura balsamicola, la cécidomyie inquiline des galles. La photo montre une galle disséquée de P. tumifex, collectée à Saint-Fortunat, le 28 octobre 2002.
2.5.3.4. Endoparasitisme La proportion de larves de P. tumifex avec endoparasitoïde, soit P. albicollis,
Pseudencyrtus sp. ou d’autres espèces non-identifiées, était très élevée (58,3 %). Nos
observations confirment que ces Pseudencyrtus sont bien des endoparasitoïdes, tout comme
les autres espèces de ce genre, en Amérique du Nord (Agriculture Canada, 1993).
Cependant nos observations n’ont pu permettre de confirmer le type de parasitisme
(endoparasitisme ou ectoparasitisme) de Q. diarthronomyiae. Le genre Quadrastichus
regroupe des espèces aux stratégies de vie très hétéroclites, certains sont endoparasitoïdes
(Castillo et al., 2006), d’autres ectoparasitoïdes (Urbaneja et al., 2003) et quelques-uns
galligènes, entre-autre sur Erythrina sp. (Fabaceae) (Kim et al., 2004).
La proportion de larves de P. tumifex contenant une ou des larves de parasitoïdes était très
élevée. La présence d’endoparasitoïdes n’a été vérifiée que sur une portion limitée de larves
lors d’observations ponctuelles, au microscope. Ces observations n’ont pu être réalisées
systématiquement pour des raisons de logistique, bien que leur fréquence a augmenté en fin

61
de saison. Des vérifications de l’endoparasitisme sur l’ensemble de la saison, auraient
donné un aperçu plus représentatif de l’abondance et des relations entre endoparasites et les
autres ennemis naturels de la cécidomyie du sapin. Par exemple, les adultes de Platygaster
sp. ont été recensés en grand nombre (soit autour de 100 individus par piège) au printemps
et le taux d’endoparasitisme par ce taxon dans les échantillons observés au microscope
dépasse les 20 % en fin de saison. Des examens au microscope de l’endoparasitisme des
larves de P. tumifex, peu de temps après les émergences et envols de Platygaster sp. au
printemps, auraient donné plus de précision sur l’endoparasitisme primaire par ce groupe,
car en fin de saison, il a pu être sous évalué dû à l’inquilinisme et aux autres
endoparasitoïdes (le multiparasitisme sera abordé plus en profondeur dans la prochaine
section). Aucun endoparasitoïde n’a été observé à l’intérieur des larves de la cécidomyie
inquiline des galles D. balsamicola.
Sur la base de notre échantillonnage, aucune relation de dépendance n’a été détectée entre
les endoparasitoïdes et le taux d’infestation des pousses par P. tumifex (Tableau 2-4).
2.5.3.5. Multiparasitisme Le multiparasitisme ainsi que les interactions entre parasitoïdes et inquilines résulte en
importante compétition pour la ressource (la galligène pour le parasitoïde et la galle pour
l’inquiline). Puisque parmi les galles mises en dormance, un seul spécimen émergeait par
galle de P. tumifex (n = 394 galles ayant produit une émergence), et qu’il n’y a pas
d’évidence dans la littérature d’émergences multiples, tant pour les Platygaster sp. (Connor
et Osgood, 1979; Sampson et al., 2006) que pour Pseudencyrtus sp. ou les ectoparasitoïdes
associés à P. tumifex (Connor et Osgood, 1979), on peut supposer jusqu’à preuve du
contraire, qu’un seul individu survit à la compétition. Il serait donc favorable pour les
ennemis naturels de P. tumifex de discerner et d’éviter les galles déjà occupées par un autre
ennemi naturel, notamment par l’inquiline. D’ailleurs, nos résultats démontrent que le
nombre d’inquilines sur une pousse a une influence sur le taux d’ectoparasitoïdes, toutes
espèces confondues (Tableau 2-3). Plus de précision sur la réponse des différentes espèces
aurait été obtenue s’il avait été possible de distinguer les différentes espèces et si nous

62
avions pu discerner s’ils évitent de pondre dans la galle où se trouve l’inquiline. Car, dans
une galle où l’inquiline est présente, la survie des ectoparasitoïdes semble menacée. En
effet, Shorthouse et West (1986) émettent l’hypothèse que l’inquiline tue la cécidomyie
galligène par le frottement de ses spinules sur la cuticule plus fragile de son hôte car
l’espace disponible dans la galle, à la fin de la croissance des galles, est restreint et les
manœuvres d’évitement deviennent impossibles. Or, si les spinules peuvent tuer la
cécidomyie galligène, il pourrait en être de même pour les ectoparasitoïdes, à moins qu’ils
n’adoptent un comportement d’évitement. Donc, si les cas observés d’ectoparasites sur la
cécidomyie galligène indiquent que certaines espèces peuvent apparemment se déplacer sur
l’inquiline, d’autres seront possiblement incapables de le faire et seront éliminés avec leur
hôte, la galligène.
Dans le cas de multiparasitisme impliquant des endoparasitoïdes et des ectoparasitoïdes,
plusieurs observations relatées dans la littérature montrent que les ectoparasitoïdes sont en
général avantagés par rapport aux endoparasitoïdes (Yu, et al. 1990, Borer, 2002), même si
dans d’autres cas, la survie des ectoparasitoïdes est plus faible en présence
d’endoparasitoïdes (Borer, 2002). Étant donné leur position physique hors de l’hôte et leur
grande compétitivité, ils pourraient même utiliser les endoparasitoïdes comme ressource
alimentaire additionnelle (hyperparasitisme) (Godfray, 1994). Les endoparasitoïdes,
confinés à l’intérieur de l’hôte, peuvent être éliminés par des réactions de défense
immunitaire de l’hôte ou, dans le cas de multiparasitisme ou de superparasitisme, par
l’attaque physique inter- ou intraspécifique entre larves ou par la suppression physiologique
(diminution d’oxygène disponible, sécrétions de toxines) (vanBaaren et Nénon, 1996;
Caltagirone, 1999).
Dans ce système, les Platygaster sp. étaient particulièrement abondants à pondre au
printemps mais peu nombreux (Tableau 2-2) suite à l’élevage des galles en laboratoire, ce
qui suggère un fort taux de mortalité hivernale et post-dormance. Étant donnée la diversité
des espèces d’endoparasitoïdes et surtout celle des ectoparasitoïdes, la mortalité de
Platygaster sp. pourrait donc être expliquée par la compétition intra-guilde, spécialement

63
en fin de saison. En effet, les larves de premier stade de Platygaster sp. restent inactives en
diapause de juin à avril l’année suivante, soit près de un an (MacGown et Osgood, 1972).
Même si ces larves ont de grandes mandibules qui devraient les aider à se défendre, leur
taille réduite et leur apparente inactivité minimisent leurs chances de survie lorsqu’elles
sont en compétition directe avec d’autres espèces dans la larve de P. tumifex (Briggs et
Latto, 2000). Les Platygaster sp. étant très spécifiques aux cécidomyies (Osgood et
Dimond, 1970; Sampson et al., 2006), ils compenseraient leur faible compétitivité par une
plus grande efficacité à trouver leurs hôtes et à y pondre leurs œufs (De Moraes et al.,
1999).
Plusieurs cas de «superparasitisme» par Platygaster sp. ont été observés (MacGown et
Osgood, 1972) et jusqu'à 13 larves de l’endoparasitoïde ont été observées dans une larve de
P. tumifex, lors de cette étude. Selon Caltagirone (1999), l’élimination d’un surnombre de
larves endoparasitoïdes est fréquente, habituellement par le combat entre larves à l’intérieur
de l’hôte, d’où la sélection de caractères tels que des mandibules acérées, des parties
sclérifiées, ou par une plus grande mobilité. La morphologie des larves P. albicollis est
représentative d’une adaptation au combat, bien qu’on leur attribue une longue période
d’inactivité (immobilité). Cependant, les données disponibles ne permettent pas de vérifier
si plusieurs Platygaster sp., provenant d’une même larve de P. tumifex, peuvent survivre et
se développer jusqu’à maturité.
Étant donné la grande diversité des parasitoïdes dans ce système, on peut supposer que
certains pourraient être hyperparasitoïdes facultatifs ou obligatoires. D’ailleurs, MacGown
et Osgood (1972) ont noté quelques cas d’hyperparasitisme (espèces non identifiées). Dans
cette étude, certaines mentions de multiparasitisme pourraient en fait être de
l’hyperparasitisme. Malheureusement, ces parasitoïdes n’ont pas encore été suffisamment
étudiés et beaucoup d’informations sur leurs biologies restent à découvrir.
Finalement, le nombre fréquent ainsi que les diverses associations de multiparasitisme
résultent en interférence. L’impact de chaque parasitoïde dans la régulation des populations

64
de P. tumifex n’est donc pas additif car certains sont éliminés aux dépens d’autres plus
compétitifs. Par exemple, dans cette étude, les Platygaster sp. semblent fréquemment
éliminés lorsqu’en compétition avec d’autres ennemis naturels de P. tumifex, alors que
D. balsamicola semble généralement y survivre.
2.5.4. Impacts des ennemis naturels sur P. tumifex Selon notre hypothèse de recherche, le nombre élevé de parasitoïdes, leur grande diversité
ainsi que l’importance de l’inquilinisme, sont importants dans la diminution des
populations de P. tumifex dans les plantations de sapins de Noël. Selon Briggs et Latto
(2000), en absence de parasitoïdes et d’autres ennemis naturels, les cécidomyies pourraient
atteindre des densités extrêmement élevées pouvant éventuellement tuer la plante.
Toutefois, ces densités sont rarement observées en nature puisque les parasitoïdes et autres
ennemis naturels y sont toujours présents.
Pour avoir un impact sur les populations de galligènes, leurs ennemis naturels devraient
être hautement spécialisés (Hufbauer, 2004). Les larves étant protégées par la galle, les
parasitoïdes doivent avoir la capacité de pénétrer les tissus végétaux pour y pondre leurs
œufs (Hufbauer, 2004). Si la galle est trop grosse, les petits parasitoïdes ayant de petits
ovipositeurs, pourraient ne pas être en mesure d’en atteindre l’intérieur (Stilling et Rossi,
1996). Pourtant, les galligènes sont hautement parasitées (Hawkins, 1993), comme c’est le
cas pour P. tumifex (Figures 2-5 et 2-6; MacGown et Osgood, 1972). Selon Hawkins
(1993), la diversité des parasitoïdes associés à un insecte serait influencée par le type de
refuge qu’il utilise. Or, selon cet auteur, le nombre moyen d’espèces de parasitoïdes
associés aux galligènes (3,8 espèces par hôte) serait comparable aux herbivores mobiles
(3,6), contrairement aux mineuses, porte-cases et enrouleurs de feuilles dont la diversité des
parasitoïdes associés à chaque espèce (respectivement 6,3 ; 6,2 et 5,5) est encore plus
élevée (Hawkins, 1993).
Dans le cas de P. tumifex, la diversité de ses parasitoïdes (minimum de cinq espèces, au
Québec et six au Maine) et les taux de parasitisme global élevés, tels qu’observé en 2002,

65
contribueraient à la réduction de ses populations. Selon nos données, ce serait surtout le fort
taux d’endoparasitisme (58 % ; Figure 2.6) qui aurait contribué le plus aux chutes
drastiques de populations de P. tumifex observées à Saint-Fortunat, en 2003. De plus,
l’inquiline pourrait également avoir un grand rôle à jouer dans la régulation des
populations. Osgood et al. (1992) avaient d’ailleurs suggéré que l’inquiline régule les
populations de P. tumifex, en s’appuyant sur la relation de densité dépendance observée
dans le temps sur plusieurs cycles successifs d’abondance au Maine. À Racine, la cause de
la chute de population observée en 2002-2003, est probablement plus reliée au haut taux
d’inquilinisme de D. balsamicola (60 %) qu’il ne l’est à Saint-Fortunat (taux d’inquilinisme
de 15 %). Il n’est pas possible d’évaluer le rôle de l’inquilinisme à Inverness car la
population de P. tumifex était déjà très basse lors de mon étude en 2002 (probablement dû
aux traitements chimiques).
Selon nos données, c’est la combinaison des parasitoïdes avec l’inquiline, malgré leur
interférence, qui en exerçant une forte pression sur P. tumifex aurait causé la chute de
population observée en 2003. Même si certaines espèces peuvent nuire au développement
des autres, l’interférence observée entre elles n’est pas suffisante pour empêcher la chute de
population de P. tumifex. L’addition des taux observés de parasitisme
(endoparasitisme = 58,26 %; ectoparasitisme = 14,22 %) et d’inquilinisme (16,43 %) puis
la soustraction des cas de multiparasitisme (total = 22,79 %) suggère que plus de 66 % des
larves de P. tumifex ont été éliminées par ces facteurs de mortalité, au cours de la saison
2002, à la plantation de Saint-Fortunat. Tous les ennemis naturels de P. tumifex pris dans
leur ensemble, pourraient également être à l’origine du maintien des populations de
P. tumifex à un faible niveau, entre les périodes épidémiques. Mais si cette hypothèse est
validée, on devra vérifier la cause des reprises des fortes densités et les épidémies de
P. tumifex, indiquant que les ennemis naturels ne seraient pas suffisants dans certaines
conditions.

66
2.6. Bibliographie Agriculture Canada. 1993. Hymenoptera of the world : an identification guide to families.
Édité par H. Goulet et J.T. Huber. 668 pages. Akar, H.Jr. 1987. Some aspects of the biology and rearing of two species of midges,
Paradiplosis tumifex and Dasineura balsamicola (Diptera : Cecidomyiidae) found in galled needles of balsam fir, Abies balsamea. Thesis. University of Maine. 42 pages.
van Baaren, J. et J-P. Nénon. 1996. Intraspecific larval competition in two solitary
parasitoids, Apoanagyrus (Epidinocarsis) lopezi and Leptomastix dactylopii. Entomologia Experimentalis et Applicata 81 : 325-333.
Berryman, A.A. 1996. What causes population cycles of forest Lepidoptera? Trends in
Ecology and Evolution 11 : 28-32. Borer, E.T. 2002. Intraguild predation in larval parasitoids : implications for coexistence.
Journal of Animal Ecology 71 : 957-965. Briggs, C.J. et J. Latto. 2000. The effect of dispersal on the population dynamics of a
gall-forming midge and its parasitoids. Journal of Animal Ecology 69 : 96-105. Caltagirone, L.E. 1999. Adaptations of insects to modes of life. Dans Ecological
entomology, édité par C.B. Huffaker et A.P. Gutierrez, publié par John Wiley and sons, États-Unis, pages 201-230.
Castillo, J., J.A. Jacas, J.E. Pena, B.J. Ulmer et D.G. Hall. 2006. Effect of temperature on
life history of Quadrastichus haitiensis (Hymenoptera : Eulophidae), an endoparasitoid of Diaprepes abbreviatus (Coleoptera : Curculionidae). Biological Control 36 : 189-196.
Chow, A. et K.M. Heinz. 2006. Control of Liriomyza langei on chrysanthemum by
Diglyphus isaea produced with a standard or modified parasitoid rearing technique. Journal of Applied Entomology 130 : 113-121.
Cloutier, C., P. Mailhot et J. Brodeur. 2006. La cécidomyie du sapin a-t-elle trop d’ennemis
naturels? Le Naturaliste Canadien 130 : 32-36. Connor, J.Y. et E.A Osgood. 1979. Host determination of Chalcidoid and Proctotrupoid
Hymenoptera associated with two species of midges (Diptera : Cecidomyiidae) found in needle galls of balsam fir. The Canadian Entomologist 111 : 945-947.

67
De Moraes, C.M., A.M. Cortesero, J.O. Stapel et W.J. Lewis. 1999. Intrinsic and extrinsic competitive interactions between two larval parasitoids of Heliothis virescens. Ecological Entomology 24 : 402-410.
Giese, R.L. et D.M. Benjamin. 1959. The biology and ecology of the balsam gall midge
in Wisconsin. Forest Science 5 : 193-208. Ginzburg, L.R. et D.E. Taneyhill, 1994. Population cycles of forest Lepidoptera : a
maternal effect hypothesis. Journal of Animal Ecology 63 : 79-92. Godfray, H.C.J. 1994. Parasitoids, Behavioral and Evolutionary Ecology. Princeton
University Press, New Jersey. 473 pages. Hagvar, E.B., N. Trandem et T. Hofsvang, 2000. Phenology and sex ratio of the
parasitoids of a cereal leaf miner Chromatomyia fuscula (Diptera : Agromyzidae). European Journal of Entomology 97 : 339-346.
Hawkins, B.A. 1993. Notes and comments: Parasitoid species richness, host mortality, and
biological control. The American Naturalist 141 : 634-641. Hufbauer, R.A. 2004. Observations of sagebrush gall morphology and emergence of
Rhopalomyia pomum (Diptera : Cecidomyiidae) and its parasitoids. Western North American Naturalist 64 : 324-330.
Jean, C., P. Mailhot, J. Brodeur et C. Cloutier. 2003. Outils de lutte intégrée contre la
cécidomyie du sapin dans les plantations d’arbres de Noël. Rapport de recherche CDAQ. 18 pages.
Kim, I.K., G. Delavare et J. LaSalle. 2004. A new species of Quadrastichus
(Hymenoptera : Eulophidae): A gall-inducing pest on Erythrina (Fabaceae). Journal of Hymenoptera Research 13 : 243-249.
LaSalle, J. 1993. North american genera of Tetrastichinae (Hymenoptera : Eulophidae).
Journal of Natural History 28 : 193-194. MacGown, M.W. 1979. Two new species of Chalcidoid wasp (Hymenoptera) parasitic on
the balsam gall midge (Diptera : Cecidomyiidae) in Maine. The Canadian Entomologist 111 : 939-943.
MacGown, M.W. et E.A Osgood. 1972. Taxonomy and biology of Chalcidoid and
Proctotrupoid Hymenoptera associated with Dasineura balsamicola (Diptera : Cecidomyiidae) in Maine. The Canadian Entomologist 104: 1259-1269.
Martineau, R. 1985. Insectes nuisibles des forêts de l’est du Canada. Éditions Marcel
Broquet Inc. Ottawa. pages 108-110.

68
Massa, B., M.C. Rizzo et V. Caleca. 2001. Natural alternative hosts of Eulophidae (Hymenoptera : Chalcidoidea) parasitoids of the citrus leafminer Phyllocnistis citrella Stainton (Lepidoptera : Gracillariidae) in the mediterranean basin. Journal of Hymenoptera Research 10 : 91–100.
Miller, G.E. et J.H. Borden, 1984, Reproductive behaviour of the Douglas fir cone gall
midge, Contarinia oregonensis (Diptera : Cecidomyiidae). The Canadian Entomologist 116 : 607-618.
Myers, J. 1990. Population cycles of western tent caterpillar : experimental introduction
and synchrony of fluctuations. Ecology 71 : 986-995. Osgood, E.A. et J.B. Dimond. 1970. Parasites of the balsam gall midge, Dasineura
balsamicola (Diptera : Cecidomyiidae). The Canadian Entomologist 102 : 182-184. Osgood, E.A. et R.J. Gagné. 1978. Biology and taxonomy of two gall midges (Diptera :
Cecidomyiidae) found in galls on balsam fir needles with description of a new species of Paradiplosis. Annals of the Entomological Society of America 71 : 85-91.
Osgood, E.A., R.L. Bradbury et F.A. Drummond. 1992. The balsam gall midge - an
economic pest of balsam fir Christmas trees. Maine Agricultural Experiment Station Technical Bulletin 151: 1-30.
Rathman, R.J., M.W. Johnson, J.A. Roseheim et B.E. Tabashnik. 1990. Carbamate and
pyrethroid resistance in the leafminer parasitoid Diglyphus begini (Hymenoptera : Eulophidae). Journal of Economic Entomology 83 : 2153–2158.
Roitberg, B.D. et R.J. Prokopy. 1987. Insects that marks host plants, an ecological,
evolutionary perspective on host-marking chemicals. Bioscience 37: 400-406. Sampson, B.J., T.A. Rinehart, O.E. Liburd, S.J. Stringer et J.M. Spiers. 2006. Biology of
parasitoids (Hymenoptera) attacking Dasineura oxycoccana and Prodiplosis vaccinii (Diptera : Cecidomyiidae) in cultivated blueberries. Annals of the Entomological Society of America 99: 113-120.
Shorthouse, J.D. et R.J. West. 1986. Role of the inquiline, Dasineura balsamicola
Diptera : Cecidomyiidae) in the balsam fir needle gall. Proceedings of the Entomological Society of Ontario 117 : 1-7.
Stilling, P. et A.M. Rossi. 1996. Complex effects of genotype and environment on insect
herbivores and their enemies. Ecology 77 : 2212-2218. Sundheim, L. et T. Rafoss. 1999. The danger to Norwegian plant health from increased
import of agricultural commodities. The Royal Norwegian Ministry of Agriculture. 46 pages.

69
Torre-Bueno, J.R. 1989. The Torre-Bueno glossary of entomology. Compilé par Stephen W. Nichols, incluant un supplément de George S. Tulloch. New York Entomological Society, New York, USA. 840 pages.
Urbaneja, A., E. Llacer, A. Garrido et J.A. Jacas. 2003. Interspecific competition between
two ectoparasitoids of Phyllocnistis citrella (Lepidoptera:Gracillariidae) : Cirrospilus brevis and the exotic Quadrastichus sp. (Hymenoptera : Eulophidae). Biological Control 28 : 243-250.
West, R.J. et J.D. Shorthouse. 1982. Morphology of the balsam fir needle gall induced by
the midge Paradiplosis tumifex (Diptera : Cecidomyiidae). Canadian Journal of Botany 60 : 131-140.
Yu, D.S., R.F. Luck et W.W. Murdoch. 1990. Competition resource partitioning and
coexistence of an endoparasitoid Encarsia perniciosi and an ectoparasitoid Aphytis melinus of the California red scale. Ecological Entomology 15 : 469-480.

Chapitre 3. Conclusions générales

71
3.1. Conclusion Cette étude nous a permis d’approfondir les connaissances relatives à la cécidomyie du
sapin, de son inquiline, de ses parasitoïdes et des interactions fréquentes entre-eux, dans
une situation de déclin des populations du ravageur, telle qu’observée au Québec
récemment.
À faible incidence, la galle occasionnée par Paradiplosis tumifex Gagné ne cause pas de
grave dommage au sapin baumier Abies balsamea Mill. L’arbre doit fournir de l’énergie
pour la formation de la galle et l’alimentation de son occupant (Bagatto et al., 1996; Stone
et Schönrogge, 2003) mais il compense ce manque en augmentant sa photosynthèse
(Bagatto et al., 1996). Le problème qu’engendre P. tumifex est plutôt d’ordre esthétique
puisque l’automne suivant, les aiguilles sur lesquelles sont formées des galles, tombent
(West et Shorthouse, 1982; Osgood et al., 1992). Les arbres fortement infestés, prêts à être
vendus sur le marché des sapins de Noël seront déclassés. Par contre, pour les jeunes
arbres, de nouvelles aiguilles poussent sur les nouvelles pousses l’année suivante ainsi que
les subséquentes, contribuant à cacher de plus en plus les rameaux dénudés d’aiguilles et
ce, jusqu’à la prochaine épidémie (Martineau, 1985).
Selon divers auteurs qui ont observé ce système (Martineau, 1985; Osgood et al., 1992), les
périodes de fortes infestations de la cécidomyie du sapin sont intermittentes et relativement
brèves, ce qui ne simplifie pas l’étude et le contrôle de ce ravageur. Pour les producteurs de
sapins, l’instabilité des populations de ce moucheron est, d’une part, avantageuse puisqu’il
ne constitue pas un problème persistant dont ils doivent s’occuper année après année. Par
contre, lorsqu’il réapparaît, suite à une longue absence, les producteurs moins expérimentés
pourraient être pris au dépourvu face à cet insecte qui, une fois à l’abri dans sa galle, est
peu exposé aux insecticides conventionnels et donc difficile à éliminer (RAP, 2002).
Nos observations ont permis de constater que les parasitoïdes et les inquilines sont
d’importants facteurs de mortalité en période épidémique et exercent une grande pression
sur les populations de P. tumifex. En effet, à la fin de la saison 2002, trois galles de

72
cécidomyies du sapin disséquées sur quatre étaient colonisées par des organismes autres
que P. tumifex. Qu’il s’agisse des inquilines qui occupent la galle et éliminent leur hôte, des
nombreux parasitoïdes (endoparasitoïdes, ectoparasitoïdes), du multiparasitisme observé ou
de l’association inquiline/parasitoïde, tous ces facteurs contribuent à la diminution
drastique de la population des galligènes. Nos résultats démontrent de l’interférence entre
les ennemis naturels (élimination de certains occupants aux dépens d’autres) mais celle-ci
n’empêche pas la régulation des populations de P. tumifex. De plus, il y a probablement
d’autres facteurs de mortalité moins évidents qui l’affectent, et que nous n’avons pas
quantifiés dans cette étude, tels que des pathogènes (virus, bactéries, champignons ou
nématodes) (Bergdahl et Mazzola, 1985), des oiseaux ou des mammifères associés à
P. tumifex (Osgood et al. 1992).
Cinq espèces de parasitoïdes ont été associées à la cécidomyie du sapin lors de cette étude,
alors que leur diversité au Québec était encore inexplorée. Mais des espèces additionnelles
de parasitoïdes sont fort probablement présentes, dont possiblement une espèce de
Platygaster sp. (Connor et Osgood, 1979). Nous avons également constaté qu’un
ectoparasitoïde (non-identifié) était parfois associé à la larve de la cécidomyie inquiline des
galles, Dasineura balsamicola (Lintner). Encore bien des questions restent sans réponse à
ce sujet, telles que la capacité de l’ectoparasite à se développer normalement jusqu’à
maturité sur l’inquiline, ainsi que l’hôte initial de l’ectoparasitoïde : était-il établi sur la
galligène et a ensuite effectué un transfert d’hôte, ou s’il s’est développé dès le début sur
l’inquiline?
Lors des expériences d’élevage de galles, la majorité (82 %) des adultes émergés étaient
des parasitoïdes. À l’inverse, lors d’expériences semblables effectuées par Akar (1987),
seulement un parasitoïde adulte a émergé alors qu’il obtenait 162 cécidomyies adultes. Ses
résultats suggèrent que la période du cycle épidémique correspondant à ses expériences
était probablement le début d’un cycle d’infestation. Si tel est le cas, nous pourrions
émettre l’hypothèse que : l’arrivée hâtive de parasitoïdes dans les plantations de sapins
pourrait réduire la période d’infestation de la cécidomyie du sapin. D’autres études et
expériences à ce sujet et des tests sur le terrain devraient être effectués pour le confirmer.

73
Dans un contexte de lutte biologique, soit « l'utilisation d'organismes vivants pour prévenir
ou réduire les dégâts causés par des ravageurs » (OILB/SROP, 1973), les parasitoïdes
présentent un bon potentiel pour agir à titre d’agent contre la cécidomyie du sapin : ils ont
une bonne capacité de recherche et ont un effet direct sur les populations. Par contre, les
défis restent de taille. Par exemple, leur efficacité pour la protection de la valeur esthétique
de l’arbre serait à vérifier. Ensuite, de nombreuses questions scientifiques devraient être
résolues pour permettre la lutte biologique, par exemple : quelle (ou quelles) espèce(s) de
parasitoïde choisir? Pourrait-on en faire l’élevage? Serait-il opérant lors d’inoculations
préventives? Ou augmentatives? Pour répondre à ces questions, la taxonomie, l’écologie et
la biologie de ces hyménoptères devront être approfondie. Des techniques d’identification
par analyse moléculaire et séquençage d’ADN mitochondrial des adultes pourraient
permettre de les identifier plus rapidement comme larves ou adultes (Sampson et al.,
2006). Des amorces PCR (réaction de polymérase en chaîne) spécifiques à chaque espèce
de parasitoïdes devront être disponibles. Des connaissances supplémentaires devront être
acquises et d’autres avenues d’exploration seraient possibles pour le remplacement des
insecticides chimiques conventionnels. L’une des solutions envisageables serait l’utilisation
de champignons entomopathogènes, tels que Uredinopsis mirabilis Magnus (Uredinales :
Pucciniastraceae) (Bergdahl et Mazzola, 1985) (Fournier et Brodeur, 2000).
Finalement, le micro-écosystème de la galle de P. tumifex est complexe et beaucoup
d’informations restent à découvrir autant pour les deux espèces de cécidomyies associées
aux galles que pour leurs parasitoïdes. Par exemple, lors de l’identification des œufs de
cécidomyies, nos observations ont révélé des différences importantes de coloration variant
de blanc translucide (tel que décrit par Giese et Benjamin, 1959) à orangé (tel que décrit
par Osgood et al., 1992). Nous n’étions pas en mesure de déterminer si ces variations
étaient dues à la maturité des œufs ou à l’espèce de cécidomyie. Du côté des parasitoïdes,
nous n’avons pas obtenu l’identité du parasitoïde qui pourrait être associé à l’inquiline. Ces
deux questions et plusieurs autres, pourraient probablement être résolues par la
combinaison d’identification moléculaire, d’élevages et d’observations contrôlées qui
devraient être entreprises.

74
3.2. Bibliographie Akar, H.Jr. 1987. Some aspects of the biology and rearing of two species of midges,
Paradiplosis tumifex and Dasineura balsamicola, (Diptera : Cecidomyiidae) found in galled needles of balsam fir, Abies balsamea. Thesis. University of Maine. 42 pages.
Bagatto, G., L.C. Paquette et J.D. Shorthouse. 1996. Influence of galls of Phanacis taraxi on carbon partitioning within common dandelion Taraxacum officinale. Entomologia Experimentalis et Applicata 79 : 111-117.
Bergdahl, D.R. et M. Mazzola. 1985. Some relationships between Uredinopsis mirabilis, Paradiplosis tumifex and Dasineura balsamicola in the needle of Abies balsamea. Canadian Journal of Forest Research 15 : 551-554.
Connor, J.Y. et E.A Osgood. 1979. Host determination of Chalcidoid and Proctotrupoid Hymenoptera associated with two species of midges (Diptera : Cecidomyiidae) found in needle galls of balsam fir. The Canadian Entomologist 111 : 945-947.
Fournier, V. et J. Brodeur. 2000. Dose-Response Susceptibility of Pest Aphids (Homoptera : Aphididae) and their Control on Hydroponically Grown Lettuce with the Entomopathogenic Fungus Verticillium lecanii, Azadirachtin, and Insecticidal Soap. Environmental Entomology 29 : 568-578.
Giese, R.L. et D.M. Benjamin. 1959. The biology and ecology of the balsam gall midge in Wisconsin. Forest Science 5 : 193-208.
Martineau, R. 1985. Insectes nuisibles des forêts de l’est du Canada. Éditions Marcel Broquet Inc. Ottawa. pages 108-110.
OILB/SROP. 1973. Statuts. Bulletin OILB-SROP. Publié par l’Organisation Internationale de Lutte Biologique et Intégrée contre les Animaux et les Plantes Nuisibles, section Regionale Ouest Paléarctique (OILB/SROP) 1973/1. 25 pages.
Osgood, E.A., R.L. Bradbury et F.A. Drummond. 1992. The balsam gall midge - an economic pest of balsam fir Christmas trees. Maine Agricultural Experiment Station Technical Bulletin 151 : 1-30.
RAP. 2002. Réseau d’information phytosanitaire: Bulletin d’information. Cécidomyie du sapin. www.agr.gouv.qc.ca/dgpar/rap ; consulté le 20 mars 2006.
Sampson, B.J., T.A. Rinehart, O.E. Liburd, S.J. Stringer et J.M. Spiers. 2006. Biology of parasitoids (Hymenoptera) attacking Dasineura oxycoccana and Prodiplosis vaccinii (Diptera : Cecidomyiidae) in cultivated blueberries. Annals of the Entomological Society of America 99: 113-120.

75
Stone, G.N. et K. Schönrogge. 2003. The adaptive significance of insect gall morphology. Trends in Ecology and Evolution 18 : 512-522.
West, R.J. et J.D. Shorthouse. 1982. Morphology of the balsam fir needle gall induced by the midge Paradiplosis tumifex (Diptera : Cecidomyiidae). Canadian Journal of Botany 60 : 131-140.


















