Vésicules extracellulaires : biomarqueurs et véhicules de ... · BDI: Beck depression inventory...
151
Vésicules extracellulaires : Biomarqueurs et véhicules de propagation de protéinopathies Mémoire Jérôme Lamontagne-Proulx Maîtrise en Neurobiologie Maître ès sciences (M.Sc.) Québec, Canada © Jérôme Lamontagne-Proulx, 2018
Transcript of Vésicules extracellulaires : biomarqueurs et véhicules de ... · BDI: Beck depression inventory...

Vésicules extracellulaires : Biomarqueurs et véhicules de propagation de protéinopathies
Mémoire
Jérôme Lamontagne-Proulx
Maîtrise en Neurobiologie Maître ès sciences (M.Sc.)
Québec, Canada
© Jérôme Lamontagne-Proulx, 2018

Vésicules extracellulaires : Biomarqueurs et véhicules de propagation de protéinopathies
Mémoire
Jérôme Lamontagne-Proulx
Sous la direction de :
Francesca Cicchetti, directrice de recherche

iii
Résumé
La maladie de Parkinson (MP) est une maladie neurodégénérative invalidante pour laquelle
le diagnostic ne peut être donné qu’une fois la dégénérescence neuronale bien entamée,
rendant impérative la découverte d’un biomarqueur ; un outil biologique permettant de
prédire l’apparition de la pathologie ou d’évaluer sa progression. Mon projet de maîtrise
visait donc l’étude des vésicules extracellulaires (VE) issues du sang comme test
diagnostique ou comme marqueur de la progression de la MP. La quantification des VE
effectuée par cytométrie de flux à haute sensibilité a révélé une augmentation spécifique des
VE dérivées d’érythrocytes (VEE) chez les parkinsoniens comparés à leurs contrôles, ainsi
qu’une forte corrélation avec la progression de la maladie. L’analyse quantitative de l’alpha-
synucléine (α-Syn), principale protéine impliquée dans la pathologie de la maladie, a montré
un niveau similaire entre les individus. Cependant, l’analyse protéomique des VEE a révélé
une modulation de certaines protéines entre les patients et les donneurs sains. Nos résultats
suggèrent que les VEE pourraient conduire au développement d’un marqueur pour suivre
l’évolution de la maladie ainsi que l’effet de nouvelles thérapies.

iv
Abstract
Parkinson’s disease (PD) is a debilitating neurodegenerative disease for which the diagnosis
can only be confirmed once the degeneration state is very advanced, making imperative the
discovery of a biomarker: a biological tool to predict the onset of pathology or its progression.
My master’s project was designed to study extracellular vesicles (EV) from the blood in order
to discover if they could be used as a diagnostic test or as a marker of disease progression.
Quantification of EV performed by high-sensitivity flow cytometry demonstrated an increase
in PD patients compared to their controls and a strong correlation with the progression of the
disease only in EV derived from erythrocytes (EEV). Quantitative analysis of α-Syn, the
main protein involved into PD pathogenesis, showed a similar level between individuals.
However, analysis of the EEV proteome reveals a modulation of some proteins between
patients and healthy donors. Our results suggest that EEV have the potential to lead to the
development of a marker abled to track disease course as well as measuring the effect of new
therapies.

v
Table des matières
Résumé .................................................................................................................................. iii
Abstract ................................................................................................................................ iv
Table des matières ................................................................................................................ v
Liste des tableaux .............................................................................................................. viii
Liste des figures ................................................................................................................... ix
Abréviations .......................................................................................................................... x
Remerciements ................................................................................................................... xiv
Avant-propos ....................................................................................................................... xv
CHAPITRE 1: INTRODUCTION ...................................................................................... 1
1.1 La maladie de Parkinson ............................................................................................... 2
1.1.1 Épidémiologie ........................................................................................................... 2
1.1.2 Étiologie .................................................................................................................... 3
1.1.2.1 Facteurs environnementaux................................................................................. 3
1.1.2.2 Facteurs génétiques ............................................................................................. 5
1.1.3 Neuropathologie ....................................................................................................... 7
1.1.3.1 Dégénérescence du système dopaminergique ..................................................... 7
1.1.3.2 L’alpha-synucléine et les corps de Lewy ............................................................ 8
1.1.3.3 Stress oxydatif et facteurs de mort cellulaire .................................................... 10
1.1.4 Diagnostic et test clinique ...................................................................................... 11
1.1.5 Biomarqueur .......................................................................................................... 16
1.1.5.1 Introduction ....................................................................................................... 16
1.1.5.2 Biomarqueurs génétiques .................................................................................. 18
1.1.5.3 Biomarqueurs par imagerie ............................................................................... 18
1.1.5.4 Biomarqueurs cliniques ..................................................................................... 19
1.1.5.5 Biomarqueurs biochimiques .............................................................................. 19
1.1.5.6 L’avenir des biomarqueurs ................................................................................ 22
1.2 Biomarqueur sanguin ................................................................................................... 22
1.2.1 Érythrocytes ........................................................................................................... 23
1.2.2 Plaquettes ................................................................................................................ 24
1.2.3 Leucocytes ............................................................................................................... 24

vi
1.3 Vésicules extracellulaires ............................................................................................. 25
1.3.1 Exosomes ................................................................................................................. 25
1.3.2 Microvésicules ........................................................................................................ 26
1.3.3 Corps apoptotiques ................................................................................................ 27
1.3.4 Rôles des VE dans la maladie de Parkinson ............................................................ 29
1.3.4.1 Études des VE dans le système nerveux central (in vitro) ................................ 30
1.3.4.2 Études des VE dans le liquide céphalo-rachidien ............................................. 30
1.3.4.3 Études des VE dans l’urine ............................................................................... 31
1.3.4.4 Études des VE dans le système circulatoire ...................................................... 31
1.4 Les objectifs de recherche ............................................................................................ 32
CHAPITRE 2: ERYTHROCYTE-DERIVED EXTRACELLULAR VESICLES: A NOVEL, ROBUST AND SPECIFIC BIOMARKER THAT MAPS TO PARKINSON'S DISEASE STAGES ............................................................................................................ 34
2.1 Résumé ........................................................................................................................... 35
2.2 Abstract ......................................................................................................................... 37
2.3 Introduction .................................................................................................................. 38
2.4 Materials and methods ................................................................................................. 39
2.4.1 Ethics statement and participant recruitment ........................................................... 39
2.4.2 Preparation of platelet-free plasma and EV labeling ............................................... 39
2.4.3 Flow cytometry quantification ................................................................................. 40
2.4.4 Production and purification of EEV ........................................................................ 40
2.4.5 C-reactive protein, free hemoglobin and α-synuclein quantification ...................... 41
2.4.6 Scanning electron microscopy ................................................................................. 41
2.4.7 Transmission electron microscopy .......................................................................... 41
2.4.8 Mass spectrometry analysis and label free protein quantification ........................... 41
2.4.9 Statistical analyses ................................................................................................. 42
2.5 Results ............................................................................................................................ 42
2.6 Discussion ...................................................................................................................... 45
2.7 Acknowledgements ....................................................................................................... 47
2.8 Tableau .......................................................................................................................... 49
2.9 Figures ........................................................................................................................... 51

vii
2.10 Matériels supplémentaires ......................................................................................... 58
2.10.1 Materials and methods ........................................................................................ 58
2.10.2 Tableaux ............................................................................................................... 62
2.10.3 Figures ................................................................................................................... 65
CHAPITRE 3: DISCUSSION ........................................................................................... 72
3.1 Caractérisation de la nature et du profil des VE ....................................................... 73
3.1.1 Biomarqueur de valeur diagnostique ....................................................................... 73
3.1.2 Biomarqueur pour la progression de la maladie ...................................................... 73
3.1.3 Validation des résultats ......................................................................................... 74
3.2 Contenu protéique des VEE ........................................................................................ 75
3.2.1 Méthodes d’activation ........................................................................................... 75
3.2.2 Quantification d’α-Syn par ELISA et microscopie ................................................. 76
3.2.3 Protéomique ........................................................................................................... 76
3.3 Perspectives d’avenir .................................................................................................... 78
3.3.1 Perspectives expérimentales .................................................................................. 78
3.3.2 Perspectives diagnostiques .................................................................................... 79
3.4 Conclusion ..................................................................................................................... 79
Bibliographie ....................................................................................................................... 80
Annexe ............................................................................................................................... 106

viii
Liste des tableaux
Tableau 1.1. Échelles d’évaluations cliniques de la maladie de Parkinson
Tableau 1.2. Comparaison des types de vésicules extracellulaires
Tableau 2.1. Participant clinical information
Tableau S2.1. Quantification of cell-derived EV – PD cohort
Tableau S2.2. Quantification of cell-derived EV – HD cohort
Tableau S2.3. Complete list of proteins identified in the EEV proteome

ix
Liste des figures
Figure 1.1. Principales catégories de biomarqueurs pour la maladie de Parkinson
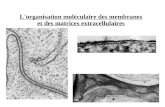
Figure 1.2. Représentation schématique des vésicules extracellulaires
Figure 2.1. EEV: a robust biomarker of PD stage
Figure 2.2. Detection of normal and phosphorylated α-Syn in EEV
Figure 2.3. Specific protein signature of EEV in PD patients
Figure 2.4. Erythrocyte and EV implication in PD: summary of the literature
Figure S2.1. Optimization of EV detection: controls for flow cytometry
Figure S2.2. LEDD and EEV
Figure S2.3. Blood counts and exclusion criteria
Figure S2.4. Proteomic analyses with and without hemoglobin
Figure S2.5. Confirmation of EEV proteins selectively modified in PD patients by Volcano
plots

x
Abréviations
ACE: Addenbrooke’s cognitive examination
α-Syn: alpha-synucléine
AU: Approximately unbiased
BDI: Beck depression inventory
BDS: Burden of disease
CISI-PD: Clinical impression of severity index for Parkinson’s disease
CMV: Corps multivésiculaire
CRP: C-reactive protein
CTRL: Control
DA: Dopamine
DAergique: Dopaminergique
EEV: Erythrocyte-derived extracellular vesicle
EV: Extracellular vesicle
FSC: Foward scatter
GABA: γ-aminobutyric acid
GFP: Green fluorescente protein
GPi: Glubus pallidus interne
H&Y: Échelle de Hoehn et Yahr
HD: Huntington’s disease
IL: Interleukine
IRM: Imagerie par résonance magnétique
LCS: Liquide cérébro-spinal
LEDD: Levodopa equivalent daily dose
LRRK2: Leucine rich kinase
MDS: Movement disorder society
MMSE: Mini-mental state examination
MP: Maladie de Parkinson
MPP+: 1-methyl-4-phenylpyridinium
MPTP: 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine
PBS: Phosphate buffered saline

xi
PD: Parkinson’s disease
PFP: Platelet-free plasma
PINK1: PTEN-induced kinase 1
PPMI: Parkinson’s progression marker initiative
PS: Phosphatidylserine
ROS: Dérivés réactifs de l’oxygène
SNpc: Substance noire pars compacta
SNr: Substance noire pars reticulata
SOD: Superoxide dismutase
STC: Sonographie transcrânienne
TEMP: Tomographie par émission de monophotonique
TEP: Tomographie par émission de positron
TFC: Total functional capacity
UPDRS: Unified Parkinson’s disease rating scale
VE: Vésicule extracellulaire
VEE: Vésicule extracellulaire dérivée d’érythrocyte

xii
Il piacere più nobile è la gioia della comprensione
The noblest pleasure is the joy of understanding
Leonardo da Vinci

xiii
À tous les patients et leur famille.
À mes parents, Mireille et Jean-Nil.

xiv
Remerciements
En premier lieu, j’aimerais sincèrement remercier Dre Francesca Cicchetti qui m’a accueilli
dans son laboratoire et qui m’a offert un projet excessivement stimulant en plus de tout son
appui professionnel. Remerciement spécial à Isabelle St-Amour pour avoir contribué de
façon majeure à ma formation et aux projets. Un grand merci également à Martine St-Pierre
pour avoir été impliqué grandement dans ma formation et avoir été patiente lors des
comportements du projet cysteamine/L-DOPA. De plus, un merci à toute l’équipe et
principalement à Katherine Coulombe et Giulia Cisbani pour leur temps de fin de semaine à
collecter du sang ainsi que leurs précieux conseils.
J’aimerais également adresser un immense merci au Dr Éric Boilard et à toute son équipe
pour leur contribution aux projets. Un merci particulier à Nathalie Cloutier pour son aide plus
importante et grandement appréciée. De plus, j’aimerais remercier Dr Roger Barker et son
équipe sans qui mes projets n’auraient pu être possibles.
Une mention spéciale pour tous les patients et leur famille pour leur don essentiel ayant
permis de réaliser mes projets.
Finalement, je voudrais remercier mes parents, Mireille et Jean-Nil, pour leur encouragement
et leur support durant toute ma maîtrise. Merci pour tout!
Je profite également de l’occasion pour remercier les évaluateurs de mon mémoire pour leur
temps et leur contribution à la finalisation de ma maîtrise.

xv
Avant-propos
Cet avant avant-propos résume les travaux auxquels j’ai contribué durant mes études de
deuxième cycle en neurobiologie. L’objectif principal était d’acquérir des connaissances
scientifiques et techniques en neuroscience et plus spécifiquement, dans le but de découvrir
un nouveau biomarqueur pour la MP.
Le premier chapitre de ce mémoire correspond à une revue de la littérature sur la MP avec
un intérêt particulier pour sa détection clinique et les différents types de biomarqueurs. Une
attention particulière est apportée quant à l’implication des cellules sanguines ainsi que des
VE dans la progression de la pathologie.
Le deuxième chapitre présente l’intégralité d’un article de recherche, dont je suis le premier
auteur, intitulé « Erythrocyte-derived extracellular vesicles: A novel, robust and specific
biomarker that maps to Parkinson’s disease stages » présentement en fin de préparation et
soumission. Il s’agit d’une étude visant à caractériser et démontrer le potentiel des VE
dérivées du sang comme biomarqueur de la maladie de Parkinson. J’ai participé de manière
importante à cette étude en étant impliqué dans la mise en place du projet (conception
expérimentale, collecte de sang), dans toutes les manipulations expérimentales (mise au
point, productions des résultats et analyse), ainsi que dans l’élaboration de toutes les figures,
l’écriture du manuscrit et la soumission à divers journaux (NEJM, JNNP, Lancet, etc).
Finalement, le troisième et dernier chapitre détaille les résultats obtenus et leurs importances
pour dans la MP. On y présente également les perspectives.
Lors de ma maîtrise, j’ai également eu la chance de contribuer à plusieurs autres projets qui
ne se retrouve pas dans ce mémoire. Le premier est intitulé « Toll-like receptor expression in
the blood and brain of patients and a mouse model of Parkinson’s Disease » et est publié dans
l’International Journal of Neuropsychopharmacology. Grâce à cette étude, j’ai pu apprendre
plusieurs techniques de laboratoire qui ont permis de finaliser le projet dès le début de ma
maîtrise. Un deuxième projet m’a permis d’apprendre la manipulation et l’expérimentation
sur modèle murin ainsi que certaines bases sur la pharmacologie parkinsonienne. Cette étude

xvi
s’intitule « The effects of cysteamine in a mouse model of levodopa-induced dyskinesias » et
est présentement en impression dans « Neuroscience Letters ». Finalement, deux autres
manuscrits sur lesquelles je suis co-premier auteur sont en préparation. Pour ces projets, j’ai
repris plusieurs techniques apprises tout au long de ma maîtrise afin de consolider les
différentes parties expérimentales débutées. Les résultats de ces deux manuscrits nous
permettront de mieux comprendre l’implication des plaquettes et de leurs VE dans la maladie
d’Huntington.

1
CHAPITRE 1: INTRODUCTION

2
1.1 La maladie de Parkinson
La maladie de Parkinson (MP) fut d’abord décrite comme étant un syndrome neurologique
par le Dr James Parkinson en 1817 dans son essai, « An essay on the shaking palsy »
(Parkinson, 2002). Cependant, des écrits indien et chinois datant de 1000 avant Jésus-Christ
suggèrent la découverte et la première description de la MP (Manyam, 1990; Zhang et al.,
2006). Suite à l’essai de Parkinson, Jean-Martin Charcot et ses étudiants ont été les premiers
d’une longue liste à contribuer de façon majeure à l’avancer de nos connaissances sur cette
maladie, notamment en caractérisant davantage les symptômes décrits en 1817 (Goetz,
2011). L’apparition et l’intensité des symptômes s’expriment de façon différente selon
l’individu, mais sont souvent représentées par une triade motrice incluant le tremblement au
repos, la rigidité et la bradykinésie (Xia and Mao, 2012). Bien que les symptômes moteurs
soient d’une importance capitale dans le développement de la maladie, plusieurs symptômes
non moteurs affectent grandement la qualité de vie des individus atteints avant même le
diagnostic clinique (Lang, 2011). L’un des premiers signes à apparaître serait lié à la perte
olfactive (Hawkes, 1995). Par la suite, on note l’apparition de différents troubles non moteurs
variant entre les patients. Parmis ces dysfonctions, on peut observer de la constipation, des
troubles du sommeil, de la fatigue, des problèmes de sudation et des perturbations de la vessie
altérant lentement, mais progressivement la qualité de vie des patients jusqu’à l’apparition
de dépression, d’anxiété, d’hallucination et de démence (Davie, 2008; Poewe, 2008; Salawu
et al., 2010; Bago Rožanković et al., 2017).
1.1.1 Épidémiologie
Aujourd’hui, la MP est la deuxième maladie neurodégénérative la plus fréquente après la
maladie d’Alzheimer (Wirdefeldt et al., 2011). Les méta-analyses à travers le monde
rapportent que l’incidence générale (par 100 000 personnes) serait de 41 cas pour les 40 à 49
ans ; 107 cas pour les 50 à 59 ans ; 428 cas pour les 60 à 69 ans ; 1087 cas pour les 70 à 79
ans ; et 1903 cas pour les plus de 80 ans. On remarque donc une augmentation importante du
nombre de cas avec l’âge. Lorsqu’on tient compte des analyses géographiques, on remarque
que les cas chez les personnes de 70 à 79 ans sont significativement plus élevés en Amérique
du Nord, en Europe et en Australie (1602 cas par 100 000) comparativement à l’Asie (646

3
cas par 100 000). Les variations génétiques et environnementales au niveau géographique
pourraient expliquer ce phénomène. En effet, la prise en charge des personnes âgées ainsi
que la méthodologie pour diagnostiquer les patients peut grandement varier entre les pays
(Pringsheim et al., 2014). Finalement, la maladie présente une plus grande atteinte chez les
hommes comparativement aux femmes, à un taux médian de 1,49. Ce débalancement serait
expliqué par les estrogènes ainsi que la susceptibilité génétique du chromosome X (Wooten
et al., 2004).
1.1.2 Étiologie
Les causes exactes de la pathologie restent pour l’instant encore très méconnues malgré
l’intensité des recherches des dernières années et le nombre de découvertes qui y sont liées.
Il semble que 10 % des cas de parkinsonisme seraient le résultat de mutations génétiques,
alors que 90 % des cas seraient définis comme étant d’origine idiopathique (de Lau and
Breteler, 2006; Lesage and Brice, 2009). Cependant, il est de plus en plus suggéré qu’une
combinaison de facteurs génétiques et environnementaux serait responsable de l’apparition
de dysfonctions mitochondriales, de stress oxydatif et d’agrégats protéiques tous impliqués
dans la MP (Greenamyre and Hastings, 2004).
1.1.2.1 Facteurs environnementaux
Les premières études de pathogenèse ont révélé une corrélation entre l’exposition prolongée
à certaines toxines environnementales et l’augmentation des cas de parkinsonisme. Depuis,
des recherches épidémiologiques ont été effectuées afin d’examiner les différents facteurs de
risque de la maladie. Les premières études ont surtout été dirigées vers les pesticides et les
herbicides. Dans cette veine, Priyadarshi et collaborateurs ont rassemblé les études publiées
entre 1989 et 1999 examinant le lien entre l’exposition aux pesticides et le développement
de la pathologie. Les résultats montrent une hétérogénéité significative entre les deux
phénomènes (Priyadarshi et al., 2000). On sait aujourd’hui que le contact quotidien avec
certains pesticides comme la roténone peut provoquer les caractéristiques anatomiques,
neurochimiques et comportementales de la MP chez le rongeur en interférant avec le
complexe 1 de la chaine respiratoire de la mitochondrie (Betarbet et al., 2000; Lapointe et

4
al., 2004; Testa et al., 2005). De plus, la contamination de l’opioïde desméthylprodine par le
1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) a montré que la MP peut être
engendrée par des composés synthétiques. En effet, plusieurs personnes ont fait l’expérience
de symptômes typiquement parkinsoniens suite à l’injection de drogues (Langston et al.,
1983).
Dans les années 90, on remarque également un intérêt croissant pour les facteurs de risque
liés à la vie rurale, aux activités agricoles et à la consommation d’eau de puits (Koller et al.,
1990; Seidler et al., 1996). Les agriculteurs et les individus vivant en milieux ruraux sont
davantage susceptibles d’être exposés aux pesticides. Il peut donc y avoir une augmentation
du facteur de risque lié à une longue exposition, bien qu’indirecte, à plusieurs neurotoxines
(Hubble et al., 1993; Lai et al., 2002).
Il a également été suggéré qu’une exposition aux métaux lourds comme le fer, le mercure, le
manganèse, le cuivre, le zinc et le plomb entraînerait une accumulation dans le cerveau
menant à la dégénérescence et à l’augmentation du stress oxydatif directement en lien avec
le risque de développer la maladie (Ngim and Devathasan, 1989; Semchuk et al., 1993; Kuhn
et al., 1998; Caudle, 2015).
Finalement, on retrouve plusieurs études ciblant les habitudes de vie comme facteur de
risque. Il existe plusieurs cibles dans cette catégorie telle que la cigarette. Bien qu’on pourrait
penser que la cigarette augmente le risque de développer la MP avec tout ces composés
chimiques et son rôle néfaste déjà connu dans plusieurs maladies, il n’en est rien. Au
contraire, plusieurs recherches montrent que le tabac diminuerait les risques de 40 à 50 % de
développer la maladie (Grandinetti et al., 1994; Hernán et al., 2001; Searles Nielsen et al.,
2012; Ritz et al., 2007). Des preuves montrent que le striatum, une structure particulièrement
affectée par la MP et présentée à la section 1.1.3.1, permet une étroite relation entre les
neurotransmetteurs cholinergiques nicotiniques et dopaminergiques (DAergique) (Zhou et
al., 2002). La nicotine et ses récepteurs jouent un rôle clé dans la régulation de l’activité
striatale et des comportements médiés par le système DAergique (Quik et al., 2009). En
neurologie expérimentale, la nicotine a montré des effets bénéfiques contre les dommages au

5
striatum et une amélioration de la fonction motrice chez certains modèles animaux de la MP
(Bordia et al., 2006). La nicotine et ses agonistes pourraient également réduire les dyskinésies
induites par la levodopa (Quik et al., 2013). On retrouve également d’autres produits pouvant
influencer les facteurs de risques tels que les diètes méditerranéennes, très riche en huile de
poisson (Archer and Kostrzewa, 2016). Ces huiles sont en grande partie composées d’acide
gras à chaîne longue polyinsaturée. Il apparaît dans la littérature que ces composées
pourraient agir comme agents neuroprotecteurs (Bousquet et al., 2011; Coulombe et al.,
2016). Il est suggéré qu’ils pourraient protéger les neurones de la toxicité, en inhibant la
production d’oxyde nitreux, en régulant les flux calciques et en augmentant l’activité des
enzymes antioxydantes. On retrouve également plusieurs autres composés impliqués dans les
habitudes de vie qui peuvent jouer sur les facteurs de risques de développer la MP (la caféine,
le soya et l’alcool) (Seidl et al., 2014).
1.1.2.2 Facteurs génétiques
Depuis 1997, un bon nombre de familles ont été identifiées comme parkinsoniennes, estimées
maintenant à environ 10 % des cas de MP (Healy et al., 2004). Bien que plusieurs
caractéristiques cliniques soient similaires aux cas sporadiques, on remarque que les cas
familiaux se manifestent généralement de manière beaucoup plus précoce (Hardy et al.,
2003).
La première mutation reliée à la MP fût découverte sur le gène SNCA codant pour la protéine
α-Syn et ce voulait être une mutation faux sens (A53T) (Polymeropoulos et al., 1997). Par la
suite, deux autres mutations furent détectées, soit A30P et E46K (Krüger et al., 1998; Zarranz
et al., 2004). Cependant, les mutations faux sens n’ont été retrouvées que dans les cas de
parkinsonisme générationnel à une rare fréquence et jamais dans les cas sporadiques (Berg
et al., 2005). La recherche a également montré que le locus contenant le gène SNCA pouvait
être, dans certains cas, dupliqué et même tripliqué chez des cas familiaux et sporadiques
(Singleton et al., 2003; Ahn et al., 2008). On remarque que l’expression de mutations sur ce
gène autosomique dominant semble provoquer l’apparition de la maladie de manière précoce
avec une progression rapide et une prévalence pour les troubles psychiatriques (Golbe et al.,

6
1996). L’étude des mutations sur le gène SNCA nous a permis de comprendre l’importance
de l’α-Syn et son agrégation pathologique dans le développement de la maladie autant chez
les cas génétiques que sporadiques (Spillantini et al., 1997; Rosborough et al., 2017).
Possédant à ce jour 20 mutations faux sens et non-sens, la protéine leucine rich repeat
kinase 2 (LRRK2) liée à une hérédité autosomique dominante semble être impliquée dans la
majorité des cas familiaux de la MP (Zimprich et al., 2004; Cookson, 2010). Responsable de
7 % des cas génétiques de la maladie et de 1 à 2 % des cas sporadiques, la mutation G2019S
est la plus fréquente (Nichols et al., 2005; Gilks et al., 2005). Bien que les fonctions de
LRRK2 ne soient connues que partiellement, la protéine mutée semble provoquer un
raccourcissement et une simplification de l’arbre dendritique des neurones (MacLeod et al.,
2006).
Davantage de mutations ont été découvertes, mais d’ordre autosomique récessive. Les
mutations récessives les plus fréquences proviendraient de la protéine Parkin (Lücking et al.,
2000; Hedrich et al., 2004). Ses mutations peuvent varier du simple changement d’acides
aminés au réarrangement d’un exon (Lücking et al., 2001). Parkin, ayant une activité E3
ligase ubiquitine, participerait à maintenir l’intégrité de la mitochondrie et à signaler
l’autophagie lorsqu’il y a dégradation de cette dernière. Les diverses mutations pourraient
donc empêcher le rôle majeur de Parkin dans la régulation de la mitochondrie (Shimura et
al., 2000; Narendra et al., 2008; Berger et al., 2009). On retrouve également dans cette
famille, le gène DJ-1, localisé dans les terminaisons synaptiques, les mitochondries et
certaines membranes d’organelles. Il serait lié à la régulation du stress oxydatif en jouant un
rôle d’antioxydant (Olzmann et al., 2007; Wang et al., 2011). Trois mutations faux-sens
(L166P, M26I, E64D) ont été découvertes sur 5 des 8 exons de ce gène (Abou-Sleiman et al.,
2003). Finalement, on retrouve le gène PTEN induced kinase 1 (PINK1) exprimé en kinase
et qui serait associé à la dynamique de la mitochondrie et à ses fonctions dans la respiration
cellulaire (Weihofen et al., 2009; Amo et al., 2011).

7
1.1.3 Neuropathologie
1.1.3.1 Dégénérescence du système DAergique
L’une des déficiences pathologiques des plus évidentes dans la MP est la perte de neurones
DAergiques au niveau de la substance noire pars compacta (SNpc) (Kordower et al., 2013;
Capriotti and Terzakis, 2016). La dégénérescence des projections DAergique de la SNpc vers
le striatum engendre une diminution importante de la dopamine (DA) dans ce sentier cérébral
(Calon et al., 2003). La partie ventrolateral de la SNpc reliée à la partie dorsale du putamen
dans le striatum est sans doute la plus touchée, comptabilisant 95 % des pertes en neurones
DAergiques (Damier et al., 1999). Des études ont d’ailleurs montré qu’une altération
pathologique menant à une perte modérée ou sévère des cellules DAergiques dans cette
région serait à l’origine de la bradykinésie et de la rigidité (Dickson et al., 2009).
À partir des années 80, la description des modèles de voies décrivant l’architecture entourant
les ganglions de la base a représenté une avancée décisive dans le domaine des neurosciences
(Penney and Young, 1986; Albin et al., 1989; Prensa et al., 2003). Aujourd’hui, nous savons
que la voie de signalisation vers les ganglions de la base via le striatum, le globus pallidus
interne (GPi) et la SN pars reticula (SNr) est principalement touchée dans la MP. L’activité
des neurones épineux moyens du striatum se traduit par la transmission du signal vers le GPi
et la SNr via une synapse γ-aminobutyric acid (GABA)ergique (voie directe). Il en résulte
une inhibition de l’activité inhibitrice dirigée vers le thalamus. Il existe également une voie
vers le globus pallidus externe via plusieurs synapses GABAergiques passant par le noyau
sous-thalamique (voie indirecte) qui vient amplifier le signal inhibiteur dirigé vers le
thalamus. On retrouve donc une voie directe qui agit de manière excitatrice sur le cortex et
une voie indirecte exerçant une fonction inhibitrice sur le cortex (Redgrave et al., 2010).
Lors de condition physiologique, la DA relâchée par la voie nigrostriale module la
transmission corticostriale au niveau des neurones épineux moyens exprimant les récepteurs
à DA, D1 ou D2, qui vont mener à l’activation ou à la suppression du mouvement,
respectivement (Kravitz et al., 2010; Tritsch and Sabatini, 2012; Calabresi et al., 2014). En
plus de jouer un rôle dans le contrôle moteur, plusieurs recherches suggèrent que la DA

8
provenant de la SNpc pourrait jouer un rôle important dans l’apprentissage moteur (Faure et
al., 2005; Bromberg-Martin et al., 2010).
Plusieurs voies ont été proposées pour expliquer la vulnérabilité et la dégénérescence des
neurones DAergiques. L’augmentation du nombre de synapses par neurone, par exemple,
pourrait accroitre la vulnérabilité de ces cellules en les surexposant à des stimulations de
d’autres populations de neurones (Hindle, 2010). De plus, un déséquilibre dans l’homéostasie
de la DA synaptique pourrait induire l’accumulation de ce neuromédiateur dans le
cytoplasme des neurones, augmentant le taux d’oxydation et de production de dérivés réactifs
de l’oxygène (ROS) (Bisaglia et al., 2007). On note également que la neuromélanine présente
dans les neurones DAergiques pourrait quant à elle lier les métaux toxiques, comme le fer,
et contribuer à la neurodégénérescence (Sian-Hülsmann et al., 2011; Zucca et al., 2017).
Finalement, il a été montré que les neurones DAergiques sont plus susceptibles à l’action des
médiateurs inflammatoires comparativement aux autres types de neurones (Herrera et al.,
2005).
1.1.3.2 L’alpha-synucléine et les corps de Lewy
Bien que les causes des maladies neurodégénératives, dont le Parkinson, soient aussi
nombreuses que leur diversité, la grande majorité d’entre elles ont en commun l’agrégation
de protéines. Les maladies neurodégénératives, appelées protéinopathies, sont également
caractérisées par de mauvais enchevêtrements de protéines principalement au niveau
cytosolique (tel qu’observé dans la MP) et au niveau extracellulaire (par exemple, la maladie
d’Alzheimer) (Rubinsztein, 2006). Dans la MP, l’α-Syn fut identifiée comme étant une
protéine importante, notamment par la découverte de mutation sur le gène SNCA, comme
discuté à la section 1.1.2.2 (Polymeropoulos et al., 1997; Spillantini et al., 1997).
L’α-Syn est une protéine soluble de 140 acides aminés identifiés dans plusieurs maladies
surnommées « Synucléinopathies » (Snead and Eliezer, 2014). La fonction ou les fonctions
physiologiques de l’α-Syn restent pour l’instant inconnu. Cependant, on reconnaît sa
présence aux terminaisons présynaptiques ainsi que son association aux vésicules
synaptiques. Lorsque la protéine est surexprimée ou sous-exprimée, on remarque une

9
déficience dans la transmission synaptique, suggérant des rôles dans la régulation du
relargage des neurotransmetteurs. L’α-Syn pourraient donc jouer sur la fonction synaptique
et/ou la plasticité synaptique (Withers et al., 1997; Lee et al., 2008; Watson et al., 2009;
Lashuel et al., 2012). Des études semblent également montrer qu’elle pourrait avoir une
interaction directe avec la DA (Souza et al., 2000; Perez et al., 2002; Butler et al., 2015;
Fakhree et al., 2016). Sous forme phosphorylée ou ubiquitinée, l’α-Syn se lie à d’autres
protéines telles que la synphiline-1, l’ubiquitine et Cdk5 pour former des agrégats
(Wakabayashi and Takahashi, 2000; Kawamata et al., 2001; Fujiwara et al., 2002; Tofaris et
al., 2003; Nonaka et al., 2005). L’α-Syn peut exister sous différentes structures incluant une
forme monomérique soluble, une forme tétramérique pliée en hélice-α résistante à
l’agrégation, une forme polymérique soluble ou une forme fibrillaire regroupé en feuillets β
pouvant mener à la formation d’enchevêtrements protéiques appelés corps de Lewy (Bartels
et al., 2011; Volpicelli-Daley et al., 2011; Fauvet et al., 2012).
Pour le moment, nous ignorons toujours le rôle des corps de Lewy dans le développement de
la maladie, bien qu’il s’agisse d’une des principales caractéristiques neuropathologiques de
la MP (Holdorff et al., 2013; Rodrigues e Silva et al., 2010). Certains pensent que ces agrégats
sont générés dans le but de protéger les neurones de la toxicité induite par la protéine
pathologique. D’autres suggèrent plutôt que l’accumulation serait elle-même toxique et
qu’elle altérerait le fonctionnement cellulaire (Harrower et al., 2005).
L’évolution des agrégats d’α-Syn au cours de la progression de la MP fût décrite par Braak
en 2003 et illustrait l’apparition des premières inclusions dans le bulbe olfactif, expliquant la
perte d’odorat comme premier symptôme du Parkinson (Braak et al., 2003). Des lésions sont
ensuite détectées au niveau du noyau dorsal des nerfs glossopharyngien et vague expliquant
plusieurs troubles non moteurs tels que l’hypotension, des troubles de déglutition et plusieurs
autres problèmes touchant le système nerveux autonome au début de la maladie (Braak et al.,
2006).
Suite aux nombreuses études post-mortem effectuées démontrant les stades évolutifs des
agrégats selon différents niveaux de la maladie, la recherche s’est penchée sur la technique

10
de propagation de la protéine. L’une des hypothèses les plus populaires est celle du prion-
like spread, suggérant une transmission de cellule à cellule où la protéine démontre des
propriétés infectieuses (Bernis et al., 2015; Brundin et al., 2016; Verma, 2016). Il a également
été démontré que les cellules pourraient utiliser leur système d’exocytose et d’endocytose
pour générer des vésicules transportant des protéines agrégées comme l’α-Syn
(Emmanouilidou et al., 2010; Danzer et al., 2012; Candelario and Steindler, 2014). Ce mode
de propagation physiologique serait d’ailleurs grandement augmenté lorsque le lysosome est
affecté, comme dans la MP (Zhang et al., 2009; Alvarez-Erviti et al., 2011). Complémentaires
à ces études, les analyses de tissus post-mortem provenant de patients ayant reçu des greffes
de cellules fœtales ont révélé des inclusions de corps de Lewy au sein des neurones greffés,
appuyant les hypothèses de propagation intercellulaire (Kordower et al., 2008; Chu and
Kordower, 2010).
1.1.3.3 Stress oxydatif et facteurs de mort cellulaire
Le stress oxydatif est un événement important dans le processus du vieillissement normal
puisqu’on remarque une diminution de la capacité à limiter et réparer les dommages oxydatifs
chez les personnes plus âgées (Sohal and Weindruch, 1996). Ce phénomène est en réalité
provoqué par un déséquilibre du ratio entre la production de ROS et une mauvaise
détoxification, pouvant être induit par une diminution d’antioxydants (Karihtala and Soini,
2007). Le neurotransmetteur DAergique joue un rôle crucial dans ce phénomène puisqu’il
produit des métabolites excessivement toxiques. L’autooxydation de la DA, la DA-quinone,
est capable de modifier de manière covalente des protéines et résidus essentiels à la survie de
la cellule (Ma et al., 2015). Il a notamment été montré que dans la pathophysiologie de la
MP, la DA-quinone modifie les monomères d’α-Syn en favorisant leur conversion en forme
fibrillaire cytotoxique (Conway et al., 2001). De plus, lors de la transformation de la DA, il
y a création de ROS très réactif via le peroxyde d’hydrogène. La formation de ROS peut
induire une dysfonction des protéines, des lipides et de l’ADN, résultant en une dégradation
membranaire et structurale de la cellule. Cette dernière peut donc être exposée à une
augmentation de l’entrée d’ion et une augmentation de la sortie de DA et de ROS, favorisant
le stress oxydatif (Lotharius and Brundin, 2002). Finalement, la DA-quinone pourrait causer

11
l’inactivation des transporteurs à DA, inhiber la tyrosine hydroxylase impliquée dans la
transformation de la DA et mener à la dysfonction des mitochondries (Kuhn et al., 1999; Lee
et al., 2002).
Une perte neuronale dans la MP peut également être associée à une inflammation chronique
contrôlée à la base par les microglies, des cellules responsables de la réponse immunitaire
innée dans le système nerveux central. Une réaction inflammatoire de ces cellules est perçue
dans la SN et le striatum de modèle murin de MP induit par le MPTP ainsi que dans la SN
de patients (idiopathique et génétique) (McGeer et al., 1988; O’Callaghan et al., 1990;
Machado et al., 2016). Les microglies sont activées comme mécanisme d’autodéfense contre
les pathogènes et débris cellulaires en réponse à une toxicité ou une blessure. Lorsqu’elles
sont activées, ces cellules relâchent des radicaux libres comme l’oxyde nitreux et l’ion
superoxyde qui peuvent contribuer au stress oxydatif dans l’environnement des neurones. Il
est proposé comme hypothèse qu’une suractivité ou une activité chronique des microglies
augmente drastiquement et de manière incontrôlée la réponse inflammatoire, menant à la
neurodégénération par un cercle vicieux d’autoactivation (Qian et al., 2010). Des études ont
montré que la SN est sujette à l’augmentation de « tumor necrosis factor » TNF-α,
d’interleukine (IL)-1β, de l’interféron-γ, de l’oxyde nitrique synthétase et du complexe
majeur d’histocompatibilité II alors que le striatum est caractérisé par une augmentation
d’IL-1β, d’IL-2, d’IL-6 et de TNF-α (Hunot and Hirsch, 2003; Stojkovska et al., 2015).
1.1.4 Diagnostic et test clinique
Le diagnostique d’un patient atteint de la MP est actuellement fait à l’examen clinique par le
médecin expérimenté. Par la suite, plusieurs tests cliniques permettant d’analyser différents
aspects de la maladie peuvent être utilisés afin d’aider le médecin a évaluer la progression de
la maladie. Une impression de la sévérité globale est utilisée à des fins de compréhension et
de mesure au niveau de la recherche fondamentale et clinique. Depuis la publication de
l’échelle de Hoehn et Yahr en 1967, l’évolution de la méthode d’évaluation de la MP n’a
cessé de croître (Hoehn and Yahr, 1967). Nous retrouvons aujourd’hui une variété de tests
cliniques touchant des aspects très spécifiques à la MP comme le Parkinson’s Fatigue Scale

12
et le Parkinson’s Disease Dyskinesia Scale alors que d’autres vont cibler de manière plus
générale la maladie comme le Unified Parkinson’s disease rating scale (UPDRS) et l’échelle
de Hoehn et Yahr (H&Y) (Brown et al., 2005; Katzenschlager et al., 2007; Fahn and Elton,
1987; Hoehn and Yahr, 1967). Avec les connaissances accrues que nous avons de la maladie,
le nombre de tests cliniques et d’échelles a largement augmenté permettant de donner une
valeur diagnostique plus précise à l’aide d’une combinaison de ces évaluations.
Le UPDRS ne fût pas la première échelle d’évaluation du parkinsonisme à être inventé, mais
elle est aujourd’hui l’une des plus utilisées (Ramaker et al., 2002). Créé par Fahn et Elton en
1987, le UPDRS suit la progression longitudinale de la MP selon 4 champs principaux (Fahn
and Elton, 1987). La première partie évalue les aptitudes mentales, comportementales et
l’humeur. Une deuxième section permet une auto-évaluation (si possible) des activités
quotidiennes. Une troisième catégorie permet au clinicien d’évaluer la motricité du patient.
Finalement, la quatrième section évalue les complications de la thérapie (exemple : les
dyskinésies) (Goetz et al., 2008). On note également une cinquième section composée du
H&Y où le score vient s’additionner au total des quatre sections précédentes. Parfois, selon
l’édition du UPDRS, certains cliniciens vont ajouter une sixième section composée de
l’échelle de Schwab and England.
Margaret M. Hoehn et Melvin D. Yahr ont publié une échelle descriptive simple combinant
une évaluation des déficits fonctionnels (invalidité) et des signes de détérioration motrice qui
aujourd’hui reste l’une des évaluations les plus utilisées pour la MP (Hoehn and Yahr, 1967).
Bien que l’échelle fût initialement construite avec 5 niveaux (1-5), on lui a ajouté des
variations de 0,5 afin d’augmenter sa précision (Jankovic et al., 1990). L’échelle de H&Y est
établie selon le principe que l’ensemble des dysfonctions touchant les parkinsoniens
impliquent une motricité bilatérale et un compromis entre balance et démarche.
L’augmentation de déficience motrice peut donc être évaluée à partir du comportement
unilatéral (Stade 1) suivi d’un comportement bilatéral, mais sans problème d’équilibre
(Stade 2), d’une perte de la stabilité posturale (Stade 3), d’une perte d’indépendance
physique (Stade 4) jusqu’à l’étape du fauteuil roulant ou l’alitement (Stade 5) (Goetz et al.,
2004).

13
Le Mini-Mental State Examination (MMSE), développé en 1975 par Folstein et McHugh, est
un questionnaire qui permet l’évaluation des fonctions cognitives et la capacité mnésique
d’un individu. Il s’agit d’un test à 30 points évaluant l’orientation spatiale et temporelle,
différentes capacités (apprentissage, concentration, mémoire et calcul), le langage, la
reconnaissance et diverses tâches complexes (Folstein et al., 1975). Il est fréquemment utilisé
pour évaluer le niveau de démence. Dans plusieurs pathologies telles que les maladies
d’Alzheimer et de Parkinson, il est utilisé pour évaluer la progression et la sévérité de la
déficience cognitive (Tombaugh and McIntyre, 1992). Une version modifiée du MMSE,
appelée Standardized MMSE fut validée en 1991 afin d’optimiser la fiabilité du test
principalement chez les patients plus âgés et plus sévèrement atteints par une déficience
cognitive (Molloy et al., 1991; Pangman et al., 2000). Le MMSE est donc devenu un outil
très utile et répandu puisqu’il a comme avantage d’être facile à utiliser, rapide (environ 5
minutes) et de ne nécessiter aucun équipement spécialisé (Harrell et al., 2000).
Dans une catégorie similaire au MMSE, on retrouve le Addenbrooke’s cognitive examination
(ACE) évaluant, par une batterie de tests d’environ 15 minutes, les types de démences aux
stades précoces chez les personnes soufrant de la maladie d’Alzheimer, de démence frontaux
temporale, de la MP, etc. (Mathuranath et al., 2000). Se basant sur le MMSE, le ACE
incorpore davantage de questions notamment au niveau des capacités d’orientation,
d’attention, de mémoire, de parole (flux et langage) et d’habiletés visuospatiales (Dudas et
al., 2005). Comme la majorité des tests cliniques, le ACE a fait l’objet d’une restructuration
permettant d’augmenter sa sensibilité et sa spécificité (Mioshi et al., 2006).
La dépression est très présente (40 %) chez les parkinsoniens et a un impact important sur la
qualité de vie des patients (Cummings, 1992; Schrag et al., 2000). Le Beck Depression
Inventory (BDI), créé par Aaron T. Beck, est l’un des tests psychométriques les plus utilisés.
Il permet de mesurer la sévérité de la dépression. Comportant 21 questions à choix de
réponses, le test peut être fait par le patient lui-même (Richter et al., 1998). L’approche de
Beck sur la « cognition négative », touchant l’entourage, le soi et l’avenir permet de définir

14
un ensemble de symptômes jouant un rôle majeur dans la dépression comme le désespoir,
l’irritabilité, la culpabilité, la fatigue, la perte de poids, etc. (Beck et al., 1961).
En 2006, l’indice clinimétrique Clinical Impression of Severity Index for Parkinson’s
Disease (CISI-PD) est proposé afin d’améliorer le diagnostic clinique de la MP. Construite
à partir du Clinical Global Impression of Severity, le CISI-PD reflète et résume l’évaluation
des autres échelles cliniques en se basant sur les symptômes moteurs, l’invalidité, les
complications moteurs et la détérioration cognitive (Martínez-Martín et al., 2006). L’objectif
étant d’incorporer des éléments d’échelles déjà existants ainsi que la perception du clinicien
afin de créer un instrument puissant pour valider la progression de la MP (Martínez-Martín
et al., 2009).
Il existe toutefois un problème considérable avec les tests diagnostiques. Les échelles
cliniques ne sont souvent utilisées que lorsque les symptômes sont avancés chez les patients.
On sait aujourd’hui que le niveau de mort neuronale du système DAergique peut atteindre
l’ordre des 60 à 70 % avant que la maladie ne soit diagnostiquée (Fearnley and Lees, 1991;
Dauer and Przedborski, 2003; Cheng et al., 2010). Pour cette raison, il est nécessaire de
remédier au manque flagrant de biomarqueurs et d’imagerie neurologique, afin de détecter
les dommages neurologiques le plus tôt possible.

15
Tableau 1.1. Échelles d’évaluations cliniques de la maladie de Parkinson
Échelle d’évaluation clinique
Abréviation Évaluation Stratification Mise à jour Références
Unified Parkinson’s disease rating scale
UPDRS 1. Expériences non motrices au quotidien
2. Expériences motrices au quotidien
3. Examination motrice 4. Complications motrices
Aucune MDS-UPDRS (Fahn and Elton, 1987; Goetz et al., 2008; Martínez-Martín et al., 2015)
Échelle de Hoehn et Yahr H&Y Symptômes moteurs Normal = 0 Léger = 1-2 Modéré = 3 Sévère = 4-5
Modifié (Hoehn and Yahr, 1967; Goetz et al., 2004)
Mini-mental state examination
MMSE 1. Orientation dans le temps et l’espace
2. Apprentissage et transcription
3. Attention et de calculs 4. Capacité mémorielle 5. Langage et identification
d’objets 6. Praxie constructive
Normal = 24-30 Léger = 19-23 Modéré = 10-18 Sévère = 0-9
SMMSE (Folstein et al., 1975; Tombaugh and McIntyre, 1992)
Addenbrooke’s cognitive examination
ACE 1. Orientation 2. Attention 3. Mémoire 4. Flux verbal 5. Langage 6. Habilité visuospatiale
Aucune, Maximum = 100
ACE-R (Mathuranath et al., 2000; Dudas et al., 2005; Mioshi et al., 2006)
Beck Depression Inventory BDI Sentiments au quotidien Normal = 1-10 Très léger = 11-16 Léger = 17-20 Modéré = 21-30 Sévère = 31-40 Extrême = 41-63
BDI-II (Beck et al., 1961; Visser et al., 2006)
Clinical impression of severity index for Parkinson’s disease
CISI-PD 1. Signe moteur 2. Invalidité 3. Complication 4. Évaluation cognitive
Normal = 0 Léger = 1-7 Modéré = 8-14 Sévère = 15-24
Aucune (Martínez-Martín et al., 2006)

16
1.1.5 Biomarqueur
1.1.5.1 Introduction
Il n’existe pour l’instant aucun biomarqueur permettant de prédire l’apparition de la MP ou
de valider le diagnostic. Bien que les médecins spécialistes soient aptes à poser le diagnostic,
de mauvais verdicts se produisent principalement au début de la maladie. En raison de la
difficulté à diagnostiquer cette maladie, la recherche d’une bonne cible comme biomarqueur
est extrêmement difficile, mais nécessaire.
Le Biomarkers Definitions Working Group définit un biomarqueur comme étant « une
caractéristique qui est mesurée de manière objective et évaluée comme un indicateur de
processus biologiques normaux, de processus pathologiques ou de réponses
pharmacologiques à une intervention thérapeutique ». Un bon biomarqueur doit idéalement
être sensible, reproductible, spécifique, peu couteux, non invasif, facile à mesurer et facile à
valider (Mayeux, 2004; Parnetti et al., 2013). En 2001, Pepe et collaborateurs ont conçu un
modèle pour le développement de biomarqueurs en cinq phases permettant de dépister le
cancer dans la population générale. Chaque phase possède un ou deux objectifs principaux
avec des mesures de résultats pertinentes, ainsi que plusieurs objectifs secondaires (Pepe et
al., 2001). Ce modèle, bien que développé pour le cancer, peut être utilisé pour d’autres
maladies telles que la MP. La première phase de recherche pour un bon biomarqueur doit
être constituée d’études exploratoires précliniques évaluant, entre autres, la spécificité et la
sensibilité de chaque cible. Le but étant de pister des marqueurs potentiellement utiles et de
les hiérarchiser. La deuxième phase vise à estimer la spécificité et la sensibilité au niveau
clinique et évaluer la capacité du marqueur à distinguer les patients des sujets sains. Il s’agit
donc de débuter le développement clinique du test. La troisième phase vise à évaluer, en
fonction du temps avant le diagnostic clinique, la capacité du biomarqueur à détecter la
maladie au niveau préclinique. Par la suite, l’étape 4 permet de déterminer les caractéristiques
de fonctionnement du test de dépistage basé sur les biomarqueurs dans une population
pertinente en étudiant le taux de détection et le taux de faux positifs. Finalement, la dernière
phase se doit de prédire l’impact du test dans la population en regardant notamment, l’impact

17
sur la détection réelle de futurs patients, le coût et la disponibilité des traitements (Pepe et
al., 2001; Frisoni et al., 2017).
Puisque dans la MP, le diagnostic est exclusivement clinique et qu’il est possible seulement
après une dégénérescence majeure des neurones de la SNpc, il est nécessaire d’avoir un
biomarqueur prodromique, capable de cibler les phases précoces ou dévaluer le risque de
développer la maladie. Un marqueur prodromique serait extrêmement utile dans la MP
puisqu’il permettrait de diagnostiquer et d’identifier la maladie avant l’apparition des
symptômes moteurs pour éventuellement appliquer des thérapies neuroprotectrices.
Malheureusement, il s’agit du type de biomarqueur le plus difficile à développer puisque les
troubles précoces de la MP, comme la constipation, ne sont pas spécifiques à cette
neuropathologie (Cooper and Chahine, 2016). Un biomarqueur détecté après l’apparition des
symptômes pourrait néanmoins nous permettre d’évaluer les stades cliniques/moteurs
(Sharma et al., 2013). Il permettrait de créer une charte de progression de la maladie et établir
l’efficacité de diverses thérapies. La recherche nous montre que ces marqueurs biologiques
pourraient être basés sur des facteurs cliniques, génétiques, protéomiques, biochimiques,
d’imagerie ou une combinaison de plusieurs (Sharma et al., 2013; Delenclos et al., 2016).
Un grand nombre de stratégies ont été déployées afin de trouver des biomarqueurs pour la
MP, étudiant tout d’abord les personnes déjà diagnostiquées (Miller and O’Callaghan, 2015).
Ces efforts permettent d’augmenter nos connaissances de la pathologie et ainsi cibler des
stades spécifiques de la maladie avec de nouveaux traitements (Cisbani et al., 2015;
Coulombe et al., 2016; Chung et al., 2017). De plus, le Progression Marker Initiative (PPMI),
une étude clinique financée par la fondation Michael J. Fox, fut créé en tant que multicentre
d’étude internationale travaillant sur l’identification de biomarqueur afin d’évaluer la
progression de la MP. Pour ce faire, le projet compile les données sur les cas rapportés de
MP et sur les essais thérapeutiques (Parkinson Progression Marker Initiative, 2011; Nalls et
al., 2016).

18
1.1.5.2 Biomarqueurs génétiques
Pendant plusieurs années, la génétique ne fut pas considérée comme un facteur d’impact
important puisque les cas familiaux ont toujours été beaucoup moins nombreux que les cas
sporadiques. Maintenant, nous savons que la forme idiopathique peut être due à un réseau de
plusieurs facteurs incluant la génétique (Alonso-Navarro et al., 2014). La découverte de
formes familiales rares et précoces de la MP a mis en évidence l’importance de la génétique
par une variété de mutations sur des gènes uniques. L’exemple le plus évident est celui du
gène SNCA qui, tel qu’expliqué à la section 1.1.2.2, peut subir des mutations ponctuelles,
des duplications ou triplications impliquant une forte pénétrance et un haut risque de
développer la pathologie (Gasser, 2009). En conséquence, la caractérisation des formes
monogéniques autosomales dominantes de la MP nous a permis d’identifier des gènes
critiques, tels que SNCA, DJ-1, PINK1, Parkin et LRKK2. On pourrait donc envisager
d’utiliser ces gènes et de leur produit protéique associé à la maladie comme biomarqueur
pour les cas sporadiques.
1.1.5.3 Biomarqueurs par imagerie
Le développement de technique de neuroimagerie nous permet d’employer diverses
méthodes dans le but d’étudier la structure et les fonctions du cerveau pour compléter les
tests diagnostics (Pavese and Brooks, 2009). La neuroimagerie utilisant la sonographie
transcrânienne (STC), la tomographie par émission de positron (TEP), la tomographie par
émission monophotonique (TEMP) et l’imagerie par résonance magnétique (IRM) peuvent
nous fournir des informations importantes afin de suivre des molécules cibles dans la
dégénérescence des cellules (Hsiao et al., 2014; Isaias et al., 2016; Toomsoo et al., 2016).
Ces approches ont comme avantage d’être non invasives et très rapides pour nous fournir une
vision de la morphologie du cerveau, dont l’intégrité du système DAergique. Par contre,
toutes ces techniques ont un coût très élevé réduisant leur utilisation dans des conditions
standards.

19
1.1.5.4 Biomarqueurs cliniques
On connaît aujourd’hui un ensemble de symptômes apparaissant très tôt durant le
développement de la maladie. Ainsi, les problèmes de sommeil paradoxal, d’aptitudes
visuospatiales, d’olfaction et de cognition pourraient permettre la mise en place de tests
fonctionnels non invasifs pouvant évaluer les facteurs de risque pour la MP (Pellicano et al.,
2007; Postuma et al., 2006; Doppler et al., 2017). On note également lors des phases
précoces, le développement de symptômes dépressifs venant d’altérations au niveau du
contrôle émotionnel, un marqueur important du processus neurodégénératif (Borgonovo et
al., 2017). Des évidences suggèrent que la constipation reliée à des problèmes gastro-
intestinaux pourrait être utilisée comme outil de diagnostic (Fasano et al., 2015). Ces
nombreux tests utilisés dans différentes études et combinés à d’autres facteurs de risque
comme l’âge pourraient servir à développer des algorithmes en vue d’un programme de
dépistage. Bien que cette approche soit prometteuse, son utilité ne peut être démontrée que
par la validation de caractéristiques prémotrices prédisant réellement la progression de la
MP. De plus, la plupart des caractéristiques envisagées ne sont pas spécifiques à la MP, ce
qui peut conduire à des diagnostics faux positifs et négatifs. Il existe donc un très grand
problème éthique face à ce type de biomarqueur.
1.1.5.5 Biomarqueurs biochimiques
Le développement de biomarqueurs pour les maladies neurodégénératives comme la MP est
principalement limité par l’accès au tissu cérébral avant l’autopsie du patient. La recherche
s’est donc penchée sur du tissu plus facile d’accès soit le liquide cérébro-spinal (LCS), le
sang, l’urine, la salive ainsi que des biopsies du tractus gastro-intestinal. Puisque les échanges
entre le cerveau et le LCS sont bien établis, ce dernier est sans doute celui qui est le plus
envisagé pour refléter les processus de dégénérescence au cerveau (Andersen et al., 2017).
Bien que le LCS possède des caractéristiques attrayantes comme biomarqueur, il n’en reste
pas moins qu’il n’est pas aussi facile d’accès comparativement à l’urine ou le sang, et ainsi
il demeure dans la catégorie des biomarqueurs invasifs. Une grande quantité de composantes
biologiques (protéines, tissus, gènes) ont été évaluées en périphérie pour leur pertinence en
tant que biomarqueur. On retrouve notamment chez le patient, une diminution de

20
catécholamine dans le LCS, une augmentation des corps de Lewy dans la glande sous-
mandibulaire et un changement métabolomique dans l’urine et dans le plasma (bilirubine,
biliverdine, tryptophane) (Goldstein et al., 2012; Beach et al., 2013; Luan et al., 2015; Hatano
et al., 2016). Pour le moment, l’α-Syn est l’aspect pathologique le plus utilisé au niveau des
tissus et des fluides dus à sa grande importance dans la maladie et à sa présence en grande
quantité dans le plasma, la salive et le LCS (Devic et al., 2011; Besong-Agbo et al., 2013;
Mondello et al., 2014). De plus, il est possible d’observer des accumulations de la protéine
sur des biopsies du tractus gastro-intestinal de patients (Shin et al., 2017). Cependant, l’avenir
de cette protéine comme biomarqueur n’est guère prometteur puisque certaines études
montrent une augmentation de la concentration alors que d’autres montrent une diminution
de sa concentration, principalement dans le plasma (Lee et al., 2006; Li et al., 2007; Chahine
et al., 2014).

21
Figure 1.1. Principales catégories de biomarqueurs pour la maladie de Parkinson
Un seul biomarqueur ne peut confirmer la progression d’une maladie aussi complexe. La
combinaison des études en génétique, en imagerie, en clinique et en biochimie est nécessaire
pour connaître et comprendre l’évolution de la MP. Figure originale, Jérôme Lamontagne-
Proulx.

22
1.1.5.6 L’avenir des biomarqueurs
Dans beaucoup de maladies, notamment la MP, la recherche de biomarqueurs est alimentée
par la progression de nos connaissances biologiques (tissues, cellules, molécules) reliées à la
pathologie. Dans la MP, l’étude des tissus cérébraux post-mortem a été la clé dans
l’identification des sentiers protéiques et des gènes permettant l’avancée des thérapies, le
développement de modèle d’étude et la compréhension du mécanisme des médicaments, etc.
Grâce à ces stratégies, le domaine des biomarqueurs a grandement évolué, augmentant les
cibles potentielles (Yadav et al., 2012; Garbayo et al., 2013; Valera and Masliah, 2013). Les
nouvelles stratégies s’orientent davantage sur les méthodes globales non ciblées comme la
génomique et la protéomique.
Finalement, l’effort des prochaines années devrait être orienté à combiner les différentes
découvertes sur les biomarqueurs avec l’analyse des grandes banques de données déjà
existante, l’analyse d’échantillons de tissus et de fluides archivés de grandes études et la
collecte d’échantillons répétés sur de longues périodes. Il est crucial d’utiliser la bio-
informatique pour combiner ces approches et monter des modèles de progression et de
diagnostic.
1.2 Biomarqueur sanguin
Bien que la recherche explore toutes les directions possibles pour trouver rapidement un
biomarqueur robuste, nous pouvons penser qu’un outil de détection au niveau du sang serait
idéal. En effet, un biomarqueur sanguin serait facilement accessible, peu invasif et peu
coûteux (Chahine et al., 2014). Les études sur le sang se sont penchées majoritairement vers
l’α-Syn, encore une fois pour son importance dans la maladie, mais également sur DJ-1, pour
son potentiel comme biomarqueur diagnostique dans le LCS, et l’acide urique, pour son
habileté à diminuer l’oxydation de la DA (Church and Ward, 1994; Polymeropoulos et al.,
1997; Hong et al., 2010). C’est d’ailleurs l’acide urique qui montre le meilleur potentiel de
biomarqueur puisque des études ont révélé une diminution de ce composé dans le plasma de
patients pouvant indiquer une croissance des risques de développer la MP (Cipriani et al.,
2010; Schwarzschild et al., 2011). Dans la recherche de cible aléatoire, ApoA1 et l’epidermal

23
growth factor semblent diminués dans le plasma de patient démontrant une augmentation des
risques pour la MP (Chen-Plotkin et al., 2011; Qiang et al., 2013).
On retrouve également pour chaque type de cellules sanguines, plusieurs études visant à
découvrir si l’une ou l’autre de ces cellules est affectée par la pathologie et si elles peuvent
être utilisées comme biomarqueur.
1.2.1 Érythrocytes
En 2007, Nakai et collaborateurs ont montré la présence d’α-Syn dans les érythroblastes, les
réticulocytes et les érythrocytes (Nakai et al., 2007). De plus, certains chercheurs soutiennent
que les érythrocytes constituent la principale source d’α-Syn dans le sang (Barbour et al.,
2008). Finalement, une étude récente rapporte que la concentration d’oligomères d’α-Syn
présent dans les érythrocytes est plus élevé chez les patients parkinsoniens (Wang et al.,
2015). Pour l’instant, aucune étude ne permet de comprendre l’implication de cette protéine
dans le système hématopoïétique.
Une augmentation de l’éryptose, une sorte de mort cellulaire programmée propre aux
érythrocytes, a également été remarquée chez les patients souffrant de la MP (Pretorius et al.,
2014). En effet, l’augmentation de la peroxydation lipidique des érythrocytes de patients
parkinsoniens observés par Sudha expliquerait la hausse de mort érythrocytaire (Sudha et al.,
2003). Ce phénomène pourrait devenir un marqueur de progression de la maladie puisque le
stress oxydatif, produit par la peroxydation, est une caractéristique importante dans la
pathogenèse de la MP (Nikam et al., 2009). Finalement, on remarque que les molécules
antioxydantes telles que la catalase, la glutathion peroxydase et la superoxyde dismutase
(SOD), responsables de la détoxification des radicaux hydroxyles, sont diminuées dans les
érythrocytes de patients, marquant, encore une fois, l’altération de ces cellules avec la
maladie (Ihara et al., 1999; Serra et al., 2001; Chen et al., 2009; Abraham et al., 2005).

24
1.2.2 Plaquettes
Les plaquettes sont largement utilisées pour la détection de changement structural,
biochimique et moléculaire dans un très grand nombre de maladies (Camacho and Dimsdale,
2000; Pretorius et al., 2008; Boilard et al., 2010). De plus, des études valident leur importance
dans certaines maladies neurodégénératives comme la MP (Shrivastava and Vivekanandhan,
2011; Gowert et al., 2014). Il a été montré que le 1-Methyl-4-Phenylpyridinium (MPP+),
utilisé comme neurotoxine pour simuler la MP chez certains animaux, peut modifier
l’ultrastructure et la morphologie des plaquettes (Factor et al., 1994). En effet, une diminution
de l’agrégation des plaquettes, une déplétion en ATP, une augmentation de l’activité de la
monoamine oxydase B, une diminution de l’activité des chaînes de transport d’électrons et
une diminution de SOD ont été observées chez des parkinsoniens (Schapira et al., 1998;
Husain et al., 2009). Une augmentation du taux de glutamate et une diminution de son
absorption dans les plaquettes ont d’ailleurs déjà été remarquées chez des patients
idiopathiques (Ferrarese et al., 1999, 2001). Des évidences suggèrent même l’utilisation des
plaquettes pour l’étude de marqueurs de méthylation et de l’α-Syn dans la MP (S-adenosyl
methionine, S-adenosyl homocysteine) (Li et al., 2002a; Obeid et al., 2009).
1.2.3 Leucocytes
Tout comme les érythrocytes et les plaquettes, plusieurs évidences suggèrent que les
leucocytes de patients parkinsoniens pourraient subir des altérations biochimiques. Il a
d’ailleurs été montré que les lymphocytes de personnes malades sont susceptibles d’être la
cible d’une mauvaise régulation des protéines du cytosquelette et d’une augmentation du
stress oxydatif (Mila et al., 2009). De plus, certaines populations de lymphocytes T semblent
modulées en périphérie. Un phénomène qui pourrait être lié aux processus inflammatoires
dans le cerveau des patients (Baba et al., 2005; Rocha et al., 2017). Finalement, des études
de cas précoces de la MP montrent une réduction des concentrations de DA au niveau
intracellulaire et d’immunoréactivité de la tyrosine hydroxylase dans les leucocytes (Caronti
et al., 1999). Les monocytes pourraient eux aussi être altérés au niveau de leur fonction et de
leurs compositions de par la prédisposition inflammatoire chez les patients (Grozdanov et al.,
2014).

25
Bien que l’implication des cellules sanguines dans la MP soit encore peu comprise, il semble
qu’elles pourraient apporter à notre compréhension de la pathologie. On peut donc penser et
suggérer que l’excrétion de particules issues d’érythrocytes, de plaquettes et de leucocytes
pourrait refléter le dégrée d’activité pathologique.
1.3 Vésicules extracellulaires
La communication intracellulaire est un mécanisme essentiel des organismes
multicellulaires. Elle peut être modulée directement par le contact de cellule à cellule ou par
le transfert de molécules sécrétées. Au cours des deux dernières décennies, un troisième
mécanisme de communication intercellulaire impliquant le transfert de vésicules entre
cellules a été découvert. Bien que l’espace extracellulaire des organismes multicellulaires
contienne des ions, des protéines et des polysaccarides, on retrouve également un très grand
nombre de vésicules mobiles appelées, vésicules extracellulaires (VE). La majorité des
cellules du corps humain ont la capacité de relâcher une variété de vésicules membranaires
dans leur environnement extracellulaire. Les VE fonctionnent comme des navettes en
transportant des protéines, des ARN messager et des micro-ARN de cellules en cellules.
1.3.1 Exosomes
Dans les années 80, les exosomes ont été décrits par Trams et al. comme étant des vésicules
exfoliées pouvant servir à des fonctions physiologiques grâce à l’activité d’ectoenzyme
(Trams et al., 1981). Par la suite, des études ont décrit et montré, par microscopie
électronique, le relâchement de vésicules lors de la maturation des réticulocytes chez le rat
et le mouton (Harding et al., 1983; Pan et al., 1985; Johnstone et al., 1987). Ces vésicules
formées d’une bicouche lipidique ont une taille similaire à celle des virus, soit de ~50 à
100 nm. Les exosomes sont relâchés par exocytose depuis les corps multivésiculaires (CMV)
par induction ou de manière constitutive (Simons and Raposo, 2009). Les CMV sont la clé
intermédiaire dans le transport endolysosomal, formé lui-même de l’invagination et la
scission de vésicules (Gruenberg and Stenmark, 2004; Piper and Katzmann, 2007; Hurley et
al., 2010). Bien qu’il subsiste encore plusieurs interrogations quant à leur genèse, on sait que

26
le endosomal sorting complex required for transport et certains sphingolipides comme le
céramide et la sphingosine -1-phosphate contribuerait au relâchement des exosomes. En
effet, le céramide favorise la flexion membranaire alors que la sphingosine -1-phosphate
régule la production constitutive d’exosomes dans le CMV (Trajkovic et al., 2008; Hurley et
al., 2010). Les exosomes ont été principalement décrits pour leurs fonctions chez les cellules
immunitaires et les tumeurs. Leurs mécanismes permettent 1) le contact direct entre leurs
molécules de surface et les cellules, 2) l’endocytose des vésicules et 3) la fusion des
membranes de l’exosome et de la cellule (Théry et al., 2009). Les exosomes peuvent
également moduler le transfert d’ARN messager, de micro ARN et de particules du virus de
l’immunodéficience humaine (Valadi et al., 2007; Izquierdo-Useros et al., 2009). On retrouve
sur la surface membranaire externe la phosphatidylsérine ainsi que des marqueurs comme
CD63, CD81, CD9, Alix et TSG101 (Sarko and McKinney, 2017; Zhao et al., 2017).
1.3.2 Microvésicules
Chargaff et West ont été les premiers à décrire les microvésicules en 1946 comme étant un
précipité ayant la possibilité de générer de la thrombine dans du plasma sans plaquettes
(Chargaff and West, 1946). Tout comme les exosomes, les microvésicules ou ectosomes sont
formées d’une bicouche lipidique. Par contre, leurs tailles se situent entre 100 et 1000 nm,
selon le type cellulaire d’où elles émergent (György et al., 2011). La formation des
microvésicules résulte d’une interaction dynamique entre les phospholipides et les protéines
du cytosquelette. La répartition des flippases et des floppases est déterminante pour le
transfert de phospholipides ; étape initiale du bourgeonnement (Akers et al., 2013). De plus,
les interactions d’actine-myosine vont mener à la contraction du cytosquelette dans le but de
finaliser le bourgeonnement (Cocucci et al., 2009). Étant enrichies d’une multitude de
protéines et de lipides provenant de la cellule mère, les fonctions des microvésicules peuvent
énormément varier. Elles peuvent participer, par exemple, à l’activité procoagulante (Leroyer
et al., 2008), à la sécrétion d’IL1beta (Boilard et al., 2010), au caractère pro-invasif des
tumeurs (Giusti et al., 2008) et à l’induction de la transformation oncogénique cellulaire
(Antonyak et al., 2011). On retrouve majoritairement à leur surface des protéines

27
membranaires issues de la cellule mère et bien souvent la phosphatidylsérine (Connor et al.,
2010).
1.3.3 Corps apoptotiques
Les corps apoptotiques possèdent un diamètre entre 1 et 5 μm (Hristov et al., 2004). Ces
vésicules plasmatiques composées de débris cellulaires sont relâchées lors du processus
apoptotique d’une cellule. Elles peuvent par ailleurs contenir des fragments du noyau et des
organelles plus ou moins intactes. Ces corps sont destinés à être phagocytés, entre autres par
les macrophages (Elmore, 2007). Le but de ces vésicules est de permettre à la cellule
apoptotique de mourir et d’être phagocytée sans déclencher de réponse inflammatoire
potentiellement nuisible pour ses voisines. Bien que le corps apoptotique possède une taille
et une quantité de matériel beaucoup plus importantes que les autres VE, le mécanisme de
suppression par les macrophages, suite au changement spécifique de la membrane, reste
similaire (Erwig and Henson, 2008). On observe, de façon similaire aux exosomes et aux
microvésicules, une translocation de la phosphatidylsérine qui se liera à l’Annexin V,
reconnu par les phagocytes (Martínez and Freyssinet, 2001).
Bien que les différences entre les exosomes et les microvésicules soient détaillées dans les
revues de littérature, les articles de recherche utilisent généralement le terme « exosomes »
ou « VE ». Les méthodes utilisées pour différencier ces deux types de vésicules ne sont
souvent pas utilisées ou pas assez précises pour les catégoriser (Maas et al., 2017).

28
Figure 1.2. Représentation schématique des vésicules extracellulaires.
Les VE sont représentés de sorte que l’on puisse comparer leur taille et leur mode de
relâchement. Figure tirée de (György et al., 2011).

29
Tableau 1.2. Comparaison des types de vésicules extracellulaires
Exosomes Microvésicules Corps apoptotiques Taille 50-100 nm 100-1000 nm 1-5 μm Mécanisme de production
Exocytose du CMV Bourgeonnement de la membrane plasmique
Formation de bulles lors de l’apoptose
Isolation • Ultracentrifugation à gradient
• Centrifugation différentielle 100 000g
• Centrifugation différentielle 20 000g
• Co-culture avec cellule
Détection • Nanosizer • Microscopie électronique • Immunobuvardage de type
Western • Spectrométrie de masse
• Nanosizer • Microscopie électronique • Cytométrie de flux
• Microscopie électronique
• Cytométrie de flux
Marqueurs Annexin V, CD63, CD81, CD9, Alix, TSG101
Annexin V, marqueurs spécifiques à la cellule
Annexin V, marqueurs spécifiques à la cellule, marqueurs nucléiques
Références (Sarko and McKinney, 2017; Barile and Vassalli, 2017)
(Porro et al., 2015; Lai et al., 2015)
(Hristov et al., 2004; György et al., 2011)
1.3.4 Rôles des VE dans la maladie de Parkinson
Il a été proposé que les VE provenant des neurones, des oligodendrocytes, des astrocytes et
des microglies puissent jouer des rôles majeurs dans la pathogenèse de maladies

30
neurodégénératives grâce à leurs diverses fonctions (Budnik et al., 2016). Ces fonctions
pourraient notamment permettre aux VE de contribuer à la propagation de protéines
pathologiques au niveau du cerveau et ainsi accentuer la progression de la maladie (Quek
and Hill, 2017).
1.3.4.1 Études des VE dans le système nerveux central (in vitro)
En raison de la nature de la MP, des mécanismes de propagation et de neurotoxicité ont
d’abord été étudiés in vitro sur des neuroblastomes (SH-SY5Y) ou des neurogliomes (H4).
Ces études démontrent que l’α-Syn pourrait être excrétée des cellules neuronales par les
exosomes via un processus dépendant du calcium, suggérant que ces vésicules seraient
impliquées dans le transport de l’α-Syn et la propagation de la pathologie (Emmanouilidou
et al., 2010). De même, il semble que le dysfonctionnement du lysosome dans le cerveau de
patients augmente la libération d’α-Syn via les exosomes et, par conséquent, accentue
l’agrégation et la transmission de la protéine à des cellules voisines (Alvarez-Erviti et al.,
2011). En effet, les exosomes permettent l’accumulation d’α-Syn dans l’espace
extracellulaire, ce qui permettrait d’accélérer l’agrégation de la protéine (Melachroinou et
al., 2013; Grey et al., 2015). Il a d’ailleurs été proposé que la libération des oligomères d’α-
Syn associée aux exosomes soit un mécanisme par lequel les cellules peuvent rejeter la
protéine toxique (Danzer et al., 2012). Finalement, on note que la mutation ATP13A2 sur le
gène Park9, impliqué dans le parkinsonisme familial et juvénile, semble réduire les niveaux
d’α-Syn intracellulaires en augmentant le relargage de la protéine par les exosomes (Kong et
al., 2014; Tsunemi et al., 2014).
1.3.4.2 Études des VE dans le liquide céphalo-rachidien
Des études montrent que les VE du LCS contiennent également l’α-Syn et ses formes
pathologiques (Kunadt et al., 2015). L’analyse de cette protéine dans des exosomes du LCS
chez des patients souffrant de neurodégénérescence révèle que ce type de vésicules a la
capacité de produire des structures en forme d’oligomère à partir de la protéine soluble
(Stuendl et al., 2016). En outre, la protéine LRRK2, identifiée dans de nombreuses formes
de parkinsonisme familial, est retrouvée dans les exosomes du LCS (Fraser et al., 2013).

31
Cependant, l’échantillonnage systématique de LCS présente un très grand défi, soit celui
d’être extrêmement invasif.
1.3.4.3 Études des VE dans l’urine
L’urine est également étudiée comme réservoir de biomarqueurs grâce à son rôle d’excrétion
des déchets et du fait qu’elle est facilement récupérable. La présence de diverses formes de
LRRK2 à l’intérieur d’exosomes provenant des cellules endothéliales du rein a d’ailleurs déjà
été rapportée. Cependant, l’expression de cette protéine dans les vésicules urinaire semble
variable dans une même population ce qui sème un doute quant à son utilité comme
biomarqueur de la MP (Fraser et al., 2013). On retrouve également la protéine DJ-1 dans les
exosomes urinaires, augmentée près de deux fois chez les patients parkinsoniens (Ho et al.,
2014). Bien que l’urine pourrait potentiellement contenir un bon marqueur de la progression
ou de la détection de la MP, il existe encore une grande lacune quant à nos connaissances sur
les vésicules de ce liquide corporel.
1.3.4.4 Études des VE dans le système circulatoire
Le plasma est un fluide facile d’accès qui peut être prélevé fréquemment. Il s’agit donc d’un
environnement propice aux études de biomarqueurs. Tout d’abord, il a été montré que les
oligomères d’α-Syn présents dans le plasma offrent une nouvelle cible pour la création de
tests de diagnostic pour les patients parkinsoniens (El-Agnaf et al., 2006). L’identification
des mécanismes de transport et de sécrétion de la protéine chez les patients montre une
augmentation de sa concentration dans les exosomes dérivées du système nerveux central
localisé dans le plasma (Shi et al., 2014). Toutefois, il a également été montré que la
concentration d’α-Syn n’est pas modulée dans les vésicules de plasma, mais que 23 protéines
associées aux exosomes sont plus abondants chez les patients (Tomlinson et al., 2015). Ces
résultats appuient l’hypothèse que l’α-Syn ne peut pas être utilisé comme biomarqueur, mais
qu’un changement de plusieurs protéines dans les vésicules pourrait servir d’outil de
diagnostic.

32
1.4 Les objectifs de recherche
L’étude des biomarqueurs dans la MP connait une expansion considérable ces dernières
années. Considérant que le diagnostic est exclusivement clinique et apparaît très tard durant
les processus dégénératifs, il est nécessaire de développer des outils permettant de cibler les
patients à un stade précoce afin de stopper la pathologie rapidement. L’objectif global de
l’étude était donc d’identifier un biomarqueur non invasif permettant 1) de discriminer un
patient d’une personne saine et 2) d’instaurer une échelle de progression au niveau biologique
pouvant appuyer les échelles cliniques. Dans le but de réaliser ce projet, nous avons quantifié
et caractérisé le protéome des vésicules extracellulaires présentes dans le plasma. Bien que
plusieurs études s’attardent sur les cellules sanguines et les VE de fluides corporels comme
potentiel biomarqueur, à notre connaissance, les vésicules plasmatiques n’ont jamais été
caractérisées dans le cadre de la MP.
Nous avons utilisé un cytomètre de flux conçu spécialement pour détecter les vésicules
(Rousseau et al., 2015). Comme les cellules sanguines semblent affectées par la pathologie,
nous pensons que la quantité de vésicules pourrait être augmentée ou diminuée, dû à
l’altération de processus biologique dans ces cellules. Nous avons donc émis une première
hypothèse suggérant que la concentration de vésicules peut non seulement varier de manière
significative entre un patient et son contrôle, mais également entre des patients de différents
stades de la maladie.
Nous avons également caractérisé les vésicules d’intérêts au niveau protéique. Bien que
plusieurs études tendent à démontrer la présence et l’importance de l’α-Syn dans les vésicules
et les cellules sanguines, la littérature n’est toujours pas en mesure de désigner cette protéine
comme biomarqueur (Bartels et al., 2011; Grey et al., 2015). De plus, plusieurs études ayant
déjà fait une analyse protéique globale laissent présager qu’une multitude de facteurs sont
capables d’influencer l’ensemble des protéines des vésicules (Bosman et al., 2008, 2012;
Tomlinson et al., 2015). De ce fait, nous avons poursuivi une deuxième hypothèse de
recherche voulant que la pathologie puisse affecter la composition protéique des VE sans
toutefois changer les concentrations d’α-Syn et ainsi identifier un profil spécifique à la MP.

33

34
CHAPITRE 2: ERYTHROCYTE-DERIVED
EXTRACELLULAR VESICLES: A NOVEL, ROBUST AND
SPECIFIC BIOMARKER THAT MAPS TO PARKINSON'S
DISEASE STAGES

35
2.1 Résumé
Contexte La production de VE est une caractéristique omniprésente des cellules eucaryotes.
Cependant, des conditions pathologiques peuvent affecter leur formation et leur constitution.
Nous avons cherché à caractériser la nature, le profil et la signature protéique des VE dans le
plasma de deux cohortes indépendantes de patients atteints de la MP et corréler les résultats
avec des mesures cliniques de la maladie. Méthodologie Les VE ont été recueillis auprès de
grandes cohortes de patients souffrant de la MP (n = 60 ; Controls, n = 37) et de patients
souffrant de la maladie d’Huntington (Pré-manifeste, n = 11 ; manifeste, n = 52 ; Contrôles,
n = 55) - pour un total de 215 participants. – Ils ont été analysés de manière exhaustive grâce
à la cytométrie de flux, la microscopie électronique et la protéomique. Résultats Nous avons
découvert qu’il existe une liaison de type affine entre le nombre de VE dérivés d’érythrocytes
(VEE) et l’échelle clinique, UPDRS. Nous avons observé que les patients atteints de la MP
au stade précoce — caractérisés par un score UPDRS inférieur à 37 — ont montré une
augmentation linéaire des VEE par rapport à l’état pathologique (corrélations = 0,82), avec
un profil presque identique chez les patients modérés avec des scores UPDRS compris entre
37 et 75 (corrélations = 0,873). Ces résultats sont obtenus grâce à un ajustement pour
certaines comorbidités. Nous n’avons pas observé ces résultats chez les patients atteints de
la maladie d’Huntington, ce qui met en évidence la spécificité de nos résultats vis-à-vis la
pathologie de la MP. D’autres analyses protéomiques des VEE ont permis d’identifiés 8
protéines dont l’expression a été significativement modifiée chez les patients atteints de la
MP aux stades précoce et modéré. Conclusion Nous avons découvert un nouveau
biomarqueur qui différencie les étapes de la MP en fonction de la quantification des VEE et
de leur cargaison unique de protéines.

36
Erythrocyte-derived extracellular vesicles:
A novel, robust and specific biomarker that maps to Parkinson's disease
stages
Jérôme Lamontagne-Proulx, BSc1, Isabelle St-Amour, PhD1, Richard Labib, PhD2, Jérémie
Pilon, BSc2, Nathalie Cloutier, PhD1, Florence Roux-Dalvai, MSc1, Antony T. Vincent,
MSc3, Sarah L Mason, PhD4, Anne-Claire Duchez, PhD1, Arnaud Droit, PhD1,5, Steve
Lacroix, PhD1,5, Nicolas Dupré, MD1,6, Mélanie Langlois, MD1,6, Sylvain Chouinard, MD7,
Roger A Barker, MD, PhD4, Eric Boilard, PhD1,8*, Francesca Cicchetti, PhD1,9*
1Centre de recherche du CHU de Québec, Québec, QC, Canada; 2Département de mathématiques et
génie industriel, École Polytechnique de Montréal, Montréal, QC, Canada; 3Institut de Biologie
Integrative et des Systemes, Universite Laval, Québec, QC, Canada; 4Department of Clinical
Neurosciences, John van Geest Centre for Brain Repair, University of Cambridge, Cambridge,
United Kingdom; 5Département de médecine moléculaire, Université Laval, Québec, QC, Canada;
6Département de médecine, Université Laval, Québec, QC, Canada; Département de neurosciences,
Hôpital de l'Enfant-Jésus, Québec, QC, Canada; 7Centre Hospitalier de l'Université de Montréal and
Centre de recherche du Centre Hospitalier de l'Université de Montréal, Hôpital Notre-Dame,
Département de médicine, Université de Montréal, Montréal, QC, Canada; 8Département de
microbiologie-infectiologie et d'immunologie, Université Laval, Québec, QC, Canada; 9Département de psychiatrie & neurosciences, Université Laval, Québec, QC, Canada
Correspondence to either:
Francesca Cicchetti, Ph.D.
Centre de Recherche du CHU de Québec
Axe Neurosciences, T2-07
2705, Boulevard Laurier
Québec, QC, G1V 4G2, Canada
Tel #: (418) 656-4141 ext. 48853
Fax #: (418) 654-2753
E-mail: [email protected]
Eric Boilard, Ph.D.
Centre de Recherche du CHU de Québec
Infectious and immune diseases, T1-49
2705, Boulevard Laurier
Québec, QC, G1V 4G2, Canada
Tel #: (418) 656-4141 ext. 46175
Fax #: (418) 654-2765
E-mail: [email protected]

37
2.2 Abstract
Background The production of extracellular vesicles (EV) is a ubiquitous feature of
eukaryotic cells, but pathological events can affect their formation and constituents. We
sought to characterize the nature, profile and protein signature of EV in the plasma of two
independent cohorts of Parkinson’s disease (PD) patients and how they correlate to clinical
measures of the disease. Methods EVs were collected from large cohorts of PD (n=60;
Controls, n=37) and Huntington’s disease (HD) patients (Pre-manifest, n=11; manifest,
n=52; Controls, n=55) - for a total of 215 participants - and exhaustively analyzed using flow
cytometry, electron microscopy and proteomics. Results We discovered that the number of
EV derived from erythrocytes (EEV) was strongly correlated to two different disease stages
as defined using the Unified Parkinson Disease Rating Scale (UPDRS). In particular, we
observed that mild PD patients – characterized by a UPDRS score lower than 37 – showed a
linear increase of EV with respect to disease state (correlations = 0.82), with a near identical
pattern in moderate patients with UPDRS scores ranging between 37 and 75 (correlations =
0.873). We did not observe this in patients with HD, highlighting the specificity of our
findings to PD process. Further proteomic analyses of the EV identified 8 proteins whose
expression was significantly altered in both mild and moderate PD patients. Conclusion We
have discovered a novel biomarker that maps onto stages of PD based on the quantification
of EV shed from erythrocytes and their unique protein cargo.

38
2.3 Introduction
There is no doubt that over the last few years, there has been a dramatic increase in new
findings that have fundamentally transformed our understanding of neurodegenerative
diseases, in particular of Parkinson’s disease (PD). One of these has been the heterogeneity
of the disease that has hindered the development of useful biomarkers, as well as the
successful translation of novel treatments. This new understanding of the different clinical
profiles seen in populations of PD patients has begun to translate into a redefinition of PD,
as is evidenced by new diagnostic criteria released by the International PD and Movement
Disorder Society (MDS) (Berg et al., 2014). This task force has also highlighted the need to
identify reliable biomarkers that can help in the diagnosis, stratification and tracking of this
condition.
In a novel approach to develop such biomarkers, we exhaustively studied circulating
extracellular vesicles (EV), known to mediate cell-to-cell communication in both
physiological and disease conditions (Quek and Hill, 2017). Their rich protein cargo, cell
type signature, and availability in bodily fluids such as blood plasma make them very
attractive biomarkers (Porro et al., 2015). We therefore characterized the nature, protein
signature and profile of EV in the plasma of two independent cohorts of PD patients and
assessed their behaviour in relation to disease states. Among the most important clinical
challenges is the assessment of drug efficacy, and more specifically in complex
neurodegenerative diseases with heterogeneous populations of patients. In PD, for example,
some patients evolve to an early dementia while others remain cognitively normal throughout
their illness and although we have some clinical and underlying genetic clues as to why this
may happen, we have no way of accurately predicting or following the course of these
different subtypes of PD. This is critical if we are to bring specific targeted therapeutics to
the clinic to treat different aspects of the disease, especially therapies that are designed to be
disease modifying which ideally should be targeted to the more rapidly progressive forms of
PD.

39
2.4 Materials and methods
2.4.1 Ethics statement and participant recruitment
Institutional review boards approved this study (CHU de Québec, #A13-2-1096; CHUM,
#14.228; Cambridge Central Regional Ethics Committee, REC #03/303 & #08/H0306/26;
and Cambridge University Hospitals Foundation Trust Research and Development
department, R&D #A085170 & #A091246) in accordance with the Declaration of Helsinki,
and written informed consents were obtained from all participants.
Blood samples were collected from 2 cohorts of patients with PD [one in Cambridge UK;
one in Quebec Canada] as well as an independent cohort of Huntington’s disease (HD)
individuals [collected in Montreal, Canada] along with age- and sex-matched healthy
Controls (Table 2.1). In the case of PD patients, the UK PDS Brain diagnostic criteria were
used which gives a diagnostic accuracy of 98.6% when applied by movement disorder
specialists (Massano and Bhatia, 2012; Hughes et al., 2002). Their clinical evaluation
included the Unified Parkinson Disease Rating Scale (UPDRS), Hoehn and Yahr (H&Y)
staging, the Mini Mental State Examination (MMSE), the Addenbrooke's Cognitive
Examination (ACE) and the Beck Depression Inventory (BDI). In the case of the HD patients,
their Unified Huntington Disease Rating Scale (UHDRS), Total Functional capacity (TFC)
and calculated values for burden of disease (BDS) were all collected and diagnosis was
confirmed by genetic testing. All clinical evaluations were conducted within 6 months of the
blood collection. Participants were further asked to fill out a questionnaire related to health
issues and medication and a full blood count was performed in all patients on the day of blood
sampling. Medications were converted into levodopa equivalent daily dose (LEDD) using
common calculator tools. It should be noted that blood sampling was conducted by the exact
same team of investigators, following identical procedures in both UK and Canada.
2.4.2 Preparation of platelet-free plasma and EV labeling
Citrated blood was centrifuged twice for 15 minutes at 2500g at room temperature. Platelet-
free plasma (PFP) was harvested and stored at -80°C within 2 hours of sampling following

40
previously published guidelines (Lacroix et al., 2012). For specific details on EV labeling,
please refer to supplementary material.
In all cases, the control samples came from healthy participants and the blood was collected
at exactly the same time as the PD and HD patients in the UK and Canadian clinics. Every
blood sample was processed immediately to avoid release of non-physiological EV. For
analyses relating to EV in the plasma, all blood samples were centrifuged twice at 2500g to
enable PFP recovery. The plasma was fractionated into 3 aliquots per individual and frozen
immediately following processing (maximum of 2 hours following blood collection). Blood
samples from Controls and PD were collected in parallel and laboratory analyses were
blinded to participant status.
2.4.3 Flow cytometry quantification
For EV quantification, we used a FACS Canto II Special Order Research Product equipped
with a forward scatter (FSC) coupled to a photomultiplier tube (FSC-PMT) and a small
particle option. Flow cytometer performance tracking was carried out daily using the BD
cytometer setup and tracking beads (BD Biosciences, San Jose, CA, USA). The size of the
EV was determined using fluorescent silica beads of 100, 500 and 1000nm (figure S2.1).
The settings for the EV detection were determined as previously described using a threshold
of 200 for SSC (also see figure S2.1) (Rousseau et al., 2015). For specific details plasma-
related EV quantification, please refer to supplementary material.
2.4.4 Production and purification of EEV
Blood was collected in heparin tubes and centrifuged for 10 minutes at 282g at room
temperature. Blood cells were washed first in PBS-2%FBS, then with 0.9% sodium chloride
solution and centrifuged for 10 minutes at 750g. To avoid leukocyte and/or platelet
contamination, the buffy coat and upper fraction of erythrocytes were removed. To preserve
erythrocytes, two volumes of glycerolyte 57 solution (57% glycerol, 142mM sodium lactate,
1mM KCl, 25mM sodium phosphate pH 6.8) were added to the pellet and stored at -80°C.
For the production of EEV, red bloods cells were thawed and EV production was induced as

41
previously described (Minetti et al., 2004). For specific details on EEV purification, please
refer to supplementary material.
2.4.5 C-reactive protein, free hemoglobin and α-synuclein quantification
The concentrations of C-reactive protein (CRP) and free hemoglobin were determined in the
PFP of all donors using the RayBio Human CRP ELISA Kit (RayBiotech, Norcross, GA,
USA) and the Hemoglobin Human ELISA kit (Abcam, Toronto, ON, Canada). To quantify
α-synuclein (α-Syn) in erythrocytes and EEV, we used the human α-Syn ELISA kit
(ThermoFisher Scientific, Waltham, MA, USA). Absorbance values were measured at
450nm using a multi-detection microplate reader (Synergy HT; BioTek; Winooski, VT,
USA). All ELISA tests were performed according to the manufacturer's instructions.
2.4.6 Scanning electron microscopy
For the visualization by scanning electron microscopy, EEV were prepared as previously
described (Duchez et al., 2015). For specific details on EEV preparation, please refer to
supplementary material.
2.4.7 Transmission electron microscopy
For the transmission electron microscopy, EEV were prepared and observed as previously
described (Duchez et al., 2015). For specific details on EEV preparation and labelling, please
refer to supplementary material.
2.4.8 Mass spectrometry analysis and label free protein quantification
The initial proteomic analyses were performed on 3 pools of 3 blood samples of each group.
Following this, we undertook a completely new set of analyses to develop a method allowing
us to isolate hemoglobin and better investigate the EEV proteome. For this, EVs from an
additional 4 individuals per group (Control, mild PD and moderate PD) were prepared as
described above. For each individual, 25μg of protein sample, according to Bradford protein

42
assay, were migrated onto an electrophoresis gel 4-12% Bis-Tris to separate hemoglobin
from higher proteins. Following gel staining using Sypro Ruby (Thermo Fischer Scientific),
the 12kDa band corresponding to the hemoglobin size was cut out and the remaining part of
the gel further fractioned into 7 slices, exposed to trypsin digestion and peptide extraction on
a MassPrep liquid handling robot (Waters, Milford, USA) according to the manufacturer’s
specifications and the protocol of Shevchenko et al. with the modifications suggested by
Havlis et al. (Shevchenko et al., 1996; Havlis et al., 2003). For specific details on protein
analysis purification, please refer to supplementary material. Spectra of peptides were
searched against a human protein database (Uniprot Complete Proteome, taxonomy Homo
sapiens – 83512 sequences) using the Andromeda search engine included in MaxQuant
software version 1.5.5.1 (Cox and Mann, 2008). MaxQuant was also used to validate proteins
and peptides at a 1% False Discovery Rate using a target/decoy database search as well as to
undertake a Label Free Quantification of the identified proteins using the ‘match between
runs’ option.
2.4.9 Statistical analyses
All statistical analyses were performed by a quakified statistician and details pertaining to
these analyses for each experiments are provided in the supplementary material.
2.5 Results
This study involved two cohorts of PD patients (n=60; n=36 Cambridge; n=24 Quebec) and
HD patients (n=63; Quebec) at all stages of diseases along with age- and sex-matched healthy
Controls (n=37 for PD; n=55 for HD) (Table 2.1) giving a total of 215 participants. First, we
evaluated the diagnostic value of EV for PD and found a notable increase in the mean number
of EEV in PD patients with respect to their age- and sex-matched healthy Controls (p=0.0319;
figure 2.1A). A closer look at the data distribution, however, revealed that only 5 patients
were responsible for this significant difference (figure 2.1A, red inset). Further investigation
using a diagnostic accuracy test, revealed that the relevant proportion of the area under the
Receiver Operating Characteristic (ROC) curve was 0.508 (data not shown), implying that

43
the number of EEV could not solely be used as a discriminant between PD patients and
healthy Controls. This was also confirmed by performing a Bayesian analysis of the data
demonstrating that the proposed predictor of the onset of PD would be correct in less than
1% of the cases (data not shown). We obtained similar result when we separated the patients
into mild and moderate states according to their UPDRS scores (data not shown).
We next evaluated the potential of EEV to map onto different PD disease stages. Analyses
revealed striking correlations between the number of vesicles derived from the red blood
cells of PD patients and their total UPDRS (figure 2.1B). Statistical linear regression analysis
was performed for each group and the R2 values obtained demonstrated that in both cases, at
least 87% of the variation in the total number of EEV/erythrocytes was due to the variation
of the UPDRS (figure 2.1B). Moreover, the results are significant with respect to the p values
obtained for each fit, since they fall below the 5% confidence level. Hence, the statistical
tests on EEV counts uncovered a clear cut-off point between mild and moderate patients,
which could not be accounted for by medication, as there was no correlation with the patients
LEDD (figure S2.2A-B). Additional analyses on the number of EEV in relation to other
clinical measures including H&Y stage, BDI, MMSE and ACE found no significant
correlations (data not shown).
In the HD cohort, the total number of EEV between pre-manifest and manifest HD was
similar to that seen in their age- and sex-matched healthy Controls (figure 2.1C). In contrast
to PD, correlation analyses failed to reveal an association between the number of EEV and
HD stage using the UHDRS score (figure 2.1D). Based on this, all our subsequent analyses
focused only on the PD cohort. It should also be noted that total blood counts did not indicate
any differences in the number of endothelial cell-, platelet-, monocyte- and granulocyte-
derived EV in the platelet-free plasma (PFP) in PD (Table S2.1) and HD patients (Table
S2.2) when compared to their respective control cohorts. Additionally, at the start of the study
and as a precautionary measure, full blood count (figure S2.3A-C) and C-reactive protein
(figure S2.3D) quantifications were obtained for all participants which revealed no
differences between groups. We did, however, observe a significant increase in EEV
concentration in the PFP of individuals with diabetes and those suffering, or having suffered,

44
from cancer (data not shown) and thus these participants were excluded from our analyses.
PFP samples with elevated free hemoglobin (>45 000ng/ml), potentially due to hemolysis
during blood sampling, were also excluded from further EEV-related analyses (figure S2.3E)
which account for the small discrepancies between the total number of participants initially
recruited and those contained in each analysis.
Having established that EEV counts map to disease state in PD, we aimed to assess whether
α-Syn – which is not only the main component of Lewy bodies but is highly expressed in
most blood cells – was differentially expressed in normal vs. diseased conditions. For this,
we opted to use scanning electron microscopy, but this did not reveal any morphological
changes between resting and activated erythrocytes in either condition (figure 2.2A). We
further used transmission electron microscopy to quantify the number of EEV containing α-
Syn and phosphorylated (serine 129) forms of the protein but again no significant differences
between PD patients and age- and sex-matched healthy Controls (figure 2.2B-C) were
observed. Quantified α-Syn levels in EEV from PD patients and Controls using commercial
ELISA kits corroborated these results (figure 2.2D).
Since our combined quantification studies (ELISA and transmission electron microscopy)
suggested that α-Syn levels could not be used as a blood marker of disease, we sought to
obtain the specific protein signature of EEV from mild and moderate PD patients (with
respect to the UPDRS scores) and their age-matched Controls. Given the significant amounts
of hemoglobin within erythrocytes that could mask the true nature of the protein signature in
EEV, we performed a label free quantitative proteomic analysis by nanoLC/MSMS (Wither
et al., 2016) using two distinct approaches: with and without hemoglobin (figure S2.4A). By
removing hemoglobin, we identified a total of 818 proteins in comparison with 356 when we
did not undertake this methodological step (refer to Table S2.3 for complete list of proteins)
– a modification which is clearly needed to give a much more accurate evaluation of the
protein content of EEV. Additionally, a Gene Ontology enrichment analysis on the ‘Cellular
Component’ ontology on the two sets of identified proteins in comparison with the whole
human proteome, revealed that our samples are enriched with elements associated with
‘vesicles’ and ‘hemoglobin complex’ (figure S2.4B).

45
Out of the 818 proteins identified in the proteome of EEV, 8 had their expression significantly
modified according to the different stages of PD (figure 2.3A-B). Hierarchical clustering,
coupled to a heatmap, allowed us to group individuals according to stages of disease (Control,
mild PD and moderate PD) and provided compelling evidence that the 8 proteins identified
could also be grouped into three categories. Proteins of group I were highly and
predominantly expressed in Controls, proteins of group II were highly and predominantly
expressed in mild PD patients and proteins belonging to group III were highly and
predominantly expressed in moderate PD patients (figure 2.3C). This data set was further
confirmed by volcano plots (figure S2.5). Of interest, two proteins of group I are associated
with the regulation system of the cell (ABHD14B, NADSYN1) and one protein significantly
expressed in moderate PD patients (ATP5A1) is involved in the regulation of mitochondrial
ATP production.
2.6 Discussion
We have identified a novel biomarker that maps to different states of PD, using two cohorts
of patients and a total of 215 participants, based on the quantification of EV shed from
erythrocytes. In particular, we have shown a remarkable correlation between the number of
EEV and the clinical expression/stages of PD using the standard UPDRS measure. Finding
this level of correlation with such a heterogeneous disorder highlights the robustness of the
biomarker we have identified. This is also unique to PD as no such changes were seen in a
second neurodegenerative disorder of the brain, HD. As these EVs contain a unique protein
cargo, we surmise that the EVs and their content will help us understand the basis for the
different disease stages in PD, with the possibility that the disease pathogenesis changes at a
critical point in its clinical course.
It has previously been reported that EV derived from the cerebrospinal fluid contain α-Syn,
its associated pathogenic species as well as the LRRK2 protein (Kunadt et al., 2015; Fraser
et al., 2013). However, routinely collecting cerebrospinal fluid presents a challenge given its
invasive nature while plasma is a very accessible fluid that further allows real time

46
monitoring, and thus has been more extensively studied from a biomarker perspective.
Indeed, α-Syn expression has been reported in platelets, leukocytes and red blood cells, with
some studies suggesting that their levels correlate with disease duration (Behari and
Shrivastava, 2013; Fuchs et al., 2008; Kim et al., 2004; Li et al., 2002b; Nakai et al., 2007).
However, none of these studies have shown that blood cells, or EV derived from blood cells,
represent a robust biomarker of disease (see summary figure 2.4) (Nakai et al., 2007; Wang
et al., 2015; Barbour et al., 2008; Pretorius et al., 2014; Nikam et al., 2009; Sudha et al.,
2003; Bartels et al., 2011; Fauvet et al., 2012; Araki et al., 2016; Helferich et al., 2015;
Renella et al., 2014; Abd-Elhadi et al., 2015). Despite the fact that a number of groups have
reported the presence of EV in various bodily fluids of PD patients (figure 2.4) (Kunadt et
al., 2015; Fraser et al., 2013; Emmanouilidou et al., 2010; Danzer et al., 2012; Alvarez-Erviti
et al., 2011; Melachroinou et al., 2013; Grey et al., 2015; Kong et al., 2014; Tsunemi et al.,
2014; Shi et al., 2014; Tomlinson et al., 2015; El-Agnaf et al., 2006; Stuendl et al., 2016; Ho
et al., 2014; Zappulli et al., 2016), we provide clear evidence that EEV can serve as a
biomarker in PD.
Although staging of PD is often done using the H&Y clinical scale, we sought to use the
UPDRS given its greater sensitivity – an approach that has recently been validated (Martínez-
Martín et al., 2015). Using these scores, we found that mild PD patients – with a UPDRS
score lower than 37 – have an increased number of EV (correlations = 0.886); and that the
exact same pattern then repeats itself with patients who had UPDRS scores between 37 and
75 (correlations = 0.873) with a marked resetting of the biomarker between these two stages.
When comparing individual patients from both correlation curves, LEDD did not seem to
account for the differences, as there was no correlation between the LEDD and the number
of EEV. At this stage, it is unclear why there is this split in correlation around the UPDRS
score and EEV counts. Building on these cross-sectional findings, ongoing longitudinal
studies in larger patient cohorts, funded by the Weston Brain Institute, will allow to further
investigate this.
Another important aspect of the test we have developed is its potential to be easily adopted
into the clinic. Most of the tests currently developed for biomarker use in patients are time

47
consuming and require appreciable quantities of biological samples. Our biomarker employs
a user-friendly clinical test which has the potential to be easily automated using limited
volumes of blood (few microliters). Finally, while proteomic studies of EEV have been
previously reported (Bosman et al., 2008, 2012), their description was limited to 270 proteins
(Shi et al., 2014). We herein report on an improved method to perform this type of analyses
in blood samples by removing hemoglobin; a large protein that can easily mask other ones
within a protein signature. Indeed, the high dynamic range of protein concentrations in
erythrocytes and their EEV, due to the abundance of hemoglobin, decreases the capacity of
the mass spectrometer to detect signals corresponding to low abundance proteins. Analyzing
the hemoglobin, separately from the other proteins of other molecular weights allowed us to
go deeper into the EEV proteome, identifying 129% more proteins than in the initial analysis.
Additionally, it is important to reiterate that our proteomic analyses were performed using
very stringent parameters, which included a minimum of 2 peptides per protein and p values
lesser than 0.05 with an absolute z score of 1.96, corresponding to values outside of the 95%
confidence range. Finally, our volcano plots and heatmap analyses revealed high specificity
of the identified proteins for each group with nodes ranging between 80 and 90%. With this,
reproducibility of our proteomics data is high despite the smaller sample size (analyses
performed on 3 pools of 3 samples per group and subsequently on 4 individual samples per
group).
Taken together, the identification of this new EEV biomarker has the potential to 1) help in
the diagnosis of PD especially at a stage of disease when disease modifying therapies could
have their biggest impact, 2) lead to a useful marker that tracks disease course, 3) provide a
therapeutic target or marker of disease modification in the development and testing of novel
disease modifying therapies and 4) offer new insights into disease pathogenesis and how it
changes through the disease stages, which in turn has important therapeutic implications.
2.7 Acknowledgements
The study was funded by The Michael J. Fox foundation and the Parkinson Society Canada
to F.C. as main PI who is also a recipient of a National Researcher career award from the

48
Fonds de Recherche du Québec en santé providing salary support and operating funds. I.S.-
A. was supported by a CIHR-Huntington Society of Canada postdoctoral fellowship. R.B.
and J.P. are supported by the Natural Science and Engineering Research Council of Canada.
R.A.B. and S.L.M. are supported by a National Institute for Health Research award of a
Biomedical Research Center to the University of Cambridge and Addenbrooke’s Hospital.
A.-C.D. is supported by the Fonds de Recherche des maladies Rhumatismales de l’Université
Laval and the Canadian Arthritis Network. E.B. is supported by the Canadian Institutes of
Health Research. N.D. MD-MSc. also funded by CIHR and by Canadian Consortium on
Neurodegeneration in Aging.
The authors would like to thank all the students and staff who helped with the blood drives
in Quebec city and Montreal, Mr. Richard Janvier for his very skilful electron microscopy
preparation and analyses and the Bioimaging platform of the Infectious Disease Research
Centre, funded by an equipment and infrastructure grant from the Canadian Foundation for
Innovation and importantly, all patients and their families for being so generous with their
time in participating to this study.
Conflicts of Interest
F.C., E.B., S.L. and I.S.-A. have filed a patent on the use of EEV as a biomarker in PD.
J.L.P., R.L., J.P., N.C., F.R.-D., A.T.V., S.L.M., A-C.D., A.D., S.L., N.D., M.L., S.C. and
R.A.B. declare no conflicts of interest.
Table 2.1. Participant clinical information PD cohort
CTRL
PD Patients – Stages of disease
Unknown Mild Moderate Severe p value
n 37 7 12 33 8 Age 66.8 69.8 66.7 71.1 75.0* 0.04 Gender F (M) 18 (19) 1 (6) 6 (6) 16 (17) 0 (8) 0.05 Disease severity Hoehn & Yahr (n) 1 ± 0.3 (12) 2 ± 0.2 (33) 3 ± 0.5 (8) <0.0001

49
2.8 Tableau
Table 2.1. Participant clinical information. (PD cohort) Disease severity levels in relation to the H&Y scale (score): Mild (1-1.5); Moderate (2-2.5); Severe (3-3.5). *p < 0.05 vs. CTRL. Statistical analyses were performed using a Welch ANOVA followed by Dunnett's multiple comparison test. Disease severity was evaluated within 6 months of blood sampling. Comorbidities were determined from medical information reported by the participant or caregiver. Cancer refers to participant having suffered from cancer in the past. (HD cohort) Disease severity levels in relation to the TFC scale: Stage 1 (11-13); Stage 2 (7-10); Stage 3 (3-6); Stage 4 (1-2); Stage 5 (0). Disease severity was evaluated within 6 months of blood sampling. Comorbidities were determined from medical information reported by the participant or caregiver. Abbreviations: ACE, Addenbrooke's Cognitive Examination; BDI,
UPDRS (n) 38 ± 11 (6) 52 ± 19 (17) 73 ± 20 (6) 0.02 BDI (n) 3 ± 2 (6) 4 ± 2 (17) 13 ± 7 (4) 0.03 MMSE (n) 29 ± 2 (7) 29 ± 1 (19) 26 ± 3 (6) 0.01 ACE (n) 96 ± 4 (6) 92 ± 7 (17) 84 ± 14 (6) 0.13 Comorbidities Depression 3 1 2 1 2 0.29 Cancer 5 0 3 4 1 0.64 Diabetes 2 0 0 1 2 0.10 Hypertension 10 1 2 10 3 0.76 Hypercholesterolemia 5 0 1 6 1 0.73 Asthma 3 1 1 5 0 0.71 Allergies 2 0 2 6 2 0.28
HD cohort
CTRL
HD Patients – Stages of disease
Pre-HD Stage 1 Stage 2 Stage 3 Stage 4 Stage 5 p value
n 55 11 15 13 12 10 2 Age 55.0 37.5 53.1 54.2 58.3 58.1 55.5 0.02 Gender F (M) 31 (22) 6 (5) 5 (10) 4 (9) 8 (4) 7 (3) 1 (1) 0.26 Disease severity CAG (n) 28.3 (3) 41.1 (10) 42.3 (13) 42.6 (12) 43.7 (7) 44.3 (7) <0.001 UHDRS (n) 2.7 (11) 15.7 (14) 34.5 (11) 42.9 (12) 55.9 (10) 67.5 (2) <0.001 TFC (n) 13 (16) 13 (11) 12.5 (15) 7.8 (13) 4.3 (12) 1.6 (10) 0 (2) <0.001 BDS (n) 206 (10) 337 (13) 356 (12) 442 (7) 465 (7) <0.001 Comorbidities Depression 8 1 1 3 6 4 1 0.0497 Cancer 0 0 0 0 0 0 0 Diabetes 3 1 1 1 1 1 0 0.99 Hypertension 4 1 2 1 1 2 0 0.92 Hypercholesterolemia 8 1 1 0 0 1 0 0.32 Asthma 0 0 1 0 0 0 0 0.65 Allergies 3 0 2 2 0 0 0 0.33

50
Beck Depression Inventory; BDS, Burden of Disease Score; CTRL, Control; HD, Huntington’s disease; H&Y, Hoehn and Yahr; MMSE, Mini-Mental State Examination; PD, Parkinson’s disease; UHDRS, Unified Huntington’s Disease Rating Scale; UPDRS, Unified Parkinson’s Disease Rating Scale; TFC, Total Functional Capacity.

51
2.9 Figures
Figure 2.1. EEV: a robust biomarker of PD stage. A. Distribution plots of CD235a+ EV/total number of erythrocytes uncovered a significant group difference between PD and healthy sex- and age-matched CTRL (PD, n=42; CTRL, n=24) although only a small number of patients (n=5) accounted for this difference. B. Robust correlations between the number of EEV/total number of erythrocytes and UPDRS scores (PD, n=20, from the UK cohort exclusively since the Quebec cohort did not have recent UPDRS scores available), displayed a clear split between mild and moderate patients, supporting the potential of EEV to act as a biomarker for disease stage. C. Distribution plots of CD235a+/total number of erythrocytes

52
revealed no differences between HD and healthy sex- and age-matched CTRL (HD, n=50; Pre-HD, n=10; CTRL, n=52) and D. no statistically significant correlations were found between the number of EEV/total number of erythrocytes and UHDRS scores (HD, n=42). This argues in favor of EEV being a specific biomarker for PD. Distributions were determined using unpaired t-test with Welch’s correction (PD) or one-way ANOVA (HD). Correlations were determined using Pearson’s correlation, *p<0.05. Abbreviations: CD235a, glycophorin A; CTRL, Controls; EEV, erythrocyte-derived extracellular vesicle; EV, extracellular vesicle; HD, Huntington’s disease; PD, Parkinson’s disease; Pre-HD, Pre-manifest; UHDRS, Unified Huntington’s Disease Rating Scale; UPDRS, Unified Parkinson’s Disease Rating Scale.

53
Figure 2.2. Detection of normal and phosphorylated α-Syn in EEV. A. Representative scanning electron micrographs of resting and activated erythrocytes (treated with calcium ionophore A23187 to generate EEV) in both PD patients and healthy sex- and age-matched CTRL. Scale bar: 2μm. B. Representative transmission electron microscopy images of
immunogold labeling for α-Syn and α-Syn pS129 in activated erythrocytes and EEV (some examples delineated by dotted lines). Arrowheads point to positive immunolabeling for either
α-Syn or α-Syn pS129. Scale bar: 100nm. C. Quantification of α-Syn in EEV as detected by

54
transmission electron microscopy and expressed as the percentage of EEVs positive for α-Syn/total number of EEV in healthy sex- and age-matched CTRL and PD patients (n=100 erythrocytes sampled in n=3 CTRL and n=3 PD). D. Quantification of α-Syn in EEV by ELISA assay in healthy sex- and age-matched CTRL, mild and moderate stage patients (n=4 erythrocytes per group; n=13 EEV per group) revealing the absence of measurable changes in α-Syn levels between PD and healthy sex- and age-matched CTRL. Statistical analyses were performed using a Mann-Whitney U test (C) or a Kruskal-Wallis ANOVA (D).
Abbreviations: α-Syn, α-synuclein; α-Syn pS129, α-synuclein phosphorylated Serine 129; CTRL, Control; EEV, erythrocyte-derived extracellular vesicle; H&Y, Hoehn and Yahr; PD, Parkinson’s disease.

55

56
Figure 2.3. Specific protein signature of EEV in PD patients. A. NanoLC-MS/MS Label-free analysis of EEV in PD patients and healthy age-matched CTRL revealed a total of 818 proteins, of which 8 had an expression that was significantly modified as a function of PD state (B). These proteins have been categorized and referenced according to the gene to which they are associated (A – see table). B. The proteins/genes have been further separated into 3 groups in relation to their variations in comparison to CTRL (Group I), mild PD (Group II) or moderate PD (Group III). C. Heatmap establishing correlations between disease stages and the abundance of the variable proteins. Cold and hot colours represent low and high correlation levels, respectively. The AU p value is indicated for each node. Protein level changes were determined by Welch’s test p value ˂ 0.05 and absolute value of z-score ˃ 1.96, *p<0.05, **p<0.01. Abbreviations: AU, Approximately Unbiased; CTRL Control; PD, Parkinson’s disease.

57
Figure 2.4. Erythrocyte and EV implication in PD: summary of the literature. The search for a reliable biomarker in PD has included investigations in neuronal cells in specific in vitro systems, in cellular elements within the circulatory system, the cerebrospinal fluid and urine with a targeted interest on the expression of α-Syn. However, none of these studies have convincingly identified a robust marker of PD. Abbreviations: αSyn, α-synuclein; CAT, Catalase; GPx, glutathione peroxidase; L1CAM, L1 cell adhesion molecule; LRRK2, Leucine-rich repeat kinase 2; RBC, red blood cells; SOD, superoxide dismutase; ✓ presence;
↑ increase; ↓ decrease.

58
2.10 Matériels supplémentaires
2.10.1 Materials and methods
Preparation of platelet-free plasma and EV labeling
For all experiments, diluted annexin-V buffer (BD Pharmingen, Mississauga, ON, Canada)
and phosphate buffered saline (PBS) were filtered on 0.22mm pore size membranes. To
quantify EV according to their cell of origin, the following surface markers were used:
CD235a+ (erythrocytes) (5µl), CD31+/CD41- (endothelial cells) (1µl), CD41+ (platelets)
(5µl), CD45+ (leukocytes) (3µl), CD45+CD14+ (monocytes) (10µl), and CD45+CD15+
(granulocytes) (2µl), with or without annexin-V staining (5µl). PFP (5µl) was incubated with
Phenylalanyl-prolyl-arginyl Chloromethyl Ketone (PPACK) 10mM (Calbiochem,
Etobicoke, ON, Canada) for 5 minutes followed by a 30-minute incubation with antibodies
and annexin-V in a final PBS volume of 100µl, all at room temperature. Finally, samples
were diluted to a final volume of 2ml prior to FACS analysis. The following antibodies were
purchased from BD Pharmingen and used for all analyses: FITC-conjugated mouse anti-
human CD235a (clone GA-R2 (HIR2), 1/20), PE-conjugated mouse anti-human CD31 (clone
WM59, 1/100), V450-conjugated mouse anti-human CD41a (clone HIP8, 1/20), V450-
conjugated mouse anti-human CD45 (clone HI30, 1/33), APC mouse anti-human CD14
(clone M5E2, 1/10), PE-conjugated mouse anti-human CD15 (clone HI98, 1/50), V450- and
PerCP-CyTM5.5-conjugated annexin-V (1/20).
Flow cytometry quantification
Between sample analyses of the PD and HD patients, the blue laser had to be replaced for
maintenance issues and therefore laser settings were recalibrated. For FSC-PMT, the
assigned voltage was 363 (PD) and 160 (HD) Volts. For SSC, the assigned voltage was 407
(PD) and 300 (HD) Volts. All other parameters were set between 450 and 500 Volts. The
acquisition of EV was performed at low speed at an approximated rate of 10μl/min. To
determine background noise levels, antibody mixes were incubated in the absence of PFP
sample and unlabeled PFP was used as a negative control. Every sample was re-identified to
ensure that the experimenter was blind to clinical status.

59
Production and purification of EEV
Briefly, the erythrocyte pellet was activated with 3 volumes of calcium ionophore solution
(150mM NaCl; 10mM Tris-HCl; 1mM CaCl2; 5μM ionophore A23187 (Sigma, St Louis,
MO)) for 30 minutes at 37°C. The activation was stopped by the addition of 5mM EDTA.
Remaining erythrocytes were pelleted at 15 000g for 20 minutes. The EEV were centrifuged
at 20 000g for 90 minutes and washed once in PBS. The EV pellet was re-suspended in PBS
and frozen at -80°C until further analyses.
Scanning electron microscopy
Preparations of erythrocytes (5μl) were fixed in 2% paraformaldehyde and 2.5%
glutaraldehyde in PBS buffer at least 24 hours before standard dehydration. Samples were
washed 3 times for 10 minutes with sodium cacodylate buffer (0.1M, pH 7.3) and fixed with
1% osmium tetroxide in sodium cacodylate buffer for 90 minutes. Subsequently, samples
were washed and processed in 50%, 70%, 90% and 100% EtOH for dehydration (10
minutes/step). Finally, samples were soaked in two subsequent baths of 100% EtOH, for 40
minutes and 10 minutes, air-dried overnight and coated with palladium. Observations were
completed using a JEOL 6360LV scanning electron microscope (JEOL, Peabody, MA,
USA).
Transmission electron microscopy
Preparations of EEV (30μl) and activated erythrocytes (5μl) were fixed in 2%
paraformaldehyde at least 24 hours before being dehydrated and sealed in LR white. Slices
of LR white were placed on a Formvar/carbon-coated grid and processed for
immunolabeling. The tissues mounted on grids were blocked in 0.5% BSA-c (Aurion,
Wageningen, The Netherlands) in HBSS and incubated for 120 minutes with rabbit anti-α-
Syn antibody (Abcam, Toronto, ON, Canada) or rabbit anti-α-Syn (phospho S129) antibody
(Abcam, Toronto, ON, Canada), both diluted at 1:250 in HBSS and washed several times
with distilled water. Finally, the grids were incubated for 60 minutes with an anti-rabbit IgG
conjugated to 6nm gold particles (EMS, Hatfield, PA, USA) diluted at 1:200, washed several
times with distilled water and then fixed in 2.5% glutaraldehyde (EMS, Hatfield, PA, USA)
in HBSS for 15 minutes. For this last step, the grids were treated with 3% uranyl acetate–

60
0.075 M oxalate (pH 7.0) (EMS, Hatfield, PA, USA) for 1 minute, which was followed by
several washes in distilled water. All staining experiments included negative controls where
the primary antibody was omitted from the incubation media. Observations were completed
with a TECNAI Spirit G2 transmission electron microscope at 80kV (FEI, Hillsboro, OR,
USA).
Mass spectrometry analysis and label free protein quantification
The extracted peptides from the 7 slices of the same individual were pooled and analyzed by
nanoLC-MS/MS. The excised hemoglobin gel slices were also analyzed in the same
conditions. One µg of each individual sample was injected on a Dionex UltiMate 3000
nanoRSLC system (Thermo Scientific) equipped with a nanoviper Acclaim Pepmap100,
C18, 3µm, 75µm x 50cm column (Thermo Scientific) connected to the nanoelectrospray
source of an Orbitrap Fusion mass spectrometer (Thermo Scientific). The peptides were
eluted at 300nL/min using an acetonitrile gradient of 90 minutes with the mass spectrometer
operating in the Data Dependent Acquisition mode. Peptide masses were measured in MS
spectra detected in the orbitrap at 120K resolution. MSMS fragmentation spectra of peptides
were generated by Higher energy Collisional Dissociation (HCD) and detected in the ion
trap.
Statistical analyses
Initial statistical analyses (unpaired t-tests or Mann-Witney when data followed normal
distribution and Shapiro-Wilk test in cases of non-normality) compared the number of each
type of blood cells (erythrocytes, platelets, leukocytes, neutrophils, monocytes, lymphocytes)
in control groups and patients of all stages of the UK to the Canadian cohorts. This revealed
no statistical differences between groups, allowing us to pool cohorts for subsequent
analyses. Statistical analyses pertaining EEV quantification were performed using The
Statistics and Machine Learning Toolbox provided by MathWorksTM under MATLABTM
platform using the MATLABR2015a version. Results obtained include the scatter plots,
classical least-square linear regressions, R-squared and p values, as well as Pearson’s
goodness-of-fit. Interval cut-off values were determined using a loop program developed in
MATLABTM. Model diagnostics, including residual behaviour and homoscedastivity, were

61
obtained using the same Toolbox. Further details on the statistical tests chosen are described
directly in the result section. For α-Syn quantification, data were first tested for normality
using the D’Agostino & Pearson normality test. Comparisons between groups were obtained
by Mann-Whitney U test or Kruskal-Wallis ANOVA and performed using Prism 6.0
(GraphPad Software, LaJolla, CA). For proteomic analyses, the ‘Intensity values’ contained
in the output ‘proteingroup.txt’ file of MaxQuant were used to quantify each identified
protein in each individual sample. The values were normalized by the median of each column
(all intensity values of proteins for one sample). The missing values were imputed with a
noise value corresponding to the 1-percentile of each sample column. For each comparison
between two groups (Control, mild PD or moderate PD), proteins with too many imputed
values where considered not quantifiable (a minimum of three not-imputed values in one of
the 2 groups are required). A protein ratio was calculated between the two groups using the
average of intensity values in each group. Finally, a statistical Welch's test was performed
between the two groups. The protein ratios were transformed into log2(ratio) then centered
by calculation of a z-score (z-score = (x - µ)/σ). A protein was considered as variant if it
fulfilled the following criteria: minimum of 2 peptides quantified, Welch’s test p value ˂ 0.05
and absolute value of z-score ˃ 1.96 (corresponding to values outside of the 95% confidence
interval). The Gene Ontology enrichment analysis on the identified proteins was performed
on the Cytoscape platform (v. 3.4.0) using the BinGO software version 3.0.3 against all
human genes with GO annotation (Uniprot-GOA generated 2015-06-22) (Maere et al., 2005).
Enrichment was calculated by a hypergeometric test and Bonferroni Family-Wise Error Rate
(FWER) was used to correct for multiple testing. The data for the resulting 8 proteins was
standardized, hierarchically clustered and visualized as a heatmap by using the statistical
framework R (R Core Team, 2016). The robustness of the nodes was evaluated by computing
Approximately Unbiased (AU) p values using the R package pvclust (10000 bootstraps,
average method and correlation-based dissimilarity matrix) (Suzuki and Shimodaira, 2006).

62
2.10.2 Tableaux
Table S2.1. Quantification of cell-derived EV – PD cohort. The quantification of EV in platelet-free plasma of PD patients and controls was performed by high-sensitivity flow cytometry. Phosphatidylserine was evaluated with Annexin V binding. The complete blood count was obtained at the time of blood sampling for 35|37 Controls and 57|59 PD and was used to calculate the EV on cell ratio. * For erythrocyte-derived EV quantification, 1 outlier (determined using Grubbs’ method with alpha=0.0001) was removed. As opposed to Figure 3A, all other samples were included in the analyses (i.e. elevated hemoglobin, diabetes and/or patients having suffered from cancer). Abbreviation: CD235a, glycophorin A; CTRL, Control; EV, extracellular vesicle; PD, Parkinson’s disease; PS, phosphatidylserine.
Table S2.1. Quantification of cell-derived EV - PD cohort
Cell types Markers Units CTRL PD p
value n Mean SEM n Mean SEM
Erythrocytes
CD235a+PS- X 103/µl 36 18.2 46.5 59 32.0 36.3 0.04 CD235a+PS+ X 103/µl 36 0.22 0.07 59 0.29 0.05 0.70 CD235a+ total X 103/µl 36 18.4 47.0 59 32.3 36.7 0.04 EV CD235a+/erythrocyte 34 0.0039 0.011 57 0.0069 0.008 0.04
Platelets
CD41+PS- X 103/µl 37 7.88 1.68 59 10.3 1.33 0.27
CD41+PS+ X 103/µl 37 15.2 3.20 59 17.9 2.53 0.51
CD41+ total X 103/µl 37 23.1 4.62 59 28.2 3.66 0.38
EV CD41+/platelet 35 0.106 0.021 57 0.125 0.016 0.49
Endothelial cells
CD31+CD41-PS- X 103/µl 37 15.8 8.04 59 11.7 6.37 0.75
CD31+CD41-PS+ X 103/µl 37 0.91 0.13 59 0.92 0.10 0.96
CD31+CD41- total X 103/µl 37 16.7 8.03 59 12.6 6.36 0.75 Leukocytes CD45+ total X 103/µl 37 10.4 2.21 59 13.8 1.75 0.26
Monocytes
CD45-CD14+PS- X 103/µl 37 1.70 0.30 59 1.62 0.24 0.85
CD45-CD14+PS+ X 103/µl 37 1.20 4.00 59 5.84 3.17 0.50
CD45+CD14+PS- X 103/µl 37 0.16 0.04 59 0.14 0.03 0.74
CD45+CD14+PS+ X 103/µl 37 0.60 0.79 59 1.47 0.63 0.59
CD14+ total X 103/µl 37 3.66 4.88 59 9.06 3.87 0.60
EV CD14+/monocyte 35 7.08 1.99 57 9.16 1.56 0.41
Granulocytes
CD45-CD15+PS- X 103/µl 37 12.3 7.96 59 16.7 6.30 0.92
CD45-CD15+PS+ X 103/µl 37 2.21 0.77 59 1.39 0.61 0.47
CD45+CD15+PS- X 103/µl 37 0.55 0.36 59 1.15 0.29 0.20
CD45+CD15+PS+ X 103/µl 37 1.01 0.30 59 1.25 0.24 0.56
CD15+ total X 103/µl 37 16.0 8.83 59 20.6 6.99 0.91
EV CD15+/granulocyte 35 3.70 0.64 57 3.16 0.50 0.53

63
Table S2.2. Quantification of cell-derived EV – HD cohort. The quantification of EV in platelet-free plasma of HD patients and controls was performed by high-sensitivity flow cytometry. Phosphatidylserine was evaluated with Annexin V binding. The complete blood count was obtained at the time of blood sampling for 53|54 CTRL, 10 pre-manifest and 48|50|51 HD patients and was used to calculate the EV on cell ratio. Statistical analyses were performed using Wilcoxon/Kruskal-Wallis followed by Wilcoxon’s multiple comparison tests. Abbreviation: CD235a, glycophorin A; CTRL, Control; EV, extracellular vesicle; HD, Huntington’s disease; PS, phosphatidylserine.
Table S2.2. Quantification of cell-derived EV - HD cohort
Cell types Markers Units CTRL HD pre-manifest HD
p value n Mean SEM n Mean SEM n Mean SEM
Erythrocytes
CD235a+PS- X 103/µl 54 15.2 2.0 10 10.3 3.5 51 14.1 1.4 0.16 CD235a+PS+ X 103/µl 54 1.1 0.2 10 0.4 0.2 51 1.1 0.1 0.04
CD235a+ total X 103/µl 54 16.4 2.0 10 10.7 3.5 51 15.3 1.5 0.09
EV CD235a+/erythrocyte 54 0.0035 0.0005 10 0.0023 0.0008 50 0.0033 0.0003 0.11
Platelets
CD41+PS- X 103/µl 54 9.2 2.2 10 4.3 1.3 50 6.1 1.0 0.78
CD41+PS+ X 103/µl 54 19.3 4.8 10 7.1 2.0 50 12.4 2.4 0.74
CD41+ total X 103/µl 54 28.4 6.9 10 11.4 3.2 50 18.6 3.4 0.70
EV CD41+/platelet 53 0.12 0.03 10 0.05 0.02 48 0.08 0.01 0.34
Endothelial cells
CD31+CD41-PS- X 103/µl 54 1.4 0.3 10 0.6 0.2 50 1.2 0.2 0.31
CD31+CD41-PS+ X 103/µl 54 0.68 0.16 10 0.25 0.06 50 0.46 0.09 0.59
CD31+CD41- total X 103/µl 54 2.1 0.4 10 0.8 0.2 50 1.7 0.3 0.26 Leukocytes CD45+ total X 103/µl 54 33.4 2.7 10 31.6 5.3 51 31.7 2.4 0.88
Monocytes
CD45-CD14+PS- X 103/µl 54 3.4 1.1 10 1.6 0.2 51 1.6 0.1 0.91
CD45-CD14+PS+ X 103/µl 54 1.8 0.3 10 0.8 0.3 51 1.5 0.2 0.14
CD45+CD14+PS- X 103/µl 54 0.18 0.07 10 0.069 0.016 51 0.056 0.008 0.34
CD45+CD14+PS+ X 103/µl 54 0.62 0.12 10 0.24 0.06 51 0.55 0.14 0.12
CD14+ total X 103/µl 54 6.0 1.3 10 2.6 0.4 51 3.7 0.4 0.08
EV CD14+/monocyte 53 12.3 2.5 10 5.7 0.6 48 8.0 1.0 0.13
Granulocytes
CD45-CD15+PS- X 103/µl 54 1.2 0.1 10 1.2 0.3 51 1.5 0.2 0.33
CD45-CD15+PS+ X 103/µl 54 0.12 0.04 10 0.18 0.08 51 0.22 0.11 0.33
CD45+CD15+PS- X 103/µl 54 0.20 0.05 10 0.07 0.02 51 0.15 0.04 0.64
CD45+CD15+PS+ X 103/µl 54 0.25 0.05 10 0.13 0.06 51 0.20 0.04 0.39
CD15+ total X 103/µl 54 1.7 0.2 10 1.6 0.4 51 0.20 0.3 0.67
EV CD15+/granulocyte 53 0.41 0.04 10 0.42 0.13 48 0.50 0.08 0.75

64
Voir Annexe Table S2.3. Complete list of proteins identified in the EEV proteome. Lists of the protein groups identified by LC-MS/MS analysis using Andromeda/MaxQuant search engine in the Uniprot Complete Proteome Homo sapiens database for the non-depleted sample (356 proteins) (A) or when hemoglobin was analyzed separately (818 proteins) (B). The lists were filtered at 1% False Discovery Rate using a target/decoy database search.

65
2.10.3 Figures
Figure S2.1. Optimization of EV detection: controls for flow cytometry. Controls for EV identification. A. To properly set the EV gate, fluorescent silica beads of 100nm (Red), 500nm (Blue) and 1000nm (Yellow) were acquired on a flow cytometer Canto II modified with a FSC-PMT small particle option. The established EV gate was used for all experiments. B. Serial dilutions of EEV (1, 2, 4 and 10) were used to confirm the linearity of the quantification. C. FSC-PMT/SSC gates of PFP stained with annexin V and respective fluorochrome-conjugated antibodies directed against erythrocyte (CD235a+), endothelial (CD31+/CD41-), platelet (CD41+) and leukocyte (CD14+CD45+, monocytes;

66
CD15+CD45+, granulocytes)-derived EVs. Controls for EV labeling. D. Treatment with the ion chelator EDTA inhibited the binding of annexin V to phosphatidylserine. E. Minimal background was observed using antibodies in the absence of PFP. This background was subtracted from all subsequent EV quantifications. F. EV sensitivity to 0.5% triton was further assessed. Abbreviations: AnnV, annexinV; EV, extracellular vesicle; EEV, erythrocyte-derived extracellular vesicle; FSC PMT-H, forward scatter photomultiplier tube; PBS, phosphate buffered saline; PFP, platelet free plasma; SSC-H, side scatter.

67
Figure S2.2. LEDD and EEV. Additional information on patients derived from each correlation has been provided (number of EEV/total number of erythrocytes and LEDD) to illustrate that LEDD cannot account for the differences we found in EEV counts. A. EV/erythrocytes ratios, UPDRS scores and LEDD values for all PD patients used in correlation analyses. B. Absence of correlation between the number of EEV and LEDD. Abbreviations: EEV, erythrocyte-derived extracellular vesicle; EV, extracellular vesicle; LEDD, Levodopa equivalent daily dose; UPDRS, Unified Parkinson’s Disease Rating Scale.

68
Figure S2.3. Blood counts and exclusion criteria. A-B. Full blood counts performed in PD (n=57), pre-manifest HD (n=10) and HD patients (n=50) revealed no significant differences for the total number of any cell type between patients and their respective healthy sex- and age-matched CTRL (n=37 for PD; n=55 for pre-HD/HD). C. Evaluated hematocrit, mean corpuscular hemoglobin and mean corpuscular volumes were similar between groups (PD, n=43; CTRL, n=24). D-E. CRP (indicative of an inflammatory response) (D) and free hemoglobin (indicative of hemolysis) (E) were further quantified in the PFP (PD, n=59; CTRL, n=37). Individuals presenting with levels exceeding 45 000ng/ml were excluded from the study to remove the possibility that these factors contributed to increased EEV concentrations. Statistical analyses were performed using Kruskal-Wallis ANOVAs (A-B) and Mann-Whitney U tests (C-E). Abbreviations: CTRL, controls; CRP, C-reactive protein; EEV, erythrocyte-derived extracellular vesicle; HD, Huntington’s disease; PD, Parkinson’s disease; PFP, platelet-free plasma.

69

70
Figure S2.4. Proteomic analyses with and without hemoglobin. To ensure the specificity of the protein signature detected in EEV, we performed proteomic analyses using two distinct methodologies. A. The first set of analyses by nanoLC-MS/MS and Label Free Quantification was performed on the complete proteome of the EEV, yielding 356 proteins. B. The second set of analyses was also performed on the entire EEV proteome but this time, isolating hemoglobin and quantifying the proteins present in both fractions. This approach uncovered 708 proteins in the EEV proteome with an additional 110 proteins in the hemoglobin fraction for a total of 818 proteins, indicating that removing hemoglobin provides a much more accurate evaluation of the protein content of EEV. To further confirm that the identified proteins were associated with the EEV proteome, a Gene Ontology enrichment analysis was performed on the Cellular Component ontology using BinGO software. The list of 356 identified proteins in the entire EEV (A) and the list of the 818 identified proteins when hemoglobin was analyzed separately (B) were compared to the whole human proteome (reference list). Circle size is proportional to the number of proteins matching the corresponding GO term and the color scale corresponds to the p value of enrichment (darker orange corresponds to greater statistical enrichments).

71
Figure S2.5. Confirmation of EEV proteins selectively modified in PD patients by Volcano plots. The protein ratios (log2(ratio)) of the three comparisons (mild PD/CTRL, moderate PD/CTRL and moderate PD/mild PD) were plotted over the corresponding Welch’s test p value (-log10(p-value)). The graphs display a V shape, as expected, and only the proteins falling outside the limits of a p value < 0.05 and absolute value of z-score > 1.96 (identified by black lines) were considered as variant proteins (red dots). Two variant proteins were excluded given that they were quantified using only one peptide. Abbreviations: CTRL: Control; PD, Parkinson’s disease.

72
CHAPITRE 3: DISCUSSION

73
3.1 Caractérisation de la nature et du profil des VE
Le premier objectif de notre étude était de caractériser la nature et le nombre de VE dans le
plasma de patients atteints de la MP afin d’évaluer leur potentiel comme biomarqueur de
diagnostic ou de progression. Nous avons ainsi récolté le sang de 60 patients et 37 contrôles
appariés pour le sexe et l’âge. Pour chaque participant, nous avons recueilli toutes les données
cliniques disponibles (évaluations cliniques, médication, facteurs de comorbidités), effectué
une analyse de formule sanguine complète et quantifié chaque sous-type de VE.
3.1.1 Biomarqueur de valeur diagnostique
Bien que l’analyse de la formule sanguine complète n’ait révélé aucune différence dans le
nombre de cellules, nous avons observé une augmentation significativement différente de
VEE entre les patients et les contrôles. Nous avons également observé une augmentation
similaire du ratio entre les VEE et les érythrocytes, démontrant l’augmentation de la
libération des VEE par les érythrocytes. La quantification des érythrocytes reste quant à elle
similaire. Cependant, un examen plus approfondi de la quantification des VEE nous a révélé
que seulement 5 patients sont responsables de cette différence significative. Ces patients se
situent à un stade plus avancé de la maladie. On peut donc conclure que le nombre de VEE
ne peut être utilisé comme caractéristique discriminatoire entre un patient et une personne
saine.
3.1.2 Biomarqueur pour la progression de la maladie
Nous avons de surcroit évalué le potentiel des VEE comme biomarqueur des différents états
de la MP. Pour ce faire, nous avons analysé les corrélations du ratio VEE par érythrocytes
par rapport à certaines échelles cliniques de la maladie telles que le UPDRS, le H&Y, le
MMSE, le BDI et le ACE. Nos résultats révèlent que le coefficient de corrélation linéaire
entre l’échelle du UPDRS et le nombre de VEE se rapproche de 1. Les régressions linéaires
montrent que 87 % de la variation du ratio VEE/érythrocyte est expliqué par la fluctuation
du UPDRS. Les intervalles de confiance montrent un seuil d’erreur de moins de 5 %. Nos
résultats révèlent également deux corrélations distinctes séparant les patients de divers stades.

74
Nous avons observé que les patients ayant un résultat inférieur à 37 au UPDRS sont
caractérisés par une augmentation du nombre de VEE au stade précoce de la maladie. Il existe
de façon similaire, une corrélation entre les scores de 37 et 75 qui définit les patients à un
stade plus avancé de la MP. Comme le UPDRS comprend plusieurs sections, nous avons
regardé si le lien entre cette échelle et la concentration de VEE est spécifique à une section
(aspects non moteurs, aspects moteurs, langage, etc.). Nos résultats montrent qu’aucune
catégorie spécifique du UPDRS ne semble associée aux fortes corrélations observées. Malgré
notre analyse exhaustive, la scission est toujours difficile à expliquer puisque le UPDRS ne
possède pas de stratification des résultats clairement détaillés comme le H&Y. La mise à jour
du UPDRS par la Société des troubles du mouvement (MDS-UPDRS) à amener la
communauté des cliniciens à réfléchir sur une méthode pour estimer la gravité de la maladie.
Martínez-Martín et collaborateur ont montré qu’un résultat de MDS-UPDRS entre 30 et 35
semble marquer un seuil de distinction entre les patients précoces et les patients modérés
(Martínez-Martín et al., 2015). Ce seuil est attribué principalement à l’évaluation motrice qui
constitue la plus grande partie de l’échelle clinique. Bien que notre analyse ait été réalisée
avec le UPDRS standard, nous pensons que la scission peut permettre de différencier deux
stades de la maladie. La diminution précipitée de la concentration de VEE au début du stade
modéré pourrait être une cause d’un changement systémique chez le patient. L’étude
exhaustive des érythrocytes dans la MP pourrait potentiellement permettre d’expliquer ce
phénomène.
3.1.3 Validation des résultats
Nous avons également reproduit la quantification totale des vésicules sur une cohorte
indépendante de patients souffrant de la maladie d’Huntington. Les résultats ne révèlent
aucune différence entre les prémanifestes, les manifestes et les contrôles sains pour tous sous-
types de vésicules incluant les VEE. Nous pouvons ainsi présumer que le nombre de VEE
corrèle spécifiquement avec le UPDRS et donc la MP.
Afin de valider nos résultats, nous avons étudié si des facteurs externes à la maladie pouvaient
influencer la quantification de VEE. En premier lieu, nous avons étudié la prise de

75
médicaments pour chaque participant. Nous n’avons tenu compte d’aucune médication non
parkinsonienne puisque les traitements entre les participants sont extrêmement variés. Chez
les parkinsoniens, nous avons regardé le lien entre la dose équivalente journalière de
lévodopa et le nombre de VEE. Les données montrent que l’augmentation de VEE n’est pas
une conséquence du changement de médications liée à la lévodopa. De plus, nous avons
observé une augmentation significative de la concentration de VEE dans le plasma des
participants atteints de diabète et de cancer, patients et contrôles confondus. Il a été rapporté
que le diabète réduit la déformabilité érythrocytaire et augmente l’agrégation de ces derniers
(Cho et al., 2008). De plus, il semble que les VE dérivés du sang soient impliqués de diverses
manières dans ces deux maladies (Tokarz et al., 2015; Żmigrodzka et al., 2016). Nous avons
donc décidé d’exclure ces participants de nos analyses. Finalement, les plasmas comportant
un niveau d’hémoglobine libre élevée (> 45 000 ng/ml), potentiellement due à l’hémolyse
lors de la collecte de sang, ont également été exclus des analyses liées aux VEE.
3.2 Contenu protéique des VEE
Le deuxième objectif de notre étude était de caractériser le contenu protéique des VEE.
3.2.1 Méthodes d’activation
Pour l’étude des VEE, il est nécessaire de les isoler et les concentrer. Nous avons essayé,
dans un premier temps, d’isoler les vésicules directement du plasma. Malheureusement, les
techniques utilisées, optimisées davantage pour les cellules, n’ont pas fonctionné avec
d’aussi petites et peu nombreuses particules. Nous avons donc tenté d’activer les érythrocytes
d’une facon la plus physiologique possible. Selon plusieurs études, les ions calcium semblent
être la clé de la biogénèse de vésicules extracellaires. Ils participent à l’activation ou
l’inhibition de protéines et phospholipides au niveau de la membrane cellulaire. Le calcium
ionophore A23187 provoque un changement dans la concentration de calcium induisant la
vésiculation. Contrairement à l’eau et la congélation, souvent utilisé pour provoquer la
libération de VEE, ce composé évite de faire éclater ou de détruire la membrane des
érythrocytes (Nguyen et al., 2016). Cependant, bien que le taux d’ion calcium change en

76
condition physiologique, notamment avec l’âge, le A23187 induit un changement d’ion
supérieur à ce qui est réellement observé fixant ainsi une limite à notre méthode in vitro.
Nous avons imagé, par microscopie électronique à balayage, des érythrocytes au repos et
activés de patients et contrôles afin de comparer la morphologie des cellules. Bien que ces
images montrent la vésiculation des érythrocytes, aucune différence entre les individus n’a
pu être observée.
3.2.2 Quantification d’α-Syn par ELISA et microscopie
Bien que l’α-Syn soit très présente dans les tissus cérébraux, des études montrent qu’on la
retrouve également en quantité importante dans le sang et plus particulièrement dans les
érythrocytes laissant présager sa présence dans les VEE (Barbour et al., 2008). Nous avons
donc regardé la concentration d’α-Syn totale et d’une forme phosphorylée (Sérine 129) dans
les VEE en microscopie électronique entre des patients à un stade modéré de la maladie et
leurs contrôles. Suite aux résultats ne montrant aucune différence entre les groupes pour les
deux types d’α-Syn, nous avons effectué une quantification de la protéine totale par ELISA
chez des patients aux stades précoce et modéré ainsi que chez les contrôles. Contrairement à
la méthode par microscopie électronique, l’ELISA est beaucoup plus quantitatif et nous
permet d’obtenir des valeurs en nanogramme par millilitre. Les résultats confirment que la
concentration d’α-Syn reste similaire entre les patients et les personnes saines. On peut donc
conclure que le niveau d’expression de cette protéine dans les VEE ne peut servir de
biomarqueur. Cependant, l’α-Syn pourrait tout de même jouer un rôle majeur au sein des
érythrocytes.
3.2.3 Protéomique
À la vue des résultats précédents, nous avons décidé d’analyser le protéome entier des VEE
de patients aux stades précoce et modéré ainsi que leurs contrôles afin de détecter une
éventuelle signature protéique entre les participants. Étant donné la grande quantité
d’hémoglobine qui subsiste dans les VEE, nous avons effectué deux types d’approches
distinctes afin d’optimiser la détection de l’ensemble du protéome des VEE. Nous avons

77
réalisé une première approche où l’ensemble du protéome fût analysé en un bloc, révélant
ainsi un total de 356 protéines. Nous avons, par la suite, utilisé une deuxième approche qui
consiste à séparer l’hémoglobine des autres protéines et de les analyser séparément. Par cette
méthode nous avons détecté 818 protéines. De plus, une analyse par Gene Ontology,
comparant nos échantillons avec l’ensemble du génome humain, montre un enrichissement,
des protéines associées aux vésicules et aux complexes d’hémoglobine.
Parmi les 818 protéines, il nous a été possible d’identifier 8 protéines significativement
modulées entre les groupes. Une heat map nous a d’ailleurs permis de mettre en évidence 3
groupes de protéines. Le premier groupe constitué les protéines (gènes) AIDA, ABHD14B
et NADSYN1 qui sont moins exprimées chez l’ensemble des patients. Les protéines (gènes)
QDPR, AKR1A1 et CNRIP1 constituent le deuxième groupe où l’expression semble être
plus importante chez les patients au stade précoce. Finalement, le troisième groupe constitué
de USP24 et ATP5A1 est significativement plus exprimé chez les patients au stade modéré.
La documentation concernant l’implication de ces protéines dans la MP est rare. Toutefois,
les ARN messagers pour la protéine CNR semblent diminués dans le striatum chez un modèle
animal de MP (Zeng et al., 1999). On remarque également que des variantes génétiques de
USP24 semblent responsables d’un risque associé à la MP (Li et al., 2006). De plus, on
retrouve plusieurs formes d’ATP synthase à des concentrations anormales chez les
parkinsoniens pouvant expliquer le changement d’expression de l’ATP5A1 (Ferrer et al.,
2007).
En terminant, la heat map et la mise en cluster hiérarchique des individus (patients précoces,
patients modérés, contrôles) permettent de les regrouper grâce à la similarité de l’expression
des 8 protéines. Ces résultats suggèrent l’utilisation de ces 8 protéines comme biomarqueur.
Cependant, on peut se demander si ces changements reflètent des modifications également
présentent dans les globules rouges. Bien que ces cellules soient anucléées, elles possèdent
plus de 2600 protéines dont toutes celles identifiés dans notre étude (Bryk and Wiśniewski,
2017). Malheureusement, on ne trouve pour l’instant, aucune étude détaillant les altérations
du protéomes d’érythrocytes dans la MP.

78
3.3 Perspectives d’avenir
3.3.1 Perspectives expérimentales
Suivant les expériences que nous avons effectuées lors de ce projet, il serait intéressant de se
pencher sur l’implication des VEE dans le développement de la pathologie. En effet, nous
observons que ce type de vésicule peut potentiellement servir d’outil pour évaluer la
progression de la maladie. Cependant, nous ignorons si elles jouent un rôle dans la
pathogenèse ou si elles ne sont qu’une conséquence de celle-ci. Des données préliminaires
visant à déterminer le rôle des VEE comme transporteur au niveau de la barrière
hématoencéphalique nous ont permis de constater que les vésicules semblent interagir avec
les cellules endothéliales. Nous avons incubé pendant 18 heures des VEE, préalablement
marqués par la green fluorescent protein (GFP), avec une barrière de cellules endothéliales
dans un modèle de chambre de Boyden tel que décrit par Prat et collaborateur (Prat et al.,
2005). Les résultats nous ont montré que certaines VEE peuvent traverser la couche de
cellules alors que d’autres sont internalisés par les cellules endothéliales. Cette étude, bien
qu’incomplète, pourrait être optimisée notamment en faisant une analyse de l’interaction des
vésicules et des cellules endothéliales en reprenant le modèle de la chambre de Boyden ou
en utilisant des systèmes microfluidiques (ibidi, BioFlux) (Ganguly et al., 2012; Hayes et al.,
2017). Selon les résultats, nous pourrions comprendre si les VEE ont la capacité de passer la
barrière hématoencéphalique et d’agir comme véhicule de propagation de la pathologie. La
conclusion de ces études pourrait nous permettre d’envisager de nouvelles voies
thérapeutiques (Hall et al., 2016).
Il serait également intéressant de regarder dans les érythrocytes si les mécanismes impliqués
dans le bourgeonnement des VEE sont altérés ou stimulés par la maladie. Comme l’α-Syn
semble réguler les vésicules synaptiques au niveau présynaptique, nous pourrions émettre
l’hypothèse d’une fonction similaire de régulation des vésicules dans les cellules sanguines
(Wang et al., 2015). De plus, l’augmentation de l’éryptose, tel que discuter à la section 1.2.1,
pourrait également être un facteur contribuant à l’augmentation du relâchement des vésicules.
Les résultats nous permettraient de mieux comprendre le rôle des globules rouges et de l’α-
Syn dans la pathologie, pouvant apporter potentiellement de nouvelles cibles thérapeutiques.

79
Finalement, l’analyse protéomique d’érythrocytes chez des patients et des personnes saines
pourrait nous indiquer si le changement protéique observé dans les VEE est également
présent dans les globules rouges.
3.3.2 Perspectives diagnostiques
Dans le but de faire valider notre biomarqueur au niveau clinique, il serait nécessaire de faire
une nouvelle étude en augmentant le nombre de patients et leurs contrôles afin de couvrir un
échantillon de population plus important. Ainsi, nous pourrions comparer davantage les
patients selon leurs médications et leurs facteurs de comorbidités. De plus, il est nécessaire
d’obtenir la contribution de ces individus afin de démontrer la valeur de notre biomarqueur
comme échelle diagnostic. Il serait intéressant de voir si ces patients suivent la corrélation
attribuée aux patients modérés ou s’ils forment une troisième corrélation distincte.
Finalement, il pourrait être pertinent d’inclure une population de patients souffrant de forme
atypique de la MP ou de maladies neurologiques dont le diagnostic est similaire à la MP par
exemple, l’atrophie multisystématisée, la paralysie supranucléaire progressive et la démence
à corps de Lewy (McKeith et al., 2005; Respondek and Höglinger, 2016; Laurens et al., 2017;
Höglinger et al., 2017).
L’obtention récente d’une demande de fonds du Weston Brain Institute par le Dr Cicchetti
va permettre de réaliser plusieurs de ces perspectives en allant chercher 200 patients
parkinsoniens et 200 personnes saines. Les résultats préliminaires de cette nouvelle cohorte
semblent reproduire la corrélation entre les VEE de patients au stade précoce et le UPDRS.
La validation des corrélations et de la signature protéique sur de grandes cohortes
représentera une avancée majeure pour la découverte de biomarqueurs dans la MP.
3.4 Conclusion
Ce projet nous a permis d’identifier un nouveau biomarqueur qui cible les états de la MP
selon deux corrélations associées au stade précoce et au stade modéré de la maladie. Les
analyses statistiques montrent en effet que la quantité de VEE dans le plasma corrèle

80
fortement avec le UPDRS. De tels résultats retrouvés dans une taille d’échantillon de 60
patients dans une maladie aussi hétérogène mettent en évidence l’importance du biomarqueur
que nous avons identifié. Le contenu protéique de ces VEE pourrait d’ailleurs nous aider à
comprendre les bases des différents stades de la maladie en plus de démontrer un point
critique de son évolution clinique.
La plupart des tests actuellement mis au point pour l’utilisation des biomarqueurs chez les
patients sont basés sur des tests ELISA. Ces tests requièrent beaucoup de temps et des
quantités importantes de matériels biologiques. Nous proposons un biomarqueur facile
d’approche pour la clinique. En effet, il ne nécessite qu’un volume de sang minimal et peut
facilement être automatisé. De plus, contrairement aux études protéomiques standards, nous
proposons une méthode plus stricte et précise qui vise à éliminer l’interférence causée par
l’hémoglobine (Bosman et al., 2012). Son potentiel est donc grandement intéressant pour les
études cliniques.
L’identification des VEE en tant que biomarqueur pourrait : 1) aider au diagnostic de la MP,
en particulier au stade précoce de la maladie lorsque les thérapies neuroprotectrices
pourraient avoir leur plus grand impact ; 2) conduire à un marqueur utile qui valide les
échelles cliniques, 3) fournir une cible thérapeutique ou un marqueur de modification de la
maladie dans le développement et l’essai de nouvelles thérapies et 4) offrir de nouveaux
aperçus sur la pathogenèse de la maladie et sont évolution à travers les stades de la maladie.
Bibliographie
Abd-Elhadi, S., A. Honig, D. Simhi-Haham, M. Schechter, E. Linetsky, T. Ben-Hur, and R. Sharon. 2015. Total and Proteinase K-Resistant α-Synuclein Levels in Erythrocytes, Determined by their Ability to Bind Phospholipids, Associate with Parkinson’s Disease. Sci. Rep. 5:11120. doi:10.1038/srep11120.
Abou-Sleiman, P.M., D.G. Healy, N. Quinn, A.J. Lees, and N.W. Wood. 2003. The role of

81
pathogenic DJ-1 mutations in Parkinson’s disease. Ann. Neurol. 54:283–286. doi:10.1002/ana.10675.
Abraham, S., C.C. Soundararajan, S. Vivekanandhan, and M. Behari. 2005. Erythrocyte antioxidant enzymes in Parkinson’s disease. Indian J. Med. Res. 121:111–115.
Ahn, T.-B., S.Y. Kim, J.Y. Kim, S.-S. Park, D.S. Lee, H.J. Min, Y.K. Kim, S.E. Kim, J.-M. Kim, H.-J. Kim, J. Cho, and B.S. Jeon. 2008. alpha-Synuclein gene duplication is present in sporadic Parkinson disease. Neurology. 70:43–49. doi:10.1212/01.wnl.0000271080.53272.c7.
Akers, J.C., D. Gonda, R. Kim, B.S. Carter, and C.C. Chen. 2013. Biogenesis of extracellular vesicles (EV): exosomes, microvesicles, retrovirus-like vesicles, and apoptotic bodies. J. Neurooncol. 113:1–11. doi:10.1007/s11060-013-1084-8.
Albin, R.L., A.B. Young, and J.B. Penney. 1989. The functional anatomy of basal ganglia disorders. Trends Neurosci. 12:366–375.
Alonso-Navarro, H., F.J. Jimenez-Jimenez, E. Garcia-Martin, and J.A.G. Agundez. 2014. Genomic and pharmacogenomic biomarkers of Parkinson’s disease. Curr. Drug Metab. 15:129–181.
Alvarez-Erviti, L., Y. Seow, A.H. Schapira, C. Gardiner, I.L. Sargent, M.J.A. Wood, and J.M. Cooper. 2011. Lysosomal dysfunction increases exosome-mediated alpha-synuclein release and transmission. Neurobiol. Dis. 42:360–367. doi:10.1016/j.nbd.2011.01.029.
Amo, T., S. Sato, S. Saiki, A.M. Wolf, M. Toyomizu, C.A. Gautier, J. Shen, S. Ohta, and N. Hattori. 2011. Mitochondrial membrane potential decrease caused by loss of PINK1 is not due to proton leak, but to respiratory chain defects. Neurobiol. Dis. 41:111–118. doi:10.1016/j.nbd.2010.08.027.
Andersen, A.D., M. Binzer, E. Stenager, and J.B. Gramsbergen. 2017. Cerebrospinal fluid biomarkers for Parkinson’s disease - a systematic review. Acta Neurol. Scand. 135:34–56. doi:10.1111/ane.12590.
Antonyak, M.A., B. Li, L.K. Boroughs, J.L. Johnson, J.E. Druso, K.L. Bryant, D.A. Holowka, and R.A. Cerione. 2011. Cancer cell-derived microvesicles induce transformation by transferring tissue transglutaminase and fibronectin to recipient cells. Proc. Natl. Acad. Sci. U. S. A. 108:4852–4857. doi:10.1073/pnas.1017667108.
Araki, K., N. Yagi, R. Nakatani, H. Sekiguchi, M. So, H. Yagi, N. Ohta, Y. Nagai, Y. Goto, and H. Mochizuki. 2016. A small-angle X-ray scattering study of alpha-synuclein from human red blood cells. Sci. Rep. 6:30473. doi:10.1038/srep30473.
Archer, T., and R.M. Kostrzewa. 2016. Exercise and Nutritional Benefits in PD: Rodent Models and Clinical Settings. Curr. Top. Behav. Neurosci. 29:333–351. doi:10.1007/7854_2015_409.
Baba, Y., A. Kuroiwa, R.J. Uitti, Z.K. Wszolek, and T. Yamada. 2005. Alterations of T-lymphocyte populations in Parkinson disease. Parkinsonism Relat. Disord. 11:493–498. doi:10.1016/j.parkreldis.2005.07.005.
Bago Rožanković, P., M. Rožanković, L. Vučak Novosel, and M. Stojić. 2017. Nonmotor

82
symptoms in de novo Parkinson disease comparing to normal aging. Clin. Neurol. Neurosurg. 155:7–11. doi:10.1016/j.clineuro.2017.02.002.
Barbour, R., K. Kling, J.P. Anderson, K. Banducci, T. Cole, L. Diep, M. Fox, J.M. Goldstein, F. Soriano, P. Seubert, and T.J. Chilcote. 2008. Red blood cells are the major source of alpha-synuclein in blood. Neurodegener. Dis. 5:55–59. doi:10.1159/000112832.
Barile, L., and G. Vassalli. 2017. Exosomes: Therapy delivery tools and biomarkers of diseases. Pharmacol. Ther. 174:63–78. doi:10.1016/j.pharmthera.2017.02.020.
Bartels, T., J.G. Choi, and D.J. Selkoe. 2011. α-Synuclein occurs physiologically as a helically folded tetramer that resists aggregation. Nature. 477:107–110. doi:10.1038/nature10324.
Beach, T.G., C.H. Adler, B.N. Dugger, G. Serrano, J. Hidalgo, J. Henry-Watson, H.A. Shill, L.I. Sue, M.N. Sabbagh, H. Akiyama, and Arizona Parkinson’s Disease Consortium. 2013. Submandibular gland biopsy for the diagnosis of Parkinson disease. J. Neuropathol. Exp. Neurol. 72:130–136. doi:10.1097/NEN.0b013e3182805c72.
Beck, A.T., C.H. Ward, M. Mendelson, J. Mock, and J. Erbaugh. 1961. An inventory for measuring depression. Arch. Gen. Psychiatry. 4:561–571.
Behari, M., and M. Shrivastava. 2013. Role of platelets in neurodegenerative diseases: a universal pathophysiology. Int. J. Neurosci. 123:287–299. doi:10.3109/00207454.2012.751534.
Berg, D., M. Niwar, S. Maass, A. Zimprich, J.C. Möller, U. Wuellner, T. Schmitz-Hübsch, C. Klein, E.-K. Tan, L. Schöls, L. Marsh, T.M. Dawson, B. Janetzky, T. Müller, D. Woitalla, V. Kostic, P.P. Pramstaller, W.H. Oertel, P. Bauer, R. Krueger, T. Gasser, and O. Riess. 2005. Alpha-synuclein and Parkinson’s disease: implications from the screening of more than 1,900 patients. Mov. Disord. Off. J. Mov. Disord. Soc. 20:1191–1194. doi:10.1002/mds.20504.
Berg, D., R.B. Postuma, B. Bloem, P. Chan, B. Dubois, T. Gasser, C.G. Goetz, G.M. Halliday, J. Hardy, A.E. Lang, I. Litvan, K. Marek, J. Obeso, W. Oertel, C.W. Olanow, W. Poewe, M. Stern, and G. Deuschl. 2014. Time to redefine PD? Introductory statement of the MDS Task Force on the definition of Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 29:454–462. doi:10.1002/mds.25844.
Berger, A.K., G.P. Cortese, K.D. Amodeo, A. Weihofen, A. Letai, and M.J. LaVoie. 2009. Parkin selectively alters the intrinsic threshold for mitochondrial cytochrome c release. Hum. Mol. Genet. 18:4317–4328. doi:10.1093/hmg/ddp384.
Bernis, M.E., J.T. Babila, S. Breid, K.A. Wüsten, U. Wüllner, and G. Tamgüney. 2015. Prion-like propagation of human brain-derived alpha-synuclein in transgenic mice expressing human wild-type alpha-synuclein. Acta Neuropathol. Commun. 3:75. doi:10.1186/s40478-015-0254-7.
Besong-Agbo, D., E. Wolf, F. Jessen, M. Oechsner, E. Hametner, W. Poewe, M. Reindl, W.H. Oertel, C. Noelker, M. Bacher, and R. Dodel. 2013. Naturally occurring α-synuclein autoantibody levels are lower in patients with Parkinson disease. Neurology.

83
80:169–175. doi:10.1212/WNL.0b013e31827b90d1. Betarbet, R., T.B. Sherer, G. MacKenzie, M. Garcia-Osuna, A.V. Panov, and J.T.
Greenamyre. 2000. Chronic systemic pesticide exposure reproduces features of Parkinson’s disease. Nat. Neurosci. 3:1301–1306. doi:10.1038/81834.
Bisaglia, M., S. Mammi, and L. Bubacco. 2007. Kinetic and structural analysis of the early oxidation products of dopamine: analysis of the interactions with alpha-synuclein. J. Biol. Chem. 282:15597–15605. doi:10.1074/jbc.M610893200.
Boilard, E., P.A. Nigrovic, K. Larabee, G.F.M. Watts, J.S. Coblyn, M.E. Weinblatt, E.M. Massarotti, E. Remold-O’Donnell, R.W. Farndale, J. Ware, and D.M. Lee. 2010. Platelets amplify inflammation in arthritis via collagen-dependent microparticle production. Science. 327:580–583. doi:10.1126/science.1181928.
Bordia, T., N. Parameswaran, H. Fan, J.W. Langston, J.M. McIntosh, and M. Quik. 2006. Partial recovery of striatal nicotinic receptors in 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-lesioned monkeys with chronic oral nicotine. J. Pharmacol. Exp. Ther. 319:285–292. doi:10.1124/jpet.106.106997.
Borgonovo, J., C. Allende-Castro, A. Laliena, N. Guerrero, H. Silva, and M.L. Concha. 2017. Changes in neural circuitry associated with depression at pre-clinical, pre-motor and early motor phases of Parkinson’s disease. Parkinsonism Relat. Disord. 35:17–24. doi:10.1016/j.parkreldis.2016.11.009.
Bosman, G.J.C.G.M., E. Lasonder, Y.A.M. Groenen-Döpp, F.L.A. Willekens, and J.M. Werre. 2012. The proteome of erythrocyte-derived microparticles from plasma: new clues for erythrocyte aging and vesiculation. J. Proteomics. 76 Spec No.:203–210. doi:10.1016/j.jprot.2012.05.031.
Bosman, G.J.C.G.M., E. Lasonder, M. Luten, B. Roerdinkholder-Stoelwinder, V.M.J. Novotný, H. Bos, and W.J. De Grip. 2008. The proteome of red cell membranes and vesicles during storage in blood bank conditions. Transfusion (Paris). 48:827–835. doi:10.1111/j.1537-2995.2007.01630.x.
Bousquet, M., F. Calon, and F. Cicchetti. 2011. Impact of ω-3 fatty acids in Parkinson’s disease. Ageing Res. Rev. 10:453–463. doi:10.1016/j.arr.2011.03.001.
Braak, H., K. Del Tredici, U. Rüb, R.A. De Vos, E.N.J. Steur, and E. Braak. 2003. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol. Aging. 24:197–211.
Braak, H., R.A.I. de Vos, J. Bohl, and K. Del Tredici. 2006. Gastric α-synuclein immunoreactive inclusions in Meissner’s and Auerbach’s plexuses in cases staged for Parkinson’s disease-related brain pathology. Neurosci. Lett. 396:67–72. doi:10.1016/j.neulet.2005.11.012.
Bromberg-Martin, E.S., M. Matsumoto, and O. Hikosaka. 2010. Dopamine in motivational control: rewarding, aversive, and alerting. Neuron. 68:815–834. doi:10.1016/j.neuron.2010.11.022.
Brown, R.G., A. Dittner, L. Findley, and S.C. Wessely. 2005. The Parkinson fatigue scale. Parkinsonism Relat. Disord. 11:49–55. doi:10.1016/j.parkreldis.2004.07.007.

84
Brundin, P., J. Ma, and J.H. Kordower. 2016. How strong is the evidence that Parkinson’s disease is a prion disorder? Curr. Opin. Neurol. 29:459–466. doi:10.1097/WCO.0000000000000349.
Bryk, A.H., and J.R. Wiśniewski. 2017. Quantitative Analysis of Human Red Blood Cell Proteome. J. Proteome Res. 16:2752–2761. doi:10.1021/acs.jproteome.7b00025.
Budnik, V., C. Ruiz-Cañada, and F. Wendler. 2016. Extracellular vesicles round off communication in the nervous system. Nat. Rev. Neurosci. 17:160–172. doi:10.1038/nrn.2015.29.
Butler, B., K. Saha, T. Rana, J.P. Becker, D. Sambo, P. Davari, J.S. Goodwin, and H. Khoshbouei. 2015. Dopamine Transporter Activity Is Modulated by α-Synuclein. J. Biol. Chem. 290:29542–29554. doi:10.1074/jbc.M115.691592.
Calabresi, P., B. Picconi, A. Tozzi, V. Ghiglieri, and M. Di Filippo. 2014. Direct and indirect pathways of basal ganglia: a critical reappraisal. Nat. Neurosci. 17:1022–1030. doi:10.1038/nn.3743.
Calon, F., M. Morissette, A.H. Rajput, O. Hornykiewicz, P.J. Bédard, and T. Di Paolo. 2003. Changes of GABA receptors and dopamine turnover in the postmortem brains of parkinsonians with levodopa-induced motor complications. Mov. Disord. Off. J. Mov. Disord. Soc. 18:241–253. doi:10.1002/mds.10343.
Camacho, A., and J.E. Dimsdale. 2000. Platelets and psychiatry: lessons learned from old and new studies. Psychosom. Med. 62:326–336.
Candelario, K.M., and D.A. Steindler. 2014. The role of extracellular vesicles in the progression of neurodegenerative disease and cancer. Trends Mol. Med. 20:368–374. doi:10.1016/j.molmed.2014.04.003.
Capriotti, T., and K. Terzakis. 2016. Parkinson Disease. Home Healthc. Now. 34:300–307. doi:10.1097/NHH.0000000000000398.
Caronti, B., G. Tanda, C. Colosimo, S. Ruggieri, C. Calderaro, G. Palladini, F.E. Pontieri, and G. Di Chiara. 1999. Reduced dopamine in peripheral blood lymphocytes in Parkinson’s disease. Neuroreport. 10:2907–2910.
Caudle, W.M. 2015. Occupational exposures and parkinsonism. Handb. Clin. Neurol. 131:225–239. doi:10.1016/B978-0-444-62627-1.00013-5.
Chahine, L.M., M.B. Stern, and A. Chen-Plotkin. 2014. Blood-based biomarkers for Parkinson’s disease. Parkinsonism Relat. Disord. 20 Suppl 1:S99-103. doi:10.1016/S1353-8020(13)70025-7.
Chargaff, E., and R. West. 1946. The biological significance of the thromboplastic protein of blood. J. Biol. Chem. 166:189–197.
Chen, C.-M., J.-L. Liu, Y.-R. Wu, Y.-C. Chen, H.-S. Cheng, M.-L. Cheng, and D.T.-Y. Chiu. 2009. Increased oxidative damage in peripheral blood correlates with severity of Parkinson’s disease. Neurobiol. Dis. 33:429–435. doi:10.1016/j.nbd.2008.11.011.
Cheng, H.-C., C.M. Ulane, and R.E. Burke. 2010. Clinical progression in Parkinson disease and the neurobiology of axons. Ann. Neurol. 67:715–725. doi:10.1002/ana.21995.
Chen-Plotkin, A.S., W.T. Hu, A. Siderowf, D. Weintraub, R. Goldmann Gross, H.I. Hurtig,

85
S.X. Xie, S.E. Arnold, M. Grossman, C.M. Clark, L.M. Shaw, L. McCluskey, L. Elman, V.M. Van Deerlin, V.M.-Y. Lee, H. Soares, and J.Q. Trojanowski. 2011. Plasma epidermal growth factor levels predict cognitive decline in Parkinson disease. Ann. Neurol. 69:655–663. doi:10.1002/ana.22271.
Cho, Y.I., M.P. Mooney, and D.J. Cho. 2008. Hemorheological disorders in diabetes mellitus. J. Diabetes Sci. Technol. 2:1130–1138. doi:10.1177/193229680800200622.
Chu, Y., and J.H. Kordower. 2010. Lewy body pathology in fetal grafts. Ann. N. Y. Acad. Sci. 1184:55–67. doi:10.1111/j.1749-6632.2009.05229.x.
Chung, Y.C., J.Y. Baek, S.R. Kim, H.W. Ko, E. Bok, W.-H. Shin, S.-Y. Won, and B.K. Jin. 2017. Capsaicin prevents degeneration of dopamine neurons by inhibiting glial activation and oxidative stress in the MPTP model of Parkinson’s disease. Exp. Mol. Med. 49:e298. doi:10.1038/emm.2016.159.
Church, W.H., and V.L. Ward. 1994. Uric acid is reduced in the substantia nigra in Parkinson’s disease: effect on dopamine oxidation. Brain Res. Bull. 33:419–425.
Cipriani, S., X. Chen, and M.A. Schwarzschild. 2010. Urate: a novel biomarker of Parkinson’s disease risk, diagnosis and prognosis. Biomark. Med. 4:701–712. doi:10.2217/bmm.10.94.
Cisbani, G., J. Drouin-Ouellet, C. Gibrat, M. Saint-Pierre, M. Lagacé, S. Badrinarayanan, M.H. Lavallée-Bourget, J. Charest, A. Chabrat, L. Boivin, M. Lebel, M. Bousquet, M. Lévesque, and F. Cicchetti. 2015. Cystamine/cysteamine rescues the dopaminergic system and shows neurorestorative properties in an animal model of Parkinson’s disease. Neurobiol. Dis. 82:430–444. doi:10.1016/j.nbd.2015.07.012.
Cocucci, E., G. Racchetti, and J. Meldolesi. 2009. Shedding microvesicles: artefacts no more. Trends Cell Biol. 19:43–51. doi:10.1016/j.tcb.2008.11.003.
Connor, D.E., T. Exner, D.D.F. Ma, and J.E. Joseph. 2010. The majority of circulating platelet-derived microparticles fail to bind annexin V, lack phospholipid-dependent procoagulant activity and demonstrate greater expression of glycoprotein Ib. Thromb. Haemost. 103:1044–1052. doi:10.1160/TH09-09-0644.
Conway, K.A., J.C. Rochet, R.M. Bieganski, and P.T. Lansbury. 2001. Kinetic stabilization of the alpha-synuclein protofibril by a dopamine-alpha-synuclein adduct. Science. 294:1346–1349. doi:10.1126/science.1063522.
Cookson, M.R. 2010. The role of leucine-rich repeat kinase 2 (LRRK2) in Parkinson’s disease. Nat. Rev. Neurosci. 11:791–797. doi:10.1038/nrn2935.
Cooper, C.A., and L.M. Chahine. 2016. Biomarkers in Prodromal Parkinson Disease: a Qualitative Review. J. Int. Neuropsychol. Soc. JINS. 22:956–967. doi:10.1017/S1355617716000503.
Coulombe, K., M. Saint-Pierre, G. Cisbani, I. St-Amour, C. Gibrat, A. Giguère-Rancourt, F. Calon, and F. Cicchetti. 2016. Partial neurorescue effects of DHA following a 6-OHDA lesion of the mouse dopaminergic system. J. Nutr. Biochem. 30:133–142. doi:10.1016/j.jnutbio.2015.11.014.
Cox, J., and M. Mann. 2008. MaxQuant enables high peptide identification rates,

86
individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 26:1367–1372. doi:10.1038/nbt.1511.
Cummings, J.L. 1992. Depression and Parkinson’s disease: a review. Am. J. Psychiatry. 149:443–454. doi:10.1176/ajp.149.4.443.
Damier, P., E.C. Hirsch, Y. Agid, and A.M. Graybiel. 1999. The substantia nigra of the human brain. II. Patterns of loss of dopamine-containing neurons in Parkinson’s disease. Brain J. Neurol. 122 ( Pt 8):1437–1448.
Danzer, K.M., L.R. Kranich, W.P. Ruf, O. Cagsal-Getkin, A.R. Winslow, L. Zhu, C.R. Vanderburg, and P.J. McLean. 2012. Exosomal cell-to-cell transmission of alpha synuclein oligomers. Mol. Neurodegener. 7:42. doi:10.1186/1750-1326-7-42.
Dauer, W., and S. Przedborski. 2003. Parkinson’s disease: mechanisms and models. Neuron. 39:889–909.
Davie, C.A. 2008. A review of Parkinson’s disease. Br. Med. Bull. 86:109–127. doi:10.1093/bmb/ldn013.
Delenclos, M., D.R. Jones, P.J. McLean, and R.J. Uitti. 2016. Biomarkers in Parkinson’s disease: Advances and strategies. Parkinsonism Relat. Disord. 22 Suppl 1:S106-110. doi:10.1016/j.parkreldis.2015.09.048.
Devic, I., H. Hwang, J.S. Edgar, K. Izutsu, R. Presland, C. Pan, D.R. Goodlett, Y. Wang, J. Armaly, V. Tumas, C.P. Zabetian, J.B. Leverenz, M. Shi, and J. Zhang. 2011. Salivary α-synuclein and DJ-1: potential biomarkers for Parkinson’s disease. Brain J. Neurol. 134:e178. doi:10.1093/brain/awr015.
Dickson, D.W., H. Braak, J.E. Duda, C. Duyckaerts, T. Gasser, G.M. Halliday, J. Hardy, J.B. Leverenz, K. Del Tredici, Z.K. Wszolek, and I. Litvan. 2009. Neuropathological assessment of Parkinson’s disease: refining the diagnostic criteria. Lancet Neurol. 8:1150–1157. doi:10.1016/S1474-4422(09)70238-8.
Doppler, K., H.-M. Jentschke, L. Schulmeyer, D. Vadasz, A. Janzen, M. Luster, H. Höffken, G. Mayer, J. Brumberg, J. Booij, T. Musacchio, S. Klebe, E. Sittig-Wiegand, J. Volkmann, C. Sommer, and W.H. Oertel. 2017. Dermal phospho-alpha-synuclein deposits confirm REM sleep behaviour disorder as prodromal Parkinson’s disease. Acta Neuropathol. (Berl.). 133:535–545. doi:10.1007/s00401-017-1684-z.
Duchez, A.-C., L.H. Boudreau, G.S. Naika, J. Bollinger, C. Belleannée, N. Cloutier, B. Laffont, R.E. Mendoza-Villarroel, T. Lévesque, E. Rollet-Labelle, M. Rousseau, I. Allaeys, J.J. Tremblay, P.E. Poubelle, G. Lambeau, M. Pouliot, P. Provost, D. Soulet, M.H. Gelb, and E. Boilard. 2015. Platelet microparticles are internalized in neutrophils via the concerted activity of 12-lipoxygenase and secreted phospholipase A2-IIA. Proc. Natl. Acad. Sci. U. S. A. 112:E3564-3573. doi:10.1073/pnas.1507905112.
Dudas, R.B., G.E. Berrios, and J.R. Hodges. 2005. The Addenbrooke’s cognitive examination (ACE) in the differential diagnosis of early dementias versus affective disorder. Am. J. Geriatr. Psychiatry Off. J. Am. Assoc. Geriatr. Psychiatry. 13:218–226. doi:10.1176/appi.ajgp.13.3.218.
El-Agnaf, O.M.A., S.A. Salem, K.E. Paleologou, M.D. Curran, M.J. Gibson, J.A. Court,

87
M.G. Schlossmacher, and D. Allsop. 2006. Detection of oligomeric forms of alpha-synuclein protein in human plasma as a potential biomarker for Parkinson’s disease. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 20:419–425. doi:10.1096/fj.03-1449com.
Elmore, S. 2007. Apoptosis: a review of programmed cell death. Toxicol. Pathol. 35:495–516. doi:10.1080/01926230701320337.
Emmanouilidou, E., K. Melachroinou, T. Roumeliotis, S.D. Garbis, M. Ntzouni, L.H. Margaritis, L. Stefanis, and K. Vekrellis. 2010. Cell-produced alpha-synuclein is secreted in a calcium-dependent manner by exosomes and impacts neuronal survival. J. Neurosci. Off. J. Soc. Neurosci. 30:6838–6851. doi:10.1523/JNEUROSCI.5699-09.2010.
Erwig, L.-P., and P.M. Henson. 2008. Clearance of apoptotic cells by phagocytes. Cell Death Differ. 15:243–250. doi:10.1038/sj.cdd.4402184.
Factor, S.A., E. Ortof, M.P. Dentinger, R. Mankes, and K.D. Barron. 1994. Platelet morphology in Parkinson’s disease: an electron microscopic study. J. Neurol. Sci. 122:84–89.
Fahn, S., and R.L. Elton. 1987. UPDRS Development Committee. The Unified Parkinson’s Disease Rating Scale. Recent Dev. Park. Dis. 2nd Edn Macmillan Healthc. Inf. Florham Park, NJ:153–163. 293–304.
Fakhree, M.A.A., N. Zijlstra, C.C. Raiss, C.J. Siero, H. Grabmayr, A.R. Bausch, C. Blum, and M.M.A.E. Claessens. 2016. The number of α-synuclein proteins per vesicle gives insights into its physiological function. Sci. Rep. 6:30658. doi:10.1038/srep30658.
Fasano, A., N.P. Visanji, L.W.C. Liu, A.E. Lang, and R.F. Pfeiffer. 2015. Gastrointestinal dysfunction in Parkinson’s disease. Lancet Neurol. 14:625–639. doi:10.1016/S1474-4422(15)00007-1.
Faure, A., U. Haberland, F. Condé, and N. El Massioui. 2005. Lesion to the nigrostriatal dopamine system disrupts stimulus-response habit formation. J. Neurosci. Off. J. Soc. Neurosci. 25:2771–2780. doi:10.1523/JNEUROSCI.3894-04.2005.
Fauvet, B., M.K. Mbefo, M.-B. Fares, C. Desobry, S. Michael, M.T. Ardah, E. Tsika, P. Coune, M. Prudent, N. Lion, D. Eliezer, D.J. Moore, B. Schneider, P. Aebischer, O.M. El-Agnaf, E. Masliah, and H.A. Lashuel. 2012. α-Synuclein in central nervous system and from erythrocytes, mammalian cells, and Escherichia coli exists predominantly as disordered monomer. J. Biol. Chem. 287:15345–15364. doi:10.1074/jbc.M111.318949.
Fearnley, J.M., and A.J. Lees. 1991. Ageing and Parkinson’s disease: substantia nigra regional selectivity. Brain J. Neurol. 114 ( Pt 5):2283–2301.
Ferrarese, C., L. Tremolizzo, M. Rigoldi, G. Sala, B. Begni, L. Brighina, G. Ricci, M.G. Albizzati, R. Piolti, F. Crosti, L. Dalprà, and L. Frattola. 2001. Decreased platelet glutamate uptake and genetic risk factors in patients with Parkinson’s disease. Neurol. Sci. Off. J. Ital. Neurol. Soc. Ital. Soc. Clin. Neurophysiol. 22:65–66.
Ferrarese, C., C. Zoia, N. Pecora, R. Piolti, M. Frigo, G. Bianchi, G. Sala, B. Begni, R. Riva, and L. Frattola. 1999. Reduced platelet glutamate uptake in Parkinson’s disease. J. Neural Transm. Vienna Austria 1996. 106:685–692. doi:10.1007/s007020050189.

88
Ferrer, I., E. Perez, E. Dalfó, and M. Barrachina. 2007. Abnormal levels of prohibitin and ATP synthase in the substantia nigra and frontal cortex in Parkinson’s disease. Neurosci. Lett. 415:205–209. doi:10.1016/j.neulet.2007.01.026.
Folstein, M.F., S.E. Folstein, and P.R. McHugh. 1975. “Mini-mental state”. A practical method for grading the cognitive state of patients for the clinician. J. Psychiatr. Res. 12:189–198.
Fraser, K.B., M.S. Moehle, J.P.L. Daher, P.J. Webber, J.Y. Williams, C.A. Stewart, T.A. Yacoubian, R.M. Cowell, T. Dokland, T. Ye, D. Chen, G.P. Siegal, R.A. Galemmo, E. Tsika, D.J. Moore, D.G. Standaert, K. Kojima, J.A. Mobley, and A.B. West. 2013. LRRK2 secretion in exosomes is regulated by 14-3-3. Hum. Mol. Genet. 22:4988–5000. doi:10.1093/hmg/ddt346.
Frisoni, G.B., M. Boccardi, F. Barkhof, K. Blennow, S. Cappa, K. Chiotis, J.-F. Démonet, V. Garibotto, P. Giannakopoulos, A. Gietl, O. Hansson, K. Herholz, C.R. Jack, F. Nobili, A. Nordberg, H.M. Snyder, M. Ten Kate, A. Varrone, E. Albanese, S. Becker, P. Bossuyt, M.C. Carrillo, C. Cerami, B. Dubois, V. Gallo, E. Giacobini, G. Gold, S. Hurst, A. Lönneborg, K.-O. Lovblad, N. Mattsson, J.-L. Molinuevo, A.U. Monsch, U. Mosimann, A. Padovani, A. Picco, C. Porteri, O. Ratib, L. Saint-Aubert, C. Scerri, P. Scheltens, J.M. Schott, I. Sonni, S. Teipel, P. Vineis, P.J. Visser, Y. Yasui, and B. Winblad. 2017. Strategic roadmap for an early diagnosis of Alzheimer’s disease based on biomarkers. Lancet Neurol. 16:661–676. doi:10.1016/S1474-4422(17)30159-X.
Fuchs, J., A. Tichopad, Y. Golub, M. Munz, K.J. Schweitzer, B. Wolf, D. Berg, J.C. Mueller, and T. Gasser. 2008. Genetic variability in the SNCA gene influences alpha-synuclein levels in the blood and brain. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 22:1327–1334. doi:10.1096/fj.07-9348com.
Fujiwara, H., M. Hasegawa, N. Dohmae, A. Kawashima, E. Masliah, M.S. Goldberg, J. Shen, K. Takio, and T. Iwatsubo. 2002. alpha-Synuclein is phosphorylated in synucleinopathy lesions. Nat. Cell Biol. 4:160–164. doi:10.1038/ncb748.
Ganguly, A., H. Zhang, R. Sharma, S. Parsons, and K.D. Patel. 2012. Isolation of human umbilical vein endothelial cells and their use in the study of neutrophil transmigration under flow conditions. J. Vis. Exp. JoVE. e4032. doi:10.3791/4032.
Garbayo, E., E. Ansorena, and M.J. Blanco-Prieto. 2013. Drug development in Parkinson’s disease: from emerging molecules to innovative drug delivery systems. Maturitas. 76:272–278. doi:10.1016/j.maturitas.2013.05.019.
Gasser, T. 2009. Genomic and proteomic biomarkers for Parkinson disease. Neurology. 72:S27-31. doi:10.1212/WNL.0b013e318198e054.
Gilks, W.P., P.M. Abou-Sleiman, S. Gandhi, S. Jain, A. Singleton, A.J. Lees, K. Shaw, K.P. Bhatia, V. Bonifati, N.P. Quinn, J. Lynch, D.G. Healy, J.L. Holton, T. Revesz, and N.W. Wood. 2005. A common LRRK2 mutation in idiopathic Parkinson’s disease. Lancet Lond. Engl. 365:415–416. doi:10.1016/S0140-6736(05)17830-1.
Giusti, I., S. D’Ascenzo, D. Millimaggi, G. Taraboletti, G. Carta, N. Franceschini, A. Pavan, and V. Dolo. 2008. Cathepsin B mediates the pH-dependent proinvasive

89
activity of tumor-shed microvesicles. Neoplasia N. Y. N. 10:481–488. Goetz, C.G. 2011. The history of Parkinson’s disease: early clinical descriptions and
neurological therapies. Cold Spring Harb. Perspect. Med. 1:a008862. doi:10.1101/cshperspect.a008862.
Goetz, C.G., W. Poewe, O. Rascol, C. Sampaio, G.T. Stebbins, C. Counsell, N. Giladi, R.G. Holloway, C.G. Moore, G.K. Wenning, M.D. Yahr, L. Seidl, and Movement Disorder Society Task Force on Rating Scales for Parkinson’s Disease. 2004. Movement Disorder Society Task Force report on the Hoehn and Yahr staging scale: status and recommendations. Mov. Disord. Off. J. Mov. Disord. Soc. 19:1020–1028. doi:10.1002/mds.20213.
Goetz, C.G., B.C. Tilley, S.R. Shaftman, G.T. Stebbins, S. Fahn, P. Martinez-Martin, W. Poewe, C. Sampaio, M.B. Stern, R. Dodel, B. Dubois, R. Holloway, J. Jankovic, J. Kulisevsky, A.E. Lang, A. Lees, S. Leurgans, P.A. LeWitt, D. Nyenhuis, C.W. Olanow, O. Rascol, A. Schrag, J.A. Teresi, J.J. van Hilten, N. LaPelle, and Movement Disorder Society UPDRS Revision Task Force. 2008. Movement Disorder Society-sponsored revision of the Unified Parkinson’s Disease Rating Scale (MDS-UPDRS): scale presentation and clinimetric testing results. Mov. Disord. Off. J. Mov. Disord. Soc. 23:2129–2170. doi:10.1002/mds.22340.
Golbe, L.I., G. Di Iorio, G. Sanges, A.M. Lazzarini, S. La Sala, V. Bonavita, and R.C. Duvoisin. 1996. Clinical genetic analysis of Parkinson’s disease in the Contursi kindred. Ann. Neurol. 40:767–775. doi:10.1002/ana.410400513.
Goldstein, D.S., C. Holmes, and Y. Sharabi. 2012. Cerebrospinal fluid biomarkers of central catecholamine deficiency in Parkinson’s disease and other synucleinopathies. Brain J. Neurol. 135:1900–1913. doi:10.1093/brain/aws055.
Gowert, N.S., L. Donner, M. Chatterjee, Y.S. Eisele, S.T. Towhid, P. Münzer, B. Walker, I. Ogorek, O. Borst, M. Grandoch, M. Schaller, J.W. Fischer, M. Gawaz, S. Weggen, F. Lang, M. Jucker, and M. Elvers. 2014. Blood platelets in the progression of Alzheimer’s disease. PloS One. 9:e90523. doi:10.1371/journal.pone.0090523.
Grandinetti, A., D.M. Morens, D. Reed, and D. MacEachern. 1994. Prospective study of cigarette smoking and the risk of developing idiopathic Parkinson’s disease. Am. J. Epidemiol. 139:1129–1138.
Greenamyre, J.T., and T.G. Hastings. 2004. Biomedicine. Parkinson’s--divergent causes, convergent mechanisms. Science. 304:1120–1122. doi:10.1126/science.1098966.
Grey, M., C.J. Dunning, R. Gaspar, C. Grey, P. Brundin, E. Sparr, and S. Linse. 2015. Acceleration of α-synuclein aggregation by exosomes. J. Biol. Chem. 290:2969–2982. doi:10.1074/jbc.M114.585703.
Grozdanov, V., C. Bliederhaeuser, W.P. Ruf, V. Roth, K. Fundel-Clemens, L. Zondler, D. Brenner, A. Martin-Villalba, B. Hengerer, J. Kassubek, A.C. Ludolph, J.H. Weishaupt, and K.M. Danzer. 2014. Inflammatory dysregulation of blood monocytes in Parkinson’s disease patients. Acta Neuropathol. (Berl.). 128:651–663. doi:10.1007/s00401-014-1345-4.

90
Gruenberg, J., and H. Stenmark. 2004. The biogenesis of multivesicular endosomes. Nat. Rev. Mol. Cell Biol. 5:317–323. doi:10.1038/nrm1360.
György, B., T.G. Szabó, M. Pásztói, Z. Pál, P. Misják, B. Aradi, V. László, E. Pállinger, E. Pap, A. Kittel, G. Nagy, A. Falus, and E.I. Buzás. 2011. Membrane vesicles, current state-of-the-art: emerging role of extracellular vesicles. Cell. Mol. Life Sci. CMLS. 68:2667–2688. doi:10.1007/s00018-011-0689-3.
Hall, J., S. Prabhakar, L. Balaj, C.P. Lai, R.A. Cerione, and X.O. Breakefield. 2016. Delivery of Therapeutic Proteins via Extracellular Vesicles: Review and Potential Treatments for Parkinson’s Disease, Glioma, and Schwannoma. Cell. Mol. Neurobiol. 36:417–427. doi:10.1007/s10571-015-0309-0.
Harding, C., J. Heuser, and P. Stahl. 1983. Receptor-mediated endocytosis of transferrin and recycling of the transferrin receptor in rat reticulocytes. J. Cell Biol. 97:329–339.
Hardy, J., M.R. Cookson, and A. Singleton. 2003. Genes and parkinsonism. Lancet Neurol. 2:221–228.
Harrell, L.E., D. Marson, A. Chatterjee, and J.A. Parrish. 2000. The Severe Mini-Mental State Examination: a new neuropsychologic instrument for the bedside assessment of severely impaired patients with Alzheimer disease. Alzheimer Dis. Assoc. Disord. 14:168–175.
Harrower, T.P., A.W. Michell, and R.A. Barker. 2005. Lewy bodies in Parkinson’s disease: protectors or perpetrators? Exp. Neurol. 195:1–6. doi:10.1016/j.expneurol.2005.06.002.
Hatano, T., S. Saiki, A. Okuzumi, R.P. Mohney, and N. Hattori. 2016. Identification of novel biomarkers for Parkinson’s disease by metabolomic technologies. J. Neurol. Neurosurg. Psychiatry. 87:295–301. doi:10.1136/jnnp-2014-309676.
Havlis, J., H. Thomas, M. Sebela, and A. Shevchenko. 2003. Fast-response proteomics by accelerated in-gel digestion of proteins. Anal. Chem. 75:1300–1306.
Hawkes, C.M. 1995. Diagnosis and treatment of Parkinson’s disease. Anosmia is a common finding. BMJ. 310:1668.
Hayes, V., I. Johnston, G.M. Arepally, S.E. McKenzie, D.B. Cines, L. Rauova, and M. Poncz. 2017. Endothelial antigen assembly leads to thrombotic complications in heparin-induced thrombocytopenia. J. Clin. Invest. 127:1090–1098. doi:10.1172/JCI90958.
Healy, D.G., P.M. Abou-Sleiman, and N.W. Wood. 2004. PINK, PANK, or PARK? A clinicians’ guide to familial parkinsonism. Lancet Neurol. 3:652–662. doi:10.1016/S1474-4422(04)00905-6.
Hedrich, K., C. Eskelson, B. Wilmot, K. Marder, J. Harris, J. Garrels, H. Meija-Santana, P. Vieregge, H. Jacobs, S.B. Bressman, A.E. Lang, M. Kann, G. Abbruzzese, P. Martinelli, E. Schwinger, L.J. Ozelius, P.P. Pramstaller, C. Klein, and P. Kramer. 2004. Distribution, type, and origin of Parkin mutations: review and case studies. Mov. Disord. Off. J. Mov. Disord. Soc. 19:1146–1157. doi:10.1002/mds.20234.
Helferich, A.M., W.P. Ruf, V. Grozdanov, A. Freischmidt, M.S. Feiler, L. Zondler, A.C. Ludolph, P.J. McLean, J.H. Weishaupt, and K.M. Danzer. 2015. α-synuclein interacts

91
with SOD1 and promotes its oligomerization. Mol. Neurodegener. 10:66. doi:10.1186/s13024-015-0062-3.
Hernán, M.A., S.M. Zhang, A.M. Rueda-deCastro, G.A. Colditz, F.E. Speizer, and A. Ascherio. 2001. Cigarette smoking and the incidence of Parkinson’s disease in two prospective studies. Ann. Neurol. 50:780–786.
Herrera, A.J., M. Tomás-Camardiel, J.L. Venero, J. Cano, and A. Machado. 2005. Inflammatory process as a determinant factor for the degeneration of substantia nigra dopaminergic neurons. J. Neural Transm. Vienna Austria 1996. 112:111–119. doi:10.1007/s00702-004-0121-3.
Hindle, J.V. 2010. Ageing, neurodegeneration and Parkinson’s disease. Age Ageing. 39:156–161. doi:10.1093/ageing/afp223.
Ho, D.H., S. Yi, H. Seo, I. Son, and W. Seol. 2014. Increased DJ-1 in urine exosome of Korean males with Parkinson’s disease. BioMed Res. Int. 2014:704678. doi:10.1155/2014/704678.
Hoehn, M.M., and M.D. Yahr. 1967. Parkinsonism: onset, progression and mortality. Neurology. 17:427–442.
Höglinger, G.U., J. Kassubek, I. Csoti, R. Ehret, H. Herbst, I. Wellach, J. Winkler, and W.H. Jost. 2017. Differentiation of atypical Parkinson syndromes. J. Neural Transm. 124:997–1004. doi:10.1007/s00702-017-1700-4.
Holdorff, B., A.M. Rodrigues e Silva, and R. Dodel. 2013. Centenary of Lewy bodies (1912-2012). J. Neural Transm. Vienna Austria 1996. 120:509–516. doi:10.1007/s00702-013-0984-2.
Hong, Z., M. Shi, K.A. Chung, J.F. Quinn, E.R. Peskind, D. Galasko, J. Jankovic, C.P. Zabetian, J.B. Leverenz, G. Baird, T.J. Montine, A.M. Hancock, H. Hwang, C. Pan, J. Bradner, U.J. Kang, P.H. Jensen, and J. Zhang. 2010. DJ-1 and alpha-synuclein in human cerebrospinal fluid as biomarkers of Parkinson’s disease. Brain J. Neurol. 133:713–726. doi:10.1093/brain/awq008.
Hristov, M., W. Erl, S. Linder, and P.C. Weber. 2004. Apoptotic bodies from endothelial cells enhance the number and initiate the differentiation of human endothelial progenitor cells in vitro. Blood. 104:2761–2766. doi:10.1182/blood-2003-10-3614.
Hsiao, I.-T., Y.-H. Weng, C.-J. Hsieh, W.-Y. Lin, S.-P. Wey, M.-P. Kung, T.-C. Yen, C.-S. Lu, and K.-J. Lin. 2014. Correlation of Parkinson disease severity and 18F-DTBZ positron emission tomography. JAMA Neurol. 71:758–766. doi:10.1001/jamaneurol.2014.290.
Hubble, J.P., T. Cao, R.E. Hassanein, J.S. Neuberger, and W.C. Koller. 1993. Risk factors for Parkinson’s disease. Neurology. 43:1693–1697.
Hughes, A.J., S.E. Daniel, Y. Ben-Shlomo, and A.J. Lees. 2002. The accuracy of diagnosis of parkinsonian syndromes in a specialist movement disorder service. Brain J. Neurol. 125:861–870.
Hunot, S., and E.C. Hirsch. 2003. Neuroinflammatory processes in Parkinson’s disease. Ann. Neurol. 53 Suppl 3:S49-58-60. doi:10.1002/ana.10481.

92
Hurley, J.H., E. Boura, L.-A. Carlson, and B. Różycki. 2010. Membrane budding. Cell. 143:875–887. doi:10.1016/j.cell.2010.11.030.
Husain, M., R. Shukla, M. Dikshit, P.K. Maheshwari, D. Nag, R.C. Srimal, P.K. Seth, and V.K. Khanna. 2009. Altered platelet monoamine oxidase-B activity in idiopathic Parkinson’s disease. Neurochem. Res. 34:1427–1432. doi:10.1007/s11064-009-9929-4.
Ihara, Y., M. Chuda, S. Kuroda, and T. Hayabara. 1999. Hydroxyl radical and superoxide dismutase in blood of patients with Parkinson’s disease: relationship to clinical data. J. Neurol. Sci. 170:90–95.
Isaias, I.U., P. Trujillo, P. Summers, G. Marotta, L. Mainardi, G. Pezzoli, L. Zecca, and A. Costa. 2016. Neuromelanin Imaging and Dopaminergic Loss in Parkinson’s Disease. Front. Aging Neurosci. 8:196. doi:10.3389/fnagi.2016.00196.
Izquierdo-Useros, N., M. Naranjo-Gómez, J. Archer, S.C. Hatch, I. Erkizia, J. Blanco, F.E. Borràs, M.C. Puertas, J.H. Connor, M.T. Fernández-Figueras, L. Moore, B. Clotet, S. Gummuluru, and J. Martinez-Picado. 2009. Capture and transfer of HIV-1 particles by mature dendritic cells converges with the exosome-dissemination pathway. Blood. 113:2732–2741. doi:10.1182/blood-2008-05-158642.
Jankovic, J., M. McDermott, J. Carter, S. Gauthier, C. Goetz, L. Golbe, S. Huber, W. Koller, C. Olanow, I. Shoulson, M. Stern, C. Tanner, W. Weiner, and P.S. Group. 1990. Variable expression of Parkinson’s disease A base‐line analysis of the DAT ATOP cohort. Neurology. 40:1529–1529. doi:10.1212/WNL.40.10.1529.
Johnstone, R.M., M. Adam, J.R. Hammond, L. Orr, and C. Turbide. 1987. Vesicle formation during reticulocyte maturation. Association of plasma membrane activities with released vesicles (exosomes). J. Biol. Chem. 262:9412–9420.
Karihtala, P., and Y. Soini. 2007. Reactive oxygen species and antioxidant mechanisms in human tissues and their relation to malignancies. APMIS Acta Pathol. Microbiol. Immunol. Scand. 115:81–103. doi:10.1111/j.1600-0463.2007.apm_514.x.
Katzenschlager, R., A. Schrag, A. Evans, A. Manson, C.B. Carroll, D. Ottaviani, A.J. Lees, and J. Hobart. 2007. Quantifying the impact of dyskinesias in PD: the PDYS-26: a patient-based outcome measure. Neurology. 69:555–563. doi:10.1212/01.wnl.0000266669.18308.af.
Kawamata, H., P.J. McLean, N. Sharma, and B.T. Hyman. 2001. Interaction of alpha-synuclein and synphilin-1: effect of Parkinson’s disease-associated mutations. J. Neurochem. 77:929–934.
Kim, S., B.S. Jeon, C. Heo, P.S. Im, T.-B. Ahn, J.-H. Seo, H.-S. Kim, C.H. Park, S.H. Choi, S.-H. Cho, W.J. Lee, and Y.-H. Suh. 2004. Alpha-synuclein induces apoptosis by altered expression in human peripheral lymphocyte in Parkinson’s disease. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 18:1615–1617. doi:10.1096/fj.04-1917fje.
Koller, W., B. Vetere-Overfield, C. Gray, C. Alexander, T. Chin, J. Dolezal, R. Hassanein, and C. Tanner. 1990. Environmental risk factors in Parkinson’s disease. Neurology. 40:1218–1221.
Kong, S.M.Y., B.K.K. Chan, J.-S. Park, K.J. Hill, J.B. Aitken, L. Cottle, H. Farghaian,

93
A.R. Cole, P.A. Lay, C.M. Sue, and A.A. Cooper. 2014. Parkinson’s disease-linked human PARK9/ATP13A2 maintains zinc homeostasis and promotes α-Synuclein externalization via exosomes. Hum. Mol. Genet. 23:2816–2833. doi:10.1093/hmg/ddu099.
Kordower, J.H., Y. Chu, R.A. Hauser, T.B. Freeman, and C.W. Olanow. 2008. Lewy body-like pathology in long-term embryonic nigral transplants in Parkinson’s disease. Nat. Med. 14:504–506. doi:10.1038/nm1747.
Kordower, J.H., C.W. Olanow, H.B. Dodiya, Y. Chu, T.G. Beach, C.H. Adler, G.M. Halliday, and R.T. Bartus. 2013. Disease duration and the integrity of the nigrostriatal system in Parkinson’s disease. Brain J. Neurol. 136:2419–2431. doi:10.1093/brain/awt192.
Kravitz, A.V., B.S. Freeze, P.R.L. Parker, K. Kay, M.T. Thwin, K. Deisseroth, and A.C. Kreitzer. 2010. Regulation of parkinsonian motor behaviours by optogenetic control of basal ganglia circuitry. Nature. 466:622–626. doi:10.1038/nature09159.
Krüger, R., W. Kuhn, T. Müller, D. Woitalla, M. Graeber, S. Kösel, H. Przuntek, J.T. Epplen, L. Schöls, and O. Riess. 1998. Ala30Pro mutation in the gene encoding alpha-synuclein in Parkinson’s disease. Nat. Genet. 18:106–108. doi:10.1038/ng0298-106.
Kuhn, D.M., R.E. Arthur, D.M. Thomas, and L.A. Elferink. 1999. Tyrosine hydroxylase is inactivated by catechol-quinones and converted to a redox-cycling quinoprotein: possible relevance to Parkinson’s disease. J. Neurochem. 73:1309–1317.
Kuhn, W., R. Winkel, D. Woitalla, S. Meves, H. Przuntek, and T. Müller. 1998. High prevalence of parkinsonism after occupational exposure to lead-sulfate batteries. Neurology. 50:1885–1886.
Kunadt, M., K. Eckermann, A. Stuendl, J. Gong, B. Russo, K. Strauss, S. Rai, S. Kügler, L. Falomir Lockhart, M. Schwalbe, P. Krumova, L.M.A. Oliveira, M. Bähr, W. Möbius, J. Levin, A. Giese, N. Kruse, B. Mollenhauer, R. Geiss-Friedlander, A.C. Ludolph, A. Freischmidt, M.S. Feiler, K.M. Danzer, M. Zweckstetter, T.M. Jovin, M. Simons, J.H. Weishaupt, and A. Schneider. 2015. Extracellular vesicle sorting of α-Synuclein is regulated by sumoylation. Acta Neuropathol. (Berl.). 129:695–713. doi:10.1007/s00401-015-1408-1.
Lacroix, R., C. Judicone, P. Poncelet, S. Robert, L. Arnaud, J. Sampol, and F. Dignat-George. 2012. Impact of pre-analytical parameters on the measurement of circulating microparticles: towards standardization of protocol. J. Thromb. Haemost. JTH. 10:437–446. doi:10.1111/j.1538-7836.2011.04610.x.
Lai, B.C.L., S.A. Marion, K. Teschke, and J.K.C. Tsui. 2002. Occupational and environmental risk factors for Parkinson’s disease. Parkinsonism Relat. Disord. 8:297–309.
Lai, F.W., B.D. Lichty, and D.M.E. Bowdish. 2015. Microvesicles: ubiquitous contributors to infection and immunity. J. Leukoc. Biol. 97:237–245. doi:10.1189/jlb.3RU0513-292RR.
Lang, A.E. 2011. A critical appraisal of the premotor symptoms of Parkinson’s disease:

94
potential usefulness in early diagnosis and design of neuroprotective trials. Mov. Disord. Off. J. Mov. Disord. Soc. 26:775–783. doi:10.1002/mds.23609.
Langston, J.W., P. Ballard, J.W. Tetrud, and I. Irwin. 1983. Chronic Parkinsonism in humans due to a product of meperidine-analog synthesis. Science. 219:979–980.
Lapointe, N., M. St-Hilaire, M.-G. Martinoli, J. Blanchet, P. Gould, C. Rouillard, and F. Cicchetti. 2004. Rotenone induces non-specific central nervous system and systemic toxicity. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 18:717–719. doi:10.1096/fj.03-0677fje.
Lashuel, H.A., C.R. Overk, A. Oueslati, and E. Masliah. 2012. The many faces of α-synuclein: from structure and toxicity to therapeutic target. Nat. Rev. Neurosci. 14:38–48. doi:10.1038/nrn3406.
de Lau, L.M.L., and M.M.B. Breteler. 2006. Epidemiology of Parkinson’s disease. Lancet Neurol. 5:525–535. doi:10.1016/S1474-4422(06)70471-9.
Laurens, B., S. Vergnet, M.C. Lopez, A. Foubert-Samier, F. Tison, P.-O. Fernagut, and W.G. Meissner. 2017. Multiple System Atrophy - State of the Art. Curr. Neurol. Neurosci. Rep. 17:41. doi:10.1007/s11910-017-0751-0.
Lee, C.S., J.H. Han, Y.Y. Jang, J.H. Song, and E.S. Han. 2002. Differential effect of catecholamines and MPP(+) on membrane permeability in brain mitochondria and cell viability in PC12 cells. Neurochem. Int. 40:361–369.
Lee, P.H., G. Lee, H.J. Park, O.Y. Bang, I.S. Joo, and K. Huh. 2006. The plasma alpha-synuclein levels in patients with Parkinson’s disease and multiple system atrophy. J. Neural Transm. Vienna Austria 1996. 113:1435–1439. doi:10.1007/s00702-005-0427-9.
Lee, S.-J., H. Jeon, and K.V. Kandror. 2008. Alpha-synuclein is localized in a subpopulation of rat brain synaptic vesicles. Acta Neurobiol. Exp. (Warsz.). 68:509–515.
Leroyer, A.S., A. Tedgui, and C.M. Boulanger. 2008. Role of microparticles in atherothrombosis. J. Intern. Med. 263:528–537. doi:10.1111/j.1365-2796.2008.01957.x.
Lesage, S., and A. Brice. 2009. Parkinson’s disease: from monogenic forms to genetic susceptibility factors. Hum. Mol. Genet. 18:R48-59. doi:10.1093/hmg/ddp012.
Li, Q.-X., B.C.V. Campbell, C.A. McLean, D. Thyagarajan, W.-P. Gai, R.M. Kapsa, K. Beyreuther, C.L. Masters, and J.G. Culvenor. 2002a. Platelet alpha- and gamma-synucleins in Parkinson’s disease and normal control subjects. J. Alzheimers Dis. JAD. 4:309–315.
Li, Q.-X., B.C.V. Campbell, C.A. McLean, D. Thyagarajan, W.-P. Gai, R.M. Kapsa, K. Beyreuther, C.L. Masters, and J.G. Culvenor. 2002b. Platelet alpha- and gamma-synucleins in Parkinson’s disease and normal control subjects. J. Alzheimers Dis. JAD. 4:309–315.
Li, Q.-X., S.S. Mok, K.M. Laughton, C.A. McLean, R. Cappai, C.L. Masters, J.G. Culvenor, and M.K. Horne. 2007. Plasma alpha-synuclein is decreased in subjects with

95
Parkinson’s disease. Exp. Neurol. 204:583–588. doi:10.1016/j.expneurol.2006.12.006. Li, Y., S. Schrodi, C. Rowland, K. Tacey, J. Catanese, and A. Grupe. 2006. Genetic
evidence for ubiquitin-specific proteases USP24 and USP40 as candidate genes for late-onset Parkinson disease. Hum. Mutat. 27:1017–1023. doi:10.1002/humu.20382.
Lotharius, J., and P. Brundin. 2002. Impaired dopamine storage resulting from alpha-synuclein mutations may contribute to the pathogenesis of Parkinson’s disease. Hum. Mol. Genet. 11:2395–2407.
Luan, H., L.-F. Liu, Z. Tang, M. Zhang, K.-K. Chua, J.-X. Song, V.C.T. Mok, M. Li, and Z. Cai. 2015. Comprehensive urinary metabolomic profiling and identification of potential noninvasive marker for idiopathic Parkinson’s disease. Sci. Rep. 5:13888. doi:10.1038/srep13888.
Lücking, C.B., V. Bonifati, M. Periquet, N. Vanacore, A. Brice, and G. Meco. 2001. Pseudo-dominant inheritance and exon 2 triplication in a family with parkin gene mutations. Neurology. 57:924–927.
Lücking, C.B., A. Dürr, V. Bonifati, J. Vaughan, G. De Michele, T. Gasser, B.S. Harhangi, G. Meco, P. Denèfle, N.W. Wood, Y. Agid, A. Brice, French Parkinson’s Disease Genetics Study Group, and European Consortium on Genetic Susceptibility in Parkinson’s Disease. 2000. Association between early-onset Parkinson’s disease and mutations in the parkin gene. N. Engl. J. Med. 342:1560–1567. doi:10.1056/NEJM200005253422103.
Ma, W., H.-T. Liu, and Y.-T. Long. 2015. Monitoring Dopamine Quinone-Induced Dopaminergic Neurotoxicity Using Dopamine Functionalized Quantum Dots. ACS Appl. Mater. Interfaces. 7:14352–14358. doi:10.1021/acsami.5b03044.
Maas, S.L.N., X.O. Breakefield, and A.M. Weaver. 2017. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 27:172–188. doi:10.1016/j.tcb.2016.11.003.
Machado, V., T. Zöller, A. Attaai, and B. Spittau. 2016. Microglia-Mediated Neuroinflammation and Neurotrophic Factor-Induced Protection in the MPTP Mouse Model of Parkinson’s Disease-Lessons from Transgenic Mice. Int. J. Mol. Sci. 17. doi:10.3390/ijms17020151.
MacLeod, D., J. Dowman, R. Hammond, T. Leete, K. Inoue, and A. Abeliovich. 2006. The familial Parkinsonism gene LRRK2 regulates neurite process morphology. Neuron. 52:587–593. doi:10.1016/j.neuron.2006.10.008.
Maere, S., K. Heymans, and M. Kuiper. 2005. BiNGO: a Cytoscape plugin to assess overrepresentation of gene ontology categories in biological networks. Bioinforma. Oxf. Engl. 21:3448–3449. doi:10.1093/bioinformatics/bti551.
Manyam, B.V. 1990. Paralysis agitans and levodopa in “Ayurveda”: ancient Indian medical treatise. Mov. Disord. Off. J. Mov. Disord. Soc. 5:47–48. doi:10.1002/mds.870050112.
Martínez, M.C., and J.M. Freyssinet. 2001. Deciphering the plasma membrane hallmarks of apoptotic cells: phosphatidylserine transverse redistribution and calcium entry. BMC Cell Biol. 2:20.

96
Martínez-Martín, P., M.J. Forjaz, E. Cubo, B. Frades, J. de Pedro Cuesta, and ELEP Project Members. 2006. Global versus factor-related impression of severity in Parkinson’s disease: a new clinimetric index (CISI-PD). Mov. Disord. Off. J. Mov. Disord. Soc. 21:208–214. doi:10.1002/mds.20697.
Martínez-Martín, P., C. Rodríguez-Blázquez, M.J. Forjaz, J. de Pedro, and Spanish-American Longitudinal PD Patient Study Group. 2009. The Clinical Impression of Severity Index for Parkinson’s Disease: international validation study. Mov. Disord. Off. J. Mov. Disord. Soc. 24:211–217. doi:10.1002/mds.22320.
Martínez-Martín, P., C. Rodríguez-Blázquez, null Mario Alvarez, T. Arakaki, V.C. Arillo, P. Chaná, W. Fernández, N. Garretto, J.C. Martínez-Castrillo, M. Rodríguez-Violante, M. Serrano-Dueñas, D. Ballesteros, J.M. Rojo-Abuin, K.R. Chaudhuri, and M. Merello. 2015. Parkinson’s disease severity levels and MDS-Unified Parkinson’s Disease Rating Scale. Parkinsonism Relat. Disord. 21:50–54. doi:10.1016/j.parkreldis.2014.10.026.
Massano, J., and K.P. Bhatia. 2012. Clinical approach to Parkinson’s disease: features, diagnosis, and principles of management. Cold Spring Harb. Perspect. Med. 2:a008870. doi:10.1101/cshperspect.a008870.
Mathuranath, P.S., P.J. Nestor, G.E. Berrios, W. Rakowicz, and J.R. Hodges. 2000. A brief cognitive test battery to differentiate Alzheimer’s disease and frontotemporal dementia. Neurology. 55:1613–1620.
Mayeux, R. 2004. Biomarkers: potential uses and limitations. NeuroRx J. Am. Soc. Exp. Neurother. 1:182–188. doi:10.1602/neurorx.1.2.182.
McGeer, P.L., S. Itagaki, B.E. Boyes, and E.G. McGeer. 1988. Reactive microglia are positive for HLA-DR in the substantia nigra of Parkinson’s and Alzheimer’s disease brains. Neurology. 38:1285–1291.
McKeith, I.G., D.W. Dickson, J. Lowe, M. Emre, J.T. O’Brien, H. Feldman, J. Cummings, J.E. Duda, C. Lippa, E.K. Perry, D. Aarsland, H. Arai, C.G. Ballard, B. Boeve, D.J. Burn, D. Costa, T. Del Ser, B. Dubois, D. Galasko, S. Gauthier, C.G. Goetz, E. Gomez-Tortosa, G. Halliday, L.A. Hansen, J. Hardy, T. Iwatsubo, R.N. Kalaria, D. Kaufer, R.A. Kenny, A. Korczyn, K. Kosaka, V.M.Y. Lee, A. Lees, I. Litvan, E. Londos, O.L. Lopez, S. Minoshima, Y. Mizuno, J.A. Molina, E.B. Mukaetova-Ladinska, F. Pasquier, R.H. Perry, J.B. Schulz, J.Q. Trojanowski, M. Yamada, and Consortium on DLB. 2005. Diagnosis and management of dementia with Lewy bodies: third report of the DLB Consortium. Neurology. 65:1863–1872. doi:10.1212/01.wnl.0000187889.17253.b1.
Melachroinou, K., M. Xilouri, E. Emmanouilidou, R. Masgrau, P. Papazafiri, L. Stefanis, and K. Vekrellis. 2013. Deregulation of calcium homeostasis mediates secreted α-synuclein-induced neurotoxicity. Neurobiol. Aging. 34:2853–2865. doi:10.1016/j.neurobiolaging.2013.06.006.
Mila, S., A.G. Albo, D. Corpillo, S. Giraudo, M. Zibetti, E.M. Bucci, L. Lopiano, and M. Fasano. 2009. Lymphocyte proteomics of Parkinson’s disease patients reveals

97
cytoskeletal protein dysregulation and oxidative stress. Biomark. Med. 3:117–128. doi:10.2217/bmm.09.4.
Miller, D.B., and J.P. O’Callaghan. 2015. Biomarkers of Parkinson’s disease: Present and future. Metabolism. 64:S40–S46. doi:10.1016/j.metabol.2014.10.030.
Minetti, G., A. Ciana, and C. Balduini. 2004. Differential sorting of tyrosine kinases and phosphotyrosine phosphatases acting on band 3 during vesiculation of human erythrocytes. Biochem. J. 377:489–497. doi:10.1042/BJ20031401.
Mioshi, E., K. Dawson, J. Mitchell, R. Arnold, and J.R. Hodges. 2006. The Addenbrooke’s Cognitive Examination Revised (ACE-R): a brief cognitive test battery for dementia screening. Int. J. Geriatr. Psychiatry. 21:1078–1085. doi:10.1002/gps.1610.
Molloy, D.W., E. Alemayehu, and R. Roberts. 1991. Reliability of a Standardized Mini-Mental State Examination compared with the traditional Mini-Mental State Examination. Am. J. Psychiatry. 148:102–105. doi:10.1176/ajp.148.1.102.
Mondello, S., R. Constantinescu, H. Zetterberg, U. Andreasson, B. Holmberg, and A. Jeromin. 2014. CSF α-synuclein and UCH-L1 levels in Parkinson’s disease and atypical parkinsonian disorders. Parkinsonism Relat. Disord. 20:382–387. doi:10.1016/j.parkreldis.2014.01.011.
Nakai, M., M. Fujita, M. Waragai, S. Sugama, J. Wei, H. Akatsu, C. Ohtaka-Maruyama, H. Okado, and M. Hashimoto. 2007. Expression of alpha-synuclein, a presynaptic protein implicated in Parkinson’s disease, in erythropoietic lineage. Biochem. Biophys. Res. Commun. 358:104–110. doi:10.1016/j.bbrc.2007.04.108.
Nalls, M.A., M.F. Keller, D.G. Hernandez, L. Chen, D.J. Stone, A.B. Singleton, and Parkinson’s Progression Marker Initiative (PPMI) investigators. 2016. Baseline genetic associations in the Parkinson’s Progression Markers Initiative (PPMI). Mov. Disord. Off. J. Mov. Disord. Soc. 31:79–85. doi:10.1002/mds.26374.
Narendra, D., A. Tanaka, D.-F. Suen, and R.J. Youle. 2008. Parkin is recruited selectively to impaired mitochondria and promotes their autophagy. J. Cell Biol. 183:795–803. doi:10.1083/jcb.200809125.
Ngim, C.H., and G. Devathasan. 1989. Epidemiologic study on the association between body burden mercury level and idiopathic Parkinson’s disease. Neuroepidemiology. 8:128–141.
Nguyen, D.B., T.B.T. Ly, M.C. Wesseling, M. Hittinger, A. Torge, A. Devitt, Y. Perrie, and I. Bernhardt. 2016. Characterization of Microvesicles Released from Human Red Blood Cells. Cell. Physiol. Biochem. Int. J. Exp. Cell. Physiol. Biochem. Pharmacol. 38:1085–1099. doi:10.1159/000443059.
Nichols, W.C., N. Pankratz, D. Hernandez, C. Paisán-Ruíz, S. Jain, C.A. Halter, V.E. Michaels, T. Reed, A. Rudolph, C.W. Shults, A. Singleton, T. Foroud, and Parkinson Study Group-PROGENI investigators. 2005. Genetic screening for a single common LRRK2 mutation in familial Parkinson’s disease. Lancet Lond. Engl. 365:410–412. doi:10.1016/S0140-6736(05)17828-3.
Nikam, S., P. Nikam, S.K. Ahaley, and A.V. Sontakke. 2009. Oxidative stress in

98
Parkinson’s disease. Indian J. Clin. Biochem. IJCB. 24:98–101. doi:10.1007/s12291-009-0017-y.
Nonaka, T., T. Iwatsubo, and M. Hasegawa. 2005. Ubiquitination of alpha-synuclein. Biochemistry (Mosc.). 44:361–368. doi:10.1021/bi0485528.
Obeid, R., A. Schadt, U. Dillmann, P. Kostopoulos, K. Fassbender, and W. Herrmann. 2009. Methylation status and neurodegenerative markers in Parkinson disease. Clin. Chem. 55:1852–1860. doi:10.1373/clinchem.2009.125021.
O’Callaghan, J.P., D.B. Miller, and J.F. Reinhard. 1990. Characterization of the origins of astrocyte response to injury using the dopaminergic neurotoxicant, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine. Brain Res. 521:73–80.
Olzmann, J.A., J.R. Bordelon, E.C. Muly, H.D. Rees, A.I. Levey, L. Li, and L.-S. Chin. 2007. Selective enrichment of DJ-1 protein in primate striatal neuronal processes: implications for Parkinson’s disease. J. Comp. Neurol. 500:585–599. doi:10.1002/cne.21191.
Pan, B.T., K. Teng, C. Wu, M. Adam, and R.M. Johnstone. 1985. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J. Cell Biol. 101:942–948.
Pangman, V.C., J. Sloan, and L. Guse. 2000. An examination of psychometric properties of the mini-mental state examination and the standardized mini-mental state examination: implications for clinical practice. Appl. Nurs. Res. ANR. 13:209–213. doi:10.1053/apnr.2000.9231.
Parkinson, J. 2002. An essay on the shaking palsy. 1817. J. Neuropsychiatry Clin. Neurosci. 14:223–236; discussion 222. doi:10.1176/jnp.14.2.223.
Parkinson Progression Marker Initiative. 2011. The Parkinson Progression Marker Initiative (PPMI). Prog. Neurobiol. 95:629–635. doi:10.1016/j.pneurobio.2011.09.005.
Parnetti, L., A. Castrioto, D. Chiasserini, E. Persichetti, N. Tambasco, O. El-Agnaf, and P. Calabresi. 2013. Cerebrospinal fluid biomarkers in Parkinson disease. Nat. Rev. Neurol. 9:131–140. doi:10.1038/nrneurol.2013.10.
Pavese, N., and D.J. Brooks. 2009. Imaging neurodegeneration in Parkinson’s disease. Biochim. Biophys. Acta. 1792:722–729. doi:10.1016/j.bbadis.2008.10.003.
Pellicano, C., D. Benincasa, V. Pisani, F.R. Buttarelli, M. Giovannelli, and F.E. Pontieri. 2007. Prodromal non-motor symptoms of Parkinson’s disease. Neuropsychiatr. Dis. Treat. 3:145–152.
Penney, J.B., and A.B. Young. 1986. Striatal inhomogeneities and basal ganglia function. Mov. Disord. Off. J. Mov. Disord. Soc. 1:3–15. doi:10.1002/mds.870010102.
Pepe, M.S., R. Etzioni, Z. Feng, J.D. Potter, M.L. Thompson, M. Thornquist, M. Winget, and Y. Yasui. 2001. Phases of biomarker development for early detection of cancer. J. Natl. Cancer Inst. 93:1054–1061.
Perez, R.G., J.C. Waymire, E. Lin, J.J. Liu, F. Guo, and M.J. Zigmond. 2002. A role for alpha-synuclein in the regulation of dopamine biosynthesis. J. Neurosci. Off. J. Soc. Neurosci. 22:3090–3099. doi:20026307.

99
Piper, R.C., and D.J. Katzmann. 2007. Biogenesis and function of multivesicular bodies. Annu. Rev. Cell Dev. Biol. 23:519–547. doi:10.1146/annurev.cellbio.23.090506.123319.
Poewe, W. 2008. Non-motor symptoms in Parkinson’s disease. Eur. J. Neurol. 15 Suppl 1:14–20. doi:10.1111/j.1468-1331.2008.02056.x.
Polymeropoulos, M.H., C. Lavedan, E. Leroy, S.E. Ide, A. Dehejia, A. Dutra, B. Pike, H. Root, J. Rubenstein, R. Boyer, E.S. Stenroos, S. Chandrasekharappa, A. Athanassiadou, T. Papapetropoulos, W.G. Johnson, A.M. Lazzarini, R.C. Duvoisin, G. Di Iorio, L.I. Golbe, and R.L. Nussbaum. 1997. Mutation in the alpha-synuclein gene identified in families with Parkinson’s disease. Science. 276:2045–2047.
Porro, C., T. Trotta, and M.A. Panaro. 2015. Microvesicles in the brain: Biomarker, messenger or mediator? J. Neuroimmunol. 288:70–78. doi:10.1016/j.jneuroim.2015.09.006.
Postuma, R.B., A.E. Lang, J. Massicotte-Marquez, and J. Montplaisir. 2006. Potential early markers of Parkinson disease in idiopathic REM sleep behavior disorder. Neurology. 66:845–851. doi:10.1212/01.wnl.0000203648.80727.5b.
Prat, A., K. Biernacki, and J.P. Antel. 2005. Th1 and Th2 lymphocyte migration across the human BBB is specifically regulated by interferon beta and copolymer-1. J. Autoimmun. 24:119–124. doi:10.1016/j.jaut.2005.01.004.
Prensa, L., S. Richard, and A. Parent. 2003. Chemical anatomy of the human ventral striatum and adjacent basal forebrain structures. J. Comp. Neurol. 460:345–367. doi:10.1002/cne.10627.
Pretorius, E., H.M. Oberholzer, E. Smit, E. Steyn, S. Briedenhann, and C.R. Franz. 2008. Ultrastructural changes in platelet aggregates of HIV patients: a scanning electron microscopy study. Ultrastruct. Pathol. 32:75–79. doi:10.1080/01913120802034793.
Pretorius, E., A.C. Swanepoel, A.V. Buys, N. Vermeulen, W. Duim, and D.B. Kell. 2014. Eryptosis as a marker of Parkinson’s disease. Aging. 6:788–819. doi:10.18632/aging.100695.
Pringsheim, T., N. Jette, A. Frolkis, and T.D.L. Steeves. 2014. The prevalence of Parkinson’s disease: a systematic review and meta-analysis. Mov. Disord. Off. J. Mov. Disord. Soc. 29:1583–1590. doi:10.1002/mds.25945.
Priyadarshi, A., S.A. Khuder, E.A. Schaub, and S. Shrivastava. 2000. A meta-analysis of Parkinson’s disease and exposure to pesticides. Neurotoxicology. 21:435–440.
Qian, L., P.M. Flood, and J.-S. Hong. 2010. Neuroinflammation is a key player in Parkinson’s disease and a prime target for therapy. J. Neural Transm. Vienna Austria 1996. 117:971–979. doi:10.1007/s00702-010-0428-1.
Qiang, J.K., Y.C. Wong, A. Siderowf, H.I. Hurtig, S.X. Xie, V.M.-Y. Lee, J.Q. Trojanowski, D. Yearout, J. B Leverenz, T.J. Montine, M. Stern, S. Mendick, D. Jennings, C. Zabetian, K. Marek, and A.S. Chen-Plotkin. 2013. Plasma apolipoprotein A1 as a biomarker for Parkinson disease. Ann. Neurol. 74:119–127. doi:10.1002/ana.23872.

100
Quek, C., and A.F. Hill. 2017. The role of extracellular vesicles in neurodegenerative diseases. Biochem. Biophys. Res. Commun. 483:1178–1186. doi:10.1016/j.bbrc.2016.09.090.
Quik, M., L.Z. Huang, N. Parameswaran, T. Bordia, C. Campos, and X.A. Perez. 2009. Multiple roles for nicotine in Parkinson’s disease. Biochem. Pharmacol. 78:677–685. doi:10.1016/j.bcp.2009.05.003.
Quik, M., A. Mallela, M. Chin, J.M. McIntosh, X.A. Perez, and T. Bordia. 2013. Nicotine-mediated improvement in L-dopa-induced dyskinesias in MPTP-lesioned monkeys is dependent on dopamine nerve terminal function. Neurobiol. Dis. 50:30–41. doi:10.1016/j.nbd.2012.09.006.
R Core Team. 2016. R: A language and environment for statistical computing. R Found. Stat. Comput.
Ramaker, C., J. Marinus, A.M. Stiggelbout, and B.J. Van Hilten. 2002. Systematic evaluation of rating scales for impairment and disability in Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 17:867–876. doi:10.1002/mds.10248.
Redgrave, P., M. Rodriguez, Y. Smith, M.C. Rodriguez-Oroz, S. Lehericy, H. Bergman, Y. Agid, M.R. DeLong, and J.A. Obeso. 2010. Goal-directed and habitual control in the basal ganglia: implications for Parkinson’s disease. Nat. Rev. Neurosci. 11:760–772. doi:10.1038/nrn2915.
Renella, R., J.S. Schlehe, D.J. Selkoe, D.A. Williams, and M.J. LaVoie. 2014. Genetic deletion of the GATA1-regulated protein α-synuclein reduces oxidative stress and nitric oxide synthase levels in mature erythrocytes. Am. J. Hematol. 89:974–977. doi:10.1002/ajh.23796.
Respondek, G., and G.U. Höglinger. 2016. The phenotypic spectrum of progressive supranuclear palsy. Parkinsonism Relat. Disord. 22 Suppl 1:S34-36. doi:10.1016/j.parkreldis.2015.09.041.
Richter, P., J. Werner, A. Heerlein, A. Kraus, and H. Sauer. 1998. On the validity of the Beck Depression Inventory. A review. Psychopathology. 31:160–168.
Ritz, B., A. Ascherio, H. Checkoway, K.S. Marder, L.M. Nelson, W.A. Rocca, G.W. Ross, D. Strickland, S.K. Van Den Eeden, and J. Gorell. 2007. Pooled analysis of tobacco use and risk of Parkinson disease. Arch. Neurol. 64:990–997. doi:10.1001/archneur.64.7.990.
Rocha, N.P., F. Assis, P.L. Scalzo, É.L.M. Vieira, I.G. Barbosa, M.S. de Souza, P.P. Christo, H.J. Reis, and A.L. Teixeira. 2017. Reduced Activated T Lymphocytes (CD4+CD25+) and Plasma Levels of Cytokines in Parkinson’s Disease. Mol. Neurobiol. doi:10.1007/s12035-017-0404-y.
Rodrigues e Silva, A.M., F. Geldsetzer, B. Holdorff, F.W. Kielhorn, M. Balzer-Geldsetzer, W.H. Oertel, H. Hurtig, and R. Dodel. 2010. Who was the man who discovered the “Lewy bodies”? Mov. Disord. Off. J. Mov. Disord. Soc. 25:1765–1773. doi:10.1002/mds.22956.
Rosborough, K., N. Patel, and L.V. Kalia. 2017. α-Synuclein and Parkinsonism: Updates

101
and Future Perspectives. Curr. Neurol. Neurosci. Rep. 17. doi:10.1007/s11910-017-0737-y.
Rousseau, M., C. Belleannee, A.-C. Duchez, N. Cloutier, T. Levesque, F. Jacques, J. Perron, P.A. Nigrovic, M. Dieude, M.-J. Hebert, M.H. Gelb, and E. Boilard. 2015. Detection and quantification of microparticles from different cellular lineages using flow cytometry. Evaluation of the impact of secreted phospholipase A2 on microparticle assessment. PloS One. 10:e0116812. doi:10.1371/journal.pone.0116812.
Rubinsztein, D.C. 2006. The roles of intracellular protein-degradation pathways in neurodegeneration. Nature. 443:780–786. doi:10.1038/nature05291.
Salawu, F.K., A. Danburam, and A.B. Olokoba. 2010. Non-motor symptoms of Parkinson’s disease: diagnosis and management. Niger. J. Med. J. Natl. Assoc. Resid. Dr. Niger. 19:126–131.
Sarko, D.K., and C.E. McKinney. 2017. Exosomes: Origins and Therapeutic Potential for Neurodegenerative Disease. Front. Neurosci. 11:82. doi:10.3389/fnins.2017.00082.
Schapira, A.H., M. Gu, J.W. Taanman, S.J. Tabrizi, T. Seaton, M. Cleeter, and J.M. Cooper. 1998. Mitochondria in the etiology and pathogenesis of Parkinson’s disease. Ann. Neurol. 44:S89-98.
Schrag, A., M. Jahanshahi, and N. Quinn. 2000. What contributes to quality of life in patients with Parkinson’s disease? J. Neurol. Neurosurg. Psychiatry. 69:308–312.
Schwarzschild, M.A., K. Marek, S. Eberly, D. Oakes, I. Shoulson, D. Jennings, J. Seibyl, A. Ascherio, and Parkinson Study Group PRECEPT Investigators. 2011. Serum urate and probability of dopaminergic deficit in early “Parkinson’s disease.” Mov. Disord. Off. J. Mov. Disord. Soc. 26:1864–1868. doi:10.1002/mds.23741.
Searles Nielsen, S., L.G. Gallagher, J.I. Lundin, W.T. Longstreth, T. Smith-Weller, G.M. Franklin, P.D. Swanson, and H. Checkoway. 2012. Environmental tobacco smoke and Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 27:293–296. doi:10.1002/mds.24012.
Seidl, S.E., J.A. Santiago, H. Bilyk, and J.A. Potashkin. 2014. The emerging role of nutrition in Parkinson’s disease. Front. Aging Neurosci. 6:36. doi:10.3389/fnagi.2014.00036.
Seidler, A., W. Hellenbrand, B.P. Robra, P. Vieregge, P. Nischan, J. Joerg, W.H. Oertel, G. Ulm, and E. Schneider. 1996. Possible environmental, occupational, and other etiologic factors for Parkinson’s disease: a case-control study in Germany. Neurology. 46:1275–1284.
Semchuk, K.M., E.J. Love, and R.G. Lee. 1993. Parkinson’s disease: a test of the multifactorial etiologic hypothesis. Neurology. 43:1173–1180.
Serra, J.A., R.O. Domínguez, E.S. de Lustig, E.M. Guareschi, A.L. Famulari, E.L. Bartolomé, and E.R. Marschoff. 2001. Parkinson’s disease is associated with oxidative stress: comparison of peripheral antioxidant profiles in living Parkinson’s, Alzheimer’s and vascular dementia patients. J. Neural Transm. Vienna Austria 1996. 108:1135–1148. doi:10.1007/s007020170003.

102
Sharma, S., C.S. Moon, A. Khogali, A. Haidous, A. Chabenne, C. Ojo, M. Jelebinkov, Y. Kurdi, and M. Ebadi. 2013. Biomarkers in Parkinson’s disease (recent update). Neurochem. Int. 63:201–229. doi:10.1016/j.neuint.2013.06.005.
Shevchenko, A., M. Wilm, O. Vorm, and M. Mann. 1996. Mass spectrometric sequencing of proteins silver-stained polyacrylamide gels. Anal. Chem. 68:850–858.
Shi, M., C. Liu, T.J. Cook, K.M. Bullock, Y. Zhao, C. Ginghina, Y. Li, P. Aro, R. Dator, C. He, M.J. Hipp, C.P. Zabetian, E.R. Peskind, S.-C. Hu, J.F. Quinn, D.R. Galasko, W.A. Banks, and J. Zhang. 2014. Plasma exosomal α-synuclein is likely CNS-derived and increased in Parkinson’s disease. Acta Neuropathol. (Berl.). 128:639–650. doi:10.1007/s00401-014-1314-y.
Shimura, H., N. Hattori, S. i Kubo, Y. Mizuno, S. Asakawa, S. Minoshima, N. Shimizu, K. Iwai, T. Chiba, K. Tanaka, and T. Suzuki. 2000. Familial Parkinson disease gene product, parkin, is a ubiquitin-protein ligase. Nat. Genet. 25:302–305. doi:10.1038/77060.
Shin, C., S.-H. Park, J.Y. Yun, J.H. Shin, H.-K. Yang, H.-J. Lee, S.-H. Kong, Y.-S. Suh, G. Shen, Y. Kim, H.-J. Kim, and B. Jeon. 2017. Fundamental limit of alpha-synuclein pathology in gastrointestinal biopsy as a pathologic biomarker of Parkinson’s disease: Comparison with surgical specimens. Parkinsonism Relat. Disord. doi:10.1016/j.parkreldis.2017.09.001.
Shrivastava, M., and S. Vivekanandhan. 2011. An insight into ultrastructural and morphological alterations of platelets in neurodegenerative diseases. Ultrastruct. Pathol. 35:110–116. doi:10.3109/01913123.2011.553350.
Sian-Hülsmann, J., S. Mandel, M.B.H. Youdim, and P. Riederer. 2011. The relevance of iron in the pathogenesis of Parkinson’s disease. J. Neurochem. 118:939–957. doi:10.1111/j.1471-4159.2010.07132.x.
Simons, M., and G. Raposo. 2009. Exosomes--vesicular carriers for intercellular communication. Curr. Opin. Cell Biol. 21:575–581. doi:10.1016/j.ceb.2009.03.007.
Singleton, A.B., M. Farrer, J. Johnson, A. Singleton, S. Hague, J. Kachergus, M. Hulihan, T. Peuralinna, A. Dutra, R. Nussbaum, S. Lincoln, A. Crawley, M. Hanson, D. Maraganore, C. Adler, M.R. Cookson, M. Muenter, M. Baptista, D. Miller, J. Blancato, J. Hardy, and K. Gwinn-Hardy. 2003. alpha-Synuclein locus triplication causes Parkinson’s disease. Science. 302:841. doi:10.1126/science.1090278.
Snead, D., and D. Eliezer. 2014. Alpha-Synuclein Function and Dysfunction on Cellular Membranes. Exp. Neurobiol. 23:292. doi:10.5607/en.2014.23.4.292.
Sohal, R.S., and R. Weindruch. 1996. Oxidative stress, caloric restriction, and aging. Science. 273:59–63.
Souza, J.M., B.I. Giasson, V.M. Lee, and H. Ischiropoulos. 2000. Chaperone-like activity of synucleins. FEBS Lett. 474:116–119.
Spillantini, M.G., M.L. Schmidt, V.M. Lee, J.Q. Trojanowski, R. Jakes, and M. Goedert. 1997. Alpha-synuclein in Lewy bodies. Nature. 388:839–840. doi:10.1038/42166.
Stojkovska, I., B. M Wagner, and B. E Morrison. 2015. Parkinson’s disease and enhanced

103
inflammatory response. Exp. Biol. Med. 240:1387–1395. Stuendl, A., M. Kunadt, N. Kruse, C. Bartels, W. Moebius, K.M. Danzer, B. Mollenhauer,
and A. Schneider. 2016. Induction of α-synuclein aggregate formation by CSF exosomes from patients with Parkinson’s disease and dementia with Lewy bodies. Brain J. Neurol. 139:481–494. doi:10.1093/brain/awv346.
Sudha, K., A.V. Rao, S. Rao, and A. Rao. 2003. Free radical toxicity and antioxidants in Parkinson’s disease. Neurol. India. 51:60–62.
Suzuki, R., and H. Shimodaira. 2006. Pvclust: an R package for assessing the uncertainty in hierarchical clustering. Bioinforma. Oxf. Engl. 22:1540–1542. doi:10.1093/bioinformatics/btl117.
Testa, C.M., T.B. Sherer, and J.T. Greenamyre. 2005. Rotenone induces oxidative stress and dopaminergic neuron damage in organotypic substantia nigra cultures. Brain Res. Mol. Brain Res. 134:109–118. doi:10.1016/j.molbrainres.2004.11.007.
Théry, C., M. Ostrowski, and E. Segura. 2009. Membrane vesicles as conveyors of immune responses. Nat. Rev. Immunol. 9:581–593. doi:10.1038/nri2567.
Tofaris, G.K., A. Razzaq, B. Ghetti, K.S. Lilley, and M.G. Spillantini. 2003. Ubiquitination of alpha-synuclein in Lewy bodies is a pathological event not associated with impairment of proteasome function. J. Biol. Chem. 278:44405–44411. doi:10.1074/jbc.M308041200.
Tokarz, A., I. Szuścik, B. Kuśnierz-Cabala, M. Kapusta, M. Konkolewska, A. Żurakowski, A. Georgescu, and E. Stępień. 2015. Extracellular vesicles participate in the transport of cytokines and angiogenic factors in diabetic patients with ocular complications. Folia Med. Cracov. 55:35–48.
Tombaugh, T.N., and N.J. McIntyre. 1992. The mini-mental state examination: a comprehensive review. J. Am. Geriatr. Soc. 40:922–935.
Tomlinson, P.R., Y. Zheng, R. Fischer, R. Heidasch, C. Gardiner, S. Evetts, M. Hu, R. Wade-Martins, M.R. Turner, J. Morris, K. Talbot, B.M. Kessler, and G.K. Tofaris. 2015. Identification of distinct circulating exosomes in Parkinson’s disease. Ann. Clin. Transl. Neurol. 2:353–361. doi:10.1002/acn3.175.
Toomsoo, T., I. Liepelt-Scarfone, R. Kerner, L. Kadastik-Eerme, T. Asser, I. Rubanovits, D. Berg, and P. Taba. 2016. Substantia Nigra Hyperechogenicity: Validation of Transcranial Sonography for Parkinson Disease Diagnosis in a Large Estonian Cohort. J. Ultrasound Med. Off. J. Am. Inst. Ultrasound Med. 35:17–23. doi:10.7863/ultra.14.12069.
Trajkovic, K., C. Hsu, S. Chiantia, L. Rajendran, D. Wenzel, F. Wieland, P. Schwille, B. Brügger, and M. Simons. 2008. Ceramide triggers budding of exosome vesicles into multivesicular endosomes. Science. 319:1244–1247. doi:10.1126/science.1153124.
Trams, E.G., C.J. Lauter, N. Salem, and U. Heine. 1981. Exfoliation of membrane ecto-enzymes in the form of micro-vesicles. Biochim. Biophys. Acta. 645:63–70.
Tritsch, N.X., and B.L. Sabatini. 2012. Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron. 76:33–50. doi:10.1016/j.neuron.2012.09.023.

104
Tsunemi, T., K. Hamada, and D. Krainc. 2014. ATP13A2/PARK9 regulates secretion of exosomes and α-synuclein. J. Neurosci. Off. J. Soc. Neurosci. 34:15281–15287. doi:10.1523/JNEUROSCI.1629-14.2014.
Valadi, H., K. Ekström, A. Bossios, M. Sjöstrand, J.J. Lee, and J.O. Lötvall. 2007. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 9:654–659. doi:10.1038/ncb1596.
Valera, E., and E. Masliah. 2013. Immunotherapy for neurodegenerative diseases: focus on α-synucleinopathies. Pharmacol. Ther. 138:311–322. doi:10.1016/j.pharmthera.2013.01.013.
Verma, A. 2016. Prions, prion-like prionoids, and neurodegenerative disorders. Ann. Indian Acad. Neurol. 19:169–174. doi:10.4103/0972-2327.179979.
Visser, M., A.F.G. Leentjens, J. Marinus, A.M. Stiggelbout, and J.J. van Hilten. 2006. Reliability and validity of the Beck depression inventory in patients with Parkinson’s disease. Mov. Disord. Off. J. Mov. Disord. Soc. 21:668–672. doi:10.1002/mds.20792.
Volpicelli-Daley, L.A., K.C. Luk, T.P. Patel, S.A. Tanik, D.M. Riddle, A. Stieber, D.F. Meaney, J.Q. Trojanowski, and V.M.-Y. Lee. 2011. Exogenous α-synuclein fibrils induce Lewy body pathology leading to synaptic dysfunction and neuron death. Neuron. 72:57–71. doi:10.1016/j.neuron.2011.08.033.
Wakabayashi, K., and H. Takahashi. 2000. [The mechanism of Lewy body formation in Parkinson’s disease]. Nihon Rinsho Jpn. J. Clin. Med. 58:2022–2027.
Wang, X., S. Yu, F. Li, and T. Feng. 2015. Detection of α-synuclein oligomers in red blood cells as a potential biomarker of Parkinson’s disease. Neurosci. Lett. 599:115–119. doi:10.1016/j.neulet.2015.05.030.
Wang, Z., J. Liu, S. Chen, Y. Wang, L. Cao, Y. Zhang, W. Kang, H. Li, Y. Gui, S. Chen, and J. Ding. 2011. DJ-1 modulates the expression of Cu/Zn-superoxide dismutase-1 through the Erk1/2-Elk1 pathway in neuroprotection. Ann. Neurol. 70:591–599. doi:10.1002/ana.22514.
Watson, J.B., A. Hatami, H. David, E. Masliah, K. Roberts, C.E. Evans, and M.S. Levine. 2009. Alterations in corticostriatal synaptic plasticity in mice overexpressing human alpha-synuclein. Neuroscience. 159:501–513. doi:10.1016/j.neuroscience.2009.01.021.
Weihofen, A., K.J. Thomas, B.L. Ostaszewski, M.R. Cookson, and D.J. Selkoe. 2009. Pink1 forms a multiprotein complex with Miro and Milton, linking Pink1 function to mitochondrial trafficking. Biochemistry (Mosc.). 48:2045–2052. doi:10.1021/bi8019178.
Wirdefeldt, K., H.-O. Adami, P. Cole, D. Trichopoulos, and J. Mandel. 2011. Epidemiology and etiology of Parkinson’s disease: a review of the evidence. Eur. J. Epidemiol. 26 Suppl 1:S1-58. doi:10.1007/s10654-011-9581-6.
Wither, M.J., K.C. Hansen, and J.A. Reisz. 2016. Mass Spectrometry-Based Bottom-Up Proteomics: Sample Preparation, LC-MS/MS Analysis, and Database Query Strategies. Curr. Protoc. Protein Sci. 86:16.4.1-16.4.20. doi:10.1002/cpps.18.
Withers, G.S., J.M. George, G.A. Banker, and D.F. Clayton. 1997. Delayed localization of

105
synelfin (synuclein, NACP) to presynaptic terminals in cultured rat hippocampal neurons. Brain Res. Dev. Brain Res. 99:87–94.
Wooten, G.F., L.J. Currie, V.E. Bovbjerg, J.K. Lee, and J. Patrie. 2004. Are men at greater risk for Parkinson’s disease than women? J. Neurol. Neurosurg. Psychiatry. 75:637–639.
Xia, R., and Z.-H. Mao. 2012. Progression of motor symptoms in Parkinson’s disease. Neurosci. Bull. 28:39–48. doi:10.1007/s12264-012-1050-z.
Yadav, S., A. Dixit, S. Agrawal, A. Singh, G. Srivastava, A.K. Singh, P.K. Srivastava, O. Prakash, and M.P. Singh. 2012. Rodent models and contemporary molecular techniques: notable feats yet incomplete explanations of Parkinson’s disease pathogenesis. Mol. Neurobiol. 46:495–512. doi:10.1007/s12035-012-8291-8.
Zappulli, V., K.P. Friis, Z. Fitzpatrick, C.A. Maguire, and X.O. Breakefield. 2016. Extracellular vesicles and intercellular communication within the nervous system. J. Clin. Invest. 126:1198–1207. doi:10.1172/JCI81134.
Zarranz, J.J., J. Alegre, J.C. Gómez-Esteban, E. Lezcano, R. Ros, I. Ampuero, L. Vidal, J. Hoenicka, O. Rodriguez, B. Atarés, V. Llorens, E. Gomez Tortosa, T. del Ser, D.G. Muñoz, and J.G. de Yebenes. 2004. The new mutation, E46K, of alpha-synuclein causes Parkinson and Lewy body dementia. Ann. Neurol. 55:164–173. doi:10.1002/ana.10795.
Zeng, B.Y., B. Dass, A. Owen, S. Rose, C. Cannizzaro, B.C. Tel, and P. Jenner. 1999. Chronic L-DOPA treatment increases striatal cannabinoid CB1 receptor mRNA expression in 6-hydroxydopamine-lesioned rats. Neurosci. Lett. 276:71–74.
Zhang, L., R. Sheng, and Z. Qin. 2009. The lysosome and neurodegenerative diseases. Acta Biochim. Biophys. Sin. 41:437–445.
Zhang, Z.-X., Z.-H. Dong, and G.C. Román. 2006. Early descriptions of Parkinson disease in ancient China. Arch. Neurol. 63:782–784. doi:10.1001/archneur.63.5.782.
Zhao, B., Y. Zhang, S. Han, W. Zhang, Q. Zhou, H. Guan, J. Liu, J. Shi, L. Su, and D. Hu. 2017. Exosomes derived from human amniotic epithelial cells accelerate wound healing and inhibit scar formation. J. Mol. Histol. 48:121–132. doi:10.1007/s10735-017-9711-x.
Zhou, F.-M., C.J. Wilson, and J.A. Dani. 2002. Cholinergic interneuron characteristics and nicotinic properties in the striatum. J. Neurobiol. 53:590–605. doi:10.1002/neu.10150.
Zimprich, A., S. Biskup, P. Leitner, P. Lichtner, M. Farrer, S. Lincoln, J. Kachergus, M. Hulihan, R.J. Uitti, D.B. Calne, A.J. Stoessl, R.F. Pfeiffer, N. Patenge, I.C. Carbajal, P. Vieregge, F. Asmus, B. Müller-Myhsok, D.W. Dickson, T. Meitinger, T.M. Strom, Z.K. Wszolek, and T. Gasser. 2004. Mutations in LRRK2 cause autosomal-dominant parkinsonism with pleomorphic pathology. Neuron. 44:601–607. doi:10.1016/j.neuron.2004.11.005.
Żmigrodzka, M., M. Guzera, A. Miśkiewicz, D. Jagielski, and A. Winnicka. 2016. The biology of extracellular vesicles with focus on platelet microparticles and their role in cancer development and progression. Tumour Biol. J. Int. Soc. Oncodevelopmental

106
Biol. Med. 37:14391–14401. doi:10.1007/s13277-016-5358-6. Zucca, F.A., J. Segura-Aguilar, E. Ferrari, P. Muñoz, I. Paris, D. Sulzer, T. Sarna, L.
Casella, and L. Zecca. 2017. Interactions of iron, dopamine and neuromelanin pathways in brain aging and Parkinson’s disease. Prog. Neurobiol. 155:96–119. doi:10.1016/j.pneurobio.2015.09.012.
Annexe
Table S2.3. Complete list of proteins identified in the EEV proteome
A. Uniprot Complete Proteome Homo sapiens database for the non-depleted sample

107
Protein ID
Protein names Gene
names
Razor + unique
peptides P02549 Spectrin alpha chain, erythrocytic 1 SPTA1 136
P11277 Spectrin beta chain, erythrocytic SPTB 123
P16157 Ankyrin-1 ANK1 70
P02730 Band 3 anion transport protein SLC4A1 38
P55072 Transitional endoplasmic reticulum ATPase VCP 33
P16452 Erythrocyte membrane protein band 4.2 EPB42 33
P11171 Protein 4.1 EPB41 33
P04040 Catalase CAT 32
Q13228 Selenium-binding protein 1 SELENBP1 29
B4DT77 Annexin;Annexin A7 ANXA7 27
Q8WUM4 Programmed cell death 6-interacting protein PDCD6IP 27
P35612 Beta-adducin ADD2 26
P68871 Hemoglobin subunit beta;LVV-hemorphin-7;Spinorphin HBB 25
Q00610 Clathrin heavy chain 1 CLTC 25
P69905 Hemoglobin subunit alpha HBA1 22
B4DVE7 Annexin A11 ANXA11 20
J3QLD9 Flotillin-2 FLOT2 20
P08758 Annexin A5;Annexin ANXA5 19
P09525 Annexin A4;Annexin ANXA4 18
O75955; Flotillin-1 FLOT1 18
P11142 Heat shock cognate 71 kDa protein HSPA8 18
P32119 Peroxiredoxin-2 PRDX2 17
P00491 Purine nucleoside phosphorylase PNP 17
P27105 Erythrocyte band 7 integral membrane protein STOM 17
P00918 Carbonic anhydrase 2 CA2 17
P23634 Plasma membrane calcium-transporting ATPase 4 ATP2B4 17
C9JIF9 Acylamino-acid-releasing enzyme APEH 17
P00915 Carbonic anhydrase 1 CA1 16
P63261 Actin, cytoplasmic 2 ACTG1 16
Q5VU58 Tropomyosin alpha-3 chain TPM3 16
P30041 Peroxiredoxin-6 PRDX6 16
E7EU23 Rab GDP dissociation inhibitor beta GDI2 16
Q00013 55 kDa erythrocyte membrane protein MPP1 16
E7EV01 Calpain-5 CAPN5 16
J3KPS3 Fructose-bisphosphate aldolase A ALDOA 15
Q08495 Dematin DMTN 15
P23276 Kell blood group glycoprotein KEL 15
P69892 Hemoglobin subunit gamma-2 HBG2 14
P60174 Triosephosphate isomerase TPI1 14
P62258 14-3-3 protein epsilon YWHAE 14
O75326 Semaphorin-7A SEMA7A 14

108
O75340 Programmed cell death protein 6 PDCD6 13
E7EV99 Alpha-adducin ADD1 13
Q5VZU9 Tripeptidyl-peptidase 2 TPP2 13
P53396 ATP-citrate synthase ACLY 13
C9J0K6 Sorcin SRI 12
P04406 Glyceraldehyde-3-phosphate dehydrogenase GAPDH 12
P07738 Bisphosphoglycerate mutase BPGM 12
B7Z3I9 Delta-aminolevulinic acid dehydratase ALAD 12
Q86X55 Histone-arginine methyltransferase CARM1 CARM1 12
P07195 L-lactate dehydrogenase B chain;L-lactate dehydrogenase LDHB 12
P23526 Adenosylhomocysteinase AHCY 12
Q32Q12 Nucleoside diphosphate kinase NME1-NME2 12
B7Z7A9 Phosphoglycerate kinase 1 PGK1 11
P11166 Solute carrier family 2, facilitated glucose transporter member 1 SLC2A1 11
P22303 Acetylcholinesterase ACHE 11
Q9NP58 ATP-binding cassette sub-family B member 6, mitochondrial ABCB6 11
P40925 Malate dehydrogenase, cytoplasmic;Malate dehydrogenase MDH1 11
P00352 Retinal dehydrogenase 1 ALDH1A1 11
F2Z2V0 Copine-1 CPNE1 11
K7EMC9 WW domain-binding protein 2 WBP2 10
F5H7S3 Tropomyosin alpha-1 chain TPM1 10
A6NN80 Annexin A6;Annexin ANXA6 10
P30043 Flavin reductase (NADPH) BLVRB 9
H7BXD5 Grancalcin GCA 9
P04083 Annexin A1;Annexin ANXA1 9
P62937 Peptidyl-prolyl cis-trans isomerase PPIA 9
P37837 Transaldolase TALDO1 9
Q06830 Peroxiredoxin-1 PRDX1 9
O75131 Copine-3 CPNE3 9
P00390 Glutathione reductase, mitochondrial GSR 9
E7EQB2 Lactotransferrin LTF 9
P07384 Calpain-1 catalytic subunit CAPN1 9
P02042 Hemoglobin subunit delta HBD 8
P30086 Phosphatidylethanolamine-binding protein 1;Hippocampal cholinergic neurostimulating peptide PEBP1 8
P35613 Basigin BSG 8
Q9H0U4 Ras-related protein Rab-1B;Putative Ras-related protein Rab-1C RAB1B;RAB1
C 8
P63092 Guanine nucleotide-binding protein G(s) subunit alpha isoforms GNAS 8
P48506 Glutamate--cysteine ligase catalytic subunit GCLC 8
P06702 Protein S100-A9 S100A9 7
Q9UBV8 Peflin PEF1 7
P17931 Galectin-3;Galectin LGALS3 7
P28066 Proteasome subunit alpha type-5 PSMA5 7
P07451 Carbonic anhydrase 3 CA3 7
E7EQ12 Calpastatin CAST 7

109
P50895 Basal cell adhesion molecule BCAM 7
P28074 Proteasome subunit beta type-5 PSMB5 7
G3V5Z7 Proteasome subunit alpha type;Proteasome subunit alpha type-6 PSMA6 7
P25786 Proteasome subunit alpha type-1 PSMA1 7
G3V1D3 Dipeptidyl peptidase 3 DPP3 7
P49247 Ribose-5-phosphate isomerase RPIA 7
Q5T9B7 Adenylate kinase isoenzyme 1 AK1 7
P25789 Proteasome subunit alpha type-4; Proteasome subunit beta type PSMA4 7
B4E022 Transketolase TKT 7
J3QS39 Ubiquitin-60S ribosomal protein L40;Ubiquitin;60S ribosomal protein L40;Ubiquitin-40S ribosomal
protein S27a;Ubiquitin;40S ribosomal protein S27a;Polyubiquitin-B;Ubiquitin;Polyubiquitin-C;Ubiquitin
UBB;RPS27A;UBC;UBA52;
UBBP4 6
H0Y7A7 Calmodulin CALM2 6
P28070 Proteasome subunit beta type-4 PSMB4 6
Q9H4G4 Golgi-associated plant pathogenesis-related protein 1 GLIPR2 6
Q9BY43 Charged multivesicular body protein 4a CHMP4A 6
P48426 Phosphatidylinositol 5-phosphate 4-kinase type-2 alpha PIP4K2A 6
P28289 Tropomodulin-1 TMOD1 6
P07911 Uromodulin;Uromodulin, secreted form UMOD 6
Q9GZP4 PITH domain-containing protein 1 PITHD1 6
P78417 Glutathione S-transferase omega-1 GSTO1 6
P25788 Proteasome subunit alpha type-3 PSMA3 6
O14818 Proteasome subunit alpha type-7;Proteasome subunit alpha type-7-like PSMA7;PSMA
8 6
P08107 Heat shock 70 kDa protein 1A/1B HSPA1A 6
H0YD13 CD44 antigen CD44 6
P61225 Ras-related protein Rap-2b;Ras-related protein Rap-2c;Ras-related protein Rap-2a RAP2B;RAP2
A;RAP2C 6
P05109 Protein S100-A8;Protein S100-A8, N-terminally processed S100A8 6
P23528 Cofilin-1 CFL1 6
Q99808 Equilibrative nucleoside transporter 1 SLC29A1 6
P84077 ADP-ribosylation factor 1;ADP-ribosylation factor 3;ADP-ribosylation factor 5;ADP-ribosylation
factor 4 ARF1;ARF3;A
RF5;ARF4 6
P31946 14-3-3 protein beta/alpha;14-3-3 protein beta/alpha, N-terminally processed YWHAB 6
C9JIS1 Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2;Guanine nucleotide-binding protein
subunit beta-4 GNB2;GNB4 6
P53990 IST1 homolog IST1 6
Q99497 Protein DJ-1 PARK7 6
F5H7U0 6-phosphogluconate dehydrogenase, decarboxylating PGD 6
B7Z7E9 Aspartate aminotransferase, cytoplasmic GOT1 6
P62834 Ras-related protein Rap-1A;Ras-related protein Rap-1b;Ras-related protein Rap-1b-like protein RAP1A;RAP1
B 6
P04899 Guanine nucleotide-binding protein G(i) subunit alpha-2 GNAI2 6
P25325 3-mercaptopyruvate sulfurtransferase;Sulfurtransferase MPST 6
Q9NP79 Vacuolar protein sorting-associated protein VTA1 homolog VTA1 6
P00492 Hypoxanthine-guanine phosphoribosyltransferase HPRT1 6
Q16531 DNA damage-binding protein 1 DDB1 6
P22314 Ubiquitin-like modifier-activating enzyme 1 UBA1 6

110
P00441 Superoxide dismutase [Cu-Zn] SOD1 5
H7BY58 Protein-L-isoaspartate O-methyltransferase;Protein-L-isoaspartate(D-aspartate) O-methyltransferase PCMT1 5
P10768 S-formylglutathione hydrolase ESD 5
P09543 2,3-cyclic-nucleotide 3-phosphodiesterase CNP 5
P06733 Alpha-enolase;Enolase ENO1 5
P26038 Moesin MSN 5
O75368 SH3 domain-binding glutamic acid-rich-like protein SH3BGRL 5
K7EQ48 Glucose-6-phosphate isomerase GPI 5
P25787 Proteasome subunit alpha type-2 PSMA2 5
P49721 Proteasome subunit beta type-2 PSMB2 5
F5H8J2 Protein disulfide-isomerase P4HB 5
C9J9P4 Phospholipid scramblase 1 PLSCR1 5
P09211 Glutathione S-transferase P GSTP1 5
B5MDF5 GTP-binding nuclear protein Ran RAN 5
P07900 Heat shock protein HSP 90-alpha HSP90AA1 5
K7EQ02 DAZ-associated protein 1 DAZAP1 5
Q13630 GDP-L-fucose synthase TSTA3 5
F5H0T1 Stress-induced-phosphoprotein 1 STIP1 5
P50502 Hsc70-interacting protein;Putative protein FAM10A5;Putative protein FAM10A4 ST13;ST13P5;
ST13P4 5
P20618 Proteasome subunit beta type-1 PSMB1 5
P62805 Histone H4 HIST1H4A 5
P51148 Ras-related protein Rab-5C RAB5C 5
H7C2G2 Ecto-ADP-ribosyltransferase 4 ART4 5
J3KQ18 D-dopachrome decarboxylase;D-dopachrome decarboxylase-like protein DDT;DDTL 5
H3BPK3 Hydroxyacylglutathione hydrolase, mitochondrial HAGH 5
B4DIT7 Protein-glutamine gamma-glutamyltransferase 2 TGM2 5
O43633 Charged multivesicular body protein 2a CHMP2A 5
B4DQH4 T-complex protein 1 subunit theta CCT8 5
Q9UN37 Vacuolar protein sorting-associated protein 4A VPS4A 5
I3L397 Eukaryotic translation initiation factor 5A-1;Eukaryotic translation initiation factor 5A-1-like EIF5A;EIF5AL
1 5
Q9Y5Z4 Heme-binding protein 2 HEBP2 5
Q9UKV8 Protein argonaute-2 AGO2 5
F5H442 Tumor susceptibility gene 101 protein TSG101 5
H3BLV0 Complement decay-accelerating factor CD55 5
P02008 Hemoglobin subunit zeta HBZ 5
E7EPV7 Alpha-synuclein SNCA 4
P63104 14-3-3 protein zeta/delta YWHAZ 4
P02724 Glycophorin-A GYPA;GPErik 4
U3KQE2 Calpain small subunit 1 CAPNS1 4
Q9NP59 Solute carrier family 40 member 1 SLC40A1 4
Q5QPM9 Proteasome inhibitor PI31 subunit PSMF1 4
C9J8U2 Nicotinate phosphoribosyltransferase NAPRT 4
P61981 14-3-3 protein gamma;14-3-3 protein gamma, N-terminally processed YWHAG 4

111
Q9UQ80 Proliferation-associated protein 2G4 PA2G4 4
E5RJR5 S-phase kinase-associated protein 1 SKP1 4
Q9NRV9 Heme-binding protein 1 HEBP1 4
Q9H444 Charged multivesicular body protein 4b CHMP4B 4
P10599 Thioredoxin TXN 4
Q5VSJ9 Blood group Rh(CE) polypeptide;Blood group Rh(D) polypeptide RHCE;RHD 4
Q9UK41 Vacuolar protein sorting-associated protein 28 homolog VPS28 4
Q9Y3I1 F-box only protein 7 FBXO7 4
P61026 Ras-related protein Rab-10 RAB10 4
Q14974 Importin subunit beta-1 KPNB1 4
P27797 Calreticulin CALR 4
P18669 Phosphoglycerate mutase 1;Probable phosphoglycerate mutase 4 PGAM1;PGA
M4 4
P54725 UV excision repair protein RAD23 homolog A RAD23A 4
P30613 Pyruvate kinase PKLR PKLR 4
P63000 Ras-related C3 botulinum toxin substrate 1;Ras-related C3 botulinum toxin substrate 3;Ras-related C3
botulinum toxin substrate 2 RAC1;RAC3;R
AC2 4
P05164 Myeloperoxidase MPO 4
F5GWY2 Bifunctional purine biosynthesis protein PURH;Phosphoribosylaminoimidazolecarboxamide
formyltransferase;IMP cyclohydrolase ATIC 4
P48637 Glutathione synthetase GSS 4
F5H5V4 26S proteasome non-ATPase regulatory subunit 9 PSMD9 4
G5E9R5 Low molecular weight phosphotyrosine protein phosphatase ACP1 3
P46976 Glycogenin-1 GYG1 3
P28072 Proteasome subunit beta type-6 PSMB6 3
P26447 Protein S100-A4 S100A4 3
F5GXQ0 BRO1 domain-containing protein BROX BROX 3
P08754 Guanine nucleotide-binding protein G(k) subunit alpha GNAI3 3
Q99436 Proteasome subunit beta type-7 PSMB7 3
P62942 Peptidyl-prolyl cis-trans isomerase FKBP1A 3
U3KQK0 Histone H2B HIST1H2B 3
J3QKR3 Proteasome subunit beta type-3 PSMB3 3
P01116 GTPase KRas;GTPase KRas, N-terminally processed;GTPase HRas;GTPase HRas, N-terminally
processed;GTPase NRas KRAS;HRAS;
NRAS 3
P13489 Ribonuclease inhibitor RNH1 3
Q08722 Leukocyte surface antigen CD47 CD47 3
Q5T123 SH3 domain-binding glutamic acid-rich-like protein 3 SH3BGRL3 3
Q8WYQ7 Galectin;Galectin-9 LGALS9 3
O75695 Protein XRP2 RP2 3
P00167 Cytochrome b5 CYB5A 3
Q9Y4D1 Disheveled-associated activator of morphogenesis 1 DAAM1 3
P11021 78 kDa glucose-regulated protein HSPA5 3
H7C1D4 Translin TSN 3
P07737 Profilin-1 PFN1 3
M0R389 Platelet-activating factor acetylhydrolase IB subunit gamma PAFAH1B3 3
A6NJA2 Ubiquitin carboxyl-terminal hydrolase 14 USP14 3

112
P10644 cAMP-dependent protein kinase type I-alpha regulatory subunit PRKAR1A 3
Q9BS40 Latexin LXN 3
G5EA52 Protein disulfide-isomerase A3 PDIA3 3
P53004 Biliverdin reductase A BLVRA 3
Q04656 Copper-transporting ATPase 1 ATP7A 3
H9KV70 Neutrophil gelatinase-associated lipocalin LCN2 3
O00299 Chloride intracellular channel protein 1 CLIC1 3
F8WF69 Clathrin light chain A CLTA 3
G3V2F7 Ubiquitin-conjugating enzyme E2 variant 1;Ubiquitin-conjugating enzyme E2 variant 2 UBE2V1;UBE
2V2 3
F8WDS9 LanC-like protein 1 LANCL1 3
P60891 Ribose-phosphate pyrophosphokinase 1 PRPS1 3
K7ESE8 Bleomycin hydrolase BLMH 3
H0YNE3 Proteasome activator complex subunit 1 PSME1 3
P16930 Fumarylacetoacetase FAH 3
F8VSD4 Ubiquitin-conjugating enzyme E2 N UBE2N 3
P07203 Glutathione peroxidase 1 GPX1 3
P62328 Thymosin beta-4;Hematopoietic system regulatory peptide TMSB4X;TMS
B4XP4 3
E5RIW3 Tubulin-specific chaperone A TBCA 3
M0R0Y2 Alpha-soluble NSF attachment protein NAPA 3
P15374 Ubiquitin carboxyl-terminal hydrolase isozyme L3 UCHL3 3
P04921 Glycophorin-C GYPC 2
H0YDI1 Lymphocyte function-associated antigen 3 CD58 2
B4E220 Aquaporin-1 AQP1 2
C9JEN3 Protein lifeguard 3 TMBIM1 2
F5H2R5 Rho GDP-dissociation inhibitor 2 ARHGDIB 2
Q53TN4 Cytochrome b reductase 1 CYBRD1 2
Q9NZD4 Alpha-hemoglobin-stabilizing protein AHSP 2
Q8NHG7 Small VCP/p97-interacting protein SVIP 2
Q5JYX0 Cell division control protein 42 homolog CDC42 2
Q71RC9 Small integral membrane protein 5 SMIM5 2
E9PNW4 CD59 glycoprotein CD59 2
P09105 Hemoglobin subunit theta-1 HBQ1 2
R4GN98 Protein S100;Protein S100-A6 S100A6 2
O75531 Barrier-to-autointegration factor;Barrier-to-autointegration factor, N-terminally processed BANF1 2
Q5T6W5 Heterogeneous nuclear ribonucleoprotein K HNRNPK 2
F5H4Q5 Vacuolar protein sorting-associated protein 37C VPS37C 2
J3QK90 NSFL1 cofactor p47 NSFL1C 2
H3BV85 BolA-like protein 2 BOLA2B;BOL
A2 2
Q9NRX4 14 kDa phosphohistidine phosphatase PHPT1 2
H3BS66 Small integral membrane protein 1 SMIM1 2
E7ESC6 Exportin-7 XPO7 2
P68402 Platelet-activating factor acetylhydrolase IB subunit beta PAFAH1B2 2
Q9BRF8 Serine/threonine-protein phosphatase CPPED1 CPPED1 2

113
P08246 Neutrophil elastase ELANE 2
E9PN50 26S protease regulatory subunit 6A PSMC3 2
E7EUC7 UTP--glucose-1-phosphate uridylyltransferase UGP2 2
B8ZZB8 CB1 cannabinoid receptor-interacting protein 1 CNRIP1 2
E9PCS3 26S proteasome non-ATPase regulatory subunit 2 PSMD2 2
P59666 Neutrophil defensin 3;HP 3-56;Neutrophil defensin 2;Neutrophil defensin 1;HP 1-56;Neutrophil
defensin 2 DEFA3;DEFA
1 2
O15400 Syntaxin-7 STX7 2
P00338 L-lactate dehydrogenase A chain LDHA 2
P61970 Nuclear transport factor 2 NUTF2 2
E7EMV0 Protein diaphanous homolog 1 DIAPH1 2
F5GY90 Porphobilinogen deaminase HMBS 2
P61020 Ras-related protein Rab-5B RAB5B 2
Q99828 Calcium and integrin-binding protein 1 CIB1 2
B4DUA0 Plastin-2 LCP1 2
C9JTY3 Protein TFG TFG 2
P27348 14-3-3 protein theta YWHAQ 2
H0YKZ7 Annexin;Annexin A2;Putative annexin A2-like protein ANXA2;ANX
A2P2 2
P08238 Heat shock protein HSP 90-beta HSP90AB1 2
J3KQP6 Ras-related protein Rab-11B;Ras-related protein Rab-11A RAB11A;RAB
11B 2
A6NMU3 Signal transducing adapter molecule 1 STAM 2
P53985 Monocarboxylate transporter 1 SLC16A1 2
F6USW4 F-actin-capping protein subunit beta CAPZB 2
O14964 Hepatocyte growth factor-regulated tyrosine kinase substrate HGS 2
P20020 Plasma membrane calcium-transporting ATPase 1;Calcium-transporting ATPase ATP2B1 2
P36959 GMP reductase 1 GMPR 2
Q9Y376 Calcium-binding protein 39 CAB39 2
Q9Y6M5 Zinc transporter 1 SLC30A1 2
Q8IZ83 Aldehyde dehydrogenase family 16 member A1 ALDH16A1 2
Q99459 Cell division cycle 5-like protein CDC5L 2
P06132 Uroporphyrinogen decarboxylase UROD 2
J3KNT0 Fascin FSCN1 2
P49189 4-trimethylaminobutyraldehyde dehydrogenase ALDH9A1 2
H3BNT7 26S proteasome non-ATPase regulatory subunit 7 PSMD7 2
P05023
Sodium/potassium-transporting ATPase subunit alpha-1;Sodium/potassium-transporting ATPase subunit alpha-3;Sodium/potassium-transporting ATPase subunit alpha-2;Sodium/potassium-
transporting ATPase subunit alpha-4;Potassium-transporting ATPase alpha chain 1;Potassium-transporting ATPase alpha chain 2
ATP1A1;ATP1A2;ATP1A3;ATP1A4;ATP4A
;ATP12A
2
P34932 Heat shock 70 kDa protein 4 HSPA4 2
K7EMV3 Histone H3 H3F3B 2
Q8IU68 Transmembrane channel-like protein 8 TMC8 2
E7ENZ3 T-complex protein 1 subunit epsilon CCT5 2
Q5TZA2 Rootletin CROCC 2
Q9P203 BTB/POZ domain-containing protein 7 BTBD7 2
Q7LBR1 Charged multivesicular body protein 1b CHMP1B 2

114
U3KQ56 Glyoxylate reductase/hydroxypyruvate reductase GRHPR 2
H0YJ11 Alpha-actinin-1;Alpha-actinin-2;Alpha-actinin-4 ACTN1;ACTN
4;ACTN2 2
Q9UDT6 CAP-Gly domain-containing linker protein 2 CLIP2 2
P09960 Leukotriene A-4 hydrolase LTA4H 2
Q5HY54 Filamin-A FLNA 2
E9PJL5 Uncharacterized protein C12orf55;Putative uncharacterized protein C12orf63 C12orf55;C12o
rf63 2
G3V2U7 Acylphosphatase;Acylphosphatase-1 FBN3;ACYP1 2
E9PQN4 Complement receptor type 1 CR1 2
Q9BSL1 Ubiquitin-associated domain-containing protein 1 UBAC1 2
Q04917 14-3-3 protein eta YWHAH 2
B7ZBP9 Serine/threonine-protein phosphatase 2A activator PPP2R4;DKFZp781M17165
2
Q8NDC0 MAPK-interacting and spindle-stabilizing protein-like MAPK1IP1L 1
P68133 Actin, alpha skeletal muscle;Actin, alpha cardiac muscle 1;Actin, gamma-enteric smooth muscle;Actin,
aortic smooth muscle
ACTA1;ACTC1;ACTG2;ACT
A2 1
P69891 Hemoglobin subunit gamma-1 HBG1 1
S4R3Y4 Protein AMBP;Alpha-1-microglobulin;Inter-alpha-trypsin inhibitor light chain;Trypstatin AMBP 1
I3L3E4 Charged multivesicular body protein 6 CHMP6 1
Q16570 Atypical chemokine receptor 1 ACKR1 1
Q5VY30 Retinol-binding protein 4;Plasma retinol-binding protein(1-182);Plasma retinol-binding protein(1-
181);Plasma retinol-binding protein(1-179);Plasma retinol-binding protein(1-176) RBP4 1
E7END7 Ras-related protein Rab-1A RAB1A 1
Q5VU59 TPM3 1
P17066 Heat shock 70 kDa protein 6;Putative heat shock 70 kDa protein 7 HSPA6;HSPA
7 1
Q04760 Lactoylglutathione lyase GLO1 1
D6RD66 WD repeat-containing protein 1 WDR1 1
K7EM02 Katanin p60 ATPase-containing subunit A-like 2 KATNAL2 1
P14209 CD99 antigen CD99 1
E9PIR7 Thioredoxin reductase 1, cytoplasmic GML;TXNRD
1 1
K7EMQ9 EIF3K 1
P15531 Nucleoside diphosphate kinase A NME1 1
H7BZT4 Small ubiquitin-related modifier 4;Small ubiquitin-related modifier 2;Small ubiquitin-related modifier 3 SUMO2;SUMO3;SUMO4
1
O00560 Syntenin-1 SDCBP 1
Q9BVM4 Gamma-glutamylaminecyclotransferase GGACT 1
K7EKH5 Fructose-bisphosphate aldolase C ALDOC 1
P49773 Histidine triad nucleotide-binding protein 1 HINT1 1
H0YBY6 Disks large-associated protein 2 DLGAP2 1
Q9Y624 Junctional adhesion molecule A F11R 1
B1AKQ8 Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-1;Guanine nucleotide-binding protein
G(I)/G(S)/G(T) subunit beta-3 GNB1;GNB3 1
K7EKN6 Urea transporter 1 SLC14A1 1
I3L0K2 Thioredoxin domain-containing protein 17 TXNDC17 1
A8MXY0 Syntaxin-4 STX4 1

115
O14773 Tripeptidyl-peptidase 1 TPP1 1
E9PNW0 Nucleosome assembly protein 1-like 1;Nucleosome assembly protein 1-like 4 NAP1L4;NAP
1L1 1
Q5TDH0 Protein DDI1 homolog 2 DDI2 1
Q96JM4 Leucine-rich repeat and IQ domain-containing protein 1 LRRIQ1 1
F5GWT9 Phosphoribosylformylglycinamidine synthase PFAS 1
F2Z3J2 26S proteasome non-ATPase regulatory subunit 5 PSMD5 1
J3QL74 Zinc finger and BTB domain-containing protein 14 ZBTB14 1
E9PJC7 CD82 antigen CD82 1
Q9H936 Mitochondrial glutamate carrier 1 SLC25A22 1
D6RD63 COP9 signalosome complex subunit 4 COPS4 1
Q6B0K9 Hemoglobin subunit mu HBM 1
Q31611 HLA class I histocompatibility antigen, alpha chain G HLA-G 1
H7BY04 Laminin subunit gamma-3 LAMC3 1
Q9UL25 Ras-related protein Rab-21 RAB21 1
H7C3P7 Ras-related protein Ral-A RALA 1
P08311 Cathepsin G CTSG 1
E9PE37 Ras-related protein Rab-2A;Ras-related protein Rab-2B RAB2B;RAB2
A 1
G3V1N2 HBA2 1
P00387 NADH-cytochrome b5 reductase 3;NADH-cytochrome b5 reductase 3 membrane-bound form;NADH-
cytochrome b5 reductase 3 soluble form CYB5R3 1
O75339 Cartilage intermediate layer protein 1;Cartilage intermediate layer protein 1 C1;Cartilage intermediate
layer protein 1 C2 CILP 1
P14324 Farnesyl pyrophosphate synthase FDPS 1
K7EKG2 Thioredoxin-like protein 1 TXNL1 1
B. Uniprot Complete Proteome Homo sapiens database when the heamoglobin was analyzed separately
Protein ID
Protein names Gene
names
Razor + unique
peptides P02549 Spectrin alpha chain, erythrocytic 1 SPTA1 197
P11277 Spectrin beta chain, erythrocytic SPTB 177
P16157 Ankyrin-1 ANK1 94
P55072 Transitional endoplasmic reticulum ATPase VCP 53
P111714 Protein 4.1 EPB41 52
P35579 Myosin-9 MYH9 44
Q8WUM4 Programmed cell death 6-interacting protein PDCD6IP 43
P02730 Band 3 anion transport protein SLC4A1 43
P16452 Erythrocyte membrane protein band 4.2 EPB42 42
P04040 Catalase CAT 40
A0A087WVQ6
Clathrin heavy chain;Clathrin heavy chain 1 CLTC 38
P35612 Beta-adducin ADD2 37
P16157 Ankyrin-1 ANK1 36
Q14254 Flotillin-2 FLOT2 33

116
P20073 Annexin A7 ANXA7 31
O75955 Flotillin-1 FLOT1 30
P53396 ATP-citrate synthase ACLY 30
P06753 TPM3 29
P49368 T-complex protein 1 subunit gamma CCT3 29
P236344 Plasma membrane calcium-transporting ATPase 4 ATP2B4 28
P11142 Heat shock cognate 71 kDa protein HSPA8 28
P60709 Actin, cytoplasmic 1;Actin, cytoplasmic 1, N-terminally processed ACTB 28
Q5T4S7 E3 ubiquitin-protein ligase UBR4 UBR4 27
P78371 T-complex protein 1 subunit beta CCT2 26
P28289 Tropomodulin-1 TMOD1 26
P50395 Rab GDP dissociation inhibitor beta GDI2 25
P27105 Erythrocyte band 7 integral membrane protein STOM 25
P68871 Hemoglobin subunit beta;LVV-hemorphin-7;Spinorphin HBB 25
P02730 Band 3 anion transport protein SLC4A1 25
Q00013 55 kDa erythrocyte membrane protein MPP1 24
J3KPS3 Fructose-bisphosphate aldolase;Fructose-bisphosphate aldolase A ALDOA 24
P00352 Retinal dehydrogenase 1 ALDH1A1 24
Q86VP6 Cullin-associated NEDD8-dissociated protein 1 CAND1 24
P49327 Fatty acid synthase FASN 24
Q13228 Selenium-binding protein 1 SELENBP1 22
P00915 Carbonic anhydrase 1 CA1 22
P50991 T-complex protein 1 subunit delta CCT4 22
P50990 T-complex protein 1 subunit theta CCT8 22
P32119 Peroxiredoxin-2 PRDX2 21
P50995 Annexin A11 ANXA11 21
P69905 Hemoglobin subunit alpha HBA1 21
P00558 Phosphoglycerate kinase 1 PGK1 20
H7BXK9 ATP-binding cassette sub-family B member 6, mitochondrial ABCB6 20
P08758 Annexin A5;Annexin ANXA5 20
P09525 Annexin A4;Annexin ANXA4 20
P07900 Heat shock protein HSP 90-alpha HSP90AA1 20
P48643 T-complex protein 1 subunit epsilon CCT5 20
P07384 Calpain-1 catalytic subunit CAPN1 20
P22314 Ubiquitin-like modifier-activating enzyme 1 UBA1 19
P04406 Glyceraldehyde-3-phosphate dehydrogenase GAPDH 19
Q08495 Dematin DMTN 19
Q99832 T-complex protein 1 subunit eta CCT7 19
P29144 Tripeptidyl-peptidase 2 TPP2 19
P30041 Peroxiredoxin-6 PRDX6 18
E9PM69 26S protease regulatory subunit 6A PSMC3 18
P40227 T-complex protein 1 subunit zeta CCT6A 18
P50570 Dynamin-2 DNM2 18
E7EQB2 Lactotransferrin;Lactoferricin-H;Kaliocin-1;Lactoferroxin-A;Lactoferroxin-B;Lactoferroxin-C LTF 18

117
P31948 Stress-induced-phosphoprotein 1 STIP1 18
E7ESC6 Exportin-7 XPO7 18
E7EV99 Alpha-adducin ADD1 17
P62258 14-3-3 protein epsilon YWHAE 17
P30613 Pyruvate kinase PKLR PKLR 17
A0A0G2JIW1 Heat shock 70 kDa protein 1B;Heat shock 70 kDa protein 1A HSPA1B;HSP
A1A 17
Q16531 DNA damage-binding protein 1 DDB1 17
P11021 78 kDa glucose-regulated protein HSPA5 17
F5H2F4 C-1-tetrahydrofolate synthase, cytoplasmic;Methylenetetrahydrofolate dehydrogenase MTHFD1 17
P07195 L-lactate dehydrogenase B chain;L-lactate dehydrogenase LDHB 16
P45974 Ubiquitin carboxyl-terminal hydrolase 5 USP5 16
O43242 26S proteasome non-ATPase regulatory subunit 3 PSMD3 16
C9J0K6 Sorcin SRI 16
P08133 Annexin A6;Annexin ANXA6 16
Q13200 26S proteasome non-ATPase regulatory subunit 2 PSMD2 16
P23276 Kell blood group glycoprotein KEL 16
P34932 Heat shock 70 kDa protein 4 HSPA4 16
A0A0A0MSI0 Peroxiredoxin-1 PRDX1 16
Q9Y230 RuvB-like 2 RUVBL2 16
Q5XPI4 E3 ubiquitin-protein ligase RNF123 RNF123 16
P68871 Hemoglobin subunit beta;LVV-hemorphin-7;Spinorphin HBB 15
P60174 Triosephosphate isomerase TPI1 15
P00491 Purine nucleoside phosphorylase PNP 15
C9JIF9 Acylamino-acid-releasing enzyme APEH 15
H7BYY1 Tropomyosin alpha-1 chain TPM1 15
P35998 26S protease regulatory subunit 7 PSMC2 15
P17987 T-complex protein 1 subunit alpha TCP1 15
P09543 2,3-cyclic-nucleotide 3-phosphodiesterase CNP 15
Q99460 26S proteasome non-ATPase regulatory subunit 1 PSMD1 15
Q9Y4E8 Ubiquitin carboxyl-terminal hydrolase 15 USP15 15
Q9C0C9 E2/E3 hybrid ubiquitin-protein ligase UBE2O UBE2O 15
P26038 Moesin MSN 15
P04083 Annexin A1;Annexin ANXA1 14
P30043 Flavin reductase (NADPH) BLVRB 14
P11166 Solute carrier family 2, facilitated glucose transporter member 1 SLC2A1 14
P00918 Carbonic anhydrase 2 CA2 14
P06733 Alpha-enolase ENO1 14
Q5TDH0 Protein DDI1 homolog 2 DDI2 14
B0QZ18 Copine-1 CPNE1 14
O75326 Semaphorin-7A SEMA7A 14
P05164 Myeloperoxidase MPO 14
Q9Y265 RuvB-like 1 RUVBL1 14
P29401 Transketolase TKT 14
I3L0N3 Vesicle-fusing ATPase NSF 14

118
Q4VB86 Protein 4.1 EPB41 14
P11277 Spectrin beta chain, erythrocytic SPTB 14
P13716 Delta-aminolevulinic acid dehydratase ALAD 13
P07738 Bisphosphoglycerate mutase BPGM 13
P48506 Glutamate--cysteine ligase catalytic subunit GCLC 13
Q99816 Tumor susceptibility gene 101 protein TSG101 13
O14818 Proteasome subunit alpha type-7 PSMA7 13
P23526 Adenosylhomocysteinase AHCY 13
P61225 Ras-related protein Rap-2b RAP2B 13
O00231 26S proteasome non-ATPase regulatory subunit 11 PSMD11 13
P11413 Glucose-6-phosphate 1-dehydrogenase G6PD 13
P00338 L-lactate dehydrogenase A chain LDHA 12
Q99808 Equilibrative nucleoside transporter 1 SLC29A1 12
A6NJA2 Ubiquitin carboxyl-terminal hydrolase;Ubiquitin carboxyl-terminal hydrolase 14 USP14 12
Q06323 Proteasome activator complex subunit 1 PSME1 12
P28074 Proteasome subunit beta type-5 PSMB5 12
B3KQV6 Serine/threonine-protein phosphatase 2A 65 kDa regulatory subunit A alpha isoform PPP2R1A 12
Q14974 Importin subunit beta-1 KPNB1 12
P25786 Proteasome subunit alpha type-1;Proteasome subunit alpha type PSMA1 12
Q86X55 Histone-arginine methyltransferase CARM1 CARM1 12
A6NG10 WW domain-binding protein 2 WBP2 12
P63092 Guanine nucleotide-binding protein G(s) subunit alpha GNAS 12
P31939 Bifunctional purine biosynthesis protein PURH;Phosphoribosylaminoimidazolecarboxamide
formyltransferase;IMP cyclohydrolase ATIC 12
P52209 6-phosphogluconate dehydrogenase, decarboxylating PGD 12
A0A087X0C8 Calpain-5 CAPN5 12
F8W9S7 GTPase-activating protein and VPS9 domain-containing protein 1 GAPVD1 12
P60842 Eukaryotic initiation factor 4A-I EIF4A1 12
P69905 Hemoglobin subunit alpha HBA1 11
Q6XQN6 Nicotinate phosphoribosyltransferase NAPRT 11
P48637 Glutathione synthetase GSS 11
H7BZ94 Protein disulfide-isomerase P4HB 11
P21980 Protein-glutamine gamma-glutamyltransferase 2 TGM2 11
P50895 Basal cell adhesion molecule BCAM 11
A0A087X2I1 26S protease regulatory subunit 10B PSMC6 11
G3V1D3 Dipeptidyl peptidase 3 DPP3 11
P05023 Sodium/potassium-transporting ATPase subunit alpha-1;Sodium/potassium-transporting ATPase
subunit alpha-3 ATP1A1;ATP1
A3 11
P40925 Malate dehydrogenase, cytoplasmic;Malate dehydrogenase MDH1 11
Q9UKV8 Protein argonaute-2 AGO2 11
P30566 Adenylosuccinate lyase ADSL 11
P20618 Proteasome subunit beta type-1 PSMB1 11
P17858 ATP-dependent 6-phosphofructokinase, liver type PFKL 11
A0A087X253 AP-2 complex subunit beta AP2B1 11
O95782 AP-2 complex subunit alpha-1 AP2A1 11

119
O00232 26S proteasome non-ATPase regulatory subunit 12 PSMD12 11
Q9BSL1 Ubiquitin-associated domain-containing protein 1 UBAC1 11
A0A087WUL0
Bifunctional ATP-dependent dihydroxyacetone kinase/FAD-AMP lyase (cyclizing);ATP-dependent dihydroxyacetone kinase;FAD-AMP lyase (cyclizing)
TKFC;DAK 11
P69891 Hemoglobin subunit gamma-1 HBG1 11
A0A087WZE4 Spectrin alpha chain, erythrocytic 1 SPTA1 11
P48426 Phosphatidylinositol 5-phosphate 4-kinase type-2 alpha PIP4K2A 10
P10644 cAMP-dependent protein kinase type I-alpha regulatory subunit PRKAR1A 10
Q9BWD1 Acetyl-CoA acetyltransferase, cytosolic ACAT2 10
P62191 26S protease regulatory subunit 4 PSMC1 10
E9PBS1 Multifunctional protein ADE2;Phosphoribosylaminoimidazole-succinocarboxamide
synthase;Phosphoribosylaminoimidazole carboxylase PAICS 10
M0R0Y2 Alpha-soluble NSF attachment protein NAPA 10
P78417 Glutathione S-transferase omega-1 GSTO1 10
P25789 Proteasome subunit alpha type-4;Proteasome subunit alpha type;Proteasome subunit beta type PSMA4 10
Q9H0U4 Ras-related protein Rab-1B;Putative Ras-related protein Rab-1C RAB1B;RAB1
C 10
Q16401 26S proteasome non-ATPase regulatory subunit 5 PSMD5 10
P30101 Protein disulfide-isomerase A3 PDIA3 10
D6RAX7 COP9 signalosome complex subunit 4 COPS4 10
O75340 Programmed cell death protein 6 PDCD6 10
Q96P70 Importin-9 IPO9 10
P38606 V-type proton ATPase catalytic subunit A ATP6V1A 10
A0A0G2JH68 Protein diaphanous homolog 1 DIAPH1 10
Q5T9B7 Adenylate kinase isoenzyme 1 AK1 10
O14980 Exportin-1 XPO1 10
P43686 26S protease regulatory subunit 6B PSMC4 10
H0YH81 ATP synthase subunit beta;ATP synthase subunit beta, mitochondrial ATP5B 10
Q04656 Copper-transporting ATPase 1 ATP7A 10
P16452 Erythrocyte membrane protein band 4.2 EPB42 10
Q9BY43 Charged multivesicular body protein 4a CHMP4A 9
P51148 Ras-related protein Rab-5C RAB5C 9
Q9NRV9 Heme-binding protein 1 HEBP1 9
Q9UNZ2 NSFL1 cofactor p47 NSFL1C 9
Q16851 UTP--glucose-1-phosphate uridylyltransferase UGP2 9
P28066 Proteasome subunit alpha type-5 PSMA5 9
A0A0C4DGQ5
Calpain small subunit 1 CAPNS1 9
A0A087X1Z3 Proteasome activator complex subunit 2 PSME2 9
Q01518 Adenylyl cyclase-associated protein 1 CAP1 9
B1AKQ8 Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-1 GNB1 9
O75131 Copine-3 CPNE3 9
P54725 UV excision repair protein RAD23 homolog A RAD23A 9
P11215 Integrin alpha-M ITGAM 9
Q93008 Probable ubiquitin carboxyl-terminal hydrolase FAF-X USP9X 9
Q96G03 Phosphoglucomutase-2 PGM2 9

120
P49721 Proteasome subunit beta type-2 PSMB2 9
Q15008 26S proteasome non-ATPase regulatory subunit 6 PSMD6 9
Q9UNQ0 ATP-binding cassette sub-family G member 2 ABCG2 9
P22303 Acetylcholinesterase;Carboxylic ester hydrolase ACHE 9
G3V5Z7 Proteasome subunit alpha type;Proteasome subunit alpha type-6 PSMA6 9
O15439 Multidrug resistance-associated protein 4 ABCC4 9
P37837 Transaldolase TALDO1 9
O14744 Protein arginine N-methyltransferase 5 PRMT5 9
P02042 Hemoglobin subunit delta HBD 9
P25788 Proteasome subunit alpha type-3 PSMA3 8
J3QS39 Ubiquitin-60S ribosomal protein L40;Ubiquitin;60S ribosomal protein L40;Ubiquitin-40S ribosomal
protein S27a;Ubiquitin;40S ribosomal protein S27a;Polyubiquitin-B;Ubiquitin;Polyubiquitin-C;Ubiquitin
UBB;RPS27A;UBC;UBA52;
UBBP4 8
O94919 Endonuclease domain-containing 1 protein ENDOD1 8
P31946 14-3-3 protein beta/alpha YWHAB 8
P60891 Ribose-phosphate pyrophosphokinase 1;Ribose-phosphate pyrophosphokinase 2;Ribose-phosphate
pyrophosphokinase 3 PRPS1;PRPS2;
PRPS1L1 8
P62195 26S protease regulatory subunit 8 PSMC5 8
A0A024RA52 Proteasome subunit alpha type;Proteasome subunit alpha type-2 PSMA2 8
P04899 Guanine nucleotide-binding protein G(i) subunit alpha-2 GNAI2 8
Q13561 Dynactin subunit 2 DCTN2 8
P53004 Biliverdin reductase A BLVRA 8
P00387 NADH-cytochrome b5 reductase 3 CYB5R3 8
P16152 Carbonyl reductase [NADPH] 1 CBR1 8
P06744 Glucose-6-phosphate isomerase GPI 8
Q99733 Nucleosome assembly protein 1-like 4 NAP1L4 8
P04792 Heat shock protein beta-1 HSPB1 8
P17612 cAMP-dependent protein kinase catalytic subunit alpha;cAMP-dependent protein kinase catalytic
subunit beta PRKACA;KIN27;PRKACB
8
P20340 Ras-related protein Rab-6A RAB6A 8
P13796 Plastin-2 LCP1 8
P52907 F-actin-capping protein subunit alpha-1 CAPZA1 8
Q14697 Neutral alpha-glucosidase AB GANAB 8
P08514 Integrin alpha-IIb;Integrin alpha-IIb heavy chain;Integrin alpha-IIb light chain, form 1;Integrin alpha-
IIb light chain, form 2 ITGA2B 8
P26641 Elongation factor 1-gamma EEF1G 8
Q9UQ80 Proliferation-associated protein 2G4 PA2G4 8
Q9Y4D1 Disheveled-associated activator of morphogenesis 1 DAAM1 8
P11166 Solute carrier family 2, facilitated glucose transporter member 1 SLC2A1 8
P63261 Actin, cytoplasmic 2;Actin, cytoplasmic 2, N-terminally processed;Actin, cytoplasmic 1;Actin,
cytoplasmic 1, N-terminally processed;Actin, gamma-enteric smooth muscle;Actin, alpha skeletal muscle;Actin, alpha cardiac muscle 1;Actin, aortic smooth muscle
ACTG1;ACTB;ACTG2;ACT
A1;ACTC1;ACTA2
8
P02008 Hemoglobin subunit zeta HBZ 8
P20073 Annexin A7 ANXA7 8
P35613 Basigin BSG 7
P49720 Proteasome subunit beta type-3 PSMB3 7
P17931 Galectin-3;Galectin LGALS3 7

121
P63104 14-3-3 protein zeta/delta YWHAZ 7
Q9H444 Charged multivesicular body protein 4b CHMP4B 7
O43396 Thioredoxin-like protein 1 TXNL1 7
P50502 Hsc70-interacting protein;Putative protein FAM10A4;Putative protein FAM10A5 ST13;ST13P4;
ST13P5 7
E7EQ12 Calpastatin CAST 7
P49189 4-trimethylaminobutyraldehyde dehydrogenase ALDH9A1 7
Q9Y3I1 F-box only protein 7 FBXO7 7
P07954 Fumarate hydratase, mitochondrial FH 7
F6S8N6 Protein-L-isoaspartate O-methyltransferase;Protein-L-isoaspartate(D-aspartate) O-methyltransferase PCMT1 7
P49247 Ribose-5-phosphate isomerase RPIA 7
P62834 Ras-related protein Rap-1A RAP1A 7
A0A087WUQ6
Glutathione peroxidase;Glutathione peroxidase 1 GPX1 7
O60256 Phosphoribosyl pyrophosphate synthase-associated protein 2 PRPSAP2 7
Q00796 Sorbitol dehydrogenase SORD 7
O00299 Chloride intracellular channel protein 1 CLIC1 7
Q99497 Protein deglycase DJ-1 PARK7 7
P17174 Aspartate aminotransferase, cytoplasmic GOT1 7
H7BXD5 Grancalcin GCA 7
E9PGT1 Translin TSN 7
C9J7K9 Phospholipid scramblase 1 PLSCR1 7
Q13618 Cullin-3 CUL3 7
O75695 Protein XRP2 RP2 7
P09960 Leukotriene A-4 hydrolase LTA4H 7
E9PLK3 Puromycin-sensitive aminopeptidase NPEPPS 7
P00492 Hypoxanthine-guanine phosphoribosyltransferase HPRT1 7
F5H4B6 Aldehyde dehydrogenase family 16 member A1 ALDH16A1 7
D6RA82 Annexin;Annexin A3 ANXA3 7
P61106 Ras-related protein Rab-14 RAB14 7
Q16775 Hydroxyacylglutathione hydrolase, mitochondrial HAGH 7
P07355 Annexin A2;Annexin;Putative annexin A2-like protein ANXA2;ANX
A2P2 7
A0A087WX08 Gamma-adducin ADD3 7
P08238 Heat shock protein HSP 90-beta HSP90AB1 7
Q8IZY2 ATP-binding cassette sub-family A member 7 ABCA7 7
P14780 Matrix metalloproteinase-9 MMP9 7
P12955 Xaa-Pro dipeptidase PEPD 7
P68371 Tubulin beta-4B chain;Tubulin beta-4A chain;Tubulin beta chain TUBB4B;TUB
B4A;TUBB 7
O15067 Phosphoribosylformylglycinamidine synthase PFAS 7
P35241 Radixin RDX 7
O60488 Long-chain-fatty-acid--CoA ligase 4 ACSL4 7
A0A0C4DGX4
Cullin-1 CUL1 7
P50148 Guanine nucleotide-binding protein G(q) subunit alpha GNAQ 7

122
Q15907 Ras-related protein Rab-11B;Ras-related protein Rab-11A RAB11B;RAB
11A 7
P30086 Phosphatidylethanolamine-binding protein 1;Hippocampal cholinergic neurostimulating peptide PEBP1 6
P55036 26S proteasome non-ATPase regulatory subunit 4 PSMD4 6
P28070 Proteasome subunit beta type-4 PSMB4 6
Q9UNS2 COP9 signalosome complex subunit 3 COPS3 6
P08754 Guanine nucleotide-binding protein G(k) subunit alpha GNAI3 6
P61006 Ras-related protein Rab-8A RAB8A 6
P61019 Ras-related protein Rab-2A RAB2A 6
H0Y8C6 Importin-5 IPO5 6
Q00577 Transcriptional activator protein Pur-alpha PURA 6
P52565 Rho GDP-dissociation inhibitor 1 ARHGDIA 6
Q9Y5Z4 Heme-binding protein 2 HEBP2 6
J3KNF4 Copper chaperone for superoxide dismutase;Superoxide dismutase [Cu-Zn] CCS 6
A0A087WXS7 ATPase ASNA1 ASNA1 6
P05089 Arginase-1 ARG1 6
O95336 6-phosphogluconolactonase PGLS 6
Q92508 Piezo-type mechanosensitive ion channel component 1 PIEZO1 6
P84077 ADP-ribosylation factor 1;ADP-ribosylation factor 3 ARF1;ARF3 6
P21281 V-type proton ATPase subunit B, brain isoform;V-type proton ATPase subunit B, kidney isoform ATP6V1B2;A
TP6V1B1 6
P04259 Keratin, type II cytoskeletal 6B KRT6B 6
Q92905 COP9 signalosome complex subunit 5 COPS5 6
P61163 Alpha-centractin ACTR1A 6
O95373 Importin-7 IPO7 6
C9JD73 Protein phosphatase 1 regulatory subunit 7 PPP1R7 6
Q99536 Synaptic vesicle membrane protein VAT-1 homolog VAT1 6
Q86UX7 Fermitin family homolog 3 FERMT3 6
C9JFE4 COP9 signalosome complex subunit 1 GPS1 6
P36959 GMP reductase 1 GMPR 6
B5MDF5 GTP-binding nuclear protein Ran RAN 6
F5GY90 Porphobilinogen deaminase HMBS 6
E7EX90 Dynactin subunit 1 DCTN1 6
H0Y512 Adipocyte plasma membrane-associated protein APMAP 6
Q32Q12 Nucleoside diphosphate kinase;Nucleoside diphosphate kinase B;Putative nucleoside diphosphate
kinase
NME1-NME2;NME2;NME1;NME2P
1
6
P13807 Glycogen [starch] synthase, muscle GYS1 6
K7ES02 Bleomycin hydrolase BLMH 6
P17213 Bactericidal permeability-increasing protein BPI 6
Q9UNM6 26S proteasome non-ATPase regulatory subunit 13 PSMD13 6
J3KQ32 Obg-like ATPase 1 OLA1 6
Q9Y490 Talin-1 TLN1 6
H0YD13 CD44 antigen CD44 6
P18669 Phosphoglycerate mutase 1;Phosphoglycerate mutase 2;Probable phosphoglycerate mutase 4 PGAM1;PGAM2;PGAM4
6

123
P23528 Cofilin-1 CFL1 6
Q5SR44 Complement receptor type 1 CR1 6
Q99436 Proteasome subunit beta type-7 PSMB7 6
P47756 F-actin-capping protein subunit beta CAPZB 6
P30740 Leukocyte elastase inhibitor SERPINB1 6
A0A024R571 EH domain-containing protein 1 EHD1 6
P30043 Flavin reductase (NADPH) BLVRB 6
P32119 Peroxiredoxin-2 PRDX2 6
C9J0K6 Sorcin SRI 6
P27105 Erythrocyte band 7 integral membrane protein STOM 6
P84077 ADP-ribosylation factor 1;ADP-ribosylation factor 3;ADP-ribosylation factor 5;ADP-ribosylation
factor 4 ARF1;ARF3;A
RF5;ARF4 6
P61981 14-3-3 protein gamma;14-3-3 protein gamma, N-terminally processed YWHAG 5
P09211 Glutathione S-transferase P GSTP1 5
P13489 Ribonuclease inhibitor RNH1 5
Q96PU5 E3 ubiquitin-protein ligase NEDD4-like NEDD4L 5
Q5SRN7 HLA class I histocompatibility antigen, A; HLA class I histocompatibility antigen, B;HLA class I
histocompatibility antigen, Cw HLA-A;HLA-
C;HLA-B 5
P02042 Hemoglobin subunit delta HBD 5
A0A087WU29 Glycophorin-A GYPA 5
P00390 Glutathione reductase, mitochondrial GSR 5
Q9UBV8 Peflin PEF1 5
Q8WVM8 Sec1 family domain-containing protein 1 SCFD1 5
F6TLX2 Glyoxalase domain-containing protein 4 GLOD4 5
H3BQF1 Adenine phosphoribosyltransferase APRT 5
Q07960 Rho GTPase-activating protein 1 ARHGAP1 5
F6XSS0 Blood group Rh(CE) polypeptide;Blood group Rh(D) polypeptide RHCE;RHD 5
A0A087WY55 Vacuolar protein sorting-associated protein VTA1 homolog VTA1 5
G5E9W8 Glycogenin-1 GYG1 5
P07451 Carbonic anhydrase 3 CA3 5
Q08722 Leukocyte surface antigen CD47 CD47 5
X6RA14 S-formylglutathione hydrolase ESD 5
Q8IUI8 Cytokine receptor-like factor 3 CRLF3 5
Q5VW32 BRO1 domain-containing protein BROX BROX 5
P61026 Ras-related protein Rab-10 RAB10 5
A0A087WWY3
Filamin-A FLNA 5
H0YGX7 Rho GDP-dissociation inhibitor 2 ARHGDIB 5
Q92783 Signal transducing adapter molecule 1 STAM 5
Q7Z6Z7 E3 ubiquitin-protein ligase HUWE1 HUWE1 5
H0YHC3 Nucleosome assembly protein 1-like 1 NAP1L1 5
Q5QPM7 Proteasome inhibitor PI31 subunit PSMF1 5
P09104 Gamma-enolase;Enolase ENO2 5
Q5T2B5 Cullin-2 CUL2 5
Q8WW22 DnaJ homolog subfamily A member 4 DNAJA4 5
P61201 COP9 signalosome complex subunit 2 COPS2 5

124
X6R433 Protein-tyrosine-phosphatase;Receptor-type tyrosine-protein phosphatase C PTPRC 5
P63000 Ras-related C3 botulinum toxin substrate 1 RAC1 5
F5GXM3 IST1 homolog IST1 5
H3BLU7 Aflatoxin B1 aldehyde reductase member 2 AKR7A2 5
P25325 3-mercaptopyruvate sulfurtransferase;Sulfurtransferase MPST 5
A0A087X0K1 Calcium-binding protein 39 CAB39 5
P23381 Tryptophan--tRNA ligase, cytoplasmic;T1-TrpRS;T2-TrpRS WARS 5
P01116 GTPase KRas;GTPase KRas, N-terminally processed KRAS 5
P30040 Endoplasmic reticulum resident protein 29 ERP29 5
P05198 Eukaryotic translation initiation factor 2 subunit 1 EIF2S1 5
Q6UX06 Olfactomedin-4 OLFM4 5
Q96KP4 Cytosolic non-specific dipeptidase CNDP2 5
Q04760 Lactoylglutathione lyase GLO1 5
J3QS39 Ubiquitin-60S ribosomal protein L40;Ubiquitin;60S ribosomal protein L40;Ubiquitin-40S ribosomal
protein S27a;Ubiquitin;40S ribosomal protein S27a;Polyubiquitin-B;Ubiquitin;Polyubiquitin-C;Ubiquitin
UBB;RPS27A;UBC;UBA52;
UBBP4 5
P06702 Protein S100-A9 S100A9 5
P15531 Nucleoside diphosphate kinase A;Nucleoside diphosphate kinase;Nucleoside diphosphate kinase B NME1;NME2;NME1-NME2
5
E7EV99 Alpha-adducin ADD1 5
P51149 Ras-related protein Rab-7a RAB7A 4
K7N7A8 Aquaporin-1 AQP1 4
P61020 Ras-related protein Rab-5B RAB5B 4
X6R4N5 Erythroid membrane-associated protein ERMAP 4
E5RJR5 S-phase kinase-associated protein 1 SKP1 4
Q9Y315 Deoxyribose-phosphate aldolase DERA 4
X6R8F3 Neutrophil gelatinase-associated lipocalin LCN2 4
O75396 Vesicle-trafficking protein SEC22b SEC22B 4
Q15102 Platelet-activating factor acetylhydrolase IB subunit gamma PAFAH1B3 4
P51665 26S proteasome non-ATPase regulatory subunit 7 PSMD7 4
Q96FZ7 Charged multivesicular body protein 6 CHMP6 4
Q9NRQ2 Phospholipid scramblase 4 PLSCR4 4
F8VWS0 60S acidic ribosomal protein P0;60S acidic ribosomal protein P0-like RPLP0;RPLP0
P6 4
O14964 Hepatocyte growth factor-regulated tyrosine kinase substrate HGS 4
A0A0A0MTJ9 Neutral cholesterol ester hydrolase 1 NCEH1 4
A0A087WY82 Junctional adhesion molecule A F11R 4
J3QSB7 Purine nucleoside phosphorylase;S-methyl-5-thioadenosine phosphorylase MTAP 4
Q5VVQ6 Ubiquitin thioesterase OTU1 YOD1 4
P36543 V-type proton ATPase subunit E 1 ATP6V1E1 4
Q9BS40 Latexin LXN 4
P47755 F-actin-capping protein subunit alpha-2 CAPZA2 4
Q9GZT8 NIF3-like protein 1 NIF3L1 4
H6UYS7 Alpha-synuclein SNCA 4
P60953 Cell division control protein 42 homolog CDC42 4
Q14773 Intercellular adhesion molecule 4 ICAM4 4

125
H0Y6T7 Nicastrin NCSTN 4
P69891 Hemoglobin subunit gamma-1 HBG1 4
P14625 Endoplasmin HSP90B1 4
Q96GD0 Pyridoxal phosphate phosphatase PDXP 4
P08311 Cathepsin G CTSG 4
Q9H9Q2 COP9 signalosome complex subunit 7b COPS7B 4
P09417 Dihydropteridine reductase QDPR 4
F8WE6 Peptidyl-prolyl cis-trans isomerase;Peptidyl-prolyl cis-trans isomerase A PPIA 4
O15173 Membrane-associated progesterone receptor component 2 PGRMC2 4
P28072 Proteasome subunit beta type-6;Proteasome subunit beta type PSMB6 4
H3BSW0 Leucine-rich repeat-containing protein 57 LRRC57 4
Q9UBW8 COP9 signalosome complex subunit 7a COPS7A 4
O00560 Syntenin-1 SDCBP 4
F5H157 Ras-related protein Rab-35 RAB35 4
Q9H479 Fructosamine-3-kinase FN3K 4
Q04917 14-3-3 protein eta YWHAH 4
C9JJ47 AP-2 complex subunit mu AP2M1 4
C9JIG9 Serine/threonine-protein kinase OSR1 OXSR1 4
Q13336 Urea transporter 1 SLC14A1 4
P46926 Glucosamine-6-phosphate isomerase 1;Glucosamine-6-phosphate isomerase;Glucosamine-6-phosphate
isomerase 2 GNPDA1;GNP
DA2 4
O43633 Charged multivesicular body protein 2a CHMP2A 4
F8VVB9 Tubulin alpha-1A chain;Tubulin alpha-1C chain;Tubulin alpha-1B chain;Tubulin alpha-3C/D
chain;Tubulin alpha-3E chain
TUBA1B;TUBA1C;TUBA1A;TUBA3C;TU
BA3E
4
A6PVN5 Serine/threonine-protein phosphatase 2A activator PPP2R4 4
B8ZZB8 CB1 cannabinoid receptor-interacting protein 1 CNRIP1 4
Q9P2R3 Rabankyrin-5 ANKFY1 4
Q86YS7 C2 domain-containing protein 5 C2CD5 4
R4GMR5 26S proteasome non-ATPase regulatory subunit 8 PSMD8 4
E7EM64 COP9 signalosome complex subunit 6 COPS6 4
J3KNI6 Integrin beta;Integrin beta-2 ITGB2 4
H0Y5R6 Uroporphyrinogen decarboxylase UROD 4
M0R165 Epidermal growth factor receptor substrate 15-like 1 EPS15L1 4
O00487 26S proteasome non-ATPase regulatory subunit 14 PSMD14 4
Q13630 GDP-L-fucose synthase TSTA3 4
P55060 Exportin-2 CSE1L 4
P20020 Plasma membrane calcium-transporting ATPase 1;Calcium-transporting ATPase ATP2B1 4
Q9NYU2 UDP-glucose:glycoprotein glucosyltransferase 1 UGGT1 4
H3BND8 Ubiquitin carboxyl-terminal hydrolase;Ubiquitin carboxyl-terminal hydrolase 7 USP7 4
Q9GZP4 PITH domain-containing protein 1 PITHD1 4
D6RD66 WD repeat-containing protein 1 WDR1 4
P48729 Casein kinase I isoform alpha CSNK1A1 4
P25685 DnaJ homolog subfamily B member 1 DNAJB1 4
P14550 Alcohol dehydrogenase [NADP(+)] AKR1A1 4

126
Q6PCE3 Glucose 1,6-bisphosphate synthase PGM2L1 4
Q9UPN7 Serine/threonine-protein phosphatase 6 regulatory subunit 1 PPP6R1 4
P62805 Histone H4 HIST1H4A 4
P62937 Peptidyl-prolyl cis-trans isomerase A PPIA 4
P30046 D-dopachrome decarboxylase;D-dopachrome decarboxylase-like protein DDT;DDTL 4
Q08495 Dematin DMTN 4
K7EIJ0 WW domain-binding protein 2 WBP2 4
E5RHP7 Carbonic anhydrase 1 CA1 4
P09105 Hemoglobin subunit theta-1 HBQ1 4
E7ESC6 Exportin-7 XPO7 4
P61225 Ras-related protein Rap-2b;Ras-related protein Rap-2c;Ras-related protein Rap-2a RAP2B;RAP2
A;RAP2C 4
O15400 Syntaxin-7 STX7 3
Q5VZR0 Golgi-associated plant pathogenesis-related protein 1 GLIPR2 3
P40199 Carcinoembryonic antigen-related cell adhesion molecule 6 CEACAM6 3
P27348 14-3-3 protein theta YWHAQ 3
P62820 Ras-related protein Rab-1A RAB1A 3
C9JEN3 Protein lifeguard 3 TMBIM1 3
Q9UDX3 SEC14-like protein 4 SEC14L4 3
Q9Y570 Protein phosphatase methylesterase 1 PPME1 3
Q96GG9 DCN1-like protein 1;DCN1-like protein DCUN1D1 3
B0YJC4 Vimentin VIM 3
Q86VN1 Vacuolar protein-sorting-associated protein 36 VPS36 3
F8WFB9 Endophilin-B2 SH3GLB2 3
A0A087WVQ9
Elongation factor 1-alpha 1;Putative elongation factor 1-alpha-like 3 EEF1A1;EEF1
A1P5 3
Q53TN4 Cytochrome b reductase 1 CYBRD1 3
P10809 60 kDa heat shock protein, mitochondrial HSPD1 3
F5H7X1 26S proteasome non-ATPase regulatory subunit 9 PSMD9 3
E9PNW4 CD59 glycoprotein CD59 3
E9PS74 Solute carrier family 43 member 3 SLC43A3 3
P53985 Monocarboxylate transporter 1 SLC16A1 3
Q8WWI5 Choline transporter-like protein 1 SLC44A1 3
E9PIR7 Thioredoxin reductase 1, cytoplasmic TXNRD1 3
Q5TD07 Ribosyldihydronicotinamide dehydrogenase [quinone] NQO2 3
Q9UL25 Ras-related protein Rab-21 RAB21 3
P27824 Calnexin CANX 3
U3KPS2 Myeloblastin PRTN3 3
Q9UKU0 Long-chain-fatty-acid--CoA ligase 6 ACSL6 3
P24666 Low molecular weight phosphotyrosine protein phosphatase ACP1 3
H7C2G2 NAD(P)(+)--arginine ADP-ribosyltransferase;Ecto-ADP-ribosyltransferase 4 ART4 3
I3L1K6 Myosin light chain 4 MYL4 3
Q9UK41 Vacuolar protein sorting-associated protein 28 homolog VPS28 3
Q9NUQ9 Protein FAM49B FAM49B 3
Q9UBQ7 Glyoxylate reductase/hydroxypyruvate reductase GRHPR 3

127
B8ZZG1 MAGUK p55 subfamily member 6 MPP6 3
Q10567 AP-1 complex subunit beta-1 AP1B1 3
O75387 Large neutral amino acids transporter small subunit 3 SLC43A1 3
Q9BTU6 Phosphatidylinositol 4-kinase type 2-alpha PI4K2A 3
J3KS22 L-xylulose reductase DCXR 3
O15498 Synaptobrevin homolog YKT6 YKT6 3
Q08211 ATP-dependent RNA helicase A DHX9 3
P50416 Carnitine O-palmitoyltransferase 1, liver isoform CPT1A 3
P08237 ATP-dependent 6-phosphofructokinase, muscle type PFKM 3
P14735 Insulin-degrading enzyme IDE 3
Q9H0R3 Transmembrane protein 222 TMEM222 3
Q14166 Tubulin--tyrosine ligase-like protein 12 TTLL12 3
Q14558 Phosphoribosyl pyrophosphate synthase-associated protein 1 PRPSAP1 3
A0A087WTB8 Ubiquitin carboxyl-terminal hydrolase;Ubiquitin carboxyl-terminal hydrolase isozyme L3 UCHL3 3
Q9NPQ8 Synembryn-A RIC8A 3
F6WQW2 Ran-specific GTPase-activating protein RANBP1 3
Q15691 Microtubule-associated protein RP/EB family member 1 MAPRE1 3
A0A0A0MR50
Cullin-4A CUL4A 3
Q96IU4 Alpha/beta hydrolase domain-containing protein 14B ABHD14B 3
Q9P0L0 Vesicle-associated membrane protein-associated protein A VAPA 3
E9PRY8 Elongation factor 1-delta EEF1D 3
Q16543 Hsp90 co-chaperone Cdc37 CDC37 3
P06702 Protein S100-A9 S100A9 3
P13639 Elongation factor 2 EEF2 3
E9PJC7 Tetraspanin;CD82 antigen CD82 3
Q01432 AMP deaminase 3 AMPD3 3
B1AUU8 Epidermal growth factor receptor substrate 15 EPS15 3
P54709 Sodium/potassium-transporting ATPase subunit beta-3 ATP1B3 3
P54727 UV excision repair protein RAD23 homolog B RAD23B 3
P36507 Dual specificity mitogen-activated protein kinase kinase 2 MAP2K2 3
Q9Y2V2 Calcium-regulated heat stable protein 1 CARHSP1 3
P31146 Coronin-1A;Coronin CORO1A 3
Q8IY17 Neuropathy target esterase PNPLA6 3
E7EQR4 Ezrin EZR 3
F8WDS9 LanC-like protein 1 LANCL1 3
X6RJP6 Transgelin-2 TAGLN2 3
A0A0J9YXM6 WD repeat-containing protein 81 WDR81 3
P62140 Serine/threonine-protein phosphatase PP1-beta catalytic subunit;Serine/threonine-protein phosphatase PPP1CB 3
H0YLJ3 Mortality factor 4-like protein 1 MORF4L1 3
E7EPV7 Alpha-synuclein SNCA 3
K7EK07 Histone H3 H3F3B 3
P05109 Protein S100-A8;Protein S100-A8, N-terminally processed S100A8 3
I3L0A0 Ubiquitin-conjugating enzyme E2 variant 1;Ubiquitin-conjugating enzyme E2 variant 2 TMEM189-
UBE2V1;UBE2V1;UBE2V2
3

128
Q6B0K9 Hemoglobin subunit mu HBM 3
O75368 SH3 domain-binding glutamic acid-rich-like protein SH3BGRL 3
A0A0J9YXB3 Ras-related protein Rap-1b;Ras-related protein Rap-1A;Ras-related protein Rap-1b-like protein RAP1B;RAP1
A 3
P60953 Cell division control protein 42 homolog CDC42 3
A0A087WTI1 Ras-related protein Rab-1B;Ras-related protein Rab-1A RAB1B;RAB1
A 3
B5MDF5 GTP-binding nuclear protein Ran RAN 3
P61088 Ubiquitin-conjugating enzyme E2 N UBE2N 3
Q00013 55 kDa erythrocyte membrane protein MPP1 3
B1AKQ8 Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-1;Guanine nucleotide-binding protein
G(I)/G(S)/G(T) subunit beta-3;Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2 GNB1;GNB2;
GNB3 3
P04406 Glyceraldehyde-3-phosphate dehydrogenase GAPDH 3
Q9UJC5 SH3 domain-binding glutamic acid-rich-like protein 2 SH3BGRL2 3
P04921 Glycophorin-C GYPC 2
Q9NP59 Solute carrier family 40 member 1 SLC40A1 2
K7EKH5 Fructose-bisphosphate aldolase;Fructose-bisphosphate aldolase C ALDOC 2
P11233 Ras-related protein Ral-A RALA 2
F8WBR5 Calmodulin CALM2;CALM3;CALM1
2
P00441 Superoxide dismutase [Cu-Zn] SOD1 2
H0YDI1 Lymphocyte function-associated antigen 3 CD58 2
H0YNE9 Ras-related protein Rab-8B RAB8B 2
J3KN67 TPM3 2
P20160 Azurocidin AZU1 2
A0A087WZZ4 Ammonium transporter Rh type A RHAG 2
F5GXS0 Complement C4-A;Complement C4-B C4B;C4A 2
Q8ND76 Cyclin-Y CCNY 2
Q14739 Lamin-B receptor LBR 2
Q15181 Inorganic pyrophosphatase PPA1 2
J3QS92 Galectin-9 LGALS9 2
I3L471 Phosphatidylinositol transfer protein alpha isoform PITPNA 2
P01111 GTPase NRas NRAS;KRAS 2
F5H4Q5 Vacuolar protein sorting-associated protein 37C VPS37C 2
A0A0A0MSW4
Phosphatidylinositol transfer protein beta isoform PITPNB 2
P67775 Serine/threonine-protein phosphatase 2A catalytic subunit alpha isoform;Serine/threonine-protein
phosphatase PPP2CA;PPP2
CB 2
P41091 Eukaryotic translation initiation factor 2 subunit 3;Putative eukaryotic translation initiation factor 2
subunit 3-like protein EIF2S3;EIF2S
3L 2
A0A087WWS7
Syntaxin-binding protein 2 STXBP2 2
F8VQX6 Methyltransferase-like protein 7A METTL7A 2
B3KT28 FAS-associated factor 1 FAF1 2
K7EIJ8 Katanin p60 ATPase-containing subunit A-like 2 KATNAL2 2
P20339 Ras-related protein Rab-5A RAB5A 2
O95456 Proteasome assembly chaperone 1 PSMG1 2
K7EK45 Polypyrimidine tract-binding protein 1 PTBP1 2

129
Q15365 Poly(rC)-binding protein 1;Poly(rC)-binding protein 3 PCBP1;PCBP3 2
Q9BSJ8 Extended synaptotagmin-1 ESYT1 2
H3BP35 Diphosphomevalonate decarboxylase MVD 2
H0Y8C4 Serine/threonine-protein phosphatase 2A 56 kDa regulatory subunit delta isoform PPP2R5D 2
K7EP09 Bifunctional coenzyme A synthase;Phosphopantetheine adenylyltransferase;Dephospho-CoA kinase COASY 2
F8VNT9 Tetraspanin;CD63 antigen CD63 2
Q8NEV1 Casein kinase II subunit alpha 3;Casein kinase II subunit alpha CSNK2A3;CS
NK2A1 2
F8VTQ5 Heterogeneous nuclear ribonucleoprotein A1 HNRNPA1 2
A0A087X2E2 Carcinoembryonic antigen-related cell adhesion molecule 8;Carcinoembryonic antigen-related cell
adhesion molecule 1;Carcinoembryonic antigen-related cell adhesion molecule 5
CEACAM8;CEACAM5;CEA
CAM1 2
H0YAS8 Clusterin;Clusterin beta chain;Clusterin alpha chain;Clusterin CLU 2
E9PP54 Tubulin-specific chaperone cofactor E-like protein TBCEL 2
P43034 Platelet-activating factor acetylhydrolase IB subunit alpha PAFAH1B1 2
Q9NT62 Ubiquitin-like-conjugating enzyme ATG3 ATG3 2
K7ERZ3 Perilipin-3 PLIN3 2
Q6DD88 Atlastin-3 ATL3 2
Q15084-3 Protein disulfide-isomerase A6 PDIA6 2
K7EQH4 ATP synthase subunit alpha, mitochondrial ATP5A1 2
G3V1U5 Vesicle transport protein GOT1B GOLT1B 2
O00186 Syntaxin-binding protein 3 STXBP3 2
Q5T6W2 Heterogeneous nuclear ribonucleoprotein K HNRNPK 2
H0YEY4 ADP-sugar pyrophosphatase NUDT5 2
P35580 Myosin-10 MYH10 2
F5H081 Solute carrier family 2, facilitated glucose transporter member 4 SLC2A4 2
Q9H1C7 Cysteine-rich and transmembrane domain-containing protein 1 CYSTM1 2
K7EJ83 Cyclin-dependent kinase 2;Cyclin-dependent kinase 3 CDK3;CDK2 2
C9J352 Importin subunit alpha-5;Importin subunit alpha-5, N-terminally processed KPNA1 2
H9KV75 Alpha-actinin-1;Alpha-actinin-4;Alpha-actinin-2;Alpha-actinin-3 ACTN1;ACTN4;ACTN3;ACT
N2 2
A0A0A0MQS1
Pyrroline-5-carboxylate reductase;Pyrroline-5-carboxylate reductase 3 PYCRL 2
D6RBY0 Rieske domain-containing protein RFESD 2
C9JC71 Low affinity immunoglobulin gamma Fc region receptor III-A FCGR3A 2
P51811 Membrane transport protein XK XK 2
C9J1G2 DnaJ homolog subfamily B member 2 DNAJB2 2
Q9UN37 Vacuolar protein sorting-associated protein 4A;Vacuolar protein sorting-associated protein
4B;Fidgetin-like protein 1 VPS4A;VPS4B
;FIGNL1 2
P61160 Actin-related protein 2 ACTR2 2
P16930 Fumarylacetoacetase FAH 2
A0A087X0K4 CUB and sushi domain-containing protein 2 CSMD2 2
P10746 Uroporphyrinogen-III synthase UROS 2
K7EQ02 DAZ-associated protein 1 DAZAP1 2
F5GYN4 Ubiquitin thioesterase OTUB1 OTUB1 2
P10599 Thioredoxin TXN 2

130
O95197 Reticulon-3 RTN3 2
E7ETB3 Aspartyl aminopeptidase DNPEP 2
P14868 Aspartate--tRNA ligase, cytoplasmic DARS 2
Q08AM6 Protein VAC14 homolog VAC14 2
Q93034 Cullin-5 CUL5 2
Q9BQA1 Methylosome protein 50 WDR77 2
A0A0B4J2G9 Ubiquitin-conjugating enzyme E2 L3 UBE2L3 2
Q04446 1,4-alpha-glucan-branching enzyme GBE1 2
Q96NA2 Rab-interacting lysosomal protein RILP 2
Q92539 Phosphatidate phosphatase LPIN2 LPIN2 2
P28482 Mitogen-activated protein kinase 1 MAPK1 2
Q7Z406 Myosin-14;Myosin-11 MYH14;MYH
11 2
Q8IU68 Transmembrane channel-like protein 8 TMC8 2
Q96BJ3 Axin interactor, dorsalization-associated protein AIDA 2
P20042 Eukaryotic translation initiation factor 2 subunit 2 EIF2S2 2
A0A0A0MQR0
Docosahexaenoic acid omega-hydroxylase CYP4F3;Phylloquinone omega-hydroxylase CYP4F2;Cytochrome P450 4F12;Phylloquinone omega-hydroxylase CYP4F11
CYP4F2;CYP4F3;CYP4F11;C
YP4F12 2
Q9BS26 Endoplasmic reticulum resident protein 44 ERP44 2
O00178 GTP-binding protein 1 GTPBP1 2
P27797 Calreticulin CALR 2
G3V0E5 Transferrin receptor protein 1;Transferrin receptor protein 1, serum form TFRC 2
P48147 Prolyl endopeptidase PREP 2
H7BZC1 Hippocalcin-like protein 1;Neuron-specific calcium-binding protein hippocalcin;Neurocalcin-delta HPCAL1;NCA
LD;HPCA 2
F8VPD4 CAD protein;Glutamine-dependent carbamoyl-phosphate synthase;Aspartate
carbamoyltransferase;Dihydroorotase CAD 2
Q9NTJ5 Phosphatidylinositide phosphatase SAC1 SACM1L 2
P26447 Protein S100-A4 S100A4 2
Q9UIW2 Plexin-A1 PLXNA1 2
H0YJS0 V-type proton ATPase subunit D ATP6V1D 2
P25445 Tumor necrosis factor receptor superfamily member 6 FAS 2
C9JEU5 Fibrinogen gamma chain FGG 2
F5H562 Copper-transporting ATPase 2;WND/140 kDa ATP7B 2
B5MCF3 Protein GUCD1 GUCD1 2
Q96TA1 Niban-like protein 1 FAM129B 2
F5GWT4 Serine/threonine-protein kinase WNK1 WNK1 2
E9PLT1 Platelet glycoprotein 4 CD36 2
Q8TDB8 Solute carrier family 2, facilitated glucose transporter member 14;Solute carrier family 2, facilitated
glucose transporter member 3 SLC2A14;SLC
2A3 2
B4DDD6 Drebrin-like protein DBNL 2
E9PII3 Band 4.1-like protein 2 EPB41L2 2
E9PNF5 Glutamine-dependent NAD(+) synthetase NADSYN1 2
K7ELL7 Glucosidase 2 subunit beta PRKCSH 2
Q9BV20 Methylthioribose-1-phosphate isomerase MRI1 2
J3KNB4 Cathelicidin antimicrobial peptide;Antibacterial protein FALL-39;Antibacterial protein LL-37 CAMP 2

131
Q5T1Z0 Phospholysine phosphohistidine inorganic pyrophosphate phosphatase LHPP 2
H3BLV0 Complement decay-accelerating factor CD55 2
Q9Y3E7 Charged multivesicular body protein 3 CHMP3 2
Q5T6H7 Xaa-Pro aminopeptidase 1 XPNPEP1 2
O94779 Contactin-5 CNTN5 2
E9PNR2 Ras and Rab interactor 1 RIN1 2
E7ESJ7 Protein FAM114A2 FAM114A2 2
A0A0A0MS99 Multidrug resistance-associated protein 1 ABCC1 2
A0A0G2JM15 Large neutral amino acids transporter small subunit 4 SLC43A2 2
H0YBF7 Arf-GAP with SH3 domain, ANK repeat and PH domain-containing protein 1;Arf-GAP with SH3
domain, ANK repeat and PH domain-containing protein 2 ASAP1;ASAP
2 2
P29992 Guanine nucleotide-binding protein subunit alpha-11 GNA11 2
F8WF69 Clathrin light chain A CLTA 2
B5MCN0 Atlastin-2 ATL2 2
F8W9F9 Serine/threonine-protein kinase WNK2;Serine/threonine-protein kinase WNK3 WNK2;WNK3 2
A2A3F3 Transient receptor potential cation channel subfamily M member 3 TRPM3 2
Q5KU26 Collectin-12 COLEC12 2
O14523 C2 domain-containing protein 2-like C2CD2L 2
Q96DG6 Carboxymethylenebutenolidase homolog CMBL 2
Q5THJ4 Vacuolar protein sorting-associated protein 13D VPS13D 2
Q02790 Peptidyl-prolyl cis-trans isomerase FKBP4;Peptidyl-prolyl cis-trans isomerase FKBP4, N-terminally
processed FKBP4 2
A0A0G2JQD2 Glutathione S-transferase theta-1 GSTT1 2
Q9NV96 Cell cycle control protein 50A;Cell cycle control protein 50B TMEM30A;T
MEM30B 2
P62330 ADP-ribosylation factor 6 ARF6 2
Q9UPU5 Ubiquitin carboxyl-terminal hydrolase 24 USP24 2
B2R4S9 Histone H2B HIST1H2BI 2
P10599 Thioredoxin TXN 2
R4GN98 Protein S100;Protein S100-A6 S100A6 2
H3BS66 Small integral membrane protein 1 SMIM1 2
Q8NHG7 Small VCP/p97-interacting protein SVIP 2
C9JIG9 Serine/threonine-protein kinase OSR1;STE20/SPS1-related proline-alanine-rich protein kinase OXSR1;STK39 2
Q5VZR0 Golgi-associated plant pathogenesis-related protein 1 GLIPR2 2
F8WD49 Anion exchange protein 3;Anion exchange protein;Anion exchange protein 2 SLC4A3;SLC4
A2 2
P61626 Lysozyme C;Lysozyme LYZ 2
Q5T123 SH3 domain-binding glutamic acid-rich-like protein 3 SH3BGRL3 2
F5H571 Ubiquitin carboxyl-terminal hydrolase 5 USP5 2
Q9H3K6 BolA-like protein 2 BOLA2;BOLA
2B 2
P05164 Myeloperoxidase;Myeloperoxidase;89 kDa myeloperoxidase;84 kDa
myeloperoxidase;Myeloperoxidase light chain;Myeloperoxidase heavy chain MPO 2
P50395 Rab GDP dissociation inhibitor beta GDI2 2
G3V2U7 Acylphosphatase;Acylphosphatase-1 ACYP1 2
J3KNB4 Cathelicidin antimicrobial peptide;Antibacterial protein FALL-39;Antibacterial protein LL-37 CAMP 2
H0Y9X3 Programmed cell death protein 6 PDCD6 2

132
A6NJA2 Ubiquitin carboxyl-terminal hydrolase;Ubiquitin carboxyl-terminal hydrolase 14 USP14 2
P35998 26S protease regulatory subunit 7 PSMC2 2
P30041 Peroxiredoxin-6 PRDX6 2
P35612 Beta-adducin ADD2 2
P00918 Carbonic anhydrase 2 CA2 2
A0A0C4DGH5
Cullin-associated NEDD8-dissociated protein 1 CAND1 2
C9JZN1 Guanine nucleotide-binding protein G(I)/G(S)/G(T) subunit beta-2;Guanine nucleotide-binding protein
subunit beta-4 GNB2;GNB4 1
I3L4X8 Integrin beta;Integrin beta-3 ITGB3 1
H7BZJ3 PDIA3 1
P17066 Heat shock 70 kDa protein 6;Putative heat shock 70 kDa protein 7 HSPA6;HSPA
7 1
J3KT70 Arf-GAP with dual PH domain-containing protein 2 ADAP2;CENT
A2 1
Q9Y6M5 Zinc transporter 1 SLC30A1 1
P54725 UV excision repair protein RAD23 homolog A RAD23A 1
F8WD59 40S ribosomal protein SA RPSA;RPSAP5
8 1
P06753 1
E9PMI6 Methylosome subunit pICln CLNS1A 1
Q9BVK6 Transmembrane emp24 domain-containing protein 9 TMED9 1
C9JJV6 Myeloid-associated differentiation marker MYADM 1
Q8IXQ3 Uncharacterized protein C9orf40 C9orf40 1
Q8NDC0 MAPK-interacting and spindle-stabilizing protein-like MAPK1IP1L 1
Q99747 Gamma-soluble NSF attachment protein NAPG 1
E7EWE1 Ubiquitin-like modifier-activating enzyme 5 UBA5 1
J3KNE3 Platelet-activating factor acetylhydrolase IB subunit beta PAFAH1B2 1
Q8IUI8 Cytokine receptor-like factor 3 CRLF3 1
Q9BTX7 Alpha-tocopherol transfer protein-like TTPAL 1
P63261 Actin, cytoplasmic 2;Actin, cytoplasmic 2, N-terminally processed ACTG1 1
P31153 S-adenosylmethionine synthase isoform type-2 MAT2A 1
O00560 Syntenin-1 SDCBP 1
Q4VB86 Protein 4.1 EPB41 1
Q8WUD1 Ras-related protein Rab-2B RAB2B;DKFZ
p313C1541 1
Q9NWV4 UPF0587 protein C1orf123 C1orf123 1
Q9Y2Z0 Suppressor of G2 allele of SKP1 homolog SUGT1 1
H0YK48 Tropomyosin alpha-1 chain TPM1 1
H3BS66 Small integral membrane protein 1 SMIM1 1
Q5VTS0 Neurensin-1 NRSN1 1
H0Y904 Multidrug resistance-associated protein 7 ABCC10 1
P08F94 Fibrocystin PKHD1 1
G3V5X4 Nesprin-2 SYNE2 1
P48507 Glutamate--cysteine ligase regulatory subunit GCLM 1
H3BUF4 Cyclin-D1-binding protein 1 CCNDBP1 1
F8WB30 Target of Myb protein 1 TOM1 1

133
E5RJI8 CA1 1
S4R3E5 Importin subunit alpha-7 KPNA6 1
E7EVS6 ACTB 1
P04206 Ig kappa chain V-III region GOL;Ig kappa chain V-III region WOL;Ig kappa chain V-III region Ti;Ig
kappa chain V-III region SIE 1
Q5SSV3 N(G),N(G)-dimethylarginine dimethylaminohydrolase 2 DDAH2 1
H0YG54 Oligoribonuclease, mitochondrial REXO2 1
P08246 Neutrophil elastase ELANE 1
P40926 Malate dehydrogenase, mitochondrial MDH2 1
P98172 Ephrin-B1 EFNB1 1
Q96DD7 Protein shisa-4 SHISA4 1
Q71RC9 Small integral membrane protein 5 SMIM5 1
P68133 Actin, alpha skeletal muscle;Actin, alpha cardiac muscle 1;Actin, gamma-enteric smooth muscle;Actin,
aortic smooth muscle
ACTA1;ACTC1;ACTG2;ACT
A2 1
H0Y9Q6 Clathrin light chain B CLTB 1
G3V5P0 KTN1 1
Q9NWX6 Probable tRNA(His) guanylyltransferase THG1L 1
Q15404 Ras suppressor protein 1 RSU1 1
B2R4S9 Histone H2B HIST1H2B 1
K7EK06 Phenylalanine--tRNA ligase alpha subunit FARSA 1
H7C3S9 COP9 signalosome complex subunit 8 COPS8 1
J3KRV4 Dual specificity mitogen-activated protein kinase kinase 3 MAP2K3 1
Q15042 Rab3 GTPase-activating protein catalytic subunit RAB3GAP1 1
P01893
Putative HLA class I histocompatibility antigen, alpha chain H;HLA class I histocompatibility antigen, Cw-6 alpha chain;HLA class I histocompatibility antigen, B-38 alpha chain;HLA class I
histocompatibility antigen, B-67 alpha chain;HLA class I histocompatibility antigen, B-82 alpha chain;HLA class I histocompatibility antigen, B-39 alpha chain;HLA class I histocompatibility antigen,
Cw-18 alpha chain;HLA class I histocompatibility antigen, Cw-7 alpha chain;HLA class I histocompatibility antigen, B-42 alpha chain;HLA class I histocompatibility antigen, B-14 alpha
chain;HLA class I histocompatibility antigen, B-8 alpha chain;HLA class I histocompatibility antigen, B-7 alpha chain
HLA-H;HLA-C;HLA-B
1
Q9UNW1 Multiple inositol polyphosphate phosphatase 1 MINPP1 1
Q6B0K9 Hemoglobin subunit mu HBM 1
O75915 PRA1 family protein 3 ARL6IP5 1
O95376 E3 ubiquitin-protein ligase ARIH2 ARIH2 1
E9PBW4 Hemoglobin subunit gamma-2 HBG2 1
C9J1X0 WD repeat-containing protein 91 WDR91 1
Q8NCV1 1
Q9Y4P8 WD repeat domain phosphoinositide-interacting protein 2 WIPI2 1
A0A087WUX6
Proteasomal ubiquitin receptor ADRM1 ADRM1 1
Q9NQS7 Inner centromere protein INCENP 1
P62805 Histone H4 HIST1H4A 1
Q8TDY2 RB1-inducible coiled-coil protein 1 RB1CC1 1
C9JFM5 Syntaxin-4 STX4 1
B0QZ43 Erlin-2;Erlin-1 ERLIN1;ERLI
N2 1
C9J8T0 Selenocysteine-specific elongation factor EEFSEC 1

134
P61224 Ras-related protein Rap-1b;Ras-related protein Rap-1b-like protein RAP1B 1
Q99828 Calcium and integrin-binding protein 1 CIB1 1
Q53GQ0 Very-long-chain 3-oxoacyl-CoA reductase HSD17B12 1
H0Y9Q9 ADP-ribosyl cyclase/cyclic ADP-ribose hydrolase 2 BST1 1
R4GN98 Protein S100;Protein S100-A6 S100A6 1
Q8TB73 Protein NDNF NDNF 1
Q6P1A2 Lysophospholipid acyltransferase 5 LPCAT3 1
F2Z2Y4 Pyridoxal kinase PDXK 1
Q9HA65 TBC1 domain family member 17 TBC1D17 1
G3V126 V-type proton ATPase subunit H ATP6V1H 1
D6RGE2 Isochorismatase domain-containing protein 1 ISOC1 1
A0A0B4J222 ADP-ribosylation factor-like protein 15 ARL15 1
F8W7W4 Androglobin ADGB 1
Q9GZR7 1
A0A087WVC4
cAMP-dependent protein kinase catalytic subunit beta PRKACB 1
A8MU39 Serine/threonine-protein phosphatase;Serine/threonine-protein phosphatase 5 PPP5C 1
G3V1N2 HBA2 1
P69892 Hemoglobin subunit gamma-2 HBG2 1
F8WEZ0 1-phosphatidylinositol 3-phosphate 5-kinase PIKFYVE 1
A0A087WZZ4 Ammonium transporter Rh type A RHAG 1
Q1JUQ3 Peptidyl-prolyl cis-trans isomerase;Peptidyl-prolyl cis-trans isomerase FKBP1A FKBP12-
Exin;FKBP1A 1
F8VWZ5 H2.0-like homeobox protein HLX 1
P62877 E3 ubiquitin-protein ligase RBX1;E3 ubiquitin-protein ligase RBX1, N-terminally processed RBX1 1
Q7Z5P9 Mucin-19 MUC19 1
F8VPB3 TPK1 1
F8WBF4 Transmembrane protein 50B TMEM50B 1
C9JL85 Myotrophin MTPN 1
Q53EQ6 Tigger transposable element-derived protein 5 TIGD5 1
E3W974 DNPEP 1
H7BYV1 Interferon-induced transmembrane protein 1;Interferon-induced transmembrane protein 2;Interferon-
induced transmembrane protein 3 IFITM2;IFITM
3;IFITM1 1
D6RC06 Histidine triad nucleotide-binding protein 1 HINT1 1
E9PNW4 CD59 glycoprotein CD59 1
H7C2Z6 Grancalcin GCA 1
Q9H1C7 Cysteine-rich and transmembrane domain-containing protein 1 CYSTM1 1
H0YK07 ATP-dependent Clp protease ATP-binding subunit clpX-like, mitochondrial CLPX 1
E9PG15 14-3-3 protein theta YWHAQ 1
K7ENK9 Vesicle-associated membrane protein 3;Vesicle-associated membrane protein 2 VAMP2;VAM
P3 1
F8WDD6 Cation channel sperm-associated protein subunit gamma CATSPERG 1
P35754 Glutaredoxin-1 GLRX 1
Q5THJ4 Vacuolar protein sorting-associated protein 13D VPS13D 1
B4E3H6 Transforming acidic coiled-coil-containing protein 1 TACC1 1
B0YJC4 Vimentin VIM 1

135
H7C5R6 AT-rich interactive domain-containing protein 4B ARID4B 1
P49755 Transmembrane emp24 domain-containing protein 10 TMED10 1
E9PMJ3 Ribonuclease inhibitor RNH1 1
B1AHA9 DNA replication licensing factor MCM5 MCM5 1
P23634 Plasma membrane calcium-transporting ATPase 4 ATP2B4 1
Q14152 Eukaryotic translation initiation factor 3 subunit A EIF3A 1
C9IY70 60S ribosomal export protein NMD3 NMD3 1
A0A087WUS7 Ig delta chain C region IGHD 1
P07384 Calpain-1 catalytic subunit CAPN1 1
G3V2C9 Guanine nucleotide-binding protein subunit gamma;Guanine nucleotide-binding protein
G(I)/G(S)/G(O) subunit gamma-2 GNG2 1
Q9Y2Y8 Proteoglycan 3 PRG3 1
V9GY70 DNAJB12 1
U5GXS0 MAM and LDL-receptor class A domain-containing protein 1 MALRD1 1
Q92625 Ankyrin repeat and SAM domain-containing protein 1A ANKS1A 1
![[halshs-00609506, v1] La loi du 20 août 2008 et ses ... · lÕ tablissement concern . Le droit syndical dans les entreprises peut galement constituer un l ment d cisi f pour accro](https://static.fdocuments.fr/doc/165x107/5fad620c74715e57454a6be9/halshs-00609506-v1-la-loi-du-20-aot-2008-et-ses-l-tablissement-concern.jpg)