
TS-tp-p2B-10 La plante domestiquée -...
13
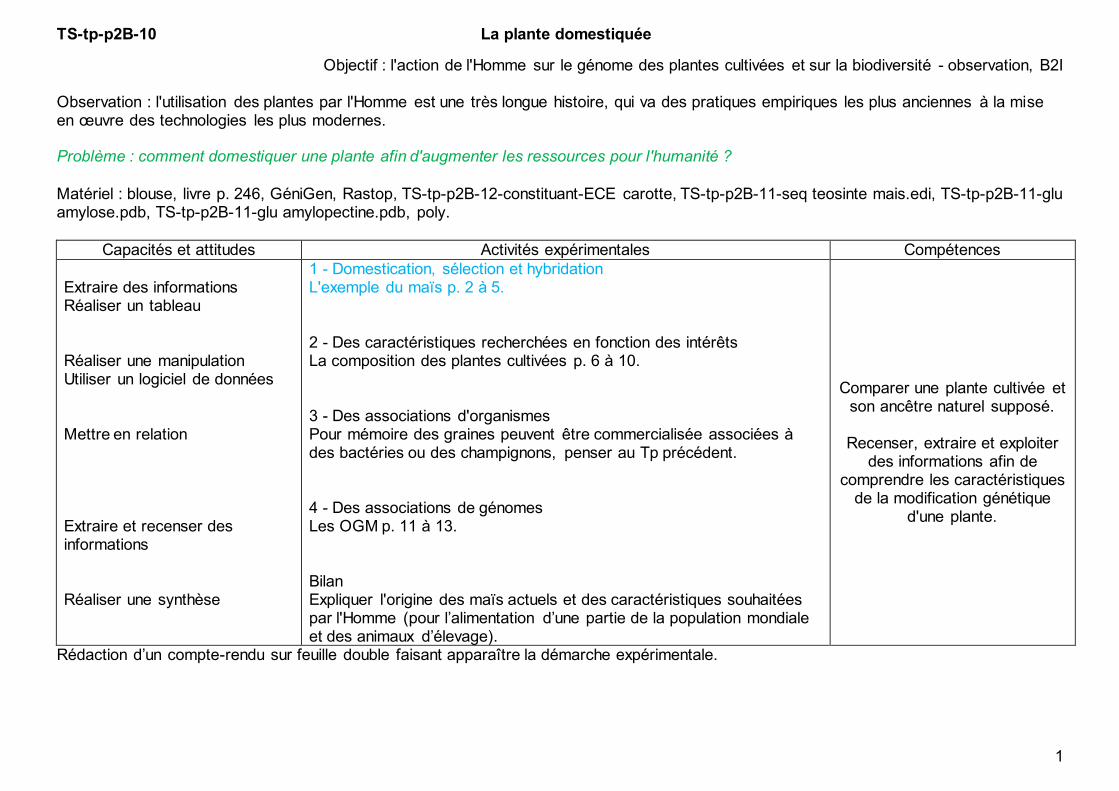
TS-tp-p2B-10 La plante domestiquée 1 Objectif : l'action de l'Homme sur le génome des plantes cultivées et sur la biodiversité - observation, B2I Observation : l'utilisation des plantes par l'Homme est une très longue histoire, qui va des pratiques empiriques les plus anciennes à la mise en œuvre des technologies les plus modernes. Problème : comment domestiquer une plante afin d'augmenter les ressources pour l'humanité ? Matériel : blouse, livre p. 246, GéniGen, Rastop, TS-tp-p2B-12-constituant-ECE carotte, TS-tp-p2B-11-seq teosinte mais.edi, TS-tp-p2B-11-glu amylose.pdb, TS-tp-p2B-11-glu amylopectine.pdb, poly. Capacités et attitudes Activités expérimentales Compétences Extraire des informations Réaliser un tableau Réaliser une manipulation Utiliser un logiciel de données Mettre en relation Extraire et recenser des informations Réaliser une synthèse 1 - Domestication, sélection et hybridation L'exemple du maïs p. 2 à 5. 2 - Des caractéristiques recherchées en fonction des intérêts La composition des plantes cultivées p. 6 à 10. 3 - Des associations d'organismes Pour mémoire des graines peuvent être commercialisée associées à des bactéries ou des champignons, penser au Tp précédent. 4 - Des associations de génomes Les OGM p. 11 à 13. Bilan Expliquer l'origine des maïs actuels et des caractéristiques souhaitées par l'Homme (pour l’alimentation d’une partie de la population mondiale et des animaux d’élevage). Comparer une plante cultivée et son ancêtre naturel supposé. Recenser, extraire et exploiter des informations afin de comprendre les caractéristiques de la modification génétique d'une plante. Rédaction d’un compte-rendu sur feuille double faisant apparaître la démarche expérimentale.
-
Upload
nguyenkien -
Category
Documents
-
view
221 -
download
0
Transcript of TS-tp-p2B-10 La plante domestiquée -...
TS-tp-p2B-10 La plante domestiquée
1
Objectif : l'action de l'Homme sur le génome des plantes cultivées et sur la biodiversité - observation, B2I Observation : l'utilisation des plantes par l'Homme est une très longue histoire, qui va des pratiques empiriques les plus anciennes à la mise en œuvre des technologies les plus modernes. Problème : comment domestiquer une plante afin d'augmenter les ressources pour l'humanité ?
Matériel : blouse, livre p. 246, GéniGen, Rastop, TS-tp-p2B-12-constituant-ECE carotte, TS-tp-p2B-11-seq teosinte mais.edi, TS-tp-p2B-11-glu amylose.pdb, TS-tp-p2B-11-glu amylopectine.pdb, poly.
Capacités et attitudes Activités expérimentales Compétences
Extraire des informations Réaliser un tableau Réaliser une manipulation Utiliser un logiciel de données Mettre en relation Extraire et recenser des informations Réaliser une synthèse
1 - Domestication, sélection et hybridation L'exemple du maïs p. 2 à 5. 2 - Des caractéristiques recherchées en fonction des intérêts La composition des plantes cultivées p. 6 à 10. 3 - Des associations d'organismes Pour mémoire des graines peuvent être commercialisée associées à des bactéries ou des champignons, penser au Tp précédent. 4 - Des associations de génomes Les OGM p. 11 à 13. Bilan Expliquer l'origine des maïs actuels et des caractéristiques souhaitées par l'Homme (pour l’alimentation d’une partie de la population mondiale et des animaux d’élevage).
Comparer une plante cultivée et son ancêtre naturel supposé.
Recenser, extraire et exploiter
des informations afin de comprendre les caractéristiques
de la modification génétique d'une plante.
Rédaction d’un compte-rendu sur feuille double faisant apparaître la démarche expérimentale.

TS-tp-p2B-10 La plante domestiquée
2
1 - Domestication, sélection et hybridation Identifier l'origine du maïs dans l'espace et le temps.
Au début du XXe siècle, un botaniste, le Russe Nicolaï Vavilov, parcourt le monde à la recherche de plantes cultivables utiles. Au cours de ses voyages, il établit que la zone d'origine d'une plante est probablement celle où poussent le plus grand nombre de variétés de celle-ci. En suivant ce raisonnement, il situe en particulier l'origine du maïs en Mésoamérique (du Mexique au Costa-Rica). Après Vavilov, plusieurs botanistes américains émettent l'hypothèse que son ancêtre sauvage est la Téosinte, une plante fourragère qui pousse notamment au Mexique et au Guatemala. Des découvertes archéologiques : Dolores Piperno, de la Smithsonian Institution, et Kent Flannery, de l'université du Michigan, ont daté de 4250 avant notre ère environ trois spécimens de maïs très primitifs trouvés à Guila Naquitz, un abri sous roche de la vallée d'Oaxaca au Mexique. Le rachis rigide de ces trois spécimens prouve qu'ils appartiennent à une espèce qui dépend de l'homme pour sa survie.
La domestication de la Téosinte était donc déjà bien avancée il y a plus de 6 000 ans.
Classification Le genre Zea comprend cinq espèces - Le maïs cultivé : Zea mays et des sous espèces Zm mays, Zm parviglumis, Zm mexicana, Zm huehuetenangensis - Les téosintes : Zea luxurians, Zea nicaraguensis, Zea diploperennis, Zea perennis Génétique Toutes les espèces et sous-espèces du genre Zea ont le même nombre de chromosomes (2n=20) sauf Zea perennis (tétraploïde 4n=40). Presque toutes les espèces de téosintes sont hybridables avec le maïs. Les Tripsacum originaires d'Amérique pouvent former des hybrides le plus souvent stériles avec Zea. - Rechercher les caractéristiques du maïs p. 246 et site AFD puis compléter le poly et le tableau.
Éléments comparés Plante Épis femelles Glumes Grains par paire ou simple Nombre de rangées de grains
Maïs
Téosinte

TS-tp-p2B-10 La plante domestiquée
3
Le maïs.
Masse des grains :
10 grains de maïs = 2,7 g
10 grains de Téosinte = 0,6 g
Les grains de Téosinte sont entourés d’une cupule =
glumes soudées. Les grains de maïs possèdent des glumes réduites
TS-tp-p2B-10 La plante domestiquée
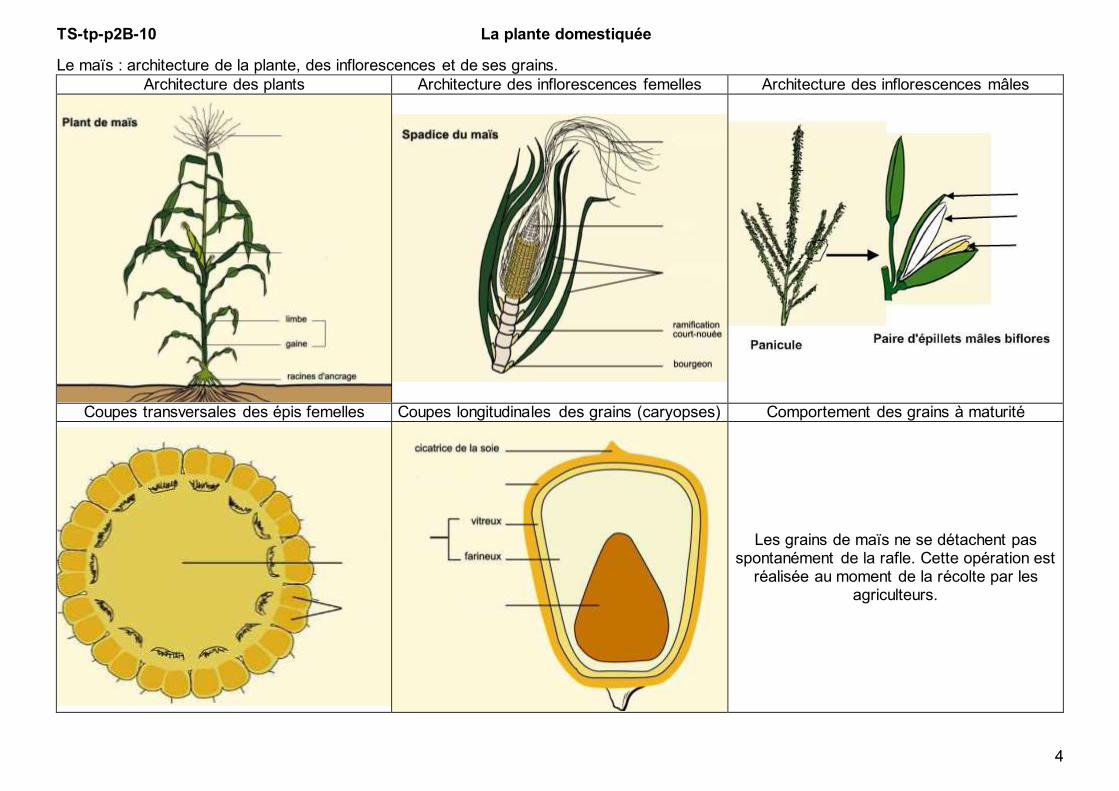
4
Le maïs : architecture de la plante, des inflorescences et de ses grains.
Architecture des plants Architecture des inflorescences femelles Architecture des inflorescences mâles
Coupes transversales des épis femelles Coupes longitudinales des grains (caryopses) Comportement des grains à maturité
Les grains de maïs ne se détachent pas spontanément de la rafle. Cette opération est
réalisée au moment de la récolte par les agriculteurs.
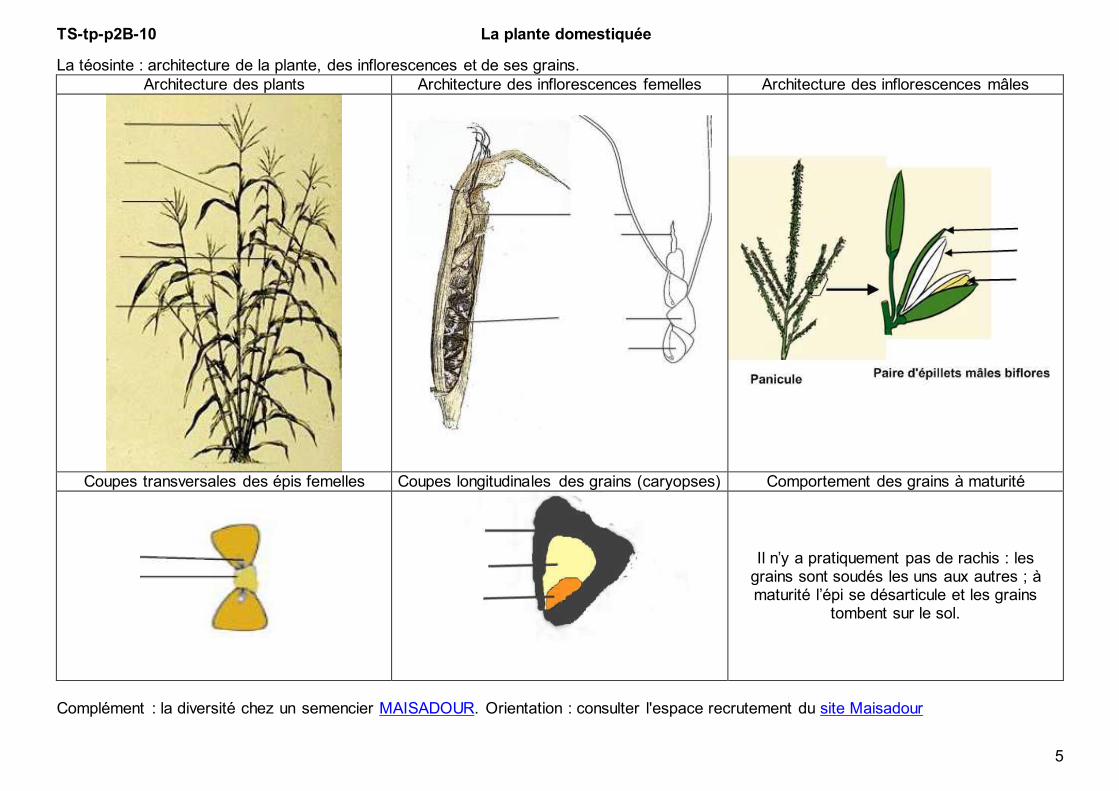
TS-tp-p2B-10 La plante domestiquée
5
La téosinte : architecture de la plante, des inflorescences et de ses grains.
Architecture des plants Architecture des inflorescences femelles Architecture des inflorescences mâles
Coupes transversales des épis femelles Coupes longitudinales des grains (caryopses) Comportement des grains à maturité
Il n’y a pratiquement pas de rachis : les grains sont soudés les uns aux autres ; à maturité l’épi se désarticule et les grains
tombent sur le sol.
Complément : la diversité chez un semencier MAISADOUR. Orientation : consulter l'espace recrutement du site Maisadour

TS-tp-p2B-10 La plante domestiquée
6
2 - Des caractéristiques recherchées en fonction des intérêts La composition des plantes cultivées. La composition de la carotte : déterminer si toutes les variétés sont à cultiver : réaliser TS-tp-p2B-12-constituant-ECE carotte. La composition du maïs Glucide : 72 à 73 % de sucre lent (amidon) + 1 à 3 % de sucre rapide (glucose, saccharose et fructose) Protéines : 8 à 11 % Lipide : 3 à 18 % (dont 13 % d'acides gras saturés) Fibre alimentaire Minéraux (mg/100 g) : P 299 ; K 324 ; Ca 48 ; Mg 108 ; Na 59 ; Fe 4,8 ; Cu 1,3 ; Mn 1,0 ; Zn 4,6 Vitamines : A et E Composition chimique des principales parties des grains de maïs (pourcentage)
Composant chimique Péricarpe Albumen Germe
Protéines 3,7 8,0 18,4 Lipides 1,0 0,8 33,2
Amidon 7,3 87,6 8,3
Sucre 0,34 0,6 10,8
Fibres 86,7 2,7 8,8
Minéraux 0,8 0,3 10,5
Les protéines du maïs : résoudre un problème scientifique. * des qualités nutritionnelles La valeur nutritive : les études sur l'animal ont montré que les protéines du maïs sont déficientes en lysine et en tryptophane, ces acides aminés sont essentiels (indispensable au fonctionnement de l'organisme). Les enfants nourris essentiellement à base de maïs présentent des troubles du développement. Quelles solutions pouvez-vous proposer pour résoudre ce problème ? Quantité des protéines Des études de l'Université d'Illinois (1974) ont montré qu'il était possible de modifier la teneur en protéines du grain de maïs par sélection, la teneur en protéines pouvait être portée de 10,9 à 26,6 % après 65 générations de sélection. La teneur en protéines de lignées autofécondées standard pouvait être accrue par croisement avec la souche HP (haute protéine) de l'Illinois puis rétrocroisement avec la lignée stable, mais avec une augmentation notable des engrais azotés. Toutefois, la qualité protéique des souches HP était inférieure à celle du maïs commun étant donné que l'accroissement des protéines était dû à une augmentation de la fraction prolamine pauvre en lysine et tryptophane.
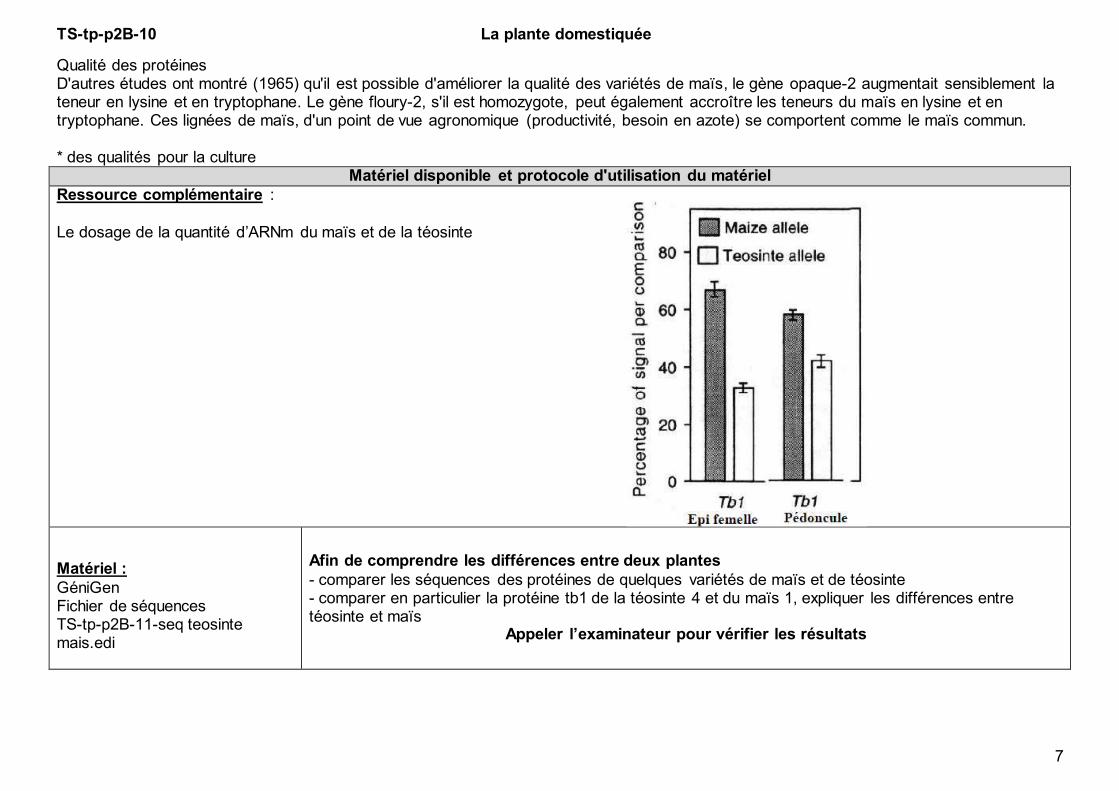
TS-tp-p2B-10 La plante domestiquée
7
Qualité des protéines D'autres études ont montré (1965) qu'il est possible d'améliorer la qualité des variétés de maïs, le gène opaque-2 augmentait sensiblement la teneur en lysine et en tryptophane. Le gène floury-2, s'il est homozygote, peut également accroître les teneurs du maïs en lysine et en tryptophane. Ces lignées de maïs, d'un point de vue agronomique (productivité, besoin en azote) se comportent comme le maïs commun. * des qualités pour la culture
Matériel disponible et protocole d'utilisation du matériel
Ressource complémentaire :
Le dosage de la quantité d’ARNm du maïs et de la téosinte
Matériel :
GéniGen Fichier de séquences TS-tp-p2B-11-seq teosinte mais.edi
Afin de comprendre les différences entre deux plantes
- comparer les séquences des protéines de quelques variétés de maïs et de téosinte - comparer en particulier la protéine tb1 de la téosinte 4 et du maïs 1, expliquer les différences entre téosinte et maïs
Appeler l’examinateur pour vérifier les résultats

TS-tp-p2B-10 La plante domestiquée
8
Complément pour faire le lien avec la première partie du cours
L’ADN sauteur du pop-corn La domestication est le passage de la forme sauvage d’une espèce à une forme cultivée. Pour de nombreuses espèces, ce processus évolutif, façonné par l’homme, trouve sa source au néolithique. La domestication entraîne la transformation héréditaire des caractères morphologiques, physiologiques ou comportementaux. Aussi, la comparaison des formes sauvages (ancestrales) et domestiquées (dérivées), et de leurs ADN respectifs, fournit-elle une situation unique pour identifier les bases génétiques de l’évolution des caractères. Le maïs que nous cultivons aujourd’hui est apparu au Mexique à partir de la téosinte, une graminée sauvage. Maïs et téosinte présentent des différences anatomiques majeures. Par exemple, la téosinte possède de nombreuses branches latérales portant de petits épis d’une douzaine de grains, alors que le maïs ne développe qu’une ou deux branches latérales dont les épis sont couverts de plus de 300 grains. En 1995 on a identifié un gène, tb1, impliqué dans la différence du nombre de branches entre maïs et téosinte. Le gène tb1 de maïs produit davantage de protéines que son homologue chez la téosinte (il est exprimé plus fortement), ce qui limite le nombre de branches latérales. Comprendre la variation
du nombre de branches revient donc à comprendre comment le gène tb1 a changé d’expression, en 2011 les chercheurs identifient un transposon inséré à proximité du gène tb1 de maïs et qui modifie son niveau d’expression. De quoi s’agit-il ? Un transposon est un petit fragment d’ADN capable de se dupliquer de façon autonome et de se déplacer physiquement dans le génome. Il en existe de nombreux types (tels que "tourist" ou "gypsy") ils se comportent comme des spams moléculaires infectant les génomes : ils constituent 50 % de notre génome et 90 % de celui du blé ! Un transposon peut altérer l’expression des gènes dans le voisinage de son site d’insertion, et ainsi modifier des caractères de l’organisme. C’est ce qui se passe pour le gène tb1 du maïs. L’étude révèle un autre point intéressant concernant l’histoire évolutive de ce transposon : cette insertion à proximité de tb1 existait déjà, mais à faible fréquence, dans la téosinte avant la domestication. Ainsi, l’apparition du maïs, ne résulte pas tant d’une nouveauté génétique que de la sélection, par l’homme, d’un variant génétique déjà présent dans les populations naturelles de téosinte. Les transposons, ces fragments d’ADN sauteurs, ne sont pas uniquement à l’origine du pop-corn de nos salles de cinéma, mais ont également joué un rôle dans la domestication de nombreuses plantes sauvages (tomate, riz). Ainsi, les transposons constituent à la fois un fardeau génétique et une source de diversification évolutive.

TS-tp-p2B-10 La plante domestiquée
9
Les glucides du maïs Matériel disponible et protocole d'utilisation du matériel
Ressource complémentaire : les glucides du maïs doc 2 p. 249
Maïs waxy Les maïs Waxy sont des maïs dont l'amidon est uniquement composé d'amylopectine. Un maïs standard contient 70 à 75 % d'amylopectine et 25 à 30 % d'amylose. C'est un caractère très recherché par l'industrie agroalimentaire : sauces, potages, plats cuisinés. Il permet d'obtenir un amidon très visqueux à chaud et ayant une texture souple au froid. D'un point de vue génétique, le caractère waxy est lié au gène waxy présent à l'état naturel dans certaines populations de maïs. C'est un caractère récessif. Pour obtenir un maïs waxy, il faut donc croiser deux parents comportant le gène waxy. Le gène a été introduit dans le génome de chaque parent par rétro croisement (les parents ne sont donc pas des organismes génétiquement modifiés). Maïs blanc Le grain de maïs blanc possède un amidon très blanc. Il permet de fabriquer la semoule entrant dans la composition de la polenta. En industrie pharmaceutique, on l'utilise pour réaliser des enrobages. En alimentation animale, on obtient des volailles à la peau blanche (le poulet de Bresse en est un exemple) ou des palmipèdes gras dont le foie est blanc. Maïs vitreux rouge (dit aussi maïs plata) Le maïs vitreux rouge possède un grain très dur, les deux tiers de la production sont destinés à la fabrication des grits de brasserie pour fabrication de la bière. Une autre partie de la production donne les flocons de céréales (éclatement à chaud). En Italie, la pigmentation des maïs vitreux rouges permet de colorer la viande pour la fabrication du jambon de Parme.
Matériel biologique :
Rastop
Afin de montrer les différences
- comparer l'amylose et l'amylopectine avec Rastop (afficher les molécules en boules et bâtonnets et les présenter cote à cote). - relier génétique, molécules et caractéristiques du maïs.
Appeler l’examinateur pour vérifier les résultats
Les lipides du maïs La teneur en huile est déterminée génétiquement. Les huiles obtenues à partir de variétés différentes présentent des compositions différentes. L'huile de maïs a une faible teneur en acides gras saturés (11 % d'acide palmitique et 2 % d'acide stéarique). En revanche, elle contient beaucoup d'acides gras polyinsaturés (acide linolénique). L'huile de maïs est relativement stable. Les populations qui consomment du maïs dégermé en tirent moins de profit en ce qui concerne l'huile et les acides gras que les populations qui consomment des produits à base de grains entiers. Quelle est la teneur du maïs en lipide où est la localisation dans la graine ? Comment fabriquer de l'huile de maïs ?
TS-tp-p2B-10 La plante domestiquée
10
4 - Des associations de génomes Principes de la transgenèse : expliquer par un schéma les étapes de la transgenèse. À partir du film (de 0 à 3 min), doc 1 p. 255, des liens OGM fabrication et Technique, Les OGM dans le monde About GM crops, what are the different positions of the countries ? Répondre à partir des liens OGM en chiffres et OGM autorisés. Des exemples d'OGM Présenter un OGM : recherche l’espèce qui a donné le gène d’intérêt, l’espèce receveuse et le caractère nouvellement acquis à partir des liens OGM exemples ou exemples suivants Les techniques du génie génétique sont actuellement utilisées dans le but de produire de nouvelles variétés de plantes cultivées. Diverses variétés de riz transgénique sont en cours d'essais : - riz résistant aux champignons parasites, qui éliminent aussi l'équivalent de la nourriture de 100 à 200 millions de personnes ; - riz enrichi en bêta-carotène précurseur de la vitamine E en carence pour certaines populations. Les tomates Le légume en première position pour sa production mondiale est la tomate. Sa maturation dépend de la production, dans le fruit, d'éthylène qui déclenche diverses réactions biochimiques, aboutissant notamment au ramollissement du fruit. Aussi, pour supporter le transport, les tomates doivent être récoltées vertes, ce qui empêche le développement des arômes qui contribuent au goût. Une tomate transgénique est apparue sur le marché en 1994 aux États-Unis. Elle ne produit plus l'enzyme responsable du ramollissement, et peut ainsi être cueillie mûre. Elle peut être récoltée à un stade de maturation plus avancé et donc offrir plus de goût au consommateur. La pomme de terre Elle occupe la quatrième place des cultures dans le monde, après le blé, le riz et le maïs. Sa production de protéines, par unité de temps et de surface, est supérieure aux autres plantes cultivées. La recherche continue pour la mise au point d'une variété transgénique résistante au mildiou, maladie due à un champignon (Phytophtora).
Deux variétés de pommes de terre transgéniques sont commercialisées, l'une résistante à la larve de doryphore, l'autre à un virus. Les agrumes Ils sont très sensibles au gel. L'objectif est donc de mettre au point des variétés d'agrumes transgéniques résistantes au gel. Les premiers essais d'expérimentation du processus ont été réalisés sur le tabac, plante modèle. On a obtenu des plants de tabac transgéniques dont les cellules sont protégées contre la formation de cristaux de glace. Pour cela le gène transféré a été isolé chez une variété de poisson (flet) qui produit une protéine permettant d'abaisser le seuil de température à partir duquel se forment des cristaux de glace.
TS-tp-p2B-10 La plante domestiquée
11
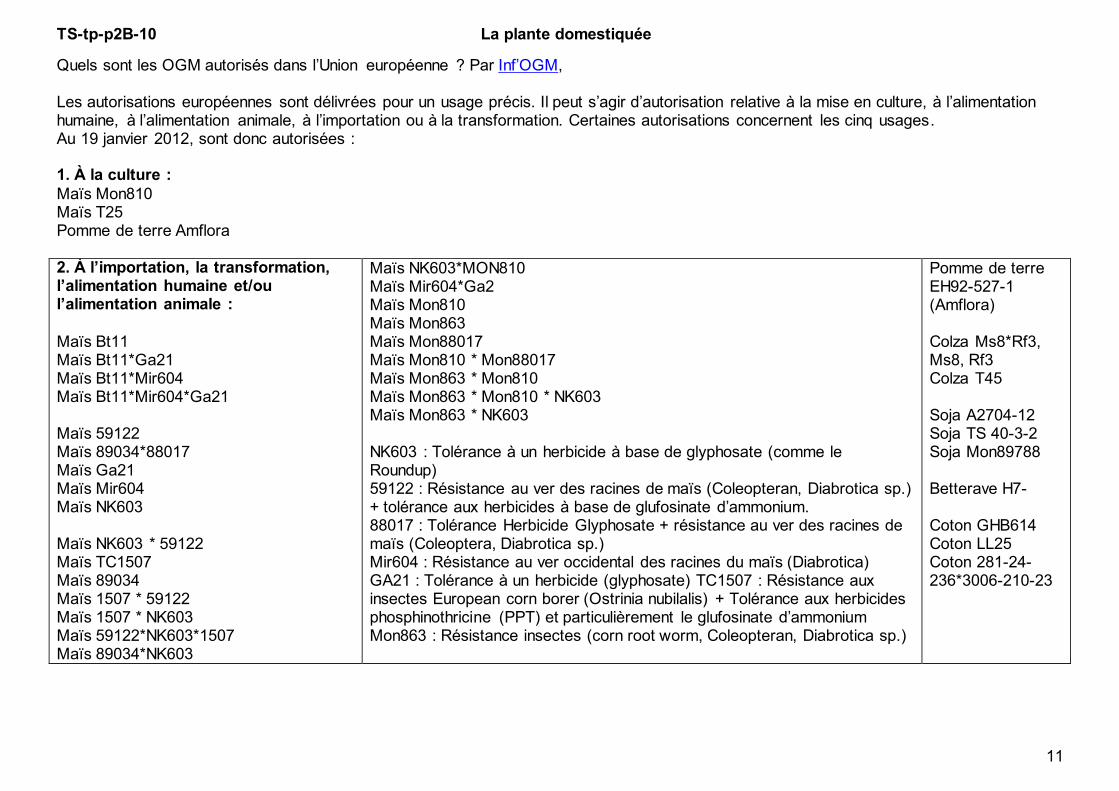
Quels sont les OGM autorisés dans l’Union européenne ? Par Inf’OGM, Les autorisations européennes sont délivrées pour un usage précis. Il peut s’agir d’autorisation relative à la mise en culture, à l’alimentation humaine, à l’alimentation animale, à l’importation ou à la transformation. Certaines autorisations concernent les cinq usages. Au 19 janvier 2012, sont donc autorisées : 1. À la culture :
Maïs Mon810 Maïs T25 Pomme de terre Amflora 2. À l’importation, la transformation, l’alimentation humaine et/ou l’alimentation animale :
Maïs Bt11 Maïs Bt11*Ga21 Maïs Bt11*Mir604 Maïs Bt11*Mir604*Ga21 Maïs 59122 Maïs 89034*88017 Maïs Ga21 Maïs Mir604 Maïs NK603 Maïs NK603 * 59122 Maïs TC1507 Maïs 89034 Maïs 1507 * 59122 Maïs 1507 * NK603 Maïs 59122*NK603*1507 Maïs 89034*NK603
Maïs NK603*MON810 Maïs Mir604*Ga2 Maïs Mon810 Maïs Mon863 Maïs Mon88017 Maïs Mon810 * Mon88017 Maïs Mon863 * Mon810 Maïs Mon863 * Mon810 * NK603 Maïs Mon863 * NK603 NK603 : Tolérance à un herbicide à base de glyphosate (comme le Roundup) 59122 : Résistance au ver des racines de maïs (Coleopteran, Diabrotica sp.) + tolérance aux herbicides à base de glufosinate d’ammonium. 88017 : Tolérance Herbicide Glyphosate + résistance au ver des racines de maïs (Coleoptera, Diabrotica sp.) Mir604 : Résistance au ver occidental des racines du maïs (Diabrotica) GA21 : Tolérance à un herbicide (glyphosate) TC1507 : Résistance aux insectes European corn borer (Ostrinia nubilalis) + Tolérance aux herbicides phosphinothricine (PPT) et particulièrement le glufosinate d’ammonium Mon863 : Résistance insectes (corn root worm, Coleopteran, Diabrotica sp.)
Pomme de terre EH92-527-1 (Amflora) Colza Ms8*Rf3, Ms8, Rf3 Colza T45 Soja A2704-12 Soja TS 40-3-2 Soja Mon89788 Betterave H7- Coton GHB614 Coton LL25 Coton 281-24-236*3006-210-23

TS-tp-p2B-10 La plante domestiquée
12
Compléments Le maïs transgénique résistant à la pyrale favorise-t-il l'apparition de résistances chez les insectes ? (Guy Riba, Josette Chaufaux, INRA : http://www.inra.fr/internet/Directions/DIC/ACTUALITES/DOSSIERS/OGM/riba.htm) Des variétés de maïs ont été transformées par des firmes privées pour produire dans leurs tissus la toxine Cry1Ab de Bacillus thuringiensis (Bt), active contre la pyrale du maïs. 1) Cette stratégie de lutte offre plusieurs avantages : - la toxine Cry1Ab n'est active que sur les insectes, aucune toxicité n'a été mise en évidence, ni pour les animaux domestiques ni pour l'homme. - la toxine est produite principalement dans les parties vertes de la plante, qui ne sont jamais consommées par l'homme ; - les premiers essais ont montré une remarquable efficacité de ces maïs (voir tableau ci-dessous) ;
Date d’infestation par la Pyrale
Maïs Bt Pourcentage de mortalité larvaire
Insecticide chimique : Chlorpyriphos-éthyl
24 juin 8 juillet 22 juillet 6 août 20 août
100 % 100 %
100 % 93 %
72 % 98 % 73 %
Comparaison de l’efficacité de différents traitements contre la pyrale de maïs : d’après Labare et cl 1992
- les larves vivent à l'intérieur de la plante, dès leur éclosion, elles sont éliminées avant d'avoir pu provoquer des dégâts, - la toxine insecticide produite dans la plante est protégée des conditions climatiques, pluie ou rayonnements ultraviolets. En 1995 la culture du maïs transgénique a été autorisée aux États-Unis, en 1996 au Canada et en novembre 1997 en France. En 1997 elle a représenté 10 % des surfaces cultivées en maïs aux États-Unis. 2) Certaines questions se sont posées avant la mise sur le marché : - quelles sont les conditions de limites d'efficacité de ces variétés ? - existe-t-il des risques de contournement par la sélection d'insectes résistants ? - y a-t-il des effets non intentionnels ? * Les limites d'efficacité La toxine insecticide produite par la plante bloque l’appareil digestif des larves de pyrale ou de sésamie (présente dans le Sud de la France). Dans les zones méridionales, une deuxième génération de chenille de pyrale et de sésamie apparaît la même année. Il faut donc estimer l’efficacité de l’insecticide produit par la plante lors des infestations tardives (voir tableau).

TS-tp-p2B-10 La plante domestiquée
13
* Les risques de contournement L'apparition de pyrales résistantes à la toxine de Bt, dans ce cas les chenilles ne seraient plus détruites, les agriculteurs devraient revenir à l’utilisation d’insecticides chimiques. Pas de risque majeur pour les autres cultures, l’infestation des haricots ou des framboisiers par la pyrale est moins massive, elle est contrôlable par plusieurs molécules chimiques. Les risques de sélection de populations de pyrale résistantes à Bt sont en principe plus élevés que dans le cas des insecticides chimiques, car tous les insectes qui ont consommé du maïs transgénique ont été en contact avec la toxine. La probabilité et la vitesse de sélection d'insectes résistants dépendent : - du nombre de générations - de la concentration de la toxine dans la plante - de la fréquence initiale et de la "force" des éventuels gènes de résistance dans les différentes populations de pyrale - de la fréquence des accouplements des survivants d'un champ transgénique avec les papillons issus des champs voisins non transgéniques - du coût biologique d'acquisition de la résistance (femelles moins fécondes, développement plus lent...). * Les effets non intentionnels On ne connaît pas d'effets non intentionnels néfastes prévisibles sur les prédateurs naturels de la pyrale. Au contraire, on peut penser que l'utilisation de maïs Bt permettra le développement d'auxiliaires (= prédateurs) jusque-là tués par les traitements chimiques. 3) Recherche de résistance chez les insectes L'objectif général des travaux effectués à l'INRA : - est d'identifier s'il existe dans les populations naturelles de pyrale des gènes susceptibles d'induire une résistance à la toxine présente dans le maïs transgénique. Il s'agit d'établir la courbe de toxicité de la toxine de Bacillus thuringiensis pour les différentes populations sauvages de pyrale, la perte de la toxicité est indicateur de l'évolution de la résistance à Bt. L'établissement d'une telle courbe dans une région donnée est indispensable avant l'installation de plantes transgéniques. - de caractériser les éventuels gènes de résistances afin de mettre au point des méthodes de détection simples avant qu'ils atteignent des fréquences permettant leur détection par des tests toxicologiques classiques. Une sélection, commencée en 1993 et poursuivie en conditions expérimentales pendant 26 générations, n'a pas permis l'obtention d'une lignée de pyrale résistante à la toxine de Bt. À chaque génération, mille chenilles ont été traitées dans le but de trouver une résistance. - d'étudier leur hérédité et leur maintien dans les populations dans différentes conditions expérimentales et ainsi d'évaluer les risques de dispersion géographique de gènes de résistance, s'ils apparaissent. Ces différents travaux donneront des éléments de biovigilance. Ils contribueront au suivi des conséquences sur l'environnement de la culture du maïs Bt. Les "marqueurs" génétiques révélés par ces études pourront de plus servir aux investigations visant à identifier les relations entre les gènes de résistance et ainsi aider à la caractérisation de marqueurs génétiques de la résistance.
















![C 200 TS-14, C 210 TS-12, C 220 TS-10, C 230 TS-9 - RUBAG...C 200 TS-14, C 210 TS-12, C 220 TS-10, C 230 TS-9 DLT 27025179 [mm] 1960 3730 2568 5403 1080 430 DLT 2702-1FR 08/16 MT.](https://static.fdocuments.fr/doc/165x107/60ae0e3c7f8c9879a34f6d52/c-200-ts-14-c-210-ts-12-c-220-ts-10-c-230-ts-9-rubag-c-200-ts-14-c-210.jpg)

