TRANSPORTS INTRACELLULAIRES DES...
16
TRANSPORTS INTRACELLULAIRES DES PROTEINES d'après le cours de Marie FLAMAND, Unité des Arbovirus & Virus des Fièvres Hémorragiques INTRODUCTION Les cellules eucaryotes ont évoluées de manière à pouvoir assurer leur équilibre homéostatique, dialoguer avec l'environnement tissulaire, et à plus grande échelle avec l'organisme tout. entier, et ainsi veiller à leur propre survie au sein de l'organisme hôte. Afin de gérer à la fois la diversité et la spécificité des échanges requis, des systèmes biochimiques complexes de synthèse et de signalisation se sont développés faisant appel en particulier à des structures subcellulaires spécialisées appelées organelles. L'existence de compartiments cellulaires, isolés des constituants cytosoliques par des membranes lipidiques, est indispensable au confinement des molécules (ions, molécules organiques, macromolécules) impliquées dans les multiples activités physiologiques de la cellule. Chaque organelle assure des fonctions distinctes mais dans un mode dynamique et la difficulté pour la cellule consiste à disposer de moyens de communication entre elles tout en maintenant leur intégrité. Dans le cadre de ce cours, nous nous intéresserons au transport des protéines dans les voies de biosynthèse/sécrétion et d'endocytose de la cellule eucaryote, aux différents modes de transport utilisés entre les différentes organelles cellulaires ainsi qu'aux mécanismes moléculaires impliqués dans la régulation et la spécification de l'adressage des protéines qui transitent ou résident dans ces régions. La cellule de mammifère sera traitée en exemple et l'utilisation de la machinerie cellulaire par les agents viraux sera abordée dans ce contexte. I- ORGANISATION CELLULAIRE
Transcript of TRANSPORTS INTRACELLULAIRES DES...

TRANSPORTS INTRACELLULAIRES DES PROTEINES
dapregraves le cours de Marie FLAMAND Uniteacute des Arbovirus amp Virus des Fiegravevres Heacutemorragiques
INTRODUCTION
Les cellules eucaryotes ont eacutevolueacutees de maniegravere agrave pouvoir assurer leur eacutequilibre homeacuteostatique dialoguer avec lenvironnement tissulaire et agrave plus grande eacutechelle avec lorganisme tout entier et ainsi veiller agrave leur propre survie au sein de lorganisme hocircte Afin de geacuterer agrave la fois la diversiteacute et la speacutecificiteacute des eacutechanges requis des systegravemes biochimiques complexes de synthegravese et de signalisation se sont deacuteveloppeacutes faisant appel en particulier agrave des structures subcellulaires speacutecialiseacutees appeleacutees organelles Lexistence de compartiments cellulaires isoleacutes des constituants cytosoliques par des membranes lipidiques est indispensable au confinement des moleacutecules (ions moleacutecules organiques macromoleacutecules) impliqueacutees dans les multiples activiteacutes physiologiques de la cellule Chaque organelle assure des fonctions distinctes mais dans un mode dynamique et la difficulteacute pour la cellule consiste agrave disposer de moyens de communication entre elles tout en maintenant leur inteacutegriteacute Dans le cadre de ce cours nous nous inteacuteresserons au transport des proteacuteines dans les voies de biosynthegraveseseacutecreacutetion et dendocytose de la cellule eucaryote aux diffeacuterents modes de transport utiliseacutes entre les diffeacuterentes organelles cellulaires ainsi quaux meacutecanismes moleacuteculaires impliqueacutes dans la reacutegulation et la speacutecification de ladressage des proteacuteines qui transitent ou reacutesident dans ces reacutegions La cellule de mammifegravere sera traiteacutee en exemple et lutilisation de la machinerie cellulaire par les agents viraux sera abordeacutee dans ce contexte
I- ORGANISATION CELLULAIRE
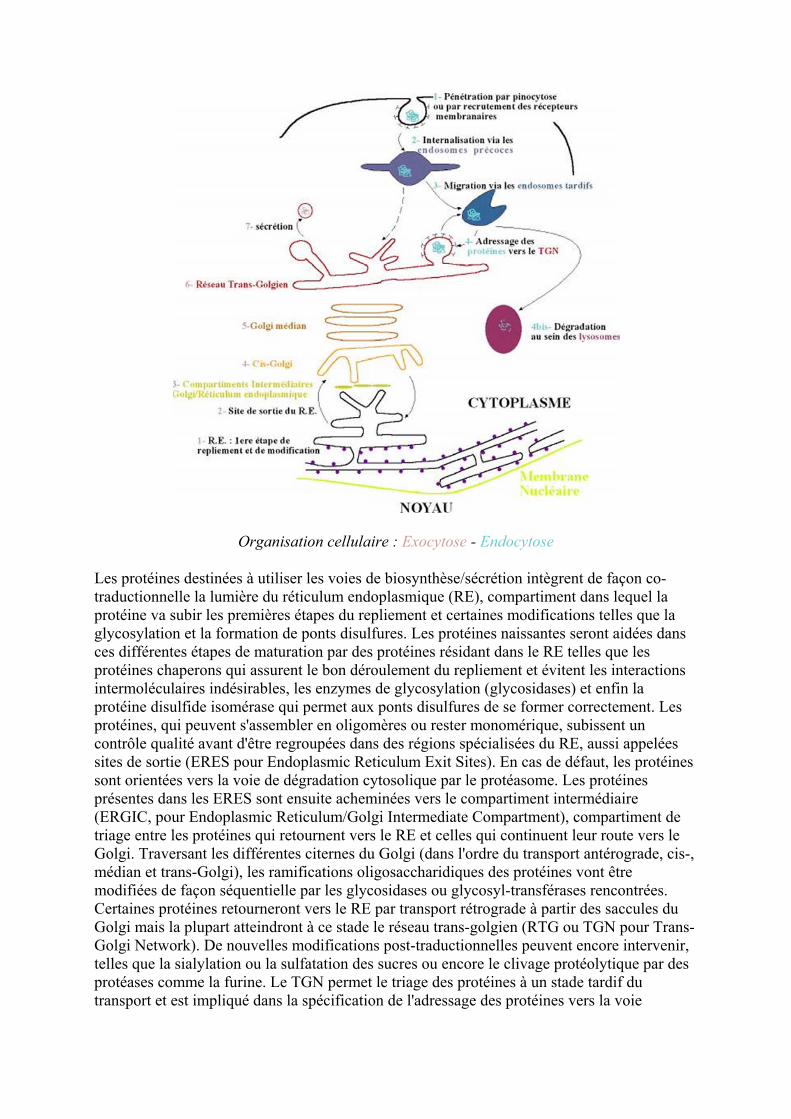
Organisation cellulaire Exocytose - Endocytose
Les proteacuteines destineacutees agrave utiliser les voies de biosynthegraveseseacutecreacutetion integravegrent de faccedilon co-traductionnelle la lumiegravere du reacuteticulum endoplasmique (RE) compartiment dans lequel la proteacuteine va subir les premiegraveres eacutetapes du repliement et certaines modifications telles que la glycosylation et la formation de ponts disulfures Les proteacuteines naissantes seront aideacutees dans ces diffeacuterentes eacutetapes de maturation par des proteacuteines reacutesidant dans le RE telles que les proteacuteines chaperons qui assurent le bon deacuteroulement du repliement et eacutevitent les interactions intermoleacuteculaires indeacutesirables les enzymes de glycosylation (glycosidases) et enfin la proteacuteine disulfide isomeacuterase qui permet aux ponts disulfures de se former correctement Les proteacuteines qui peuvent sassembler en oligomegraveres ou rester monomeacuterique subissent un controcircle qualiteacute avant decirctre regroupeacutees dans des reacutegions speacutecialiseacutees du RE aussi appeleacutees sites de sortie (ERES pour Endoplasmic Reticulum Exit Sites) En cas de deacutefaut les proteacuteines sont orienteacutees vers la voie de deacutegradation cytosolique par le proteacuteasome Les proteacuteines preacutesentes dans les ERES sont ensuite achemineacutees vers le compartiment intermeacutediaire (ERGIC pour Endoplasmic ReticulumGolgi Intermediate Compartment) compartiment de triage entre les proteacuteines qui retournent vers le RE et celles qui continuent leur route vers le Golgi Traversant les diffeacuterentes citernes du Golgi (dans lordre du transport anteacuterograde cis- meacutedian et trans-Golgi) les ramifications oligosaccharidiques des proteacuteines vont ecirctre modifieacutees de faccedilon seacutequentielle par les glycosidases ou glycosyl-transfeacuterases rencontreacutees Certaines proteacuteines retourneront vers le RE par transport reacutetrograde agrave partir des saccules du Golgi mais la plupart atteindront agrave ce stade le reacuteseau trans-golgien (RTG ou TGN pour Trans-Golgi Network) De nouvelles modifications post-traductionnelles peuvent encore intervenir telles que la sialylation ou la sulfatation des sucres ou encore le clivage proteacuteolytique par des proteacuteases comme la furine Le TGN permet le triage des proteacuteines agrave un stade tardif du transport et est impliqueacute dans la speacutecification de ladressage des proteacuteines vers la voie
lysosomale ou vers des domaines speacutecialiseacutes de la membrane plasmique En fonction des voies emprunteacutees agrave partir du TGN les proteacuteines seront seacutecreacuteteacutees de faccedilon constitutive ou reacuteguleacutee et pourront ecirctre envoyeacutees vers les reacutegions apicales ou basales distinctement dans le cas des cellules polariseacutees Ce compartiment est aussi particuliegraverement important car il fait le lien entre les voies dexocytose et dendocytose Les proteacuteines provenant du milieu extracellulaire peacutenegravetre dans la cellule cible soit par un meacutecanisme non speacutecifique de pinocytose soit par recrutement par des reacutecepteurs membranaires Linternalisation des moleacutecules se fait par lintermeacutediaire dendosomes preacutecoces qui font le tri entre les proteacuteines agrave recycler vers la membrane plasmique (cest en geacuteneacuteral le cas pour les reacutecepteurs libeacutereacutes de leur ligand) et celles qui migreront via les endosomes tardifs vers les lysosomes A partir des structures endosomales il existe eacutegalement une voie dadressage des proteacuteines vers le TGN Les lysosomes constituent le principal site de deacutegradation des moleacutecules (proteacuteines acides nucleacuteiques sucres et phospholipides) mais aussi de structures plus importantes (fragments dorganelles corps eacutetrangers) Lhydrolyse des substrats se fait agrave pH acide sous laction denzymes speacutecialiseacutees et les produits de deacutegradation sont alors reconduits vers le cytosol pour ecirctre reacuteutiliseacutes ou relargueacutes dans le milieu extracellulaire
II- DIFFERENTS MODES DE TRANSPORT DES PROTEINES
Les proteacuteines qui cheminent agrave linteacuterieur de la cellule en provenance ou non du milieu exteacuterieur sont confronteacutees au problegraveme du passage des barriegraveres lipidiques assurant linteacutegriteacute de la cellule et de ses diffeacuterents compartiments Au cours de leacutevolution plusieurs meacutecanismes se sont mis en place pour permettre aux proteacuteines de traverser les membranes sans modifier le contenu de lenceinte et risquer de pertuber les fonctions essentielles de la cellule Ces meacutecanimes sont communs pour certains aux bacteacuteries levures et cellules de mammifegraveres et font intervenir de nombreuses proteacuteines ayant gardeacute un certain degreacute dhomologie entre les diffeacuterents organismes
A - Translocation Ce type de transport correspond agrave un passage direct des proteacuteines au travers de la membrane via des complexes proteacuteiques formant des pores speacutecifiques de chaque organelle Il a lieu de faccedilon cotraductionnelle dans les membranes du RE des cellules de mammifegravere ou post-traductionnellement pour limport dans les mitochondries les peacuteroxisomes ou le noyau apregraves reconnaissance dune seacutequence dadressage speacutecifique vers la membrane cible Dans la plupart des cas la seacutequence signal est cliveacute une fois le chargement termineacute Pour le transfert des proteacuteines deacutejagrave replieacutees des proteacuteines chaperon coopegraverent avec le translocon pour leur deacutepliement transitoire
Translocation des glycoproteacuteines solubles
Translocation des glycoproteacuteines membranaires de type 1
Translocation des glycoproteacuteines membranaires de type 2
La translocation de proteacuteines au travers des membranes du RE a eacuteteacute caracteacuteriseacutee et implique la reconnaissance dune seacutequence signal hydrophobe en la partie N-terminale de la proteacuteine en cours de synthegravese par le complexe SRP (pour Signal Recognition Particle) son attachement agrave la membrane via un reacutecepteur pour le SRP ainsi que la liaison entre le ribosome et le complexe proteacuteique Sec61p qui assure le passage du brin proteacuteique au travers de la membrane
Rocircle de la proteacuteine SRP dans la translocation
Il existe eacutegalement des exemples de reacutetro translocation du RE vers le cytosol lorsquune proteacuteine mal replieacutee est orienteacutee par les proteacuteines chaperon telles que Bip ou calnexine vers la voie de deacutegradation par le proteacuteasome La chaine polypeptidique est deacuteplieacutee et le translocon
Sec61p alors programmeacute pour le retrotransport de la proteacuteine Une fois dans le cytosol la proteacuteine est deacuteglycosyleacutee polyubiquitinyleacutee et deacutegradeacutee par le proteacuteasome
B - Diffusion Ce mode de deacuteplacement sappuie sur la fluiditeacute des membranes qui permet la diffusion lateacuterale de ses constituants
Membrane cytoplasmique
Il pourrait ecirctre impliqueacute dans la constitution de sous-compartiments se distinguant du reste de lorganelle aux plans biochimiques et fonctionnels comme par exemple le rassemblement aux sites ERES de proteacuteines destineacutees agrave lexport ou lexistence de microdomaines agrave la membrane plasmique De mecircme il a eacuteteacute eacutevoqueacute que le mouvement anteacuterograde le long des citernes du Golgi pourrait avoir lieu selon ce principe et se poursuivrait tant que les moleacutecules ne seraient pas retenues ou renvoyeacutees dans les voies reacutetrogrades du transport Pour les proteacuteines transmembranaires reacutesidant preacutefeacuterentiellement au niveau de certaines reacutegions du Golgi (telles que les enzymes de la biosynthegravese de sucres) larrecirct de la progression pourrait deacutependre de la capaciteacute des proteacuteines agrave interagir avec la nature environnante des lipides avec un eacuteleacutement deacuteterminant qui pourrait ecirctre la longueur de la reacutegion transmembranaire de la proteacuteine ou encore de la capaciteacute de la proteacuteine sous certaines conditions de milieu agrave former des structures oligomeacuteriques de grandes tailles agrave mobiliteacute reacuteduite De faccedilon plus geacuteneacuterale la mobiliteacute des constituants membranaires paraicirct un eacuteleacutement important de leacutequilibre dynamique dont la cellule fait preuve
C - Transport veacutesiculo-tubulaire Ce mode de transport est largement utiliseacute par les proteacuteines empruntant les voies dendo- et dexocytose Il a fait lobjet ces derniegraveres anneacutees de nombreux travaux aussi bien par des approches geacuteneacutetiques chez la levure que par des reconstitutions de systegravemes membranaires in vitro De grandes avanceacutees ont eacuteteacute reacutealiseacutees en particulier dans la compreacutehension de meacutecanismes moleacuteculaires assurant la reacutegulation du traffic veacutesiculaire et sa speacutecificiteacute agrave la fois lors de la formation des veacutesicules et de leur chargement que lors de leur adressage vers lorganelle cible une fois formeacutees Lincroyable activiteacute cellulaire se fait de maniegravere coordonneacutee gracircce agrave un ensemble de proteacuteines membranaires ou cytosoliques precirctes agrave ecirctre activeacutees recruteacutees et recycleacutees en permanence et dont le jeu des interactions deacutefinit la nature des eacutechanges Suivant le trajet et les moleacutecules agrave transporter les partenaires veacutesiculaires diffegraverent
Traffic veacutesiculo-tubulaire intracellulaire
Le transport de moleacutecules entre deux compartiments peut toutefois se deacutecomposer en une succession deacutevegravenements eacuteleacutementaires Le processus de formation des veacutesicules neacutecessite le recrutement de complexes cytosoliques ou proteacuteines manteau Ce recrutement se fait via des GTPases telles que les proteacuteines Arf dynamine et peut-ecirctre Rab dans certains cas qui sassocieraient agrave la membrane apregraves eacutechange de leur GDP par du GTP eacutechange catalyseacute par la proteacuteine membranaire GEF (guanine nucleotide exchange factor) Lassemblage contigu de plusieurs complexes manteau agrave la membrane du compartiment donneur conduit localement agrave sa deacuteformation puis agrave un bourgeonnement du cocircteacute du manteau La veacutesicule dont la taille est geacuteneacuteralement comprise entre 50 et 100 nm se deacutetache du compartiment donneur emportant un chargement bien deacutefini Le contenu est en effet trieacute par des reacutecepteurs speacutecifiques pour les proteacuteines solubles (ou proteacuteines cargo) tels que les lectines ERGIC53 ou VIP36 ou par des signaux contenus dans la partie cytosolique des proteacuteines transmembranaires
Activation des membranes du compartiment donneur et recrutement des proteacuteines manteaux
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209
Organisation cellulaire Exocytose - Endocytose
Les proteacuteines destineacutees agrave utiliser les voies de biosynthegraveseseacutecreacutetion integravegrent de faccedilon co-traductionnelle la lumiegravere du reacuteticulum endoplasmique (RE) compartiment dans lequel la proteacuteine va subir les premiegraveres eacutetapes du repliement et certaines modifications telles que la glycosylation et la formation de ponts disulfures Les proteacuteines naissantes seront aideacutees dans ces diffeacuterentes eacutetapes de maturation par des proteacuteines reacutesidant dans le RE telles que les proteacuteines chaperons qui assurent le bon deacuteroulement du repliement et eacutevitent les interactions intermoleacuteculaires indeacutesirables les enzymes de glycosylation (glycosidases) et enfin la proteacuteine disulfide isomeacuterase qui permet aux ponts disulfures de se former correctement Les proteacuteines qui peuvent sassembler en oligomegraveres ou rester monomeacuterique subissent un controcircle qualiteacute avant decirctre regroupeacutees dans des reacutegions speacutecialiseacutees du RE aussi appeleacutees sites de sortie (ERES pour Endoplasmic Reticulum Exit Sites) En cas de deacutefaut les proteacuteines sont orienteacutees vers la voie de deacutegradation cytosolique par le proteacuteasome Les proteacuteines preacutesentes dans les ERES sont ensuite achemineacutees vers le compartiment intermeacutediaire (ERGIC pour Endoplasmic ReticulumGolgi Intermediate Compartment) compartiment de triage entre les proteacuteines qui retournent vers le RE et celles qui continuent leur route vers le Golgi Traversant les diffeacuterentes citernes du Golgi (dans lordre du transport anteacuterograde cis- meacutedian et trans-Golgi) les ramifications oligosaccharidiques des proteacuteines vont ecirctre modifieacutees de faccedilon seacutequentielle par les glycosidases ou glycosyl-transfeacuterases rencontreacutees Certaines proteacuteines retourneront vers le RE par transport reacutetrograde agrave partir des saccules du Golgi mais la plupart atteindront agrave ce stade le reacuteseau trans-golgien (RTG ou TGN pour Trans-Golgi Network) De nouvelles modifications post-traductionnelles peuvent encore intervenir telles que la sialylation ou la sulfatation des sucres ou encore le clivage proteacuteolytique par des proteacuteases comme la furine Le TGN permet le triage des proteacuteines agrave un stade tardif du transport et est impliqueacute dans la speacutecification de ladressage des proteacuteines vers la voie
lysosomale ou vers des domaines speacutecialiseacutes de la membrane plasmique En fonction des voies emprunteacutees agrave partir du TGN les proteacuteines seront seacutecreacuteteacutees de faccedilon constitutive ou reacuteguleacutee et pourront ecirctre envoyeacutees vers les reacutegions apicales ou basales distinctement dans le cas des cellules polariseacutees Ce compartiment est aussi particuliegraverement important car il fait le lien entre les voies dexocytose et dendocytose Les proteacuteines provenant du milieu extracellulaire peacutenegravetre dans la cellule cible soit par un meacutecanisme non speacutecifique de pinocytose soit par recrutement par des reacutecepteurs membranaires Linternalisation des moleacutecules se fait par lintermeacutediaire dendosomes preacutecoces qui font le tri entre les proteacuteines agrave recycler vers la membrane plasmique (cest en geacuteneacuteral le cas pour les reacutecepteurs libeacutereacutes de leur ligand) et celles qui migreront via les endosomes tardifs vers les lysosomes A partir des structures endosomales il existe eacutegalement une voie dadressage des proteacuteines vers le TGN Les lysosomes constituent le principal site de deacutegradation des moleacutecules (proteacuteines acides nucleacuteiques sucres et phospholipides) mais aussi de structures plus importantes (fragments dorganelles corps eacutetrangers) Lhydrolyse des substrats se fait agrave pH acide sous laction denzymes speacutecialiseacutees et les produits de deacutegradation sont alors reconduits vers le cytosol pour ecirctre reacuteutiliseacutes ou relargueacutes dans le milieu extracellulaire
II- DIFFERENTS MODES DE TRANSPORT DES PROTEINES
Les proteacuteines qui cheminent agrave linteacuterieur de la cellule en provenance ou non du milieu exteacuterieur sont confronteacutees au problegraveme du passage des barriegraveres lipidiques assurant linteacutegriteacute de la cellule et de ses diffeacuterents compartiments Au cours de leacutevolution plusieurs meacutecanismes se sont mis en place pour permettre aux proteacuteines de traverser les membranes sans modifier le contenu de lenceinte et risquer de pertuber les fonctions essentielles de la cellule Ces meacutecanimes sont communs pour certains aux bacteacuteries levures et cellules de mammifegraveres et font intervenir de nombreuses proteacuteines ayant gardeacute un certain degreacute dhomologie entre les diffeacuterents organismes
A - Translocation Ce type de transport correspond agrave un passage direct des proteacuteines au travers de la membrane via des complexes proteacuteiques formant des pores speacutecifiques de chaque organelle Il a lieu de faccedilon cotraductionnelle dans les membranes du RE des cellules de mammifegravere ou post-traductionnellement pour limport dans les mitochondries les peacuteroxisomes ou le noyau apregraves reconnaissance dune seacutequence dadressage speacutecifique vers la membrane cible Dans la plupart des cas la seacutequence signal est cliveacute une fois le chargement termineacute Pour le transfert des proteacuteines deacutejagrave replieacutees des proteacuteines chaperon coopegraverent avec le translocon pour leur deacutepliement transitoire
Translocation des glycoproteacuteines solubles
Translocation des glycoproteacuteines membranaires de type 1
Translocation des glycoproteacuteines membranaires de type 2
La translocation de proteacuteines au travers des membranes du RE a eacuteteacute caracteacuteriseacutee et implique la reconnaissance dune seacutequence signal hydrophobe en la partie N-terminale de la proteacuteine en cours de synthegravese par le complexe SRP (pour Signal Recognition Particle) son attachement agrave la membrane via un reacutecepteur pour le SRP ainsi que la liaison entre le ribosome et le complexe proteacuteique Sec61p qui assure le passage du brin proteacuteique au travers de la membrane
Rocircle de la proteacuteine SRP dans la translocation
Il existe eacutegalement des exemples de reacutetro translocation du RE vers le cytosol lorsquune proteacuteine mal replieacutee est orienteacutee par les proteacuteines chaperon telles que Bip ou calnexine vers la voie de deacutegradation par le proteacuteasome La chaine polypeptidique est deacuteplieacutee et le translocon
Sec61p alors programmeacute pour le retrotransport de la proteacuteine Une fois dans le cytosol la proteacuteine est deacuteglycosyleacutee polyubiquitinyleacutee et deacutegradeacutee par le proteacuteasome
B - Diffusion Ce mode de deacuteplacement sappuie sur la fluiditeacute des membranes qui permet la diffusion lateacuterale de ses constituants
Membrane cytoplasmique
Il pourrait ecirctre impliqueacute dans la constitution de sous-compartiments se distinguant du reste de lorganelle aux plans biochimiques et fonctionnels comme par exemple le rassemblement aux sites ERES de proteacuteines destineacutees agrave lexport ou lexistence de microdomaines agrave la membrane plasmique De mecircme il a eacuteteacute eacutevoqueacute que le mouvement anteacuterograde le long des citernes du Golgi pourrait avoir lieu selon ce principe et se poursuivrait tant que les moleacutecules ne seraient pas retenues ou renvoyeacutees dans les voies reacutetrogrades du transport Pour les proteacuteines transmembranaires reacutesidant preacutefeacuterentiellement au niveau de certaines reacutegions du Golgi (telles que les enzymes de la biosynthegravese de sucres) larrecirct de la progression pourrait deacutependre de la capaciteacute des proteacuteines agrave interagir avec la nature environnante des lipides avec un eacuteleacutement deacuteterminant qui pourrait ecirctre la longueur de la reacutegion transmembranaire de la proteacuteine ou encore de la capaciteacute de la proteacuteine sous certaines conditions de milieu agrave former des structures oligomeacuteriques de grandes tailles agrave mobiliteacute reacuteduite De faccedilon plus geacuteneacuterale la mobiliteacute des constituants membranaires paraicirct un eacuteleacutement important de leacutequilibre dynamique dont la cellule fait preuve
C - Transport veacutesiculo-tubulaire Ce mode de transport est largement utiliseacute par les proteacuteines empruntant les voies dendo- et dexocytose Il a fait lobjet ces derniegraveres anneacutees de nombreux travaux aussi bien par des approches geacuteneacutetiques chez la levure que par des reconstitutions de systegravemes membranaires in vitro De grandes avanceacutees ont eacuteteacute reacutealiseacutees en particulier dans la compreacutehension de meacutecanismes moleacuteculaires assurant la reacutegulation du traffic veacutesiculaire et sa speacutecificiteacute agrave la fois lors de la formation des veacutesicules et de leur chargement que lors de leur adressage vers lorganelle cible une fois formeacutees Lincroyable activiteacute cellulaire se fait de maniegravere coordonneacutee gracircce agrave un ensemble de proteacuteines membranaires ou cytosoliques precirctes agrave ecirctre activeacutees recruteacutees et recycleacutees en permanence et dont le jeu des interactions deacutefinit la nature des eacutechanges Suivant le trajet et les moleacutecules agrave transporter les partenaires veacutesiculaires diffegraverent
Traffic veacutesiculo-tubulaire intracellulaire
Le transport de moleacutecules entre deux compartiments peut toutefois se deacutecomposer en une succession deacutevegravenements eacuteleacutementaires Le processus de formation des veacutesicules neacutecessite le recrutement de complexes cytosoliques ou proteacuteines manteau Ce recrutement se fait via des GTPases telles que les proteacuteines Arf dynamine et peut-ecirctre Rab dans certains cas qui sassocieraient agrave la membrane apregraves eacutechange de leur GDP par du GTP eacutechange catalyseacute par la proteacuteine membranaire GEF (guanine nucleotide exchange factor) Lassemblage contigu de plusieurs complexes manteau agrave la membrane du compartiment donneur conduit localement agrave sa deacuteformation puis agrave un bourgeonnement du cocircteacute du manteau La veacutesicule dont la taille est geacuteneacuteralement comprise entre 50 et 100 nm se deacutetache du compartiment donneur emportant un chargement bien deacutefini Le contenu est en effet trieacute par des reacutecepteurs speacutecifiques pour les proteacuteines solubles (ou proteacuteines cargo) tels que les lectines ERGIC53 ou VIP36 ou par des signaux contenus dans la partie cytosolique des proteacuteines transmembranaires
Activation des membranes du compartiment donneur et recrutement des proteacuteines manteaux
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

lysosomale ou vers des domaines speacutecialiseacutes de la membrane plasmique En fonction des voies emprunteacutees agrave partir du TGN les proteacuteines seront seacutecreacuteteacutees de faccedilon constitutive ou reacuteguleacutee et pourront ecirctre envoyeacutees vers les reacutegions apicales ou basales distinctement dans le cas des cellules polariseacutees Ce compartiment est aussi particuliegraverement important car il fait le lien entre les voies dexocytose et dendocytose Les proteacuteines provenant du milieu extracellulaire peacutenegravetre dans la cellule cible soit par un meacutecanisme non speacutecifique de pinocytose soit par recrutement par des reacutecepteurs membranaires Linternalisation des moleacutecules se fait par lintermeacutediaire dendosomes preacutecoces qui font le tri entre les proteacuteines agrave recycler vers la membrane plasmique (cest en geacuteneacuteral le cas pour les reacutecepteurs libeacutereacutes de leur ligand) et celles qui migreront via les endosomes tardifs vers les lysosomes A partir des structures endosomales il existe eacutegalement une voie dadressage des proteacuteines vers le TGN Les lysosomes constituent le principal site de deacutegradation des moleacutecules (proteacuteines acides nucleacuteiques sucres et phospholipides) mais aussi de structures plus importantes (fragments dorganelles corps eacutetrangers) Lhydrolyse des substrats se fait agrave pH acide sous laction denzymes speacutecialiseacutees et les produits de deacutegradation sont alors reconduits vers le cytosol pour ecirctre reacuteutiliseacutes ou relargueacutes dans le milieu extracellulaire
II- DIFFERENTS MODES DE TRANSPORT DES PROTEINES
Les proteacuteines qui cheminent agrave linteacuterieur de la cellule en provenance ou non du milieu exteacuterieur sont confronteacutees au problegraveme du passage des barriegraveres lipidiques assurant linteacutegriteacute de la cellule et de ses diffeacuterents compartiments Au cours de leacutevolution plusieurs meacutecanismes se sont mis en place pour permettre aux proteacuteines de traverser les membranes sans modifier le contenu de lenceinte et risquer de pertuber les fonctions essentielles de la cellule Ces meacutecanimes sont communs pour certains aux bacteacuteries levures et cellules de mammifegraveres et font intervenir de nombreuses proteacuteines ayant gardeacute un certain degreacute dhomologie entre les diffeacuterents organismes
A - Translocation Ce type de transport correspond agrave un passage direct des proteacuteines au travers de la membrane via des complexes proteacuteiques formant des pores speacutecifiques de chaque organelle Il a lieu de faccedilon cotraductionnelle dans les membranes du RE des cellules de mammifegravere ou post-traductionnellement pour limport dans les mitochondries les peacuteroxisomes ou le noyau apregraves reconnaissance dune seacutequence dadressage speacutecifique vers la membrane cible Dans la plupart des cas la seacutequence signal est cliveacute une fois le chargement termineacute Pour le transfert des proteacuteines deacutejagrave replieacutees des proteacuteines chaperon coopegraverent avec le translocon pour leur deacutepliement transitoire
Translocation des glycoproteacuteines solubles
Translocation des glycoproteacuteines membranaires de type 1
Translocation des glycoproteacuteines membranaires de type 2
La translocation de proteacuteines au travers des membranes du RE a eacuteteacute caracteacuteriseacutee et implique la reconnaissance dune seacutequence signal hydrophobe en la partie N-terminale de la proteacuteine en cours de synthegravese par le complexe SRP (pour Signal Recognition Particle) son attachement agrave la membrane via un reacutecepteur pour le SRP ainsi que la liaison entre le ribosome et le complexe proteacuteique Sec61p qui assure le passage du brin proteacuteique au travers de la membrane
Rocircle de la proteacuteine SRP dans la translocation
Il existe eacutegalement des exemples de reacutetro translocation du RE vers le cytosol lorsquune proteacuteine mal replieacutee est orienteacutee par les proteacuteines chaperon telles que Bip ou calnexine vers la voie de deacutegradation par le proteacuteasome La chaine polypeptidique est deacuteplieacutee et le translocon
Sec61p alors programmeacute pour le retrotransport de la proteacuteine Une fois dans le cytosol la proteacuteine est deacuteglycosyleacutee polyubiquitinyleacutee et deacutegradeacutee par le proteacuteasome
B - Diffusion Ce mode de deacuteplacement sappuie sur la fluiditeacute des membranes qui permet la diffusion lateacuterale de ses constituants
Membrane cytoplasmique
Il pourrait ecirctre impliqueacute dans la constitution de sous-compartiments se distinguant du reste de lorganelle aux plans biochimiques et fonctionnels comme par exemple le rassemblement aux sites ERES de proteacuteines destineacutees agrave lexport ou lexistence de microdomaines agrave la membrane plasmique De mecircme il a eacuteteacute eacutevoqueacute que le mouvement anteacuterograde le long des citernes du Golgi pourrait avoir lieu selon ce principe et se poursuivrait tant que les moleacutecules ne seraient pas retenues ou renvoyeacutees dans les voies reacutetrogrades du transport Pour les proteacuteines transmembranaires reacutesidant preacutefeacuterentiellement au niveau de certaines reacutegions du Golgi (telles que les enzymes de la biosynthegravese de sucres) larrecirct de la progression pourrait deacutependre de la capaciteacute des proteacuteines agrave interagir avec la nature environnante des lipides avec un eacuteleacutement deacuteterminant qui pourrait ecirctre la longueur de la reacutegion transmembranaire de la proteacuteine ou encore de la capaciteacute de la proteacuteine sous certaines conditions de milieu agrave former des structures oligomeacuteriques de grandes tailles agrave mobiliteacute reacuteduite De faccedilon plus geacuteneacuterale la mobiliteacute des constituants membranaires paraicirct un eacuteleacutement important de leacutequilibre dynamique dont la cellule fait preuve
C - Transport veacutesiculo-tubulaire Ce mode de transport est largement utiliseacute par les proteacuteines empruntant les voies dendo- et dexocytose Il a fait lobjet ces derniegraveres anneacutees de nombreux travaux aussi bien par des approches geacuteneacutetiques chez la levure que par des reconstitutions de systegravemes membranaires in vitro De grandes avanceacutees ont eacuteteacute reacutealiseacutees en particulier dans la compreacutehension de meacutecanismes moleacuteculaires assurant la reacutegulation du traffic veacutesiculaire et sa speacutecificiteacute agrave la fois lors de la formation des veacutesicules et de leur chargement que lors de leur adressage vers lorganelle cible une fois formeacutees Lincroyable activiteacute cellulaire se fait de maniegravere coordonneacutee gracircce agrave un ensemble de proteacuteines membranaires ou cytosoliques precirctes agrave ecirctre activeacutees recruteacutees et recycleacutees en permanence et dont le jeu des interactions deacutefinit la nature des eacutechanges Suivant le trajet et les moleacutecules agrave transporter les partenaires veacutesiculaires diffegraverent
Traffic veacutesiculo-tubulaire intracellulaire
Le transport de moleacutecules entre deux compartiments peut toutefois se deacutecomposer en une succession deacutevegravenements eacuteleacutementaires Le processus de formation des veacutesicules neacutecessite le recrutement de complexes cytosoliques ou proteacuteines manteau Ce recrutement se fait via des GTPases telles que les proteacuteines Arf dynamine et peut-ecirctre Rab dans certains cas qui sassocieraient agrave la membrane apregraves eacutechange de leur GDP par du GTP eacutechange catalyseacute par la proteacuteine membranaire GEF (guanine nucleotide exchange factor) Lassemblage contigu de plusieurs complexes manteau agrave la membrane du compartiment donneur conduit localement agrave sa deacuteformation puis agrave un bourgeonnement du cocircteacute du manteau La veacutesicule dont la taille est geacuteneacuteralement comprise entre 50 et 100 nm se deacutetache du compartiment donneur emportant un chargement bien deacutefini Le contenu est en effet trieacute par des reacutecepteurs speacutecifiques pour les proteacuteines solubles (ou proteacuteines cargo) tels que les lectines ERGIC53 ou VIP36 ou par des signaux contenus dans la partie cytosolique des proteacuteines transmembranaires
Activation des membranes du compartiment donneur et recrutement des proteacuteines manteaux
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209
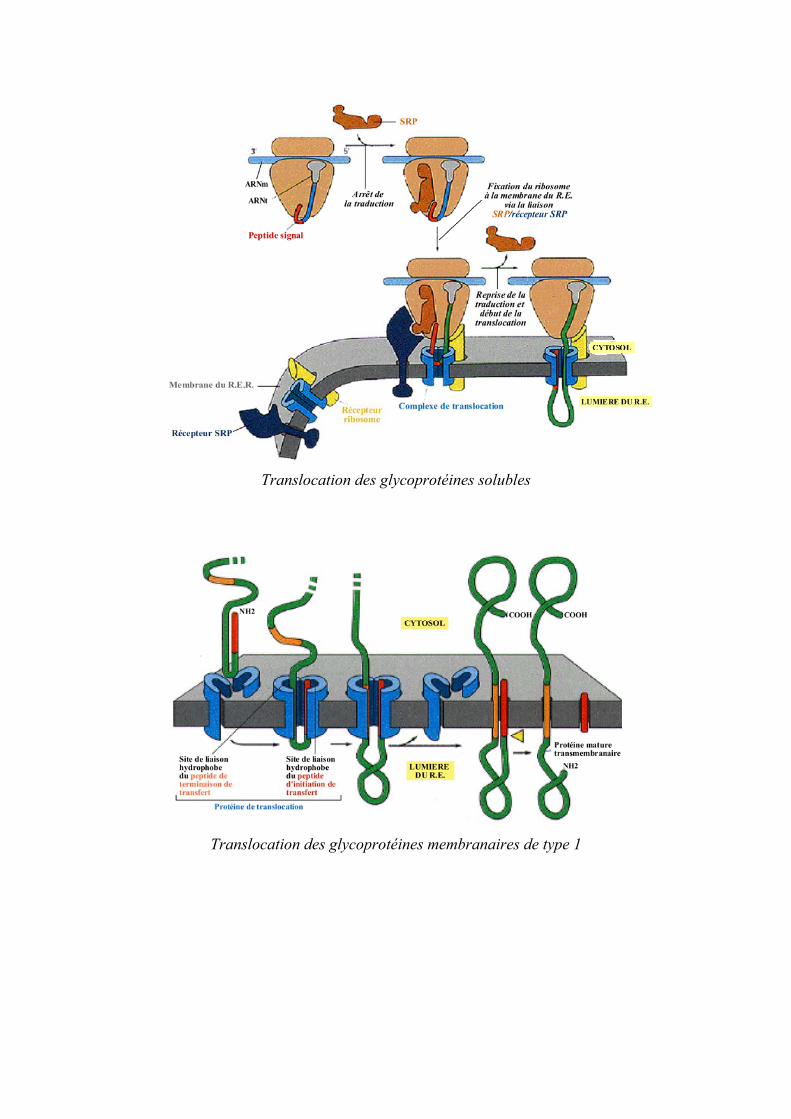
Translocation des glycoproteacuteines solubles
Translocation des glycoproteacuteines membranaires de type 1
Translocation des glycoproteacuteines membranaires de type 2
La translocation de proteacuteines au travers des membranes du RE a eacuteteacute caracteacuteriseacutee et implique la reconnaissance dune seacutequence signal hydrophobe en la partie N-terminale de la proteacuteine en cours de synthegravese par le complexe SRP (pour Signal Recognition Particle) son attachement agrave la membrane via un reacutecepteur pour le SRP ainsi que la liaison entre le ribosome et le complexe proteacuteique Sec61p qui assure le passage du brin proteacuteique au travers de la membrane
Rocircle de la proteacuteine SRP dans la translocation
Il existe eacutegalement des exemples de reacutetro translocation du RE vers le cytosol lorsquune proteacuteine mal replieacutee est orienteacutee par les proteacuteines chaperon telles que Bip ou calnexine vers la voie de deacutegradation par le proteacuteasome La chaine polypeptidique est deacuteplieacutee et le translocon
Sec61p alors programmeacute pour le retrotransport de la proteacuteine Une fois dans le cytosol la proteacuteine est deacuteglycosyleacutee polyubiquitinyleacutee et deacutegradeacutee par le proteacuteasome
B - Diffusion Ce mode de deacuteplacement sappuie sur la fluiditeacute des membranes qui permet la diffusion lateacuterale de ses constituants
Membrane cytoplasmique
Il pourrait ecirctre impliqueacute dans la constitution de sous-compartiments se distinguant du reste de lorganelle aux plans biochimiques et fonctionnels comme par exemple le rassemblement aux sites ERES de proteacuteines destineacutees agrave lexport ou lexistence de microdomaines agrave la membrane plasmique De mecircme il a eacuteteacute eacutevoqueacute que le mouvement anteacuterograde le long des citernes du Golgi pourrait avoir lieu selon ce principe et se poursuivrait tant que les moleacutecules ne seraient pas retenues ou renvoyeacutees dans les voies reacutetrogrades du transport Pour les proteacuteines transmembranaires reacutesidant preacutefeacuterentiellement au niveau de certaines reacutegions du Golgi (telles que les enzymes de la biosynthegravese de sucres) larrecirct de la progression pourrait deacutependre de la capaciteacute des proteacuteines agrave interagir avec la nature environnante des lipides avec un eacuteleacutement deacuteterminant qui pourrait ecirctre la longueur de la reacutegion transmembranaire de la proteacuteine ou encore de la capaciteacute de la proteacuteine sous certaines conditions de milieu agrave former des structures oligomeacuteriques de grandes tailles agrave mobiliteacute reacuteduite De faccedilon plus geacuteneacuterale la mobiliteacute des constituants membranaires paraicirct un eacuteleacutement important de leacutequilibre dynamique dont la cellule fait preuve
C - Transport veacutesiculo-tubulaire Ce mode de transport est largement utiliseacute par les proteacuteines empruntant les voies dendo- et dexocytose Il a fait lobjet ces derniegraveres anneacutees de nombreux travaux aussi bien par des approches geacuteneacutetiques chez la levure que par des reconstitutions de systegravemes membranaires in vitro De grandes avanceacutees ont eacuteteacute reacutealiseacutees en particulier dans la compreacutehension de meacutecanismes moleacuteculaires assurant la reacutegulation du traffic veacutesiculaire et sa speacutecificiteacute agrave la fois lors de la formation des veacutesicules et de leur chargement que lors de leur adressage vers lorganelle cible une fois formeacutees Lincroyable activiteacute cellulaire se fait de maniegravere coordonneacutee gracircce agrave un ensemble de proteacuteines membranaires ou cytosoliques precirctes agrave ecirctre activeacutees recruteacutees et recycleacutees en permanence et dont le jeu des interactions deacutefinit la nature des eacutechanges Suivant le trajet et les moleacutecules agrave transporter les partenaires veacutesiculaires diffegraverent
Traffic veacutesiculo-tubulaire intracellulaire
Le transport de moleacutecules entre deux compartiments peut toutefois se deacutecomposer en une succession deacutevegravenements eacuteleacutementaires Le processus de formation des veacutesicules neacutecessite le recrutement de complexes cytosoliques ou proteacuteines manteau Ce recrutement se fait via des GTPases telles que les proteacuteines Arf dynamine et peut-ecirctre Rab dans certains cas qui sassocieraient agrave la membrane apregraves eacutechange de leur GDP par du GTP eacutechange catalyseacute par la proteacuteine membranaire GEF (guanine nucleotide exchange factor) Lassemblage contigu de plusieurs complexes manteau agrave la membrane du compartiment donneur conduit localement agrave sa deacuteformation puis agrave un bourgeonnement du cocircteacute du manteau La veacutesicule dont la taille est geacuteneacuteralement comprise entre 50 et 100 nm se deacutetache du compartiment donneur emportant un chargement bien deacutefini Le contenu est en effet trieacute par des reacutecepteurs speacutecifiques pour les proteacuteines solubles (ou proteacuteines cargo) tels que les lectines ERGIC53 ou VIP36 ou par des signaux contenus dans la partie cytosolique des proteacuteines transmembranaires
Activation des membranes du compartiment donneur et recrutement des proteacuteines manteaux
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

Translocation des glycoproteacuteines membranaires de type 2
La translocation de proteacuteines au travers des membranes du RE a eacuteteacute caracteacuteriseacutee et implique la reconnaissance dune seacutequence signal hydrophobe en la partie N-terminale de la proteacuteine en cours de synthegravese par le complexe SRP (pour Signal Recognition Particle) son attachement agrave la membrane via un reacutecepteur pour le SRP ainsi que la liaison entre le ribosome et le complexe proteacuteique Sec61p qui assure le passage du brin proteacuteique au travers de la membrane
Rocircle de la proteacuteine SRP dans la translocation
Il existe eacutegalement des exemples de reacutetro translocation du RE vers le cytosol lorsquune proteacuteine mal replieacutee est orienteacutee par les proteacuteines chaperon telles que Bip ou calnexine vers la voie de deacutegradation par le proteacuteasome La chaine polypeptidique est deacuteplieacutee et le translocon
Sec61p alors programmeacute pour le retrotransport de la proteacuteine Une fois dans le cytosol la proteacuteine est deacuteglycosyleacutee polyubiquitinyleacutee et deacutegradeacutee par le proteacuteasome
B - Diffusion Ce mode de deacuteplacement sappuie sur la fluiditeacute des membranes qui permet la diffusion lateacuterale de ses constituants
Membrane cytoplasmique
Il pourrait ecirctre impliqueacute dans la constitution de sous-compartiments se distinguant du reste de lorganelle aux plans biochimiques et fonctionnels comme par exemple le rassemblement aux sites ERES de proteacuteines destineacutees agrave lexport ou lexistence de microdomaines agrave la membrane plasmique De mecircme il a eacuteteacute eacutevoqueacute que le mouvement anteacuterograde le long des citernes du Golgi pourrait avoir lieu selon ce principe et se poursuivrait tant que les moleacutecules ne seraient pas retenues ou renvoyeacutees dans les voies reacutetrogrades du transport Pour les proteacuteines transmembranaires reacutesidant preacutefeacuterentiellement au niveau de certaines reacutegions du Golgi (telles que les enzymes de la biosynthegravese de sucres) larrecirct de la progression pourrait deacutependre de la capaciteacute des proteacuteines agrave interagir avec la nature environnante des lipides avec un eacuteleacutement deacuteterminant qui pourrait ecirctre la longueur de la reacutegion transmembranaire de la proteacuteine ou encore de la capaciteacute de la proteacuteine sous certaines conditions de milieu agrave former des structures oligomeacuteriques de grandes tailles agrave mobiliteacute reacuteduite De faccedilon plus geacuteneacuterale la mobiliteacute des constituants membranaires paraicirct un eacuteleacutement important de leacutequilibre dynamique dont la cellule fait preuve
C - Transport veacutesiculo-tubulaire Ce mode de transport est largement utiliseacute par les proteacuteines empruntant les voies dendo- et dexocytose Il a fait lobjet ces derniegraveres anneacutees de nombreux travaux aussi bien par des approches geacuteneacutetiques chez la levure que par des reconstitutions de systegravemes membranaires in vitro De grandes avanceacutees ont eacuteteacute reacutealiseacutees en particulier dans la compreacutehension de meacutecanismes moleacuteculaires assurant la reacutegulation du traffic veacutesiculaire et sa speacutecificiteacute agrave la fois lors de la formation des veacutesicules et de leur chargement que lors de leur adressage vers lorganelle cible une fois formeacutees Lincroyable activiteacute cellulaire se fait de maniegravere coordonneacutee gracircce agrave un ensemble de proteacuteines membranaires ou cytosoliques precirctes agrave ecirctre activeacutees recruteacutees et recycleacutees en permanence et dont le jeu des interactions deacutefinit la nature des eacutechanges Suivant le trajet et les moleacutecules agrave transporter les partenaires veacutesiculaires diffegraverent
Traffic veacutesiculo-tubulaire intracellulaire
Le transport de moleacutecules entre deux compartiments peut toutefois se deacutecomposer en une succession deacutevegravenements eacuteleacutementaires Le processus de formation des veacutesicules neacutecessite le recrutement de complexes cytosoliques ou proteacuteines manteau Ce recrutement se fait via des GTPases telles que les proteacuteines Arf dynamine et peut-ecirctre Rab dans certains cas qui sassocieraient agrave la membrane apregraves eacutechange de leur GDP par du GTP eacutechange catalyseacute par la proteacuteine membranaire GEF (guanine nucleotide exchange factor) Lassemblage contigu de plusieurs complexes manteau agrave la membrane du compartiment donneur conduit localement agrave sa deacuteformation puis agrave un bourgeonnement du cocircteacute du manteau La veacutesicule dont la taille est geacuteneacuteralement comprise entre 50 et 100 nm se deacutetache du compartiment donneur emportant un chargement bien deacutefini Le contenu est en effet trieacute par des reacutecepteurs speacutecifiques pour les proteacuteines solubles (ou proteacuteines cargo) tels que les lectines ERGIC53 ou VIP36 ou par des signaux contenus dans la partie cytosolique des proteacuteines transmembranaires
Activation des membranes du compartiment donneur et recrutement des proteacuteines manteaux
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

Sec61p alors programmeacute pour le retrotransport de la proteacuteine Une fois dans le cytosol la proteacuteine est deacuteglycosyleacutee polyubiquitinyleacutee et deacutegradeacutee par le proteacuteasome
B - Diffusion Ce mode de deacuteplacement sappuie sur la fluiditeacute des membranes qui permet la diffusion lateacuterale de ses constituants
Membrane cytoplasmique
Il pourrait ecirctre impliqueacute dans la constitution de sous-compartiments se distinguant du reste de lorganelle aux plans biochimiques et fonctionnels comme par exemple le rassemblement aux sites ERES de proteacuteines destineacutees agrave lexport ou lexistence de microdomaines agrave la membrane plasmique De mecircme il a eacuteteacute eacutevoqueacute que le mouvement anteacuterograde le long des citernes du Golgi pourrait avoir lieu selon ce principe et se poursuivrait tant que les moleacutecules ne seraient pas retenues ou renvoyeacutees dans les voies reacutetrogrades du transport Pour les proteacuteines transmembranaires reacutesidant preacutefeacuterentiellement au niveau de certaines reacutegions du Golgi (telles que les enzymes de la biosynthegravese de sucres) larrecirct de la progression pourrait deacutependre de la capaciteacute des proteacuteines agrave interagir avec la nature environnante des lipides avec un eacuteleacutement deacuteterminant qui pourrait ecirctre la longueur de la reacutegion transmembranaire de la proteacuteine ou encore de la capaciteacute de la proteacuteine sous certaines conditions de milieu agrave former des structures oligomeacuteriques de grandes tailles agrave mobiliteacute reacuteduite De faccedilon plus geacuteneacuterale la mobiliteacute des constituants membranaires paraicirct un eacuteleacutement important de leacutequilibre dynamique dont la cellule fait preuve
C - Transport veacutesiculo-tubulaire Ce mode de transport est largement utiliseacute par les proteacuteines empruntant les voies dendo- et dexocytose Il a fait lobjet ces derniegraveres anneacutees de nombreux travaux aussi bien par des approches geacuteneacutetiques chez la levure que par des reconstitutions de systegravemes membranaires in vitro De grandes avanceacutees ont eacuteteacute reacutealiseacutees en particulier dans la compreacutehension de meacutecanismes moleacuteculaires assurant la reacutegulation du traffic veacutesiculaire et sa speacutecificiteacute agrave la fois lors de la formation des veacutesicules et de leur chargement que lors de leur adressage vers lorganelle cible une fois formeacutees Lincroyable activiteacute cellulaire se fait de maniegravere coordonneacutee gracircce agrave un ensemble de proteacuteines membranaires ou cytosoliques precirctes agrave ecirctre activeacutees recruteacutees et recycleacutees en permanence et dont le jeu des interactions deacutefinit la nature des eacutechanges Suivant le trajet et les moleacutecules agrave transporter les partenaires veacutesiculaires diffegraverent
Traffic veacutesiculo-tubulaire intracellulaire
Le transport de moleacutecules entre deux compartiments peut toutefois se deacutecomposer en une succession deacutevegravenements eacuteleacutementaires Le processus de formation des veacutesicules neacutecessite le recrutement de complexes cytosoliques ou proteacuteines manteau Ce recrutement se fait via des GTPases telles que les proteacuteines Arf dynamine et peut-ecirctre Rab dans certains cas qui sassocieraient agrave la membrane apregraves eacutechange de leur GDP par du GTP eacutechange catalyseacute par la proteacuteine membranaire GEF (guanine nucleotide exchange factor) Lassemblage contigu de plusieurs complexes manteau agrave la membrane du compartiment donneur conduit localement agrave sa deacuteformation puis agrave un bourgeonnement du cocircteacute du manteau La veacutesicule dont la taille est geacuteneacuteralement comprise entre 50 et 100 nm se deacutetache du compartiment donneur emportant un chargement bien deacutefini Le contenu est en effet trieacute par des reacutecepteurs speacutecifiques pour les proteacuteines solubles (ou proteacuteines cargo) tels que les lectines ERGIC53 ou VIP36 ou par des signaux contenus dans la partie cytosolique des proteacuteines transmembranaires
Activation des membranes du compartiment donneur et recrutement des proteacuteines manteaux
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209
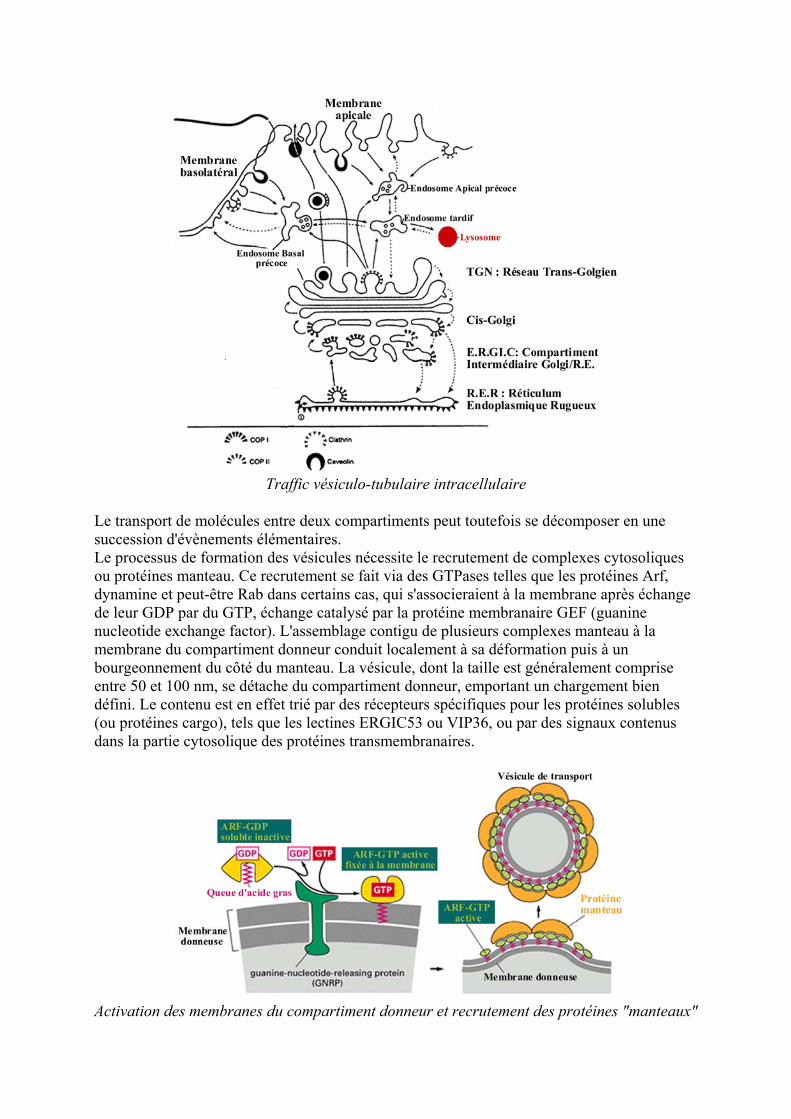
Traffic veacutesiculo-tubulaire intracellulaire
Le transport de moleacutecules entre deux compartiments peut toutefois se deacutecomposer en une succession deacutevegravenements eacuteleacutementaires Le processus de formation des veacutesicules neacutecessite le recrutement de complexes cytosoliques ou proteacuteines manteau Ce recrutement se fait via des GTPases telles que les proteacuteines Arf dynamine et peut-ecirctre Rab dans certains cas qui sassocieraient agrave la membrane apregraves eacutechange de leur GDP par du GTP eacutechange catalyseacute par la proteacuteine membranaire GEF (guanine nucleotide exchange factor) Lassemblage contigu de plusieurs complexes manteau agrave la membrane du compartiment donneur conduit localement agrave sa deacuteformation puis agrave un bourgeonnement du cocircteacute du manteau La veacutesicule dont la taille est geacuteneacuteralement comprise entre 50 et 100 nm se deacutetache du compartiment donneur emportant un chargement bien deacutefini Le contenu est en effet trieacute par des reacutecepteurs speacutecifiques pour les proteacuteines solubles (ou proteacuteines cargo) tels que les lectines ERGIC53 ou VIP36 ou par des signaux contenus dans la partie cytosolique des proteacuteines transmembranaires
Activation des membranes du compartiment donneur et recrutement des proteacuteines manteaux
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209
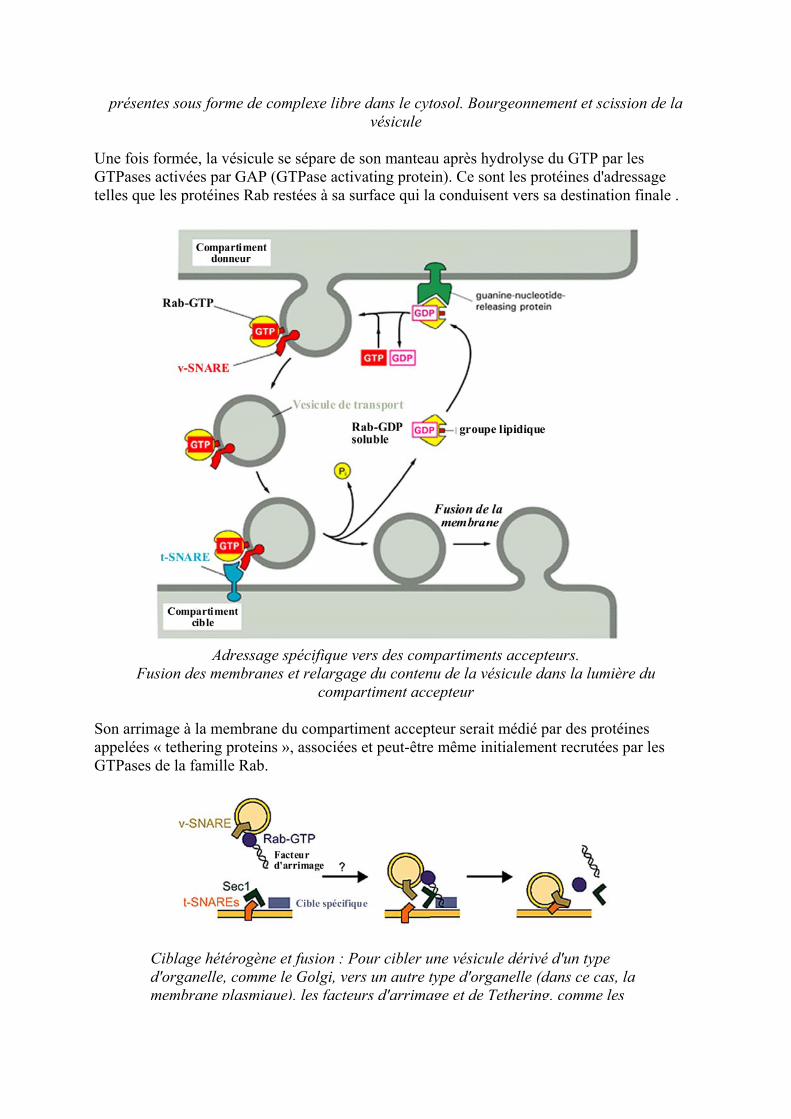
preacutesentes sous forme de complexe libre dans le cytosol Bourgeonnement et scission de la veacutesicule
Une fois formeacutee la veacutesicule se seacutepare de son manteau apregraves hydrolyse du GTP par les GTPases activeacutees par GAP (GTPase activating protein) Ce sont les proteacuteines dadressage telles que les proteacuteines Rab resteacutees agrave sa surface qui la conduisent vers sa destination finale
Adressage speacutecifique vers des compartiments accepteurs
Fusion des membranes et relargage du contenu de la veacutesicule dans la lumiegravere du compartiment accepteur
Son arrimage agrave la membrane du compartiment accepteur serait meacutedieacute par des proteacuteines appeleacutees laquo tethering proteins raquo associeacutees et peut-ecirctre mecircme initialement recruteacutees par les GTPases de la famille Rab
Ciblage heacuteteacuterogegravene et fusion Pour cibler une veacutesicule deacuteriveacute dun type dorganelle comme le Golgi vers un autre type dorganelle (dans ce cas la membrane plasmique) les facteurs darrimage et de Tethering comme les
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209
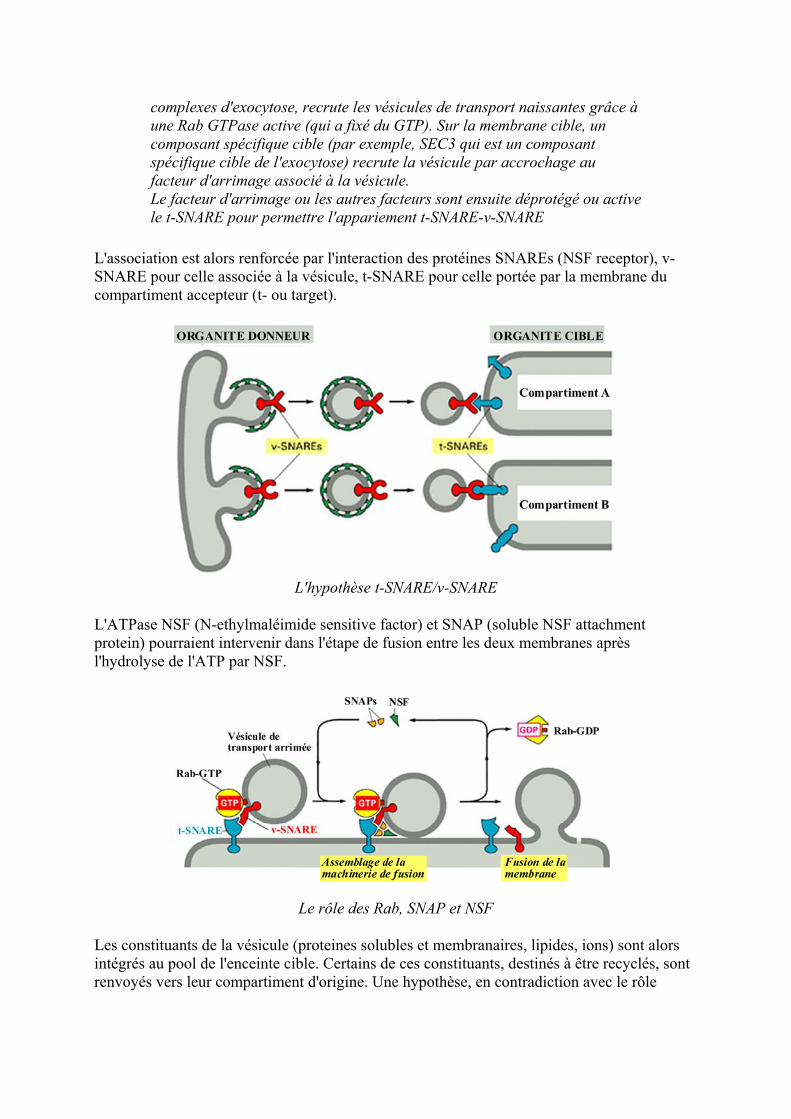
complexes dexocytose recrute les veacutesicules de transport naissantes gracircce agrave une Rab GTPase active (qui a fixeacute du GTP) Sur la membrane cible un composant speacutecifique cible (par exemple SEC3 qui est un composant speacutecifique cible de lexocytose) recrute la veacutesicule par accrochage au facteur darrimage associeacute agrave la veacutesicule Le facteur darrimage ou les autres facteurs sont ensuite deacuteproteacutegeacute ou active le t-SNARE pour permettre lappariement t-SNARE-v-SNARE
Lassociation est alors renforceacutee par linteraction des proteacuteines SNAREs (NSF receptor) v-SNARE pour celle associeacutee agrave la veacutesicule t-SNARE pour celle porteacutee par la membrane du compartiment accepteur (t- ou target)
Lhypothegravese t-SNAREv-SNARE
LATPase NSF (N-ethylmaleacuteimide sensitive factor) et SNAP (soluble NSF attachment protein) pourraient intervenir dans leacutetape de fusion entre les deux membranes apregraves lhydrolyse de lATP par NSF
Le rocircle des Rab SNAP et NSF
Les constituants de la veacutesicule (proteines solubles et membranaires lipides ions) sont alors inteacutegreacutes au pool de lenceinte cible Certains de ces constituants destineacutes agrave ecirctre recycleacutes sont renvoyeacutes vers leur compartiment dorigine Une hypothegravese en contradiction avec le rocircle
suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

suggeacutereacute plus haut pour NSF SNAP serait que NSF ninterviendrait quapregraves la fusion pour dissocier les proteines du complexe v t-SNARE et permettre leur recyclage (fig 25)
III- MECANISME MOLECULAIRE DU TRAFFIC VESICULAIRE DANS LES VOIES DE BIOSYNTHESESECRETION ET DENDOCYTOSE
A - Les veacutesicules formeacutees par les coatomegraveres COP Les veacutesicules formeacutees par les coatomegraveres COP sont chargeacutees du transport des moleacutecules dans les compartiments preacutecoces des voies de seacutecreacutetion soit entre le RE et le Golgi soit entre les citernes golgiennes Deux types de coatomegraveres ont pu ecirctre identifieacutes COPI et Il Les coatomegraveres COPII sont impliqueacutes dans le transport anteacuterograde des proteacuteines du RE vers lERGIC tandis que les complexes COPI sont associeacutes aux veacutesicules assurant les eacutechanges intra-golgiens ou le transport reacutetrograde du Golgi vers le RE
Le recrutement des coatomegraveres sur les membranes du compartiment donneur est reacuteguleacutee par la GTPase Sarl pour COPII et Arf1 pour COPI Les proteacuteines v-SNAREs peuvent eacutegalement coopeacuterer avec les GTPases en interagissant directement avec les proteacuteines du manteau comme cela a pu ecirctre mis en eacutevidence pour COPII Les coatomegraveres sont initialement preacutesents dans le cytosol sous forme de complexes dont les sous-uniteacutes proteacuteiques peuvent varier Lexistence de sous-complexes permet une reacutegulation fine du chargement et de ladressage final de la veacutesicule et en particulier de distinguer les transports anteacutero- et reacutetrogrades de COPI Certains signaux de tri impliqueacutes lors du recrutement veacutesiculaire ont eacuteteacute caracteacuteriseacutes en particulier ceux porteacutes par les proteacuteines destineacutees agrave ecirctre recycleacutees vers le RE Les proteacuteines membranaires comme P24 ou ERGIC53 qui pourraient fonctionner comme des transporteurs de proteacuteines cargo portent un motif C-terminal de type KKXX ou KXKXX exposeacute dans le cytosol et reconnu par le coatomegravere COPI Les proteacuteines solubles qui possegravedent le signal K(ou H)DEL de recapture (retrieval) sont reconnues par un reacutecepteur speacutecifique au niveau du Golgi et eacutegalement renvoyeacutees vers la lumiegravere du RE Par ailleurs la lectine ERGIC53 pourrait servir de transporteur des glycoproteacuteines entre le RE et lERGIC
B - Les veacutesicules agrave clathrine Les veacutesicules agrave clathrine identifieacutees degraves le milieu des anneacutees 70 par microscopie eacutelectronique participent agrave lendocytose de tregraves nombreux ligands apregraves interaction avec leur reacutecepteurs et au transport de certaines proteacuteines comme les enzymes lysosomales du TGN vers les endosomes Les oligomegraveres de clathrine ninterragissent pas directement avec la membrane cible mais requiert la fixation preacutealable de complexes proteacuteiques (Adaptor Protein
complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

complex) appeleacutes APl pour les veacutesicules deacuteriveacutees du TGN et AP2 pour celles formeacutees agrave partir de la membrane plasmique
Les proteacuteines manteau servent agrave initier le bourgeonnement de la veacutesicule et agrave lui imprimer
sa forme
Les complexes adapteurs de clathrine comportent diffeacuterentes sous-uniteacutes posseacutedant des homologies de seacutequence avec certaines sous-uniteacutes des coatomegraveres COP Comme dans le cas des laquoCOP-coatedraquo veacutesicules la proteacuteine Arf participe au recrutement des complexes APl sur la membrane Pour AP2 il semble que ce soit une autre GTPase la dynamine qui intervienne La dynamine sert eacutegalement lors de la scission entre la veacutesicule et la membrane plasmique par formation dun collier agrave la base de la veacutesicule naissante
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209
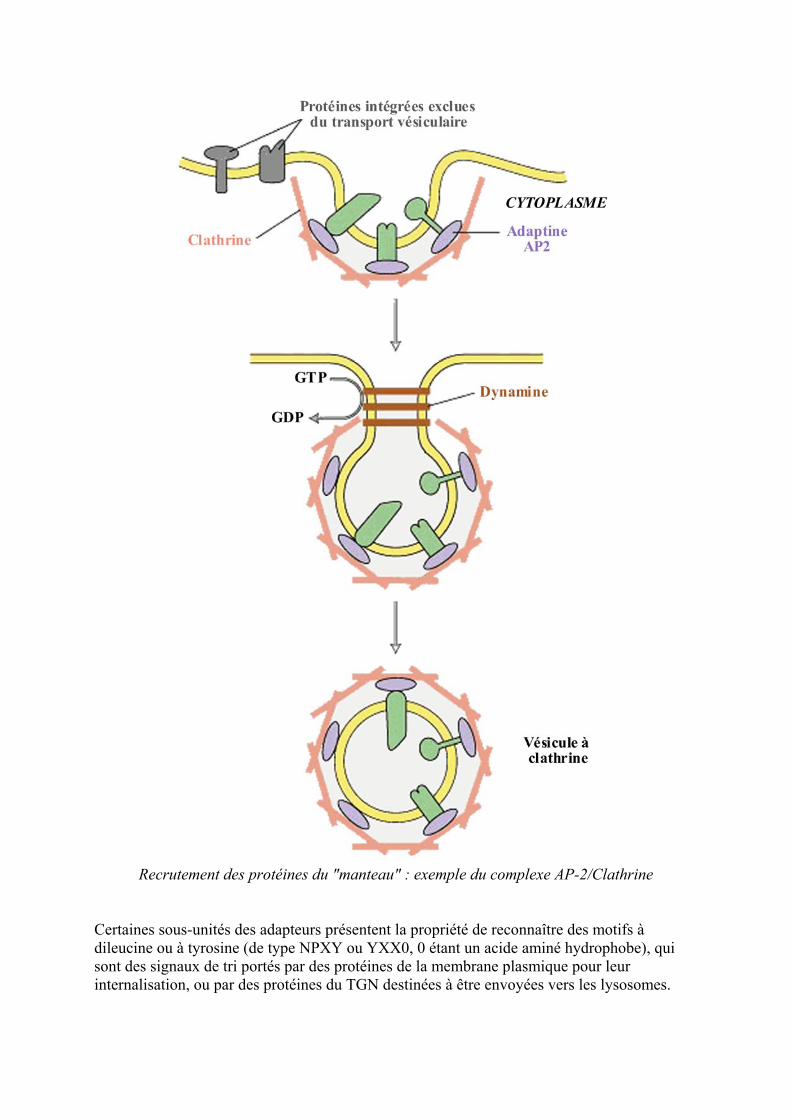
Recrutement des proteacuteines du manteau exemple du complexe AP-2Clathrine
Certaines sous-uniteacutes des adapteurs preacutesentent la proprieacuteteacute de reconnaicirctre des motifs agrave dileucine ou agrave tyrosine (de type NPXY ou YXX0 0 eacutetant un acide amineacute hydrophobe) qui sont des signaux de tri porteacutes par des proteacuteines de la membrane plasmique pour leur internalisation ou par des proteacuteines du TGN destineacutees agrave ecirctre envoyeacutees vers les lysosomes
Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

Certaines proteacuteines cargo solubles disposent aussi dun signal dadressage lysosomal le mannose 6 phosphate reconnu par le reacutecepteur membranaire MRP (mannose 6P receptor)
C - Autres types de veacutesicules Le complexe AP3 est associeacute agrave des veacutesicules qui circuleraient entre le TGN et les lysosomes ainsi quagrave des veacutesicules synaptiques Il nest cependant pas clair si AP3 comme APl ou AP2 interagit avec la clathrine pour former le manteau de ces diffeacuterents types de veacutesicules Les caveolines repreacutesentent une famille grandissante de proteacuteines associeacutees agrave des veacutesicules autres que celles recouvertes par APclathrine ou par les coatomegraveres Plusieurs types de caveolines ont eacuteteacute identifieacutees associeacutees suivant le cas agrave la membrane plasmique ou agrave des compartiments golgiens Les proteacuteines veacutesiculaires impliqueacutees dans le recrutement et ladressage apical ou basolateacuteral des proteacuteines nont pas eacuteteacute caracteacuteriseacutees de mecircme que les signaux reconnus pour le ciblage nont pas encore eacuteteacute clairement identifieacutes Ces derniers pourraient ecirctre des acides amineacutes transmembranaires particuliers des motifs sucreacutes ou encore certains lipides associeacutes aux proteacuteines
IV- EXEMPLES DINTERACTION ENTRE DES PROTEINES VIRALES ET CELLULAIRES
Les proteacuteines G du virus de la stomatite veacutesiculaire (mutant thermosensible saccumulant dans le RE agrave tempeacuterature non permissive) ou HA du virus de la grippe ont servi de modegravele pour leacutetude du repliement et du transport des proteacuteines (oligomeacuterisation mode et cineacutetique de transport dans les voies dexocytose signaux drsquoadressage basolateacuteral ou apical en cellules polariseacutees) Plus reacutecemment le rocircle de plusieurs proteacuteines virales agissant sur le transport de proteacuteines cellulaires a pu ecirctre deacutemontreacute Les eacutetudes pour la plupart encore assez descriptives visent agrave mettre en eacutevidence les cibles moleacuteculaires impliqueacutees dans ces interactions
A - Modification de lexpression de proteacuteines agrave la surface cellulaire Plusieurs proteacuteines virales semble agir sur le transport de proteacuteines cellulaires aussi importantes que des reacutecepteurs aux interleukines (Il) ou des moleacutecules du complexe majeur dhistocompatibiliteacute (MHC) impliqueacutees dans la reacuteponse de 1 hocircte agrave linfection Ainsi la proteacuteine p21I de l oncoreacutetrovirus humain HTL V-1 diminue lexpression de reacutecepteurs agrave lIp tandis que la proteacuteine vpu du virus de limmunodeacuteficience humaine HIV-1 diminue lexpression de surface des moleacutecules du MHC de classe 1 De plus la mecircme proteacuteine vpu agit de concert avec Nef pour retenir les moleacutecules CD4 dans le RE et les envoyer vers la voie de deacutegradation du proteacuteasome
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209
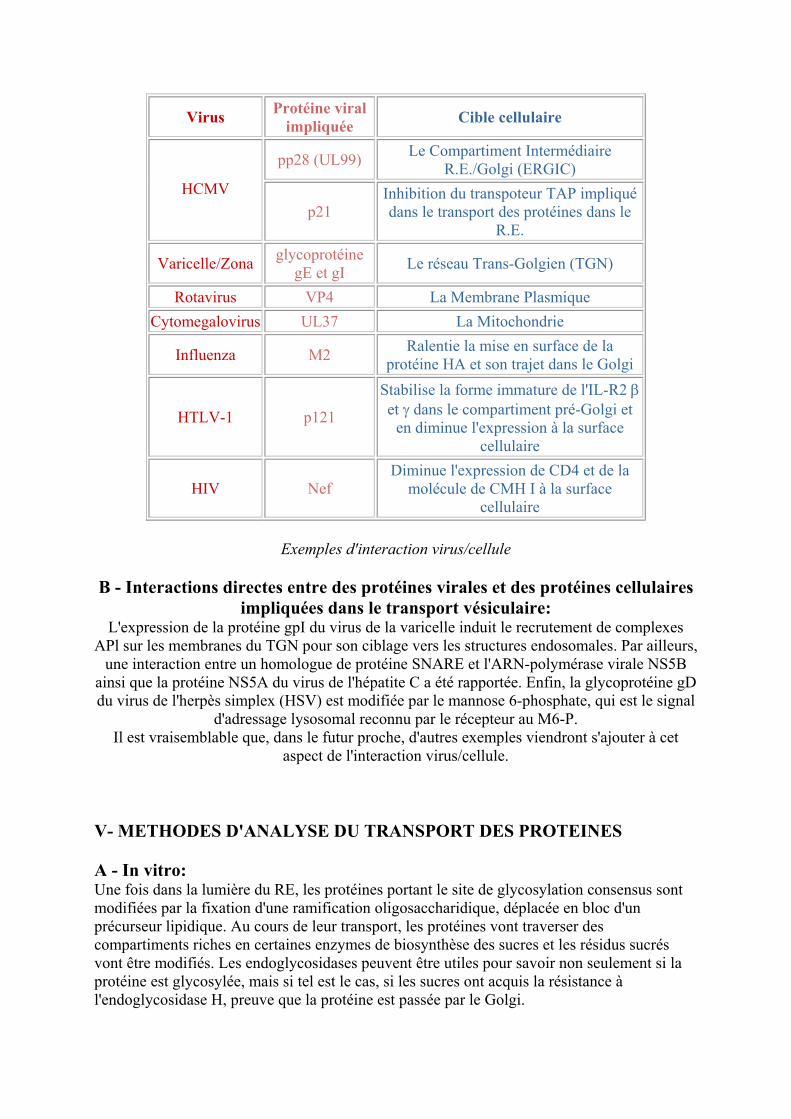
Virus Proteacuteine viral impliqueacutee Cible cellulaire
pp28 (UL99) Le Compartiment Intermeacutediaire REGolgi (ERGIC)
HCMV p21
Inhibition du transpoteur TAP impliqueacute dans le transport des proteacuteines dans le
RE
VaricelleZona glycoproteacuteine gE et gI Le reacuteseau Trans-Golgien (TGN)
Rotavirus VP4 La Membrane Plasmique Cytomegalovirus UL37 La Mitochondrie
Influenza M2 Ralentie la mise en surface de la proteacuteine HA et son trajet dans le Golgi
HTLV-1 p121
Stabilise la forme immature de lIL-R2 β et γ dans le compartiment preacute-Golgi et
en diminue lexpression agrave la surface cellulaire
HIV Nef Diminue lexpression de CD4 et de la
moleacutecule de CMH I agrave la surface cellulaire
Exemples dinteraction viruscellule
B - Interactions directes entre des proteacuteines virales et des proteacuteines cellulaires impliqueacutees dans le transport veacutesiculaire
Lexpression de la proteacuteine gpI du virus de la varicelle induit le recrutement de complexes APl sur les membranes du TGN pour son ciblage vers les structures endosomales Par ailleurs
une interaction entre un homologue de proteacuteine SNARE et lARN-polymeacuterase virale NS5B ainsi que la proteacuteine NS5A du virus de lheacutepatite C a eacuteteacute rapporteacutee Enfin la glycoproteacuteine gD du virus de lherpegraves simplex (HSV) est modifieacutee par le mannose 6-phosphate qui est le signal
dadressage lysosomal reconnu par le reacutecepteur au M6-P Il est vraisemblable que dans le futur proche dautres exemples viendront sajouter agrave cet
aspect de linteraction viruscellule
V- METHODES DANALYSE DU TRANSPORT DES PROTEINES
A - In vitro Une fois dans la lumiegravere du RE les proteacuteines portant le site de glycosylation consensus sont modifieacutees par la fixation dune ramification oligosaccharidique deacuteplaceacutee en bloc dun preacutecurseur lipidique Au cours de leur transport les proteacuteines vont traverser des compartiments riches en certaines enzymes de biosynthegravese des sucres et les reacutesidus sucreacutes vont ecirctre modifieacutes Les endoglycosidases peuvent ecirctre utiles pour savoir non seulement si la proteacuteine est glycosyleacutee mais si tel est le cas si les sucres ont acquis la reacutesistance agrave lendoglycosidase H preuve que la proteacuteine est passeacutee par le Golgi
Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

Des approches biochimiques peuvent eacutegalement ecirctre entreprises pour isoler les organelles cellulaires les unes des autres sur des gradients (de Ficoll par exemple) Il est alors possible danalyser speacutecifiquement les contenus des diffeacuterents compartimentss en fonction de traitements auxquels la cellule aura preacutealablement eacuteteacute soumise
B - In v ivo Les techniques utiliseacutees sur des cellules fixeacutees par des agents fixateurs (tels que la paraformaldeacutehyde et la glutaraldeacutehyde) ou sur des cellules vivantes sont beaucoup plus fines Il est possible deacutetudier le transport par des meacutethodes de colocalisation entre une proteacuteine dinteacuterecirct et une proteacuteine cellulaire reacutesidant dans un endroit particulier de la cellule (marqueurs cellulaires) Lanalyse dune colocalisation eacuteventuelle peut ecirctre reacutealiseacutee dans un premier temps en microscopie confocale agrave fluorescence puis si neacutecessaire en microscopie eacutelectronique avec des anticorps marqueacutes avec des billes dor Certains inhibiteurs permettent daccumuler des proteacuteines comme la proteacuteine VSV -G fusionneacutee agrave la proteacuteine GFP (green fluorescent protein) dans certains compartiments et de suivre son transport une fois linhibition leveacutee en videacuteomicroscopie agrave fluorescence Ainsi divers agents chimiques sont utiliseacutes tels que la Breacutefeldine A qui inhibe le recrutement des manteaux de type COP 1 et conduit agrave laccumulation des proteacuteines dans le RE etou lERGIC ou encore le Nocodazole qui induit la deacutepolymeacuterisation des microtubules et conduit au fractionnement du Golgi dans des structures de type ERES Enfin il est possible de bloquer le transport entre les ERES et lERGIC en incubant les cellules de mammifegraveres agrave 15degC et de bloquer le transport agrave partir du TGN agrave 20degC
VI- VIDEOMICROSCOPIE
Les eacutetudes en videacuteomicroscopie sur le transport des proteacuteines dans des cellules vivantes peuvent ecirctre consulteacutees sur les sites suivants http dirnichdnihgovcbmbpb7labobhtml http dirnichdnihgovcbmbpb1labobhtml
BIBLIOGRAPHIE
Essentiel de la Biologie Cellulaire Alberts B 1999 Garland Eds
Farquhar MG amp Hauri H-P Protein sorting and vesicular traffic in the Golgi apparatus 1997 p 63-129 dans laquoThe Golgi Apparatusraquo Berger EG amp Roth J Eds
Halban PA Irminger J-C Sorting and processing of secretory pro teins 1994 Biochem J vol 299 P 1-18
Lippincott-Schwartz J Cole NB amp Donaldson JG Building a secretory apparatus raie of Arf1COPI in Golgi biogenesis and maintenance 1998 Histochem CeU Biol vol 109 P 449-62
Le Borgne R amp Hoflack B Mechanisms of protein sorting and coat assembly insights from the clathrin-coated vesicle pathway 1998 Curr Op CeU Biol vol 10 P 499-503 Kirchhausen T Bonifacino J S amp Riezman H Linking cargo to vesicle formation receptor tail interactions with coat pro teins 1997 Curr Op CeU Biol vol 9 P 488-95
Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209

Hay JC amp Scheller RH SNAREs and NSF in targeted membrane fusion 1997 Curr Op CeU Biol vol 9 P 505-12
Chavrier P amp Goud B The raie of ARF and Rab GTPases in membrane transport 1999 Curr Op CeU Biol vol Il p 466-75
Traub LM amp Kornfeld S The Trans-Golgi network a late secretory sorting station 1997 Curr Op CeU Biol vol 9 p 527-33
Goda Y SNAREs and regulated exocytosis 1997 PNAS vol 94 P 769-72
Le Borgne R amp Hoflack B Protein transport from the secretory to the endocytic pathway in mammalian cells 1998 Biochim Biophys Acta vol 1404 p 195-209