
TRABAJO DE MAESTRIA CINDY NIÑO
55
PONTIFICIA UNIVERSIDAD JAVERIANA FACULTAD DE CIENCIAS MAESTRÍA EN CIENCIAS BIOLÓGICAS EFECTO DEL USO, MANEJO Y REGIÓN GEOGRÁFICA SOBRE LOS GENES DEL CICLO DE CARBONO Y NITRÓGENO DE LA COMUNIDAD MICROBIANA EDÁFICA EN DOS REGIONES DEL VALLE DEL CAUCA CINDY JOHANNA NIÑO CAMACHO TRABAJO DE GRADO Presentado como requisito parcial Para optar al título de MAGISTER EN CIENCIAS BIOLÓGICAS DIRECTOR ZIV ARBELI PhD. CODIRECTOR ALEJANDRO CARO QUINTERO PhD. BOGOTA D.C. COLOMBIA 2019
Transcript of TRABAJO DE MAESTRIA CINDY NIÑO

PONTIFICIA UNIVERSIDAD JAVERIANA
FACULTAD DE CIENCIAS
MAESTRÍA EN CIENCIAS BIOLÓGICAS
EFECTO DEL USO, MANEJO Y REGIÓN GEOGRÁFICA SOBRE LOS GENES
DEL CICLO DE CARBONO Y NITRÓGENO DE LA COMUNIDAD MICROBIANA
EDÁFICA EN DOS REGIONES DEL VALLE DEL CAUCA
CINDY JOHANNA NIÑO CAMACHO
TRABAJO DE GRADO
Presentado como requisito parcial
Para optar al título de
MAGISTER EN CIENCIAS BIOLÓGICAS
DIRECTOR ZIV ARBELI PhD.
CODIRECTOR ALEJANDRO CARO QUINTERO PhD.
BOGOTA D.C.
COLOMBIA
2019

NOTA DE ADVERTENCIA
"La Universidad no se hace responsable por los conceptos emitidos por sus alumnos en sus trabajos de tesis. Solo velará por que no se publique nada contrario al dogma y a la moral católica y porque las tesis no contengan ataques personales contra persona alguna, antes bien se vea en ellas el anhelo de buscar la verdad y la justicia".
Artículo 23 de la Resolución No13 de Julio de 1946.


EFECTO DEL USO, MANEJO Y REGIÓN GEOGRÁFICA SOBRE LOS GENES DEL
CICLO DE CARBONO Y NITRÓGENO DE LA COMUNIDAD MICROBIANA
EDÁFICA EN DOS REGIONES DEL VALLE DEL CAUCA
CINDY JOHANNA NIÑO CAMACHO
APROBADO
CONCEPCIÓN PUERTA B. PhD ALBA ALICIA TRESPALACIOS Ph.D.
DECANA DIRECTORA DEL POSGRADO
FACULTAD DE CIENCIAS FACULTAD DE CIENCIAS

DEDICATORIA
Es mi deseo como sencillo gesto de agradecimiento, dedicarle mi Trabajo de Grado, a mis padres Pedro y Eudalia, mis hermanos Laura y Santiago y mi tía Martha por brindarme la confianza, consejos, oportunidad y recursos para lograrlo, por estar siempre en esos momentos difíciles.

AGRADECIMIENTOS A la Pontificia Universidad Javeriana y toda su planta de docente, a mi Director Ziv Arbeli y mi codirector Alejandro Caro por la dedicación y apoyo que ha brindado a este trabajo, por el respeto a mis sugerencias e ideas y por la dirección y el rigor que ha facilitado a las mismas, por su orientación y atención a mis consultas sobre metodología, gracias por la confianza ofrecida. Al grupo de investigación USBA y sus integrantes por recibirme, ofrecerme sus instalaciones y ayudarme en la realización de este proyecto. Gracias a mi gran amigo Julián quien siempre me ha prestado un gran apoyo moral y humano, necesarios en los momentos difíciles de este trabajo y esta profesión. Gracias a Mario Leandro y Juliana por toda su colaboración en el final de este proceso. Por, sobre todo, gracias a mis padres, a mis hermanos Laura y Santiago y a mi tía Martha, por su paciencia, comprensión y solidaridad con este proyecto.

RESUMEN
La intensificación de las actividades agrícolas y pecuarias asociada a la creciente demanda mundial de alimentos conducen a cambios en la estructura física y bioquímica de los suelos, como resultado de la adopción de monocultivos y la simplificación de las interacciones biológicas. Esto desencadena en la reducción de la biodiversidad y aumento en la emisión de gases de efecto invernadero. Una de los indicadores de degradación del suelo es la pérdida de la diversidad y de funciones de las comunidades microbianas. En la última década se han empleado como más frecuencia las técnicas de secuenciación de ADN para explorar la composición y función microbiana del suelo, como la secuenciación de marcadores moleculares para evaluar la composición taxonómica (e.g., 16S rARN) y secuenciación metagenómica por shotgun para estudiar los genes funcionales de las comunidades y su potencial funcional. No obstante, la secuenciación metagenómica shotgun del suelo es difícil debido a la mega diversidad de las comunidades microbianas del suelo que requiere de una altísima inversión económica en cuanto a secuenciación y hace imposible emplear esta técnica de forma frecuente en los estudios de suelo. En los últimos años se han desarrollado metodologías como el PICRUSt que permiten realizar predicciones funcionales de genes a partir de secuencias de 16S rARN. Estas herramientas basan sus predicciones en la presencia de genomas completos y disponibles relacionados cercanamente a los taxones identificados por el 16S rRNA. En consecuencia, estos métodos funcionan mejor en ambientes con alto número de genomas secuenciados como los estudios del microbioma del ser humano que en estudios con poca representatividad en las bases de datos como los estudios del suelo. Por lo tanto, el presente estudio pretende establecer si las herramientas de predicción como PICRUSt pueden ser utilizadas en suelo para establecer el potencial funcional de las comunidades microbianas y así establecer el efecto del uso y manejo del suelo en la composición y funciones microbianas. Para esto se comparó la predicción de genes por PICRUSt con datos de secuenciación metagenómica. Una vez establecidas que funciones poseen una alta correlación entre la predicción y el metagenoma se evaluó el efecto de distintos usos y manejos agrícolas sobre dichas funciones. Para establecer el efecto de estas prácticas en el suelo como un paso importante para diseñar mejores prácticas agrícolas a futuro. En el presente trabajo se utilizaron datos de secuenciación que fueron obtenidos en un trabajo de grado anterior. Estos datos se obtuvieron a partir de muestras de suelo colectadas en dos regiones del departamento de Valle del Cauca, Colombia, con diferentes usos y manejo del suelo: silvopastoril intensivo, pasto convencional, caña de azúcar con manejo convencional y agroecológico y bosque seco tropical. PICRUSt predijo 12111 anotaciones funcionales, mientras que el análisis metagenómico identificó solo 6188 anotaciones funcionales. La correlación de Spearman entre la abundancia de todos los genes obtenidos por PICRUSt y por secuenciación metagenómica

fue relativamente baja. Esta misma correlación para los genes relacionados con los ciclos de Nitrógeno, Carbono fue considerablemente más alta. Además, se calculó el Índice de Taxón Secuenciado Más Cercano (NSTI), que está relacionada con la incertidumbre de predicción, varían entre 0.25 y 0.05 en los diferentes sitios de muestreo. Para evaluar el efecto del uso y manejo del suelo, así como la región geográfica sobre la abundancia de genes funcionales, se realizó un análisis de coordenadas principales (PCoA), para mostrar el ordenamiento de las muestras en los ciclos de carbono y nitrógeno, además se realizó un análisis estadístico PERMANOVA para determinar los efectos del uso, manejo y región sobre los mismos ciclos. Asimismo, se comparó la abundancia de diferentes genes de los ciclos del nitrógeno y carbono. Se observó una agrupación de todos los genes por manejo y región en la predicción de PICRUSt. En el ciclo del nitrógeno se encontró una separación representativa por región, en el ciclo del carbono se observó una separación entre los suelos de bosques y suelos intervenidas. Las abundancias de genes relacionados con la metanógenesis fueron mayores en sistemas con manejo convencional y bajo en sistemas con manejo agroecológico con un valor P significativo de 0.016, en el metabolismo de carbohidratos valores altos en bosque y todos los sistemas de El Cerrito con valor P 0.001. En oxidación de metano no se encontraron diferencias significativas. En la fijación de carbono valores altos en Bosque y cañas y bajos en silvopastoril y Pasturas, encontrando diferencias significativas del uso del suelo con valor P 0.0186. En cuanto al ciclo del nitrógeno, la fijación de nitrógeno se encontró valores altos en los sistemas de El Cerrito y bajos en Bugalagrande, concordando con diferencias significativas en cuanto a región valor P 2e-16. En reducción asimilatoria de nitratos fue alta en bosque, seguido por altos en manejo agroecológico y bajos en manejo convencional validado por la diferencia significativa del manejo del suelo valor P 0.0034. En la nitrificación se encontró valores bajos en suelos con uso de ganadería y altos en cañas y bosque, afirmando resultados de diferencia significativa del uso del suelo valor P 0.012. Por último, en la desnitrificación los valores fueron muy variables concordando con los resultados de análisis estadísticos donde no se encontró diferencia significativa. PALABRAS CLAVES: Metagenómica, PICRUSt, agroecológico y agricultura convencional

SUMMARY The intensification of agricultural and livestock activities associated with the growing global demand for food, leads to changes in the physical and biochemical structure of soils, because of the adoption of monocultures and the simplification of biological interactions. The reduction of biodiversity and an increase in the emission of greenhouse gases. One of the indicators of soil degradation is the loss of diversity and functions of microbial communities. In the last decade, DNA sequencing techniques have been used more frequently to explore soil composition and microbial function, such as molecular marker sequencing to assess taxonomic composition (eg, 16S rRNA) and metagenomic sequencing by shotgun to study functional genes of the communities and their functional potential. However, the shotgun metagenomic sequencing of the soil is difficult due to the mega diversity of the microbial communities of the soil that requires a very high economic investment in terms of sequencing and makes it impossible to use this technique frequently in soil studies. In recent years, methodologies such as PICRUSt have been developed that allow functional predictions of genes from 16S rRNA sequences. These tools base their predictions on the presence of complete and available genomes closely related to the taxa identified by the 16S rRNA. Consequently, these methods work better in environments with a high number of sequenced genomes such as human microbiome studies than in studies with little representation in databases such as soil studies. Therefore, the present study aims to establish whether prediction tools such as PICRUSt can be used in soil to establish the functional potential of microbial communities and thus establish the effect of soil use and management on microbial composition and functions. For this, the prediction of genes by PICRUSt was compared with metagenomic sequencing data. Once established which functions have a high correlation between prediction and metagenome, the effect of different agricultural uses and management on these functions was evaluated. To establish the effect of these practices on the soil as an important step to designing better agricultural practices in the future. In the present work, sequencing data were used that were obtained in a previous grade work. These data were obtained from soil samples collected in two regions of the department of Valle del Cauca, Colombia, with different uses and soil management: intensive silvopastoral, conventional grass, sugarcane with conventional and agro-ecological management and tropical dry forest. PICRUSt predicted 12111 functional annotations, while the metagenomic analysis identified only 6188 functional annotations. Spearman's correlation between the abundance of all genes obtained by PICRUSt and by metagenomic sequencing was relatively low. This same correlation for the genes related to the nitrogen, carbon cycles was considerably higher.

In addition, the Nearest Sequenced Taxon Index (NSTI), which is related to the uncertainty of prediction, was calculated varying between 0.25 and 0.05 at different sampling sites. To assess the effect of land use and management, as well as the geographic region on the abundance of functional genes, a principal coordinate analysis (PCoA) was performed, to show the ordering of the samples in the carbon and nitrogen cycles, in addition A PERMANOVA statistical analysis was performed to determine the effects of use, management, and region on the same cycles. Likewise, the abundance of different genes of the nitrogen and carbon cycles was compared. A grouping of all genes by management and region was observed in the prediction of PICRUSt. In the nitrogen cycle, a representative separation by region was found, in the carbon cycle a separation between forest soils and intervened soils was observed. The abundances of genes related to methanogenesis were higher in systems with conventional management and low in systems with agroecological management with a significant P value of 0.016, in carbohydrate metabolism high values in the forest and all El Cerrito systems with P-value 0.001. In methane oxidation, no significant differences were found. In the fixation of carbon high values in Forest and reeds and low in silvopastoral and Pastures, finding significant differences in land use with P-value 0.0186. Regarding the nitrogen cycle, nitrogen fixation was found high in El Cerrito systems and low in Bugalagrande, agreeing with significant differences in value region P 2e-16. In the assimilatory reduction of nitrates was high in the forest, followed by high in agroecological management and low in conventional management validated by the significant difference in soil management value P 0.0034. In nitrification, low values were found in soils with the use of livestock and high in reeds and forest, affirming results of significant difference in land use value P 0.012. Finally, in denitrification, the values were very variable according to the results of statistical analysis where no significant difference was found. KEYWORDS: Metagenomics, PICRUSt, agroecological and conventional

1. INTRODUCCIÓN
La agricultura convencional ha conducido a procesos de degradación del suelo, dentro de
los cuales se encuentran la pérdida de materia orgánica, erosión, inundaciones,
salinización, deslizamientos, compactación y contaminación (Acosta et al., 2010; Allison
et al., 2005; Meyer et al., 2008) Es el suelo, el requisito más esencial para la producción
de alimentos, y es así que existe gran debate sobre su uso y sostenibilidad, particularmente
en relación a la agricultura.
La sostenibilidad del suelo hace referencia a la conservación de los recursos naturales.
Dado que la población mundial va en aumento, los sistemas alternativos deben ser
altamente productivos, y de no cumplirse, esto conlleva a mayor desplazamiento de
ecosistemas naturales. Una alternativa que se ha dado para tratar de mitigar los efectos
del manejo convencional es la implementación de manejos de tipo agroecológico como
los sistemas de cultivo de caña agroecológica y silvopastoril, con la intención de
disminuir o evitar la degradación del suelo coadyuvando a la conservación de sus
características iniciales (Garrigues et al., 2012; Vallejo et al., 2010).
Los microorganismos del suelo son componentes integrales de una amplia gama de
procesos ecosistémicos, incluidos el ciclo de nutrientes, el almacenamiento de carbono
en el suelo, la producción de gases de efecto invernadero, la degradación de xenobióticos
y contaminantes, al igual que del mantenimiento de su estructura (Xue et al., 2010;
Groffman y Bohlen, 1999). A pesar de su importancia, las interacciones microbianas no
suelen ser fáciles de clasificar. Se han realizado estudios extensos sobre él cuáles y qué
tipo de microorganismos son más abundantes o diversos en suelo de acuerdo a los
cambios de factores ambientales y el uso de suelo (Lovera & Cuenca, 2007; León-
Gamboa et al., 2010; Wagg et al., 2014). Sin embargo, aún no se cuenta con el suficiente
conocimiento para elaborar modelos predictivos, ni para entender cómo el cambio en la
comunidad microbiana afecta la función del suelo.
El estudio de la influencia de la comunidad microbiana sobre el suelo, ha sido complejo
y es por tanto que se hace necesario el uso de herramientas moleculares para su análisis,
tales como análisis filogenéticos basados en 16 ARNr y secuenciación metagenómica

(Elsacker, 2011). Estos generan miles de datos, por lo que el esfuerzo para el análisis de
los datos ha conllevado su vez al uso de herramientas bioinformáticas como es el caso de
QIIME (Quantitative Insigths Into Microbial Ecology), MG RAST (Metagenomic Rapid
Annotations) (Liu, et al., 2013) y muchas más. Uno de los últimos avances
bioinformáticos que permiten utilizar información de la mayoría de bases de datos
genómicas es PICRUST (Phylogenetic Investigation of Communities by Reconstruction
of Unobserved States), este, es un programa que partir de datos de 16S ARNr, y por medio
de algoritmos busca genomas de referencia publicados en todas las bases del mundo y
predice el metagenoma de una amplia gama de muestras asociadas a diferentes ambientes.
La exactitud de la predicción depende de los genomas de referencia que estén incluidos
en las bases de datos por esto el programa ha sido usado en gran medida en muestras de
microbiomas humanos y en muestras de cultivo de suelo en donde existe una gran
variedad de genomas de referencia (Langille et al., 2013).
En Colombia, en la región del Valle del Cauca, se han establecido sistemas
agroecológicos como la caña orgánica y los sistemas silvopastoriles intensivos
(policultivos). De hecho los sistemas silvopastoriles intensivos se han desarrolladas en
Colombia y México, donde la reserva natural El Hatico (El Cerrito, Valle del Cauca) fue
unas de las fincas pioneras de establecer estos sistemas y se convirtió en un modelo con
numerosos estudios (Calle et al., 2012, 2013; Vallejo 2010, 2012; Cubillos 2016) En
donde se ha evaluado el efecto de las propiedades fisicoquímicas sobre actividades
enzimáticas al compararlo con un sistema de pastura convencional mostrando como
resultados un aumento de las actividades enzimáticas en los sistemas silvopastoril de 12
años, además se concluye que los sistemas silvopastoriles promueven la sostenibilidad
del ecosistema al mejorar la capacidad de los suelos para realizar la descomposición y
mineralización de nutrientes resultado reflejado por las actividades hidrolíticas (Vallejo
et al., 2010). En otros estudios en la misma región se ha estudiado el impacto de la
conversión de pasturas convencionales a sistemas silvopastoriles, en las comunidades
microbianas del suelo y las propiedades fisicoquímicas del suelo por medio de perfiles
FAME, indicando que la estructura y la composición de la comunidad microbiana del
suelo cambian según los sistemas de manejo del suelo (Vallejo et al., 2012). En resultados
de Cubillos et al, 2016 estudio realizado en la misma zona se encontró que las huellas
digitales de electroforesis en gel de gradiente desnaturalizante (DGGE) del gen 16S ARNr
y el gen amoA indicaron que las comunidades de bacterias del suelo variaron entre usos

del suelo, con mayores similitudes entre bosque y silvopastoril en comparación con
pasturas convencionales. Un último estudio realizado en esta zona por Sáenz, 2014 (datos
no publicados) evaluó el efecto del uso del suelo en dos regiones del Valle del Cauca
sobre las comunidades microbianas por medio de 16S ARNr de sistemas de pastoreo y
cultivos de caña en donde encontró que la estructura y composición de las comunidades
microbianas se ve afectada por el uso del suelo más que por el manejo y la región, además
realizó un análisis metagenómico en solo 12 muestras por lo tanto estos datos no fueron
usados por la falta de réplicas.
Este trabajo tuvo como objetivo evaluar el efecto del uso (ganadería, caña de azúcar y
bosque seco tropical) y manejo (convencional vs agroecológico) del suelo sobre la
abundancia de genes implicados en los ciclos de carbono y nitrógeno en el Valle del
Cauca y así aportar al conocimiento para de pronto en el futuro resolver los siguientes
preguntas: ¿Cómo las diferentes prácticas y usos agrícolas modifican el potencial
funcional de los microorganismos del suelo? y como estos cambios afectan la fertilidad y
la sostenibilidad del suelo.

2. MARCO TEÓRICO Y ESTADO DEL ARTE
2.1. Efecto de la agricultura sobre el suelo
La actividad agrícola es uno de los problemas más extendidos del siglo XXI (Vermeulen
et al., 2012). Para seguir el ritmo de la creciente población humana, el área total de tierras
cultivadas en todo el mundo ha aumentado más del 500% en las últimas cinco décadas,
con un aumento del 700% en el uso de fertilizantes (Elsacker et al., 2011). La
intensificación agrícola ha planteado una amplia gama de problemas ambientales, que
incluyen una pobre eficiencia en el uso de nutrientes, mayores emisiones de gases de
efecto invernadero, eutrofización de aguas subterráneas, degradación de la calidad del
suelo y erosión del suelo (Calle & Murgueitio, 2013).
El uso del suelo para la agricultura conduce a la pérdida de carbono orgánico, una menor
cobertura y profundidad de las raíces, además de presión física por el pisoteo constante
del ganado, en sistemas de ganadería, lo cual, produce cambios en la estructura del suelo,
genera compactación, disminución en la estabilidad de los agregados, en la porosidad del
suelo, la aireación, la capacidad de infiltración, acelerando los procesos de erosión, y
deterioro de la calidad y fertilidad del suelo (Jangid et al.,2008).
Los sistemas agrícolas alternativos como los sistemas agroecológicos, los cuales se han
planteado para mitigar el efecto de la agricultura al cambio climático. Dado que las
prácticas agroecológicas incluyen una integración de varias herramientas, labranza
reducida, no uso de fertilizantes sintéticos ni pesticidas y diversidad de cultivos como
sistemas silvopastoril en donde se integran pastos, hierbas y árboles lo cual aporta
beneficios a los suelos mejorando el ciclo de nutrientes y las interacciones biológicas,
aumenta la fertilidad del suelo y reduce las emisiones del dióxido de carbono, óxido
nitroso y metano (Fromin et al., 2002). Estos sistemas se han acogido para tratar de
mantener la sostenibilidad ambiental y los servicios de los ecosistemas,
Dentro de algunos ejemplos de sistemas de agricultura convencional y agricultura
agroecológica se encuentran:

2.1.1. Cultivos de Caña
Los cultivos de caña convencional que utiliza el arado, fertilizantes inorgánicos,
plaguicidas y la quema de los residuos de cultivos tienen efectos negativos sobre las
reservas de carbono, los agregados del suelo, la estructura del suelo, la compactación, la
biomasa, las actividades enzimáticas, y la fertilidad (Galdos, et al., 2009). El manejo
agronómico ecológico de la caña de azúcar se diferencia en que los residuos cortados no
son quemados y se introducen al suelo junto con los residuos del bagazo como abono para
posteriores cultivos. Como consecuencia, se aumenta el contenido de carbono del suelo
y se disminuyen el uso de fertilizantes inorgánicos.
2.1.2. Sistemas Silvopastoriles
Los sistemas silvopastoriles intensivos son un ejemplo de sistemas agroecológicos.
Fueron desarrollados en Colombia y México como respuesta a problemas ambientales y
a la baja sostenibilidad de las actividades ganaderas convencionales. Estos sistemas
intensivos integran árboles y altas densidades de arbustos y hierbas, mientras que las
entradas de labranza y agroquímicos no son necesarias (Pezzopane et al., 2019; Plieninger
& Huntsinger 2018; Cubillos et al., 2012;). La introducción de árboles y arbustos para
sistemas de pastoreo aumenta la biomasa por encima y por debajo del suelo lo que
proporciona mayor entrada de material orgánico y otros nutrientes, estos además mejoran
la estructura del suelo (Pezzopane et al., 2019). Los sistemas silvopastoriles mejoran las
propiedades del suelo, reducen la necesidad de fertilizantes externos, pesticidas y riego,
a la vez contribuyen al secuestro de carbono, además de reducir al mínimo el estrés
generado por el ganado y preservar la biodiversidad del suelo (Drenovsky et. al., 2014).
2.2. Importancia de los microorganismos en el suelo
Las comunidades microbianas desempeñan un papel indispensable en los ecosistemas y
prestan una amplia gama de servicios. En los agroecosistemas, los microorganismos

modulan una serie de procesos, incluidos el ciclo de nutrientes, la descomposición de la
materia orgánica, la estabilización de agregados del suelo, las interacciones simbióticas y
patógenas con las plantas, por lo tanto, desempeñan un papel esencial en la productividad
y la sostenibilidad de los agroecosistemas (Bender et al., 2016). La actividad agrícola con
alto uso de recursos y baja diversidad de cultivos puede afectar los microorganismos
asociados con el suelo y la planta, y por consiguiente impactar los servicios de los
ecosistemas (Bender et al., 2016).
Los ciclos biogeoquímicos son las vías a través de las cuales los nutrientes y los minerales
se utilizan, reciclan o almacenan en el suelo. Varios macro y micronutrientes han sido
identificados como cruciales para la salud del suelo y las plantas; algunos de ellos son:
Nitrógeno, Fósforo, Potasio, Calcio, Magnesio y Azufre (macronutrientes), Boro, Cobre,
Hierro, Cloruro, Manganeso, Molibdeno y Zinc (micronutrientes). Estos nutrientes se
clasifican como "micro" o "macro" dependiendo de las concentraciones que se consideran
necesarias para apoyar el crecimiento de las plantas (Leff et al., 2018).
Las actividades enzimáticas del suelo pueden proporcionar una instantánea de los
procesos funcionales que ocurren en él, pero no siempre es evidente qué microorganismos
son responsables de estos procesos. Los métodos microbiológicos y filogenéticos para la
identificación de especies microbianas, por lo tanto, van de la mano con las mediciones
de la actividad enzimática y proporcionan una vista más detallada del funcionamiento del
microbioma del suelo.
2.3. Efecto del uso del suelo sobre la comunidad edáfica del suelo
El cambio en el uso de la tierra es uno de los principales trastornos ambientales a nivel
mundial, que contribuye en gran medida al cambio climático, la pérdida de servicios de
los ecosistemas y la diversidad de especies microbianas. La conversión del uso de la tierra
se ha considerado como una actividad antropogénica intensiva que altera en gran medida
el funcionamiento del ecosistema del suelo.
Las prácticas de manejo y uso del suelo pueden influir en los roles funcionales de los
microorganismos del suelo mediante la modificación de la cantidad y calidad de los
insumos de materia orgánica, por ejemplo las prácticas agrícolas intensivas contribuyen

a la pérdida de la materia orgánica del suelo, causa problemas ambientales como la
erosión del suelo, la desertificación, eutrofización lo cual altera las actividades
microbianas y, en última instancia, afecta la calidad del suelo ( Deng et al., 2018; Ghoshal,
2017).
La variación de los tipos de residuos de plantas, como hojarasca y raíces finas, la dinámica
de nutrientes (e.g., carbono orgánico y el nitrógeno) y la composición de especies de
plantas durante los cambios en los usos del suelo, son factores determinantes en la
cantidad y calidad de las entradas de sustratos al suelo. Estos factores a su vez afectan la
dinámica de la comunidad microbiana (Zhao et al., 2014).
Los estudios de metagenómica del suelo están aumentando en todo el mundo y han
ayudado a determinar la potencial función de los microorganismos en el suelo. Los
perfiles metabólicos encontrados en algunos estudios han demostrado que el tipo de
manejo agrícola afecta el metabolismo de aminoácidos y carbohidratos, lípidos e
isoprenoides en el suelo (Fierer et al., 2012).
2.4. Herramientas moleculares y bioinformáticas que ayudan a evaluar la
diversidad funcional de los microorganismos
Los métodos tradicionales para medir la actividad microbiana en el suelo involucran
estimaciones de biomasa utilizando técnicas como la fumigación-incubación, la
fumigación-extracción y la respiración inducida por sustrato o la extracción (Sparling et
al, 1981; Oades & Jenkinson 1979) Estos métodos proporcionaban información sobre la
abundancia y la actividad de comunidades microbianas en un hábitat (Taylor et al., 2002).
Pero muchos de estos métodos bioquímicos de estimación de la biodiversidad microbiana
han sido reemplazados por los moleculares. Estas técnicas se centran en clasificar
microorganismos según su contenido genómico y no propiedades fisiológicas o
bioquímicas.
La reasociación e hibridación de ADN fue uno de los primeros métodos moleculares
utilizados para la identificación de especies y sirvió como estándar para definiciones
posteriores de identidad de secuencia y delimitación de especies. Este método se basa en

que la tasa de reasociación del ADN desnaturalizado dependerá de la similitud de las
secuencias presentes. Bajo condiciones específicas, el valor de media asociación del ADN
puede usarse como un índice de diversidad (Cho & Tiedje, 2001)
La reacción en cadena de la polimerasa o PCR, permitió el desarrollo de varias técnicas
moleculares para evaluar la diversidad y la función microbiana que involucraron la
clonación o amplificación de regiones específicas de ADN procariota. Una de las técnicas
más populares en esta categoría es la electroforesis en gel de gradiente desnaturalizante o
DGGE. El ADN extraído de muestras de suelo se une a una pinza GC de 35-40 pb y se
separa en un gel de poliacrilamida con una concentración creciente de desnaturalizantes,
esto permite que el ADN se desnaturalice y se funda en dominios específicos de secuencia
que luego migran diferencialmente a través del gel (Muyzer et al., 1993).
Otra herramienta que utiliza polimorfismos de ADN procarióticos es el polimorfismo de
longitud de fragmentos de restricción (RFLP). El ADN ribosomal amplificado por PCR
se digiere con una enzima de restricción de corte par de 4 bases. Se detectan diferentes
longitudes de fragmentos utilizando electroforesis en gel de poliacrilamida de agarosa o
no desnaturalizante, y estos patrones de bandas se usan para identificar cambios en la
estructura de la comunidad bacteriana (Liu et al., 1997). Esto permite un patrón de bandas
más simple, así como una mejor resolución taxonómica, ya que cada banda etiquetada
puede considerarse como una unidad taxonómica-OTU (Tiedje et al., 1999).
El gen 16S ARNr es quizás la herramienta más popular utilizada para la identificación
taxonómica de microorganismos en estudios ambientales, debido a una variedad de
razones. La combinación de tecnología de secuenciación rentable y la importancia
taxonómica del gen 16S ARNr son responsables de un progreso sin precedentes en el
campo de la ecología microbiana del suelo.
Actualmente se utilizan diferentes herramientas bioinformáticas para el análisis de datos
metagenómicos microbianos, los más comunes son MG-RAST o MetaGenomic Rapid
Annotation (Meyer et al., 2008), MEGAN o MEtaGenome ANalyzer (Huson et al., 2007)
QIIME o Quantitative Insights Into Microbial Ecology) (Caporaso et al., 2011). Una de
los últimos avances en herramientas bioinformáticas que permiten predecir el potencial
funcional a partir de la filogenia de los microorganismos es PICRUST, el cual predice

metagenomas a partir de 16S, de una amplia gama de muestras asociadas a diferentes
ambientes, basa sus predicciones en genomas de referencia publicados (Langille et al.,
2013). PICRUST por medio de algoritmos, predijo el metagenoma del microbioma
humano, al compararlos con los datos metagenómicos, se observó que la predicción tiene
resultados confiables (Langille et al., 2013). Además para el desarrollo de predicciones
de metagenomas del suelo, ecosistemas marinos, entre otros, en donde los genomas de
referencia son muy pocos, PICRUST desarrollo el índice de taxones secuenciado más
cercano (NSTI), el cual permite cuantificar la disponibilidad de representantes de
genomas más cercanos para cada muestra del microbioma, el NSTI suma las distancias
filogenéticas para cada organismos de la tabla OTU a su pariente más cercano con un
genoma de referencia secuenciado (Langille et al., 2013).

3. OBJETIVOS
3.1. General
Evaluar el efecto del uso (ganadería, caña de azúcar y bosque seco tropical), manejo
(convencional vs agroecológico) y la región geográfica del suelo sobre la abundancia de
genes implicados en los ciclos de carbono y nitrógeno en dos regiones del Valle del
Cauca.
3.2. Específicos
● Evaluar la exactitud de la predicción de PICRUSt del metagenoma del suelo
● Evaluar el efecto del uso (sistemas de pastoreo y cultivos de caña de azúcar) y manejo
(convencional vs agroecológico) y la región geográfica del suelo sobre la abundancia
de genes implicados en los ciclos del Nitrógeno y Carbono.

4. MATERIALES Y MÉTODOS
4.1. Diseño Experimental
Las muestras fueron recolectadas en el departamento del Valle del Cauca, en los
municipios de: El Cerrito y Bugalagrande, donde se llevó a cabo dentro y alrededor de la
reserva natural “El Hatico” (N 3º 38’ 39”, W 76º 19’ 12”), y la finca agroecológica “La
Lucerna” (4º 14’ 54”, W 76º 8’ 40”) respectivamente. Estas se encuentran a una distancia
aproximada de 75 kilómetros entre sí (Figura 1).
Fig. 1. Ubicación de las dos fincas del Valle del Cauca.
La abundancia de genes implicados en los ciclos de carbono y nitrógeno fue evaluada en
cinco distintos usos del suelo: sistemas silvopastoril intensivos, pasturas convencionales,
caña de azúcar ecológica, caña de azúcar convencional y bosque seco tropical. En cada
municipio, se contó con 3 parcelas físicamente separadas (3 réplicas) de cada tratamiento
(salvo caña convencional en Bugalagrande, en donde se tomaron dos réplicas).
Adicionalmente se tomaron 3 muestras de suelo de un bosque seco tropical en El Cerrito
como referencia de suelo sin intervención agrícola esto a que existe solo un bosque en El
Cerito (y no hay uno en Bugalagrande), este fue dividido en tres franjas, para así obtener
tres muestras (Sáenz, 2014) (Figura 2).

Se describen a continuación cada uno de los tratamientos
4.1.1. Bosque Seco tropical
El bosque seco tropical de más de 80 años de edad, ocupa 14 ha en la finca “El Hatico”,
ubicada en el Municipio de El Cerrito, en donde la comunidad vegetal que predomina el
paisaje pertenece a: Anacardium escelsum, Ficus sp, Bombacopsis sp., Myrcia
popayamensis y Ceiba pentandra. Este fue dividido en tres franjas. Desde la zona sur
hasta la zona norte, para así obtener tres muestras (Sáenz, 2014).
4.1.2. Pastura con manejo convencional
Las pasturas convencionales muestreadas en los municipios de Cerrito y Bugalagrande
están compuestas de monocultivos de pastos los cuales en época de lluvia reciben
adiciones de fertilizantes del tipo NPK combinado con estiércol y se aplican herbicidas
como Picloran para controlar malezas. Las pasturas se encontraban en descanso del
pastoreo de toro de lidia (El Cerrito) y ganado lechero (Bugalagrande), las muestras
fueron tomadas fuera del dosel de los árboles presentes en el terreno en baja densidad
(Sáenz, 2014).
4.1.3. Sistemas silvopastoril intensivo (manejo agroecológico)
Los sistemas silvopastoriles tenían edades intermedias de establecimiento entre 10-12
años según estudios previos de clasificación (Vallejo et. al., 2010; Vallejo et. al., 2012).
El estrato inferior de los sistemas se compone de las poáceas: Cynodon plectostachyrus y
Paniccum maximum var tanzania y Mombasa. El nivel intermedio es un sotobosque de
arbustos de Leucaena leucocephala (10.000 por hectárea), con una distancia de 1,3 m
entre los surcos, El estrato superior se compone de árboles nativos, que han quedado
dentro de los sistemas, dentro de estos sistemas pastan de 4-7 cabezas /ha de ganado
durante tres o cuatro días, además estos sistemas tenían aproximadamente 25 días de
descanso del pastoreo lechero (Sáenz, 2014; Cubillos et al., 2012; 2016; Calle et al., 2012;
2013).

4.1.4. Caña de azúcar
En El Cerrito y en Bugalagrande se muestrearon cultivos de caña con manejo
convencional y agroecológico. El manejo agronómico ecológico de la caña de azúcar se
diferencia en gran medida del convencional por la reutilización de los residuos del bagazo
como abono para posteriores cultivos, además no se hace quema de residuos cortados.
Las submuestras fueron tomadas en las calles del cultivo; en el caso de la caña con manejo
agroecológico únicamente se tomaron en las calles que no contenían residuos de cortes
anteriores (Sáenz, 2014).
4.2. Amplificación y secuenciación de la región V4 del gen 16S ARNr, control de
calidad y filtración de las secuencias
La amplificación de la región V4 del gen 16S ARNr a las 26 muestras se realizó en un
trabajo anterior (Sáenz, 2014), usando los primers 515F y 806R (Coparaso et al., 2012).
La secuenciación se realizó con la plataforma Miseq de Illumina (2x250pb) siguiendo las
recomendaciones del protocolo de secuenciación de Coparaso et al., 2012.
Las secuencias obtenidas de estas muestras no fueron ensambladas ya que la calidad de
las secuencias del sentido Reverse fue menor de lo esperado, y no permitió ensamblaje,
fue así que solo se utilizaron las secuencias que corresponden al sentido Forward. (Sáenz,
2014).
4.3. Preparación de librerías metagenómicos
La secuenciación de ADN se realizó a 12 muestras, dos (2) réplicas de bosque seco
tropical y dos de sistemas de sistemas silvopastoril y dos de pasturas, una (1) de caña
agroecológica y caña convencional con respecto al El Cerrito, y en Bugalagrande se
tomaron una (1) réplica de todos los sistemas. (Figura 2). El ADN metagenómico se
extrajo utilizando el kit de aislamiento de ADN PowerSoil ™ (Mo Bio, EE. UU.)
siguiendo las instrucciones del fabricante. La concentración de ADN se determinó
mediante el kit de ensayo Quant-iT dsDNA BR y un fluorómetro Qubit (Invitrogen

GmbH, Karlsruhe, Alemania). La verificación de la pureza del ADN se evaluó
espectrofotométricamente (Nano Drop 1000, ThermoFisher Scientific, EE. UU.). En
total, 1 µg de ADN por muestra se fragmentó usando la enzima Alu I durante 5 h
(Invitrogen,) dirigiéndose a fragmentos de ~ 500 pb. Las bibliotecas metagenómicas se
construyeron utilizando el kit de preparación de muestras TruSeq DNA LT (Set A,
Illumina®). Las perlas AMPure XP fueron reemplazadas por el kit de limpieza de PCR
UltraClean ™ (Mo Bio, EE. UU.). La selección del tamaño de los productos de ligadura
se realizó según las instrucciones del fabricante y dirigidas a insertos de 400-500 pb. El
kit de extracción de gel Purelink ™ Quick (Thermo Fisher) se utilizó para extraer y
purificar fragmentos de gel de agarosa. Las bibliotecas finales fueron secuenciadas por
medio de Hiseq Illumina en las instalaciones de ADN de la Universidad de Iowa, EE.
UU. La extracción de ADN y preparación de librerías fueron realizados por Johan S.
Sáenz (datos aún no publicados).
Fig. 2. Diseño de muestreo en El Cerrito y Buga la Grande en el Valle del Cauca para
análisis de PICRUSt, en cuadro rojo diseño de muestreo para análisis metagenomas.

4.4. Procesamiento de las secuencias de secuenciación metagenómica y
secuenciación de la región V4 del gen 16S ARNr
Las secuencias obtenidas por secuenciación metagenómica fueron anotadas en el servidor
de análisis metagenómicos MG-RAST siguiendo el flujo de análisis del programa para
obtener al final una anotación de la abundancia relativa de genes asociadas a funciones
de diferentes subsistemas de las 12 muestras evaluadas
Las secuencias obtenidas por secuenciación de la región V4 del gen 16S ARNr, fueron
agrupados en OTUS con secuencias con un 97% de similitud, y los OTUS con una
frecuencia menor de 50 considerando todas las muestras fueron descartados ya que
pueden representar errores intrínsecos de la secuenciación. Las tablas OTUs fueron re
muestreadas a una profundidad de 10000 secuencias por muestra con la finalidad de
normalizar el número de secuencias para todas las muestras. Estas fueron tomadas como
la entrada de la herramienta bioinformática PICRUSt y analizadas siguiendo el flujo
metodológico del programa: normalización, obtención del índice NSTI (Nearest
Sequenced Taxon Index) y la anotación mediante la base de datos KEGG se realizó una
predicción de las anotaciones funcionales.
4.5. Análisis de los datos
Para evaluar los datos de predicción de genes de PICRUSt se realizó un análisis de
correlación de Spearman usando el paquete estadístico R-studio versión 1.1.463 de los
datos obtenidos por secuenciación metagenómica y los datos predichos por PICRUSt.
Como la profundidad de secuenciación de los datos de PICRUSt y la secuenciación
metagenómica no era la misma se hizo una normalización, en el caso de MG-RAST se
obtiene el número de genes asignado a funciones, mientras que en PICRUSt se obtiene
una predicción del número de funciones, por lo tanto para comparar las dos
aproximaciones de las diferentes muestras, se normalizaron a la librería de menor tamaño
en nuestro caso (1000000), seguido de esto como en PICRUSt se obtuvieron mayor
número de lecturas, lo cual ve reflejado en que el número de anotaciones funcionales fue
mayor (12111) en comparación con las anotaciones funcionales obtenidas por medio de
MG-RAST (6188), se realizó una aleatorización por número de lecturas del total de
anotaciones funcionales obtenidas por PICRUSt para obtener al final el mismo número

de lecturas que en MG-RAST. Además, para observar el ordenamiento de todas las
anotaciones funcionales en todas las muestras tanto por PICRUSt como por secuenciación
metagenómica se realizó un análisis de coordenadas principales (PcoA) en donde se
visualizó el ordenamiento de las diferentes muestras obtenidas de PICRUSt y MG-RAST
mediante el programa STAMP.
Para identificar el efecto del uso del suelo (sistemas de pastoreo y cultivos de caña de
azúcar) y manejo (convencional vs agroecológico) sobre la abundancia de anotaciones
funcionales implicadas en el ciclo del Nitrógeno y el ciclo del Carbono obtenidos del total
de anotaciones funcionales de PICRUSt se realizó un Análisis de Coordenadas
Principales (PcoA) para así visualizar el ordenamiento de los diferentes usos y manejos
del suelo y su relación con las anotaciones funcionales englobadas en el ciclo del N y C,
además para identificar si existían diferencias significativas entre el uso, manejo y región
y las abundancias de anotaciones funcionales involucradas en los ciclos anteriores se
realizó un PERMANOVA de tres factores ya que los datos obtenidos no cumplieron
supuestos de normalidad y homogeneidad. Adicionalmente se realizó un análisis de
medias de cada sistema en el paquete estadístico R studio versión 1.1.463.
5. RESULTADOS
5.1. Comparación de la Predicción de genes obtenidos por PICRUSt y
Secuenciación Metagenómica.
Análisis del índice NSTI (índice de taxones secuenciados más cercanos) de las
anotaciones funcionales obtenidas por medio de PICRUSt
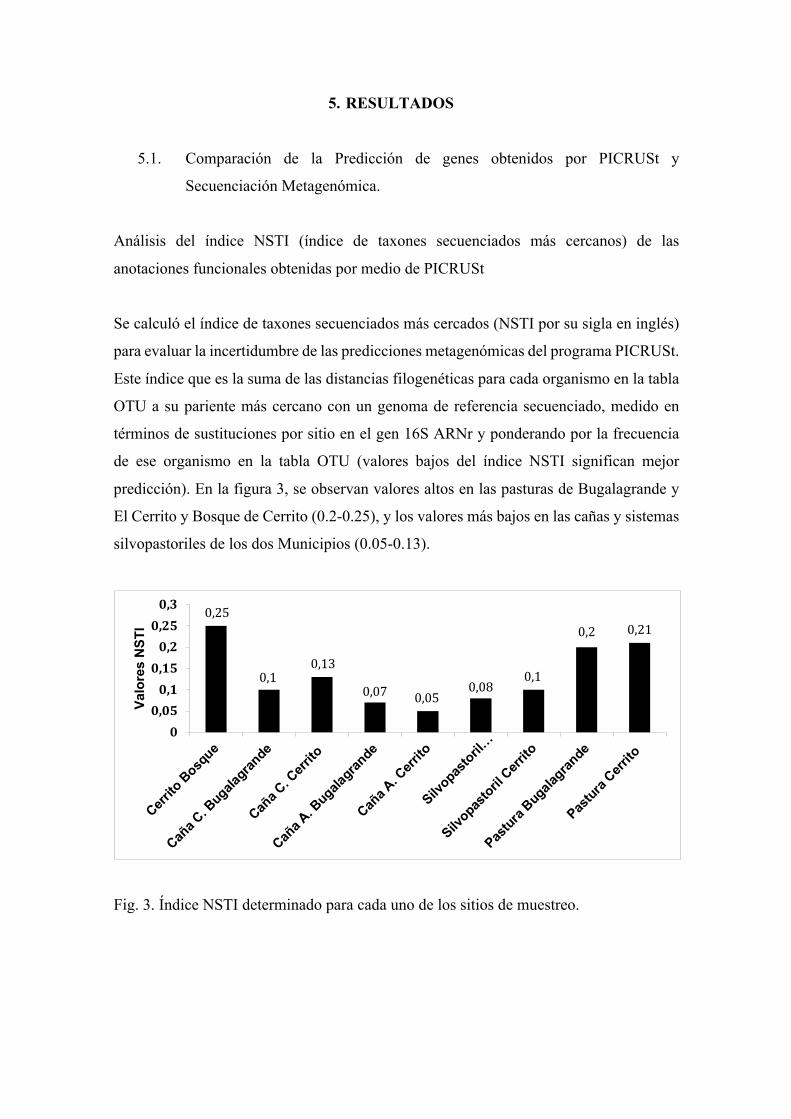
Se calculó el índice de taxones secuenciados más cercados (NSTI por su sigla en inglés)
para evaluar la incertidumbre de las predicciones metagenómicas del programa PICRUSt.
Este índice que es la suma de las distancias filogenéticas para cada organismo en la tabla
OTU a su pariente más cercano con un genoma de referencia secuenciado, medido en
términos de sustituciones por sitio en el gen 16S ARNr y ponderando por la frecuencia
de ese organismo en la tabla OTU (valores bajos del índice NSTI significan mejor
predicción). En la figura 3, se observan valores altos en las pasturas de Bugalagrande y
El Cerrito y Bosque de Cerrito (0.2-0.25), y los valores más bajos en las cañas y sistemas
silvopastoriles de los dos Municipios (0.05-0.13).
Fig. 3. Índice NSTI determinado para cada uno de los sitios de muestreo.
0,25
0,10,13
0,07 0,050,08
0,1
0,2 0,21
00,050,10,150,20,250,3
Cerrito
Bosque
Caña C
. Bugala
grande
Caña C
. Cerr
ito
Caña A
. Bugala
grande
Caña A
. Cerr
ito
Silvopas
toril…
Silvopas
toril Cerr
ito
Pastura
Bugalagran
de
Pastura
Cerrito
Valo
res
NST
I

5.2. Anotaciones funcionales obtenidas en PICRUSt y secuenciación
metagenómica
En base de las secuencias del amplicón del gen 16S ARNr, PICRUSt predijo 12111
anotaciones funcionales específicas (Nivel 3, ejemplo: nitrogenasa), involucradas en 181
anotaciones generales (Nivel 1, ejemplo: metabolismo Nitrógeno). Por otro lado,
mediante secuenciación metagenómica y realizando anotación de dichas secuencias por
MG-RAST, se obtuvieron 28 anotaciones generales (Nivel 1) y 6188 anotaciones
específicas (Nivel 3). Todas las anotaciones se obtuvieron mediante la base de datos de
KEGG. Las anotaciones generales que se encontraron tanto en PICRUSt como por
secuenciación metagenómica se describen en la figura 4. De manera general se observan
valores similares de la abundancia de genes entre sistemas tanto de PICRUSt como de
secuenciación metagenómica, con algunas excepciones como el metabolismo de carbono,
nitrógeno, aminoácidos.
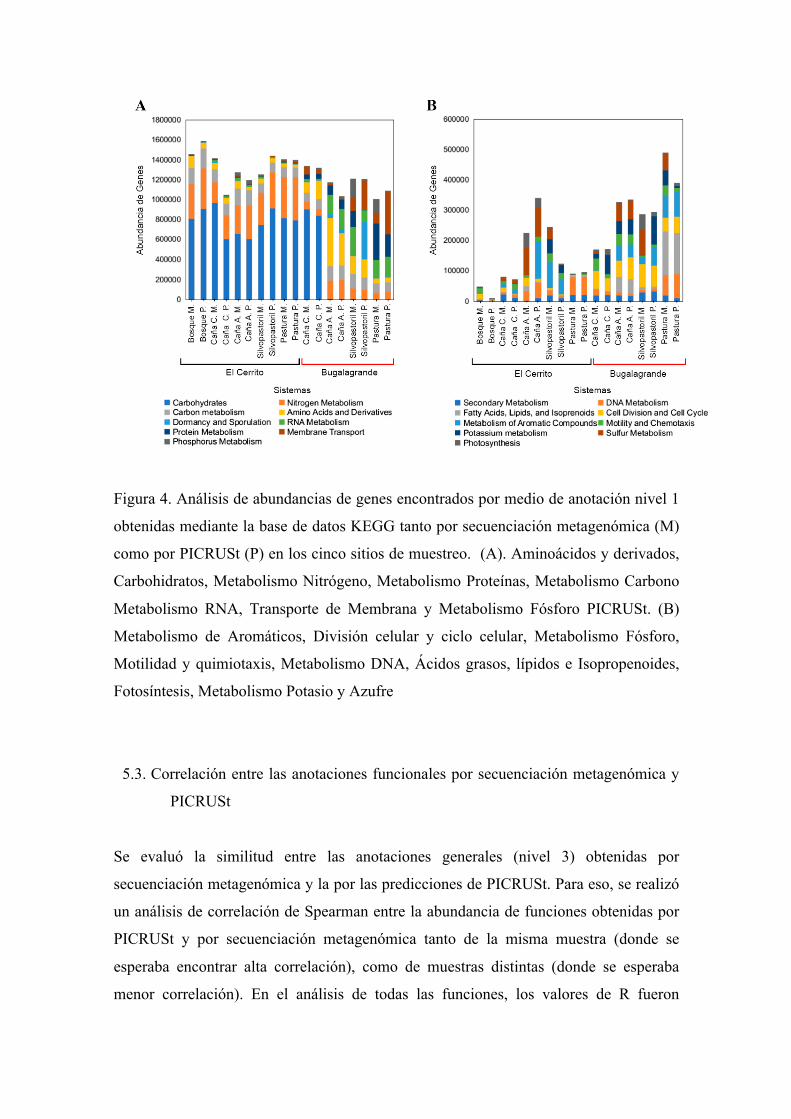
Figura 4. Análisis de abundancias de genes encontrados por medio de anotación nivel 1
obtenidas mediante la base de datos KEGG tanto por secuenciación metagenómica (M)
como por PICRUSt (P) en los cinco sitios de muestreo. (A). Aminoácidos y derivados,
Carbohidratos, Metabolismo Nitrógeno, Metabolismo Proteínas, Metabolismo Carbono
Metabolismo RNA, Transporte de Membrana y Metabolismo Fósforo PICRUSt. (B)
Metabolismo de Aromáticos, División celular y ciclo celular, Metabolismo Fósforo,
Motilidad y quimiotaxis, Metabolismo DNA, Ácidos grasos, lípidos e Isopropenoides,
Fotosíntesis, Metabolismo Potasio y Azufre
5.3. Correlación entre las anotaciones funcionales por secuenciación metagenómica y
PICRUSt
Se evaluó la similitud entre las anotaciones generales (nivel 3) obtenidas por
secuenciación metagenómica y la por las predicciones de PICRUSt. Para eso, se realizó
un análisis de correlación de Spearman entre la abundancia de funciones obtenidas por
PICRUSt y por secuenciación metagenómica tanto de la misma muestra (donde se
esperaba encontrar alta correlación), como de muestras distintas (donde se esperaba
menor correlación). En el análisis de todas las funciones, los valores de R fueron

relativamente bajos, tanto en comparaciones de la misma muestra (R = 0,31 - 0,5) así
como entre diferentes muestras (R = 0.2 – 0.58) (Fig. 5A). Los valores de correlación más
altos en la misma muestra se obtuvieron para las anotaciones funcionales del ciclo de
nitrógeno (R=0,79) a excepción de una réplica de pastura con manejo convencional del
El Cerrito superando los valores entre distintas muestras (Fig. 5B). Los valores R de
correlación de spearman de funciones del ciclo del carbono en la misma muestra
estuvieron entre (0,7-0,8), con 3 excepciones de menor valores (0.53, 0.61 y 0.67). Salvo
estas excepciones, los valores de R de la misma muestra fueron mayores que los valores
entre muestras distintas. (Fig. 5C). Por último, los valores de spearman obtenidos para el
ciclo del Fósforo fueron los más bajos entre 0,282 y 0,609 (Fig. 5D). Por este motivo se
decidió no analizar las funciones englobadas en el ciclo del Fósforo sino solo las
funcionales englobadas en el ciclo del Carbono y Nitrógeno obtenidas por PICRUSt.
Además, se realizó un diagrama de barras donde se muestran las abundancias de genes
agrupados en los ciclos de carbono y nitrógeno obtenidos mediante secuenciación
metagenómica y PICRUSt, esto para evidenciar la veracidad de los datos obtenidos por
medio de las metodologías. En general, las abundancias de genes del ciclo de carbono y
del nitrógeno fueron similares entre PICRUSt y secuenciación metagenómica (Fig. 6 y
7), con excepción en las funciones agrupadas en la nitrificación en donde se obtuvieron
valores mayores en los sistemas silvopastoril y pastura de El Cerrito por medio de
PICRUSt y menores por secuenciación metagenómica y valores menores en caña
agroecológica de Bugalagrande en PICRUSt y mayores en secuenciación metagenómica
(Fig. 6C)
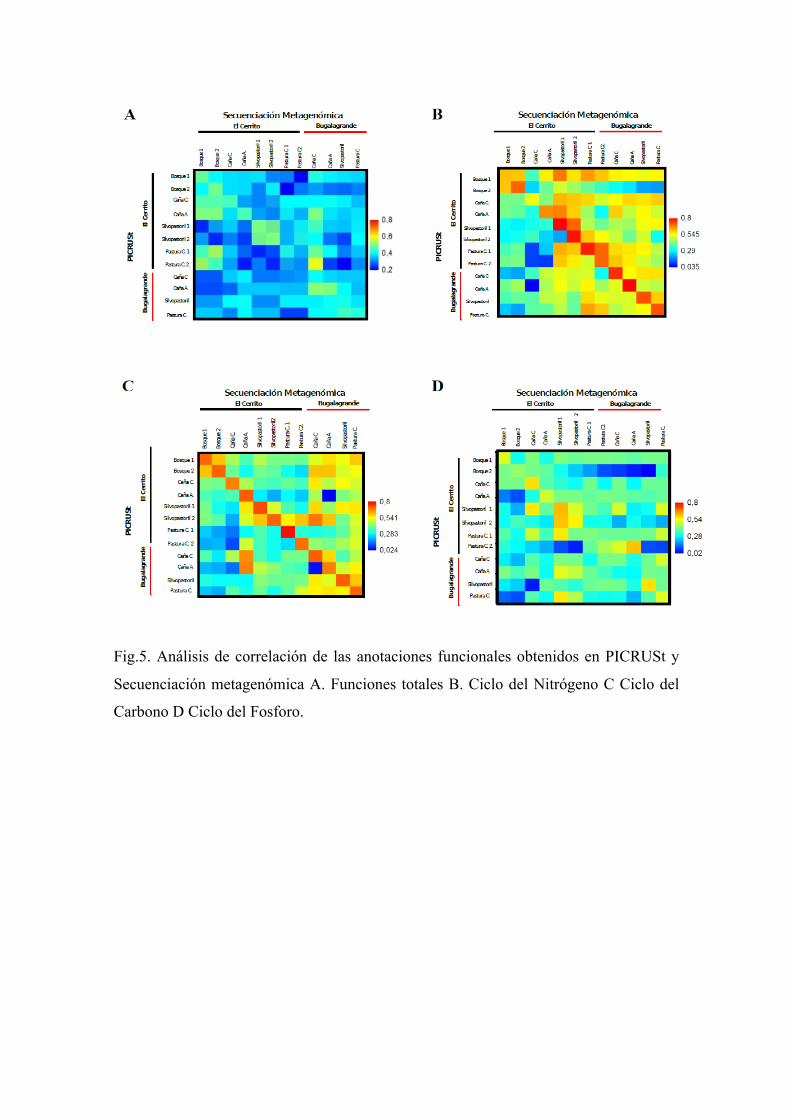
Fig.5. Análisis de correlación de las anotaciones funcionales obtenidos en PICRUSt y
Secuenciación metagenómica A. Funciones totales B. Ciclo del Nitrógeno C Ciclo del
Carbono D Ciclo del Fosforo.
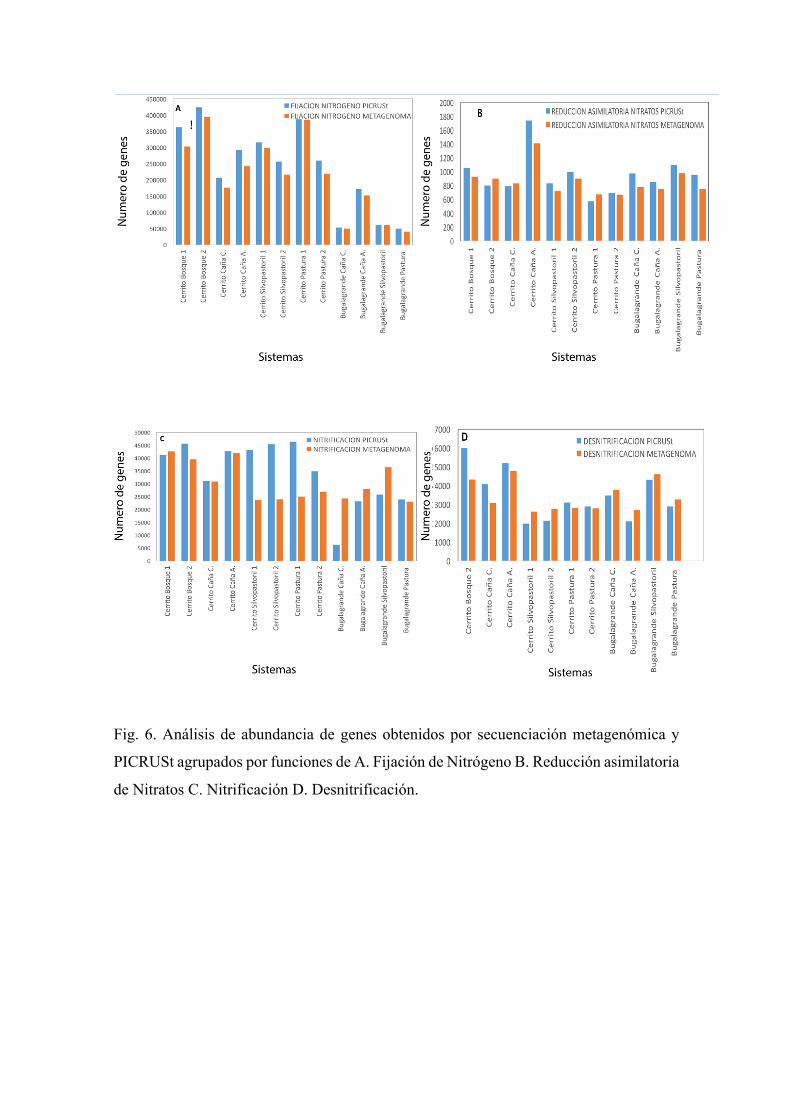
Fig. 6. Análisis de abundancia de genes obtenidos por secuenciación metagenómica y
PICRUSt agrupados por funciones de A. Fijación de Nitrógeno B. Reducción asimilatoria
de Nitratos C. Nitrificación D. Desnitrificación.
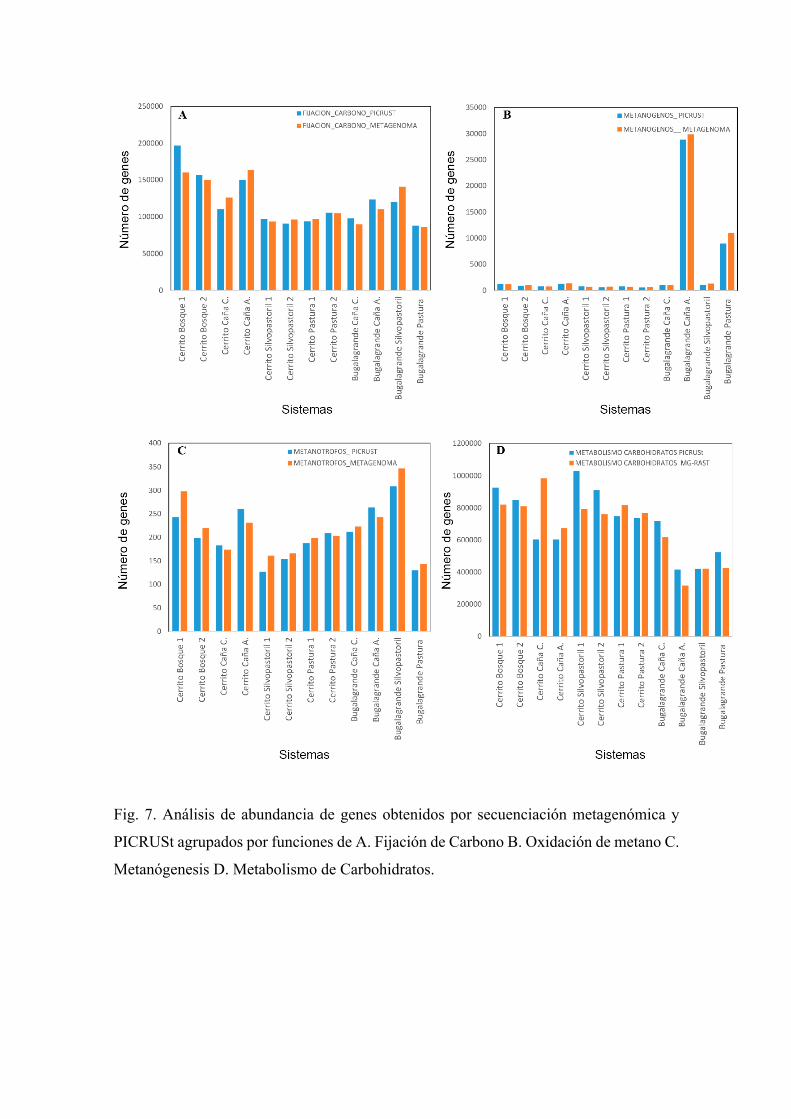
Fig. 7. Análisis de abundancia de genes obtenidos por secuenciación metagenómica y
PICRUSt agrupados por funciones de A. Fijación de Carbono B. Oxidación de metano C.
Metanógenesis D. Metabolismo de Carbohidratos.
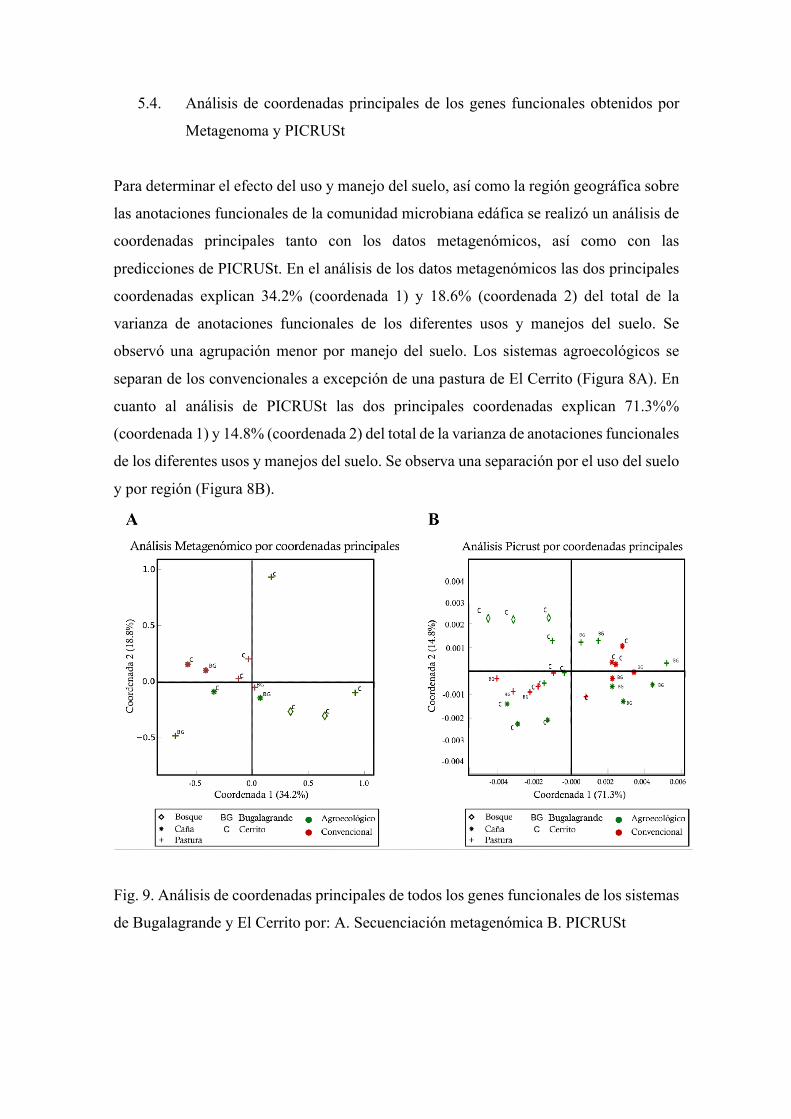
5.4. Análisis de coordenadas principales de los genes funcionales obtenidos por
Metagenoma y PICRUSt
Para determinar el efecto del uso y manejo del suelo, así como la región geográfica sobre
las anotaciones funcionales de la comunidad microbiana edáfica se realizó un análisis de
coordenadas principales tanto con los datos metagenómicos, así como con las
predicciones de PICRUSt. En el análisis de los datos metagenómicos las dos principales
coordenadas explican 34.2% (coordenada 1) y 18.6% (coordenada 2) del total de la
varianza de anotaciones funcionales de los diferentes usos y manejos del suelo. Se
observó una agrupación menor por manejo del suelo. Los sistemas agroecológicos se
separan de los convencionales a excepción de una pastura de El Cerrito (Figura 8A). En
cuanto al análisis de PICRUSt las dos principales coordenadas explican 71.3%%
(coordenada 1) y 14.8% (coordenada 2) del total de la varianza de anotaciones funcionales
de los diferentes usos y manejos del suelo. Se observa una separación por el uso del suelo
y por región (Figura 8B).
Fig. 9. Análisis de coordenadas principales de todos los genes funcionales de los sistemas
de Bugalagrande y El Cerrito por: A. Secuenciación metagenómica B. PICRUSt

5.5. Análisis de coordenadas principales de las anotaciones funcionales obtenidas
por PICRUSt involucrados en el ciclo del Carbono y Nitrógeno
En el ciclo del Carbono se observan tres grupos: el primero es de suelo del Bosque, el
segundo es una leve separación por manejo del suelo y el tercero es el que contiene los
sistemas silvopastoriles de Bugalagrande con una réplica de pastura convencional de
Bugalagrande y una de caña con manejo convencional y de El Cerrito. En cuanto al ciclo
del Nitrógeno se observa una separación representativa por región, están separados los
sistemas de Bugalagrande y El Cerrito, además se observa separación leve de subgrupos
en la región de EL Cerrito por uso del suelo y en Bugalagrande por manejo del suelo y
una separación de Bosque.
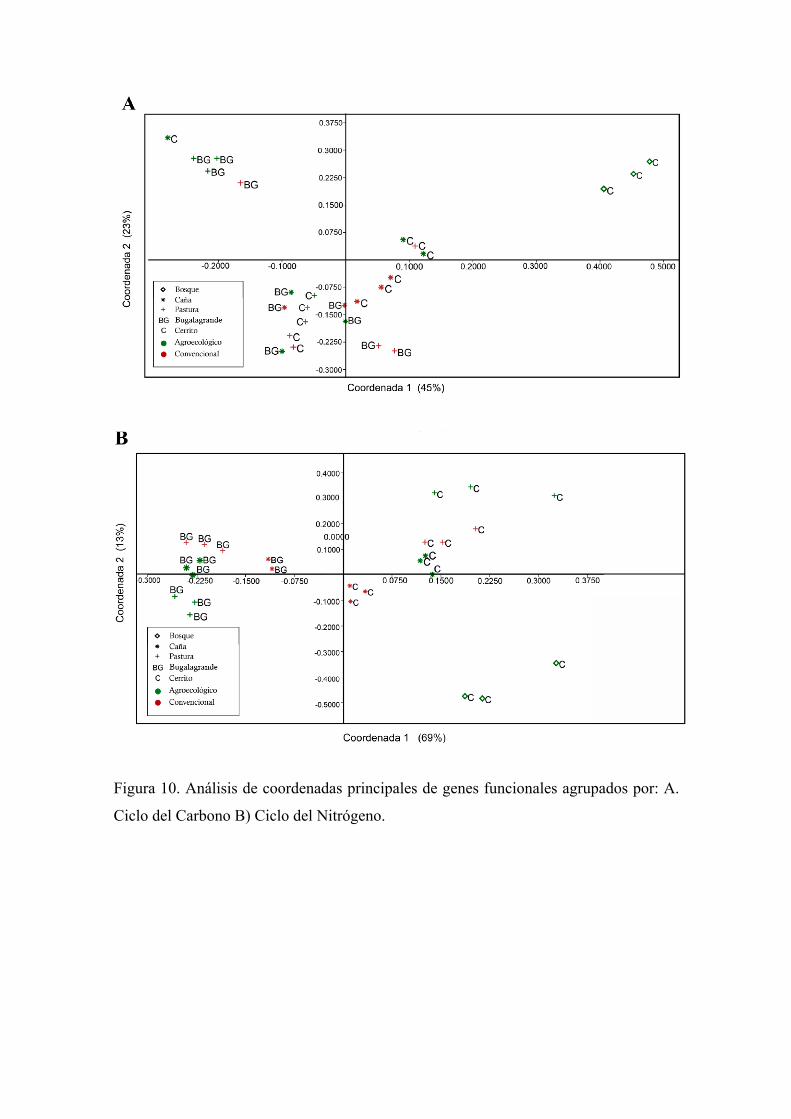
Figura 10. Análisis de coordenadas principales de genes funcionales agrupados por: A.
Ciclo del Carbono B) Ciclo del Nitrógeno.

5.6. Análisis de PERMANOVA de los genes funcionales correspondientes al
ciclo del Carbono.
En los resultados de predicción de anotaciones funcionales obtenidas por PICRUSt se
encontraron diferentes funciones biológicas implicadas en el ciclo del Carbono como: la
metanogénesis, oxidación de metano, fijación de carbono y metabolismo de carbohidratos
(Anexos 1-2-3).
Para evaluar el efecto del uso y manejo del suelo, y región geográfica sobre las funciones
biológicas edáficas encontradas en el ciclo del Carbono se realizó un análisis de
PERMANOVA factorial de interacción entre las tres variables independientes (Manejo,
Uso y Región) para cada una de las diferentes funciones biológicas encontradas. La
abundancia de genes relacionados con metanogénesis es mayor en sistemas con manejo
convencional como Caña y Pasturas de Bugalagrande y bajos para los mismos sistemas
en El Cerrito y valores bajos para todos los sistemas con manejo agroecológico
independientemente de las regiones, concordando con esto se encontró diferencia
significativa entre la interacción del manejo y la región con un Valor P significativos
0,016 y 0,005 (Figura 11C).
En los resultados de metabolismo de carbohidratos se encontraron valores altos para los
sistemas de El Cerrito incluyendo Bosque y bajos para todos los sistemas de
Bugalagrande, concordando con los resultados de diferencia estadística de la región con
valor P de 0,001 (Figura 11B). En cuanto la oxidación de metano (metanotrofos) se
observan valores bajos en las cañas Agroecológicas tanto de Bugalagrande y Cerrito y
altos en las cañas convencionales, en cuanto a los valores de los sistemas de pasturas y
silvopastoril los resultados son muy variables, en los análisis estadísticos no se
encontraron diferencias significativas (Figura 11D). En la fijación de carbono el valor
más alto se muestra en Bosque seguido por valores altos en los sistemas de uso de cañas
y bajos en los de ganadería (pastura y silvopastoril) independiente de la región y el manejo
del suelo, validando estos resultados se encontró diferencia significativa en el uso del
suelo con un valor P significativo 0,0186 (Figura 11A).
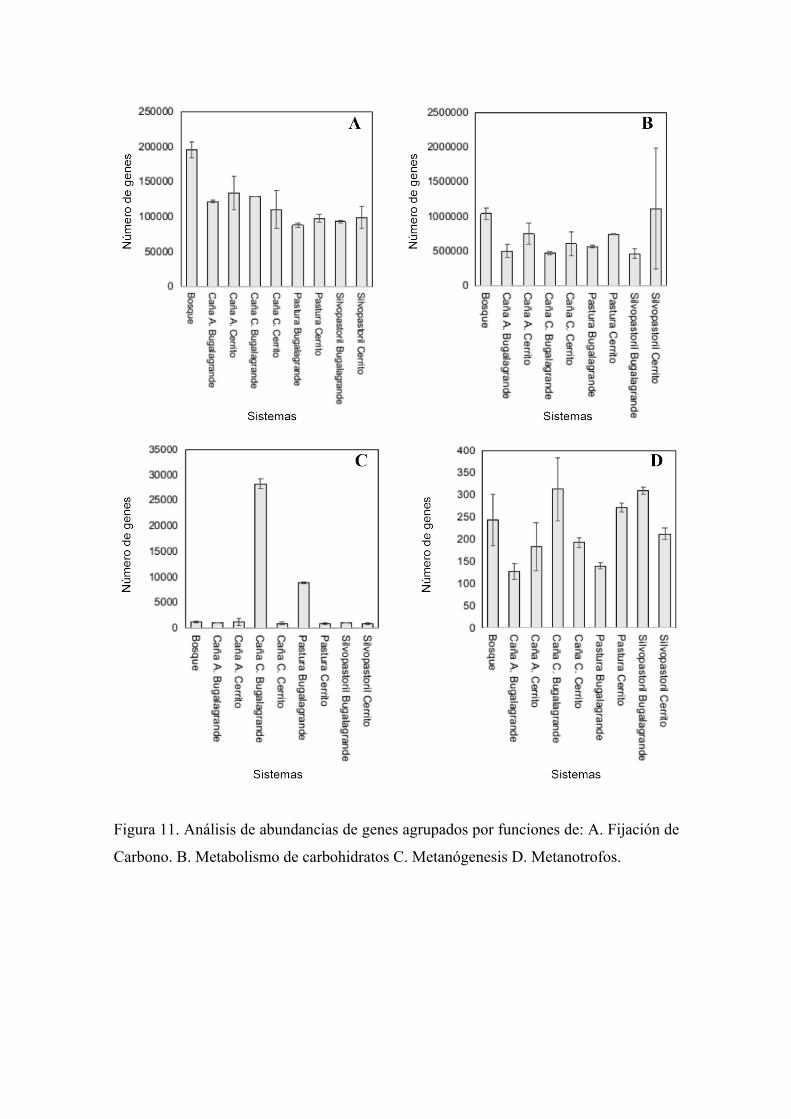
Figura 11. Análisis de abundancias de genes agrupados por funciones de: A. Fijación de
Carbono. B. Metabolismo de carbohidratos C. Metanógenesis D. Metanotrofos.

5.7. Análisis de las anotaciones funcionales correspondientes al ciclo del
Nitrógeno.
Tomando los resultados obtenidos en PICRUSt se dividió el ciclo del Nitrógeno en cuatro
partes con base en la anotación de KEGG (desnitrificación, fijación de nitrógeno,
nitrificación y reducción de nitrato vía asimilatoria) para evaluar si existen diferencia
significativa del uso y manejo del suelo sobre las funciones relacionadas con el ciclo del
Nitrógeno se realizó un análisis de PERMANOVA de interacción entre las tres variables
independientes (Manejo-Uso-Región) para cada una de las diferentes funciones
biológicas.
En la fijación de Nitrógeno el valor mayor se encontró en Bosque seguido por valores
altos de las medias en todos los sistemas de El Cerrito y bajos en Bugalagrande,
concordando con la diferencia altamente significativa (P = 2e-16) que se encontró en
cuanto a la región (Figura 12A). Los resultados de reducción asimilatoria de nitratos fue
más alta en bosque seguidos por manejo agroecológico y bajos en manejo convencional
validando los resultados de diferencia significativa del manejo del suelo valor P 0,0034
(Figura 12D). En la nitrificación se encontró valores bajos en suelos de uso ganadería
(pasturas y silvopastoril) y altos en cañas y bosque lo cual afirma los resultados de los
análisis estadísticos de diferencia significativa del uso del suelo (P=0,012) (Figura 12C).
Por último, la desnitrificación de manera general los resultados fueron muy variables los
valores más altos fueron en caña agroecológica de El Cerrito y Silvopastoril de Buga la
grande, los valores más bajos fueron en caña convencional, pastura de El Cerrito y
bosque, concordando con los resultados de análisis estadísticos donde no se encontró
diferencia Figura 12B).
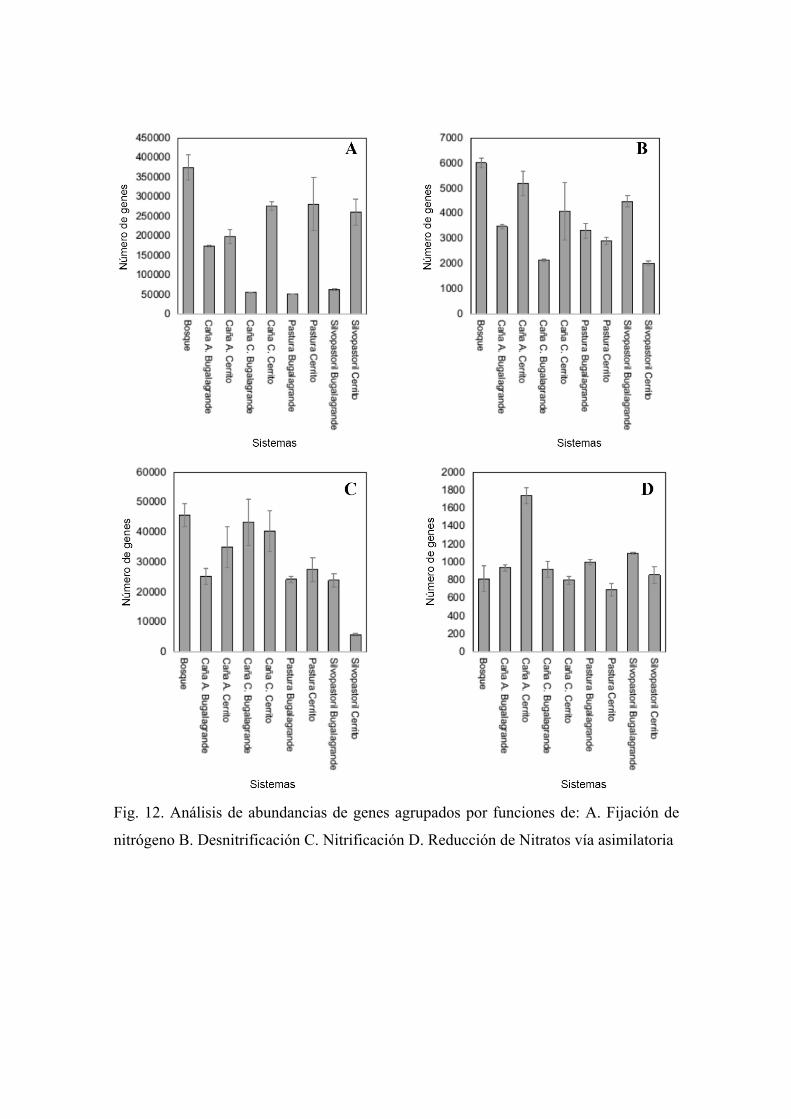
Fig. 12. Análisis de abundancias de genes agrupados por funciones de: A. Fijación de
nitrógeno B. Desnitrificación C. Nitrificación D. Reducción de Nitratos vía asimilatoria

6. DISCUSIÓN
6.1. Comparación Predicción de genes obtenidos por PICRUSt y Secuenciación
Metagenómica.
En este estudio se analizaron anotaciones funcionales correspondientes a cinco sistemas:
caña agroecológica, caña convencional, silvopastoril, pastura y bosque de dos regiones:
El Cerrito y Bugalagrande. Las anotaciones obtenidas fueron asignadas funcionalmente
mediante la base de datos KEGG (Kyoto Encyclopedia of Genes and Genomes), se
obtuvieron por secuenciación metagenómica en solo 12 muestras y en PICRUSt se
realizaron en todas las muestras por triplicado. En secuenciación metagenómica se
encontraron 6188 anotaciones funcionales y en PICRUSt el cual es un programa
bioinformático que permite predecir genes funcionales a partir de la secuencia del gen
16S ARNr se obtuvieron 12111 anotaciones funcionales. Este diferencia se puede atribuir
a la mayor profundidad que permite la secuenciación de un gen marcador (16S ARNr) en
comparación de la secuenciación de toda el metagenoma, lo cual muestra una ventaja del
programa, permitiendo un incremento del casi el 100% de las anotaciones funcionales
Además como PICRUSt es un programa de predicción se evaluó el índice NSTI el cual
representa la suma de las distancias filogenéticas de cada organismo en la tabla OTU a su
pariente más cercano con un genoma de referencia secuenciado, medido en términos de
sustituciones por sitio en el gen 16S ARNr y ponderado por las frecuencias de ese
organismo en la tabla OTU (Langille et al., 2013). En nuestro estudio se encontraron
valores entre 0.25 y 0.01 que corresponden a rangos similares encontrados en otros
estudios como el de Langille et al., 2013 donde evaluaron 12 muestras de suelo
encontrando valores similares a los de este estudio, concluyendo que los datos se
encuentran en una predicción media según los rangos de Langille et al., 2013. En otros
estudios donde se han analizado muestras de suelo, pero en cultivos de enriquecimiento
se han obtenido valores entre (0.18-0.03) donde 0.18 corresponde a la muestra original
de suelo y bajó a 0.03 después de que la muestra fue enriquecida., Debido a lo anterior en
donde el programa PICRUSt se ha usado muy pocas veces en muestras de suelo y que los
resultados de este estudio estuvieron dentro de un rango medio de predicción se decidió
hacer un análisis adicional de correlación de Spearman entre los datos de secuenciación
metagenómica y PICRUSt, análisis que se ha realizado en muy pocos estudios de suelo
donde se ha usado PICRUSt.

Las anotaciones funcionales de PICRUST y MG-RAST en su mayoría se encuentran
relacionadas con el mantenimiento de la función y la estructura celular y, por lo tanto, las
realizan todas las especies de la comunidad microbiana (Hariharan, 2015). De especial
interés, sin embargo, son ciertas funciones relacionadas con la salud y productividad del
suelo como: el metabolismo del nitrógeno, carbono y fósforo. En cuanto a la abundancia
de anotaciones funcionales relacionadas con el metabolismo del nitrógeno y carbono
fueron altos en bosques y pasturas tanto en la predicción de PICRUSt como en la
secuenciación metagenómica, lo cual corresponde con estudios anteriores en donde el
nitrógeno y carbono suelen ser altos en los sistemas de praderas y bosques (Ratnayake et
al., 2011)
El análisis comparativo que agrupan todas las funciones generales obtenidas por
PICRUSt y secuenciación metagenómica estuvieron entre (R=0.31 - 0,5), al compararlo
con otros estudios como el de (Millares et al., 2015) donde se correlacionan estos dos
métodos con muestras de microbiomas humanos los resultados son (0,7-0,8), esto debido
a que existen más genomas de referencia (Figura 5). Los ciclos de Carbono (0,56-0,8) y
Nitrógeno (0,56-0,8) presentaron valores más altos en comparación con los valores
obtenidos de la correlación de todas las anotaciones funcionales. Además, los resultados
arrojados en los diagramas de barras de las anotaciones funcionales agrupados en los
mismos ciclos son muy similares en PICRUSt y secuenciación metagenómica
permitiendo sugerir que los datos obtenidos por predicción de PICRUSt de estos dos
ciclos son confiables para ser usados en este estudio (Figuras 11 y 12). En comparación
con los resultados del ciclo del fosforo donde la correlación fue muy baja y por esto se
decidió no usarlos en este estudio. Con estos resultados se podría plantear que el programa
predice mejores grupos de funciones de ciclos biogeoquímicos que se han estudiado más
a fondo y por lo tanto van a existir más enzimas predichas en las bases de datos en donde
PICRUSt se basa para hacer las predicciones.
Debido a lo anterior se decidió usar solo los genes funcionales agrupados en los ciclos
del carbono y del nitrógeno obtenidos por PICRUSt y además por la falta de réplicas en
secuenciación metagenómica no se tomaron los datos para los siguientes análisis.

En conclusión se puede inferir que el programa PICRUSt proporciona una buena
clasificación de la asignación funcional cuando se cuenta con una gran base de datos de
genomas de referencia, ya que en los estudios del microbioma humano el programa
realiza una buena predicción, caso contrario en nuestro caso en donde los genomas de
referencia en suelos son muy pocos, por esto otros tipos de análisis estadísticos como los
de correlación permiten verificar la confiabilidad de los datos, siendo este el primer
estudio en muestras de suelo en donde se hace este tipo de análisis para poder confirmar
la veracidad de la predicción. Sin embargo, las nuevas tecnologías y los avances en
herramientas bioinformáticas, como las lecturas de secuenciación más largas, las
evoluciones en las bases de datos de anotación, las herramientas para predecir mejor las
estructuras de genes y operones, y las herramientas que interrogan los datos a nivel de
polimorfismo de un solo nucleótido serán esenciales para proporcionar anotaciones
funcionales más detalladas y precisas (Xue et al., 2014).
La abundancia y presencia de anotaciones funcionales involucradas en el ciclo del
nitrógeno está relacionado con el estudio preliminar de Sáenz, 2014 donde determinó la
abundancia y diversidad de microorganismos y encontró géneros como: Bradyrhizobium
y Heliobacterium, los cuales, aunque no son los únicos que realizan fijación de nitrógeno
en el estudio de Sáenz 2014 se encontraron dentro de los más abundantes. contribuyentes
en la fijación de nitrógeno y expresan genes nif (nifK, nifH y nifD) que codifican la
proteína nitrogenasa determinante en la salud y productividad del suelo (Jones &
Bradford 2001; Hariharan, 2015) otros como Pseudomonas y Bacillus los cuales fueron
abundantes en el estudio de Sáenz, 2014 que están involucrados en la desnitrificación
conversión de nitrato en nitrógeno a partir de la expresión de genes como: norB, norC y
nirK, los cuales son usados como marcadores genéticos para las principales enzimas de
la desnitrificación como la nitrato reductasa, nitrito reductasa y óxido nitroso (Wertz et
al., 2012), y por último las especies Nitrospira y Nitrococcus encontradas en el estudio
anterior de Sáenz, 2014 y en secuenciación metagenómica productores del gen amo
principal catalizador de la oxidación de amoniaco (nitrificación) cuentan con el potencial
de oxidar tanto los compuestos de nitrógeno orgánicos como inorgánicos (Hariharan,
2015; 2017; Francis et al., 2003; Dionisi et al., 2002; Purkhold et al., 2000; Rotthauwe et
al., 1997). Así mismo la abundancia de genes relacionados con el metabolismo del
carbono también correlacionado con el estudio preliminar de Sáenz, 2014 en donde
encontró Proteobacteria y Acidobacteria los cuales son los más abundantes y se

encuentran asociados a la fijación de carbono (Hariharan, 2015). Otra parte importante
involucrada en el ciclo del carbono es el metabolismo del metano estrechamente
relacionado con los gases de efecto invernadero y el cambio climático, del cual se
encontraron anotaciones funcionales en este estudio correspondiente a estudios
preliminares de (Sáenz, 2014) en donde encontraron géneros de metanogénicas y
metanótrofas como Methanococcus y Methanobacterium. El proceso de oxidación del
metano convierte el metano en formaldehido y se lleva a cabo por metanotrofos. Los
metanotrofos a menudo se clasifican como Tipo I (generalmente Gammaproteobacteria,
utilizan la vía de RuMP para la asimilación del carbono) y Tipo II (Alfaproteobacteria,
usan la serina para la misma función), dos grupos encontrados en el estudio preliminar de
Sáenz, 2014. La metanol deshidrogenasa es una enzima importante involucrada en la
oxidación del metano, y las subunidades 1 y 2 de la metanol deshidrogenasa asociada al
citocromo c fueron predichas por PICRUSt y secuenciación metagenómica, mdh1 y mdh2
está conectada a familia metanotrofía como Bradyrhizobiaceae (Fitriyanto et al., 2011).
Otro conjunto importante de genes involucrados en la oxidación del metano son los genes
pmo, como: pmoA, pmoB y pmoC estos genes codifican para las subunidades A, B y C
de la metano monooxigenasa asignados a Methanococcus y otros miembros como
Methanobacterium.. Todas las anteriores familias encontradas en el estudio preliminar de
Sáenz, 2014. La metanogénesis es el proceso de formación de metano. El gen mtrH (que
expresa la subunidad H de la tetrahidrometanopterina S-metiltransferasa encontrada en
este estudio), esta enzima ayuda en la producción de metano a partir de acetato y está
vinculada exclusivamente a Methylococcus (Pester et al., 2010).
6.2. Efecto del uso del suelo (sistemas de pastoreo y cultivos de caña de azúcar) y
manejo (convencional vs agroecológico) y la región sobre la abundancia de
genes implicados en el ciclo del Nitrógeno
El nitrógeno es uno de los principales factores que limita la producción de los cultivos,
es un componente importante de las proteínas y los aminoácidos y por lo tanto es un
nutriente esencial para todos los organismos. El nitrógeno existe en varios estados de
oxidación y formas químicas, los cambios en las diferentes transformaciones suelen ser
catalizadas por microorganismos. La identificación de los genes involucrados en este

ciclo y su abundancia en cualquier muestra de suelo es clave para estimar el potencial
funcional de los microorganismos en los suelos.
La actividad de fijación de nitrógeno (conversión de di nitrógeno atmosférico en
amoníaco). La fijación biológica de nitrógeno es la fuente natural más grande de
nitrógeno para la mayoría de los ecosistemas terrestres, en este proceso los
microorganismos diazotrofos simbióticos y de vida libre reducen el gas di nitrógeno
atmosférico a formas reactivas y biológicamente disponibles (Keeler et al., 2009). La
nitrogenasa está muy extendida en bacterias y arqueas, y les proporciona una ventaja
competitiva en entornos que carecen de nitrógeno disponible. Existen tres tipos de
nitrogenasas: hierro-hierro (FeFe), vanadio-hierro (VFe) y molibdeno-hierro (MoFe)
(Lloyd et al., 2009; Soon & Arshad, 2005). La fijación de nitrógeno suele ser más alta
en los Bosques ya que estos se caracterizan por poseer zonas hotpots que van la van a
incrementar, lo cual se obtuvo en este estudio donde la fijación de nitrógeno fue más alta
en el Bosque esto debido posiblemente a que microorganismos como los diazótrofos de
vida libre que son la forma más dominante de bacterias fijadoras de nitrógeno suelen estar
en alta abundancia en los bosques (Hoorman et al., 2010). Otro factor que puede influir
en el aumento de fijación de nitrógeno en el bosque es la no presencia de fertilizantes
agrícolas los cuales pueden disminuir la fijación de nitrógeno (Barberán et al., 2012; Soon
& Arshad, 2005). Por último, se observó un efecto significativo de la región sobre la
fijación de nitrógeno esto debido posiblemente a la alteración de factores
medioambientales como: la deficiencia o toxicidad de fósforo y la acidez del suelo, donde
en el estudio anterior de Sáenz, 2014 se observó que el fósforo se encuentra en menor
cantidad en Bugalagrande y el pH es bajo en la misma zona.
El proceso de reducción de nitrato puede ser asimilatorio o desasimilatorio. La
desasimilación solo se lleva a cabo en ambientes anaeróbicos. El proceso de asimilación
fue más común en estos sistemas. El manejo tuvo un efecto sobre la actividad de la
reducción de nitrato asimilatoria, el cual fue más alto en todos los sistemas agroecológicos
y el Bosque concordando con estudios anteriores como Selbie et al. 2015; GBowles et al.,
2014 quienes sugieren que el uso de fertilizantes tipo NPK usados en la actividad agrícola
aumentan el contenido de amonio lo cual disminuye la reducción asimilatoria de nitrato,
lo cual podría aumentar el nitrato y este no es del todo asimilable por los microorganismos

provocando así una lixiviación del nitrato (Piotrowska & Wilczewski, 2014), además
concuerda con lo reportado por Sáenz, 2014 el cual encontró que los fertilizantes usados
en estos sistemas aumentan el contenido de amonio.
El proceso de nitrificación (amoníaco a nitrito y posteriormente el nitrito se oxida a
nitrato). Este proceso clave juega un papel importante en la regulación de la
disponibilidad de nitrógeno y la pérdida de nitrógeno en los ecosistemas terrestres puede
causar la lixiviación de nitrato a las aguas subterráneas y producir N2O directamente. La
nitrificación en los suelos se divide en nitrificación autótrofa y heterótrofa. La primera es
llevada a cabo por bacterias quimio autotróficas que oxidan el amonio y la segunda por
ciertas bacterias con el potencial de oxidar tanto compuestos orgánicos como inorgánicos
(Kuypers et al., 2018; Hariharan, 2015). La nitrificación desempeña un papel clave en
determinar cuánto y qué formas de Nitrógeno inorgánico del suelo están disponibles para
las plantas. El aumento del proceso de nitrificación aumenta las pérdidas de Nitrógeno de
los ecosistemas a través de la lixiviación de nitratos y la emisión de óxido de nitrógeno
(Assémien et al., 2017). Se encontraron valores menores en los sistemas silvopastoril y
pasturas y bajos en sistemas de cañas agroecológicas y convencionales resultados
contradictorios se encontraron en el estudio anterior de Cubillos et al., 2016 en donde se
encuentra que la nitrificación es más alta en los sistemas de pasturas. Los altos valores
presentados en las cañas puede deberse correlacionarse con los valores altos de amoniaco
en estos sistemas resultados reportados por Sáenz, 2014, además la aplicación de
fertilizantes inorgánicos y los altos niveles de amonio aumentan la producción de
nitrificación (Groenigen et al., 2005; Pajares et al., 2016; Paungfoo-Lonhienne et al.,
2017; Coskun et al., 2017).
La desnitrificación es un proceso de respiración esencialmente anaeróbico, en el que el
nitrógeno principalmente en forma de NO3- el cual se utiliza como aceptor de electrones
en la oxidación de carbono (Hofstra & Bouwman, 2005; Clough et al., 2017). En la
desnitrificación se pueden generar “hots spots” los cuales son puntos de actividad
microbiana que se producen como resultado de la heterogeneidad de las condiciones del
suelo y posiblemente pueden determinar los estados de oxigenación determinando así el
proceso de desnitrificación, debido a esto es probable no encontrar efecto ni de la región,

manejo y uso del suelo, lo cual se obtuvo en este estudio en donde no se encontró
diferencia significativa (Hofstra & Bouwman, 2005), esto concuerda con este estudio en
donde las tres variables (región, manejo y uso) influyeron en la desnitrificación. Los tipos
de cultivos y abonos, los tipos de fertilizantes, la tasa de aplicación de nitrógeno, método
y tiempo de aplicación influye en el período de disponibilidad de nitrógeno y la forma en
el que el nitrógeno se vuelve disponible es determinante en la desnitrificación (Hofstra &
Bouwman, 2005). La entrada de nitrógeno a partir de residuos orgánicos de los cultivos
varía entre tipos de cultivos y como resultado de esta gestión los factores relacionados
van a influir significativamente en la desnitrificación (Hofstra & Bouwman, 2005; Selbie
et al., 2015).
6.3. Efecto del uso del suelo (sistemas de pastoreo y cultivos de caña de azúcar) y
manejo (convencional vs agroecológico) sobre la abundancia de genes
implicadas en el ciclo del Carbono.
El carbono es el mayor componente en la célula y el elemento químico clave en los
compuestos de naturaleza orgánica, elemento que circula entre los océanos, la atmósfera,
el suelo y el subsuelo, instancias que conforman los depósitos, reservorios o almacenes
de carbono. El paso entre los depósitos ocurre mediante procesos de orden químico, físico
y biológico. El intercambio de carbono entre el reservorio terrestre y el atmosférico es el
resultado de procesos naturales como la fotosíntesis y la respiración, así como de la
emisión de gases de origen antrópico. Los microorganismos del suelo se consideran
actualmente como una de las estrategias más prometedoras para mejorar la disponibilidad
y el almacenamiento de carbono en el suelo, por lo que es importante evaluar los genes
que intervienen en el ciclo del carbono (Hariharan, 2015).
El ciclo del carbono se inicia con la fijación del dióxido de carbono atmosférico, la cual
se ha atribuido principalmente a microorganismos quimiolitotróficos que sintetizan el
carbono atmosférico dióxido de carbono en material orgánico. Los microorganismos del
suelo esencialmente transfieren entre compartimentos ambientales para cumplir su
función fundamental: la supervivencia a través de la reproducción. En este estudio se
encontró un efecto del uso del suelo, esto debido posiblemente a que la disponibilidad de
otros elementos clave como el nitrógeno y fósforo son esenciales para la fijación de
carbono, concordando con resultados donde en los sistemas de pastoreo y bosque se

encontraron valores más altos de fósforo total y nitrógeno total que en los sistemas de
cañas reportado en el estudio de Sáenz, 2014 (Orellana et al. 2012; Sperling et al., 2017;
Ratnayake, Seneviratne & Kulasooriya, 2011).
El intercambio de metano en el suelo con la atmósfera está regulado por arqueas
productoras de metano es decir metanógenos y bacterias oxidantes de metano (MOB), es
decir metanotrofos. Los metanotrofos son bacterias Gram negativas que utilizan el
metano como fuente de carbono y energía y pueden oxidar hasta el 90% del metano
producido por los metanógenos (Zhang et al., 2018). La metanogénesis fue alta en caña
convencional y pastura debido posiblemente a la adición de nitrógeno inorgánico en el
suelo, lo cual aumenta la deposición de nitrógeno y la escorrentía lo cual a su vez aumenta
producción de metano y disminuye la oxidación de metano la cual fue baja en estos
mismos sistemas (Zou et al., 2005; Liu et al., 2015), además la adición de estiércol puede
aumentar las emisiones de metano, aumentando a su vez el nitrógeno y fósforo en suelo
que contribuyen al aumento de la metanogénesis (Cai et al., 2016).

7. CONCLUSIONES
● La exactitud de la predicción de PICRUSt varía en distintos genes funcionales.
Por ejemplo, la predicción de genes asociados a los ciclos de nitrógeno y carbono
fue relativamente exacta, mientras que la exactitud de la predicción de los genes
agrupados en ciclo del fósforo fue relativamente baja.
● Las abundancias de los diferentes genes asociados con el ciclo de nitrógeno están
afectadas principalmente por la región.
● Las abundancias de los diferentes genes asociados con el ciclo de carbono están
determinados por sistemas intervenidos vs. el bosque.
● Diferentes genes están afectados por diferentes factores. Por ejemplo, la fijación
de nitrógeno está afectado principalmente por la región mientras que los genes
asociados con la nitrificación están afectados principalmente por el uso de suelo.

8. REFERENCIAS
● Acosta-Martínez, V., & Tabatabai, M. A. 2000. Enzyme activities in a limed
agricultural soil. Biology and Fertility of Soils, 31(1), 85–91. ● Allison, S. D., & Vitousek, P. M. 2005. Responses of extracellular enzymes to
simple and complex nutrient inputs. Soil Biology and Biochemistry, 37(5), 937– 944.
● Banerjee, S., Walder, F., Buchi, L., Meyer, A., Gattinger, A., Keller, T., Charles, R., & Van der Heijden, M. 2019. Agricultural intensification reduces microbial network complexity and the abundance of keystone taxa on roots. The ISME Journal. 8.
● Barberán A, Bates ST, Casamayor EO, Fierer N. 2012. Using network analysis to explore co-occurrence patterns in soil microbial communities. ISME J. 6:343–51.
● Bates, S. T., Berg-Lyons, D., Caporaso, J. G., Walters, W. a, Knight, R., & Fierer, N. 2011. Examining the global distribution of dominant archaeal populations in soil. The ISME Journal, 5(5), 908–17.
● Bender SF, Wagg C, van der Heijden MGA. 2016. An underground revolution: biodiversity and soil ecological engineering for agricultural sustainability. Trends Ecol Evol. 31:440–52.
● Bowles, T. M., Acosta-Martínez, V., Calderón, F., & Jackson, L. E. 2014. Soil enzyme activities, microbial communities, and carbon and nitrogen availability in organic agroecosystems across an intensively-managed agricultural landscape. Soil Biology and Biochemistry, 68, 252–262.
● Cai, Y., Yue, W., Xu, Yang, Z., Rong, Q. 2016. Sustainable urban wáter resources management considering life-cycle environmental impacts of wáter utilization under uncertainly. Resour converv Recycl. 108:21-40.
● Calle Z, Murgueitio E. 2012. Integrating forestry, sustainable cattle-ranching and landscape restoration 63:31–40
● Calle, M. Baer, I., Robouch, P., Cordeiro, F., Emteborg, H., Baxter, M., Brereton, N., Raber, G., Velez, D., Devesa, V., Rubio, R., Llorente-Mirandes, T., Raab, A., Feldmann, J., Sloth, J., Rasmussen, R., D´Amato M., Cubadda, F. 2012. It is posible to agree on a value for inorganic arsenic in food? The outcome of IMEP-112. Anal Bioanal Chem. 8: 2475-2488.
● Caporaso JG, Lauber CL, Walters W a, Berg-Lyons D, Huntley J, Fierer N, Owens SM, Betley J, Fraser L, Bauer M, Gormley N, Gilbert J a, Smith G, Knight R. 2012. Ultra-highthroughput microbial community analysis on the Illumina HiSeq and MiSeq platforms.
● Caporaso JG, Lauber CL, Walters W a, Berg-Lyons D, Lozupone C a, Turnbaugh PJ, Fierer N, Knight R. 2011. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. U. S. A. 108:4516–22.
● Clough, T., Lanigan, G., De Klein, C., Samad, S., Morales, S., Rex, D., Bakken, L., Johns, C., Condron, L., Grant, J., Richards, K. 2017. Influence of soil moisture on codenitrification fluxes from a urea-affected pasture soil. Nature. 7: 2185.

● Coskun, D., Britto, D., Shi, W., Kronzucker, H. 2017. Nitrogen transformations in modern agricultura and the role of biological nitrification inhibition. Nature plants. 3: 17074.
● Cubillos A. 2012. Efecto de sistemas silvopastoriles sobre las comunidades bacterianas edáficas en la reserva natural El Hatico (Valle del Cauca, Colombia).
● Cubillos, A., Vallejo, V., Arbeli, Z., Teran, W., Dick, R., Molina, C., Molina, E. & Roldan, F. 2016. Effect of the conversion of conventional pasture to intensive silvopastoril systems on edaphic bacterial and ammonia oxidizer communities in Colombia.
● Cho, J., & Tiedje, J. M. 2001. Bacterial Species Determination from DNA-DNA Hybridization by Using Genome Fragments and DNA Microarrays. Appl Environ Microbiol., 67(8), 3677-82.
● Dionisi, H. M., Layton, A. C., Harms, G., Igrid, R., Robinson, K. G., Sayler, G. S., & Gregory, I. R. 2002. Quantification of Nitrosomonas oligotropha -Like Ammonia-Oxidizing Bacteria and Nitrospira spp. from Full-Scale Wastewater Treatment Plants by Competitive PCR. Applied and Environmental Microbiology, 68(1), 245–253.
● Drenovsky R, Steenwerth K, Jackson L, Scow KM. 2014. Land use and climatic factors structure regional patterns in soil microbial communities. Natl. Inst. Heal. 19:27–39.
● Elsacker S Van. 2011. Effects of land use and soil management on soil quality in the Mekong Delta, Vietnam. Universiteit Gent.
● Fitriyanto, N. A., Fushimi, M., Matsunaga, M., Pertiwiningrum, A., Iwama, T., & Kawai, K. 2011. Molecular structure and gene analysis of Ce3+-induced methanol dehydrogenase of Bradyrhizobium sp. MAFF211645. Journal of Bioscience and Bioengineering, 111(6), 613–617.
● Francis, C. A., O’Mullan, G. D., & Ward, B. B. 2003. Diversity of ammonia monooxygenase (amoA) genes across environmental gradients in Chesapeake Bay sediments. Geobiology, 1, 129–140.
● Fromin N, Hamelin J, Tarnawski S, Roesti D, Forestier N, Gillet F, Aragno M, Rossi P. 2002. Statistical analysis of denaturing gel electrophoresis (DGGE) fingerprinting patterns. Evironmental Microbiol. 4:634–643
● Galdos MV, Cerri CC, Cerri CEP. 2009. Soil carbon stocks under burned and unburned sugarcane in Brazil. Geoderma 153:347–352.
● Garrigues E, Corson MS, Angers D a., van der Werf HMG, Walter C. 2012. Soil quality in Life Cycle Assessment: Towards development of an indicator. Ecol. Indic. 18:434–442.
● Groenigen, K., Hastings, A., Forristal, D., Roth, B., Jones, M., Smith. 2011. Soil C storage as affected by tillage and Straw management: As assessment using field measurements and model predictions. Agriculture, Ecosystems and Environment. 140: 218-225.
● Groffman, P. M., & Bohlen, P. J. 1999. Soil and sediment biodiversity: cross-system comparisons and large-scale effects. BioScience, 49(2), 139–148.

● Hariharan, J. 2015. Predictive profiling of soil microbes under different tillages and crop. The Ohio State University.
● Hofstra, N., & Bouwman, A. F. 2005. Denitrification in agricultural soils: Summarizing published data and estimating global annual rates. Nutrient Cycling in Agroecosystems, 72(3), 267-278.
● Hoorman, J., Islam., R. 2010. Understanding soil microbes and nutrient recycling. Agriculture and Natural Resource.
● Huson, D. H., Auch, A. F., Qi, J., & Schuster, S. C. 2007. MEGAN analysis of metagenomic data. Genome Research, 17(3), 377–86.
● Jangid K, Williams M a., Franzluebbers AJ, Sanderlin JS, Reeves JH, Jenkins MB, Endale DM, Coleman DC, Whitman WB. 2008. Relative impacts of land-use, management intensity and fertilization upon soil microbial community structure in agricultural systems. Soil Biol. Biochem. 40:2843–2853
● Jones, T. H., & Bradford, M. A. 2001. Assessing the functional implications of soil biodiversity in ecosystems. Ecological Research, 16(5), 845–858.
● Keeler, B. L., Hobbie, S. E., & Kellogg, L. E. 2009. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: implications for litter and soil organic matter decomposition. Ecosystems, 12(1), 1–15.
● Langille, M. G. I., Zaneveld, J., Caporaso, J. G., McDonald, D., Knights, D., Reyes, J. A, & Huttenhower, C. 2013. Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nature Biotechnology, 31, 814–21.
● Leff, J., Bardgett, R., Wilkinson, A. 2018. Predicting the structure of soil communities from plant community taxonomy, phylogeny, and traits. ISME J. 12: 1794-1805.
● León-Gamboa, A.L., Ramos, C. & García, M.R., 2010.- Efecto de plantaciones de pino en la artropofauna del suelo de un bosque altoandino. Revista de Biología Tropical, 58: 1031-1048
● Li C, Yan K, Tang L, Jia Z, Li Y. 2014. Change in deep soil microbial communities due to long-term fertilization. Soil Biol. Biochem. 75:264–272.
● Liu Y, Gao M, Wu W, Tanveer SK, Wen X, Liao Y. 2013. The effects of conservation tillage practices on the soil water-holding capacity of a non-irrigated apple orchard in the Loess Plateau, China. Soil Tillage Res. 130:7–12.
● Liu, W., Marsh, T. L., Cheng, H., & Forney, L. J. 1997. Characterization of Microbial Diversity by Determining Terminal Restriction Fragment Length Polymorphisms of Genes Encoding 16S rRNA. Applied and Environmental Microbiology, 63(11), 4516–4522.
● Lori M, Symnaczik S, Mäder P, De Deyn G, Gattinger A. 2017. Organic farming enhances soil microbial abundance and activity A meta-analysis and meta-Regression. PLoS ONE.12:1–25.
● Lloyd, D., Soldat, D., & Balser, T. C. 2009. Understanding microbes helps explain nutrient cycling. Turfgrass Trends.

● Meyer, F., Paarmann, D., D’Souza, M., Olson, R., Glass, E. M., Kubal, M., & Edwards, R. A. 2008. The metagenomics RAST server - a public resource for the automatic phylogenetic and functional analysis of metagenomes. BMC Bioinformatics, 9, 386.
● Muyzer, G., de Waal E.C., & Uitterlinden, A.G., 1993. Profiling of complex microbial populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Applied and Environmental Microbiology, 59(3), 695–700.
● Oades, J. M., & Jenkinson, D. S. 1979. Adenosine triphosphate content of the soil microbial biomass. Soil Biology and Biochemistry, 11(2), 201–204.
● Paungfoo-Lonhienne C., Wang, W., Yeoh, Y., Halpin, N. 2017. Legume crop rotation suppressed nitrifying microbial community in a sugarcane cropping soil. Nature. 7: 16707.
● Pester, M., Bittner, N., Deevong, P., Wagner, M., & Loy, A. 2010. A “rare biosphere” microorganism contributes to sulfate reduction in a peatland. The ISME Journal, 4(12), 1591–1602.
● Pezzopane, J.R.M., Nicodemo, M.L.F., Bosi, C. Garcia, A.R. Lulu, J.2019 Animal thermal comfort indexes in silvopastoral systems with different tree arrangements. J. Therm. Biol., 79: 103-111.
● Purkhold, U., Pommerening-Röser, a., Juretschko, S., Schmid, M. C., Koops, H. P., & Wagner, M. 2000. Phylogeny of all recognized species of ammonia oxidizers based on comparative 16S rRNA and amoA sequence analysis: Implications for molecular diversity surveys. Applied and Environmental Microbiology, 66(12), 5368–5382.
● Ratnayake, R.R., Seneviratne, G., Kulasooriya, S.A., 2011. Effects of land use and management practices on quantitative changes of soil carbohydrates. J. Natn. Sci. Foundation Sri Lanka 39(4): 345 – 353.
● Rotthauwe, J. H., Witzel, K. P., & Liesack, W. 1997. The ammonia monooxygenase structural gene amoa as a functional marker: Molecular fine-scale analysis of natural ammonia-oxidizing populations. Applied and Environmental Microbiology, 63(12), 4704–4712.
● Sáenz, J. 2014. Efecto del uso del suelo sobre comunidades microbianas de sistemas de pastoreo y cultivos de caña de azúcar del Valle del Cauca.
● Sait, M., Hugenholtz, P., & Janssen, P. H. 2002. Cultivation of globally distributed soil bacteria from phylogenetic lineages previously only detected in cultivation-independent surveys. Environmental Microbiology, 4(11), 654–666.
● Scheu, S. 2003. Effects of earthworms on plant growth: patterns and perspectives. Pedobiologia, 47(5-6), 846–856.
● Selbie, D. R., Buckthought, L. E. & Shepherd, M. A. 2015. The Challenge of the Urine Patch for Managing Nitrogen in Grazed Pasture Systems. Adv. Agron.129, 229–292.
● Soon, Y. K., & Arshad, M. a. 2005. Tillage and liming effects on crop and labile soil nitrogen in an acid soil. Soil and Tillage Research, 80, 23–33.

● Sparling, G. P., Ord, B. G., & Vaughan, D. 1981. Microbial biomass and activity in soils amended with glucose. Soil Biology and Biochemistry, 13(2), 99–104.
● Sperling, M., Piontek, J., Engel, A., Wilshire, K., Niggemann, J., Gerdts, G. Wichels, A. 2017. Combined carbohydrates support rich communities of particle-asociated marine bacterioplankton. Microbiology.
● Stoecker, K., Bendinger, B., Schöning, B., Nielsen, P. H., Nielsen, J. L., Baranyi, C., & Wagner, M. 2006. Cohn’s Crenothrix is a filamentous methane oxidizer with an unusual methane monooxygenase. Proceedings of the National Academy of Sciences of the United States of America, 103(7), 2363–2367.
● Taylor, J. P., Wilson, B., Mills, M. S., & Burns, R. G. 2002. Comparison of microbial numbers and enzymatic activities in surface soils and subsoils using various techniques. Soil Biology and Biochemistry, 34(3), 387–401.
● Tiedje, J. M., Asuming-Brempong, S., Nüsslein, K., Marsh, T. L., & Flynn, S. J. 1999. Opening the black box of soil microbial diversity. Applied Soil Ecology, 13(2), 109–122.
● Vallejo VE, Arbeli Z, Terán W, Lorenz N, Dick RP, Roldan F. 2012. Effect of land management and Prosopis juliflora (Sw.) DC trees on soil microbial community and enzymatic activities in intensive silvopastoral systems of Colombia. Agric. Ecosyst. Environ. 150:139–148
● Vallejo VE, Roldan F, Dick RP. 2010. Soil enzymatic activities and microbial biomass in an integrated agroforestry chronosequence compared to monoculture and a native forest of Colombia. Biol. Fertil. Soils 46:577–587.
● Vermeulen, S. J., Campbell, B. M., & Ingram, J. S. I. 2012. Climate Change and Food Systems. Annual Review of Environment and Resources, 37(1), 195–222.
● Wagg, S., Bender, F., Widmer, F. & Van der Heijden, A. 2014. Soil biodiversity and soil community composition determie ecosystem multifunctionality. PNAS 14: 5266-5270.
● Wertz, J. T., Kim, E., Breznak, J. A., Schmidt, T. M., & Rodrigues, J. L. M. 2012. Genomic and physiological characterization of the Verrucomicrobia isolate Diplosphaera colitermitum gen. nov., sp. nov., reveals microaerophily and nitrogen fixation genes. Applied and Environmental Microbiology, 78(5), 1544–1555.
● Xue, D., Huang, X., Yao, H., & Huang, C.2010. Effect of lime application on microbial community in acidic tea orchard soils in comparison with those in wasteland and forest soils
● Xue, Z., Malmer, D., Langille, M., Way, S. & Knight, R. 2014. Which is more important for clasifying microbial communities: who´s there or what they can do? ISME journal. 8 2357–2359.
● Zhao J, Ni T, Li Y, Xiong W, Ran W, Shen B, Shen Q, Zhang R. 2014. Responses of bacterial communities in arable soils in a rice-wheat cropping system to different fertilizer regimes and sampling times. PLoS One 9.



















