Thème : Génétique et évolutionhboucher-ts3.e-monsite.com/medias/files/theme-genetique-et... ·...
29
1 Thème : Génétique et évolution Chapitre 1 : Brassage génétique, innovation génétique et évolution des génomes Introduction Les espèces qui utilisent la reproduction sexuée conservent leurs plan d’organisation au cour des générations mais se caractérisent aussi par leur très grandes diversité génétique et phénotypique Les deux mécanismes fondamentaux de la reproduction sexuée sont à l’origine de cette diversité : La méiose permet la formation des gamètes La fécondation permet la rencontre des gamètes Comment ces mécanismes contribuent-il à la diversité génétique des individus ? I. Méiose et production de gamètes haploïdes variés. 1) La méiose dans le cycle de développement d’un organisme diploïde. Activité 1 : Découvrir le rôle de la méiose dans le cycle de développement d’un organisme diploïde. Bilan cours a) Place de la méiose dans le cycle de développement des diploïdes.
Transcript of Thème : Génétique et évolutionhboucher-ts3.e-monsite.com/medias/files/theme-genetique-et... ·...

1
Thème : Génétique et évolution
Chapitre 1 : Brassage génétique, innovation génétique et
évolution des génomes
Introduction
Les espèces qui utilisent la reproduction sexuée conservent leurs plan d’organisation au cour des
générations mais se caractérisent aussi par leur très grandes diversité génétique et phénotypique
Les deux mécanismes fondamentaux de la reproduction sexuée sont à l’origine de cette diversité :
La méiose permet la formation des gamètes
La fécondation permet la rencontre des gamètes
Comment ces mécanismes contribuent-il à la diversité génétique des individus ?
I. Méiose et production de gamètes haploïdes variés.
1) La méiose dans le cycle de développement d’un organisme diploïde.
Activité 1 : Découvrir le rôle de la méiose dans le cycle de développement d’un organisme diploïde.
Bilan cours
a) Place de la méiose dans le cycle de développement des diploïdes.

2
1 : cellule a 2N chromosome monochromatidiens
2 : synthèse d’ADN = duplication (phase S)
3 : cellule a 2N chromosome bichromatidiens (cellule diploïde)
4 : première division de méiose, passage a 2 cellules à N chromosomes bichromatidiens
(cellules haploïde)
5 : seconde division de méiose, passage à 4 cellules a N chromosomes monochromatidiens
(cellules haploïde)
Un organisme diploïde possède des paires de chromosomes, ces cellules sont dites à 2N
chromosome n étant le nombre de paire de chromosome. Dans le cycle de développement dans
organisme diploïde, la méiose est une division qui permet la formation des cellules sexuelles ou
gamètes males et femelles, qui sont haploïdes. La méiose permet de diviser par deux la quantité
Schéma : Le cycle de développement d’un organisme diploïde
Schéma : Quantité d’ADN dans une lignée cellulaire pendant la
méiose

3
d’ADN présente dans les gamètes (voir graphique ci-dessus.) ces gamètes ne possèdent plus de paire
de chromosomes et sont donc des cellules à N chromosomes.
La méiose permet donc d’obtenir l’haploïdie, c'est-à-dire permet le passage d’un état diploïde a un
état haploïde. Chez les diploïdes la méiose précède obligatoirement la fécondation qui reforme de
nouveaux organismes diploïdes. Ceci est indispensable pour le caryotype d’une espèce se maintienne
au cours des générations.
La méiose est obligatoirement précédée par une réplication semi-conservative de l’ADN, qui permet
de doubler la quantité d’ADN et de passer de chromosomes à une chromatide à des chromosomes à
deux chromatides.
b) Les étapes de la méiose
La méiose est composée de successions de deux divisions.
La première division est une division réductionnelle car elle sépare toutes les paires de
chromosomes, c’est donc elle qui permet de passer d’une cellule 2N à une cellule N
Prophase : les chromosomes sont visibles et possèdent deux chromatides chacun. Les
homologues (les chromosomes d’une même paire) s’apparient, ils forment des bivalents ou
tétrades. Au cours de cette étape des échanges de portion de chromatides peuvent se
produire entre deux homologues d’une même paire : se sont les « crossing-over ».
Métaphase : les chromosomes homologues se placent de part et d’autre de la plaque
équatoriale. Contrairement à la mitose ils ne se divisent pas au niveau du centromère.
Anaphase : les chromosomes homologues se séparent et migrent aux pôles de la cellule.
Télophase : deux cellules haploïdes à N chromosome se forment
La seconde division est une division équationnelle qui équivaut à une mitose concernant les deux
cellules à N chromosomes issues de la première division de méiose.
Prophase : les chromosomes à deux chromatides sont bien individualisés
Métaphase : tous les chromosomes se rangent à l’équateur de la cellule et ils se coupent au
niveau du centromère.
Anaphase : deux lots identiques de N chromosomes à une chromatide migrent aux pôles de
la cellule.
Télophase : Quatre cellules à N chromosomes s’individualisent.
Le mécanisme du crossing-over entre deux chromosomes homologues en prophase I de méiose
Début de Prophase I de méiose (Appariement chromosomes homologues)
Fin de prophase I de méiose (Formation de chromosomes recombinés)
Crossing-over

4
Conclusion : bilan de la méiose
Exercice d’application page 30 ex 7
1) 4 : prophase I 2Q 2N
3 : métaphase I 2Q 2N
8 : anaphase I2Q 2N
5 : télophase I Q N
2 : Prophase II Q N
6 : Métaphase II Q N
7 : Anaphase II Q N
1 : Télophase II Q/2 N
2) Les brassages génétiques de la méiose et la production de gamètes
haploïdes variés
a) Le brassage intra-chromosomiques

5
P
Génotype → e
Phénotype →
_ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _
Gamètes ↓ ↘ Gènes eb+ ; vg+ eb ; vg
F1
Génotype→ eb+//eb ; vg+//vg
Phénotypes de tous les F1 →
2ème croisement
Parent F1 femelle x double récessif mal
Génotype eb+//eb ; vg+// vg eb//eb ; vg//vg
Phénotype
Descendants : à compter !
Gènes : couleurs du corps (2 allèles)
Clair eb+
Ebène eb
Gènes : longueur des ailles (2 allèles)
Longues vg+
Vestigiales vg
1 2

6
Les résultats obtenu montre 4 sortes de descendent, 2 de phénotypes parental en grande proportion
et 2 de phénotypes recombiné en faible proportion. Ils sont issu du croisement test (ou test cross),
donc leur proportion dépendent de la proportion des gamètes du parent hétérozygote. Les gènes
sont donc situé sur la même paire de chromosomes et un brassage intra chromosomique à eu lieu
par crossing-over, lors de la prophase 1 de méiose, lorsque les chromosomes homologues étaient
appariés.
Voir livre page 25 : schéma des gènes liés

7
Bilan : Le brassage intra chromosomique concerne des gènes porté par la même paire de
chromosomes : se sont des gènes liés. Au cours de la première division de méiose, lorsque pendant la
prophase les chromosomes homologues s’apparient et que leur chromatides s’enchevêtre, des
chiasmas s’établissent, ce qui permet des crossing-over avec échange de matériel génétique entre les
chromatides de chromosomes homologues. On obtient alors des chromosomes recombiné c'est-à-
dire présentant une nouvelle combinaison allélique originale.
Schéma théorique

8
Une formule mathématique permet de prévoir le nombre potentiel de gamètes produit par ce
brassage. Ce nombre dépend du nombre de gènes porté par une paire de chromosomes et du
nombre de paire de chromosomes. Pour une seule paire de chromosome donné comportant x gènes
le nombre de gamètes : 2x x est le nombre de gènes à l’état hétérozygotes
Les crossing-over sont donc responsable du brassage intra chromosomiques, il contribuent a la
diversité des gamètes en permettant la création de nouvelles combinaison alléliques. Ceci permet
d’augmenter la diversité génétique des individus
Remarques lors des croisements la proportion des gamètes avec des chromosomes recombinés est
toujours très inférieure à la proportion des gamètes de types parentales. En effet les méioses avec
crossing-over sont toujours plus rares que celles sans crossing-over. Se sont les croisements test ou
test-cross qui révèlent un éventuel brassage intra chromosomique
Définition : le test-cross consiste à croiser un individu de génotype inconnu (généralement un F1)
avec un individu de phénotype récessif : sont génotype est alors parfaitement connu, il est
obligatoirement homozygote récessif pour tout les gènes considéré.
Dans le cas d’un brassage intra chromosomique affectant des gènes lié le croisement test le révélera
en montrant des proportions inégales de phénotype pour les descendants : il y aura une majorité de
phénotypes parentaux et une minorité de phénotypes recombiné
b. Le brassage inter-chromosomique

9
Le brassage inter chromosomique concerne des gènes indépendant (des gènes non liés) porté par
des paires différentes de chromosomes. Lors de la 1ère division de méiose, toutes les paires de
chromosomes homologues se séparent de façon aléatoire, donnant naissance a des gamètes
haploïdes génétiquement différents en quantité équiprobables.

10
Les résultats d’un croisement test révèlent donc un brassage inter chromosomique en montrant des
proportions égales de phénotype pour les descendants
Voir livre page 25 figure 3

11
Une formule mathématique permet d’estimer le nombre de gamètes différents possibles produit par
ce type de brassage
Nombre possible de gamètes différent = 2N pour N= nombre de paire de chromosomes
Le brasage inter chromosomique qui au cours de la méiose se produit après le brassage intra
chromosomique amplifie encore le mélange des allèles. Ceci accentue encore la diversité des
gamètes produit et par conséquent la diversité des individus
Pour estimer le nombre potentiel de gamètes différent produit par l’association des deux brassages
on utilise la formule suivante :
Nombre de gamètes = 2NX pour N= nombre de paire de chromosomes X= nombre de gènes
hétérozygotes liés
II. Fécondation et production d’organismes diploïdes variés
1) Place de la fécondation dans le cycle de développement d’un organisme
diploïde
Voir Schéma Le cycle de développement d’un organisme diploïde
Dans le cycle d’un organisme diploïde, la fécondation survient peut après la méiose. Au cours de la
fécondation, un gamète mal et un gamète femelle fusionne, cela forme un zygote ou cellule œuf
diploïde. Les noyaux males et femelles fusionnent également : c’est la caryogamie voir page 21 doc
1b

12
2) Le brassage génétique de la fécondation amplifie la diversité génétique
des individus Exercice doc 2 page 20-21
Questions 2,4,5,6 page 20
Question 3
A P Ap aP a p
AP AP AP AP AP
Ap AP Ap AP Ap
aP AP AP aP aP
a p AP Ap aP ap
AP= 9/16= 56.25% Ap= 3/16=18.75% aP= 3/16=18.75% ap= 1/16=6.25%
Question 4
9 cas différents
Bilan : Lors de la fécondation, la rencontre au hasard de gamètes génétiquement différents donne à
tous zygotes une combinaison originale d’allèles. Il existe une formule mathématique pour
déterminer le nombre de zygotes possible obtenu par fécondation.
Nombre zygotes :
La fécondation amplifie donc les brassages alléliques de la méiose, elle est elle-même responsable
d’une immense diversité génétique possible pour les zygotes. Chaque zygote contient donc une
combinaison unique et nouvelle d’allèles. Seul une fraction d’entre eux est viable et se développe.
III. Méiose, innovation génétique et évolution des génomes
1) Anomalie de disjonctions de chromosomes en méiose et modification
des caryotypes.
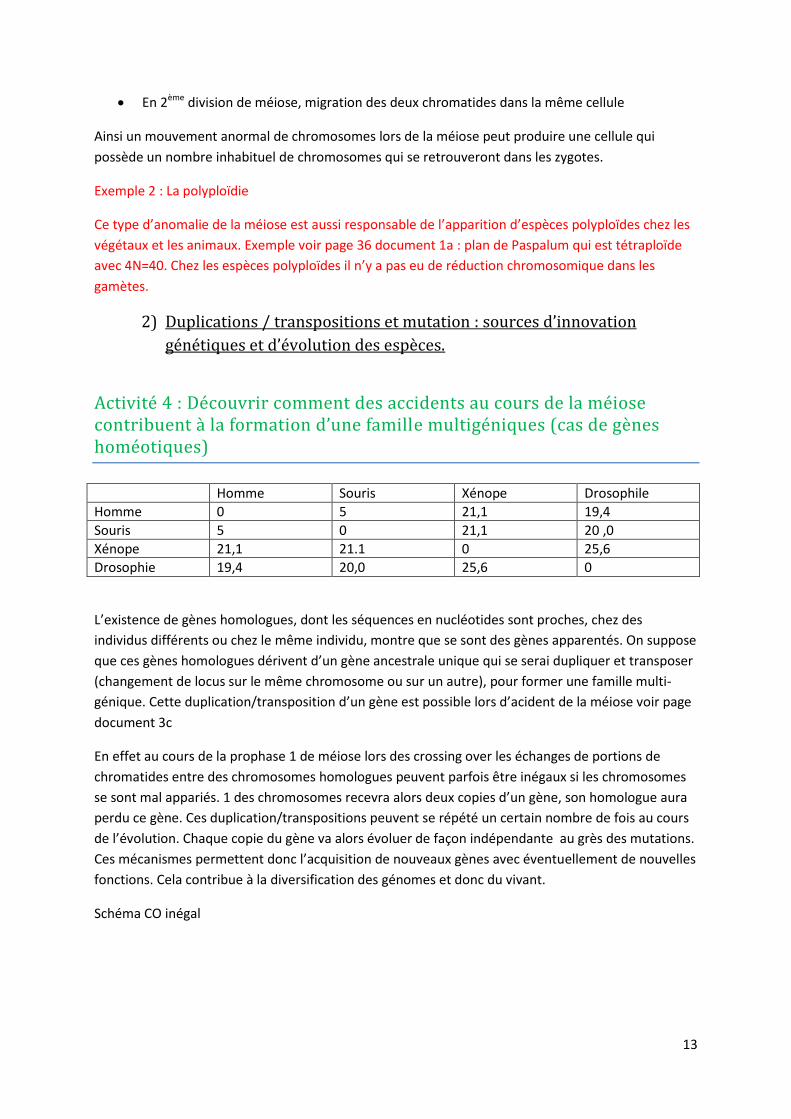
Exemple 1 : Trisomie 21 / syndrome de Down/ mongolisme
Le caryotype montre 3 chromosomes 21 au lieu de 2. Une mauvaise disjonction des chromosomes
homologues au cours de la méiose peut être responsable de cette anomalie.
En 1ère division de méiose, une non-séparation des chromosomes homologues.
13
En 2ème division de méiose, migration des deux chromatides dans la même cellule
Ainsi un mouvement anormal de chromosomes lors de la méiose peut produire une cellule qui
possède un nombre inhabituel de chromosomes qui se retrouveront dans les zygotes.
Exemple 2 : La polyploïdie
Ce type d’anomalie de la méiose est aussi responsable de l’apparition d’espèces polyploïdes chez les
végétaux et les animaux. Exemple voir page 36 document 1a : plan de Paspalum qui est tétraploïde
avec 4N=40. Chez les espèces polyploïdes il n’y a pas eu de réduction chromosomique dans les
gamètes.
2) Duplications / transpositions et mutation : sources d’innovation
génétiques et d’évolution des espèces.
Activité 4 : Découvrir comment des accidents au cours de la méiose contribuent à la formation d’une famille multigéniques (cas de gènes homéotiques)
Homme Souris Xénope Drosophile
Homme 0 5 21,1 19,4
Souris 5 0 21,1 20 ,0
Xénope 21,1 21.1 0 25,6
Drosophie 19,4 20,0 25,6 0
L’existence de gènes homologues, dont les séquences en nucléotides sont proches, chez des
individus différents ou chez le même individu, montre que se sont des gènes apparentés. On suppose
que ces gènes homologues dérivent d’un gène ancestrale unique qui se serai dupliquer et transposer
(changement de locus sur le même chromosome ou sur un autre), pour former une famille multi-
génique. Cette duplication/transposition d’un gène est possible lors d’acident de la méiose voir page
document 3c
En effet au cours de la prophase 1 de méiose lors des crossing over les échanges de portions de
chromatides entre des chromosomes homologues peuvent parfois être inégaux si les chromosomes
se sont mal appariés. 1 des chromosomes recevra alors deux copies d’un gène, son homologue aura
perdu ce gène. Ces duplication/transpositions peuvent se répété un certain nombre de fois au cours
de l’évolution. Chaque copie du gène va alors évoluer de façon indépendante au grès des mutations.
Ces mécanismes permettent donc l’acquisition de nouveaux gènes avec éventuellement de nouvelles
fonctions. Cela contribue à la diversification des génomes et donc du vivant.
Schéma CO inégal

14
Conclusion du chapitre
La reproduction sexuée aboutie a une grande variabilité des individus, grâce aux brassages
génétiques de la méiose et de la fécondation. Certains accidents lors de la méiose peuvent faire
apparaitre des caryotypes anormaux, mais ils peuvent aussi contribuer à l’évolution des êtres vivants
en favorisant la diversité des génomes avec la formation des familles multi-géniques.

15
Chapitre 2 : Mécanisme de diversification des êtres vivants et
évolution de la biodiversité.
Introduction
L’association des mutations et du brassage génétique au cours de la méiose et de la fécondation ne
suffi pas a expliqué la totalité de la diversification génétique des êtres vivants. En outre une
diversification des êtres vivants n’est pas toujours lié à une diversification génétique.
Questions
Quels sont les autres mécanismes génétique ou non qui contribue a l’évolution des êtres vivants et
de la biodiversité ?
Comment cette biodiversité peut-elle évoluer au cours du temps et conduire a la formation de
nouvelles espèces ?
I. Mécanismes impliqués dans la diversifications des êtres
vivants
A) D’autres mécanismes impliqués dans la diversification du vivant
1) Association de génomes et diversification du vivant
Activité 1 : Découvrir comment l’hybridation chez les végétaux conduit à l’évolution des
génomes et à la diversification du vivant.
1)
2) Sur chaque espèce de spartina on retrouve des points communs
Espèce importée Spartina 2n=62
Espèce initiale Spartina maritima
2n=60
Spartina maritima
2n=60 sexué
Spartina alterniflora
2n=62 sexué
Spartina x townsendii n=61
asexué ; hybridation: croisement
entre deux espèces différentes
Spartina anglica
2n=122 sexué
Méiose
Méiose Gamètes
N=30
Gamètes N=31
Hybridation : Spartiva x townsendii
N=61 Méiose impossible
Doublement accidentel des chromosomes
Spartina anglica 2n=122
Méiose possible

16
Bilan : La plupart des plantes et des animaux sont diploïdes cependant certaines espèces peuvent
être polyploïdes c’est à dire possédé leurs chromosomes en plus de 2 exemplaires (voir doc 1 page
36)
Plusieurs mécanismes peuvent expliquer cette polyploïdie :
Un individu d’une espèce subit une anomalie lors de sa méiose ce qui peut conduire à un doublement du nombre de chromosomes dans les gamètes (cf chapitre 1 : non disjonction des paires de chromosomes homologues) l’autopolyploïdie.
Deux individus appartenant a des espèces différentes peuvent s’hybrider (allo ploïdie). Le descendant hérite alors d’un lot chromosomique de chaque parent. Comme les chromosomes proviennent d’espèces différentes, ils ne sont pas homologues et leur appariement en méiose est donc impossible. Ceci explique la stérilité fréquente des hybrides. Si un événement accidentel de doublement des chromosomes suit une hybridation, chaque chromosome retrouve un homologue. La méiose redevient possible et la fertilité de l’espèce est rétablie.
La polyploïdie joue un rôle très important dans l’évolution des plantes, elle est source de variabilité génétique ce qui contribue à la diversification des espèces végétales. De nombreuses espèces de plantes y compris des plantes cultivé sont des polyploïdes (exemple : blé, colza et tabac)
2) Transfert entre espèces et diversification du vivant
Bilan : Des transferts horizontaux de gènes entre espèces très différentes existent et peuvent contribuer à la diversification des génomes et des êtres vivants. Ces transferts de gènes plus fréquemment réalisé par des virus ou bactéries peuvent apportés d’éventuels avantages aux organismes qui les reçoivent. Exemple : chez les primates c’est le transfert d’un gène viral (codant pour une protéine : la syncytine) qui a permis l’acquisition du placenta. Des études récentes on montrer que des rétrovirus on transféré horizontalement le gène codant de la syncytine à des mammifères probablement entre - 40 et -25 million d’années
3) Modification de l’expression des gènes et diversification du vivant
Activité 2 : Expliquer l’absence de pattes chez les serpents.
Connaissances : s’agissant des gènes impliqués dans le développement, des formes vivantes très différentes peuvent résulter de variations dans la chronologie de l’intensité d’expression de gènes communs plus que d’une différence génétique. Capacité : saisir et interpréter des informations, les mettre en relation logique dans un but explicatif. (Entrainement à la question de type IIB) Attitude : manifester sens de l’observation, curiosité, esprit critique. Comprendre la nature provisoire, en devenir, du savoir scientifique Bilan : L’homologie des gènes homéotiques révèlent qu’ils appartiennent à la même famille multi génique et qu’il on une origine commune. Ils ont été préservés au cours de l’évolution. Ainsi des gènes homéotiques très comparables peuvent êtres présents chez des animaux très différents pour programmer leur développement. Ce sont donc des variations dans la chronologie, l’intensité et la localisation d’expression de ces gènes qui varient d’une espèce à l’autre, plus qu’une modification génétique entre eux. Ces variations se traduisent par des modifications des organes formés à l’issue du développement embryonnaire. Ces modifications de l’expression des gènes homéotiques au cours de l’évolution ont ainsi pu mener à des innovations qui on été retenues ou non, permettant d’assurer la diversité du vivant.

17
B) Mécanisme de diversification du vivant sans modification des génomes.
1) Association de type symbiose et diversification du vivant.
Question document 6 page
Sur la photographie on voit deux éléments des filaments (mycéliums) en beige (champignon) et des
sphères (cyanobactéries) vertes (algues) des études génétiques montrent que deux génomes
différents se trouvent dans le lichen : le premier appartient a une algue et le second à un
champignon.les algues peuvent fixer le CO2 et donc procédé à la photosynthèse (autotrophe : se
nourri seul) et le champignon n’a pas cette capacité de fixer le CO2 (il est hétérotrophe).
Les algues vont transformer le CO2 en matière organique et les champignons vont alors se nourrir de
cette matière organique. De ce fait on constate du carbone 14 dans les champignons. 30 à 35% de la
matière organique de l’algue est absorbé par le champignon.
En échange, les champignons vont protéger les algues et donner des sels minéraux (prélevé de la
roche qu’ils dissout à l’aide d’acides) aux algues.
Bilan : les individus appartenant à des espèces différentes peuvent vivre en association étroite et
durable une telle association profitable pour les deux partenaires est qualifié de symbiose. La
symbiose peut aboutir à l’acquisition de nouvelles fonctions ex : assimilation de l’azote
atmosphérique par les plantes vivant en symbioses avec certaines bactéries (voir livre doc 1 page 40).
La symbiose peut aussi assurer la survie préférentielle d’organismes dans un milieu donné c’est donc
une association qui contribue à la diversité du vivant.
2) Diversification du comportement et du vivant.
Questions 1 et 2 page 42-43
Bilan : chez les vertébrés notamment les oiseaux (acquisition du chant) et les mammifères (utilisation
d’outils) le développement de comportement nouveau peut se produire sans modification du
génotype et être transmis d’une génération a une autre par apprentissage et imitation. Cela
concerne surtout les jeunes d’une population qui imitent les plus âgés.
Cela est aussi source de diversité des êtres vivants
II. De la diversification des êtres vivants à l’évolution de la
biodiversité.
A) Evolution de la diversité des populations au cours du temps. Exercice 1 page 58
Pendant la sécheresse de 1977, le nombre de graines a baissé car les arbres et arbustes source de ces
graines on maqué d’eau pour les créé et leur dureté provient aussi du manque d’eau
Lorsque l’abondance des graines augmente et leurs dureté diminue, on observe une hausse du
nombre de pinçons et leur hauteur de bec est normale et stable alors que lorsque la quantité de
graines diminue et que leur dureté augmente, on observe une diminution du nombre de pinson et la
hauteur de leur bec augmente

18
L’évolution des populations dépend de l’hérédité et du milieu de vie. En effet, l’évolution s’adapte au
changement du milieu (sélection naturelle)
Bilan : on défini une population comme un ensemble d’individu de la même espèce, qui vivant à
proximité des un des autres se reproduisent majoritairement entre eux. On observe à l’intérieur
d’une population une variabilité génétique en effet les individus ne possèdent pas les même
combinaisons d’allèles pour leurs gènes. Une population est considérée comme une étape de
l’évolution. En effet divers mécanismes sont à l’origine de l’évolution des populations :
Action de l’environnement : il induit une sélection naturelle, les conditions du milieu et la
concurrence entre les individus exerce une pression sur les populations qui favorise la survie
des êtres les plus adapté à ces conditions. Par conséquent, ils se reproduisent d’avantages et
assurent mieux la transmission de leurs allèles.
L’effet du hasard : il intervient de façons multiples :
o Le hasard des mutations : il provoque une diversification génétique des individus
dans une population. De nouveaux allèles à l’origine de nouveaux caractères
apparaissent
o Le hasard lors de la reproduction : il peut provoquer une dérive génétique à partir
des allèles transmis lors de la reproduction sexuée.
L’évolution est donc la transformation des populations qui résulte de ses différences de survies et du
nombre de descendant.
B) Evolution de la biodiversité et apparition de nouvelles espèces
Activité 4 : expliquer l’apparition d’une nouvelle espèce de moustiques
1) Espèces : ensemble des individus qui se distinguent des autres par des caractères communs
et ont une descendance fertile.
Bilan :
a) Définir la notion d’espèce
La définition de l’espèce est délicate et peut reposer sur des critères variés qui permettent
d’apprécier le caractère plus ou moins distinct de deux populations. On peut définir l’appartenance
d’individus à la même espèce en se basant sur différents critères :
Critère de ressemblance : deux individus appartiennent à la même espèce s’ils se
ressemblent
Critère d’interfécondité : deux individus appartiennent à la même espèce s’ils peuvent se
reproduire entre eux et avoir une descendance fertile.
A partir du critère d’interfécondité, Ernest Mayr à proposer une définition biologique de l’espèce :
selon lui une espèce est une population ou un ensemble de populations don les individus peuvent
effectivement ou potentiellement se reproduire entre eux et engendré une descendance viable et
féconde.
Le concept d’espèce est modifié au cours de l’histoire de la biologie :

19
Conception pré-évolutionniste : l’espèce est une entité permanente et stable elle est défini
par rapport à un individu type. Tout les individus qui lui ressemble et qui sont interfécond
avec lui appartiennent à la même espèce. Toute variation par rapport à l’individu type est
considérée comme une anomalie.
Conception évolutionniste (XIXème siècle ): on admet la variabilité des individus au sein de
l’espèce, l’idée d’une filiation entre les espèce s’impose.
Actuellement : une espèce peut être considéré comme une population d’individu
suffisamment isolé génétiquement des autres populations pour qu’un isolement
reproducteur ai pu apparaitre.
b) La spéciation et l’évolution des espèces
La diversité du vivant ou biodiversité est en partie décrite comme une diversité d’espèces qui
sont susceptibles d’évolué au cours du temps. Au sein d’une même espèce, deux population
isolé par une barrière géographique ou comportementale on des échanges génétiques lié à la
reproduction sexué réduit et accumulent des différences génétiques importantes. Au delà
d’un certain seuil, ces différences peuvent empêcher les individus des deux populations de se
reproduire, il y a alors isolement reproducteur. Quand l’isolement reproducteur entre deux
populations est associé à un isolement génétique, il n’y a plus d’échanges d’allèles entres
elles. Chaque population est alors considéré comme une nouvelle espèce qui continuera à
évolué séparément sous l’effet du hasard et de la sélection naturelle. Ce processus
conduisant à la formation de nouvelles espèces est appelé spéciation. Une population
identifiée comme constituant d’une espèce n’est défini que durant une période limité. On dit
qu’une espèce disparait si l’ensemble des individus concerné disparait ou cesse d’être isolé
génétiquement.
Conclusion du chapitre
La biodiversité telle qu’on l’observe à une période donné est à la foi le résultat et une étape
de l’évolution. En effet il existe de divers mécanismes qui engendrent une modification de la
diversité génétique des populations au cours du temps

20
Chapitre 3 : un regard sur l’évolution de l’homme
Introduction :
Notre espèce « homo sapiens » a une histoire évolutive comme toutes les autres espèces. les
similitudes avec les autres primates comme les grands singe suggère une parenté commune. Quelles
sont les grandes étapes de cette évolution qui ont conduit à l’émergence de notre ligné humaine et
plus particulièrement du genre homo.
I. parenté de l’homme et des grands singes
1) Comparaison des caryotypes et des génomes entre l’homme et
chimpanzé Doc 1 page 76
Bilan : Le caryotype de l’homme possède 22 paires d’autosome plus une paire de gonosomes. On
observe qu’au niveau de caryotype du chimpanzé il y a 23 paires d’autosomes et 1 paire de
gonosomes. Le chimpanzé possède donc 1 paire de chromosome supplémentaire les comparaison
précises des 2 caryotypes révèlent que 13 paires de chromosomes sont vigoureusement identiques
chez les 2 espèces les autres paires se distinguent par quelques réarrangement chromosomiques et
en conduit à la délétion ou à l’addition de gènes. La différence du nb de chromosomes entre les 2
espèces est expliquée par la fusion de deux chromosomes du chimpanzé pour formé le chromosome
2 humain. La grande similitude des 2 caryotypes indique la parenté et l’origine commune entre
l’homme et le chimpanzé
Comparaison de génotype : (voir doc 2 page 77) l’homme et le chimpanzé présentent une
très grande proximité génétique. Depuis 2005 on sais qu’il y a 98,77% de similitudes
génétiques entre homme et chimpanzé ce qui confirme une proche parenté.
2) Développement et acquisition du phénotype humain sous le contrôle
génétique et de l’environnement La grande proximité génétique entre l’homme et les grands singes pose le problème de la
différence phénotypique évidente
Questions 1, 2, 3 pages 78-79
Bilan : Lorsque l’on compare les phases du développement pré et post natale chez l’homme et le
chimpanzé on observe que toutes les phases du développement sont allongé chez l’homme
notamment la période prénatale importante ou il y a multiplication des neurones (2 semaines chez le
chimpanzé contre 8 semaine chez l’homme) la naissance plus tôt chez le chimpanzé 8 mois au lieu de
9. Apres la naissance le trou occipital qui permet de relier la colonne vertébrale au crane migre d’une
position centrale vers l’arrière chez le chimpanzé. Cela conditionnera donc son mode de locomotion
(quadrupède chez le chimpanzé et bipède pour l’homme). On peut également observer que la
maturité sexuelle survient également plus tôt chez le chimpanzé 8 ans au lieu de 16 ans ainsi chez
l’homme les phases de développement semble tes ralenti par rapport à celle du singe, ce qui permet
le maintien de la bipédie (trou occipital central) et une morphologie crânienne proche de celle du

21
fœtus si ce n’est la taille. Chez le singe adulte, le crane a subit des modifications importantes avec un
développement accéléré qui n’a pas permit une croissance importante du crane (capacité crânienne
450cm3)
Questions 4, 5 pages 76-77
Bilan : Lorsque l’on compare les gènes de développement impliqué dans le développement cérébral
chez le chimpanzé et l’homme on observe une homologie de ces gènes de développement.
Néanmoins il présente des mutations qui permettent d’expliquer les différences de développement.
Ce sont donc ces différences dans les gènes de développement et dans leurs expressions qui
conduisent a l’acquisition d’un phénotype différent chez les deux espèces. En effet les gènes du
développement conditionnent l’ordre d’acquisition des caractères mais aussi la durée et le rythme de
croissance. Les conditions environnementales ont également une influence sur le développement
(nutrition, apprentissage, communication).
II. L’homme un primate parmi d’autres
1) Caractéristiques et diversité des primates actuel et fossiles
Activité 1 : Déterminer la parenté de l’homme avec d’autres primates
Bilan : Les primates actuels et fossiles présentes une très grande diversité (voir doc 2 page 81).
Néanmoins ils partagent des caractères homologues à l’état dérivé, qui leurs sont propres et qui
traduisent leur parenté et leur origines communes :
pouce opposables aux autres doigts
présence d’ongles au lieu de griffes
vision binoculaire bien développé
cortex cérébrale relativement bien développé
Les premiers primates fossiles datent de -65 a -50 millions d’années. Très diversifier, ils n’étaient ni
identique à l’homme actuel ni aux autres grands singes actuels. Les premier grands primates fossiles
dont nous faisons parti sont apparu il y a 20 millions d’années ils se distinguent par leur taille
importante, un nez dont les narines sont séparé par une fine cloison et par leur absence de queue.
La diversité des grands primates connus par les fossiles fut très grande (40 genres différents environ
il y a 30 millions d’années) aujourd’hui cette diversité très réduite on ne distingue plus que quelques
genres différents (orant outan, gorilles, homo, chimpanzé,..). Aujourd’hui les grands primates à
l’exception de l’homme vivent dans les forêts tropicales (chaude et humide) d’Afrique et d’Asie ce
qui explique la rareté de découverte de formes fossiles.
2) Caractéristiques de l’ancêtre commun de l’homme et du chimpanzé Voir doc 3 page 81
Bilan : Parmi les grands singes (hominoïdes) c’est l’homme et le chimpanzé qui présente la plus
grande parenté et qui partage un ancêtre commun le plus récent. La date de divergence entre les
deux ligné est actuellement situé à -7 millions d’années. On a pu établir cette date grâce aux données
génétiques mais aussi grâce à la découverte fossile de Toumaï daté à priori de -7 millions d’années et

22
qui ^présenterais déjà les caractères dérivé propre à la ligné humaine (bipédie envisagé). Cela di
aucun fossile ne peut être à coup sur considéré comme un ancêtre de l’homme et du chimpanzé.
III. Emergence et évolution du genre Homo
1) Les caractères propres au genre Homo TP phylogène
Exercice : http://evolution.biologique.free.fr/histoire/homme/comparai.htm
Utiliser les donné du site internet pour construire un tableau de comparaison des caractères
anatomique de l’homme et du chimpanzé
En déduire les critères dérivés propres a l’homme
Bilan : L’homme actuel se distingue d’un grand singe comme le chimpanzé par un certain nombre de
caractères. Des caractères crâniens tel-que sont volume cérébral. Sont volume cérébral important
1550cc contre 450cc pour le chimpanzé, sa face aplatie au lieu d’être prognathe, son trou occipital
centré au lieu d’être arrière. La posture droite de l’homme autorise sa bipédie alors que le chimpanzé
à une posture recourbé engendrant une locomotion principalement quadrupède. De plus il a une
capacité de langage, il réalise une utilisation intensive d’outils contrairement aux grands singes.
Toutes ces différences justifient que l’homme appartienne à un genre différent de celui des grands
singes : le genre Homo.
Le genre homo regroupe : l’homme (homo sapiens) et plusieurs espèces fossiles, plus proches
parents de l’homme actuel que des chimpanzés.
On distingue notamment :
Homo habilis daté de -2,5 à -2,8 millions d’années
Homo ergaster daté de -2,8 millions d’années à -150 mille ans
Homo néanderthalensis daté de -200 milles ans à -30 mille ans
Homo sapiens daté de -195 milles ans à nos jours.
Les premiers représentants du genre homo on été retrouvé en Afrique, à partir de -1 ,8 millions
d’années le genre homo connait une forte extension en Europe et en Asie.
Ces diverses espèces possèdent des caractères dérivé qui leur sont propres et qui se sont mis en
place progressivement :
Un dimorphisme sexuel peut marquer entre les squelettes masculins et féminins
Une face réduite : le prognathisme à progressivement régressé.
Une mandibule parabolique
Un volume cérébrale supérieur ou égal à 600cc (atteint avec homo habilis)
Une bipédie permanente et stricte permettant une aptitude à la couse à pied. Ceci exigent
les caractères anatomique suivant :
o Un trou occipital en position centrale

23
o Colonne relativement droite
o Bassin cour et évasé
o Membre inférieur plus long que les membres supérieurs
o Développement d’une voute plantaire
o Fémur en position évasé
Production d’outils complexe (ce caractère n’étant toute fois non exclusive au genre Homo)
Des pratiques cultuelles ( sculptures et peinture)
2) L’évolution du genre Homo : une évolution contrastée.
Bilan : Les diverses espèce du genre homo connu actuellement (plus de 10 espèces en 2012)
présentent des caractères dérivé commun qui traduisent leurs parenté étroite. Néanmoins cette
diversité d’espèce rend très difficile la reconstitution des parentés entre elles d’autant plus que
certaines espèces ont coexisté.
Les découvertes paléontologiques souvent incomplètes (doc 3 page 81) ne facilitent pas la tache.
L’évolution de l’homme est qualifié de buissonnante (schéma bilan page 87).
Conclusion du chapitre :
L’homme actuel est issu d’une évolution qui débute il y a -65 million d’années avec
l’apparition des premier primates. Les caractères dérivés qu’il partage avec tous les primates
traduisent leur grande parenté. Cependant c’est avec le chimpanzé qu’il partage la parenté
la plus étroite. A partir de -7 million d’années les deux ligné ont divergé des différences dans
le développement, sous l’action des gènes et de l’environnement ont permis l’émergence de
la ligné humaine puis celle du genre Homo dont nous sommes issus.

24
Chapitre 4 : les relations entre organisation et mode de vie,
résultat de l’évolution : exemple de la vie fixé chez les
plantes.
Introduction :
Les végétaux chlorophylliens peuplent tous les écosystèmes et comprennent en particulier des
plantes à fleurs, les arbres, les arbustes et des plantes herbacées. Ils sont fixés dans le sol où ils
puisent des matières minérales et produisent, grâce à la photosynthèse, de la matière organique à
partir de cette matière minérale et du CO2 prélevé dans l’atmosphère.
Chez les végétaux, la reproduction sexuée se fait comme chez les animaux par fécondation entre un
gamète mal et un gamète femelle aboutissant à une cellule œuf qui formera un nouvel individu. Chez
les plantes à fleurs cette phase demande une pollinisation qui formera des graines permettant la
colonisation du milieu.
En quoi l’organisation des plantes à fleurs est elle adapté à leur vie fixé à l’interface entre le sol et
l’atmosphère ?
Comment la reproduction des plantes à fleurs est elle organisé en fonction de cette vie fixé ?
I. Organisation des plantes a fleur et vie fixée
1) Morphologie générale d’une plante a fleur
Bilan : Les plantes à fleurs présentent une grande diversité de forme de taille et de milieu de vie : les
plantes dites ligneuse (arbre et arbuste) se distinguent de plantes herbacée par la présence dans
certaines cellules de leurs tiges de lignine qui les rend rigides. Cependant leur appareil végétatif est
organisé de la même façon, il est composé de racines, de tiges et de feuilles dont l’organisation
permet des échanges avec leur milieu et la circulation de matière entre les différents organes.

25
2) Echanges entre la plante et son environnement
Les plantes à fleur on développé des systèmes d’échanges avec l’atmosphère et le sol. Ces dispositifs
présentent une surface importante et une organisation fonctionnelle particulière
a) Les racines et le sol
L’extrémité des racines est recouverte de poils absorbant qui sont des cellules allongée plongeant
dans le sol où elles prélèvent l’eau et les sels minéraux indispensable à la photosynthèse. La densité
et la longueur des poils absorbants ainsi que la ramification des racines peuvent augmenter en cas de
carence minérale dans le sol (voir livre page 98 figure 1)
b) Les feuilles et l’atmosphère
(Voir page 98 doc 3) le limbe des feuilles possède surtout sur sa face inferieur des stomates qui
permettent l’entrée du CO2 atmosphérique nécessaire à la photosynthèse. Ils s’ouvrent sur un tissu
végétal lacuneux qui permet une bonne circulation des gaz dans la feuille. La photosynthèse se
réalise dans toutes les cellules chlorophylliennes, très denses dans la partie supérieure du limbe

26
fortement éclairée. Le nombre et l’orientation des feuilles permettent d’optimisé la réception de
l’énergie lumineuse (voir page 99 doc 3)
Le nombre de stomate ainsi que leur ouverture peuvent varier selon les conditions de
l’environnement.
Remarque : l’indice stomatique, c'est-à-dire le nombre de stomate sur le nombre de cellules peut
être utilisé chez des feuilles fossilisées pour étudier les variations de la teneur de l’atmosphère en
CO2 au cours des temps géologique
3) Circulation de matière dans la plante
Bilan cour : Les substances prélevées dans les milieux extérieurs ou issus de la photosynthèse
permettent d’assuré l’alimentation de la plante. Elles sont distribuées dans l’ensemble de la plante
sous deux formes :
Sève brut (ascendante) qui est une solution diluée d’eau et de sels minéraux absorbé dans le
sol, et qui circulent dans les vaisseaux du xylème
Sève élaboré (descendante) qui contient les molécules organiques fabriqué dans les feuilles
par la photosynthèse et qui circulent dans les vaisseaux du phloème ou tubes criblés
(Voir documents page 100-101) les vaisseaux du xylème et du phloème forment deux réseau
continu dans toute la plante entre les racines et les feuilles dans les quels on retrouve les deux type
de vaisseaux.
L’organisation des feuilles permet d’optimiser la circulation des gaz autour des cellules
chlorophyllienne grâce au parenchyme lacuneux qui communique avec l’atmosphère par les

27
stomates. Le parenchyme palissadique localisé sur la face supérieur assure un bon éclairement des
cellules
4) La protection des plantes à fleurs Anatomique
morphologique biologique Chimique mécanique
Type de plante Lavande Acacia Marronnier
Acacia Vesce des champs
Peuplier Pavot Acacia Menthe
L’oyat Acacia haricot
Type d’agression Herbivores Conditions climatiques Insectes
Herbivore Insectes acariens
Herbivore Insecte Bactéries Champignon
Herbivore Conditions climatiques
Mécanisme de défense
Allongement des épines des acacias Nombreux poils sur les feuilles qui rendent le limbe inaccessible pour les prédateurs Passage de la mauvaise saison grâce aux bourgeons
Symbiose entre acacia et fourmis Les fourmis défendent la plante des agressions avec des piqures douloureuses sur les agresseurs
Acacia : Sécrétion de tanin et l’éthylène sert pour provoquer la sécrétion de tanin chez les autres espèces de type acacia Sécrétion de morphine Sécrétion de menthol
Oyat : Ouverture et fermeture de la plante pour un stockage d’eau au fond d’une crypte très hydrophile et déformable Acacia allongement des épines Haricot : passage de mauvaises saisons sous forme de graine
Bilan cour : En relation avec leur vie fixé les plantes on développé des mécanismes de défense contre
les agressions du milieu. La lutte contre les organismes prédateurs ou pathogènes se fait par des
défenses morpho-anatomique, chimiques et biologiques. Ces défenses peuvent être constitutives
(présente naturellement dans la plante), ou induite (défenses intensifier en cas d’attaque par un
animal prédateur). Certaines plantes développent aussi des mécanismes de défense indirects en
attirant les prédateurs des herbivores.
Les plantes font face aux variations des conditions du milieu en synchronisant leur activité biologique
avec les saisons. Ainsi en automne les arbres entrent en vie ralentie, leur métabolisme et les
échanges avec les milieux sont réduits. Des organes spatialisés assurent le passage de la mauvaise
saison comme par exemple les bougeons qui passent l’hiver protégé par des écailles
Recopier schéma de la page 109 circulations de la matière dans la plante.

28
II. Reproduction des plantes à fleurs et vie fixée
1) L’organisation des fleurs
Activité 4 : fleur de Lisianthus
Bilan cour : Les fleurs présentent une organisation commune en verticilles concentriques :
Le verticille 1 ou calice est constitué par les sépales
Le verticille 2 ou corole est constitué par les pétales
Le ou les verticilles 3 sont constitués par les étamines
Le verticille 4 est constitué par le pistil
Certaines pièces florales sont directement impliquer dans la reproduction : le pistil est l’organe
femelle contenant les ovules, les étamines sont les organes males dont les anthères contiennent les
grains de pollens. La mise en place des pièces florales se réalise sons l’action de gènes du
développement, la mutation de l’un de ces gènes entrainant la formation d’une fleur anormale (voir
page 103 doc C et D)
2) De la fleur au fruit : la pollinisation
Bilan : La majorité des fleurs produisent à la foi des gamètes males et des gamètes femelles. La
pollinisation peut avoir lieu à l’intérieur de la même fleur (autopollinisation) ou entre fleur
différentes (pollinisation croisée). Dans ce dernier cas les grains de pollens produit par une fleur se
dépose sur le stigmate du pistil d’une autre fleur de la même espèce.
Les fleurs pollinisées par les insectes émettent différents signaux qui attirent les
pollinisateurs. Leurs grains de pollen sont assez gros et ornementé, ce qui facilite leur
adhésion au corps des insectes qui visitent les fleurs. Ils y trouvent de la nourriture sous
forme de pollen et de nectar. Le pollen prélevé sur une fleur peut alors être déposé sur une
autre fleur visitée.
On peut observée une coévolution entre l’organisation de ses plantes et les organes des
insectes permettant de repéré les fleurs et de s’y nourrir (ex page 104-105 doc 2) ceci
procure un avantage sélectif pour la plante qui se reproduit plus efficacement, et pour
l’insecte qui y trouve des ressources alimentaires. Ce phénomène est particulièrement
remarquable pour les plantes pollinisées par un insecte précis comme le couple
orchidée/sphinx
Les fleurs pollinisées par le vent sont généralement de petites tailles et produisent en grande
quantité du pollen dont les grains sont petits et lisses. Leurs étamines et leur pistils sont bien
exposé au vent (exemple les graminées ou poacées)
La pollinisation croisée favorise le brassage génétique au sein de l’espèce (voir thème 1A)
Une fois les grains de pollens posé sur le stigmate, un afflux d’eau leur permet de germée : chaque
grain développe un tube pollinique qui grandi dans le style jusqu’au ovules. Le gamète mal contenu

29
dans le tube féconde alors le gamète femelle contenu dans un ovule (voir page 106-107). Apres la
fécondation l’ovaire devient un fruit chaque ovule fécondé devient une graine.
3) La dispersion des graines Bilan : Celle-ci permet la pérennité de l’espèce et la colonisation par la plante de nouveaux
territoires. Elle peut être assuré par le vent ou par les animaux (voir page 107 doc 3). Les graines
dispersé par le vent sont petites et légères et peuvent posséder des dispositifs qui améliorent leurs
portance (exemple pissenlit). Les graines dispersées par les animaux sont généralement dans des
fruits charnus, coloré et sucré, attractif pour les animaux. Ceux-ci les mangent et regrettent les
graines par exemples dans leurs excréments : c’est une dispersion active. D’autres graines sont dans
des fruits qui adhèrent au pelage : c’est une dispersion passive.
Le premier cas montre encore une collaboration entre la plante qui est dispersé et l’animal qui se
nourri. On observe aussi dans certains cas que les graines germent mieux après passage dans le tube
digestif d’un animal et/ou lorsqu’elles germent loin de leur plante mère. Cette collaboration est aussi
le résultat d’une coévolution. Conclusion générale
Conclusion du chapitre :
Au cours de leur évolution, les plantes ont mis au point de nombreux dispositifs leur
permettant de réalisé les diverses fonction des êtres vivants tout en menant une vie fixé.
Elles ont parfois utilisé l’aide des animaux avec lesquels elles ont établis des relations à
bénéfices réciproques grâce à une longue coévolution. Voir bilan du livre page 111.