Research Article The Macrophage Galactose-Type Lectin-1 (MGL1) Recognizes Taenia...
17
Research Article The Macrophage Galactose-Type Lectin-1 (MGL1) Recognizes Taenia crassiceps Antigens, Triggers Intracellular Signaling, and Is Critical for Resistance to This Infection Daniel Montero-Barrera, 1 Héctor Valderrama-Carvajal, 1 César A. Terrazas, 2 Saúl Rojas-Hernández, 3 Yadira Ledesma-Soto, 1 Laura Vera-Arias, 1 Maricela Carrasco-Yépez, 4 Lorena Gómez-García, 5 Diana Martínez-Saucedo, 1 Mireya Becerra-Díaz, 1 and Luis I. Terrazas 1 1 Unidad de Biomedicina, Facultad de Estudios Superiores-Iztacala, Universidad Nacional Aut´ onoma de M´ exico (UNAM), Avenue de los Barrios 1, Los Reyes Iztacala, 54090 Tlalnepantla, MEX, Mexico 2 Department of Pathology, e Ohio State University Medical Center, Columbus, OH, USA 3 Secci´ on de Estudios de Postgrado e Investigaci´ on, Escuela Superior de Medicina, Instituto Polit´ ecnico Nacional, Mexico 4 Proyecto CyMA, UIICSE, FES Iztacala, UNAM, 54090 Tlalnepantla, MEX, Mexico 5 Instituto Nacional de Cardiolog´ ıa “Ignacio Ch´ avez,” 14080 Mexico, DF, Mexico Correspondence should be addressed to Luis I. Terrazas; [email protected] Received 9 August 2014; Revised 14 October 2014; Accepted 15 October 2014 Academic Editor: Abraham Landa Copyright © 2015 Daniel Montero-Barrera et al. is is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited. C-type lectins are multifunctional sugar-binding molecules expressed on dendritic cells (DCs) and macrophages that internalize antigens for processing and presentation. Macrophage galactose-type lectin 1 (MGL1) recognizes glycoconjugates expressing Lewis X structures which contain galactose residues, and it is selectively expressed on immature DCs and macrophages. Helminth parasites contain large amounts of glycosylated components, which play a role in the immune regulation induced by such infections. Macrophages from MGL1 −/− mice showed less binding ability toward parasite antigens than their wild-type (WT) counterparts. Exposure of WT macrophages to T. crassiceps antigens triggered tyrosine phosphorylation signaling activity, which was diminished in MGL1 −/− macrophages. Following T. crassiceps infection, MGL1 −/− mice failed to produce significant levels of inflammatory cytokines early in the infection compared to WT mice. In contrast, MGL1 −/− mice developed a 2-dominant immune response that was associated with significantly higher parasite loads, whereas WT mice were resistant. Flow cytometry and RT-PCR analyses showed overexpression of the mannose receptors, IL-4R, PDL2, arginase-1, Ym1, and RELM- on MGL1 −/− macrophages. ese studies indicate that MGL1 is involved in T. crassiceps recognition and subsequent innate immune activation and resistance. 1. Introduction Pattern recognition receptors (PRRs) function to rapidly detect pathogen invasion as well as to control innate immune activation leading to inflammation. e PRRs of dendritic cells (DCs) and macrophages (Ms), among others, recognize and bind to conserved pathogen motifs such as LPS in gram- negative bacteria, flagellin, viral RNA, and several molecules in intracellular protozoa [1], including cyclophilin from Toxoplasma gondii [2] and GIPL from Trypanosoma cruzi [3]. Among the PRRs, the Toll-like receptor (TLR) family is the main group of receptors known to be involved in maturation and inducing inflammatory cytokines in DCs and Ms[1]. Other types of PRRs are the C-type lectin receptors (CLRs), which have selective affinity for a carbohydrate or group of carbohydrates in a Ca 2+ -dependent manner; CLRs also play a role in recognizing different pathogens, including bacteria, fungal, and viral glycoconjugates [4–7]. However, there is a less-defined function for CLRs in response to helminth infections. Hindawi Publishing Corporation BioMed Research International Volume 2015, Article ID 615865, 16 pages http://dx.doi.org/10.1155/2015/615865
Transcript of Research Article The Macrophage Galactose-Type Lectin-1 (MGL1) Recognizes Taenia...

Research ArticleThe Macrophage Galactose-Type Lectin-1 (MGL1)Recognizes Taenia crassiceps Antigens Triggers IntracellularSignaling and Is Critical for Resistance to This Infection
Daniel Montero-Barrera1 Heacutector Valderrama-Carvajal1 Ceacutesar A Terrazas2
Sauacutel Rojas-Hernaacutendez3 Yadira Ledesma-Soto1 Laura Vera-Arias1
Maricela Carrasco-Yeacutepez4 Lorena Goacutemez-Garciacutea5 Diana Martiacutenez-Saucedo1
Mireya Becerra-Diacuteaz1 and Luis I Terrazas1
1Unidad de Biomedicina Facultad de Estudios Superiores-Iztacala Universidad Nacional Autonoma de Mexico (UNAM)Avenue de los Barrios 1 Los Reyes Iztacala 54090 Tlalnepantla MEX Mexico2Department of Pathology The Ohio State University Medical Center Columbus OH USA3Seccion de Estudios de Postgrado e Investigacion Escuela Superior de Medicina Instituto Politecnico Nacional Mexico4Proyecto CyMA UIICSE FES Iztacala UNAM 54090 Tlalnepantla MEX Mexico5Instituto Nacional de Cardiologıa ldquoIgnacio Chavezrdquo 14080 Mexico DF Mexico
Correspondence should be addressed to Luis I Terrazas literrazascampusiztacalaunammx
Received 9 August 2014 Revised 14 October 2014 Accepted 15 October 2014
Academic Editor Abraham Landa
Copyright copy 2015 Daniel Montero-Barrera et alThis is an open access article distributed under the Creative CommonsAttributionLicense which permits unrestricted use distribution and reproduction in anymedium provided the originalwork is properly cited
C-type lectins are multifunctional sugar-binding molecules expressed on dendritic cells (DCs) and macrophages that internalizeantigens for processing and presentation Macrophage galactose-type lectin 1 (MGL1) recognizes glycoconjugates expressing LewisX structureswhich contain galactose residues and it is selectively expressed on immatureDCs andmacrophagesHelminth parasitescontain large amounts of glycosylated components which play a role in the immune regulation induced by such infectionsMacrophages from MGL1minusminus mice showed less binding ability toward parasite antigens than their wild-type (WT) counterpartsExposure ofWTmacrophages to T crassiceps antigens triggered tyrosine phosphorylation signaling activity which was diminishedin MGL1minusminus macrophages Following T crassiceps infection MGL1minusminus mice failed to produce significant levels of inflammatorycytokines early in the infection compared to WT mice In contrast MGL1minusminus mice developed a Th2-dominant immune responsethat was associated with significantly higher parasite loads whereasWTmice were resistant Flow cytometry and RT-PCR analysesshowed overexpression of the mannose receptors IL-4R120572 PDL2 arginase-1 Ym1 and RELM-120572 on MGL1minusminus macrophages Thesestudies indicate that MGL1 is involved in T crassiceps recognition and subsequent innate immune activation and resistance
1 Introduction
Pattern recognition receptors (PRRs) function to rapidlydetect pathogen invasion as well as to control innate immuneactivation leading to inflammation The PRRs of dendriticcells (DCs) andmacrophages (M120601s) amongothers recognizeand bind to conserved pathogen motifs such as LPS in gram-negative bacteria flagellin viral RNA and several moleculesin intracellular protozoa [1] including cyclophilin fromToxoplasma gondii [2] andGIPL fromTrypanosoma cruzi [3]
Among the PRRs the Toll-like receptor (TLR) family is themain group of receptors known to be involved in maturationand inducing inflammatory cytokines in DCs and M120601s [1]Other types of PRRs are the C-type lectin receptors (CLRs)which have selective affinity for a carbohydrate or group ofcarbohydrates in a Ca2+-dependent manner CLRs also playa role in recognizing different pathogens including bacteriafungal and viral glycoconjugates [4ndash7] However there is aless-defined function for CLRs in response to helminthinfections
Hindawi Publishing CorporationBioMed Research InternationalVolume 2015 Article ID 615865 16 pageshttpdxdoiorg1011552015615865
2 BioMed Research International
It has been largely demonstrated that helminths displaya variety of glycan moieties on their antigens which mayparticipate in the induction of Th2 responses [8 9] and inimmune downregulation both characteristics of helminthinfections [10] Nevertheless given the complexity of thesemolecules it may also be expected that some helminthglycoconjugates display immunostimulatory activities [11]
Some helminth parasites have been reported to be recog-nized by different CLRs for example dendritic cell-specificICAM-3 grabbing integrin (DC-SIGN) mannose receptor(MR) macrophage galactose-type lectin (MGL) and SIGN-related 1 (SIGNR1) bind different glycans on soluble eggantigens from Schistosoma mansoni in vitro [12 13] Trichurismuris a nematode is also recognized by the MR but suchCLRs appear to play a limited role in protection or suscep-tibility in both parasitic infections given that mice lackingthe MR or SIGNR1 mount a normal immune response andclear these infections as usual [14]
Taenia crassiceps is a cestode that is useful for understand-ing the host-parasite relationship in cysticercosis Glycocon-jugates of T crassiceps and other cestodes have been shown toinduce strongTh2-biased responses in vivo [15] and tomodu-late both human and mouse DC activity in vitro [16ndash18] Wehave previously reported that most of the T crassiceps anti-gens bind to concanavalin A indicating that they are glycosy-lated with glucose mannose or galactose [15 17 19]The nat-ural candidates for such carbohydrate recognition are CLRssuch as the MR MGL and DC-SIGN As mentioned beforeDC-SIGN and the MR have been shown to be irrelevant forSchistosoma orTrichuris protectionWe therefore focused ourstudy onMGL1MGL1 ismainly found onM120601s and immatureDCs although it was recently reported to be expressed onmast cells [4] and it recognizes glycoconjugates expressinggalactose and N-acetyl-galactosamine residues Because theability of MGL1 to act as an autonomous receptor has beenappreciated only recently the molecular regulation of its sig-naling pathway is partly known [20] but its biological role inresponse to helminth infections is still unrevealed Recentlyit has been shown that upon strong MGL engagementwith polyclonal anti-MGL antibodies this receptor is ableto signal through tyrosine phosphorylation and induce ERKsignaling [20] however whether MGL signals after a ldquonatu-ralrdquo ligation is also unknown
To clarify the role ofMGL1 in the recognition and defenseagainst a helminth infection we infected wild-type andMGL1-deficient mice (MGL1minusminus) with Taenia crassiceps Ourdata demonstrated that MGL1-deficient mice exhibited redu-ced parasite clearance despite strong induction of Th2 res-ponses MGL1minusminus macrophages displayed reduced intracellu-lar signaling in response to Taenia antigens and lower TNF-120572 and NO production Taken together our data suggest thatMGL1 plays a key role in driving macrophage responses invivo and thus may act as an important mediator of resistanceto this helminth infection
2 Materials and Methods
21 Mice Six- to eight-week-old female MGL1minusminus mice on aC57BL6 background were donated by Glycomics Consor-tium (USA) MGL1minusminus mice have been backcrossed for more
than 7 generations on a C57BL6NHsd genetic backgroundwith mice from Harlan Laboratories (Mexico) In someexperiments BALBc mice were purchased from Harlan Lab-oratories (Mexico) All mice were maintained in a pathogen-free environment at the FES-Iztacala UNAM animal facili-ties according to the Faculty Animal Care and Use Commit-tee and government guidelines (official Mexican regulationNOM-062-ZOO-1999) which are in strict accordance withthe recommendations in the Guide for the Care and Useof Laboratory Animals of the National Institutes of HealthUSA
22 Parasites and Infection Metacestodes of T crassicepswere harvested from the peritoneal cavity of female BALBcmice after 2 to 3 months of infection The cysticerci werewashed four times in sterile phosphate-buffered saline (PBS)(015M pH 72) Experimental infection was achieved byintraperitoneal (ip) injectionwith 20 small (approx 2mm indiameter) nonbudding cysticerci ofT crassiceps suspended in03mL of PBS per mouse
23 Lectin-Blot Analysis on T crassiceps Soluble Antigens Tcrassicepswas lysed by sonicationwith one 10-s pulse at 100Wof amplitude (Fisher Sonic Dismembrator model 300) Theresulting suspension was centrifuged at 4∘C for 1 h at 2700 gand the pellet was discarded and we keep the supernatantSoluble antigen was concentrated and quantified by theBradford method A total of 20120583g of protein of T crassicepssoluble antigens (TcSol) was separated by SDS-PAGE (10)and electroblotted (400mA for 1 h) onto a nitrocellulosemembrane [21]
For the detection of carbohydrates with residues of gala-ctose or N-acetylgalactosamine strips of nitrocellulose wereblocked with 1 BSA diluted in PBS-T (10mM sodium phos-phate buffer 150mM NaCl and 05 Tween 20) and incu-bated overnight at 4∘C The membranes were treated with abiotinylated lectin (Hp or RCA) at a concentration of 5120583gmLdiluted 1 500 in PBS-T and incubated overnight at 4∘C(Sigma-Aldrich St LouisMOUSA)Thenext day themem-branes were incubated with SBHP the strips were washedwith PBS and enzyme activities were then detected by asubstrate solution (01 H
2
O2
175 methanol 015 4-chloro-120572-naphthol and 825 PBS) following a 15-min incu-bation The proteins that reacted with the lectins wereidentified on the nitrocellulose strips
24 FITC Labeling of T crassiceps Antigens T crassiceps solu-ble products (TcSol) were labeled with FITC (Sigma) accord-ing to the manufacturerrsquos instructions Briefly 1mgmL TcSolwas prepared in 01M sodium bicarbonate buffer pH 9 FITCwas dissolved in DMSO at 1mgmL and 500120583L of the FITCsolutionwas slowly added to TcSol in 5120583L aliquots while gen-tly stirring The TcSol-FITC solution was incubated for 8 h at4∘C in the dark and washed 3 times with PBS protein wasquantified by the Bradford assay and maintained at 4∘C
25 Cell Preparations and Culture Conditions The spleen wasremoved from infected mice under sterile conditions Single-cell suspensions were prepared by gently teasing apart the
BioMed Research International 3
spleen in RPMI-1640 supplemented with 10 fetal bovineserum 100 units of penicillinstreptomycin 2mMglutamine25mM HEPES buffer and 1 nonessential amino acids (allfrom GIBCO BRL Grand Island New York) The cells werecentrifuged and the erythrocytes were lysed by resuspendingthe cells in Boylersquos solution (017M Tris and 016M ammo-nium chloride) Following two washes the viable cells werecounted by trypan blue exclusion with a Neubauer hemocy-tometer and the splenocytes were adjusted to 3times 106 cellsmLin the same medium Aliquots (100 120583L) of the adjusted cellsuspensions were placed into 96-well flat bottom cultureplates (Costar Cambridge Massachusetts) and stimulatedwith a soluble extract ofT crassiceps (25120583gmL) orwith plate-bound anti-CD3 antibody (1 120583gmL) at 37∘C for 72 h Thesupernatants from these cultures were analyzed for IL-4 andIFN-120574 (PeproTech Mexico) production by ELISA
26 Cytokine and Nitric Oxide Production by PeritonealMacrophages Peritoneal exudate cells (PECs) were obtainedfrom uninfected thioglycolate-injected mice and from micefollowing 2 4 and 8 weeks of T crassiceps infection PECswere adjusted to 5 times 106mL in supplemented RPMI andplated in 6-well plates (Costar) After 2 hours at 37∘C and 5CO2
nonadherent cells were removed and adherent cellswere gently scrapped using cold PBS and readjusted to 1 times106mL Viability at this point was gt90 These cells weregt90 macrophages according to FACS analysis (F480+BioLegend SanDiego CA)Then 1mL of the cell suspensionwas plated and cells were activated in 24-well plates (Costar)with LPS (1120583gmL E coli 111B4 Sigma St Louis MO) fol-lowed by incubation for 24 h TNF-120572 IL-12 (PeproTech) andnitric oxide (Griess reaction) were examined in supernatants
27 Generation of Bone Marrow Derived M120601s (BMDM120601s)BMDM120601s fromWTorMGL1minusminusmicewere obtained as previ-ously described [22] Briefly bone marrow cells were isolatedby flushing femurs and tibias with culture media Bonemarrow cells were plated at 1 times 106 cellsmL in medium sup-plemented RPMI medium supplemented with 10 SFB andpenicillinstreptomycin plus 20 ngmL murine recombinantmacrophage colony-stimulating factor (M-CSF PeprotechMexico)Onday 3 freshmedia containingM-CSFwas addedOn day 7 nonadherent cells were removed and adherent cellswere detached and centrifuged at 1500 rpm at 4∘C for 15minAdherent cells (F480+) were resuspended in freshmedia andcultured for an additional 24 h in 12-well plates
28 In Vitro Stimulation of BMDM120601s and Protein ExtractionBone marrow cells were differentiated as described aboveOn day 7 the M120601s were harvested and plated at 25 times 106cellsmL in six-well plates The cells were used 24 hours afterplating to eliminate any residual effects from M-CSF M120601swere stimulatedwith theTcSol (50120583gmL) for 5 15 or 30minAfter stimulation the cells were centrifuged at 1500 rpm for5min and washed with PBS The cells were then lysed withlysis buffer for 15min and the lysates were quantified usingthe BCA assay (Thermo Scientific) and then frozen at minus80∘Cuntil further use
29 Western Blot Assays Whole-cell lysates were resolved bySDS-PAGE (40 120583g of protein was loaded into each well) using10 acrylamide mini-gels followed by electrophoretic trans-fer to PVDF membranes (Immobilon-P MILLIPORE) for2 hMembraneswere blockedwith 5 fat-freemilk in PBS for2 h and incubated with primary antibodies overnight Thedetection step was performed with peroxidase-coupled anti-mouse IgG and anti-rabbit IgG (BioLegend 1 2000) andanti-goat IgG (Santa Cruz Biotechnology) for 2 h The pri-mary antibodies included antiphosphotyrosine (Santa CruzBiotechnology) and 120573-Actin (BioLegend) All primary anti-bodies were diluted 1 500 in 1 fat-free milk in PBS Blotswere developed with the ECL detection system according tothemanufacturerrsquos instructions (Amersham) Blots are repre-sentative of two separate experiments
210 Detection of Nitric Oxide Production Nitric oxide pro-duction by macrophages was assayed by determining theincrease in nitrite concentration [23] by the Griess reactionadapted to microwell plates (Costar) Briefly 50120583L of culturesupernatant was mixed with an equal volume of Griessreagent and incubated for 10min at room temperature in thedark the absorbancewasmeasured at 570 nm in an automaticmicroplate reader (Organon Technika Microwell System)Values were quantified using serial dilutions of sodiumnitrite
211 Flow Cytometry Analysis Peritoneal exudate cells(PECs) were obtained from naıve or infected mice by inject-ing 10mL of ice-cold PBS For flow cytometry single cellsuspensions of the PECs obtained during the sacrifice werestained with anti-F480 anti-PDL2 anti-IL-4R120572 anti-MGL1and anti-MGL2 antibodies (Biolegend San Diego CA) for30min at 4∘CThe cells werewashed twice and analyzed usingthe FACSCalibur system and Cell Quest software (BectonDickinson USA)
212 Reverse Transcription (RT)-PCR Analyses The levels ofarginase 1 (Arg-1) Ym-1 and resistin-likemolecule-120572 (RELM-120572) mRNA transcripts in adherent peritoneal macrophages(which were allowed to adhere for 2 h at 37∘C and 5 CO
2
)were determined using RT-PCRAt the indicated time pointsadherent peritoneal macrophages from T crassiceps-infectedMGL1++ and MGL1 minusminus mice were aseptically removed andwithout any further stimulation (basal) or LPS + IFN-120574-stimulation (1 120583g and 20 ngmL resp) were processed forRNA extraction using TRIzol reagent (Invitrogen CarlsbadCA USA) and the propanol-chloroform techniqueThe RNAwas quantified and 3120583g of RNA was reverse transcribedusing the Superscript II First Strand Synthesis Kit (Invit-rogen) and an oligo dT primer as recommended by themanufacturer Once cDNA was obtained conventional PCRwas performed The PCR reactions contained (in a 25 120583Lfinal volume) 5X PCR Buffer blue 10mM dNTP 40 nM eachof the forward and reverse primers (Table 1) 1 U Taq DNApolymerase (Sacace Biotechnologies Italy) and 2 120583L of thecDNA The program used for the amplification of each genecontained an initial denaturation step at 95∘C for 5min 35
4 BioMed Research International
Table 1 The following primer pairs were used in this study
Gen Sequence Melting temperature Number of cycles
GAPDH Forward -CTCATGACCACAGTCCATGC-Reverse -CACATTGGGGGTAGGAACAC- 54∘C 35
Arg-1 Forward -CAGAAGAATGGAAGAGTCAG-Reverse -CAGATATGCAGGGAGTCACC- 54∘C 35
Ym-1 Forward -TCACAGGTCTGGCAATTCTTCTG-Reverse -TTTGTCCTTAGGAGGGCTTCCTC- 56∘C 35
Fizz-1 Forward -GGTCCCAGTGCATATGGATGAGACCATAG-Reverse -CACCTCTTCACTCGAGGGACAGTTGGCAGC- 65∘C 35
iNOS Forward -GCCACCAACAATGGCAACAT-Reverse -AAGACCAGAGGCAGCACATC- 60∘C 30
cycles of 95∘C for 40 s the indicated melting temperaturefor 50 s and 72∘C for 40 s and a final extension step at 72∘Cfor 4min All reactions were performed in a thermal cycler(Corbett Research Australia) Finally to observe the ampli-fied products a 15 agarose gel was prepared and sampleswere loaded with blue juice buffer containing SYBR Green(Invitrogen) The gels were visualized using a Fujifilm FLA5000 scanner (Fuji Japan) with FLA 5000 image reader V21software to capture the imagesThe specificity of the PCRwasverified by the absence of signal in the no-template controls ofmacrophage samplesThe sequences of the primers used havebeen previously reported [24]
213 Antibody ELISAs Peripheral blood was collected at 2-wk intervals from tails snips of the T crassiceps-infectedMGL1++ and MGL1minusminus mice T crassiceps-specific IgG1 andIgG2a levels were determined by ELISA as previously des-cribed The results are expressed as the maximal serum dilu-tion (endpoint titer) at which Optical Density was detected(OD) Total IgE production was detected by Opt-ELISA fromBiolegend
214 Statistical Analysis Comparisons between thewild-typeand MGL1minusminus groups in this study were performed usingStudentrsquos unpaired 119905 test 119875 lt 005was considered significantThe statistical significance of the serum titers was deter-mined by nonparametric tests using the Mann-Whitney119880Wilcoxon Rank tests
3 Results
31 Taenia crassiceps Soluble Antigens Express GlycoconjugatesContaining N-Acetylgalactosamine and Galactose ResiduesTo detect the expression of glycoconjugates on soluble anti-gens of T crassiceps a lectin-blot assay was performed usingHelix pomatia (specific for N-acetylgalactosamine residues)and Ricinus comunis (specific for galactose residues) lectinsSeveral glycoconjugates were recognized by both lectinsthose of 310 287 250 210 182 161 79 77 75 49 47 and 41 kDa(Figure 1(a)) whereas two bands at 51 and 39KDa were onlyrecognized by H pomatia However using only streptavidinperoxidase (Figure 1(b)) two glycoconjugates were recog-nized on TcSol antigens those of 250 and 124 kDaTherefore
49
80
124
209
35
124
250
797775
514947
4139
287310
210182161
124
250
310
210182161
797775
4947
41
287
RCAHPMW
(a)
250
124
SBHP
(b)
Figure 1 Lectin-blot analysis of Taenia crassiceps soluble antigensT crassiceps soluble proteins were subjected to electrophoresison a 10 SDS-polyacrylamide gel transferred to nitrocellulosemembrane and revealed with a couple of different biotinylatedlectins (a) MW-molecular weight HP-Helix pomatia and RCA-Ricinus communis (b) The strips were incubated only with SBHP-streptavidin-bound horseradish peroxidase
the presence of these two biotin-containing proteins mostlikely corresponds to biotin-dependent carboxylases recog-nized by streptavidin alone [25 26] These findings indicatethat TcSol antigens express high levels of glycoconjugateswith N-acetylgalactosamine and galactose residues
32 MGL1minusminus Macrophages Display Reduced Binding to Tae-nia crassiceps Soluble Antigens The innate recognition ofpathogen-associated molecules by antigen-presenting cells iscrucial to the initiation of the immune response Differentstudies have shown that helminth-derived molecules can berecognized by C-type lectins [27 28] In our model wepreviously reported that most TcSol bound to concanavalin
BioMed Research International 5
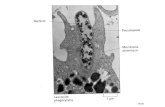
A indicating that TcSol are glycosylated with glucose man-nose or galactose [17] whereas Jang Lee et al reported thatthe main N-glycan structures in T crassiceps were mannosefucose galactose and GlcNAc They also found a rare Fuc1205721rarr 3GlcNAc antenna on T crassiceps molecules [29] Thusthe natural candidates for carbohydrate recognition on Tcrassiceps are CLRs such as the MR MGL and DC-SIGNThus we hypothesized that one of the possible receptors thatbind the glycomolecules on TcSol may be MGL given itsspecificity to recognize both N-acetylgalactosamine andgalactose residues To investigate the role of MGL1 in TcSolrecognition TcSol antigens were labeled with FITC toperform binding assays TcSol-FITC efficiently bound toMGL1++ macrophages at 37∘C (approximately 30 of cellswere positive) indicating that TcSol-FITC can be recognizedand internalized by these cells (Figures 2(a) and 2(b)) Incu-bation of M120601s and TcSol at 4∘C showed reduced fluorescence(5ndash10) indicating a low level of binding Next we similarlyexposed MGL1minusminus M120601s to TcSol-FITC at 37∘C and foundsignificant reductions in both the percentage of TcSol-FITC-positive cells (approximately 10) and the mean fluorescenceintensity (MFI) of TcSol-FITC on these cells (Figure 2(c))These data suggest that M120601s can recognize TcSol via MGL
33 The Absence of MGL1 on Macrophages Impairs TcSol-Triggered Global Tyrosine Phosphorylation The fact thatTcSol binding was impaired in MGL1minusminusM120601s led us to ques-tion whether the interaction with MGL-TcSol triggers anintracellular signaling pathway Recent advances in theunderstanding of the intracellular signaling induced bystrong antibody-mediated crosslinking of MGL have high-lighted a possible role for the ERK pathway [20] To deter-mine whether TcSol can induce intracellular signaling weexposed BMDM120601s fromMGL1++ or MGL1minusminus mice to TcSolfor 5 15 and 30min and found that tyrosine phosphorylationwas enhanced in MGL1++ macrophages interestingly tyro-sine phosphorylation was detected as early as 5min andremained for at least 30min (Figure 3(a)) In contrastMGL1minusminus BMDM120601s that were similarly exposed to TcSol dis-played a lower and less-sustained tyrosine phosphorylationin several protein bands (Figure 3(a) arrows) To further ana-lyze this response we stimulated bothMGL1++ andMGL1minusminusBMDM120601s with LPS (500 ngmL) or TcSol again for 30minAs observed in Figure 3(b) alsowedetected a lack of responseto LPS and a lower tyrosine phosphorylation in response toTcSol (Figures 3(b) and 3(c) resp) in MGL1minusminus BMDM120601s
34 Mice Lacking MGL1 Are Highly Susceptible to Taenia cras-siceps Infection After determining that T crassiceps antigenscan be recognized by MGL1 and that macrophages deficientin MGL1 are partially refractory to TcSol stimulation weinvestigated the in vivo role of this molecule in experimentalcysticercosis using MGL-deficient mice To approach thisquestion we compared the course of T crassiceps infection inMGL1minusminus with that in MGL1++ mice As a control for sus-ceptibility we used BALBc mice which have consistently
been reported as a highly susceptible strain to T crassicepsinfection [30]
We examined the kinetics of parasite growth from 2 to 8weeks after T crassiceps infection Early in the infection (2weeks) both groups of mice displayed comparable parasiteburdens (Figure 4(a)) Interestingly as infection progressedby the fourth week of infection the number of larvae in theperitoneal cavity increased significantly in MGL1minusminus micecompared to MGL1++ mice which successfully reduced thenumber of parasites by week 8 after infection (Figures 4(a)and 4(b)) In fact MGL1minusminus mice exhibited a parasite burdenthat was very similar to that in the susceptible strain of mice(BALBc) which were consistently observed as very suscepti-ble toT crassiceps infection (Figure 4(a))These findings sug-gest that theMGL1-mediated signaling pathway is involved inresistance during T crassiceps infection on a resistant geneticbackground such as C57BL6
35 MGL1minusminus Mice Display an Altered Immune Responseto Taenia crassiceps A few studies have demonstrated thatMGL can mediate signaling after a strong stimulus such aspolyclonal anti-MGL antibodies but its role in modulatingprotective immunity against helminth parasites is unknownTherefore wemeasured levels ofTh1-associated IgG2a as wellas Th2-associated IgG1 and total IgE antibodies in MGL1minusminus
andMGL1++mice at different timepoints following infectionwith T crassiceps
Early in infection T crassiceps-infected MGL1++ andMGL1minusminus mice displayed comparable levels of T crassicepsAg-specific Th1-associated IgG2a antibodies (data notshown) However by week 8 after infection MGL1++ micedisplayed significantly higher titers of specific IgG2aantibodies againstT crassiceps antigens (Figure 5(a)) By con-trast no clear differences were observed in Th2-associatedIgG1 production MGL1++ mice displayed high titers ofanti-T crassiceps-specific IgG1 that were similar to thosein MGL1minusminus mice at week 8 after infection (Figure 5(b))AlthoughTh2-associated IgE has been shown to play a role inmediating immunity against certain helminths we found thatT crassiceps-infected MGL1++ mice harbored a lower para-site burden despite producing significantly lower levels ofIgE compared to similarly infected MGL1minusminus mice whichdisplayed higher levels of total IgE (Figure 5(c))
Additionally we also compared the cytokine produc-tion by splenocytes from these mice in response to either25 120583gmL TcSol or 1 120583gmL plate-bound anti-CD3 antibodyAnti-CD3 (data not shown) or TcSol-stimulated splenocytesfrom both strains produced similar levels of IFN-120574 at 2 wkafter infection (Figure 6(a)) However 4 and 8 weeks afterinfection the IFN-120574 production by the spleen cells ofMGL1minusminus mice decreased and did not reach the level pro-duced by MGL1++ splenocytes (Figure 6(a)) Similarly asearly as 2 wk after infection the splenocytes from theMGL1minusminus and MGL1++ mice produced comparable levels ofIL-4 (Figure 6(b)) However by weeks 4 and 8 after infectionMGL1minusminus mice produced significantly more IL-4 than the
6 BioMed Research International
F480 F480
TcSo
l-FIT
C
TcSo
l-FIT
C
100 101 102 103 104 100 101 102 103 104
29 6
100
101
102
103
104
100
101
102
103
104MGL1++ 37∘C MGL1minusminus 37∘C
(a)
TcSo
l-FIT
C (
)
lowast
50
40
30
20
10
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(b)
lowast
MFI
500
400
300
200
100
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(c)
Figure 2 Peritoneal M120601s recognize Taenia crassiceps soluble products through MGL1 M120601s from naıve MGL1++ or MGL1minusminus mice wereincubated with fluorescently labeled TcSol and fluorescence was analyzed by flow cytometry (a) Representative dot blots indicate M120601s fromeither MGL1++ or MGL1minusminus mice incubated for 30 minutes with TcSol at 37∘C (b) Percentage of TcSol-FITC positive naıve peritoneal cellsexposed for 30min (c) Data are represented as the mean florescence intensity (MFI) of naıve M120601s exposed to TcES-FITC Basal levels ofautofluorescence were subtracted from all treatments Data are representative of three independent experiments lowast119875 lt 005
MGL1++ splenocytes in response to anti-CD3 whereasantigen-specific higher IL-4 production in MGL1minusminus spleno-cytes was only observed at week 4 after infection comparedwithMGL1++ cells (Figures 6(b) and 6(c))Thus as the infec-tion progressed splenocytes from MGL1minusminus mice producedsignificantly greater levels of IL-4 compared with splenocytes
fromT crassiceps-infectedMGL1++mice in response to anti-CD3
36 Differential Cytokine Production in MGL1minusminus andMGL1++ Peritoneal Macrophages To determine whetherMGL1minusminus mice had a systemic defect in innate activation we
BioMed Research International 7
TcSol
170130
9870
55
42
35
MW
50120583gmL5998400 15998400 30998400 5998400 15998400 30998400
MGL1++ MGL1minusminus
minusminus
(a)
170130
98705542
35
25
TcSolLPS 30998400
3099840030998400 30998400
MGL1++MGL1minusminus
120573-Actin
minusminus
minus
minus minus
minus minusminus
(b)
MGL1++MGL1minusminus
Relat
ive p
hosp
hory
latio
n le
vel
TcSolLPS 30998400
30998400
08
06
04
02
00
3099840030998400minus
minus
minus
minus minus
minus minus
minus
P-Y LPS and TcSol
(c)
Figure 3 MGL1minusminus macrophages display deficient tyrosine phosphorylation in response to TcSol (a) MGL1++ and MGL1minusminus bone marrow-derived M120601 were synchronized in starvation conditions over night and then were stimulated or not with TcSol (50120583gmL) for 5 15 or30min Next total cell extracts were obtained and resolved by electrophoresis transferred to nitrocellulose and probed with monoclonalantiphosphotyrosine antibody Lane 1 serum-starved MGL1++ M120601s Lanes 2ndash4 serum-starved MGL1++ M120601s following TcSol exposure for5 15 and 30min respectively Lane 5 serum-starved MGL1minusminus M120601s Lanes 6ndash8 serum-starved MGL1minusminus M120601s following TcSol exposure for5 15 and 30min respectively Lanes 2ndash4 show an upregulation of the tyrosine phosphorylation of total proteins which was sustained forat least 30min (arrows) Lanes 6ndash8 show an unsustained tyrosine phosphorylation of proteins after similar TcSol stimulation of MGL1minusminusM120601s (b) Similar experiment using LPS (500 ngmL) andor TcSol stimulation for 30min (c) Densitometry of western blot placed in (b)Proteins were visualized using a goat anti-mouse secondary conjugated to HRP and a chemiluminescence detection system Western blotsare representative of two independent experiments
examined the responsiveness of macrophages fromMGL1minusminusmice to proinflammatory stimuli Macrophages were isolatedfrom the peritoneal cavities of both strains of infected miceand either left unstimulated (basal) or stimulated for 24 hwith LPS (1120583gmL) and IFN-120574 (20 ngmL) The supernatantswere collected and analyzed for IL-12 TNF-120572 and NOproduction As shown in Figure 7 macrophages fromMGL1minusminus mice obtained during the early phase of infectionwith T crassiceps (2 wks) produced lower levels of IL-12 and
TNF-120572 compared with those from MGL1++ mice (Figures7(a) and 7(b)) As the infection became chronicmacrophagesfrom infected MGL1minusminus mice produced decreased levels ofTNF-120572 (Figure 7(b)) and theNO levels dropped significantly(Figure 7(c)) in both groups but even more in MGL1minusminusmacrophagesThese patterns ofmacrophage responsewere incontrast to those observed in MGL1++ macrophages whichshowed a better proinflammatory response throughout theinfection with higher production of TNF-120572 and NO in late
8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

2 BioMed Research International
It has been largely demonstrated that helminths displaya variety of glycan moieties on their antigens which mayparticipate in the induction of Th2 responses [8 9] and inimmune downregulation both characteristics of helminthinfections [10] Nevertheless given the complexity of thesemolecules it may also be expected that some helminthglycoconjugates display immunostimulatory activities [11]
Some helminth parasites have been reported to be recog-nized by different CLRs for example dendritic cell-specificICAM-3 grabbing integrin (DC-SIGN) mannose receptor(MR) macrophage galactose-type lectin (MGL) and SIGN-related 1 (SIGNR1) bind different glycans on soluble eggantigens from Schistosoma mansoni in vitro [12 13] Trichurismuris a nematode is also recognized by the MR but suchCLRs appear to play a limited role in protection or suscep-tibility in both parasitic infections given that mice lackingthe MR or SIGNR1 mount a normal immune response andclear these infections as usual [14]
Taenia crassiceps is a cestode that is useful for understand-ing the host-parasite relationship in cysticercosis Glycocon-jugates of T crassiceps and other cestodes have been shown toinduce strongTh2-biased responses in vivo [15] and tomodu-late both human and mouse DC activity in vitro [16ndash18] Wehave previously reported that most of the T crassiceps anti-gens bind to concanavalin A indicating that they are glycosy-lated with glucose mannose or galactose [15 17 19]The nat-ural candidates for such carbohydrate recognition are CLRssuch as the MR MGL and DC-SIGN As mentioned beforeDC-SIGN and the MR have been shown to be irrelevant forSchistosoma orTrichuris protectionWe therefore focused ourstudy onMGL1MGL1 ismainly found onM120601s and immatureDCs although it was recently reported to be expressed onmast cells [4] and it recognizes glycoconjugates expressinggalactose and N-acetyl-galactosamine residues Because theability of MGL1 to act as an autonomous receptor has beenappreciated only recently the molecular regulation of its sig-naling pathway is partly known [20] but its biological role inresponse to helminth infections is still unrevealed Recentlyit has been shown that upon strong MGL engagementwith polyclonal anti-MGL antibodies this receptor is ableto signal through tyrosine phosphorylation and induce ERKsignaling [20] however whether MGL signals after a ldquonatu-ralrdquo ligation is also unknown
To clarify the role ofMGL1 in the recognition and defenseagainst a helminth infection we infected wild-type andMGL1-deficient mice (MGL1minusminus) with Taenia crassiceps Ourdata demonstrated that MGL1-deficient mice exhibited redu-ced parasite clearance despite strong induction of Th2 res-ponses MGL1minusminus macrophages displayed reduced intracellu-lar signaling in response to Taenia antigens and lower TNF-120572 and NO production Taken together our data suggest thatMGL1 plays a key role in driving macrophage responses invivo and thus may act as an important mediator of resistanceto this helminth infection
2 Materials and Methods
21 Mice Six- to eight-week-old female MGL1minusminus mice on aC57BL6 background were donated by Glycomics Consor-tium (USA) MGL1minusminus mice have been backcrossed for more
than 7 generations on a C57BL6NHsd genetic backgroundwith mice from Harlan Laboratories (Mexico) In someexperiments BALBc mice were purchased from Harlan Lab-oratories (Mexico) All mice were maintained in a pathogen-free environment at the FES-Iztacala UNAM animal facili-ties according to the Faculty Animal Care and Use Commit-tee and government guidelines (official Mexican regulationNOM-062-ZOO-1999) which are in strict accordance withthe recommendations in the Guide for the Care and Useof Laboratory Animals of the National Institutes of HealthUSA
22 Parasites and Infection Metacestodes of T crassicepswere harvested from the peritoneal cavity of female BALBcmice after 2 to 3 months of infection The cysticerci werewashed four times in sterile phosphate-buffered saline (PBS)(015M pH 72) Experimental infection was achieved byintraperitoneal (ip) injectionwith 20 small (approx 2mm indiameter) nonbudding cysticerci ofT crassiceps suspended in03mL of PBS per mouse
23 Lectin-Blot Analysis on T crassiceps Soluble Antigens Tcrassicepswas lysed by sonicationwith one 10-s pulse at 100Wof amplitude (Fisher Sonic Dismembrator model 300) Theresulting suspension was centrifuged at 4∘C for 1 h at 2700 gand the pellet was discarded and we keep the supernatantSoluble antigen was concentrated and quantified by theBradford method A total of 20120583g of protein of T crassicepssoluble antigens (TcSol) was separated by SDS-PAGE (10)and electroblotted (400mA for 1 h) onto a nitrocellulosemembrane [21]
For the detection of carbohydrates with residues of gala-ctose or N-acetylgalactosamine strips of nitrocellulose wereblocked with 1 BSA diluted in PBS-T (10mM sodium phos-phate buffer 150mM NaCl and 05 Tween 20) and incu-bated overnight at 4∘C The membranes were treated with abiotinylated lectin (Hp or RCA) at a concentration of 5120583gmLdiluted 1 500 in PBS-T and incubated overnight at 4∘C(Sigma-Aldrich St LouisMOUSA)Thenext day themem-branes were incubated with SBHP the strips were washedwith PBS and enzyme activities were then detected by asubstrate solution (01 H
2
O2
175 methanol 015 4-chloro-120572-naphthol and 825 PBS) following a 15-min incu-bation The proteins that reacted with the lectins wereidentified on the nitrocellulose strips
24 FITC Labeling of T crassiceps Antigens T crassiceps solu-ble products (TcSol) were labeled with FITC (Sigma) accord-ing to the manufacturerrsquos instructions Briefly 1mgmL TcSolwas prepared in 01M sodium bicarbonate buffer pH 9 FITCwas dissolved in DMSO at 1mgmL and 500120583L of the FITCsolutionwas slowly added to TcSol in 5120583L aliquots while gen-tly stirring The TcSol-FITC solution was incubated for 8 h at4∘C in the dark and washed 3 times with PBS protein wasquantified by the Bradford assay and maintained at 4∘C
25 Cell Preparations and Culture Conditions The spleen wasremoved from infected mice under sterile conditions Single-cell suspensions were prepared by gently teasing apart the
BioMed Research International 3
spleen in RPMI-1640 supplemented with 10 fetal bovineserum 100 units of penicillinstreptomycin 2mMglutamine25mM HEPES buffer and 1 nonessential amino acids (allfrom GIBCO BRL Grand Island New York) The cells werecentrifuged and the erythrocytes were lysed by resuspendingthe cells in Boylersquos solution (017M Tris and 016M ammo-nium chloride) Following two washes the viable cells werecounted by trypan blue exclusion with a Neubauer hemocy-tometer and the splenocytes were adjusted to 3times 106 cellsmLin the same medium Aliquots (100 120583L) of the adjusted cellsuspensions were placed into 96-well flat bottom cultureplates (Costar Cambridge Massachusetts) and stimulatedwith a soluble extract ofT crassiceps (25120583gmL) orwith plate-bound anti-CD3 antibody (1 120583gmL) at 37∘C for 72 h Thesupernatants from these cultures were analyzed for IL-4 andIFN-120574 (PeproTech Mexico) production by ELISA
26 Cytokine and Nitric Oxide Production by PeritonealMacrophages Peritoneal exudate cells (PECs) were obtainedfrom uninfected thioglycolate-injected mice and from micefollowing 2 4 and 8 weeks of T crassiceps infection PECswere adjusted to 5 times 106mL in supplemented RPMI andplated in 6-well plates (Costar) After 2 hours at 37∘C and 5CO2
nonadherent cells were removed and adherent cellswere gently scrapped using cold PBS and readjusted to 1 times106mL Viability at this point was gt90 These cells weregt90 macrophages according to FACS analysis (F480+BioLegend SanDiego CA)Then 1mL of the cell suspensionwas plated and cells were activated in 24-well plates (Costar)with LPS (1120583gmL E coli 111B4 Sigma St Louis MO) fol-lowed by incubation for 24 h TNF-120572 IL-12 (PeproTech) andnitric oxide (Griess reaction) were examined in supernatants
27 Generation of Bone Marrow Derived M120601s (BMDM120601s)BMDM120601s fromWTorMGL1minusminusmicewere obtained as previ-ously described [22] Briefly bone marrow cells were isolatedby flushing femurs and tibias with culture media Bonemarrow cells were plated at 1 times 106 cellsmL in medium sup-plemented RPMI medium supplemented with 10 SFB andpenicillinstreptomycin plus 20 ngmL murine recombinantmacrophage colony-stimulating factor (M-CSF PeprotechMexico)Onday 3 freshmedia containingM-CSFwas addedOn day 7 nonadherent cells were removed and adherent cellswere detached and centrifuged at 1500 rpm at 4∘C for 15minAdherent cells (F480+) were resuspended in freshmedia andcultured for an additional 24 h in 12-well plates
28 In Vitro Stimulation of BMDM120601s and Protein ExtractionBone marrow cells were differentiated as described aboveOn day 7 the M120601s were harvested and plated at 25 times 106cellsmL in six-well plates The cells were used 24 hours afterplating to eliminate any residual effects from M-CSF M120601swere stimulatedwith theTcSol (50120583gmL) for 5 15 or 30minAfter stimulation the cells were centrifuged at 1500 rpm for5min and washed with PBS The cells were then lysed withlysis buffer for 15min and the lysates were quantified usingthe BCA assay (Thermo Scientific) and then frozen at minus80∘Cuntil further use
29 Western Blot Assays Whole-cell lysates were resolved bySDS-PAGE (40 120583g of protein was loaded into each well) using10 acrylamide mini-gels followed by electrophoretic trans-fer to PVDF membranes (Immobilon-P MILLIPORE) for2 hMembraneswere blockedwith 5 fat-freemilk in PBS for2 h and incubated with primary antibodies overnight Thedetection step was performed with peroxidase-coupled anti-mouse IgG and anti-rabbit IgG (BioLegend 1 2000) andanti-goat IgG (Santa Cruz Biotechnology) for 2 h The pri-mary antibodies included antiphosphotyrosine (Santa CruzBiotechnology) and 120573-Actin (BioLegend) All primary anti-bodies were diluted 1 500 in 1 fat-free milk in PBS Blotswere developed with the ECL detection system according tothemanufacturerrsquos instructions (Amersham) Blots are repre-sentative of two separate experiments
210 Detection of Nitric Oxide Production Nitric oxide pro-duction by macrophages was assayed by determining theincrease in nitrite concentration [23] by the Griess reactionadapted to microwell plates (Costar) Briefly 50120583L of culturesupernatant was mixed with an equal volume of Griessreagent and incubated for 10min at room temperature in thedark the absorbancewasmeasured at 570 nm in an automaticmicroplate reader (Organon Technika Microwell System)Values were quantified using serial dilutions of sodiumnitrite
211 Flow Cytometry Analysis Peritoneal exudate cells(PECs) were obtained from naıve or infected mice by inject-ing 10mL of ice-cold PBS For flow cytometry single cellsuspensions of the PECs obtained during the sacrifice werestained with anti-F480 anti-PDL2 anti-IL-4R120572 anti-MGL1and anti-MGL2 antibodies (Biolegend San Diego CA) for30min at 4∘CThe cells werewashed twice and analyzed usingthe FACSCalibur system and Cell Quest software (BectonDickinson USA)
212 Reverse Transcription (RT)-PCR Analyses The levels ofarginase 1 (Arg-1) Ym-1 and resistin-likemolecule-120572 (RELM-120572) mRNA transcripts in adherent peritoneal macrophages(which were allowed to adhere for 2 h at 37∘C and 5 CO
2
)were determined using RT-PCRAt the indicated time pointsadherent peritoneal macrophages from T crassiceps-infectedMGL1++ and MGL1 minusminus mice were aseptically removed andwithout any further stimulation (basal) or LPS + IFN-120574-stimulation (1 120583g and 20 ngmL resp) were processed forRNA extraction using TRIzol reagent (Invitrogen CarlsbadCA USA) and the propanol-chloroform techniqueThe RNAwas quantified and 3120583g of RNA was reverse transcribedusing the Superscript II First Strand Synthesis Kit (Invit-rogen) and an oligo dT primer as recommended by themanufacturer Once cDNA was obtained conventional PCRwas performed The PCR reactions contained (in a 25 120583Lfinal volume) 5X PCR Buffer blue 10mM dNTP 40 nM eachof the forward and reverse primers (Table 1) 1 U Taq DNApolymerase (Sacace Biotechnologies Italy) and 2 120583L of thecDNA The program used for the amplification of each genecontained an initial denaturation step at 95∘C for 5min 35
4 BioMed Research International
Table 1 The following primer pairs were used in this study
Gen Sequence Melting temperature Number of cycles
GAPDH Forward -CTCATGACCACAGTCCATGC-Reverse -CACATTGGGGGTAGGAACAC- 54∘C 35
Arg-1 Forward -CAGAAGAATGGAAGAGTCAG-Reverse -CAGATATGCAGGGAGTCACC- 54∘C 35
Ym-1 Forward -TCACAGGTCTGGCAATTCTTCTG-Reverse -TTTGTCCTTAGGAGGGCTTCCTC- 56∘C 35
Fizz-1 Forward -GGTCCCAGTGCATATGGATGAGACCATAG-Reverse -CACCTCTTCACTCGAGGGACAGTTGGCAGC- 65∘C 35
iNOS Forward -GCCACCAACAATGGCAACAT-Reverse -AAGACCAGAGGCAGCACATC- 60∘C 30
cycles of 95∘C for 40 s the indicated melting temperaturefor 50 s and 72∘C for 40 s and a final extension step at 72∘Cfor 4min All reactions were performed in a thermal cycler(Corbett Research Australia) Finally to observe the ampli-fied products a 15 agarose gel was prepared and sampleswere loaded with blue juice buffer containing SYBR Green(Invitrogen) The gels were visualized using a Fujifilm FLA5000 scanner (Fuji Japan) with FLA 5000 image reader V21software to capture the imagesThe specificity of the PCRwasverified by the absence of signal in the no-template controls ofmacrophage samplesThe sequences of the primers used havebeen previously reported [24]
213 Antibody ELISAs Peripheral blood was collected at 2-wk intervals from tails snips of the T crassiceps-infectedMGL1++ and MGL1minusminus mice T crassiceps-specific IgG1 andIgG2a levels were determined by ELISA as previously des-cribed The results are expressed as the maximal serum dilu-tion (endpoint titer) at which Optical Density was detected(OD) Total IgE production was detected by Opt-ELISA fromBiolegend
214 Statistical Analysis Comparisons between thewild-typeand MGL1minusminus groups in this study were performed usingStudentrsquos unpaired 119905 test 119875 lt 005was considered significantThe statistical significance of the serum titers was deter-mined by nonparametric tests using the Mann-Whitney119880Wilcoxon Rank tests
3 Results
31 Taenia crassiceps Soluble Antigens Express GlycoconjugatesContaining N-Acetylgalactosamine and Galactose ResiduesTo detect the expression of glycoconjugates on soluble anti-gens of T crassiceps a lectin-blot assay was performed usingHelix pomatia (specific for N-acetylgalactosamine residues)and Ricinus comunis (specific for galactose residues) lectinsSeveral glycoconjugates were recognized by both lectinsthose of 310 287 250 210 182 161 79 77 75 49 47 and 41 kDa(Figure 1(a)) whereas two bands at 51 and 39KDa were onlyrecognized by H pomatia However using only streptavidinperoxidase (Figure 1(b)) two glycoconjugates were recog-nized on TcSol antigens those of 250 and 124 kDaTherefore
49
80
124
209
35
124
250
797775
514947
4139
287310
210182161
124
250
310
210182161
797775
4947
41
287
RCAHPMW
(a)
250
124
SBHP
(b)
Figure 1 Lectin-blot analysis of Taenia crassiceps soluble antigensT crassiceps soluble proteins were subjected to electrophoresison a 10 SDS-polyacrylamide gel transferred to nitrocellulosemembrane and revealed with a couple of different biotinylatedlectins (a) MW-molecular weight HP-Helix pomatia and RCA-Ricinus communis (b) The strips were incubated only with SBHP-streptavidin-bound horseradish peroxidase
the presence of these two biotin-containing proteins mostlikely corresponds to biotin-dependent carboxylases recog-nized by streptavidin alone [25 26] These findings indicatethat TcSol antigens express high levels of glycoconjugateswith N-acetylgalactosamine and galactose residues
32 MGL1minusminus Macrophages Display Reduced Binding to Tae-nia crassiceps Soluble Antigens The innate recognition ofpathogen-associated molecules by antigen-presenting cells iscrucial to the initiation of the immune response Differentstudies have shown that helminth-derived molecules can berecognized by C-type lectins [27 28] In our model wepreviously reported that most TcSol bound to concanavalin
BioMed Research International 5
A indicating that TcSol are glycosylated with glucose man-nose or galactose [17] whereas Jang Lee et al reported thatthe main N-glycan structures in T crassiceps were mannosefucose galactose and GlcNAc They also found a rare Fuc1205721rarr 3GlcNAc antenna on T crassiceps molecules [29] Thusthe natural candidates for carbohydrate recognition on Tcrassiceps are CLRs such as the MR MGL and DC-SIGNThus we hypothesized that one of the possible receptors thatbind the glycomolecules on TcSol may be MGL given itsspecificity to recognize both N-acetylgalactosamine andgalactose residues To investigate the role of MGL1 in TcSolrecognition TcSol antigens were labeled with FITC toperform binding assays TcSol-FITC efficiently bound toMGL1++ macrophages at 37∘C (approximately 30 of cellswere positive) indicating that TcSol-FITC can be recognizedand internalized by these cells (Figures 2(a) and 2(b)) Incu-bation of M120601s and TcSol at 4∘C showed reduced fluorescence(5ndash10) indicating a low level of binding Next we similarlyexposed MGL1minusminus M120601s to TcSol-FITC at 37∘C and foundsignificant reductions in both the percentage of TcSol-FITC-positive cells (approximately 10) and the mean fluorescenceintensity (MFI) of TcSol-FITC on these cells (Figure 2(c))These data suggest that M120601s can recognize TcSol via MGL
33 The Absence of MGL1 on Macrophages Impairs TcSol-Triggered Global Tyrosine Phosphorylation The fact thatTcSol binding was impaired in MGL1minusminusM120601s led us to ques-tion whether the interaction with MGL-TcSol triggers anintracellular signaling pathway Recent advances in theunderstanding of the intracellular signaling induced bystrong antibody-mediated crosslinking of MGL have high-lighted a possible role for the ERK pathway [20] To deter-mine whether TcSol can induce intracellular signaling weexposed BMDM120601s fromMGL1++ or MGL1minusminus mice to TcSolfor 5 15 and 30min and found that tyrosine phosphorylationwas enhanced in MGL1++ macrophages interestingly tyro-sine phosphorylation was detected as early as 5min andremained for at least 30min (Figure 3(a)) In contrastMGL1minusminus BMDM120601s that were similarly exposed to TcSol dis-played a lower and less-sustained tyrosine phosphorylationin several protein bands (Figure 3(a) arrows) To further ana-lyze this response we stimulated bothMGL1++ andMGL1minusminusBMDM120601s with LPS (500 ngmL) or TcSol again for 30minAs observed in Figure 3(b) alsowedetected a lack of responseto LPS and a lower tyrosine phosphorylation in response toTcSol (Figures 3(b) and 3(c) resp) in MGL1minusminus BMDM120601s
34 Mice Lacking MGL1 Are Highly Susceptible to Taenia cras-siceps Infection After determining that T crassiceps antigenscan be recognized by MGL1 and that macrophages deficientin MGL1 are partially refractory to TcSol stimulation weinvestigated the in vivo role of this molecule in experimentalcysticercosis using MGL-deficient mice To approach thisquestion we compared the course of T crassiceps infection inMGL1minusminus with that in MGL1++ mice As a control for sus-ceptibility we used BALBc mice which have consistently
been reported as a highly susceptible strain to T crassicepsinfection [30]
We examined the kinetics of parasite growth from 2 to 8weeks after T crassiceps infection Early in the infection (2weeks) both groups of mice displayed comparable parasiteburdens (Figure 4(a)) Interestingly as infection progressedby the fourth week of infection the number of larvae in theperitoneal cavity increased significantly in MGL1minusminus micecompared to MGL1++ mice which successfully reduced thenumber of parasites by week 8 after infection (Figures 4(a)and 4(b)) In fact MGL1minusminus mice exhibited a parasite burdenthat was very similar to that in the susceptible strain of mice(BALBc) which were consistently observed as very suscepti-ble toT crassiceps infection (Figure 4(a))These findings sug-gest that theMGL1-mediated signaling pathway is involved inresistance during T crassiceps infection on a resistant geneticbackground such as C57BL6
35 MGL1minusminus Mice Display an Altered Immune Responseto Taenia crassiceps A few studies have demonstrated thatMGL can mediate signaling after a strong stimulus such aspolyclonal anti-MGL antibodies but its role in modulatingprotective immunity against helminth parasites is unknownTherefore wemeasured levels ofTh1-associated IgG2a as wellas Th2-associated IgG1 and total IgE antibodies in MGL1minusminus
andMGL1++mice at different timepoints following infectionwith T crassiceps
Early in infection T crassiceps-infected MGL1++ andMGL1minusminus mice displayed comparable levels of T crassicepsAg-specific Th1-associated IgG2a antibodies (data notshown) However by week 8 after infection MGL1++ micedisplayed significantly higher titers of specific IgG2aantibodies againstT crassiceps antigens (Figure 5(a)) By con-trast no clear differences were observed in Th2-associatedIgG1 production MGL1++ mice displayed high titers ofanti-T crassiceps-specific IgG1 that were similar to thosein MGL1minusminus mice at week 8 after infection (Figure 5(b))AlthoughTh2-associated IgE has been shown to play a role inmediating immunity against certain helminths we found thatT crassiceps-infected MGL1++ mice harbored a lower para-site burden despite producing significantly lower levels ofIgE compared to similarly infected MGL1minusminus mice whichdisplayed higher levels of total IgE (Figure 5(c))
Additionally we also compared the cytokine produc-tion by splenocytes from these mice in response to either25 120583gmL TcSol or 1 120583gmL plate-bound anti-CD3 antibodyAnti-CD3 (data not shown) or TcSol-stimulated splenocytesfrom both strains produced similar levels of IFN-120574 at 2 wkafter infection (Figure 6(a)) However 4 and 8 weeks afterinfection the IFN-120574 production by the spleen cells ofMGL1minusminus mice decreased and did not reach the level pro-duced by MGL1++ splenocytes (Figure 6(a)) Similarly asearly as 2 wk after infection the splenocytes from theMGL1minusminus and MGL1++ mice produced comparable levels ofIL-4 (Figure 6(b)) However by weeks 4 and 8 after infectionMGL1minusminus mice produced significantly more IL-4 than the
6 BioMed Research International
F480 F480
TcSo
l-FIT
C
TcSo
l-FIT
C
100 101 102 103 104 100 101 102 103 104
29 6
100
101
102
103
104
100
101
102
103
104MGL1++ 37∘C MGL1minusminus 37∘C
(a)
TcSo
l-FIT
C (
)
lowast
50
40
30
20
10
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(b)
lowast
MFI
500
400
300
200
100
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(c)
Figure 2 Peritoneal M120601s recognize Taenia crassiceps soluble products through MGL1 M120601s from naıve MGL1++ or MGL1minusminus mice wereincubated with fluorescently labeled TcSol and fluorescence was analyzed by flow cytometry (a) Representative dot blots indicate M120601s fromeither MGL1++ or MGL1minusminus mice incubated for 30 minutes with TcSol at 37∘C (b) Percentage of TcSol-FITC positive naıve peritoneal cellsexposed for 30min (c) Data are represented as the mean florescence intensity (MFI) of naıve M120601s exposed to TcES-FITC Basal levels ofautofluorescence were subtracted from all treatments Data are representative of three independent experiments lowast119875 lt 005
MGL1++ splenocytes in response to anti-CD3 whereasantigen-specific higher IL-4 production in MGL1minusminus spleno-cytes was only observed at week 4 after infection comparedwithMGL1++ cells (Figures 6(b) and 6(c))Thus as the infec-tion progressed splenocytes from MGL1minusminus mice producedsignificantly greater levels of IL-4 compared with splenocytes
fromT crassiceps-infectedMGL1++mice in response to anti-CD3
36 Differential Cytokine Production in MGL1minusminus andMGL1++ Peritoneal Macrophages To determine whetherMGL1minusminus mice had a systemic defect in innate activation we
BioMed Research International 7
TcSol
170130
9870
55
42
35
MW
50120583gmL5998400 15998400 30998400 5998400 15998400 30998400
MGL1++ MGL1minusminus
minusminus
(a)
170130
98705542
35
25
TcSolLPS 30998400
3099840030998400 30998400
MGL1++MGL1minusminus
120573-Actin
minusminus
minus
minus minus
minus minusminus
(b)
MGL1++MGL1minusminus
Relat
ive p
hosp
hory
latio
n le
vel
TcSolLPS 30998400
30998400
08
06
04
02
00
3099840030998400minus
minus
minus
minus minus
minus minus
minus
P-Y LPS and TcSol
(c)
Figure 3 MGL1minusminus macrophages display deficient tyrosine phosphorylation in response to TcSol (a) MGL1++ and MGL1minusminus bone marrow-derived M120601 were synchronized in starvation conditions over night and then were stimulated or not with TcSol (50120583gmL) for 5 15 or30min Next total cell extracts were obtained and resolved by electrophoresis transferred to nitrocellulose and probed with monoclonalantiphosphotyrosine antibody Lane 1 serum-starved MGL1++ M120601s Lanes 2ndash4 serum-starved MGL1++ M120601s following TcSol exposure for5 15 and 30min respectively Lane 5 serum-starved MGL1minusminus M120601s Lanes 6ndash8 serum-starved MGL1minusminus M120601s following TcSol exposure for5 15 and 30min respectively Lanes 2ndash4 show an upregulation of the tyrosine phosphorylation of total proteins which was sustained forat least 30min (arrows) Lanes 6ndash8 show an unsustained tyrosine phosphorylation of proteins after similar TcSol stimulation of MGL1minusminusM120601s (b) Similar experiment using LPS (500 ngmL) andor TcSol stimulation for 30min (c) Densitometry of western blot placed in (b)Proteins were visualized using a goat anti-mouse secondary conjugated to HRP and a chemiluminescence detection system Western blotsare representative of two independent experiments
examined the responsiveness of macrophages fromMGL1minusminusmice to proinflammatory stimuli Macrophages were isolatedfrom the peritoneal cavities of both strains of infected miceand either left unstimulated (basal) or stimulated for 24 hwith LPS (1120583gmL) and IFN-120574 (20 ngmL) The supernatantswere collected and analyzed for IL-12 TNF-120572 and NOproduction As shown in Figure 7 macrophages fromMGL1minusminus mice obtained during the early phase of infectionwith T crassiceps (2 wks) produced lower levels of IL-12 and
TNF-120572 compared with those from MGL1++ mice (Figures7(a) and 7(b)) As the infection became chronicmacrophagesfrom infected MGL1minusminus mice produced decreased levels ofTNF-120572 (Figure 7(b)) and theNO levels dropped significantly(Figure 7(c)) in both groups but even more in MGL1minusminusmacrophagesThese patterns ofmacrophage responsewere incontrast to those observed in MGL1++ macrophages whichshowed a better proinflammatory response throughout theinfection with higher production of TNF-120572 and NO in late
8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 3
spleen in RPMI-1640 supplemented with 10 fetal bovineserum 100 units of penicillinstreptomycin 2mMglutamine25mM HEPES buffer and 1 nonessential amino acids (allfrom GIBCO BRL Grand Island New York) The cells werecentrifuged and the erythrocytes were lysed by resuspendingthe cells in Boylersquos solution (017M Tris and 016M ammo-nium chloride) Following two washes the viable cells werecounted by trypan blue exclusion with a Neubauer hemocy-tometer and the splenocytes were adjusted to 3times 106 cellsmLin the same medium Aliquots (100 120583L) of the adjusted cellsuspensions were placed into 96-well flat bottom cultureplates (Costar Cambridge Massachusetts) and stimulatedwith a soluble extract ofT crassiceps (25120583gmL) orwith plate-bound anti-CD3 antibody (1 120583gmL) at 37∘C for 72 h Thesupernatants from these cultures were analyzed for IL-4 andIFN-120574 (PeproTech Mexico) production by ELISA
26 Cytokine and Nitric Oxide Production by PeritonealMacrophages Peritoneal exudate cells (PECs) were obtainedfrom uninfected thioglycolate-injected mice and from micefollowing 2 4 and 8 weeks of T crassiceps infection PECswere adjusted to 5 times 106mL in supplemented RPMI andplated in 6-well plates (Costar) After 2 hours at 37∘C and 5CO2
nonadherent cells were removed and adherent cellswere gently scrapped using cold PBS and readjusted to 1 times106mL Viability at this point was gt90 These cells weregt90 macrophages according to FACS analysis (F480+BioLegend SanDiego CA)Then 1mL of the cell suspensionwas plated and cells were activated in 24-well plates (Costar)with LPS (1120583gmL E coli 111B4 Sigma St Louis MO) fol-lowed by incubation for 24 h TNF-120572 IL-12 (PeproTech) andnitric oxide (Griess reaction) were examined in supernatants
27 Generation of Bone Marrow Derived M120601s (BMDM120601s)BMDM120601s fromWTorMGL1minusminusmicewere obtained as previ-ously described [22] Briefly bone marrow cells were isolatedby flushing femurs and tibias with culture media Bonemarrow cells were plated at 1 times 106 cellsmL in medium sup-plemented RPMI medium supplemented with 10 SFB andpenicillinstreptomycin plus 20 ngmL murine recombinantmacrophage colony-stimulating factor (M-CSF PeprotechMexico)Onday 3 freshmedia containingM-CSFwas addedOn day 7 nonadherent cells were removed and adherent cellswere detached and centrifuged at 1500 rpm at 4∘C for 15minAdherent cells (F480+) were resuspended in freshmedia andcultured for an additional 24 h in 12-well plates
28 In Vitro Stimulation of BMDM120601s and Protein ExtractionBone marrow cells were differentiated as described aboveOn day 7 the M120601s were harvested and plated at 25 times 106cellsmL in six-well plates The cells were used 24 hours afterplating to eliminate any residual effects from M-CSF M120601swere stimulatedwith theTcSol (50120583gmL) for 5 15 or 30minAfter stimulation the cells were centrifuged at 1500 rpm for5min and washed with PBS The cells were then lysed withlysis buffer for 15min and the lysates were quantified usingthe BCA assay (Thermo Scientific) and then frozen at minus80∘Cuntil further use
29 Western Blot Assays Whole-cell lysates were resolved bySDS-PAGE (40 120583g of protein was loaded into each well) using10 acrylamide mini-gels followed by electrophoretic trans-fer to PVDF membranes (Immobilon-P MILLIPORE) for2 hMembraneswere blockedwith 5 fat-freemilk in PBS for2 h and incubated with primary antibodies overnight Thedetection step was performed with peroxidase-coupled anti-mouse IgG and anti-rabbit IgG (BioLegend 1 2000) andanti-goat IgG (Santa Cruz Biotechnology) for 2 h The pri-mary antibodies included antiphosphotyrosine (Santa CruzBiotechnology) and 120573-Actin (BioLegend) All primary anti-bodies were diluted 1 500 in 1 fat-free milk in PBS Blotswere developed with the ECL detection system according tothemanufacturerrsquos instructions (Amersham) Blots are repre-sentative of two separate experiments
210 Detection of Nitric Oxide Production Nitric oxide pro-duction by macrophages was assayed by determining theincrease in nitrite concentration [23] by the Griess reactionadapted to microwell plates (Costar) Briefly 50120583L of culturesupernatant was mixed with an equal volume of Griessreagent and incubated for 10min at room temperature in thedark the absorbancewasmeasured at 570 nm in an automaticmicroplate reader (Organon Technika Microwell System)Values were quantified using serial dilutions of sodiumnitrite
211 Flow Cytometry Analysis Peritoneal exudate cells(PECs) were obtained from naıve or infected mice by inject-ing 10mL of ice-cold PBS For flow cytometry single cellsuspensions of the PECs obtained during the sacrifice werestained with anti-F480 anti-PDL2 anti-IL-4R120572 anti-MGL1and anti-MGL2 antibodies (Biolegend San Diego CA) for30min at 4∘CThe cells werewashed twice and analyzed usingthe FACSCalibur system and Cell Quest software (BectonDickinson USA)
212 Reverse Transcription (RT)-PCR Analyses The levels ofarginase 1 (Arg-1) Ym-1 and resistin-likemolecule-120572 (RELM-120572) mRNA transcripts in adherent peritoneal macrophages(which were allowed to adhere for 2 h at 37∘C and 5 CO
2
)were determined using RT-PCRAt the indicated time pointsadherent peritoneal macrophages from T crassiceps-infectedMGL1++ and MGL1 minusminus mice were aseptically removed andwithout any further stimulation (basal) or LPS + IFN-120574-stimulation (1 120583g and 20 ngmL resp) were processed forRNA extraction using TRIzol reagent (Invitrogen CarlsbadCA USA) and the propanol-chloroform techniqueThe RNAwas quantified and 3120583g of RNA was reverse transcribedusing the Superscript II First Strand Synthesis Kit (Invit-rogen) and an oligo dT primer as recommended by themanufacturer Once cDNA was obtained conventional PCRwas performed The PCR reactions contained (in a 25 120583Lfinal volume) 5X PCR Buffer blue 10mM dNTP 40 nM eachof the forward and reverse primers (Table 1) 1 U Taq DNApolymerase (Sacace Biotechnologies Italy) and 2 120583L of thecDNA The program used for the amplification of each genecontained an initial denaturation step at 95∘C for 5min 35
4 BioMed Research International
Table 1 The following primer pairs were used in this study
Gen Sequence Melting temperature Number of cycles
GAPDH Forward -CTCATGACCACAGTCCATGC-Reverse -CACATTGGGGGTAGGAACAC- 54∘C 35
Arg-1 Forward -CAGAAGAATGGAAGAGTCAG-Reverse -CAGATATGCAGGGAGTCACC- 54∘C 35
Ym-1 Forward -TCACAGGTCTGGCAATTCTTCTG-Reverse -TTTGTCCTTAGGAGGGCTTCCTC- 56∘C 35
Fizz-1 Forward -GGTCCCAGTGCATATGGATGAGACCATAG-Reverse -CACCTCTTCACTCGAGGGACAGTTGGCAGC- 65∘C 35
iNOS Forward -GCCACCAACAATGGCAACAT-Reverse -AAGACCAGAGGCAGCACATC- 60∘C 30
cycles of 95∘C for 40 s the indicated melting temperaturefor 50 s and 72∘C for 40 s and a final extension step at 72∘Cfor 4min All reactions were performed in a thermal cycler(Corbett Research Australia) Finally to observe the ampli-fied products a 15 agarose gel was prepared and sampleswere loaded with blue juice buffer containing SYBR Green(Invitrogen) The gels were visualized using a Fujifilm FLA5000 scanner (Fuji Japan) with FLA 5000 image reader V21software to capture the imagesThe specificity of the PCRwasverified by the absence of signal in the no-template controls ofmacrophage samplesThe sequences of the primers used havebeen previously reported [24]
213 Antibody ELISAs Peripheral blood was collected at 2-wk intervals from tails snips of the T crassiceps-infectedMGL1++ and MGL1minusminus mice T crassiceps-specific IgG1 andIgG2a levels were determined by ELISA as previously des-cribed The results are expressed as the maximal serum dilu-tion (endpoint titer) at which Optical Density was detected(OD) Total IgE production was detected by Opt-ELISA fromBiolegend
214 Statistical Analysis Comparisons between thewild-typeand MGL1minusminus groups in this study were performed usingStudentrsquos unpaired 119905 test 119875 lt 005was considered significantThe statistical significance of the serum titers was deter-mined by nonparametric tests using the Mann-Whitney119880Wilcoxon Rank tests
3 Results
31 Taenia crassiceps Soluble Antigens Express GlycoconjugatesContaining N-Acetylgalactosamine and Galactose ResiduesTo detect the expression of glycoconjugates on soluble anti-gens of T crassiceps a lectin-blot assay was performed usingHelix pomatia (specific for N-acetylgalactosamine residues)and Ricinus comunis (specific for galactose residues) lectinsSeveral glycoconjugates were recognized by both lectinsthose of 310 287 250 210 182 161 79 77 75 49 47 and 41 kDa(Figure 1(a)) whereas two bands at 51 and 39KDa were onlyrecognized by H pomatia However using only streptavidinperoxidase (Figure 1(b)) two glycoconjugates were recog-nized on TcSol antigens those of 250 and 124 kDaTherefore
49
80
124
209
35
124
250
797775
514947
4139
287310
210182161
124
250
310
210182161
797775
4947
41
287
RCAHPMW
(a)
250
124
SBHP
(b)
Figure 1 Lectin-blot analysis of Taenia crassiceps soluble antigensT crassiceps soluble proteins were subjected to electrophoresison a 10 SDS-polyacrylamide gel transferred to nitrocellulosemembrane and revealed with a couple of different biotinylatedlectins (a) MW-molecular weight HP-Helix pomatia and RCA-Ricinus communis (b) The strips were incubated only with SBHP-streptavidin-bound horseradish peroxidase
the presence of these two biotin-containing proteins mostlikely corresponds to biotin-dependent carboxylases recog-nized by streptavidin alone [25 26] These findings indicatethat TcSol antigens express high levels of glycoconjugateswith N-acetylgalactosamine and galactose residues
32 MGL1minusminus Macrophages Display Reduced Binding to Tae-nia crassiceps Soluble Antigens The innate recognition ofpathogen-associated molecules by antigen-presenting cells iscrucial to the initiation of the immune response Differentstudies have shown that helminth-derived molecules can berecognized by C-type lectins [27 28] In our model wepreviously reported that most TcSol bound to concanavalin
BioMed Research International 5
A indicating that TcSol are glycosylated with glucose man-nose or galactose [17] whereas Jang Lee et al reported thatthe main N-glycan structures in T crassiceps were mannosefucose galactose and GlcNAc They also found a rare Fuc1205721rarr 3GlcNAc antenna on T crassiceps molecules [29] Thusthe natural candidates for carbohydrate recognition on Tcrassiceps are CLRs such as the MR MGL and DC-SIGNThus we hypothesized that one of the possible receptors thatbind the glycomolecules on TcSol may be MGL given itsspecificity to recognize both N-acetylgalactosamine andgalactose residues To investigate the role of MGL1 in TcSolrecognition TcSol antigens were labeled with FITC toperform binding assays TcSol-FITC efficiently bound toMGL1++ macrophages at 37∘C (approximately 30 of cellswere positive) indicating that TcSol-FITC can be recognizedand internalized by these cells (Figures 2(a) and 2(b)) Incu-bation of M120601s and TcSol at 4∘C showed reduced fluorescence(5ndash10) indicating a low level of binding Next we similarlyexposed MGL1minusminus M120601s to TcSol-FITC at 37∘C and foundsignificant reductions in both the percentage of TcSol-FITC-positive cells (approximately 10) and the mean fluorescenceintensity (MFI) of TcSol-FITC on these cells (Figure 2(c))These data suggest that M120601s can recognize TcSol via MGL
33 The Absence of MGL1 on Macrophages Impairs TcSol-Triggered Global Tyrosine Phosphorylation The fact thatTcSol binding was impaired in MGL1minusminusM120601s led us to ques-tion whether the interaction with MGL-TcSol triggers anintracellular signaling pathway Recent advances in theunderstanding of the intracellular signaling induced bystrong antibody-mediated crosslinking of MGL have high-lighted a possible role for the ERK pathway [20] To deter-mine whether TcSol can induce intracellular signaling weexposed BMDM120601s fromMGL1++ or MGL1minusminus mice to TcSolfor 5 15 and 30min and found that tyrosine phosphorylationwas enhanced in MGL1++ macrophages interestingly tyro-sine phosphorylation was detected as early as 5min andremained for at least 30min (Figure 3(a)) In contrastMGL1minusminus BMDM120601s that were similarly exposed to TcSol dis-played a lower and less-sustained tyrosine phosphorylationin several protein bands (Figure 3(a) arrows) To further ana-lyze this response we stimulated bothMGL1++ andMGL1minusminusBMDM120601s with LPS (500 ngmL) or TcSol again for 30minAs observed in Figure 3(b) alsowedetected a lack of responseto LPS and a lower tyrosine phosphorylation in response toTcSol (Figures 3(b) and 3(c) resp) in MGL1minusminus BMDM120601s
34 Mice Lacking MGL1 Are Highly Susceptible to Taenia cras-siceps Infection After determining that T crassiceps antigenscan be recognized by MGL1 and that macrophages deficientin MGL1 are partially refractory to TcSol stimulation weinvestigated the in vivo role of this molecule in experimentalcysticercosis using MGL-deficient mice To approach thisquestion we compared the course of T crassiceps infection inMGL1minusminus with that in MGL1++ mice As a control for sus-ceptibility we used BALBc mice which have consistently
been reported as a highly susceptible strain to T crassicepsinfection [30]
We examined the kinetics of parasite growth from 2 to 8weeks after T crassiceps infection Early in the infection (2weeks) both groups of mice displayed comparable parasiteburdens (Figure 4(a)) Interestingly as infection progressedby the fourth week of infection the number of larvae in theperitoneal cavity increased significantly in MGL1minusminus micecompared to MGL1++ mice which successfully reduced thenumber of parasites by week 8 after infection (Figures 4(a)and 4(b)) In fact MGL1minusminus mice exhibited a parasite burdenthat was very similar to that in the susceptible strain of mice(BALBc) which were consistently observed as very suscepti-ble toT crassiceps infection (Figure 4(a))These findings sug-gest that theMGL1-mediated signaling pathway is involved inresistance during T crassiceps infection on a resistant geneticbackground such as C57BL6
35 MGL1minusminus Mice Display an Altered Immune Responseto Taenia crassiceps A few studies have demonstrated thatMGL can mediate signaling after a strong stimulus such aspolyclonal anti-MGL antibodies but its role in modulatingprotective immunity against helminth parasites is unknownTherefore wemeasured levels ofTh1-associated IgG2a as wellas Th2-associated IgG1 and total IgE antibodies in MGL1minusminus
andMGL1++mice at different timepoints following infectionwith T crassiceps
Early in infection T crassiceps-infected MGL1++ andMGL1minusminus mice displayed comparable levels of T crassicepsAg-specific Th1-associated IgG2a antibodies (data notshown) However by week 8 after infection MGL1++ micedisplayed significantly higher titers of specific IgG2aantibodies againstT crassiceps antigens (Figure 5(a)) By con-trast no clear differences were observed in Th2-associatedIgG1 production MGL1++ mice displayed high titers ofanti-T crassiceps-specific IgG1 that were similar to thosein MGL1minusminus mice at week 8 after infection (Figure 5(b))AlthoughTh2-associated IgE has been shown to play a role inmediating immunity against certain helminths we found thatT crassiceps-infected MGL1++ mice harbored a lower para-site burden despite producing significantly lower levels ofIgE compared to similarly infected MGL1minusminus mice whichdisplayed higher levels of total IgE (Figure 5(c))
Additionally we also compared the cytokine produc-tion by splenocytes from these mice in response to either25 120583gmL TcSol or 1 120583gmL plate-bound anti-CD3 antibodyAnti-CD3 (data not shown) or TcSol-stimulated splenocytesfrom both strains produced similar levels of IFN-120574 at 2 wkafter infection (Figure 6(a)) However 4 and 8 weeks afterinfection the IFN-120574 production by the spleen cells ofMGL1minusminus mice decreased and did not reach the level pro-duced by MGL1++ splenocytes (Figure 6(a)) Similarly asearly as 2 wk after infection the splenocytes from theMGL1minusminus and MGL1++ mice produced comparable levels ofIL-4 (Figure 6(b)) However by weeks 4 and 8 after infectionMGL1minusminus mice produced significantly more IL-4 than the
6 BioMed Research International
F480 F480
TcSo
l-FIT
C
TcSo
l-FIT
C
100 101 102 103 104 100 101 102 103 104
29 6
100
101
102
103
104
100
101
102
103
104MGL1++ 37∘C MGL1minusminus 37∘C
(a)
TcSo
l-FIT
C (
)
lowast
50
40
30
20
10
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(b)
lowast
MFI
500
400
300
200
100
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(c)
Figure 2 Peritoneal M120601s recognize Taenia crassiceps soluble products through MGL1 M120601s from naıve MGL1++ or MGL1minusminus mice wereincubated with fluorescently labeled TcSol and fluorescence was analyzed by flow cytometry (a) Representative dot blots indicate M120601s fromeither MGL1++ or MGL1minusminus mice incubated for 30 minutes with TcSol at 37∘C (b) Percentage of TcSol-FITC positive naıve peritoneal cellsexposed for 30min (c) Data are represented as the mean florescence intensity (MFI) of naıve M120601s exposed to TcES-FITC Basal levels ofautofluorescence were subtracted from all treatments Data are representative of three independent experiments lowast119875 lt 005
MGL1++ splenocytes in response to anti-CD3 whereasantigen-specific higher IL-4 production in MGL1minusminus spleno-cytes was only observed at week 4 after infection comparedwithMGL1++ cells (Figures 6(b) and 6(c))Thus as the infec-tion progressed splenocytes from MGL1minusminus mice producedsignificantly greater levels of IL-4 compared with splenocytes
fromT crassiceps-infectedMGL1++mice in response to anti-CD3
36 Differential Cytokine Production in MGL1minusminus andMGL1++ Peritoneal Macrophages To determine whetherMGL1minusminus mice had a systemic defect in innate activation we
BioMed Research International 7
TcSol
170130
9870
55
42
35
MW
50120583gmL5998400 15998400 30998400 5998400 15998400 30998400
MGL1++ MGL1minusminus
minusminus
(a)
170130
98705542
35
25
TcSolLPS 30998400
3099840030998400 30998400
MGL1++MGL1minusminus
120573-Actin
minusminus
minus
minus minus
minus minusminus
(b)
MGL1++MGL1minusminus
Relat
ive p
hosp
hory
latio
n le
vel
TcSolLPS 30998400
30998400
08
06
04
02
00
3099840030998400minus
minus
minus
minus minus
minus minus
minus
P-Y LPS and TcSol
(c)
Figure 3 MGL1minusminus macrophages display deficient tyrosine phosphorylation in response to TcSol (a) MGL1++ and MGL1minusminus bone marrow-derived M120601 were synchronized in starvation conditions over night and then were stimulated or not with TcSol (50120583gmL) for 5 15 or30min Next total cell extracts were obtained and resolved by electrophoresis transferred to nitrocellulose and probed with monoclonalantiphosphotyrosine antibody Lane 1 serum-starved MGL1++ M120601s Lanes 2ndash4 serum-starved MGL1++ M120601s following TcSol exposure for5 15 and 30min respectively Lane 5 serum-starved MGL1minusminus M120601s Lanes 6ndash8 serum-starved MGL1minusminus M120601s following TcSol exposure for5 15 and 30min respectively Lanes 2ndash4 show an upregulation of the tyrosine phosphorylation of total proteins which was sustained forat least 30min (arrows) Lanes 6ndash8 show an unsustained tyrosine phosphorylation of proteins after similar TcSol stimulation of MGL1minusminusM120601s (b) Similar experiment using LPS (500 ngmL) andor TcSol stimulation for 30min (c) Densitometry of western blot placed in (b)Proteins were visualized using a goat anti-mouse secondary conjugated to HRP and a chemiluminescence detection system Western blotsare representative of two independent experiments
examined the responsiveness of macrophages fromMGL1minusminusmice to proinflammatory stimuli Macrophages were isolatedfrom the peritoneal cavities of both strains of infected miceand either left unstimulated (basal) or stimulated for 24 hwith LPS (1120583gmL) and IFN-120574 (20 ngmL) The supernatantswere collected and analyzed for IL-12 TNF-120572 and NOproduction As shown in Figure 7 macrophages fromMGL1minusminus mice obtained during the early phase of infectionwith T crassiceps (2 wks) produced lower levels of IL-12 and
TNF-120572 compared with those from MGL1++ mice (Figures7(a) and 7(b)) As the infection became chronicmacrophagesfrom infected MGL1minusminus mice produced decreased levels ofTNF-120572 (Figure 7(b)) and theNO levels dropped significantly(Figure 7(c)) in both groups but even more in MGL1minusminusmacrophagesThese patterns ofmacrophage responsewere incontrast to those observed in MGL1++ macrophages whichshowed a better proinflammatory response throughout theinfection with higher production of TNF-120572 and NO in late
8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

4 BioMed Research International
Table 1 The following primer pairs were used in this study
Gen Sequence Melting temperature Number of cycles
GAPDH Forward -CTCATGACCACAGTCCATGC-Reverse -CACATTGGGGGTAGGAACAC- 54∘C 35
Arg-1 Forward -CAGAAGAATGGAAGAGTCAG-Reverse -CAGATATGCAGGGAGTCACC- 54∘C 35
Ym-1 Forward -TCACAGGTCTGGCAATTCTTCTG-Reverse -TTTGTCCTTAGGAGGGCTTCCTC- 56∘C 35
Fizz-1 Forward -GGTCCCAGTGCATATGGATGAGACCATAG-Reverse -CACCTCTTCACTCGAGGGACAGTTGGCAGC- 65∘C 35
iNOS Forward -GCCACCAACAATGGCAACAT-Reverse -AAGACCAGAGGCAGCACATC- 60∘C 30
cycles of 95∘C for 40 s the indicated melting temperaturefor 50 s and 72∘C for 40 s and a final extension step at 72∘Cfor 4min All reactions were performed in a thermal cycler(Corbett Research Australia) Finally to observe the ampli-fied products a 15 agarose gel was prepared and sampleswere loaded with blue juice buffer containing SYBR Green(Invitrogen) The gels were visualized using a Fujifilm FLA5000 scanner (Fuji Japan) with FLA 5000 image reader V21software to capture the imagesThe specificity of the PCRwasverified by the absence of signal in the no-template controls ofmacrophage samplesThe sequences of the primers used havebeen previously reported [24]
213 Antibody ELISAs Peripheral blood was collected at 2-wk intervals from tails snips of the T crassiceps-infectedMGL1++ and MGL1minusminus mice T crassiceps-specific IgG1 andIgG2a levels were determined by ELISA as previously des-cribed The results are expressed as the maximal serum dilu-tion (endpoint titer) at which Optical Density was detected(OD) Total IgE production was detected by Opt-ELISA fromBiolegend
214 Statistical Analysis Comparisons between thewild-typeand MGL1minusminus groups in this study were performed usingStudentrsquos unpaired 119905 test 119875 lt 005was considered significantThe statistical significance of the serum titers was deter-mined by nonparametric tests using the Mann-Whitney119880Wilcoxon Rank tests
3 Results
31 Taenia crassiceps Soluble Antigens Express GlycoconjugatesContaining N-Acetylgalactosamine and Galactose ResiduesTo detect the expression of glycoconjugates on soluble anti-gens of T crassiceps a lectin-blot assay was performed usingHelix pomatia (specific for N-acetylgalactosamine residues)and Ricinus comunis (specific for galactose residues) lectinsSeveral glycoconjugates were recognized by both lectinsthose of 310 287 250 210 182 161 79 77 75 49 47 and 41 kDa(Figure 1(a)) whereas two bands at 51 and 39KDa were onlyrecognized by H pomatia However using only streptavidinperoxidase (Figure 1(b)) two glycoconjugates were recog-nized on TcSol antigens those of 250 and 124 kDaTherefore
49
80
124
209
35
124
250
797775
514947
4139
287310
210182161
124
250
310
210182161
797775
4947
41
287
RCAHPMW
(a)
250
124
SBHP
(b)
Figure 1 Lectin-blot analysis of Taenia crassiceps soluble antigensT crassiceps soluble proteins were subjected to electrophoresison a 10 SDS-polyacrylamide gel transferred to nitrocellulosemembrane and revealed with a couple of different biotinylatedlectins (a) MW-molecular weight HP-Helix pomatia and RCA-Ricinus communis (b) The strips were incubated only with SBHP-streptavidin-bound horseradish peroxidase
the presence of these two biotin-containing proteins mostlikely corresponds to biotin-dependent carboxylases recog-nized by streptavidin alone [25 26] These findings indicatethat TcSol antigens express high levels of glycoconjugateswith N-acetylgalactosamine and galactose residues
32 MGL1minusminus Macrophages Display Reduced Binding to Tae-nia crassiceps Soluble Antigens The innate recognition ofpathogen-associated molecules by antigen-presenting cells iscrucial to the initiation of the immune response Differentstudies have shown that helminth-derived molecules can berecognized by C-type lectins [27 28] In our model wepreviously reported that most TcSol bound to concanavalin
BioMed Research International 5
A indicating that TcSol are glycosylated with glucose man-nose or galactose [17] whereas Jang Lee et al reported thatthe main N-glycan structures in T crassiceps were mannosefucose galactose and GlcNAc They also found a rare Fuc1205721rarr 3GlcNAc antenna on T crassiceps molecules [29] Thusthe natural candidates for carbohydrate recognition on Tcrassiceps are CLRs such as the MR MGL and DC-SIGNThus we hypothesized that one of the possible receptors thatbind the glycomolecules on TcSol may be MGL given itsspecificity to recognize both N-acetylgalactosamine andgalactose residues To investigate the role of MGL1 in TcSolrecognition TcSol antigens were labeled with FITC toperform binding assays TcSol-FITC efficiently bound toMGL1++ macrophages at 37∘C (approximately 30 of cellswere positive) indicating that TcSol-FITC can be recognizedand internalized by these cells (Figures 2(a) and 2(b)) Incu-bation of M120601s and TcSol at 4∘C showed reduced fluorescence(5ndash10) indicating a low level of binding Next we similarlyexposed MGL1minusminus M120601s to TcSol-FITC at 37∘C and foundsignificant reductions in both the percentage of TcSol-FITC-positive cells (approximately 10) and the mean fluorescenceintensity (MFI) of TcSol-FITC on these cells (Figure 2(c))These data suggest that M120601s can recognize TcSol via MGL
33 The Absence of MGL1 on Macrophages Impairs TcSol-Triggered Global Tyrosine Phosphorylation The fact thatTcSol binding was impaired in MGL1minusminusM120601s led us to ques-tion whether the interaction with MGL-TcSol triggers anintracellular signaling pathway Recent advances in theunderstanding of the intracellular signaling induced bystrong antibody-mediated crosslinking of MGL have high-lighted a possible role for the ERK pathway [20] To deter-mine whether TcSol can induce intracellular signaling weexposed BMDM120601s fromMGL1++ or MGL1minusminus mice to TcSolfor 5 15 and 30min and found that tyrosine phosphorylationwas enhanced in MGL1++ macrophages interestingly tyro-sine phosphorylation was detected as early as 5min andremained for at least 30min (Figure 3(a)) In contrastMGL1minusminus BMDM120601s that were similarly exposed to TcSol dis-played a lower and less-sustained tyrosine phosphorylationin several protein bands (Figure 3(a) arrows) To further ana-lyze this response we stimulated bothMGL1++ andMGL1minusminusBMDM120601s with LPS (500 ngmL) or TcSol again for 30minAs observed in Figure 3(b) alsowedetected a lack of responseto LPS and a lower tyrosine phosphorylation in response toTcSol (Figures 3(b) and 3(c) resp) in MGL1minusminus BMDM120601s
34 Mice Lacking MGL1 Are Highly Susceptible to Taenia cras-siceps Infection After determining that T crassiceps antigenscan be recognized by MGL1 and that macrophages deficientin MGL1 are partially refractory to TcSol stimulation weinvestigated the in vivo role of this molecule in experimentalcysticercosis using MGL-deficient mice To approach thisquestion we compared the course of T crassiceps infection inMGL1minusminus with that in MGL1++ mice As a control for sus-ceptibility we used BALBc mice which have consistently
been reported as a highly susceptible strain to T crassicepsinfection [30]
We examined the kinetics of parasite growth from 2 to 8weeks after T crassiceps infection Early in the infection (2weeks) both groups of mice displayed comparable parasiteburdens (Figure 4(a)) Interestingly as infection progressedby the fourth week of infection the number of larvae in theperitoneal cavity increased significantly in MGL1minusminus micecompared to MGL1++ mice which successfully reduced thenumber of parasites by week 8 after infection (Figures 4(a)and 4(b)) In fact MGL1minusminus mice exhibited a parasite burdenthat was very similar to that in the susceptible strain of mice(BALBc) which were consistently observed as very suscepti-ble toT crassiceps infection (Figure 4(a))These findings sug-gest that theMGL1-mediated signaling pathway is involved inresistance during T crassiceps infection on a resistant geneticbackground such as C57BL6
35 MGL1minusminus Mice Display an Altered Immune Responseto Taenia crassiceps A few studies have demonstrated thatMGL can mediate signaling after a strong stimulus such aspolyclonal anti-MGL antibodies but its role in modulatingprotective immunity against helminth parasites is unknownTherefore wemeasured levels ofTh1-associated IgG2a as wellas Th2-associated IgG1 and total IgE antibodies in MGL1minusminus
andMGL1++mice at different timepoints following infectionwith T crassiceps
Early in infection T crassiceps-infected MGL1++ andMGL1minusminus mice displayed comparable levels of T crassicepsAg-specific Th1-associated IgG2a antibodies (data notshown) However by week 8 after infection MGL1++ micedisplayed significantly higher titers of specific IgG2aantibodies againstT crassiceps antigens (Figure 5(a)) By con-trast no clear differences were observed in Th2-associatedIgG1 production MGL1++ mice displayed high titers ofanti-T crassiceps-specific IgG1 that were similar to thosein MGL1minusminus mice at week 8 after infection (Figure 5(b))AlthoughTh2-associated IgE has been shown to play a role inmediating immunity against certain helminths we found thatT crassiceps-infected MGL1++ mice harbored a lower para-site burden despite producing significantly lower levels ofIgE compared to similarly infected MGL1minusminus mice whichdisplayed higher levels of total IgE (Figure 5(c))
Additionally we also compared the cytokine produc-tion by splenocytes from these mice in response to either25 120583gmL TcSol or 1 120583gmL plate-bound anti-CD3 antibodyAnti-CD3 (data not shown) or TcSol-stimulated splenocytesfrom both strains produced similar levels of IFN-120574 at 2 wkafter infection (Figure 6(a)) However 4 and 8 weeks afterinfection the IFN-120574 production by the spleen cells ofMGL1minusminus mice decreased and did not reach the level pro-duced by MGL1++ splenocytes (Figure 6(a)) Similarly asearly as 2 wk after infection the splenocytes from theMGL1minusminus and MGL1++ mice produced comparable levels ofIL-4 (Figure 6(b)) However by weeks 4 and 8 after infectionMGL1minusminus mice produced significantly more IL-4 than the
6 BioMed Research International
F480 F480
TcSo
l-FIT
C
TcSo
l-FIT
C
100 101 102 103 104 100 101 102 103 104
29 6
100
101
102
103
104
100
101
102
103
104MGL1++ 37∘C MGL1minusminus 37∘C
(a)
TcSo
l-FIT
C (
)
lowast
50
40
30
20
10
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(b)
lowast
MFI
500
400
300
200
100
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(c)
Figure 2 Peritoneal M120601s recognize Taenia crassiceps soluble products through MGL1 M120601s from naıve MGL1++ or MGL1minusminus mice wereincubated with fluorescently labeled TcSol and fluorescence was analyzed by flow cytometry (a) Representative dot blots indicate M120601s fromeither MGL1++ or MGL1minusminus mice incubated for 30 minutes with TcSol at 37∘C (b) Percentage of TcSol-FITC positive naıve peritoneal cellsexposed for 30min (c) Data are represented as the mean florescence intensity (MFI) of naıve M120601s exposed to TcES-FITC Basal levels ofautofluorescence were subtracted from all treatments Data are representative of three independent experiments lowast119875 lt 005
MGL1++ splenocytes in response to anti-CD3 whereasantigen-specific higher IL-4 production in MGL1minusminus spleno-cytes was only observed at week 4 after infection comparedwithMGL1++ cells (Figures 6(b) and 6(c))Thus as the infec-tion progressed splenocytes from MGL1minusminus mice producedsignificantly greater levels of IL-4 compared with splenocytes
fromT crassiceps-infectedMGL1++mice in response to anti-CD3
36 Differential Cytokine Production in MGL1minusminus andMGL1++ Peritoneal Macrophages To determine whetherMGL1minusminus mice had a systemic defect in innate activation we
BioMed Research International 7
TcSol
170130
9870
55
42
35
MW
50120583gmL5998400 15998400 30998400 5998400 15998400 30998400
MGL1++ MGL1minusminus
minusminus
(a)
170130
98705542
35
25
TcSolLPS 30998400
3099840030998400 30998400
MGL1++MGL1minusminus
120573-Actin
minusminus
minus
minus minus
minus minusminus
(b)
MGL1++MGL1minusminus
Relat
ive p
hosp
hory
latio
n le
vel
TcSolLPS 30998400
30998400
08
06
04
02
00
3099840030998400minus
minus
minus
minus minus
minus minus
minus
P-Y LPS and TcSol
(c)
Figure 3 MGL1minusminus macrophages display deficient tyrosine phosphorylation in response to TcSol (a) MGL1++ and MGL1minusminus bone marrow-derived M120601 were synchronized in starvation conditions over night and then were stimulated or not with TcSol (50120583gmL) for 5 15 or30min Next total cell extracts were obtained and resolved by electrophoresis transferred to nitrocellulose and probed with monoclonalantiphosphotyrosine antibody Lane 1 serum-starved MGL1++ M120601s Lanes 2ndash4 serum-starved MGL1++ M120601s following TcSol exposure for5 15 and 30min respectively Lane 5 serum-starved MGL1minusminus M120601s Lanes 6ndash8 serum-starved MGL1minusminus M120601s following TcSol exposure for5 15 and 30min respectively Lanes 2ndash4 show an upregulation of the tyrosine phosphorylation of total proteins which was sustained forat least 30min (arrows) Lanes 6ndash8 show an unsustained tyrosine phosphorylation of proteins after similar TcSol stimulation of MGL1minusminusM120601s (b) Similar experiment using LPS (500 ngmL) andor TcSol stimulation for 30min (c) Densitometry of western blot placed in (b)Proteins were visualized using a goat anti-mouse secondary conjugated to HRP and a chemiluminescence detection system Western blotsare representative of two independent experiments
examined the responsiveness of macrophages fromMGL1minusminusmice to proinflammatory stimuli Macrophages were isolatedfrom the peritoneal cavities of both strains of infected miceand either left unstimulated (basal) or stimulated for 24 hwith LPS (1120583gmL) and IFN-120574 (20 ngmL) The supernatantswere collected and analyzed for IL-12 TNF-120572 and NOproduction As shown in Figure 7 macrophages fromMGL1minusminus mice obtained during the early phase of infectionwith T crassiceps (2 wks) produced lower levels of IL-12 and
TNF-120572 compared with those from MGL1++ mice (Figures7(a) and 7(b)) As the infection became chronicmacrophagesfrom infected MGL1minusminus mice produced decreased levels ofTNF-120572 (Figure 7(b)) and theNO levels dropped significantly(Figure 7(c)) in both groups but even more in MGL1minusminusmacrophagesThese patterns ofmacrophage responsewere incontrast to those observed in MGL1++ macrophages whichshowed a better proinflammatory response throughout theinfection with higher production of TNF-120572 and NO in late
8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 5
A indicating that TcSol are glycosylated with glucose man-nose or galactose [17] whereas Jang Lee et al reported thatthe main N-glycan structures in T crassiceps were mannosefucose galactose and GlcNAc They also found a rare Fuc1205721rarr 3GlcNAc antenna on T crassiceps molecules [29] Thusthe natural candidates for carbohydrate recognition on Tcrassiceps are CLRs such as the MR MGL and DC-SIGNThus we hypothesized that one of the possible receptors thatbind the glycomolecules on TcSol may be MGL given itsspecificity to recognize both N-acetylgalactosamine andgalactose residues To investigate the role of MGL1 in TcSolrecognition TcSol antigens were labeled with FITC toperform binding assays TcSol-FITC efficiently bound toMGL1++ macrophages at 37∘C (approximately 30 of cellswere positive) indicating that TcSol-FITC can be recognizedand internalized by these cells (Figures 2(a) and 2(b)) Incu-bation of M120601s and TcSol at 4∘C showed reduced fluorescence(5ndash10) indicating a low level of binding Next we similarlyexposed MGL1minusminus M120601s to TcSol-FITC at 37∘C and foundsignificant reductions in both the percentage of TcSol-FITC-positive cells (approximately 10) and the mean fluorescenceintensity (MFI) of TcSol-FITC on these cells (Figure 2(c))These data suggest that M120601s can recognize TcSol via MGL
33 The Absence of MGL1 on Macrophages Impairs TcSol-Triggered Global Tyrosine Phosphorylation The fact thatTcSol binding was impaired in MGL1minusminusM120601s led us to ques-tion whether the interaction with MGL-TcSol triggers anintracellular signaling pathway Recent advances in theunderstanding of the intracellular signaling induced bystrong antibody-mediated crosslinking of MGL have high-lighted a possible role for the ERK pathway [20] To deter-mine whether TcSol can induce intracellular signaling weexposed BMDM120601s fromMGL1++ or MGL1minusminus mice to TcSolfor 5 15 and 30min and found that tyrosine phosphorylationwas enhanced in MGL1++ macrophages interestingly tyro-sine phosphorylation was detected as early as 5min andremained for at least 30min (Figure 3(a)) In contrastMGL1minusminus BMDM120601s that were similarly exposed to TcSol dis-played a lower and less-sustained tyrosine phosphorylationin several protein bands (Figure 3(a) arrows) To further ana-lyze this response we stimulated bothMGL1++ andMGL1minusminusBMDM120601s with LPS (500 ngmL) or TcSol again for 30minAs observed in Figure 3(b) alsowedetected a lack of responseto LPS and a lower tyrosine phosphorylation in response toTcSol (Figures 3(b) and 3(c) resp) in MGL1minusminus BMDM120601s
34 Mice Lacking MGL1 Are Highly Susceptible to Taenia cras-siceps Infection After determining that T crassiceps antigenscan be recognized by MGL1 and that macrophages deficientin MGL1 are partially refractory to TcSol stimulation weinvestigated the in vivo role of this molecule in experimentalcysticercosis using MGL-deficient mice To approach thisquestion we compared the course of T crassiceps infection inMGL1minusminus with that in MGL1++ mice As a control for sus-ceptibility we used BALBc mice which have consistently
been reported as a highly susceptible strain to T crassicepsinfection [30]
We examined the kinetics of parasite growth from 2 to 8weeks after T crassiceps infection Early in the infection (2weeks) both groups of mice displayed comparable parasiteburdens (Figure 4(a)) Interestingly as infection progressedby the fourth week of infection the number of larvae in theperitoneal cavity increased significantly in MGL1minusminus micecompared to MGL1++ mice which successfully reduced thenumber of parasites by week 8 after infection (Figures 4(a)and 4(b)) In fact MGL1minusminus mice exhibited a parasite burdenthat was very similar to that in the susceptible strain of mice(BALBc) which were consistently observed as very suscepti-ble toT crassiceps infection (Figure 4(a))These findings sug-gest that theMGL1-mediated signaling pathway is involved inresistance during T crassiceps infection on a resistant geneticbackground such as C57BL6
35 MGL1minusminus Mice Display an Altered Immune Responseto Taenia crassiceps A few studies have demonstrated thatMGL can mediate signaling after a strong stimulus such aspolyclonal anti-MGL antibodies but its role in modulatingprotective immunity against helminth parasites is unknownTherefore wemeasured levels ofTh1-associated IgG2a as wellas Th2-associated IgG1 and total IgE antibodies in MGL1minusminus
andMGL1++mice at different timepoints following infectionwith T crassiceps
Early in infection T crassiceps-infected MGL1++ andMGL1minusminus mice displayed comparable levels of T crassicepsAg-specific Th1-associated IgG2a antibodies (data notshown) However by week 8 after infection MGL1++ micedisplayed significantly higher titers of specific IgG2aantibodies againstT crassiceps antigens (Figure 5(a)) By con-trast no clear differences were observed in Th2-associatedIgG1 production MGL1++ mice displayed high titers ofanti-T crassiceps-specific IgG1 that were similar to thosein MGL1minusminus mice at week 8 after infection (Figure 5(b))AlthoughTh2-associated IgE has been shown to play a role inmediating immunity against certain helminths we found thatT crassiceps-infected MGL1++ mice harbored a lower para-site burden despite producing significantly lower levels ofIgE compared to similarly infected MGL1minusminus mice whichdisplayed higher levels of total IgE (Figure 5(c))
Additionally we also compared the cytokine produc-tion by splenocytes from these mice in response to either25 120583gmL TcSol or 1 120583gmL plate-bound anti-CD3 antibodyAnti-CD3 (data not shown) or TcSol-stimulated splenocytesfrom both strains produced similar levels of IFN-120574 at 2 wkafter infection (Figure 6(a)) However 4 and 8 weeks afterinfection the IFN-120574 production by the spleen cells ofMGL1minusminus mice decreased and did not reach the level pro-duced by MGL1++ splenocytes (Figure 6(a)) Similarly asearly as 2 wk after infection the splenocytes from theMGL1minusminus and MGL1++ mice produced comparable levels ofIL-4 (Figure 6(b)) However by weeks 4 and 8 after infectionMGL1minusminus mice produced significantly more IL-4 than the
6 BioMed Research International
F480 F480
TcSo
l-FIT
C
TcSo
l-FIT
C
100 101 102 103 104 100 101 102 103 104
29 6
100
101
102
103
104
100
101
102
103
104MGL1++ 37∘C MGL1minusminus 37∘C
(a)
TcSo
l-FIT
C (
)
lowast
50
40
30
20
10
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(b)
lowast
MFI
500
400
300
200
100
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(c)
Figure 2 Peritoneal M120601s recognize Taenia crassiceps soluble products through MGL1 M120601s from naıve MGL1++ or MGL1minusminus mice wereincubated with fluorescently labeled TcSol and fluorescence was analyzed by flow cytometry (a) Representative dot blots indicate M120601s fromeither MGL1++ or MGL1minusminus mice incubated for 30 minutes with TcSol at 37∘C (b) Percentage of TcSol-FITC positive naıve peritoneal cellsexposed for 30min (c) Data are represented as the mean florescence intensity (MFI) of naıve M120601s exposed to TcES-FITC Basal levels ofautofluorescence were subtracted from all treatments Data are representative of three independent experiments lowast119875 lt 005
MGL1++ splenocytes in response to anti-CD3 whereasantigen-specific higher IL-4 production in MGL1minusminus spleno-cytes was only observed at week 4 after infection comparedwithMGL1++ cells (Figures 6(b) and 6(c))Thus as the infec-tion progressed splenocytes from MGL1minusminus mice producedsignificantly greater levels of IL-4 compared with splenocytes
fromT crassiceps-infectedMGL1++mice in response to anti-CD3
36 Differential Cytokine Production in MGL1minusminus andMGL1++ Peritoneal Macrophages To determine whetherMGL1minusminus mice had a systemic defect in innate activation we
BioMed Research International 7
TcSol
170130
9870
55
42
35
MW
50120583gmL5998400 15998400 30998400 5998400 15998400 30998400
MGL1++ MGL1minusminus
minusminus
(a)
170130
98705542
35
25
TcSolLPS 30998400
3099840030998400 30998400
MGL1++MGL1minusminus
120573-Actin
minusminus
minus
minus minus
minus minusminus
(b)
MGL1++MGL1minusminus
Relat
ive p
hosp
hory
latio
n le
vel
TcSolLPS 30998400
30998400
08
06
04
02
00
3099840030998400minus
minus
minus
minus minus
minus minus
minus
P-Y LPS and TcSol
(c)
Figure 3 MGL1minusminus macrophages display deficient tyrosine phosphorylation in response to TcSol (a) MGL1++ and MGL1minusminus bone marrow-derived M120601 were synchronized in starvation conditions over night and then were stimulated or not with TcSol (50120583gmL) for 5 15 or30min Next total cell extracts were obtained and resolved by electrophoresis transferred to nitrocellulose and probed with monoclonalantiphosphotyrosine antibody Lane 1 serum-starved MGL1++ M120601s Lanes 2ndash4 serum-starved MGL1++ M120601s following TcSol exposure for5 15 and 30min respectively Lane 5 serum-starved MGL1minusminus M120601s Lanes 6ndash8 serum-starved MGL1minusminus M120601s following TcSol exposure for5 15 and 30min respectively Lanes 2ndash4 show an upregulation of the tyrosine phosphorylation of total proteins which was sustained forat least 30min (arrows) Lanes 6ndash8 show an unsustained tyrosine phosphorylation of proteins after similar TcSol stimulation of MGL1minusminusM120601s (b) Similar experiment using LPS (500 ngmL) andor TcSol stimulation for 30min (c) Densitometry of western blot placed in (b)Proteins were visualized using a goat anti-mouse secondary conjugated to HRP and a chemiluminescence detection system Western blotsare representative of two independent experiments
examined the responsiveness of macrophages fromMGL1minusminusmice to proinflammatory stimuli Macrophages were isolatedfrom the peritoneal cavities of both strains of infected miceand either left unstimulated (basal) or stimulated for 24 hwith LPS (1120583gmL) and IFN-120574 (20 ngmL) The supernatantswere collected and analyzed for IL-12 TNF-120572 and NOproduction As shown in Figure 7 macrophages fromMGL1minusminus mice obtained during the early phase of infectionwith T crassiceps (2 wks) produced lower levels of IL-12 and
TNF-120572 compared with those from MGL1++ mice (Figures7(a) and 7(b)) As the infection became chronicmacrophagesfrom infected MGL1minusminus mice produced decreased levels ofTNF-120572 (Figure 7(b)) and theNO levels dropped significantly(Figure 7(c)) in both groups but even more in MGL1minusminusmacrophagesThese patterns ofmacrophage responsewere incontrast to those observed in MGL1++ macrophages whichshowed a better proinflammatory response throughout theinfection with higher production of TNF-120572 and NO in late
8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

6 BioMed Research International
F480 F480
TcSo
l-FIT
C
TcSo
l-FIT
C
100 101 102 103 104 100 101 102 103 104
29 6
100
101
102
103
104
100
101
102
103
104MGL1++ 37∘C MGL1minusminus 37∘C
(a)
TcSo
l-FIT
C (
)
lowast
50
40
30
20
10
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(b)
lowast
MFI
500
400
300
200
100
0
MGL1++
MGL1minusminus
4 4 3737
(∘C)
(c)
Figure 2 Peritoneal M120601s recognize Taenia crassiceps soluble products through MGL1 M120601s from naıve MGL1++ or MGL1minusminus mice wereincubated with fluorescently labeled TcSol and fluorescence was analyzed by flow cytometry (a) Representative dot blots indicate M120601s fromeither MGL1++ or MGL1minusminus mice incubated for 30 minutes with TcSol at 37∘C (b) Percentage of TcSol-FITC positive naıve peritoneal cellsexposed for 30min (c) Data are represented as the mean florescence intensity (MFI) of naıve M120601s exposed to TcES-FITC Basal levels ofautofluorescence were subtracted from all treatments Data are representative of three independent experiments lowast119875 lt 005
MGL1++ splenocytes in response to anti-CD3 whereasantigen-specific higher IL-4 production in MGL1minusminus spleno-cytes was only observed at week 4 after infection comparedwithMGL1++ cells (Figures 6(b) and 6(c))Thus as the infec-tion progressed splenocytes from MGL1minusminus mice producedsignificantly greater levels of IL-4 compared with splenocytes
fromT crassiceps-infectedMGL1++mice in response to anti-CD3
36 Differential Cytokine Production in MGL1minusminus andMGL1++ Peritoneal Macrophages To determine whetherMGL1minusminus mice had a systemic defect in innate activation we
BioMed Research International 7
TcSol
170130
9870
55
42
35
MW
50120583gmL5998400 15998400 30998400 5998400 15998400 30998400
MGL1++ MGL1minusminus
minusminus
(a)
170130
98705542
35
25
TcSolLPS 30998400
3099840030998400 30998400
MGL1++MGL1minusminus
120573-Actin
minusminus
minus
minus minus
minus minusminus
(b)
MGL1++MGL1minusminus
Relat
ive p
hosp
hory
latio
n le
vel
TcSolLPS 30998400
30998400
08
06
04
02
00
3099840030998400minus
minus
minus
minus minus
minus minus
minus
P-Y LPS and TcSol
(c)
Figure 3 MGL1minusminus macrophages display deficient tyrosine phosphorylation in response to TcSol (a) MGL1++ and MGL1minusminus bone marrow-derived M120601 were synchronized in starvation conditions over night and then were stimulated or not with TcSol (50120583gmL) for 5 15 or30min Next total cell extracts were obtained and resolved by electrophoresis transferred to nitrocellulose and probed with monoclonalantiphosphotyrosine antibody Lane 1 serum-starved MGL1++ M120601s Lanes 2ndash4 serum-starved MGL1++ M120601s following TcSol exposure for5 15 and 30min respectively Lane 5 serum-starved MGL1minusminus M120601s Lanes 6ndash8 serum-starved MGL1minusminus M120601s following TcSol exposure for5 15 and 30min respectively Lanes 2ndash4 show an upregulation of the tyrosine phosphorylation of total proteins which was sustained forat least 30min (arrows) Lanes 6ndash8 show an unsustained tyrosine phosphorylation of proteins after similar TcSol stimulation of MGL1minusminusM120601s (b) Similar experiment using LPS (500 ngmL) andor TcSol stimulation for 30min (c) Densitometry of western blot placed in (b)Proteins were visualized using a goat anti-mouse secondary conjugated to HRP and a chemiluminescence detection system Western blotsare representative of two independent experiments
examined the responsiveness of macrophages fromMGL1minusminusmice to proinflammatory stimuli Macrophages were isolatedfrom the peritoneal cavities of both strains of infected miceand either left unstimulated (basal) or stimulated for 24 hwith LPS (1120583gmL) and IFN-120574 (20 ngmL) The supernatantswere collected and analyzed for IL-12 TNF-120572 and NOproduction As shown in Figure 7 macrophages fromMGL1minusminus mice obtained during the early phase of infectionwith T crassiceps (2 wks) produced lower levels of IL-12 and
TNF-120572 compared with those from MGL1++ mice (Figures7(a) and 7(b)) As the infection became chronicmacrophagesfrom infected MGL1minusminus mice produced decreased levels ofTNF-120572 (Figure 7(b)) and theNO levels dropped significantly(Figure 7(c)) in both groups but even more in MGL1minusminusmacrophagesThese patterns ofmacrophage responsewere incontrast to those observed in MGL1++ macrophages whichshowed a better proinflammatory response throughout theinfection with higher production of TNF-120572 and NO in late
8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 7
TcSol
170130
9870
55
42
35
MW
50120583gmL5998400 15998400 30998400 5998400 15998400 30998400
MGL1++ MGL1minusminus
minusminus
(a)
170130
98705542
35
25
TcSolLPS 30998400
3099840030998400 30998400
MGL1++MGL1minusminus
120573-Actin
minusminus
minus
minus minus
minus minusminus
(b)
MGL1++MGL1minusminus
Relat
ive p
hosp
hory
latio
n le
vel
TcSolLPS 30998400
30998400
08
06
04
02
00
3099840030998400minus
minus
minus
minus minus
minus minus
minus
P-Y LPS and TcSol
(c)
Figure 3 MGL1minusminus macrophages display deficient tyrosine phosphorylation in response to TcSol (a) MGL1++ and MGL1minusminus bone marrow-derived M120601 were synchronized in starvation conditions over night and then were stimulated or not with TcSol (50120583gmL) for 5 15 or30min Next total cell extracts were obtained and resolved by electrophoresis transferred to nitrocellulose and probed with monoclonalantiphosphotyrosine antibody Lane 1 serum-starved MGL1++ M120601s Lanes 2ndash4 serum-starved MGL1++ M120601s following TcSol exposure for5 15 and 30min respectively Lane 5 serum-starved MGL1minusminus M120601s Lanes 6ndash8 serum-starved MGL1minusminus M120601s following TcSol exposure for5 15 and 30min respectively Lanes 2ndash4 show an upregulation of the tyrosine phosphorylation of total proteins which was sustained forat least 30min (arrows) Lanes 6ndash8 show an unsustained tyrosine phosphorylation of proteins after similar TcSol stimulation of MGL1minusminusM120601s (b) Similar experiment using LPS (500 ngmL) andor TcSol stimulation for 30min (c) Densitometry of western blot placed in (b)Proteins were visualized using a goat anti-mouse secondary conjugated to HRP and a chemiluminescence detection system Western blotsare representative of two independent experiments
examined the responsiveness of macrophages fromMGL1minusminusmice to proinflammatory stimuli Macrophages were isolatedfrom the peritoneal cavities of both strains of infected miceand either left unstimulated (basal) or stimulated for 24 hwith LPS (1120583gmL) and IFN-120574 (20 ngmL) The supernatantswere collected and analyzed for IL-12 TNF-120572 and NOproduction As shown in Figure 7 macrophages fromMGL1minusminus mice obtained during the early phase of infectionwith T crassiceps (2 wks) produced lower levels of IL-12 and
TNF-120572 compared with those from MGL1++ mice (Figures7(a) and 7(b)) As the infection became chronicmacrophagesfrom infected MGL1minusminus mice produced decreased levels ofTNF-120572 (Figure 7(b)) and theNO levels dropped significantly(Figure 7(c)) in both groups but even more in MGL1minusminusmacrophagesThese patterns ofmacrophage responsewere incontrast to those observed in MGL1++ macrophages whichshowed a better proinflammatory response throughout theinfection with higher production of TNF-120572 and NO in late
8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

8 BioMed Research International
2 4 80
200
400
600
800
Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowastlowast
BALBcMGL1++
MGL1minusminus
(a)
2 4 8Weeks after infection
Num
ber o
f par
asite
sm
ouse
lowast
lowastlowast
800
600
400
200
150
100
50
0
MGL1++
MGL1minusminus
(b)
Figure 4 MGL1minusminus mice do not efficiently control T crassiceps infection (a) Course of ip T crassiceps infection in MGL1minusminus MGL1++ andBALBc mice after infection with 20 cysticerci (b) An independent experiment of the course of ip T crassiceps infection in MGL1minusminus andMGL1++ mice that were similarly infected Data are expressed as the mean plusmn SE of 6 mice per group lowast119875 lt 001 comparing MGL1minusminus versusMGL1++ at the same time point
infections compared to MGL1minusminus macrophages (Figures 7(b)and 7(c))
37 MGL1minusminus Mice Recruit Alternatively Activated Macro-phages Next the macrophage polarization genotype wasevaluated in adherent peritoneal cells at 4 and 8 weeks afterinfection RT-PCR was performed to identify mRNA tran-scripts of AAM120601s markers Infected MGL1minusminus mice showedhigher expression of arginase-1 Relm-120572 and Ym-1 thantheir MGL1++ counterparts (Figure 8(a)) Moreover we alsoanalyzed themacrophage surface expression of othermarkersassociated with alternative activation by flow cytometry Sur-face expression of theMR IL-4R120572 and PD-L2 has previouslybeen associated with AAM120601s in distinct helminth infections[31] Indeed we found differences between the two strainsin the expression of these molecules during T crassicepsinfection Interestingly during chronic infection (8 weeks)the macrophages from MGL1minusminus mice expressed twice asmuch PD-L2 as macrophages from MGL1++ mice whichdownregulated PD-L2 expression (Figures 8(b) and 8(c))TheMR and IL-4R120572 expression were also been significantlyelevated inmacrophages fromT crassiceps-infectedMGL1minusminus
mice when compared with MGL1++ mice that expressedlower percentages of both markers (Figures 8(b) and 8(c))
Given that two related MGL molecules do exist in miceMGL1 and MGL2 with different carbohydrate specificities
we performed flow cytometry on peritoneal cells for thedetection of MGL1 and MGL2 in order to further determinethe specific role for MGL1 and not for MGL2 in susceptibilityto T crassiceps As shown in Figure 8(d) peritoneal macro-phages (F480+) from MGL1++ infected mice did increaseMGL1 as well as MGL2 expression after 8 weeks of infectionwhereasMGL1minusminusmacrophages were able to expressMGL2 ata similar level than MGL1++ mice These data suggest a veryfine discrimination between MGL1 and MGL2 to recognizeglycan structures on T crassiceps and its antigens
4 Discussion
The role of CLRs in the recognition of microbial fungaland parasitic glycoconjugates has become evident in the lastfew years [5] The knowledge in this area has passed fromunderstanding them as ldquosimple endocytic receptorsrdquo toimportant molecules involved in innate immunity with thecapacity to trigger intracellular signaling pathways to modifycellular responses to different pathogens [32] Although therole for CLRs in immunity against fungal and bacterialinfections is widely recognized their role in immunity duringhelminth infections is much less known [13] Therefore inthis study we focused on the role of MGL1 and MGL1-mediated signaling in immunity to the cestode T crassiceps
BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 9
IgG2
aO
D405
nm
Sera 1dilution
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
lowast
lowast
lowast
(a)
Sera 1dilution
Spec
ific I
gG1
OD
405
nm
05
04
03
02
01
0010 100 1000 10000 100000
mMGL++
mMGLminusminus
(b)
0 2 4 8 0
2000
4000
6000
8000
10000
12000
Tota
l IgE
(ng
mL)
Weeks after infection
lowast
lowast
lowast
(c)
Figure 5 Antibody production during T crassiceps infection inMGL1minusminus andMGL1++ mice (a) Anti-T crassiceps-specific IgG2a (b) anti-Tcrassiceps-specific IgG1 (c) total IgE Values are the mean plusmn SE (119899 = 6 animals) and are representative of three independent experimentslowast
119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the same time point
It has been previously reported that T crassiceps antigensare rich in structures recognized by concanavalin A Here wefound that in addition to this and to the previously reportedfucose [17 29] glycosylation moieties in T crassiceps such asgalactose and N-acetylgalactosamine are also importantBecause the carbohydrates in T crassiceps antigens as well asin those of other helminths are considered important fortheir modulatory activities [17] we investigated the role of
MGL in the ability to recognize glycosylated structures onTcSol as well as the possible intracellular signaling pathwaysthat they may trigger The lower recognition of TcSol byMGL1minusminusM120601s strongly suggested a role for this C-type lectinin the host-parasite interaction during experimental cysticer-cosis Concomitantly the decreased ability of MGL1minusminus mac-rophages to bind TcSol was associated with weak phos-photyrosine-mediated intracellular signaling Therefore we
10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

10 BioMed Research International
2 4 80
Ant
igen
spec
ific I
FN-120574
(pg
mL)
1000
800
600
400
200
0
lowast
lowast
MGL1++
MGL1minusminus
Weeks after infection
(a)
2 4 8Weeks after infection
0
Ant
igen
spec
ific I
L-4
(pg
mL)
1500
1000
500
0
lowast
mMGL++
mMGLminusminus
(b)
2 4 8Weeks after infection
0
Ant
i CD3
IL-4
(pg
mL)
8000
6000
4000
2000
0
lowast
lowast
mMGL++
mMGLminusminus
(c)
Figure 6 Kinetics of in vitro cytokine production by TcSol-stimulated spleen cells from MGL1minusminus and MGL1++ mice (a) Antigen-specificIFN-120574 production (b) antigen-specific IL-4 production and (c) polyclonal IL-4 production (anti-CD3) in response to in vitro stimulationwith TcSol (25 120583gmL) by splenocytes after 72 h Data are representative of 2 independent experiments lowast119875 lt 005
performed in vivo assays to determine whether MGL1 recog-nition of T crassiceps plays a critical role in the resistance tothis helminth infection Thus we demonstrate for the firsttime that mice deficient in the CLR macrophage galactose-type lectin 1 (MGL1) have increased susceptibility to experi-mental cysticercosis caused by the cestode T crassiceps witha reduction in the levels of the proinflammatory cytokines
IL-12 IFN-120574 and TNF-120572 as well as in NO production Theseobservations are in line with those reported using anti-MGLantibody engagement on DCs where MGL-engaged DCsproduced higher levels of IL-10 than did DCs treated with thecontrol isotype [20] In contrast higher levels of IL-4 and IgEwere detected in these mice compared to similarly infectedMGL1++ mice AlthoughTh2-associated responses and high
BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 11
12000
10000
8000
4000
2000
0
2 4 8
IL-1
2(p
gm
L)
MGL1++
MGL1minusminus
Weeks after infection
(a)
TNF-
120572(p
gm
L)
4000
3000
2000
1000
0
lowast
lowast
lowast
MGL1++
MGL1minusminus
0 2 4 8BasalWeeks after infection
(b)
Nitr
ates
(120583M
)
100
75
50
25
0
0 2 4 8BasalWeeks after infection
lowast
lowastlowast
MGL1++
MGL1minusminus
(c)
Figure 7 Peritoneal macrophages from MGL1minusminus and MGL1++ T crassiceps-infected mice display different responses Macrophages wereobtained at different time points after infection and stimulated with LPS (1120583gmL) plus IFN-120574 (5 ngmL) for 48 h supernatants were analyzedfor (a) IL-12 (b) TNF-120572 and (c) NO production Data are expressed as in Figure 2 lowast119875 lt 005 Data are representative of 2 independentexperiments lowast119875 lt 005
12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

12 BioMed Research International
lowastlowast
lowastlowast
Arg
inas
eG
APD
H ra
tio
Ym-1
GA
PDH
ratio
Relm
-120572G
APD
H ra
tio
iNO
SG
APD
H ra
tio
20
15
10
05
00
20
15
10
05
00
25
20
15
10
05
00
3
2
1
0
lowast
lowast
lowast
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
MG
L1++
cont
rol
MG
L1minusminus
cont
rol
MG
L1++
T cr
assic
eps
MG
L1minusminus
T cr
assic
eps
(a)
MGL1++ MGL1minusminus
MGL1++ MGL1minusminus
100 101 102 103 104
100 101 102 103 104 100 101 102 103 104
100 101 102 103 104
100
101
102
103
104
100
101
102
103
104
100
101
102
103
104
F48
0FI
TC
FSC-H
PD-L
2
PD-L1PD-L1
PD-L
2
PDL2
PE
PDL2
PE
PDL1 APC PDL1 APC
50
40
30
20
10
0
Cou
nts
IL4RIL-4R120572
2521 512
(b)
Figure 8 Continued
BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 13
lowast
lowast80
60
40
20
0
PDL2+ MR+
Posit
ive c
ells
()
MGL1++
MGL1minusminus
IL-4R120572
(c)
MG
L1
MG
L1
MG
L1
MG
L1M
GL1
and
2
MG
L1an
d2
MG
L1an
d2
MG
L1an
d2
66 167 161 32
97 361 21 13210000 10000 10000 10000
10000100001000010000
10000 10000 10000 10000
10000100001000010000
F480 F480 F480 F480
F480 F480 F480 F480
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
MGL1++ control MGL1++ T crassiceps MGL1minusminus control MGL1minusminus T crassiceps
(d)
Figure 8 MGL1minusminus mice recruit alternatively activated macrophages Peritoneal macrophages were obtained fromMGL1minusminus and MGL1++ Tcrassiceps-infectedmice and processed for RT-PCRor flow cytometry analyses (a) RT-PCR showing the expression of arginase-1 Ym1 RELM-120572 and iNOS on macrophages obtained at 8 weeks after infection with T crassiceps (b) Representative dot plot and histograms demonstratingincreased expression of PD-L2 and IL-4R120572 onMGL1minusminusmacrophages (c) Percentages of expression of different surfacemarkers on peritonealmacrophages (d) Dot plots from PECs showing the expression of MGL1 and MGL2 from uninfected (control) and 8 weeks-infected Tcrassiceps MGL1++ and MGL1minusminus mice numbers on the quadrants indicate average percentage of positive cells Values are the mean plusmn SE(119899 = 6 animals) and are representative of two independent experiments lowast119875 lt 005 comparing MGL1minusminus versus MGL1++ mice at the sametime point
14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

14 BioMed Research International
levels of IgE correlate with protection in gastrointestinalhelminth infections [33] our data suggest that IgEmay have alimited role inmediating protective immunity againstT cras-siceps Notably despite the greater Th2-associated antibodyresponse in MGL1minusminus mice these mice displayed greater sus-ceptibility toT crassicepsThese data agreewith those reportedby others [34 35] and indicate a new and finely tuned role forMGL1 in activating innate and adaptive immune responsesagainst this parasiteThese results also suggest that theMGL1-dependent signaling pathway is not only required for inflam-matory cytokine production in the early phase of the hostresponse toT crassiceps infection but also thatMGLmay playa critical role in the development of Th1-adaptive immunityboth of which may be essential for limiting infection andreducing pathology during experimental cysticercosis
In vivo MGL1minusminus mice significantly increased recruit-ment of alternatively activated macrophages as determinedby mRNA transcripts for arginase-1 Ym-1 and FIZZ1 andhigher expression of the MR IL-4R120572 PD-L2 and MGL2 intheir membranes Thus a clear alternative activation statusin macrophages was observed only in MGL1minusminus mice Thesefindings are in line with previous reports showing alterna-tively activated macrophages associated with susceptibility inthis model [24] and also indicate that MGL1 expression is notnecessary for the induction of AAM120601s [36] Together thesedata indicate a role for MGL1 in the regulation of inflam-mation in a model of experimental cysticercosis and suggestthat MGL1 may be a critical innate factor in the response tohelminths and may be to other parasites
Although many helminth parasites express a large num-ber of carbohydrates few CLRs have been associated withhelminth recognition and in vivo function for exampleDectin-2 DC-SIGN SIGNR3 SIGNR1 and MGL have beenshown to recognize several components of S mansoni [13]but their roles in vivo are unclear On the other hand the MRhas been associated with uptake of cercariae [12] but no oneCLR has been demonstrated to be essential for resistance tosuch infection Moreover in another helminth infection theabsence of the MR during Trichuris muris challenge wasirrelevant for either an immune response or resistance tothis worm infection [14] However MRminusminusmice were recentlydemonstrated to exhibit increased survival to Mesocestoidescorti infection a neurocysticercosis model mainly throughthe downregulation of inflammatory responses [37] Theseobservations highlight the role of different CLRs in address-ing distinct helminth infections and also suggest that it ispossible to identify differential roles of CLRs depending uponthe type of helminth challenge
The specificity of rodent MGL1 has been addressed inat least two papers [38 39] where was shown MGL1 has aprimary specificity for the Lewis X determinant and it mayadditionally bind biantennary glycans ending in LacNAc orLacDiNAc We have previously shown that Tcsol containsglycoconjugates binding ConA may be N-glycans and the Tcrassicepsmetacestode N-glycans described by Jang Lee et al[29] include biantennary and triantennary LacNAc-termin-ated structures that could explain our MGL1 bindingobserved Alternatively or complementarily the Fuc alpha1-3GlcNAc antenna might constitute MGL1 targets as the motif
is related to Lewis X The fact that MGL2 expression was un-affected by MGL1 deletion suggest that such targets may beuseful to trigger better immunity against this helminth
Here we add to the knowledge of the role of CLRs inmediating possible immune activation and resistance in vivoto the helminthT crassiceps whichmaymediate cellular acti-vation via tyrosine phosphorylation These data agree withthose recently reported byMishra et al [37] who described apotential role for the MR in inducing proinflammatory res-ponses to the cestode M corti Thus it is possible thatsome CLRs are involved in proinflammatory responseswhereas other CLRs may induce regulatory responses as hasbeen observed in vitro for different helminth antigen-derivedmolecules that are rich in carbohydrate residues [40 41] Infact several helminth-derived glycoconjugates have beenshown to downregulate the inflammatory response of DCs inresponse to different TLR ligands [41 42] and a possiblesignaling through the c-RAF-dependent pathway has recentlybeen proposed as a modulator of TLR-mediated inflamma-tory responses [40] All these data support the idea previouslysuggested by Tealersquos group [11] indicating that given thecomplexity of helminth-derived glycomolecules it may beexpected that they play distinct roles including roles in strongregulatory mechanisms as well as the induction of protectiveresponses which in turn are likely dependent on the specificCLRs that bind such glycoconjugates
In summary in this study we have demonstrated for thefirst time that cestode-derived molecules are recognized byMGL1 on M120601s and that such interaction triggers phospho-tyrosine-mediated intracellular signaling which most likelyinduces inflammatory responses in M120601s and affects theimmune response as well as possible resistance toT crassicepsinfection
Conflict of Interests
The authors declare that there is no conflict of interestsregarding the publication of this paper
Acknowledgments
This work was supported by grants from Direccion Generalde Apoyo al Personal Academico-UNAM IN213512 Institutode Ciencia y Tecnologıa del Distrito Federal 3192010 andPINV11-16 (3012001) and Consejo Nacional de Ciencia yTecnologıa 167799 It is part of the requirements to obtain thePhDdegree in the postgraduate programDoctorado enCien-cias Biomedicas UNAM for Daniel Montero-Barrera whowas supported by a fellowship from CONACYT-Mexico
References
[1] S Akira and K Takeda ldquoToll-like receptor signallingrdquo NatureReviews Immunology vol 4 no 7 pp 499ndash511 2004
[2] J Aliberti J G Valenzuela V B Carruthers et al ldquoMolecularmimicry of a CCR5 binding-domain in themicrobial activationof dendritic cellsrdquo Nature Immunology vol 4 no 5 pp 485ndash490 2003
BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

BioMed Research International 15
[3] C Brodskyn J Patricio R Oliveira et al ldquoGlycoinositolphos-pholipids from Trypanosoma cruzi interfere with macrophagesand dendritic cell responsesrdquo Infection and Immunity vol 70no 7 pp 3736ndash3743 2002
[4] K V Vukman A Ravida A M Aldridge and S M OrsquoNeillldquoMannose receptor and macrophage galactose-type lectin areinvolved in Bordetella pertussis mast cell interactionrdquo Journalof Leukocyte Biology vol 94 no 3 pp 439ndash448 2013
[5] A Vazquez-Mendoza J C Carrero and M Rodriguez-SosaldquoParasitic infections a role for C-type lectins receptorsrdquoBioMedResearch International vol 2013 Article ID 456352 11 pages2013
[6] G D Brown and S Gordon ldquoImmune recognition of fungal 120573-glucansrdquo Cellular Microbiology vol 7 no 4 pp 471ndash479 2005
[7] E P McGreal J L Miller and S Gordon ldquoLigand recognitionby antigen-presenting cell C-type lectin receptorsrdquo CurrentOpinion in Immunology vol 17 no 1 pp 18ndash24 2005
[8] R M Maizels and M Yazdanbakhsh ldquoImmune regulation byhelminth parasites cellular andmolecularmechanismsrdquoNatureReviews Immunology vol 3 no 9 pp 733ndash744 2003
[9] M Okano A R Satoskar K Nishizaki M Abe and D A HarnJr ldquoInduction ofTh2 responses and IgE is largely due to carbo-hydrates functioning as adjuvants on Schistosomamansoni eggantigensrdquo Journal of Immunology vol 163 no 12 pp 6712ndash67171999
[10] S Tundup L Srivastava andDAHarn Jr ldquoPolarization of hostimmune responses by helminth-expressed glycansrdquo Annals ofthe New York Academy of Sciences vol 1253 pp E1ndashE13 2012
[11] J I Alvarez J Rivera and J M Teale ldquoDifferential release andphagocytosis of tegument glycoconjugates in neurocysticerco-sis implications for immune evasion strategiesrdquo PLoS NeglectedTropical Diseases vol 2 no 4 article e218 2008
[12] R A Paveley S A Aynsley J D Turner et al ldquoThe MannoseReceptor (CD206) is an important pattern recognition receptor(PRR) in the detection of the infective stage of the helminthSchistosoma mansoni and modulates IFN120574 productionrdquo Inter-national Journal for Parasitology vol 41 no 13-14 pp 1335ndash13452011
[13] N S PrasanphanichM LMickum JHeimburg-Molinaro andR D Cummings ldquoGlycoconjugates in host-helminth interac-tionsrdquo Frontiers in Immunology vol 4 article 240 2013
[14] M L Deschoolmeester L Martinez-Pomares S Gordon andK J Else ldquoThe mannose receptor binds Trichuris muris excre-torysecretory proteins but is not essential for protective immu-nityrdquo Immunology vol 126 no 2 pp 246ndash255 2009
[15] L Gomez-Garcıa I Rivera-Montoya M Rodrıguez-Sosa andL I Terrazas ldquoCarbohydrate components of Taenia crassi-cepsmetacestodes displayTh2-adjuvant and anti-inflammatoryproperties when co-injected with bystander antigenrdquo Parasitol-ogy Research vol 99 no 4 pp 440ndash448 2006
[16] J K Nono K Pletinckx M B Lutz and K Brehm ldquoExcretorysecretory-products of echinococcus multilocularis larvae ind-uce apoptosis and tolerogenic properties in dendritic cells invitrordquo PLoS Neglected Tropical Diseases vol 6 no 2 Article IDe1516 2012
[17] C A Terrazas L Gomez-Garcıa and L I Terrazas ldquoImpairedpro-inflammatory cytokine production and increased Th2-biasing ability of dendritic cells exposed to Taenia excretedsecreted antigens a critical role for carbohydrates but not forSTAT6 signalingrdquo International Journal for Parasitology vol 40no 9 pp 1051ndash1062 2010
[18] L I Terrazas F Sanchez-Munoz M Perez-Miranda et alldquoHelminth excretedsecreted antigens repress expression ofLPS-induced let-7i but not mir-146a and mir-155 in humandendritic cellsrdquoBioMedResearch International vol 2013 ArticleID 972506 6 pages 2013
[19] L Gomez-Garcıa L M Lopez-Marın R Saavedra J L ReyesM Rodrıguez-Sosa and L I Terrazas ldquoIntact glycans fromcestode antigens are involved in innate activation of myeloidsuppressor cellsrdquo Parasite Immunology vol 27 no 10-11 pp395ndash405 2005
[20] C Napoletano I G Zizzari A Rughetti et al ldquoTargeting ofmacrophage galactose-type C-type lectin (MGL) induces DCsignaling and activationrdquo European Journal of Immunology vol42 no 4 pp 936ndash945 2012
[21] H Towbin T Staehelin and J Gordon ldquoElectrophoretic trans-fer of proteins frompolyacrylamide gels to nitrocellulose sheetsprocedure and some applicationsrdquo Proceedings of the NationalAcademy of Sciences of theUnited States of America vol 76 no 9pp 4350ndash4354 1979
[22] M B Lutz N Kukutsch A L J Ogilvie et al ldquoAn advancedculture method for generating large quantities of highly puredendritic cells from mouse bone marrowrdquo Journal of Immuno-logical Methods vol 223 no 1 pp 77ndash92 1999
[23] P Migliorini G Corradin and S B Corradin ldquoMacrophageNOminus2
production as a sensitive and rapid assay for the quantita-tion of murine IFN-120574rdquo Journal of Immunological Methods vol139 no 1 pp 107ndash114 1991
[24] J L Reyes C A Terrazas J Alonso-Trujillo N van Rooijen AR Satoskar and L I Terrazas ldquoEarly removal of alternativelyactivated macrophages leads to Taenia crassiceps cysticercosisclearance in vivordquo International Journal for Parasitology vol 40no 6 pp 731ndash742 2010
[25] S Jitrapakdee M St Maurice I Rayment W W Cleland J CWallace and P V Attwood ldquoStructure mechanism and regula-tion of pyruvate carboxylaserdquoThe Biochemical Journal vol 413no 3 pp 369ndash387 2008
[26] L K Fritz-Laylin S E Prochnik M L Ginger et al ldquoThegenome of Naegleria gruberi illuminates early eukaryotic ver-satilityrdquo Cell vol 140 no 5 pp 631ndash642 2010
[27] E van Liempt S J van Vliet A Engering et al ldquoSchistosomamansoni soluble egg antigens are internalized by human den-dritic cells through multiple C-type lectins and suppress TLR-induced dendritic cell activationrdquo Molecular Immunology vol44 no 10 pp 2605ndash2615 2007
[28] L Guasconi M C Serradell A P Garro L Iacobelli and D TMasih ldquoC-type lectins on macrophages participate in theimmunomodulatory response to Fasciola hepatica productsrdquoImmunology vol 133 no 3 pp 386ndash396 2011
[29] J Jang Lee S DissanayakeM Panico H RMorris A Dell andS M Haslam ldquoMass spectrometric characterisation of Taeniacrassicepsmetacestode N-glycansrdquo Molecular and BiochemicalParasitology vol 143 no 2 pp 245ndash249 2005
[30] J L Reyes C A Terrazas L Vera-Arias and L I Terrazas ldquoDif-ferential response of antigen presenting cells from susceptibleand resistant strains of mice to Taenia crassiceps infectionrdquo Infe-ction Genetics and Evolution vol 9 no 6 pp 1115ndash1127 2009
[31] L I Terrazas D Montero C A Terrazas J L Reyes and MRodrıguez-Sosa ldquoRole of the programmed Death-1 pathway inthe suppressive activity of alternatively activated macrophagesin experimental cysticercosisrdquo International Journal for Para-sitology vol 35 no 13 pp 1349ndash1358 2005
16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

16 BioMed Research International
[32] J C Hoving G J Wilson and G D Brown ldquoSignalling C-typelectin receptors microbial recognition and immunityrdquo CellularMicrobiology vol 16 no 2 pp 185ndash194 2014
[33] J E Allen and R M Maizels ldquoDiversity and dialogue in immu-nity to helminthsrdquo Nature Reviews Immunology vol 11 no 6pp 375ndash388 2011
[34] L I Terrazas A R Satoskar M Rodriguez-Sosa and JMorales-Montor ldquoImmunology and cell biology of parasiticdiseases 2013rdquo BioMed Research International vol 2013 ArticleID 101268 4 pages 2013
[35] A N Peon A Espinoza-Jimenez and L I Terrazas ldquoImmu-noregulation by taenia crassiceps and its antigensrdquo BioMedResearch International vol 2013 Article ID 498583 13 pages2013
[36] G Raes L Brys B K Dahal et al ldquoMacrophage galactose-type C-type lectins as novel markers for alternatively activatedmacrophages elicited by parasitic infections and allergic airwayinflammationrdquo Journal of Leukocyte Biology vol 77 no 3 pp321ndash327 2005
[37] P K Mishra E G Morris J A Garcia A E Cardona and JM Tealea ldquoIncreased accumulation of regulatory granulocyticmyeloid cells in mannose receptor C type 1-deficient mice cor-relates with protection in amousemodel of neurocysticercosisrdquoInfection and Immunity vol 81 no 4 pp 1052ndash1063 2013
[38] M Tsuiji M Fujimori Y Ohashi et al ldquoMolecular cloning andcharacterization of a novel mouse macrophage C-type lectinmMGL2 which has a distinct carbohydrate specificity frommMGL1rdquo The Journal of Biological Chemistry vol 277 no 32pp 28892ndash28901 2002
[39] P J Coombs M E Taylor and K Drickamer ldquoTwo categoriesof mammalian galactose-binding receptors distinguished byglycan array profilingrdquo Glycobiology vol 16 no 8 pp 1Cndash7C2006
[40] C A Terrazas M Alcantara-Hernandez L Bonifaz L I Ter-razas and A R Satoskar ldquoHelminth-excretedsecreted prod-ucts are recognized by multiple receptors on DCs to block theTLR response and bias Th2 polarization in a cRAF dependentpathwayrdquoTheFASEB Journal vol 27 no 11 pp 4547ndash4560 2013
[41] E J Klaver L M Kuijk L C Laan et al ldquoTrichuris suis-inducedmodulation of human dendritic cell function is glycan-mediatedrdquo International Journal for Parasitology vol 43 no 3-4pp 191ndash200 2013
[42] P G Thomas M R Carter O Atochina et al ldquoMaturation ofdendritic cell 2 phenotype by a helminth glycan uses a Toll-likereceptor 4-dependent mechanismrdquoThe Journal of Immunologyvol 171 no 11 pp 5837ndash5841 2003
Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology

Submit your manuscripts athttpwwwhindawicom
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Anatomy Research International
PeptidesInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporation httpwwwhindawicom
International Journal of
Volume 2014
Zoology
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Molecular Biology International
GenomicsInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
The Scientific World JournalHindawi Publishing Corporation httpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioinformaticsAdvances in
Marine BiologyJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Signal TransductionJournal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
BioMed Research International
Evolutionary BiologyInternational Journal of
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Biochemistry Research International
ArchaeaHindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Genetics Research International
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Advances in
Virolog y
Hindawi Publishing Corporationhttpwwwhindawicom
Nucleic AcidsJournal of
Volume 2014
Stem CellsInternational
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
Enzyme Research
Hindawi Publishing Corporationhttpwwwhindawicom Volume 2014
International Journal of
Microbiology