Plusieurs types de division cellulaire Cellules somatiques Cellules humaines en culture Embryon de...
22
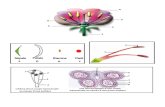
ieurs types de division cellula Cellules somatique s Cellules humaines en culture Embryon de Xenope après 4-5 divisions au stade 8-10 blastomères (ou cellules) Première division méiotique chez un Ovocyte humain, Xenope, etoile de mer… Cellules embryonnaire s Cellules germinales : ovocytes 1mm 1mm Vésicule germinale (noyau) 1mm Globule polaire division Levures Figure 1.1
-
Upload
enide-larue -
Category
Documents
-
view
109 -
download
3
Transcript of Plusieurs types de division cellulaire Cellules somatiques Cellules humaines en culture Embryon de...

Plusieurs types de division cellulaire
Cellulessomatiques
Cellules humainesen culture
Embryon de Xenope après 4-5 divisions au stade 8-10 blastomères
(ou cellules)Première division méiotique chez un Ovocyte humain, Xenope, etoile de
mer…
Cellulesembryonnaires
Cellules germinales : ovocytes
1mm
1mm
Vésicule germinale(noyau)
1mm
Globule polaire
division
Levures
Figure 1.1

Embryon précoce de Drosophile 8 minutes
Embryon précoce de Xenope 30 minutes
Embryon précoce d’Oursin 60 minutes
Schizosaccharomyces pombe ou levure du boulanger ou « fission yeast » 60 minutes
Saccharromyces cerevisiae ou levure de bière ou « budding yeast » 90 minutes
Cellule humaine en culture 18 heures
Figure 1.2 : Durée des divisions cellulaires dans divers types de cellules.

Figure 1.3.1 : Evènements observables durant les phases du cycle cellulaire.
Cytocinèse
Télo
phas
e
An
aph
ase
Mét
aph
ase
Prophase
Prom
étap
hase
Croissance
Croissance et réplication de l’ADN
Croissance et Derniers préparatifs
en vue de la division
SG2
MG1
Phase Go
Point de restriction

Interphase Métaphase Interphase
noyau
réticulumendoplasmique
golgi
Dans les cellules HeLa environ 130 vésicules (ou fragments de golgi) d’un diamètre moyen de 60 nm se dispersent dans le cytoplasme.
Des vésicules mixtes de membranes denoyau et de reticulum de forment.
En fin d’anaphase les membranes vésiculaires mixtes de noyau et de réticulum se reforment progressivement à partir de leur organisation autour de la chromatine. Les vésicules de golgi fusionnent pour reformer l’organite.
Figure 1.3.2. : Devenir des organites au cours de la division cellulaire.

Figure 1.4 : Variations des types de cycle cellulaire et des cellules produites.
G1 S G2 M
S M
M
S
Nombre de chromosomesdivisés par deux
Nombre de chromosomesmultipliés par deux
Volume cellulairedivisé par deux
A
B
C
D

Figure 1.5 : Succession des types de cycle cellulaire.
G1 S G2 MS MM M
MéioseMitose de
l’embryon précoceMitose de
l’embryon et adulte

Figure 1.6 : Place du Cycle cellulaire dans les destins cellulaires.
Cycle de divisioncellulaire
C
A
B
D
F
E
Quiescence ouphase G0
Différenciation
Z
Z
Apoptose (ou mort cellulaire programmée)
Sénescence post-réplicative
Cellule musculaire multinucléée
Cellules polarisées en brossede l’épithelium du tube digestif
Fragmentation de le membrane plasmique Fragmentation
de l’ADN nucléaire
Dégradation lente des fonctions cellulaires suivie de nécrose cellulaire

A
B
G1, S ou G2 M M
G1 S S
+ =
+ =
Figure 1.7 : Dominance cellulaire : Expériences de Rao et Johnson, 1970.
ADN en réplication (incorporation de BrdU)
ADN Métaphasique condenséADN Interphasique: Chromatine

Figure 1.8 : La maturation méiotique chez quelques espèces et les étapes de fertilisation provoquant l’accomplissement de la méiose.
VG
Fertilisation chez :Ascaris (nématode)Spisula (bivalve)Octopus (poulpe)Urechis (ver marin)
Fertilisation chez :Chaetopteres (ver marin tubulaire)Mytilus (moule)Patella (patelle ou Bernique)Artemia (crustacé marin)Drosphila (mouche du vinaigre)Ascidia (tunicier marin ou violet)
Fertilisation chez Presque tous les vertébrés
Fertilisation chez :Arbacia (oursin)Tubularia (hydrozoaire)
Chromosomes Fuseau
1er globule polaire
2ème globule polaire
Noyau haploïde= pronucleus
Vésicule germinale=VG= noyau de l’ovocyte
Rupture de la vésicule germinale= GVBD
Prophase I= Arrêt en G2
= Ovocyte immature
GVBD= Germinal Vesicle Break Down
= Ovocyte mature
Metaphase I Metaphase II Pronucleus= Phase G1
Arrêt méiotique primaire Arrêts méiotiques secondaires

Figure 1.9 : Déclenchement hormonal de la maturation méiotique chez quelques espèces modèles.
GVBD
GVBD
GVBD
17, 20-dihydroxy4-pregnen 3-one(Nagama et Adachi, 1985)
Progestérone(Fortune et al., 1975 ;Godeau et al., 1978)
1-Methyladenine(Kanatani et al., 1969)
Poisson rouge
Rana pipiens; Xénope
Etoile de mer
Protéases (fluide digestif)(Peaucellier, 1977)
Sabellaria alveolata
GVBD
VG
VG
VG
VG
Vésicule germinale=VG= noyau de l’ovocyte
Rupture de la vésicule germinale= GVBD
Phase G2 Phase MI
Déclencheur de la maturation méiotique

GVBD
ProgestéroneVG
G2 M
Figure 1.10 : Emergence du concept MPF (Maturation Promoting Factor) après transfert de cytoplasme d’ovocytes activés par l’hormone de maturation chez diverses espèces.
GVBD
ProgestéroneVG
G2 M
GVBD
1-MethyladenineVG
G2 M
VG
G2
GVBD
M
VG
G2
GVBD
M
VG
G2
GVBD
M
Crapaud (Bufo sp.)(Dettlaf, 1964)
Grenouille (Rana pipiens)(Masui et Markert, 1971;Smith et Ecker, 1971)
Etoile de mer(Kishimoto et Kanatani, 1976)
MPF
MPF
MPF
Hormone induisant la maturation méiotique
Vésicule germinale=VG= noyau de l’ovocyte
Rupture de la vésicule germinale= GVBD
A
B
C

Incubation dans un milieu contenant du cycloheximide = blocage des traductions de protéines.
VG
G2
GVBD
M
Figure 1.11.1 : Découvertes des propriétés du MPF (Maturation Promoting Factor) par Yoshio Masui et Clement Markert chez Rana pipiens (1971).
CVG
G2
Progestérone
VG
G2
Pas de maturation méiotique
VG
G2
VG
G2
VG
G2
Pas de maturation méiotique
A
B GVBD
ProgestéroneVG
G2 M
VG
G2

GVBD
M
Figure 1.11.2 : Découvertes des propriétés du MPF (Maturation Promoting Factor) par Yoshio Masui et Clement Markert chez Rana pipiens (1971).
D
E
GVBD
ProgestéroneVG
G2 M
VG
G2
Traitement du cytoplasme contenant
le MPF par des enzymes (DNase
RNAse) ou dialyse.
GVBD
ProgestéroneVG
G2 M
VG
G2
Traitement du cytoplasme contenant
le MPF par des Protéases ou la
chaleur.
VG
G2
Pas de maturation méiotique

GVBD100%
Progestérone 1-Methyladenine
Masui et Markert, 1971Grenouille (Rana pipiens)
VG
G2 M
VG
G2 M
GVBD69%
VG
G2 M
GVBD91%
VG
G2 M
GVBD75%
GVBD100%
Kishimoto et Kanatani, 1976Etoile de mer
VG
G2 M
VG
G2 M
GVBD100%
VG
G2 M
GVBD88%
VG
G2 M
GVBD93%
Figure 1.12 : Résumé des expériences de « transferts de cytoplasmes d’ovocytes activés » en série dans deux modèles d’étude (le pourcentage d’ovocytes receveurs qui atteignent le stade GVBD est indiqué).
A B

Figure 1.13 : Principe du test MPF de Masui et Markert Rana pipiens (1971).
G2 MI MII fertilisation 1ère Mitose 2ème mitose
VG
MPF ?
VG
G2 M
[MPF]= concentration
de MPF2,5
2,0
1,5
1,0
0,5
Divisions embryonnaires
Maturation de l’ovocyte
Progestérone Fertilisation
10 20 30 40 50 60 minutes
Embryon 2 cellules

Figure 1.14.1 : Principe des tests MPF et CSF de Masui et Markert chez Rana pipiens (1971).
GVBD
M
GVBD
ProgestéroneVG
G2 M
VG
G2
TestMPF
TestCSF
fertilisation
fertilisation
MPF
CSF
contrôle
MPF
?

Figure 1.14.2 : Activité MPF et CSF mesurables au cours de la maturation méiotique par transferts de cytoplasmes dans des ovocytes ou des blastomères d’embryons précoces de. grenouille (Rana pipiens ) par Masui et Markert (1971).
Act
ivit
é M
PF
Act
ivit
é C
SF
---
---
Arrêt G2 Meïose I Meïose II 1er cycle 2ème cycle embryonaire embryonaire
G2 MI MII fertilisation
VG
Temps

Figure 1.15 : Chronogramme de l’identification du MPF.
Année Modèle méiotique Modèle somatique Modèle embryonnaire
Découverte de la transformation automatique d’un ADN exogène en ADN d’ovocyte (Dominance) après microinjection (Oursin, Amphibien).
Synchronisation du cycle cellulaire chez Tetrahylmena.
Découverte de l’hormone de maturation des amphibiens.
Découverte de l’hormone de maturation de l’Etoile de mer.
1922-1929
1964
1967
1969
1970 Dominance des phases S et M du cycle cellulaire (Cellules HeLa).
1964-1971 Découverte du MPF (Amphibiens).
1971 Découverte du CSF (Amphibiens).
Premiers mutants cdc de S. cerevisiae (Levure)1974
Premiers mutants cdc de S. pombe (levure)1975
Gene Cdc2 requis pour l’entrée en phase M de S. pombe (Levure)1976 Découverte du MPF d’Etoile de mer.
Elévation des phosphorylations de protéines au cours de l’entrée en méiose (Etoile de mer).
1976
1978-1982
1980 Nécessité d’ATP pour l’activité MPF.
Universalité du MPF en méiose.
1979-1982
1982-1986 Identification et clonage de la 1ère Cycline (Oursin).
Découverte du MPF dans cellules HeLa et chez S. cerevisiae
Les gènes cdc2 (S. pombe) et CDC28 (S. cerevisiae) sont équivalents.
1982
Oscillations parralèles du MPF et des phosporylations de protéines (Etoile de mer).
1983
Nécessité de synthèses de protéines au MPF pour pouvoir se régénérer (Xenope).1984
Découverte du MPF en mitose (Xenope).
1985-1986Les gènes CDC28 et cdc2 codent pour une protéine kinase (Levures).
Clonage du gène cdc2 humain.1987
Purification du MPF de Xenope
Purification du MPF (Xenope) .Purification de l’Histone H1 kinase (Etoile de mer).
1988
Identification de cdc2 dans le complexe MPF-H1 kinase de Xenope et d’Etoile de mer (western blot).
Le MPF = complexe stoechiométrique entre une sous-unité cycline et une sous-unité P34cdc2 (Etoile de mer).
1989
1990-1991 Homologues de cdc2 identifiés dans divers modèles et adoption d’une nouvelle nomenclature des protéines da la famille de cdc2= CDK1.
2001 Paul Nurse, Leland Hartwell et Tim Hunt partagent ensemble le prix Nobel de Médecine pour leurs découvertes respectives des régulateurs de la division cellulaire.

Figure 1.16.1 : Expérience d’identification de la première cycline dans l’embryon d’Oursin (Arbacia punctulata), Tim Hunt 1983.
Fertilisation des ovocytes dans l’eau de mer en présence de [35S]methionine pour marquer les néo-synthèses de protéines.
Collecte d’échantillons toutes les 10 minutes et traitement dans l’acide trichloroacetique pour fixation puis migration sur gel SDS-PAGE, séchage et autoradiogramme.
Cycline
Protéines radioactivesSDS-PAGE
Densitométrie de la cycline
Le second pic de synthèse de Cycline est moins important à cause de la dilution de la [35S]methionine rajoutée au début de l’expérience.
3
2
1
Minutes 10 30 50 70 90 110 130

Figure 1.16.2 : Expérience d’identification de la première cycline dans l’embryon d’Oursin (Arbacia punctulata), Tim Hunt 1983.
Fertilisation des ovocytes dans l’eau de mer en présence de [35S]methionine pour marquer les néo-synthèses de protéines.
Collecte d’échantillons toutes les 10 minutes et traitement comme pour la Figure 1.17.1.
Quantité de Cycline (- -) % d’embryons en division ( )
2
1
0
0 1 2 Heures après fertilisation
50
25
0

Figure 1.17 : Mutants cdc de Levure (Schizosaccharomyces pombe) isolés par Paul Nurse à partir de 1976 : Réseau de régulation en amont de cdc2.
CDC2 kinase
CDC2 kinase
CDC2 kinase
CDC2 kinase
Légende : Effet inhibiteur Délétion du gène
Effet activateur Surexpression du gène
Phénotype de division normal
Phénotype de division Wee
Phénotype de division cdc25
Phénotype de division de typecatastrophe mitotique= Entrée en phase M avant la fin de la phase S = Totale désynchronisation des phases S et M.
wee1 cdc25

Figure 1.18 : Clonage de l’homologue humain de cdc2 par complémentation focntionnelle, Lee et Nurse, 1987.
Banque d’ADNc humains(Labo. Hirota Okayama)
Culture de mutant cdc2ts de S. pombeÀ la température permissive= divisions
Transfection des levures cdc2ts avec l’ADN de la banque
Observation des colonies de Levures cdc2ts à T° restrictive
Repiquage des colonies de levures qui arrivent à pousser
Observation au microscoped’une colonie de Levure à T° restrictive.
Levure de grande taille ayant perdu le plasmide et incapable de se diviser = phénotype cdc2ts.
Levures de taille normale sans phénotype cdc2ts= phénotype normal.
Isolement et séquençage du plasmide de la banque d’ADNc Humain présent dans le clone de Levure.
N- -C
Cdc2 LevureN- -C
Cdc2 Humain
Identification du produit codé par l’ADNc responsable de la complémentation fonctionnelle de la mutation cdc2ts (perte de fonction) par comparaison de séquence aminoacides.
63% d’identité