
Les Enzymesd1n7iqsz6ob2ad.cloudfront.net/document/pdf/5385f11ebfa1a.pdf · protéique et certaines...
29
Les Enzymes I. Généralités A. Définition 1. Les enzymes sont des protéines douées d’activité catalytique spécifique a. Naturellement produites b. Action hors des cellules qui les produisent c. Thermolabiles 2. On désigne sous le nom de substrat ce qui va être transformé et produit ce qui découle de la réaction a. b. A une réaction correspond une seule enzyme c. En fait il existe des iso-enzymes élaborées par le même organisme qui catalysent la même réaction mais dont la structure protéique et certaines propriétés physico- chimiques sont légèrement différentes. B. Notion de catalyse 1. Réaction d’estérification a. hydrolyse b. 2. A l’équilibre a.
Transcript of Les Enzymesd1n7iqsz6ob2ad.cloudfront.net/document/pdf/5385f11ebfa1a.pdf · protéique et certaines...

Les Enzymes
I. Généralités
A. Définition
1. Les enzymes sont des protéines douées d’activité catalytique spécifique
a. Naturellement produites
b. Action hors des cellules qui les produisent
c. Thermolabiles
2. On désigne sous le nom de substrat ce qui va être transformé et produit ce qui découle de la réaction
a.
b. A une réaction correspond une seule enzyme
c. En fait il existe des iso-enzymes élaborées par le même organisme qui catalysent la même réaction mais dont la structure protéique et certaines propriétés physico-chimiques sont légèrement différentes.
B. Notion de catalyse
1. Réaction d’estérification
a.hydrolyse
b.
2. A l’équilibre
a.

b.
c.constante caractéristique de l’équilibre à une température donnée
3. Modalités de l’action catalytique
C. Moments historiques de l’enzymologie
D. Répartition des enzymes chez les êtres vivants
1. Dans les organes et les tissus
a. D’une façon générale, la répartition des enzymes est analogue à celle des substrats qu’elles peuvent transformer
b. Localisation dans les mêmes organes chez les espèces voisines
c. Différences immunologiques et de structure à marqueurs tissulaires
2. Dans la cellule
a. A l’intérieur de la cellule les enzymes sont regroupées e unités fonctionnelles
b. Ex : mitochondries, lysozomes
II. Constitution des enzymes
A. Rappels
1. Dénaturation
a. Passage d’une structure ordonnée à une structure désordonnée : précipitation
b. Rupture des liaisons non - covalentes et des ponts S-S
1. Pont hydrogène

2. Liaisons hydrophobes
3. Liaisons électrostatiques
c. La dénaturation est réversible au départ, ilexiste un état intermédiaire :
2. Explications principales
a. pH extrêmes
b. Solvants organiques et détergents
c. Action du b-mercapto éthanol
B. Structure générale des enzymes
1. Protéine monomérique
a. Réaction directement sur la protéine
b. Site actif = quelques acides aminés
c. Arrangement spatial fondamental
2. Structure quaternaire : oligomères
a. Autant de sites actifs que de monomères
3. Autres éléments
a. Cations = cofacteurs métalliques
b. Substances organiques : coenzymes
c. Nécessite cofacteurs + coenzyme en général
C. Propriétés générales des coenzymes
1. Caractères communs
a. Jamais protéique

b. Molécule organique de faible masse moléculaire
c. Toujours thermostable
d. Non responsable de la spécificité
e. Apparaissent dans les termes de la réaction
f. Retrouvent leur état général in vivo
g. Systèmes conjugués (alternances des types de liaisons)
2. Les deux types de coenzymes
a. Dissociables : coenzymes = transporteur
b. Non dissociables : très solidement lié à l’apoprotéine
D. Le site actif
1. Définition
a. Le site actif est une zone constituée de quelques acides aminés localisés dans la zone interne hydrophobe de la protéine et au niveau de laquelle s’exerce la catalyse
b. Le site actif est constitué des acides aminés qui
1. Lient les substrats à site de fixation / reconnaissance
2. Assurent la catalyse à site catalytique
c. Généralités
1. Grande diversité
2. Caractéristiques : petits, crevasses, hydrophobe, a.a. polaires

3. Arguments : substrats petits, enzymes autocatalytiques
2. Nature des groupes catalytiques du site actif
a. Seuls certains acides aminés peuvent intervenir dans la catalyse à acides aminéspolaires (His, Ser, Cys, Tyr, Lys)
3. Théories
a. Ajustement induit
b. Distorsion dans le substrat
4. Exemples de mode d’action du site actif
a. 4 genres de phénomènes
1. Positionnement de la molécule du substrat
2. Catalyse covalente : formation d’un acylenzyme (S-E)
3. Catalyse générale acide - base (mvt de H+)
4. Catalyse nucléophile – électrophile : interactions entre charges positives partielles et charges négatives partielles
b. Trypsine
1. Ser 183, His 43, (Asp 182)
2. S + E à AcylE + HX
3. AcylE + H2O + HY
c. Chymotrypsine
1. Hydrolyse à droite de Phe, Trp, Tyr
2. Asp 102, His 52, Ser 195, Gly 193
3. Réseau de relais de charges

d. Métaloenzymes : mécanisme de catalyse covalente
e. Notion de centre catalytique
1. Le centre catalytique d’une enzyme est l’ensemble des groupements fonctionnels de l’enzyme qui participent à l’action catalytique
III. Spécificité des enzymes
A. Spécificité de réaction
1. Spécificité
2.La spécificité est uniquement déterminée par l’apoenzyme
3. Notion d’isoenzyme
a. Enzymes catalysant la même réaction maisqui ont des structures légèrement différentes
b. Combinaison de 2 monomères différents
B. Spécificité de substrat
1. Spécificité étroite
a. Substrat

1. Isoméries Z & E avec double liaison
2. Isoméries optiques C*
b. Produit
1. Apparition de C*
2. Spécificité large
a. Spécificité de groupe
1. 1 enzyme « 1 groupe de substances transformées
2. Ex : b-galactosidase
b. Spécificité basse
1. Action de nombreuses substances
2. L’enzyme ne reconnaît que la liaison : on parle souvent de spécificité de liaison
3. Ex : broméline
IV. Interprétation de la catalyse enzymatique
A. Notion de complexe activé
1. Une réaction se réalise
a. Avec une vitesse appréciable entre deux espèces moléculaires A et B, pour un nombre de chocs efficaces par unité de temps suffisant
b. Dans des conditions optimales
1. Orientation des molécules
2. Minimum d’énergie
2. Complexe activé

a. Si lors d’une collision, les molécules possèdent G* (barrière énergétique), elles forment une association temporaire = complexe activé et peuvent donner A + B
b. La vitesse dépend donc du nombre de complexes activés
3. Evolution de l’enthalpie libre au cours d’une réaction
a. Intervention de la potentialité et de la vitesse
1. Spontanéité = enthalpie libre DG = G2 – G1
2. Energie d’activation : Ea = G* - G1
B. Rôle des catalyseurs et des enzymes
a. G* diminue et Ea diminue pour une réaction donnée
b. Ea diminue avec un catalyseur chimique
c. Ea diminue fortement avec un catalyseur enzymatique
2 Pourquoi Ea diminue avec les enzymes
a. L’enzyme offre à la réaction une voie différente de la voie chimique
b. Avec des enzymes, on a plusieurs étapes successives de Ea voisines à apparition de plusieurs intermédiaires réactionnels. Ea globalest donc la différence entre S et le plus haut niveau
2 Cinétique enzymatique= étude des vitesses de réaction enzymatique
1 Vitesse initiale
1. Généralités

a. Pendant la première période : on a un couple qui est une droite : tangente et courbe se confondent, la pente est appelée la vitesse initiale
b. Lorsque la vitesse est constante : ordre deréaction 0
c. L’augmentation de l’ordre est due à la disparition du substrat et l’apparition des produits
d. aA + bB à P
1. a et b sont les coeff. de stoechiométrie
2. a + b : molécularité = ordre global de la réaction
3. a : ordre partiel pour A
2. Les 2 ordres des réactions
a. Ordre classique
1. nA à nA’
2. avec n : ordre de réaction
3.
b. Ordre initial
1. nA à n’A’
2.
3.
3. Ordres classiques principaux
a. Ordre 0 : [P] = kt
b. Odre 1

1. A à A’
2. ;
3.
c. Ordre 2
1. A + B à C
2.
d. Cas général
1.
e. Ordre 1 réversible
1.
2.
3.
2 Variations de vi
1. En fonction de la [enzyme]
a. Les réactions enzymatiques sont du 1er ordre par rapport à la [enzyme]
b.
2. En fonction de la [substrat]
1. Pour doser des enzymes, on préfère utiliser la vitesse maximum qui ne dépend uniquement que de la concentration en enzyme
2 Etude cinétique de la réaction enzymatique

1. Généralités
a. Pour expliquer la spécificité, on est obligé d’admettre que les enzymes forment un complexe transitoire avec le substrat, où ily aura complémentarité entre le substrat et le site actif
b. à complexe enzyme – substrat spécifique transitoire
2. Equations et constantes de Michaelis
a. Simplification de l’hypothèse générale
b. Constante de Michaelis
1.
2. Hypothèse de l’état stationnaire :
3. Loi de conservation de l’enzyme :
4. Constante de Mickaelis :
c. Equation de Mickaelis
1.
2. Méthode de King et Altman
3. Equation des vitesses
intégrées :
3. Représentation de v0=f([S]0)
a.
b. Valeur particulière : [S]0 = Km

4. Détermination de Km et vM
a. vi = f([S])
b. Représentation logarithmique : vi = f(Log [S0])
c. Représentation en coordonnées réciproques
1. Méthode de Limewover et
Burck :
2. Méthode de Eodie-Holster :
3. Méthode de Dixon / Woolf :
d. Méthode de Eisenthal – Cornish
Bowden :
e. Equation des vitesses intégrées :
5. Interprétation de Km et de vM
a. Km :L’expression de Km est dans certains cas beaucoup plus compliquée que celle vue précédemment
b. vM
1. Quand nous sommes très près de vM, l’enzyme est saturée, tous les sites actifs sont occupés
2. Activité (z)
i. L’activité est proche de vM, elle représente une quantité d’enzymes qui donne cette vitesse

ii. Exprimée en quantité de substrat qui disparaît par unité de temps
iii. En unités d’activité catalytique : qS / min
3. Concentration d’activité catalytique
i. = concentration catalytique = catc= activité volumique
ii. Exprimée en U / L
4. Activité catalytique spécifique : ACS
i. C’est l’activité de l’échantillon diviséepar la masse de protéine qui permet cette activité
ii. Exprimée en U / mg ; cat / mg
iii. Elle représente un critère de pureté
5. Activité catalytique molaire : ACM
i. C’est l’activité catalytique de solutiondivisée par la quantité d’enzymes
ii. = nombre de moles de substrat transformées divisées par unité de temps
6. Constante catalytique :
7. Activité catalytique par centre actif : = ACM / n
c. Mise en évidence du complexe enzyme – substrat
1. Théorie de l’interaction protéine – ligand
i. La dialyse à l’équilibre permet de déterminer Ka et le nombre n de sitespour le ligand

ii. Une protéine P avec n sites récepteurs :
iii. Equation de
Scatchard :
2 Influence de la température
1. Influence de l’augmentation de la température sur la cinétique
a. Utilisation des basses températures
1. On peut refroidir brusquement un mélangeréactionnel pour stopper une réaction enzymatique
2. Par contre, il faut éviter un temps trop longet préférer un petit volume
b. Relation empirique du Q10
1. Augmentation de 10°C, v x 2 à Q10 = 2
2.
c. Relation réelle : loi d’Arrhénius (théorie des collisions)
1.
2.
3. Calcul de Ea de la réaction
globale :
2. Influence de la température sur la dénaturation
a.
b. La dénaturation ne commence qu’après q1

c. La valeur de k à une température donnée va varier suivant les conditions du milieu (pH, force ionique, …)
d. La vitesse d’inactivation va s’amplifier avec le degré de purification de l’enzyme et varie avec l’origine et la nature de l’enzyme
3 Influence du pH
1. Explications
a. Action du pH sur le substrat : il peut exister sous différentes formes +/- attaquables par les enzymes
b. Action du pH sur les enzymes :
1. Sur la stabilité conformationnelle (dénaturation)
2. Sur le site de fixation du substrat
3. Sur la catalyse
2. Action du pH vM et Km
a. Action du pH sur vM :
b. Action du pH sur vM et Km :
c. Action du pH sur Km : peu de variation en fonction du pH
d. Le pH0 varie en fonction de l’origine de l’enzyme, du substrat transformé
e. L’action du pH varie en fonction de la présence de substances associées et de la durée de la réaction
f. On détermine l’activité au pH0 (tamponné)
4 Les effecteurs Mickaeliens de l’activité enzymatique

1. Généralités
a. Action primaire : action directe de l’effecteur sur l’enzyme
b. Action secondaire = indirecte, l’effecteur modifie l’activité en réagissant avec un activateur ou un inhibiteur présent dans lemilieu
c. Effet réversible : l’effet disparaît dès que l’effecteur est éliminé : diminution de la vitesse d’inhibition
d. Effet irréversible : diminution de la vitessed’activation
2. Action des inhibiteurs
a. Combinaison entre enzyme et inhibiteur non indépendante = association exclusive
1. Inhibition compétitive : l’enzyme fixe le substrat ou l’inhibiteur, jamais les deux à la fois à complexes binaires ES / EI
2. Généralités : ET = E + ES + EI
3. Mécanisme
4. Cinétique
i. Pas de I :
ii. Avec IC :
iii. Détermination de Ki
iv. Représentation de
Dixon :

v. Principaux cas d’inhibition compétitive : excès de produit / substrat
b. Combinaisons entre l’enzyme et l’inhibiteur indépendantes : associations non-exclusives
1. Inhibition non-compétitive : récepteurs pour I et S différentes à complexes ternaires ESI
i. Modifications cinétiques
Pas de variation de Km
Représentation de
Dixon :
ii. Cas de l’inhibition mixte (à éviter !)
iii. Mécanisme
2. Inhibition incompétitive : récepteurs pour I et S différents à I se fixe après S, complexes ternaires ESI
1. Modifications cinétiques
Diminution de vM et Km
Mécanisme
2. Action des activateurs Mickaeliens
a. Augmentation des vitesses de réaction

b. Activateurs non modifiés
c. Ions métalliques
d. Associations réversibles
1 Association au hasard
2 Mécanisme séquencé
2 Les effecteurs allostériques(action sur les enzymes allostériques non mickaeliennes)
1. Caractéristiques cinétiques et structurales des enzymes allostériques
a. Généralités
1 Km apparent : vM / 2
2 Coopérativité : dès qu’il y a fixation d’un S,il y a modification de la structure du monomère qui a fixé S, cette modification se transmet aux sous-unités voisines qui vont fixer S plus facilement.
b. Définition des enzymes allostériques
1 Oligomères de type quaternaire, axe de symétrie
2 Effet coopératif entre les molécules de S : affinité croissante
i. Enzymes de type K : effet coopératif homotrope (même catégorie de S), sigmoïde
ii. Enzymes de type v : pas de coopération, effet hétérotrope (entre sites différents), hyperbolique
2 Les deux états conformationnels
i. [S] faible, état défavorable T

ii. [S] élevé, état favorable R
iii. Le passage de T à R se fait par transition allostérique
3 Les deux théories
i. Théorie de Koshland
ii. Théorie de Monod Wyman Changeux MWC
4 Les ligands vont stabiliser l’enzyme dans l’état T ou R suivant la nature du ligand
b. Les deux catégories d’enzymes allostériques
1 Type K
i. v = f([S]) sigmoïde
ii. Effets coopératifs
iii. Km varie ; vM ne varie pas
iv. Représentation de
Hill :
Coeff. directeur : , indice de coopérativité
y = 0 à x = Km apparent
2 Type V
i. VM varie, Km ne varie pas
ii. Disparition de la sigmoïdicité
iii. Coopérativités homotropes
2. Action des effecteurs allostériques
a. Exemples caractéristiques

1 Biosynthèse de la L-Ile à partir de la Thr
2 Synthèse de nucléotides pyrimidiques par l’aspartate transcarbamylase
b. Les inhibiteurs allostériques
1 Les activateurs allostériques
3. Action des agents de désensibilisation
a. La sensibilité à un effecteur allostérique peut disparaître sous l’effet d’un traitement chimique (Hg / urée / F.ion.) ou physique (congél. / décongèl.)
b. La désensibilisation entraîne une normalisation de la cinétique à courbe hyperbolique
c. Multiplication de la coopérativité, suppression des interactions
4. Importance des enzymes allostériques dans les chaînes de biosynthèse
a. Chaînes de biosynthèse linéaire
b. Chaînes ramifiées
1 Inhibition concertée
2 Inhibition cumulative
3 Inhibition différentielle
4 Inhibition étagée
2 Les coenzymes
1. Rappels
a. Molécules organiques nécessaires à l’activité enzymatique
1 Activateurs : liés à l’apoprotéine

2 Transporteurs : co-substrats
b. Jamais de nature protéique
c. Toujours régénérés
2. Les vitamines
a. Tous les coenzymes dérivent des vitamines
b. Une vitamine est une substance organiquequi doit être fournie dans l’alimentation entrès petite quantité, d’une façon continue, pour permettre le fonctionnement de l’organisme.
3. Classification
4. Coenzymes des oxydoréductases
a. Les nicotinamides nucléotides
1 Vitamine impliquée dans la structure
i. Amide nicotinique = B3 = PP
ii. Provitamine = acide nicotinique
iii. B3 à NAD+ / NADP+
2 La structure du NAD+
3 Les propriétés
i. Absorbance max : l = 260, 340 nm
ii. Fluorescence (émission à 460 nm)
iii. Travail en milieu tamponné pour la stabilité
iv. Réactions d’oxydoréduction
b. Les flavines nucléotides
1 Vitamine impliquée dans la structure

i. B2 = riboflavine = lactoflavine
ii. = colorant E101 (jaune)
iii. très sensible à la lumière
2 Les coenzymes qui dérivent de la vitamine B2
i. FMN : Flavine MonoNucléotide
ii. FAD : Flavine Adénine Dinucléotide
3 Mécanismes d’action
i. Coenzymes très fortement liés à la protéine = flavoprotéine
ii. Deux formes semi-quinoniques
c. Les coenzymes hématiniques
1 Fortement liés aux protéines
2 Structure tétrapyrolyque centrée sur un atome de Fe2+
3 Fonction : CytFe3+ ó CytFe2+
d. Autres enzymes d’oxydoréduction
1 Acide lipoïque
2 à intervention dans les décarboxylations
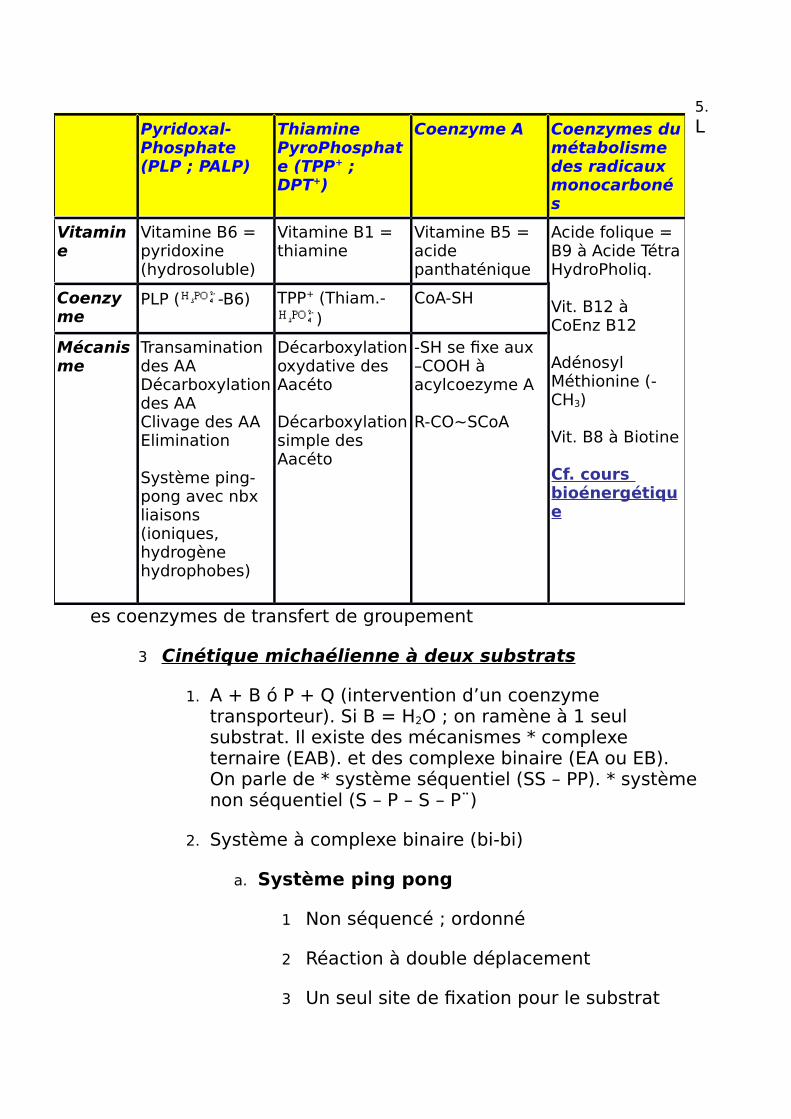
5.L
es coenzymes de transfert de groupement
3 Cinétique michaélienne à deux substrats
1. A + B ó P + Q (intervention d’un coenzyme transporteur). Si B = H2O ; on ramène à 1 seul substrat. Il existe des mécanismes * complexe ternaire (EAB). et des complexe binaire (EA ou EB). On parle de * système séquentiel (SS – PP). * systèmenon séquentiel (S – P – S – P¨)
2. Système à complexe binaire (bi-bi)
a. Système ping pong
1 Non séquencé ; ordonné
2 Réaction à double déplacement
3 Un seul site de fixation pour le substrat
Pyridoxal-Phosphate (PLP ; PALP)
Thiamine PyroPhosphate (TPP+ ; DPT+)
Coenzyme A Coenzymes dumétabolisme des radicaux monocarbonés
Vitamine
Vitamine B6 = pyridoxine (hydrosoluble)
Vitamine B1 = thiamine
Vitamine B5 = acide panthaténique
Acide folique = B9 à Acide TétraHydroPholiq.
Vit. B12 à CoEnz B12
Adénosyl Méthionine (-CH3)
Vit. B8 à Biotine
Cf. cours bioénergétique
Coenzyme
PLP ( -B6) TPP+ (Thiam.-)
CoA-SH
Mécanisme
Transamination des AADécarboxylationdes AAClivage des AAElimination
Système ping-pong avec nbx liaisons (ioniques, hydrogène hydrophobes)
Décarboxylationoxydative des Aacéto
Décarboxylationsimple des Aacéto
-SH se fixe aux –COOH à acylcoezyme A
R-CO~SCoA

b. Cinétique
1
2
3
c. Mécanisme de Theroell et Chance
1 E + A ó EA || EA + B ó E + P + Q
3. Cinétique à complexe ternaire
a. Mécanismes d’association aléatoire substrat – enzyme (bi-bi aléatoire)
1 Cas général : associations de A et de B avec E dépendantes
i.
ii. Représentation de Cleland
iii.
iv. Représentation secondaire :
2 Cas simplifié : association indépendante à KA et KB
i. Représentation de Cleland

ii. Représentation
secondaire :
b. Mécanisme ordonné (bi-bi ordonné)
1 E + A ó EA ó EAB à E +P + Q
2 Représentation de Cleland
i. Equation des vitesses
ii. Détermination des
constantes :
3 Détermination du mécanisme réactionnel
i. Graphiquement :
ii. Inhibiteurs par excès de produit
iii.
Mécanismes
Produits Substrat variable
Ordonné
P
A B
B non - saturant
B saturant A non - A saturant

bi-bi
Theorell / Chance bi-bi
Ping-pong
Q
PQ
PQ
saturant
NCC
NCC
NCC
C
ICC
-C
-C
-
NCNC
CNC
CNC
C
NC-
C-
C-
-
iv. D’après l’action des analogues structuraux
B’ IC de E pour B, A cst, B’ analogue structural de B
B fixe, A variable en présence de B’ à mécanisme aléatoire à complexe ternaire (INC de E pour A, B fixe)un site pour S, un site pour I
Ou à mécanisme ordonné à complexe ternaire (II pour A, B fixe)un site pour A, un site pour B’ =site de B
4 Remarque : réversibilité des réactionsenzymatiques (Réaction de Maldane, cinétiques à deux substrats)

Substrats Produits Enzyme
on appellera A et B les SUBSTRATS d'une réaction ;
on appellera P et Q les PRODUITS d'une réaction ;
on appellera E et E' les différentes formes de l'enzyme pour les systèmes impliquant une isomérisation de celle ci.
Formeenzymatique
toute forme sous laquelle se trouve l'enzyme (libre, complexée au(x) substrat(s) ; complexée au(x) produit(s) ; ...) ;
toute forme qui ne peut pas se dissocier selon une réaction unimoléculaire (ou s'isomériser dans une telle forme) est appelée forme enzymatique stable.
Complexecentral
complexe entre l'enzyme ET TOUS les substrats OU complexe entre l'enzyme ET TOUS les produits ;
exemple : dans la réaction EAB <==> E +P, EAB est un complexe central ;
le nombre minimal de complexes centraux est donc 2 ; un tel complexe ne participe qu'à des réactions unimoléculaire (relarguage d'un substrat ou d'un produit) ;
exception : dans les systèmes à double déplacement ("ping pong"), iln'existe pas de complexe (enzyme tous les substrats) ou de complexe(enzyme tous les produits).
Complexeterminal
(ou abortif)
complexe enzyme ligand (substrat ou produit) qui ne participe pas à la formation de produit(s) de la réaction ;
une partie de l'enzyme est donc inactive et le ligand est considéré comme un inhibiteur.

Séquentiel
système où TOUS les substrats doivent se fixer à l'enzyme AVANT que n'importe lequel des produits ne soit relargué ;
exemple : système au hasard ou système ordonné.
Nonséquentiel
système où un PRODUIT est relargué ENTRE les additions successives des substrats ;
exemple : système à double déplacement ("ping pong").
Theorell -Chance
système proposé par H. Theorell & B. Chance (1951) pour un mécanisme ordonné SANS complexe central (le premier produit est relargué par la fixation du dernier substrat)).
Nomenclature
dessystèmes
Bi Bi : système impliquant 2 substrats et 2 produits ; exemple : Bi Bi au hasard ;
Bi Uni (ou Uni Bi) : système impliquant 2 substrats et 1 produit (ou l'inverse); exemple : Bi Bi ordonné ;
Iso : système impliquant une isomérisation de l'enzyme (changement de conformation E <=> E'); exemple : Iso ping pong ;
Modèleshoméomorp
hes
modèles différents qui fournissent des équations de vitesse formellement identiques.
exemple : le système Bi Bi au hasard analysé avec l'hypothèse du quasi équilibre et le système Bi Bi ordonné analysé avec l'hypothèse de l'état stationnaireconduisent à la même expression de la vitesse de la réaction.
Postulat 1
toute réaction élémentaire ne peut être que mono ou bimoléculaire.
en d'autres termes, deux molécules de substrats ne peuvent pas se fixer simultanément (les complexes ternaires, quaternaires ... ne peuvent être formés que par additions successives de ligand).
Postulat 2
lorsqu'une réaction élémentaire est bimoléculaire, la réaction réverse est
ki . [S] Ei <=======> Ei+1 ki . [P]

monomoléculaire. On ne peut donc pas avoir de réaction du type :
Complexeterminal
(ou abortif)
complexe enzyme ligand (substrat ou produit) qui ne participe pas à la formation de produit(s) de la réaction ;
une partie de l'enzyme est donc inactive et le ligand est considéré comme un inhibiteur.
ABZYMESLe concept d'anticorps catalytique, ou abzyme, fut énoncé dès les années 1940 par Linus
Pauling. La réaction chimique de transformation d'une molécule en une autre passe par
un état de transition intermédiaire, qui représente une barrière énergétique à franchir.
Pauling suggéra qu'une enzyme, pour favoriser une réaction, abaisse cette barrière
énergétique en stabilisant préférentiellement l'état de transition de la réaction plutôt que la
molécule de substrat. Par opposition, un anticorps stabilise la même molécule dans son état
énergétique de base. Linus Pauling émit l'idée totalement novatrice que l'obtention d'un
anticorps dirigé contre l'état de transition d'une réaction plutôt que contre le substrat
permettrait la préparation d'anticorps porteurs d'une activité catalytique. Le problème est que
ces états de transition ont une durée


















