
Influence de la croissance somatique sur le cycle spermatogénétique de Lithobius forficatus L....
11
GENERAL AND COMPARATIVE ENDOCRINOLOGY 33,412-422 (1977) Influence de la Croissance Somatique sur le Cycle Spermatogbnbtique de Lithobius forficatus L. (Myriapode Chilopode) MICHEL DESCAMPS Wniversitt; des Sciences et Techniques, Laboratoire de Biologie Animafe, L.A. associt; act CNRS n”. 148 (“Endocrinologie compar6e des Invert&b&“), B.P. 36,5%50 Wleneuve D’hscq. France Accepted May 18, 1977 L’influence de la croissance somatique SW le cycle spermatog&tique a Cti BtudiCe chez le Chilopode Lithobiusforficutus L. en provoquant le diclenchement de la mue par injection d’ecdysone ou par ablation d’appendices (locomoteurs ou antennaires). Dans tous les cas, la spermiogenise n’est que retardde par rapport aux tkmoins. L’injection d’ecdysone entraine en outre des multiplications goniales et parfois l’avortement d’une premiire poussCe sper- matog&tique: il en rCsulte une digkrescence de nombreuses cellules germinales. The relationship between somatic growth and spermatogenesis has been studied in Lithobius forjicatus (Myriapoda Chilopoda) by inducing moulting either by injection of ecdysone or by removal of antennae or limbs. Both treatments result in a delay in the appearance of spermatids and spermatozoa in the testes. Morover, injection of ecdysone induces an increase in the size of the got&J population and sometimes a dramatic increase in the number of degenerating cells. Dans de prkidents travaux nous avons Ctudik Ie cycle spermatogkrktique de Lithobius forficatus (Joly et Descamps, 1969) et montrk I’influence de diffkents fac- teurs externes (Descamps, 1971) ou endo- crines (Descamps, 1974, 1975). Chez ce Chilopode, le cycle sper- matog&ktique ne pksente pas de varia- tions importantes en relation avec ie cycle de mue. Toutefois, afin de mettre en Cvi- dence une iventuelle competition entre la croissance et la spermatogenkse, nous avons provoquk une mue expkimentale: (1) par injection d’ecdysone. Nous avons eu recours h cette technique car l’absence de donnkes prkises sur la localisation de la glande de mue interdit toute expkrience d’implantation. (2) par section d’appendices, loco- moteurs ou antennaires. 1 Nous remericions les Iaboratoires Hoffmann- Laroche (BFde) qui nous ont fourni gracieusement les ichantillons d’ecdysone. MATERIEL ET TECHNIQUES Nos expdriences ont ttt r&listes sur des Lithobius forfcatus maturus senior (Verhoeff, 1905), de 3 g 4 jours apri?s leur rtcolte. L’ecdysone’ (milange d’cx et de B ecdysone) est dissoute dans le liquide de Ringer adapt& aux Ar- thropodes (Ephrussi et Beadle, 1936) alcoolisi Q 5%; elle est utilisbe B une concentration de 0,90 pg (y) pour 10 ~1 de solution. Les doses inject&es (0,45 y d’ecdysone ou 0,90 $ sont susceptibtes de provoquer l’exuviation (Joly, 1964): les animaux pesant de 100 & 120 mg, chaque individu reFoit done respectivement 4-4,5, ou 8-9 pg d’ecdysone par gramme de poids frais. Ces doses sont comparables B celles qui sont utilisees chez les Insectes et les Crustaces (Krishnakumaran et Schneiderman, 1970). Les injections sont uniques (0,45 et 0,90 y) ou ri- p&es (0,90 7). Dans ce demier cas, elles sont &par&es par des laps de temps de 30 jours environ, d&i assez long, mais qui s’explique par le fait que chez L. forficatus, l’augmentation du pourcentage de mues n’est significative que 25 jours environ apr&s l’intervention exptrimentale (Joly, 1964, 1966): des in- jections trap rapprochCes entraZnent d’ailleurs une augmentation importante de la mortal&& Les injec- tions uniques ont iti pratiquees durant la croissance spermatocytaire ou les mitoses de maturation. 412 Copyright @ 1977 by Academic Press. Inc. All rights of reprtiduction in any form reserved ISSN OQI644RO
-
Upload
michel-descamps -
Category
Documents
-
view
213 -
download
0
Transcript of Influence de la croissance somatique sur le cycle spermatogénétique de Lithobius forficatus L....

GENERAL AND COMPARATIVE ENDOCRINOLOGY 33,412-422 (1977)
Influence de la Croissance Somatique sur le Cycle Spermatogbnbtique de Lithobius forficatus L. (Myriapode Chilopode)
MICHEL DESCAMPS
Wniversitt; des Sciences et Techniques, Laboratoire de Biologie Animafe, L.A. associt; act CNRS n”. 148 (“Endocrinologie compar6e des Invert&b&“),
B.P. 36,5%50 Wleneuve D’hscq. France
Accepted May 18, 1977
L’influence de la croissance somatique SW le cycle spermatog&tique a Cti BtudiCe chez le Chilopode Lithobiusforficutus L. en provoquant le diclenchement de la mue par injection d’ecdysone ou par ablation d’appendices (locomoteurs ou antennaires). Dans tous les cas, la spermiogenise n’est que retardde par rapport aux tkmoins. L’injection d’ecdysone entraine en outre des multiplications goniales et parfois l’avortement d’une premiire poussCe sper- matog&tique: il en rCsulte une digkrescence de nombreuses cellules germinales.
The relationship between somatic growth and spermatogenesis has been studied in Lithobius forjicatus (Myriapoda Chilopoda) by inducing moulting either by injection of ecdysone or by removal of antennae or limbs. Both treatments result in a delay in the appearance of spermatids and spermatozoa in the testes. Morover, injection of ecdysone induces an increase in the size of the got&J population and sometimes a dramatic increase in the number of degenerating cells.
Dans de prkidents travaux nous avons Ctudik Ie cycle spermatogkrktique de Lithobius forficatus (Joly et Descamps, 1969) et montrk I’influence de diffkents fac- teurs externes (Descamps, 1971) ou endo- crines (Descamps, 1974, 1975).
Chez ce Chilopode, le cycle sper- matog&ktique ne pksente pas de varia- tions importantes en relation avec ie cycle de mue. Toutefois, afin de mettre en Cvi- dence une iventuelle competition entre la croissance et la spermatogenkse, nous avons provoquk une mue expkimentale:
(1) par injection d’ecdysone. Nous avons eu recours h cette technique car l’absence de donnkes prkises sur la localisation de la glande de mue interdit toute expkrience d’implantation.
(2) par section d’appendices, loco- moteurs ou antennaires.
1 Nous remericions les Iaboratoires Hoffmann- Laroche (BFde) qui nous ont fourni gracieusement les ichantillons d’ecdysone.
MATERIEL ET TECHNIQUES
Nos expdriences ont ttt r&listes sur des Lithobius forfcatus maturus senior (Verhoeff, 1905), de 3 g 4 jours apri?s leur rtcolte.
L’ecdysone’ (milange d’cx et de B ecdysone) est dissoute dans le liquide de Ringer adapt& aux Ar- thropodes (Ephrussi et Beadle, 1936) alcoolisi Q 5%; elle est utilisbe B une concentration de 0,90 pg (y) pour 10 ~1 de solution. Les doses inject&es (0,45 y d’ecdysone ou 0,90 $ sont susceptibtes de provoquer l’exuviation (Joly, 1964): les animaux pesant de 100 & 120 mg, chaque individu reFoit done respectivement 4-4,5, ou 8-9 pg d’ecdysone par gramme de poids frais. Ces doses sont comparables B celles qui sont utilisees chez les Insectes et les Crustaces (Krishnakumaran et Schneiderman, 1970).
Les injections sont uniques (0,45 et 0,90 y) ou ri- p&es (0,90 7). Dans ce demier cas, elles sont &par&es par des laps de temps de 30 jours environ, d&i assez long, mais qui s’explique par le fait que chez L. forficatus, l’augmentation du pourcentage de mues n’est significative que 25 jours environ apr&s l’intervention exptrimentale (Joly, 1964, 1966): des in- jections trap rapprochCes entraZnent d’ailleurs une augmentation importante de la mortal&& Les injec- tions uniques ont iti pratiquees durant la croissance spermatocytaire ou les mitoses de maturation.
412 Copyright @ 1977 by Academic Press. Inc. All rights of reprtiduction in any form reserved ISSN OQI644RO

Certains individus re$oivent une injection de IO ~1 de Ringer: ce sont les “operes-temoins.”
L’amputation affecte la moitie des appendices locomoteurs (15 sur 30): dans ces conditions I’animal peut continuer a se dtplacer. La section antennaire est realisie bilatetalement, entre le scape et le second article.
Une strie timoin accompagne chaque serie ex- perimentale. Cette precaution est rendue necessaire par les variations annuelles du cycle sper- matoginetique en fonction des conditions climatiques (Joly et Descamps, 1969: Descamps, 1971: cf. Figs. la, Id, 2a, co~espondant chacune B une ant&e de recolte differente).
Tow les animaux (350 environ) sont places dans les mCmes conditions d’tlevage: temperature du laboratoire (18-20”), rythme nycthimiral normal, et nourriture reguliere. Dans tous les cas le taux de mortaliti reste faible (inferieur a 10%).
Les series de fixation ont lieu tous les 15 jours a &son de 2 ou 3 animaux chez ies temoins et de 3 ri 4 chez les operes; elles pe~ettent d’btabiir ie cycle spe~atog~n~tique et de suivre l’ivolution des popu- lations cellulaires. Les techniques d’etudes his- tologique et quantitative ont ttC exposees anteri- eurement (Descamps, 1974).
cycle spermatoginltique et des populations cellulaires cornparables a ceux des temoins.
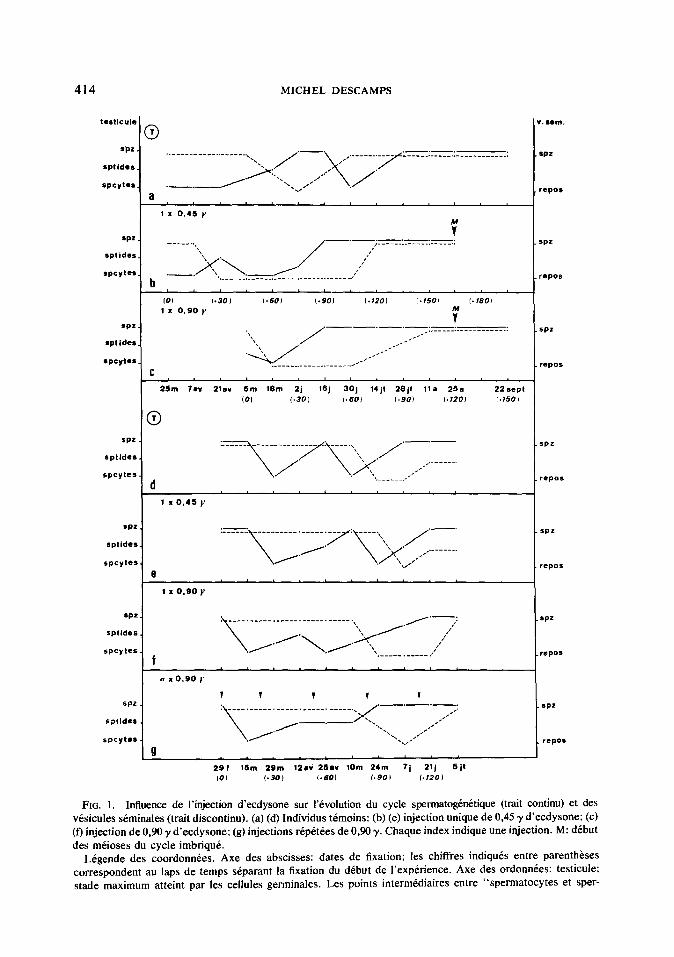
fl) Z~jec~~u~ unique. L’injection d’ec- dysone provoque un retard dans l’appari- tion des spermatozoides (de 1.5 a 60 jours par rapport aux temoins, cf. Tableau 4).
La premiere phase spermatog&rCtique est parfois precedke d’une poussee sper- matogenetique incomplete (Figs. lb et f). I1 peut arriver que les divisions de maturation de la seconde phase spe~atog~n~tique complete se produisent alors que l’evacua- tion des faisceaux de spermatozoides vers les visicules seminales n’est pas achevee; nous parlerons alors de phase sper- matogenetique imbriquke (Figs. lb et c). Ce phknombne est egalement observe dans d’autres cas et n’est pas lie a l’injection d’ecdysone.
RfiSULTATS (A) Injections D’Ecdysone
L’injection d’ecdysone provoque une nette augmentation du pourcentage de mues (temoins, 10%; injections uniques, res- pectivement, 33% (0, 45 y) et 62% (0, 90 y); injections rep&&es, 87%); ces resultats confirment ceux qui ont CtC obtenus par Joly (1964). Nous n’avons retenu que les mues survenues au minimum 25 jours apres l’inte~ention experimentale; en effet, les processus exuviatoires sont d&clenches de fagon irkversible 20 jours environ avant une mue naturelle (Joly, 1966).
Les differences constaties dans les dif- fkrentes series experimentales dependent de deux facteurs principaux: l’age des spermatocytes et la dose d’ecdysone injec- t&e. En effet, 0,45 y suffisent pour induire une phase spermatogenCtique incomplete qui avorte rapidement dans un testicule ren- fermant des spermatocytes en tours de croissance (Fig. lb): cette dose n’entraine qu’un retard dans le declenchement des phenomenes spermatogenktiques dans un testicule ne presentant encore que de tres jeunes spermatocytes (Fig. le). Dans ce dernier cas, 090 y sont necessaires pour obtenir une phase spe~atog~n~tique in- compl&te (Fig. If).
Nous envisagerons successivement l’influence des injections uniques ou rC- pet&es. Dans chacune des skies ex- pkimentales, le testicuk des animaux t& moins renferme des spermatocytes ou des spermatozoi’des, suivant la periode ope- rat&e. L’Cvolution du cycle est voisiue de celle qui est observee lors du cycle nature1 et comporte deux poussees spermato- genetiques (Figs. la et d).
Enfin, une injection d’ecdysone realisee pendant les divisions de maturation les fait avorter (Fig. lc).
L’etude des populations cellulaires met en evidence une multiplication goniale im- portante encore constatke quinze jours apres le debut de l’experience et une grande abondance de cellules dig&rCrescentes d’origine spermatocytaire en relation avec les phases spermatogknetiques avortees. Ces demieres entrainent une diminution importante, quoique passagere, de la popu- . .
. Ianon spermatocytaire. Les “o&G-tCmoins” tksentent un
CYCLE SPERMATOGkNkTIQUE DE LITHOBIUS 413
414 MICHEL DESCAMPS
SP2
rpt ides
spcytcr
6PZ
sptidea
spcytar
SPZ
sptidcr
spcyttr
SPl
sptidc!
spcyte
,p; sptidc!
rpcyter
SPl
sptldes
spcyt*s
. - - _ -. ._ _ _ - _ . _ _ _ _ _ _.. ,\.\ /- ,,,. _ -_--_._--- ..L=: -___-______.:
/’ /:...
.-.-. ‘. .’ . ..I’
,,,.,. ,,. y -----
\
a
1 x 0.45 y
F .--.---I ------.
.-.k\.- ,. / ,.- _ - . _ _ _ - - _ _ _ .- - _ _ _ . .
,/” b I .___ - __.______: _____..______.______. *
lOI 1.30 I I.601 (.SOJ f.120) I.150) f.180) 1x o.soy
i
.t/‘-‘-’
-.-.-. ‘\ ,/’
,.~ _____._____---_--.: ‘\ ‘\ _.-*
/- __--
: ___________ - _._____... _ ”
25m 7s~ 21~ 5m 19m 2j ‘61 301 14jt 28jt lla 25a 22 sept (01 1.30) I.601 1.90) (.I201 1.160)
d 1 * 0.45 y
.-. .-__--.. _--_.._-__ _ ___ __ ___
\
.
/ /---
_.
/’ \
“‘\ /- ,/<: /. I.------.
: ____._____._____.___--.-----.
\
.-.
.A .A’ \
l,” /”
.A \ ,/ I
f .___--. ----- .*
n x 0.90 )’ 1 1 1 f 1
\
.-.-.-. - - _ - . - - - - - - - - _ _ _ _ _ _ _ -- _ _ -. . .,,,.-.- 4.
,/ .’ l’ *.
..\. /’ ,.d’
._, 4
291 1Sm 29m 124v 256r 10m 24m 7j 211 sjt I01 f.30) t.601 1.901 I.120)
FIG. 1. Influence de I’mjection d’ecdysone sur I’bvolution du cycle spermatogknetique (trait continu) et des vesicules semimales (trait discontinu). (a) (d) Individus temoins: (b) (e) injection unique de 0,45 y d’ecdysone; (c) (f) injection de 0,90 y d’ecdysone; (g) injections repktees de 030 y. Chaque index indique une injection. M: debut des meioses du cycle imbrique.
Ltgende des coordonnees. Axe des abscisses: dates de fixation; les chiffres indiques entre parentheses correspondent au laps de temps separant la fixation du debut de I’experience. Axe des ordonnees: testicule: stade maximum atteint par les cellules germinales. Les points intermediaires entre “spermatocytes et sper-
CYCLE SPERMATOCtiNkTiQUE DE LfTHOBiUS 415
testicule 0
i SPZ - .-. .-.-.
------‘5
sptides- ‘\ /
---.--Y”
,,/
,,.I
spcytes. ‘\ .’ ‘,F-F----.- I’
a abl. pattes
SPZ- i
I-. .-. .___“__ ..____...___-..,
sptides- .-.+.--.--. / ,,/ ,A.
spcytes- ‘\ ‘I .______._____ 4’ I’
b abl. antennes
SPZ-
sptides-
spcytes-
‘i’ .-.-. .____ I_ ._____...____..\
‘\ .A .------~-‘-~ /’
‘. *. ,/
5. .,____- . ..- --__.I ,’
1 c 14m 28m 11s 25~ Qm 23m 6j 20j 4 jt
(01 c.301 660) &SO1 I*?ZO/
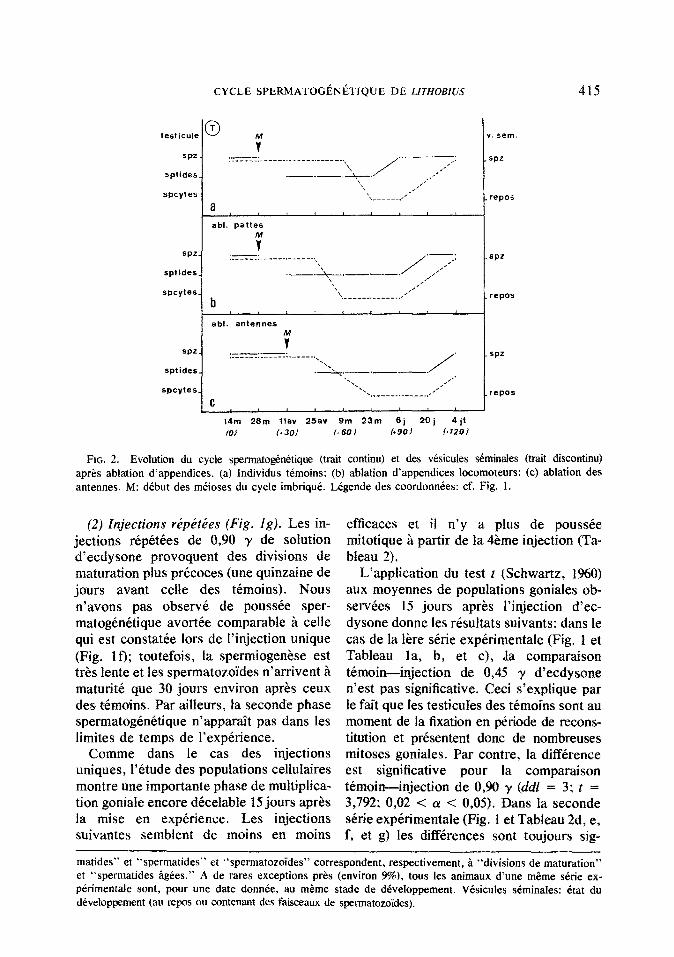
FIG. 2. Evolution du cycle spermatog&ktique (trait continu) et des vbicules dminales (trait discontinu) aprks ablation d’appendices. (a) Individus tkmoins: (b) ablation d’appendices locomoteurs: (c) ablation des antennes. M: dibut des mkioses du cycle imbriquk. LCgende des coordonnkes: cf. Fig. 1.
(2) Injections r&b&es (Fig. ig). Les in- jections &pCtCes de 0,90 y de solution d’ecdysone provoquent des divisions de maturation plus prkoces (une quinzaine de jours avant celle des ttmoins). Nous n’avons pas observk de pousske sper- matogknitique avortke comparable B celle qui est constatie lors de I’injection unique (Fig. If); toutefois, la spermiogenkse est t&s iente et les spermatozoides n’arrivent B maturitk que 30 jours environ aprks ceux des tkmoins. Par ailleurs, fa seconde phase spermatog&&ique n’apparait pas dans les limites de temps de l’expkrience.
Comme dans le cas des injections uniques, l’ktude des populations cellulaires montre une importante phase de multiplica- tion goniale encore dkcelable 15 jours aprks la mise en expkience. Les injections suivantes semblent de moins en moins
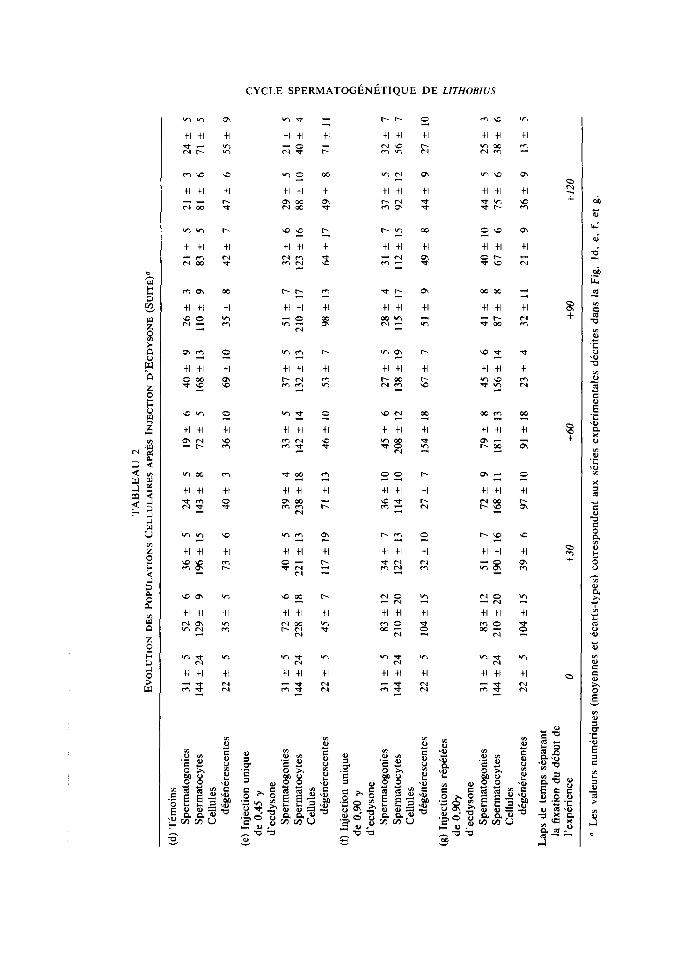
efficaces et il n’y a plus de poussCe mitotique B partir de la 4kme injection (Ta- bleau 2).
L’application du test t (Schwartz, l%O) aux moyennes de populations goniales ob- servees 15 jours aprks l’injection d’ec- dysone donne les rksultats suivants: dans le cas de la l&-e s&e expirimentale (Fig. 1 et Tableau 1 a, b, et c), *la comparaison t~moin-injection de 0,45 y d’ecdysone n’est pas significative. Ceci s’explique par le fait que fees testicules des tkmoins sont au moment de la fixation en pkriode de recons- titution et prksentent done de nombreuses mitoses goniales. Par contre, la diffkrence est signifkative pour la comparaison tkmoin-injection de 0,90 y (&I = 3: t = 3,792: 0,02 < (Y < 0,OS). Dans la seconde sCrie expkimentale (Fig. 1 et Tableau 26, e, f, et g) les difkences sont toujours sig-
matides” et “spermatides” et “spermatozoides” correspondent, respectivement, & “divisions de maturation” et “spermatides Sggtes.” A de rares exceptions p&s (environ %), tous les animaux d’une mame sOtie ex- pirimentale sont, pour une date don&e, au mime stade de dtveloppement. VBsicules shminales: itat du dCveloppement (au repos ou contenant des faisceaux de spermatozoides).

416 MICHEL DESCAMPS
Fir-- m
tl tl fl
KS F;
‘o* 0‘
tl fl fl
CYCLE SPERMATOGtiNkTIQUE DE LITHOBIUS
r-r- 0
$1 +I fl
:z F;
IAN at
+I fl +I
!G;8 $
r-m 00
$1 tl tl
$;!I 9
vt- Q\
+I tl $1
g?jz G;
m m r-
$1 fl +I
Ki,m 5;
wt-4 2
tl tl $1
q z
zz t-.
+I fl +I
g:n &
t-m 2
$1 +I +I
G%,M 2
2% 2
fl fl tl
22 23 t-4 -
";: v,
+I tl t-I
";f G
mw b-1
tl fl +I m
s4 -
vlw Q\
tl +I +I
SF :
ow o\
fl +I +I
%G z
OQOO 4
tl +I fl
G6 2
Wb d
+I +I +I
%si N"
cam cc - 3
+I tl fl
FS z
m- M ,o
fl +I +I
F% ST
r-w w
tl $1 +I
z E z
2:: 2
tl +I $1 mo 8 CO-
t-4 -
"-& v,
$1 fl tl
zs M
8

418 MICHEL DESCAMPS
nificatives (temoins, 0,45 y; ddl = 3; t = 3,725: 0,02 < o! < 0,05; temoins, 090 y; ddl = 5; t = 3,739; 0,Ol < fy < 0,02).
(B) Ablation D’Appendices
Les pourcentages de mues observes apres ablation d’appendices locomoteurs (temoins 15%: 0pCres 46%) ou antennaires (temoins 15%: opires 66%) confirment les r&&tats respectifs de Cameron (1926) et Joly (1%6) et de Joly et Lehouelleur (1972). Nous n’avons tenu compte que des exuvia- tions intervenant aprbs le delai minimum de 25 jours apt-k les diverses interventions operatoires.
Au moment de la recolte le testicule des animaux temoins renferme des sper- matozdides. La duke des series ex- perimentales (120 jours) n’a permis l’obser- vation que d’une seule phase sper- matogenetique.
Chez les tcmoins (Fig. 2a) la phase sper- matog~n~tique est imbriquee darts la pre- cedente. L’evolution des populations cel- lulaires est neanmoins comparable h celle qui est habituellement observke. Un tel phenomene est Cgalement decelable dans les skies correspondantes d’individus opbes,
(I) Ablation des appendices locomoteurs (Fig. 26). Comme chez les temoins, les di- visions de maturation commencent 15 jours apt-es la mise en elevage experimen- tal. Cependant les spermatozoi:des mclrs apparaissent avec une quinzaine de jours de retard par rapport aux temoins.
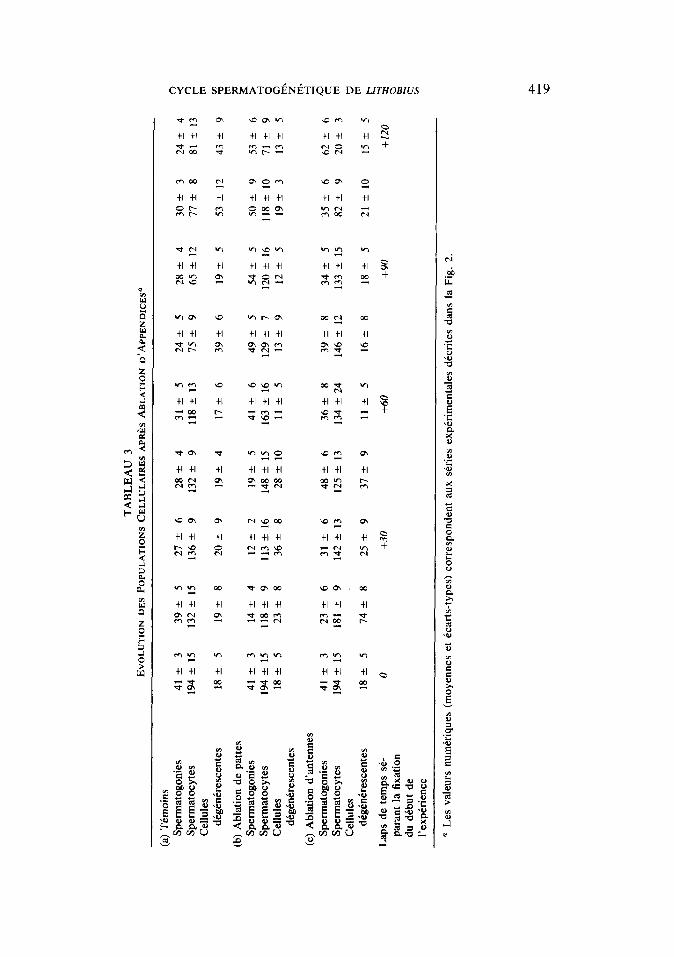
L’~volution des populations cellulaires n’est pas significativement differente de celle qui est observee chez les animaux te- moins {Tableau 3).
(2) Ablation d’antennes (Fig. 2~). Par rapport aux temoins l’ablation des antennes provoque un retard, respectivement de 15 et 30 jours dans le ddclenchement des di- visions de maturation et dans l’apparition des spermatozdides.
Les populations cellulaires suivent une
evolution comparable a celle qui est obser- v&e chez les temoins (Tableau 3).
Les r&mltats relatifs aux dj~~rentes se- ries experimentales (injection d’ecdysone et ablation d’appendices) ont CtC reunis dans le Tableau 4.
DISCUSSION
Chez les Arthropodes, a l’exception des Symphyles (Juberthie-Jupeau, 1%3), il ne semble pas exister de liaison stricte entre croissance et cycle spermatogenetique. Toutefois, chez Lforficatus, le leger retard constate exp~rimen~iement dans le de- clenchement de la spermiogencse semble pouvoir etre interprete comme la const- quence d’une competition entre croissance somatique et spermatogenese; une telle in- teraction a CtC signalee chez les Crustaces (Dtmeusy, 1965; Rouquette, 1970).
L’injection d’ecdysone ou I’ablation d’appendices n’empechent pas le deroule- ment tardif d’un cycle spe~atog~n~tique complet. Un tel retard de la gametogenese est a rapprocher de celui qui est observe chez la femelle (Herbaut, 1977). Notons que lorsque l’injection ou l’amputation ne provoquent pas le declenchement de la mue dans la duke de temps de l’experience, les individus presentent une spermatogenbse comparable a celle des animaux ayant exuvie. II semblerait done que les modifications physiologiques soient impor- tantes et non l’exuviation par elle-m&me. La mime remarque s’applique aussi aux animaux appartenant aux premieres series de fixation chez lesquels la mue n’est pas encore intervenue.
Les deux types d’interventions ex- p~~rnenta~es semblent avoir des consi- quences differentes: l’appendicectomie n’entraine ni multiplication goniale, ni a- vortement de la premiere poussee sper- matogenetique, ni digenerescence cel- lulaire. Ces divers processus semblent done en relation avec l’action de l’ecdysone.
L’induction de mitoses goniales cons-
CYCLE SPERMATOGkNkTIQUE DE YTHOBIUS 419
m
$1
3
2
tl
CI
m
tl Q\
w
+I
2
w
tl t-
w
+I Q\
Q\
+I
w
m
+I Q\
VI
$1 00
wo\‘A
+I +I +I
cl!?=
Q\0m
+I +I $1 OoOQ\ vi--
m w m
fl tl +I *0cI 'Or44
ml--a
tl $I +I
3P-i~
wwvi
tl tl +I
;;g=
lnvl0 e-
$1 $I fl
5?qg
NWCC
+I +I +I NrnW -3m
-TOY00
$1 $I fl
b-9:
mmrl
+I tl +I
Gzx;s
ci .M L 2
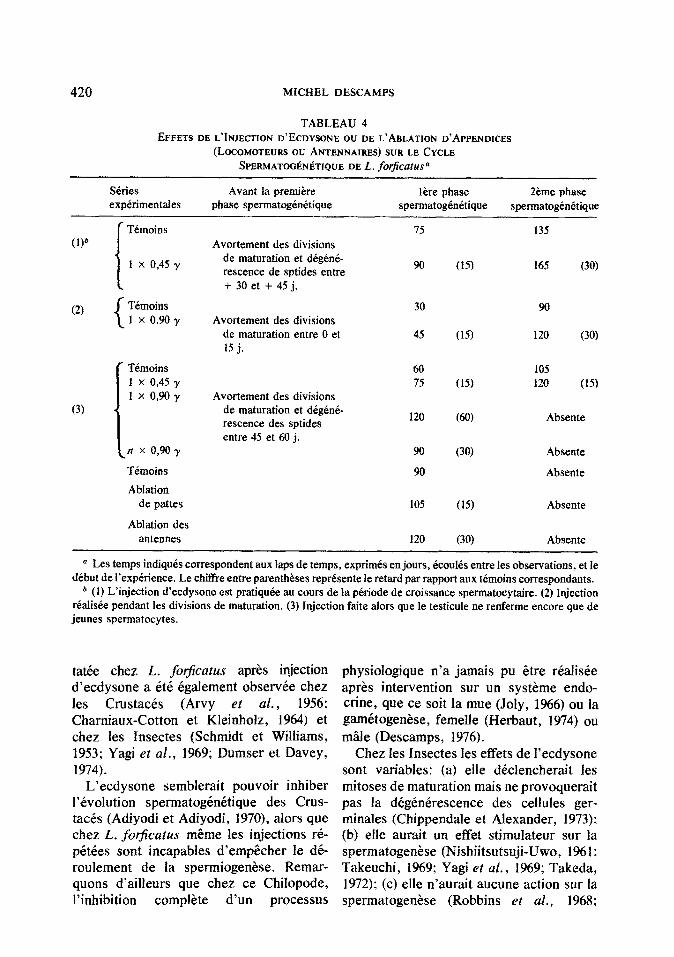
420 MICHEL DESCAMPS
TABLEAU 4 EFFETS DE L~NJECTION D'ECDYSONE ou DE L'ABLATJON D'APPENDICES
(LOCOMOTEURS ou ANTENNAIRES) SUR LE CYCLE SPERMATOG~N~TIQUE DEL. forficatusa
Skies experimentales
Avant la premiere phase spermatogenitique
lhre phase spermatogtnitique
Z&me phase spermatogendtique
Timoins (1)”
1 x 0,45 y
(2) Temoins 1 x 09oy
Avortement des divisions de maturation et degene- rescence de sptides entre + 30 et f 45 j.
Avortement des divisions de maturation entre 0 et 15 j.
30
4.5
(3)
r Temoins 1 x 0,4.5 y 1 x 0,90 y
ifI x 0,9oy
Temoins
Ablation de pattes
Ablation des antennes
Avortement des divisions de maturation et dig&t& rescence des sptides entre 45 et 60 j.
60 7.5
120
90
90
105
165 (30)
90
120 (30)
105 120 (15)
Absente
Absente
Absente
Absente
Absente
a Les temps indiquis correspondent aux laps de temps, exprimds en jours, Bcoults entre fes observations, et Ie debut de I’expirience. Le chiffre entre parentheses reprCsente le retard par rapport aux temoins correspondants.
* (1) L’injection d’ecdysone est pratiqude au tours de la p&ode de croissance spermatocytaire. (2) Injection reahsee pendant les divisions de maturation. (3) Injection faite alors que le testicule ne renferme encore que de jeunes spermatocytes.
tat&e chez L. jyficatus apt& injection d’ecdysone a et& Cgalement observee chez les Crustaces (Arvy et al., 1956; Charniaux-Cotton et Kleinholz, 1964) et chez les Insectes (Schmidt et Williams, 1953: Yagi et al., 1969; Dumser et Davey, 1974).
L’ecdysone semblerait pouvoir inhiber l’evolution spermatogenetique des Crus- taces (Adiyodi et Adiyodi, 1970), alors que chez L. f~~cfft~~ meme les injections r-e- pet&es sont incapables d”empicher le di- roulement de la spermiogenbse. Remar- quons d’ailleurs que chez ce Chilopode, l’inhibition complete d’un processus
physiologique n’a jamais pu etre realisee apres intervention sur un systeme endo- crine, que ce soit la mue (Joly, 1966) ou la gametogenese, femelle (Herbaut, 1974) ou mfle (Descamps, 1976).
Chez les Insectes les effets de l’ecdysone sont variables: (a) elle declencherait les mitoses de maturation mais ne provoquerait pas la diginerescence des cellules ger- minales (Chippendale et Alexander, 1973): (b) elle aurait un effet stimulateur sur la spermatogenese (Nishiitsutsuji-Uwo, 1961: Takeuchi, 1969; Yagi et al., 1969; Takeda, 1972); (c) elle n’aurait aucune action sur la spermatogenese (Robbins et al., 1968;

CYCLE SPERMATOG~~~TIQUE DE L~T~~B~~S 421
Herman et Barker, 1976) ou ne serait pas Gcessaire (Demal, 1961: Lender et Duveau-Hagege, 1963): (d) elle ne jouerait qu’un rale indirect en facilitant l’action d’un facteur macromol~culaire (Kambysel- lis et Williams, 1971). Signalons toutefois que certains des travaux citCs ont tti: rCalisis in vitro: il est possible que d’autres facteurs intetfkrent lors d’Ctudes in viva. L’~nhibition du d~veloppement des produits ggnitaux, ou leur d&eloppement anormal, apt-&s action de l’ecdysone, n’ont, B notre connaissance, Ctt mis en ividence que lors de l’ovogedse [Robbins et al., 1968: Wright et Kaplanis, 1970; Engelmann, 1971; Laverdure, 1976 (a ecdysone)~.
Une remarque s’impose; tous les auteurs sont unanimes pour affirmer que chez les Arthropodes, l’ecdysone est t&s rapide- ment mitabolisCe apr&s injection. Chez L. forecasts, son action n’est observable qu’aprks un iaps de temps relativement long (Joly, 1966; Herbaut, 1974: Descamps, 1976): il semble que son action sur les dif- f&-ents processus physiologiques ne soit qu’indirecte.
Signalons enfin que now ne pouvons jus- qu’h p&sent expliquer l’absence, chez L. fovficatus, apr&s injections rCpCtkes d’ec- dysone, de phase spermiogCnitique avortCe comparable B celle qui a et6 d&rite apr&s injection unique.
BIBLIOGRAPHIE Adiyodi, K. G.. and Adiyodi, R. G. (1970). Endocrine
control of reproduction in decapod Crustacea. Bid. Rev. 45, 121-166.
Arvy, L., Echalier, G., et Gabe, M. (1956). Organe Y et gonade chez Car&ides maenas L. Ann. Sci. Nat. Zool. Riot. Anim. 18, 263-267.
Cameron, J. A. (1926). Regeneration in Scutigera for- ceps. J. Exp. 2001. 11, 169-179.
Chamiaux-Cotton, H., and Kleinholz, L. H. (1964). Hormones in invertebrates other than insects. In “Hormones” (G. Pincus, K. V. Thimann, and E. 33. Astwood, eds.), Vol. IV, pp. 135-198. Academic Press, New York.
Chippendale, G. M., and Alexander, B. R. (1973). Spermatogenesis of the southwestern corn borer, Diatraea grandiosella. II. Resumption in diapause larvae. Ann. Entomol. Sot. Amer. 66, 761-768.
Demal, J. (1961). Problemes concernant la morphogenese in vitro chez les insectes. Bull. Sot. 2001. Fr. 96, 522-533.
Dimeusy, N. (1965). Croissance somatique et fonction de reproduction chew la femelle du Dicapode Brachyoure Carcinus maenas Linni. Arch. 2001. Exp. Gen. 106, 625-664.
Descamps, M. (1971). Le cycle spermatog&&ique chez Lithobius for3catus L. (Myriapode Chilopode). II. Influence de facteurs extemes sur 1’Cvolution du testicule et des vCsicules siminales. Arch. Zool. Exp. Gen. 112, 731-746.
Descamps, M. (1974). Etudedu contritleendocrinien du cycle spermatog&tique chez Lithobius fo$icatus L. (Myriapode Chilopode). Rale de lapars inter- cerebra/is. Gen. Camp. Endocrinol. 24, 191-202.
Descamps, M. (1975). Etude du contrtile endocrinien du cycle spermatog&&ique chez Lithobius forficatus L. ~My~~ Chose). R& due complexe “cel- Iules neurodc&ices des lobes frontaux du protocerebron-glandes c&&braIes.” Gen. Comp. Endocrinol. 25, 346-357.
Descamps, M. (1976). Contribution B l’itude du cycle spermatogtn&ique et de son dbterminisme chez Lithobius foficatus L. (Myriapode Chilopode). These Doet. Etat. Univ. Sci. et Techn. Lille.
Dumser, J. B., and Davey, K. G. (1974). Endo- crinological and other factors influencing testis development in Rhodnius prolixus. Canad. .I. 2001. 52, 101 I-1022.
Engelmann, F. (1971). Endocrine control of insect re- production, a possible basis for insect controi. Acta Phytopathol. Acad, Sci. Hangar. 6, 211- 217.
Ephrussi, B., and Beadle, G. W. (1936). A technic of transplantation for Drosophila. Amer. Natur. 70, 2 18-225.
Herbaut, C. (1974). Contribution Q l’itude du cycle ovoginitique et de son d~te~jnisme chez Lithobius forficatus L. (Myriapode Chilopode). Thise Doct. Etat. Univ. Sci. et Techn. Lille.
Herbaut, C. (1977). Influence de la croissance somatique sur l’ovogen&se chez Lithobius
forfcatus L. (Myriapode Chilopode). Arch. Zool. Exp. Gen. 118, 63-72.
Herman, W. S., and Barker, J. F. (1976). Ecdysteron ant~onism, mimicry, and synergism of juvenile hormone action on the monarch butterfly repro- ductive tract. J. Insect Physiol. 22, 643448.
Joly, R. (1964). Action de I’ecdysone sur le cycle de mue de Lithobius forficatus L. (Myriapode Chilopode). C.R. SOC. Biol. 158, 548-550.
JOEY, R. (1966). contribution 5 l’&ude du cycle de mue et de son ddterminisme chez les Myriapodes Chilopodes. Bull. Biol. France Belg. Ser. C 3, 379-480.
Joly, R., et Descamps, M. (1969). Evolution du tes- ticule, des vCsicules siminales et cycle sper-

422 MICHEL DESCAMPS
matog&&ique chez Lithobius fo$catus L. (My~apode Chilopode). Arch. Zool. Exp. Gen. 110, 341-348.
Joly, R., et Lehouelleur, J. (1972). Effet de la section antennaire sur le dtclenchement de la mue chez Lithobius forfcatus L. (Myriapode Chilopode). Gen. Camp. Endocrinol. 19, 320-324.
Jube~hie-Jupeau, L. (1963). Recherches sur la repro- duction et la mue des Symphyles. Arch. Zoof. Exp. Gen. 102, 1-172.
Kambysellis, M. P., and Williams, C. M. (1971). In vitro development of insect tissues. II. The role of ecdysone in the spermatogenesis of silkworms. Biof. Bull. 141, 541-5.52.
K~shnakumaran, A., and Schneiderman, H. A. (1970). Control of molting in mandibulate and chelicerate arthropods by ecdysones. Biol. Bull. 139, 520-538.
Laverdure, A. M. (1976). Action de l’cr- et de la P-ecdysone sur l’ivolution de l’ovaire nymphal de Tenebrio molitor. Etude en culture in vitro. Ac- tualit6s sur les hormones. Colloque No. 251. CN.R.S. Ed., pp. 367-374.
Lender, T. H., et Duveau-Hagege (1%3). La survie et la differentiation en culture in vitro des gonades de larves de dernier age de Galleria mellonella (LepidoptenxPyralididae). Develop. Biol. 6, l-22.
Nishjitsutsuji-Uwo, 5. (1961). Further studies on the developmental features of the male germ cells in the diapausing and non-diapausing swallowtails, Papilio xuthus. Annot. Zool. Japon. 34, 30-37.
Robbins, W. E., Kaplanis, J. N., Thompson, M. H., Shortino, T. J., Cohen, C. F., and Joyner, S. C. (1968). Ecdysones and analogs: Effects on de-
velopment and reproduction of insects. Science 161, 1158-l 160.
Rouquette, M. (1970). Etude du tissu ovarien chez le Crabe Pachygrapsus marmoratus (Fabricius). Premiers resultats concemant les roles de la tem- perature et des pidoncules oculaires. Bull. Sot. ZooI. Fr. 95, 23.5-240.
Schmidt, E. L., and Williams, C. M. (1953). Physiol- ogy of insect diapause. V. Assay for the growth and differentiation hormone of Lepidoptera by the method of tissue culture. Biol. Bull. 105,174-187.
Schwartz, D. (1960). “Methodes Statistiques a I’Usage des Medecins et des Biologistes.” Ed. Medicales Flammarion, Paris.
Takeda, N. (1972). Effect of ecdysterone on spermato- genesis in the diapausing slug moth pharate pupa, Monema flavescens. J. Insect Physiol. 18, 571- 580.
Takeuchi, S. (1969). Endocrinological studies on spermatogenesis in the silkworm, Bombyx mori. Devel. Growth Differ. 11, 8-28.
Verhoeff, K. W. (19.50). Uber die Ent~cklungstufen der Steinlaiifer, Lithobiiden und Beitriige Zur Kenntnis der Chilopoden. Zool. Jahrb. (Suppl.) 7, 195-298.
Wright, J. E., and Kaplanis, J. N. (1970). Ecdysone and ecdysone analogues: Effects on fecundity of the stable fly, Stomoxys calcftrans. Ann. En- tomol. Sot. Amer. 65, 622-623.
Yagi, S., Kondo, E., and Fukaya, M. (1969). Hormone effect on cultivated insect tissues. I. Effect of ecdysone on cultivated testes of diapausing rice stem borer larvae (LepidopteraPyralidae). Appl. Entomol. Zool. 4, 70-78.


















