Hérédité cytoplasmique.pdf
22
Génétique 2 Hérédité cytoplasmique 1866 : Mendel publie ses lois de l'hérédité à partir d'études menées chez le Pois Pisum salivum : ➢ Première loi : uniformité des h ybrides en première génération ➢ Deuxième loi : ségrégation des caractère en seconde génération ➢ Troisième loi : Disjonction indépendante de caractère en F2 Les chromosomes, support de l'hérédité... Dans les années 1950-1960, on a découvert de l'ADN et des ribosomes dans les mitochondries et les chloroplastes Comment expliquer la présence d'ADN dans les mitochondrie et chloroplaste ? Selon Margulis et Taylor , les cellules eucaryotes proviennent de l'association de plusieurs proca- ryotes : on parle de théorie endosymbiotique • Absorption d'une bactérie par une cellule eucaryote primitive et formation d'une cellule eu- caryote hétérotrophe. Les bactéries absorbées deviennent des mitochondries et pratiquent la respiration • Réalisation d'une cellule eucaryote autotrophe par absorption d'une bactérie photosyn- thétique par une cellule eucaryote hétérotrophe. Ces bactéries deviennent des chloroplastes, ses membranes internes ont une origine bactérienne Quels sont les faits qui étayent cette théorie ? ✔ La taille des mitochondries et des chloroplastes est semblable à celle des bactéries ✔ Chacun de ces organites possède un matériel génétique (ADN) qui lui est propre ✔ Chacun de ces organites possède le matériel nécessaire pour la synthèse protéique(ARNt, ri- bosomes, poly mérases) ✔ Chacun de ces organites peut se diviser par étranglement médian (après avoir dupliqué le matériel génétique) La ressemblance entre un chloroplaste de cellule eucaryote actuelle et d'une bactérie photosyn- thétique (cyanobactérie) est confortée par plusieurs caractères : ✔ L'ADN du chloroplaste est circulaire et non associé à des histones comme chez les bactéries ✔ Cet ADN code pour une partie des protéines des chloroplastes (organites semi autonomes) ✔ Une partie de la synthèse de protéines chloroplastiques s'effectu e dans le chloroplaste, grâce a la présence de ribosomes qui présentent des analogies avec les ribosomes bactériens ✔ La division des chloroplastes suit un r ythme indépendant de la division du noyau ✔ Tour chloroplaste provient d'un plaste préexistant ✔ Chez les plantes supérieurs, les deux membranes de l'enveloppe du chloroplaste sont différentes : la membrane interne ainsi que la membranes des thylakoïdes présentent des analogies avec les membranes bactériennes 1
-
Upload
coursenlignelille1 -
Category
Documents
-
view
222 -
download
0
Transcript of Hérédité cytoplasmique.pdf

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 1/22
Génétique 2
Hérédité cytoplasmique
1866 : Mendel publie ses lois de l'hérédité à partir d'études menées chez le Pois Pisum salivum :➢ Première loi : uniformité des hybrides en première génération➢ Deuxième loi : ségrégation des caractère en seconde génération➢ Troisième loi : Disjonction indépendante de caractère en F2
Les chromosomes, support de l'hérédité...Dans les années 1950-1960, on a découvert de l'ADN et des ribosomes dans les mitochondries et leschloroplastes
Comment expliquer la présence d'ADN dans les mitochondrie et chloroplaste ?
Selon Margulis et Taylor, les cellules eucaryotes proviennent de l'association de plusieurs proca-ryotes : on parle de théorie endosymbiotique
• Absorption d'une bactérie par une cellule eucaryote primitive et formation d'une cellule eu-caryote hétérotrophe. Les bactéries absorbées deviennent des mitochondries et pratiquent larespiration
• Réalisation d'une cellule eucaryote autotrophe par absorption d'une bactérie photosyn-thétique par une cellule eucaryote hétérotrophe. Ces bactéries deviennent des chloroplastes,ses membranes internes ont une origine bactérienne
Quels sont les faits qui étayent cette théorie ?
✔ La taille des mitochondries et des chloroplastes est semblable à celle des bactéries✔ Chacun de ces organites possède un matériel génétique (ADN) qui lui est propre✔ Chacun de ces organites possède le matériel nécessaire pour la synthèse protéique(ARNt, ri-
bosomes, polymérases)✔ Chacun de ces organites peut se diviser par étranglement médian (après avoir dupliqué le
matériel génétique)
La ressemblance entre un chloroplaste de cellule eucaryote actuelle et d'une bactérie photosyn-thétique (cyanobactérie) est confortée par plusieurs caractères :
✔ L'ADN du chloroplaste est circulaire et non associé à des histones comme chez les bactéries✔ Cet ADN code pour une partie des protéines des chloroplastes (organites semi autonomes)✔ Une partie de la synthèse de protéines chloroplastiques s'effectue dans le chloroplaste, grâce
a la présence de ribosomes qui présentent des analogies avec les ribosomes bactériens✔ La division des chloroplastes suit un rythme indépendant de la division du noyau✔ Tour chloroplaste provient d'un plaste préexistant✔ Chez les plantes supérieurs, les deux membranes de l'enveloppe du chloroplaste sont
différentes : la membrane interne ainsi que la membranes des thylakoïdes présentent desanalogies avec les membranes bactériennes
1

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 2/22
Génétique 2
Certains gênes contenue dans l'ADN des plastes ou des mitochondries peuvent être transférés augénomes nucléaire, mais les gènes ne sont jamais perdus
Conclusions
➢ Il existe un génome nucléaire et des génomes extra-nucléaires !➢ Il existe une hérédité mendélienne et une hérédité non mendélienne !
DOLLY : Le fait que le clonage de Dolly ait eu un tel succès est dû au fait que ce fut le premier mammifère né sans une fécondation d'un ovule par un spermatozoïde ait eu lieu, par la technique dela constitution d'un embryon par fusion en laboratoire d'un noyau cellulaire et d'un ovule énucléé :la brebis ainsi née possède un patrimoine nucléaire identique à un individu déjà existant. Cependant,Dolly n'est pas le clone parfait de Belinda. En effet, dans l'ovule utilisé, comme dans toutes lescellules, se trouvaient des organites, les mitochondries (les « usines énergétiques » de la cellule),qui possèdent leur propre patrimoine génétique. Dolly est donc aussi le fruit de la brebis qui a donnéson ovule.Les chercheurs se sont vite aperçus qu'elle vieillissait rapidement. L'arthrite prématurée de Dolly aété attribuée au fait que la cellule prise sur Belinda alors âgée de 6 ans disposait de télomèresraccourcis. Cela est dû au fait que la longueur des télomères est réduite après chaque divisioncellulaire. De fait, Dolly avait un patrimoine génétique vieux de 6 ans dès sa naissance.
3 thèmes seront abordés dans le cours :
- Les stérilités mâles cytoplasmique chez le végétaux
- L'hérédité chloroplastique
- Les pathologies mitochondriales humaines
Chapitre 1 : Les stérilités mâles cytoplasmique chez levégétaux
Qu'est ce la stérilité mâle cytoplasmique ?C'est un système de stérilité mâle qui met en cause à la fois le cytoplasme par des gènes mitochon-driaux et le noyau par des gènes nucléaires particuliers (de stérilité ou de restauration fertilité). Lemécanisme de stérilité est porté par le génome mitochondrial. Si dans le noyau appartient des gènesde restauration de la fertilité, alors la plante sera fertile
La SMC (stérilité mâle cytoplasmique) a donc un déterminisme nucleo-cytoplasmique
Plante monoécie : sur le même pied, présence de fleur mâle et femelle
SMC :• Elle existe à l'état naturel ! Des individus mâles stériles et des individus hermaphrodites
peuvent, par exemple, coexister au sein d'une population naturelle : on parle de gynodioécie• Elle concerne aujourd'hui plus de 150 espèces
2
7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 3/22
Génétique 2
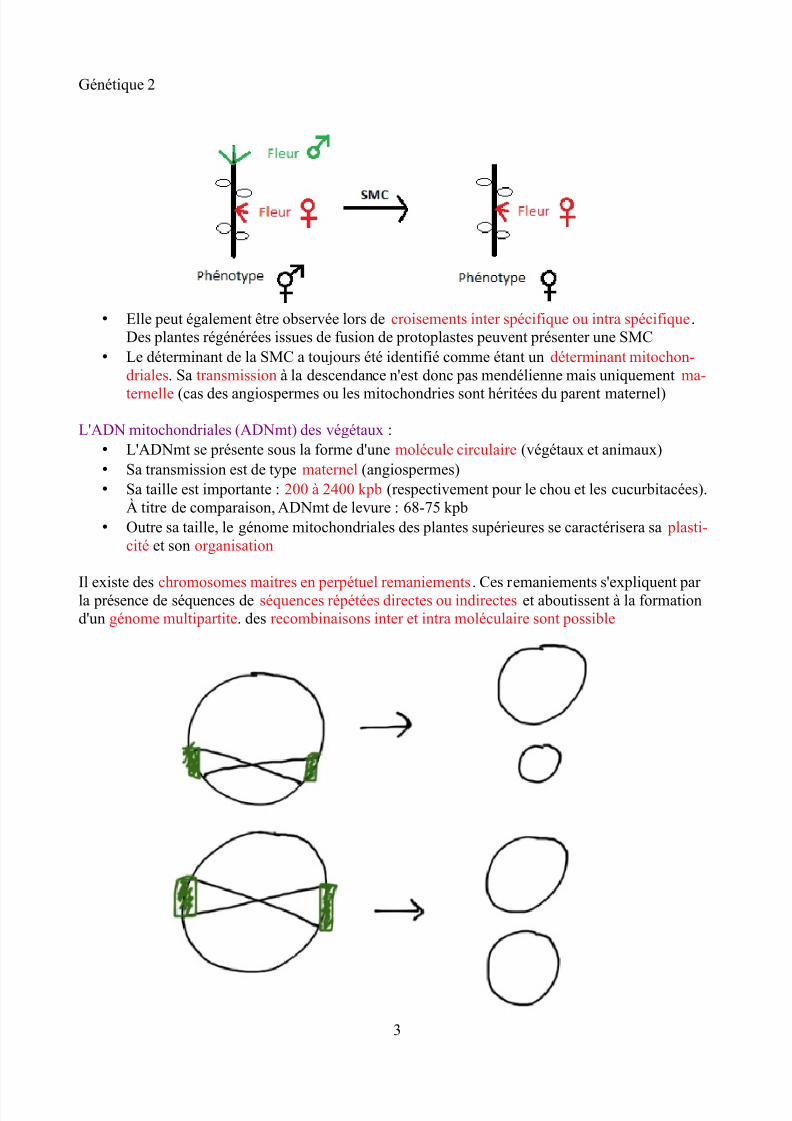
• Elle peut également être observée lors de croisements inter spécifique ou intra spécifique.Des plantes régénérées issues de fusion de protoplastes peuvent présenter une SMC
• Le déterminant de la SMC a toujours été identifié comme étant un déterminant mitochon-driales. Sa transmission à la descendance n'est donc pas mendélienne mais uniquement ma-
ternelle (cas des angiospermes ou les mitochondries sont héritées du parent maternel)
L'ADN mitochondriales (ADNmt) des végétaux :• L'ADNmt se présente sous la forme d'une molécule circulaire (végétaux et animaux)• Sa transmission est de type maternel (angiospermes)• Sa taille est importante : 200 à 2400 kpb (respectivement pour le chou et les cucurbitacées).
À titre de comparaison, ADNmt de levure : 68-75 kpb• Outre sa taille, le génome mitochondriales des plantes supérieures se caractérisera sa plasti-
cité et son organisation
Il existe des chromosomes maitres en perpétuel remaniements. Ces remaniements s'expliquent par la présence de séquences de séquences répétées directes ou indirectes et aboutissent à la formationd'un génome multipartite. des recombinaisons inter et intra moléculaire sont possible
3
7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 4/22
Génétique 2
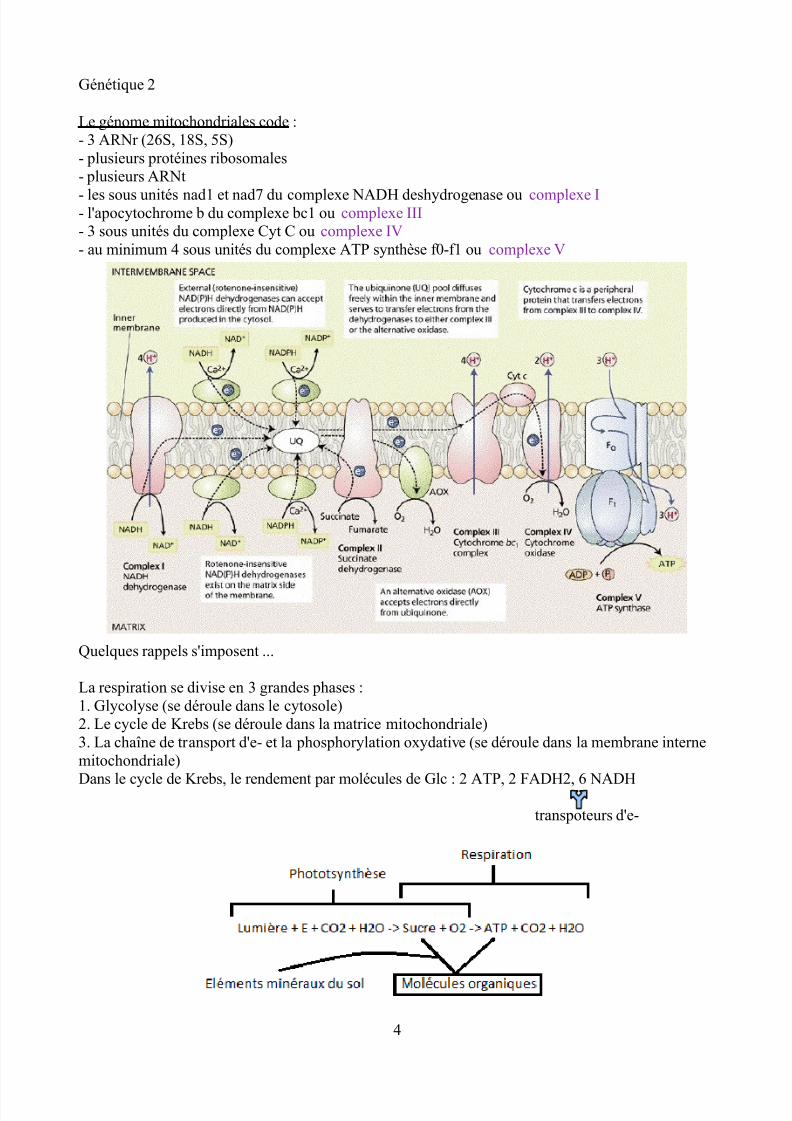
Le génome mitochondriales code :- 3 ARNr (26S, 18S, 5S)- plusieurs protéines ribosomales- plusieurs ARNt
- les sous unités nad1 et nad7 du complexe NADH deshydrogenase ou complexe I- l'apocytochrome b du complexe bc1 ou complexe III- 3 sous unités du complexe Cyt C ou complexe IV- au minimum 4 sous unités du complexe ATP synthèse f0-f1 ou complexe V
Quelques rappels s'imposent ...
La respiration se divise en 3 grandes phases :1. Glycolyse (se déroule dans le cytosole)2. Le cycle de Krebs (se déroule dans la matrice mitochondriale)3. La chaîne de transport d'e- et la phosphorylation oxydative (se déroule dans la membrane internemitochondriale)
Dans le cycle de Krebs, le rendement par molécules de Glc : 2 ATP, 2 FADH2, 6 NADHtranspoteurs d'e-
4

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 5/22
Génétique 2
Le génome mitochondrial code, notamment, diverses sous unités de la chaîne respiratoire (ellemême composés de 4 complexes et de l'ATP synthase)
L'altération de ces gènes ou d'autres gènes mitochondriaux, peut conduire à la SMC
Intérêt de la SMC dans les schémas de sélection :
Il est intéressant, en amélioration végétales, de créer des variétés de type hybrides, lesquelles présentent souvent des performances agronomiques supérieures au meilleur de leur parent (par ex-emple un rendement ou une résistance accrue)
On parle de vigueur hybride ou d'effet hétérosis
→ La plante fille aura une meilleur résistance que les 2 parents
L'effet d'hétérosis se traduit par la supériorité pour de nombreux caractères de l'individu hybride (vi-gueur, rendement, résistance aux maladies, précocité...) sur la moyenne des 2 parents ou sur le meil-leur des 2 parents
Il faut éviter l'autofécondation des plantes → on castre donc les fleurs mâles du maïs qui est censéêtre la "femelle" → la technique fonctionne bien mais fastidieuse car il faut arracher les fleurs
Schéma de sélection "classique" chez le maïs :
Parent 1 Parent 2Sélection des parents géniteurs
Sélection des descendants en vérifiant qu'ils possèdent lecaractère intéressant des parents
Obtention de lignées pures (autofécondation)
Hybridation (croisement des lignées pures)
La SMC peut être utilisé dans les schémas de sélection pour réaliser des croisements contrôlés etéviter, par exemple, la castration de la plante
5
7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 6/22
Génétique 2
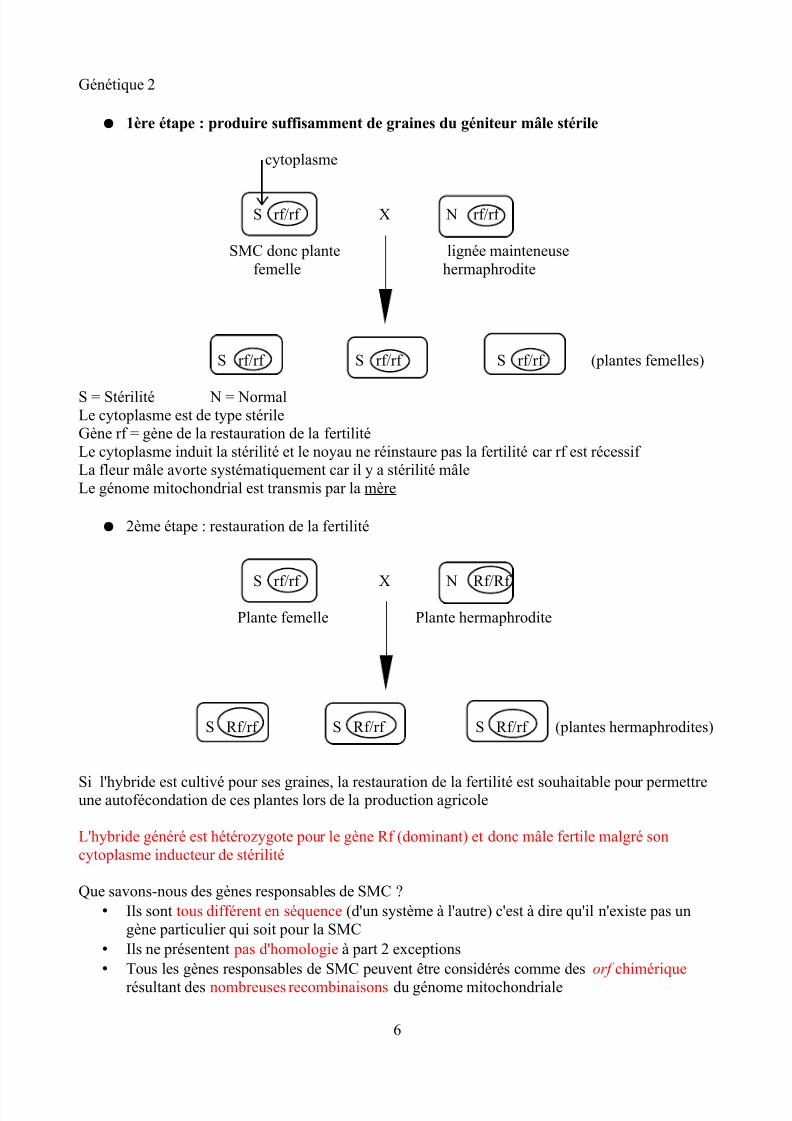
● 1ère étape : produire suffisamment de graines du géniteur mâle stérile
cytoplasme
S rf/rf X N rf/rf
SMC donc plante lignée mainteneusefemelle hermaphrodite
S rf/rf S rf/rf S rf/rf (plantes femelles)
S = Stérilité N = NormalLe cytoplasme est de type stérileGène rf = gène de la restauration de la fertilitéLe cytoplasme induit la stérilité et le noyau ne réinstaure pas la fertilité car rf est récessif La fleur mâle avorte systématiquement car il y a stérilité mâleLe génome mitochondrial est transmis par la mère
● 2ème étape : restauration de la fertilité
S rf/rf X N Rf/Rf Plante femelle Plante hermaphrodite
S Rf/rf S Rf/rf S Rf/rf (plantes hermaphrodites)
Si l'hybride est cultivé pour ses graines, la restauration de la fertilité est souhaitable pour permettreune autofécondation de ces plantes lors de la production agricole
L'hybride généré est hétérozygote pour le gène Rf (dominant) et donc mâle fertile malgré soncytoplasme inducteur de stérilité
Que savons-nous des gènes responsables de SMC ?• Ils sont tous différent en séquence (d'un système à l'autre) c'est à dire qu'il n'existe pas un
gène particulier qui soit pour la SMC• Ils ne présentent pas d'homologie à part 2 exceptions• Tous les gènes responsables de SMC peuvent être considérés comme des orf chimérique
résultant des nombreuses recombinaisons du génome mitochondriale
6

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 7/22
Génétique 2
• Ces gènes sont co-transcrits avec un gène mitochondrial essentiel. Cette contrainte reste unecaractéristique essentielle du maintien de la SMC dans les populations naturelles
Que savons-nous des polypeptides codés par ces gènes ?
– La plupart du temps, ils sont présents dans tous les tissus de la plante– Les protéines de SMC sont généralement associées aux membranes mitochondriales
4 exemples :
SMC Texas SMC IS112C(graminée)
SMC PET-1(tournesol)
SMC Ogura(radis)
La SMC Texas chez le MaïsLe cytoplasme T (T pour Texas) confère :
– La SMC– La sensibilité à des toxines de champignons
→ toxine BmT de Bipolaris maydis → toxine PmT de Phyllasticta maydis
– La sensibilité à une insecticide→ le méthomyl
→ 13 kDa
Individu stérile Individu fertile
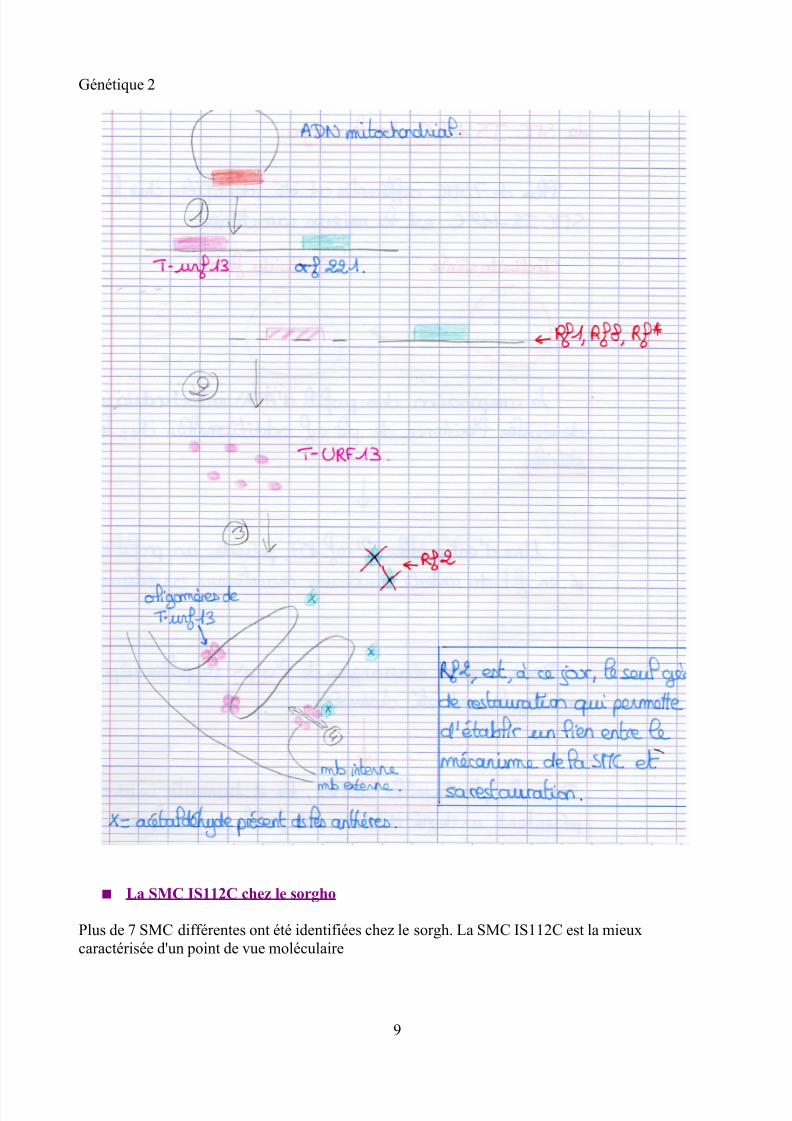
La protéine T-URF 13 est codée par un gène mitochondrial chimérique composée de :➢ la partie 3' non codante de l'ARN ribosomique 26S➢ 9 codons d'origine inconnue➢ une partie de la région codante de l'ARN ribosomique 26S
Le gène T-URF 13 est co-transcrit avec l'orf221 identifié comme codant une protéine spécifique dela mitochondrie végétale
La protéine T-URF 13 est localisée dans la membrane interne des mitochondries et s'associe enoligomères. Chaque monomère de la protéine T-URF 13 est constitué de 3 hélices alpha.L'association de 6 à 8 monomères de T-URF permettrait la formation d'un pore
Il est établit que la toxine de Boipolaris maydis interagit spécifiquement avec la protéine T-URF 13conduisant à l'altération du potentiel membranaire par l'ouverture de ce pore
Bmt
7

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 8/22
Génétique 2
Chez les plantes mâles stériles, il doit y avoir un facteur inconnu « x » (spécifique des tissus del'anthère et similaire à la toxine BmT) viendrait se fixer sur le pore formé par T-URF 13
Cette association provoquerait l'ouverture du pore aboutissant à la destruction du potentielélectrochimique de protons et ainsi à la mort cellulaire
STERILITE (mâle)
Plusieurs gènes de restauration de fertilité (=rf) ont été identifiés chez le maïs (gène nucléaire)– Rf1, Rf8, Rf*– Rf2
La restauration du phénotype mâle fertile nécessite la présence des gène Rf1 (ou Rf8, Rf*)et Rf2
Les gènes Rf1, Rf8, Rf* affectent chacun le profil de la transcription (ADN à ARNm) du gène T-URF 13 d'une façon différente, mais conduisant tous à la diminution de la quantité de transcrits (on
aura donc moins de protéines T-URF, donc moins de formation de pore au niveau de la membrane
interne de la mitochondrie donc moins de « chance » de voir les pores s'ouvrir)
L'effet le plus drastique est celui de Rf1 qui permet une diminution de 80 % de la quantité detranscrits du gène T-urf13 dans les tissus végétatifs
Le gène Rf2 code une enzyme, un aldéhyde déshydrogénase. Cette protéine jouerait un rôle
important dans la détoxification des intermédiaires de réaction issus de la fermentation alcooliquecomme l'acétaldéhyde dans les anthères.
Cette donnée suggère que le composé X soit l'acétaldéhyde
BmT ↔ X = Acétaldéhyde
La protéine Rf2 empêcherait donc la fixation de l'acétaldéhyde sur la protéine T-urf13 permettant degarder l'intégralité mitochondriale pendant la gamétogénèse
8
7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 9/22
Génétique 2
La SMC IS112C chez le sorgho
Plus de 7 SMC différentes ont été identifiées chez le sorgh. La SMC IS112C est la mieuxcaractérisée d'un point de vue moléculaire
9
7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 10/22
Génétique 2
La comparaison des profils d'ADN mitochondriaux a permis de révéler l'existence de plusieurs orf
additionnelles chez les plantes stériles
Une d'entre elles, l'orf107 présente un profil de transcription différent en fonction du contexte
nucléaire mainteneur ou restaurateur
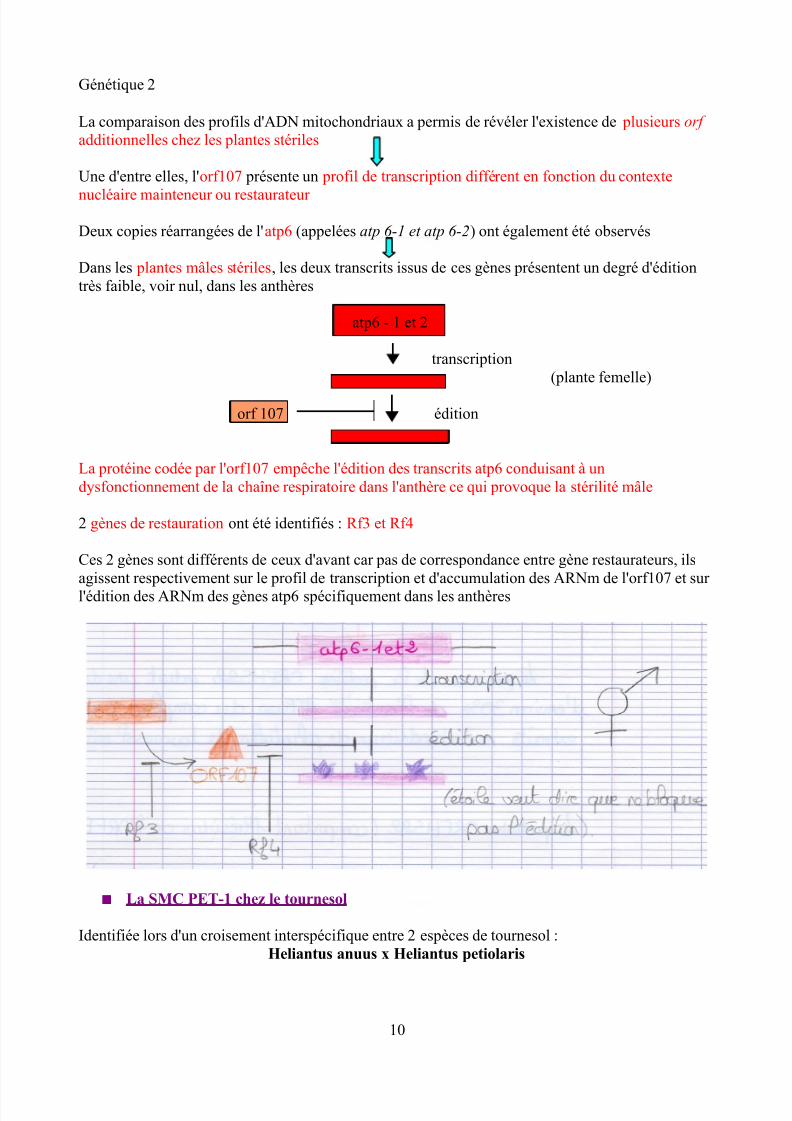
Deux copies réarrangées de l'atp6 (appelées atp 6-1 et atp 6-2) ont également été observés
Dans les plantes mâles stériles, les deux transcrits issus de ces gènes présentent un degré d'éditiontrès faible, voir nul, dans les anthères
atp6 - 1 et 2
transcription(plante femelle)
orf 107 édition
La protéine codée par l'orf107 empêche l'édition des transcrits atp6 conduisant à undysfonctionnement de la chaîne respiratoire dans l'anthère ce qui provoque la stérilité mâle
2 gènes de restauration ont été identifiés : Rf3 et Rf4
Ces 2 gènes sont différents de ceux d'avant car pas de correspondance entre gène restaurateurs, ilsagissent respectivement sur le profil de transcription et d'accumulation des ARNm de l'orf107 et sur l'édition des ARNm des gènes atp6 spécifiquement dans les anthères
La SMC PET-1 chez le tournesol
Identifiée lors d'un croisement interspécifique entre 2 espèces de tournesol :Heliantus anuus x Heliantus petiolaris
10

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 11/22
Génétique 2
La comparaison des génomes mitochondriaux a permis des modifications dans une région de 17kbautour du gène codant pour la sous-unité alpha de l'ATPase (atp1). Cette région comprends uneinversion de 12kb et une insertion de 5kb
Ces modifications touchent la région 3' du gène atp1 donnant naissance à une orf supplémentaire :l'orf H522
Cette orf est co-transcrite avec l'atp1. Par ailleurs, ses premiers codons sont homologues avec l'orf Balors que le reste de la séquence est d'origine inconnue
L'orfH522 code une protéine de 15KDa intrinsèque aux membranes mitochondriales
La présence de la protéine ORFH522 induit une diminution d'environ 30% de l'activité ATPsynthase (complexe V) dans les extraits mitochondriaux de plantules de tournesol mâle stériles
La protéine ORFH522 (comportant 18 résidus de l'ORFB en position N-terminale) entre en
compétition avec la protéine ORFB dans l'assemblage ou le fonctionnement du complexe F1-F0synthase
La SMC Ogura
Cette SMC a été découverte en 1968 par Ogura au Japon chez une variété asiatique de Radius(Raphanus sativus)
Les observations cytologiques réalisées sur les plants de radis mâles stériles ont montré que :– Certaines structures avortes dont les microspores–
Une dégénérescence prématurée de l'assise tapétaleLes selectionneurs du chou et du colza, qui recherchaient un système fiable de production dessemences hybrides, ont entrepris le transfert de la SMC Ogura, dans ces Brassica cultivées, par descroisements interspécifiques
11

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 12/22
Génétique 2
Ils ont réussi ce qu'ils voulaient mais :- déficience en chlorophylle à basse température- faible production de nectar - fertilité femelle altérée
→ donc plantes inintéressantesLe problème vient des chloroplastes qui sont d'origine maternelle (du radis)
Pour éliminer ces caractères délétères, des fusions de protoplastes ont été entreprises (car on peut pas retirer les chloroplastes alors obligé de faire cette fusion pour générer non pas des hybrides maisdes cybrides)
Parmi les plantes obtenues après régénération (on parle de cybrides) les plantes mâles stériles ne présentant plus les caractères indésirables ont été sélectionnées
De nombreuses différences entre les génomes mitochondriaux des cybrides ont été observés,
suggérant qu'un grand nombre d'événement de recombinaison avait eu lieu entre les génomes parentaux
Une fusion « + » n'est pas un croisement « x »
La comparaison des profits de restriction d'ADNmt des cybrides obtenus a montré que denombreuses différences pouvaient être observées
Un de ces cybrides présentait un comportement particulier : une forte proportion de sa descendancemontrait une réversion vers la fertilité (interprétée comme une instabilité de son génomemitochondrial)
L'absence de transcrits de l'orf138 chez les révertants fertiles a confirmé la corrélation entrel'orf138 et la SMC ogura
12

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 13/22
Génétique 2
Chapitre 2 : Hérédité cytoplasmique
Le proplastide est l'organisme d'où dérive le chloroplaste. Dans les cellules méristématiques(cellules de croissance), le chloroplaste existe sous une forme réduite → le proplastideLes thylakoïdes sont issus de la membrane interne du proplastide qui s'invagine et s'assemble envésicule
Expérience de CORRENS (1909) sur la panachure des feuilles de Mirabilisjalapa (La belle de nuit)→ Chez le parent femelle, une structure est transmise par celle-ce. Transfert du chloroplaste du
parent femelle vers l'hybride F1, ce transfert donne la couleur. L'hybridation F1 a le même phénotype que celui du parent femelle
➢ Le chloroplaste et la mitochondrie du seigle sont hérités des 2 parents (hérédité biparentale)
➢ Chez Actinidia : chloroplaste → hérédité paternelle et mitochondrie → hérédité maternelle→ Grande diversité, l'hérédité dépend de la plante prise en compte
ADN chloroplaste :– Petit (environ 150 kb) et circulaire– Nombreuses copies(50-100)– Peu de gène (environ 120)
.ARNr
.ARNt pour la traduction plastidiale
.Gènes pour le ribosome chloroplastique
.4 gènes codant des sous-unités de l'ARN polymérase
.Un gène pour la grande sous unité de la RUBISCO
.9 gènes pour les photosynthèses I et II
.6 gènes pour l'ATP synthase
Les gènes de l'ADNcp du Maïs :
Gène codant pour la grande sous unité de la RUBISCO = Rbc L
Le génome chloroplastique du maïs :
IRA et IRB = 2 régions répétées (diffère d'une plante à l'autre)Il y a toujours le même nombre. Ces 2 régions donnent lieu à des taille de génomes différents → 2sous génome : un petit et un grand
L'hérédité des chloroplastes pouvait être paternel, maternelle ou biparentalLes chloroplastes sont présent dans les tissus verts, mais ils sont absents dans les pollens (gamètemâle). Les chloroplastes sont généralement hérités par voie maternelle
Cette caractéristique a suggéré la possibilité de créer des plantes transgéniques en insérant le gèned’intérêt dans le génome du plaste
Pas de transfert du gène par le pollen
13

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 14/22
Génétique 2
Technique de transformation du chloroplaste :
Après bombardement des feuilles avec l'ADN contenant le gène d’intérêt et la recombinaisonhomologue (insertion) avec l'ADN chloroplastique, on obtient une plante transformée
Canon à particule : Des micro-projectiles d'or enrobés d'ADN sont projetés sur une feuille placée
dan l'enceinte sous vide du canon. Les feuilles sont découpées en petits fragments placés sur unmilieu de sélection contenant la spectinomycine. Les cellules renfermant des chloroplastestransformés vont se diviser et donner des cals puis des plantules qui pourront être repiquées sur sollorsqu'elles auront raciné. Après traitement au PEG en présence d'ADN, les protoplastes ensuspension dan le milieu liquide se divisent et forment des micro-cals. Les plantules qui sedéveloppent à partir du cal sont des transformants chloroplastiques qui pourront être repiqués sur sol lorsque les racines seront développées
Quelques applications de la transformation du chloroplaste :
Résistance au glyphosate
Daniell et al (1998) a transformé avec succès des plants de tabac avec le gène de résistance auglyphosate inséré dans le génome du chloroplaste. Les plantes sont résistantes et le gène ne peutêtre transféré par le pollen à d'autres végétaux
Résistance aux insectes grâce à la toxine Bt
Les toxines de Bacillus thuringensis (Bt) sont toxiques pour les insectes après ingestion. Kota et al(1999) ont observé que l'expression de la toxine Bt dan les chloroplastes permet de protéger les
plantes des attaques (i.e. pyrale Ostrinia nubilalis)
14
7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 15/22
Génétique 2
Un exemple de coopération des génomes : la RUBISCO :
• protéine la plus importante dan la biosphère• elle catalyse la carboxylation du RuBP lors d la 1ère étape du cycle de Calvin
• purifiée par Wildman et Bonner en 1947• fonction élucidée par Quayle et al en 1954• jusqu'à 65% des protéines foliaires
Structure de la RUBISCO
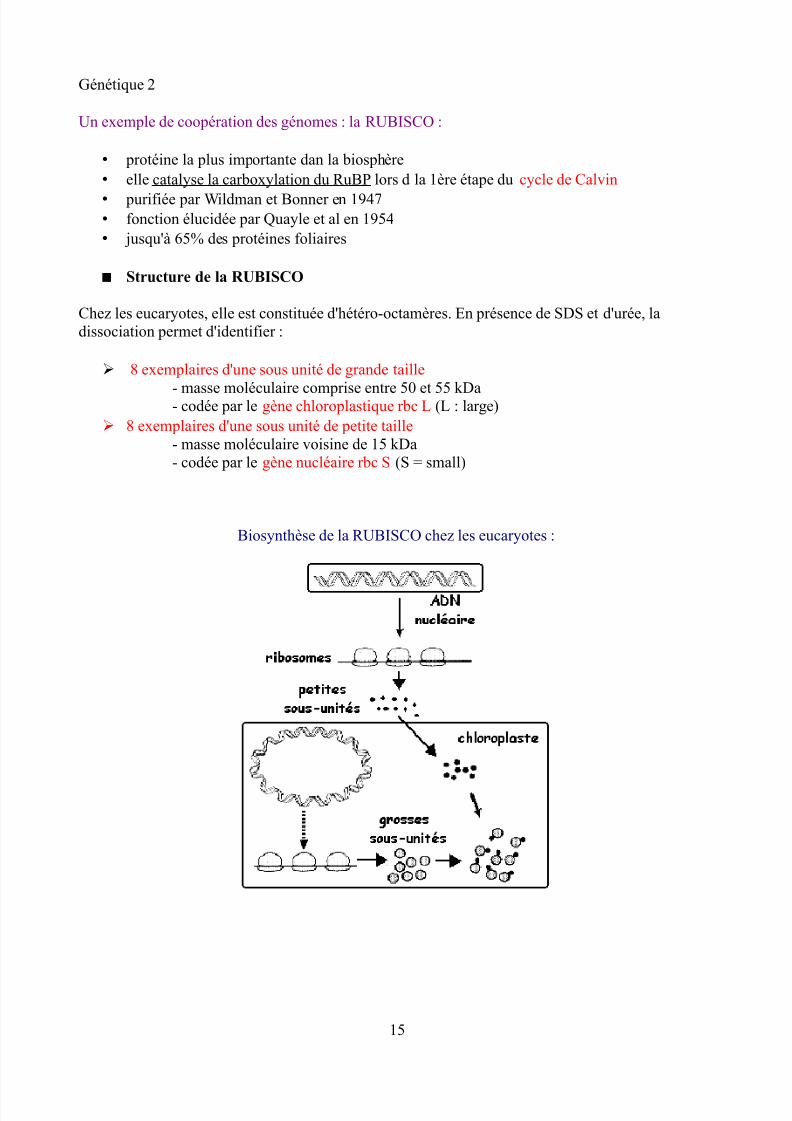
Chez les eucaryotes, elle est constituée d'hétéro-octamères. En présence de SDS et d'urée, ladissociation permet d'identifier :
➢ 8 exemplaires d'une sous unité de grande taille
- masse moléculaire comprise entre 50 et 55 kDa- codée par le gène chloroplastique rbc L (L : large)➢ 8 exemplaires d'une sous unité de petite taille
- masse moléculaire voisine de 15 kDa- codée par le gène nucléaire rbc S (S = small)
Biosynthèse de la RUBISCO chez les eucaryotes :
15

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 16/22
Génétique 2
Chapitre 3 : Les maladies humaines transmises par les
mitochondries
Chaine respiratoire : composée de plusieurs complexes. Les protons via l'ATPase produisent del’énergie. Mitochondrie siège de la respiration et du métabolismes énergétique
Il existe 2 cas de figures :- défaillance du génome nucléaire → défaillance de la chaine respiratoire- défaillance du génome mitochondrial → perturbe le métabolisme énergétique
On s’intéresse dans ce cours, aux défaillance du génome mitochondriale
On distingue 4 types de maladies génétiques : ✗ les maladies héréditaires → elles sont transmises selon un mode héréditaire suivant les lois
de Mendel : dominant, récessif, autosomique ou lié à l'X ✗ les maladies par aberration chromosomique → ex : trisomie 21 ✗ les maladies mitochondriales ✗ les maladies multifactorielles → ex : diabète, hypertension artérielle ...
Les mécanismes conduisant à l'apparition de ces pathologies sont différents :- pour les 3 premières → mutations génétiques ou chromosomiques- pour les maladies multifactorielles → gènes de susceptibilité
Hérédité maternelle des mitochondries :
Pénétration du noyau spermatique dans l'ovocyte. Les mitochondries paternelles ne sont pasretrouvés dan l'embryon formé
Caractéristiques Génome nucléaire Génome mitochondrial
Taille environ 3,3 x 10 pb 16569 pb
Nombre de moléculed'ADN/cellule
23 dans cellules haploïdes/46dans cellules diploïdes
plusieurs milliers de copies par cellule
Nombre de gène codant environ 20 000 à 30 000 37 (13 polypeptides, 22 ARNr et ARNt)
Densité gènes 1 pour 40 000 pb 1 pour 450 pb
Introns absent
% ADN codant 3,00% 93,00%
Usage codon code génétique universel AUA → méthionine ; TGA→ tryptophane ; AGA et AGG
→ codon stop
Protéines associés pas d'histone
Mode d'hérédité exclusivement maternelle
16

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 17/22
Génétique 2
Les molécules d'ADNmt sont circulaires et codent :- 13 polypeptides (sous unités de la chaîne respiratoire)- 22 ARNt- 2 ARNr
Carte génétique mitochondrial :
Le génome code pour des ARNt, les ARNr et quelque protéines comme le cytochrome b et l'ATPsynthase (ATPase). Les 2 brins complémentaires de l'ADN mitochondrial sont représentés par convention comme brin léger et brin lourd
Pour que la chaine de respiratoire mitochondriale fonctionne, il faut une collaboration entre génomemitochondrial et génome nucléaire
Complexe I Complexe II Complexe III Complexe IV Complexe V
ADNmt 7 0 1 3 2
ADN nucléaire 36 4 10 10 14
Les mitochondries se divisent en permanence indépendamment des mitoses et se répartissentaléatoirement entre les cellules filles
normal
anormal cellule mère
cas intermédiaire
cellules filles
La répartition des mitochondries se fait déjà dan l'ovocyte. Ensuite, se produit des mitosessuccessives. Une cellule peut avoir une majorité de "normal" ou "anormal" → aléatoire
Une population de mitochondries génétiquement hétérogène
Une population de cellules elles-mêmes hétérogènes sur le plan de l'hérédité cytoplasmique
Certaines cellules contiennent un seul type demitochondries mutées ou non mutées : on parle
d'homoplasmie
D'autres contiennent les 2 types demitochondries en proportions variables : on
parle d'hétéroplasmie
17

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 18/22
Génétique 2
Lorsque l'on a des mitochondries anormales :➢ on peut altérer le complexe de la chaîne respiratoire auquel appartient le peptide et donc
altération de la chaîne respiratoire entière➢ altération de la synthèse protéique
→ Organes affectés par la défaillance de la chaîne respiratoire : muscles, cœur, cerveau ... →organes nécessitant un apport énergétique important
Ce qui est important, c'est la balance entre homoplasmie et hétéroplasmie
L'hétéroplasmie rend compte du phénomène de "spécificité tissulaire" qui affecte souvent lesmaladies mitochondriales
Chaque tissu a un seuil spécifique de mitochondries mutées au deçà duquel il peut être cliniquementnormal
Le rôle central des mitochondries et la complexité de la chaîne respiratoire expliquent la sévérité etla grande diversité des cytopathies mitochondriales parmi les maladies mitochondriales
Ces pathologies interviennent dans des organes qui nécessitent des apports importants en ATP
Exemple : cœur, muscles striés, système nerveux, reins, foie, pancréas
Des symptômes et des causes multiples ...
La diversité et la complexité des symptômes posent le problème du diagnostique ...Des gènes nucléaires et mitochondriaux peuvent êtres impliqués ...
On distingue 2 grandes catégories de maladies mitochondriales :➢ implication de mutation/délétions dan l'un des 22 gènes plus 2 gènes intervenant dan la
synthèse protéique➢ anomalies concernant les polypeptides = la structure de la chaîne respiratoire
On a trouvé qu'une grande délétion (jusqu'à 1/4 de la cellule) peut causer la perte d'un grand nombrede gènes :
➢ si ne concerne que quelques mitochondries → pas grave➢ si concerne une majorité de mitochondries → synthèse protéique plus chaîne respiratoire
très défaillante → mort possible
La "constellation" d'anomalies associées à une pathologie mitochondriale :• le gène mitochondrial muté• le niveau de ségrégation après la réplication d'un génome mitochondrial muté au cours des
1ères étapes du développement embryonnaire• l'abondance du gène mitochondrial muté dans un tissu donné• le niveau critique de la quantité d'ADN muté nécessaire à la manifestation clinique de
l'anomalie dans un tissu
18

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 19/22
Génétique 2
Réarrangement de l'ADNmt :
Délétion Syndrome de PearsonDuplications Syndrome de Kearns-Saye (KSS)
KSS : principale symptôme = ophtalmoplégie externe progressive (=PEO) (problème mouvementde l’œil)
• dégénérescence de la couche pigmentaire de la rétine• trouble de la conduction cardiaque• ataxie• surdité• démence• diabète
La maladie se situe généralement avant 20 ans. En générale, le sujet meurt avant 20 ansEn général, délétion de grande taille (plus de 1000 pb) de l'ADNmt également des délétions. 1/3 des patients présentaient une délétion de 4977 pb de l'ADNmt → cassure entre les gènes de l'ATP 8 etde NDS
proportion importante de cellules présentant des mitochondries anormales en dessous de 85% de mitochondries anormales → toujours défaillance mais moins grave lorsque les ADNmt porteur d'anomalies se trouvent dans les cellules souches du sang →
Syndrome de Pearson
La même origine peut avoir des impacts multiples causées par :
✗ homoplasmie/hétéroplasmie ✗ ségrégation (initiale au cours de l’embryogenèse)
Pour le Syndrome de Pearson, incapacité pour ces cellules sanguines à utiliser le fer dans lasynthèse d'hémoglobine → maladie très précoce et mortelle
Mutations ponctuelles de l'ADNmt affectant les ARNt :
MELAS = encéphalopathie mitochondriale avec acidose lactique et accidents vasculaires cérébrauxMaladie mitochondriale la plus fréquenteSyndromes :
• accidents vasculaires cérébraux précoces (avant 40 ans)• ataxie• contractions musculaires brusques• démence• surdité• vomissements récurrents• migraines• PEO• laptose = relâchement des paupières•
diabète
19

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 20/22
Génétique 2
Acidose lactique = augmentation de la concentration en acide lactique et entraîne la chute du pH etle pouvoir tampon des fluides corporels. Le pyruvate n'est plus métabolisé et s'accumule et deretrouve dans les fluides corporels, le pyruvate est alors transformé en acide lactique et c'est cetacide qui s’accumule
Une mutation est prépondérante dans cette maladie MTTL1*MELAS3243GMT = mitochondrial ; T = ARNt ; L1 = leucine ; 3243G = positionD'autres mutations ponctuelles peuvent être responsables de ce syndrome MELAS mais la mutationci-dessus représente 80% des cas de MELAS
✗ Si plus de 90% des cellules musculaires comportent des mitochondries portant la mutation→ forme la plus grave
✗ Si moins de 90% des cellules musculaires comportent des mitochondries portant lamutation → forme moins grave donc symptôme les moins graves (vomissements,migraines...)
MERRF = Épilepsie myochronique associée à la myopathie des fibres rouges en haillonsMaladie rare. Associe des symptômes multiples :
• crise d’épilepsie• trouble du mouvement• perte de cellule musculaires• démence• surdité• dégénérescence des nerfs spinaux
Agrégat de grandes taille de mitochondries anormales dans les cellules musculaires → se visualise par des fibres rouges en haillons → permet d'identifier plus facilement la maladie
Ce syndrome apparait plutôt durant l'enfance et implique plusieurs mutations ponctuelles :➢ une mutation ponctuelle principale = MTTK*MERRF 8344G➢ une 2ème (plus marginale) MTTK*MERRF 8356G
✗ Si plus de 90% des mitochondries sont porteuses de mutations → MERRF avec lessymptômes les plus graves
✗ Si moins de 90% des mitochondries sont porteuses de mutations → MERRF avec les
symptômes les moins gravesCes symptômes peuvent atteindre divers systèmes physiologiques :
• vue• audition• système endocrinien• cœur • reins• système digestif et excréteur
20

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 21/22
Génétique 2
Plusieurs gènes d'ARNt mitochondriaux peuvent êtres affectés et être à l'origine de plusieurs pathologies. Des mutations différentes peuvent aboutir à des manifestations cliniques similaires.Des mutations différentes sur un même ARNt peuvent aboutir à des syndromes différents
Mutation de gènes codant pour des protéines de la chaine respiratoire :
NARP = Rétinine pigmentaire avec neuropathie et ataxieMILS = la maladie de Leign
NARP : atteint jeunes adultes• faiblesse musculaire• démence• retard• convulsion•
rétinine pigmentaireMILS : maladie infantile grave
• dégénérescence de certains neurones → ceux du bulbe rachidien et des noyaux cérébraux
Ces 2 syndromes sont induit par les mutation qui touche l'ATP6 :- MTATP6*NARP 8993G- MTATP6*NARP 8933C
Entre 70 et 80% → manifestation du syndrome NARPPlus de 90% de mitochondries anormales → manifestation du syndrome MILS
LHON = la neuropathie optique héréditaire de Leber
Affecte essentiellement l’œil. Trouble de la vision, puis s'aggrave et le patient perd la vision d'unœil (sans douleur) puis perte du 2ème œil. Ou alors les 2 sont affectés en même tempsIl y a dégénérescence du nerf optique plus les neurones de la rétine
Quelques symptômes périphériques :• tremblement• troubles cardiaques• perte du tonus musculaire
Apparait à 25 ans environ ou au delà de 70 ans. 5 fois plus d'hommes que de femmes affectés
18 mutations affectant 9 gènes : ND1 ; ND2 ; ND4 ; ND5 ; ND6 ; Cytb ; CO1 ; CO3 ; ATP6Dont 4 mutations principales, qui, à elles seules peuvent provoquer la manifestation clinique duLHON
Mutations primaires : 1 des 4 mutations principalesMutations secondaires : les autres mutations
21

7/23/2019 Hérédité cytoplasmique.pdf
http://slidepdf.com/reader/full/heredite-cytoplasmiquepdf 22/22
Génétique 2
Les mutations secondaires peuvent provoquer la maladie à la condition d'avoir une association avecd'autres mutations ou l'implication de mutation sur certains gènes nucléaires ou associés avec unstress environnemental (= facteurs épigénique)
90% des cas de LHON primaire sont causées par 3 mutations → MTND4*LHON 11778A ;MTND1*LHON ...
C'est toujours le complexe 1 de la chaine respiratoire qui est touchéeMTND4*LHON 11778A ralentit la vitesse d'oxydation
ND1 baisse l'efficacité du complexe 1 de 80%
Clinique 1 :• MELAS• MERFF• LHON• Pearson• MIDD (diabète, surdité)• NARP• MILS• ets ...
Clinique 2 :• Association inexpliquée de symptômes• N'importe quel organe ou tissu atteint• Plusieurs organes touchés sans origine embryologique ou fonctions biologiques communes• Évolution imprévisible
Diagnostique :• ADNmt extrait du muscle, sang, fibroblaste ...• Recherche délétion, duplication
- Pearson- Kearns-Sayre
• Recherche mutation ponctuelle (PCR)- LHON- MELAS
- MERFF• Séquençage• Estimation du degrés d'hétéroplasmie
22