
Génétique des populations - Association des …€¦ · GENETIQUE MEDICALE – Génétique des...
10
GENETIQUE MEDICALE – Génétique des populations 30/10/2014 RETAILLEAU Eloïse L2 CR : Julie Chapon Génétique Médicale Pr Philip 10 pages Génétique des populations A. Loi de Hardy-Weinberg Elle s'applique dans certaines conditions : Dans une population de dimension infinie, où les mariages se font au hasard (PANMIXIE), où il n'existe ni migration d'une autre population, ni sélection contre un phénotype particulier, où le taux de mutations est constant, les proportions des différents génotypes restent constantes d'une génération à l'autre. Cette loi permet de calculer la fréquence des hétérozygotes dans une population. Dans un système biallélique, on a trois génotypes possibles que sont AA, Aa et aa. Dans cette population idéale dans laquelle la loi s'applique, les fréquences des différents génotypes sont toujours les mêmes : AA : p² Aa : 2pq aa : q² Mais au niveau des fréquences alléliques, p est la proportion d'allèles A et q est la proportion d'allèle a. Facteurs influençant l'équilibre de Hardy-Weinberg : • Mutations • Sélection • Dérive génétique Modifient les fréquences alléliques p et q • Petites populations • Migrations 1/10 Plan A. Loi de Hardy-Weinberg B. Facteurs influençant les fréquences alléliques I. Mutations II. Sélection III. Dérive génétique C. Causes du maintien des maladies génétiques dans les grandes populations I. L'équilibre mutation-sélection II. L'avantage sélectif des hétérozygotes D. Fréquence élevée de certaines maladies très rares dans de petites populations E. Consanguinité I. Calcul du coefficient II. Conséquence de la consanguinité sur un individu III. Les conséquences de la consanguinité à l'échelle d'une population
Transcript of Génétique des populations - Association des …€¦ · GENETIQUE MEDICALE – Génétique des...

GENETIQUE MEDICALE – Génétique des populations
30/10/2014RETAILLEAU Eloïse L2CR : Julie ChaponGénétique MédicalePr Philip10 pages
Génétique des populations
A. Loi de Hardy-Weinberg
Elle s'applique dans certaines conditions :Dans une population de dimension infinie, où les mariages se font au hasard (PANMIXIE), où il n'existe ni migration d'une autre population, ni sélection contre un phénotype particulier, où le taux de mutations est constant, les proportions des différents génotypes restent constantes d'une génération à l'autre.
Cette loi permet de calculer la fréquence des hétérozygotes dans une population.
Dans un système biallélique, on a trois génotypes possibles que sont AA, Aa et aa.
Dans cette population idéale dans laquelle la loi s'applique, les fréquences des différents génotypes sont toujours les mêmes :AA : p²Aa : 2pqaa : q²Mais au niveau des fréquences alléliques, p est la proportion d'allèles A et q est la proportion d'allèle a.
Facteurs influençant l'équilibre de Hardy-Weinberg :• Mutations • Sélection• Dérive génétique Modifient les fréquences alléliques p et q • Petites populations• Migrations
1/10
Plan
A. Loi de Hardy-Weinberg B. Facteurs influençant les fréquences alléliques
I. Mutations II. Sélection
III. Dérive génétique C. Causes du maintien des maladies génétiques dans les grandes populations
I. L'équilibre mutation-sélection II. L'avantage sélectif des hétérozygotes
D. Fréquence élevée de certaines maladies très rares dans de petites populations E. Consanguinité
I. Calcul du coefficientII. Conséquence de la consanguinité sur un individuIII. Les conséquences de la consanguinité à l'échelle d'une population

GENETIQUE MEDICALE – Génétique des populations
• La consanguinité modifie la répartition des génotypes mais ne modifie pas la fréquence allélique.
B. Facteurs influençant les fréquences alléliques
I. Mutations
Le terme de mutation peut vouloir dire deux chose. C'est quelque chose de physique, un changement de base, une délétion, c'est une modification de la séquence d'ADN. Mais la mutation sert aussi à définir l'évènement de ce changement.
Définition : C'est une modification brusque et définitive du message héréditaire.Nous nous attacherons ici aux mutations géniques et non chromosomiques
Les mutations peuvent affecter :• une cellule germinale : lorsqu'elle se produit à la formation de la gamète, il va y avoir changement de la
séquence ADN de l'embryon• une cellule somatique
La réplication de l'ADN est une copie de 3 milliards de bases ce qui implique une forte probabilité de mutation.
Les mutations sont :• Spontanées, liées à des erreurs de réplication de l'ADN. On définit un taux de mutation par cellule et
par génération. Ces mutations sont physiologiques.• Induites (par la radio ou chimioinduites)
Il existe deux types de mutations géniques au niveau d'une population :
• Les mutations non-récurrentes : Si une mutation est unique ou très rare, la probabilité qu'elle disparaisse est très grande du fait des fluctuations d'échantillonnage. De plus, une mutation unique qui n'entraîne pas d'avantage sélectif pour le mutant ne peut pas produire d'effet permanent dans une population.
• Les mutations récurrentes : Si une mutation est récurrente, on parle de pression de mutation. C'est le même évènement qui va se produire chez des individus différents. C'est l'exemple de l'achondroplasie (nanisme) qui est une chondrodysplasie liée dans 95% des cas à une même mutation génique. Or 80% des cas sont des mutations de novo au niveau de ce même gène. Il s'agit d'un point chaud de mutation récurrente.
Soit une gène A :Dans le cas des mutations récurrentes, une fraction de l'allèle A1 va être transformée en allèle A2. Il y a un mécanisme inverse avec la transformation de A2 en A1μ = probabilité de mutation de A1 vers A2
μ ν A1 -----------> A2 A2 --------------> A1
p0 μp0 q0 νq0
La fraction de A2 transformée en A1 par la mutation sera de νq.La fraction de A1 transformée en A2 par la mutation sera de μp.
2/10

GENETIQUE MEDICALE – Génétique des populations
En fait, n (ν) est habituellement beaucoup moins fréquent que m (μ). Donc, l'allèle A1 devrait tendre à diminuer au profit de A2 (p diminue au profit de q). Pour maintenir l'équilibre, il y a donc un autre mécanisme, la sélection. Il y a un taux remarquablement stable des fréquences alléliques dans la population. En effet, le taux de mutation va être contrecarré par la sélection.
II. Sélection
On parle de sélection naturelle lorsque différents génotypes ne sont pas également viables et féconds. Les malades ont moins d'enfants que la population générale, voire pas d'enfants.S'il y a différence de fécondité entre les génotypes, il y a sélection.
A chaque génotype, on peut associer un coefficient s ou coefficient de sélection, compris entre 0 et 1.
Dans une maladie létale à la naissance ou au bout de quelques mois ou génétiquement létale (les individus peuvent survivre mais ne se reproduisent pas). C'est le cas de la dystrophie musculaire de Duchenne pour laquelle les malades ont une espérance de vie de 20 à 30 ans mais n'ont pas d'enfants. Dans ce cas :
s = 1
La valeur adaptative (f) d'un génotype est définie comme son efficacité à produire des descendants. Cette valeur adaptative est mesurée en valeur relative, 1 symbolisant la valeur adaptative du génotype optimum.
s = 1 – f
Dans une maladie génétiquement létale, f = 0 car s = 1
Calcul de la valeur adaptative : ( les chiffres ne sont pas à apprendre)
Dans le cas de la maladie de Becker : maladie qui commence vers l'âge de 15-20 ans et entraine la perte de la capacité à marcher vers 40 ans. Les malades ont moins de descendants que la population générale.Sur une période de 100 années fertiles, il y a eu 4,959 descendants pour les Becker contre 7,418 pour leurs frères germains. Ils ont 30% d'enfants en moins par rapport aux autres.Le coefficient de reproduction (ou valeur adaptative) est donc de 0,67.
Dans le cas de la neurofibromatose (taches café au lait, tumeur du nerf optique), ce coefficient est de 0,50.Dans le cas de la polykystose rénale, ce coefficient est de 0,80.
III. Dérive génétique
Dans l'équilibre de Hardy-Weinberg, la population doit être de dimension infinie.
Tous les individus participent de manière égale à la génération suivante. Les individus hétérozygotes Aa ont un risque sur 2 de transmettre leur mutation dans le cas d'une maladie dominante. Mais avec les lois du hasard, un individu peut très bien avoir 7 enfants sains ou 7 enfants malades.
Dans tous les cas il y a équilibre au niveau populationnel.
Dans les grandes populations (plusieurs centaines de milliers d'habitants), les variations (liées au hasard) du nombre d'enfants produits par des individus de génotypes différents, n'ont pas d'effet significatif sur la fréquence des gènes.
3/10

GENETIQUE MEDICALE – Génétique des populations
Dans les petites populations , ces variations peuvent avoir un effet considérable :
• Si un gène particulier n'est retrouvé que chez un petit nombre d'individus (1/500), si ces individus n'ont pas d'enfants ou, que par chance (hasard), ces enfants n'héritent pas de ce gène, le gène en question va complètement disparaître de la population (éteint : fréquence = 0) et son allèle va devenir fixé (fréquence = 1)
• La part de la dérive génique aléatoire dépend de la taille de la population. Plus la population est petite, plus les variations liées au hasard ont une influence.
C. Causes du maintien des maladies génétiques dans les grandes populations (malgré la sélection)
Pourquoi malgré la sélection existe t-il un taux remarquablement stable de mutations ? Taux d'hétérozygotes dans la population = 1/25 pour la mucoviscidose par exemple.
I. L'équilibre mutation-sélection
Les nouvelles mutations sont compensées par la sélection. Si les individus malades n'ont pas d'enfants, la perte des allèles non transmis est compensée par les mutations de novo. Il est certain qu'il y a un taux de mutation qui va compenser cette faible fécondité.
a) Dans les maladies dominantes
La valeur adaptative est comprise entre 0 (maladie génétiquement létale) et 1 (les individus ont une fertilité normale).
La proportion d'allèles éliminés, c'est à dire qui ne sont pas transmis à la génération suivante, est compensée par l'apparition de nouvelles mutations (de novo).
Pour une maladie génétiquement létale (f=0), les individus atteints n'ont jamais de descendants et tous les nouveaux cas résultent de nouvelles mutations.
4/10

GENETIQUE MEDICALE – Génétique des populations
AA Aa aaExeptionnel Malades Sains
2pq éliminés à chaque génération Nouvelles mutations
Lorsque la fertilité est diminuée (à cause de la barrière sociale par exemple), un certain nombre de cas sont liés à de nouvelles mutations.
AA Aa aaExeptionnel Malades Sains
Nouvelles mutations
Exemple : Les individus atteints d'achondroplasie ont environ 1 cinquième d'enfants par rapport à la population générale. Leur fertilité (valeur adaptative) est donc de 0,20. De plus, 80% des achondroplastes naissent de parents normaux et sont la conséquence de nouvelles mutations.Il existe donc un équilibre entre mutations et sélection qui permet le maintien des fréquences alléliques stables dans la population générale.
Nanisme tanatophore : mutation du FGFR3 (Fibroblast growth factor receptor 3) entrainant une forme tellement grave de chondrodysplasie que les enfants meurent à la naissance.
Dans les pathologies génétiques, il y a beaucoup plus de mutations de novo accidentelles que de mutations transmises.
b) Dans une maladie récessive liée au chromosome X génétiquement létale (ex : dystrophie musculaire de Duchenne)
Les mâles atteints (fréquence = q) n'ont pas de descendance.Seuls les gènes présents chez les femmes hétérozygotes (ou conductrices) (fréquence = 2q) sont transmis à la génération suivante.
Aa aConductrices Malades
2pq q
Nouvelles mutations
Ainsi, 1/3 des gènes mutés sont perdus à chaque génération et sont remplacés par des mutations de novo.Donc 1/3 des cas sont dus à des nouvelles mutations.Les cas sporadiques s'expliquent par le fait qu'il y a 2/3 de chances que la mère soit hétérozygote.
c) Age paternel et mutations
L'age paternel va favoriser les mutations dominantes de novo.Certaines mutations de novo ont une origine paternelle exclusive.
Pour beaucoup d'entre elles, l'âge moyen des pères à la naissance est significativement plus élevé que l'âge paternel moyen dans la population générale.
5/10

GENETIQUE MEDICALE – Génétique des populations
Explication classique : Les cellules souches donnant naissance aux spermatozoïdes continuent à se diviser toute la vie, favorisant ainsi l'accumulation progressive d'erreurs de réplication dans les gonades masculines. Le nombre d'ovocytes est fixé à la naissance mais pour un homme la division cellulaire dans la gonade continue tout au long de la vie.
Explication « moderne »: Certaines mutations confèrent aux spermatogonies un avantage sélectif, favorisant la sélection clonale.
Toutes les mutations d'origine paternelle exclusives sont des mutations qui, quand elles se produisent au niveau germinal donnent une malformation et une déficience intellectuelle alors qu'au niveau somatique, elles donnent des cancers.
Au niveau d'une spermatogonies, il va y avoir une prolifération des spermatogonies atteintes de cette mutation par rapport aux autres en raison d'un avantage sélectif, ce qui explique que ces maladies soient majoritairement d'origine paternelle.
Dans les maladies fréquentes (mucoviscidose), si les individus malades n'ont pas d'enfants, il devrait y avoir une diminution des mutations dans la population générale. Or ce n'est pas le cas en raison de l'avantage sélectif des hétérozygotes.Phénomène démontré pour les mutations des gènes FGFR (Aper, Crouzon, achondroplasie...), mutations retrouvées dans certains cancers.
II. L'avantage sélectif des hétérozygotes
Maladies récessives fréquentes dans leur génotype.Pas de mutations de novo.
Les hétérozygotes ont une capacité supérieure de défense vis à vis de maladies par rapport aux homozygotes (exemple de la drépanocytose).
Les hétérozygotes résistent plus aux maladies donc ont une fertilité supérieure à celle des homozygotes normaux et transmettent donc à la génération suivante un excès d'allèles mutés.
AA Aa aa Sains Hétérozygotes Malades
q² éliminé à chaque génération
Exemples :
Drépanocytose : Maladie très fréquente en Afrique, très grave. Cependant, les hétérozygotes ont une résistance accrue au paludisme. Ils ont une forme de globules rouges qui rend la pénétration du plasmodium difficile.
Mucoviscidose : Les hétérozygotes pour la mutation Fdel508 seraient plus résistants au choléra et autres maladies infectieuses intestinales (épidémies de choléra jusqu'au Moyen-Age). Les individus hétérozygotes pour la mucoviscidose étaient plus résistants et se sont donc plus développés que les homozygotes.
6/10

GENETIQUE MEDICALE – Génétique des populations
D. Fréquence élevée de certaines maladies très rares dans de petites populations : L'effet de fondateur
L'effet de fondateur : Lorsqu'une nouvelle population est créée par essaimage à partir d'une population d'origine, si l'un des fondateurs de ce nouveau groupe est porteur d'un allèle rare, celui-ci peut, sous l'effet de la dérive génétique, être fixé à une fréquence relativement élevée. Ceci est à la base de l'effet fondateur.
Chez les Bédouins (nomades de Palestine), il y a une fréquence d'hétérozygotes pour le CIPA (maladie récessive causée par une mutation proche de celle impliquée dans la mucoviscidose) plus importante car l'hétérozygotie leur donne une résistance accrue vis à vis des agents infectieux. Cependant ca ne ressemble pas vraiment à un effet fondateur mais plus à une dérive génétique car il y a plusieurs mutations différentes. Les enfants sont des hétérozygotes composites, ce qui leur confère une résistance particulière.
Chez les Amish, très petite population, isolat religieux d'origine européenne (Jura et Suisse) qui s'est fixé en Pennsylvanie, le syndrome d'Ellis Van Creveld, récessif autosomique, est très fréquent (q = 0,07), alors qu'il est extrêmement rare dans la populations générale. 5% de la population Amish est hétérozygote pour cette mutation.
E. Consanguinité
Définition : Est dit consanguin l'enfant issu de 2 sujets apparentés. Par extension l'épithète peut concerner une union mais pas les parents.
Deux sujets sont apparentés lorsqu'ils ont au moins un ancêtre commun véritable. Certains gènes qu'ils portent viennent donc de la réplication d'un gène de cet ancêtre et sont dit identiques.Elle favorise le fait d'être homozygote pour une mutation.
Coefficient de consanguinité : C'est la probabilité que chez un individu diploïde pris au hasard, deux allèles soient identiques par descendance mendélienne, c'est à dire dérivent d'un allèle ancestral unique.
En l'absence de mutation nouvelle : f (ou ϕ ) = coefficient de parenté des deux parents = coefficient de consanguinité.
7/10
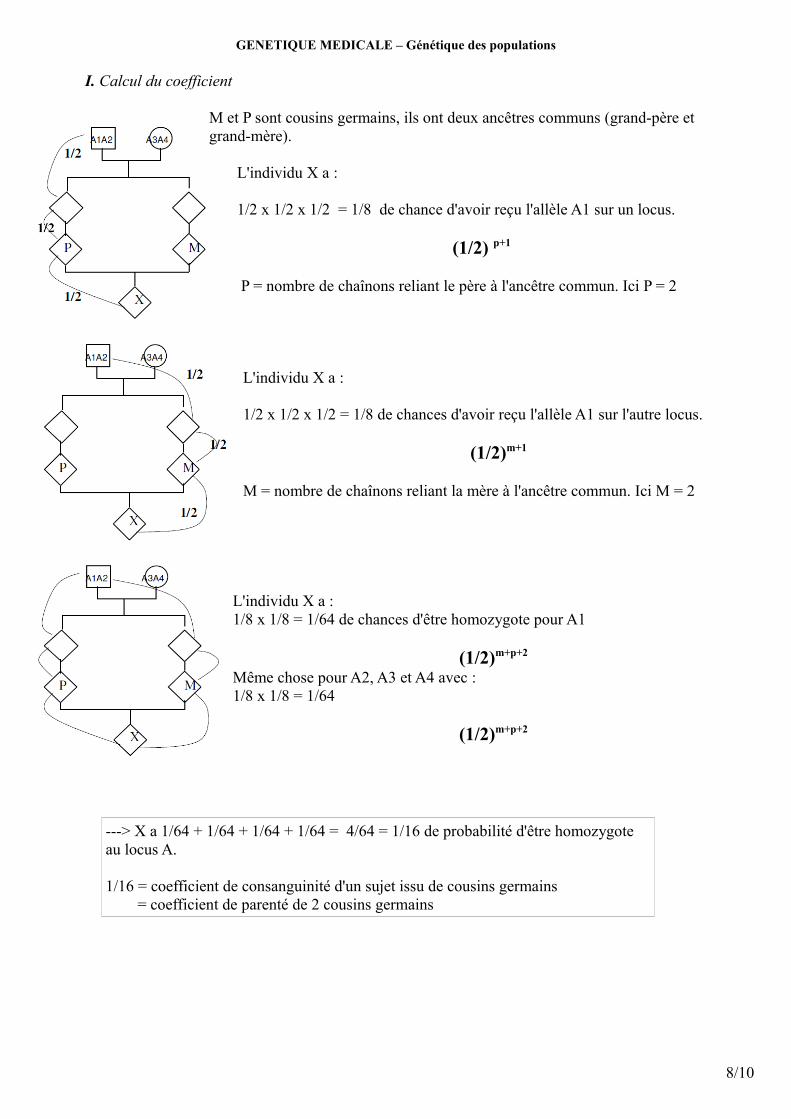
GENETIQUE MEDICALE – Génétique des populations
I. Calcul du coefficient
M et P sont cousins germains, ils ont deux ancêtres communs (grand-père et grand-mère).
L'individu X a : 1/2 x 1/2 x 1/2 = 1/8 de chance d'avoir reçu l'allèle A1 sur un locus.
(1/2) p+1
P = nombre de chaînons reliant le père à l'ancêtre commun. Ici P = 2
L'individu X a :
1/2 x 1/2 x 1/2 = 1/8 de chances d'avoir reçu l'allèle A1 sur l'autre locus.
(1/2)m+1
M = nombre de chaînons reliant la mère à l'ancêtre commun. Ici M = 2
L'individu X a :1/8 x 1/8 = 1/64 de chances d'être homozygote pour A1
(1/2)m+p+2
Même chose pour A2, A3 et A4 avec : 1/8 x 1/8 = 1/64
(1/2)m+p+2
---> X a 1/64 + 1/64 + 1/64 + 1/64 = 4/64 = 1/16 de probabilité d'être homozygote au locus A.
1/16 = coefficient de consanguinité d'un sujet issu de cousins germains = coefficient de parenté de 2 cousins germains
8/10

GENETIQUE MEDICALE – Génétique des populations
Pour l'ancêtre A1A2 (arrière grand-père) : L'individu X a 1/64 + 1/64 = 1/32 de chances d'être homozygote à ce locus.
(1/2)m+p+1
Même chose pour l'ancêtre A3A4 (arrière grand-mère) :1/8 x 1/8 = 1/64 pour A31/8 x 1/8 = 1/64 pour A41/64 + 1/64 = 1/32 de chances d'être homozygote à ce locus
(1/2)m+p+1
Pour chaque allèle, X a 1/2m+p+2 de chances d'être homozygote.Pour chaque ancêtre, X a 1/2m+p+1 (=2 x 1/2m+p+2 ) de chances d'être homozygote à ce locus.
ϕ = Σ 1/2m+p+1
Σ = somme (on calcule 1/2m+p+1 pour chaque ancêtre).
Coefficients de consanguinité :• frère-sœur = 1/4 • oncle-nièce = 1/8• cousins germains = 1/16• cousins issus de germains = 1/64
II. Conséquence de la consanguinité sur un individu
Le risque pour un individu consanguin d'être homozygote pour une maladie de fréquence q2 (risque dans la population générale d'être atteint d'une maladie), c'est :
• le risque qu'il soit homozygote du fait de sa consanguinité = ϕ.q+
• le risque qu'il soit homozygote du fait du hasard = (1 – ϕ).q²
Donc le risque qu'il soit homozygote est := ϕq + (1 - ϕ).q²= ϕq + q² - ϕq= q² - ϕq² + ϕq= q² + ϕq(1 – q)= q2 + ϕ pq
C'est la probabilité d'avoir un enfant homozygote dans un mariage consanguin. Cela augmente donc le risque d'avoir une maladie récessive.En général q est très petit donc p est proche de 1.
Exemple entre des cousins germains dans le cas de la phénylcétonurie :q² = 1/10 000q = 1/100
R (risque relatif) = 1/10 000 + 1/16 x 1/100 = 1/10 000 + 1/1600 = 1/1600
9/10

GENETIQUE MEDICALE – Génétique des populations
Risque relatif d'avoir un enfant malade dans un couple consanguin par rapport à un couple non consanguin :
= risque lié à la consanguinité
risque de la population générale
Pour la phénylcétonurie chez des cousins germains : 1 /1600
1 /10000= 6
Si la fréqeunce est de 1/1000000 → 1/100000 + 1/16 x 1/1000 = 1/16000Le RR est alors de environ 60
Le risque relatif est d'autant plus important que la maladie est rare. On trouve une plus grande proportion d'individus apparentés parmi les parents d'enfants atteints de maladies récessives.
III. Les conséquences de la consanguinité à l'échelle d'une population
Fi = coefficient moyen de parenté dans un population non panmictique (consanguine)Fi = moyenne des coefficients de parenté des différents couples de la génération i
Soit un locus di-allélique avec les allèles A1 et A2, de fréquences respectives p et q chez les parents. Soit F le coefficient moyen de parenté à la génération des parents.
A la génération suivante :
f(A1A1) = Fp + (1-F)p² soit p2 + Fp (1-p)f(A2A2) = Fq + (1-F)q² soit q2 + Fq(1-q)f(A1A2) = Fpq + (1-F)2pq soit 2pq – 2Fpq
Il y a donc un accroissement de la fréquence des homozygotes, et une diminution de la fréquence des hétérozygotes (fréquences génotypiques). Dans une population consanguine, il y a moins d'hétérozygotes et plus d'homozygotes.La consanguinité ne change ni p ni q mais elle change la répartition des génotypes.
La consanguinité favorise donc les maladies récessives autosomiques.
Cependant, elle n'a pas d'effet sur les anomalies chromosomiques (amniosynthèse inutile chez la femme enceinte), les maladies dominantes et les maladies liées au chromosome X.
CR : selon la prof, ces 2 dernières lignes sont les seules à retenir pour notre pratique de médecin... mais pas pour l'examen où il faudra tout savoir !
10/10


















