
Evaluation de l ˇactivitØ sulfhydryl oxydase/quelescine Q6 ... · Evaluation de l ˇactivitØ...
84
République Algérienne Démocratique et Populaire Ministère de l’Enseignement Supérieur et de la Recherche Scientifique UNIVERSITE D’ORAN FACULTE DES SCIENCES DEPARTEMENT DE BIOLOGIE Mémoire présenté en vue de l’obtention du Diplôme de Magister en Biologie Animale. Option : Embryogenèse et Oncogenèse Evaluation de l’activité sulfhydryl oxydase/quelescine Q6 (SQX/Q6) humaine dans les lignées cellulaires du cancer broncho-pulmonaire. Présenté par : Mademoiselle SEDDIKI Lamia Soutenue le : Devant le Jury composé de : Pr. EL KEBIR Fatima Zohra Président Université D’Oran Dr. SAHRAOUI Tewfik Directeur Université D’Oran Pr. SLIMANI Miloud Examinateur Université D’Oran Pr. AOUES Abdelkader Examinateur Université D’Oran Année Universitaire 2008-2009
Transcript of Evaluation de l ˇactivitØ sulfhydryl oxydase/quelescine Q6 ... · Evaluation de l ˇactivitØ...

République Algérienne Démocratique et PopulaireMinistère de l’Enseignement Supérieur et de la Recherche Scientifique
UNIVERSITE D’ORAN
FACULTE DES SCIENCES DEPARTEMENT DE BIOLOGIE
Mémoire présenté en vue de l’obtention du Diplôme de Magister en BiologieAnimale.
Option : Embryogenèse et Oncogenèse
Evaluation de l’activité sulfhydryl oxydase/quelescine Q6(SQX/Q6) humaine dans les lignées cellulaires du cancer
broncho-pulmonaire.
Présenté par :
Mademoiselle SEDDIKI Lamia
Soutenue le :
Devant le Jury composé de :
Pr. EL KEBIR Fatima Zohra Président Université D’Oran
Dr. SAHRAOUI Tewfik Directeur Université D’Oran
Pr. SLIMANI Miloud Examinateur Université D’Oran
Pr. AOUES Abdelkader Examinateur Université D’Oran
Année Universitaire 2008-2009

1
Remerciements
Je remercie Dr Sahraoui Toufik pour m’avoir encadrée tout le long de ce travail et pour
le temps qu’il m’a consacré pour réussir ce stage.
Je tiens à remercie tout les membres de jurys qui ont accepté de juger mon travail.
Je tiens à remercie vivement tout les membres de ma famille de leur présence, de leur
soutien :
Mon père, Ma mère et tous mes frères et sœurs.
Et enfin je remercie toute personne qui m’a aidé de loin ou de près dans la réalisation de
ce travail

2
SUMMARY
The broncho-pulmonary cancer is a major problem of public health because of its prognosis and its
more frequency, from where tobacco and the pollution are the first reasons. Our work carried on a
functional survey of the quiescin– sulfhydryl oxidize/ (SOx/Q6). The results of the epidemiological
Oran region shows that tobacco is the leading cause of development of lung cancer in this region, plus
the majority of patients present with advanced disease. The quiescin– sulfhydryl oxidase is the
enzymes that catalyze the formations of the bridges disulfide. Their secretion has been studied
according to the cellular formation. While using the DTT as substrate, we valued oxydative activity in
the supernageant of human lung fibroblasts (WI38) and of lung the epithelial tumoral cells lines non–
small– cells. The analyses of western - blot shows two types of SOx/Q6 a long and the short one at the
WI38. However, the analyses of western - blot of the middle with 10% FBS show the presence of the
bovine SOx/Q6, it is confirmed by the oxydative activity.
On the other hand, the results show that the secretion of the SOx/Q6 varies according to the tumoral
cells lines either : i) a secretion that increases at the time of the proliferation, ii)a secretion that
increases at the time of the confluence, iii) secretion that increases the after - confluence.
Keywords: Sulfyhydryl oxydase. Human quiescin Q6. Oxidative activity. Secretion. Lung cancer.

3
RESUME
Le cancer broncho-pulmonaire est un problème majeur de santé publique en raison de son pronostic
et de sa grande fréquence, d’où le tabac et la pollution sont les premières causes. Notre travail a porté
en premiers sur l’étude épidémiologique de la région oranaise et en second sur l’étude fonctionnelle de
la sulfhydryl oxydase/quiescine Q6 (SOx/Q6). L’ensemble des résultats épidémiologique de la région
oranaise montre que le tabac est la première cause de développement du cancer broncho-pulmonaire
dans la région, de plus la majorité des patients se présentent au stade avancé de la maladie.
Les sulfhydryls oxydases/ quiescine Q6 sont des enzymes qui catalysent les formations des ponts
disulfure. Leur sécrétion a été étudiée en fonction de la formation cellulaire. En utilisant le DTT
comme substrat, nous avons évalué l’activité oxydasique sécrétée dans les surnageants de fibroblastes
pulmonaire (WI38) et des lignées cellulaires tumorales épithéliales non à petites cellules. Les analyses
de western–blot montrent deux types de SOx/Q6 une longue et l’autre courte chez les WI38.
Cependant, les analyses de western –blot du milieu avec 10% SVF montrent la présence de la SOx/Q6
bovine, cela est confirmé par l’activité oxydasique.
D’autre part, les résultats montrent que la sécrétion de la SOx/Q6 varie en fonction des lignées
cellulaires tumorales soit une sécrétion qui augmente au moment de la prolifération, soit une sécrétion
qui augmente au moment de la confluence, soit sécrétion qui augmente l’après–confluence.
Mot clé: Sulfhydryl oxydase. Quiescine Q6. Activité oxydasique. Sécrétion. Cancer broncho-
pulmonaire

4
Sommaire INTRODUCTION .................................................................................................................................... 9
CHAPITRE I ETUDE BIBLIOGRAPHIQUE ....................................................................................... 11
I.1 Anatomie et structure des poumons ................................................................................................... 12
I.1.1 Physiologie des poumons ........................................................................................................... 13
I. 1.1.1 Les échanges gazeux ........................................................................................................... 13
I.1.1.2. La ventilation ....................................................................................................................... 13
I.1.1.3 Les centres respiratoires ....................................................................................................... 13
I.1.1.4 Les voies nerveuses .............................................................................................................. 14
I.1.1.5 La mécanique ....................................................................................................................... 14
I.1.2 Embryologie ............................................................................................................................... 15
I.1.3 Histologie .................................................................................................................................... 18
I.1.3.1 Conduction de l’air .............................................................................................................. 19
I.1.4 Le poumon périphérique ............................................................................................................. 21
I.1.4.1 Zone de conduction .............................................................................................................. 21
I. 1.4.2 Zone de transition ................................................................................................................ 22
I.1.4.3 Zone d’échanges gazeux ...................................................................................................... 22
I.1.4.4 L’épithélium alvéolaire ........................................................................................................ 23
I.2.Etiologie et épidémiologie du cancer des poumons .......................................................................... 25
I.2.1 Définition du cancer ................................................................................................................... 25
I.2.2 Le cancer broncho-pulmonaire ................................................................................................... 26
I.2.3 Etiologie ...................................................................................................................................... 26
I.2.3.1 Les principales causes du cancer broncho-pulmonaire ........................................................ 26
I.2.4 Epidémiologie descriptive du cancer des poumons .................................................................... 31
I.2.4.1 Cancer des poumons en Algérie ........................................................................................... 31
I.2.4.2 Cancer des poumons en France ............................................................................................ 32
I.2.4.3. Cancer des poumons en Canada .......................................................................................... 32
I.2.5. Épidémiologie analytique .......................................................................................................... 33
I.3.Anatomie pathologique des cancers broncho-pulmonaire ................................................................. 35
I.3.1 Classification des cancers broncho-pulmonaire ......................................................................... 35
I.3.1.1 Cancer bronchique épidermoïde ........................................................................................... 35

5
I.3.1.2 Cancer bronchique à petites cellules .................................................................................... 35
I.3.1.3.Adénocarcinome bronchique est divisé en 4 sous-groupes histologiques ........................... 37
I.3.1.4. Epithéliome bronchique à grandes cellules ......................................................................... 38
I.3.1.5 Carcinomes adénosquameux ................................................................................................ 38
I.3.2 Diagnostic du cancer broncho-pulmonaire ................................................................................. 39
I.3.2.1. Les signes cliniques ............................................................................................................. 39
I.3.2.2. Signes radiologiques ............................................................................................................ 41
I.4. Introduction à la famille de la sulfhydryl oxydase ........................................................................... 42
I.4.1. La famille de la sulfhydryl oxydase ........................................................................................... 42
I.4.2. Structure de la sulfhydryl oxydase/Quiescine Q6 .................................................................. 45
CHAPITRE II MATERIELS ET METHODES ..................................................................................... 47
II.1 L’étude épidémiologique du cancer pulmonaire de la région oranaise ........................................ 48
II.2 Matériel biologique ...................................................................................................................... 49
II.2.1 Lignées cellulaires .................................................................................................................. 49
II.3. Préparation d’anticorps spécifiques dirigés contre la sulfhydryl oxydase de rat ........................ 49
II.3.1 Extraction des anticorps anti-sulfhydryl oxydase de rat à partir de jaune d’œuf de poule .... 49
II.3.2 Purification des anticorps spécifiques anti-sulfhydryl oxydase de rat par chromatographie
d’affinité .......................................................................................................................................... 50
II.3.3 Electrophorèse sur gel polyacrylamide 12% en conditions dénaturantes (SDS-PAGE) ....... 51
II.3.4 Western blot et révélation par chimioluminescence .............................................................. 51
II.4 Mesure de l’activité oxydasique des sulfhydryl oxydases ........................................................... 52
II.4.1 Principe de dosage de l’oxydation du DTT par le DTNB ...................................................... 52
II.4.2 Mesure de l’activité oxydasique des surnageants de culture ................................................. 53
II.4.3 Analyse de l’activité protéasique à cystéine .......................................................................... 54
II.5 Analyses statistiques ................................................................................................................. 54
RESULTATS .......................................................................................................................................... 55
DISCUSSION ......................................................................................................................................... 69
CONCLUSION ET PERSPECTIVES .................................................................................................. 75
REFERENCES BIBLIOGRAPHIQUES ................................................................................................ 77
Annexe ................................................................................................................................................... 82

6
Abréviations
ADN : Acide DésoxyriboNucléique
ARNm : Acide Ribonucléique Messager
ATCC : American Type Culture Collection
dNTP : désoxynucléosides 5’ triphosphate
DTNB : Acide 5,5’-dithiobis-2-nitrobenzoïque
DTT : Dithiothréitol
ERV : Essential for Respiration and Viability
IFN- : Interféron gamma
MEM : Minimum Essential Medium
Mr : Masse moléculaire relative
NADPH : Nicotinamide Adénine Dinucléotide Phosphate, forme réduite
pb : paire de base
PCR : Polymerase Chain Reaction
PDI : Protéine Disulfure Isomérase
RPMI 1640: Roswell Park Memorial Institute
rSOx: Sulfhydryl oxydase de rat
SOx/Q6: Sulfhydryl oxydase quiecine Q6
SDS-PAGE: Sodium dodecyl sulphate-polyacrylamide gel electrophoresis
SVF : Sérum du Veau Fœtal
TAE : Tris Acétate EDTA
TRX : Thiorédoxine
TNM : T : la taille de la tumeur, N : l’existence d’adénopathies satellites, M : métastases

7
Liste des figures
Figure 1 : Description anatomique des poumons
Figure 2 : Description anatomique des poumons et des bronches
Figure 3 : Schéma représente les modifications morphologiques des poumons en état
d’inspiration et d’expiration
Figure 4 : Une coupe microscopique d’un tissu pulmonaire périphérique.
Figure 5 : Représentation de la structure de la barrière alvéolaire.
Figure 6 : Poumon périphérique : zone d'échanges gazeux
Figure 7: Les étapes d’évolution d’une tumeur maligne.
Figure 8 : Dissection d'un poumon cancéreux.
Figure 9 : Effet multiplicateur du tabagisme et de l’exposition à l’amiante sur le risque relatif de
cancer du poumon.
Figure 10 : Schéma d’activation du récepteur EGFR par l’EGF
Figure 11 : Taux de mortalité due au cancer du poumon, hommes et femmes
Figure 12 : Biopsie d'un poumon souffrant de carcinome à petites cellules
Figure 13 : Adénocarcinome bronchique iconographie.
Figure 14 : Structures protéiques de la thiorédoxine, de la protéine disulfure isomérase, de la
sulfhydryl oxydase/Quiéscine Q6
Figure 15 : Schéma représentant l’activité de la sulfhydryl oxydase/QuiescineQ6 (SOx/Q6).
Figure 16 : Purification des anticorps IgY anti SOx/Q6 par chromatographie.
Figure 17 : Réaction d’oxydation du DTT catalysée par la sulfhydryl oxydase de rat.
Figure 18 : Réaction du DTNB avec les thiols libres.
Figure 19 : Répartition des cas recensés en fonction de la localisation du cancer pulmonaire.
Figure 20 : Répartition des cas étudiés en fonction de l’âge.
Figure 21 : La répartition des cas étudiés selon le stade d’évolution du cancer broncho-
pulmonaire.
Figure 22 : Répartition des cas en fonction du type histologique du cancer broncho-pulmonaire
les plus représentatifs de la région oranaise.
Figure 23 : Purification des IgY anti-rSOx spécifiques par chromatographie d’affinité.
Figure 24 : Analyse de la pureté des IgY spécifiques par affinité sur gel de polyacrylamide à
12%.
Figure 25 : Analyse par western blot du seuil de détection de rSOx par les IgY spécifiques.
Figure 26 : Analyse par Western blot de la reconnaissance de la sulfhydryl oxydase/quiescine Q6
humaine par les IgY spécifiques.

8
Figure 27: Analyse par Western blot de la présence de sulfhydryl oxydase dans le sérum de veau
fœtal.
Figure 28 : Activité oxydasique de la sulfhydryl oxydase de rat.
Figure 29 : Activité oxydasique du milieu de culture contenant le sérum de veau fœtal.
Figure 30 : Les activités oxydasiques corrigées des surnageants des lignées cellulaires
pulmonaires
Figure 31 : Les activités oxydasiques corrigées des surnageants des lignées cellulaires
pulmonaires.
Figure 32 : Comparaison des activités oxydasiques entre les lignées cellulaires et la WI38.
Figure 33 : Comparaison des activités oxydasiques entre les lignées cellulaires.
Figure 34 : Comparaison des activités oxydasiques entre les lignées cellulaires.

9
INTRODUCTION
Le cancer broncho-pulmonaire est la première cause de mortalité par le cancer dans le monde, dont
15 000 en Algérie, 22 000 en France et 19 000 au Canada. En Algérie les chiffres s’avèrent
inquiétants. La première cause en est le tabagisme.
Les études ont montré une relation positive et fortement significative entre le cancer broncho-
pulmonaire et le tabagisme. La fumée de cigarette est constituée de 4 000 produits chimiques, dont la
plupart sont cancérigènes. L’exposition permanente des cellules des poumons (les pneumocytes) à la
fumée engendre des changements moléculaires irréversibles tel que le dérèglement des systèmes
détoxification et antioxydants.
La mortalité causée par le cancer du poumon est trois fois plus élevée chez les femmes que chez les
hommes. Cependant la mortalité chez les hommes a stagné, ou a régressé légèrement par apport aux
femmes [1]. La gravité de la maladie est due au diagnostique tardif au stade métastatique. La majorité
(80 %) des cas de cancer broncho-pulmonaire correspond à des tumeurs non à petites cellules.
Par ailleurs, les connaissances épidémiologiques étiologiques, et histologiques se sont affinées. Les
données biologiques concernant la cancérogenèse, les mécanismes d’invasion locale et de métastases à
distance ont eu leurs retombées dans le domaine des cancers intra thoraciques.
Environs 80% des cas de cancer broncho-pulmonaire correspondent à des tumeurs non à petites
cellules (NSCLC). On distingue en particulier l’adénocarcinome et le carcinome épidermoïde qui sont
les formes les plus représentées. Les profils d’expression géniques de ces deux types majeurs de
cancer font apparaître des changements moléculaires avec en particulier une surexpression de gènes de
détoxification ou antioxydants [2].
Les sulfhydryls oxydases SOx sont des enzymes qui catalysent la réactiond’oxydation de résidus thiol
par l’oxygène moléculaire pour former des ponts disulfures. Les substrats des ceux type d’enzyme
peuvent être deux protéines ou deux péptides et dans ce cas le pont disulfure permet leur dimérisation.
De plus, les ponts disulfures peuvent être formé à partir de résidus cystéyls d’une et permet son
repliement.
La SOx/Q6 est impliquée dans le cancer broncho-pulmonaire non à petites cellules. Dans ce contexte,
l’objectif de notre travail, d’estimer l’expression de SOx/Q6 dans les lignées tumorales pulmonaires
modèles par analyse du niveau de sécrétion de SOx/Q6 humaine à différents stades de prolifération par
mesure de l’activité oxydasique.
Dans ce contexte, notre travail est réparti en deux. La première partie de ce mémoire est la partie
bibliographique composée de trois chapitres, le premier chapitre est consacré aux généralités sur les
poumons et les bronches du point de vu embryologique, anatomique et physiologique, le deuxième
chapitre traite les phénomènes lésionnels bénins et malins, et un dernier chapitre traite l’étiologie et
épidémiologie du cancer des poumons.

10
La deuxième partie, la partie paratique est consacrée au travail pratique, l’evaluation de l’activité
enzymatique de la SOx/Q6, puis une exposition des resultats suivie de la discussion. Et nous terminons
par une conclusion générale et les perspectives.

11
CHAPITRE I ETUDE BIBLIOGRAPHIQUE

12
I.1 Anatomie et structure des poumons
Les deux poumons sont situés dans la cage thoracique et séparés l'un de l'autre par un
espace, le médiastin (où se trouvent le cœur, l'œsophage et les gros vaisseaux).
Chaque poumon a grossièrement une forme conique, avec
un apex qui remonte jusqu'à la base du cou,
une face latérale convexe qui s'applique contre la paroi thoracique,
une base concave qui repose sur le diaphragme,
et une face interne plaquée contre le médiastin qui reçoit en sa partie médiane, le hile, la
bronche souche, branche de division de la trachée, les vaisseaux sanguins et les nerfs.
Chaque poumon est entouré d'une enveloppe, la plèvre, composée de deux feuillets étroitement
appliqués l'un sur l'autre et se réunissant au niveau du hile, en formant une sorte de sac sans
ouverture autour d'une cavité virtuelle, la cavité pleurale. Le feuillet externe de la plèvre étant
lui-même accolé à la paroi thoracique, la cavité pleurale qui permet le glissement des poumons
lors de chaque mouvement respiratoire. [3]
Les poumons sont anatomiquement séparés en lobes par des " scissures ", visibles sur leur face
externe.
Le poumon droit compte trois lobes, supérieur, inférieur et moyen,
Le poumon gauche deux seulement, supérieur et inférieur.
Figure 1 : Description anatomique des poumons
A : Poumons dans la cage thoracique. 1) lobe supérieur droit ; 2) lobe moyen droit ; 3) lobe
inférieur droit ; 4) lobe supérieur gauche ; 5) lobe inférieur gauche. B : ensemble de poumons :
(1) trachée, (2) bronches souches (3) [Encarta 2008]

13
Les artères pulmonaires, issues du cœur droit et contenant du sang pauvre en oxygène mais
chargé en gaz carbonique, pénètrent dans les poumons au niveau du hile comme les bronches
souches. Elles se divisent en artères lobaires puis segmentaires et, de façon parallèle à la
ramification bronchique, par divisions successives, elles se terminent à la surface des alvéoles
en un fin réseau de capillaires. Le sang libère le gaz carbonique et se charge d'oxygène, le
transport gazeux se faisant à travers la barrière alvéolo-capillaire. Au-delà, les capillaires se
réunissent à nouveau pour former le réseau veineux pulmonaire, qui aboutit aux veines
pulmonaires, lesquelles se jettent dans l'oreillette gauche. [3]
I.1.1 Physiologie des poumons
I. 1.1.1 Les échanges gazeux
L’oxygène et le gaz carbonique se déplacent entre l’aire et le sang par simple diffusion, c’est à-
dire d’une zone de haute pression à une zone de faible pression partielle. La loi de Fick établit
que la quantité de gaz qui traverse une couche de tissu est proportionnelle à la surface de cette
couche et inversement proportionnelle à son épaisseur. La barrière gaz-sang est extrêmement
mince, et sa surface est comprise entre 50 et 100 m². Elle est par conséquent bien adaptée à sa
fonction d’échange gazeux.
I.1.1.2. La ventilation
C'est un processus qui va permettre les échanges gazeux entre l'air et les alvéoles pulmonaires.
La réserve d'oxygène dans les poumons est à peine suffisante pour couvrir les besoins de
l'organisme pendant deux minutes. Au bout de deux minutes, on va avoir des lésions plus ou
moins importantes de certains organes et surtout au cerveau. Il faut renouveler l'air en
permanence contenu dans les poumons. Cette fonction de ventilation doit se faire sans arrêt
tout au long de la vie, elle est vitale. La fonction de ventilation est dite automatique. Elle est
sous la dépendance de cellules nerveuses appelées centres respiratoires.
I.1.1.3 Les centres respiratoires
Ils sont situés au niveau du bulbe rachidien et de la protubérance annulaire. On distingue les
centres inspiratoires et les centres expiratoires. Ces deux centres sont sous la dépendance d'un
système régulateur. Les cellules des centres inspiratoires ont une activité rythmique et adresse
régulièrement des incitations motrices aux nerfs des muscles inspiratoires. Ces cellules sont
aussi sous la dépendance de différents facteurs qui contrôlent, régularisent et adoptent l'activité
des centres. Facteurs: émotions, vomissements, volonté, hyperthermie, effort.
14
I.1.1.4 Les voies nerveuses
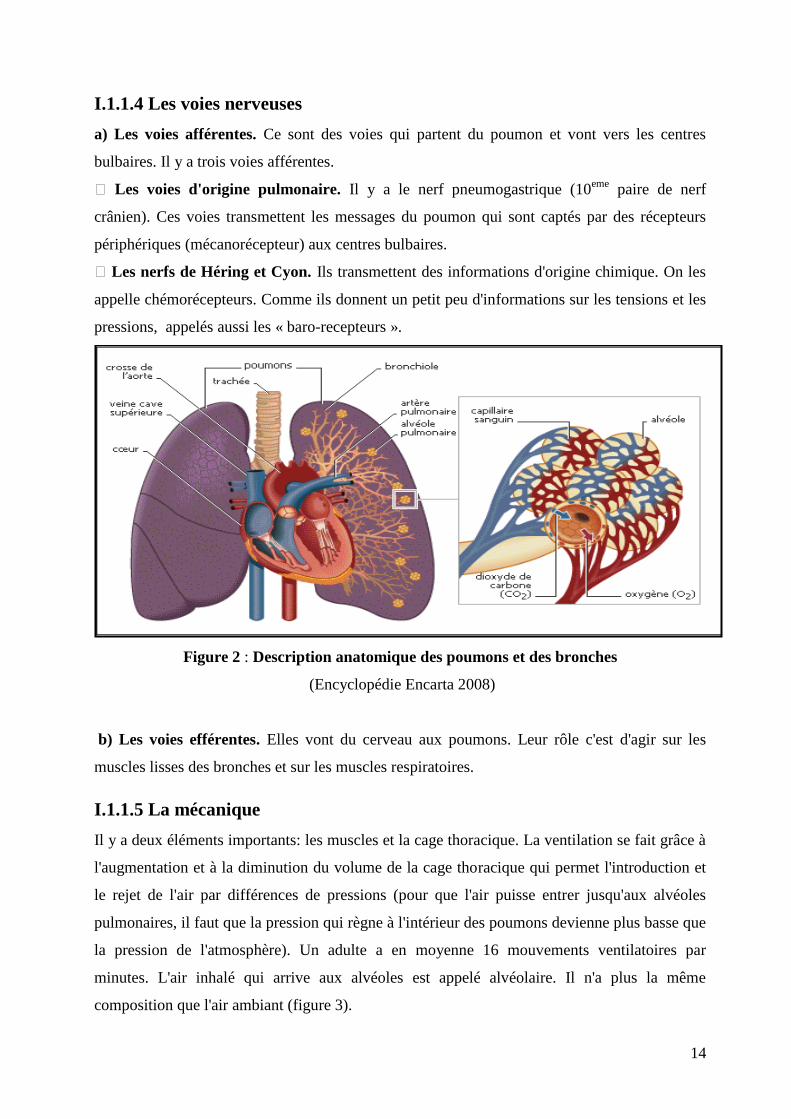
a) Les voies afférentes. Ce sont des voies qui partent du poumon et vont vers les centres
bulbaires. Il y a trois voies afférentes.
Les voies d'origine pulmonaire. Il y a le nerf pneumogastrique (10eme
paire de nerf
crânien). Ces voies transmettent les messages du poumon qui sont captés par des récepteurs
périphériques (mécanorécepteur) aux centres bulbaires.
Les nerfs de Héring et Cyon. Ils transmettent des informations d'origine chimique. On les
appelle chémorécepteurs. Comme ils donnent un petit peu d'informations sur les tensions et les
pressions, appelés aussi les « baro-recepteurs ».
Figure 2 : Description anatomique des poumons et des bronches
(Encyclopédie Encarta 2008)
b) Les voies efférentes. Elles vont du cerveau aux poumons. Leur rôle c'est d'agir sur les
muscles lisses des bronches et sur les muscles respiratoires.
I.1.1.5 La mécanique
Il y a deux éléments importants: les muscles et la cage thoracique. La ventilation se fait grâce à
l'augmentation et à la diminution du volume de la cage thoracique qui permet l'introduction et
le rejet de l'air par différences de pressions (pour que l'air puisse entrer jusqu'aux alvéoles
pulmonaires, il faut que la pression qui règne à l'intérieur des poumons devienne plus basse que
la pression de l'atmosphère). Un adulte a en moyenne 16 mouvements ventilatoires par
minutes. L'air inhalé qui arrive aux alvéoles est appelé alvéolaire. Il n'a plus la même
composition que l'air ambiant (figure 3).

15
Figure 3 : Schéma représente les modifications morphologiques des poumons en état
d’inspiration et d’expiration
I.1.2 Embryologie
Le poumon des vertébrés prend son origine embryologique dans une excroissance de
l'œsophage, la vessie natatoire, et est donc formé d'endoderme (doublé secondairement de
mésoderme pour la vascularisation). Le poumon des Oiseaux est très modifié formant une série
de tubes entre deux sacs aériens où l'air circule dans un seul sens (contrairement aux autres
poumons où l'air fait des mouvements de va-et-vient au cours d'un cycle inspiration/expiration).
Il existe également des « poumons » chez les invertébrés qui ne sont pas homologues des
poumons de vertébrés, puisqu'ils sont formés par invagination de l'ectoderme. On les trouve
chez les Chélicérates (araignée, scorpion) et chez les Gastéropodes Pulmonés (escargot) où
c'est la cavité palléale qui joue le rôle de poumon, communiquant avec l'extérieur par un petit
orifice appelé pneumostome. D'une manière générale, les poumons permettent une respiration
en milieu aérien tout en évitant la déshydratation [3].
- Embryologie des voies aériennes profondes
On se limitera à une étude succincte du développement embryonnaire des voies aériennes
profondes, le développement des voies aériennes supérieures ayant été inclus dans la mise en
place du splanchnocrâne.
- Organogénèse
Cette période s’étend de la fin de S3 (après la mise en place de l’embryon tridermique) jusqu’à
la fin du deuxième mois. Durant cette courte période, environ 95 % des organes sont
préformés. Au niveau de l’appareil respiratoire, cela se traduit par la mise en place des voies de
conduction [9].

16
- Arbre broncho-pulmonaire
Il dérive de l’entoblaste du tube digestif primitif antérieur. Dès J22 apparaît sur la face ventrale
du tube digestif primitif antérieur une évagination, en forme de gouttière, le diverticule
respiratoire ou bourgeon bronchique. Ce diverticule se développe en direction ventrale et
caudale tout en se divisant par dichotomie. Toutes les structures des épithéliales pulmonaires
dérivent de l’entobalste.
- Le tissu conjonctif
Il dérive naturellement du mésoblaste. Du fait des plicatures embryonnaires, le tube digestif
primitif est composé de deux tubes coaxiaux, l’entoblaste, central, entouré d’un tube de
mésoblaste. Ce mésoblaste périentoblastique est tout naturellement entraîné par le diverticule
respiratoire et sera à l’origine de toutes les structures conjonctives des voies aériennes
profondes :
Voies aériennes : chorion de la muqueuse, les cartilages
Mésenchyme pulmonaire : mésenchyme axial associé à l’arbre bronchique, les grandes
cloisons conjonctives intra-pulmonaire
Conjonctif du parenchyme : interstitium interalvéolaire
La plèvre provient des lames latérales du mésoblaste :
- Splanchnopleure : feuillet viscéral
- Somatopleure : feuillet pariétal
- La vascularisation
Le diverticule respiratoire entraîne aussi avec lui la vascularisation du tube digestif primitif.
Cette vascularisation, issue du système aortique dorsal (4ème et 6ème arcs aortiques), va
accompagner le développement de l’arbre bronchique, mais lui restera restreinte, préfigurant la
circulation bronchique (qui appartient à la grande circulation).
Ultérieurement, un deuxième réseau, issu du tube cardiaque, va se développer. Il va migrer tout
au long de l’arbre bronchique pour coloniser plus tard le parenchyme. Il constitue la circulation
pulmonaire ou petite circulation [9].
- Histogénèse
Cette période va de la fin de S8 (2ème mois) jusqu’à la naissance. Elle correspond à la période
foetale, caractérisée par une croissance pondérale et une maturation histologique qui se traduit
par une différenciation cellulaire, chaque cellule développant les fonctions spécifiques du tissu
auquel elle appartient.

17
Au niveau de l’appareil respiratoire, cette période correspond à l’achèvement de la mise en
place des voies de conduction et surtout par la différenciation de la zone d’échange.
- Arbre bronchique
La croissance, avec division dichotomique, se poursuit jusqu’à S16 ou l’on atteint le stade
bronchiole terminale (16ème génération de division). Cela correspond à un envahissement des
cavités pleurales, formées dès S5 par le septum transversum, les plis pleuro-péricardiques et les
membranes pleuro-péritonéales)
- Zone d’échange
De S16 à S18, chaque bronchiole terminale va donner naissance à deux ou plusieurs
bronchioles respiratoires.
A S28 (6ème mois), les cellules épithéliales des ramifications distales de cet arbre bronchique
s’aplatissent. Elles se différencient en pneumocytes (I et II) qui sont les cellules spécifiques de
l’épithélium alvéolaires. Ces portions distales constituent alors les bronches trapues,
précurseurs des futurs sacs alvéolaires. Cette différenciation commence au niveau de l’apex et
se propage jusqu’au niveau des bases (sens cranio-caudal). Un mois plus tard, les pneumocytes
II ont définitivement acquis la capacité de synthétiser et de sécréter le surfactant alvéolaire.
L’appareil respiratoire est désormais fonctionnel ; il ne restera plus, à la naissance, qu’à laisser
pénétrer l’air dans l’appareil respiratoire (1er cri), ce qui va déployer les sacs alvéolaires, grâce
au surfactant alvéolaire, et à résorber le peu de liquide qui les remplissait durant la vie in utero
[3].
- Réseau vasculaire pulmonaire :
Lors de la différenciation des bronches trapues, le réseau pulmonaire se densifie dans le
mésoblaste voisin, donnant un réseau capillaire très dense entourant chaque sac alvéolaire, à
S36.
Néanmoins, ce réseau n’acquière sa configuration définitive qu’après la naissance (au delà de 7
– 9 ans). Dans les parois inter-alvéolaires épaisses cohabitent deux réseaux capillaires
adjacents, un pour chaque alvéole. Au cours de la phase d’hypertrophie, l’étirement des parois
oblige les deux réseaux à fusionner pour donner le réseau terminal.
- Le tissu conjonctif
Le tissu conjonctif est définitivement structuré.
- Période post-natale
Cette période est caractérisée par la succession de trois phases : les deux premières
correspondent à une évolution permettant le passage progressif du poumon d’un nouveau-né à
un poumon adulte.

18
- Phase d’hyperplasie alvéolaire
Durant cette période, on va continuer à fabriquer du parenchyme pulmonaire, jusqu’à l’âge de 7
– 10 ans, pour accompagner la croissance staturo-pondérale de l’enfant (tableau 1).
Tableau 1 : nombre d’alvéoles et de bronchioles respiratoires [3].
Naissance Adulte
Nb d’Alvéoles 24 X 106 280 – 300 X 106
Nb de Bronchioles respiratoires 1,5 X 106 14 X 10
On va donc décupler le nombre d’éléments, avec une période de croissance rapide jusqu’à l’âge
de 5 ans où l’on est aux environs de 260 X 106 d’alvéoles (environ 90% du potentiel), suivi
d’une phase d’amortissement qui prend fin entre 8 et 10 ans.
- Phase d’hypertrophie alvéolaire
Le programme d’hyperplasie étant épuisé, la croissance pulmonaire sera complétée par une
hypertrophie alvéolaire.
-Phase de régression
Dès l’âge de 20 ans, on observe un début de bronchiolisation très progressive, qui va
s’accentuer sensiblement chez le sujet âgé.
I.1.3 Histologie
En histologie, l’appareil respiratoire est composé de trois types d’éléments :
- de l’air : particulièrement évident à l’observation de coupe histologique de poumon
- du sang, qui représente 50% du volume du parenchyme (Interstitium interalvéolaire)
- des tissus
Les tissus constitutifs du poumon sont composés uniquement d’épithéliums et de tissus
conjonctifs, une structure histologique relativement simple pour résoudre les contraintes
mécaniques qui lui sont imposées (Figure 4).

19
Figure 4 : Une coupe microscopique d’un tissu pulmonaire périphérique.
(Coloration hématéine éosine safran, grossissement x 100).
La structure de l’arbre bronchique est étroitement adaptée à sa fonction : conduction et
conditionnement de l’air [9].
I.1.3.1 Conduction de l’air
Les voies aériennes se divisent de façon dichotomique : chaque bronche mère donne 2
bronches filles. Il y a 23 générations de voie aérienne dont les 16 premières sont
"conductrices". Ainsi à la trachée succèdent 2 bronches souches, 5 lobaires, 20 segmentaires et
environ 3000 bronchioles lobulaires.
La structure histologique de la paroi des voies aériennes est très bien adaptée à la conduction
car elle permet d’associer rigidité, flexibilité et extensibilité. Elle est formée de différentes
couches dont la composition et la répartition varient de la trachée à la bronchiole afin de
s’adapter au mieux au diamètre des voies aériennes et à leur fonction.
La muqueuse tapisse la lumière bronchique. Elle est composée d’un épithélium de type
respiratoire reposant sur une membrane basale et d’un chorion conjonctif constitué de fibres
collagènes et élastiques, contenant des vaisseaux sanguins et lymphatiques. Les fibres
élastiques permettent une certaine flexibilité et un retour à la normale après dilatation des
bronches.
Entre le chorion et le cartilage s’insère un muscle lisse, absent au niveau de la trachée mais
présent sur tout l’arbre bronchique. Au niveau de la trachée, le muscle est situé à la partie
postérieure du mur cartilagineux. La contraction musculaire permet de réguler le flux aérien
lors de l’inspiration et de l’expiration.
Entre le muscle et le cartilage, la sous-muqueuse contient outre les trajets vasculaires et
lymphatiques, des glandes séromuqueuses s’ouvrant dans la lumière bronchique.

20
Enfin l’adventice (la couche la plus externe) est constituée d’un tissu conjonctivo-adipeux
lâche contenant vaisseaux et nerfs [9].
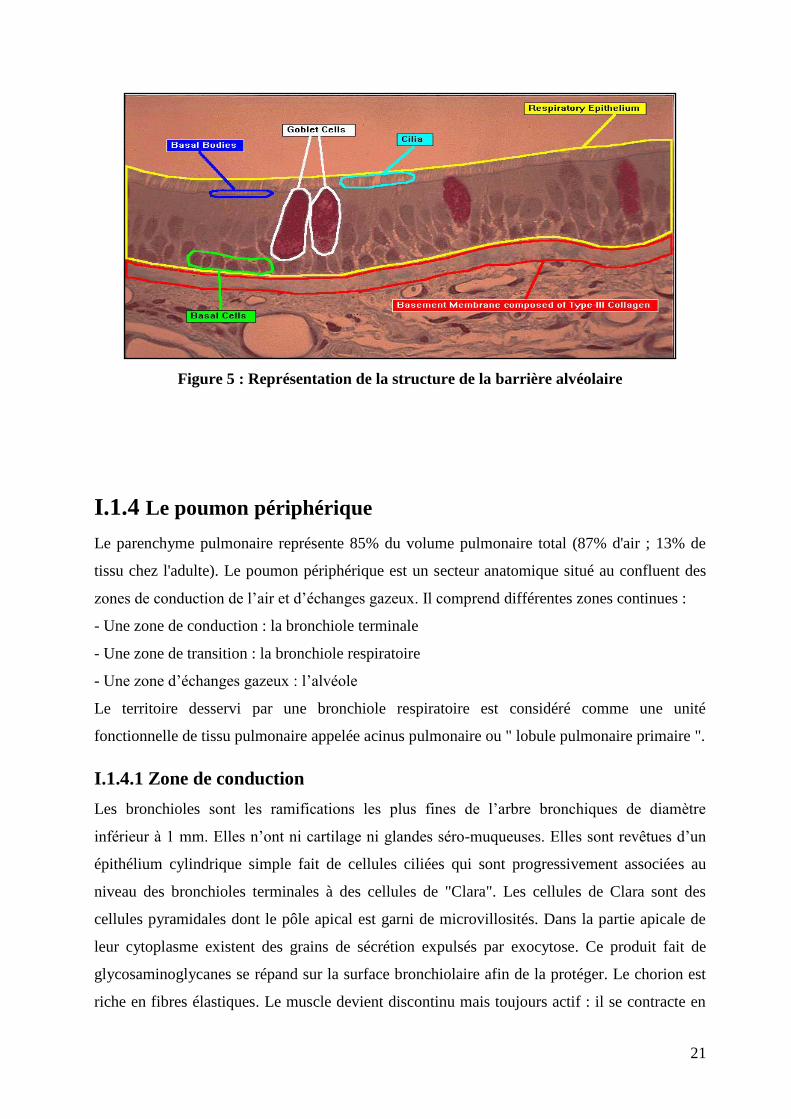
I.1.3. 2 Composition de l’épithélium respiratoire
L’épithélium respiratoire est composé de 5 types de cellules :
1-Les cellules ciliées sont les plus abondantes. Ces cellules ciliées jouent un rôle fondamental
dans l’épuration pulmonaire.
2-Les cellules caliciformes (1 pour environ 5 cellules ciliées) ont un noyau refoulé vers la base
par une vacuole supranucléaire contenant du mucigène qui participe, avec les secrétions des
glandes séro-muqueuses à la formation du mucus. Celui-ci permet d’emprisonner les impuretés
de l’air inspiré et d’absorber certains gaz (SO2, ozone…). Les cellules ciliées persistent dans
les bronches de petit calibre au delà des cellules caliciformes. Le nombre de cellules
caliciformes augmente avec celui des impuretés dans l’air inspiré. Le nouveau-né en est
dépourvu. A l’opposé, il existe une hyperplasie des cellules caliciformes chez les fumeurs.
3-Les cellules basales sont des petites cellules situées dans la partie profonde de l’épithélium
contre la membrane basale. Elles sont très importantes dans le renouvellement cellulaire car
elles sont capables de remplacer n’importe quel autre type de cellule bronchique.
4-Les cellules à bordures en brosse ne sont visibles qu’en microscopie électronique.
5-Les cellules chromaffines ou argentaffines sont peu nombreuses, visibles en microscopie
électronique ou sur coupe histologique après un immunomarquage à l’aide de marqueurs
neuro-endocrines (comme la chromogranine A ou la Synaptophysine). Ces cellules se groupent
pour former des corps neuro-épithéliaux : chémorécepteurs capables de détecter les variations
de teneur en O2 ou CO2 de l’air.
Le rôle des glandes séro-muqueuses est lui aussi important dans le conditionnement de l’air.
Celui du mucus est décrit ci-dessus. La sécrétion des glandes séreuses humidifie l’air
préalablement purifié par le mucus et les cils.
21
Figure 5 : Représentation de la structure de la barrière alvéolaire
I.1.4 Le poumon périphérique
Le parenchyme pulmonaire représente 85% du volume pulmonaire total (87% d'air ; 13% de
tissu chez l'adulte). Le poumon périphérique est un secteur anatomique situé au confluent des
zones de conduction de l’air et d’échanges gazeux. Il comprend différentes zones continues :
- Une zone de conduction : la bronchiole terminale
- Une zone de transition : la bronchiole respiratoire
- Une zone d’échanges gazeux : l’alvéole
Le territoire desservi par une bronchiole respiratoire est considéré comme une unité
fonctionnelle de tissu pulmonaire appelée acinus pulmonaire ou " lobule pulmonaire primaire ".
I.1.4.1 Zone de conduction
Les bronchioles sont les ramifications les plus fines de l’arbre bronchiques de diamètre
inférieur à 1 mm. Elles n’ont ni cartilage ni glandes séro-muqueuses. Elles sont revêtues d’un
épithélium cylindrique simple fait de cellules ciliées qui sont progressivement associées au
niveau des bronchioles terminales à des cellules de "Clara". Les cellules de Clara sont des
cellules pyramidales dont le pôle apical est garni de microvillosités. Dans la partie apicale de
leur cytoplasme existent des grains de sécrétion expulsés par exocytose. Ce produit fait de
glycosaminoglycanes se répand sur la surface bronchiolaire afin de la protéger. Le chorion est
riche en fibres élastiques. Le muscle devient discontinu mais toujours actif : il se contracte en

22
fin d’expiration et se relâche à l’inspiration. En cas de crise d’asthme la contraction se prolonge
et l’inspiration devient difficile. A noter la présence tout au long des bronchioles suivant les
trajets lymphatiques d’un tissu lymphoïde. Celui ci peut devenir hyperplasique au cours de tout
type d’agression bronchiolaire. La présence de follicules lymphoïdes est alors responsable
d’une lésion appelée "bronchiolite folliculaire". [5]
I. 1.4.2 Zone de transition
Les bronchioles respiratoires est une zone est extrêmement vulnérable car elle est située entre
les voies de conduction et d’échanges gazeux. Ainsi des lésions atteignant la bronchiole
respiratoire sont fréquentes chez les fumeurs. Il existe 3 générations de bronchioles
respiratoires. Leur structure histologique est identique à celle de la bronchiole terminale à ceci
près que plus elles sont périphériques plus leur paroi est interrompue par la présence
d’alvéoles. Leur chorion contient des cellules musculaires disperses (Figure 6).
Figure 6 : Poumon périphérique : zone d'échanges gazeux
I.1.4.3 Zone d’échanges gazeux
- L’acinus pulmonaire et la barrière alvéolo-capillaire
Au fur et à mesure le nombre d’alvéoles s’ouvrant dans la bronchiole respiratoire est tellement
important qu’il n'y a plus réellement d’épithélium bronchiolaire : on parle alors de canal
alvéolaire. Un acinus (ou lobule pulmonaire primaire) est constitué d’une bronchiole
respiratoire donnant naissance de 3 à 6 canaux alvéolaires. Chaque canal alvéolaire lui même se
divise de 2 à 3 fois. Chaque canal alvéolaire final s’ouvre au niveau de l’atrium sur 2 ou 3 sacs
alvéolaires. A ce niveau le muscle disparaît, l’armature est uniquement faite de fibres
collagènes et élastiques. Les fibres élastiques permettent aux alvéoles de s’élargir lors de

23
l’inspiration. Les alvéoles se contractent ensuite "passivement" lors de l’expiration. Les fibres
collagènes évitent une trop grande distension des murs alvéolaires et donc leur rupture.
Les alvéoles sont des "sacs" de 200µm de diamètre. Leur structure histologique est adaptée à
leur fonction essentielle : l’échange oxygène/gaz carbonique. La surface d’échange de
l’ensemble des 300 millions d’alvéoles représente à peu près 140 m2 ; 12000L d'air filtrés face
à 6000 L de sang perfusés par jour. Dans chaque alvéole on distingue la lumière alvéolaire
bordée par un épithélium alvéolaire et le "mur" ou cloison inter alvéolaire (interstitium) où
circule un réseau capillaire abondant.
I.1.4.4 L’épithélium alvéolaire
L’épithélium alvéolaire se compose de 2 types de cellules dites type I et II
1-Les pneumocytes de type I recouvrent 97% de la paroi alvéolaire. Leur rôle principal est
d’être une barrière protégeant l’alvéole mais perméable aux gaz.
2-Les pneumocytes de type II sont des cellules volumineuses.
Elles produisent et secrètent le surfactant. Cette substance est composée de protéines et de
phospholipides.
Il recouvre la surface alvéolaire et diminue la tension de surface. Ainsi la force d’inspiration
nécessaire pour dilater l’alvéole est moins grande. De plus en l’absence de surfactant l’alvéole
a tendance à se collaber. Chez l’enfant prématuré une insuffisance de surfactant peut-être
responsable de la maladie des membranes hyalines. Le surfactant a aussi un rôle antibactérien.
Elles ont des capacités de présentation antigénique et de sécrétion de cytokines et donc rôle
important dans les processus inflammatoires pulmonaires
A l’état normal les principaux éléments cellulaires retrouvés dans la ‘lumière alvéolaire’ sont
les macrophages qui se déplacent sur la paroi alvéolaire, immergés dans le surfactant.
La barrière alvéolo-capillaire (air-sang) de 0,1 à 0,5 µm d’épaisseur, est formée par différentes
couches :
- Le surfactant (phospholipides et protéines)
- Un film aqueux de quelques microns les pneumocytes
- L'interstitium pulmonaire : myofibroblastes
- La membrane basale résultant de la fusion de la membrane basale des pneumocytes d’un côté
et des cellules endothéliales de l’autre.
- Les cellules endothéliales forment la paroi des capillaires ; elles ont une fonction métabolique
importante (angiotensine, prostaglandine, bradykinine, sérotonine).

24
Cette barrière est perméable aux gaz ainsi l’oxygène qui est amené dans la lumière alvéolaire
lors de l’inspiration passe à travers la barrière dans la circulation capillaire et sera transporté
dans tout l’organisme par la circulation vasculaire. A l’inverse, le gaz carbonique, ramené aux
poumons par la circulation capillaire, passe à travers la barrière alvéolo-capillaire et est éliminé
lors de l’expiration.
En cas d’agression le poumon périphérique réagit dans un premier temps par un élargissement
des murs alvéolaires en rapport avec un afflux d’éléments inflammatoires (polynucléaires,
lymphocytes). Puis ces lésions peuvent avoir une évolution fibrosante : les murs alvéolaires
sont alors siège d’une hyperplasie de fibres musculaires puis d’une fibrose collagène.

25
I.2.Etiologie et épidémiologie du cancer des poumons
I.2.1 Définition du cancer
La tumeur cancéreuse ou le cancer est un terme générique qui désigne toutes les tumeurs
malignes qu’elles soient d’épithéliale ou conjonctive. La tumeur cancéreuse est définie come
étant une prolifération indéfinie d’une ou plusieurs cellules appelées clone. Ces clones
échappent aux mécanismes de régulation proliférative, envahissent et détruisent les tissus
normaux et provoquent la mort des patients par extension des cellules tumorales à tout
l’organisme [5].
Figure 7 : Les étapes d’évolution d’une tumeur maligne. 1) cellules génétiquement
altérée, 2) hyperplasie, 3) Dysplasie, 4) carcinome in situ, 5) cancer invasif [6].
Figure 8: Dissection d'un poumon cancéreux. Les tissus
blancs sont des tissus cancéreux, les tissus noirs d’une
personne fumeuse [6].

26
I.2.2 Le cancer broncho-pulmonaire
Le cancer broncho-pulmonaires se développe dans les bronches et dans les poumons. Il existe
deux types de cancer broncho-pulmonaire :
1. Cancer broncho-pulmonaire non à petites cellules (CBNPC) une forme de cancer la plus
répandue, qui progresse moins rapidement que les autres types, on trouve trois variétés :
- Carcinomes épidermoïde
- Adénocarcinome,
- Carcinome à grandes cellules.
2. Cancer broncho-pulmonaire à petites cellules (CBPC), forme de cancer plus rare qui
progresse plus rapidement que celui non à petites cellules, il existe trois variétés :
- Carcinome à petite cellules,
- Carcinome mixte à petites et grandes cellules,
- Carcinome mixte à petites cellules.
I.2.3 Etiologie
I.2.3.1 Les principales causes du cancer broncho-pulmonaire
Avec prés de 22 700 décès en 2000, le cancer du poumon est au premier rang des causes de
décès par cancer pour l’homme et au troisième pour la femme.
Le cancer broncho-pulmonaire est habituellement causé par le tabagisme ou l’exposition à la
fumée secondaire, mais il peut avoir d’autres causes. La principale cause est le tabagisme, ce
qui représente 80% des cancers du poumon recensés, dont 90% chez les hommes et moins de
70 % chez les femmes.
I.2.3.1.1 Les facteurs de risque environnementaux et professionnels
Les facteurs de risque professionnels et à moindre titre, environnementaux sont actuellement
considérés comme la seconde cause bronchique chez les hommes.
I.2.3.1.1.1 Le tabac
Depuis les années cinquante, il a été démontré formellement que le tabagisme était un facteur
de risque indéniable de néoplasie pulmonaire [19]. Il en est même le premier facteur de
risque, quel que soit le type histologique, même s’il favorise davantage le développement de
carcinomes épidermoïde et d’adénocarcinomes à petites cellules. Il n’y a pas de seuil minimal
de consommation en dessous duquel la consommation tabagique ne présenterait pas un

27
facteur de risque de cancer du poumon [20]. On sait que l’élément déterminant pour le risque
de carcinogenèse est plus la durée d’exposition au tabac que le nombre total de cigarettes
fumées [7].
Le fait de commencer à fumer jeune accroît le risque carcinogénique [21]. On voit apparaître
un décalage d’une vingtaine d’années entre les courbes de consommation du tabac et
l’incidence du cancer du poumon. Ceci est lié à l’histoire naturelle de la carcinogenèse
pulmonaire [8]. La fraction des cancers bronchiques attribuable au tabac est évaluée à 91 %
chez l’homme et à 57 % chez la femme. Le risque de développer un cancer du poumon chez
l’homme fumeur est 24 fois plus important que chez le non-fumeur; chez la femme, par
contre, il est 9 fois plus important chez la fumeuse que chez la non-fumeuse. On note une
diminution du risque chez les patients ayant arrêté de fumer par rapport à ceux chez qui le
tabagisme persiste. Cette diminution est d’autant plus importante que la durée d’abstinence
tabagique est longue. On estime qu’une personne ayant arrêté depuis 10 ans à cinq fois moins
de chance de développer un cancer bronchique qu’une personne persistant dans ce
comportement [5].
Un model schématique communément admis décrit l’évolution de l’épidémie tabagique. Cette
épidémie se décompose en quatre phases. Dans une société, le tabagisme s’installe d’abord
chez les hommes dont le niveau socio-économique est élevé. Puis il envahit les hommes dont
le statut socio-économique est plus faible. L’épidémie tabagique croît jusqu’à un maximum
puis subit une régression. C’est ce qui s’est passé aux Etats-Unis et en Europe où la
proportion d’hommes fumeurs est depuis quelques années en régression. L’épidémie atteint
de la même manière la femme, mais avec quelques années de décalage. Ceci explique qu’en
France l’épidémie des maladies liées au tabac a sûrement atteint son pic probable chez
l’homme, alors que chez la femme l’épidémie des maladies liées au tabac ne fait que débuter
[22]. Si la plupart des pays occidentaux se trouvent dans cette phase de décroissance, les pays
en voie de développement sont, quant à eux, toujours en phase de croissance de
consommation tabagique.
C’est non seulement la consommation active de tabac, mais aussi l’inhalation passive de la
fumée exhalée par les fumeurs actifs qui accentuent le risque cancérigène pour le poumon. A
cet égard, il est intéressant de noter que, depuis 2002, l’OMS (Organisation Mondiale de la
Santé) a reconnu le rôle cancérigène pour le poumon de l’exposition passive à la fumée de
tabac, même si le risque est nettement moindre que chez le fumeur actif.

28
I.2.3.1.1.2 L’exposition à l’amiante
De par ses propriétés d’isolant thermique et son incroyable résistance, l’amiante a connu de
multiples utilisations industrielles. De nombreux ouvriers sont ou ont été exposés à l’amiante
(production de garniture de frein, extraction de minerais, installation de matériel friable à base
d’amiante, électriciens, plombiers, maçons,..). Une étude française de 2003 montre que 12 %
des cancers du poumon chez les hommes de plus de 55 ans et environ 7 % dans la tranche
d’âge de 35 à 55 ans peuvent être attribués à une exposition professionnelle à l’amiante [23].
On note un effet synergique du tabagisme et de l’exposition à l’amiante sur le risque de
développer un cancer bronchique [24] (figure 9).
Figure 9 : Effet multiplicateur du tabagisme et de l’exposition à
l’amiante sur le risque relatif de cancer du poumon [24].
I.2.3.1.1.3 Autres expositions à risque
En 1997, la silice a été classée comme cancérigène par le Comité International de Recherche
sur le Cancer. Récemment, l’exposition au cadmium chez des ouvriers flamands a été
reconnue comme responsable de cancer du poumon [19]. Par ailleurs arsenic, nickel, chrome,
gaz moutarde sont autant d’autres facteurs reconnus.
- Le radon
Le radon est un gaz radioactif naturel émis par certains sols et, en particulier, par les sols
schisteux auquel le poumon est exposé par inhalation. Une méta-analyse de 2003 montre une

29
augmentation de 24 % du risque de cancer de poumon chez les patients exposés dans leur
maison à une radiation au radon de 150 Bq/m3 [20].
I.2.3.1.1.4 La pollution atmosphérique
La mortalité par cancer du poumon est plus importante en milieu urbain qu’en milieu rural
[10]. Les hydrocarbures polycycliques aromatiques provenant de la combustion du fuel fossile
et des émissions du diesel plus abondant dans l’air urbain pourraient expliquer cette
observation [22].
-L’alimentation
Une analyse regroupant 8 études prospectives concernant 3.206 cas détectés de cancer du
poumon parmi 430.281 sujets suivis pendant 6 à 16 ans a démontré, pour une consommation
tabagique identique, une réduction de 16 à 23 % du cancer bronchique chez les individus
consommant régulièrement des fruits. L’effet protecteur de la consommation de légumes est
moins important [23].
Co morbidités
Les patients porteurs d’une broncho-pneumopathie chronique obstructive (BPCO) ont un
risque relatif significativement plus important de développer un cancer du poumon, quel que
soit le type histopathologique de celui-ci [24]. Les fibroses et les cicatrices pulmonaires
augmentent aussi le risque du cancer du poumon, particulièrement celui d’adénocarcinome.
I.2.3.2 Prédisposition génétique
La relation entre tabagisme et cancer du poumon est indiscutable. Mais, comme seuls 10 à 15
% des fumeurs réguliers et que des personnes n’ayant jamais fumé développeront cette
pathologie, on pense qu’il existe une certaine sensibilité génétique aux agents cancérigènes
environnementaux. Quelques études se sont concentrées sur le polymorphisme génique au
niveau des voies métaboliques d’activation des substances cancérigènes, des voies de
détoxification et des mécanismes de réparation de l’ADN. Pour se lier à l’ADN et induire des
lésions géniques, les hydrocarbures polycycliques aromatiques (présents dans les cigarettes)
doivent être métabolisés par des enzymes du cytochrome P450. Le CYP1A1 et le CYP2D6
font partie de celles-ci. Pour le CYP1A1, le polymorphisme MspI et une mutation au niveau
du codon 462 de l’exon 7 du gêne codant pour cette enzyme (cette dernière mutation appelée
Ile462Val entraîne le remplacement de la valine par l’isoleucine à proximité du site protéique
liant l’hème) étaient associés à un risque accru de développer un cancer du poumon. Les
sujets porteurs du phénotype CYP2D6 métaboliseur rapide de la débrisoquine pourraient être

30
à risque accru de développer une néoplasie pulmonaire, mais les résultats des études portant
sur ce sujet sont contradictoires.
La gluthation S-transférase est une enzyme détoxifiant les métabolites réactifs des
hydrocarbures aromatiques polycycliques. Certains polymorphismes géniques de cette
enzyme ont été associés au cancer du poumon. Une méta-analyse a montré que les patients ne
possédant pas l’allèle GSTM1 étaient plus à risque que ceux le possédant. La conjugaison de
la présence de l’allèle CYP1A1 Ile462Val et de l’absence d’allèle GSTM1 augmenterait de
quatre fois le risque de développer un cancer du poumon chez le sujet non fumeur [15].
I.2.3.2.1 Modifications géniques au sein de la tumeur
Certaines études ont montré que le récepteur au facteur de croissance épithélial (EGFR) était
régulièrement modifié dans les néoplasies épithéliales [26]. L’EGFR est une tyrosine kinase
de la famille ErbB. Ce récepteur se trouve sous forme de monomère lorsqu’il n’est pas activé
par le ligand. La fixation du ligand sur le domaine extra-membranaire de la protéine entraîne
la dimérisation de celle-ci et l’activation via la consommation d’ATP d’une voie de
signalisation intracellulaire importante pour la survie et la multiplication cellulaire [18] (10).
Le gène EGFR est un oncogène capable d’induire une néoplasie lorsqu’il est muté. En fait, le
système EGF/EGFR peut faciliter la cancérisation par trois mécanismes : la surstimulation de
l’EGFR par un excès de production d’EGF [27], la surexpression d’EGFR [28] ou l’activation
excessive et spontanée par mutation de l’EGFR [29]. Une étude ayant analysé les mutations
EGFR sur 759 pièces tumorales bronchopulmonaires montrait que le gène codant pour cette
protéine était muté dans 149 pièces tumorales (19,6 %). Ces mutations étaient plus fréquentes
chez les non-fumeurs, les femmes, les patients atteints d’adénocarcinomes et ceux provenant
de l’Est Asiatique. L’EGFR est codé par un gène situé au niveau du chromosome 7p12. Les
principales mutations se trouvent entre l’exon 18 et l’exon 21 [17, 30].

31
Figure 10 : Schéma d’activation du récepteur EGFR par l’EGF. La liaison de l’EGF
entraîne une dimérisation du récepteur qui stimule le domaine intracellulaire de l’EGFR. Via
l’ATP, plusieurs cascades cellulaires sont activées : la prolifération tumorale d’un côté (sys
RAS-RAF), l’activation antiapoptotique d’un autre côté (sys Akt et Stat 3/5) [28].
I.2.4 Epidémiologie descriptive du cancer des poumons
I.2.4.1 Cancer des poumons en Algérie
Il est la première cause de mortalité par cancer chez les hommes en Algérie. Le Dr Ameur
Soltane, président de l’Association algérienne du cancer des poumons, a révélé que 2 500
à 3 500 nouveaux cas de cancer des poumons sont enregistrés chaque année en Algérie, et
qu’entre 80 et 87% des personnes atteintes du cancer des poumons meurent dans les cinq ans
qui suivent le diagnostic. Il a ajouté qu’un cas sur quatre peut subir une opération. Cela
intervient au moment où le nombre de décès annuel est de 15000, selon le Dr Maghraoui du
service de chirurgie thoracique de l’hôpital d’Oran .Ce dernier affirme que le tabac est à
l’origine de 90% des cas, alors que 10 % de ce type de cancers sont liés aux facteurs
professionnels dont l’amiante. Rien que dans le secteur sanitaire, il a été recensé 96 cas de
cancer à l’amiante entre 1998 et 2005. Il convient enfin de signaler que la prévalence du
cancer a connu une évolution rapide en Algérie et le coût de sa prise en charge est de plus en
plus élevé. (Amina Azoune. Le jeune indépendant - Algérie - 06/04/2006).
Le Pr. Berrabah du CHU d’Oran, a estimé que «le gros problème qui se pose en Algérie est
l’insuffisance de la prévention». «Le tabac qui était l’apanage des hommes se féminise ;
beaucoup de femmes fument, ce qui fait craindre une flambée du cancer des poumons chez la
femme d’ici 10 à 15 ans», a-t-il noté.
Dans notre pays, ce type de cancer est en nette progression, et ce, malgré les progrès
scientifiques réalisés durant les 20 dernières années dans la connaissance des mécanismes

32
intimes du cancer bronchique et l’amélioration de sa prise en charge, notamment par la
découverte de nouvelles drogues.
I.2.4.2 Cancer des poumons en France
Le réseau français des registres des cancers (Francim) et l’Institut national de la santé et de la
recherche médicale (Inserm) estiment le nombre total de nouveaux cas de cancers en France,
en 2000, à 280 000, dont 58 % survenant chez les hommes. Pour la même année, le nombre
de décès par cancer est estimé à 150 000 dont 61 % chez l’homme.
Le cancer du poumon vient en 4e position pour son incidence (28 000 nouveaux cas) après le
cancer du sein (42 000), de la prostate (40 000), et du colon et rectum (36 000) et en première
place pour sa mortalité. 23 152 cancers du poumon ont été diagnostiqués chez l’homme en
2000 et 4 591 chez la femme. Compte tenu de l’efficacité modeste des traitements, le cancer
du poumon est la première cause de mort par cancer avec environ 27 000 décès en 2000, dont
23 000 chez l’homme et 4 500 chez la femme (Inserm, année 2000). Selon l'assurance
maladie, 14 237 nouveau cas en France métropolitaine en 2002.
Tableau 1 : Taux d’incidence (standardisé au monde) du cancer du poumon pour 100 000
hommes et femmes (source Cnamts 1997- 2002)
Hommes Femmes
1997 41,3 7,3
1998 41,5 7,7
1999 40,4 7,9
2000 40,8 8,6
2001 40,9 9,1
2002 40,1 9,6
I.2.4.3. Cancer des poumons en Canada
Au Canada, en 2005, on estime que 22 200 personnes ont reçu un diagnostic de cancer du
poumon, soit 12 000 hommes et 10 200 femmes.
Au cours de la même période, 10 700 hommes et 8 300 femmes ont été emportés par cette
maladie. Le cancer du poumon est, de loin, le plus mortel des cancers au pays.
Chaque semaine, 427 Canadiens apprennent qu’ils sont atteints de ce cancer, tandis que
365 en meurent.

33
Au Canada, 1 homme sur 11 (soit 8,8 %) court le risque d’être atteint du cancer du poumon au
cours de sa vie. Chez les femmes, ce risque est de 1 femme sur 17 (soit 5,9 %).
Le risque de succomber à cette maladie, chez les Canadiens, est de 1 homme sur 12, et de
1 femme sur 20. Tant pour les hommes que pour les femmes, la principale cause du cancer du
poumon est le tabagisme. (Institut national du cancer du Canada, Statistiques canadiennes sur
le cancer 2005).
I.2.5. Épidémiologie analytique
Le cancer broncho-pulmonaire primitif est la première cause de décès par cancer chez
l'homme et ce dans tous les pays développés. On estime le nombre de nouveaux cas par an à
28000 en France et à plus de 150000 aux Etats-Unis. L'incidence et la prévalence sont proches
ce qui témoigne du pronostic désastreux de cette maladie. Le cancer broncho-pulmonaire est
donc un véritable problème de santé publique. Paradoxalement, une de ses particularités est
d'être accessible à la prévention puisque son principal facteur de risque, le tabagisme est
identifié.
I.2.5.1. L'âge
L'âge médian de survenue d'un cancer broncho-pulmonaire est de 60 ans et 75 % des patients
ont entre 50 et 70 ans au moment du diagnostic.
I.2.5.2. Le sexe
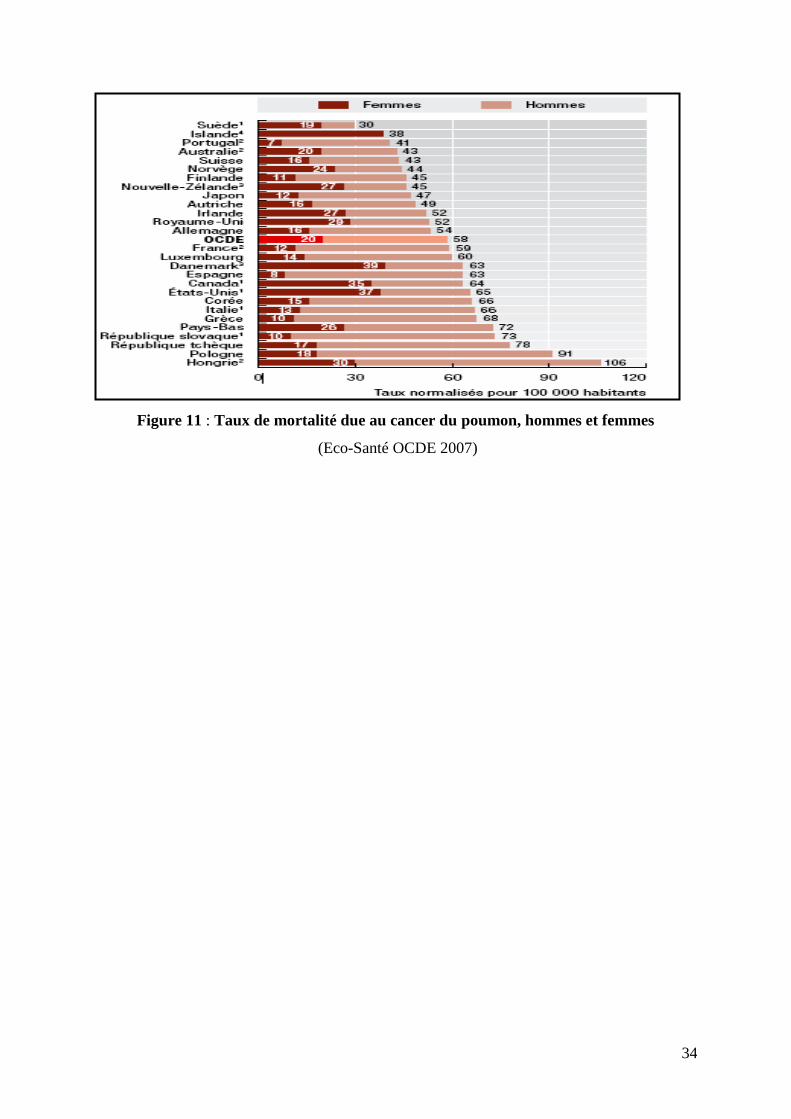
Si l’on concentre l’analyse sur des types spécifiques de cancer, le cancer du poumon reste le
plus mortel chez les hommes dans tous les pays de l’OCDE (à l’exception de la Suède) et
demeure l’une des principales causes de décès par cancer chez les femmes. La consommation
de tabac est le facteur de risque le plus important pour le cancer du poumon.
En 2004, les pays qui affichaient les taux les plus élevés de décès par cancer du poumon
étaient les pays d’Europe centrale et orientale (la Hongrie, la Pologne, la Républiques tchèque
et slovaque), les Pays-Bas, la Grèce et la Corée. Dans tous ces pays, la proportion de fumeurs
chez les hommes a été de tout temps (et reste) relativement élevée. C’est en Suède que le taux
de décès par cancer du poumon est le plus faible; ce pays est également l’un de ceux dans
lesquels les hommes fument le moins (figure 11).
34
Figure 11 : Taux de mortalité due au cancer du poumon, hommes et femmes
(Eco-Santé OCDE 2007)

35
I.3.Anatomie pathologique des cancers broncho-
pulmonaire
I.3.1 Classification des cancers broncho-pulmonaire
La dernière classification de l’OMC éditée en 1999, préserve les nomenclatures consacrées
par le temps et les nouvelles acquisitions liées aux progrès des techniques qui permettent de
compléter et d’affiner la simple analyse morphologique. Cette classification est représentée
dans tableau 3.
I.3.1.1 Cancer bronchique épidermoïde
Il se localise préférentiellement dans une bronche lobaire ou segmentaire et se présente
volontiers sous la forme d'une tumeur végétative en "chou-fleur" obstruant plus ou moins la
lumière bronchique. En même temps que la tumeur progresse dans la lumière bronchique elle
envahit la sous-muqueuse et le tissu péri-bronchique. Lorsque la tumeur est périphérique une
nécrose centrale est habituelle ; lorsque la tumeur est proximale il s'y associe une pneumonie
obstructive.
Histologiquement cette tumeur se caractérise par l'existence de ponts intercellulaires, de
kératinisation et par la formation de "perles" cornées.
L'ultrastructure des formes bien différenciées comporte des cellules à cytoplasme abondant
avec un réticulum endoplasmique, un appareil de Golgi et des mitochondries peu importants
mais des ribosomes et des filaments intermédiaires abondants. Ces derniers s'agrègent
volontiers pour former des desmosomes.
I.3.1.2 Cancer bronchique à petites cellules
Il se localise généralement près des voies aériennes proximales. La tumeur est mal limitée et
s'étend dans la sous -muqueuse et les tissus de connexion. L'extension endo-bronchique est ici
beaucoup plus rare et lorsqu'il existe une obstruction bronchique elle est généralement due à
une compression extrinsèque. L'invasion des vaisseaux et des lymphatiques est très précoce et
les relais ganglionnaires hilaires et médiastinaux sont inévitablement envahis.

36
Tableau II Classification histologique des carcinomes broncho-pulmonaires extrait de
l’OMS, 1999.
1. Carcinome épidermoïde
1.1 Papillaire
1.2 A cellulaires claires
1.3 A petites cellules
1.4 Basaloïde
2. Carcinome à petites cellules
2.1 Carcinome à petites cellules
3. Adénocarcinome
3.1 Acineux
3.2 Papillaire
3.3 Broncho-alvéolaire
3.3.1 Non mucineux
3.3.2 Mucineux
3.3.3 Mixte (mucineux et non mucineux)
4. Carcinome à grande cellules
4.1 Carcinome neuroendocrine à grandes cellules
4.1.1 Carcinome neuroendocrine à grandes cellules composites
4.2. Carcinome basoloïde
4.3 Carcinome lympho-épithéliale like
4.4 Carcinome à cellules claires
4.5 Carcinome à grande cellules de phénotype rhabdoïde
5. Carcinome adéno-squameux
6. Carcinome pléiomorphe, sarcomatoïde ou avec éléments sarcomateux
6.1 Carcinome à cellules fusiforme et / ou géantes
6.1.1 Carcinome pléiomorphe
6.1.2 Carcinome à cellules fusiforme
6.1.3 Carcinome à cellules géantes
6.2 Carcinosarcome
6.3 Blastome
7. Tumeurs carcinoïde
7.1 Carcinoïde typique
7.2 Carcinoïde atypique
8. Carcinome du type glandes salivaires
8.1 Carcinome muco-épidermique
8.2 Carcinome adénoïde kystique
8.3 Autres
9. Carcinomes inclassables

37
Figure 12: Biopsie d'un poumon souffrant de carcinome à petites cellules.
Le sous-type oat-cell est fait de cellules uniformes mesurant 1 à 1,5 fois la taille d'un
lymphocyte. Le cytoplasm est quasi absent. Les noyaux sont hyperchromatiques, contenant
une chromatine fine et dispersée et de petits nucléoles.
Le sous-type intermédiaire est fait de cellules plus grandes, rondes ou polygonales, contenant
un cytoplasme un peu plus abondant. Les noyaux sont moins hyperchromatiques. Les cellules
de ces deux sous-types sont arrangées en rubans ou en rosettes. Ces tumeurs sont volontiers
très nécrotiques. Parfois il existe deux composantes, l'une correspondant à un cancer
bronchique à petites cellules typique, l'autre à un cancer épidermoïde ou à un adénocarcinome.
Il s'agit alors de la variante composite.
En microscopie électronique, la présence de granules neurosécrétoires est très évocatrice du
diagnostic de carcinome bronchique à petites cellules.
I.3.1.3.Adénocarcinome bronchique est divisé en 4 sous-groupes
histologiques
Acinaire, papillaire, bronchiolo-alvéolaire et carcinome solide avec formation de mucus. La
signification clinique de cette subdivision est très discutée avec l'exception de la forme diffuse
de l'épithélioma bronchiolo-alvéolaire (figure 12).

38
Figure 13 : Adénocarcinome bronchique iconographie.
L'adénocarcinome se localise volontiers en périphérie dans les régions souspleurales. Il est
habituellement bien limité bien qu'il puisse exister des aspects spiculés, irréguliers. Lorsqu'il
s'agit d'un bronchiolo-alvéolaire, il n'y a pas de destruction de l'architecture alvéolaire et les
alvéoles sont littéralement tapissés par les cellules cancéreuses ; radiologiquement l'image est
alors celle d'une pneumonie. L'adénocarcinome montre une différenciation glandulaire de
degré variable pouvant ou non former des papilles.
En microscopie électronique l'aspect varie selon le degré et le type de différenciation.
Les cellules sont columnaires ou cuboïdales avec des microvillosités de surface, un réticulum
endoplasmique et un appareil de Golgi relativement abondant, et des granules sécrétoires.
En immuno-histochimie le marquage pour l'antigène carcino-embryonnaire (ACE) est très
fréquent de même que pour les cytokératines de bas poids moléculaire.
I.3.1.4. Epithéliome bronchique à grandes cellules
L'épithéliome bronchique à grandes cellules est essentiellement un diagnostic d'exclusion et
s'applique aux tumeurs qui n'ont pas l'aspect d'un cancer bronchique à petites cellules et qui
n'ont ni différenciation épidermoïde ni différenciation glandulaire. La proportion d'épithélioma
à grandes cellules varie donc largement selon le degré de sophistication des colorations et des
techniques anatomo-pathologiques et selon la quantité de tissu disponible pour le diagnostic. Il
s'agit en général de tumeurs périphériques volumineuses. Histologiquement les tumeurs
consistent en des nappes de cellules contenant un cytoplasme éosinophile abondant.
I.3.1.5 Carcinomes adénosquameux
Les carcinomes adénosquameux sont très rares surtout si l'analyse est faite en microscopie
optique. Si on se fonde sur la microscopie électronique, la fréquence peut aller jusque 40 %.
Le pronostic semble être identique à celui de l'adénocarcinome [31].

39
I.3.2 Diagnostic du cancer broncho-pulmonaire
I.3.2.1. Les signes cliniques
I.3.2.1.1 Signes en rapport avec la tumeur bronchique
1- La toux est le plus fréquent des symptômes inauguraux. Elle n'amène cependant que
rarement le patient à consulter car il la considère comme banale et due au tabagisme. Cette
toux est liée à la stimulation des récepteurs endobronchiques par des phénomènes mécaniques
(compression par la tumeur) ou des phénomènes inflammatoires. Ces récepteurs sont moins
fréquents en périphérie au niveau des bronchioles qu'au niveau des grosses bronches.
2 - L'expectoration :
L'apparition d'une expectoration purulente avec ou sans fièvre peut être due à une infection en
amont d'une sténose ou à une nécrose tumorale. Celle-ci est surtout le fait des carcinomes
épidermoïdes. Classiquement le carcinome bronchiolo-alvéolaire serait responsable d'une
sécrétion muqueuse abondante. Ce phénomène est en fait rare et tardif.
3 - La dyspnée :
En cas de tumeur à développement central, la dyspnée sera liée à l'obstruction par un bourgeon
néoplasique ou à la compression extrinsèque d'une grosse bronche. En cas d'obstruction d'une
grosse bronche on pourra entendre un ronchus fixe, un wheezing voire un cornage si la tumeur
est située dans la trachée.
4 - L'hémoptysie :
N'est pas un symptôme très fréquent mais elle alerte davantage le patient.
I.3.2.1.2 Signes en rapport avec l'extension loco-régionale
Le syndrome cave supérieur associant des céphalées, une cyanose de la face, un oedème en
pèlerine, une turgescence des jugulaires, une circulation collatérale thoracique antérieure est
relativement fréquent et en rapport avec l'engainement ou la compression de la veine cave
supérieure par la tumeur ou par une adénopathie latéro-trachéale droite.
- Une compression oesophagienne par la tumeur ou par une adénopathie sera à l'origine d'une
dysphagie.
- Les épanchements pleuraux sont généralement secondaires à l'envahissement de la plèvre
viscérale et donc néoplasiques, mais parfois ils sont simplement liés à une atélectasie.

40
- Une tamponnade, une arythmie récente signent l'envahissement péricardique par la tumeur ou
ses extensions lymphatiques.
- Un hoquet ou une paralysie phrénique sont en rapport avec une compression du nerf
phrénique.
- Des douleurs pariétales thoraciques lorsqu'elles sont fixées, insomniantes, évoqueront
l'envahissement de la paroi par la tumeur.
- Une dysphonie témoigne généralement d'une compression du nerf récurrent gauche sous la
crosse de l'aorte, par une adénopathie ou par la tumeur.
- Une adénopathie sus-claviculaire généralement de consistance élastique, mobile ou fixée au
plan profond peut révéler un cancer bronchique.
- Le syndrome de Pancoast-Tobias reconnait le plus souvent une étiologie cancéreuse et
associe des douleurs scapulaires puis des douleurs de type radiculaire C8 - D1 irradiant
jusqu'au 5e doigt, un syndrome de Claude-Bernard Horner homolatéral avec myosis,
enophtalmie, et rétrécissement de la fente palpébrale. Ces signes sont liés au développement
d'un cancer de l'apex envahissant le plexus brachial, le ganglion stellaire sympathique et
détruisant les 2 premiers arcs costaux.
Tous les signes en rapport avec une extension loco-régionale éliminent d'emblée toute
possibilité chirurgicale à l'exception parfois de l'envahissement pariétal [31].
I.3.2.1.3 Signes en rapport avec l'extension métastatique
Les sites métastatiques les plus fréquents sont le foie, l'os et le système nerveux central. C'est
ainsi qu'une hépatomégalie douloureuse parfois marronnée avec ou sans perturbation des
fonctions hépatiques, des douleurs osseuses, des manifestations neurologiques (déficitaires ou
épileptiques), des nodules sous-cutanés violacés peuvent révéler un cancer bronchique primitif.
Les métastases surrénaliennes et gastro-intestinales sont fréquentes mais ne s'accompagnent
généralement pas de symptômes cliniques.
I.3.2.1.4 Signes en rapport avec des syndromes paranéoplasiques sont fréquents
Certains ne sont rencontrés que dans le cancer bronchique non à petites cellules: syndrome de
Pierre-Marie ou ostéoarthropathie hypertrophiante pneumique, hypercalcémie alors que
d'autres sont spécifiques du cancer bronchique à petites cellules : hyponatrémie du syndrome
de Schwartz-Bartter en rapport avec une sécrétion inappropriée d'hormone antidiurétique,
syndrome de Cushing, syndrome de Lambert-Eaton. D'autres enfin se voient indifféremment
dans tous les types histologiques : syndromes hématologiques, cutanés etc...

41
I.3.2.1.5 Autres cas
Certains cancers bronchiques notamment ceux se présentant comme un nodule solitaire en
plein parenchyme sont parfois détectés lors d'un examen radiographique pulmonaire de
rencontre et n'occasionnent aucun signe clinique.
I.3.2.2. Signes radiologiques
Ils sont très variables mais souvent évocateurs et permettent de différencier les cancers
centraux développés dans le tiers interne des champs pulmonaires, des cancers périphériques.
Les premiers se présentent souvent sous la forme d'une opacité polycyclique hilaire, ganglio-
tumorale accompagnée ou non d'une atélectasie ou d'un trouble de ventilation d'un ou deux
lobes voire de tout un poumon. Parfois l'opacité hilaire se poursuit vers le médiastin supérieur
par une opacité latérotrachéale voire même une ombre en cheminée signant l'envahissement
ganglionnaire médiastinal.
Ces images sont volontiers le fait des cancers bronchiques à petites cellules mais aussi de
certains épidermoïdes ou des indifférenciés à grandes cellules, beaucoup plus rarement des
adénocarcinomes.
Les seconds se présentent parfois sous forme d'un nodule solitaire, à contours spiculés,
grossièrement arrondi et correspondent volontiers à des adénocarcinomes. Lorsque la masse
tumorale est plus importante elle est fréquemment le siège d'une nécrose centrale donnant une
image de pseudo-abcès dont les parois sont épaisses et les limites internes irrégulières. Cette
image est quasi-pathognomonique d'un cancer épidermoïde.
Les cancers périphériques proches de la paroi peuvent s'accompagner d'une lyse costale, d'un
épanchement pleural.
Les cancers bronchiolo-alvéolaires diffus sont à l'origine d'une image de comblement
alvéolaire non systématisée comportant un bronchogramme aérique et pouvant s'étendre dans
les deux champs pulmonaires.
Le syndrome de Pancoast-Tobias se traduit volontiers par une coiffe apicale accompagnée de
la lyse des premiers arcs costaux [31].

42
I.4. Introduction à la famille de la sulfhydryl oxydase
Le cancer bronchique est la première cause de décès par cancer dans le monde. La première
cause en est le tabagisme. A Oran seul 88 nouveaux patients sont détectés chaque année et
présentent un mauvais pronostic. La gravité de la maladie est due au diagnostique tardif au
stade métastatique. La majorité des cas de cancer broncho-pulmonaire correspond à des
tumeurs non à petites cellules (NSCLC). On distingue en particulier l’adénocarcinome et le
carcinome épidermoïde qui sont les formes les plus représentées. Les profils d’expression
géniques de ces deux types majeurs de cancer font apparaître des changements moléculaires
avec en particulier une surexpression de gènes de détoxification ou anti-oxydants [32].
I.4.1. La famille de la sulfhydryl oxydase
Au niveau cellulaire ou tissulaire, la balance du niveau des réducteurs et des oxydants est en
équilibre. Un déficit ou une surproduction de l’un des acteurs moléculaires, va influencer le
comportement de la cellule [33].Les deux systèmes antioxydants les mieux connus sont, le
système glutathion réduit/ glutathion oxydé (2GSH/GSSG) et le système thiorédoxine réduite/
thiorédoxine oxydé (TRX(SH)2/TRXSS). Une déficience de l’un d’entre eux ou une
augmentation importante du taux de pro- oxydants et de radicaux libres (ROS, NO) induit des
lésions de l’ADN, dont des mutations dans les gènes pro- et anti-oncogène et entraîne de ce
fait une prolifération cellulaire [34].
La super-famille de la Thiorédoxine comprend la thiorédoxine (TRX1), la Protéine Disulfure
Isomérase (PDI), auxquelles sont associées des protéines reliées fonctionnellement comme la
thiorédoxine réductase. La thiorédoxine est une petite protéine ubiquitaire (Mr11500)
contenant un motif très conservé Cystéine-Glycine-Proline-Cystéine (CGPC), qui constitue
son site actif. Elle est impliquée dans l’activité de facteurs de transcription (NFB, AP-1,
ASK1) ce qui va moduler l’expression de gènes cibles et les fonctions cellulaires associées
(prolifération, apoptose, résistance aux agents antitumoraux). Une surexpression de la
thiorédoxine dans les cellules cancéreuses favorise la prolifération cellulaire et augmente la
résistance d’une variété de cellules à la chimiothérapie [35].
La protéine-disulfure isomérase (PDI) est très abondante dans le réticulum endoplasmique où
elle catalyse l’oxydation, la réduction des ponts disulfure ainsi que leur isomérisation en
utilisant le site actif Cystéine-Glycine-Histidine-Cystéine (CGHC) [36]. Elle est caractérisée

43
par la présence de deux domaines thiorédoxine fonctionnels similaires et une séquence de
rétention dans le réticulum endoplasmique Lysine-Aspartate-Glutamate-Leucine (KDEL).

44
SOx/Q6
’courte ’
SOx/Q6
’longue ’
PDI
Thiorédoxine
SOxN
CGPC
CGHC CGHC
N N
C73GHC75 CRDC CSAC512455 515452
C73GHC75 CRDC CSAC
NN
452 455 515512
CGHC CRDC CSAC
N
SOx/Q6
’courte ’
SOx/Q6
’longue ’
PDI
Thiorédoxine
SOxN
CGPC
CGHC CGHCCGHC CGHC
N N
C73GHC75 CRDC CSAC512455 515452
N N
C73GHC75 CRDC CSACC73GHC75 CRDC CSACC73GHC75 CRDC CSAC512455 515452
C73GHC75 CRDC CSAC
NN
452 455 515512C73GHC75 CRDC CSAC
NN
C73GHC75 CRDC CSACC73GHC75 CRDC CSACC73GHC75 CRDC CSAC
NN
452 455 515512
CGHC CRDC CSAC
N
CGHC CRDC CSACCGHC CRDC CSACCGHC CRDC CSAC
N
Figure 14 : Structures protéiques de la thiorédoxine, de la protéine disulfure
isomérase, de la sulfhydryl oxydase/Quiéscine Q6 (SOx/Q6) et de la sulfhydryl
oxydase de neuroblastome (SOxN). Les domaines thiorédoxine sont présentés en orange
et les domaines ERV1 en vert, les séquences transmembranaires en noir, l’extension C
terminale en violet. N: site de glycosylation potentiel (Seddiki, 2009).
Figure 15: Schéma représentant l’activité de la sulfhydryl oxydase/QuiescineQ6
(SOx/Q6). La SOx/Q6 forme un pont disulfure en présence d’oxygène moléculaire
en produisant du peroxyde d’hydrogène.

45
I.4.2. Structure de la sulfhydryl oxydase/Quiescine Q6
Récemment, la nouvelle famille des sulfhydryl oxydases FAD dépendantes a été décrite. les
premiers membres de cette famille ont été nommés sulfhydryl oxydase/Quiescine Q6
(SOx/Q6) [37], [38]. En effet, l’expression du gène SOx/Q6 humain est spécifiquement induit
dans les fibroblastes WI38 lors de leur entrée en quiescence [39]. Ceci laisse suggérer une
implication potentielle de ce gène dans la prolifération cellulaire. Un second membre de cette
famille a été récemment décrit à partir des cellules de neuroblastome et appelé sulfhydryl
oxydase dérivé de neuroblastome (SOxN), cette protéine joue un rôle dans la sensibilisation
des cellules à l’interféron gamma (IFN-) un inducteur de l’apoptose. La protéine est localisée
au niveau de la membrane plasmique et au niveau la membrane nucléaire [40].
Ces sulfhydryl oxydases (SOx) sont des flavoenzymes qui oxydent les groupements thiols des
protéines et des petites molécules pour former des ponts disulfures selon la réaction
2R- SH + O2 R-S-S-R+ H2O2.
Le gène de la quiescine Q6 humaine produit deux transcrits majeurs, un transcrit long de 5600
nucléotides codant pour une forme longue de la protéine (Mr de 80000) et un transcrit court
de 4800 nucléotides codant pour une forme courte (Mr 60000) (figure 14).
La structure protéique déduite des ADNc des sulfhydryl oxydases/ Quiescine Q6 présente un
domaine thiorédoxine de type PDI dans la région N terminale et un domaine catalytique ERV
(essentiel à la respiration et la viabilité) dans la région C terminale. SOx/Q6 longue possède
une extension de 143 acides aminés en position C terminale qui possède une séquence trans-
membranaire potentielle.
Les quatre protéines citées précédemment, la TRX, PDI, SOxN et la SOx/Q6 possédent sur le
plan structural au moins un domaine thiorédoxine mais ont sur le plan fonctionnel des actions
antagonistes qui varient entre l’oxydation et la réduction, ce qui suggère qu’il puisse y avoir
des interactions fonctionnelles entre ces protéines.
Les études montrent la présence de SOx/Q6 dans de nombreux tissus humains. Cette protéine
peut être aussi sécrétée par les cellules [37].Ceci suggère que SOx/Q6 puisse être impliquée
dans la formation de ponts disulfure d’une large variété de protéines au sein de la cellule
(figure 15) [41] et que cette molécule puisse aussi participer au dépôt et l’état de réticulation
de la matrice extra cellulaire (deux protéines ou deux peptides) par la catalyse de ponts
disulfure [41].
Dans cette optique, nous allons vérifier l’existence de la protéine Sox/Q6 dans les différentes
lignées cellulaires broncho-pulmonaires et les variétés de cette protéine par western blot, et

46
ainsi que son activité oxydasique dans les surnageants. Cependant, on ne peut pas exclure la
sécrétion d’autres sulfhydryl oxydases.
Ces résultats pouvant être comparés à ceux de l’expression des ARN messagers dans les
différentes lignées tumorales étudiées obtenus.

47
CHAPITRE II MATERIELS ET METHODES

48
II.1 L’étude épidémiologique du cancer pulmonaire de la région
oranaise
Une étude rétrospective a porté sur 173 patients atteints d’un cancer du poumon et traités au
service de radiothérapie du CHU Oran, entre Janvier 2004 et Décembre 2007. Les résultats
portent les caractéristiques épidémiologiques, cliniques, anatomo-pathologiques d’où :
160 cas masculins (92,5% des cas) et 13 cas féminins (7,5% des cas)
Les résultats ont été traités de la façon suivante :
- La répartition selon la localisation du cancer dans les poumons (droit et gauche)
- La répartition des cas en fonction des tranches d’âges
- La répartition des patients selon la classification du TNM et par type histologique

49
II.2 Matériel biologique
II.2.1 Lignées cellulaires
Les travaux ont été réalisés à partir d’une lignée de fibroblaste pulmonaire
embryonnaire humain WI38, et de cellules épithéliales pulmonaires tumorales humaines
(Calu-1, H1838, H358, H460, H23, A549, H520) (annexe 2). Ces lignées proviennent de
l’ATCC (American Type Culture Collection). Elles sont cultivées dans leurs milieux de
culture appropriés (MEM, RPMI 1640 (GIBCO)) avec 10% de sérum du veau fœtal (SVF)
(référence, 10106-169, Biological Industries) à 37°C et 5% de CO2.
II.2.1.2 Les surnageants de culture cellulaire
Les surnageants de culture ont été récupérés à pré-confluence soit trois jours après
ensemencement (T1), à confluence soit cinq jours après ensemencement (T2) et à post-
confluence soit sept jours après ensemencement (T3). Les échantillons sont conservés à -20°C
puis concentrés 20 fois avec des concentrateurs Vivaspin 2 mL (Vivasciences) ayant un seuil
de rétention de 10 000 par centrifugation répétées de 3 minutes à 5000g à 4°C (Universal 16
R). Un cocktail d’inhibiteurs de protéases (Sigma, P8340) est ajouté aux surnageants
concentrés dans la proportion de 1/50.
Des témoins SVF ont été réalisés parallèlement. Le milieu de culture RPMI avec 10% de SVF
a été incubé à 37°C et 5% de CO2 pendant 3, 5 et 7 jours avant concentration.
II.3. Préparation d’anticorps spécifiques dirigés contre la sulfhydryl
oxydase de rat
II.3.1 Extraction des anticorps anti-sulfhydryl oxydase de rat à partir de
jaune d’œuf de poule
Les IgY totales anti-sulfhydryl oxydase de rat sont extraites de jaunes d’œuf, provenant
de poules immunisées (399 et 400) contre la sulfhydryl oxydase de rat (rSOx), avec le
″Eggcellent Chicken IgY Purification kit″ (Pierce, 44918). La purification est basée sur une
délipidation suivi d’une précipitation. Les anticorps sont conservés en tampon phosphate 50
mM, pH 7,4, NaCl 0,5 M, à -20°C. La concentration en IgY obtenue est analysée par
spectrophotométrie à 280 nm (SHIMADZU, UV-160A) et par électrophorèse en gel de
polyacrylamide.

50
II.3.2 Purification des anticorps spécifiques anti-sulfhydryl oxydase de rat
par chromatographie d’affinité
Toutes les étapes de purification sont réalisées à 4°C. Le gel de Sépharose activé au bromure
de cyanogène (CNBr-actived Sepharose 4B, Pharmacia) sur lequel a été couplé de la
sulfhydryl oxydase de rat (rSOx) (colonne disponible au laboratoire) est équilibré avec du
tampon phosphate 0,05 M, pH 7,4, NaCl 0,5 M.
Les IgY totales purifiées à partir de l’œuf 399 soit 35,73 mg d’IgY dans 9 mL sont mises en
contact avec le gel 1mL pendant 2 heures sous agitation à 4°C. La colonne est ensuite lavée en
tampon phosphate 0,05 M, pH 7,4, NaCl 0,5 M. L’élution des IgY non fixées est suivie par
mesure de l’absorbance à 280 nm. Les IgY spécifiques sont alors éluées avec un tampon
glycine 100 mM, pH 3,0, contenant 15 % de D (+) tréhalose (Sigma, T-5251), un ose
protégeant les IgY de la dénaturation et neutralisées immédiatement avec du tampon Tris 1M,
pH 8,0. La colonne est conservée dans du tampon borate 1M, NaCl 0,5, pH 7,4 à + 4°C.
Figure 16: Purification des anticorps IgY anti SOx/Q6 par chromatographie. Le gel de Sépharose activé au bromure de cyanogène sur lequel a été couplé de la sulfhydryl
oxydase de rat (rSOx) est équilibré avec du tampon phosphate 0,05 M, pH 7,4, NaCl 0,5 M.
Les IgY totales purifiées sont mises en contact avec le gel.L’élution des IgY non fixées est
suivie par mesure de l’absorbance à 280 nm. Les IgY spécifiques sont alors éluées avec un
tampon glycine à pH 3.(Seddiki, 2009)

51
II.3.3 Electrophorèse sur gel polyacrylamide 12% en conditions
dénaturantes (SDS-PAGE)
Cette technique nous a permis d’analyser les fractions d’élution de la chromatographie
d’affinité en visualisant directement les protéines par coloration du gel au bleu de Coomassie.
Le SDS est un détergeant anionique qui se fixe sur les protéines et leur donne une charge
globale négative. Le même nombre de molécules de SDS se fixe par unité de masse de
protéines ; la charge des protéines est proportionnelle à leur masse moléculaire relative (Mr).
Soumis à un champ électrique, les complexes SDS-protéine migrent de la cathode vers
l’anode en fonction inverse de leur masse moléculaire relative.
Les fractions à déposer ont été préalablement diluées en tampon échantillon réducteur (voir
annexe), chauffées à 100°C pendant 3 minutes puis centrifugées. Les échantillons ont été
analysés sur gel à 12%. La migration a été faite à 10 mA dans le gel de concentration, et à 20
mA dans le gel de séparation. Le Magic Mark (Invitrogen) est un marqueur de taille allant de
12 000 à 20 000 et est visualisé lors de la révélation par luminescence.
II.3.4 Western blot et révélation par chimioluminescence
La qualité des IgY spécifiques est analysée par la technique du western blot. Des échantillons
contenant rSOx ou hQ6 sont soumis à une électrophorèse sur gel de polyacrylamide-SDS 12%
comme décrit précédemment. Après la migration, le gel est soumis à un électrotransfert en
milieu liquide sur membrane de nitrocellulose Hybond-ECL (Amersham Pharmacia Biotech)
avec le système de transfert Mini-Trans-Blot Cell (Bio-Rad) pendant 15 heures à 4°C, à 40 V
en tampon Tris 24 mM, glycine 192 mM, éthanol 20% (Générateur Bio-Rad Power Pac 3000).
Le gel de polyacrylamide-SDS est coloré au bleu de Coomassie et la membrane au rouge
Ponceau pour contrôler la qualité du transfert. La membrane est placée dans du rouge Ponceau
pendant 10 minutes, puis décolorée dans des bains successifs d’eau ultra-pure.
La membrane est ensuite incubée dans du tampon PBS (tampon phosphate de sodium 10 mM,
pH 7,4, NaCl 0,15 M) Tween 20 0,1%, lait écrémé déshydraté 10%, pendant une heure à 37°C
afin de saturer les sites de fixation non spécifiques. La membrane est ensuite incubée avec
37,5 µL de la fraction contenant les IgY spécifiques anti-sulfhydryl oxydase de rat, dilués dans
10 mL de tampon PBS, Tween 20 0,1%, lait 5% à température ambiante, pendant 1 heure sous
agitation. Puis, la membrane est lavée 3 fois 5 minutes dans le PBS, Tween 20 0,1%, et
incubée à température ambiante, pendant 1 heure sous agitation avec des IgG de lapin anti
IgY de poulet conjugués à la peroxydase (Sigma, A9046) dilués au 1/15000 dans du PBS,
Tween 20 0,1%, lait 5%. A nouveau, la membrane est lavée 3 fois 10 minutes. Les protéines

52
ayant fixé les anticorps sont alors révélés par chimioluminescence avec un kit substrat ECL
(Amersham Pharmacia Biotech). En présence d’H2O2, la peroxydase libère H2O +1/2O2.
L’oxygène oxyde le luminol en milieu alcalin et le luminol oxydé, instable, revient à son état
fondamental stable en émettant de la lumière à 428 nm qui va impressionner un film
photographique (précipitation des sels d’argent).
II.4 Mesure de l’activité oxydasique des sulfhydryl oxydases
II.4.1 Principe de dosage de l’oxydation du DTT par le DTNB
Pour réaliser cette mesure, nous avons utilisé comme substrat le dithiothréitol (DTT)
(Référence 708984 lot 85849121, Boehringer) qui possède deux groupements SH. La
sulfhydryl oxydase, mise en présence de DTT réduit, catalyse la formation d’un disulfide
interne suivant la réaction décrite dans la figure 16.
Figure 17 : Réaction d’oxydation du DTT catalysée par la sulfhydryl oxydase de rat.
L’oxydation du DTT réduit par la sulfhydryl oxydase conduit à la formation d’un pont
disulfure (catalague Boehringer).
La quantité de thiols libres présente dans le milieu est évaluée grâce au réactif des thiols
l’acide 5,5’-dithiobis 2-nitrobenzoïque (DTNB). Le DTNB (référence D-8130, lot 54H0479,
Sigma) réagit avec les thiols libres pour former un disulfide mixte libérant ainsi l’acide 5-thio-
2-nitrobenzoïque (TNB) (figure 17), chromophore qui possède un maximum d’absorption à
412 nm avec un coefficient d’extinction molaire de 13600 M-1
.cm-1
. Cette méthode nous
permet de mesurer la disparition des thiols libres présents dans le milieu réactionnel en
fonction du temps et de déduire la vitesse d’oxydation.
OH
OH
s
s
+ H2O2 + O2 Thiol
oxydase
DTT réduit DTT oxydé
SH OH
SH OH

53
+
Acide 5-thio-2-nitrobenzoïque Disulfide mixte
(Chromophore : coloration jaune)
Figure 18 : Réaction du DTNB avec les thiols libres.
II.4.2 Mesure de l’activité oxydasique des surnageants de culture
Les mesures de l’activité oxydasique sur les surnageants de culture concentrés ont été
effectuées dans les conditions suivantes:
Surnageant de culture
Concentré 20 fois
Témoin sulfhydryl
oxydase (rSOx)
Témoin DTT
Volume de tampon 34,6 µL 37,6 µL 39,6 µL
Volume de rSOx 0 µL 2 µL 0 µL
Volume d’échantillon 5 µL 0µL 0 µL
Volume de DTT 200 mM 0.4 µL 0.4 µL 0.4 µL
Volume total 40 µL 40 µL 40 µL
La sulfhydryl oxydase purifiée à partir des sécrétions des vésicules séminales de rat a été
utilisée comme témoin d’activité (6,6 10-7
M). Un témoin DTT seul et un témoin DTNB sont
réalisés lors de chaque étude d’activité. La concentration finale en DTT est 2 mM. Le tampon
utilisé est un tampon Tris 0,1 M, pH 8,14, NaCl 0,15 M. Chaque mélange réactionnel est
incubé au bain-marie à 25°C et nous avons prélevé 5 µL de milieu réactionnel à différents
temps. Les 5 µL alors sont ajoutés immédiatement à un mélange préparé extemporanément de
600 µL de tampon (tampon Tris 0,1 M, pH 8,14, NaCl 0,15 M) et 7 µL de DTNB 20 mM
(référence D-8130, lot 54H04019, SIGMA). L’absorbance à 412 nm est mesurée avec un
spectrophotomètre CARY 100 SCAN.
+ R-SH H
R
H
O
H
H
DTNB

54
II.4.3 Analyse de l’activité protéasique à cystéine
Afin de tester une éventuelle présence d’activité protéasique à cystéine dans les
surnageants potentiellement activé par l’incubation avec le substrat DTT 2mM, nous avons
utilisé le substrat fluorigénique Z-FR-AMC (Benzyloxycarbonyl-L-phénylalanyl-L-arginyl-7-
amino 4 méthyl-coumarine) (Calbiochem). L’activité a été mesurée par la fluorescence
détectée due à la libération le fluorophore AMC. La fluorescence est mesurée avec un
spectrofluomètre, lecteur de microplaques Spectramax Gemini (Molecular Devices, France).
Les mesures ont été effectuées avec une microplaque de 96 puits contenant par puits 100 µL
de tampon d’acétate de sodium 0,2 M, DTT 4 mM, pH 5,0 auquel est rajouté 80 µL
d’échantillon de surnageants non concentrés, puis 25 µL de substrat (10 mM dilué dans le
DMF (diméthylformamide). L’incubation s’étend sur deux heures et la lecture s’effectue toutes
les 30 secondes à une longueur d’onde d’excitation de 350 nm et une longueur d’onde
d’émission de 460 nm. Le surnageant utilisé pour ce test provient de la lignée cellulaire H1838
issus d’adénocarcinome pulmonaire aux différents temps, T1, T2, T3, le témoin est le milieu
seul sans SVF. Les temps T1, T2 et T3 correspondent aux temps de 3, 5 et 7 jours après
ensemencement respectivement.
II.5 Analyses statistiques
L’analyse statistique a été réalisée par le test one-way ANOVA et post test de Tukey's
Multiple Comparaison Test : L’activité oxydasique des SOx/Q6 dans les différents surnageants
en fonction du temps de culture des cellules et pour la comparaison entre les résultats de
l’activité oxydasique entre les lignées cellulaires.

55
RESULTATS

56
Résultats et discussion de l’étude
épidémiologique

57
I.1.L’étude épidémiologique du cancer pulmonaire de la région
oranaise
Une étude rétrospective a porté sur 173 patients atteints d’un cancer du poumon et traités au
service de radiothérapie du CHU Oran, entre Janvier 2004 et Décembre 2007.
I.1.La mise en évidence des données épidémiologiques
I.1.1.La répartition du cancer pulmonaire en fonction la localisation
La nature a fait que l’Homme est une paire de poumon. Cependant la taille des poumons n’est
pas identique (figure 2), le poumon gauche est plus petit que le poumon droit.
Dans cette optique nous avons étudié lequel des deux poumons est le plus touché par le cancer
chez les patients. Les résultats montrent que 57% le poumon droit atteint et 43% pour le
poumon gauche (figure19). On n’a pas pu obtenir une hypothèse convaincante pour expliquer
cette tendance.
Figure 19: Répartition des cas recensés en fonction de la localisation
du cancer pulmonaire.
I.1.2.La répartition des cas étudiés en fonction des tranches d’âge
Les données recueillies du service radiothérapie montrent que le plus grand pourcentage
(30%) des patients malades fait parti de la tranche d’âge de 50 et 60 ans. Ce qui correspond à
une durée de 25 à 35 ans de tabagisme, bien sûre si on considère que le facteur inducteur est la
fumée. En revanche les jeunes moins de 40 ans, la pathologie est moins observée avec 4% ce
qui pourrait s’expliquer par une exposition à la fumée secondaire (tabagisme passif) puis le
tabagisme, soit une exposition à un autre facteur cancérigène, ou même une prédisposition à ce
type de cancer (tableau 3, figure 20).
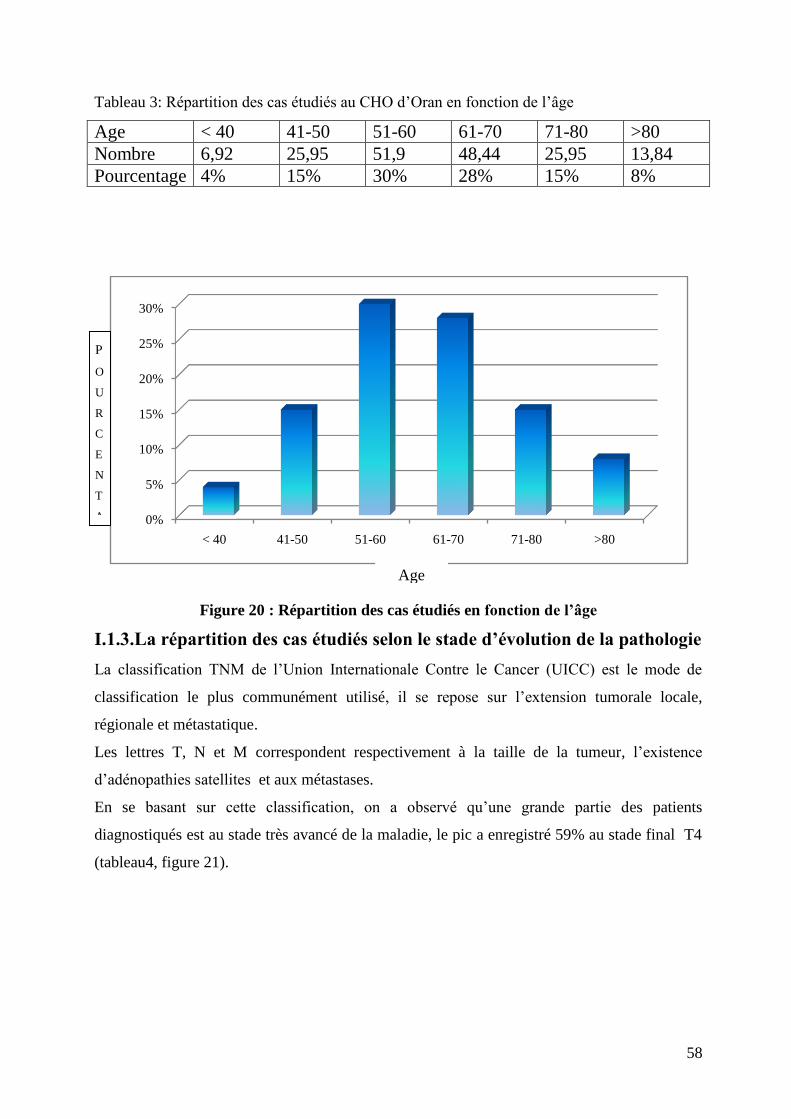
58
Tableau 3: Répartition des cas étudiés au CHO d’Oran en fonction de l’âge
Age < 40 41-50 51-60 61-70 71-80 >80
Nombre 6,92 25,95 51,9 48,44 25,95 13,84
Pourcentage 4% 15% 30% 28% 15% 8%
Figure 20 : Répartition des cas étudiés en fonction de l’âge
I.1.3.La répartition des cas étudiés selon le stade d’évolution de la pathologie
La classification TNM de l’Union Internationale Contre le Cancer (UICC) est le mode de
classification le plus communément utilisé, il se repose sur l’extension tumorale locale,
régionale et métastatique.
Les lettres T, N et M correspondent respectivement à la taille de la tumeur, l’existence
d’adénopathies satellites et aux métastases.
En se basant sur cette classification, on a observé qu’une grande partie des patients
diagnostiqués est au stade très avancé de la maladie, le pic a enregistré 59% au stade final T4
(tableau4, figure 21).
0%
5%
10%
15%
20%
25%
30%
< 40 41-50 51-60 61-70 71-80 >80
Age
P
O
U
R
C
E
N
T
A
G
E
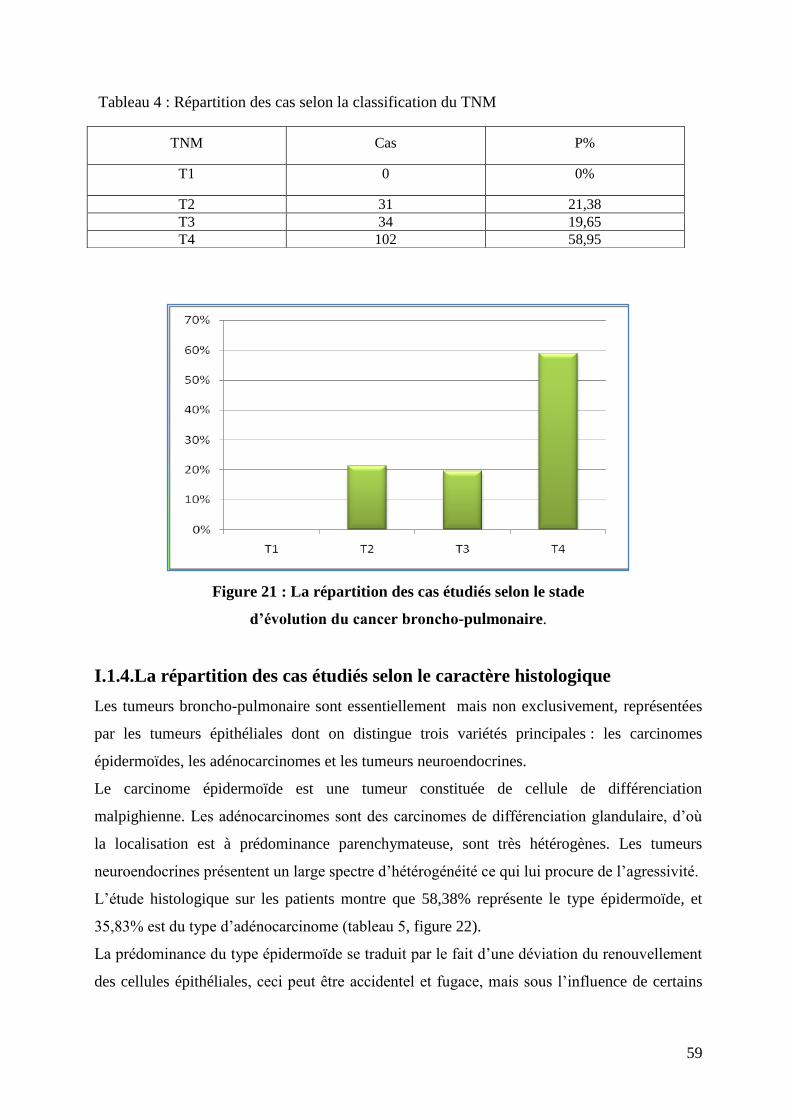
59
Tableau 4 : Répartition des cas selon la classification du TNM
Figure 21 : La répartition des cas étudiés selon le stade
d’évolution du cancer broncho-pulmonaire.
I.1.4.La répartition des cas étudiés selon le caractère histologique
Les tumeurs broncho-pulmonaire sont essentiellement mais non exclusivement, représentées
par les tumeurs épithéliales dont on distingue trois variétés principales : les carcinomes
épidermoïdes, les adénocarcinomes et les tumeurs neuroendocrines.
Le carcinome épidermoïde est une tumeur constituée de cellule de différenciation
malpighienne. Les adénocarcinomes sont des carcinomes de différenciation glandulaire, d’où
la localisation est à prédominance parenchymateuse, sont très hétérogènes. Les tumeurs
neuroendocrines présentent un large spectre d’hétérogénéité ce qui lui procure de l’agressivité.
L’étude histologique sur les patients montre que 58,38% représente le type épidermoïde, et
35,83% est du type d’adénocarcinome (tableau 5, figure 22).
La prédominance du type épidermoïde se traduit par le fait d’une déviation du renouvellement
des cellules épithéliales, ceci peut être accidentel et fugace, mais sous l’influence de certains
TNM Cas P%
T1 0 0%
T2 31 21,38
T3 34 19,65
T4 102 58,95
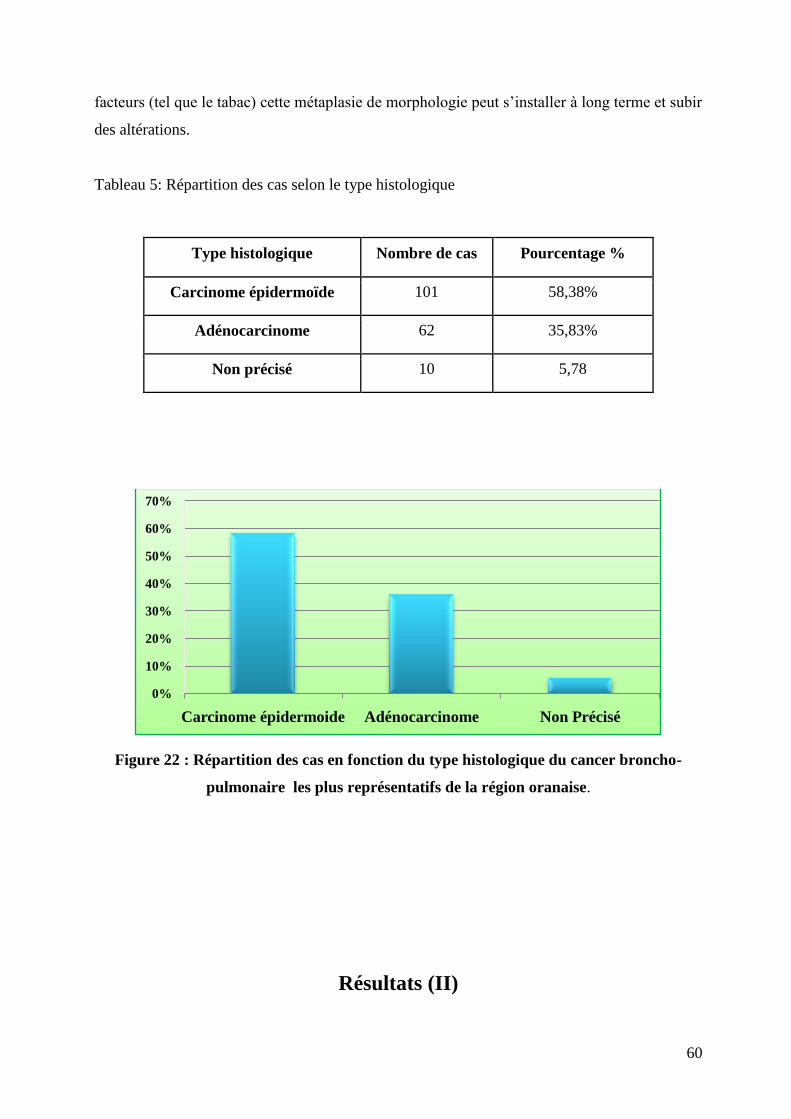
60
facteurs (tel que le tabac) cette métaplasie de morphologie peut s’installer à long terme et subir
des altérations.
Tableau 5: Répartition des cas selon le type histologique
Type histologique Nombre de cas Pourcentage %
Carcinome épidermoïde 101 58,38%
Adénocarcinome 62 35,83%
Non précisé 10 5,78
Figure 22 : Répartition des cas en fonction du type histologique du cancer broncho-
pulmonaire les plus représentatifs de la région oranaise.
Résultats (II)
0%
10%
20%
30%
40%
50%
60%
70%
Carcinome épidermoide Adénocarcinome Non Précisé

61
II.1 Purification des anti-corps IgY spécifiques de la sulfhydryl oxydase de rat
A partir des œufs 399 et 400, nous avons extrait respectivement 52,8 mg et 23,37 mg d’IgY.
Les IgY spécifiques représentent entre 1 et 2 % de ces IgY totales soit entre 528 µg et 1 mg
pour l’œuf 399.
Nous avons entrepris la purification des IgY spécifiques anti-rSOx à partir de l’extrait de l’œuf
399 par chromatographie d’affinité sur le gel rSOx-Sepharose. Le profil d’élution à 280 nm
montre 2 pics de protéines. Un pic correspondant aux IgY non retenues et un deuxième pic
correspondant aux IgY spécifiques (figure 23).
Les interactions antigène- anticorps sont affaiblies par l’exposition à un pH acide, ce principe
est souvent utilisé pour la dissociation des complexes antigène-anticorps lors des
chromatographies d’affinité.
Cependant, la littérature nous indique qu’une instabilité particulière des IgY à pH acide
pouvait entraîner leur dénaturation. Au cours du protocole le risque de dénaturation existe
aussi bien pour les IgY spécifiques éluées que la rSOx couplée au gel. Nous avons choisi des
conditions qui permettaient de stabiliser très rapidement ces protéines et nous avons utilisé un
sucre très soluble, le D(+) tréhalose connu pour sa propriété stabilisatrice des protéines à des
concentrations relativement modérées [43].
Nous avons coloré ces fractions après électrophorèse en gel de polyacrylamide au bleu de
Coomassie. Nous avons ainsi pu sélectionner les fractions fixées analysées (figure 24) pour la
détection spécifique de la sulfhydryl oxydase. Nous avons aussi déterminé le seuil de détection
de nos IgY purifiées, il apparaît que les anticorps peuvent reconnaître une quantité de
sulfhydryl oxydase de rat de 5 ng (figure 25).
Par western blot, les résultats montrent que les anticorps spécifiques IgY rSOx sont capables
de reconnaître la SOx/Q6 humaine dans les surnageants de fibroblaste pulmonaire WI38 (Mr
84000) et dans le liquide séminal (Mr 64000) (figure 26). On peut donc considérer que la
bande de Mr 64000 observée dans le liquide séminal et la bande Mr 84000 observée dans les
surnageants de culture des fibroblastes WI38 correspondent respectivement aux formes courte
et longue de la quiescine Q6 humaine. Cependant, les résultats indiquent aussi que les
anticorps IgY spécifiques non seulement ont la capacité de reconnaître la quiescine Q6
humaine mais aussi la quiescine Q6 bovine présente dans le SVF et qui apparaît à une masse.

62
60000 80000
1 2 3 4 5 6 7 8 9
Figure 23: Purification des IgY anti-rSOx spécifiques par chromatographie d’affinité. Les IgY
extraites du jaune d’œuf de poule, reprises dans 9 ml de tampon phosphate, ont été ajoutées au gel de
Sepharose couplé avec rSOx sous agitation pendant 2 heures après lavage en tampon borate. Les IgY
spécifiques ont été éluées avec du tampon glycine 100 mM, pH 3,0 contenant 15% de D(+) tréhalose et
neutralisées immédiatement avec du Tris 1M, pH 8,0.
Figure 24: Analyse de la pureté des IgY spécifiques par affinité sur gel de polyacrylamide à 12%.
Le gel de polyacrylamide à 12% coloré en bleu de Coomassie. Puits 1: étalon MagicMark, puits 2
fraction non fixée n°1 (12 µl), puits 3 fraction non fixée n° 21 (12 µl), puits 4), 5), 6) ,7), 8) fractions
fixées 1, 2, 3, 4, 5 (12 µl), puits 9 IgY totales (1µl).
Purification d'Igy spécifique rSOx
0
0,5
1
1,5
2
2,5
10 30 50 70 90
volume
DO
à 2
80 Tp glycine
pH 3

63
moléculaire relative de 84000 (figure 27) et peut potentiellement interférer avec les mesures
d’activité oxydasiques.
II.2 Analyse des activités oxydasiques des surnageants de culture cellulaire
Nous avons déterminé l’activité oxydasique dans les surnageants de culture de différentes
lignées cellulaires tumorales pulmonaires en fonction de l’état de confluence.
En raison des faibles quantités de sulfhydryl oxydase sécrétées par les cellules en culture nous
avons dû concentrer 20 fois chaque échantillon avant de procéder à l’analyse de son activité.
Nous avons employé une méthode colorimétrique avec le DTNB, celui-ci réagit avec les thiols
libres présents dans le milieu réactionnel. En présence de sulfhydryl oxydase, le DTT à 2 mM
disparaît progressivement du milieu réactionnel. La pente de la représentation de la
concentration en thiols libres en fonction du temps est proportionnelle à l’activité oxydasique
de l’échantillon.
Dans un premier temps, nous avons analysé l’activité de la sulfhydryl oxydase de rat (rSOx)
(6,6 10-7
M), enzyme purifiée à partir du liquide des vésicules séminales de rat. Nous
atteignons une oxydation totale du DTT au bout de 20 minutes (figure 26). Cette expérience
nous sert de témoin vis-à-vis de l’activité sulfhydryl oxydase sécrétée par les différentes
lignées cellulaires tumorales.

64
Figure 25 : Analyse par western blot du seuil de détection de rSOx par les IgY spécifiques.
1) étalon MagicMark, puits 2), 3), 4), 5) rSOx à des dépôts de 40, 20, 10, 5 ng respectivement.
Figure 26: Analyse par Western blot de la reconnaissance de la sulfhydryl oxydase/quiescine Q6
humaine par les IgY spécifiques. 1): étalon MagicMark, 2) rSOx purifiée (0,04µg), 3) liquide séminal
humain (3 µL), et 4) surnageant de culture des fibroblastes WI38 concentré 20 fois (3µL).
Figure 27: Analyse par Western blot de la présence de sulfhydryl oxydase dans le sérum de veau
fœtal. 1), 7 µL de surnageant de culture des fibroblastes WI38 concentré 20 fois, 2) milieu de culture
RPMI contenant 10% de SVF concentré 20 fois (7µL), 3) SVF pur (2µL).
66000
Puits 1 2 3 4 5
Puits 1 2 3 4
66000
84000
Puits 1 2 3
84000

65
rSOx
0
0,1
0,2
0,3
0,4
0,5
0,6
0 10 20 30
Temps (minutes)
A 4
12
nm
Figure 28: Activité oxydasique de la sulfhydryl oxydase de rat. La sulfhydryl oxydase de rat a été
incubée avec 2 mM de DTT, à 25°C. la concentration en thiols est exprimé en absorbance (A) à 412
nm.
Figure 29 : Activité oxydasique du milieu de culture contenant le sérum de veau fœtal. Analyse de
l’activité oxydasique dans le SVF seul en fonction des temps d’incubation T0, T1, T2, T3. 5µL de SVF
concentré 20 fois a été incubé avec 2 mM de DTT, à 25°C. L’activité est exprimée en ∆A412
nm/minute. Les T1, T2 et T3 correspondent aux temps d’incubation de 3, 5 et 7 jours respectivement.
(= p<0,01 ; = p<0,001).
T0 T1 T2 T30.0000
0.0005
0.0010
0.0015
0.0020
0.0025
0.0030
0.0035
Temps (Jours)
A
412 n
m/
min
ute
SVF
***
***
**

66
Le milieu de culture des cellules contient 10% de SVF, nous avons donc avant d’exploiter les
résultats, mesuré l’activité oxydasique du SVF. Pour cela nous avons incubé le SVF dans les
mêmes conditions que les milieux conditionnés par les lignées cellulaires soit en étuve à CO2
à 37°C pendant 3, 5 et 7 jours. La présence d’une activité oxydasique dans le SVF a été
observée, cette activité présente une tendance à baisser avec le temps d’incubation, cette
baisse s’est avéré très significatif (p<0,001) (figure 29). Au vu de cette baisse d’activité au
cours du temps, nous avons effectué un contrôle dans le but de vérifier si l’activité sulfhydryl
oxydase dans les surnageants pouvait être dégradée par des protéases présentes en utilisant le
substrat Z-FR-AMC. C’est un substrat qui est clivé en particulier par les protéases à cystéine.
Le substrat a été rajouté aux surnageants de la lignée cellulaire H1838, et au milieu de culture
seul. Après incubation, les courbes de fluorescence obtenues montrent une absence d’activité
protéasique dans les surnageants analysés. Cette activité associée au milieu de culture a été
soustraite des valeurs d’activité oxydasique obtenues pour les surnageants de culture pour
obtenir une valeur plus représentative de la sécrétion de SOx/Q6 cellulaire.
Les résultats montrent que l’activité de la sulfhydryl oxydase sécrétée dans les surnageants
de culture cellulaire des lignées WI38, Calu-1, H1838, H358, H460, H23, A549, H520
(figure 30 et 31) présente une augmentation en fonction du temps d’incubation, indiquant une
capacité de synthèse et de sécrétion conservée pour les différentes lignées.
Une comparaison de la sécrétion et d’activité oxydasique entre les lignées cellulaires : WI38
et les autres lignées cellulaires est de significatif soit très significatif. D’autre part la
comparaison entre les lignées H358 et H1838 avec les autres lignées les résultats sont
également entre significatif et très significatif (p<0,05 et p<0,001, n=3).

67
Figure 30: Les activités oxydasiques corrigées des surnageants des lignées cellulaires
pulmonaires. Les valeurs d’activité oxydasique du SVF sont soustraites des valeurs brutes des
surnageants de cultures cellulaires. Les lignées cellulaire ont été incubées à 37°C soit à un temps de 3
jours (T'1), soit 5 jours (T'2), soit à 7 jours (T'3) dans du milieu culture contenant 10% de SVF. Les
activités oxydasiques sont exprimées ∆ A412nm/ minute (=p<0,05 ;= p<0,01 ; =
p<0,001).
H1838
T'1 T'2 T'30.000
0.001
0.002
0.003
0.004
0.005
***
***
Temps (Jours)
A
412 n
m/
min
T'1 T'2 T'30.000
0.005
0.010
0.015
0.020
WI38
***
***
Temps (Jours)
A
412 n
m/
min
H23
T'1 T'2 T'30.000
0.001
0.002
0.003
0.004
0.005
0.006
0.007
Temps (Jours)
A
412 n
m/
min
T'1 T'2 T'30.000
0.001
0.002
0.003
0.004
0.005
H549
***
Temps (Jours)
A
412 n
m/
min
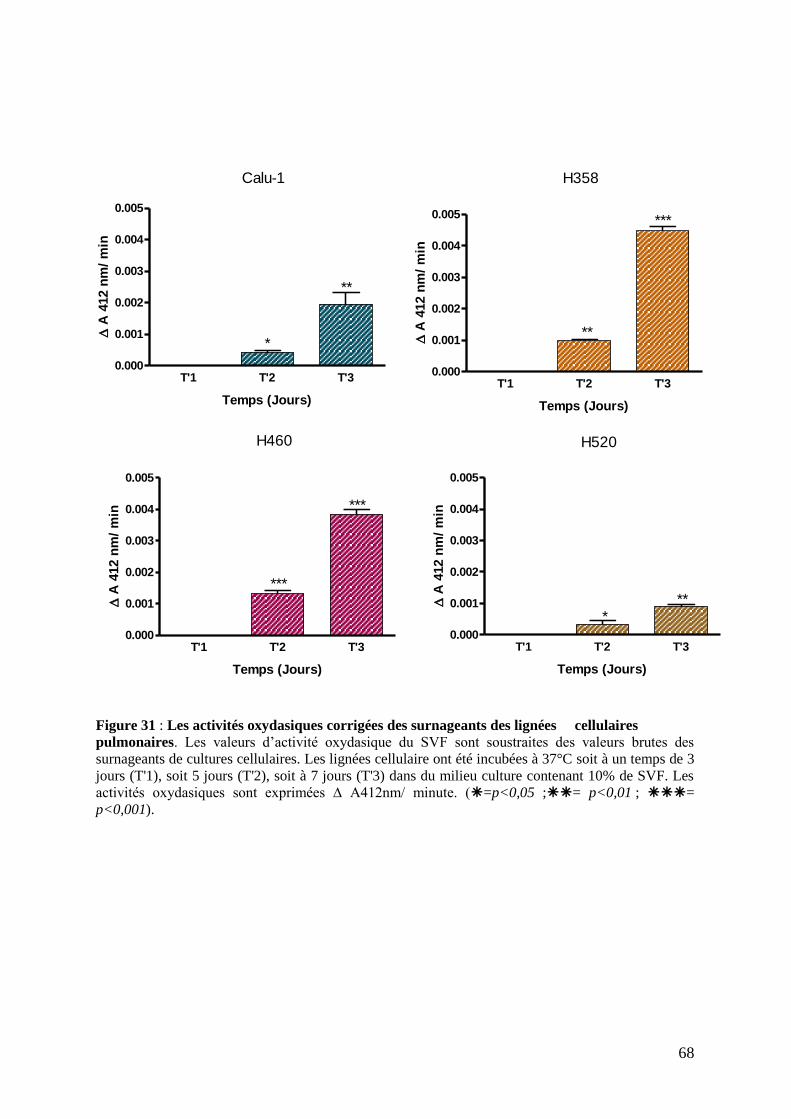
68
Figure 31 : Les activités oxydasiques corrigées des surnageants des lignées cellulaires
pulmonaires. Les valeurs d’activité oxydasique du SVF sont soustraites des valeurs brutes des
surnageants de cultures cellulaires. Les lignées cellulaire ont été incubées à 37°C soit à un temps de 3
jours (T'1), soit 5 jours (T'2), soit à 7 jours (T'3) dans du milieu culture contenant 10% de SVF. Les
activités oxydasiques sont exprimées ∆ A412nm/ minute. (=p<0,05 ;= p<0,01 ; =
p<0,001).
H460
T'1 T'2 T'30.000
0.001
0.002
0.003
0.004
0.005
***
***
Temps (Jours)
A
412 n
m/
min
H520
T'1 T'2 T'30.000
0.001
0.002
0.003
0.004
0.005
***
Temps (Jours)
A
412 n
m/
min
Calu-1
T'1 T'2 T'30.000
0.001
0.002
0.003
0.004
0.005
*
**
Temps (Jours)
A
412 n
m/
min
H358
T'1 T'2 T'30.000
0.001
0.002
0.003
0.004
0.005
**
***
Temps (Jours)
A 4
12 n
m/
min

69
DISCUSSION
Des résultats de la sulfhydryl oxydase

70
Discussion des résultats de la sulfhydryl oxydase
Le peu d’étude effectuée sur la protéine SOx/Q6 montre son implication dans le cancer
broncho-pulmonaire non à petites cellules. Dans ce contexte, le but de ce travail est de
vérifier l’expression de SOx/Q6 dans les lignées tumorales pulmonaires et analyser le niveau
de sécrétion de la SOx/Q6 humaine à différents stades de prolifération par mesure de
l’activité oxydasique.
Les sulfhydryl oxydases FAD dépendante, ont été identifiées et caractérisées par leur activité
oxydasique dans divers tissus et en particulier dans les vésicules séminales de rat [44]. Leur
identification avec la quiescine Q6 a été plus récente [45], [38]. Les quiescines sont des gènes
qui s’expriment dans la lignée fibroblastique pulmonaire humaine WI38 lors de son entrée en
quiescence [39]. Parmi ces gènes se trouvent des gènes de chaînes de protéines de la matrice
extracellulaire (collagènes, décorine) ainsi la SOx/Q6. Un second ADNc issus d’une
expression pulmonaire a aussi été décrit et nommé ″Cell Growth Inhibiting Factor″ (CGIF).
Ces deux ADNc correspondent à deux formes d’épissage issues du gène SOx/Q6. L’ADNc
quiescine Q6 correspond à la forme longue codant pour une protéine de Mr 80000 et l’ADNc
CGIF correspond à la forme courte codant pour une protéine de Mr 66000.
De nombreuses questions restent à résoudre concernant ces enzymes, celle de leurs fonctions
cellulaires et celle de leur implication dans les pathologies pulmonaires. Il est mené en
parallèle avec des études sur l’expression des messagers dans ces mêmes lignées, ainsi que
des études sur l’implication de ce gène dans le cancer broncho-pulmonaire au sein du groupe
thématique. A l’aide des anticorps anti-sulfhydryl oxydase de rat obtenus chez la poule et
purifiés, nous avons pu mettre en évidence les deux formes protéique de quiescine Q6
humaine par immunodétection, soit la forme longue (Mr 84000) dans le surnageant de
fibroblaste pulmonaire WI38, soit la forme courte (Mr 66000) dans le liquide séminal
humain. Ces anticorps sont capables de reconnaître les deux formes de la SOx/Q6 qui ne
semblent pas co-exprimées dans les mêmes échantillons biologiques.
Par ailleurs, en utilisant les anticorps purifiés, nous avons pu détecter une forme longue de
Mr proche de 84000 présentes dans le SVF. Cette observation corrèle avec les résultats déjà
décrits dans la littérature [46]. Ces résultats suggèrent aussi que la sulfhydryl oxydase bovine
porte des épitopes communs avec les sulfhydryls oxydases/ quiescine Q6 humaine et de rat,
ce qui est représentatifs de l’homologie que possèdent entre eux les différents membres de la
famille des SOx/Q6. Cependant, la présence de SOx/Q6 bovine dans le SVF risque
d’interférer avec le dosage de l’activité oxydasique sécrété, par les cellules.

71
Nous avons effectivement observé au cours des expériences que le sérum de veau fœtal seul
présente une faible activité oxydasique avec le substrat DTT. Cette activité décroît avec le
temps d’incubation en étuve à CO2. Nous n’avons observé aucune activité protéolytique dans
les surnageants non concentrés issus de la lignée cellulaire ce qui suggère que la protéolyse
ne semble pas influencer sur le niveau d’activité oxydasique mesuré.
La présence de la sulfhydryl oxydase/ quiescine Q6 bovine dans le milieu de culture à 10 %
de SVF interfère cependant avec les mesures d’activité de SOx/Q6 sécrétée par les cellules.
Pour cette raison nous avons soustrait l’activité oxydasique des milieux de culture des
activités brutes pour faire ressortir l’activité propre des lignées cellulaires.
Nous avons observé une accumulation progressive de l’activité oxydasique en fonction du
temps d’incubation. Cependant, les niveaux d’activités sont variables en fonction des lignées
cellulaires. Les lignées cellulaires tumorales pulmonaires sécrètent de façon nettement plus
faible que les fibroblastes WI38 (p<0,001) (figure 32). Cependant la comparaison de
l’activité oxydasique entre les lignées cellulaires montre que les H358 et H1838 la plus forte
sécrétion au stade 3 de façon significatif (p<0,05) à très significatif (p<0,01;p<0,001)
(figure 33).
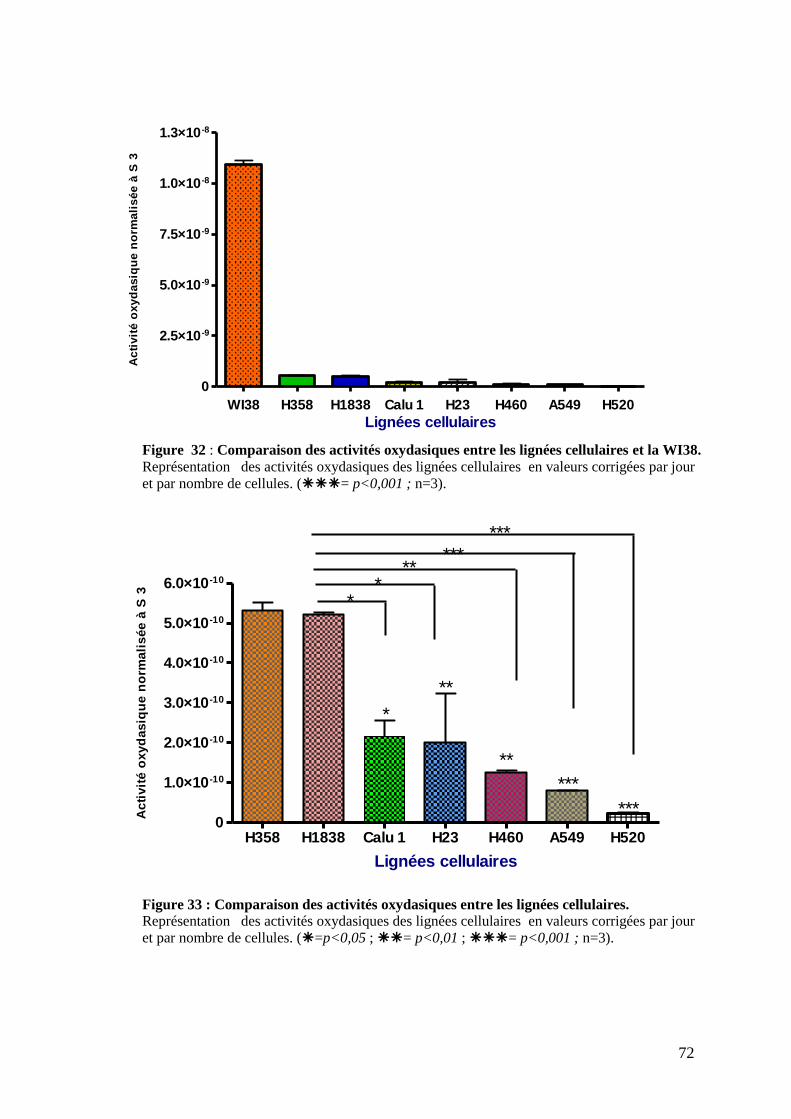
72
Figure 32 : Comparaison des activités oxydasiques entre les lignées cellulaires et la WI38.
Représentation des activités oxydasiques des lignées cellulaires en valeurs corrigées par jour
et par nombre de cellules. (= p<0,001 ; n=3).
Figure 33 : Comparaison des activités oxydasiques entre les lignées cellulaires.
Représentation des activités oxydasiques des lignées cellulaires en valeurs corrigées par jour
et par nombre de cellules. (=p<0,05 ; = p<0,01 ; = p<0,001 ; n=3).
WI38 H358 H1838 Calu 1 H23 H460 A549 H520
0
2.5×10-9
5.0×10-9
7.5×10-9
1.0×10-8
1.3×10-8
Lignées cellulaires
Ac
tivit
é o
xyd
asiq
ue n
orm
ali
sée à
S 3
H358 H1838 Calu 1 H23 H460 A549 H5200
1.0×10-10
2.0×10-10
3.0×10-10
4.0×10-10
5.0×10-10
6.0×10-10
*
**
**
***
***
**
*****
***
Lignées cellulaires
Ac
tivit
é o
xyd
asiq
ue n
orm
ali
sée à
S 3
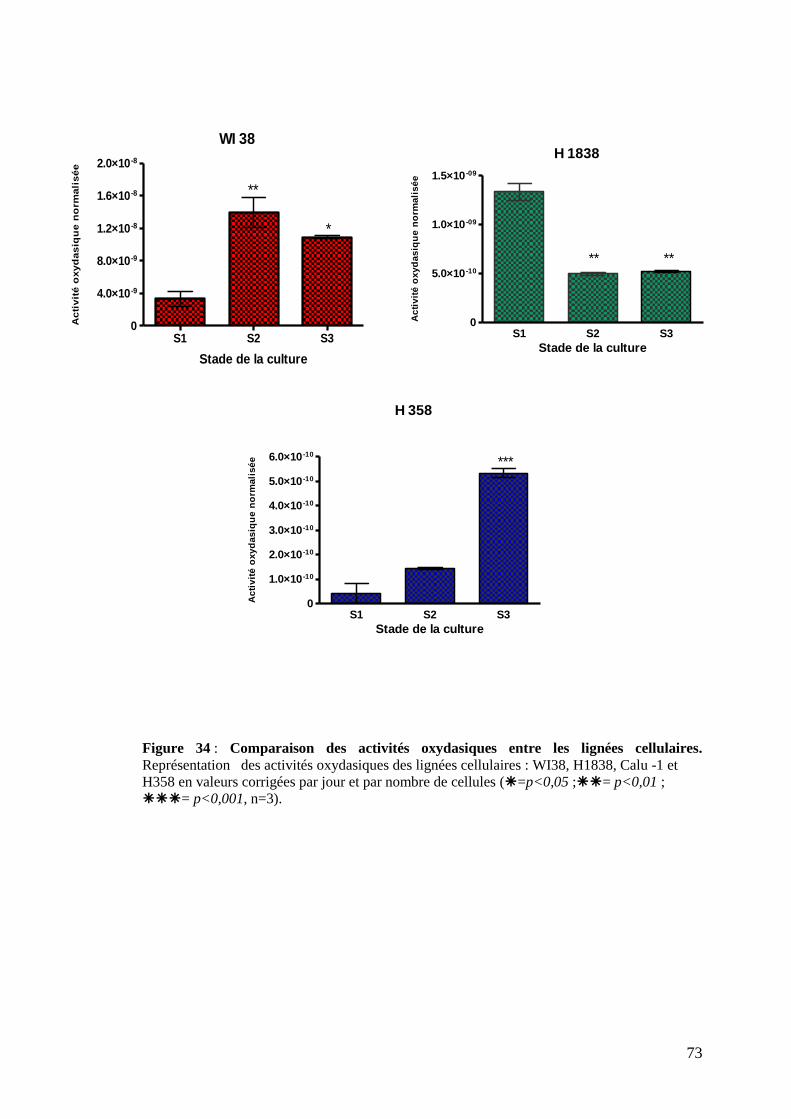
73
Figure 34 : Comparaison des activités oxydasiques entre les lignées cellulaires.
Représentation des activités oxydasiques des lignées cellulaires : WI38, H1838, Calu -1 et
H358 en valeurs corrigées par jour et par nombre de cellules (=p<0,05 ;= p<0,01 ;
= p<0,001, n=3).
WI 38
S1 S2 S30
4.0×10-9
8.0×10-9
1.2×10-8
1.6×10-8
2.0×10-8
**
*
Stade de la culture
Ac
tiv
ité
ox
yd
as
iqu
e n
orm
ali
sé
e
H 1838
S1 S2 S30
5.0×10-10
1.0×10-09
1.5×10-09
** **
Stade de la culture
Ac
tiv
ité o
xyd
as
iqu
e n
orm
ali
sé
eH 358
S1 S2 S30
1.0×10-10
2.0×10-10
3.0×10-10
4.0×10-10
5.0×10-10
6.0×10-10
***
Stade de la culture
Ac
tiv
ité o
xy
das
iqu
e n
orm
ali
sé
e

74
La présentation de l’accumulation de l’activité au cours du temps a été normalisée par jour et
par le nombre de cellules afin d’essayer de voir si la production était sensible à l’état de
passage à l’état de quiescence.
Nous avons pu déterminer trois comportements de sécrétion de l’activité oxydasique chez les
cellules étudiées. Un comportement observé chez la lignée cellulaire WI38 où la sécrétion est
induite par la confluence, un comportement présenté par les lignées H1838, H23 et Calu1,
caractérisé par une forte sécrétion au moment de la croissance. Alors que les lignées cellulaires
H358, H460, H520, A549 enregistrent une augmentation importante de la sécrétion post-
confluence (figure 34).
On peut supposer que le moment de sécrétion de SOx/Q6 et la modulation de son niveau
diffèrent d’un groupe de lignées cellulaires à un autre. Un groupe a tendance à une sécrétion
liée à la prolifération, un groupe a tendance à une sécrétion régulée par la confluence comme
les fibroblastes WI38, un autre a tendance à une sécrétion plus tardive comme les cellules
H358 après confluence. Enfin, un troisième groupe semble avoir perdu la modulation de la
sécrétion vis-à-vis de la confluence. Ces résultats suggèrent probablement que l’expression de
SOx/Q6 est au besoin de la cellule de produire des protéines contenants un ou plusieurs ponts
disulfures. Dans l’intestin grêle, par immunohistologie, les cellules neuroendocrines qui
sécrètent des peptides contenant des ponts disulfures, présentent un marquage de QSOx1
intense [47].

75
CONCLUSION ET PERSPECTIVES

76
Conclusion et perspectives
Toutes les études ont confirmées l’effet néfaste du tabac sur la santé du fumeur et sur son
entourage par le tabagisme passif. La fumée de la cigarette et la pollution induisent des
dérèglements du métabolisme cellulaire engendrant ainsi le cancer broncho-pulmonaire.
Les données épidémiologiques de la littérature indiquent que le pourcentage du cancer
broncho-pulmonaire est en croissance continue. A Oran, le pourcentage du cancer broncho-
pulmonaire est de 17,5 % parmi les différents types de cancers. Une estimation de 88 cas
annuel est enregistrée à Oran toute seule, dans un état avancé de la maladie, de plus la durée de
survie est estimée de 5 ans.
Pour une meilleure survie des patients, il faut améliorer leurs prise en charge la plus adaptée
possible aux différentes caractéristiques des patients. Celle-ci repose sur une analyse précise
des facteurs pronostiques.
Dans cette optique, nous avons vérifié l’existence de la protéine SOx/Q6 dans les surnageants
des différentes lignées cellulaires broncho-pulmonaires et les variétés de cette protéine, ainsi
que sa production et son activité extracellulaire.
Les résultats obtenus d’activité sont en corrélation avec les résultats de western blot montrent
que la SOx/Q6 essentiellement la forme longue contribue à l’activité oxydasique dans les
surnageants, issus des lignées broncho-pulmonaire. Cependant, on ne peut pas exclure la
sécrétion d’autres sulfhydryl oxydases. Nous somme donc en présence d’une protéine
d’expression ubiquitaire même dans les lignées tumorales pulmonaires dont il faudra envisager
l’inactivation de la SOx/Q6 pour aborder le rôle cellulaire.
Nous envisageons donc d’étudier l’effet de l’inactivation par siRNA de SOx/Q6 sur le
comportement des lignées tumorales pulmonaires (migration, invasion par exemple). Une autre
étape consistera à produire les mutants des domaines thiorédoxine et ERV de SOx/Q6. Enfin,
la compréhension de la non-expression de la forme courte dans ces cellules tumorales
pulmonaires humaines contribuera à la compréhension des mécanismes de régulation de
l’expression des SOx/Q6 et à l’analyse de leur implication dans le cancer broncho-pulmonaire.

77
REFERENCES BIBLIOGRAPHIQUES

78
Références bibliographiques
[1] Quoix E. Novel epidemiology in lung cancer - non-smokers, women and cannabis] Rev
Mal Respir. 2007 Oct;24(8 Pt 2):6S10-5.
[2] Nacht M., Dracheva T., Gao Y., Fujii T., Chen Y., Player A., Akmaev V., Cook B.,
Dufault M., Zhang M., Zhang W., Guo M., Curran J., Han S., Sidransky D., Buetow K.,
Madden SL., Jen J.(2001) Molecular characteristics of non-small cell lung cancer. Proc Natl
Acad Sci U S A., 98(26):15203-8.
[3] Rouëssé J, Turpin F. 1994. Abrégé oncologie. 3 édition Masson.
[5] Trédaniel Jean. Le Cancer du Poumon, première edition, Paris Masson 2004, pp 11.
[6] Vogelstein B, Kinzler KW. Cancer genes and the pathways they control. Nat Med. 2004.
Aug;10(8):789-99.
[7] Doll R, Peto R. Cigarette smoking and bronchial carcinoma : dose and time relationships
among regular smokers and lifelong non smokers. J Epidemiol Commun Health, 1978, 32,
303-313.
[8] Stewart BW, Kleihues P. WHO. World Cancer Report. Lyon, 2003.
[9] Yaker A. Profile de morbidité cancéreuse en Algérie de 1966- 1975.SNED Médecine.
1975. 6; 4-47
[10] Wynder EL, Graham EA. Tobacco smoking as a possible etiologic factor in bronchogenic
carcinoma : a study of six hundred and heighty-four proved cases. JAMA, 1950, 143, 329-346.
[11] Churg A. Lung cancer cell typeand occupationnal exposure, in Samet Jm (ed) :
Epidemiology of lung lung cancer. New York, Marcel Dekker, 1994. pp 413-436
[12] Arveux P. Les cancers de la trachée, des bronches et du poumon. In : Salem G, Rican S,
Jougla E. Atlas de la santé en France vol 1- les causes de décès. John Libbey eurotext, paris
1999. 72-74.
[13] Eco-santé OCDE (2007). Les statistiques de mortalité sont extraites de la base de
données sur la mortalité de l’OMS, et sont normalisées par rapport à l’âge en fonction de la
structure de la population des pays de l’OCDE en 1980.
[14] Charloux A., Quoix E, Wolkove N, et al.— The increasing incidence of lung
adenocarcinoma : reality or artefact ? A review of the epidemiology of lung adenocarcinoma.
Int J Epidemiol, 1997. 26, 14-23.
[15] Alberg AJ, Brock MV, Samet JM. Epidemiology of lung cancer : looking to the future. J
clin oncol, 2005. 23, 3175-3185.

79
[16] Quoix E. Bronchial epidermoid cancers and adenocarcinomas. Epidemiology, pathology,
etiology, diagnosis, evolution, treatment Rev Prat. 2000. Dec 15;50(20):2295-301.
[17] Pao W, Miller VA. Epidermal Growth Factor Receptor Mutations, Small-Molecule
Kinase Inhibitors, and Non-Small-Cell Lung cancer : Current Knowledge and Future
Direction. J Clin Oncol, 2005. 23, 2556-2568.
[18] Jorissen RN, Walker F, Pouliot N, et al. Epidermal growth factor receptor : Mechanisms
of activation and signalling. Exp Cell Res, 2003. 284, 31-53.
[19] Nawot T, Plusquin M, et al. Environmental exposure to cadmium and risk of cancer : a
prospective population- based study. Lancet, 2006. 17, 119-126.
[20] Fielding JE, Phenow KJ. Health effects of involuntary smoking. N Engl j Med, 1988, 319,
1452-1460.
[20] Pavia M, Bianco A, Pileggi C, Italo F. Angillo : métaanalysis of residential exposure to
radon gas and lung cancer. Bull World Health Organ, 2003. 81.
[21] Peto R, Darby S, Deo H et al. Smoking, smoking cessation, and lung cancer in the UK
since 1950 : combination of national statistics with two case-control studies. Br med J. 2000.
321, 323-329.
[22] Heminki K, Pershagen G. Cancer risk of air polution : epidemiological evidence. Environ
Health Perspect. 1994. 102, 187- 192.
[23] Imbrenon E. Estimation du nombre de cas de certains cancers attribuables à des facteurs
professionnels en France. Département santé travail-Institut de veille sanitaire, Saint Maurice,
2003. 28.
[22] Trédaniel Jean. L’épidémie tabagique. Rev mal respire. 2006. 23, 16S7-16S10.
[23] Smith-Warner SA, Spiegelman D, Yaun SS et al. Fruits, Vegetables and lung cancer : a
pooled analysis of seven cohort studies. Int J Cancer. 2003. 107, 1001-1011.
[24] Hammond EC, Selikoff IJ, Seidman H. Asbestos exposure, cigarettes smoking and death
rates. Ann N Y Acad Sci. 1979. 330, 473-490.
[25] Tockman MS. Other host factors and lung cancer susceptibility. In samet J, éd
Epidemiology of lung cancer. Lung biology in Health and Disease. 1994. 74, 397-412.
[26] Mendelson J, Baselga J. Status of epidermal growth factor receptor antagonists in the
biology and treatment of cancer. J Clin Oncol. 2003. 21, 2787-2799.
[27] Sandgren EP, Luetteke NC, Palmiter RD, et al. Overexpression of TGF alpha in
transgenic mice : induction of epithelial hyperplasia, pancreatic metaplasia, and carcinoma of
the breast. Cell.1990. 61, 1121-1135.
[28] Di fiore PP, Pierce JH, Fleming TP, et al. Overexpression of the human EGF receptor

80
confers an EGFdependent transformed phenotype to NIH 3T3 cells. Cell. 1987. 51, 1063-
1070.
[29] Downward J, Yarden Y, Mayes E, et al. Close similarity of epidermal growth factor
receptor and v-erb-B oncogene protein sequences. Nature. 1984. 307, 521-527.
[30] Gabrecht T, Andrejevic-Blant S, Wagnières G.Blue-violet excited autofluorescence
spectroscopy and imaging of normal and cancerous human bronchial tissue after formalin
fixation. Photochem Photobiol. 2007. Mar-Apr;83(2):450-8.
[31] Quoix E. Schraub S. Cancers broncho-pulmonaires primitifs : épidémiologie, étiologie,
anatomie pathologique, diagnostic, traitement. 2003. Faculté de médecine, université de Louis
Pasteur. Strasbourg.
[32] Nacht M., Dracheva T., Gao Y.et al. Molecular characteristics of non-small cell lung
cancer. Proc Natl Acad Sci U S A. 2001, 98(26):15203-8.
[33] Schafer FQ., Buettner GR. Redox environment of the cell as viewed through the redox
state of the glutathione disulfide/glutathione couple. Free Radic Biol Med. 2001. 30(11):1191-
212.
[34] Ghosh, R., Mitchell., D L. Effect of oxidative DNA damage in promoter elements on
transcription factor binding. Nucleic Acids Res. 1999. 7:3213–3218.
[35] Soini Y., Kahlos K., Napankangas U., Kaarteenaho-Wiik R., Saily M., Koistinen P.,
Paaakko P., Holmgren A., Kinnula VL. Widespread expression of thioredoxin and thioredoxin
reductase in non-small cell lung carcinoma., Clin Cancer Res. 2001.7(6):1750-7.
[36] Freedman RB., Hirst TR, Tuite MF. Protein disulphide isomerase: building bridges in
protein folding. Trends Biochem Sci. 1994. 19(8):331-336.
[38] Coppock, D L., Cina-Poppe, D., and Gilleran, S. The quiescin Q6 gene (QSCN6) is a
fusion of two ancient gene families: thioredoxin and ERV1. Genomics. 1998. 54: 460-468.
[39] Benayoun. B., Esnard-Feve.A., Castella S. Rat seminal vesicale FAD-dependent
sulfhydryl oxydase. Biochemical charcterization and molecular cloning of a member of the
new sulfhydryl oxydase/quiescin Q6 gene fa mily.. J Biol chem. 2001. 276: 13830-7
[40] Coppock DL., Kopman C, Scandalis S, Gilleran S. Preferential gene expression in
quiescent human lung fibroblasts. Cell Growth Differ. 1993, 4(6):483-93.
[41] Wittke I., Wiedemeyer R., Pillmann A., Savelyeva. Neuroblastoma-Derived Sulfhydryl
Oxidase, a New Member of the Sulfhydryl Oxidase /Quiescin6 Family, Regulates
Sensitization to Interferon –Induced Cell Death in Human Neuroblastoma Cells. Cancer
Research, 2003. 63 : 7742–7752.

81
[42] Thorpe C., Hoober KL., Raje S. et al. Sulfhydryl oxidases: emerging catalysts of protein
disulfide bond formation in eukaryotes. Arch Bioche Biophys. 2002. 405 : 1–12
[43] Li-chan EC. Applications of egg immunoglobulins in immunoaffinity. chromatography.
Egg Nutrition and Biotechnology. J. Sim, S. 2000. 323-339.
[44] Ostrowski M C., and Kistler, W.S et al. Properties of a flavoprotein sulfhydryl oxidase
from rat seminal vesicle secretion .Biochemistry.1980, 19: 2639-2645.
[45] Hoober KL, Glynn NM, Burnside J, Coppock DL, Thorpe C., Homology between egg
white sulfhydryl oxidase and quiescin Q6 defines a new class of flavin-linked sulfhydryl
oxidases. J Biol Chem. 1999. 274(45):31759-62.
[46] Zanata SM., Luvizon AC., Batista DF., High levels of active quiescin Q6 sulfhydryl
oxidase (QSOX) are selectively present in fetal serum. Redox Rep. 2005. 10(6):319-23.
[47] Thorpe C., Coppock D.L. Generating dislfides in multicellulare organisme: emerging
roles for a new flavoprotein family. J Biol Chem. 2007. 282:13929-33

82
Annexe 1
Réactifs pour SDS-PAGE
Réactifs : - Tris ( Euromedex, 26-128-3094B)
- SDS en solution 20% (Euromedex, EU0660B)
- Polyacrylamide 40% w/v (Qbiogene, Biac 3751)
- TEMED (Euromedex, 50406)
- Glycine (Euromedex, 26-128-6405)
- HCl en solution à 37% (Merck, 1L 1000)
Solutions :
1) Tris-SDS pH 8,8 (Tris 0,75 M, SDS 0,2 % w/v)
- Tris ………………………22,7 g
- SDS 20% …………………2,5 mL
- HCl ……………………….ajuster à pH 8,8
- H2O………………………..qsp 250 mL
2) Tris-SDS pH 6,8 (Tris 0,25 M, SDS 0,2% w/v)
- Tris …………………………..7,6 g
- SDS 20%.................................2,5 mL
- HCl …………………………..ajuster à pH 8,8
- H2O………………………….qsp 250 mL
3) Tampon échantillon non réducteur :
- Tris 0,25 M, SDS 0,2% pH 6,8....5 mL
- SDS 20%.……………………….4 mL
- Glycérol 30%……………………6 mL
- Bleu de bromophénol …………...40 µL
- H2O………………………….qsp 20 mL
Pour obtenir un tampon échantillon réducteur, nous ajoutons 10 % de mercaptoéthanol (Sigma, M-
7154) lors de la préparation des échantillons.
4) Tampon d’éléctrode Tris-glycine (Glycine 0,192M, Tris 0,05M)
- Glycine…………………………...28,8 g
- Tris …………………………..........6 g
- SDS 20% …………………………5 mL
- H2O……………………………….qsp 1000 mL
5) Persulfate d’ammonium 10%
- Persulfate d’ammonium………......0,1 g
- H2O……………………………….1 mL
Gel de séparation (polyacrylamide 12%)
- Acrylamide ……………………......1,2 mL
- Tampon pH 8,8 …………………….2 mL
- TEMED……………………………0,005 mL
- Persulfate d’ammonium…………...0,033 mL
- H2O………………………………..0,8 mL
Gel de concentration (polyacrylamide 5%)
- Acrylamide …………………….......0,250 mL
- Tampon pH 6,8 …………………....1 mL
- TEMED……………………………0,005 mL
- Persulfate d’ammonium…………...0,100 mL
- H2O………………………………...0,850 mL
100 µL de Persulfate 10% et 5µL TEMED à ajouter au moment de couler le gel
Solution de coloration : rouge Ponceau
-Rouge ponceau …………………… 2 g
-Acide trichloracétique…………….. 30 g
-H2O……… ………………………. 1 L
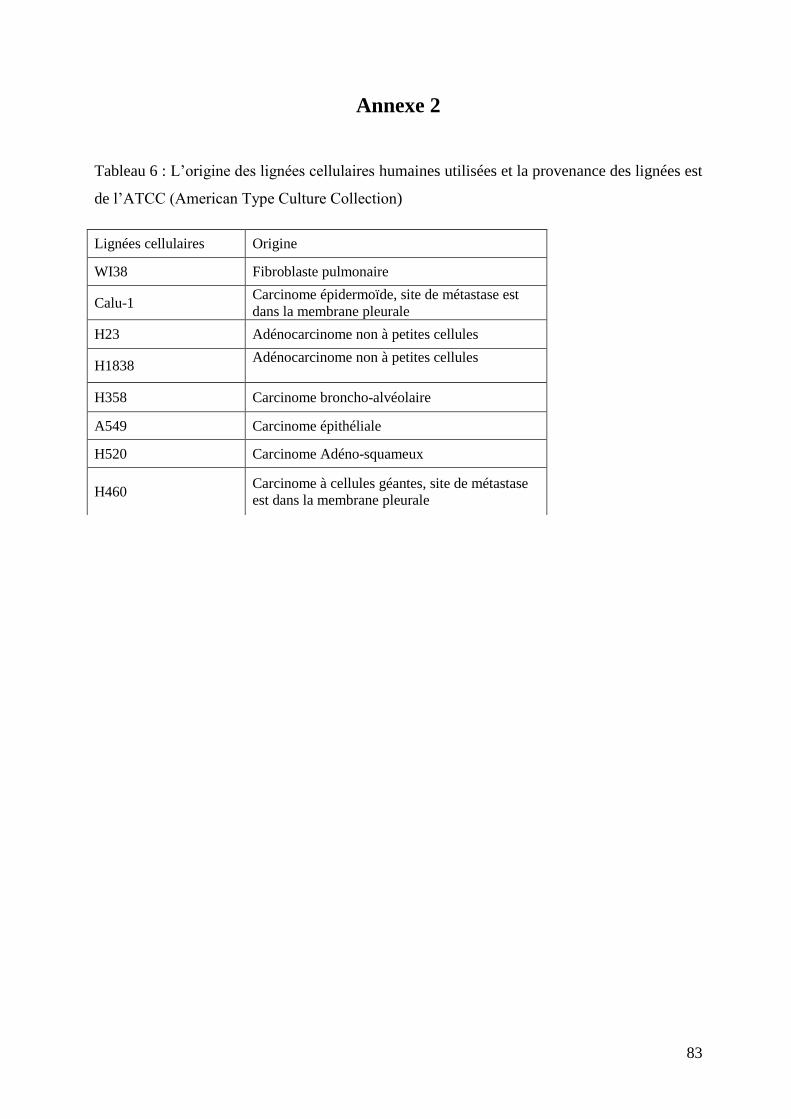
83
Annexe 2
Tableau 6 : L’origine des lignées cellulaires humaines utilisées et la provenance des lignées est
de l’ATCC (American Type Culture Collection)
Lignées cellulaires Origine
WI38 Fibroblaste pulmonaire
Calu-1 Carcinome épidermoïde, site de métastase est
dans la membrane pleurale
H23 Adénocarcinome non à petites cellules
H1838 Adénocarcinome non à petites cellules
H358 Carcinome broncho-alvéolaire
A549 Carcinome épithéliale
H520 Carcinome Adéno-squameux
H460 Carcinome à cellules géantes, site de métastase
est dans la membrane pleurale









![1Introduction - ITU!MSW-F.docx · Web view2013-04-18~26 Incheon, Corée (Rép. de) Q6/16 [rapport] Q6/16 de l'UIT-T, JCT-VC & JCT3V 2013-05-06~10 Johannesburg, Afrique du Sud ...](https://static.fdocuments.fr/doc/165x107/5ff9c5f20a4fe81f807a3033/1introduction-itu-msw-fdocx-web-view-2013-04-1826-incheon-core-rp.jpg)







