
ET DE LA RECHERCHE SCIENTIFIQUE · Remerciements En mon nom personnel, Mes premiers remerciements...
113
REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE MINISTERE DE L’ENSEIGNEMENT SUPERIEUR ET DE LA RECHERCHE SCIENTIFIQUE UNIVERSITE D’ORAN ES-SENIA FACULTE DES SCIENCES DEPARTEMENT DE BIOLOGIE LABORATOIRE DE MICROBIOLOGIE APPLIQUEE MEMOIRE Présenté par Mr. BENADA Mhamed POUR L’OBTENTION DU DIPLOME DE MAGISTER Spécialité : Microbiologie Option : Phytiatrie Phytopathologie Soutenu le : Devant le jury composé de : Année Universitaire : 2009 - 2010 Mr : KIHEL Mebrouk Professeur à l'Université d'Oran Président Mr : SLIMANI Miloud Professeur à l'Université d'Oran Examinateur Mme : SADKI Keira Maître de Conférences à l'Université d'Oran Examinateur Mr : HENNI Jamal Eddine Professeur à l'Université d'Oran Rapporteur Melle : BENICHOU Soumaya lalia Maître Assistante Classe A à l'Université d'Oran Co-rapporteur « Etude de la variabilité phénotypique du champignon Alternaria dauci pathogènes des Apiacées en vue d’améliorer les méthodes de lutte.»
Transcript of ET DE LA RECHERCHE SCIENTIFIQUE · Remerciements En mon nom personnel, Mes premiers remerciements...

REPUBLIQUE ALGERIENNE DEMOCRATIQUE ET POPULAIRE
MINISTERE DE L’ENSEIGNEMENT SUPERIEUR
ET DE LA RECHERCHE SCIENTIFIQUE
UNIVERSITE D’ORAN ES-SENIA
FACULTE DES SCIENCES
DEPARTEMENT DE BIOLOGIE
LABORATOIRE DE MICROBIOLOGIE APPLIQUEE
MEMOIRE
Présenté par
Mr. BENADA Mhamed
POUR L’OBTENTION DU DIPLOME DE MAGISTER
Spécialité : Microbiologie
Option : Phytiatrie Phytopathologie
Soutenu le : Devant le jury composé de :
Année Universitaire : 2009 - 2010
Mr : KIHEL Mebrouk Professeur à l'Université d'Oran Président
Mr : SLIMANI Miloud Professeur à l'Université d'Oran Examinateur
Mme : SADKI Keira Maître de Conférences à l'Université d'Oran Examinateur
Mr : HENNI Jamal Eddine Professeur à l'Université d'Oran Rapporteur
Melle : BENICHOU Soumaya lalia Maître Assistante Classe A à l'Université d'Oran Co-rapporteur
« Etude de la variabilité phénotypique du champignon
Alternaria dauci pathogènes des Apiacées en vue d’améliorer
les méthodes de lutte.»

Résumé
Les maladies fongiques, transmissibles par les semences, sont responsables d’importantes
pertes souvent difficiles à estimer à ce stade de la culture. Parmi celles-ci les alternarioses
sont particulièrement redoutées dans la mesure où elles concernent un nombre important de
plantes de grande culture et ont, de ce fait, un impact économique non négligeable. On trouve
notamment sur les semences de carottes et autres Apiacées (persil, cerfeuil, celeri…), un
complexe d’espèces du genre Alternaria responsables de brûlures foliaires et de taches
racinaires (Alternaria dauci, A. radicina, A. petroselini). Ces espèces sont présentes de façon
récurrente sur les Apiacées et elles peuvent, selon le stade d’attaque, affecter la qualité
germinative des semences, provoquer la mort des plantules issues de la germination des
graines (baisse de rendement) ou bien provoquer des taches nécrotiques sur différents organes
des plantes adultes les rendant ainsi impropres à la commercialisation.
Dans notre travail, qui se divise en trois parties ; nous avons élaboré :
En premier partie, une caractérisation biométrique, et cela pour retrouver les meilleures
conditions physico-chimiques de la croissance mycélienne.
En deuxième partie, est une lutte chimique en a utilisant deux substances chimiques (triazole)
synthétisées au sein du laboratoire de synthèse organique appliquée (université d’Oran).
En troisième partie, une étude du pouvoir pathogène de notre champignon, en comparant le
degré d’agressivité entre trois souches consernant des variétés différentes.
L’étude des conditions physico-chimiques a montré que : nous obtenons meilleur croissance
1) sur milieu PSA (Pomme de Terre Saccharose) dont la source de carbone est le saccharose,
par rapport aux autres milieux utilisés.
2) sur un pH de milieu situé à 06.
3) sur une température d’incubation comprise entre 22°C et 28°C.
L’étude du polymorphisme par la révélation de trois systèmes isoenzymatique à savoir : les
estérases, les phosphatases acides, le catalase et la peroxydase, montre une similarité des
profils électrophorétiques chez les souches d’Alternaria dauci.
Nos souches ont montré une sensibilité envers les substances chimiques testées.
Le test de la pathogénicité a révélé la virulence de ces souches vis-à-vis des trois variétés de
la carotte avec un degré qui se diffère d’une variété à l’autre et d’une souche à l’autre.
Mots clés : Alternaria dauci, Alternariose, triazol, polymorphisme isoenzymatique.
i

Abstract
The fungal diseases, transmissible by the seeds, are responsible of huge loses which are
frequently difficult the estimate (evaluate) at this stage of culture. Among these, the
alternariose are particularly feared because they concern a great number of large scale culture
plants and have therefore a considerable economic impact.
We find, mainly in the seeds of carrots and other Apiacèes (parsley, chervil, celery), a
complexe of species of the type Alternaria responsible of foliar burnings and root stains
(Alternaria dauci, A. radicina, A. petrosilini).
These species are found in a recurrent way in the Apiacèe and can, according to the stage of
attack, affect the germinative quality of seeds, cause the dearth of seedlings resulting from the
germination or cause necrotic stains improper for commercialization.
In our work, which is divided in three parts, we proceeded:
First to a biometrical characterization to find the best physicals-chemicals conditions of
mycotic development.
The second part is dedicated to the chemical fight where we used two synthetised chemical
substances in the laboratory of organic and applied synthesis of the universite of Oran Es-
Senia (Triazol).
The third part consists of the study of pathogenic ability of the fungus, companing the degree
of aggressiveness of the three different varieties.
The study of physical chemical conditions showed that:
1) In the environnement PSA, where the saccharose is the best source of carbon.
2) The pH is 06
3) The optimal temperature between 22°C and 28°C.
The stady of isoenzymatic polymorphism of three systems nomely: The esterases, the
phosphatase acids, the peroxydase and catalisis showed a similarity of the electrophoretic
profiles of the strain of Alternaria dauci showing a sensibility towards the chemical
substances used.
The pathogenecity test revealed the virulence of our strain towards the two varieties of carrot
with different degrees between the varieties and strains.
Key words: Alternariose dauci, Alternariosis, triazol, phytopathology.
ii

الملخص
، مه تيه هزي نهمحصىل مهمح خسائش ػه انمسؤونح ذؼذ انثزوس تىاسؽح انمرىمهح انفؽشيح األمشاض
، وانري ذؤدي إنً خسائش انىثاذاخ أوىاع مه كثيش ػذد ذعش انري ، Alternariose وجذاألمشاض
. Alternariaمضيج مه وىع.... الرصاديح كثيشج، حيس وجذ ػهً تزوس انجضس،انكشفس وانثمذووس
: انجزسيح انثمغ و األوساق احرشاق ػه انمسؤونح
. (Aternaria dauci, A. radicina, A. petroselini)
خالل دساسرىا انري ذىمسم إنً شالز مشاحم لمىا في انمشحهح األونً تذساسح انفضيىنىجيح و رنك
، وانمشحهح انصاويح مخصصح نهمكافحح انكيماويح ظذ انفؽشياخ إليجاد انظشوف انمالئمح نىمى انفؽش
وانمشحهح انصانصح هي دساسح انمذسج , Triazol) ( ذؽثيمي ذشكية مخثش في مشكثح تاسرؼمال مادج كيماويح
. انرمشيعيح نهفؽش ػهً شالشح أوىاع مه انجضس
مصذس ػهً انمحرىيPSA وسػ انضسع دومى جي انذساسح انفيضيىنىجيح نهفؽش، كشفد ػه
. C22 °C2°8 ننهىمى تي، و دسجح حشاسج مىاسثح pH 6 في دسجح حمىظح سكاسوص شكم ػهً كشتىن
، إوضيم األسرشج و حمط إوضيم األستؼحدساسح ذؼذد انىشاغ اإلوضيمي، ػه ؼشيك إظهاس األوظمح
.Alternaria dauci مه و انثيشوكسيذاص، أظهش ذىاسك األشكال نهؼيىاخ انمؼضونح و انكراالصانفىسفىس
in مثيذ انفؽش2تاسرؼمال Alternaria dauci مه جهرها ذجاسب انمكافحح انكيميائيح، ظذ
vitro مذمجح في انىسػ PSAجامذ ترشكيض مخرهفح (500 و 250-100-50ppm ) ذكشف وجىد ،
. مصثػ نهزي انمىاد ػهً ومى انفؽش
شالشح أوىاع مه ي دساسح انمذسج انرمشيعيح نهفؽش أظهشخ اخرالف في دسجح ػذائيح انفؽش ػم
. انجضس
مفتاحيةال الكلمات
.اإلوضيمي انىشاغ انمكافحح انكيميائيح، انمذسج انرمشيعيح . Alternaria dauci, Alternariose, triazol,
iii

Remerciements
En mon nom personnel,
Mes premiers remerciements iront à Dieu le tout puissant pour m’avoir
aidé à suivre mes études universitaires et à les corroborer par le présent mémoire.
Mes vifs remerciements s’adressent à ma très chère mère qui a été toujours
présente quand il le faut ainsi que mes sœurs.
Que Mr. HENNI J.E., Professeur à la Faculté des Sciences, Département
de Biologie à l’Université d’Oran, veuille bien accepter mes remerciements les
plus sincères pour l’accueil qu’il m’a réservé durant la réalisation de ma thèse au
sein de son Laboratoire de Phytopathologie, à l’Université d’Oran au
Département de Biologie, et la mise à ma disposition des moyens nécessaires
pour le déroulement correct de mon travail.
Je remercie Melle. BENICHOU S.L., Maître assistant à l’Université
d’Oran, pour son suivi efficace et rigoureux. Je la remercie chaleureusement pour
son guide précieux et ses conseils judicieux. Qu’il trouve ici l’expression de ma
gratitude et de mon profond respect.
Je remercie chaleureusement Mr. KARKACHI N., Maître assistant à
l’Université d’Oran, pour son aide, sa disponibilité qu’il a constamment montrée
à mon égard et ses conseils judicieux.
Je tiens à remercier vivement Mme KEDAR Née HAMINI N Maître
assistant à l’Université d’Oran et Mme GHARBI KARKACHI. S, Maître
assistante à l’Université Mohamed Boudiaf d’Oran (U.S.T.O.) pour leurs aides

précieuses, ses rigueurs scientifiques, ses conseils avisés et ses encouragements
permanents.
Je suis très sensible à l’honneur que me fait Mr. KIHAL. M., Professeur à
l’Université d’Oran, qui a bien voulu accepter de présider ce Jury. Qu’il veuille
bien accepter l’expression de mes sincères remerciements.
Je remercie Mr. SLIMANI M., Professeur à l’Université d’Oran et Mme.
SADKI.K., Maître de Conférences à l’Université d’Oran, qui ont acceptés
d’examiner ce travail. Qu’ils trouvent ici le témoignage de mon profond respect.
Je tiens à remercier tout particulièrement Mr. GUESSAS B., Maître de
Conférences à l’Université d’Oran, qui m’a toujours aidé par ces conseils.
Mes remerciements s’adressent également à toute l’équipe du Laboratoire
de microbiologie appliquée et tous mes encouragements à ma promotion de
Magister.
Je suis particulièrement reconnaissant à mes collègues Mohamed D., Yezli
W, Zabouri Y, Si mohamed A, Amina A., Fouzia T, Narimenne B pour leur
disponibilité exceptionnelle, leur aide physique, leurs encouragements et surtout
leur soutien moral.
Je remercie enfin, tout ce qui ont contribué de prés ou de loin et qui ont
pris une part active à la réalisation de ce travail, ainsi que tous ceux qui j’aurais
pu oublier et qui se reconnaitront dans la collaboration apportée à la réalisation
de ce travail.
Benada M’hamed

Dédicace
Au nom d’ALLAH ;
Celui qui fait miséricorde, le très miséricordieux, qui m’a donné la
force de concevoir ce travail et que le salut et la bénédiction de dieu soit
sur notre prophète Mohamed.
Je dédie ce travail à la mémoire de mon père qui sont départ a
laisser un grand vide dans ma vie que dieu bénit son âme, je le dédié à la
femme la plus chère du monde, la plus proche de mon cœur « ma mère »
que dieu la garde pour moi, à mes adorables sœurs ainsi leurs enfants et
à mes beau frères, à mes amis : Boualem, Djamel , Mansour, Mourad,
Filalli, Kamel, Mohamed et Youssef.
Une grande dédicace s’adresse à Melle BENICHOU S.L (co-
encadreur) en qui j’ai trouvé aide et encouragements.
A tous que j’aime et qui m’aiment.
Merci

Liste des Abréviations
% : pourcent
°C : Degré Celsius
µl : microlitre
µm : micromètre
Cm : centimètre
CMI50 : Concentration Minimale Inhibitrice
Est : Estérase
g/L : grammes / Litre
M : Molaire
mA : milliampère
mg : milligramme
ml : millilitre
mm : millimètre
nm : nanomètre
PAC : Phosphatase Acide
PAGE : Poly Acrylamide Gel Electrophoresis
PSA : Potato Saccharose Agar
pH : potentiel d'Hydrogène
Pox : Peroxydase
ppm : partie par million
q.s.p. : quantité suffisante pour
Glu : Glucose
Sac : Saccharose
Fru : Fructos
Lac : Lactose
Tém : Témoin
L.C : Lumiére continu
O. C : Obscurité continue
Photo. 12h : Photopériode de 12 heures
A.dauci : Alternaria dauci
vi

Tables des Matières
Résumé .................................................................................................................................................... i
Abstract .................................................................................................................................................. ii
iii .................................................................................................................................................. انمهخص
Liste des abréviations ............................................................................................................................vi
Liste des tableaux ............................................................................................................................... xiii
Liste des Planches ............................................................................................................................. xiv
Liste des Histogrammes ................................................................................................... xv
Glossaire ............................................................................................................................................ xvi
Introduction....................................................................................................................................... 2
Première Partie : Synthèse Bibliographique
Chapitre I : La carotte……………………………………………………………………………. 5
I. Carotte (Daucus carota)……………………………………………………………………………. 6
I.1 Origine et classification (systématique)………………………………………………………. 6
I.2 Production mondial de la carotte …………………………………………………………… 7
I.3 Caractéristique de la plante…………………………………………………………………. 7
I.4 Valeurs nutritif de la carotte…………………………………………………………………. 7
I.5 Principales maladies fongique de la carotte…………………………………………………. 8
Chapitre II : L’agent pathogène………………………………………………………………… 10
II. Alternariose …………………………………………………………………………………… 11
II.1 Historique du genre Alternaria ..................................................................................................1 11
II.2 Généralité sur le genre Alternaria …………………………………………………………… 11
II.3 Classification du genre Alternaria caractéristiques distinctives ………………………….. 13
II.4 Caractéristiques distinctives …………………………………………………………………. 13

II.5 Systématique du genre Alternaria : position taxonomique et morphologie des conidies et des
colonies……………………………………………………………………………………….
13
II.6 Le genre Alternaria ………………………………………………………………………….. 14
II.7 Mode d’infection de la plante ………………………………………………………………. 15
II.8 Déterminants du pouvoir pathogène des champignons appartenant au genre Alternaria…….. 15
II.8.1 Fixation du champignon à la cuticule de la plante
……………………………………………
15
II.8.2 Pénétration des tissus végétaux par le champignon…………………………………. 16
II.8.3 Colonisation de la plante et protection vis à vis des défenses de l’hôte…………….. 16
II.8.4 Colonisation de la plante et production de facteurs nécrogènes par le
Champignon………………………………………………………………………………
17
II.9 Symptomatologie de la maladie des brûlures foliaires…………………………………….. 18
II.10 Cycle parasitaire et gamme d’hôtes d’A. dauci……………………………………………. 18
II.11 Importance économique de la maladie des brûlures foliaires …………………………… 22
II.12 Méthode de lutte contre Alternaria dauci…………………………………………………. 22
II.12.1 Les méthodes culturales……………………………………………………………. 22
II.12.2 L’utilisation des agents antagonistes ………………………………………………. 22
II.12.3 La lute génétique……………………………………………………………………. 23
II.12.4 L’utilisation des fongicides ……………………………………………………….. 25
II.12.5 La lutte intégrée……………………………………………………………………... 27
III. Connaissances actuelles sur les déterminants du pouvoir pathogène d’A. dauci…………….. 27
V. Objectifs de l’étude.…………………………………………................................................... 28
Deuxième Partie : Matériels et Méthodes
Chapitre I : Etude du parasite et de la plante hôte…………………………………................ 30

I. Origine des souches ………………………………………………………………………... 31
II. Étude des caractères culturaux …………………………………………………………… 31
III. Conditions de culture ………………………………………………………………………. 32
III.1 Influence de la température d’incubation ………………………………………... 32
III.2 Photopériodicité………………………………………………………………….. 32
III.3 Estimation e la croissance mycélienne sur le milieu gélosé ……………………… 32
IV. Etude physiologique des souches d’Alternaria dauci …………………………………….…. 34
IV.1 Influence du pH du milieu ...………………………………………………………... 34
IV.2 Influence des sources carbonées sur la croissance mycélienne …………………. 34
IV.3 Influence des sources azotées sur la croissance mycélienne …………………….. 34
IV.4 Conservation des souches ………………………………………………………… 35
V. Evaluation du pouvoir pathogène ………………………………………………....................... 35
V.1 Conditions de culture de Daucus carota ……………………………………………. 35
V.2 Culture d’Aternaria dauci …………………………………………………………. 35
V.3 Préparation de l’inoculum…………………………………………………………... 35
V.4 Inoculation des plantes hôtes ……………………………………………………….. 36
V.5 Réisolement du parasite……………………………………………………………… 36
Chapitre II : Etude du polymorphisme isoenzymatique ……………………………………. 37
I. L’étude des polymorphismes isoenzymatiques d’Alternaria dauci ……………………….. 38
I.1 Préparation des extraits mycéliens ………………………………………………....... 38
I.1.1 Culture du champignon ……………………………………………………………... 38
I.1.2 Obtention des extraits ……………………………………………………………… 38
I.1.3 Électrophorèse ………………………………………………………………………. 38
I.1.3.* Préparation des gels pour le système PAGE………………………………… 38
I.1.3.* Le dépôt des échantillons …………………………………………………… 39
I.1.3.*La migration et mise sous tension …………………………………………… 39
I.1.3.*.Révélation des systèmes isoenzymatiques ………………………………….. 39
I.1.3.* Isoenzymes étudiés…………………………………………………………... 39
-Les Estérases …….……….…………………………………………………... 39
- Les Phosphatases Acides ….………………….……………………………... 40
- Les Peroxydases……………………………………………………………... 40
- Les Catalases………………………………………………………………… 40
Chapitre III : Essais in vitro de recherche de moyen de lutte ……………………………… 42

I. Lutte chimique contre Alternaria dauci ……………………………………………………… 43
I.1 Les fongicides testés ………………………………………………………………… 43
I.2 Préparation des solutions de fongicides ……………………………………………... 44
Troisième partie : Résultats et Discussions
I. Aspect morphologique ……………………………………………………………………….. 47
II. Caractères microscopiques…………………………………………………………………… 47
III. Effet des différents milieux de culture …………………………………………………….. 47
IV. Etude physiologique d’Alternaria dauci. …………………………………………………… 50
IV.1 Influence de la température ………………………………………………………... 50
IV.2 Influence du pH du milieu …………………………………………………………. 50
IV.3 Influence des sources carbonées…………………………………………………. 50
IV.4 Influence des sources azotées ……………………………………………………... 50
IV.5 Influence de la lumière …………………………………………………………….. 51
V. Pouvoir pathogènes…………………………………………………………………………... 57
VI Électrophorèse …………………………………………………………………………......... 61
VI.1 Estérases …………….……………………………………………………………… 61
VI.2 Phosphatases ………………………………………………………………………… 61
VI.3 Peroxydases ………………………………………………………………………... 61
VI.4 catalases……………………………………………………………………………... 61
VII Lutte chimique contre les souches d’Alternaria dauci ………………………………………. 64
Discussions ……………………………………………………………………………………... 69
Conclusion………………………………………………………………………………………. 72
Perspectives……………………………………………………………………………………... 73
Références Bibliographiques………………………………………………………………. 75
Annexe ………………………………………………………………………………………… 83

Liste des Tableaux
Tableau n° 01 – Production mondiale des carottes ……………………………………................ 07
Tableau n° 02 – Collection des souches utilisées………………………………………………... 31
Tableau n° 03 – Echelle de notation des lésions nécrotiques après inoculation d’Alternaria
dauci sur les feuilles 2 et 3 de carotte ……………………………………………………………
58
Tableau n° 04 – Diamètre moyen (cm) des souches d’Alternaria dauci confronté avec la
substance A……………….……………………………………………………………………….
65
Tableau n° 05 – Diamètre moyen (cm) des souches d’Alternaria dauci confronté avec la
substance B………………………………………………………………………………………...
66

Liste des Planches
Planche n° 01 – Aspect des conidies de quelque espèce appartenant au genre Alternaria ……………. 12
Planche n° 02 – : Cycle parasitaire des champignons du genre Alternaria en interaction avec
différentes plantes-hôtes …………………………………………………………………………….
20
Planche n° 03 – symptôme d’Alternariose sur les feuilles de la carotte …………………………………. 21
Planche n° 04 – Aspect microscopique de spore d’Alternaria dauci…………………………………… 21
Planche n° 05 – Schéma de la méthode de mesure du diamètre dans l es deux sens
perpendiculaires …………………………………………………………………………………...
33
Planche n° 06 –Aspect macroscopique d’Alternaria dauci ……………………………………………. 48
Planche n° 07 – Aspect microscopique d’Alternaria dauci grossissement (X 40)………………………. 48
Planche n° 08 – Aspect du mycélium d’Alternaria dauci sur les quatre milieux différents ………… 49
Planche n° 09 – Effet des différentes températures sur la croissance mycélienne ……………………… 53
Planche n° 10 – Effet du différents pH sur la croissance mycélienne………………………………. 53
Planche n° 11 – Influence de la source de carbone sur la croissance d’Alternaria dauci ……………… 55
Planche n° 12 – Influence de la source d’azote sur la croissance d’Alternaria dauci ………………….. 55
Planche n° 13 – Influence d’éclairage sur la croissance d’Alternaria dauci ……………………………. 56
Planche n° 14 - symptômes d’Alternariose sur les feuilles de carotte causé par la
soucheP2……………………………………………………………………………………………
59
Planche n° 15 – symptômes d’Alternariose sur les feuilles de carotte causé par la
souche B4…………………………………………………………………………………………..
14
59
Planches n° 16 – symptômes d’Alternariose sur les feuilles de carotte causé par la
souche WAL1 …………………………….……………………………………………………….
15
59
Planches n° 17 – résultat de réisolement à partir des feuilles …………………………………….. 60
Planches n° 18 – observation microscopique de résultat de réisolement à partir des feuilles ……… 60
Planche n° 19 – Profil électrophorétique des estérases chez les souches d’Alternaria dauci …..…. 62
Planche n° 20 – Profil électrophorétique des phosphatases chez les souches
d’Alternaria dauci …………………………………………………………………………………...
16
62
Planche n° 21 – Profil électrophorétique des catalase chez les souches d’Alternaria dauci….…… 63
Planche n° 22 – Profil électrophorétique des catalase chez les souches d’Alternaria dauci …………….. 63
Planche n° 23 – Effet de la substance A à différentes concentrations sur la croissance
mycélienne des souches d’Alternaria dauci ………………………………..……………………….
67
Planche n° 24 – Effet de la substance B à différentes concentrations sur la croissance
mycélienne des souches d’Alternaria dauci ……………………………………….…………………
19
67

Liste des Histogrammes
Histogramme n° 01 – Représentation graphique de l’influence des différents milieux de
culture sur la croissance d’Alternaria dauci apres 7jours d’incubation ………………………….
49
Histogramme n° 02 – Représentation graphique de l’influence des températures sur la
croissance d’Alternaria dauci après 7 joursd’incubation ……………………………………………….
52
Histogramme n° 03 – Représentation graphique de l’influence du pH du milieu sur la croissance
d’Alternaria dauci après 7 jours d’incubation …………………………………………………………….
52
Histogramme n° 04: Représentation graphique de l’influence de la source de carbone sur
la croissance d’Alternaria dauci après 7 joursd’incubation….……………………………………....
54
Histogramme n° 05: Représentation graphique de l’influence de la source d’azote sur la
croissance d’Alternaria dauci après 7 jours d’incubation ……………………………………………...
54
Histogramme n° 06: Représentation graphique de l’influence d’éclairage sur la croissance
d’Alternaria dauci après 7 jours d’incubation ……………………………………………………….
56
Histogramme n° 07: Représentation graphique du % d’inhibition des souches d‘Alternaria
dauci confronté à la substance A. ……………………………………………….………………….
65
Histogramme n° 08: Représentation graphique du % d’inhibition des souches d‘Alternaria
dauci confronté à la substance B …………………………………………………………………..…
66

Glossaire
Ombelle : Mode d’inflorescence dans le quel les pédoncules floraux partent tout
d’un même point et arrive à peu prés au même niveau
Bisannuelle : Qui se produit tout les deux ans.
Bipennatiséquées : Pourvu de deux ailes.
Oblongues: Plus long que large.
Ellipsoïde : Solide engendré par la révélation d’une ellipse autour de l’un de ses
axes.
Purpurine: Matière colorante rouge que contient la garance.

Introduction

Introduction
La carotte est le légume racine le plus consommé dans le monde à cause de son intérêt
énergétique, parmi les maladies fongiques qui touchent la carotte, on trouve l’Alternariose qui
est due au champignon Alternaria dauci.
Alternaria dauci est mondialement présent dans toutes les zones de production de la
carotte. Il s’agit aujourd’hui de la maladie la plus destructive de cette culture (Farrar et al.,
2004).
A. dauci est fréquent sur les semences de carotte en France, mais aussi dans de nombreux
autres pays (Champion, 1997). Il peut infecter la carotte à tous les stades de développement.
Bien que les semences, parfois fortement contaminées, sont à l’origine des « manques à la
levée », cependant, ce n’est souvent que lorsque les plantes sont bien développées, que la
maladie est la plus visible, au début les taches sur les feuilles sont petites, brunâtres et
entourées d’un halo de décoloration jaune (Champion, 1997), ainsi les infestations sévères
peuvent engendrer une défoliation complète de la plante (Farrar et al., 2004).
Parmi les moyens de lutte, la lutte chimique est la plus disponible, la plus rapide, la plus
efficace et la plus économique (Henni, 1987).
L’objectif de ce travail est d’évaluer l’efficacité de deux nouvelles substances chimiques
(des triazol), synthétisés au sein du laboratoire de Synthèse organique Appliquée, Université
d’Oran Es-Senia.
Notre étude est subdivisée en trois parties :
La première partie, nous avons effectués une caractérisation biométrique :
a/ pour déterminer les meilleures, conditions physico-chimiques de la croissance mycélienne,
en utilisant différentes températures d’incubation, pH du milieu, lumière, source de carbone
du milieu.
b/ la caractérisation culturale sur les différents milieux de culture.
c/ l’étudie du profils isoenzimatiques d’Alternaria dauci en se qui concerne : les estérase, les
phosphatase, les peroxydase et les catalase.
La deuxième partie est consacrée à la lutte chimique sur milieu solide, des laquelle on a étudié
le taux d’inhibition des différentes souches, et voir l’efficacité de ces deux substances de
triazol.
-2-

Introduction
La troisième partie : nous avons testé l’agressivité de trois souches sur trois variétés de
carotte presto et boléro (variété sensible) et texto (variété plus ou moins résistante).Ces trois
souches sont la P2, la B4 et la WAL1. D’autre part, on a étudié le taux d’agressivité de ces
trois souches en faisant une comparaison entre les souches WAL1 (souche algérienne) avec
les deux autres souches (P2 considéré comme très agressive et la B4 comme agressivité
moyenne).
-3-

Première Partie :
Synthèse Bibliographique

Chapitre I :
La carotte

Synthèse Bibliographique Chapitre I : La carotte
I. Carotte (Daucus carota) :
I.1 Origine et classification (systématique) :
L'ancêtre sauvage de la carotte (Daucus carota) provient d'Afghanistan, qui reste le
centre de la diversité de cette plante. la carotte sauvage est une parente de la carotte du jardin.
Au début la carotte du jardin produit sa partie comestible la première année, fruits, fleurit la
deuxième année. Si on la laisse monter en graine, elle retourne rapidement à son prototype
sauvage qui est théoriquement comestible, mais trop amer et fibreux pour être mangé. Au
mésolithique final, elle est non différenciée du panais. On peut distinguer deux variétés de
carotte : la carotte de l'Est et la carotte de l'Ouest. (Péron 2006).
La carotte de l'Est a été domestiquée au Xe siècle et même peut-être plus tôt en Asie
Centrale, probablement dans ce qui est l'Afghanistan présent. Les carottes de l'Est, qui sont
encore présentes aujourd'hui, sont souvent violettes ou jaunes et ont parfois une racine
branchée. La couleur violette est due à la présence d'anthocyanes. (Péron 2006).
La carotte de l'Ouest est apparue à la Renaissance, au XVIIe siècle, en Hollande, suite à
des sélections naturelles. Cette première carotte charnue, appelé la « Longue Orange ».
Les variétés maraîchères sont cultivées comme des plantes annuelles, car ce qui
intéresse le jardinier est la racine tubérisée et non la graine. La plupart des variétés présentent
une couleur orange caractéristique. Il existe aussi des carottes fourragères blanches. Des
variétés anciennes, actuellement en voie d'extinction, sont aussi de couleur rouge, violette ou
jaune.
Classification : Systématique (Guignard j.L, 2001).
Règne : Plantae.
Emb: Spermaphytes.
S/emb: Angiospermes.
Classe : Eudicots ou Eudicotylèdones.
S/Classe : Astèridèes.
Ordre : Apiales.
Famille : Apiacèes ou Ombellifères.
Genre : Daucus.
Espèce : carota.
-6-

Synthèse Bibliographique Chapitre I : La carotte
I.2 Production mondial de la carotte :
Tableau n° 01 : Production mondiale des carottes (statistiques de la F.A.O., 2007).
Production en tonnes. Chiffres 2004 et 2007 Données de FAOSTAT
(FAO)
Chine 8 292 500 35 % 9105 000 33,8 %
Fédération de Russie 1 825 000 8 % 1900 000 7,1 %
États-Unis 1 900 110 8 % 1600 000 6,0 %
Pologne 800 000 3 % 902 100 3,4 %
Royaume-Uni 650 000 3 % 859 300 3,2 %
Japon 660 000 3 % 750 000 2,8 %
France 469 280 2 % 710 000 2,6 %
Ukraine 500 000 2 % 625 000 2,3 %
Espagne
605 000 2,3 %
Italie 580 000 2 % 548 537 2,0 %
Autres pays 9 747 500 34 % 9304 007 32,1 %
Total 23601214 100 %
I.3 Caractéristique de la plante :
Plante bisannuelle de 30-80 cm., à rameaux étalés ; feuilles molles, les inférieures
oblongues, bipennatiséquées, à segments ovales ou oblongs, incisés-dentés ; fleurs blanches
ou rosées, celles de la circonférence rayonnantes, la centrale presque toujours purpurine ;
ombelles grandes,à 20-40 rayons grêles, arqués-convergents à la maturité ; involucelle à
folioles linéaires-acuminées, membraneuses au bord, entières ou trifides; fruit ellipsoïde, à
aiguillons en alêne, distincts à la base, égalant environ sa largeur. (Péron, 2006).
-7-
Synthèse Bibliographique Chapitre I : La carotte
I.4 Valeurs nutritif de la carotte :
L'apport énergétique de la carotte crue est de 31 kilocalories par 100 g (= 130 kilojoules
par 100 g). Elle contient 89.2g d'eau, 0.8 g de protéines, 0,3g de lipides et jusqu'à 6.6g de
Glucide dont 6.4g de sucre et 0.2g d’Amidon.
Les carottes sont riches en Béta carotène (provitamine A) 10000µg et 7µg de vitamine C.
Avec 25 g. on touvre plus de la moitié du besoin quotidien de vitamine A (204% pour 100g).
Elle contient aussi de la vitamine B1 et B2.
Les minéraux apportés sont principalement calcium 27mg, magnésium 10mg, potassium
286mg phosphore 16mg et fer 0.3mg.
La teneur en fibres alimentaires est importante 2.6g. Celles-ci sont composées en majorité de
pectine et de cellulose.
I.5 Principales maladies fongiques de la carotte :(Messian, 1991).
Les principales maladies cryptogamiques sont le mildiou, l'oïdium et l'alternariose.
Maladie
Agent causal Symptômes
I. Maladie des
feuilles
Bactériose américaine =bactérial
light
Xanthosomas
campesti pv
.carota
Taches graisseuses entourés
d’un halo jaune.
Mildiou Plasmospora
crustosa
Taches jaunes sur feuilles et
desséchement.
Septoriose Septoria carota Jaunissement du feuillage et
desséchement
Oïdium Erisphe heracki. Poudre blanche sale constituée
de conidies.
Cercosporiose Cercospora carota Taches circulaires claires et
bordées de brun.
Alternariose Alternaria dauci Taches brunâtres sur les bords
des feuilles.
-8- -8-
Synthèse Bibliographique Chapitre I : La carotte
I. Maladie des racines
Cavity-spot Pythium violae Nécrose sous forme de lésions
cicatrisées+ fendillement puis
éclatement longitudinales.
Rhizoctone violet ou bleu Rhizoctonia
violaceae
Formation d’un réseau, puis
feutrage velouté de couleur
pourpre ou bleuâtre
Pourriture blanche Sclerotinia
xclerotiorum
Feutrage blanc.
Pouriture noire Alternaria tenuis Nécrose noire dans la racine.
-9-

Chapitre II :
L’agent pathogène

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II. Alternariose :
II.1 Historique du genre Alternaria :
En 1816, Ness décrit pour la première fois un champignon présentant des spores
produites en chaîne portant un bec filiforme qu’il appelle Alternaria tenuis. Le genre
Alternaria a été successivement décrit par Groves et Skolko (1944), Neergaard (1945), Joly
(1964) et Simmons (1967, 1986, 1992).
Wiltshire en 1993, propose de regrouper dans le genre Alternaria toutes les espèces
dont les spores présentent un bec, sans tenir compte de la formation ou non des chaines
(Koenig,1995).
II.2 Généralité sur le genre Alternaria :
Les Alternaria, hyphomycetes dèmaties, sont caractérisés par la production des spores
d’un type particulier, les « dictyospores ».Les espèces correspondantes à ce genre produisent,
au moins théoriquement, leurs dictyospores en chaines, selon un mode centrifuge ; chaque
spore bourgeonnant une spore fille montre à sa partie apical, un rétrécissement en forme de
bec.
Ce caractère n’existe pas obligatoirement chez toutes les spores ; la dernière formée de
chaque chaine en étant généralement dépourvue (Joly, 1964).
A l’exception de quelques espèces aux conidies produitent en chaines, exemple :
Alternaria tenuis sur tomate.
La plupart des Alternaria, parasites des plantes maraichères, ont de très grandes spores
solitaires, pourvues le plus souvent d’un appendice filiformes c’est le cas des Alternaria
parasites des Allium (ail, oignon), de la carotte et de la tomate. Les conidies d’Alternaria sont
très résistantes à la sècheresse, elles résistent à plus d’un an à l’état sec et sont doués d’une
très grande longétivité (Messiaen et al ,1991).
-11-
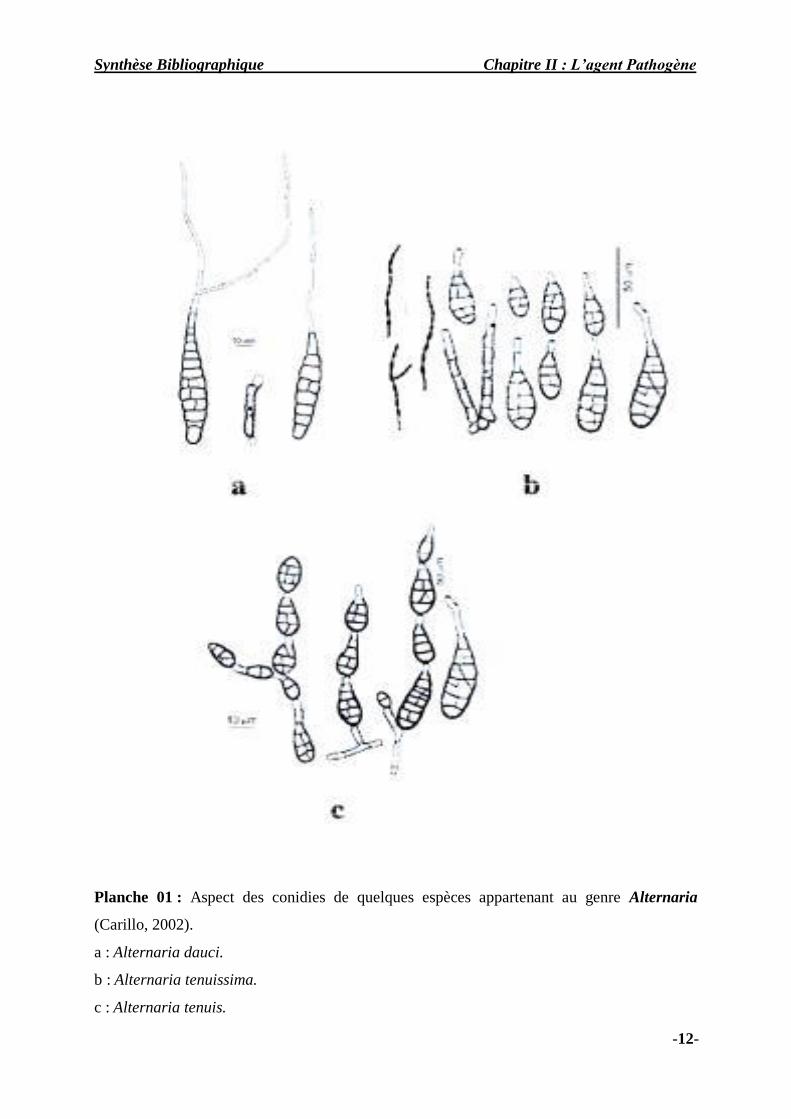
Synthèse Bibliographique Chapitre II : L’agent Pathogène
Planche 01 : Aspect des conidies de quelques espèces appartenant au genre Alternaria
(Carillo, 2002).
a : Alternaria dauci.
b : Alternaria tenuissima.
c : Alternaria tenuis.
-12-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.3 Classification du genre Alternaria :
La classification chez les Alternaria est fondée, sur la longueur des chaines de spores
et la taille de cette derniere d’après Neergaard(1945) in Joly (1964), Grillot(1981),
Nechev(1989), Ainsworth et Al (1961) :
Embranchement : Thallophytes.
Sous-embranchement : Septomycetes.
Classe : Deutéromycètes.
Ordre : Mucèdinales.
Famille : Dèmatiaceae.
Genre : Alternaria.
Espèces : dauci.
II.4 Caractéristiques distinctives :
Regroupé dans le tableau suivant, d’après Viennot, 1949 ; Joly, 1964 et Chou et Wu, 2002. :
Classe : Deutéromycètes =
Champignon imparfait =
Adélomycètes
Organisme dont la reproduction sexuée inconnue se présente
sous deux formes :
L’appareil végétatif constitué par un mycélium cloisonné et
l’appareil de reproduction asexué qui forme des spores de
dissémination.
Ordre : Mucédinales =
Moniliales = hyphes.
Caractérisé par des conidiophores généralement bien développé,
séparé les uns des autres.
Mycélium et conidies hyalins ou blanchâtres
Famille : Dematiacès. Conidies et conidiophores, colorés de teinte sombre et fortement
pigmentée, mycélium et spores de couleur brune.
Sous famille :
Dictyospores
Conidiophores simples ou ramifiés, spores muriformes.
Genre : Alternaria Caractérisé par des conidies pluricellulaires en masse fortement
colorée et pédiculée, regroupe de nombreuse espèces saprophytes
ou parasites.
-13-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.5 Systématique du genre Alternaria : Position taxonomique et morphologie des
conidies et des colonies
Alternaria dauci (Kühn) Groves & Skolko 1944 appartient à la famille des
Pléosporacées, à l’ordre des Pleosporales et à la classe des Ascomycètes. A. dauci possède
une reproduction exclusivement asexuée sous forme de conidies produites en abondance par
un conidiophore érigé. Les conidiophores sont de couleur brun-olive, flexueux et produisent
généralement une seule conidie. Comme chez toutes les espèces d’Alternaria, les conidies
d’A. dauci sont mélanisées. Elles sont cloisonnées transversalement et longitudinalement
(Planche 03.). Les conidies matures possèdent un long bec terminal (80 – 250 x 5 µm)
pouvant se ramifier en 2 ou 3 branches (Champion, 1997).
Les colonies d’A. dauci sur milieu gélosé sont pourvues d’un mycélium aérien court
gris foncé au centre et plus clair à la périphérie. Le contour des colonies est d’aspect ondulé.
Une pigmentation brun-rougeâtre, plus ou moins intense peut apparaître dans le milieu de
culture (Champion, 1997).
II.6 Le genre Alternaria :
Le genre est devisé en trois sections sur la base de la formation ou non de dictyospores en
chaines, et si ces chaines sont courtes ou longues (Chou et Wu ,2002).
-Caractéristiques afférentes aux trois sections :
-première section : comprend les espaces phytopathogénes qui forment des conidies
isolées avec des long filaments, on observe rarement la formation de 2 ou 3 conidies.
Exemple d’Alternaria dauci (Konstantinovka, 2002), (Planche 01 : 1a).
-deuxième section : regroupe les espèces qui forment les conidies en chaines non
ramifiées. L’exemple d’Alternaria gaisen qui forment les conidies en charnues et
ovoïdes en chaines par rapport à un court conidiospore secondaire, né dans la base de
la chaine d’une cellule intercalé dans une conidie. En culture les chaines se tiennent en
position verticale (Robert, 2002), (Planche 01: 1b).
-troisième section : Les espèces produisent des conidies en chaines ramifiées.
L’exemple d’Alternaria alternata qui produit des chaines de 10 ou plusieurs conidies
à partir de cellules conidiales fondamentales ou apicales (Andersen ,2001).Les chaines
en cultures paraissent denses et isolées en paquets (Robert ,2002), (Planche 01: 1c).
-14-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.7 Mode d’infection de la plante :
En dépit des taxonomies et les différences pathogènicites entre les espèces
d’Alternaria, elles causent des modèles d’infections semblables.
Les dictyospores sont facilement disséminées par le vent .Leur introduction dans de
nouveaux sols, ne se réalise qu’en partie grâce aux déplacements aériens (Joly, P, 1964).
Les spores conservées dans des conditions favorables, produisent un ou plusieurs
tubes de germination. Par la suite, ces tubes pénètrent dans des stomates, cuticule ou blessures
avec ou sans la formation du petit appressorium. Pour les espèces moins virulentes, les
blessures et stomates sont indispensables, en revanche, les espèces plus virulentes peuvent
pénétrer directement (Rotem, 1994).
Les processus enzymatiques dans les infections des Alternaria sont essentiellement
semblables à ceux des autres maladies fongiques. Les cuticules, qui consistent en une
combinaison de cutine (un polyester de l’acide gras de l’hydroxyle) et cires, comprend la
premières ligne de défense, qui ne résiste pas à la pénétration du pathogène fongique (Bart et
Thomma, 1999).
II.8 Déterminants du pouvoir pathogène des champignons appartenant au genre
Alternaria :
Les champignons du genre Alternaria appartiennent à la catégorie des nécrotrophes.
Ils tuent les cellules végétales (notamment par l’action de toxines) en avant du front de
développement de la lésion qu’ils provoquent et colonisent les tissus végétaux morts en quasi-
saprophytes (Agrios, 1997). Le processus infectieux mis en place par les champignons du
genre Alternaria pour infecter leur plante-hôte peut se diviser en 3 stades : la fixation, la
pénétration, la colonisation (protection vis à vis des défenses de l’hôte et production de
facteurs nécrogènes).
II.8.1 Fixation du champignon à la cuticule de la plante :
L’hydrophobicité de la cuticule est indispensable pour l’adhésion des spores, mais la
nature des propriétés physiques de surface intervenant dans ce phénomène est encore mal
connue. Chez les champignons du genre Alternaria, l’adhésion à la plante-hôte passe par la
formation d’un mucilage fibreux au niveau du tube germinatif, constituté de polysaccharides
(mannose et rhamnose) ou de glycoprotéines (Hatzipapas et al., 2002). Des lipases identifiées
à la surface des spores non germées d’A. brassicicola seraient impliquées dans les
mécanismes d’adhésion à la plante-hôte (Berto et al., 1999).
-15-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.8.2 Pénétration des tissus végétaux par le champignon :
Les champignons phytopathogènes, possèdent deux stratégies non exclusives de
pénétration dans les tissus végétaux : mécanique et enzymatique. Chez Alternaria spp., la
formation d’appressoria vrai n’a pas été mise en évidence (McRoberts et Lennard, 1996). La
formation de structures de type « appressoria-like » non mélanisées (Otani et al., 1998 a et b ;
Cramer et Lawrence, 2003) semble indiquer que la pénétration enzymatique est la plus
évidente chez les champignons du genre Alternaria. Le rôle crucial de lipases de type sérine
estérase dans la dégradation de la cuticule semble clairement établi chez A. brassicicola
(Berto et al., 1997). L’addition d’anticorps anti-lipases, à une suspension de conidies, a pour
effet de supprimer presque totalement l’apparition des symptômes (Berto et al., 1999).
Différentes cutinases induites, par la présence de monomères de cutines en surface du végétal,
semblent également impliquées dans la pénétration du champignon dans sa plante-hôte (Trail
et Köller, 1993, Yao et Köller, 1995). A. citri semble dépendant de la présence
d’endopolygalacturonases pour pénétrer son hôte, puisque le mutant déficient en cette enzyme
est altéré dans sa capacité à établir une infection. (Isshiki et al., 2001).
II.8.3 Colonisation de la plante et protection vis à vis des défenses de l’hôte :
Pour se protéger des phytoalexines et phytoanticipines synthétisées par la plante-
hôte, les espèces d’Alternaria mettent en place différents mécanismes de détoxication
(enzymatique, par conjugaison, systèmes d’efflux). A. solani possède des enzymes à activité
tomatinase qui le protège de la tomatine (phytoanticipine), une saponine synthétisée par la
tomate (Sandrock et vanEtten, 1998). A. brassicicola possède un mécanisme de dégradation
enzymatique faisant intervenir des enzymes de type cyanide hydratase. Ces enzymes
protègent le champignon vis à vis des isothiocyanates en les transformant en produits moins
toxiques donc tolérables (Cramer et Lawrence, 2003). La modification, par conjugaison, est
un autre mécanisme de détoxication des phytoanticipines, réalisée par la gluthation S-
transférase. Cette enzyme, mise en évidence chez A. brassicicola, métabolise les
isothiocyanates pour les rendre moins toxiques (Sellam et al., 2005). La surexpression de
gènes codant pour des pompes, à efflux de type ABC transporteur, a été observée chez A.
brassicicola en présence de phytoalexines (camalexine, brassinine) de Brassicacées. Ces ABC
transporteurs empêchent l’accumulation de phytoalexines à des niveaux toxiques pour la
cellule fongique (Sellam et al., 2005).
-16-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.8.4 Colonisation de la plante et production de facteurs nécrogènes par le
champignon :
La mort des cellules hôtes est provoquée par des toxines, produites par le
champignon, qui affecte généralement la perméabilité membranaire ou le fonctionnement de
certaines enzymes. Ces toxines sont des métabolites secondaires, de faible poids moléculaire,
efficaces à de très faibles concentrations. Deux catégories de toxines sont synthétisées par les
champignons du genre Alternaria : toxines non spécifiques de l’hôte (TNSH) et toxines
spécifiques de l’hôte (TSH).
Les toxines non spécifiques de l’hôte (TNSH0) :
Les TNSH sont des déterminants secondaires du pouvoir pathogène et interviennent
dans la composante quantitative du pouvoir pathogène (agressivité). Elles ne sont pas
nécessaires à l’initiation du processus infectieux mais contribuent à augmenter l’intensité des
symptômes de la maladie (Thomma, 2004). Les TNSH les plus courantes se répartissent en 4
groupes : dérivés du dibenzo-a-pyrone (alternariol : A. alternata), dérivés du pérylène
(altertoxine : A. alternata), dérivés de l’acide tétramique (acide ténuazonique : A. alternata, A.
tenuissima), polyketide (zinniol : présent chez tous les Alternaria à grandes spores et long bec
dont A. dauc ; les gènes impliqués dans la biosynthèse de ces NHST ne sont pas identifiés à
ce jour (Cotty et Misaghi, 1984).
Les toxines spécifiques de l’hôte (TSH)
Les toxines spécifiques de l’hôte sont les déterminants primaires du pouvoir
pathogène. Elles représentent la composante qualitative du pouvoir pathogène (virulence) et
sont indispensables à l’initiation de la maladie. Les TSH sont également nommées « agent de
compatibilité », car elles sont directement à l’origine du spectre d’hôte du champignon
(Walton, 1996). L’espèce Alternaria alternata comprend plus d’une dizaine de pathotypes
producteurs de TSH. Les TSH sont généralement des peptides cycliques ou des polyketides.
La biosynthèse des peptides cycliques ne se fait généralement pas par la voie ribosomique
classique mais par le biais d’enzymes de type Peptide Synthase Non Ribosomique (NRPS).
Les NRPS sont des enzymes géantes monomériques, modulaires et multifonctionelles. De
même, les polyketides sont synthétisés par des enzymes modulaires nommées polyketides
synthases. Les gènes codant pour la synthèse et la sécrétion de certaines TSH sont regroupés
en clusters et portés par un petit chromosome surnuméraire appelé « chromosome
dispensable » (Johnson et al., 2001 ; Hatta et al., 2002). Ce type de chromosome ne porte pas
de gènes nécessaires à la croissance basale du champignon, mais des gènes conférant un
avantage adaptatif vis à vis de l’expression du processus infectieux ou des conditions
environnementales.
-17-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.9 Symptomatologie de la maladie des brûlures foliaires :
La maladie des brûlures foliaires a été décrite pour la première fois en 1855 en
Allemagne. A. dauci peut infecter la carotte à tous les stades de développement. Les
semences, parfois fortement contaminées, sont à l’origine de « manques à la levée ».
Cependant ce n’est souvent que lorsque les plantes sont bien développées, que la maladie est
la plus visible. Les taches sur les feuilles sont d’abord petites, brunâtres et entourées d’un halo
de décoloration jaune (Champion, 1997) (Planche.03). Les infestations sévères peuvent
engendrer une défoliation complète de la plante (Farrar et al., 2004). Sur porte-graines, la
brûlure des feuilles apparaît au stade de la floraison. Les feuilles les plus âgées se dessèchent,
se crispent alors que les extrémités prennent une teinte grisâtre. Dès que la floraison est
accomplie, on retrouve le champignon sur les enveloppes florales à partir desquelles il
contamine les graines. Des contradictions existent en ce qui concerne la localisation d’A.
dauci dans les tissus de la graine. D’après Strandberg (1983), le champignon reste dans les
téguments et ne pénètre pas dans l’embryon, ni dans l’albumen. Or d’après Champion (1997),
la localisation du champignon dans la graine est parfois très profonde et l’albumen ainsi que
l’embryon peuvent être envahis.
II.10 Cycle parasitaire et gamme d’hôtes d’A. dauci :
Le cycle d’A. dauci est similaire à ceux des autres espèces d’Alternaria responsable
de maladies foliaire (Farrar et al., 2004). Un temps pluvieux ou la présence de rosée sont des
facteurs clés pour l’initiation de l’épidémie (Strandberg, 1977 ; Strandberg, 1987).
Les sources d’inoculum peuvent avoir plusieurs origines (carottes sauvages,
adventices, repousses de carotte ou semences) mais se sont les débris de carotte infectées qui
constituent la source d’inoculum prédominante (Farrar et al., 2004). A. dauci peu survivre
dans les débris de carotte infectées au champ pendant près d’un an si le sol est maintenu sec.
Les conidies des Alternaria sont capable de germer sans apport de nutriments
extérieurs cependant la sécrétion d’exsudats par la plante peu faciliter l’installation du
champignon. Ces exsudats sont composés d’acides organiques, d’amino-acides, de substances
de croissance, de vitamines, de composés phénoliques et de carbohydrates. La sécrétion
d’exsudats s’effectue au niveau de blessures dues aux insectes, à la pluie ou la rosée (Rotem,
1994).
Les conidies et les conidiophores sont produits dans des intervalles de températures
compris entre 8 et 28°C, en présence d’une humidité relative de 96 à 100% ou en présence
d’eau libre (rosée) en surface des organes végétaux (Strandberg, 1977). Les conidiophores
produits ressortent souvent par les stomates (c’est le cas notamment d’A. helianthi sur
-18-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
tournesol) mais il peuvent également se développer en d’autres site de la feuille tels
que les vaisseaux, trichomes ou à partir de mycélium croissant de façon sous-cuticulaire
(Allen et al., 1983 dans Rotem, 1994). Witsh et Wagner (1955) ont démonté que la
sporulation d’A. dauci s’effectuait en deux phases. Tout d’abord, la lumière stimule la
formation des conidiophores et deuxièmement les conidies sont formées à l’obscurité et sont
inhibées par la lumière (Rotem, 1994).
Une fois les conidies libérées, elles sont dispersées par des vents (Roy, 1969). La
dispersion des spores augmente lorsque l’humidité relative diminue et elle peut être
interrompue par une période prolongée de vents forts, d’humidité sur les feuilles et de
températures fraîches (Langenberg et al., 1977).
Les conidies déposées en surface des feuilles de carotte vont germer et infecter la
plante en conditions favorables de température et d’humidité. A 24°C, les dégâts sont plus
sévères si la feuille reste humide (Roy, 1969) pendant une période prolongée (72 heures
d’humidité en conditions saturées). A l’inverse, le taux d’infection est fortement réduit par des
températures nocturnes inférieures à 12°C (strandberg, 1988).
La période infectieuse est définie par le nombre de jours durant lesquels les conidies
sont produites. La période infectieuse des Alternaria est longue car elle commence sur feuilles
vivantes et se poursuit après la mort des tissus infectées (Rotem, 1994). Des conditions
environnementales favorables à la maladie qui se prolongent sur une longue période peuvent
conduire à un développement rapide de la maladie et engendrer des pertes de rendement
importantes.
-19-

Synthèse Bibliographique Chapitre II : L’agent Pathogène

Pla
nch
e 02 : C
ycle p
arasitaire des c
ham
pig
nons d
u g
enre A
lternaria en
interac
tion av
ec
différen
tes plan
tes-hô
tes (insp
irer d
u cy
cle d’A
grio
s, 1997).

Synthèse Bibliographique Chapitre II : L’agent Pathogène
Planche n° 03 : symptôme d’Alternariose sur les feuilles de la carotte.
Planche n° 04 : Aspect microscopique de spore d’Alternaria dauci. (Gr400).
-20-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.11 Importance économique de la maladie des brûlures foliaires :
A. dauci est mondialement présent dans toutes les zones de production de la carotte. Il
s’agit aujourd’hui de la maladie la plus destructive sur cette culture (Farrar et al., 2004). A.
dauci est fréquent sur semences de carotte en France et aussi dans de nombreux autres pays
(Champion, 1997). En France, A. dauci est chaque année responsable du refus de lots de
semences pour la certification. Cependant, l’impact économique de la brûlure foliaire dans
notre pays n’a jamais fait l’objet d’étude complète, et le nombre d’hectares touchés par la
maladie n’est pas évalué. En cultures de carottes de consommation, A. dauci entraîne des
pertes de rendements importantes en perturbant la récolte mécanique par préhension des fanes
lorsque la plante est atteinte par la brûlure foliaire, les feuilles fragilisées se cassent au
moment de la récolte et la racine reste dans le sol. En 2003, environ 80% des champs de
carotte de la région de Hatay en Turquie étaient infectés par A. dauci (Soylu et al., 2005). En
Israël, A. dauci est responsable de perte de rendement allant de 40 à 60% (Ben-Noon et al.,
2001).
II.12 Méthode de lutte contre Alternaria dauci :
Différents moyens de défense sont utilisés :
II.12.1 Les méthodes culturales :
Elles consistent, d’une part a éliminer les refuges parasitaires, les mauvaises herbes et
les débits végétaux, que l’on considère comme des sources de contamination et qui permettent
le suivi des organes de conservation du parasites en l’absence de l’hôte. D’autre part, la
pratique d’assolement de longue rotation permettant de réduire l’activité pathogénique du
parasite (Williams et al.,1989 in Henni, 1998).
Ce sont surtout les méthodes d’irrigation qui peuvent influer sur la propagation des
maladies foliaires (Blancard, 1988). L’irrigation par aspersion favorisera l’épidémie si elle se
pratique tôt le matin, ou tard le soir. Par contre une irrigation entre 11h et 15h ne favorisera
pas l’infection, si le feuillage sèche rapidement au soleil et pourra même freiner la
dissémination des Alternaria (Messiaen et al.,1991).
-21-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
II.12.2 L’utilisation des agents antagonistes :
Appeler aussi la lutte biologique, c’est une méthode alternative a la lutte chimique
qui domine encore largement le marché, cette lutte propose de combattre les ravageurs des
cultures en introduisant dans le milieu où il se développe un de leurs ennemis, un organisme
antagoniste. Souvent couplée à des techniques de conduite culturale, elle constitue une arme
efficace et écologique (Trotin-Caudal et Millot, 1993).
L’activité biologique des antagonistes vis-à-vis des champignons phytopathogénes
peut s’exprimer sous trois formes :
Une action destructrice ou inhibitrice de la germination des spores :
Owens et al., (1969) ont repartie certaines substances provenant de la décomposition
des matériaux organiques dans le sol par des saprophytes détruisent apparemment le
pathogène.
Une élimination des refuges parasitaires :
Selon Leach (1939), certains saprophytes contribueraient à l’élimination de ces
refuges en favorisant la décomposition des débits végétaux matériaux organiques, ce
qui réduit les disponibilités en éléments nutritifs nécessaires à la multiplication et à
l’extension du parasite dans le sol.
Une stimulation de la résistance chez la plante hôte :
Cette activité biologique efficace est basée sur la prémunition, un phénomène de
résistance induit ou acquis, par lequel une plante inoculée par une souche peu ou pas
pathogènes se trouve protégée des effets d’une deuxième inoculation par une autre
souche virulente. L’inoculation par une souche peu pathogène, stimule et déclenche
chez la plante la synthèse des phytoalexines (Rucci, 1986).
Selon Henni (1987), malgré les résultats prometteurs, la lutte biologique ne peut être
applicable en grande culture ou considérée comme une fin en soi, mais peut être
envisagée comme une action complémentaire à l’utilisation des variétés résistantes en
premier lieu ou éventuellement associée à une lutte chimique, vue que :
- L’action biologique est habituellement tardive et restreinte aux sols dans lesquels
des facteurs favorables à l’antagoniste utilisé sont réunis.
-22-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
- L’importation d’agents antagonistes dans un sol conduit à long terme à une
stabilisation de leur action, un équilibre biologique naturel entre partenaires.
- Leur utilisation rotationnelle est liée aux possibilités de leur culture artificielle et à
l’intensité de leur fructification, conditions nécessaires à leur vulgarisation en
masse à un moment donné.
II.12.3 La lutte génétique :
Les premiers travaux dans ce domaine remontent au début des années 90 chez la
pomme de terre et la tomate. Les gènes qui étaient utilises et qui le sont, encore
probablement aujourd’hui, sont des gènes qui code pour des enzymes impliqués dans la
dégradation des parois cellulaires des champignons : la chitinase et la glucanasec (Rioux et
Voldeng ,1999).
Etant donné que la majorité des résistances génétiques des plantes ne dépend que de
quelques gènes. Bien que la transgénèse semble tout indiquée pour développer des plantes
résistantes aux maladies, toutefois, ce n’est qu’avec une meilleure compréhension des
différents mécanismes impliqués en pathogènes et l’isolement de gènes de résistances des
plantes que nous pouvons réaliser de réelle progrès dans le développement de plantes
résistantes aux maladies que se soit à l’aide de la transgénèse ou autres méthodes (Rioux et
Voldeng, 1999).
Les sélectionneurs visent à cumuler le maximum de résistances, déjà bien connues et
utilisées avec des caractères agronomiques et de qualité des fruit .Cependant des défauts
affectant la fertilité des plantes résistantes à certaines maladies sont connus (Philouze et
Laterrot, 1992).On constate aussi qu’en très peu d’années, après la vulgarisation des variétés
résistantes, l4 de nouvelles races plus virulentes du parasite sont apparues (Messiaen,1981).
La création de variétés de carottes résistantes à A. dauci par sélection classique est
l’un des objectifs majeurs des sélectionneurs. L’analyse de croisements diallèliques révèle que
la résistance à A. dauci est surtout de type horizontal avec quelques gènes dominants (Simon
et Strandberg, 1998). Plus récemment, les travaux de thèse de Pawelec (2005) à l’UMR
GenHort ont permis d’identifier 10 QTL potentiellement impliqués dans la résistance de la
carotte à la brûlure foliaire lors des stades précoces de l’interaction hôte-pathogène.
-23-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
Certains auteurs ont eu recours à la transgénèse pour la création de variétés de carottes
résistantes à A. dauci. Takaichi et Oeda (2000) ont mis au point une lignée L12-1 transformée
avec un gène de lysozyme humain, protéine capable de cliver les liaisons ß-glycosidiques
de la chitine de la paroi fongique. Cette lignée de carotte transgénique produit abondamment
le lysoszyme (0,5% des protéines solubles totales) et possède un haut niveau de résistance à A.
dauci (Takaichi et Oeda, 2000). Récemment, des carottes transgéniques résistantes à A. dauci
ont été obtenues par transformation avec une protéine « thaumatin-like » (PR protéine)
identifiée chez le riz. Deux lignées ont été obtenues avec cette méthode et leur niveau de
résistance est bien corrélé au niveau d’expression de la protéine « thaumatin-like » (Punja,
2005) dont la localisation extracellulaire lui confère un rôle dans les réactions précoces de la
résistance à d’A. dauci. Toutefois dans le contexte actuel, l’utilisation de telles variétés
transgéniques pour la consommation humaine est difficilement envisageable.
II.12.4 L’utilisation des fongicides :
Malgré tout ce qui ce dit de nos jour sur les fongicides, ils constituent néanmoins de
outil indispensables à l’’agriculture. Avant la 2eme
guerre mondiale, le rendement était plus
faible que lui que l’on obtient aujourd’hui. Les causes sont multiples : manque de
mécanisation, cultivars à faible productivité, sans oublier la quantité limitée de fongicides
disponibles .De la fin des années 40 jusqu’au années 80, suite à la mise en place de pratiques
culturales de plus en plus mécanisées, de l’apparition de cultivars à productivité élevée et de
fongicides organiques, possédant un effet curatif, il a été possible d’accroitre de manière
substantielle les rendements agricoles (Bacon,2002).
La lutte chimique contre les maladies des plantes provoquée par les champignons
parasites est aussi vieille que l’algriculture. L’historique peut être essentiellement distingué
en trois époques :
1 -La période du <<soufre>>qui va des temps anciens à 1882.
2- La période du <<cuivre>> de 1882 à 1934 environ.
3- La période des <<fongicides organiques>> de 1934 à nos jours.
Bien que le terme fongicide dérive de : fongique (champignon) et cid (tuer ou
détruire), néanmoins, actuellement, sur pouvoir fongicide ou fongistatique (inhiber la
germination ou la sporulation) désigne tout composé ayant le pouvoir de détruire (action
-24-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
fongicide) ou d’inhiber la germination ou la sporulation (action fongistatique) des
champignons.
Beaucoup de ces produits n’agissent efficacement que de façon préventive c’est –à –
dire en empêchant la pénétration du parasite dans l’hôte; en agissant sur le métabolisme
énergétique ou sur la biosynthèse des glucides, des protéinés, des lipides et des acides
nucléiques (Bacon, 2002).
On distingue deux classes de fongicides (Deyson et Delcourt, 1980) :
1-Fongicides de contact ou inhibiteur (non systématiques) : se sont des fongicides qui ne
pénètrent pas dans l’hôte .Ils détruisent les champignons externes avant que ceux –ci ne
pénètrent dans l’hôte.
2—Fongicides systémiques : se sont des fongicides qui pénètrent dans les cellules,
circulent à l’intérieure de l’hôte pour tuer ou inhiber les champignons.
Selon Erwin 1977 (in Henni,1998), les fongicides curatifs peuvent agir de trois
façons :
a. En neutralisant la toxine émise par le parasite ;
b. En modifiant le métabolisme de l’hôte de telle manière qu’il devient plus résistant à
l’infection ou à la progression de parasite ;
c. En agissant directement sur le parasite après avoir pénétrer dans la plante et avoir été
transporté vers les tissus infectés.
Les fongicides offrent une longue protection, néanmoins, il faut recourir le plus
souvent à l’autres types de fongicides dans le traitement, car les organismes pathogènes
réussissent à développer une résistance contre les fongicides (Messiaen et al ; 1991).
Les stratégies pour la gestion des fongicides, selon Bacon (2002)
Ne pas utiliser un fongicides seul, (jamais un uni site seul) ;
Favoriser la diversité des produits, utiliser des fongicides de classes différentes ;
Réduire le nombre d’application des fongicides, en appliquant les fongicides aux
périodes propices, suite au dépistage de la maladie ;
Maintenir les doses préscrites par le manufacturier ;
Eviter tant que possible l’utilisation d’éradiquant (la plupart du temps uni site).
-25-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
On utilisera de préférence des fongicides d’efficacité générale : mancozébe,
propinébe, chlorothalonil… l’Alternaria est spécialement sensible à l’iprodione (Messiaen et
al ., 1991).
Cependant selon Henni (1987), malgré l’efficacité de ces produit chimiques, des
contingences socio-économiques de chaque pays ne permettent pas leur génération.
En effet
leur utilisation fait l’objet de législations qu’il convient de respecter en vue de protéger
l’homme, les animaux utiles et les microorganismes qui contribuent à l’équilibre du milieu.
Nous pensons que la lutte chimique se présente plus comme complément à la recherche de
variétés résistantes que comme une alternative en soi. En règle générale, la dose et le nombre
de traitements sont liés à :
-la sensibilité de la plante et à l’agressivité du parasite. Une variété plus résistante
permet de diminuer la dose et le nombre de traitements, ce qui fait baisser le cout de
la culture ;
-L’environnement, Walter (1971) a remarqué que la sensibilité de certains
champignons phytopathogénes est plus élèvées à des températures inférieures à 27
°C, qu’à des températures supérieures à 27°C, bien que les champignons se
développent optimalement à des températures supérieures à 27°C.
II.12.5 la lutte intégrée :
En cultures de tomate sous serre, il existe des risques permanents d’attaques par des
champignons pathogènes et des insectes piqueurs telles Aleurodes .En effet, la serre, milieu
confiné et climatisé, est très propice au développement de ravageurs et pathogènes.
Pour lutter contre les maladies provoquées par ces organismes, la protection intégrée
est largement utilisée depuis une quinzaine d’année .Elle consiste à associer différents
moyennes de lutte en réservant l’usage des produits chimiques aux situations pour lesquelles
il n’existe pas d’autres solutions ; elle intègre les moyens « naturels » de protection des
cultures comme la lutte biologique , l’action sur les facteurs climatiques sous serre , les
techniques culturales (Trottin-Caudal et Millot,1993).
-26-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
IV. Connaissances actuelles sur les déterminants du pouvoir pathogène d’A. dauci
A. dauci est un champignon nécrotrophe et contrairement à d’autres champignons du
même genre (A. brassicae, A. brassicicola, A. alternata).peu d’information sont disponibles
en ce qui concerne l’expression de son pouvoir pathogène malgré l’importance économique
de la maladie. Aucune étude n’a été conduite sur le mode de fixation d’A. dauci sur sa plante-
hôte. Aucune information n’est disponible concernant les mécanismes de protection et/ou de
détoxication mis en œuvre par ce champignon envers les composés de défense produits par la
carotte (Boedo, 2006). En présence d’A. dauci, la carotte produit une phytoalexine appelée 6-
méthoxymelléine (Dugdale et al., 2000 ; Fan et al., 2000 ; Mecier et al., 2000),
dihydroisocoumarine hautement toxique envers les bactéries et champignons phytopathogènes
(Kurosaki, 1996 ; Marinelli et al., 1996). Les données bibliographiques concernant le mode de
pénétration des hyphes mycéliennes d’A. dauci dans les tissus végétaux sont divergentes.
La bibliographie et littérature donne peu d’informations sur la production de toxines
spécifiques de l’hôte (TSH) par A. dauci. Une TNSH appelée zinniol qui altère la perméabilité
des membranes plasmiques et chloroplastiques (Thuleau et al., 1998) a été identifiée chez A.
dauci. L’application de zinniol purifié sur des feuilles de carotte provoque des symptômes
similaires à ceux causés par A. dauci (Barash et al., 1981). Les travaux de thèse de Montillet
(1986) montrent que le zinniol est présent dans les tissus infectés de la carotte pendant le
développement de l’alternariose. Au cours de la cinétique d’infection, lorsque la teneur en
zinniol augmente les symptômes s’aggravent. Cependant, l’importance de cette toxine dans le
processus d’installation du champignon sur son hôte n’est pas connue. Aucun dosage du
zinniol n’a été réalisé sur des spores en germination et on ne sait pas à quel moment du
processus infectieux cette toxine est synthétisée par A. dauci (germination, percée de la
cuticule, colonisation des tissus).
-27-

Synthèse Bibliographique Chapitre II : L’agent Pathogène
V. Objectifs de l’étude :
La présente thèse a été réalisée à l’université d’Oran au sein de l’unité de phytopathologie du
laboratoire de microbiologie appliquée dirigée par le professeur KIHAL.les travaux réalisés
ont été dirigés par le professeur HENNI Jamel Eddine et co-dirigés par Melle BENICHOU
Soumaya Lalia maître Assistante classe A.
Les objectifs de la présente thèse s’articulent autour de trois axes :
1- L’analyse de la variabilité phénotypique au sein de l’espèce A.dauci par l’analyse de la
sensibilité aux fongicides.
2- Mises en évidence du profil enzymatique de nos souches.
3- L’étude du pouvoir pathogènes sur différentes variétés de carotte.
Les expérimentations pour les deux premiers axes ont été réalisées avec une collection de
souche d’A.dauci d’origine géographique différente avec une souche Algérienne afin de faire
une comparaison (P2, B4, CBS1, CBS2, 10631, C5356-7, C5356-3, ALL, B7, WAL1).
En ce qui concerne le troisième axe, trois souches ont été utilisée P2 pour sa forte sporulation
et agressivité, la B4 pour sa faible sporulation et la WAL1 souche originaire de walhassa,
pour évaluer le facteur provenance géographique.
A terme, l’objectif majeur serait l’acquisition de connaissances fondamentales et leur
application au sein de travaux de recherche appliqué.
-28-

Deuxième Partie :
Matériels et Méthodes

Chapitre I :
Etude du parasite et de la
plante hôte

Matériels et Méthodes Chapitre I : Etude du parasite et de la plante hôte
I. Origine des souches :
Les souches d’Alternaria utilisées dans ces travaux proviennent de plusieurs pays du
monde, la majorité des isolats utilisés par l’analyse sont issue d’isolements effectués par le
GEVES-SNFS à partir des semences. Ces isolats nous ont été procurés de l’UMR PaVé de la
faculté des sciences à l’université d’Angers en France sous la direction du Pr Philippe
SIMONEAU. La souche wal1 à été isolé au sein de notre laboratoire de microbiologie
appliquée, à partir de feuille de carotte contaminée provenant de walhassa.
Tableau n°02 : Collections des souches utilisées.
code souche Origine géographique Identification morphologique
P2 France Alternaria dauci
B4 France Alternaria dauci
10631 Australie Alternaria dauci
CBS1 Souche de référence Alternaria dauci
B7 France Alternaria dauci
CBS2 Souche de référence Alternaria dauci
ALL Allemagne Alternaria dauci
C-5356-3 France Alternaria dauci
C-5356-7 France Alternaria dauci
WAL1 Walhassa W.A.Témouchent
(Algérie)
Alternaria dauci
II. Etude des caractères culturaux :
La culture sur différents milieux est nécessaire pour savoir sur le quel on aura une bonne
croissance mycélienne. Cette étude est réalisée par le dépôt d’un implant mycélien de 6 mm
de diamètre (de chaque isolat âge de 5 jours), au centre des boites de Petri, contenant chacune
un milieu différent. 3 répétitions de chaque souche pour chaque milieu ont été effectuées. Les
différents milieux utilises (Annexe) sont :
un milieu organique (PSA),
un milieu minéral (Czapeck),
un milieu riche (Mathur).
un milieu malth agar.

Matériels et Méthodes Chapitre I : Etude du parasite et de la plante hôte
L’examen des boites s’effectue à l’œil nu, des caractères culturaux observés selon
Koenig 1995 ; Guiraud et Galzy, 1980, sont la vitesse de croissance mycélienne, la surface, la
consistance, la pigmentation et sa variation du mycélium en fonction du temps de la colonie.
Cette étude est réalisée pour toutes les souches.
III. Conditions de culture :
III.1 la température d’incubation des cultures :
La température influence la croissance du champignon ainsi que son métabolisme
(action sur la vitesse de réactions chimiques et biochimiques) (Henri et al, 1995).
Pour déterminer la température optimale, des fragments mycéliens d’une préculture âgée
de 5 jours de 5 mm de diamètre sont posés au milieu de la boite de Pétri contenant un seul
milieu choisi : milieu PSA. Les boites sont incubées à différentes températures : 04 °C, 28 °C,
37 °C et 45 °C pendant 7 jours. La lecture des diamètres est effectuée à l’ordre de : 3, 7 et 14
éme jour.
Sauf indication, contraires, pour toutes nos expérimentations, nous avons utilisé une
température à 22°C.
III.2 Photopériodicité :
Le type et la durée d’éclairement joue un rôle important sur le développement des
champignons phytopathogènes. Les cultures sont incubées sous : un éclairage, une
photopériode continu de 12heures et une obscurité. La mesure de la croissance mycélienne a
été effectuée le : 3, 7 et 14 ème jours.
III.3 Estimation e la croissance mycélienne sur le milieu gélosé :
L’évaluation de la croissance mycélienne de chaque souche est mesurée
quotidiennement par deux diamètres perpendiculaires (Planche n°5) pendant 3, 7 et 14jours
d’incubation.
-32-

Matériels et Méthodes Chapitre I : Etude du parasite et de la plante hôte
Planche n° 05 : Schéma de la méthode de mesure du diamètre dans les deux sens
perpendiculaires (Blaise et al., 2001).
Boite de Pétri
contenant le
milieu de culture
Implants du
mycélium
-33-

Matériels et Méthodes Chapitre I : Etude du parasite et de la plante hôte
IV. Etude physiologique des souches d’Alternaria dauci :
Les champignons se multiplient à partir des aliments ou nutriments présents dans les
milieux de cultures, ils ont tous un certain nombre de besoins communs : nécessité d’eau, de
source d’énergie, de source de carbone, de source d’azote et d’élément minéraux (Henri et al,
1995).
Tous ces facteurs sont testés pour toutes les souches, à l’ordre de 3 répétitions pour
chaqu’une et pour chaque facteur.
IV.1 Influence du pH du milieu :
Le pH optimal de la croissance mycélienne des moisissures se situe entre : 2 et 9 (Regnault,
1990).
Pour préciser le pH optimal de la croissance mycélienne d’Alternaria dauci, les souches
sont cultivées sur milieu PSA solide tamponné à différents pH : 4 par le Tampon Acétate de
Sodium, 8 par le Tampon Phosphate ainsi que le pH 6 (annexe).
IV.2. Influence de source carbonée sur la croissance mycélienne :
Pour assurer leur développement (croissance et reproduction), les moisissures comme tout
microorganismes, utilisent les sources de carbone et d’énergie qu’elles trouvent dans leur
environnement et qu’elles dégradent à l’aide d’enzymes exocellulaires appropriées (Rosse,
1978).
De nombreux auteurs tel que : Moreau (1968), ont montré que la plupart des sources
carbonées peuvent être utilisé par les champignons. Jacquet (1979), a montré que la nature de
la source carbonée à une influence sur la croissance et la reproduction des champignons.
Pour chercher la source de carbone la plus assimilable par nos souches, le milieu utilisé
est le milieu PSA dans lequel le saccharose utilisé pour les boites témoins, était remplacé par
d’autres sources de carbone : le Glucose comme monosaccharide, le Lactose et le Fructose
comme disaccharide.
IV.3. Influence des sources azotées sur la croissance mycélienne :
Selon Jacquet (1979), la plupart des organismes fongiques utilisent les sources azotées
sous forme d’azote nitrique ou d’azote ammoniacal, ce qui nous a conduit à rechercher la
meilleure source azotée pour la croissance de nos souches.
Le milieu utilisé est le milieu Czapek, dont la source d’azote NaNo3 (Nitrate de Sodium)
est remplacée par le KNO3 (Nitrate de Potassium) et le NH4SO4 (Sulfate d’Ammonium),
comme deux sources d’azote.
-34-

Matériels et Méthodes Chapitre I : Etude du parasite et de la plante hôte
IV.4 Conservation des souches :
La conservation des souches d’Alternaria dauci se fait dans des tubes à essai contenant
milieu incline (PSA), à une température de -4°C et ceci pour une utilisation ultérieure.
V. Evaluation du pouvoir pathogéne :
V.1 Conditions de culture de Daucus carota :
Les semis de carotte sont réalisés en serre dans des pots contenant un substrat spécial
pour jeunes semis compose de tourbe. Dans chaque pot on a réalisé 6 puits de profondeur
d’environ de 2 a 3 cm. Dans chaque puits, on met 5 graines, ensuite on recouvre avec de la
tourbe, les pots sont ensuite recouvrés par des sachets humides pendant 48heures pour que
l’atmosphère reste humide. La température se situe entre 22 et 23°c avec une photopériode,
selon le besoin.
L’arrosage est effectué manuellement chaque jour avec de l’eau du robinet. Les variétés de
carottes utilisées sont boléro, presto et texto. Presto possède un rendement élevé et elle est
sensible à la brulure foliaire causée par A. dauci, alors que texto est reconnue pour sa
tolérance à ce genre de champignon.
V.2 Culture d’Aternaria dauci :
Pour l’ensemble de nos expériences nous avons utilisé les souches P2, B4 et WAL1
d’A .dauci (la WAL1 est une souche algérienne, elle est utilisée pour faire une comparaison
avec les souches P2 qui est connue pour son agressivité et la B4 avec une agressivité
moyenne). Ces souches sont cultivées sur milieu PSA (voir annexe). Les boites ensemencées
sont placées sous une photopériode à 22°c.
V.3 Préparation de l’inoculum :
10 ml d’eau distillé sont mis sur la surface de la culture, puis le mycélium est racle (cette
suspension est filtrée à travers une double épaisseur de gaz. La concentration est calibrée à
l’aide d’une cellule Malassez jusqu’à obtention de 105 spores/ml).
La suspension obtenue est filtrée à travers une double épaisseur de gaze. La concentration est
calibrée à l’aide d’une cellule Malassez jusqu'à obtention de 105 spores/ ml.
V.4 Inoculation des plantes hôtes :
Les plantes sont inoculées manuellement par pulvérisateur, l’atmosphère doit être saturé en
humidité, pour cela les pots sont recouverts par des sachets et il est nécessaire de repasser 2
fois par jours pendant les 2 jours qui suivent, pour asperger les plantes.
Parallèlement et pour les deux lots de plantes témoins, sont pulvérisés avec de l’eau distillée
stérile.
-35-

Matériels et Méthodes Chapitre I : Etude du parasite et de la plante hôte
V.5 Réisolement du parasite :
Après l’inoculation, la recherche du parasite dans la plante infectée a pour objectif de
s’assurer que les symptômes notés sont l’effet du pathogène. Ce contrôle est effectué sur
toutes les plantes inoculées, le champignon est recherché dans les feuilles présentant les
symptômes. Ces derniers sont rincées avec une solution d’Hypochlorite de Sodium diluée à
2% pendant 03 minutes, pour une désinfection superficielle et suivie de trois bains successifs
d’eau distillée stérile, ensuite, elles sont déposées en boites de Pétri contenant le milieu PSA.
-36-

Chapitre II :
Etude du polymorphisme
isoenzymatique

Matériels et Méthodes Chapitre II : Etude du polymorphisme isoenzymatique
I. L’étude des polymorphismes isoenzymatiques d’Alternaria dauci :
I.1 Préparation des extraits mycéliens :
I.1.1 Culture du champignon :
Des erlenmeyers de 250 ml, contenant 100 ml de milieu PSA liquide, sont ensemencées
par 5 boutures mycéliennes (de 6 mm de diamètre), prélevées de la zone périphérique d’une
préculture de 5jours du champignon sur milieu PSA. Les cultures à raison de 2 fioles pour
chaque isolat sont incubées pendant 10 jours sous photopériodicité.
I.1.2 Obtention des extraits :
Le mycélium est récupéré après filtration du milieu de culture à l’aide d’une passoire
stérile. Il est ensuite broyé dans un mortier maintenu au froid dans de la glace, additionné du
sable de Fontaine bleu et du Tampon Phosphate (0,1 M - pH 7,1) (P/P/V), jusqu’à l’obtention
d’une pâte fine et homogène. Le broyât est récupéré dans des tubes puis centrifugé à10 000 g
pendant 20 minutes dans une centrifugeuse réfrigérée (4 °C) (type Biofuge).
Le surnageant est réparti rapidement dans des tubes eppendorf par fraction de 100 µl. Il
peut être utilisé directement pour l’électrophorèse, comme il peut être conservé au
congélateur.
I.1.3 Électrophorèse :
I.1.3.* Préparation des gels pour le système PAGE :
L’électrophorèse se fait sur gel de polyacrylamide. Les deux gels (de séparation à 10% et
de concentration à 5%) sont préparés selon la méthode de Laemmli (1970), modifiée de la
manière suivante (Annexe)
La composition des tampons utilisés pour la préparation des gels et pour la migration est
donnée en Annexe.
Le gel se présente sous forme de matériel gélifié, formé par la polymérisation d’un
mélange d’acrylamide et de bis-acrylamide. Le gel de concentration est coulé en haut du gel
de séparation, pour permettre une entrée homogène de l’échantillon dans le gel de séparation.
Des pistes individuelles (puits) sont réalisées à l’aide d’un peigne, qui sépare le gel de
concentration en portions égales, destinées à la migration de chaque échantillon.
I.1.3.* Le dépôt des échantillons :
En terme de polymérisation des 02 gels , le peigne est enlevé délicatement sans
avoir à déchirer les gels .Les plaques en verre sont placées dans la cuve de
l’électrophorèse contenant le tampon de migration .On à 02 plaques de verre , et dans
chaque plaques on à 10 puits , On met les 10 extraits enzymatiques dans une plaque pour
-38-

Matériels et Méthodes Chapitre II : Etude du polymorphisme isoenzymatique
chaque révélation d’enzyme. On dépose 20 µl du tampon de charge dans chaque puits
sur lequel on ajoute 50 µl de chaque extrait enzymatique .
Chaque extrémité du gel sera mise en contact avec le tampon de migration qui est
soumis dans un potentiel électrique, permettant la propagation d’un courant dans le
gel ; ce courant entrainera les molécules constituant l’échantillon.
I.1.3* La migration et la mise sous tension :
La migration doit se dérouler en chambre froide à 4°C sous tension constante de 150 v
avec une intensité de 50 ma , durant 2 à 3 heures.
La migration est jugée suffisante lorsque l’un des composants du tampon de charge, le
bleu de bromophénol, atteint le front de migration du gel .
I.1.3.* Révélation des systèmes isoenzymatiques :
La révélation des isoenzymes est effectuée par incubation du gel (après démoulage) dans
un mélange réactionnel, contenant le substrat de l’enzyme étudiée. Trois systèmes
enzymatiques sont étudiés : Estérases (Est), Phosphatases acide (PAC) et Peroxydases (Pox).
L’annexe, nous indique les mélanges réactionnels pour la révélation de ces dernières.
I.1.3.* Isoenzymes étudiés :
Le terme Isoenzyme a été introduit en premier par Markert et Moller (1959), pour faire
référence à de multiples formes moléculaires d'une enzyme, avec une semblable ou identique
spécificité du substrat, qui se produit dans le même organisme.
- Les Estérases :
Une estérase est une enzyme de l'hydrolase qui fait les esters en un acide et un alcool,
sous une réaction chimique avec l'eau appelée l'hydrolyse.Il existe une grande gamme
d’estérases, mais qui diffèrent entre elles par la spécificité du substrat, la structure de la
protéine et la fonction biologique (Baaziz, 2000).
- Les Phosphatases Acides:
Sous le terme de Phosphatase Acide, sont regroupées des hydrolases. Ces hydrolases
catalysent la déphosphorylation d'esters orthophosphoriques organiques dont le pH d'activité
optimale est inférieur à 7. Elles possèdent également une activité transférasique, assurant le
transfert d'un phosphate sur le groupement hydroxyle d'un alcool (Moss, 1984).
-39-

Matériels et Méthodes Chapitre II : Etude du polymorphisme isoenzymatique
- Les Peroxydases :
Une Peroxydase est une enzyme, qui contient parfois un hème, qui catalyse une réaction
de la forme : ROOR' + donneur d'électrons (2 e-) + 2H
+ → ROH + R'OH
Ce type d'enzyme a notamment pour fonction de décomposer les peroxydes, dérivés toxiques
de l'oxygène (comme par exemple l'eau oxygénée ou peroxyde d'hydrogène), ce qui lui vaut
son nom. Se sont des enzymes très répondues chez les êtres vivants. Chez les plantes, elles
catalysent l’oxydation de différents substrats (des amines aromatiques, des phénols,...), elles
interviennent ainsi dans différents phénomènes physiologiques tels que l’enracinement, la
lignification, et la résistance aux stress biotiques et abiotiques (Qacif et al., 2006).
Certaines interviennent dans le métabolisme de l'iode et sont nécessaires à l'élaboration des
hormones thyroïdiennes. D'autres participent au transfert du chlore dans certaines cellules du
système immunitaire, permettant ainsi la production d'hypochlorite (la vulgaire eau de Javel),
servant à la destruction des microorganismes ingérés. D'autres microorganismes utilisent des
Peroxydases pour fabriquer des antibiotiques. Ces enzymes sont donc parmi les plus
universelles du monde vivant.
- Les Catalase :
La catalase est une enzyme catalysant la dismutation de l'eau oxygénée (peroxyde
d'hydrogène) : 2H2O2 → O2 + 2H2O
Elle est formée de quatre chaînes polypeptidiques d’environ 500 acides aminés, comportant
chacune un groupe hème. Ces hèmes et leur environnement protéique sont les sites actifs de
cette enzyme. Pour catalyser la réaction, l’atome de fer du groupement hème de la catalase
réalise une coupure hétérolytique de la liaison O-O du peroxyde d'hydrogène, créant de ce fait
une molécule d’eau et un groupement Fe(IV)=O hautement oxydant ; ce dernier peut ensuite
oxyder une autre molécule de peroxyde d'hydrogène pour donner du dioxygène. Ce processus
est illustré plus spécifiquement par les équations suivantes :
H2O2 + Fe (III)-E → H2O +O=Fe (IV)-E (1)
H2O2 + O=Fe (IV)-E → O2 + Fe (III)-E +H2O (2)
-40-

Chapitre III :
Essais in vitro de recherches
de moyens de lutte

Matériels et Méthodes Chapitre III : Essais in vitro de recherches de moyens de lutte
I. Lutte chimique contre ALternaria dauci. :
Les méthodes du laboratoire qui permettent d’estimer les propriétés d’un produit in vitro
sont nombreuses, mais reposent toutes sur le même principe, celui de confronter le fongicide
et le champignon sur un support artificiel (Henni, 1987).
I.1 Les fongicides testés :
Deux substances antifongiques récemment synthétisées par le Laboratoire de Synthèse
Organique Appliquée de l’Université Es-sénia Oran ont étaient testées vis-à-vis les souches
d’Alternaria dauc. Ces substances sont des triazol.
Les Formules des deux substances utilisées sont les suivantes :
o
1[4-methoxyphenol].4methyl alcool [1, 2,3]-triazol. (Substance A).
o
1[4-Bromophenol].4methyl alcool [1, 2,3]-triazol. (Substance B).
Substance Fongicide Laboratoire Solubilité
Produit A Laboratoire de Chimie
Université ES-Sénia Oran
Acétone
Produit B Laboratoire de Chimie
Université ES-Sénia Oran
Acétone
H3CO NO
OH
N
N
Br NO
OH
N
N
-42-

Matériels et Méthodes Chapitre III : Essais in vitro de recherches de moyens de lutte
I.2 Préparation des solutions de fongicides :
Chaque produit à tester est soluble dans un solvant organique qui est l’acétone. (Tableau
ci-dessus).
Dans un premier temps, une solution mère de chaque produit est préparée à une concentration
de 105 ppm (partie par million), puis des dilutions seront effectuées avec un milieu liquide à
base de pomme de terre (PSA), pour arriver à chaque fois à la concentration voulue (50, 100,
250 et 500 ppm). Le 0 ppm comme étant le témoin, ne contient que le solvant incorporé dans
le milieu PSA.
Les différentes dilutions sont obtenues en respectant la loi suivante :
Le test est effectué sur un milieu solide. Après avoir calculé le V1, ce dernier sera complété
avec milieu PSA jusqu'à l’obtention du V2, ensuite le complexe (PSA-substance) sera
autoclave à une atmosphère 20 minutes à 120°c, sachant que, les fongicide se dénature à une
température supérieure à 120°c. en suite on coule ce milieu dans des boite de Petri à raison de
15ml par boite. Après solidification ; des implants mycéliens de 6 mm de diamètre sont
prélevés de la périphérie d’une préculture de 7 jours de nos souches d’Alternaria dauci, et
sont déposés au centre de la boite de Pétri contenant le mélange avec les 04 concentrations
pour chaque substance active à étudier (Braffio et al., 2004).
Pour chaque concentration et chaque produit, trois répétitions sont réalisées. L’incubation a
été pendant 7 jours et la lecture est réalisée les 3 éme et 7 éme jours.
Le pourcentage d’inhibition se calcule selon la formule suivante (Diallo et Diouf, 2000) :
C1 V1 = C2 V2
C1 : Concentration de la solution mère V1 : Volume pris de la solution mère
C2 : Concentration à préparer V2 : Volume final voulu du milieu PSA
Taux d’inhibition (%) = ((D témoin – D test) / D témoin) x 100
D : Diamètre
-43-

Troisième Partie :
Résultats et Discussions

Résultats

Résultats
I. Aspect morphologique de nos souches :
Après quelques jours d’incubation, l’aspect des colonies est circulaires, le mycélium aérien
cotonneux à duveteux de couleur vert olive, sombre ou noire, la face inferieur des colonies est
de la même pigmentation. (Planche : 06).
II. Caractères microscopiques :
L’observation microscopique des souches a révélé la présence de conidie de type
dictyospores de couleur brune avec de longue appendice, ce qui confirme que les souches
isolées appartiennent à l’éspece Alternaria dauci. (Planche : 07).
III. Effet des différents milieux de culture :
Nos souches ont une meilleure croissance sur le milieu PSA avec un aspect du
mycélien aérien cotonneux et une pigmentation naturelle verte olive foncée. La pigmentation
de la colonie au verso est plus sombre. On note ici que les souches P2 et 10631 ont une
meilleur croissance après 7 jours d’incubation (Histogramme : 01 et planche : 08).
Pour le milieu GZAPECK on a une croissance mycélienne de vitesse moyenne sauf pour
la souche 10631 qui a une forte croissance de 5.9cm après 7jours d’incubation, la
pigmentation de la colonie est noire au centre et vert jauni au contour ce qui montre la perte
de la sporulation.
Pour le milieu mathure on trouve une vitesse de croissance moyenne, sauf pour la P2 qui
atteint 6.13cm aprés 7 jours d’incubation avec pigmentation verte au centre et plus claire tout
autour.
Les résultats sur milieu malt gélosé agar ont révèlé que la vitesse de croissance est faible
par rapport aux autres milieux, sauf pour la P2 (6.1cm après 7 jours d’incubation), la
pigmentation de la colonie est verte foncée au centre et claire tout autour.
-46- -46-
-46-
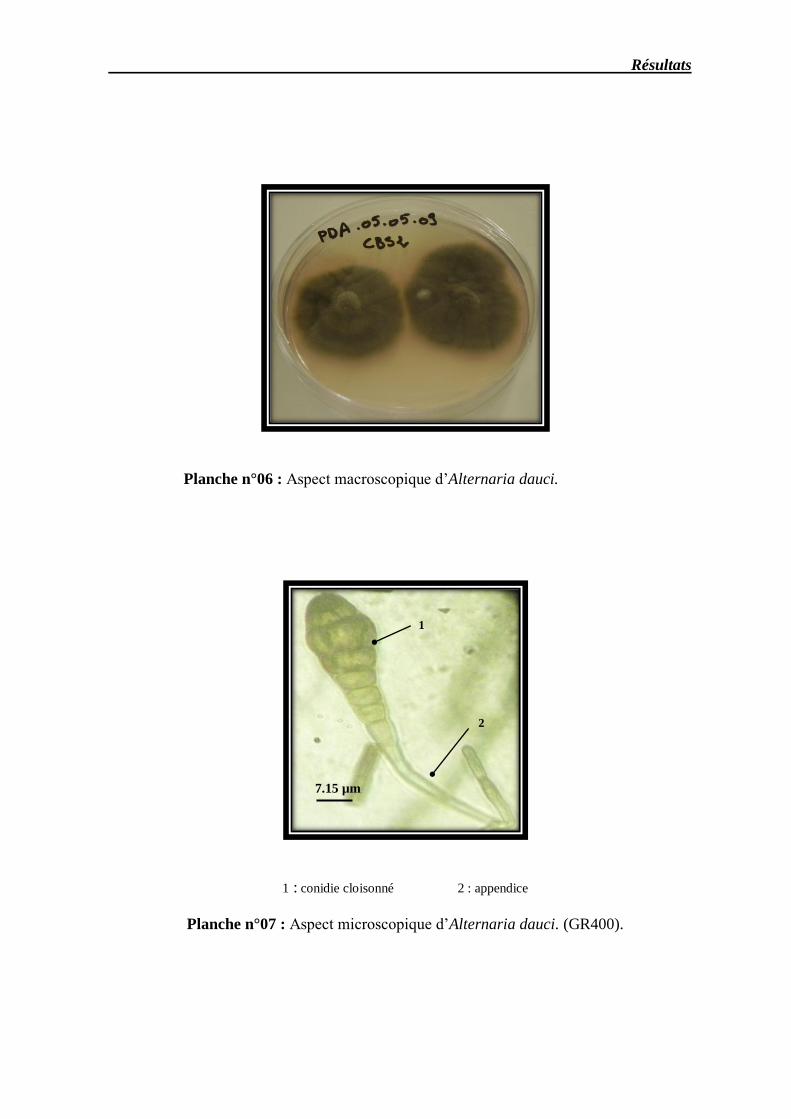
Résultats
Planche n°06 : Aspect macroscopique d’Alternaria dauci.
1 : conidie cloisonné 2 : appendice
Planche n°07 : Aspect microscopique d’Alternaria dauci. (GR400).
1
2
7.15 µm

Histogramme n° 01 : Représentation graphique de l’influence des différents milieux de
culture sur la croissance mycélienne d’Alternaria dauci après 7jours d’incubation.
(Résumé des donnés, annexe2).
Planche n°08 : Aspect de colonie d’Alternaria dauci sur les quatre milieux de culture après 7
jours d’incubation.
Résultats
0
1
2
3
4
5
6
7
8
PDA
Malt
Czapeck
Mathur
Gzapeck
Malth agar
PDA
Mathur
-48-

Résultats
IV. Etude physiologique d’Alternaria dauci :
IV.1 Influence de la température :
La croissance mycélienne est nulle à 37°C et 45°C et elle est faible à 4°C, elle se situe
entre 1.5cm à 2cm (après 7jours incubation), la colonie est de couleur blanche, qui indique la
perte de sporulation.
Par contre nous avons une meilleure croissance mycélienne entre 22°C et 28°C, pour
toutes les souches avec un diamètre qui atteint jusqu'à 6.78cm après 7 jours d’incubation,
ceci se rapproche de la bibliographie qui fixe la température optimale à la croissance des
Alternaria à 30°C (Koeing, 1995). (Hist. : 02 et planche : 09).
IV.2 Influence du pH du milieu :
On remarque qu’à pH 4 la croissance mycélienne est nulle après 3 jours d’incubation, et
après 7 jours d’incubation on a eu une croissance faible pour la P2 et la 10631.
Pour pH 6 on remarque une bonne croissance mycélienne pour toutes les souches avec une
gamme allant de 5.4cm à 6.85cm après 7 jours d’incubation.
Pour pH 8 on a une faible croissance mycélienne entre 2cm à 4.5cm après 7 jours
d’incubation. (Hist. : 03 et planche : 10).
IV.3 Influence des sources carbonées :
Les résultats représentés dans Histogramme : 04 et planche : 11) montrent que la
croissance mycélienne est presque identique sur le milieu PSA avec différentes sources de
carbones (Glucose, Fructose et Lactose) avec un diamètre qui atteint presque 7cm après 7
jours d’incubations.
IV.4 Influence des sources azotées :
La croissance mycéliennes de nos souches est presque identiques sur le milieu Gzapeck
avec comme sources d’azotes le nitrates de sodium (NaNO3) et le nitrate de potassium avec
un diamètre qui atteint jusqu'à 6cm après7jours d’incubation avec même aspect
morphologique, ce qui correspond à la bibliographie dont les meilleurs aliments décris pour
les champignons sont constitués de l’azote nitrique et de certaines substances azotées
organiques (Joly, 1964). (Hist. : 05 et planche : 12).
Pour Gzapeck modifié par l’addition du sulfate d’ammonium (NH 4SO4 ), il a donné des
colonies presque identiques pour toutes les souches, de couleur vert clair avec alentour de
couleur rosâtre, due à une absence de sporulation, cela peut s’expliquer par le fait que les
aliments azotés, plus complexes, vent entrainer une variation du pH non négligeable,
-49-

Résultats
provoquant des irrégularités de croissances et un arrêt de la sporulation dès que le pH décent
au dessous d’une valeur proche de pH3.2 (Joly, 1964) avec un diamètres de croissances qui
est faible.
IV.5 Influence de la lumière :
Nous avons obtenue une bonne croissance mycélienne dans les différentes conditions de
lumière (lumière continue, obscurité continue et photopériode) avec un diamètre jusqu'à 8cm
après 7 jours d’incubations avec même couleur de colonies sauf qu’elle est un peu claire dans
l’obscurité continue. (Hist. : 06 et planche : 13).
-50-
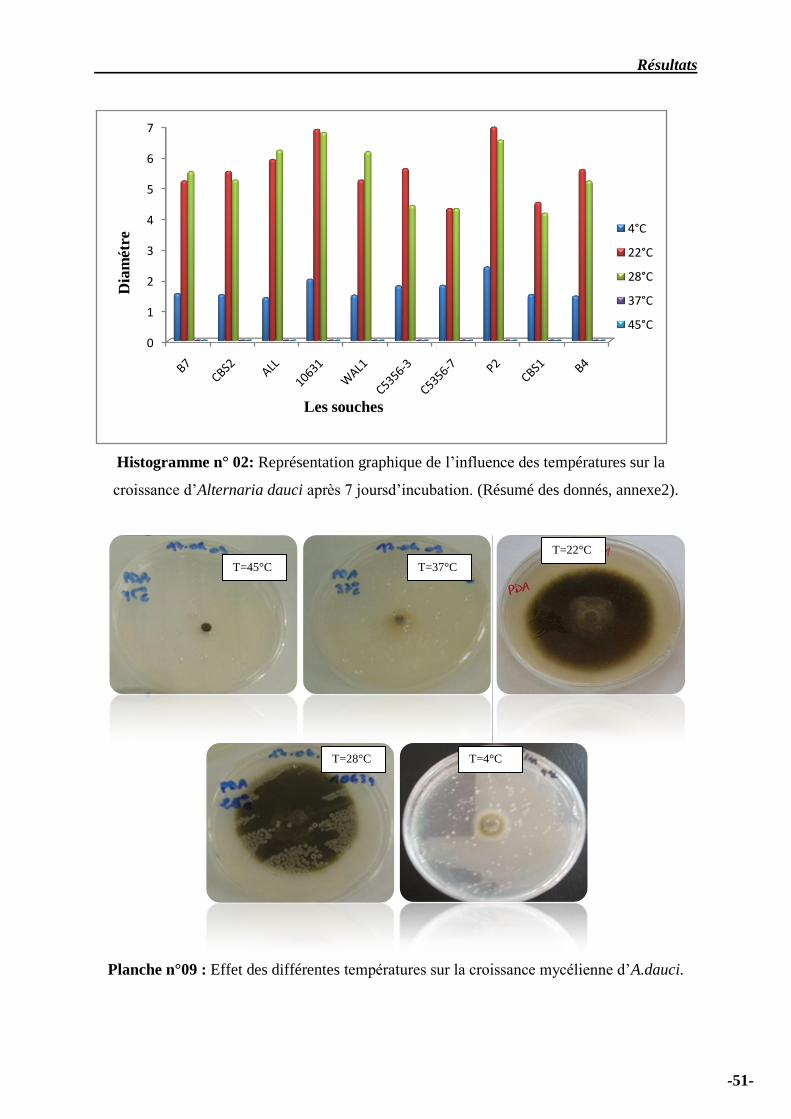
Résultats
Histogramme n° 02: Représentation graphique de l’influence des températures sur la
croissance d’Alternaria dauci après 7 joursd’incubation. (Résumé des donnés, annexe2).
Planche n°09 : Effet des différentes températures sur la croissance mycélienne d’A.dauci.
0
1
2
3
4
5
6
7D
iam
étr
e
Les souches
4°C
22°C
28°C
37°C
45°C
T=45°C
T=37°C
T=22°C
T=28°C
T=4°C
-51-

Résultats
Histogramme n° 03: Représentation graphique de l’influence du pH du milieu sur la
croissance d’Alternaria dauci après 7 jours d’incubation. (Résumé des donnés, annexe2).
Planche n°10 : Effet du différents pH du milieu sur la croissance mycélienne d’A.dauci.
0
1
2
3
4
5
6
7D
iam
étr
e
Les souches
pH04
pH06
pH08
pH=4
pH=6
pH=8
-52-
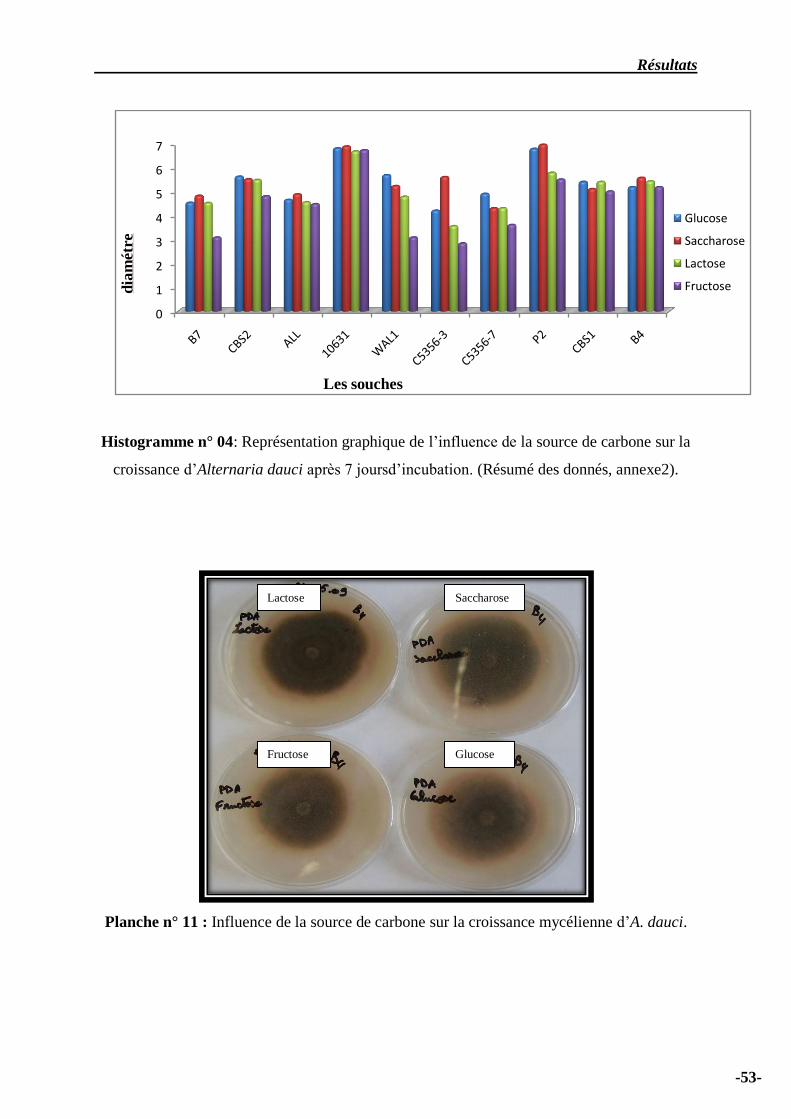
Résultats
Histogramme n° 04: Représentation graphique de l’influence de la source de carbone sur la
croissance d’Alternaria dauci après 7 joursd’incubation. (Résumé des donnés, annexe2).
Planche n° 11 : Influence de la source de carbone sur la croissance mycélienne d’A. dauci.
0
1
2
3
4
5
6
7
dia
métr
e
Les souches
Glucose
Saccharose
Lactose
Fructose
Lactose Saccharose
Fructose Glucose
-53-
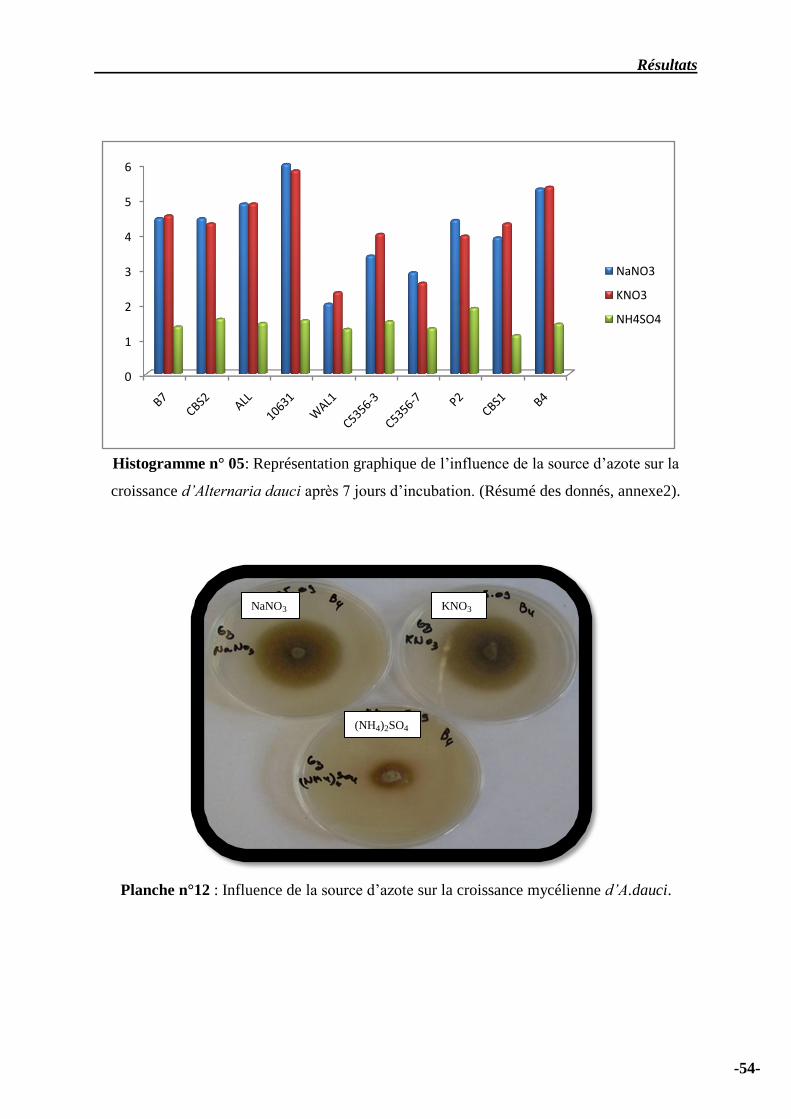
Résultats
Histogramme n° 05: Représentation graphique de l’influence de la source d’azote sur la
croissance d’Alternaria dauci après 7 jours d’incubation. (Résumé des donnés, annexe2).
Planche n°12 : Influence de la source d’azote sur la croissance mycélienne d’A.dauci.
0
1
2
3
4
5
6
NaNO3
KNO3
NH4SO4
NaNO3 KNO3
(NH4)2SO4
-54-
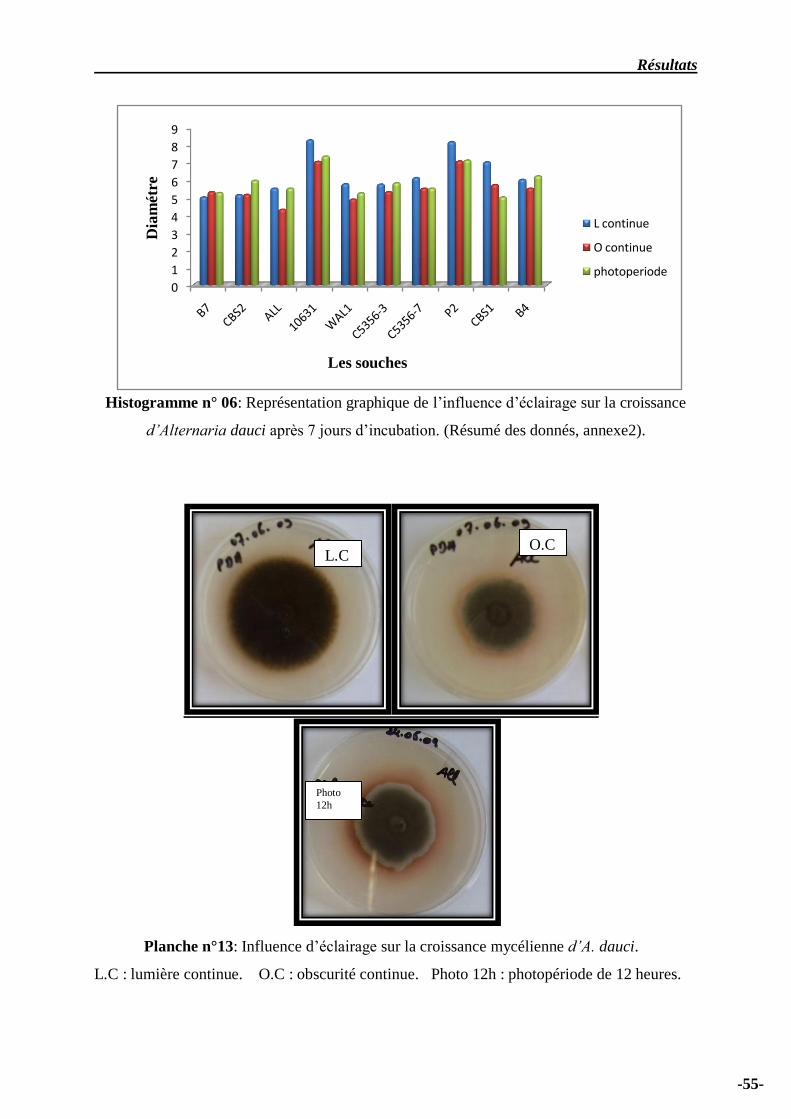
Résultats
Figure n° Influence de la source d’azote sur la croissance d’Alternaria dauci.
Histogramme n° 06: Représentation graphique de l’influence d’éclairage sur la croissance
d’Alternaria dauci après 7 jours d’incubation. (Résumé des donnés, annexe2).
Planche n°13: Influence d’éclairage sur la croissance mycélienne d’A. dauci.
L.C : lumière continue. O.C : obscurité continue. Photo 12h : photopériode de 12 heures.
0
1
2
3
4
5
6
7
8
9
Dia
métr
e
Les souches
L continue
O continue
photoperiode
Photo
12h
L.C O.C
-55-

Résultats
V. Pouvoir pathogène :
Les symptômes de brûlure foliaire sont petites, d’une couleur brunâtre et entouré d’un
halo de décoloration jaune (Champion, 1997). (Planche : 14, 15,16).
Nous avons débuté les notations des symptômes, 01 mois après l’inoculation, en utilisant
l’échelle de notation des lésions nécrotiques après inoculation d’Alternaria dauci (Pawelec et
al.,2006) (tableau n° 05 ) qui se base sur la recherche de surface foliaire nécrosée, la notation
exprimée en pourcentage:
Pour la variété Presto :
Inoculée avec la souche P2, on a plus de 60% de la surface foliaire nécrosée ce qui donne une
échelle 5.
Inoculée avec la souche B4, on a < 50% de la surface foliaire nécrosée ce qui donne une
échelle 3.
Inoculée avec la souche WAL1, on a presque 20% de la surface foliaire nécrosée ce qui
donne une échelle 1.
Pour la variété Boléro :
Inoculée avec la souche P2, on a > 50% de la surface foliaire nécrosée ce qui donne une
échelle 5.
Inoculée avec la souche B4, on a environ 30% de la surface foliaire nécrosée ce qui donne
une échelle 3.
Inocule avec la souche WAL1, on a presque 20% de la surface foliaire nécrosée ce qui donne
une échelle 1.
Pour la variété texto :
Inoculée avec la souche P2, on a > 40% de la surface foliaire nécrosée ce qui donne une
échelle 3.
Inoculée avec la souche B4, on a >25% de la surface foliaire nécrosée ce qui donne une
échelle 3.
Inoculée avec la souche WAL1, on a < 20% de la surface foliaire nécrosée ce qui donne une
échelle 1.
-56-

Résultats
Les symptômes de brulure foliaire constatée sur nos plantules, sont dus à la
contamination de ces derniers par nos souches inoculées.
Le réisolement à partir des feuilles infectées, confirme qu’il s’agit bien du champignon
Alternaria dauci, et que ce sont bien nos souches que l’on retrouve P2, B4 et WAL1.
(Planche :17,18).
Tableau n°03 : Echelle de notation des lésions nécrotiques après inoculation d’Alternaria
dauci sur les feuilles 2 et 3 de carotte (D’après Pawelec et al. , 2006 modifie).
Note Critères de classification
0 Absence de symptôme
1 Moins de 25% de la surface foliaire nécrosée
3 25% à 50% de la surface foliaire nécrosée
5 50% à 75%de la surface foliaire nécrosée
7 Plus de 75% de la surface foliaire nécrosée
9 Feuilles entièrement détruites
-57-

Résultats
Planche n°14:symptômes d’Alternariose sur les feuilles de carotte causé par la souche P2
après 21jours.
Planche n°15:symptômes d’Alternariose sur les feuilles de carotte causé par la souche B4
après 21jours.
Planche n°16:symptômes d’Alternariose sur les feuilles de carotte causé par la souche WAL1
après 21jours.
-58-
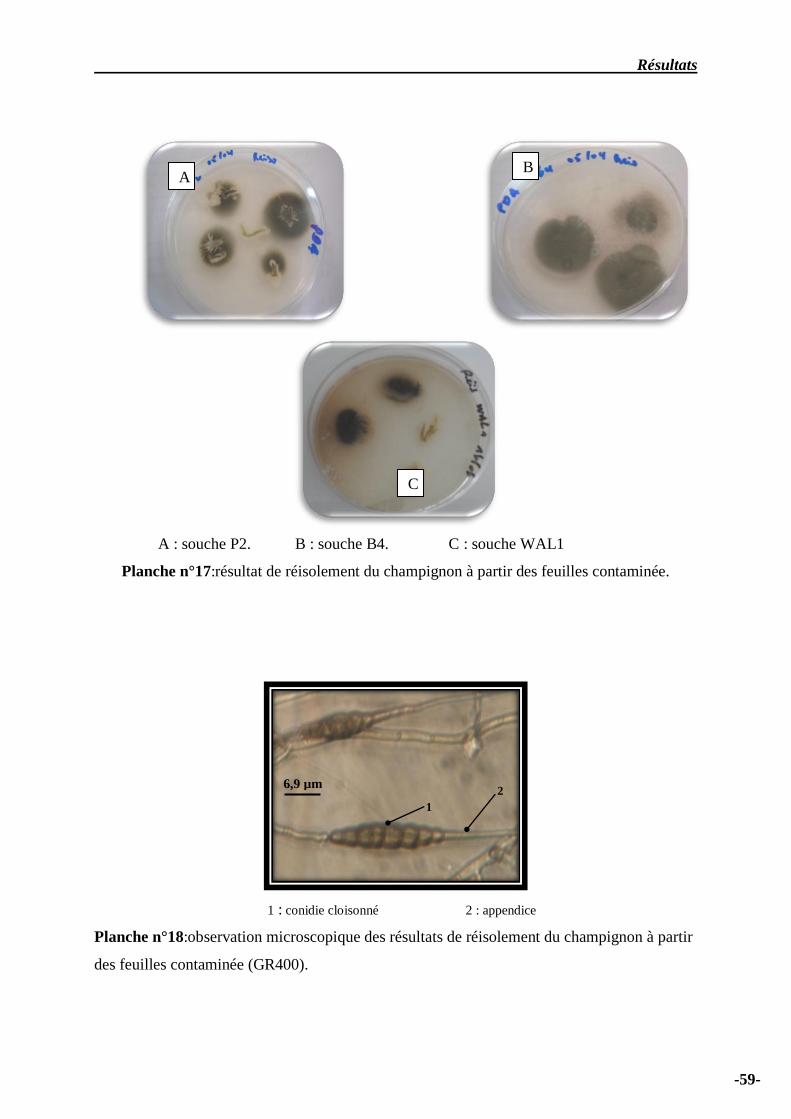
Résultats
A : souche P2. B : souche B4. C : souche WAL1
Planche n°17:résultat de réisolement du champignon à partir des feuilles contaminée.
1 : conidie cloisonné 2 : appendice
Planche n°18:observation microscopique des résultats de réisolement du champignon à partir
des feuilles contaminée (GR400).
A B
1
2 6,9 µm
C
-59-

Résultats
VI. Électrophorèse :
Le gel de polyacrylamide a servi pour l’étude isoenzymique des souches isolées
d’Alternaria dauci (WAL1, C5356-7, C5357-3, CBS2, B4, 10631, ALL, P2, CBS1, B7).
Quatre systèmes enzymatiques ont été étudiés.
VI.1 Estérases :
Ce système révèle un profil électrophorétique de 02 bandes chez tous les isolats
d’A.dauci Les bandes sont visibles, d’une intensité variable et d’une couleur mauve (Planche
n°19)
VI.2 Phosphatases :
Le profil électrophorétique des phosphatases des isolats d’A.dauci révèle la présence de deux
bandes lisibles et visibles d’une intensité variable et d’une couleur bleu (Planche n°20).
VI.3 Peroxydases :
Le profil électrophorétique des peroxydases des isolats d’A.dauci révèle la présence d’une
bande lisible et visible d’une intensité variable et d’une couleur rouge brique (Planche n°21).
VI.4 Catalase :
Le résultat positive des catalases se manifeste par l’apparition des bulles d’airs, donc le profil
électrophorétique des catalases des isolats d’A.dauci révèle la présence, de deux bandes
lisibles et visibles, constituée de bulle d’air (Planche n°22).
-60-

Résultats
Planche n°19: Profil électrophorétique des estérases Alternaria dauci
Planche n°20: Profil électrophorétique des phosphatases Alternaria dauci
WAL1 C5356-7 C5356-3 CBS2 B4 10631 ALL P2
CBS1 B7 B7
WAL1 C5356-7 C5356-3 CBS2 B4 10631 ALL P2 CBS1
B7
-61-

Résultats
Planche n°21: Profil électrophorétique des peroxydases Alternaria dauci
Planche n°22: Profil électrophorétique des catalase Alternaria dauci.
WAL1 C-7 C-3 CBS2 B4 10631 ALL P2 CBS1 B7
WAL1 C-7 C-3 CBS2 B4 10631 ALL P2
CBS1 B7
-62-

Résultats
VII. Lutte chimique contre les souches d’Alternaria dauci :
Les 02 substances chimiques testés in vitro contre A.dauci ont révèle l’efficacité de ces
substances, déterminée par le calcul du diamètre moyen de la colonie, pendant les 07 jours
d’incubation (Tableaux n°06) ainsi que par le calcule du taux d’inhibition de chaque produit
et à chaque concentration testée (voir annexe).
Les résultats obtenus sont illustrés par les Histogramme n°07 et 08 et Planche n° 19 et 20.
L’histogramme n°07 montre l’efficacité de la substance A, ainsi que L’histogramme n°08
pour la substance B.
Dans le cas des deux substances, les résultats montrent qu’en augmentant la concentration
on a une augmentation du taux d’inhibition qui est supérieur à 50% pour la majorité des
souches à la concentration 500ppm et qui peut atteindre jusqu'à 80% pour certaines souches.
Néanmoins, il est noté qu’en plein champ, les concentrations, utilisées sont inférieures à
10ppm.
En effet, les concentrations >, nécessitent des quiritaux de fongicides pour 1 hectare et
beaucoup de solvant.
-63-

Résultats
Tableau n°04 : Diamètre moyen (cm) des souches d’Alternaria dauci confronté à la
substance A
Substance A [0] [50] [100] [250] [500]
P2 5,2 4,4 4,2 3,2 2,6
B4 4,8 4 3,6 3 1,8
10631 5,3 4,6 4,4 2,7 2,2
CBS1 5 4,2 3,5 2,5 1,2
B7 3,7 3,1 2,8 2,2 1,1
CBS2 4 3,1 3 2,1 1,5
ALL 2,5 2 1,6 1,3 1
C5356-3 3,7 3,5 3,2 2,5 1,3
C5356-7 3,2 3,1 2,8 2,1 1,6
WaL1 5 4,8 4,5 3,1 1,7
Histogramme n° 07: Représentation graphique du % d’inhibition des souches d‘Alternaria
dauci confronté à la substance A. (Résumé des donnés, annexe2).
0
10
20
30
40
50
60
70
80
% d
'inhib
itio
n
les souches
%in[0]
%in[50]
%in[100]
%in[250]
%in[500]
-64-

Résultats
Tableau n°05 : Diamètre moyen (cm) des souches d’Alternaria dauci confronté à la
substance B
Substance B [0] [50] [100] [250] [500]
P2 5,5 5 3,7 3 2
B4 4,5 4,2 3,2 2,2 1,7
10631 5,6 5,5 3,4 2,8 1,9
CBS1 5 4,6 4,1 2,9 1
B7 3,7 3,1 2,8 2,2 1,1
CBS2 4 3,1 3 2,1 1,1
ALL 2,4 2,2 2,3 1,6 1,2
C5356-3 3,2 2,8 2,7 1,9 1,5
C5356-7 3,6 2,9 2,3 2,2 1,7
WAL1 5 4,6 4 2,6 2
Histogramme n° 08: Représentation graphique du % d’inhibition des souches d‘Alternaria
dauci confronté à la substance B. (Résumé des donnés, annexe2).
0
20
40
60
80
% d
'inhib
itio
n
Les souches
%in[0]
%in[50]
%in[100]
%in[250]
%in[500]
-65-
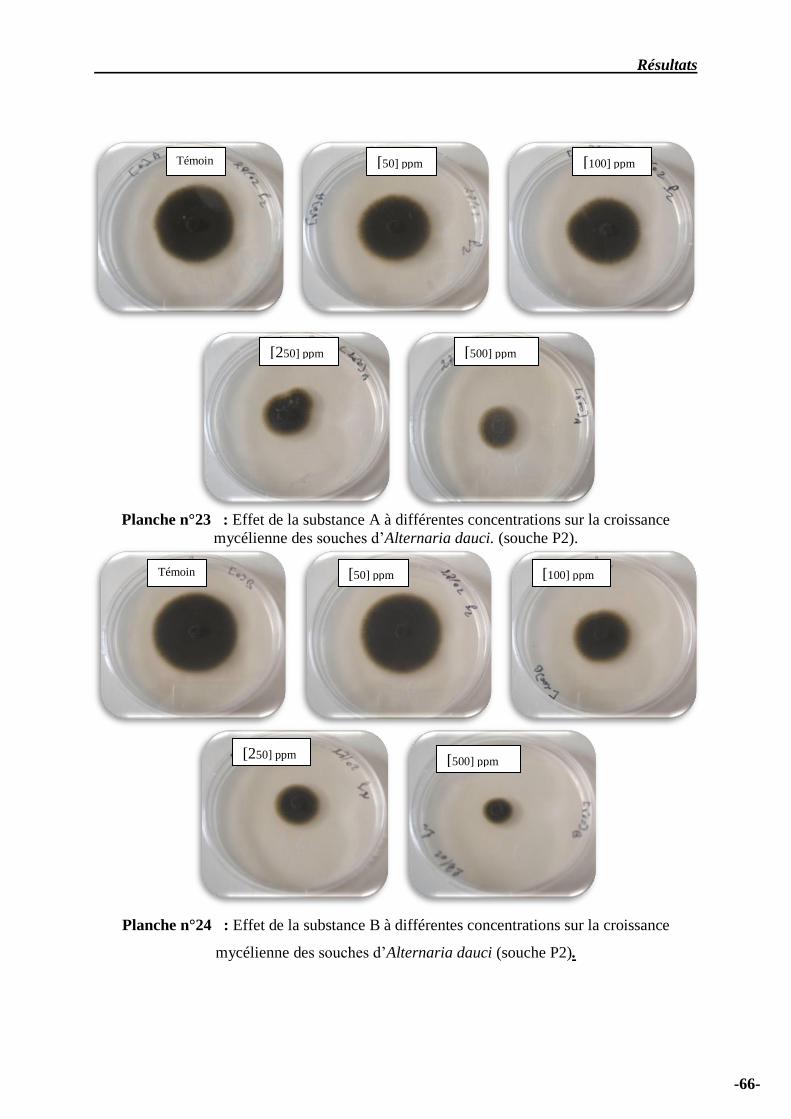
Résultats
Planche n°23 : Effet de la substance A à différentes concentrations sur la croissance
mycélienne des souches d’Alternaria dauci. (souche P2).
Planche n°24 : Effet de la substance B à différentes concentrations sur la croissance
mycélienne des souches d’Alternaria dauci (souche P2).
Témoin [50] ppm [100] ppm
[250] ppm [500] ppm
Témoin [50] ppm [100] ppm
[250] ppm [500] ppm
-66-

Discussions

Discussions
Notre travail de recherche s’inscrit dans le cadre de l’étude de la caractérisation
morphologique et physiologique de l’espèce Alternaria dauci, l’agent causal de
l’ Alternariose de la carotte, où on a cherché les meilleures conditions de croissance selon les
conditions qu’on peut favoriser, avec comme objectif : essayer un moyen de lutte in vitro,
par des substances chimiques (Synthétisées au laboratoire de Chimie de synthése organique et
appliquée à l’université d’Oran Es-senia ) d’un côté et de faire une étude de degré de
l’agréssivité et faire une comparaison entre les souches d’un autre côté.
1. L’étude morphologique : , l’aspect des colonies est circulaire, couleur vert olive sombre
ou noire, le mycélium aérien est rampant, la face inférieur des colonies est vert olive sombre
ou noire.
2. L’étude microscopique des souches nous a démontrés que le mycélium est cloisonné, les
conidies de type dictyospores de couleur brune avec de longue appendic,e ce qui confirme que
ces souches isolées sont Alternaria dauci (Konstantinovka, 2002).
Du point de vue morphologique, les variations s’avèrent importantes vis-à-vis des quatre
milieux utilisés (PSA, Czapeck, Mathur, Malt gélosé), avec une meilleure croissance et
meilleur sporulation sur le milieu PSA, ce qui a été décrit comme étant un très bon milieu par
Joly 1964.
3. S’agissant de l’étude physiologique,
3.1 Le test d’influence de la source carbonée sur la croissance mycélienne d’Alternaria
dauci a montré que le champignon se développe sur les quatre substrats et que la croissance
est presque la même, avec une légère préférence pour le saccharose.
3.2 L’influence de la source azotée sur la croissance mycélienne d’Alternaria dauci a montré
que la croissance mycélienne de nos souches isolées est presque identique sur le milieu
Gzapeck avec comme sources d’azotes le nitrate de sodium (NaNO3) et le nitrate de
potassium, ce qui correspond à la bibliographie dont les meilleurs aliments décris pour les
champignons sont constitués par l’azote nitrique et certaines substances azotées organiques
(Joly, 1964).
3.3 Influence de la température : La croissance mycélienne est nulle à 37°C et 45°C et
faible à 4°C, de couleur blanche, ce qui indique la perte de sporulation. La croissance est
bonne entre 22°C et 28°C pour toutes les souches avec un diamètre qui atteint jusqu'à 6.78cm,
-68-

Discussions
ceci se rapproche de la bibliographie qui fixe la température optimale à la croissance des
Alternaria à 30°C (koeing, 1995).
3.4 Influence du pH du milieu : On remarque qu’à pH=4 la croissance mycélienne est nulle
après 3 jours d’incubation, et après 7 jours on a eu une croissance faible pour la P2 et 10631.
Pour pH=6 on remarque une bonne croissance mycélienne pour toutes les souches et elle est
faible pour pH=8.
3.5 Influence de la lumière : La croissance mycélienne est bonne dans les 3 conditions de la
lumière (lumière continue, obscurité continue, et photopériodicité) avec la même couleur des
colonies, sauf qu’elle est un peu claire dans l’obscurité continue.
4. L’étude du pouvoir pathogène : a révèle que notre souche (WAL1) est moins virulente
par rapport à la souche P2 et elle a la même échèle que la soucheB4 (même résultat pour les
trois variétés de la carotte, presto, boléro et texto), avec une échelle qui est 5 pour la P2, 3
pour la B4 et 3 pour la WAL1ce qui confirme la virulence de notre souches vis-à-vis de la
carotte.
5. L’étude du polymorphisme isoenzymatique, par la révélation des estérases, des
phosphatases acides, des peroxydases ainsi que les catalases de nos souches d’Alternaria
dauci, nous a montré une similarité des profils électrophorétiques entre ces derniers pour
chaque système révélé.
A la lumière des résultats obtenus dans l’étude morphologique, microscopique et
physiologique (citées précédemment), nous avons lancé nos essais de lutte chimique contre
Alternaria dauci où on a utilisé deux substances chimiques qui sont synthétisé au sein du
laboratoire de synthèse organique et appliqué université d’Oran Es-senia et on a eu les
résultats suivants :
Dans le cas des deux substances, les résultats montre qu’en augmentant la concentration on a une
augmentation du taux d’inhibition, qui est supérieur à 50% pour la majorité des souches à la
concentration 500ppm et qui peut atteindre jusqu'à presque 80% pour certaines souches. Pour la
souche WAL1 elle est inhibé à 66% pour substance A et à 60% pour la substance B.
-69-

Conclusion et Perspectives

Conclusion et perspective
Alternaria dauci, agent de la brûlure foliaire peut détruire jusqu’à 99% du feuillage de la
plante entraînant une diminution du rendement pouvant atteindre 78% (Ben-Noon et al.,
2001).
Alternaria dauci est un véritable fléau pour les cultures. Il est indéniable que ces dégâts se
répercutent au niveau économique. L’approfondissement des connaissances sur Alternaria
dauci est un enjeu majeur de la recherche dans le but d’apporter des solutions de lutte contre
cette maladie que pourront utiliser les producteurs.
Les diverses expériences menées durant cette thèse ont permis de compléter l’analyse de la
diversité phénotypique des isolats de notre collection de souches, celles-ci sont réalisées à
différents stades du développement du champignon :
- Nous avons obtenu une meilleur croissance mycélienne et germination conidial sur
milieu de culture PSA, le mycélium est rampant de couleur vert olive allant jusqu’au
noir, les conidies sont cloisonnées avec un long appendice.
- L’étude des facteurs physico-chimiques nous a montré que la température optimale
pour la croissance de nos souches est comprise entre 22°C et 28°C, la croissance est
optimale à pH 6. Pour la source de carbone où nous avons eux une croissance
semblable dans les quartes sources de carbone utilisées. La meilleure source d’azote
pour la croissance de nos souches est le nitrate de sodium (NaNO3) et pour la lumière
on a la même croissance dans les trois temps d’exposition à la lumière.
- L’agressivité différentielle a été estimés sur trois variétés de carotte avec trois souches
(P2, B4 et WAL1), nos souches se sont montrés virulente vis-à-vis de ces variétés
avec un degré qui diffère d’une variété à l’autre et d’une souche à l’autre.
- L’étude du polymorphisme isoenzymatique, par la révélation des estérases, des
phosphatases acides, des peroxydases ainsi que les catalases de nos isolats
d’Alternaria dauci, nous a montré une similarité des profils électrophorétiques entre
les souches pour chaque système révélé.
- En ce qui concerne l’activité des deux substances chimiques sur la croissance
mycélienne de nos souches, les résultats obtenue montrent que ces substances ont un
effet croissant en augmentant les concentrations où on a l’augmentation du taux
d’inhibition, à [500] ppm on a un taux d’inhibition de 50% est plus, la souche WAL1
est inhibé à 66% par la substance A et 60% par la substance B.
-71-

Conclusion et perspective
En perspectives pour approfondir et compléter cette étude il serait intéresse de réaliser:
Une étude de l’efficacité des substances chimiques sur la sporulation du champignon.
Passer à l’étape in vivo pour voir leur effet sur la plante et voir s’il agit de manière
néfaste sur la plante et s’il a le même effet sur le champignon que le test in vitro.
Utiliser des techniques de biologie moléculaire qui nous permetent de détecté le
champignon dans les semences avant de les utilisées.
Faire appel à la génétique pour nous permettre de rechercher des variétés de carotte
résistantes à ce type de champignon.
La quantification des paramètres étudiés dans notre thèse en utilisant des outils
moléculaires permettant d’apprécier le degré de colonisation des tissus végétaux
infectés.
-72-

Références Bibliographiques

Références Bibliographiques
1. Agrios, G. N. (1997). Plant pathology. 4ème
édition. Academic Press, USA. 635 pp.
2. Ainsworth, b et Hawksworth, D.L. (1961). A dictionary of the fungy. Ed.
hawksworth.c.a.b. 747p.
3. Andersen, B. (2001). Chemical and morphological segregation of Alternaria tenuis, A.
gaisen and A. longipes. Mycological Research 105 :291-299.
4. Baaziz, M., 2000. Date palm culture in the Maghreb countries. Constraints and scientific
researches. Date palm International Symposium. Windhoek, Namibia, 22-25
5. Bacon, R. (2002).gestion de la résistance aux fongicides. Agriculture et agroalimentaire
Canada
6. Barash, I., Mor, H., Netzer, D., Kashman, Y. (1981). Production of zinniol by Alternaria
dauci and its phytotoxic effet on carrot. Physiological Plant Pathology. 19 : 7-16.
7. Bart, P. et Thomma, H.J. (1993). Alternaria sp. From general saprophyte te specific
parasite. Molecukar plant pathology (4), 225-236.
8. Ben-noon, E., Shtienberg, D., Shelvin, E., Vintal, H., Dinoor, A. (2001). Optimization of
chemical suppression of Alternaria dauci, the causal agent of Alternaria leaf blight in
carrots. Plant Disease. 85 : 1149-1156.
9. Berto, P., Belingheri, L., Dehorter, B. (1997). Production and purification of a novel
extracellular lipase from Alternaria brassicicola. Biotechnology Letters. 19 (6) : 533-536.
10. Berto, P., Commenil, P., Belingheri, L., Dehorter, B. (1999). Occurrence of a lipase in
spores of Alternaria brassicicola with a crucial role in the infection of cauliflower
leaves. FEMS Microbiology Letters. 180 : 183-189.
11. Blaise, K., Couture, L., Dostaler, D., et Berner, L., (2001). Variabilité phénétique du
Colletotrichum graminicola du sorgho. Can.J.Plant Pathol. 23.
12. Blancard, D. (1988). Maladies de la tomate : Observer, identifier et lutter. Ed. INRA,
Paris. P : 119, 125, 131, 164-165.
13. Boedo, C. (2006). Caractérisation des étapes précoces de la brûlure foliaire due à
Alternaria dauci, chez une variété de carotte sensible et une variété tolérante à la maladie
mémoire de Master.
14. Braffio, C.A., Kivrak, E. et Heller, W.E., (2004). Vulgarisation en cultures maraîchères,
Mise en évidence de champignon du genre Alternaria résistants à l’iprodione utilisé pour
la désinfection des semences de carottes. Le Marîcher.
15. Carillo, L. (2002). Los hongos de los Alimentos y forrajes :Alternaria.
-74-

Références Bibliographiques
16. Champion, R. (1997). Identifier les champignons transmis par les semences. Editions
INRA, Paris. 391 pp.
17. Chou, H.H et Wu, W.S. (2002). Phylogenetic analysis of internal transcriber spacer
regions of the genus Alternaria and the significance of filament-beaked conidia.
106 :164-169.
18. Cotty, P.J. et Misaghi, I. J. (1984). Zinniol production by Alternaria species.
Phytopathology. 74 : 785-788.
19. Cramer. R. A. et Lawrence, C. B. (2003). Identification of Alternaria brassicicola genes
expressed in planta during pathogenesis of Arabidopsis thaliana. Fungal Genetics and
Biology. 41 : 115-128.
20. Deyson, G et Delcourt, A. (1980). Cryptogamie (mycologie) générale et appliqué. Ed.
Sedes et C.D.U.
21. Diallo, B., et Diouf, A., (2000). Etude de l’activité analgésique du Pilostigma reticulatum
(Nguiguis). Odontostomatologie tropicale 92.
22. Dugdale, L. J., Mortimer, A. M., Isaact, S., Collin, H. A. (2000). Disease response of
carrot and carrot somaclones to Alternaria dauci. Plant Pathology. 49 : 57-67.
23. Erwin, C.D. (1977). Control of vascular pathogens. Antifungal compounds vol. 1, M.R.
Siegel et H.D. Sister, Ed. Marcel Dekker inc., New york, 163-2230.
24. Fan, X., Mattheis, J. P., Roberts, R. G. (2000). Biosynthesis of phytoalexin in carrot root
requires ethylene action. Physiologia plantarum. 110: 450-454.
25. Farrar, J. J., Pryor, B. M., Davis, R. M. (2004). Alternaria diseases of carrot. Plant
Disease. 88: 776-784.
26. Grillot, A. (1981). Protection des végétaux. Cour semestre 4, tome1, 95p.
27. Groves, J.W.and Skolko, A.J.(1944).Notes on seed-bome fungi ;II.Alternaria.Can. j.
bot.221, 219-234.
28. Guignard, J.L. (2001):Systématique moléculaire, Ed :Masson, Paris.
29. Guiraud, H et Glazy, P. (1980). L’analyse microbiologie dans l’industrie
alimentaire.Collection Génie Alimentaire.Ed. Usine nouvelle.Paris.P :43-53.
30. Hatta, R., Ito, K., hosaki, Y., Tanaka, T., Tanaka, A., Yamamoto, M., Akimitsu, K. et
Tsuge, T. (2002). A conditionally dispensable chromosome controls host-specific
pathogenicity in the fungal pathogen Alternaria alternata. Genetics. 161 : 59-70.
-75-

Références Bibliographiques
31. Hatzipapas, P., Kalosaka, K., Dara, A. and Christias, C. (2002). Spore germination and
appressorium formation in the entomophagic Alternaria alternata. Mycologycal
Research. 106 : 1349-1359.
32. Henni, J..E. (1998). Morphologie, pouvoir pathogène et diversité génétique chez
Fusarium oxysporium f.sp.lycopersici.172p.
33. Henni. J.E. (1987). Evaluation de l’efficacité de certains champignons antagonistes vis-à-
vis de Verticillium dahliae. Crypt. Mycol. P :8,203-207.
34. Henri, L., Alponse, M et jose, D. (1995). Cours de microbiologie générale. P7, 8,91-100,
163-167.
35. Isshiki,A.,Akimitsu, K., Yamamoto, M. et Yamamoto, H. (2001). Endopolygalacturonase
is essential for Citrus Black Rot caused by Alternaria citri but not Brown Spot caused by
Alternaria alternata. The American Phytopathological Society. 14 : 749-757.
36. Jacquet, M., (1979): Journée française d’étude et d’information .Acta. ENGPC.les
maladies des plantes. 745 p.
37. Johnson, L. J., Johnson, R. D., Akamatsu, H., Salamiah, A., Otani, H., Kohmoto, K. et
Kodama, M. (2001). Spontaneous loss of a conditionally dispensable chromosome from
Alternaria alternata apple pathotype leads to loss of toxin production and pathogenicity.
Currents of Genetics. 40 : 65-72.
38. Joly, P. (1964). Le genre Alternaria. Ed.P . le chevalier. Paris. P : 7, 8, 91-100, 163-167.
39. Koeing, H.(1995).Guide de mycologie médicale.Ellipdes.P :13, 171, 172.
40. Konstantinova, P. (2002). Development of secific primers for detection and identification
of Alternaria sp in carrot material by PCR and comparaison with blotter and plating
assays. Mycological research. 106 : 23-33.
41. Kurosaki, F. (1996). Partial purification and characterization of 6-hydroxymellein-O-
methyltransferase from elicitor treated carrot cells. Phytochemistry. 4 : 1023-1027.
42. Laemmli, U.K. (1970): Cleavage of structural proteins during the assembly of
bacteriophage T4. Nature 227: 680-685.
43. Langenberg, W. J., Sutton, J. C. et Gillespie, T. J. (1977). Relation of weather variables
and periodicities of airborne spores of Alternaria dauci. Phytopathology. 67: 879-883.
44. Leach, R. (1939).Biologie control and ecology of Amillaria mella (Vah1) Fr.
Trans.br.Myc. Soc. 23, 320-329.
45. Marinelli, F., Zanelli, U., Nutironchi, V. (1996). Toxicity of 6-methoxymellein and 6-
hydroxymellein to the producing carrot cells. Phytochemistry. 3 : 641-643.
-76-

Références Bibliographiques
46. Markert, C.L., Moller, F., (1959). Proc. Nat. Acad. Sci. USA 45:753-63.
47. Mcroberts, N. et Lennard, J. H. K. (1996). Pathogen behaviour and plant cell reactions in
interactions between Alternaria species and leaves of host and nonhost plants. Plant
Pathology. 45: 742-752.
48. Mecier, J., Roussel, D., Charles, M. T., Arul, J. (2000). Systemic and local responses
associated with UV and pathogen induced resistance to Botryits cinerea in stored carrot.
Phytopathology. 90: 981-986.
49. Messiaen, C.M. (1981).Les variétés résistantes : Méthode de lutte contre les maladies de
la plante maraichère. Paris.
50. Messiaen, C.M., Blancard, D., Rouxel, F., Laform.R. (1991).Les maladies des plantes
maraichéres, 3éme Edition, I.N.R.A.,Paris.
51. Montillet, J-L. (1986). Dosage radioimmunologique du zinniol : application à l’étude de
cette toxine dans l’alternariose de la carotte. Thèse de doctorat, Université de Toulouse.
73 pp.
52. Moreau, C., (1968) : Moisissures toxiques dans l’alimentation. Encyclopédie
mycologique t.xxxv.p : 10-37.
53. Moss, D.W., (1984). Acid phosphatases. In Methods of enzymatic analysis. Volume IV.
Enzymes 2: esterases, glycosidases, lyases, ligase (Bergmeyer J ed) Verlag Chemie,
Weinheim, 92-106.
54. Nechev, G. (1989). Cours de phyopathologie.Netzer, D., Kenneth, R. G. (1969).
Persistance and transmission of Alternaria dauci (Kuhn) Groves & Skolko in the semi-
arid conditions of Israel. Annual Applied of Biology. 63 : 289-294.
55. Neergaard, P. (1945). Danish Species of Alternaria and stremphylium. London, UK :
Oxford University Press.
56. Nees, V.E. (1817).System der pilze and Schwamme.234p.In Joly.(1964).
57. Netzer, D.,Kenneth,R. G.(1969). Persistance and transmission of Alternaria dauci (Kuhn)
Groves & Skolko in the semi-arid conditions of Israel. Annual Applied of Biology. 63:
289-294.
58. Otani, H., Kohnobe, A., Kodama, M. and Kohmoto, K. (1998b). Production of a host-
specific toxin by germinating spores of Alternaria brassicicola. Physiological and
Molecular Plant Pathology. 52 : 285-295.
59. Otani, H., Kohnobe, A., Kodama, M. et Kohmoto, K. (1998a). Involvement of host
factors in the production of a protein host-specific toxin produced by Alternaria
brassicicola. Molecular Genetics of Host Specific Toxins in Plant Disease. 13: 63-69.
-77-

Références Bibliographiques
60. Owens,L.D., Cilbert, R.C., Griebel, C.E et Menzies, J.D. (1969). Sooty molde infecting
citrus in chile : Identification of plant volatiles that stimulate microbes respiration an
growth in soil. Phytopathology 59, 1468-1472.
61. Pawelec A., Dubourg C. et Briard M. (2006). Evaluation of carrot resistance to Alternaria
leaf blight in controlled environments. Plant Pathology. 55 : 68-72.
62. Pawelec, A. (2005). Caractérisation des déterminants génétiques et moléculaires de la
résistance à Alternaria dauci chez la carotte. Thèse de doctorat, Université d’Angers. 98
pp.
63. Peron, J-Y, (2006).Production légumières. Edition Lavoisier, 2eme
edition. Page 198-205.
64. Philouzr, J et Laterrot, H. (1992). La tomate amélioration des espéces végétales cultivés :
Objevtifs et critéres de seléction. Ovrage collectifs coordpnées par : Gallais, A et
bannterot, H. Ed .INRA. paris : 379-391.
65. Pryor, B. M., Strandberg, J. O., Davis, R. M., Nunez, J. J., Gilbertson, R. L. (2002).
Survival and persistence of Alternaria dauci in carrot cropping systems. Plant Disease.
86: 1115-1122.
66. Punja, Z. K. (2005). Transgenic carrots expressing a thaumatin-like protein display
enhanced resistance to several fungal pathogens. Canadian Journal of Plant Pathology.
27: 291-296.
67. Qacif, N., Bendiab, K. et Baaziz, M., (2006). Enzymologie et métabolisme. Aspects
qualitatif des peroxydases du palmier dattier (Phoenix dactylifera L.) étudiés chez des
pieds mâles et femelles. Deuxième Congrés Internationalde Biochimie. Agadir, Maroc. 9-
12.
68. Regnault, J.P., (1990) : Microbiologie générale .Ed, Vigot, Paris et Décarie, Montréal,
844p.
69. Rioux,S., Voldeng, S.F. (1999). Des plantes résistantes aux maladies : mythe ou réalité ?.
Cerom n°99.01.
70. Robert, R.G. (2002). Fragment pattern analysis and morphological segregation of small-
spored Alternaria species and species group. Mycological Research. 104 :151-160.
71. Rose, A.H. (1978). Responses to the chemical environnement. Academic Press.Londes
72. Rotem, J. (1994).The Genus Alternaria: Biology, Epidemiology and pathogenicity. St
Paul, MN, USA: APS press.
73. Rucci, T. (1986). Study of pathogenicity of causal agents wilt in tomate. Bulletin ;
Shkenkave Bujqesore, 25(2) : 91-97.
-78-

Références Bibliographiques
74. Sandrock, R. W. et vanetten, H. D. (1998). Fungal sensitivity to and enzymatic
degradation of the phytoanticipin alpha-tomatine. Phytopathology. 88 : 137-143.
75. Sellam, A., Poupard, P. et Simoneau, P. (2005). Physiological and molecular responses of
Alternaria brassicicola to exposure to host plant antimicrobial compounds. The 23th
Fungal Genetics. USA Californie. Fungal Genetics Newletters. 52.
76. Simmons E. G. (1967). Typification of Alternaria, Stemphylium and Ulocladium.
Mycologia. 59: 67-92
77. Simmons E. G. (1986). Alternaria themes and variations (22-26). Mycotaxon 25: 287-309
78. Simmons E. G. (1992). Alternaria taxonomy : current status, viewpoint ; challenge. In
Alternaria biology, Plant Diseases and Metbolites, EDS. J. Chekkovsci and A. Visconti.
Elsevier Science Publishers, Amsterdam. 135 p.
79. Simmons, E. G. (1995). Alternaria themes and variations. Mycotaxon. 55: 55-163.
80. Simon, P. W. et Strandberg, J. O. (1998). Diallel analysis of resistance in carrot to
Alternaria leaf blight. Journal of American Society of Horticultural. 123: 412-415.
81. Soteros, J. J. (1979). Pathogenicity and control of Alternaria radicina and A. dauci in
carrots. New Zealand Journal of Agricultural Research. 22 : 191-196.
82. Soylu, S., Kurt, S., Soylu, E. M. et Tok, F. M. (2005). First report of Alternaria leaf
blight caused by Alternaria dauci on carrot in Tukey. Plant pathology. 54 : 252.
83. Stranberg, J. O. (1977). Spore production and dispersal of Alternaria dauci.
Phytopathology. 67: 1262-1266.
84. Strandberg, J. O. (1983). Infection and colonization of inflorescences and mericarps of
carrot by Alternaria dauci. Plant Disease. 67 : 1351-1353.
85. Strandberg, J. O. (1987). Isolation, storage and inoculum production methods for
Alternaria dauci. Phytopathology. 77: 1008-1012.
86. Takaichi, M., oeda, K. (2000). Transgenic carrots with enhanced resistance against two
major pathogens, Erysiphe heraclei and Alternaria dauci. Plant Science. 153 : 135-144.
87. Thomma, B. P. H. J. (2004). Alternaria spp. : from general saprophyte to specific
parasite. Molecular Plant Pathology. 4 : 225-236.
88. Thuleau, P., Graziana, A., Rossignol, M., Kauss, H., Auriol, P., Ranjeva, R. (1998).
Binding of the phytotoxin zinniol stimulates the entry of calcium into plant protoplasts.
Proceedings of the National Academy of Science of the USA. 85: 5932-5935.
-79-

Références Bibliographiques
89. Trail, F. et Köller, W. (1993). Diversity of cutinases from plant pathogenic fungi:
purification and characterization of two cutinases from Alternaria brassicicola.
Physiological and Molecular Plant Pathology. 42 : 205-220.
90. Trottin-Caudal, Y et Millot, P. (1993). Lutte intégrée contre les ravageurs sur tomate sous
abri. Situation et perspectives. Infos-ctifl. n°96.33.
91. Viennot, B. (1949). Les champignons parasites des plantes cultivés. Ed. Masson. et Cie,
Paris.
92. Walter, J.C. (1971). Fusarium wilts of tomato. Ann. Phyt. Soc. St Paul, Minnesta monog.
n°6.
93. Walton, J. (1996). Host-selective toxins: agents of compatibility. The Plant Cell. 8: 1723-
1733.
94. Yao, C. et Köller, W. (1995). Diversity of cutinases from plant pathogenic fungi :
different cutinases are expressed during saprophytic and pathogenic stages of Alternaria
brassicicola. Physiological and Molecular Plant Pathology. 8 : 122-130.
-80-

Annexes

Annexes
Annexe I : Composition des milieux de culture
- Tous les milieux de culture sont stérilisés à l’autoclave à 120 °C pendant 20 minutes sous
une pression de 1 bar.
* Milieu PSA (g/ l) : (pomme de terre saccharose agar) d’après Larpent et Gourand, 1985.
Pomme de terre …………………….. 200 g
Saccharose …………………….. 20 g
Agar agar …………………….. 20 g
Eau Distillée q.s.p. …………………….. 1000 ml
- La pomme de terre est épluchée, coupée en petits dés puis cuite dans un litre d’eau distillée.
Après cuisson, le filtrat est récupéré, ajusté à 1000 ml puis ajuster le pH à 5,5.
* Milieu Czapek (g/ l): d’après Loquin1984.
Saccharose …………………….. 30 g
KH2PO4 …………………….. 1 g
KCl …………………….. 0,5 g
Na2No3 …………………….. 2 g
MgSO4 7H2O …………………….. 0,5 g
FeSO4 …………………….. 0,1 g
Agar agar …………………….. 20 g
Eau Distillée q.s.p. …………………….. 1000 ml
pH = 5,6
* Milieu Mathur (g/ l):
Extrait de levure …………………….. 0,5 g
KH2PO4 …………………….. 2,72 g
Peptone …………………….. 1,5 g
MgSO4 7H2O …………………….. 1,23 g
Glucose …………………….. 2,8 g
Agar agar …………………….. 20 g
Eau Distillée q.s.p. …………………….. 1000 ml
pH = 5,6
-82-

Annexes
* Milieu Malth agar : d’après Koenig 1995.
Extrait de Malth …………………….. 20 g
Glucose …………………….. 20 g
Peptone …………………….. 1 g
MgSO4 7H2O …………………….. 1,23 g
Glucose …………………….. 2,8 g
Agar …………………….. 20 g
Eau Distillée q.s.p. …………………….. 1000 ml
pH = 5,6
* Gélose 2 % :
Agar agar …………………….. 20 g
Eau Distillée q.s.p. …………………….. 1000 ml
Les sources de carbone utilisé :
Sucres : Formules :
Glucose C6H12O6
Saccharose C12H22O11
Fructose C6H12O6
Lactose C12H22O11
Les sources d’azote utilisé :
Sources d’azote : Formules :
Nitrate de sodium NaNO3
Nitrate de potassium KNO3
Sulfate d’ammonium NH4SO4
-83-

Annexes
Annexe II : Composition des solutions d’électrophorèse
Composition des gels de polyacrylamide.
Solutions
Gel de séparation
10%
Gel de concentration
5%
Solution d’acrylamide 30%
Solution de bis-acrylamide 1%
Tampon de séparation pH = 8,8
Tampon de concentration pH = 6,8
Solution SDS 10% (pour le système SDS-
PAGE)
Eau distillée
Temed
Persulfate d’Ammonium 10%
5 ml
5 ml
1,95 ml
/
0,15 ml
4,1 ml
50 µl
100 µl
2,5 ml
3,9 ml
/
3,75 ml
0,15 ml
4,65 ml
50 µl
100 µl
* Tampon de charge (PAGE) :
Glycérol …………………….. 5 g
Bleu de bromophénol 1% …………………….. 1 g
Eau Distillée q.s.p. …………………….. 10 ml
Le tampon est réparti à raison de 0,5 ml par tube à hémolyse avant d’être conservé au
congélateur jusqu’au moment de l’emploi.
Solution d’acrylamide : 30 % :
Acrylamide …………………. 30g
Eau distillée ………………… 1000ml
-84-

Annexes
Solution de bis –acrylamide 1% :
bis –acrylamide………………. 1g
Eau distillée ………………… 1000ml
* Tampon de migration :
(Tris : 0,25 M ; Glycine : 1,92 M ; SDS : 0,1 % ; pH=8,3)
Tris …………………….. 3,03 g
Glycine …………………….. 14,4 g
SDS (pour le système SDS-PAGE) …………………….. 1 g
Eau Distillée q.s.p. …………………….. 1000 ml
Le tampon est conservé au réfrigérateur.
* Tampon de séparation (Tris-HCl : 1,5 M ; pH = 8,8) :
Tris …………………….. 36,3 g
Eau Distillée q.s.p. …………………….. 100 ml
Le pH est ajusté avec environ 24 ml d’HCl 1N, puis complété à 200 ml avec de l’eau
distillée. Après filtration la solution est conservée à 4°C.
* Tampon de concentration (Tris-HCl : 0,5 M ; pH = 6,8) :
Tris …………………….. 3 g
Eau Distillée q.s.p. …………………….. 20 ml
Le pH est ajusté avec environ 24 ml d’HCl 1N, puis complété à 200 ml avec de l’eau
distillée. Après filtration la solution est conservée à 4°C.
-85-

Annexes
Les mélanges réactionnels pour les révélations enzymatiques.
NB : Les chiffres écrits en gras entre parenthèses (1 à 6) renvoient à la liste des solutions de
révélation en Annexe.
Enzyme
Mélange réactionnel
Estérases Scandalios (1969)
Un ester carboxylique + H2O
Est un alcool + un anion
acide carboxylique
Fast Blue RR 40 mg
Tampon Phosphate (0,1 M; pH = 7,1) (1) 50 ml
Bien dissoudre, Ajouter:
β-Naphtyl Acétate 2% (0,02g dans 1 ml Acétone) 2
ml
α-Naphtyl Acétate 2% (0,02g dans 1 ml Acétone) 2
ml
Rincer et Fixer.
Phosphatase Acide (PAC)
Pai, Endo et Oka (1975)
α-Naphtyl Phosphate
PAC α-Naphtol Acétate
α-Naphtyl Acide Phosphate 50 mg
Fast Garnet GBC 50 mg
Tampon Acétate (0,05 M; pH = 5) (2) 50 ml
Incuber 3 heures à 40° C, Rincer et Fixer.
Peroxydase
Shaw et Prasad
Pox
2 H2O2 2 H2O2 + O2
3 Amino, 9 Ethyl Carbazole 40 mg
Diméthyl formamide 5 ml
Tampon Acétate (0,05 M ; pH = 5) 92,5 ml
Solution de Ca Cl2 (0,1 M) (3) 2 ml
Solution de H2O2 à 30% 30 µl
Incuber 30 à 60 minutes, Rincer et Fixer.
Catalase
Brewer (1970)
Cat
2 H2O2 2 H2O2 O2
H2O2 I-
I2
Solution Thiosulfate (0,06 M) (4) 30 ml
Solution H2O2 à 1% (5) 70 ml
Mélanger les deux solutions puis les verser sur le gel
et attendre 2 minutes.
Ajouter la solution IK (0,09 M) (6)
Photographier immédiatement.
-86-

Annexes
* Tampon Na-phosphate 0,1 M – pH = 7,1 (1) :
Solution A : KH2PO4, H2O …………… 2,76 g dans 100 ml H2O
Solution B : Na2HPO4, 2 H2O …………… 7,12 g dans 100 ml H2O
Ajuster le pH de la solution A à 7,1 avec la solution B.
* Tampon Acétate 0,05 M – pH = 5 (2) :
Acétate de Sodium, 3H2O …………………….. 3,4 g
Eau Distillée q.s.p. …………………….. 475 ml
Ajuster à pH = 5 avec HCl 1N, Compléter à 500 ml avec H2O.
* Solution CaCl2 0,1 M (3) :
CaCl2 …………………….. 1,1 g
Eau Distillée q.s.p. …………………….. 100 ml
* Solution thiosulfate de Na 0,06 M (4) :
thiosulfate de Na …………………….. 0.075 g
Eau Distillée q.s.p. …………………….. 50 ml
*H2O2 (5): H2O2 …………………….. 3.3 ml
H2O …………………….. 100 ml
* Solution IK 0,09 M (6) :
IK …………………….. 0.15 g
Eau Distillée q.s.p. …………………….. 100 ml
-87-

Annexes
Annexe III : Calculs des diamètres
I/ Etude morphologique des souches d’Alternaria dauci :
1/ Diamètre quotidien des souches d’Alternaria dauci sur différents milieux : après 03jours
d’incubation
3° Jour PDA Malt Czapek Mathur
B7 2,1 1,15 1,68 1,67
CBS2 2,25 1,32 1,75 1,9
ALL 2,5 1,2 1,8 1,5
10631 3,1 2 2,6 2,55
WAL1 2,58 1,88 1,83 1,3
C5356-3 2,48 1,65 2,3 1,48
C5356-7 2 1,5 1,8 1,5
P2 3,43 2,63 2,15 2,6
CBS1 2,5 1,6 2,3 2,34
B4 2,42 1,4 1,7 1,85
1I/ Diamètre quotidien des souches d’Alternaria dauci sur différents milieux : après 07jours
d’incubation
7° Jour PDA Malt Czapeck Mathur
B7 5,25 1,95 4,6 3,68
CBS2 5,95 2,68 3,92 4,55
ALL 5,5 2,2 4,7 3,1
10631 7,33 4,77 6,1 5,83
WAL 5,23 2,85 3,23 3
C5356-3 5,8 3,03 4,48 2,93
C5356-7 5,5 3 3,5 3,4
P2 7,1 6,13 3,93 5,9
CBS1 5 5,43 3,9 5,05
B4 6,2 2,73 4,28 4,48
I1I/ Diamètre quotidien des souches d’Alternaria dauci sur différents milieux : après
14jours d’incubation
14° Jour PDA Malt Czapeck Mathur
B7 8 4,48 7,6 7,13
CBS2 8,5 5,37 7,47 7,25
ALL 7,5 4,1 7,8 6,3
10631 8,5 7,8 8,35 7,68
WAL 8,08 4,2 5,63 5,93
C5356-3 8,5 4,2 5,98 5,23
C5356-7 8,3 5 6 6,5
P2 8,5 8,5 6,68 8,5
CBS1 8,5 8,5 6,3 8,5
B4 8,5 5,2 7,58 7,13
-88-

Annexes
II. Etude des caractères physico-chimiques des souches d’Alternaria dauci :
1/a. Diamètre des souches d’Alternaria dauci avec les différentes sources de carbone : après
03jours
3° Jour Glucose Saccharose Lactose Fructose
B7 2,53 1,23 2,45 2,4
CBS2 2,45 2,48 2,62 2,45
ALL 2,45 2,22 2,25 2,33
10631 2,97 3,1 3,08 2,9
WAL1 2,8 2,43 2,13 1,73
C5356-3 2,15 2,6 1,8 1,75
C5356-7 2,2 1,8 2 1,9
P2 3,18 3,35 2,65 2,73
CBS1 2,6 2,7 2,6 2,6
B4 2,6 2,65 2,78 2,65
1/b. Diamètre des souches d’Alternaria dauci avec les différentes sources de carbone : après
07jours
7° Jour Glucose Saccharose Lactose Fructose
B7 4,53 3,08 4,52 4,82
CBS2 5,62 4,77 5,48 4,8
ALL 4,65 4,05 4,88 4,47
10631 6,8 6,88 6,67 6,72
WAL1 5,68 5,23 4,78 3,08
C5356-3 4,2 5,6 3,55 2,83
C5356-7 4,9 4,3 4,3 3,6
P2 6,78 6,95 5,78 5,5
CBS1 5,4 5,1 5,4 5
B4 5,18 5,57 5,42 5,18
1/c. Diamètre des souches d’Alternaria dauci avec les différentes sources de carbone : après
14jours
14° Jour Glucose Saccharose Lactose Fructose
B7 6,87 6 6,85 7,3
CBS2 8,07 7,7 7,97 7,93
ALL 7,78 6,8 7,75 7,68
10631 8,5 8,5 8,5 8,5
WAL1 8,5 8,5 8 5,3
C5356-3 5,88 7,85 6,13 5
C5356-7 8 7,1 7,7 7
P2 8,5 8,5 8,5 8,5
CBS1 8 7,6 7,9 7,8
B4 7,93 8,5 7,78 7,93
-89-
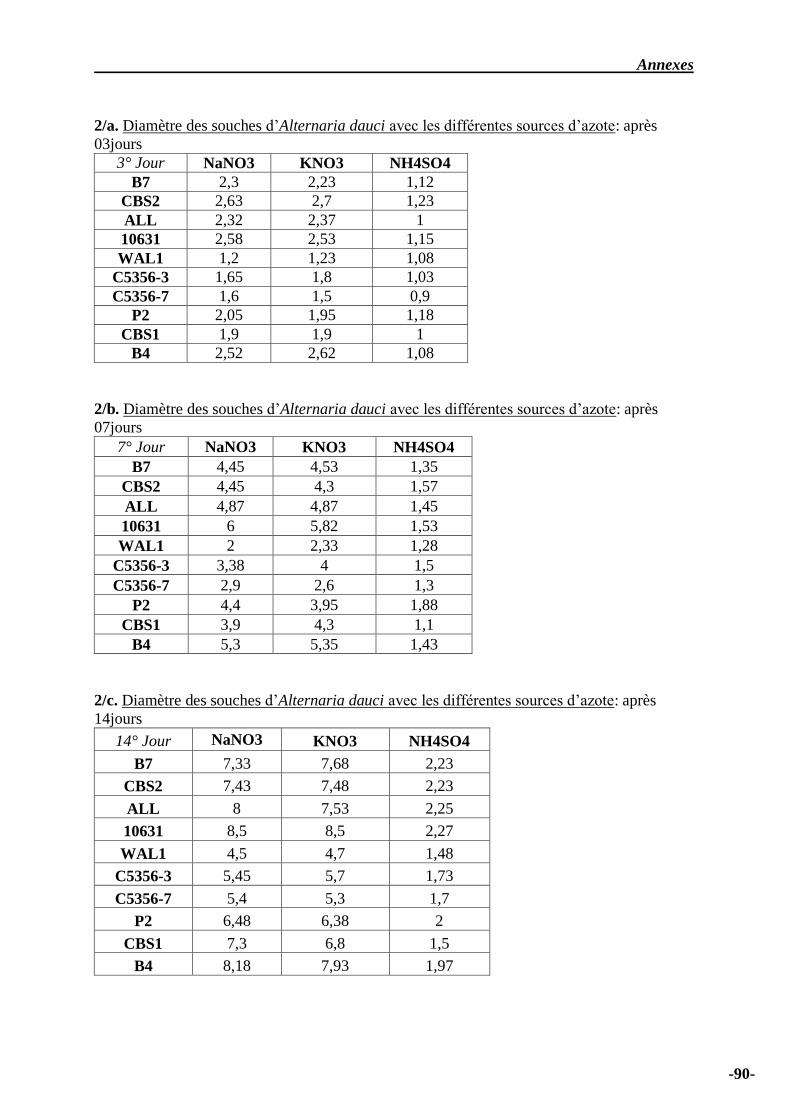
Annexes
2/a. Diamètre des souches d’Alternaria dauci avec les différentes sources d’azote: après
03jours
3° Jour NaNO3 KNO3 NH4SO4
B7 2,3 2,23 1,12
CBS2 2,63 2,7 1,23
ALL 2,32 2,37 1
10631 2,58 2,53 1,15
WAL1 1,2 1,23 1,08
C5356-3 1,65 1,8 1,03
C5356-7 1,6 1,5 0,9
P2 2,05 1,95 1,18
CBS1 1,9 1,9 1
B4 2,52 2,62 1,08
2/b. Diamètre des souches d’Alternaria dauci avec les différentes sources d’azote: après
07jours
7° Jour NaNO3 KNO3 NH4SO4
B7 4,45 4,53 1,35
CBS2 4,45 4,3 1,57
ALL 4,87 4,87 1,45
10631 6 5,82 1,53
WAL1 2 2,33 1,28
C5356-3 3,38 4 1,5
C5356-7 2,9 2,6 1,3
P2 4,4 3,95 1,88
CBS1 3,9 4,3 1,1
B4 5,3 5,35 1,43
2/c. Diamètre des souches d’Alternaria dauci avec les différentes sources d’azote: après
14jours
14° Jour NaNO3 KNO3 NH4SO4
B7 7,33 7,68 2,23
CBS2 7,43 7,48 2,23
ALL 8 7,53 2,25
10631 8,5 8,5 2,27
WAL1 4,5 4,7 1,48
C5356-3 5,45 5,7 1,73
C5356-7 5,4 5,3 1,7
P2 6,48 6,38 2
CBS1 7,3 6,8 1,5
B4 8,18 7,93 1,97
-90-
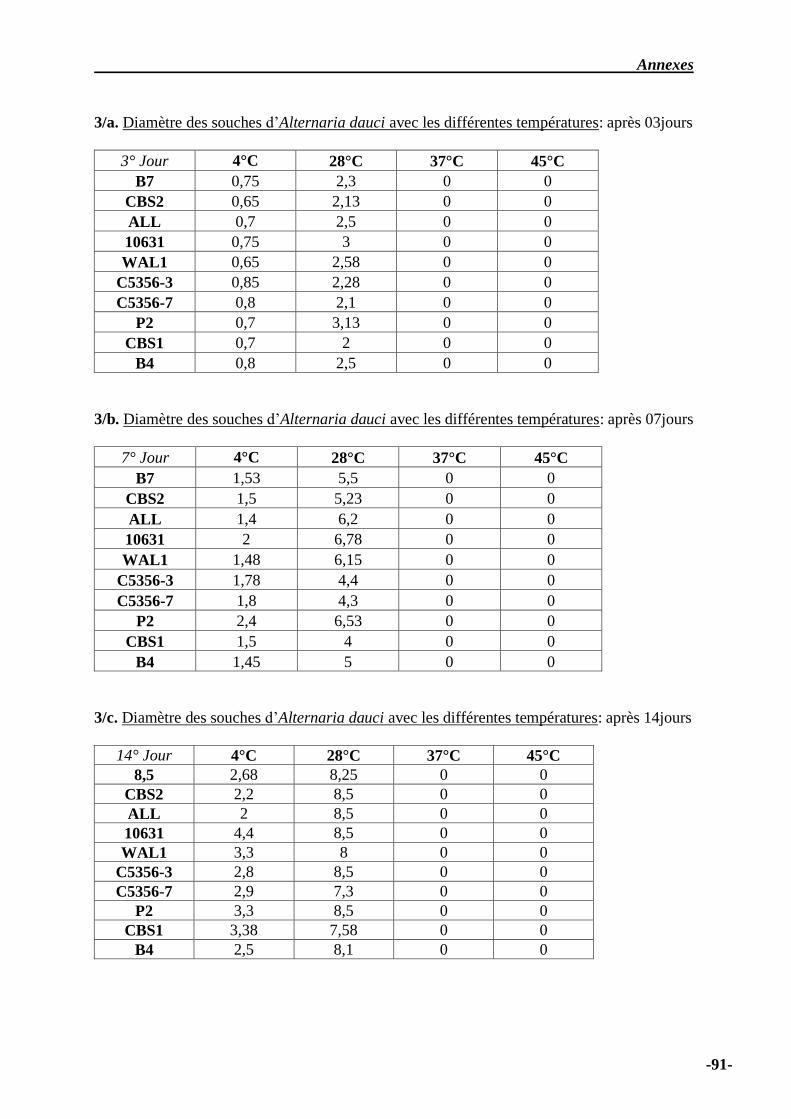
Annexes
3/a. Diamètre des souches d’Alternaria dauci avec les différentes températures: après 03jours
3° Jour 4°C 28°C 37°C 45°C
B7 0,75 2,3 0 0
CBS2 0,65 2,13 0 0
ALL 0,7 2,5 0 0
10631 0,75 3 0 0
WAL1 0,65 2,58 0 0
C5356-3 0,85 2,28 0 0
C5356-7 0,8 2,1 0 0
P2 0,7 3,13 0 0
CBS1 0,7 2 0 0
B4 0,8 2,5 0 0
3/b. Diamètre des souches d’Alternaria dauci avec les différentes températures: après 07jours
7° Jour 4°C 28°C 37°C 45°C
B7 1,53 5,5 0 0
CBS2 1,5 5,23 0 0
ALL 1,4 6,2 0 0
10631 2 6,78 0 0
WAL1 1,48 6,15 0 0
C5356-3 1,78 4,4 0 0
C5356-7 1,8 4,3 0 0
P2 2,4 6,53 0 0
CBS1 1,5 4 0 0
B4 1,45 5 0 0
3/c. Diamètre des souches d’Alternaria dauci avec les différentes températures: après 14jours
14° Jour 4°C 28°C 37°C 45°C
8,5 2,68 8,25 0 0
CBS2 2,2 8,5 0 0
ALL 2 8,5 0 0
10631 4,4 8,5 0 0
WAL1 3,3 8 0 0
C5356-3 2,8 8,5 0 0
C5356-7 2,9 7,3 0 0
P2 3,3 8,5 0 0
CBS1 3,38 7,58 0 0
B4 2,5 8,1 0 0
-91-

Annexes
4/a. Diamètre des souches d’Alternaria dauci avec les différent pH: après 03jours
3° Jour pH04 pH06 pH08
B7 0 2,4 2,1
CBS2 0 2,08 2,08
ALL 0 2,7 1,9
10631 0 2,03 2,03
WAL1 0 3,03 1,08
C5356-3 0 2,83 1,1
C5356-7 0 2 1,3
P2 0 3,35 1,13
CBS1 0 2,88 2
B4 0 3 1,78
4/b. Diamètre des souches d’Alternaria dauci avec les différent pH: après 07jours
7° Jour pH04 pH06 pH08
B7 0 5,4 3,7
CBS2 0 5,65 4,08
ALL 0 5,4 3,9
10631 0 6,2 4,5
WAL1 0 6 2,55
C5356-3 0 6,08 2,08
C5356-7 0 5,5 3,1
P2 1,95 6,85 2,1
CBS1 0 5,95 4,38
B4 0 6,25 3,5
4/c. Diamètre des souches d’Alternaria dauci avec les différent pH: après 14jours
14° Jour pH04 pH06 pH08
B7 0 8 5,6
CBS2 0 8,5 7
ALL 0 7,3 6,1
10631 6 8,5 8
WAL1 2,45 8,5 4,75
C5356-3 1,8 8 4
C5356-7 0 8,3 5,8
P2 3,2 8,5 5
CBS1 0 8,5 7
B4 1,5 7,3 7,2
-92-

Annexes
5/a. Diamètre des souches d’Alternaria dauci avec les différentes lumières: après 03jours
3° Jour L continue O continue photopériode
B7 2,4 2,4 2,1
CBS2 2,63 2,73 2,25
ALL 2,8 2,2 2,5
10631 3,75 2,98 3,1
WAL1 2,73 2,28 2,58
C5356-3 2,9 2,5 2,48
C5356-7 2,4 1,9 2
P2 3,98 3,1 3,43
CBS1 3,5 2,5 2,5
B4 3,05 2,58 2,42
5/b. Diamètre des souches d’Alternaria dauci avec les différentes lumières: après 07jours
7° Jour L continue O continue photopériode
B7 5 5,3 5,25
CBS2 5,13 5,15 5,95
ALL 5,5 4,3 5,5
10631 8,25 7,03 7,33
WAL1 5,75 4,88 5,23
C5356-3 5,73 5,3 5,8
C5356-7 6,1 5,5 5,5
P2 8,15 7,05 7,1
CBS1 7 5,7 5
B4 6 5,5 6,2
5/c. Diamètre des souches d’Alternaria dauci avec les différentes lumières: après 14jours
14° Jour L continue O continue photopériode
B7 7,6 7,5 8
CBS2 8 7,78 8,5
ALL 8,1 7,6 7,5
10631 8,5 8,5 8,5
WAL1 8,5 8 8,08
C5356-3 8,25 7,5 8,5
C5356-7 8,5 8,5 8,3
P2 8,5 8,5 8,5
CBS1 8,5 7,68 8,5
B4 8,25 7,88 8,5
-93-
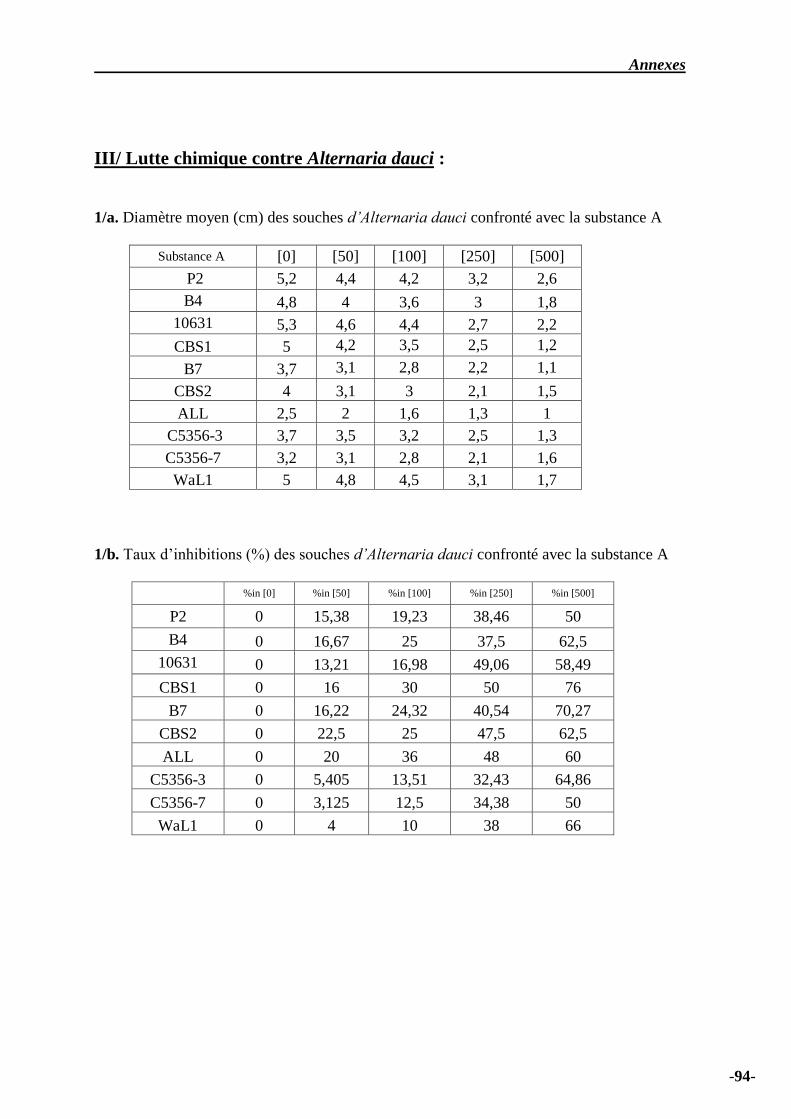
Annexes
III/ Lutte chimique contre Alternaria dauci :
1/a. Diamètre moyen (cm) des souches d’Alternaria dauci confronté avec la substance A
Substance A [0] [50] [100] [250] [500]
P2 5,2 4,4 4,2 3,2 2,6
B4 4,8 4 3,6 3 1,8
10631 5,3 4,6 4,4 2,7 2,2
CBS1 5 4,2 3,5 2,5 1,2
B7 3,7 3,1 2,8 2,2 1,1
CBS2 4 3,1 3 2,1 1,5
ALL 2,5 2 1,6 1,3 1
C5356-3 3,7 3,5 3,2 2,5 1,3
C5356-7 3,2 3,1 2,8 2,1 1,6
WaL1 5 4,8 4,5 3,1 1,7
1/b. Taux d’inhibitions (%) des souches d’Alternaria dauci confronté avec la substance A
%in [0] %in [50] %in [100] %in [250] %in [500]
P2 0 15,38 19,23 38,46 50
B4 0 16,67 25 37,5 62,5
10631 0 13,21 16,98 49,06 58,49
CBS1 0 16 30 50 76
B7 0 16,22 24,32 40,54 70,27
CBS2 0 22,5 25 47,5 62,5
ALL 0 20 36 48 60
C5356-3 0 5,405 13,51 32,43 64,86
C5356-7 0 3,125 12,5 34,38 50
WaL1 0 4 10 38 66
-94-
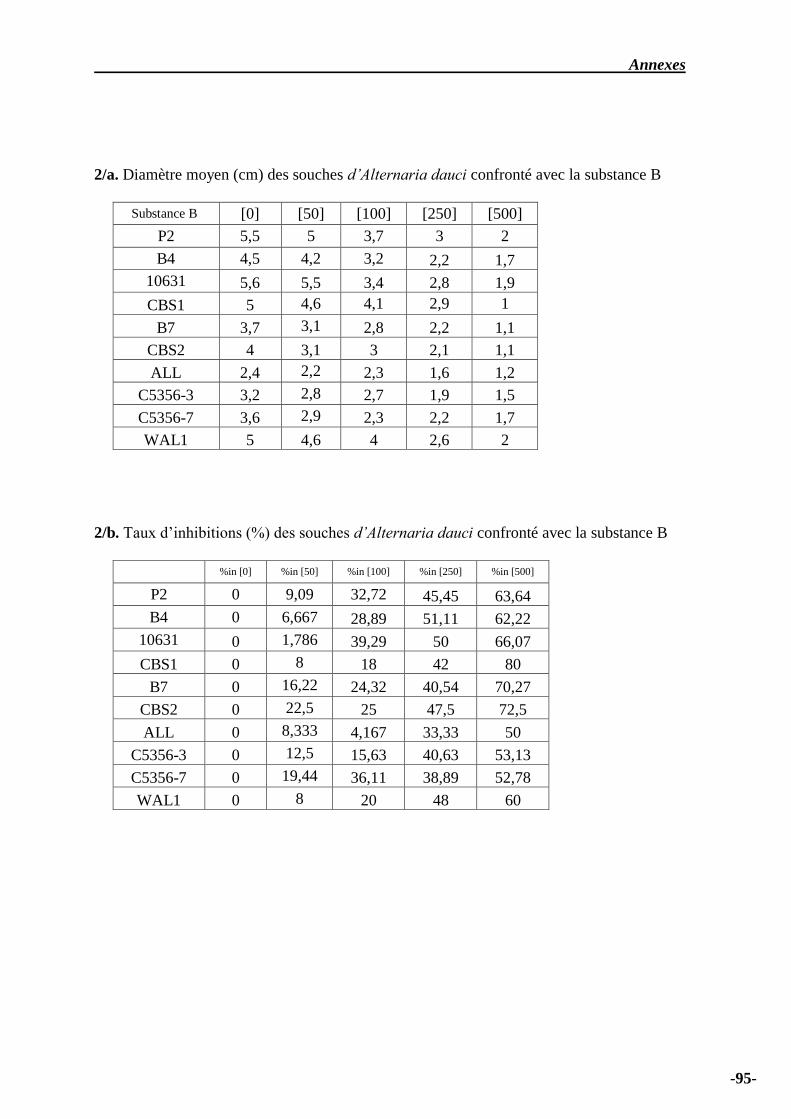
Annexes
2/a. Diamètre moyen (cm) des souches d’Alternaria dauci confronté avec la substance B
Substance B [0] [50] [100] [250] [500]
P2 5,5 5 3,7 3 2
B4 4,5 4,2 3,2 2,2 1,7
10631 5,6 5,5 3,4 2,8 1,9
CBS1 5 4,6 4,1 2,9 1
B7 3,7 3,1 2,8 2,2 1,1
CBS2 4 3,1 3 2,1 1,1
ALL 2,4 2,2 2,3 1,6 1,2
C5356-3 3,2 2,8 2,7 1,9 1,5
C5356-7 3,6 2,9 2,3 2,2 1,7
WAL1 5 4,6 4 2,6 2
2/b. Taux d’inhibitions (%) des souches d’Alternaria dauci confronté avec la substance B
%in [0] %in [50] %in [100] %in [250] %in [500]
P2 0 9,09 32,72 45,45 63,64
B4 0 6,667 28,89 51,11 62,22
10631 0 1,786 39,29 50 66,07
CBS1 0 8 18 42 80
B7 0 16,22 24,32 40,54 70,27
CBS2 0 22,5 25 47,5 72,5
ALL 0 8,333 4,167 33,33 50
C5356-3 0 12,5 15,63 40,63 53,13
C5356-7 0 19,44 36,11 38,89 52,78
WAL1 0 8 20 48 60
-95-


















