

Dynamique et Réplication de l’ADN. - jn0.free.fr20Physiopath/Ron%E9os%20Canc%E9... · Lien...
68
Dynamique et Réplication de l’ADN. D.Burnouf, UPR9002, IBMC, 03 88 41 70 02 [email protected] UPR 9002
Transcript of Dynamique et Réplication de l’ADN. - jn0.free.fr20Physiopath/Ron%E9os%20Canc%E9... · Lien...

Dynamique et Réplication de l’ADN.
D.Burnouf, UPR9002, IBMC, 03 88 41 70 02 [email protected]
UPR
9002

Plan
• I - Structure et dynamique de l’ADN et de la chromatine.
• II- ADN polymérases: classification, structure et mécanisme.
• III- Acteurs de la réplication. • IV- Réplication d’un ADN endommagé:
modèles.

Quelques chiffres.
• E.coli: 4.7 10 pb. – #4000 gènes
• S.cerevisiae:13.4 10 pb – 14 chromosomes – # 6-7000 gènes – 70% du génome est codant
• Homo sapiens:3.3 10 pb – 23 chromosomes – ~21 000 gènes – 2% du génome est codant.
6
6
9
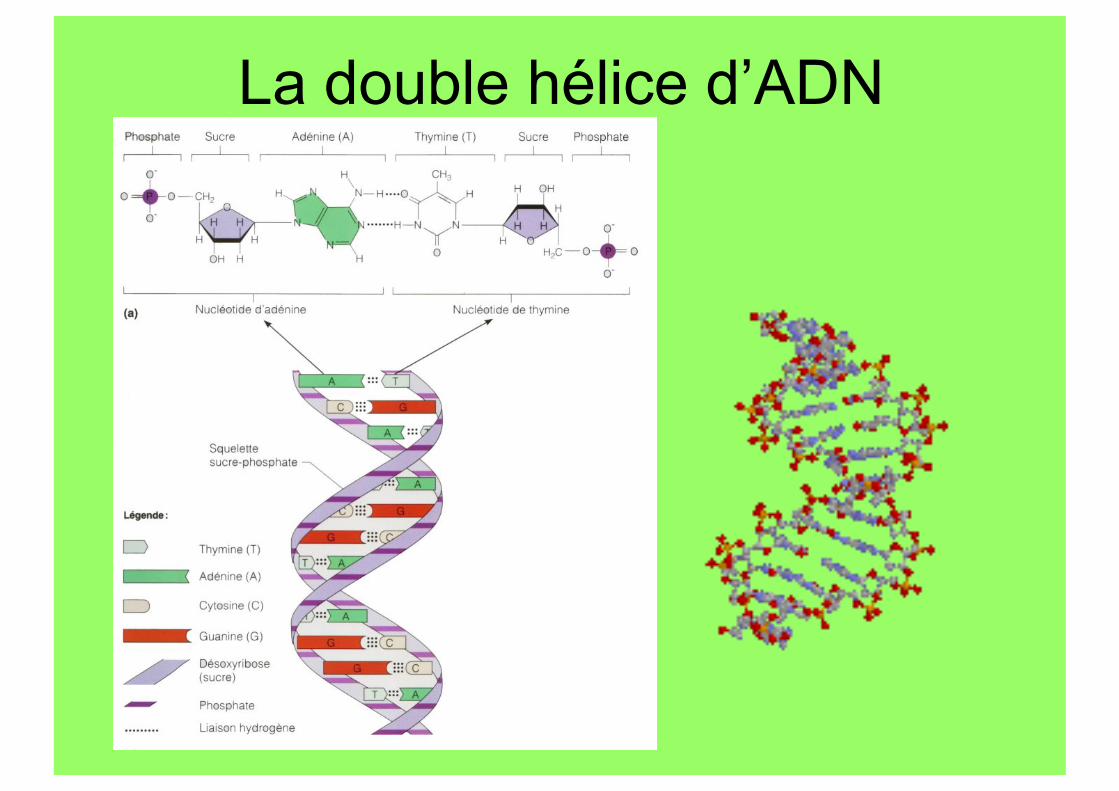
La double hélice d’ADN
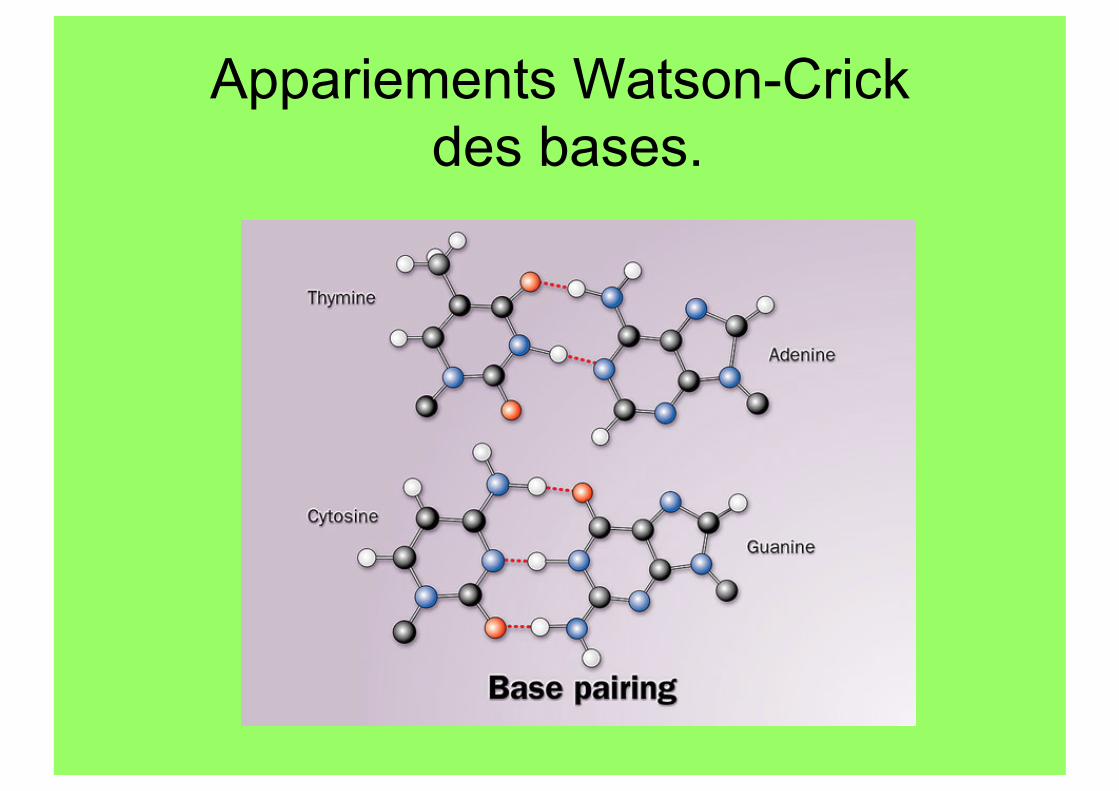
Appariements Watson-Crick des bases.
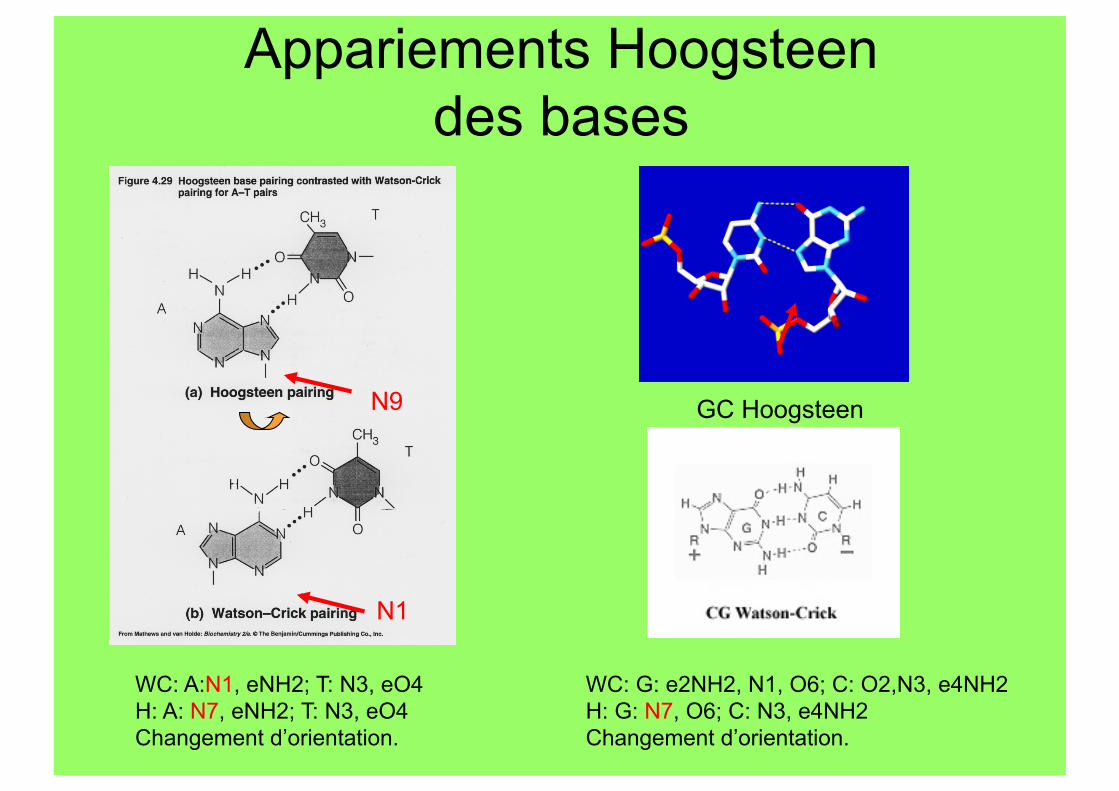
Appariements Hoogsteen des bases
GC Hoogsteen N9
N1
WC: A:N1, eNH2; T: N3, eO4 H: A: N7, eNH2; T: N3, eO4 Changement d’orientation.
WC: G: e2NH2, N1, O6; C: O2,N3, e4NH2 H: G: N7, O6; C: N3, e4NH2 Changement d’orientation.
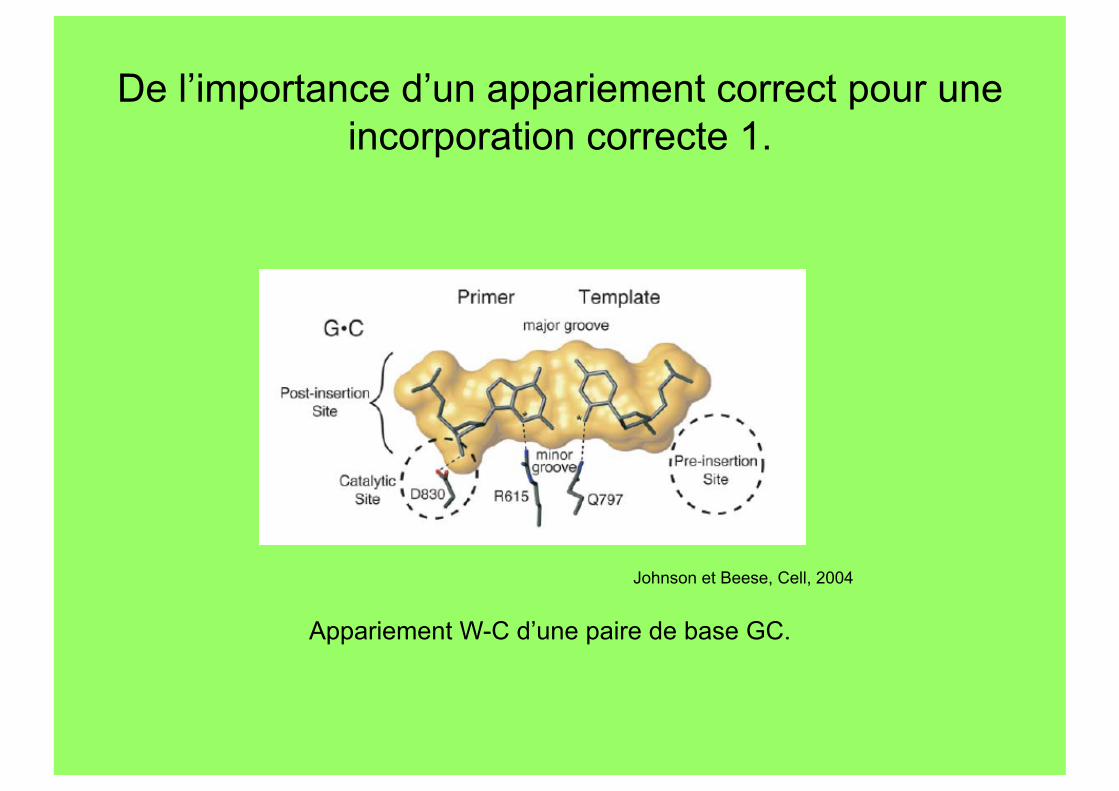
De l’importance d’un appariement correct pour une incorporation correcte 1.
Johnson et Beese, Cell, 2004
Appariement W-C d’une paire de base GC.

Perturbations géométriques liées à un mésappariement.
De l’importance d’un appariement correct pour une incorporation correcte 2.

Topologie de l’ADN.
Forme B

Autres conformations de l’ADN Formes A et Z.
Pb/tour: 11 12 10 Diamètre: 23A 18A 20A

II. Dynamique de la chromatine.
Permanente et nécessaire.
Assurer les fonctions fondamentales: réplication/réparation transcription condensation et ségrégation cycle cellulaire division cellulaire
régulation
Différents mécanismes: Modification des histones remodelage de la chromatine incorporation d’histones variants modulation topologie de l’ADN
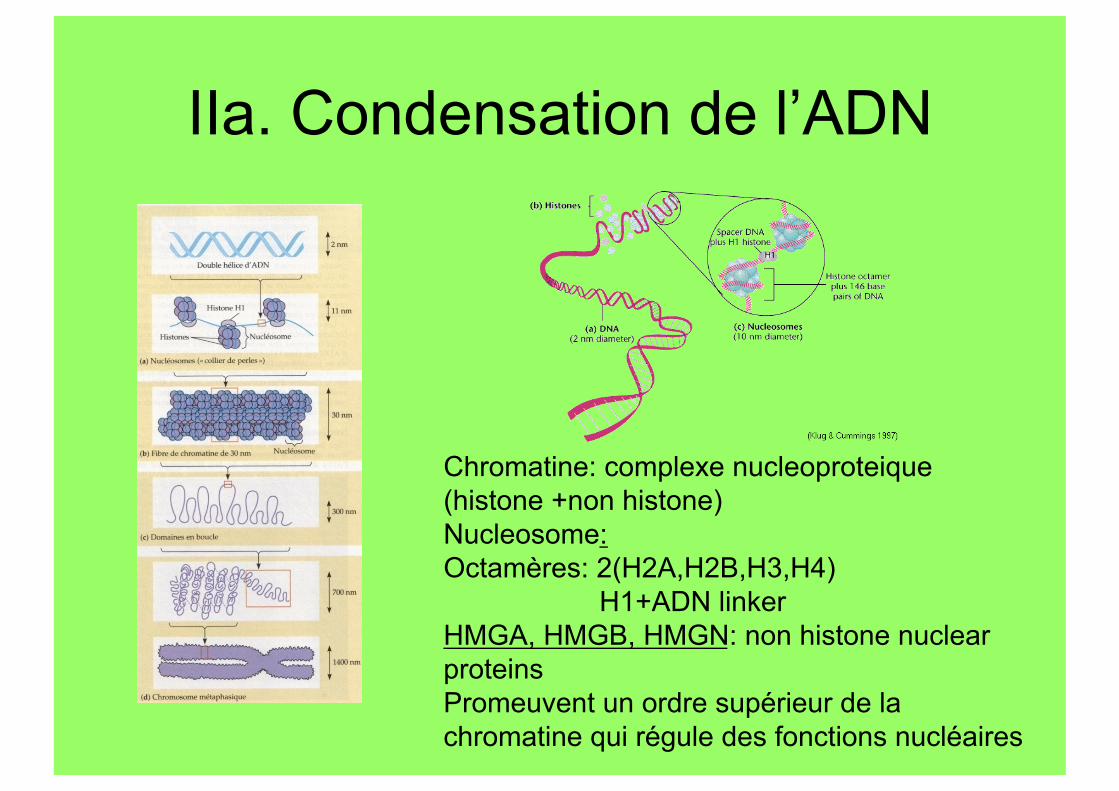
IIa. Condensation de l’ADN
Chromatine: complexe nucleoproteique (histone +non histone) Nucleosome: Octamères: 2(H2A,H2B,H3,H4)
H1+ADN linker HMGA, HMGB, HMGN: non histone nuclear proteins Promeuvent un ordre supérieur de la chromatine qui régule des fonctions nucléaires
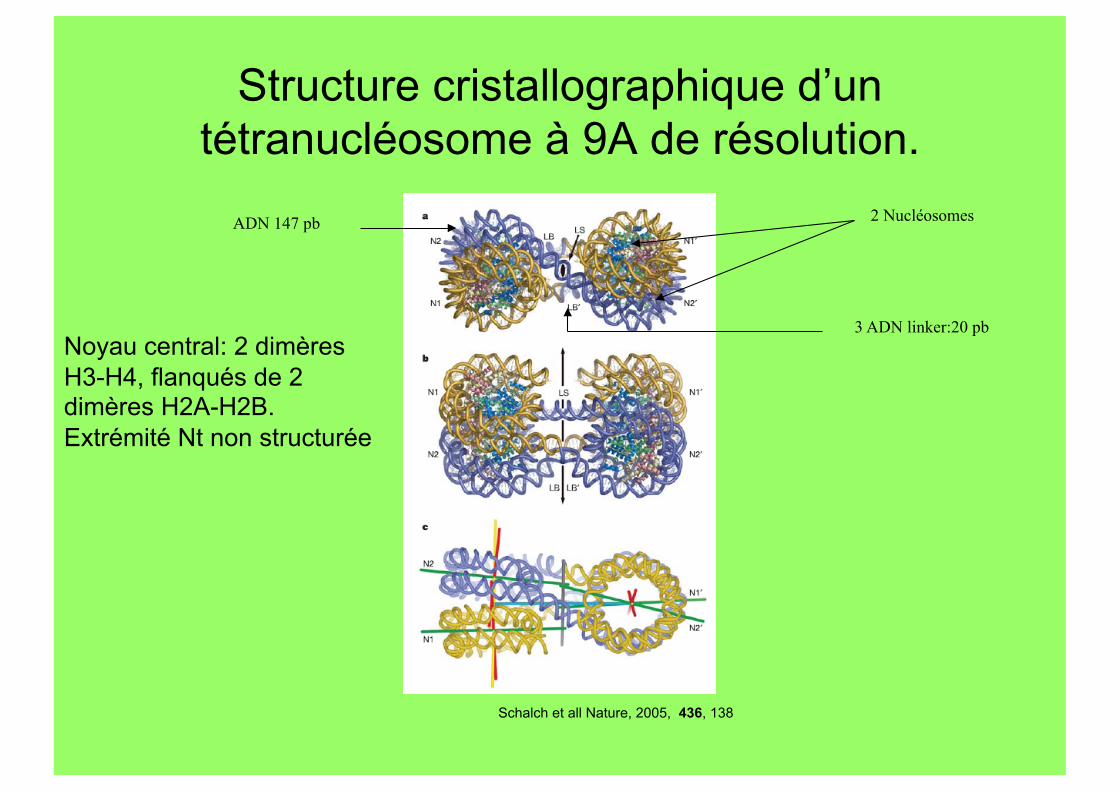
Structure cristallographique d’un tétranucléosome à 9A de résolution.
Schalch et all Nature, 2005, 436, 138
2 Nucléosomes
3 ADN linker:20 pb
ADN 147 pb
Noyau central: 2 dimères H3-H4, flanqués de 2 dimères H2A-H2B. Extrémité Nt non structurée

IIb. Métabolisme des histones. • Hautement régulé (excès toxique) • Couplé à la réplication
* contrôle de l’expression des gènes d’histones. * mécanisme de dégradation contrôlé par check point (Rad 53). • Contrôle des pools par formation de complexes de haut PM • Contrôle de l’import nucléaire par des karyophérines
=> régulation de la structure de la chromatine et de l’expression des gènes.
Les histones chaperones jouent un rôle central dans la dynamique de la chromatine.

Dynamique des nucléosomes lors de la réplication.
La chaperone Asf1 est un accepteur-donneur d’histones H3-H4 qui régule la progression de la fourche de réplication. Une autre chaperone FACT, s’associe à H2A-H2B.
Nombreux complexes impliqués: chaperones, karyopherins (import nucléaire), acetyl transférases, Chromatin Assembly Factor, Nucleosome Assembly Factor.
Groth, Science, 2007

IIC. Histones variants.
• Isoformes des histones conventionnels (sauf H4) mais • Timing d’expression différent (non couplé à la réplication) • Expression réduite • Associée à des séquences d’ADN spécifiques • Parfois expression tissus spécifique
• Incorporés dans les nucléosomes => • altère la structure: new structures et fonctionnalités • Régulation (transcription, repair,….) • Modulation de la structure de la chromatine(>0 ou <0) • Mécanismes de dépositions différents selon les variants et mal connus

Variants de H2A:
– Famille la plus importante: 4 sous types
• H2A.Z: rôles contradictoires: – hétérochromatine: favorise l’assemblage et l’organisation en structures compactes – déstabilise l’euchromatine, rôle dans la régulation de la transcription
• H2A.X: très conservé, 25% de H2A. Distribution sur tout le génome. kinasé (SQ,Ct) par ATM si DNA dommages, interagit avec translocases et Xtine remodeleurs près des sites de lésions.
• macroH2A: rare, région NHR, 2 formes, assemblage de l’hétérochromatine, bloque l’action des remodeleurs et des acétylases d’histones.
• H2A.Bbd: altère et déstabilise la structure des nucléosomes et bloque l’action des remodeleurs: favorise la transcription. X actif spécifique=> rôle épigénétique

Mode d’action supposé de mH2A sur la transcription.

• Variants de H3: 2 formes – CENH3: centromère, gène essentiel,organisation du kinétochore – H3.3: activation de la transcription, spécifique de HIRA
• Variants de H2B – Peu connus – Rôle lors de la gamétogenèse(H2BFWT)
• Histone variants et cancer. – H2A.X-/-: défaut de réparation, immune déficients,radiosensible, Xsome
instables (translocations, amplifications) – +P53 => leucémie, sarcomes, lymphomes, …
• => H2A.X= suppresseur de tumeur
– Lien entre CENH3 et aneuploïdie: • surexpression de CENH3 liée à genome instabilité (Xsomes multicentriques, …)

IId. Torsion de l’ADN.
relaxée
linéaire
superenroulée
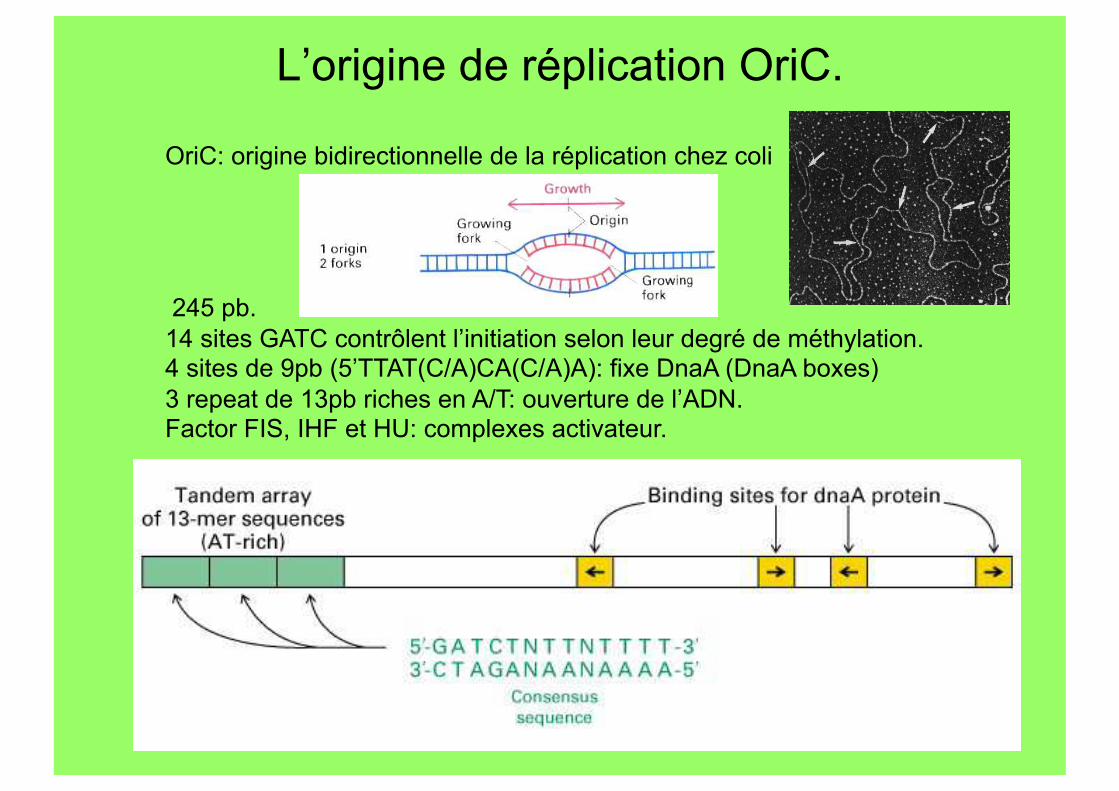
L’origine de réplication OriC.
OriC: origine bidirectionnelle de la réplication chez coli
245 pb. 14 sites GATC contrôlent l’initiation selon leur degré de méthylation. 4 sites de 9pb (5’TTAT(C/A)CA(C/A)A): fixe DnaA (DnaA boxes) 3 repeat de 13pb riches en A/T: ouverture de l’ADN. Factor FIS, IHF et HU: complexes activateur.
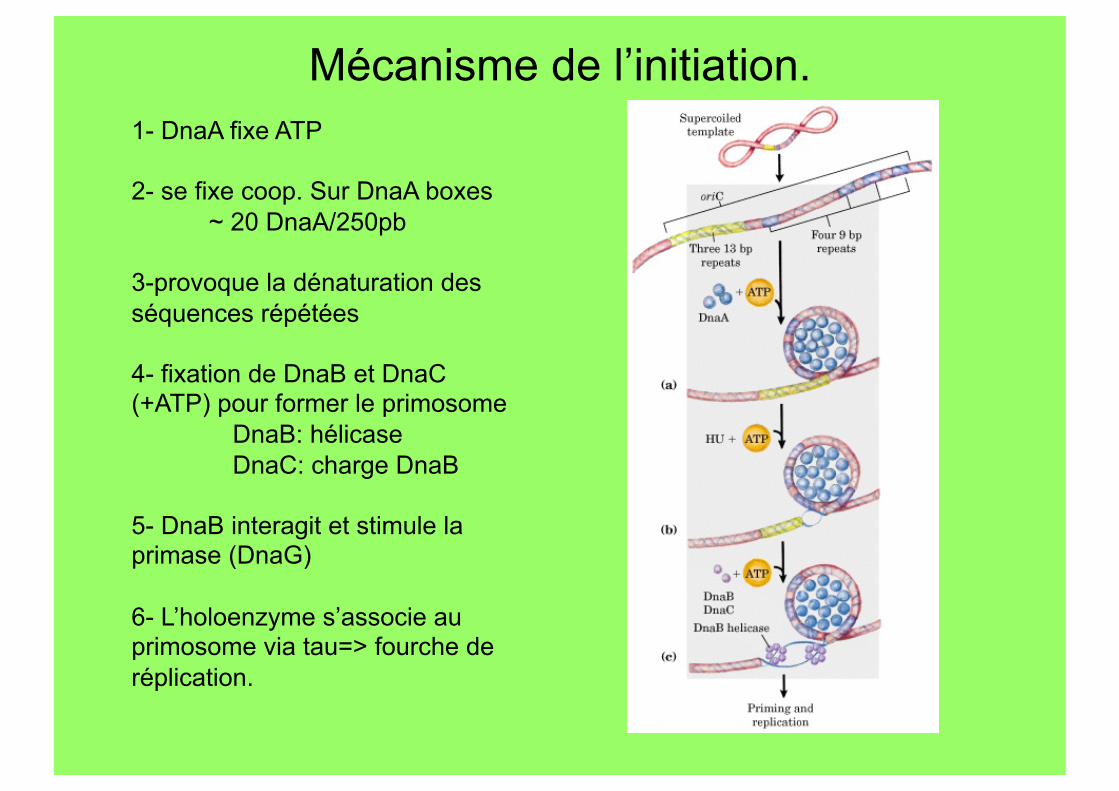
Mécanisme de l’initiation. 1- DnaA fixe ATP
2- se fixe coop. Sur DnaA boxes ~ 20 DnaA/250pb
3-provoque la dénaturation des séquences répétées
4- fixation de DnaB et DnaC (+ATP) pour former le primosome
DnaB: hélicase DnaC: charge DnaB
5- DnaB interagit et stimule la primase (DnaG)
6- L’holoenzyme s’associe au primosome via tau=> fourche de réplication.
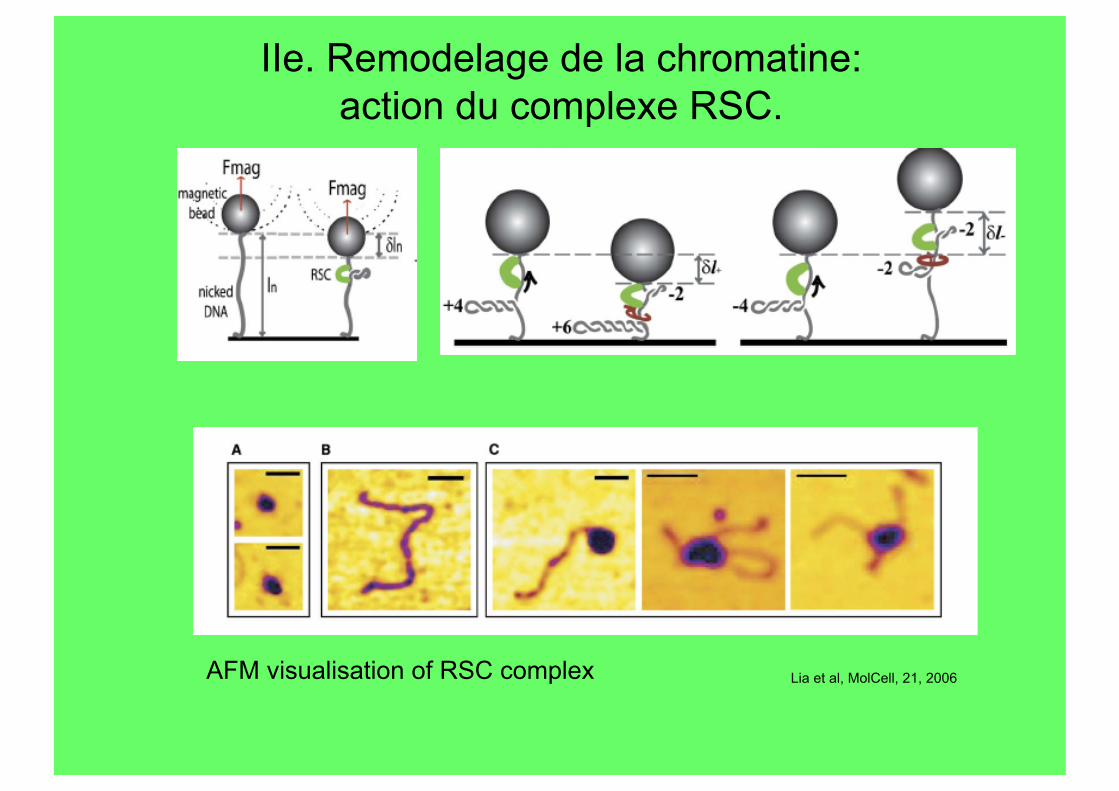
IIe. Remodelage de la chromatine: action du complexe RSC.
Lia et al, MolCell, 21, 2006 AFM visualisation of RSC complex
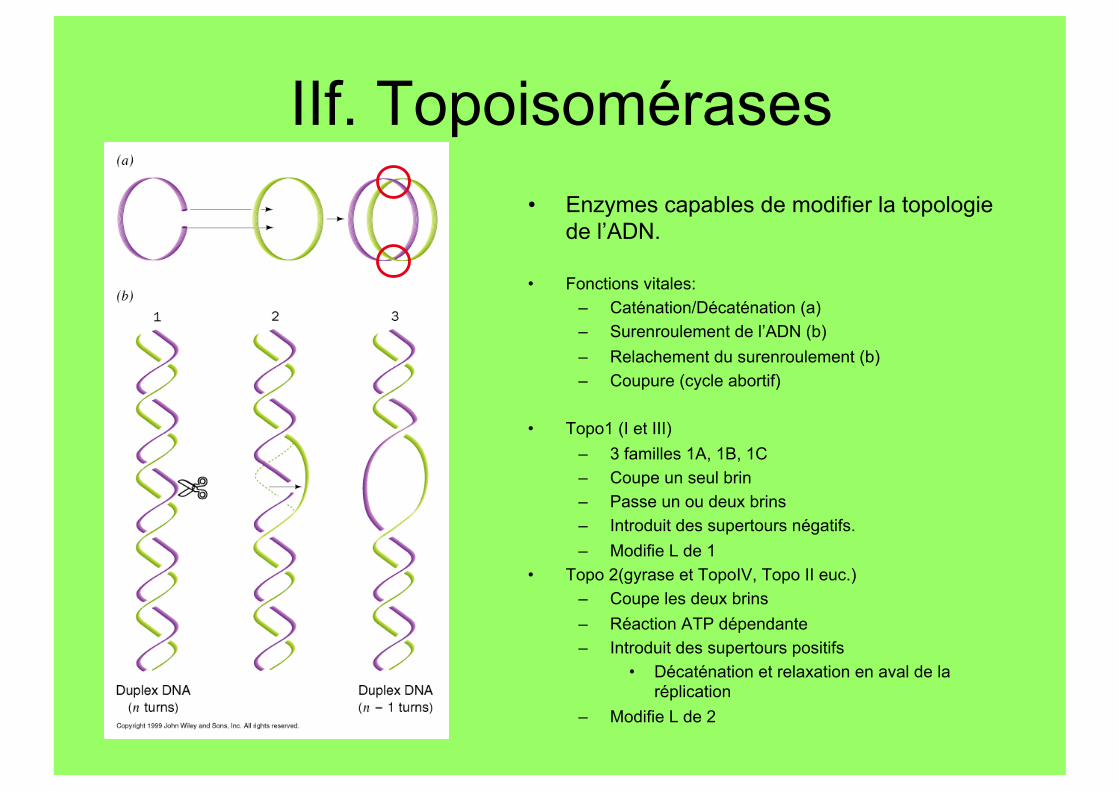
IIf. Topoisomérases • Enzymes capables de modifier la topologie
de l’ADN.
• Fonctions vitales: – Caténation/Décaténation (a) – Surenroulement de l’ADN (b) – Relachement du surenroulement (b) – Coupure (cycle abortif)
• Topo1 (I et III) – 3 familles 1A, 1B, 1C – Coupe un seul brin – Passe un ou deux brins – Introduit des supertours négatifs. – Modifie L de 1
• Topo 2(gyrase et TopoIV, Topo II euc.) – Coupe les deux brins – Réaction ATP dépendante – Introduit des supertours positifs
• Décaténation et relaxation en aval de la réplication
– Modifie L de 2
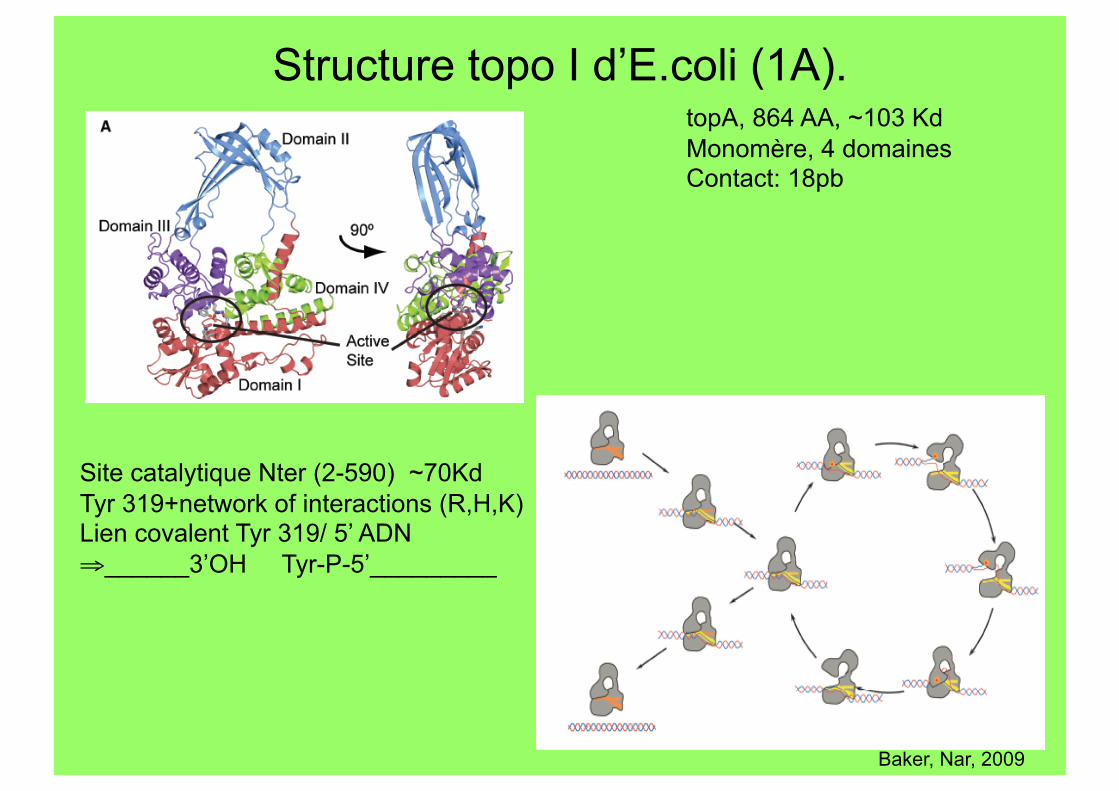
Structure topo I d’E.coli (1A).
Site catalytique Nter (2-590) ~70Kd Tyr 319+network of interactions (R,H,K) Lien covalent Tyr 319/ 5’ ADN ⇒ ______3’OH Tyr-P-5’_________
topA, 864 AA, ~103 Kd Monomère, 4 domaines Contact: 18pb
Baker, Nar, 2009
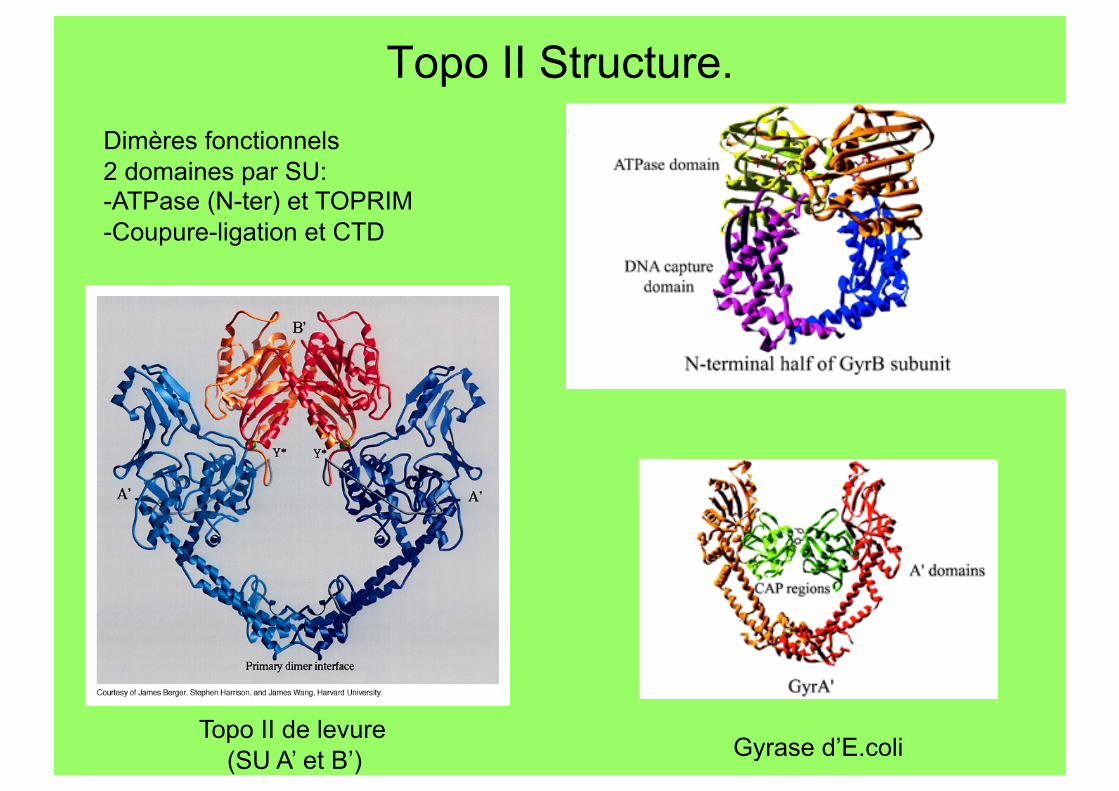
Topo II Structure.
Topo II de levure (SU A’ et B’) Gyrase d’E.coli
Dimères fonctionnels 2 domaines par SU: - ATPase (N-ter) et TOPRIM - Coupure-ligation et CTD
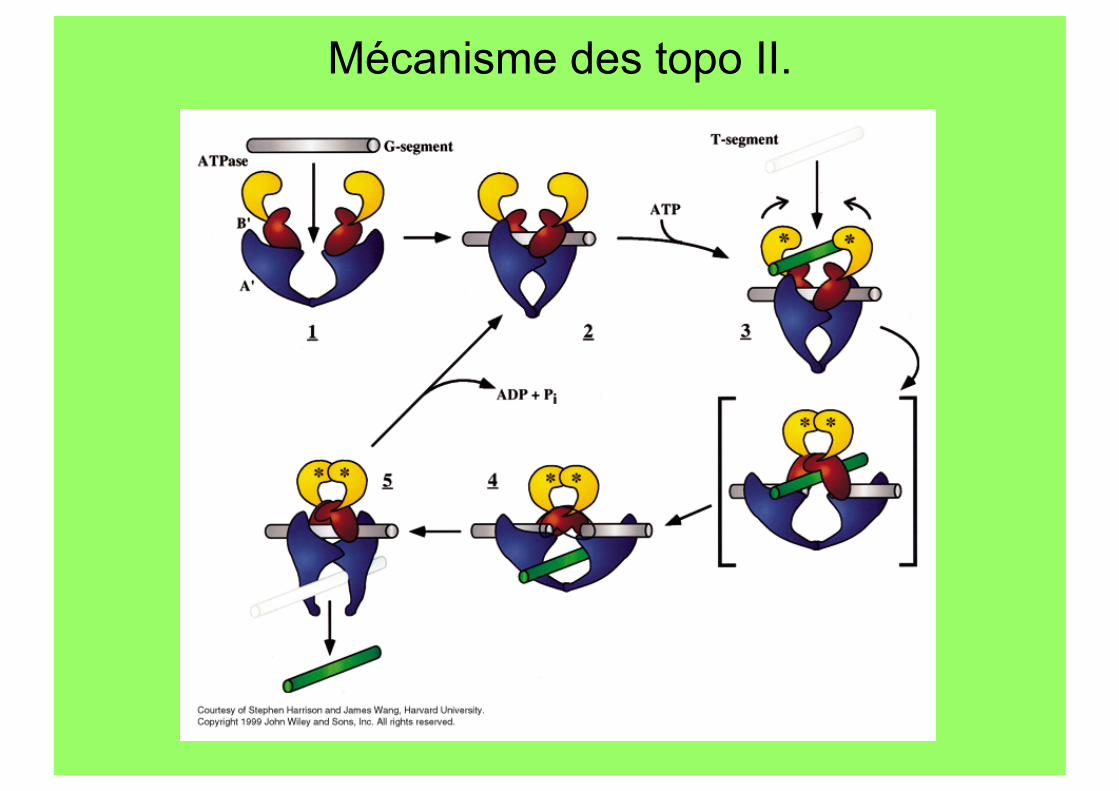
Mécanisme des topo II.

III. ADN Polymérases.
Classification. Structure.
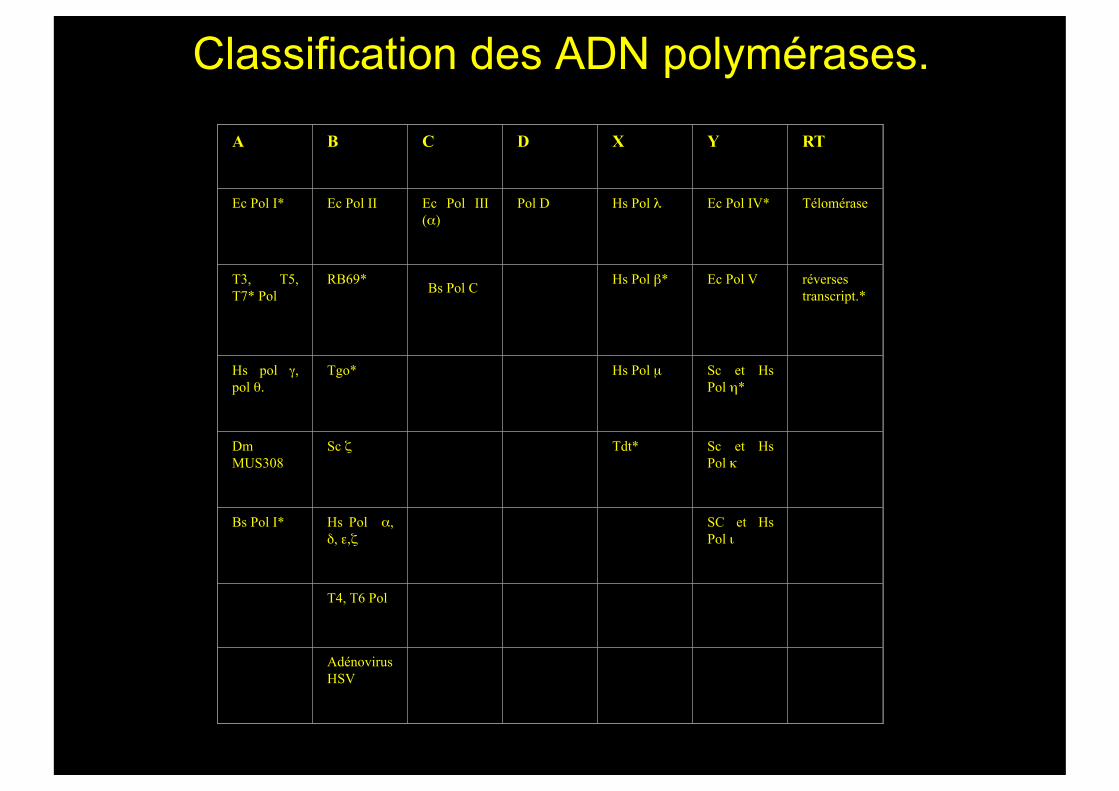
Classification des ADN polymérases.
A B C D X Y RT
Ec Pol I* Ec Pol II Ec Pol III (α)
Pol D Hs Pol λ Ec Pol IV* Télomérase
T3, T5, T7* Pol
RB69* Hs Pol β* Ec Pol V réverses transcript.*
Hs pol γ, pol θ.
Tgo* Hs Pol µ Sc et Hs Pol η*
Dm MUS308
Sc ζ Tdt* Sc et Hs Pol κ
Bs Pol I* Hs Pol α, δ, ε,ζ
SC et Hs Pol ι
T4, T6 Pol
Adénovirus HSV
Bs Pol C
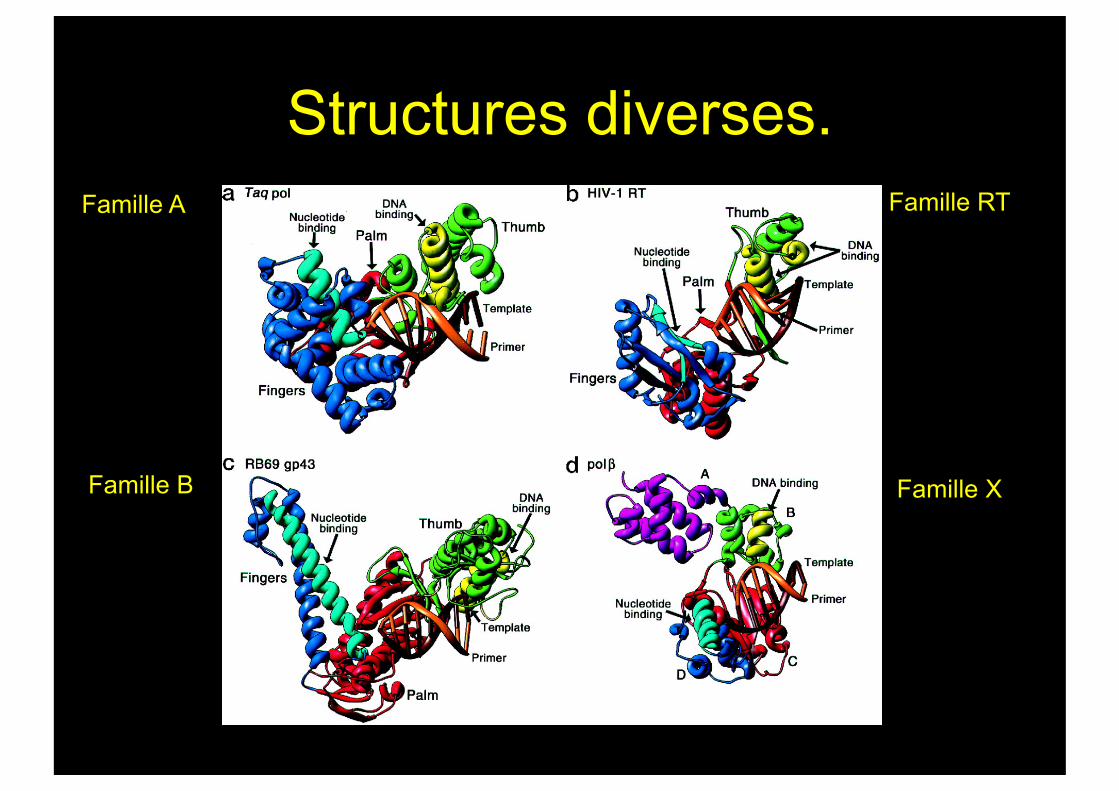
Structures diverses. Famille A
Famille B
Famille RT
Famille X

DNA pol I de B.stéarothermophilus (BF). Famille A

Structure de la SU α de Pol III. Famille C.
Lamers, Cell, 2006.
2.3A
Domaine supplémentaire
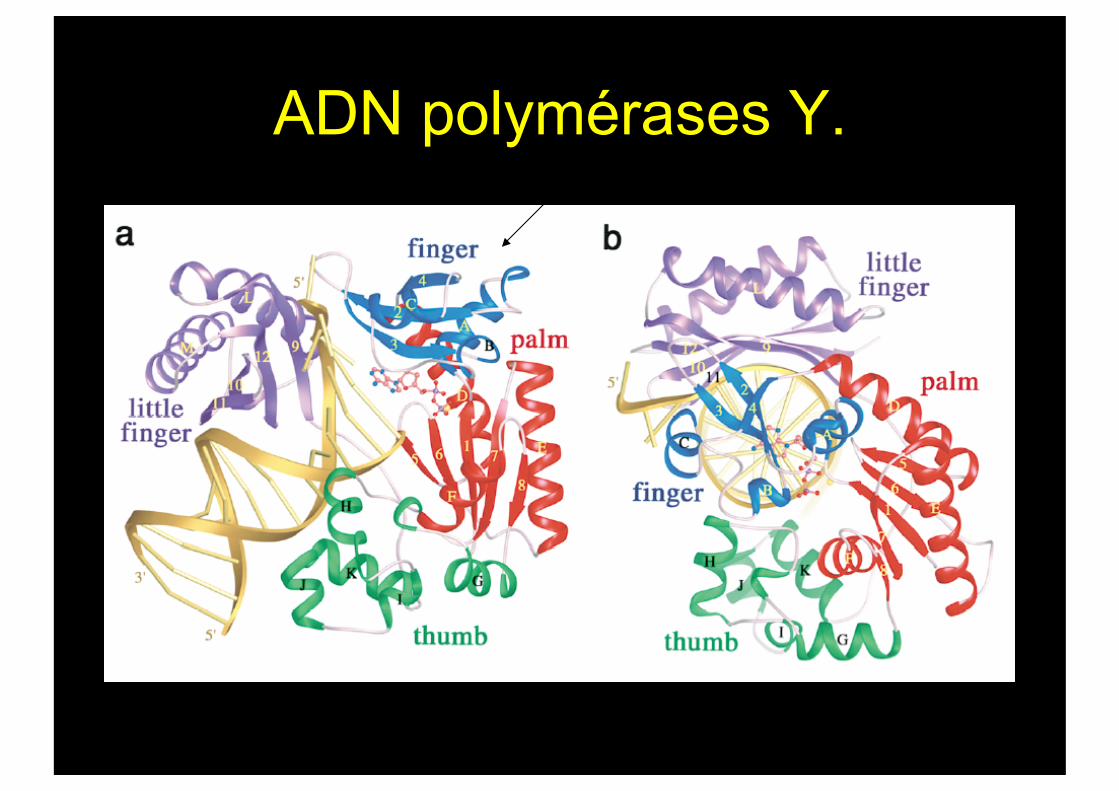
ADN polymérases Y.
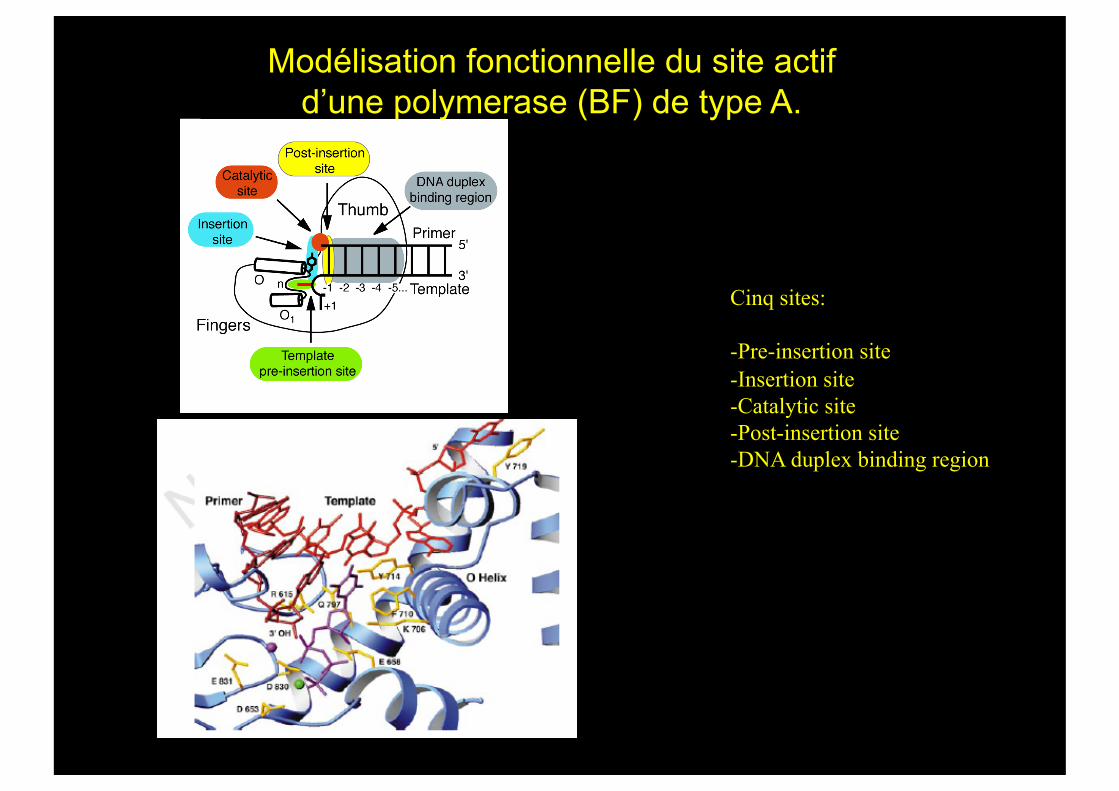
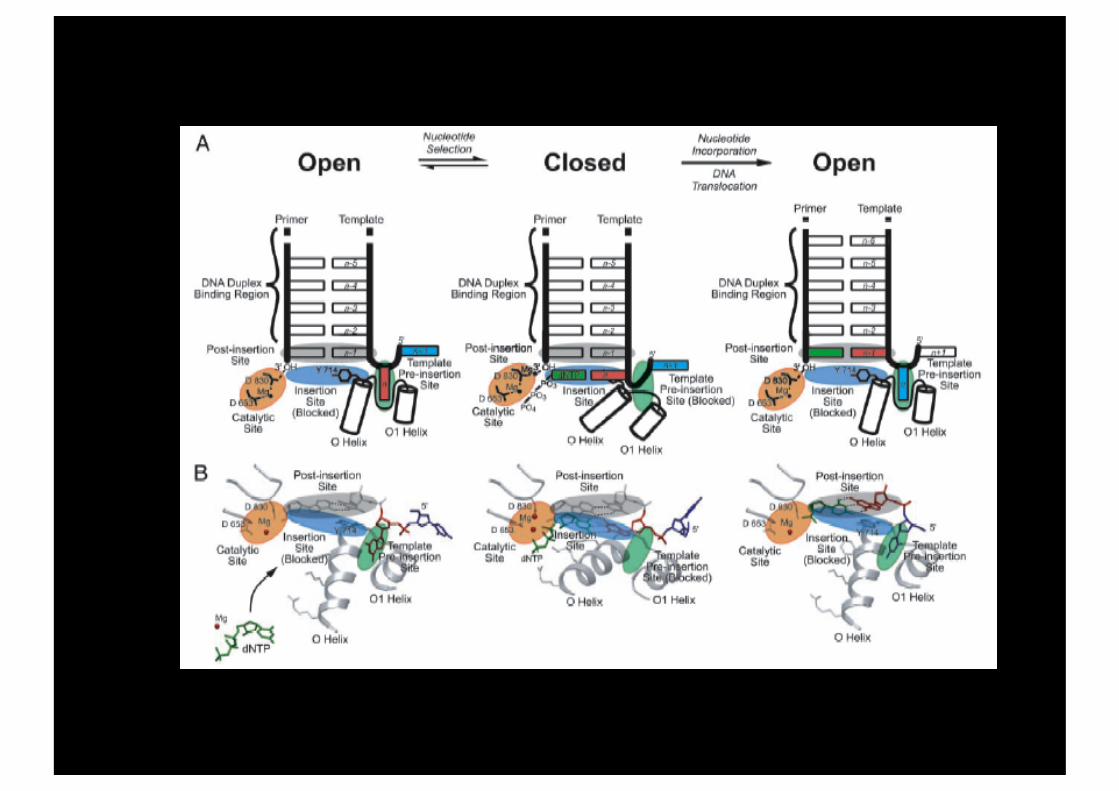
Modélisation fonctionnelle du site actif d’une polymerase (BF) de type A.
Cinq sites:
- Pre-insertion site - Insertion site - Catalytic site - Post-insertion site - DNA duplex binding region
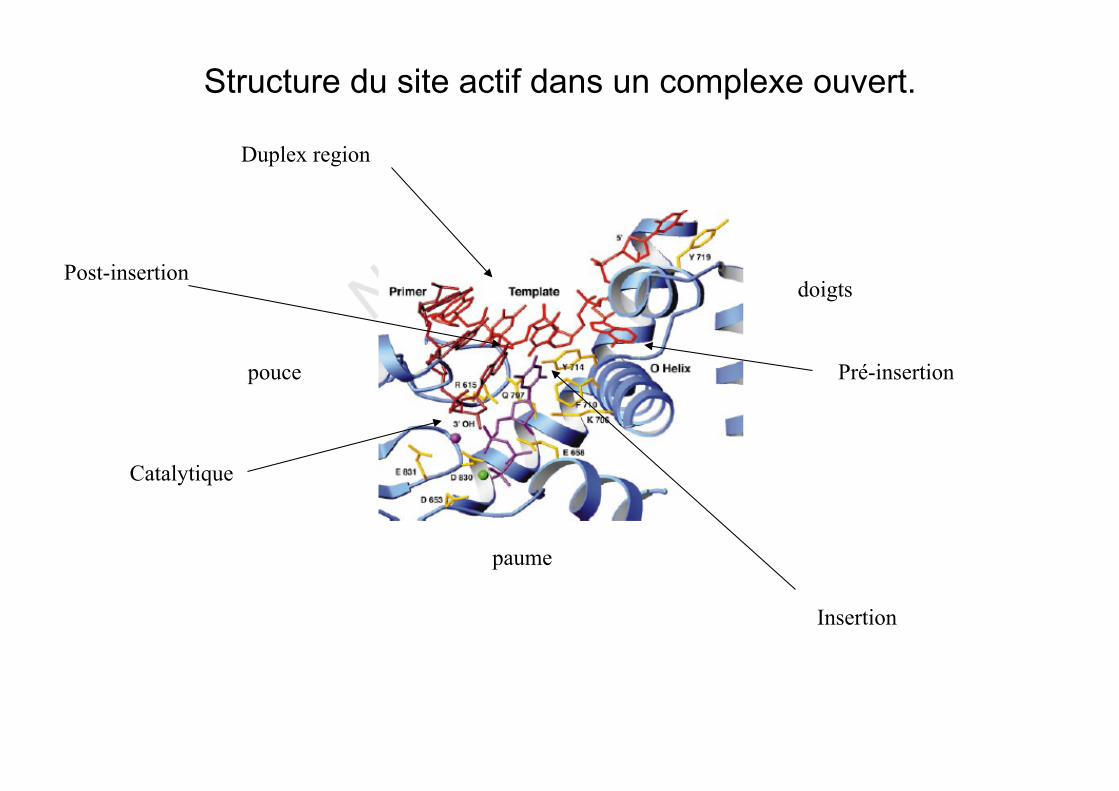
Structure du site actif dans un complexe ouvert.
paume
pouce
doigts
Pré-insertion
Insertion
Catalytique
Post-insertion
Duplex region
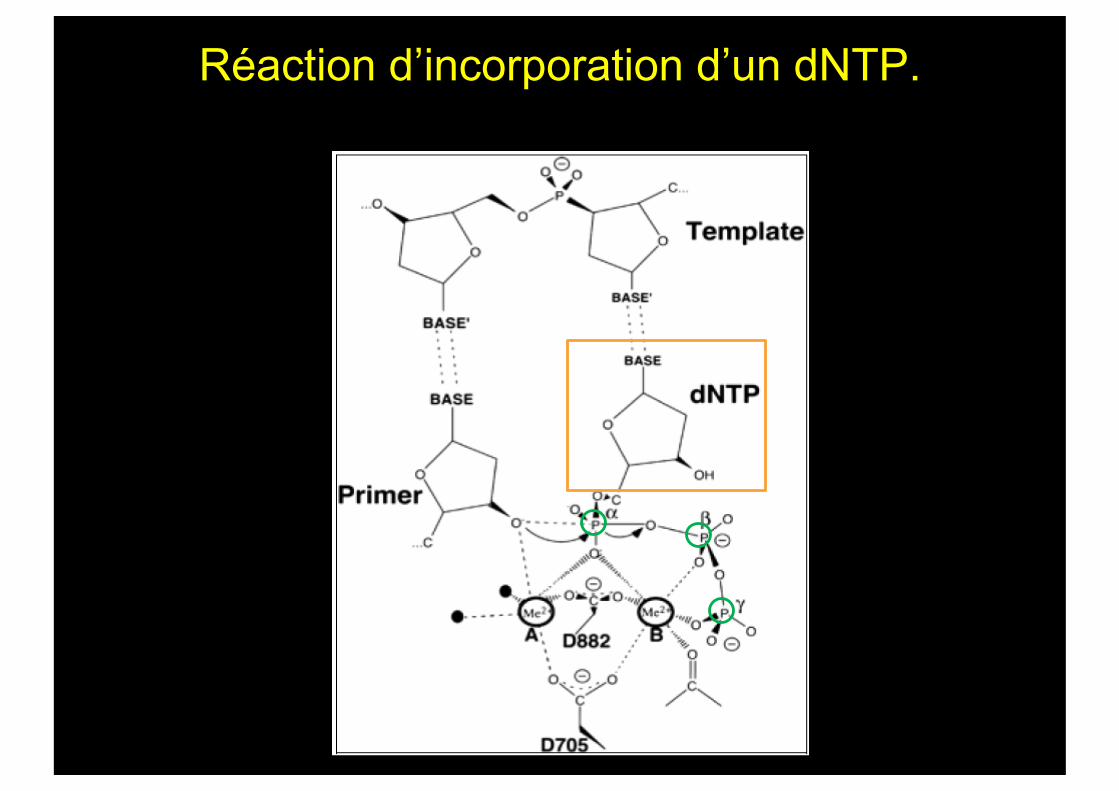
Réaction d’incorporation d’un dNTP.
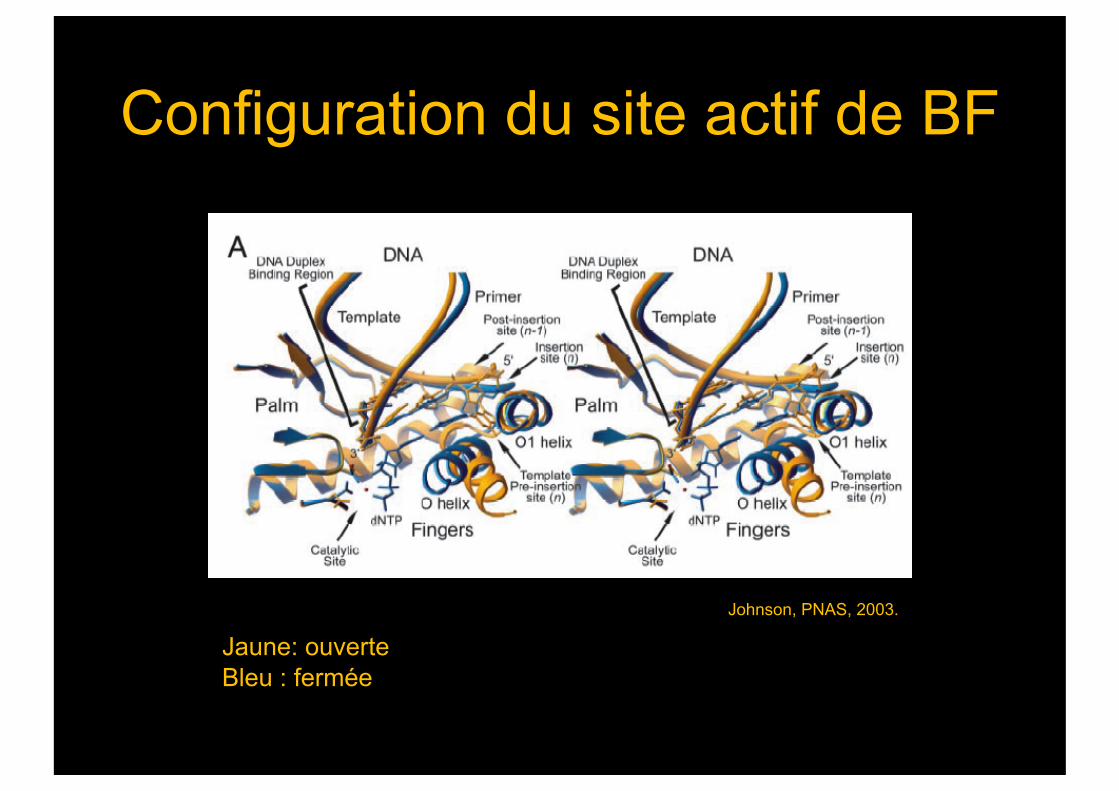
Configuration du site actif de BF
Jaune: ouverte Bleu : fermée
Johnson, PNAS, 2003.

III. ADN Polymérases.
Composants du complexe réplicatif.
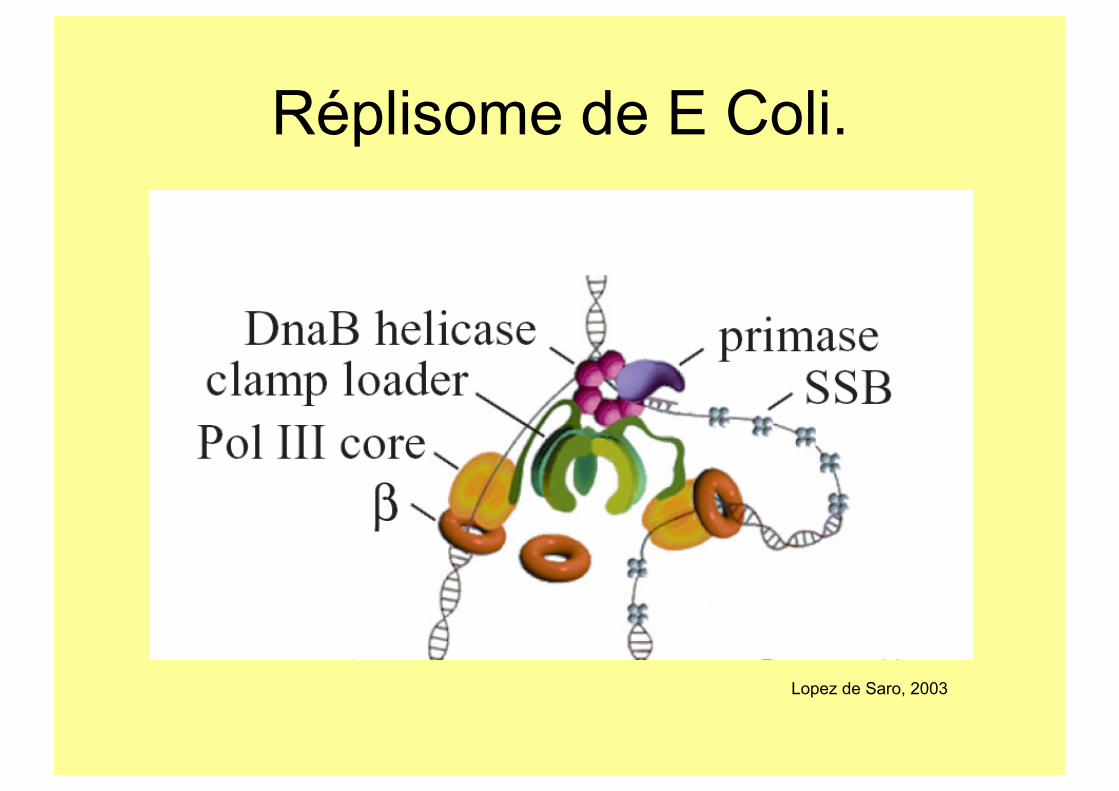
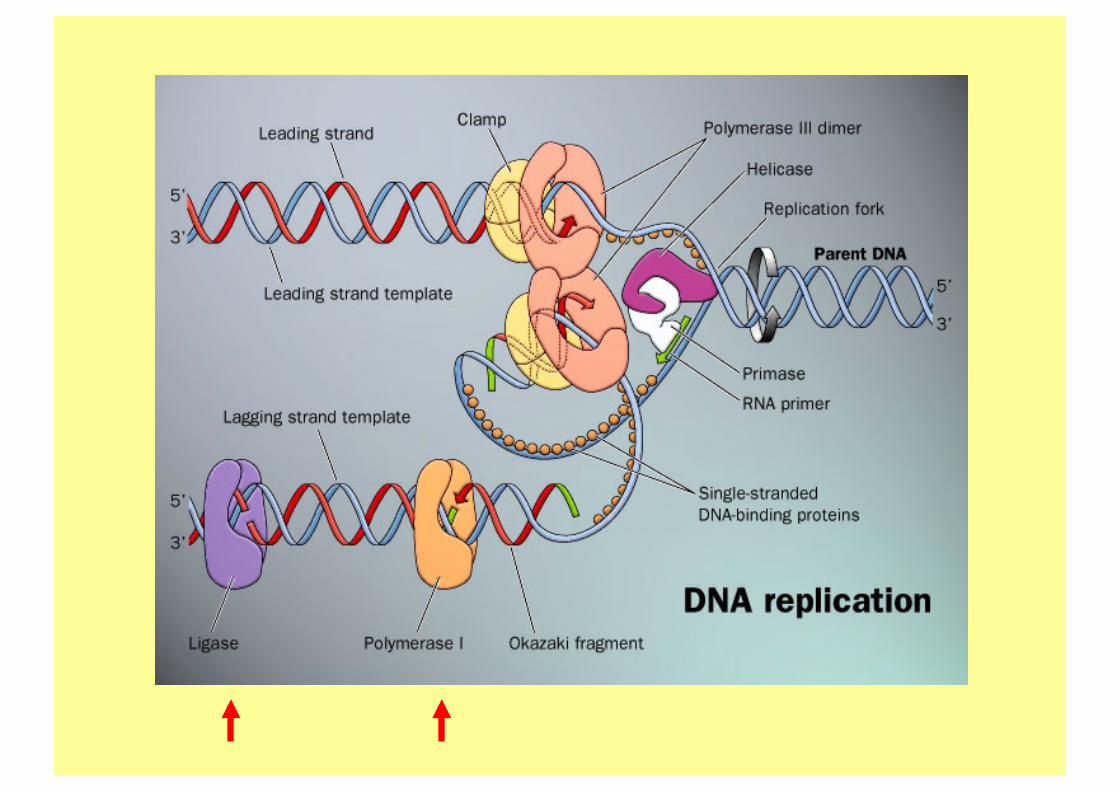
Réplisome de E Coli.
Lopez de Saro, 2003

Hélicase DnaB 52,3 Kd
Homohexamère: 300Kd
Lie 6 ATPs ADP:ouvre la DH
20 mol/cell
Se déplace 5’ 3’ (fourche)
Interagit avec τ
1000 N/s
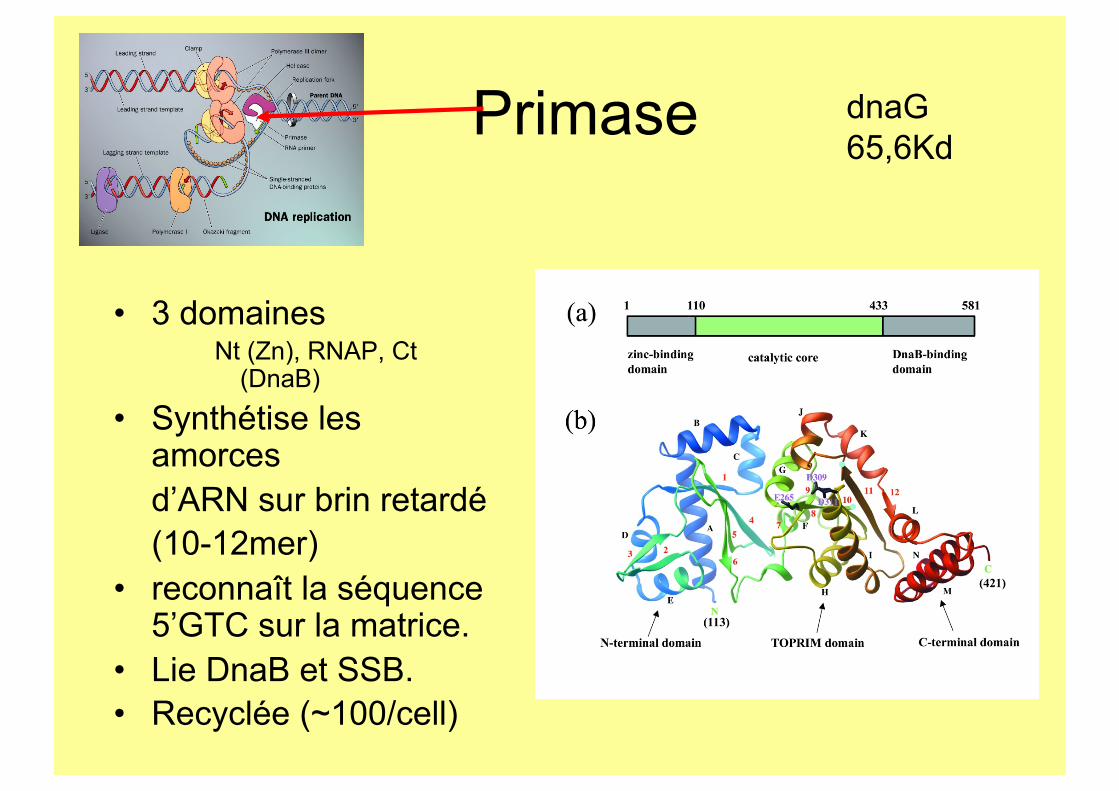
Primase
• 3 domaines Nt (Zn), RNAP, Ct
(DnaB)
• Synthétise les amorces d’ARN sur brin retardé (10-12mer)
• reconnaît la séquence 5’GTC sur la matrice.
• Lie DnaB et SSB. • Recyclée (~100/cell)
dnaG 65,6Kd
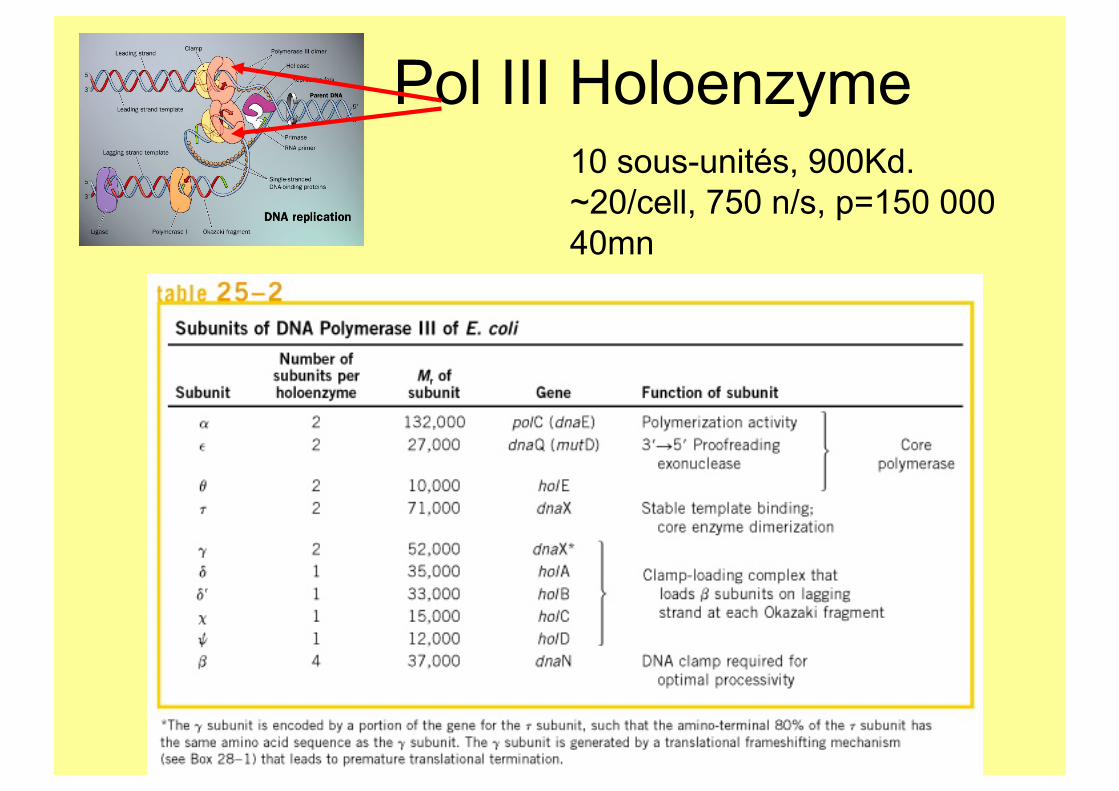
Pol III Holoenzyme 10 sous-unités, 900Kd. ~20/cell, 750 n/s, p=150 000 40mn
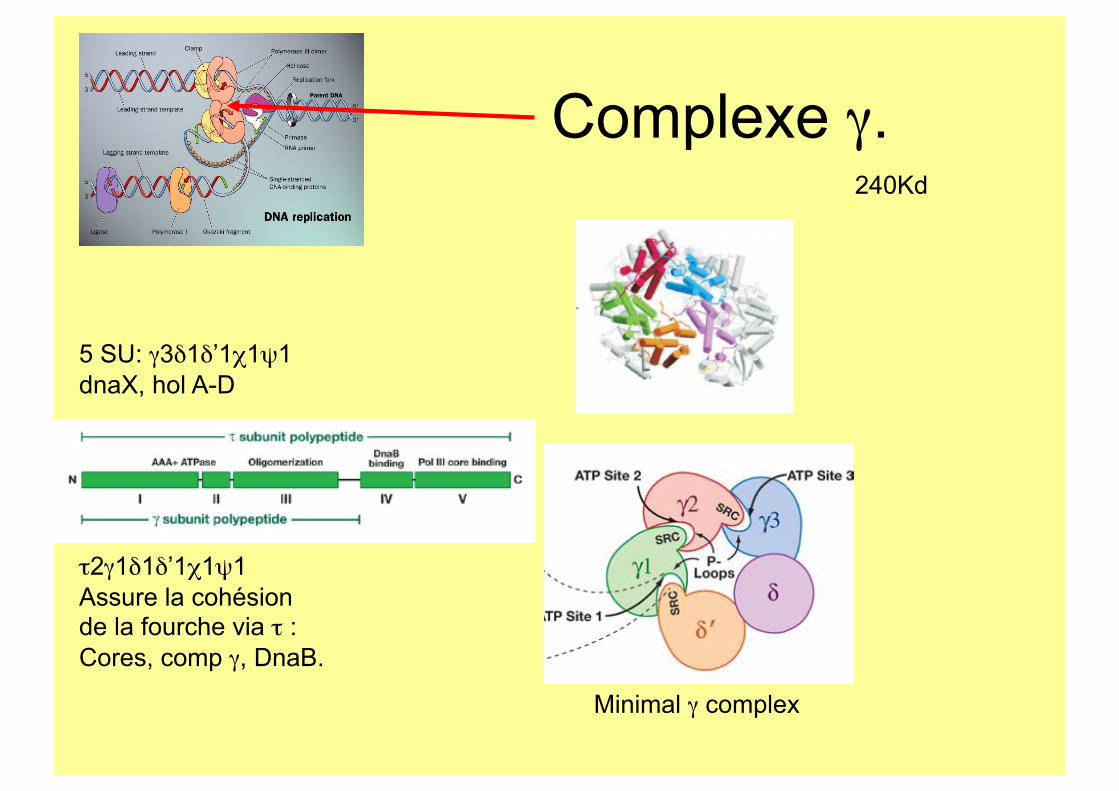
Complexe γ.
5 SU: γ3δ1δ’1χ1ψ1 dnaX, hol A-D
τ2γ1δ1δ’1χ1ψ1 Assure la cohésion de la fourche via τ : Cores, comp γ, DnaB.
240Kd
Minimal γ complex

Facteur de processivité
dnaN Homodimère 80Kd 300 dimères/cell
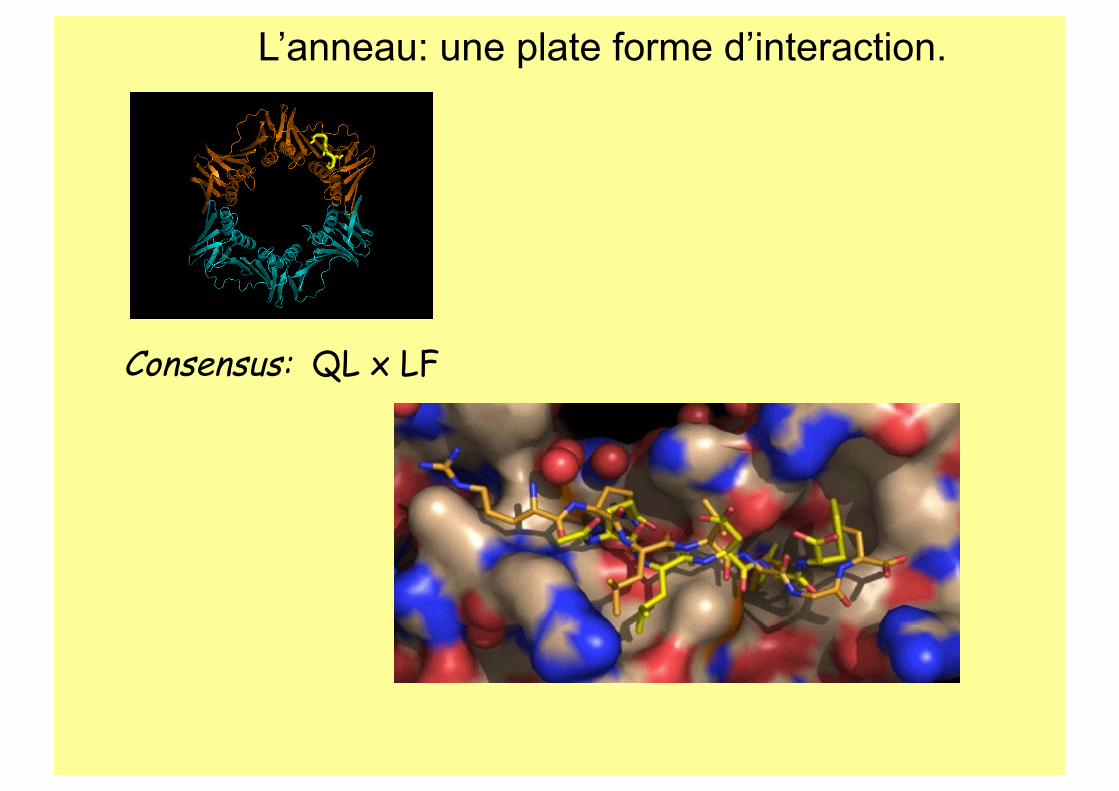
L’anneau: une plate forme d’interaction.
Consensus: QL x LF
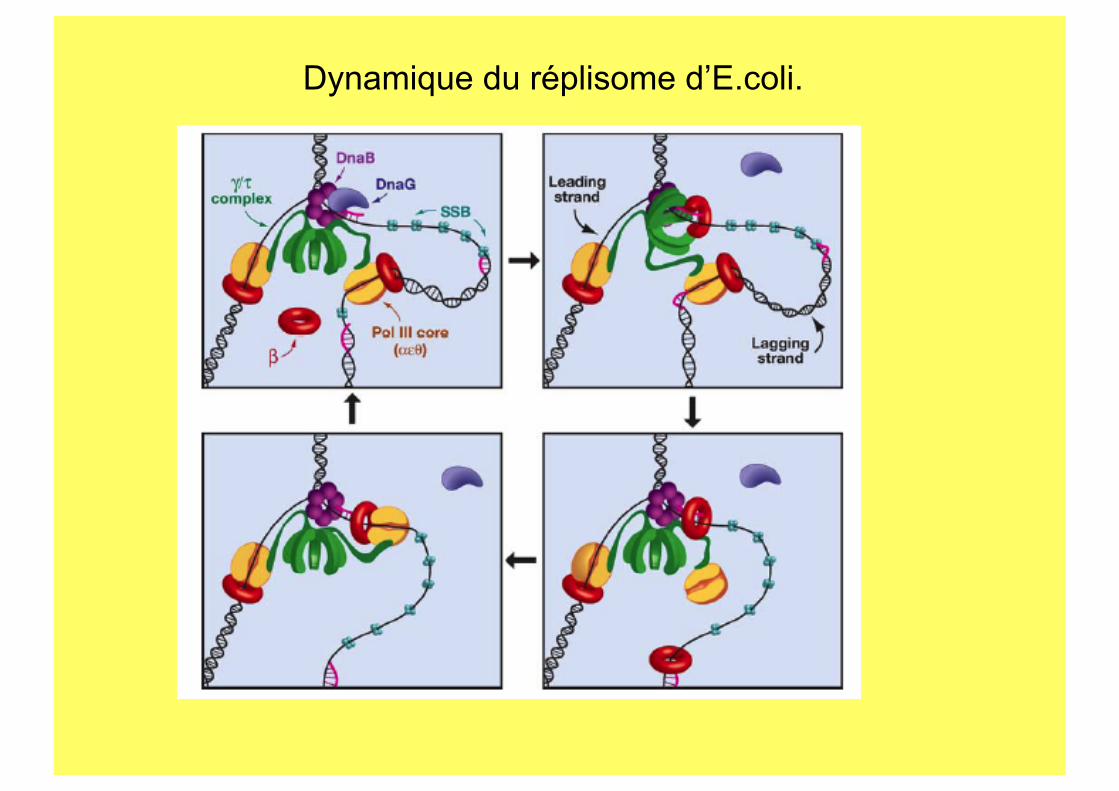
Dynamique du réplisome d’E.coli.
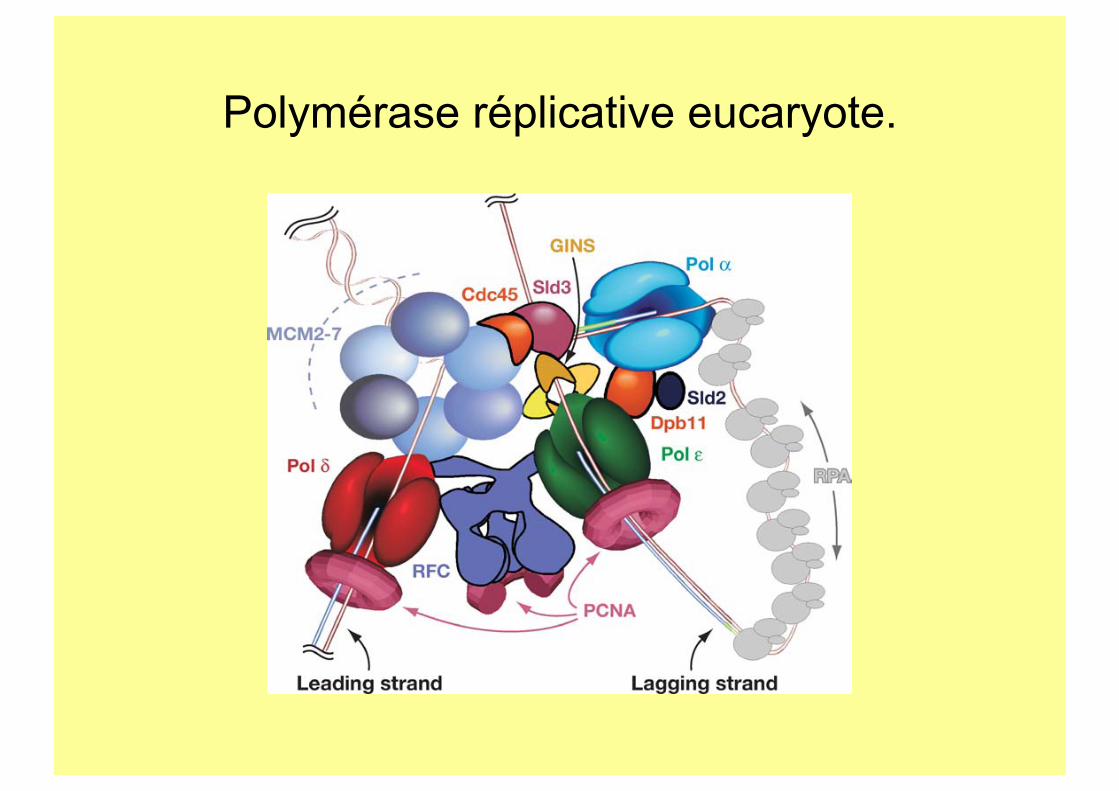
Polymérase réplicative eucaryote.

VI. Réplication d’un ADN endommagé: Mécanismes de translésion.
réponse au stress
Reparation
cell
Dommages ADN
Tolerance des Dommages
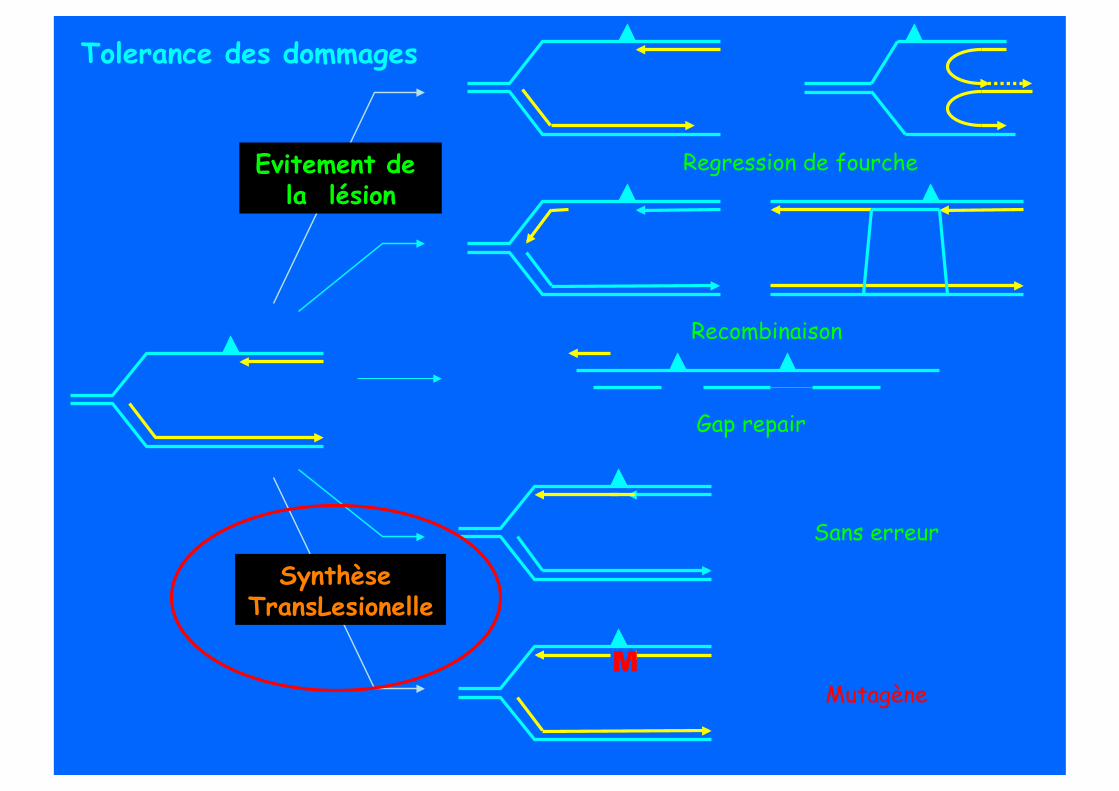
Sans erreur
Mutagène
Synthèse TransLesionelle
M
Tolerance des dommages
Regression de fourche
Recombinaison
Evitement de la lésion
Gap repair

Polymérases translésionnelles
H.Sapiens E.coli
Pol II (polB)
Pol V (umuC)
Pol IV (dinB)
Pol ι (rad30B)* Pol κ (DINB1)
Pol ζ (rev3/rev7)*
*:specifique eucaryotes
REV1 (dCMP)��
Y Y
B
Un même organisme dispose de plusieurs polymérases TLS.
Pol η (rad30A)*
B
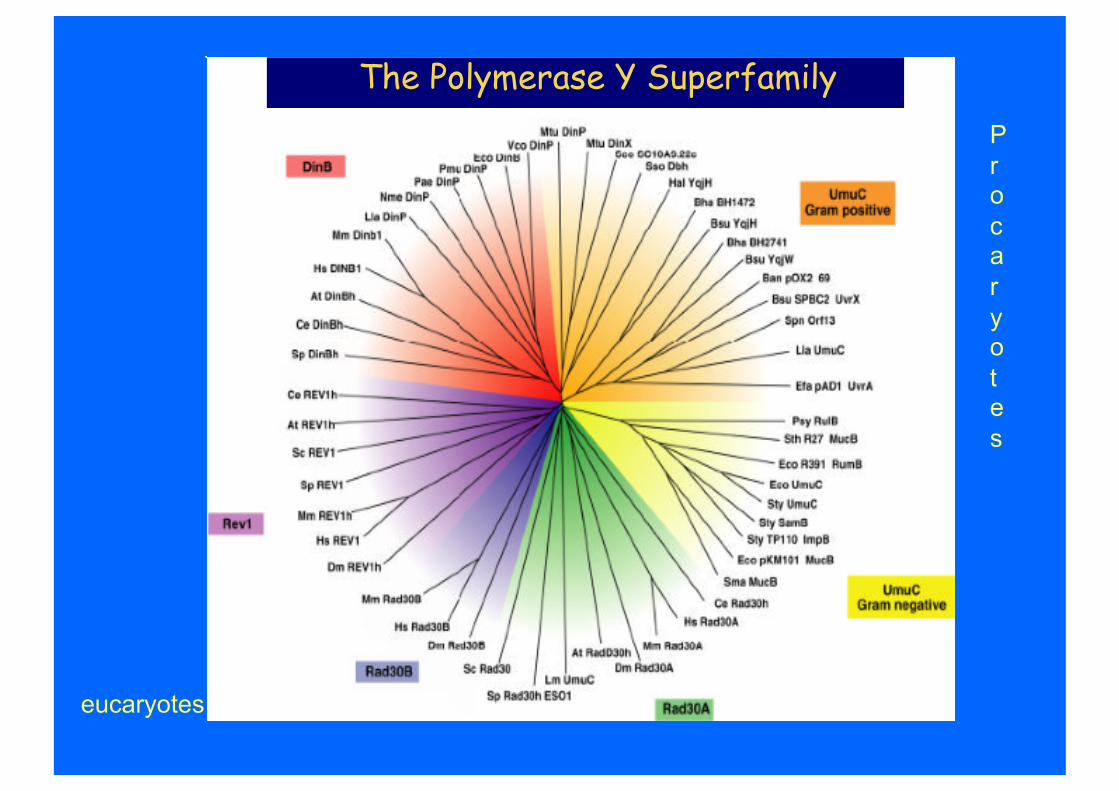
The Polymerase Y Superfamily
eucaryotes
P r ocaryotes

Propriétés des polymérases.
• Réplicatives
• fidèles (10-5-10-6) • correction de lecture • Processives (>50kb) • Site actif contraint
Translésionnelles.
• Peu fidèles (10-2-10-4) • Pas de correction de
lecture • Peu processives (qques
nt) • Site actif large • Plusieurs par cellule

TLS chez les procaryotes.
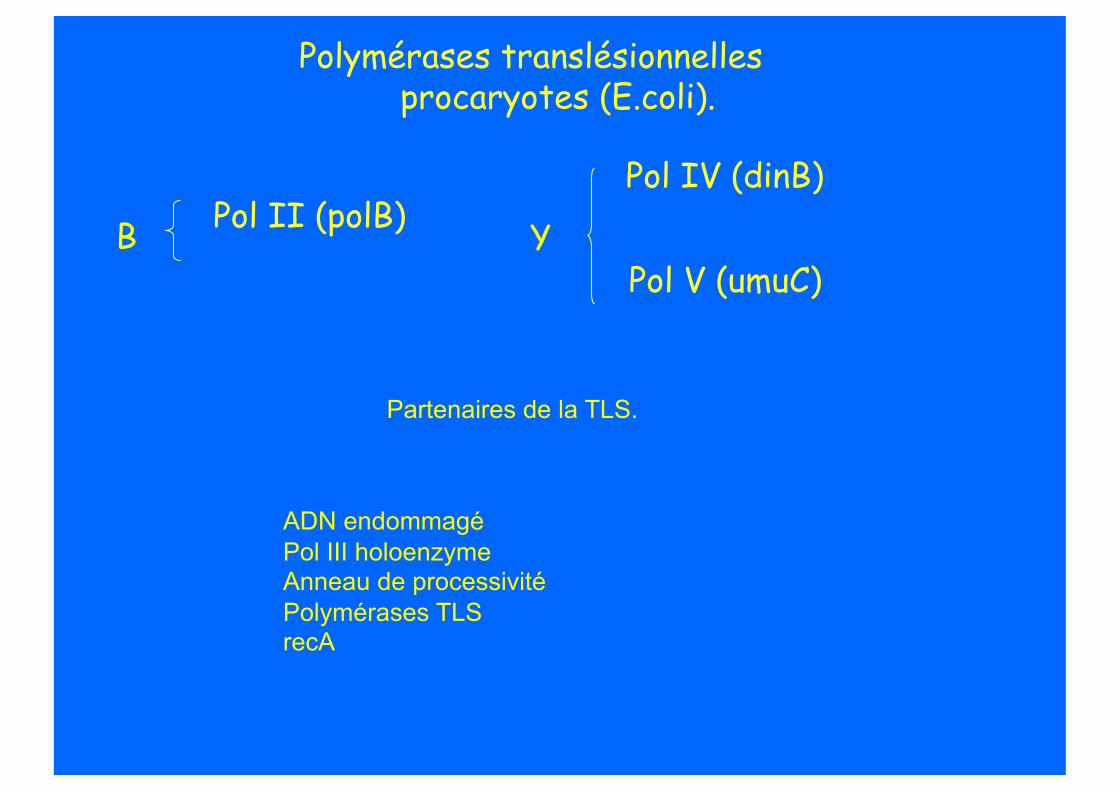
Polymérases translésionnelles procaryotes (E.coli).
Pol V (umuC)
Pol IV (dinB)
Y Pol II (polB) B
Partenaires de la TLS.
ADN endommagé Pol III holoenzyme Anneau de processivité Polymérases TLS recA
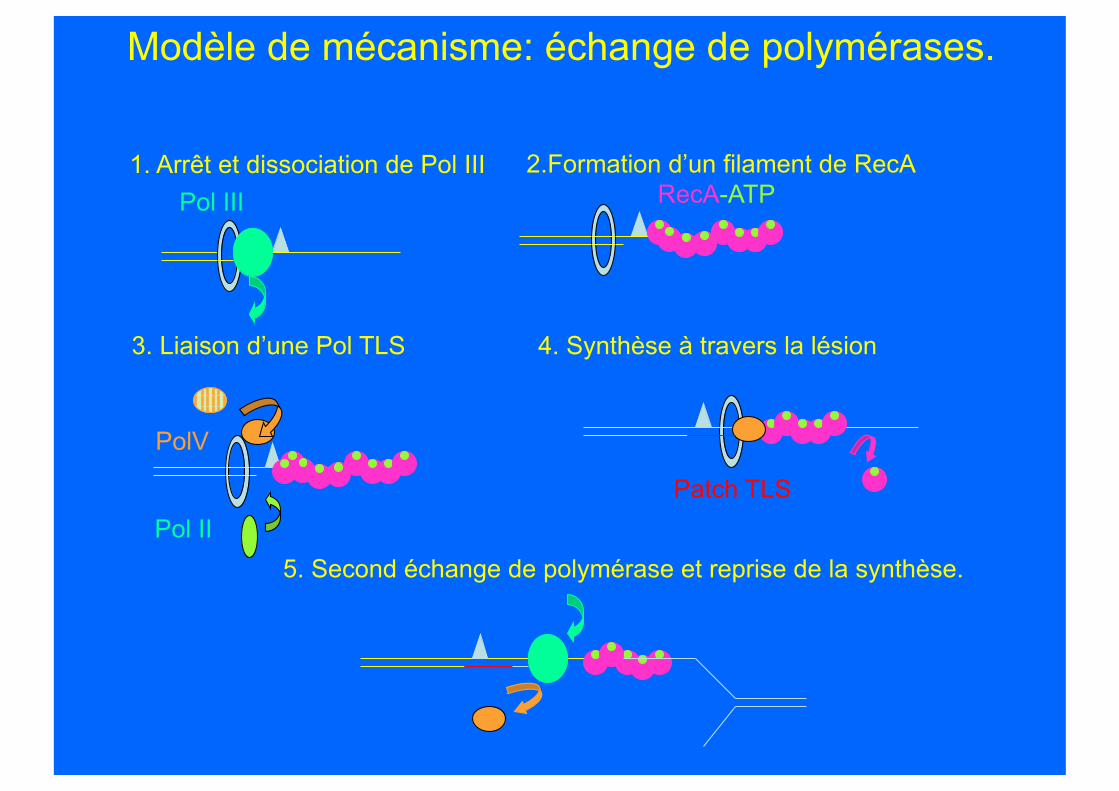
Modèle de mécanisme: échange de polymérases.
Pol III RecA-ATP 1. Arrêt et dissociation de Pol III
4. Synthèse à travers la lésion 3. Liaison d’une Pol TLS
2.Formation d’un filament de RecA
PolV
Pol II Patch TLS
5. Second échange de polymérase et reprise de la synthèse.
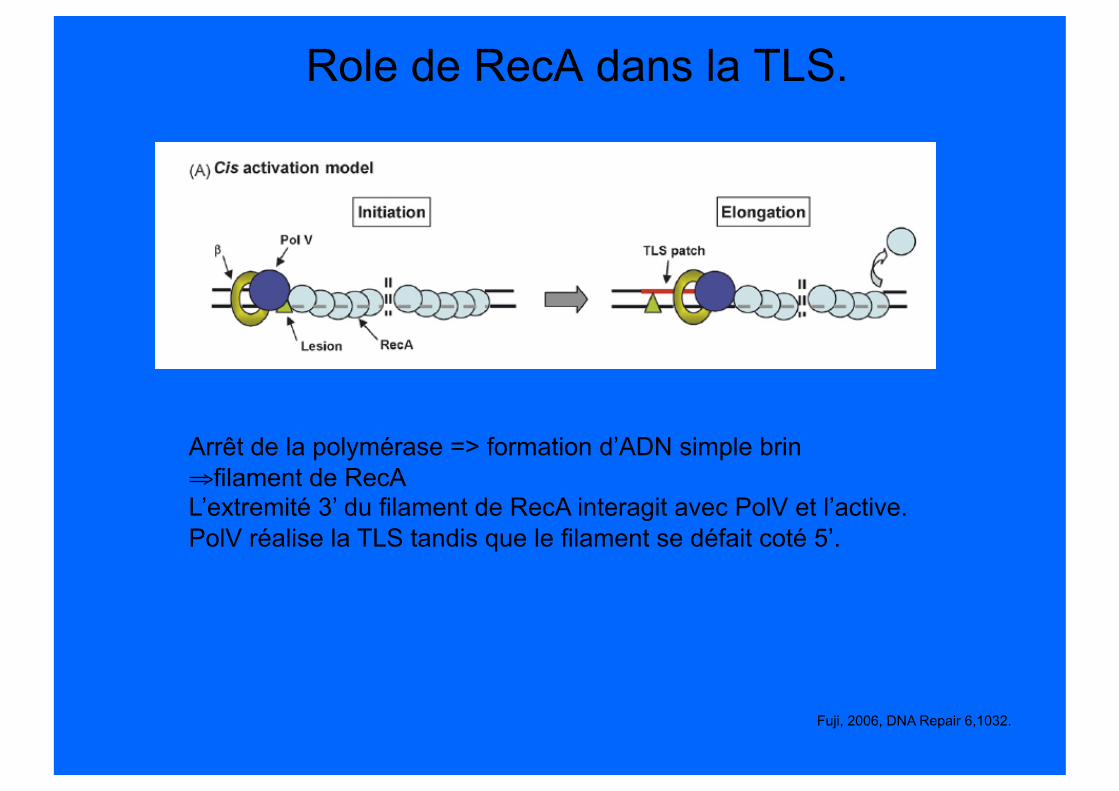
Role de RecA dans la TLS.
Fuji, 2006, DNA Repair 6,1032.
Arrêt de la polymérase => formation d’ADN simple brin ⇒ filament de RecA L’extremité 3’ du filament de RecA interagit avec PolV et l’active. PolV réalise la TLS tandis que le filament se défait coté 5’.

TLS chez les eucaryotes.
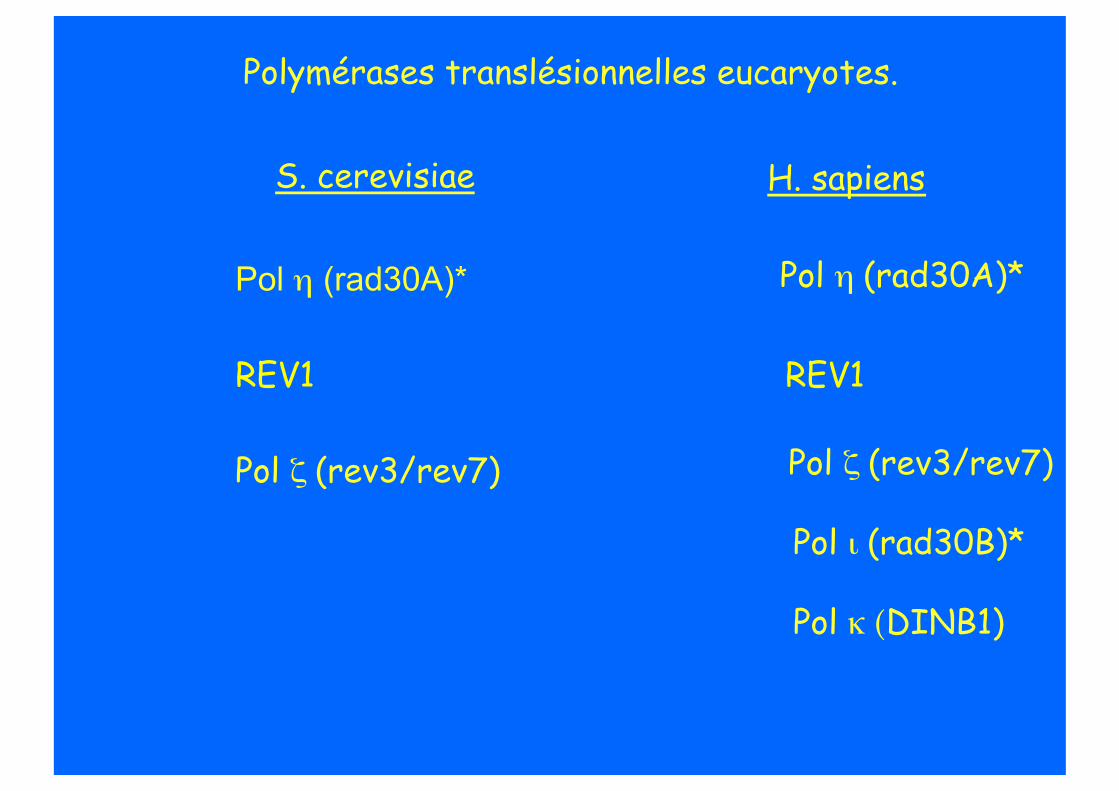
Polymérases translésionnelles eucaryotes.
H. sapiens S. cerevisiae
Pol ι (rad30B)*
Pol κ (DINB1)
Pol η (rad30A)*
REV1
Pol η (rad30A)*
REV1
Pol ζ (rev3/rev7) Pol ζ (rev3/rev7)

Pol η.
• XPV groupe variant (20%) • TLS de dimères de pyrimidine
(cyclobutane), incorpore A • => rôle important de polη pour maintenir un
taux de mutation faible • Si polη mutée-> polι ou/et polκ la
remplacent mais mutations.

Rôle des autres polymérases.
• Moins précis. – Pol ι, pol κ: action concertée
ι: incorpore en face de certaines lésions mais pas d’extension. Fonction mal connue
κ: analogue de PolIV, role in NER?
– Pol ζ (Rev3-Rev7): mutagène. – Rev1(dCMP transferase), agirait avec pol ζ, η ι et κ: rôle de plate forme?
• Localisation dans le noyau (replication factories).

Régulation de la TLS par ubiquitination.
• Modification post traductionnelle • Fixation d’ubiquitine: 8.5 Kd, 76AA
• Mono-Ub • Poly-Ub
– 48: dégradation – 63: régulation
mqifvktltgktitlevepsdtienvkakiqdkegippdqqrlifagKqledgrtlsdyniqKestlhlvlrlrgg1 48 63 76
2 étapes: activation: fixation sur une C d’une enzyme de conjugaison (Rad6, Mms2/Ubc13)
ligation: transfert sur un K activée du susbtrat (Rad18, Rad5)

Mode d’activation de PCNA (cellules de mammifères).
PCNA PCNA K164 Rad6
Rad18
MonoUbiquitination sur L164
Rad6, Mms2/Ubc13:Ub Conjugating Enzyme Rad18, Rad5: Ub Ligase
PCNA
K63
Mms2/Ubc13 Rad5
TLS Voie majeure
Voie d’évitement de la lésion sans erreur
Voie rare (1/20)
Dommages À l’ADN
UV, Cis Pt, MMS…
Arrêt de la réplication
USP1
Mécanisme inconnu

Régulation de la TLS.
• Les Pol TLS ont un site de liaison de l’ubiquitine -> stabilisation du complexe avec PCNA.
• Les pol interagissent ensembles • Co-localisation dans les factories (faibles interactions)
UBM: ι et REV1 UBZ: η et κ Cruciaux: interaction avec PCNA
fonction TLS role dans la localisation
Lehmann, DNA Repair, 2007

Modèle de TLS eucaryote.
1- arrêt de la polymérase 2- formation de SS 3- recrutement de rad6/rad18 et clivage de UPS1 4-mono ubiq de PCNA 5- => augmente l’affinité des polY (site de liaison Ub) pour PCNA 6- fixation d’une pol TLS sur PCNA 7- bypass de la lésion ou échange de pol 8- dissociation et recharge de la pol réplicative
Lehmann, DNA Repair, 2007


















