
Diabète sucré félin : stratégies nutritionnelles par T.A. Lutz; A5205 ...
42
DMV, Responsable des Éditions Scientifiques, Communication, Groupe Royal Canin DMV, PhD, Dipl. ACVN, Dipl. ECVCN Directeur Scientifique Nutrition-Santé pour le Centre de Recherche Royal Canin BVSc (Hons) PhD, Dipl. ACVIM, Dipl. ACVN Directrice Scientifique Royal Canin aux États-Unis Pascale Pibot Vincent Biourge Denise Elliott Nutrition Encyclopédie de la Clinique Féline Fermez cette fenêtre pour retourner sur IVIS Ce livre est reproduit sur le site d'IVIS avec l'autorisation de Royal Canin. IVIS remercie Royal Canin pour son soutien.
Transcript of Diabète sucré félin : stratégies nutritionnelles par T.A. Lutz; A5205 ...

DMV, Responsable desÉditions Scientifiques,
Communication,Groupe Royal Canin
DMV, PhD, Dipl. ACVN, Dipl.
ECVCNDirecteur ScientifiqueNutrition-Santé pour
le Centre de RechercheRoyal Canin
BVSc (Hons) PhD,Dipl. ACVIM, Dipl. ACVNDirectrice
Scientifique Royal Canin aux
États-Unis
Pascale Pibot Vincent Biourge Denise Elliott
NutritionE n c y c l o p é d i e d e l a
Clinique Féline
Fermez cette fenêtre pour retourner sur IVIS
Ce livre est reproduit sur le site d'IVIS avec l'autorisation de Royal Canin. IVIS remercie Royal Canin pour son soutien.

Diab
ète
181
Diabète sucré félin :stratégies nutritionnelles
Thomas A. LUTZDMV, PhD
181
AGNE : acide gras non estérifiéAGPI : acide gras polyinsaturéAGPI n-3 : acide gras polyinsaturé oméga-3DS : diabète sucréDS1 : diabète sucré de type 1DS2 : diabète sucré de type 2DSID : diabète sucré insulinodépendantDSNID : diabète sucré non insulinodépendantFT : fibres totales
GK : glucokinaseGLP-1 : glucagon-like peptide-1GLUT (1, 2 ou 4) : transporteur du glucose detype 1, 2, ou 4HGPIV : épreuve d’hyperglycémie provoquéepar voie intraveineuseIAPP : islet amyloid polypeptideIGF-1 : insulin-like growth factor 1IL-1b : interleukine béta
MS : matière sèchePIG : peptide insulinotrope glucose-dépen-dant ou polypeptide gastrique inhibiteurPPAR g : peroxisome proliferator-activatedreceptor gammaTNF-a : tumor necrosis factor alphaTSA : test de stimulation à l’arginineTSG : test de stimulation au glucagonTSI : test de stimulation à l’insuline
ABRÉVIATIONS UTILISÉES DANS CE CHAPITRE
1- Prévalence du diabète sucré félin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183
2- Tableau clinique . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
3- Spécificités du métabolisme félin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 184
4- Classification du diabète sucré . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 185
5- Introduction au diabète sucré félin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 186
6- Aspects physiologiques du contrôle de l’homéostasie nutritionnelle . . . . . . . . . . . . . . . . . 184
7- Physiopathologie du diabète sucré félin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 489
8- Diabète sucré transitoire . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 199
9- Conséquences à long terme du diabète sucré . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 200
10- Diagnostic du diabète sucré félin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 201
11- Stratégies thérapeutiques . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 204
12- Aspects diététiques du traitement du diabète sucré félin . . . . . . . . . . . . . . . . . . . . . . . . . 206
13- Influence du niveau de protéines sur la fonction rénale . . . . . . . . . . . . . . . . . . . . . . . . . . 211
14- Recommandations pratiques pour le chat diabétique . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 211
Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 212
Questions fréquemment posées . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 213
Références . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 214
Informations nutritionnelles Royal Canin . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 218

182
Le diabète sucré est une maladie endocrinienne fréquente du chat. Sa prévalence a augmenté au cours
des 30 dernières années : elle est d’environ 1 cas pour 200 chats.Cette augmentation pourrait être directement corrélée à celle de l’obésité chez le chat. Le diabète félin ressemble en beaucoupde points au diabète sucré de type 2 de l’homme (DS2), notamment du point de vue de la physiopathologie, des facteursde risques et des stratégies thérapeutiques. Les recommandations nutritionnelles pour le chat diabétique ont évolué au cours de ces dernières années et l’accent est maintenant mis sur desrégimes relativement riches en protéines et pauvres en glucides. La définition d’un aliment “riche en protéines et pauvre en glucides” varie selon les auteurs, mais d’une façon générale, ces termes font référence à un taux protéique de 50% ou plus sur matière sèche et à une part glucidique inférieure à 15%. Les valeurs sont précisées dans ce chapitre à chaque fois que cela est nécessaire. Ce type de régime, associé à une thérapieinsulinique rigoureuse et bien conduite, a conduit à une augmentation des cas de rémission du diabète sucré félin. Ce chapitre fait le point sur la physiopathologie du diabète sucré félin et ses stratégies thérapeutiques, en tenant compte des besoins nutritionnels du chat et des recommandations au sujet des aliments hyperprotéiques et hypoglucidiques.
Diabète sucré félin : stratégies nutritionnelles
Thomas A. LUTZDMV, PhDThomas Lutz est diplômé de la faculté vétérinaire de l’Université de Berlin depuis 1989. Il obtient son doctorat de médecine vétérinaire à
l’Institut de Physiologie vétérinaire de l’Université de Zurich en 1991. En 1995, il effectue un PhD sur le diabète sucré félin à l’Université
du Queensland (Brisbane, Australie) et revient en 1999 à l’Institut de Physiologie vétérinaire de l’Université de Zurich en tant que direc-
teur de recherches. Depuis 2004, il est Professeur de Physiologie vétérinaire appliquée à Zurich. Ses domaines de recherche privilégiés
concernent la régulation neuroendocrinienne de la consommation alimentaire et le diabète sucré félin. Il a publié plus de 80 articles scienti-
fiques dans des revues indexées.
Diab
ète

1 -Prévalence du diabète sucré félinLe diabète sucré (DS) est une maladie endocrinienne fréquentedu chat. Sa prévalence se situe environ entre 1: 400 et 1: 100(Panciera et coll, 1990; Rand et coll, 1997). Une étude rétrospec-tive réalisée à partir des cas recensés dans des écoles vétérinairesmontre que la prévalence du diabète félin a augmenté plus de 10fois au cours des 30 dernières années : moins d’un cas sur mille en1970, plus de douze cas pour mille en 1999 (Prahl et coll,2003;2007). Sur la même période cependant, le taux de mortali-té a chuté de plus de 40 % à moins de 10 %, prouvant que leschats diabétiques peuvent être traités avec succès. Ceci peut enpartie s’expliquer par une meilleure compréhension de la physio-pathologie du diabète sucré. Les facteurs de risque à prendre encompte pour le développement de cette maladie sont l’âge, le sta-tut sexuel et l’obésité (Tableau 1).
Lien entre diabète sucré félin et obésité
Ce dernier facteur de risque est probablement responsable del’augmentation de la prévalence du DS félin de nos jours carl’obésité féline est directement corrélée à la résistance à l’insuli-ne (Scarlett et coll, 1994; Hoenig, 2006; 2007; voir aussi Physiopa-thologie du diabète félin), et l’obésité est très fréquente dans la popu-lation féline actuelle : 20 à probablement 35-40 % des chats sonten surpoids ou obèses (Baral et coll, 2003; Lund et coll, 2005; Diezet Nguyen, 2006; German, 2006).
Influence de l’âge
Le DS félin touche en général des chats d’âge mûr et avancé, avec un pic après 7 ans. Les chats demoins d’1 an ont 50 fois moins de chance de développer un diabète que les chats de plus de 10 ans(Prahl et coll, 2003).
Influence du statut sexuel
Le risque de développer un diabète semble plus élevé pour les chats mâles que pour les femelles. Bienque cette situation soit similaire en médecine humaine au moins avant l’âge moyen de la ménopause,la raison de cette prédisposition liée au sexe chez le chat est pour le moment inconnue. Elle n’est pro-bablement pas directement liée à l’imprégnation hormonale puisque la plupart des chats mâles sontcastrés, et que la castration ne fait pas varier le risque de développement du diabète lorsque le poids etl’âge sont comparables (Prahl et coll, 2003).
Influence de la race
Peu d’études se sont intéressées à d’éventuelles prédispositions raciales pour le diabète félin. Bien qu’uneétude rétrospective américaine ne montre aucune augmentationde la prévalence chez certaines races de chats, mais à l’inverse unrisque plus faible chez les chats de race par rapport aux chats demaison (Prahl et coll, 2003), quelques données australiennes révè-lent une prévalence plus importante chez le Burmese (Rand etcoll, 1997) (Figure 1). Une prédisposition similaire a été relevéeen Angleterre (McCann et coll, 2007). En l’absence d’autresétudes du même type ailleurs qu’en Australie et en Angleterre, iln’est pas possible de généraliser cette observation.
1 -
Prév
alen
ce d
u di
abèt
e su
cré
félin
Figure 1 - Chat BurmeseUne étude australienne indique que les chats Burmese sont prédisposés génétiquement au développement du diabète sucré (Rand et coll, 1997).Cependant, d’éventuelles prédispositionsraciales sont toujours discutées.
183
Diab
ète
TABLEAU 1 - FACTEURS DE RISQUE POUR LE DÉVELOPPEMENTDU DIABÈTE SUCRÉ (DS) CHEZ LE CHAT
(Nelson, 2005; Rand et Marshall, 2005 ; McCann et coll, 2007)
ÂgeLe DS félin est plus fréquent
chez le chat âgé
SexeLe DS affecte plus souvent
les chats mâles que les femelles
StérilisationFacteur de risque indirect :
la stérilisation favorise l’obésité
ObésitéLes chats obèses ont un risque augmenté de développer un DS
Activité physiqueLe DS félin survient plus volontiers
chez des chats sédentaires
Race Burmese ?
Traitementsacétate de mégestrol,
glucocorticoïdes
Maladie sous-jacente infection systémique, stomatite

2 -Tableau clinique(voir également: Nelson, 2005)
La plupart des chats diabétiques ont plus de 7 ans. Les signes cliniquesclassiques incluent une polyurie osmotique qui se développe suite à l’hy-perglycémie, une polydipsie secondaire et souvent une polyphagie. Uneforte proportion de chats diabétiques est obèse au moment du diagnostic(Figure 2). Une perte de poids peut survenir malgré l’augmentation del’appétit, mais les chats sont souvent en surpoids lorsqu’ils sont présentésau vétérinaire pour la première fois. La maigreur est exceptionnelle.
Certains chats diabétiques sont léthargiques suite à la déshydratation. Laneuropathie diabétique peut entraîner une faiblesse des postérieurs et unedémarche plantigrade (Figure 3). Une atrophie des muscles des posté-rieurs peut être présente. La lipidose hépatique se traduit par une hépa-tomégalie. Les complications infectieuses telles que stomatites ou cystitesse développent ultérieurement.
3 -Spécificités du métabolisme félin
Adaptation à un régime carnivore
Le chat est un carnivore, ce qui le distingue du chien qui est omnivore.Ses proies naturelles (par exemple, les souris), contiennent 70 à 80 %d’eau. Par rapport à la matière sèche (MS), ce type de régime contientenviron 55-60 % de protéines, 35 % de matières grasses, mais moins de10 % de glucides. En revanche, la plupart des aliments industriels secspour chats contiennent des taux élevés de glucides, principalement repré-sentés par l’amidon des céréales. Ce taux est plafonné autour de 40 % surMS dans le cas de croquettes hyperdigestibles pour chats. Les chats nour-
2 -
Tabl
eau
clin
ique
184
Figure 2 - Chat obèse diabétique (10 kg) de 11 ansLes chats obèses ont un risque accru de développer un diabètesucré.
Ave
c l’a
imab
le a
utor
isatio
n du
Pro
f. C
.R
eusc
h,
Vet
suiss
e-Fa
culty
Uni
vers
ity o
f Zur
ich
(Ave
c l’a
imab
le a
utor
isatio
n du
Pro
f. C
.R
eusc
h,V
etsu
isse-
Facu
lty U
nive
rsity
of Z
uric
h).
0
1
2
3
4
5
Diab
ète
Figure 3 - Chat plantigrade à cause d’une neuropathie diabétique. La démarche plantigrade est typique de la neuropathie diabétique du chat.
FIGURE 4A - ABSENCE D’HYPERGLYCÉMIE POSTPRANDIALE CHEZ DES CHATSRECEVANT UN ALIMENT RICHE EN PROTÉINES (54% DE PROTÉINES
ET 8% DE GLUCIDES SUR MATIÈRE SÈCHE)
Après 24 heures de jeûne, les chats ont accès à un repas test correspondant à 50% de leursbesoins quotidiens. Ce repas est proposé pendant 10 minutes pendant lesquelles toute la nourriture est ingérée. Ce graphique montre la glycémie de 10 chats sains justeavant et après la présentation du repas.
Glyc
émie
(mm
ol/L
)
valeurde
base
fin durepas
15min
30min
1 h 2 h 5 h
Temps

ris avec un aliment riche en protéines (54 % MS) ne montrent pas d’hy-perglycémie postprandiale (Martin et Rand, 1999) (Figure 4A), sauf lorsquedes quantités assez élevées de sucres simples sont ajoutées (Figure 4B). Ceciexplique probablement en partie pourquoi les aliments hyperprotéiques,c’est-à-dire proches du régime carnivore initial du chat, facilitent le contrô-le de l’homéostasie du métabolisme nutritionnel chez le chat diabétique(voir ci-dessous).
Les chats ont généralement un besoin élevé en acides aminés indispensables,comme l’arginine et la taurine. Un déficit en taurine a été incriminé en tantque facteur prédisposant au développement du DS. Cependant, le fait quela taurine soit utilisée pour la prévention ou le traitement de la rétinopathieou de la neuropathie diabétiques (Franconi et coll, 2006) ne permet pas for-cément d’établir un lien de cause à effet. Aucune preuve expérimentalen’existe à ce jour permettant une telle association chez le chat.
Intensité de la néoglucogénèse
Chez le chat, la néoglucogénèse à partir des acides aminés ne faiblit pasmême si l’ingéré protéique est insuffisant (Rogers et coll, 1977).
L’activité des enzymes de la néoglucogénèse est bien plus forte chez le chatque chez le chien (Washizu et coll, 1998; Washizu et coll, 1999; Takeguchi etcoll, 2005). D’un autre côté, la faible expression hépatique de la glucokina-se (GK) ou la faible activité enzymatique hépatique du chat entraîne uneinsuffisance fonctionnelle de la GK (Washizu et coll, 1999; Schermerhorn,2005; Tanaka et coll, 2005). Cependant, la régulation de l’activité de la GKchez le chat semble différer de celle d’autres espèces car l’activité de la pro-téine régulatrice de la GK est très faible (Schermerhorn, 2005), ce qui, dansd’autres espèces entraînerait une forte activité de la GK. L’activité des autresenzymes clés de la glycolyse, comme l’hexokinase, qui compense peut-êtrepartiellement la faible activité de la GK, est plus forte chez le chat que chezle chien (Washizu et coll, 1999).
4 -Classification du diabète sucré
Les différentes formes de diabète sucré ont été décrites de plusieurs façons chez l’homme et dans d’autresespèces. La terminologie suivante est basée sur la physiopathologie sous-jacente et sera utilisée danstout ce chapitre. Le diabète sucré primaire peut être divisé en un diabète sucré de type 1 (DS1) et undiabète sucré de type 2 (DS2) (Tableau 2).
Chez l’homme, il était d’usage de les nommer respectivement diabète juvénile et diabète de l’adulte.Cependant, l’augmentation massive de l’obésité chez l’enfant fait que jusqu’à 50 % des enfants diabé-tiques souffrent maintenant d’un DS2 alors qu’ils n’étaient de 5-10 % auparavant. Ces termes nedevraient donc plus être utilisés.
Diabète sucré insulinodépendant (DSID) et diabète sucré non-insulinodépendant (DSNID) sont destermes purement descriptifs qui se réfèrent à la nécessité ou non de traiter le patient diabétique avecde l’insuline pour contrôler le métabolisme. La pathophysiologie sous-jacente n’apparaît pas dans cestermes qui ne sont donc pas utilisés dans ce chapitre.
D’un point de vue physiopathologique, le diabète le plus fréquent du chat ressemble au DS2 del’homme (pour rappel, voir Henson et O’Brien, 2006). Il est développé dans le paragraphe suivant. Desmodifications histologiques évoquant un syndrome de type DS1 au sein des îlots pancréatiques sontdécrites chez le chat (Nakayama et coll, 1990), mais cela n’est pas fréquent. De plus, les chats ne for-
4-C
lass
ifica
tion
du d
iabè
te s
ucré
FIGURE 4B - INFLUENCE DU RÉGIME ALIMENTAIRESUR LA GLYCÉMIE POSTPRANDIALE CHEZ LE CHAT
Absence d’hyperglycémie postprandiale chez 12 chats recevantun aliment riche en protéines (54% protéines et 8% de glucides sur matière sèche), sauf si 20% de glucose est ajouté.
0
2
-20 30 60 90 120
150
180
210
240
270
300
330
360
4
6
8
10
12
14
16
18
aliment additionné de glucose (20%)aliment riche en protéines
Temps après la prise du repas (min)
Glyé
mie
(mm
ol/L
)
185
Diab
ète
De par son régime carnivore traditionnel, le chat a développé unecapacité de néoglucogénèse intense à partir des acides aminés.
© L
enfa
nt/R
C
repa
s

5 -
Intr
oduc
tion
au d
iabè
te s
ucré
fél
in
ment pas d’auto-anticorps contre des antigènes des cellules b ou contre l’insuline (Hoenig et coll, 2000),ce qui va à l’encontre de l’hypothèse d’une forme de diabète auto-immune typique du DS1. Enfin, ilest maintenant admis que la physiopathologie du DS2 fait également intervenir un processus inflam-matoire et à médiation immunitaire (Donath et coll, 2005). La présence d’un processus inflammatoiren’exclut donc pas un DS2.
5 - Introduction au diabète sucré félinProblèmes majeurs liés au diabète sucré félin
D’un point de vue physiopathologique, le diabète félin et le DS2 de l’homme sont des maladies endo-criniennes comparables. Afin de mieux comprendre les modifications sous-jacentes, des références àdes données issues de modèles expérimentaux, le plus souvent de rongeurs, sont faites dans ce para-graphe.
Les problèmes majeurs rencontrés chez le chat diabétique et lors de DS2 chez l’homme sont :- résistance à l’insuline entraînant une modification de l’utilisation des nutriments dans les tissus sen-
sibles à l’insuline.- perturbation de la fonction des cellules b du pancréas, entraînant des anomalies de la sécrétion d’in-
suline et d’amyline.- dépôt d’amyloïde au sein des îlots pancréatiques résultant de la précipitation de l’amyline (islet amy-
loid polypeptide) (Figure 5).
D’autres problèmes seront également discutés plus loin. Le fait que le dysfonctionnement des cellulesb précède ou non la perturbation de l’activité de l’insuline lors de DS2 fait encore débat. Ces deux ano-malies sont généralement présentes au moment du diagnostic et contribuent à détériorer la situationmétabolique. La glucotoxicité les rend également responsables de la pérennisation de la maladie quiest couramment observée.
Diabète sucré félin et génétique
Chez l’homme, les facteurs génétiques prédisposant les individus au développement du DS2 font l’ob-jet de recherches intenses. Plusieurs mutations et polymorphismes génétiques ont été associés à unrisque accru de développement du DS2 chez certains individus diabétiques (pour une revue complète,voir e.g. Barroso, 2005; Malecki, 2005). Cependant, l’augmentation massive du DS2 chez l’homme n’estpas le résultat d’un changement profond du contexte génétique, mais plus la conséquence d’un chan-gement de mode de vie incluant l’abondance de nourriture et le manque d’activité physique, qui lerend plus vulnérable à l’obésité et donc au DS2. Un contexte génétique favorable par le passé peutdonc avoir des effets néfastes aujourd’hui.
186
Diab
ète
TABLEAU 2 - CLASSIFICATION DU DIABÈTE SUCRÉ FÉLIN
Type de diabèteFréquence
chez le chatAnomalies majeures
DS primaire
DS de type 1 (DS1) rare destruction auto-immune des cellules b du pancréas
DS de type 2 (DS2) au moins 90%
des casdysfonctionnement des cellules b‚ insulinorésistance,
amyloïdose des îlots pancréatiques
Autres causes de DS
(autrefois appeléesDS secondaires)
Maladie antagoniste
Infection
environ 10 % des cas
insulinorésistance
Pancréatite,tumeur pancréatique destruction des cellules b
Acromégalie effet antagoniste de la GH
Induction par les stéroïdes ex : chats traités par la progestérone (acétate de mégestrol)
DS = diabète sucré GH = hormone de croissance

6 -
Asp
ects
phy
siol
ogiq
ues
du c
ontr
ôle
de l’
hom
éost
asie
nut
ritio
nnel
le
Les connaissances relatives au rôle possible de la génétique sur le développement du diabète sont bienmoins avancées chez le chat que chez l’homme. Certains chats pourraient présenter une prédispositionà l’intolérance au glucose : en effet, chez les chats ayant développé une diminution plus importante dela sensibilité à l’insuline en prenant du poids, la sécrétion basale d’insuline est plus élevée et la sensi-bilité à l’insuline est plus faible pendant la 1re phase de réponse (Appleton et coll, 2001b). Des résultatssimilaires sont décrits par Wilkins et coll (2004). De plus, quelques études suggèrent une prédispositionraciale des chats Burmese au DS (Rand et coll, 1997). Malgré ces pistes, le mode de transmission et lanature des gènes qui pourraient être impliqués ne sont pas connus.
6 -Aspects physiologiques du contrôle de l’homéostasie nutritionnelle
Avant de détailler la physiopathologie du dia-bète félin, quelques aspects du rôle des facteurshormonaux clés sont brièvement résumés. Chezl’animal en bonne santé, la sécrétion pancréa-tique d’insuline est contrôlée par de nombreuxnutriments (Figures 6 et 7). L’action de l’insu-line sur les tissus cibles s’exerce par l’intermé-diaire d’un récepteur à l’insuline. La fixation del’une sur l’autre active la tyrosine kinase liée à cerécepteur et entraîne des effets immédiats(comme la translocation du transporteur de glu-cose sensible à l’insuline GLUT4 et la modifica-tion de l’activité des enzymes métaboliques) etdes effets retardés sous l’influence de la trans-cription génique. Ces derniers sont liés au fac-teur de transcription PPARg (peroxisome prolife-rator-activated receptor gamma). Ce facteur detranscription est la cible des thiazolidinédionesantidiabétiques qui augmentent la sensibilité àl’insuline.
187
Diab
ète
FIGURE 5 - DÉRÉGLEMENTS PHYSIOLOGIQUES MAJEURS LORS DE DIABÈTE FÉLIN
Mode de vieEnvironnement
GénétiqueStérilisation
Acides gras + triglycérides augmentés
Détériorationdes cellules b
Insulino-résistance
Diminution de l’expression de la GLUT 4 (transporteursensible à insuline)
Amyloïdose pancréatique +sécrétion inadéquate d’insulineet d'amyline
OBÉSITÉ DIABÈTE
Dérivés de lasulfonylurée K+
Ca2+
Ca2+
CaMK
Glut2
Insuline
ATP Sécrétion d’insuline
Phosphorylationdes protéines
Glucose
FIGURE 6 - RÉGULATION DE LA SÉCRÉTION D’INSULINEPAR LES CELLULES B DU PANCRÉAS
Le glucose pénètre dans les cellules b du pancréas grâce au transporteur du glucoseGLUT2. Il est métabolisé via la glycolyse et le cycle de Krebsdans les mitochondries.L’adénosine triphosphate (ATP)entraîne la fermeture des canaux K+ sensibles à l’ATP qui sont également les structurescibles des dérivés de la sulfonylurée. Il en résulte une dépolarisation qui ouvre les canaux Ca2+ sensibles au voltage ; le flux de Ca2+
entraîne l’activation des kinases dépendantes du Ca2+ (CaMK)et enfin la sécrétion d’insuline.

6 -
Asp
ects
phy
siol
ogiq
ues
du c
ontr
ôle
de l’
hom
éost
asie
nut
ritio
nnel
le
188
Diab
ète
Détection du glucosepar le pancréas chez le chat
Une forte augmentation de la sécrétion d’insu-line suit l’administration intraveineuse ou oralede glucose chez le chat. De la même manière,l’administration intraveineuse d’acides aminéscomme l’arginine augmente la sécrétion d’insu-line chez le chat. Dans des conditions naturellesd’alimentation, la sécrétion d’insuline induitepar les nutriments semble très efficace car l’hy-perglycémie postprandiale n’existe pas si lerégime est riche en protéines (Figures 4 A et B).La contribution respective des acides aminés oudu glucose à l’augmentation postprandiale dutaux d’insuline circulante reste à éclaircir bienque les voies de détection des nutriments dans lepancréas du chat ont été partiellement élucidées(Schermerhorn, 2006). Malgré la faible activitéde la GK hépatique, il existe une GK pancréa-tique chez le chat et son activité semble compa-rable à celle d’autres espèces. La GK est un deséléments clés qui déterminent la sensibilité auglucose (Schuit et coll, 2001). Parmi eux, des uni-tés de canaux potassiques sensibles à l’ATP(Figures 6 & 7), Kir6.2 et SUR1, sont égale-ment caractérisées chez le chat (Schermerhorn,2006).
Potentialisation de la stimulation de la sécrétion d’insuline par les incrétines
La stimulation de la sécrétion d’insuline par les nutriments est potentialisée par des hormones incré-tines, les plus importantes étant le glucagon-like peptide-1 (GLP-1) et le polypeptide insulinotropiqueglucose-dépendant (PIG, connu auparavant sous le nom de polypeptide inhibiteur gastrique). Les incré-tines sont des hormones sécrétées en réponse aux nutriments et qui potentialisent la sécrétion pan-créatique d’insuline. Les incrétines engendrent une réponse insulinique plus forte lorsque du glucoseest administré par voie orale comparé à la voie parentérale (pour rappels : Drucker, 2001).
Chez les hommes et chez le rat, la sécrétion de GLP-1 fait suite à l’ingestion d’un repas et son taux san-guin augmente en phase postprandiale. Une partie de la sécrétion de GLP-1 iléal est due à un effet directdu glucose intraluminal sur les cellules L par un mécanisme sensible au glucose. Les nutriments stimu-leraient également indirectement la libération iléale de GLP-1 car le taux plasmatique de GLP-1 aug-mente quelques minutes après le repas, c’est-à-dire bien avant qu’un nutriment quelconque n’ait atteintl’iléon (Drucker, 2001). L’effet insulinotropique puissant du GLP-1 est gluco-dépendant et disparaîtlorsque le taux plasmatique du glucose devient inférieur à 4,5 mmol/L environ (80 mg/dL). Le GLP-1n’engendre donc généralement pas d’hypoglycémie. Le GLP-1 potentialise la libération d’insuline sti-mulée par le glucose, probablement en interagissant avec les canaux K+ ATP-dépendants (voir ci-dessuset Figure 6), mais aussi par des effets liés directement à la sécrétion d’insuline.
Il semblerait que le GLP-1 stimule également la biosynthèse d’insuline et la synthèse des cellules sen-sibles au glucose, en particulier le transporteur de glucose GLUT2 et la glucokinase. Enfin, le GLP-1a également des effets trophiques sur les cellules b et leurs précurseurs, en en stimulant leur différen-ciation et leur prolifération. Cet effet s’accompagne d’une inhibition de l’apoptose des cellules b quisemble jouer un rôle majeur dans le développement du DS2 de l’homme (Donath et coll, 2005) et pro-
Le métabolisme des AA et des AG conduit à la formation d’ATP, comme le glucose (voir Figure 6). Certains acides aminés comme l’arginine entraînentune dépolarisation directe (transport électrogène) de la membrane des cellules bet induit un flux de Ca2+. Les acides gras activés (AG-CoA) peuvent également libérer du Ca2+ depuis le milieu intracellulaire. CPT-1 : carnitine palmitoyl transférase-1
FIGURE 7 - RÉGULATION DE LA SÉCRÉTION D’INSULINE PAR LES ACIDES AMINÉS(AA) ET LES ACIDES GRAS (AG) DANS LES CELLULES B DU PANCRÉAS
AG
AG
Na+
ATP
CTP-1
AG-CoA
AA
AA
Insuline
K+
Ca2+
Ca2+
Ca2+
CaMK
Sécrétion d’insuline
Phosphorylationdes protéines

7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
189
Diab
ète
bablement du DS félin. Comme l’amyline, le GPL-1 diminue lalibération de glucagon. Cette action dépend du glucose car ceteffet existe en cas d’eu- ou d’hyperglycémie mais pas lors d’hypo-glycémie lorsque l’action du glucagon est alors nécessaire et impor-tante.
L’amyline pancréatique
Les cellules b du pancréas produisent également de l’amyline quiest co-synthétisée et co-sécrétée avec l’insuline en réponse à desstimuli appropriés (Lutz et Rand, 1996). Un déficit en amyline eten ses effets métaboliques pourrait jouer un rôle dans le dévelop-pement du DS2 de l’homme et du DS du chat. Ces effets sont indé-pendants de la tendance des amylines humaine et féline à formerdes dépôts d’amyloïde, un autre facteur important contribuant audéveloppement du DS félin (voir plus bas; O’Brien, 2002). L’amy-line produit au moins trois effets hormonaux qui contribuent à larégulation du métabolisme des nutriments :- inhibition de la prise de nourriture (Lutz, 2005)- modulation de la sécrétion du glucagon pancréatique en rédui-
sant une hyperglycémie postprandiale excessive (Edelman etWeyer, 2002)
- régulation de la vidange gastrique (Edelman et Weyer, 2002).
L’importance physiologique de ces effets est clairement démontréeà la fois chez l’homme et chez les rongeurs mais aucun n’est à cejour confirmé chez le chat. Une étude préliminaire réalisée sur deschats sains montre cependant que l’amyline pourrait diminuer letaux de glucagon circulant (Furrer et coll, 2005; voir également ci-dessous et Figure 16). Chez l’homme, l’analogue de l’amyline, lepramlintide (Symlin ND) est maintenant autorisé comme traite-ment adjuvant de l’insuline pour les patients diabétiques, en tantque réducteur de la sécrétion de glucagon et d’inhibiteur de lavidange gastrique.
7 -Physiopathologie du diabète sucré félin
Insulinorésistance et diabète sucré félin
Après la perturbation de la fonction des cellules b du pancréas, la deuxième caractéristique majeure duDS2 de l’homme et du DS félin est l’insulinorésistance. Cette sensibilité à l’insuline inférieure à la nor-male se caractérise par une réponse diminuée des tissus cibles à un taux fixe d’insuline. Chez les indi-vidus insulinorésistants, la capture insulinodépendante du glucose est réduite de manière importante.Si une sécrétion accrue d’insuline peut partiellement compenser l’insulinorésistance, une intoléranceau glucose ou une hyperglycémie franche se développent ultérieurement car l’hyperinsulinémie ne peutse prolonger ou conduit à l’épuisement des cellules b (Figure 8).
> Mise en évidence de l’insulinorésistance
Les tests classiques permettant de mesurer la sensibilité à l’insuline sont l’épreuve de l’hyperglycémieprovoquée par voie intraveineuse (HGPIV) (O’Brien et coll, 1985; Appleton et coll, 2001a,b) ou le testde sensibilité à l’insuline (TSI; Feldhahn et coll, 1999; Appleton et coll, 2001a,b). Le HGPIV mesure leglucose sanguin et la concentration d’insuline suite à une dose intraveineuse de glucose. Chez le chatnormal, la demi-vie du glucose plasmatique (glucose T1/2) ne doit pas dépasser environ 75-80 min(Lutz et Rand, 1996; Appleton et coll, 2001a,b). Le TSI mesure, lui, directement l’effet hypoglycémiantde l’insuline (Appleton et coll, 2001a,b).
FIGURE 8 - CERCLE VICIEUX DE L’INSULINORÉSISTANCE, DU DYSFONCTIONNEMENT DES CELLULES b
ET DE LA GLUCOTOXICITÉ, QUI CONDUIT PARFOISÀ L’ÉPUISEMENT DES CELLULES b ET AU DIABÈTE FRANC
L’insulinorésistance, parfois causée par l’obésité ou une prédispositiongénétique, et les lésions éventuelles des cellules b qui provoquent unediminution de la sécrétion d’insuline, conduisent à l’intolérance au glucose et l’hyperglycémie secondaire. Les cellules b restantes sont alors soumises à une pression de sécrétion augmentée. De plus, la glucotoxicité perturbe progressivement le fonctionnement des cellules b et renforce l’insulinorésistance. Finalement, les cellules pancréatiques ne parviennent plus à produire suffisamment d’insuline,d’où la déclaration du diabète sucré.
ObésitéPrédisposition génétique
Type d’alimentAutres maladies
HyperinsulinémieStress accru sur les cellules b restantes
Glucotoxicité
Insulinorésistance
Intolérance au glucoseHyperglycémie
Pathologie sous-jacente,
anomalie des cellules b

7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
Au cours de l’HGPIV, les chats diabétiques ou en phase “prédiabétique” présentent des concentrationsde glucose supérieures et une demi-vie prolongée du glucose. L’insulinémie à jeun est augmentée danscertaines études (Nelson et coll, 1990) mais pas dans d’autres (Lutz et Rand, 1996).
> Mécanismes de l’insulinorésistance
La diminution de la tolérance au glucose des chats diabétiques est secondaire à une réponse insuliniquediminuée (O’Brien et coll, 1985) à et une réduction de la sensibilité à l’insuline. Cette dernière est 6fois plus faible chez le chat diabétique que chez le chat normal (Feldhahn et coll, 1999). Les mécanismesprécis de l’insulinorésistance ne sont encore connus ni chez l’homme ni chez le chat (Reaven, 2005;Reusch et coll, 2006b) mais, comme chez l’homme, les causes principales de l’insulinorésistance félinesont l’obésité et la sédentarité. La sensibilité à l’insuline est beaucoup plus faible chez les chats obèsesque chez les chats normaux (voir plus bas).
> Facteurs favorisant l’insulinorésistance
Les causes génétiques du dysfonctionnement des récepteurs ou des post-récepteurs n’ont pas été étu-diées dans le détail chez le chat, mais des outils moléculaires sont maintenant disponibles et vont per-mettre d’approfondir la connaissance des mécanismes responsables de l’insulinorésistance périphérique.L’attention se porte surtout sur les transporteurs de glucose au sein des tissus sensibles à l’insuline et surles cytokines actives sécrétées par le tissu adipeux (Brennan et coll, 2004; Hoenig et coll, 2007a; Zini etcoll, 2006).
L’existence d’une différence de sensibilité à l’insuline entre mâles et femelles est moins claire. D’uncôté, les chats mâles montreraient une sensibilité à l’insuline plus faible et des concentrations basalesd’insuline plus élevées que les femelles (Appleton et coll, 2001a; Rand et Marshall, 2005). La dernièreétude a été réalisée chez des animaux maigres recevant un aliment relativement riche en glucides maistous les chats, mâles et femelles, étaient stérilisés au moment de l’étude. Il est donc improbable quecette différence de sensibilité à l’insuline puisse s’expliquer par un effet direct des hormones sexuelles.Ces différences pourraient cependant s’expliquer soit par un effet très précoce des hormones sexuellesavant la castration, soit par un effet indirect des hormones sexuelles.
D’un autre côté, l’obésité est le facteur de risque majeur d’insulinorésistance et la prise de poids aprèscastration survient plus rapidement chez les femelles que chez les mâles (Martin et Siliart, 2005). Cecicontraste quelque peu avec une étude réalisée par Hoenig et coll (2007b) qui montre que l’insuline
entraîne une augmentation de l’oxydation du glucose chez les mâles castrés alorsque l’oxydation des acides gras est plus importante chez les femelles stérili-
sées. Cette différence métabolique liée au sexe favoriserait donc uneaccumulation plus rapide de masse grasse chez les mâles que chez
les femelles, ce qui expliquerait le risque plus élevé de DS chezle mâle castré. Cependant, les mêmes auteurs ont égalementmontré que le statut sexuel n’est pas un facteur de risque à lui
seul en comparant les paramètres de la cinétique du glu-cose chez des chats maigres et obèses (Hoenig et coll,
2007a,b).
Les glucocorticoïdes et les progestagènes sontdes hormones directement antagonistes de l’in-suline et représentent d’autres causes possiblesd’insulinorésistance. De plus, au moins chezd’autres espèces, les glucocorticoïdes augmen-
tent la prise alimentaire et contribuent donc audéveloppement de l’obésité. L’hyperthyroïdie et
l’excès d’hormone de croissance (acromégalie)diminuent la tolérance au glucose, probablement
suite au développement d’une insulinorésistancepériphérique (Hoenig et Ferguson, 1989; Feldman et
Nelson, 2004).
© Y
ves
Lanc
eau/
RC
(C
hart
reux
)
190
D’autres études devront déterminer l’influence
du sexe sur la sensibilité à l’insuline et le développement
du diabète sucré félin.
Diab
ète
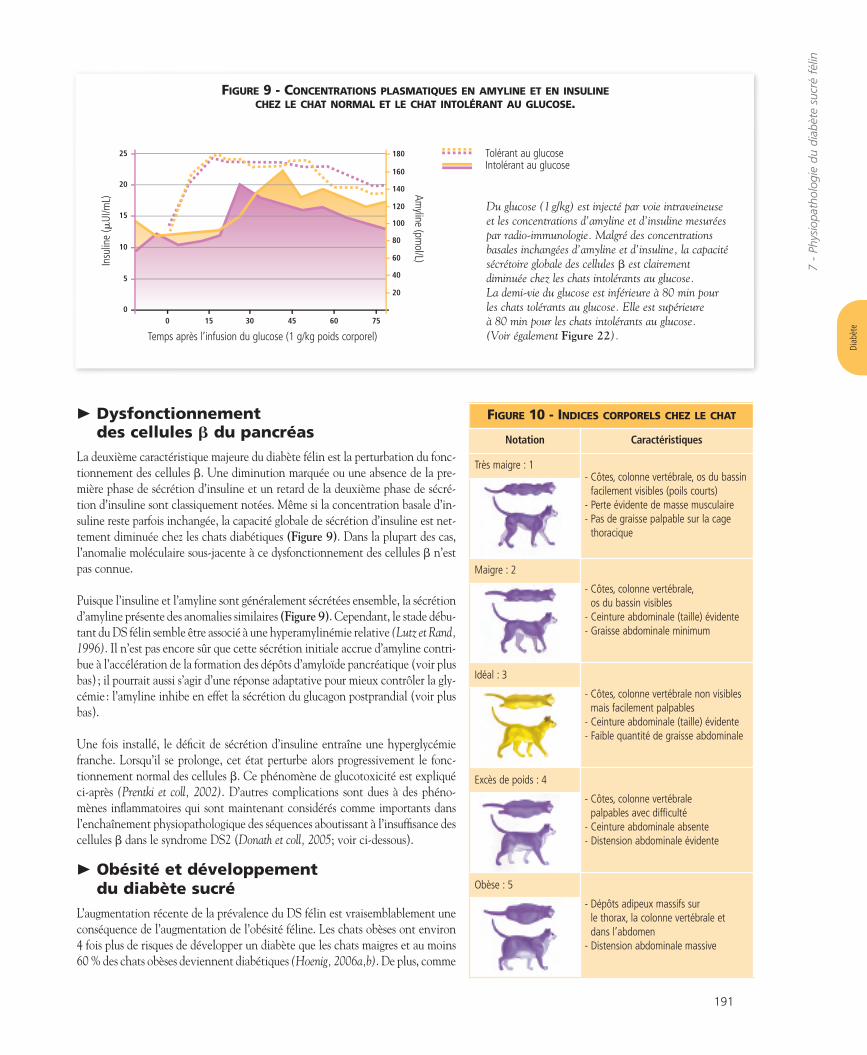
Dysfonctionnement des cellules b du pancréas
La deuxième caractéristique majeure du diabète félin est la perturbation du fonc-tionnement des cellules b. Une diminution marquée ou une absence de la pre-mière phase de sécrétion d’insuline et un retard de la deuxième phase de sécré-tion d’insuline sont classiquement notées. Même si la concentration basale d’in-suline reste parfois inchangée, la capacité globale de sécrétion d’insuline est net-tement diminuée chez les chats diabétiques (Figure 9). Dans la plupart des cas,l’anomalie moléculaire sous-jacente à ce dysfonctionnement des cellules b n’estpas connue.
Puisque l’insuline et l’amyline sont généralement sécrétées ensemble, la sécrétiond’amyline présente des anomalies similaires (Figure 9). Cependant, le stade débu-tant du DS félin semble être associé à une hyperamylinémie relative (Lutz et Rand,1996). Il n’est pas encore sûr que cette sécrétion initiale accrue d’amyline contri-bue à l’accélération de la formation des dépôts d’amyloïde pancréatique (voir plusbas) ; il pourrait aussi s’agir d’une réponse adaptative pour mieux contrôler la gly-cémie : l’amyline inhibe en effet la sécrétion du glucagon postprandial (voir plusbas).
Une fois installé, le déficit de sécrétion d’insuline entraîne une hyperglycémiefranche. Lorsqu’il se prolonge, cet état perturbe alors progressivement le fonc-tionnement normal des cellules b. Ce phénomène de glucotoxicité est expliquéci-après (Prentki et coll, 2002). D’autres complications sont dues à des phéno-mènes inflammatoires qui sont maintenant considérés comme importants dansl’enchaînement physiopathologique des séquences aboutissant à l’insuffisance descellules b dans le syndrome DS2 (Donath et coll, 2005; voir ci-dessous).
Obésité et développement du diabète sucré
L’augmentation récente de la prévalence du DS félin est vraisemblablement uneconséquence de l’augmentation de l’obésité féline. Les chats obèses ont environ4 fois plus de risques de développer un diabète que les chats maigres et au moins60 % des chats obèses deviennent diabétiques (Hoenig, 2006a,b). De plus, comme
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
FIGURE 9 - CONCENTRATIONS PLASMATIQUES EN AMYLINE ET EN INSULINECHEZ LE CHAT NORMAL ET LE CHAT INTOLÉRANT AU GLUCOSE.
Du glucose (1 g/kg) est injecté par voie intraveineuseet les concentrations d’amyline et d’insuline mesuréespar radio-immunologie. Malgré des concentrationsbasales inchangées d’amyline et d’insuline, la capacitésécrétoire globale des cellules b est clairement diminuée chez les chats intolérants au glucose. La demi-vie du glucose est inférieure à 80 min pour les chats tolérants au glucose. Elle est supérieure à 80 min pour les chats intolérants au glucose. (Voir également Figure 22).
FIGURE 10 - INDICES CORPORELS CHEZ LE CHAT
Notation Caractéristiques
Très maigre : 1- Côtes, colonne vertébrale, os du bassin
facilement visibles (poils courts)- Perte évidente de masse musculaire- Pas de graisse palpable sur la cage
thoracique
Maigre : 2
- Côtes, colonne vertébrale,os du bassin visibles
- Ceinture abdominale (taille) évidente- Graisse abdominale minimum
Idéal : 3
- Côtes, colonne vertébrale non visiblesmais facilement palpables
- Ceinture abdominale (taille) évidente- Faible quantité de graisse abdominale
Excès de poids : 4
- Côtes, colonne vertébrale palpables avec difficulté
- Ceinture abdominale absente- Distension abdominale évidente
Obèse : 5
- Dépôts adipeux massifs sur le thorax, la colonne vertébrale et dans l’abdomen
- Distension abdominale massive
0
5
20
40
60
80
100
120
140
160
180
0 15 30 45 60 75
10
15
20
25 Tolérant au glucoseIntolérant au glucose
Temps après l’infusion du glucose (1 g/kg poids corporel)
Insu
line
(mUI
/mL)
Amyline (pm
ol/L)
191
Diab
ète
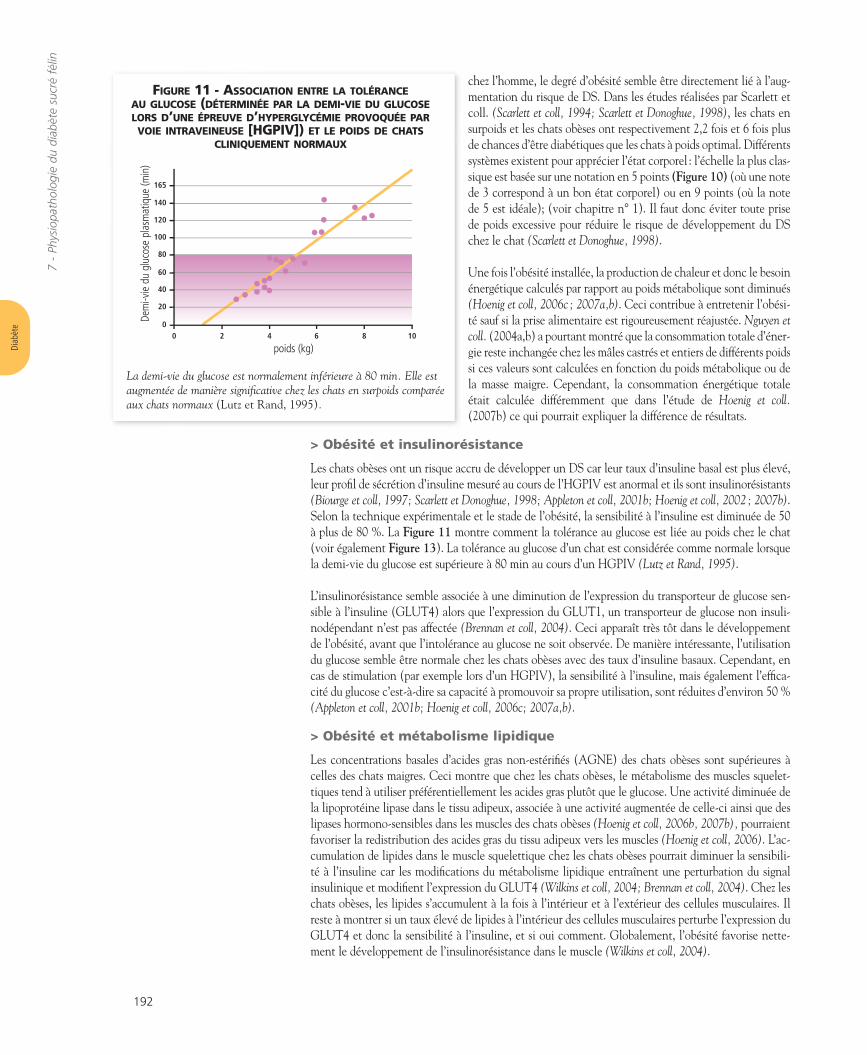
chez l’homme, le degré d’obésité semble être directement lié à l’aug-mentation du risque de DS. Dans les études réalisées par Scarlett etcoll. (Scarlett et coll, 1994; Scarlett et Donoghue, 1998), les chats ensurpoids et les chats obèses ont respectivement 2,2 fois et 6 fois plusde chances d’être diabétiques que les chats à poids optimal. Différentssystèmes existent pour apprécier l’état corporel : l’échelle la plus clas-sique est basée sur une notation en 5 points (Figure 10) (où une notede 3 correspond à un bon état corporel) ou en 9 points (où la notede 5 est idéale); (voir chapitre n° 1). Il faut donc éviter toute prisede poids excessive pour réduire le risque de développement du DSchez le chat (Scarlett et Donoghue, 1998).
Une fois l’obésité installée, la production de chaleur et donc le besoinénergétique calculés par rapport au poids métabolique sont diminués(Hoenig et coll, 2006c; 2007a,b). Ceci contribue à entretenir l’obési-té sauf si la prise alimentaire est rigoureusement réajustée. Nguyen etcoll. (2004a,b) a pourtant montré que la consommation totale d’éner-gie reste inchangée chez les mâles castrés et entiers de différents poidssi ces valeurs sont calculées en fonction du poids métabolique ou dela masse maigre. Cependant, la consommation énergétique totaleétait calculée différemment que dans l’étude de Hoenig et coll.(2007b) ce qui pourrait expliquer la différence de résultats.
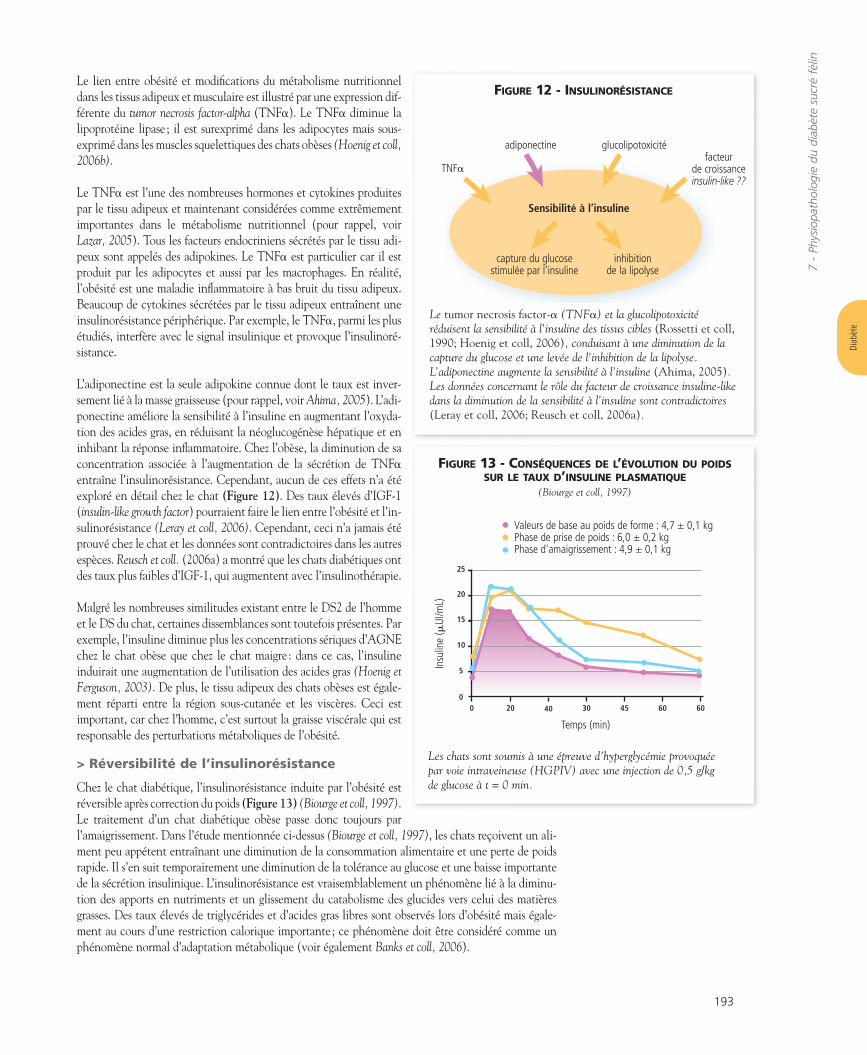
> Obésité et insulinorésistance
Les chats obèses ont un risque accru de développer un DS car leur taux d’insuline basal est plus élevé,leur profil de sécrétion d’insuline mesuré au cours de l’HGPIV est anormal et ils sont insulinorésistants(Biourge et coll, 1997; Scarlett et Donoghue, 1998; Appleton et coll, 2001b; Hoenig et coll, 2002; 2007b).Selon la technique expérimentale et le stade de l’obésité, la sensibilité à l’insuline est diminuée de 50à plus de 80 %. La Figure 11 montre comment la tolérance au glucose est liée au poids chez le chat(voir également Figure 13). La tolérance au glucose d’un chat est considérée comme normale lorsquela demi-vie du glucose est supérieure à 80 min au cours d’un HGPIV (Lutz et Rand, 1995).
L’insulinorésistance semble associée à une diminution de l’expression du transporteur de glucose sen-sible à l’insuline (GLUT4) alors que l’expression du GLUT1, un transporteur de glucose non insuli-nodépendant n’est pas affectée (Brennan et coll, 2004). Ceci apparaît très tôt dans le développementde l’obésité, avant que l’intolérance au glucose ne soit observée. De manière intéressante, l’utilisationdu glucose semble être normale chez les chats obèses avec des taux d’insuline basaux. Cependant, encas de stimulation (par exemple lors d’un HGPIV), la sensibilité à l’insuline, mais également l’effica-cité du glucose c’est-à-dire sa capacité à promouvoir sa propre utilisation, sont réduites d’environ 50 %(Appleton et coll, 2001b; Hoenig et coll, 2006c; 2007a,b).
> Obésité et métabolisme lipidique
Les concentrations basales d’acides gras non-estérifiés (AGNE) des chats obèses sont supérieures àcelles des chats maigres. Ceci montre que chez les chats obèses, le métabolisme des muscles squelet-tiques tend à utiliser préférentiellement les acides gras plutôt que le glucose. Une activité diminuée dela lipoprotéine lipase dans le tissu adipeux, associée à une activité augmentée de celle-ci ainsi que deslipases hormono-sensibles dans les muscles des chats obèses (Hoenig et coll, 2006b, 2007b), pourraientfavoriser la redistribution des acides gras du tissu adipeux vers les muscles (Hoenig et coll, 2006). L’ac-cumulation de lipides dans le muscle squelettique chez les chats obèses pourrait diminuer la sensibili-té à l’insuline car les modifications du métabolisme lipidique entraînent une perturbation du signalinsulinique et modifient l’expression du GLUT4 (Wilkins et coll, 2004; Brennan et coll, 2004). Chez leschats obèses, les lipides s’accumulent à la fois à l’intérieur et à l’extérieur des cellules musculaires. Ilreste à montrer si un taux élevé de lipides à l’intérieur des cellules musculaires perturbe l’expression duGLUT4 et donc la sensibilité à l’insuline, et si oui comment. Globalement, l’obésité favorise nette-ment le développement de l’insulinorésistance dans le muscle (Wilkins et coll, 2004).
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
192
Diab
ète 0
20
40
60
0 2 4 6 108
80
100
140
120
165
FIGURE 11 - ASSOCIATION ENTRE LA TOLÉRANCEAU GLUCOSE (DÉTERMINÉE PAR LA DEMI-VIE DU GLUCOSELORS D’UNE ÉPREUVE D’HYPERGLYCÉMIE PROVOQUÉE PARVOIE INTRAVEINEUSE [HGPIV]) ET LE POIDS DE CHATS
CLINIQUEMENT NORMAUX
La demi-vie du glucose est normalement inférieure à 80 min. Elle estaugmentée de manière significative chez les chats en surpoids comparéeaux chats normaux (Lutz et Rand, 1995).
poids (kg)
Dem
i-vie
du
gluc
ose
plas
mat
ique
(min
)
Le lien entre obésité et modifications du métabolisme nutritionneldans les tissus adipeux et musculaire est illustré par une expression dif-férente du tumor necrosis factor-alpha (TNFa). Le TNFa diminue lalipoprotéine lipase ; il est surexprimé dans les adipocytes mais sous-exprimé dans les muscles squelettiques des chats obèses (Hoenig et coll,2006b).
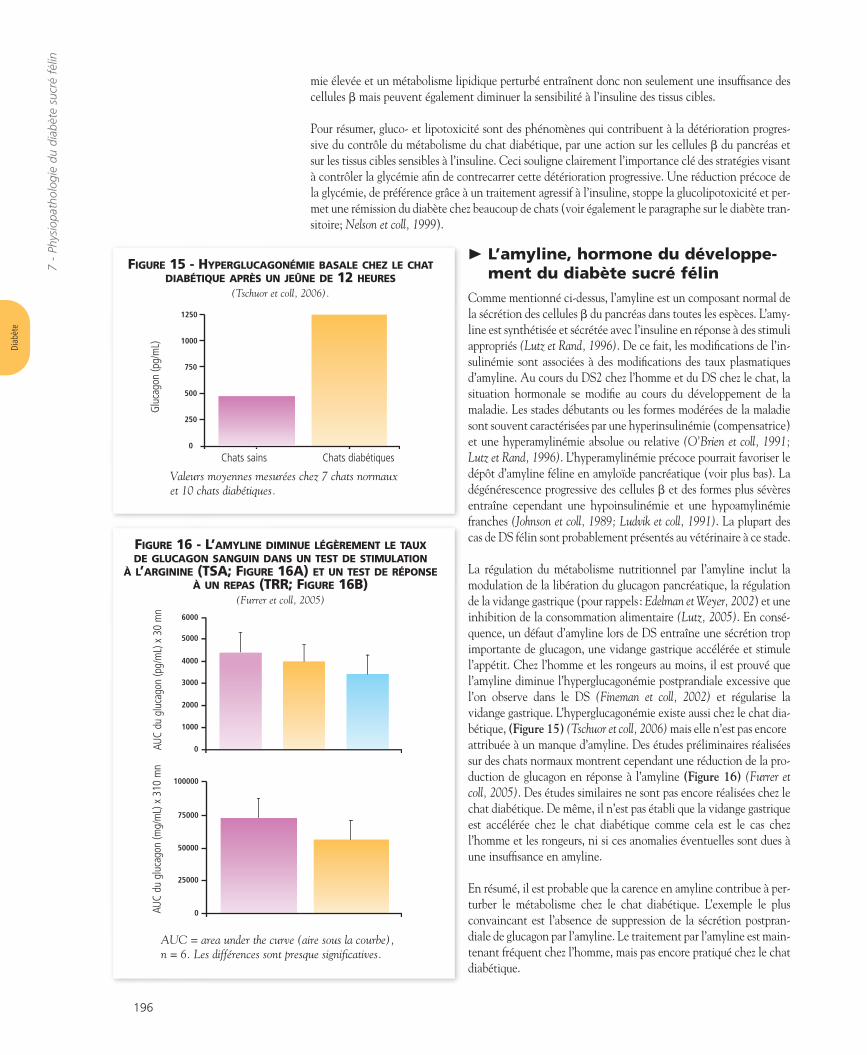
Le TNFa est l’une des nombreuses hormones et cytokines produitespar le tissu adipeux et maintenant considérées comme extrêmementimportantes dans le métabolisme nutritionnel (pour rappel, voirLazar, 2005). Tous les facteurs endocriniens sécrétés par le tissu adi-peux sont appelés des adipokines. Le TNFa est particulier car il estproduit par les adipocytes et aussi par les macrophages. En réalité,l’obésité est une maladie inflammatoire à bas bruit du tissu adipeux.Beaucoup de cytokines sécrétées par le tissu adipeux entraînent uneinsulinorésistance périphérique. Par exemple, le TNFa, parmi les plusétudiés, interfère avec le signal insulinique et provoque l’insulinoré-sistance.
L’adiponectine est la seule adipokine connue dont le taux est inver-sement lié à la masse graisseuse (pour rappel, voir Ahima, 2005). L’adi-ponectine améliore la sensibilité à l’insuline en augmentant l’oxyda-tion des acides gras, en réduisant la néoglucogénèse hépatique et eninhibant la réponse inflammatoire. Chez l’obèse, la diminution de saconcentration associée à l’augmentation de la sécrétion de TNFaentraîne l’insulinorésistance. Cependant, aucun de ces effets n’a étéexploré en détail chez le chat (Figure 12). Des taux élevés d’IGF-1(insulin-like growth factor) pourraient faire le lien entre l’obésité et l’in-sulinorésistance (Leray et coll, 2006). Cependant, ceci n’a jamais étéprouvé chez le chat et les données sont contradictoires dans les autresespèces. Reusch et coll. (2006a) a montré que les chats diabétiques ontdes taux plus faibles d’IGF-1, qui augmentent avec l’insulinothérapie.
Malgré les nombreuses similitudes existant entre le DS2 de l’hommeet le DS du chat, certaines dissemblances sont toutefois présentes. Parexemple, l’insuline diminue plus les concentrations sériques d’AGNEchez le chat obèse que chez le chat maigre : dans ce cas, l’insulineinduirait une augmentation de l’utilisation des acides gras (Hoenig etFerguson, 2003). De plus, le tissu adipeux des chats obèses est égale-ment réparti entre la région sous-cutanée et les viscères. Ceci estimportant, car chez l’homme, c’est surtout la graisse viscérale qui estresponsable des perturbations métaboliques de l’obésité.
> Réversibilité de l’insulinorésistance
Chez le chat diabétique, l’insulinorésistance induite par l’obésité estréversible après correction du poids (Figure 13) (Biourge et coll, 1997).Le traitement d’un chat diabétique obèse passe donc toujours parl’amaigrissement. Dans l’étude mentionnée ci-dessus (Biourge et coll, 1997), les chats reçoivent un ali-ment peu appétent entraînant une diminution de la consommation alimentaire et une perte de poidsrapide. Il s’en suit temporairement une diminution de la tolérance au glucose et une baisse importantede la sécrétion insulinique. L’insulinorésistance est vraisemblablement un phénomène lié à la diminu-tion des apports en nutriments et un glissement du catabolisme des glucides vers celui des matièresgrasses. Des taux élevés de triglycérides et d’acides gras libres sont observés lors d’obésité mais égale-ment au cours d’une restriction calorique importante; ce phénomène doit être considéré comme unphénomène normal d’adaptation métabolique (voir également Banks et coll, 2006).
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
193
Diab
ète
FIGURE 12 - INSULINORÉSISTANCE
0
5
200 40 30 45 60 60
10
15
20
25
FIGURE 13 - CONSÉQUENCES DE L’ÉVOLUTION DU POIDSSUR LE TAUX D’INSULINE PLASMATIQUE
(Biourge et coll, 1997)
Les chats sont soumis à une épreuve d'hyperglycémie provoquée par voie intraveineuse (HGPIV) avec une injection de 0,5 g/kg de glucose à t = 0 min.
Temps (min)
Valeurs de base au poids de forme : 4,7 ± 0,1 kgPhase de prise de poids : 6,0 ± 0,2 kgPhase d'amaigrissement : 4,9 ± 0,1 kg
Insu
line
(mUI
/mL)
adiponectine glucolipotoxicité
Sensibilité à l’insuline
TNFa
inhibition de la lipolyse
facteur de croissance insulin-like ??
capture du glucose stimulée par l’insuline
Le tumor necrosis factor-a (TNFa) et la glucolipotoxicité réduisent la sensibilité à l’insuline des tissus cibles (Rossetti et coll,1990; Hoenig et coll, 2006), conduisant à une diminution de lacapture du glucose et une levée de l’inhibition de la lipolyse.L’adiponectine augmente la sensibilité à l’insuline (Ahima, 2005).Les données concernant le rôle du facteur de croissance insuline-likedans la diminution de la sensibilité à l’insuline sont contradictoires(Leray et coll, 2006; Reusch et coll, 2006a).

Bien que la prise de poids des chats castrés soit observée depuis longtemps, les raisons n’en sont connuesque depuis quelques années. Chez les chats castrés, l’augmentation du poids corporel associée à la dimi-nution de la sensibilité à l’insuline est liée à la fois à l’augmentation de la prise alimentaire et à la dimi-nution des besoins énergétiques (Root et coll, 1996; Biourge et coll, 1997; Fettman et coll, 1997; Harperet coll, 2001; Hoenig et Ferguson, 2002; Kanchuk et coll, 2002; Kanchuk et coll, 2003). La diminutiondes besoins énergétiques est cependant controversée car elle n’est pas observée dans toutes les étudeschez les chats mâles (Kanchuk et coll, 2003). Des procédures différentes peuvent expliquer ces résultats :Kanchuk et coll (2003) exprime la dépense énergétique par rapport à la masse maigre, selon l’hypothè-se que la prise de poids chez les chats suralimentés résulte principalement de l’augmentation d’un tissuadipeux inactif du point de vue métabolique (Kanchuk et coll, 2003; voir également Martin et coll, 2001).Dans tous les cas, les chats castrés risquent plus de devenir obèses.
Notions générales de glucotoxicité, lipotoxicité et glucolipotoxicité
Le système de détection du glucose dans le pancréas semble être similaire dans l’espèce féline et dansd’autres espèces. Les Figures 6 et 7 montrent que le glucose et les acides gras libres (ou les AGNE) sti-mulent normalement la sécrétion d’insuline. Le glucose renforce également l’expansion normale de lamasse des cellules b. Ces deux mécanismes induits par le glucose (capture par le GLUT2 et proliféra-tion cellulaire) semblent être directement liés au travers de signaux intracellulaires distincts (rappelsdans Prentki et Nolan, 2006). L’action du glucose sur la prolifération des cellules b est renforcée par lesincrétines comme la GLP-1 et les acides gras libres. La GLP-1 protège donc les cellules b de l’apoptoseet stimule la croissance des cellules b.
Comme l’étude de Prentki et coll. (2002) le rappelle, des concentrations de glucose inférieures à 10mmol/L (180 mg/dL) ne sont normalement pas toxiques pour les cellules b du pancréas. L’hyperglycé-mie postprandiale physiologique entraîne en fait une prolifération des cellules b (Donath et coll, 2005).De la même manière, des concentrations physiologiques élevées et isolées d’acides gras ne sont pastoxiques, du moins lorsque le taux de malonyl-CoA, un produit secondaire du métabolisme du glucosequi inhibe la récupération des acides gras par les mitochondries en vue d’une b-oxydation, est faible.Les acides gras favorisent la sécrétion d’insuline en augmentant le Ca2+ et le diacylglycérol (Figure 7).Les problèmes ne surviennent que lorsque l’hyperglycémie et l’augmentation des acides gras se produi-sent en même temps et sur des périodes prolongées. Alors que la sécrétion d’insuline est initialementaccrue par le glucose et les acides gras-CoA à longue chaîne (Figures 6 et 7), elle baisse ensuite à causede l’élévation marquée du glucose, des acides gras activés et des molécules de signal lipidique qui déclen-chent également une apoptose. Ces effets sont respectivement appelés glucotoxicité et lipotoxicité. Leterme de glucolipotoxicité a été choisi car la lipotoxicité est plus marquée lors d’hyperglycémie sous-jacente (Prentki et Nolan, 2006).
Peu d’aspects de la gluco- et lipotoxicité ont été étudiés chez le chat. Il n’en reste pas moins que, àcause des nombreuses similitudes entre les modèles de DS2 de rongeurs et surtout le DS2 de l’hommeet le DS du chat (Henson et O’Brien, 2006), plusieurs points discutés dans le paragraphe suivant sontprobablement également vrais pour le chat.
La réduction de la masse des cellules b induite par l’hyperglycémie chronique et la glucotoxicité, résulted’un déséquilibre entre la néogénèse cellulaire, la prolifération et l’apoptose des cellules b (Donath etcoll, 2005). Au cours de la phase chronique d’hyperglycémie et d’hyperlipidémie, le glucose, les acidesgras saturés et les triglycérides s’accumulent dans les cellules b et induisent la libération de cytokines,entraînant la diminution de la sécrétion d’insuline et l’apoptose des cellules b. Au niveau cellulaire, laglucotoxicité est liée à un dysfonctionnement mitochondrial qui peut être responsable d’un stress oxy-datif des cellules b du pancréas, lié à l’augmentation du métabolisme oxydatif du glucose (Prentki etNolan, 2006). Les composés réactifs de l’oxygène peuvent être “détoxifiés” mais cela coûte en ATP etdiminue d’autant la sécrétion d’insuline (Figures 6 et 7).
Le dysfonctionnement du métabolisme lipidique, du cycle des triglycérides et des acides gras librescontribue également à l’insuffisance des cellules b. Une accumulation d’acides gras-CoA à longuechaîne a une influence directe sur les canaux K ATP-sensibles impliqués dans la sécrétion insulinique
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
194
Diab
ète
Le concept de glucotoxicité ou de glucolipotoxicité n’est pas nouveau(Rossetti et coll, 1990), mais les causeset les mécanismes sous-jacents sontbeaucoup mieux connus depuisquelques années. La glucotoxicité et lalipotoxicité sont induites par un troubledu couplage stimulus-sécrétion quientraîne irrémédiablement une insuffisance des cellules b. Les deuxphénomènes surviennent suffisammentrapidement pour que l’hyperglycémieprésente pendant seulement quelquesjours modifie à la baisse le système detransport du glucose et que l’élévationdes acides gras libres pendant 24 heures diminue la sécrétion d’insuline.
GLUCOTOXICITÉ ET LIPOTOXICITÉ

après stimulation par le glucose. De plus, des taux élevés de malonyl-CoA intracellulaires diminuentla capture d’acides gras par les mitochondries et induisent donc un détournement du métabolisme del’oxydation des acides gras vers leur estérification et l’accumulation de lipides. La production intra-cellulaire d’ATP, importante pour le couplage stimulus-sécrétion, est de ce fait diminuée (Prentki etNolan, 2006).
Ces dernières années, les preuves d’un lien direct entre glucotoxicité, lipotoxicité et inflammation desîlots ont également été rassemblées. L’interleukine 1-b (IL-1b) est considérée comme l’une des molé-cules clés (Donath et coll, 2005). Bien qu’une régulation à la hausse de l’IL-1b ait maintenant étédécrite chez plusieurs modèles animaux de DS2, d’autres études sont attendues sur le lien entre hyper-glycémie et inflammation (Prentki et Nolan, 2006). À notre connaissance, de telles études n’ont pasencore été réalisées chez le chat.
Gluco- et lipotoxicité chez le chat
En 1948, Dohan et Lukens, dans «Diabète expérimental produit par l’administration de glucose»,décrivent les effets d’une hyperglycémie persistante sur les îlots de Langerhans, notamment la dégra-nulation des cellules b suivie d’une dégénération des îlots. Plusieurs chats développent un diabète sucréidentifié à l’époque par une glucosurie importante.
> Glucotoxicité
La glucotoxicité contribue clairement à l’insuffisance des cellules b chez le chat mais elle est réversiblesi l’hyperglycémie disparaît. Si elle est maintenue, la perte des cellules b peut devenir permanente.Chez le chat sain, une hyperglycémie persistante de 30 mmol/L environ (540 mg/dL), induite par uneperfusion continue de glucose, abolit presque totalement la sécrétion d’insuline en 3 à 7 jours. L’histo-logie du pancréas révèle des changements importants dans la morphologie des cellules b: vacuolisa-tion, dépôts de glycogène, perte de coloration à l’insuline et pyknose. Cependant, même ces modifica-tions profondes sont réversibles après l’arrêt de l’hyperglycémie (Rand et Marshall, 2005). Des donnéesnon publiées de l’auteur montrent aussi clairement qu’une hyperglycémie d’environ 25 mmol/L(450 mg/dL), pendant seulement 10 jours, suffit à diminuer considérablement la capacité de sécrétiond’insuline par les cellules b du pancréas chez des chats sains.
> Lipotoxicité
La lipotoxicité n’a pas été étudiée en détail chez le chat.Cependant, Hoenig (2002) a émis l’hypothèse d’un rôlepathogène possible de la lipotoxicité chez le chat diabé-tique. Comme cela est décrit dans le cycle glucose/acide gras(cycle de Randle; Randle, 1998), le glucose inhibe l’oxyda-tion des acides gras et vice versa (Figure 14). Puisque lesconcentrations d’AGNE sont élevées chez le chat obèse etqu’il est prédisposé au diabète sucré, il est probable que lesAGNE abaissent le métabolisme du glucose dans les cellulesb. Cependant, le métabolisme du glucose est nécessaire à lasécrétion d’insuline, qui serait donc réduite. Une étude réa-lisée par la même équipe montre que chez le chat les acidesgras saturés sont préjudiciables au contrôle de la glycémiealors que les acides gras polyinsaturés oméga-3 (AGPI n-3)auraient des effets bénéfiques (Wilkins et coll, 2004).
Bien que moins étudiés, des mécanismes semblables à ceuxdéjà décrits pour les cellules b du pancréas semblent égale-ment jouer un rôle dans la glucolipotoxicité des tissus ciblesde l’insuline. Comme cité précédemment, l’accumulationde lipides intramyocellulaires diminue la sensibilité des cel-lules musculaires à l’insuline (Wilkins et coll, 2004; voir éga-lement Hoenig, 2002). Chez le chat diabétique, une glycé-
L’apport de glucose stimule l’oxydation du glucose, le stockage du glucose etdes lipides et inhibe l’oxydation des acides gras (A). Cette dernière comprometl’oxydation du glucose (B) et peut renforcer le stockage du glucose sous formede glycogène si les réserves de glycogène sont faibles.
FIGURE 14 - CONCEPT SIMPLIFIÉ DU CYCLE GLUCOSE/ACIDES GRAS(cycle de Randle ; Randle, 1998).
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
195
Diab
ète
© Y
. La
ncea
u/R
C
A BOxydation du glucose Lipolyse
Oxydation des acides gras
Acétyl-CoA NADH
Activation de pyruvate déshydrogénase (PDH) kinase
Inhibition de l’oxydation du glucose
Pyruvate
Citrate
Malonyl-CoA
Inhibition de l'entrée des radicauxacyl-CoA dans les mitochondries
Inhibition de l’oxydation des acides gras
Le premier article traitant de la glucotoxicité chez le chat date de 1948.
mie élevée et un métabolisme lipidique perturbé entraînent donc non seulement une insuffisance descellules b mais peuvent également diminuer la sensibilité à l’insuline des tissus cibles.
Pour résumer, gluco- et lipotoxicité sont des phénomènes qui contribuent à la détérioration progres-sive du contrôle du métabolisme du chat diabétique, par une action sur les cellules b du pancréas etsur les tissus cibles sensibles à l’insuline. Ceci souligne clairement l’importance clé des stratégies visantà contrôler la glycémie afin de contrecarrer cette détérioration progressive. Une réduction précoce dela glycémie, de préférence grâce à un traitement agressif à l’insuline, stoppe la glucolipotoxicité et per-met une rémission du diabète chez beaucoup de chats (voir également le paragraphe sur le diabète tran-sitoire; Nelson et coll, 1999).
L’amyline, hormone du développe-ment du diabète sucré félin
Comme mentionné ci-dessus, l’amyline est un composant normal dela sécrétion des cellules b du pancréas dans toutes les espèces. L’amy-line est synthétisée et sécrétée avec l’insuline en réponse à des stimuliappropriés (Lutz et Rand, 1996). De ce fait, les modifications de l’in-sulinémie sont associées à des modifications des taux plasmatiquesd’amyline. Au cours du DS2 chez l’homme et du DS chez le chat, lasituation hormonale se modifie au cours du développement de lamaladie. Les stades débutants ou les formes modérées de la maladiesont souvent caractérisées par une hyperinsulinémie (compensatrice)et une hyperamylinémie absolue ou relative (O’Brien et coll, 1991;Lutz et Rand, 1996). L’hyperamylinémie précoce pourrait favoriser ledépôt d’amyline féline en amyloïde pancréatique (voir plus bas). Ladégénérescence progressive des cellules b et des formes plus sévèresentraîne cependant une hypoinsulinémie et une hypoamylinémiefranches (Johnson et coll, 1989; Ludvik et coll, 1991). La plupart descas de DS félin sont probablement présentés au vétérinaire à ce stade.
La régulation du métabolisme nutritionnel par l’amyline inclut lamodulation de la libération du glucagon pancréatique, la régulationde la vidange gastrique (pour rappels : Edelman et Weyer, 2002) et uneinhibition de la consommation alimentaire (Lutz, 2005). En consé-quence, un défaut d’amyline lors de DS entraîne une sécrétion tropimportante de glucagon, une vidange gastrique accélérée et stimulel’appétit. Chez l’homme et les rongeurs au moins, il est prouvé quel’amyline diminue l’hyperglucagonémie postprandiale excessive quel’on observe dans le DS (Fineman et coll, 2002) et régularise lavidange gastrique. L’hyperglucagonémie existe aussi chez le chat dia-bétique, (Figure 15) (Tschuor et coll, 2006) mais elle n’est pas encoreattribuée à un manque d’amyline. Des études préliminaires réaliséessur des chats normaux montrent cependant une réduction de la pro-duction de glucagon en réponse à l’amyline (Figure 16) (Furrer etcoll, 2005). Des études similaires ne sont pas encore réalisées chez lechat diabétique. De même, il n’est pas établi que la vidange gastriqueest accélérée chez le chat diabétique comme cela est le cas chezl’homme et les rongeurs, ni si ces anomalies éventuelles sont dues àune insuffisance en amyline.
En résumé, il est probable que la carence en amyline contribue à per-turber le métabolisme chez le chat diabétique. L’exemple le plusconvaincant est l’absence de suppression de la sécrétion postpran-diale de glucagon par l’amyline. Le traitement par l’amyline est main-tenant fréquent chez l’homme, mais pas encore pratiqué chez le chatdiabétique.
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
196
FIGURE 15 - HYPERGLUCAGONÉMIE BASALE CHEZ LE CHATDIABÉTIQUE APRÈS UN JEÛNE DE 12 HEURES
(Tschuor et coll, 2006).
Valeurs moyennes mesurées chez 7 chats normaux et 10 chats diabétiques.
0
500
750
250
1000
1250
0
1000
2000
3000
4000
6000
5000
0
50000
75000
25000
100000
Diab
ète
AUC = area under the curve (aire sous la courbe), n = 6. Les différences sont presque significatives.
AUC
du g
luca
gon
(mg/
mL)
x 3
10 m
nAU
C du
glu
cago
n (p
g/m
L) x
30
mn
FIGURE 16 - L’AMYLINE DIMINUE LÉGÈREMENT LE TAUXDE GLUCAGON SANGUIN DANS UN TEST DE STIMULATION
À L’ARGININE (TSA; FIGURE 16A) ET UN TEST DE RÉPONSEÀ UN REPAS (TRR; FIGURE 16B)
(Furrer et coll, 2005)
Chats sains Chats diabétiques
Glu
cago
n (p
g/m
L)
Le glucagon pancréatique, une hormone du développement du diabète sucré félin
Le glucagon pancréatique a été négligé en tant que facteur pathogène du développement du DS à causede la priorité absolue donnée à la carence en insuline. Néanmoins, une inhibition insuffisante de lasécrétion de glucagon, particulièrement en période postprandiale immédiate, semble être un facteurmajeur de l’hyperglycémie postprandiale (Figure 15) (O’Brien et coll, 1985; Furrer et coll, 2005; Tschuoret coll, 2006). L’hyperglucagonémie diabétique semble être directement liée à l’insuffisance en amylineet par conséquence à la désinhibition de la sécrétion du glucagon. Ceci pourrait également être vraichez le chat (Figure 16) (Furrer et coll, 2005); il reste alors à déterminer à quel point.
Amyloïdose pancréatique
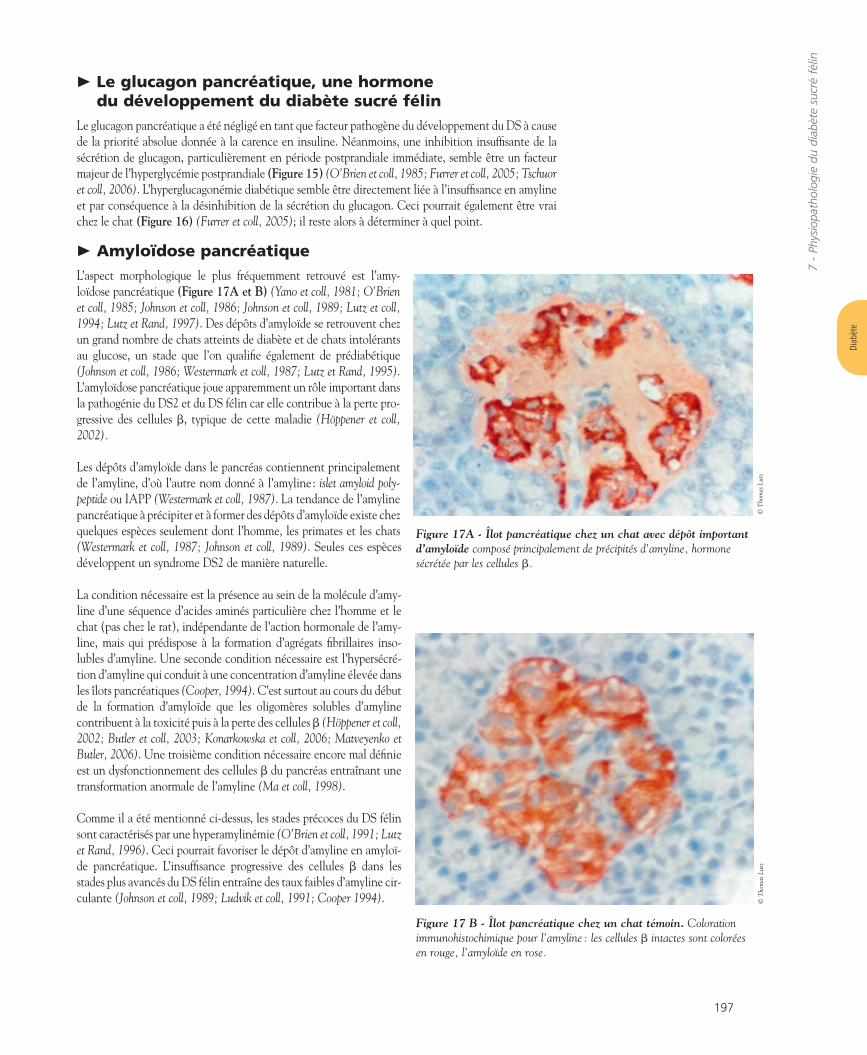
L’aspect morphologique le plus fréquemment retrouvé est l’amy-loïdose pancréatique (Figure 17A et B) (Yano et coll, 1981; O’Brienet coll, 1985; Johnson et coll, 1986; Johnson et coll, 1989; Lutz et coll,1994; Lutz et Rand, 1997). Des dépôts d’amyloïde se retrouvent chezun grand nombre de chats atteints de diabète et de chats intolérantsau glucose, un stade que l’on qualifie également de prédiabétique(Johnson et coll, 1986; Westermark et coll, 1987; Lutz et Rand, 1995).L’amyloïdose pancréatique joue apparemment un rôle important dansla pathogénie du DS2 et du DS félin car elle contribue à la perte pro-gressive des cellules b, typique de cette maladie (Höppener et coll,2002).
Les dépôts d’amyloïde dans le pancréas contiennent principalementde l’amyline, d’où l’autre nom donné à l’amyline : islet amyloid poly-peptide ou IAPP (Westermark et coll, 1987). La tendance de l’amylinepancréatique à précipiter et à former des dépôts d’amyloïde existe chezquelques espèces seulement dont l’homme, les primates et les chats(Westermark et coll, 1987; Johnson et coll, 1989). Seules ces espècesdéveloppent un syndrome DS2 de manière naturelle.
La condition nécessaire est la présence au sein de la molécule d’amy-line d’une séquence d’acides aminés particulière chez l’homme et lechat (pas chez le rat), indépendante de l’action hormonale de l’amy-line, mais qui prédispose à la formation d’agrégats fibrillaires inso-lubles d’amyline. Une seconde condition nécessaire est l’hypersécré-tion d’amyline qui conduit à une concentration d’amyline élevée dansles îlots pancréatiques (Cooper, 1994). C’est surtout au cours du débutde la formation d’amyloïde que les oligomères solubles d’amylinecontribuent à la toxicité puis à la perte des cellules b (Höppener et coll,2002; Butler et coll, 2003; Konarkowska et coll, 2006; Matveyenko etButler, 2006). Une troisième condition nécessaire encore mal définieest un dysfonctionnement des cellules b du pancréas entraînant unetransformation anormale de l’amyline (Ma et coll, 1998).
Comme il a été mentionné ci-dessus, les stades précoces du DS félinsont caractérisés par une hyperamylinémie (O’Brien et coll, 1991; Lutzet Rand, 1996). Ceci pourrait favoriser le dépôt d’amyline en amyloï-de pancréatique. L’insuffisance progressive des cellules b dans lesstades plus avancés du DS félin entraîne des taux faibles d’amyline cir-culante (Johnson et coll, 1989; Ludvik et coll, 1991; Cooper 1994).
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
197
Diab
ète
Figure 17A - Îlot pancréatique chez un chat avec dépôt importantd’amyloïde composé principalement de précipités d’amyline, hormonesécrétée par les cellules b.
Figure 17 B - Îlot pancréatique chez un chat témoin. Colorationimmunohistochimique pour l’amyline : les cellules b intactes sont coloréesen rouge, l’amyloïde en rose.
© T
hom
as L
utz
© T
hom
as L
utz

Importance des dépôts d’amyloïde dans le pancréas du chat
Le dépôt d’amyloïde dans les îlots du pancréas est la caractéristique his-tologique principale du diabète félin mais il est intéressant de noterqu’il se retrouve également chez des chats non diabétiques. Certainsproduisent même une quantité importante d’amyloïde sans pour autantmanifester de signes cliniques (Figure 18) (Lutz et coll, 1994). La pré-valence de l’amyloïde pancréatique augmente avec l’âge (Figure 19),tout comme la prévalence du diabète félin. Il faut cependant consta-ter que les chats diabétiques forment des dépôts d’amyloïde bien plusimportants que les chats normaux et que l’étendue des dépôts estdirectement corrélée à la sévérité des signes cliniques du DS félin(O’Brien et coll, 1985; Johnson et coll, 1989; Lutz et coll, 1994). L’as-sociation entre la quantité d’amyloïde pancréatique et l’intoléranceau glucose est retrouvée lors de l’évaluation de la demi-vie du glucoseplasmatique au cours d’un test HGPIV (Figure 20).
Malheureusement, bien que l’amyloïde pancréatique joue un rôleimportant dans la physiopathologie du DS félin, ce n’est pas confir-mé in vivo. Elle ne constitue donc pas aujourd’hui un marqueur utilepour le pronostic de cette maladie.
D’une manière générale, les études sur des rongeurs transgéniquesmontrent clairement que l’amyloïde dérivée de l’amyline joue un rôleimportant dans le développement et la progression du DS2. Les oli-gomères solubles d’amyline de faible poids moléculaire dans lesespèces possédant une séquence d’acides aminées amyloïdogéniquessont responsables de l’apoptose des cellules b (pour rappels : voir Muffet coll, 2004). Néanmoins, les événements primaires à l’origine de laformation de ces oligomères toxiques dans le DS2 restent à élucider.
Lien entre hyperglycémie et amyloïdose pancréatique
Actuellement, un lien entre les principaux facteurs entraînant l’in-suffisance des cellules b (gluco-lipotoxicité et amyloïde dérivée del’amyline) n’a pas été mis en évidence chez le chat. Cependant, il sepeut que des modifications du milieu intracellulaire induites par untaux de glucose ou d’acides gras élevé (stress intracellulaire) créentdes conditions favorisant la formation et la précipitation des fibrillesd’amyloïde. La forme la plus toxique pour les cellules b sont lesfibrilles d’amyline composées d’oligomères de faible poids moléculai-
re, probablement formées au cours du stade précoce de la maladie. Toute thérapie visant à contrôler laglycémie, et donc à réduire la sollicitation excessive des cellules b du pancréas le plus tôt possible dansle développement de la maladie, peut donc favoriser la rémission du diabète observée dans les DS tran-sitoires (voir plus bas).
Diminution de la sensibilité à l’insuline chez les chats malades
Comme chez l‘homme, l’homéostasie glucidique est souvent altérée chez les chats atteints de maladiesdiverses : inflammation sévère, néoplasie, septicémie, infection virale, stade terminal de maladie rénaleet insuffisance cardiaque chronique. Une association entre l’augmentation de la synthèse des cytokinespro-inflammatoires et la présence d’hormones contre régulant l’insuline serait une cause sous-jacente.Ceci a été vérifié chez les chats atteints d’insuffisance cardiaque congestive avec des taux élevés deTNFa (Meurs et coll, 2002).
7 -
Phys
iopa
thol
ogie
du
diab
ète
sucr
é fé
lin
198
FIGURE 19 - LES DÉPÔTS D’AMYLOÏDEAUGMENTENT AVEC L’ÂGE
(Lutz et coll, 1994)
0
5
10
0 5 10 15
15
20
25
30
Âge (années)
% d
u vo
lum
e d’
amylo
ïde
par
rapp
ort
au v
olum
e de
s îlo
ts p
ancr
éatiq
ues
Les jeunes chats cliniquement normaux ont peu ou pas du tout de dépôt d’amyloïde pancréatique détectable.
Diab
ète
00 5 10 20 30 40 50 60 70 80 90 100
10
20
30
40
% volume de l’amyloïde
Nom
bre
de c
hats
(%)
Certains chats montrent d’importants dépôts d’amyloïdesans pour autant développer des signes cliniques de diabètesucré (Lutz et coll, 1994). Le pourcentage du volumed’amyloïde est mesuré par rapport au volume total des îlots (=100 %).
FIGURE 18 - FRÉQUENCE DE DÉPÔTS D’AMYLOÏDECHEZ 84 CHATS NORMAUX

De plus, stomatites, lésions pulmonaires (Mexas et coll, 2006) et infec-tions urinaires (Jin et Lin, 2005) semblent être plus fréquentes chez lechat diabétique. Les chats gravement malades ont une hyperglycémiede stress marquée mais ils ne présentent pas toujours d’hyperinsuliné-mie concomitante, témoin d’une insulinorésistance (Chan et coll,2006).
Les mécanismes exacts associant une perturbation de l’homéostasie duglucose et les diverses maladies ne sont pas connus chez le chat maisferaient intervenir plusieurs cytokines. Une étude préliminairerécente a montré que la perfusion pendant 10 jours d’un lipopolysac-charide, constituant de la membrane cellulaire d’une bactérie Gramnégatif et à l’origine de la libération de diverses cytokines, conduit àune moins bonne tolérance au glucose (non publié). Ces troublesseraient également associés à des taux réduits d’adiponectine, l’hor-mone du tissu adipeux qui semble être un facteur important dans larégulation de la sensibilité à l’insuline des tissus cibles (Hoenig et coll,2007a). En plus d’un effet sur les tissus sensibles à l’insuline, diversescytokines pourraient directement diminuer la sécrétion pancréatiqueendocrine.
Enfin, il faut admettre que nous sommes face au problème de l’œuf etde la poule. D’un côté, hyperglycémie et DS abaissent les défensesimmunitaires, par exemple dans la sphère urogénitale (e.g., Lederer etcoll, 2003; Bailiff et coll, 2006). De l’autre côté, infections et inflam-mations sont associées à une insulinorésistance, peut-être par le biaisdu TNFa, qui conduit irrémédiablement au DS (Figure 21).
8 -Diabète transitoireLe DS transitoire est relativement fréquent chez le chat diabétique.Environ 20 % des chats diabétiques rentrent d’habitude dans cettecatégorie (Nelson et coll, 1999; Nelson, 2005). Cependant, la propor-tion de chats atteints de diabète transitoire a augmenté récemment(voir plus bas). Le diabète transitoire se caractérise par une rémissionspontanée, c’est-à-dire que les signes cliniques comme la polyurie etla polydipsie disparaissent, la glycémie se normalise et la glucosuriedisparaît également. Ceci survient en général 1 à 4 mois après le débutdu traitement (Nelson et coll, 1999). Le traitement antidiabétique peutalors être arrêté. Une fois le DS guéri, la sécrétion d’insuline induitepar le glucose se normalise. Néanmoins, la densité des cellules b esttoujours diminuée et la pathologie des îlots pancréatiques toujours pré-sente. La plupart de ces cas constituent en fait des stades subcliniquesde DS (Nelson et coll, 1999).
Conditions nécessaires pour observer une rémission du diabète félin
Les conditions nécessaires à la rémission du diabète ne sont pas encore complètement connues. Ilsemble évident qu’un certain nombre de cellules b fonctionnelles doit encore exister (Nelson et coll,1999). La résolution précoce de l’hyperglycémie et donc la réduction, voire la disparition de la gluco-toxicité serait un facteur important. Un contrôle strict de la glycémie, associé à un régime approprié(voir plus bas) peut rompre le cercle vicieux de l’hyperglycémie, du dysfonctionnement des cellules bdu pancréas et de la diminution de la sensibilité à l’insuline. Puisque la glucotoxicité est réversible dansun premier temps, il est permis de penser que plus la glycémie est maîtrisée tôt, plus les chances derémission sont importantes. Cependant, il manque encore des données solides confirmant cette hypo-thèse.
8 -
Dia
bète
tra
nsito
ire
FIGURE 21 - AUTO-ENTRETIEN DU DIABÈTE
FIGURE 20 - LA QUANTITÉ D’AMYLOÏDE PANCRÉATIQUEEST POSITIVEMENT CORRÉLÉE À LA DEMI-VIE DU GLUCOSE
(GLUCOSE T1/2) MESURÉE PAR UN HGPIV(Lutz et coll, 1994)
0
10
20
30
0 20 40 60 80 100 120 140
40
50
60
70
80
Volu
me
de Is
let a
mylo
id (%
)
Glucose T1/2 dans le plasma (min)
199
Diab
ète
Diabète sucré
HyperglycémieInsulinorésistance
Baisse des défensesimmunitaires
Infections et inflammations (sphère
urogénitale)

Différence entre diabète permanent et diabète transitoire?
Il est difficile de prédire le caractère transitoire d’un diabète chez le chat par l’épreu-ve de l’hyperglycémie provoquée par voie intraveineuse (HGPIV) et par le test
de stimulation par le glucagon. Une étude prospective a évalué la prédic-tibilité de la rémission du diabète chez des chats par un test de stimu-
lation à l’arginine (TSA; Tschuor et coll, 2006). Ce test est utiliséavec succès chez l’homme avec DS2. La glycémie de base est signi-
ficativement plus élevée et la réponse à l’insuline significative-ment abaissée chez le chat diabétique comparé au chat nor-
mal. Le glucagon basal et la réponse du glucagon à l’argini-ne sont significativement plus élevés chez le chat diabé-
tique. Malgré ces différences évidentes entre chat diabé-tique et chat normal, aucune différence significativen’est retrouvée entre diabète transitoire et diabète per-manent pour aucun des paramètres mesurés (glucose,insuline, glucagon). Le TSA n’est donc pas un testcapable de différencier le diabète transitoire du dia-bète permanent chez le chat de manière prospective(Tschuor et coll, 2006); (voir plus bas et Figure 25).
L’hypothèse selon laquelle les niveaux d’IGF-1 pour-raient aider à prévoir un diabète sucré transitoire chez le
chat a récemment été explorée mais cette idée a cependantété écartée (Alt et coll, 2007).
Dans les cas de rémission chez les chats diabétiques, une rechute clinique est toujours possible. La patho-logie des îlots du pancréas existe généralement chez les chats atteints de diabète transitoire. La proba-bilité de déclarer un diabète franc est donc certainement plus élevée que chez les chats auparavantsains. Les médicaments antagonistes de l’insuline (glucocorticoïdes, acétate de mégestrol) ou l’obésitépourraient constituer des facteurs de stress favorisant la récidive. Il est en général impossible de prédiresi et quand les signes cliniques vont revenir et il faut donc surveiller les chats en rémission de très près.Le diabète de certains chats évoluerait d’un stade subclinique à clinique en 3 ans (Nelson et coll, 1999).
Évolution du taux de rémission diabétique
La proportion de chats manifestant un diabète transitoire semble avoir augmenté, atteignant parfois70 % des cas dans certaines études, ce qui pourrait être lié aux recommandations diététiques récentesd’utiliser un régime riche en protéines et pauvre en glucides. Il reste à savoir si l’amélioration consta-tée est liée à la haute teneur protéique (49-57 % sur MS dans les études de Frank et coll, 2001; Maz-zaferro et coll, 2003), à la faible teneur glucidique (18 % dans l’étude par Bennett et coll, 2006), ou auxdeux (voir également plus bas). Nous avons également confirmé que le taux de rémission des chats dia-bétiques est plus élevé qu’avant s’ils sont nourris avec une alimentation riche en protéines (environ54 % de protéines, 8 % de glucides sur MS; Tschuor et coll, 2006). Dans notre étude, une rémission aété constatée chez environ 50 % des chats traités à l’insuline en 4 semaines. De manière étonnante, larémission a précédé la perte significative de poids.
9 -Conséquences à long terme de l’hyperglycémie diabétique
L’hyperglycémie chronique a des effets délétères sur les cellules b du pancréas qui produisent l’insuline etsur les tissus cibles de l’insuline (voir ci-dessus, glucotoxicité). Mais l’hyperglycémie prolongée apparaîtaussi comme la cause majeure des complications fréquentes du diabète félin, que sont les neuropathie,néphropathie et rétinopathie diabétiques. Ces affections reposent sur deux mécanismes sous-jacents: uneglycolysation des protéines et une perturbation osmotique liée à l’accumulation des polyols.
9 -
Con
séqu
ence
s à
long
ter
me
de l’
hype
rgly
cém
ie d
iabé
tique
200
Diab
ète
La mesure de la glucosurie avec unebandelette urinaire, dans une litièrerécemment souillée par un mélanged’eau et d’urine, permet de surveillerfacilement l’évolution d’un diabètesucré transitoire vers un stade subclinique ou clinique.
© Yves Lanceau/Royal Canin - Sacré de Birmanie

Glycolysation des protéines et accumulation des polyols
Une des modifications précoces du DS est une augmentation non spécifique de la glycosylation nonenzymatique des protéines, qui entraîne une agrégation anormale des fibrilles de collagène, la produc-tion de radicaux superoxyde, et un endommagement du tissu conjonctif et des membranes basales. L’ac-cumulation du sorbitol, non perméable aux membranes cellulaires, est responsable de cette perturba-tion osmotique cellulaire. L’aldose réductase transforme le glucose en sorbitol. Dans des conditions nor-males, le sorbitol est présent en petites quantités mais l’hyperglycémie entraîne parfois une accumula-tion importante de sorbitol lorsque la voie d’utilisation du glucose par l’hexokinase est saturée.
Neuropathie, rétinopathie et cataracte diabétiques
La prévalence exacte des neuropathie, néphropathie et rétinopathie diabétiques est inconnue chez lechat. La neuropathie diabétique se traduit par une faiblesse des postérieurs et une plantigradie typique(Figure 3). Cette affection semble présenter beaucoup de similitudes avec la neuropathie diabétiquechez l’homme (Misizin et coll, 2007).
Si le contrôle de la glycémie est mis en place de manière intensive et rapide après le diagnostic, plu-sieurs de ces anomalies disparaissent et les aplombs redeviennent normaux. Même si la néphropathieet la rétinopathie diabétiques existent aussi chez le chat, elles sont rarement observées en pratique.Expérimentalement, une hyperglycémie induite pendant plusieurs années peut provoquer des modifi-cations de la rétine, mais leur mise en évidence nécessite des techniques particulières (communicationpersonnelle du Dr M. Richter, Unité d’Ophtalmologie, Faculté VetSuisse de l’Université de Zurich).
De même, et contrairement au chien, les cataractes diabétiques sont aussi très rares chez le chat (Figure 22). La formation de sorbitol chez les vieux chats diabétiques serait beaucoup plus faible quechez les chiens et les jeunes chats car l’activité de l’aldose réductase est plus faible chez les chats âgés(Richter et coll, 2002). L’excès de sorbitol est responsable de lésions rétiniennes. Bien que le DS soit trèsrare chez les jeunes chats, ceux-ci présentent souvent une opacité cristallinienne comme les chiens dia-bétiques, probablement à cause de l’activité importante de l’aldose réductase (Richter et coll, 2002). Uneétude a cependant contredit cette idée commune de faible prévalence de la cataracte diabétique chezle chat, en montrant que l’opacité cristallinienne se retrouve beaucoup plus fréquemment que ce quel’on pensait auparavant (Williams et Heath, 2006). De plus, elle survient plus tôt chez le chat diabé-tique que chez le chat non diabétique.
10 -Diagnostic du diabète sucré félinLe diagnostic du DS devrait toujours inclure l’évaluation des signes cliniques clés des formes non com-pliquées de diabète, c’est-à-dire polyurie, polydipsie, polyphagie, prise de poids. La présence d’un ouplusieurs de ces signes cliniques n’est pas pour autant pathognomonique. Il faut compléter cet examenpar des analyses de laboratoire.
10 -
Dia
gnos
tic d
u di
abèt
e su
cré
félin
Figure 22 - Cataracte chez un chat diabétique.
Ave
c l’a
imab
le a
utor
isatio
n du
Pro
f. B
. Sp
iess
, V
etsu
isse-
Facu
lty U
nive
rsity
of Z
uric
h.
201
Diab
ète

Hyperglycémie à jeun
L’hyperglycémie à jeun est une des clés du diabète du chatmais ce signe pris isolément n’est pas fiable à cause duphénomène d’hyperglycémie de stress (Figure 23), plusfréquente chez le chat que chez le chien. La glycémie d’unchat stressé dépasse parfois 20 mmol/L (360 mg/dL) (Lalu-ha et coll, 2004). Il faut donc savoir repérer une simplehyperglycémie de stress avant d’instaurer une thérapie àl’insuline (voir plus bas). De la même manière, la mesurede la glucosurie peut induire en erreur. Bien qu’elle soitnormalement présente chez le chat diabétique et absentechez le chat normal, l’hyperglycémie de stress peut êtresuffisamment importante pour entraîner une fuite de glu-cose dans les urines.
Insulinémie
Les chats diabétiques ne sont pas capables de sécréter suf-fisamment d’insuline pour maintenir leur glycémie dansles limites usuelles. Cette insuffisance est cependant àrelativiser puisque l’insulinémie peut paraître normale,même si elle est en réalité faible au regard de la glycémie.La mesure de l’insuline à jeun est donc en général inutileà moins que l’hypoinsulinémie absolue ne soit très impor-tante. De plus, ce test coûte cher et sa mesure spécifiquechez le chat reste d’accès limité.
La proinsuline ou le ratio insuline/proinsuline pourraientrespectivement être utiles au diagnostic du DS chez lechat. Chez l’homme, des taux à jeun élevés de proinsu-line traduisent des lésions des cellules b et la proinsulinepourrait servir de marqueur précoce du dysfonctionne-ment cellulaire. La séquence d’acides aminés de la proin-suline féline a été publiée récemment. Il est donc possibleque des tests deviennent disponibles pour permettre lediagnostic précoce du DS félin (Hoenig et coll, 2006a). Deplus, la sécrétion de proinsuline semble élevée chez leschats obèses.
Fructosamine et hémoglobine glycosylées
Comme il a été mentionné précédemment, ni la mesure du glucose à jeun, ni celle du glucose urinairene sont des marqueurs fiables du DS félin. La fructosamine et l’hémoglobine glycosylées sont mainte-nant les deux marqueurs fréquemment utilisés pour l’évaluation au long cours de la glycémie dans lediagnostic et le suivi du DS félin (Tableaux 3 et 4). Ce sont des produits de la fixation non spécifique,irréversible et non-enzymatique du glucose à des résidus d’acides aminés.
- La fructosamine est la somme des protéines sériques glycosylées ; elle est mesurée par des tests colo-rimétriques.
- L’hémoglobine glycosylée, et surtout la fraction A1c de l’hémoglobine glycosylée (HbA1c), est unproduit de la glycosylation de l’hémoglobine et du glucose mesuré par chromatographie ; l’hémoglo-bline glycosylée est un marqueur diagnostique rarement utilisé chez le chat.
Le taux de fructosamine et d’hémoglobine glycosylée sont directement proportionnels à la concentra-tion moyenne de glucose sanguin au cours du temps. Les deux dépendent également du taux de renou-vellement moyen de ces deux protéines, plus court pour les protéines sériques que pour l’hémoglobine.La concentration sérique de fructosamine reflète donc l’hyperglycémie moyenne des 10-14 derniers
10 -
Dia
gnos
tic d
u di
abèt
e su
cré
félin
0
40
10
160 240 400 560 720 880320 480 800640
30
20
Fructosamine sérique (µmol/L)
Glu
cose
(µm
ol/L
)
FIGURE 23 - CONCENTRATIONS SÉRIQUES DE FRUCTOSAMINECHEZ UN CHAT NORMOGLYCÉMIQUE ET LORS D’HYPERGLYCÉMIE
D’ORIGINE DIABÉTIQUE OU D’HYPERGLYCÉMIE DE STRESS(D’après Prof. C. Reusch, Vetsuisse-Faculté Université de Zurich)
TABLEAU 3 - COMPARAISON DE LA FRUCTOSAMINEET DE L’HÉMOGLOBINE GLYCOSYLÉE POUR L’ÉVALUATION
D’UNE HYPERGLYCÉMIE PERSISTANTE
Caractéristiquescommunes
Fructosamine Hémoglobine glycosylée
- Proviennent de la fixation irréversible, non-enzymatique et non-spécifique du glucose à des résidus d’acides aminés.
- Sont directement proportionnelles à la glycémie moyenne au cours du temps.
- Dépendent du taux de renouvellement moyen de leur protéine respective, plus rapide pour les protéines sériques que pour l’hémoglobine.
Caractéristiquespropres
- La fructosamine se réfère à la somme des protéines sériques glycolysées qui peut être mesurée par des tests colorimétriques.
- Marqueur de la glycémie moyenne des 10-14 derniers jours.
- Modifiée par des changements detaux de protéines sériques.
- L’hémoglobine glycosylée est un produit de la glycosylationde l’hémoglobine et du glucose.Elle est mesurée par chromatographie.
- Marqueur du taux moyen de glucose sanguin des 4-8 dernières semaines.
- Modifiée par la concentrationd’hémoglobine.
Groupe témoinHyperglycémie de stressChats diabétiquesLimite supérieure des valeursusuelles (365 µmol/L)
202
Diab
ète
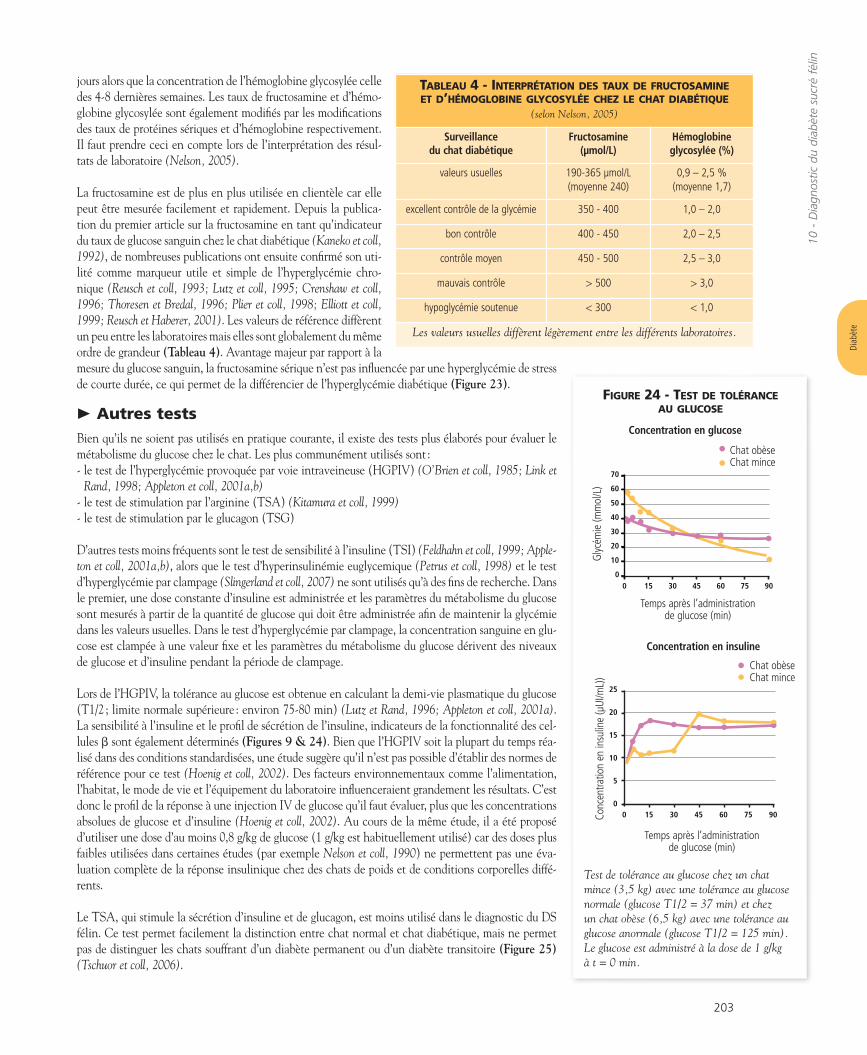
jours alors que la concentration de l’hémoglobine glycosylée celledes 4-8 dernières semaines. Les taux de fructosamine et d’hémo-globine glycosylée sont également modifiés par les modificationsdes taux de protéines sériques et d’hémoglobine respectivement.Il faut prendre ceci en compte lors de l’interprétation des résul-tats de laboratoire (Nelson, 2005).
La fructosamine est de plus en plus utilisée en clientèle car ellepeut être mesurée facilement et rapidement. Depuis la publica-tion du premier article sur la fructosamine en tant qu’indicateurdu taux de glucose sanguin chez le chat diabétique (Kaneko et coll,1992), de nombreuses publications ont ensuite confirmé son uti-lité comme marqueur utile et simple de l’hyperglycémie chro-nique (Reusch et coll, 1993; Lutz et coll, 1995; Crenshaw et coll,1996; Thoresen et Bredal, 1996; Plier et coll, 1998; Elliott et coll,1999; Reusch et Haberer, 2001). Les valeurs de référence diffèrentun peu entre les laboratoires mais elles sont globalement du mêmeordre de grandeur (Tableau 4). Avantage majeur par rapport à lamesure du glucose sanguin, la fructosamine sérique n’est pas influencée par une hyperglycémie de stressde courte durée, ce qui permet de la différencier de l’hyperglycémie diabétique (Figure 23).
Autres tests
Bien qu’ils ne soient pas utilisés en pratique courante, il existe des tests plus élaborés pour évaluer lemétabolisme du glucose chez le chat. Les plus communément utilisés sont :- le test de l’hyperglycémie provoquée par voie intraveineuse (HGPIV) (O’Brien et coll, 1985; Link et
Rand, 1998; Appleton et coll, 2001a,b)- le test de stimulation par l’arginine (TSA) (Kitamura et coll, 1999)- le test de stimulation par le glucagon (TSG)
D’autres tests moins fréquents sont le test de sensibilité à l’insuline (TSI) (Feldhahn et coll, 1999; Apple-ton et coll, 2001a,b), alors que le test d’hyperinsulinémie euglycemique (Petrus et coll, 1998) et le testd’hyperglycémie par clampage (Slingerland et coll, 2007) ne sont utilisés qu’à des fins de recherche. Dansle premier, une dose constante d’insuline est administrée et les paramètres du métabolisme du glucosesont mesurés à partir de la quantité de glucose qui doit être administrée afin de maintenir la glycémiedans les valeurs usuelles. Dans le test d’hyperglycémie par clampage, la concentration sanguine en glu-cose est clampée à une valeur fixe et les paramètres du métabolisme du glucose dérivent des niveauxde glucose et d’insuline pendant la période de clampage.
Lors de l’HGPIV, la tolérance au glucose est obtenue en calculant la demi-vie plasmatique du glucose(T1/2 ; limite normale supérieure : environ 75-80 min) (Lutz et Rand, 1996; Appleton et coll, 2001a).La sensibilité à l’insuline et le profil de sécrétion de l’insuline, indicateurs de la fonctionnalité des cel-lules b sont également déterminés (Figures 9 & 24). Bien que l’HGPIV soit la plupart du temps réa-lisé dans des conditions standardisées, une étude suggère qu’il n’est pas possible d’établir des normes deréférence pour ce test (Hoenig et coll, 2002). Des facteurs environnementaux comme l’alimentation,l’habitat, le mode de vie et l’équipement du laboratoire influenceraient grandement les résultats. C’estdonc le profil de la réponse à une injection IV de glucose qu’il faut évaluer, plus que les concentrationsabsolues de glucose et d’insuline (Hoenig et coll, 2002). Au cours de la même étude, il a été proposéd’utiliser une dose d’au moins 0,8 g/kg de glucose (1 g/kg est habituellement utilisé) car des doses plusfaibles utilisées dans certaines études (par exemple Nelson et coll, 1990) ne permettent pas une éva-luation complète de la réponse insulinique chez des chats de poids et de conditions corporelles diffé-rents.
Le TSA, qui stimule la sécrétion d’insuline et de glucagon, est moins utilisé dans le diagnostic du DSfélin. Ce test permet facilement la distinction entre chat normal et chat diabétique, mais ne permetpas de distinguer les chats souffrant d’un diabète permanent ou d’un diabète transitoire (Figure 25)(Tschuor et coll, 2006).
10 -
Dia
gnos
tic d
u di
abèt
e su
cré
félin
203
FIGURE 24 - TEST DE TOLÉRANCEAU GLUCOSE
TABLEAU 4 - INTERPRÉTATION DES TAUX DE FRUCTOSAMINEET D’HÉMOGLOBINE GLYCOSYLÉE CHEZ LE CHAT DIABÉTIQUE
(selon Nelson, 2005)
Surveillance du chat diabétique
Fructosamine(µmol/L)
Hémoglobine glycosylée (%)
valeurs usuelles 190-365 µmol/L(moyenne 240)
0,9 – 2,5 %(moyenne 1,7)
excellent contrôle de la glycémie 350 - 400 1,0 – 2,0
bon contrôle 400 - 450 2,0 – 2,5
contrôle moyen 450 - 500 2,5 – 3,0
mauvais contrôle > 500 > 3,0
hypoglycémie soutenue < 300 < 1,0
Les valeurs usuelles diffèrent légèrement entre les différents laboratoires.
Diab
ète
0
10
150 30 45 60 75 90
20
30
40
50
60
70
00 15 30 45 60 75 90
5
15
10
20
25
Chat obèseChat mince
Chat obèseChat mince
Temps après l’administration de glucose (min)
Conc
entra
tion
en in
sulin
e (µ
UI/m
L))
Glyc
émie
(mm
ol/L
)
Test de tolérance au glucose chez un chatmince (3,5 kg) avec une tolérance au glucosenormale (glucose T1/2 = 37 min) et chez un chat obèse (6,5 kg) avec une tolérance auglucose anormale (glucose T1/2 = 125 min).Le glucose est administré à la dose de 1 g/kg à t = 0 min.
Concentration en glucose
Concentration en insuline
Temps après l’administration de glucose (min)

11 -
Str
atég
ies
thér
apeu
tique
s
204
00 2 74 9 15 25 30
5
15
10
20
25
30
00 2 74 9 15 25 30
5
15
10
20
25
30
FIGURE 25 - TEST DE STIMULATION À L’ARGININE
Cependant, ce test ne permet pas de différencier un diabète transitoire d’un diabète permanent (Tschuor et coll, 2006).
Dans un test de stimulation à l’arginine (injection d’arginine à t = 0 min; 0,2 g/kg), les concentrations de glucose sanguin sont significativement plus faibles chez les chats normaux que chez les chats diabétiques.
11 -Stratégies thérapeutiquesLes points clés du traitement du diabète félin
Au-delà de la simple disparition des signes cliniques (polyurie, polydipsie), traditionnellement consi-dérée comme suffisante, il est aussi possible d’obtenir une rémission spontanée du DS félin, c’est-à-direune transition vers une forme subclinique de DS. La rémission du DS est principalement due à la dis-parition de la glucotoxicité une fois l'hyperglycémie contrôlée. L’objectif clé du traitement du chat dia-bétique est de ramener la glycémie entre 5 et 15 mmol/L (90-270 mg/dL).
Un autre élément clé réside dans le moment où la glycémie commence à diminuer, qui doit intervenirle plus tôt possible, car les modifications induites par la glucotoxicité dans les îlots pancréatiques sontau départ réversibles, mais deviennent ensuite irréversibles (Prentki et Nolan, 2006). Ceci n’est pas una-nimement démontré, mais l’auteur pense qu’un traitement précoce conduit à un pourcentage plus élevéde rémission chez les chats diabétiques.
Il faut donc traiter tôt et de manière intense. De nos jours, le traitement inclut un aspect diététique,et tout particulièrement l’utilisation d’aliments riches en protéines (> 50 % sur MS), et pauvres en glu-cides (< 15 % sur MS).
L’insuline en tant qu’agent hypoglycémiant
L’insulinothérapie est de loin la méthode la plus efficace pour obtenir un bon contrôle de la glycémiechez le chat diabétique. L’insuline féline n’est pas disponible mais l’insuline d’origine animale (bovineou porcine), l’insuline recombinante humaine et un analogue de synthèse de l’insuline humaine sontutilisés pour le traitement des chats diabétiques (Goossens et coll, 1998; Marshall et Rand, 2002; Wea-ver et coll, 2006). Les différentes formes d’insuline utilisées sont résumées dans le Tableau 5.
Les protocoles de traitement médical du chat diabétique sont largement décrits, voir Nelson (2005).L’insuline est normalement injectée par voie sous-cutanée, excepté lors du traitement d’une crise dia-bétique aiguë (diabète acidocétosique) où l’insuline peut être administrée par voie intramusculaire ouveineuse. La plupart des chats diabétiques ont besoin de deux injections quotidiennes car la durée d’ac-tion des préparations d’insuline est courte dans cette espèce comparée à l’homme.
Diab
ète
Glu
cose
(mm
ol/L
)
Chats en bonne santé Chats diabétiques Différence significative (p < 0,05)Arginine 0,2 g/kg
Diabète félin transitoireDiabète félin permanent Différence significative (p < 0,05)Arginine 0,2 g/kg
Glu
cose
(mm
ol/L
)
Temps (min) Temps (min)
p = 0,548p = 0,22
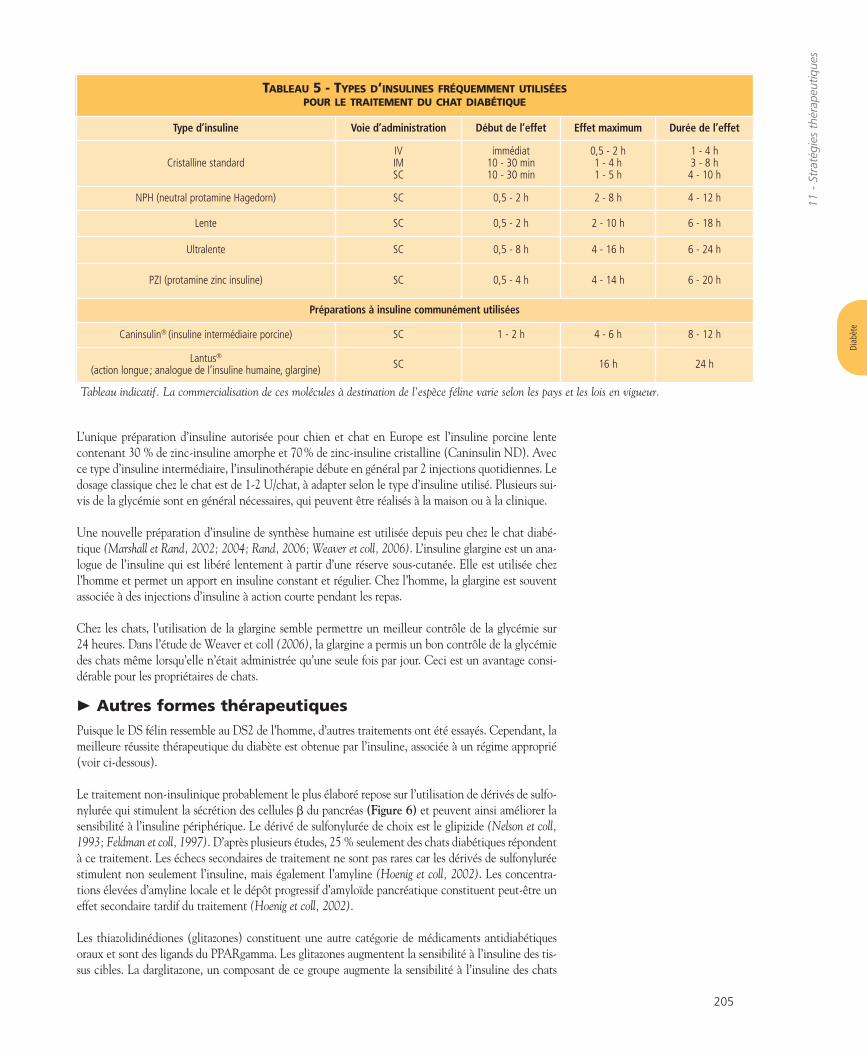
L’unique préparation d’insuline autorisée pour chien et chat en Europe est l’insuline porcine lentecontenant 30 % de zinc-insuline amorphe et 70% de zinc-insuline cristalline (Caninsulin ND). Avecce type d’insuline intermédiaire, l’insulinothérapie débute en général par 2 injections quotidiennes. Ledosage classique chez le chat est de 1-2 U/chat, à adapter selon le type d’insuline utilisé. Plusieurs sui-vis de la glycémie sont en général nécessaires, qui peuvent être réalisés à la maison ou à la clinique.
Une nouvelle préparation d’insuline de synthèse humaine est utilisée depuis peu chez le chat diabé-tique (Marshall et Rand, 2002; 2004; Rand, 2006; Weaver et coll, 2006). L’insuline glargine est un ana-logue de l’insuline qui est libéré lentement à partir d’une réserve sous-cutanée. Elle est utilisée chezl’homme et permet un apport en insuline constant et régulier. Chez l’homme, la glargine est souventassociée à des injections d’insuline à action courte pendant les repas.
Chez les chats, l’utilisation de la glargine semble permettre un meilleur contrôle de la glycémie sur24 heures. Dans l’étude de Weaver et coll (2006), la glargine a permis un bon contrôle de la glycémiedes chats même lorsqu’elle n’était administrée qu’une seule fois par jour. Ceci est un avantage consi-dérable pour les propriétaires de chats.
Autres formes thérapeutiques
Puisque le DS félin ressemble au DS2 de l’homme, d’autres traitements ont été essayés. Cependant, lameilleure réussite thérapeutique du diabète est obtenue par l’insuline, associée à un régime approprié(voir ci-dessous).
Le traitement non-insulinique probablement le plus élaboré repose sur l’utilisation de dérivés de sulfo-nylurée qui stimulent la sécrétion des cellules b du pancréas (Figure 6) et peuvent ainsi améliorer lasensibilité à l’insuline périphérique. Le dérivé de sulfonylurée de choix est le glipizide (Nelson et coll,1993; Feldman et coll, 1997). D’après plusieurs études, 25 % seulement des chats diabétiques répondentà ce traitement. Les échecs secondaires de traitement ne sont pas rares car les dérivés de sulfonyluréestimulent non seulement l’insuline, mais également l’amyline (Hoenig et coll, 2002). Les concentra-tions élevées d’amyline locale et le dépôt progressif d’amyloïde pancréatique constituent peut-être uneffet secondaire tardif du traitement (Hoenig et coll, 2002).
Les thiazolidinédiones (glitazones) constituent une autre catégorie de médicaments antidiabétiquesoraux et sont des ligands du PPARgamma. Les glitazones augmentent la sensibilité à l’insuline des tis-sus cibles. La darglitazone, un composant de ce groupe augmente la sensibilité à l’insuline des chats
11 -
Str
atég
ies
thér
apeu
tique
s
205
TABLEAU 5 - TYPES D’INSULINES FRÉQUEMMENT UTILISÉESPOUR LE TRAITEMENT DU CHAT DIABÉTIQUE
Type d’insuline Voie d’administration Début de l’effet Effet maximum Durée de l’effet
Cristalline standardIVIMSC
immédiat10 - 30 min10 - 30 min
0,5 - 2 h1 - 4 h1 - 5 h
1 - 4 h3 - 8 h4 - 10 h
NPH (neutral protamine Hagedorn) SC 0,5 - 2 h 2 - 8 h 4 - 12 h
Lente SC 0,5 - 2 h 2 - 10 h 6 - 18 h
Ultralente SC 0,5 - 8 h 4 - 16 h 6 - 24 h
PZI (protamine zinc insuline) SC 0,5 - 4 h 4 - 14 h 6 - 20 h
Préparations à insuline communément utilisées
Caninsulin® (insuline intermédiaire porcine) SC 1 - 2 h 4 - 6 h 8 - 12 h
Lantus®
(action longue; analogue de l’insuline humaine, glargine) SC 16 h 24 h
Tableau indicatif. La commercialisation de ces molécules à destination de l’espèce féline varie selon les pays et les lois en vigueur.
Diab
ète

obèses (Hoenig et Ferguson, 2003). L’utilité de ces médicaments en routine dans le traitement du DSfélin reste cependant à démontrer.
La metformine améliore la sensibilité à l’insuline principalement par inhibition de la néoglucogénèsehépatique et la glycogénolyse. Bien que la metformine puisse avoir des effets métaboliques intéressantschez le chat diabétique, son utilisation comme traitement de routine est largement controversée. Peudes chats traités se sont améliorés. La metformine n’apporte a priori pas d’avantage par rapport au trai-tement conventionnel (Nelson et coll, 2004).
L’hyperglycémie postprandiale est une caractéristique clé du DS. Diminuer l’absorption intestinale post-prandiale du glucose apparaît donc comme une alternative logique à la thérapeutique traditionnelle.L’acarbose, un inhibiteur compétitif de l’amylase pancréatique et des glucosidases de la membrane intes-tinale a été proposé (Nelson, 2005). Bien que l’acarbose puisse ralentir l’absorption gastrointestinaledu glucose, la prescription d’un régime riche en protéines s’avère bien plus efficace.
Options thérapeutiques futures
Les effets métaboliques bénéfiques de l’amyline et du GLP-1 ont été décrits précédemment : inhibitionde la vidange gastrique et de la libération postprandiale du glucagon (pour l’amyline, voir Figure 16).Tous ces effets n’ont pas encore été étudiés chez le chat. Le pramlintide (Symlin®), un analogue del’amyline qui est associé à l’insuline, et l’exendine-4, un agoniste du GLP-1 (Byetta®) sont maintenantutilisés pour le traitement du diabète humain. Aucune de ces molécules n’a encore été utilisée chez lechat et il n’est pas sûr qu’elles offrent des avantages considérables par rapport aux traitements actuels.
Les composés chimiques qui activent la glucokinase ont suscité de l’intérêt pour le traitement du dia-bète (Schermerhorn, 2006). Leur utilité en clinique est peu probable dans un avenir proche.
12 -Aspects diététiques du traitement du diabète sucré félin
Il se peut que l’aliment idéal du chat diabétique n’existe pas encore. Cependant, la diététique du dia-bète félin a largement évolué ces dernières années. Le contrôle de la glycémie a été
surtout facilité par l’introduction et la recommandation de régimes riches enprotéines (> 45 % des calories) et pauvres en glucides (< 20 % des calo-
ries).
Il peut sembler évident de donner au chat un régime riche en protéines,proche de son régime naturel. Néanmoins, la reconnaissance de cette uti-lité a révolutionné le traitement diététique du diabète qui reposait aupa-ravant sur la prescription d’un régime riche en glucides digestibles (sur-tout de l’amidon) (> 30 % des calories) et en fibres (> 50 g de fibres
totales (FT)/ 1000 kcal). Ces recommandations avaient probablement étéextrapolées de celles faites pour l’homme et le chien diabétiques, mais ellesne sont plus d’actualité, surtout en ce qui concerne l’amidon.
Objectifs généraux de la thérapie nutritionnelle du chat diabétique
(voir également: Biourge, 2005)Le DS félin étant une maladie similaire au DS2 de l’homme, un des axes
principaux du traitement et de la prévention réside dans le maintien d’unpoids corporel idéal. Comme il est décrit ci-dessous, les aliments richesen protéines sont d’une utilité particulière pour l’alimentation des chatsdiabétiques. Cependant, leur efficacité est encore meilleure lorsque cetype de régime est associé à une thérapie de réduction de la glycémie. Àcet effet, le traitement par l’insuline est le plus efficace, qui permet le
contrôle de la glucotoxicité (voir ci-dessus). Les meilleurs résultats sont
12 -
Asp
ects
dié
tétiq
ues
du t
raite
men
t du
dia
bète
suc
ré f
élin
© Y
ves
Lanc
eau/
RC
(Si
amoi
s)
L’un des axes principaux du traitement et de la prévention
du DS félin réside dans le maintiend’un poids corporel idéal.
206
Diab
ète

obtenus avec des injections biquotidiennes d’insuline. Sans insulinothérapie (ou d’autres traitementsvisant à diminuer la glycémie), les chances de traiter avec succès le chat diabétique sont très faibles,au moins au cours de la phase initiale du traitement. L’association entre un traitement insulinique etun régime adéquat renforce la probabilité de rémission du diabète et peut permettre ensuite d’inter-rompre l’administration d’insuline. Afin de bien contrôler le métabolisme et d’éviter le risque d’hy-poglycémie secondaire, la programmation des injections et la composition des repas doivent être lesplus régulières possible.
Les trois buts principaux de la gestion nutritionnelle du chat diabétique sont :1. Contrôler l’excès de poids2. Réduire l’hyperglycémie postprandiale3. Stimuler la sécrétion endogène d’insuline.
Prévenir ou corriger l’obésité
L’obésité est directement liée à l’insulinorésistance qui prédispose les chats au diabète (Scarlett et coll,1994; Scarlett et Donoghue, 1998). La prévention de l’obésité doit donc constituer un des objectifs prin-cipaux de l’alimentation des chats.
Les vétérinaires doivent conseiller aux propriétaires de restreindre l’apport calorique des chats immé-diatement après la castration, en utilisant des aliments à faible densité énergétique, c’est-à-dire conte-nant une quantité limitée de matières grasses. Les aliments secs riches en matières grasses (> 40 % descalories) sont associés à une prise de poids et au développement de l’obésité dans de nombreuses études,particulièrement chez les chats castrés (Scarlett et coll, 1994; Scarlett et Donoghue, 1998). En revanche,un aliment modérément gras (25 % des calories) et au contenu modéré en amidon (35 % des calories)permet de réduire la prise de poids consécutive à la castration, comparé à un aliment sec riche enmatières grasses (> 40 % des calories) (Nguyen et coll, 2004a).
La perte de poids est favorisée par un aliment riche en protéines (45 % sur MS; 25 % glucides sur MS)plus que par un aliment contenant plus de glucides (28 % protéines, 38 % glucides sur MS) (Hoeniget coll, 2007a). Il est important d’adapter la consommation au besoin énergétique, même si ces ali-ments sont proches du régime carnivore naturel du chat, car, au moins à court terme, les régimes richesen protéines en libre-service n’entraînent cependant pas de perte de poids significative. Mais si la quantité d’aliment est restreinte et que les chats perdent du poids, les régimes riches en protéinesont un effet bénéfique additionnel en favorisant la perte de masse grasse (Mazzaferro et coll, 2003;Hoenig et coll, 2007a).
Une augmentation modérée du niveau de fibres (25 à 30 g de FT/1000 kcal) permet de modérer la den-sité énergétique du régime et de réduire les niveaux de lipides et de glucides. Il faut ajuster les quanti-tés distribuées à l’état corporel (Nguyen et coll, 2004a,b). En moyenne, le besoin énergétique quotidienest d’environ 45-55 kcal/kg. La plupart des chats étant stérilisés et sédentaires, il faut éviter les régimestrès appétents et très énergétiques. Il faut que le propriétaire comprenne clairement que toute prise depoids au-dessus de la normale augmente le risque de développement du DS et doit donc être évitée(Scarlett et Donoghue, 1998). Une fois installée, l’obésité est un facteur de risque majeur pour le déve-loppement du DS car elle entraîne une diminution de la sensibilité à l’insuline (Biourge et coll, 1997;Appleton et coll, 2001b). Chez les chats obèses insulinorésistants, le profil de sécrétion d’insuline estmodifié avant même que la tolérance au glucose ne soit affectée (Hoenig, 2002).
Minimiser les pics de glucose postprandiaux
En plus d’agir sur le poids, l’alimentation peut intervenir sur le métabolisme. Un niveau d’amidon élevé(50 % des calories) entraîne une glycémie postprandiale, particulièrement si la source glucidique a unindex glycémique élevé (Figure 26). L’hyperglycémie stimule les cellules b du pancréas à sécréter plusd’insuline. Ce stress peut devenir ingérable pour le pancréas chez les chats en surpoids qui souffrentd’insulinorésistance. Cependant, aucune étude à l’heure actuelle ne montre que les régimes riches englucides sont directement liés au développement de l’insulinorésistance ou du DS.
12 -
Asp
ects
dié
tétiq
ues
du t
raite
men
t du
dia
bète
suc
ré f
élin
207
PRINCIPES DE FORMULATIOND'UN RÉGIME POURCHAT DIABÉTIQUE
Le régime idéal du chat diabétique doit être :- modérément énergétique
(< 4000 kcal/kg de MS),- faible en matières grasses (< 30 %
des calories),- mais riche en protéines
(> 45 % calories).
Diab
ète
Les chats diabétiques nourris avec un aliment hyperprotéique (protéines = 57 % de MS; glucides = 8 %)contrôlent mieux leur homéostasie que les chats nourris avec un aliment riche en glucides (protéines =40 %, glucides = 24 %; Frank et coll, 2001). Les régimes riches en protéines sont utiles pour diminuerl’hyperglycémie postprandiale (Figure 4) (par exemple, 63 % protéines, 8 % glucides dans l’étude deKettelhut et coll, 1980; respectivement 54 % et 8 % dans l’étude de Tschuor et coll, 2006).
En plus de l’équilibre protido-glucidique du régime, l’origine de l’amidon est également importante. Lasource de glucose présente dans les aliments pour chats diabétiques doit être complexe et avoir un indexglycémique bas (ex : orge, blé). Le riz, qui possède un index glycémique plus élevé que le maïs, entraîneune augmentation plus importante du glucose postprandial et de l’insulinémie (Rand et coll, 2004).
Puisque les aliments actuels pour chats diabétiques contiennent peu d’amidon, l’importance de ce fac-teur est sans doute à relativiser. L’index glycémique jouait un rôle beaucoup plus important dans lesrégimes riches en glucides que dans les régimes actuels. Ni le rôle spécifique de l’index glycémique dansles régimes hypoglucidiques, ni l’effet de l’origine de l’amidon ont été explorés.
Stimuler la sécrétion endogène d’insuline
Un régime riche en protéines aide également à atteindre ce troisième but car la réponse des cellules bdu pancréas aux acides aminés chez le chat diabétique est généralement maintenue plus longtemps quela réponse au glucose (Kitamura et coll, 1999). L’arginine a un effet puissant sur la sécrétion pancréa-tique d’insuline.
Intérêt des aliments riches en protéines pour le traitement du diabète sucré félin
L’introduction des régimes riches en protéines dans l’alimentation du chat diabétique a constitué ungrand pas en avant dans le traitement du DS félin. Plusieurs études montrent que les régimes riches enprotéines améliorent la situation métabolique chez les chats diabétiques obèses.
- Hoening (2006 a, b) montre que la sensibilité à l’insuline n’est pas normalisée chez les chats obèsesnourris avec un régime riche en glucides et ayant perdu du poids mais un régime riche en protéines(45 % MS) améliore la sensibilité à l’insuline des chats obèses. Cependant, les chats diabétiques nesont pas inclus dans cette étude.
12 -
Asp
ects
dié
tétiq
ues
du t
raite
men
t du
dia
bète
suc
ré f
élin
208
Diab
ète
FIGURE 26 - QU’EST-CE QUE L’INDEX GLYCÉMIQUE?
glycémie
temps
Glucose (étalon)
Aliment test
Chez l’homme, il est mesuré de la manière suivante :- Consommation d’un aliment contenant 50 g de glucides
en moins de 13 minutes- Suivi de la glycémie pendant les 2 à 3 heures après consommation :
mesure de l’aire sous la courbe (area under the curve : AUC)- Essai répété 2 fois chez 8 à 10 individus- Index Glycémique (IG) = rapport entre l’AUC de l’aliment testé
par rapport à l’AUC obtenu avec 50 g de glucose (méthode de Wolever)
- Classification des résultats :< 55 : IG basentre 55 et 70 : IG moyen> 70: IG élevé
Chez l’homme, cette méthode ne constitue pas une bonne évaluation pratique des aliments car les résultats peuvent varier selon l’environnementdes glucides au sein de l’aliment (composition du repas), la méthode de cuisson et les individus. Chez l’animal, la maîtrise de la composition des aliments permet d’obtenir des résultats plus fiables.

- Un aliment humide riche en protéines (57 % sur MS et 50 % des calories), pauvre en glucides diges-tibles (8 % sur MS et 13 % des calories) (Frank et coll, 2001) se révèle avantageux par rapport à unaliment plus riche en glucides digestibles (24 % MS et 23 % des calories) et riche en fibres (56 gFT/1000 kcal). La dose d’insuline peut être diminuée jusqu’à 50 % chez les chats diabétiques rece-vant un aliment riche en protéines. Pour 3 chats sur 9, l’insuline devient même facultative (Frank etcoll, 2001; Bennett et coll, 2006).
- À notre connaissance (Tschuor et coll, 2006), l’utilisation d’un régime humide riche en protéines(54 % MS) et pauvre en amidon conduit à un bien meilleur taux de rémission diabétique (50-70 %)que ce qui était décrit auparavant. Ceci survient d’ailleurs avant que la perte de poids ne soit trèsapparente. Bien que les régimes riches en protéines soient connus pour faciliter la perte de poids chezle chat (Szabo et coll, 2000; Michel et coll, 2005), celle-ci ne semble donc pas être nécessaire à l’amé-lioration des chats diabétiques.
Intérêt des aliments riches en protéines pour la prévention du diabète sucré félin
Il a été suggéré que les cellules b du pancréas du chat ne sont pas adaptées à un régime hyperglucidiqueet que ce type de régime peut être nuisible pour eux. (Il est important de préciser que les qualificatifsde “riche en protéines” et “riche en glucides” ne sont pas toujours utilisés uniformément. En général,un aliment “riche en protéines” contient au moins 50 % de protéines sur MS, alors qu’un aliment “richeen glucides” contient entre 30 et 50 % de glucides). L’influence à long terme d’un régime hypergluci-dique sur la probabilité de chats sains de développer un diabète reste cependant à définir. Une publi-cation rapporte que la sensibilité à l’insuline est diminuée et que l’hyperinsulinémie prévaut chez leschats recevant un aliment riche en glucides par rapport à ceux recevant un aliment riche en protéines(Hoenig, 2002). D’un autre côté, une étude chez des chats de poids normal n’a mis en évidence aucu-ne différence entre un aliment riche en protéines (environ 57 % MS pour les protéines et 22 % MSpour les glucides) et un aliment modérément riche en protéines (protéines = 32 % MS, glucides = 49 %MS) concernant la concentration et la sensibilité à l’insuline au cours d’un test d’hyperglycémie pro-voquée par voie intraveineuse ou de stimulation à l’arginine (Leray et coll, 2006). Des études plus appro-fondies quant à l’influence directe de la richesse respective en protéines ou riche en glucides d’un ali-ment sur le développement de l’insulinorésistance, de l’insuffisance des cellules b et éventuellementdu DS chez le chat sont nécessaires.
Les mécanismes sous-jacents permettant d’expliquer les avantages d’un régime hyperprotéique ne sont pasclairement connus. Ils pourraient être liés à une diminution du taux d’IGF-1 (Leray et coll, 2006; Alt et coll.(2007) notent aussi qu’un niveau bas d’IGF-1 chez des chats diabétiques peut se normaliser avec le traite-ment par l’insuline). Cependant, l’étude de Leray et coll. ne relève aucun effet particulier lié à un alimentsec riche en protéines (50 % des calories) sur la sensibilité à l’insuline des chats de poidsnormal. Ceci diffère des autres espèces. Actuellement, le rôle bénéfique d’un aliment richeen protéines pour prévenir le développement du diabète sucré n’est toujours pas établi.
Teneurs respectives en amidon et en fibres de l’aliment du chat diabétique
L’aliment traditionnel du chat diabétique contenait des taux relativement élevés d’ami-don (> 30 % des calories) et de fibres (> 50 g FT/1000 kcal). Les fibres végétales sontconsidérées comme avantageuses car elles retardent la vidange gastrique et l’absorptionintestinale du glucose, elles augmentent la sensibilité à l’insuline et elles améliorent lecontrôle du métabolisme des nutriments par le biais des hormones digestives (Nelson etcoll, 2000). Les fibres solubles visqueuses étaient jusqu’ici considérées comme très inté-ressantes car elles retardent le transport du glucose à la surface de la muqueuse gas-trointestinale (Nelson, 2005).
Une étude compare l’impact sur la gestion du diabète de deux régimes humides à teneurprotéique correspondant à environ 40 % de l’énergie. L’un contient de faibles quanti-tés d’amidon (12 % de l’énergie) et de fibres (0,1 g/100 kcal) et l’autre contient desquantités modérées d’amidon (26 % de l’énergie) et plus de fibres (environ 5 g/100 kcal)
12 -
Asp
ects
dié
tétiq
ues
du t
raite
men
t du
dia
bète
suc
ré f
élin
209
Diab
ète
Les graines de psyllium sont traditionnellement utilisées pour leursvertus “coupe-faim” dans le cadre des régimes amaigrissants. Grâceà leur aptitude à capter l’eau, les mucilages forment un gel volumi-neux en présence d’eau dans l’estomac et ralentissent la vidange del’estomac.

(Bennett et coll, 2006). Le taux le plus élevé de rémission du diabète est obtenu avec le premier régime(> 60 % contre environ 40 %). Un faible apport en glucides semble donc être avantageux et compensela relativement faible teneur en fibres de ce régime.
Une étude réalisée par Nelson et coll. (2000) montre qu’un régime contenant des quantités impor-tantes de fibres (13 % MS) est plus intéressant qu’un régime contenant des quantités faibles de fibres(2 % MS), pour des quantités de protéines similaires (44 % MS). Cependant, le régime riche en fibrescontenait aussi moins d’amidon (27 % contre 38 % MS). L’effet conjugué de tous ces éléments peutfort bien être responsable des avantages observés.
Globalement, il est maintenant admis que l’aliment idéal d’un chat diabétique doit être riche en pro-téines et pauvre en amidon. Dans ces conditions, le taux de fibres peut être moins important qu’aupa-ravant. Cependant, en ralentissant le transit gastro-intestinal, les fibres diététiques ont un rôle nonnégligeable dans l’alimentation des chats diabétiques. De plus, la richesse en fibres de l’aliment tend àdiluer la concentration énergétique, ce qui aide à contrôler la consommation alimentaire des chats.
Rôle de certains acides gras
Le rôle des différents acides gras chez les chats obèses a aussi étéenvisagé. Une étude compare un aliment riche en acides graspolyinsaturés oméga-3 (AGPI n-3; 20,1 % de matières grasses surMS; les AGPI n-3 représentent 9,6 % des matières grasses), avecun aliment témoin contenant des taux réduits de AGPI n-3(19,8 % de matières grasses ; les AGPI n-3 représentent 1,5 %).L’aliment riche en AGPI n-3 améliore le contrôle à long termede la glycémie et diminue l’insulinémie (Wilkins et coll, 2004).En revanche, les acides gras saturés sont considérés commenéfastes au maintien d’une glycémie normale. Ces observationspourraient reposer sur l’activation ou l’expression accrue dePPAR g et donc une augmentation de la sensibilité à l’insuline.
Oligo-éléments et antioxydants
Le chrome a été considéré comme un cofacteur essentiel pourl’activité de l’insuline. Le mécanisme exact par lequel le chromeaugmente la sensibilité à l’insuline n’est pas connu. Cependant,les données sont contradictoires et ne permettent pas deconclure. L’utilisation du chrome ne peut donc pas être recom-mandée. À notre connaissance, l’effet du chrome sur les chatsdiabétiques n’a pas été publié. Ses effets bénéfiques semblentnégligeables par rapport à d’autres traitements.
Le vanadium, un autre oligo-élément, semble posséder des effetssimilaires à ceux du chrome, mais par des mécanismes différents.Seuls des résultats préliminaires sont disponibles et suggèrent quele vanadium aurait des effets bénéfiques chez le chat diabétique.La dose recommandée est de 0,2 mg/kg par jour, administré avecde la nourriture ou de l’eau (Nelson, 2005).
L’hyperglycémie chronique entraîne une glucotoxicité quicontribue à la dégénérescence progressive des cellules b et à l’in-sulinorésistance. Ceci est dû en partie à l’augmentation du stressoxydatif intracellulaire. A notre connaissance, la généralisationde l’utilisation d’antioxydants pour réduire ces effets n’a pas faitl’objet d’études correctement contrôlées chez le chat. Cepen-dant, ces composés n’étant pas dangereux, leur inclusion dans lesaliments peut être envisagée.
12 -
Asp
ects
dié
tétiq
ues
du t
raite
men
t du
dia
bète
suc
ré f
élin
Les acides gras -trans (AGT) sont une forme particulière de graisses insaturées. Laplupart des acides gras naturels insaturés ont une configuration-cis, mais dans le casdes AGT, la configuration spatiale est différente car les atomes d’hydrogène sesituent à l’opposé de la double liaison. Les viandes de ruminants et les produits lai-tiers sont naturellement riches en AGT: la transformation de la forme -cis vers laforme -trans a lieu dans le rumen. Des taux élevés d’AGT peuvent également êtreobtenus lors de l’hydrogénation industrielle ou de la désodorisation d’huiles végé-tales. Les matières grasses issues de ruminants contiennent entre 5 et 8 % d’AGTmais le taux peut s’élever à 45 % dans les huiles partiellement hydrogénées.
AGT et nutrition humaineLes risques sanitaires potentiels associés à la consommation d’AGT ont récemmentfocalisé l’attention du public. Les AGT alimentaires augmenteraient en effet la résis-tance à l’insuline et représenteraient alors un facteur de risque de développementdu diabète sucré de type 2 (DS2). Le remplacement des AGT par d’autres acides graspolyinsaturés serait donc une mesure bénéfique pour la prévention du diabète. Enconséquence, plusieurs pays imposent maintenant un étiquetage clair concernantles AGT. Certains pays, tel que le Danemark, restreignent même la vente des huileset graisses contenant un taux élevé de AGT (soit plus de 2 % d’AGT au Danemark).Aux États-Unis, les AGT doivent être déclarés séparément sur les étiquettes des pro-duits alimentaires.
Tous les AGT ne se valent pasIl est très important d’insister sur le fait que tous les AGT ne se valent pas. Ceux quisont obtenus à la suite du traitement industriel des huiles végétales ont des effetsnégatifs qui ne se retrouvent pas chez les AGT produits par la fermentation micro-bienne du rumen. Parmi ces derniers, au moins un, l’acide C18 trans-vaccen joueraitmême un rôle positif sur la santé. Cet acide gras peut en effet être métabolisé enacide linoléique conjugué, dont les propriétés anti-diabétiques et anti-tumorales ontdéjà été montrées chez l’animal.
Les AGT dans l’alimentation des chiens et des chatsActuellement, il n’existe pas de raison de croire que la présence d’AGT issus deviandes de ruminants dans les aliments pour chiens et chats puisse avoir un quel-conque effet nuisible. Il n’existe encore aucune étude sur ce sujet. La distinction entrel’influence des AGT d’origine naturelle ou industrielle reste également à étudier.
ACIDES GRAS-TRANSPatricia A. Schenck, DVM, PhD
210
Diab
ète Dans la configuration trans,
les atomes d’hydrogène se situent à l’opposé de la double liaison.
Configuration cis
CONFIGURATIONS CIS ET TRANSDES ACIDES GRAS
Configuration trans

13 - Influence du niveau de protéines sur la fonction rénale
La question de l’impact sur la fonction rénale de la distribution à long terme d’aliments riches en pro-téines a été soulevée. Cependant, il faut souligner qu’il n’existe pas de preuve d’une détérioration dela fonction rénale des chats normaux ou ayant une maladie rénale débutante lorsque ceux-ci reçoiventun aliment riche en protéines (Finco et coll, 1998). Les régimes riches en protéines sont cependantcontre-indiqués chez les chats azotémiques alors que la néphropathie est relativement fréquente chezles chats diabétiques (Nelson, 2005); de plus, il est important de surveiller la phosphatémie pour contrô-ler la progression de la maladie rénale. À notre connaissance, cependant, aucune étude n’a approfon-di cette question.
Au cas où un chat diabétique présenterait un dysfonctionnement rénal et une azotémie, le taux pro-téique de l’aliment doit être réduit afin de minimiser le risque d’urémie. Dans ce type de situation,l’ajout de médicaments comme l’acarbose, qui limite l’absorption gastro-intestinale des glucides, peutêtre envisagé. Cependant, les données manquent pour étayer cette recommandation.
Malgré l’amélioration nette de la gestion des chats diabétiques depuis l’introduction des régimes richesen protéines et pauvres en glucides, plusieurs questions restent sans réponse.- Quel sont les éléments clés, les protéines ou les glucides? En d’autres termes, est-ce le caractère hyper-
protéique ou hypoglucidique le plus important?- Certains acides aminés particuliers comme l’arginine offrent-ils des avantages? L’origine des protéines
pourrait-elle jouer un rôle (Leray et coll, 2006) ?- Quelles sont les conséquences à long terme de ces régimes par rapport au risque d’acidocétose diabé-
tique ou de néphropathie diabétique? Jusqu’à présent, rien n’indique que l’utilisation prolongée deces régimes entraîne une défaillance de la fonction rénale chez les chats normaux ou chez les chatsavec une maladie rénale débutante (Finco et coll, 1998).
- Quelles sont les conséquences sur à long terme des régimes riches en protéines sur le poids et la com-position corporelle?
14 -Recommandations pratiques pour le chat diabétique
Présentation de l’aliment
Les aliments diététiques pour chats diabétiques sont aujourd’hui disponibles soit sous forme humide(boîtes de conserve) soit sous forme sèche (croquettes). L’amélioration de la cuisson-extrusion des ali-ments secs permet en effet de fabriquer des aliments secs riches en protéines et pauvres en glucides. Dumoment que cet équilibre protido-glucidique est respecté, aucune donnée ne permet de préférer l’uneou l’autre forme.
Rythme d’alimentation
La plupart des chats diabétiques peuvent être nourris deux fois par jour, avec une injection d’insulinejuste avant ou après les repas. Ce rythme de ce régime est différent du rythme d’alimentation naturel,pour lequel jusqu’à 15 petits repas par jour sont possibles. Néanmoins, le glucose postprandial n’aug-mente que légèrement avec les régimes riches en protéines, comparés aux régimes riches en glucides(Kettelhut et coll, 1980; Kienzle, 1994; Martin et Rand, 1999). La programmation des injections d’insu-line selon les repas semble donc moins importante. Une étude non publiée montre que la program-mation de l’injection d’insuline, supposée être optimisée par rapport à l’activité de l’insuline (45minutes avant un repas contre le début du repas) a peu d’effets sur le contrôle du métabolisme (Alt,2006). La composition de l’aliment est en réalité beaucoup plus importante que l’heure des repas.Cependant, il est nécessaire que l’aliment soit disponible lorsque l’insuline débute son action afin d’em-pêcher une hypoglycémie potentiellement fatale.
13 -
Influ
ence
du
nive
au d
e pr
otéi
nes
sur
la f
onct
ion
réna
le
211
Diab
ète
Selon notre expérience, la plupart deschats accepte facilement les alimentsriches en protéines et pauvres en glucides. Pour beaucoup d’entre eux,l’appétit est même augmenté dans lesstades initiaux du traitement.
© S
téph
anie
Vid
al

Surveillance médicale
Il faut surveiller une possible hypoglycémie chez les chats diabétiques traités par l’insuline etrecevant une alimentation riche en protéines et pauvre en glucides.
Ce point renforce l’idée que les chats doivent être régulièrement suivis au cours du traite-ment. Pour cela, la glycémie peut être mesurée à la maison avec des glucomètres portables(Figure 27; Reusch et coll, 2006), en association avec à une mesure régulière de la fructosa-mine au laboratoire. Les propriétaires doivent également connaître les signes cliniques d’unehypo- ou d’une hyperglycémie. Tout au long du traitement, et ensuite lorsque l’insulinothé-rapie n’est plus nécessaire (diabète transitoire), les propriétaires peuvent facilement détec-ter une glucosurie avec des bandelettes, dans la litière du chat légèrement imprégnée d’eau.Cette information permet de pouvoir revoir le traitement insulinique si nécessaire.
Si le traitement de normalisation de la glycémie est efficace, avec l’insuline et un régimeriche en protéines et pauvre en glucides, beaucoup de chats entrent en rémission et n'ontpas besoin d'être traités toute leur vie. L’administration d’insuline peut alors être arrêtée. Ilest recommandé de maintenir le régime riche en protéines pendant la rémission et de conti-nuer un suivi régulier pour repérer une récidive éventuelle des signes cliniques de diabètesucré. Dans ce cas, un traitement spécifique doit être instauré immédiatement.
ConclusionLe diabète sucré félin est un trouble métabolique fréquent dont la prévalence a augmenté aucours des 30 dernières années. Ceci a certainement un lien avec le problème de l’obésité denos chats domestiques. Cependant, le traitement s’est beaucoup amélioré et le taux de mor-talité du diabète sucré a énormément baissé au cours des 10-20 dernières années. Puisquel’anomalie physiopathologique majeure est la carence ou la résistance à l’insuline, la plupartdes chats diabétiques ont traditionnellement été traités avec de l’insuline. Celle-ci constituetoujours le traitement de choix car elle permet de contrôler le métabolisme du glucose etd’aider à réduire la glucolipotoxicité. Parfois les signes cliniques disparaissent complètement.Au cours de ces dernières années, le principe d’associer l’insulinothérapie à un régime richeen protéines (>50 %) et pauvre en glucides (<15 %) est devenu une évidence. Le taux derémission a beaucoup augmenté depuis l’introduction de ces régimes dans la thérapie. Globalement, le diabète sucré félin est réellement une maladie qui peut et doit être traitée.
Con
clus
ion
Figure 27 - Suivi à la maison de la glycémie chez le chat.
Du sang capillaire est prélevé à l’oreille.
Le glucose est mesuré à l’aide d’un glucomètreportable.
Ave
c l’a
imab
le a
utor
isatio
n du
Pro
f. C
. R
eusc
h,
Vet
suiss
e-Fa
culty
Uni
vers
ity o
f Zur
ich
Ave
c l’a
imab
le a
utor
isatio
n du
Pro
f. C
. R
eusc
h,
Vet
suiss
e-Fa
culty
Uni
vers
ity o
f Zur
ich
Ave
c l’a
imab
le a
utor
isatio
n du
Pro
f. C
. R
eusc
h,
Vet
suiss
e-Fa
culty
Uni
vers
ity o
f Zur
ich
212
Diab
ète
Que
stio
ns f
réqu
emm
ent
posé
es
213
Quel est le moyen le plus efficace de traiter un chat diabétique?
L’expérience de ces quelques dernières années est clairement en faveur d’une insulinothérapieintense (souvent 2 fois par jour), associée à une alimentation riche en protéines et pauvre en glucides.
Les chats diabétiques présentent-ilsune hyperglycémie postprandiale?
Les chats qui reçoivent un aliment riche en protéines présentent peu ou pas d’hyperglycémie post-prandiale. Plus la teneur en glucides digestibles de l’aliment est élevée, plus l’hyperglycémie post-prandiale est importante.
Quel est l’effet des différents aliments sur la glycémie moyenne?
En général, il est plus facile de maintenir une glycémie à peu près normale chez les chats diabé-tiques traités à l’insuline lorsqu’ils sont nourris avec un aliment riche en protéines et pauvre englucides. L’hyperglycémie est presque absente et la glycémie moyenne est réduite.
Combien de temps avant ou après l’injection d’insuline faut-il nourrirun chat diabétique?
L'injection peut avoir lieu juste après le repas mais il n’existe pas de recommandations précises àce sujet. Aucune différence métabolique majeure n’est observée entre un repas qui suit immédia-tement l’injection ou différé de 45 minutes.
Quel est le rythme d’alimentationidéal pour le chat diabétique?
Si le poids n’est pas un problème, la nourriture peut être laissée en libre-service. Si le chat est obèse,la nourriture doit être restreinte. Dans cette situation, deux repas par jour, suivis d’une injectiond’insuline, sont recommandés.
Que faire lorsqu’un chat diabétiquene mange pas après son injection d’insuline?
Dans une situation d’urgence, lorsque le chat diabétique a reçu sa dose d’insuline et ne mange pas,il faut lui proposer rapidement des glucides à absorption rapide, tel que du miel, afin d’empêcherune hypoglycémie potentiellement fatale. Une telle situation d’urgence peut être prévenue enadministrant l’insuline seulement après le repas. Ceci s’avère cependant difficile à mettre en pra-tique pour certains propriétaires.
Peut-on varier l’alimentation du chatdiabétique?
Idéalement, les chats diabétiques doivent toujours recevoir une alimentation riche en protéines,même si une rémission survient. Certaines publications font état de récidives d’hyperglycémie enquelques jours lorsque le régime est changé pour un aliment riche en glucides. Étant donné la situa-tion métabolique des chats et l’intérêt particulier des régimes riches en protéines et pauvres en glu-cides, on ne peut que recommander leur utilisation à long terme, même après résolution des signescliniques.
L’activité physique joue-t-elle un rôledans le traitement?
Il s’avère difficile de surveiller l’activité physique d’un chat. Cependant, il est recommandé que lechat ait une activité régulière, pour que la consommation alimentaire et la dépense énergétiquesoient en concordance avec le traitement insulinique et le régime.
L’aliment du chat diabétique doit-ilcontenir un taux élevé de fibres alimentaires?
Les fibres ont été longtemps recommandées pour les chats diabétiques. Cependant, un niveau élevéde fibres n’apparaît pas comme le facteur le plus important. Les régimes riches en protéines etpauvres en glucides semblent très efficaces. À l’heure actuelle, il n’est pas encore certain que lesfibres apportent un bénéfice supplémentaire. Dans tous les cas cependant, la faible densité éner-gétique des régimes riches en fibres facilite le contrôle du poids.
Que faut-il faire pour obtenir unpoids corporel optimal chez les chatsdiabétiques?
La plupart des chats diabétiques sont obèses. Le traitement doit donc tendre à réduire le poids cor-porel. Une baisse du poids corporel de 1,5 % par semaine semble raisonnable (voir chapitre n° 1).Lorsqu’ils sont nourris avec un aliment riche en protéines, les chats perdent de la masse grasse,mais conservent leur masse maigre.
Peut-on prévenir le diabète sucré?
La prévention de l’obésité apparaît comme le facteur le plus important pour diminuer le risque dediabète. Ceci est particulièrement vrai pour les chats castrés, car ils mangent davantage et dépen-sent moins d’énergie. Le risque d’obésité est 3 à 4 fois plus élevé chez les chats castrés et le risquede diabète est 4 fois plus élevé chez les chats obèses.
Q R
Questions fréquemment posées à propos des alimentsriches en protéines et pauvres en glucides
Diab
ète

Réfé
renc
es
214
Diab
ète
Références
Ahima RS. Central actions of adipocyte hormones.Trends Endocrinol Metab 2005; 16: 307-313.
Alt N. The effect of feeding time on the quality ofmetabolic control, day-to-day variability of blood glucose curves and evaluation of IGF-1 levels in catswith diabetes mellitus. Dissertation, University ofZurich, 2006.
Alt N, Kley S, Tschuor F, et al Evaluation of IGF-1 levels in cats with transient and permanentdiabetes mellitus. Res Vet Sci 2007; doi:10.1016/j.rvsc.2007.01.014.
Appleton DJ, Rand JS, Sunvold GD. Plasma leptinconcentrations in cats: reference range, effect ofweight gain and relationship with adiposity as measuredby dual energy X-ray absorptiometry. J Fel MedSurg 2000; 2: 191-199.
Appleton DJ, Rand JS, Priest J, et al. Determinationof reference values for glucose tolerance, insulin tolerance, and insulin sensitivity tests in clinically normal cats. Am J Vet Res 2001a; 62: 630-636.
Appleton DJ, Rand JS, Sunvold GD. Insulin sensitivity decreases with obesity, and lean cats withlow insulin sensitivity are at greatest risk of glucoseintolerance with weight gain. J Fel Med Surg2001b; 3: 211-228.
Bailiff NL, Nelson RW, Feldman EC, et al.Frequency and risk factors for urinary tract infectionin cats with diabetes mellitus. J Vet Intern Med2006; 20: 850-855.
Banks WA, Farr SA, Morley JE. The effects of highfat diets on the blood-brain barrier transport of leptin: failure or adaptation? Physiol Behav 2006;88: 244-248.
Baral RM, Rand JS, Catt MJ. Prevalence of felinediabetes mellitus in a feline private practice. J VetIntern Med 2003; 17: 433.
Barroso I. Genetics of type 2 diabetes. Diabet Med2005; 22: 517-535.
Bennett N, Greco DS, Peterson ME, et al.Comparison of low carbohydrate-low fiber diet and a moderate carbohydrate-high fiber diet in the management of feline diabetes mellitus. J Fel MedSurg 2006; 8: 73-84.
Biourge V, Nelson RW, Feldman EC, et al. Effectof weight gain and subsequent weight loss on glucosetolerance and insulin response in healthy cats. J VetIntern Med 1997; 11: 86-91.
Biourge VC. Feline diabetes mellitus: nutritionalmanagement. Waltham Focus 2005; 15: 36-40.
Brennan CL, Hoenig M, Ferguson DC. GLUT4but not GLUT1 expression decreases early in thedevelopment of feline obesity. Domest AnimEndocrinol 2004; 26: 291-301.
Butler AE, Janson J, Soeller WC, et al. Increasedbeta-cell apoptosis prevents adaptive increase in beta-cell mass in mouse model of type 2 diabetes:evidence for role of islet amyloid formation ratherthan direct action of amyloid. Diabetes 2003; 52:2304-14.
Chan DL, Freeman LM, Rozanski EA, et al.Alterations in carbohydrate metabolism in critically illcats. J Vet Emerg Crit Care 2006; 16: S7-S13.
Cooper GJS. Amylin compared with calcitonin-generelated peptide: structure, biology and relevance tometabolic disease. Endocr Rev 1994; 15: 163-201.
Crenshaw KL, Peterson ME, Heeb LA, et al.Serum fructosamine concentration as an index of glycemia in cats with diabetes mellitus and stress hyperglycemia. J Vet Intern Med 1996; 10: 360-364.
Diez M, Nguyen P. The epidemiology of canine and feline obesity. Waltham Focus 2006; 16: 2-8.
Dohan FC, Lukens FDW. Experimental diabetesproduced by the administration of glucose.Endocrinol 1948; 42: 244-262.
Donath MY, Ehses JA, Maedler K, et al.Mechanisms of beta-cell death in type 2 diabetes.Diabetes 2005; 54 suppl. 2: S108-S113.
Drucker DJ. Minireview: the glucagon-like peptides.Endocrinol 2001; 142: 521-527.
Edelman SV, Weyer C. Unresolved challenges with insulin therapy in type 1 and type 2 diabetes:potential benefit of replacing amylin, a second beta-cell hormone. Diab Technol Therapeut 2002;4: 175-189.
Elliott DA, Nelson RW, Reusch CE, et al.Comparison of serum fructosamine and blood glycosylated hemoglobin concentrations for assessment of glycemic control in cats with diabetesmellitus. J Am Vet Med Assoc 1999; 214: 1794-1798.
Feldhahn JR, Rand JS, Martin G. Insulin sensitivityin normal and diabetic cats. J Fel Med Surg 1999;1: 107-115.
Feldman EC, Nelson RW, Feldman MS. Intensive50-week evaluation of glipizide administration in 50cats with previously untreated diabetes mellitus. JAm Vet Med Assoc 1997; 210: 772-777.
Feldman EC, Nelson RW. Feline diabetes mellitus.In: Feldman EC, Nelson RW, eds. Canine andFeline Endocrinology and Reproduction. 3rd ed. St.Louis: Saunders, 2004; 539-579.
Fettman MJ, Stanton CA, Banks LL, et al. Effectsof neutering on bodyweight, metabolic rate and glucose tolerance of domestic cats. Res Vet Sci1997; 62: 131-136.
Finco DR, Brown SA, Brown CA, et al. Protein andcalorie effects on progression of induced chronic renalfailure in cats. Am J Vet Res 1998; 59: 575-82.
Fineman M, Weyer C, Maggs DG, et al. The human amylin analog, pramlintide, reducespostprandial hyperglucagonemia in patients with type2 diabetes mellitus. Horm Metab Res 2002; 34:504-508.
Franconi F, Loizzo A, Ghirlanda G, et al. Taurinesupplementation and diabetes mellitus. Curr OpinClin Nutr Metab Care 2006; 9 : 32-36.
Frank G, Anderson W, Pazak H, et al. Use of high-protein diet in the management of feline diabetesmellitus. Vet Therapeut 2001; 2: 238-245.
Furrer D, Tschuor F, Reusch C, et al. Effect ofamylin on plasma concentrations of glucose, insulinand glucagon in an arginine stimulation and meal response test in cats, in: Proceedings ECVIMForum 2005.
German AJ. The growing problem of obesity in dogsand cats. J Nutr 2006; 136: 1940S-1946S.
Goossens MMC, Nelson RW, Feldman EC, et al.Response to insulin treatment and survival in 104 catswith diabetes mellitus (1985-1995). J Vet InternMed 1998; 12: 1-6.
Harper EJ, Stack DM, Watson TDG, et al. Effectsof feeding regimens on bodyweight, composition andcondition score in cats following ovariohysterectomy.J Small Anim Pract 2001; 42: 433-438.
Henson MS, O’Brien TD. Feline models of type 2diabetes mellitus. ILAR J 2006; 47: 234-242.
Hoenig M. Comparative aspects of diabetes mellitusin dogs and cats. Mol Cell Endocrinol 2002; 197:221-229.

Réfé
renc
es
215
Diab
ète
Hoenig M. The cat as a model for human nutritionand disease. Curr Opin Nutr Metab Care 2006a;9: 584-588.
Hoenig M. Unique aspects of metabolism in obesecats, in Proceedings. 24th ACVIM Forum 2006b;432.
Hoenig M, Alexander S, Holson J, et al. Influenceof glucose dosage on interpretation of intravenousglucose tolerance tests in lean and obese cats. J VetIntern Med 2002; 16: 529-532.
Hoenig M, Caffall ZF, McGraw RA, et al.Cloning, expression and purification of feline proinsulin. Domest Anim Endocrinol 2006a; 30:28-37.
Hoenig M, Ferguson DC. Effect of darglitazone onglucose clearance and lipid metabolism in obese cats.Am J Vet Res 2003; 64: 1409-1413.
Hoenig M, Ferguson DC. Effects of neutering onhormonal concentrations and energy requirements in male and female cats. Am J Vet Res 2002; 63:634-639.
Hoenig M, Ferguson DC. Impairment of glucosetolerance in hyperthyroid cats. J Endocrinol 1989;121: 249-251.
Hoenig M, Hall G, Ferguson D, et al. A felinemodel of experimentally induced islet amyloidosis.Am J Pathol 2000; 157: 2143-2150.
Hoenig M, McGoldrick JB, deBeer M, et al.Activity and tissue-specific expression of lipases andtumor-necrosis factor alpha in lean and obese cats.Domest Anim Endocrinol 2006b; 30: 333-344.
Hoenig M, Thomaseth K, Brandao J, et al.Assessment and mathematical modeling of glucose turnover and insulin sensitivity in lean and obese cats.Domest Anim Endocrinol 2006c; 31: 373-389.
Hoenig M, Thomaseth K, Waldron M, et al. Fattyacid turnover, substrate oxidation, and heat productionin lean and obese cats during the euglycemic hyperinsulinemic clamp. Domest Anim Endocrinol2007b; 32: 39-338.
Hoenig M, Thomaseth K, Waldron M, et al. Insulinsensitivity, fat distribution and adipocytokine responseto different diets in lean, and obese cats before andafter weight loss. Am J Physiol Regul Integr CompPhysiol 2007a; 292: R227-R234.
Höppener JWM, Nieuwenhuis MG, Vroom TM, etal. Role of islet amyloid in type 2 diabetes mellitus:consequence or cause? Mol Cell Endocrinol 2002;197: 205-212.
Jin Y, Lin D. Fungal urinary tract infections in thedog and cat: a retrospective study (2001-2004). JAm Anim Hosp Assoc 2005; 41: 373-381.
Johnson KH, Hayden DW, O'Brien TD, et al.Spontaneous diabetes mellitus - islet amyloid complexin adult cats. Am J Pathol 1986; 125: 416-419.
Johnson KH, O’Brien TD, Betsholtz C, et al. Isletamyloid, islet-amyloid polypeptide and diabetes mellitus. New Engl J Med 1989; 321, 513-518.
Kanchuk ML, Backus RC, Calvert CC, et al.Neutering induces changes in food intake, bodyweight, plasma insulin and leptin concentrations in normal and lipoprotein lipase-deficient male cats. J Nutr 2002; 132: 1730S-1732S.
Kanchuk ML, Backus RC, Calvert CC, et al.Weight gain in gonadectomized normal and lipoproteinlipase-deficient male domestic cats results fromincreased food intake and not decreased energy expenditure. J Nutr 2003; 133: 1866-1874.
Kaneko JJ, Kawamoto M, Heusner AA, et al.Evaluation of serum fructosamine concentration asan index of blood glucose control in cats with diabetesmellitus. Am J Vet Res 1992; 53: 1797-1801.
Kettelhut IC, Foss MC, Migliorini RH. Glucosehomeostasis in a carnivorous animal (cat) and in ratsfed a high-protein diet. Am J Physiol 1980; 239:R437-R444.
Kienzle E. Blood sugar levels and renal sugar excre-tion after the intake of high carbohydrate diets in cats. J Nutr 1994; 124: 2563S-2567S.
Kitamura T, Yasuda J, Hashimoto A. Acute insulinresponse to intravenous arginine in nonobese healthycats. J Vet Intern Med 1999; 13: 549-556.
Konarkowska B, Aitken J, Kistler J, et al. Theaggregation potential of human amylin determines itscytotoxicity towards islet beta-cells. FEBS J 2006;273: 3614-3624.
Laluha P, Gerber B, Laluhova D, et al. Stresshyperglycemia in sick cats: a retrospective study over4 years. Schweiz Arch Tierheilk 2004; 146: 375-383.
Lazar MA. How obesity causes diabetes: not a talltale. Science 2005; 307: 373-375.
Lederer R, Rand JS, Hughes IP, et al. Chronic orrecurring medical problems, dental disease, repeatedcorticosteroid treatment, and lower physical activityare associated with diabetes in burmese cats. J VetIntern Med 2003; 17: 433.
Leray V, Siliart B, Dumon H, et al. Protein intakedoes not affect insulin sensitivity in normal weightcats. J Nutr 2006; 136:2028S-2030S.
Link KRJ, Rand JS. Reference values for glucosetolerance and glucose tolerance status in cats. J Am Vet Med Assoc 1998; 213: 492-496.
Ludvik B, Lell B, Hartter E, et al. Decrease of stimulated amylin release precedes impairment ofinsulin secretion in type II diabetes. Diabetes 1991;40: 1615-1619.
Lund EM, Armstrong PJ, Kirk CA, et al.Prevalence and risk factors for obesity in adult catsfrom private US veterinary practices. Intern J ApplRes Vet Med 2005; 3: 88-96
Lutz TA, Ainscow J, Rand JS. Frequency of pancreatic amyloid deposition in cats from south-eastern Queensland. Aus Vet J 1994; 71:254-256.
Lutz TA, Rand JS. Pathogenesis of feline diabetesmellitus. Vet Clin North Am Small Anim Pract1995; 25: 527-552.
Lutz TA, Rand JS, Ryan E. Fructosamine concentrations in hyperglycemic cats. Can Vet J1995 ; 36 : 155-159.
Lutz TA, Rand JS. Plasma amylin and insulin concentrations in normoglycemic and hyperglycemiccats. Can Vet J 1996 ; 37 : 27-34.
Lutz TA, Rand JS. Detection of amyloid depositionin various regions of the feline pancreas by differentstaining techniques. J Comp Pathol 1997; 116:157-170.
Lutz TA. The pancreatic hormone amylin as a centrally acting satiating hormone. Curr DrugTargets 2005; 6: 181-189.
Ma Z, Westermark GT, Johnson KH, et al.Quantitative immunohistochemical analysis of isletamyloid polypeptide (IAPP) in normal, impaired glucose tolerant, and diabetic cats. Amyloid 1998;5: 255-261.
Malecki MT. Genetics of type 2 diabetes mellitus.Diabetes Res Clin Pract 2005; 68 suppl. 1 : S10-S21.
Marshall RD, Rand JS. Comparison of the pharmacokinetics and pharmacodynamics of onceversus twice daily administration of insulin glarginein normal cats. J Vet Intern Med 2002; 16: 373.

Réfé
renc
es
216
Diab
ète
Marshall RD, Rand JS. Insulin glargine and a highprotein-low carbohydrate diet are associated with highremission rates in newly diagnosed diabetic cats. J Vet Intern Med 2004; 18: 401.
Martin GJW, Rand JS. Food intake and blood glucose in normal and diabetic cats fed ad libitum. J Fel Med Surg 1999; 1: 241-251.
Martin L, Siliart B, Dumon H, et al. Leptin, body fat content and energy expenditure in intact and gonadectomized adult cats: a preliminary study. J Anim Physiol Anim Nutr 2001; 85; 195-199.
Martin L, Siliart B. Hormonal consequences of neutering in cats. Waltham Focus 2005; 15: 32-35.
Matveyenko AV, Butler PC. Beta-cell deficit due toincreased apoptosis in the human islet amyloid polypeptide transgenic (HIP) rat recapitulates themetabolic defects present in type 2 diabetes.Diabetes 2006; 55: 2106-2114.
Matveyenko AV, Butler PC. Islet amyloid polypeptide (IAPP) transgenic rodents as models fortype 2 diabetes. ILAR J 2006; 47: 225-233.
Mazzaferro EM, Greco DS, Turner AS, et al.Treatment of feline diabetes mellitus using an alpha-glucosidase inhibitor and a low-carbohydratediet. J Fel Med Surg 2003; 5: 183-189.
McCann TM, Simpson KE, Sham DJ, et al. Felinediabetes mellitus in the UK: the prevalence within aninsured cat population and a questionnaire-basedputative risk factor analysis. J Fel Med Surg 2007;9: 289-299.
Meurs KM, Fox PR, Miller MW, et al. Plasmaconcentrations of tumor necrosis factor-alpha in catswith congestive heart failure. Am J Vet Res 2002;63: 640-642.
Mexas AM, Hess RS, Hawkins EC, et al.Pulmonary lesions in cats with diabetes mellitus. J Vet Intern Med 2006; 20: 47-51.
Michel KE, Bader A, Shofer FS, et al. Impact oftime-limited feeding and dietary carbohydrate contenton weight loss in group-housed cats. J. Fel Med Surg2005; 7: 349-355.
Mizisin AP, Nelson RW, Sturges BK, et alComparable myelinated nerve pathology in feline andhuman diabetes mellitus. Acta Neuropathol 2007;113: 431-442.
Muff R, Born W, Lutz TA, et al. Biological importance of the peptides of the calcitonin family asrevealed by disruption and transfer of correspondinggenes. Peptides 2004; 25: 2027-2038.
Nakayama H, Uchida K, Ono K, et al. Pathologicalobservation of six cases of feline diabetes mellitus.Jpn J Vet Sci 1990; 52: 819-825.
Nelson RW, Himsel CA, Feldman EC, et al.Glucose tolerance and insulin response in normal-weight and obese cats. Am J Vet Res 1990;51: 1357-1362.
Nelson RW, Feldman EC, Ford SL, et al. Effect ofan orally administered sulfonylurea, glipizide, fortreatment of diabetes mellitus in cats. J Am VetMed Assoc 1993; 203: 821-827.
Nelson RW, Griffey SM, Feldman EC, et al.Transient clinical diabetes mellitus in cats: 10 cases(1989-1991). J Vet Intern Med 1999; 13: 28-35.
Nelson RW, Scott-Moncrieff JC, Feldman EC, et al. Effect of dietary insoluble fiber on control ofglycemia in cats with naturally acquired diabetes mellitus. J Am Vet Med Assoc 2000; 216: 1082-1088.
Nelson RW, Spann D, Elliott DA, et al. Evaluationof the oral antihyperglycemic drug metformin in normaland diabetic cats. J Vet Intern Med 2004; 18: 18-24.
Nelson RW. Diabetes mellitus. In: Ettinger SJ,Feldman EC, eds. Textbook of Veterinary InternalMedicine. 6th ed. St. Louis: Elsevier Saunders,2005; 1563-1591.
Nguyen P, Leray V, Dumon H, et al. High proteinintake affects lean body mass but not energy expenditure in nonobese neutered cats. J Nutr2004a; 134 suppl.: 2084S-2086S.
Nguyen P, Dumon HJ, Siliart BS, et al. Effects ofdietary fat and energy on body weight and compositionafter gonadectomy in cats. Am J Vet Res 2004b;65: 1708-1713.
O’Brien TD, Hayden DW, Johnson KH, et al. High dose intravenous glucose tolerance test and serum insulin and glucagon levels in diabetic and non-diabetic cats: relationships to insular amyloidosis. Vet Pathol 1985; 22: 250-261.
O'Brien TD, Westermark P, Johnson KH. Isletamyloid polypeptide and insulin secretion from isolated perfused pancreas of fed, fasted, glucose-treated and dexamethasone treated rats.Diabetes 1991; 40: 1701-1706.
O’Brien TD. Pathogenesis of feline diabetes mellitus.Mol Cell Endocrinol 2002; 197: 213-219.
Panciera DL, Thomas CB, Eicker SW, et al.Epizootiologic patterns of diabetes mellitus in cats:333 cases (1980-1986). J Am Vet Med Assoc1990; 197: 1504-1508.
Petrus DJ, Jackson MW, Kemnitz JW, et al.Assessing insulin sensitivity in the cat: evaluation of the hyperinsulinemic euglycemic clamp and theminimal model analysis. Res Vet Sci 1998; 65:179-181.
Plier ML, Grindem CB, MacWilliams PS, et al.Serum fructosamine concentration in nondiabetic anddiabetic cats. Vet Clin Pathol 1998; 27: 34-39.
Prahl A, Glickman L, Guptill L, et al. Time trendsand risk factors for diabetes mellitus in cats. J VetIntern Med 2003; 17: 434.
Prahl A, Guptill L, Glickman NW, et al. Timetrends and risk factors for diabetes mellitus in catspresented to veterinary teaching hospitals. J Fel MedSurg 2007, doi:10.1016/j.jfms.2007.02.004.
Prentki M, Joly E, El-Assad W, et al. Malonly-CoA signaling, lipid partitioning, and glucolipotoxicity. Diabetes 2002; 51 suppl. 3:S405-S413.
Prentki M, Nolan CJ. Islet beta cell failure in type 2diabetes. J Clin Invest 2006; 116: 1802-1812.
Rand JS, Bobbermien LM, Hendrikz JK, et al. Overrepresentation of Burmese cats with diabetes mellitus.Aust Vet J. 1997; 75: 402-405..
Rand JS, Fleeman LM, Farrow HA, et al. Canineand feline diabetes mellitus: nature or nurture? J Nutr 2004; 134 suppl.: 2072S-2080S.
Rand JS, Marshall RD. Diabetes mellitus in cats.Vet Clin North Am Small Anim Pract 2005; 35:211-224.
Rand J. Editorial: Glargine, a new long-acting insulin analog for diabetic cats. J Vet Intern Med2006; 20: 219-220.
Randle PJ. Regulatory interactions between lipidsand glucides: the glucose fatty acid cycle after 35years. Diabetes Metab Rev 1998; 14: 263-283.
Reaven GM. The insulin resistance syndrome: definition and dietary approaches to treatment. An Rev Nutr 2005; 25: 391-406.

Reusch CE, Liehs MR, Hoyer M, et al.Fructosamine. A new parameter for diagnosis etmetabolic control in diabetic dogs and cats. J VetIntern Med 1993; 7: 177-182.
Reusch CE, Haberer B. Evaluation of fructosaminein dogs and cats with hypo- or hyperproteinaemia, azotaemia, hyperlipidaemia and hyperbilirubinaemia.Vet Rec 2001; 148: 370-376.
Reusch CE, Kley S, Casella M, et al.Measurements of growth hormone and insulin-likegrowth factor 1 in cats with diabetes mellitus. Vet Rec 2006a; 158: 195-200.
Reusch CE, Tschuor F, Kley S, et al. Diabetes mellitus in the cat: a review. Schweiz ArchTierheilk 2006b; 148: 130-138.
Richter M, Guscetti F, Spiess B. Aldose reductaseactivity and glucose-related opacities in incubatedlenses from dogs et cats. Am J Vet Res 2002; 63:1591-1597.
Rogers QR, Morris JG, Freedland RA. Lack ofhepatic enzymatic adaptation to low and high levelsof dietary protein in the adult cat. Enzyme 1977;22: 348-356.
Root MV, Johnston SD, Olson PN. Effect of prepuberaland postpuberal gonadectomy on heat production measured by indirect calorimetry in male and femaledomestic cats. Am J Vet Res 1996; 57: 371-374.
Rossetti L, Giaccari A, DeFronzo RA. Glucosetoxicity. Diabetes Care 1990; 13: 610-630.
Scarlett JM, Donoghue S, Saidla J. Overweight cats:prevalence and risk factors. Int J Obes 1994; 18suppl. 1: S22-S28.
Scarlett JM, Donoghue S. Associations betweenbody condition and disease in cats. J Am Vet MedAssoc 1998; 212: 1725-1731.
Schermerhorn T. Clinical aspects of feline hepaticglucose metabolism, in: Proceedings 23rd ACVIMForum 2005; 556-558.
Schermerhorn T. Pancreatic glucose sensing in dogsand cats, in: Proceedings 24th ACVIM Forum2006; 433-435.
Schuit FC, Huypens P, Heimberg H, et al. Glucosesensing in pancreatic beta-cells. Diabetes 2001; 50:1-11.
Slingerland LI, Robben JH, vanHaeften TW, et al.Insulin sensitivity and beta-cell function in healthycats: assessment with the use of the hyperglycemicglucose clamp. Horm Metab Res 2007; 39: 341-346.
Szabo J, Ibrahim WH, Sunvold GD, et al. Influenceof dietary protein and lipid on weight loss in obese ovariohysterectomized cats. Am J Vet Res 2000;61: 559-565.
Takeguchi A, Urabe S, Tanaka A, et al. Activitiesof enzymes in some types of peripheral leucocytesmay reflect the differences in nutrient metabolism between dogs and cats. Res Vet Sci 2005; 78: 21-24.
Tanaka A, Inoue A, Takeguchi A, et al.Comparison of expression of glucokinase gene and activities of enzymes related to glucose metabolism inlivers between dogs and cats. Vet Res Commun2005; 29: 477-485.
Thoresen SI, Bredal WP. Clinical usefulness of fructosamine measurements in diagnosing and monitoring feline diabetes mellitus. J Small AnimPract 1996; 37: 64-68.
Tschuor F, Furrer D, Kaufmann, et al. Intravenousarginine stimulation test in cats with transient et non-transient diabetes mellitus, in: Proceedings24th ACVIM Forum 2006.
Washizu T, Kuramoto E, Abe M, et al. A comparison of the activities of certain enzymesrelated to energy metabolism in leukocytes in dogsand cats. Vet Res Commun 1998; 22: 187-192.
Washizu T, Tanaka A, Sako T, et al. Comparisonof the activities of enzymes related to glycolysis andgluconeogenesis in the liver of dogs et cats. Res VetSci 1999; 67: 205-206.
Weaver KE, Rozanski EA, Mahony OM, et al. Useof glargine and lente insulins in cats with diabetesmellitus. J Vet Intern Med 2006; 20: 234-238.
Westermark P, Wernstedt C, O'Brien TD, et al.Islet amyloid in type 2 human diabetes mellitus andadult diabetic cats contains a novel putative polypep-tide hormone. Am J Pathol 1987; 127: 414-417.
Wilkins C, Long RC, Waldron M, et al. Assessmentof the influence of fatty acids on indices of insulinsensitivity and myocellular lipid content by use ofmagnetic resonance spectroscopy in cats. Am J VetRes 2004; 65: 1090-1099.
Williams DL, Heath MF. Prevalence of feline cataract: results of a cross-sectional study of 2000normal animals, 50 cats with diabetes and one hundredcats following dehydrational crises. Vet Ophthal2006; 9: 341-349.
Yano BL, Hayden DW, Johnson KH. Feline insularamyloid. Ultrastructural evidence for intracellularformation by nonendocrine cells. Lab Invest 1981;45: 149-56.
Zini E, Linscheid P, Franchini M, et al. Expressionof genes involved in glucose metabolism in non-diabetic diseased cats. Proceedings EuropeanCollege of Veterinary Internal Medicine 2006.
Réfé
renc
es
217
Diab
ète

Informations nutritionnelles Royal Canin
218
Diab
ète
L’arginine est un acide aminé indis-pensable chez le chat pour la syn-thèse de très nombreuses protéines.Il joue également un rôle dans desfonctions essentielles : - c’est un intermédiaire obligatoire
pour la synthèse de l’urée- il est précurseur de la synthèse du
monoxyde d’azote et d’amines bio-gènes
- enfin, il stimule la sécrétion de plu-sieurs hormones comme l’insuline,le glucagon et la gastrine.
Conséquences d’une carence en arginine
Si la croissance d’un chaton dépendévidemment d’un apport suffisant enarginine dans l’alimentation, il fautaussi rappeler que le chat adulte estextrêmement sensible à un déficitmême court en arginine. Lorsqu’ilreçoit un aliment riche en acides ami-nés, mais sans arginine, des signesd’intoxication ammoniacale appa-raissent 1 à 3 heures après la consom-mation du repas carencé : ptyalisme,vomissements, ataxie, hyperesthésie
et troubles nerveux (Morris et Rogers,1978). Dans certaines conditions l’in-toxication peut être mortelle.
La forte dépendance du chat àl’égard de l’arginine peut s’expliquerpar son excellente adaptation à unrégime carnivore, l’arginine étant eneffet abondante dans les protéinesd’origine animale. D’autre part, sil’ornithine et la citrulline sont desprécurseurs possibles de l’argininechez d’autres mammifères, le taux deconversion est trop bas chez le chatpour couvrir les besoins en arginine.
SCHÉMA DE LA SYNTHÈSE DE L’URÉE
Chez le chat, l'arginine est indispensable à la synthèse de l'urée.
Niveaux d’apports recommandés en argininechez le chat
Pour un chat adulte, le NRC (2006)recommande un niveau au moinségal à 0,77 % MS (pour une EM d’envi-ron 4000 kcal/kg) soit 1,93 g/1000 kcal.Plus le régime est riche en protéines,plus le niveau d’arginine doit êtreproportionnellement élevé (+ 0,02 gd’arginine par g de protéines au-des-sus du niveau minimal de 20 %).
L’arginine, un promoteurde la sécrétion insulinique
Chez les chats, les acides aminés etsurtout l’arginine stimulent considé-rablement la sécrétion pancréatiqued’insuline (Curry et coll, 1982).L’arginine agit en entraînant unedépolarisation directe de la mem-brane des cellules b du pancréas etinduit un flux d’ions calcium.
Cet effet stimulateur de la sécrétiond’insuline existe même en présenced’une glucotoxicité. Lors d’hypergly-
cémie chronique, la sécrétion d’insu-line en réponse à l’arginine est nor-male ou augmentée, alors que lasécrétion d’insuline en réponse auglucose est diminuée (Kitamura etcoll, 1999).
Ces observations pourraient expli-quer l’intérêt des régimes très richesen protéines chez les chats diabé-tiques. Un régime riche en arginine(>7 g/1000 kcal) contribue à stimulerla sécrétion insulinique et favorisel’obtention d’une rémission de lamaladie.
Gros plan sur :
l’arginine
Carbamyl-phosphate
Ornithine
Arginine
Citrulline
Urée
Iso-urée
Acide aspartique
ATP
AMP+PP
Acide argino-succinique
Acide fumarique

Informations nutritionnelles Royal Canin
219
Diab
ète
© C
.H
erm
elin
e/D
iffom
édia
(B
urm
ese)
Les maladies endocriniennes repré-sentent de plus en plus la nouvellefrontière de la médecine féline. Avecl’hyperthyroïdie, le diabète sucré estl’une des endocrinopathies les plusfréquentes chez le chat. Depuisquelques années, le nombre desujets obèses augmente régulière-ment dans la population féline etl’incidence des cas de diabète sucrésuit la même ascension.
L’obésité
Elle représente le principal facteur derisque: elle est responsable de ladiminution de la sensibilité périphé-rique à l’insuline. L’hyperinsulinémiecompensatrice peut alors entraînerun épuisement des cellules ß du pan-créas.
L’âge
La maladie survient le plus souventchez des individus d’âge mûr asso-ciant insulinorésistance (défaut d’ac-tion périphérique de l’insuline) etdéficit d’insulinosécrétion. Un pic estobservé vers 10-12 ans.
Le sexe
Les mâles semblent présenter unrisque accru vis-à-vis du diabète (enassociation avec l’obésité).
Les médicaments
Les glucocorticoïdes et les progesta-tifs de synthèse sont à l’origine d’unediminution de la sensibilité à l’insu-line.
Les maladies endocriniennes
Ex: acromégalie et hypercorticisme,maladies rares chez le chat.
Le diabète sucré est une maladiehétérogène caractérisée par unehyperglycémie marquée qui est laconséquence d’une anomalie desécrétion et/ou d’action de l’insuline.Par analogie avec ce qui est observéchez l’homme, la plupart des chatssemblent souffrir du diabète detype 2.
Les signes le plus souvent signaléspar les propriétaires sont listés ci-des-sous :- polyuro-polydipsie- perte de poids évoluant depuis plu-
sieurs semaines- anorexie- faiblesse, léthargie- vomissements- polyphagie
Parfois, malpropreté et troubleslocomoteurs (ex: difficulté à sauter)constituent des motifs de consulta-tion. Tous ces signes sont souventpeu marqués et s’installent très pro-gressivement.
Facteurs de risque de diabète sucré chez le chat
Sémiologie
Le diabète sucré est le plus souventobservé chez des chats d’âge muravec un pic vers 10-12 ans.
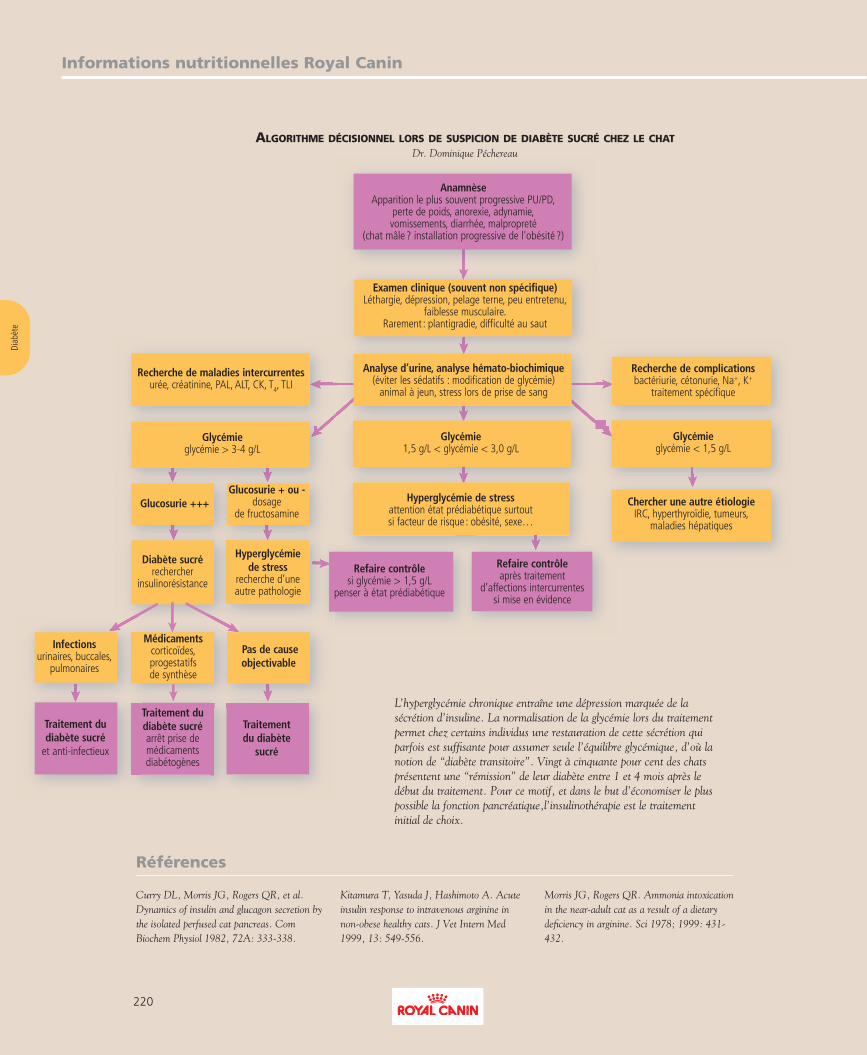
Informations nutritionnelles Royal Canin
220
Diab
ète
AnamnèseApparition le plus souvent progressive PU/PD,
perte de poids, anorexie, adynamie,vomissements, diarrhée, malpropreté
(chat mâle? installation progressive de l'obésité?)
Examen clinique (souvent non spécifique)Léthargie, dépression, pelage terne, peu entretenu,
faiblesse musculaire.Rarement : plantigradie, difficulté au saut
Analyse d’urine, analyse hémato-biochimique(éviter les sédatifs : modification de glycémie)
animal à jeun, stress lors de prise de sang
Recherche de maladies intercurrentesurée, créatinine, PAL, ALT, CK, T4, TLI
Recherche de complicationsbactériurie, cétonurie, Na+, K+
traitement spécifique
Glycémieglycémie < 1,5 g/L
Chercher une autre étiologieIRC, hyperthyroïdie, tumeurs,
maladies hépatiques
Refaire contrôleaprès traitement
d’affections intercurrentes si mise en évidence
Refaire contrôlesi glycémie > 1,5 g/L
penser à état prédiabétique
Traitement du diabète
sucré
Traitement dudiabète sucréarrêt prise demédicaments diabétogènes
Traitement dudiabète sucré
et anti-infectieux
Glycémie1,5 g/L < glycémie < 3,0 g/L
Glycémieglycémie > 3-4 g/L
Glucosurie + ou -dosage
de fructosamine
Diabète sucrérechercher
insulinorésistance
Infectionsurinaires, buccales,
pulmonaires
Médicamentscorticoïdes,progestatifs de synthèse
Pas de causeobjectivable
Hyperglycémie de stress
recherche d’uneautre pathologie
Hyperglycémie de stressattention état prédiabétique surtout si facteur de risque : obésité, sexe…
Glucosurie +++
L’hyperglycémie chronique entraîne une dépression marquée de la sécrétion d’insuline. La normalisation de la glycémie lors du traitementpermet chez certains individus une restauration de cette sécrétion qui parfois est suffisante pour assumer seule l’équilibre glycémique, d’où lanotion de “diabète transitoire”. Vingt à cinquante pour cent des chats présentent une “rémission” de leur diabète entre 1 et 4 mois après ledébut du traitement. Pour ce motif, et dans le but d’économiser le pluspossible la fonction pancréatique,l’insulinothérapie est le traitement initial de choix.
Curry DL, Morris JG, Rogers QR, et al.Dynamics of insulin and glucagon secretion bythe isolated perfused cat pancreas. ComBiochem Physiol 1982, 72A: 333-338.
Kitamura T, Yasuda J, Hashimoto A. Acuteinsulin response to intravenous arginine innon-obese healthy cats. J Vet Intern Med1999, 13: 549-556.
Morris JG, Rogers QR. Ammonia intoxicationin the near-adult cat as a result of a dietarydeficiency in arginine. Sci 1978; 1999: 431-432.
Références
ALGORITHME DÉCISIONNEL LORS DE SUSPICION DE DIABÈTE SUCRÉ CHEZ LE CHATDr. Dominique Péchereau

Informations nutritionnelles Royal Canin
221
Diab
ète
Traitement initial
Règles diététiques
Insulinothérapie (à privilégier),choix insuline (Caninsulin, NPH) + approche alimentaire
Glycémie < 4 g/Lcommencer par 0,25 U/kg 2 fois/jour
Glycémie > 4 g/Lcommencer par 0,5 U/kg 2 fois/jour
Traitement pendant 2-3 semainessurveillance : glycémie “à la maison”, glucosurie, fructosamine, appétit, consommation d’eau
Adaptation dose d’insulinenécessité d’une surveillance par le propriétaire (si glucosurie négative, diminuer la dose d’insuline)
Rythme de l’insulinothérapie
Donner la préférence à deux injec-tions journalières d’insuline, en véri-fiant l’activité de l’insuline choisie.• Ne jamais commencer par plus de
0,5 U/kg, 2 fois par jour (au moinsles 2 premières semaines).
• Bien veiller à l’éducation du pro-priétaire, pour permettre la réalisa-tion d’un traitement efficace: pra-tique de l’injection, respect de laposologie, site d’injection, signes àsurveiller (en particulier ceux del’hypoglycémie).
Suivi du traitement
Évolution de la glycémieLe propriétaire peut surveiller lui-même la glycémie grâce à un “gluco-mètre”. L’objectif est de maintenir laglycémie entre 1,2- 1,6 g/L. Si ellechute en-dessous de 1,2 g/L, il fautdiminuer la dose d’insuline.Contrôle de la quantité d’eau bue Paramètre très fiable en pratiquepour surveiller la polydipsie.Contrôle régulier de la glucosurie Il est important d’apprendre au pro-priétaire à bien utiliser et à lire unebandelette urinaire. Chez le chat, leseuil d’excrétion rénale du glucose se
situe entre 2 et 2,7 g/L de glycémie.Le suivi de la glucosurie permet sou-vent de bien gérer la posologie del’insuline lors de “diabète transi-toire”. Si la glucosurie reste négativepour plusieurs prélèvements consé-cutifs, la dose d’insuline peut êtrediminuée.
Dosage de la fructosamine et del’hémoglobine glycosyléeL’analyse de ces paramètres simplifiele contrôle par le propriétaire. Lafructosamine doit être maintenue endessous de 500 µmol/L et l’hémoglo-bine glycosylée en-dessous de 3 %.
aContrôler l’excès de poids
L’obésité constituant un facteurmajeur de risque d’insulinorésis-tance, il est primordial de proposerun aliment qui respecte les principessuivants : concentration énergétiqueet niveau de matières grasses modé-rés et richesse en protéines pourfavoriser une condition corporelleidéale ainsi que le maintien de lamasse maigre. La supplémentationen L-carnitine est également àconseiller pour faciliter l’utilisationdes acides gras et donc la perte depoids.
Minimiser la stimulationdes cellules b par le glucose
Les régimes très riches en protéines(> 45 % de la matière sèche ou MS)et contenant un niveau modéréd’amidon (< 20 % MS) issu d’unesource à faible index glycémique per-mettent de limiter les pics d’hyper-glycémie post-prandiale. De telsrégimes luttent contre l’insulino-résistance. La présence de fibres depsyllium, des fibres solubles quiretardent la vidange gastrique etrégularisent le transit digestif, estégalement intéressante pour ralentirl’assimilation du glucose.
Stimuler la sécrétion d’insuline endogène
Plusieurs acides aminés, l’arginine enparticulier, sont des promoteurs de lasécrétion endogène d’insuline chezle chat. Cela constitue un argumentsupplémentaire en faveur de l’utilisa-tion de régimes hyperprotéiques lorsde diabète sucré chez le chat.
Le respect de ces règles diététiquespeut permettre d’obtenir une dimi-nution des quantités d’insuline àadministrer, voire une rémission de lamaladie.


















