
Comportement hydrique du frêne (Fraxinus excelsior L) dans ...
18
HAL Id: hal-00882798 https://hal.archives-ouvertes.fr/hal-00882798 Submitted on 1 Jan 1992 HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci- entific research documents, whether they are pub- lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés. Comportement hydrique du frêne (Fraxinus excelsior L) dans une formation montagnarde mésoxérophile G Carlier, Jp Peltier, L Gielly To cite this version: G Carlier, Jp Peltier, L Gielly. Comportement hydrique du frêne (Fraxinus excelsior L) dans une formation montagnarde mésoxérophile. Annales des sciences forestières, INRA/EDP Sciences, 1992, 49 (3), pp.207-223. hal-00882798
Transcript of Comportement hydrique du frêne (Fraxinus excelsior L) dans ...

HAL Id: hal-00882798https://hal.archives-ouvertes.fr/hal-00882798
Submitted on 1 Jan 1992
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Comportement hydrique du frêne (Fraxinus excelsior L)dans une formation montagnarde mésoxérophile
G Carlier, Jp Peltier, L Gielly
To cite this version:G Carlier, Jp Peltier, L Gielly. Comportement hydrique du frêne (Fraxinus excelsior L) dans uneformation montagnarde mésoxérophile. Annales des sciences forestières, INRA/EDP Sciences, 1992,49 (3), pp.207-223. �hal-00882798�

Article original
Comportement hydrique du frêne(Fraxinus excelsior L) dans une formation
montagnarde mésoxérophile
G Carlier, JP Peltier L Gielly
Université Joseph Fourier, laboratoire de biologie alpine,BP 53 X, 38041 Grenoble cedex, France
(Reçu le 19 novembre 1991; accepté le 20 janvier 1992)
Résumé — L’article décrit les variations diurnes et saisonnières du potentiel hydrique foliaire, de laconductance stomatique et de la transpiration des feuilles d’un frêne dans une station montagnardemésoxérophile située dans la zone intermédiaire des Alpes nord-occidentales. Le potentiel hydriquede base est très proche du potentiel hydrique du sol à 30 cm de profondeur et en corrélation étroiteavec lui. En cas de sécheresse estivale sévère, le potentiel de base peut s’abaisser à -4,8 MPa et lepotentiel minimum à -5,5 MPa sans dommage pour les feuilles. En conditions favorables à l’alimen-tation en eau, la conductance stomatique maximale atteint 200 mmol[H2O] m-2s-1, valeur bien infé-rieure à celle des frênes des forêts alluviales de plaine. La conductance maximale ne se maintientque brièvement car une fermeture stomatique partielle a lieu au plus tard à midi (heure solaire) etéventuellement dès 7 h. En outre, la conductance maximale diminue quand le sol s’assèche. Enconséquence, le frêne de montagne consomme moins d’eau que le frêne de forêt alluviale. Cepen-dant, sa survie au cours des périodes de sécheresse est attribuée à la stratégie de tolérance plusqu’à celle d’évitement.
Fraxinus excelsior L = frêne / stomate / potentiel hydrique / sécheresse
Summary — Water relations of ash (Fraxinus excelsior L) in a mesoxerophilic mountainstand. This paper reports on the diurnal and seasonal variations in water potential, stomatal conduc-tance, and transpiration of the leaves of an ash tree in a mesoxerophilic mountain stand situated inthe intermediate zone of the north-western Alps. The pre-dawn water potential is close to the soilwater potential at 30 cm depth and is closely correlated with the latter. In the case of pronouncedsummer drought the pre-dawn water potential and minimum water potential of leaves may fall as lowas -4.8 MPa and -5.5 MPa, respectively. Under good water supply conditions the maximum stoma-tal conductance may reach 200 mmol (H2O) m-2.s-1, a value much lower than that of ash leaves inalluvial forests. The maximum stomatal conductance holds for only a short time, as stomata partiallyclose at noon (solar time) at the latest, and in some cases as early as 7 am. Moreover, the maxi-mum stomatal conductance decreases as the soil dries up. Accordingly ash water uptake is less inmountain stands than in alluvial forests. The survival of ash during drought is, however, attributed totolerance strategy rather than to avoidance strategy.
Fraxinus excelsior L = ash / stoma / water potential / water stress
*
Correspondance et tirés à part

INTRODUCTION
La croissance et la productivité du frênedépendent principalement de l’alimentationen eau à partir du sol (Devauchelle et
Lévy, 1977). Les sols doivent être bien
pourvus en eau, ni trop tassés, ni trop argi-leux, afin que les racines puissent s’enfon-cer jusqu’à 1 m ou 1,50 m (Le Goff et
Lévy, 1984). Le besoin en eau du frêne(masse d’eau consommée par unité dematière sèche produite) est plus élevé
(Braun, 1977) et la transpiration est plusintense (Ladefoged, 1963) que pourd’autres arbres présents dans les mêmesformations que le frêne, comme l’érable
sycomore.
Dans les Alpes nord-occidentales lefrêne se trouve principalement dans les fo-rêts alluviales à nappe phréatique de pro-fondeur modérée (profondeur moyenne :0,6-2,4 m; Pautou, 1970). Ce frêne deforêt alluviale est sujet à une transpirationtrès élevée pouvant atteindre 12,4 mmolm-2.s-1 (Besnard et Carlier, 1990), ce quiest dû à une conductance stomatique éle-vée (maximum : 800 mmol m-2.s-1), peusensible à la température et à l’humiditéatmosphérique. Cependant, le potentielhydrique foliaire ne s’abaisse jamais au-dessous de -2 MPa, valeur assez banalepour un feuillu de région tempérée (Hinck-ley et al, 1978).
En plus des frênaies alluviales, il existe,aux étages collinéen et montagnard infé-rieur, des frênaies à noisetier (Corylusavellana L) et à l’étage montagnard supé-rieur, des frênaies à bouleau (Betula pen-dula Roth) souvent installées sur deséboulis à sol peu épais et donc épisodi-quemert soumises à la sécheresse esti-vale (Pautou et al, 1991). Le présent tra-vail se rapporte à un frêne d’une telleformation montagnarde mésoxérophile.L’objectif est d’obtenir des informations
quantitatives sur les potentiels hydriques
atteints dans le sol et dans les feuilles, surla transpiration et la conductance stomati-que, afin de les comparer aux valeurs réa-lisées chez les frênes de forêt alluviale etde rechercher les modalités de l’adaptationdu frêne montagnard à son biotope.
MÉTHODES D’ÉTUDE
Site d’étude
Le site choisi se trouve sur la commune d’Huez
(Isère) à 1 350 m d’altitude. Les coordonnées
géographiques sont les suivantes : 45°4’34"N,6°3’21 "E. Du point de vue phytogéographique etbioclimatique, le site appartient à la zone
«intermédiaire» des Alpes nord-occidentales,définie par un angle de Gams compris entre 40°et 50° (Ozenda, 1985). La formation végétaleest une prairie de fauche (abandonnée) à Arrhe-natherum elatius (L) J et C Presl et Trisetum fla-vescens (L) PB largement colonisée par Agropy-rum campestre Godr et Gr et bocagée de frênes(Fraxinus excelsior L) et de quelques chênessessiles (Quercus petræa [Mattuschka] Lie-
blein). Les frênes sont traités en taillis; les cé-pées sont distantes de 10-15 m; les brins lesplus forts ont 25-60 ans et 8-12 m de haut. Lapente, orientée vers le SE, est d’environ 10%.
Le sol est un limon sableux remplissant lesinterstices d’un éboulis de gros blocs schisteux,qui n’ont pas permis de creuser à plus de 60cm; l’enracinement observé ne dépasse pas 40cm de profondeur.
Potentiel hydrique du sol
Le potentiel hydrique du sol a été mesuré, selonla méthode du point de rosée, au moyen desondes hygrométriques Wescor PCT 55 placéeshorizontalement (Bruckler, 1984) aux profon-deurs de 5, 15, 30, 45 et 60 cm et reliées à unmicrovoltmètre Wescor HR-33 T. Les mesuresétaient faites avant le lever du soleil. Lors-
qu’elles ont été répétées au cours d’une mêmejournée, il n’a pas été constaté de variation, saufà 5 cm où l’échauffement diurne rend les me-sures invalides.

Conductance stomatique et potentielhydrique foliaire
La conductance stomatique des folioles a étédéterminée à l’aide d’un poromètre Li-Cor 1600,uniquement sur la face adaxiale (Besnard et
Carlier, 1990). La résistance, affichée en s.cm-1,est convertie en conductance gs (mmol m-2s-1)suivant Körner et Cochrane (1985). Immédiate-ment après la mesure porométrique la folioleétait détachée et son potentiel hydrique ψf me-
suré à l’aide d’une bombe de pression (Scholan-der et al, 1965). Toutes les heures 5 folioles ap-partenant à des feuilles différentes étaienttraitées successivement. De telles séries ho-raires étaient répétées 11 à 14 fois au cours dela journée. Toutes les folioles utilisées étaientadultes et ont été choisies dans le quadrant sudd’une même cépée, entre 2,5 et 3,5 m de hau-teur sur des brins de 15 cm de diamètre aumoins et d’environ 9 m de haut.
Conditions microclimatiques
Au début et à la fin de chaque série horaire, latempérature et l’humidité de l’air étaient détermi-nées à proximité du feuillage (mais à l’abri dusoleil) à l’aide d’un thermo-hygromètre Coreci,régulièrement recalé suivant la notice du cons-tructeur. L’éclairement (ou irradiance, en μmol[photons] m-2.s-1) reçu par chaque foliole, per-pendiculairement au limbe, était mesuré à l’aided’un capteur Li-Cor 190-SB installé sur le poro-mètre.
Les précipitations indiquées sont celles duposte météorologique de Bourg d’Oisans, dis-tant de 4 km.
Calcul de la transpiration
La transpiration mesurée par le poromètre estconsidérée comme différente de la transpirationréelle de la foliole à l’air libre, car l’atmosphèrede la cuvette est plus chaude et plus sèche quel’air ambiant. La transpiration (Ecal, en mmol.m-2.
s-1) a donc été calculée en tenant compte de lapression de vapeur d’eau dans l’air, de la pres-sion de vapeur saturante à la température de la
foliole et de la résistance stomatique donnéepar le poromètre; la résistance de la couche li-mite a été prise égale à celle existant dans leporomètre, soit 0,2 s.cm-1.
Les valeurs retenues pour la conductance
stomatique, la transpiration et le potentiel hydri-que sont les moyennes de chaque série horaire.
Relations entre RWC et potentiels
Les relations entre la teneur en eau relative
(RWC %, Turner et Kramer, 1980) et les poten-tiels hydrique ψf et osmotique ψs ont été déter-minées, à des dates choisies, sur des lots de fo-lioles. Les folioles ont d’abord été saturées (ψf =
0, RWC = 100%) par séjour de 48 h en atmos-phère saturée, à 5 °C, la base du limbe étant im-mergée dans de l’eau distillée. Plusieurs foliolessaturées ont été tuées par immersion dansl’azote liquide et leur potentiel osmotique à l’étatsaturé ψso a été déterminé dans des chambresen inox munies de sondes psychrométiquesWescor PST 55 reliées à un microvoltmètreWescor PR 55 (une foliole par chambre). Les fo-lioles restantes ont été amenées à différents ni-veaux de RWC et on a mesuré leur potentiel hy-drique ψf puis, après passage dans l’azote
liquide, leur potentiel osmotique ψs. Dans unesérie décroissante de RWC, le premier RWCpour lequel ψs = ψf indique, par défaut, le débutde perte de turgescence; le potentiel correspon-dant est ψsl.
Tests statistiques
Les corrélations ont été éprouvées par le test dePearson ou, lorsque celui-ci est illicite, par letest de Kendall (Sokal et Rohlf, 1981).
RÉSULTATS
Les conditions hydriques du milieuet le potentiel foliaire de base (fig 1)
Les mesures ont commencé le 16 août1988 à la suite d’une période de 6 se-

maines caractérisée par des précipitationsréduites; le sol était sec (ψsol = -2,73 MPaà 30 cm de profondeur le 16 août 1988) etle gradient des potentiels hydriques crois-sant vers la profondeur. La pluie du 20août 1988 a fait remonter les potentiels, in-stantanément en surface et plus progressi-vement en profondeur.
La campagne de mesures de 1989 acommencé fin mai, sur sol bien humide ensurface avec un gradient de potentiel légè-rement décroissant vers la profondeur.L’été n’a comporté que des précipitationsminimes de juillet à septembre, l’assèche-ment du sol à partir de la surface entraî-nant l’inversion du gradient. En fin de sai-son, le sol était extrêmement sec (ψsol = -5,09 MPa à 30 cm de profondeur le 27
septembre 1989). L’été 1990 a connu unesécheresse moins poussée (ψsol = -1,95MPa à -30 cm le 7 septembre 1990).
Les mesures physiologiques ont doncpu être faites dans un intervalle très largede conditions hydriques édaphiques. Le
potentiel de base des feuilles ψb suit lesvariations de potentiel du sol. En fait il est,sauf rares exceptions, compris entre les
potentiels du sol à 30 et à 45 cm de pro-fondeur (fig 1). Le ψb est très étroitement
corrélé au ψsol à -30 cm (r = 0,984; n =11; P < 0,01). Les corrélations de ψb avec
les ψsol aux autres profondeurs sont moinsbonnes. Il n’y a pas de corrélation entre ψbet le ψsol à -60 cm, profondeur à laquelle il
n’a pas été observé de racines.

Même quand ψb était au plus bas (-3,16MPa le 18 août 1988 et -4,80 MPa le 27
septembre 1989) les feuilles paraissaientturgescentes et ne présentaient pas designes de dommages. Cependant quel-ques frênes voisins ont subi des dom-
mages se manifestant pas le brunissementde certaines feuilles.
Schémas des variations diurnesdes grandeurs physiologiques
Toutes les journées de mesures se sontdéroulées par temps bien à très bien enso-leillé. La figure 2 donne 4 exemples de va-riations des grandeurs physiologiques,choisis pour couvrir l’ensemble des condi-tions hydriques rencontrées, des plus mo-dérées (ψb = -0,48 MPa le 31 mai 1989)aux plus sévères (ψb = -4,8 MPa le 27
septembre 1989). Les valeurs extrêmes
pour toutes les journées sont portées autableau I.
La conductance stomatique, faible maisrarement nulle avant le lever du soleil, aug-mente avec l’éclairement et culmine dans
la matinée (fig 2). Le maximum de conduc-tance tend à être plus précoce en périodesèche qu’en période humide; par exempleil s’est produit à 8 h le 18 août 1988 (ψb =
-3,16 MPa) et à 12 h le 31 mai 1989 (ψb =
-0,48 MPa). La conductance décroît en-suite de 30 à 75%, en 1-2 h, tandis quel’éclairement continue à augmenter. Ellereste alors plus ou moins constante ou dé-croît doucement jusqu’à la fin de la jour-née, où la fermeture des stomates s’accen-tue. Le 27 septembre 1989 (ψb = -4,8MPa) les stomates se sont très peu (et ce-pendant significativement) ouverts et lemoment du maximum de conductance est
imprécis.Le maximum de transpiration se produit
en milieu de journée. Il est toujours posté-rieur au maximum de conductance, ce quirésulte évidemment du fait que la tempéra-ture et le VPD atmosphériques continuentà augmenter alors que la conductance di-minue. Quant au minimum de potentiel hy-drique foliaire, il est atteint en milieu de
journée ou plus tard. Il coïncide le plussouvent avec le maximum de transpirationà ± 1 h près (ce qui représente le pas detemps des mesures). Cependant les 18 et


31 août 1988, le minimum de ψf est nette-
ment plus tardif mais cela correspond àdes courbes de variation de la transpira-tion très aplaties.
Relation de la conductanceavec l’éclairement reçu par les feuilles
Si, pour chaque journée, on traçait le
graphe des points représentant la conduc-tance en fonction de l’éclairement sanstenir compte de leur ordre chronologique,ces points s’ajusteraient, avec une disper-sion notable, à des courbes d’allure hyper-bolique comportant un plateau quasi hori-zontal atteint pour des éclairements
compris entre 400 et 800 μmol (pho-tons)m-2.s-1. Mais si on relie les
moyennes horaires dans l’ordre chronolo-
gique (fig 3) on obtient des boucles quipeuvent être décomposées en 5 segmentscomme suit :- a) une phase d’ouverture d’une durée de2-3 h; nous appelons «efficacité lumi-neuse» la pente moyenne de ce segment;- b) un plateau correspondant approximati-vement à la conductance maximale et pou-vant se réduire à celle-ci (par exemple le18 août 1988, fig 3);- c) un décrochement constitué par la di-minution de conductance sous éclairementconstant ou croissant et réalisé en 1-2 h;- d) une phase de «conductance stabili-
sée», sous éclairement décroissant, quidure plusieurs h jusqu’à ce que l’éclaire-ment retombe au voisinage de 200 μmol(photons) m-2.s-1;- e) la fermeture finale dont le tracé se
confond avec celui de l’ouverture matinale.
Cette phase n’est observée que si les me-sures sont prolongées suffisamment tard.
Tout se passe comme s’il existait 2 rela-tions distinctes de gs avec l’éclairement,l’une s’appliquant le matin jusqu’au décro-chement (a, b) et l’autre après le décroche-ment (d, e). Le 27 septembre 1989 lesconductances sont tellement faibles queles 2 relations sont confondues. Le pas-sage d’une relation à l’autre suggère l’inter-vention, directe ou indirecte, de facteursautres que l’éclairement.
Relations de la conductance avecdes variables autres que l’éclairement
L’étude des conditions microclimatiques(température, VPD) existant au moment où

la conductance est maximale ou au débutdu «décrochement» (les 2 étant occasion-nellement confondus) ne fait apparaîtreaucune coïncidence remarquable (résul-tats non présentés). En revanche plu-sieurs corrélations significatives, voire trèssignificatives, ont été mises en évidence(tableau II):- la conductance maximale gsmax diminuetrès significativement avec le potentiel debase ψb (τ = 0,778**); elle diminue aussiavec le potentiel hydrique foliaire qui lui
est concomitant; mais cette relation est
moins étroite que la précédente (τ =
0,666*);- la «conductance stabilisée» est trèsétroitement corrélée au potentiel de base(τ = 0,911**); cette relation peut n’être pasindépendante de la précédente car la
conductance maximale et la conductancestabilisée sont elles-mêmes en étroite cor-rélation (τ=0,778**);
- l’efficacité lumineuse décroît avec le ψb(τ = 0,688*); cette relation est indépen-dante de celle qui relie gsmax à ψb , car
gsmax n’est pas corrélée avec l’efficacité lu-mineuse (autrement dit, ce n’est pas parceque la conductance augmente vite à partirdu lever du soleil qu’elle atteindra une va-leur maximale élevée).
Ainsi la sécheresse entraîne une dimi-nution de l’ensemble des valeurs de la
conductance et une détérioration de la ré-
ponse des stomates à l’éclairement.
Relation entre le potentiel ψfet la transpiration Ecal (fig 4)
Les courbes exprimant cette relation s’éta-gent exactement dans l’ordre des poten-tiels de base, ce qui montre qu’en toute cir-constance le ψf dépend d’abord desconditions édaphiques et ensuite seule-ment de la transpiration.


L’augmentation de la transpiration aucours de la journée, même si elle est mi-nime (ex: le 27 septembre 1989), est tou-jours accompagnée d’une chute de poten-tiel. L’écart Δψ entre le potentiel de base etle potentiel minimal ne varie pas significati-vement d’une journée à l’autre (tableau I)alors que la transpiration décroît de façonévidente quand la sécheresse s’accentue.
En conditions de sécheresse modérée
(ψb > -1,5 MPa) le potentiel minimal coïn-cide presque exactement avec la transpi-ration maximale et, à transpiration égale,le potentiel est sensiblement le mêmeavant e après ce point extrême : la rela-tion entre ψf et Ecal ne comporte pasd’hystérésis. Quand les conditions sont
plus sévères (ψb < -1,5 MPa) le potentielminimal peut être postérieur à la transpira-tion maximale et un effet d’hystérésis peutapparaître.
Toutes les régressions linéaires de ψfsur Ecal sont significatives. Dans quelquescas, les points s’ajustent mieux à une rela-tion quadratique, donc curvilinéaire, qu’àune relation linéaire; mais en aucun cas il
n’y a de différence significative entre les 2coefficients de corrélation. En consé-
quence c’est la pente de la relation linéairequi est considérée comme représentant larésistance hydraulique sol-feuille (Reich etHinckley, 1989). L’inverse de cette résis-tance est la conductance hydraulique sol-feuille (tableau II). De façon hautement si-gnificative, la conductance hydrauliquesol-feuille diminue (fig 5) avec le potentielde base ψf(τ= 0,943**).
Relation entre le potentiel hydriquefoliaire ψf et la conductance
stomatique gs (fig 6)
Dans tous les cas, l’augmentation de gsest accompagnée d’une chute de ψf. Lesdifférences concernent les événements
postérieurs au maximum de gs. En bonnesconditions d’humidité du sol (le 31 mai1989 ainsi que le 14 juin 1989, non pré-senté) le ψf ne décroît pas, ou à peine,au-dessous de la valeur qu’il a atteinte aumoment du maximum de gs; la réductionde la conductance freine suffisamment la
transpiration pour stopper la chute de ψf.Au contraire, quand les conditions hydri-ques édaphiques sont plus sévères (ex: le12 septembre 1988 et le 18 août 1988) leψf continue de décroître pendant que lesstomates se ferment et cette décroissance
supplémentaire peut atteindre -0,7 MPa;la réduction de conductance ne freine passuffisamment la transpiration pour stopperla chute de ψf . En conditions de séche-resse extrême (le 27 septembre 1989) le
blocage de ψf réapparaît.Le caractère le plus remarquable de la
relation de ψf à gs (fig 6) est que la mêmeconductance stomatique peut être réaliséepour des potentiels extrêmement diffé-
rents, suivant les conditions hydriques dusol. Ceci suggère qu’un ajustement osmoti-que se produit au cours des périodes desécheresse.

Indices de l’ajustement osmotique
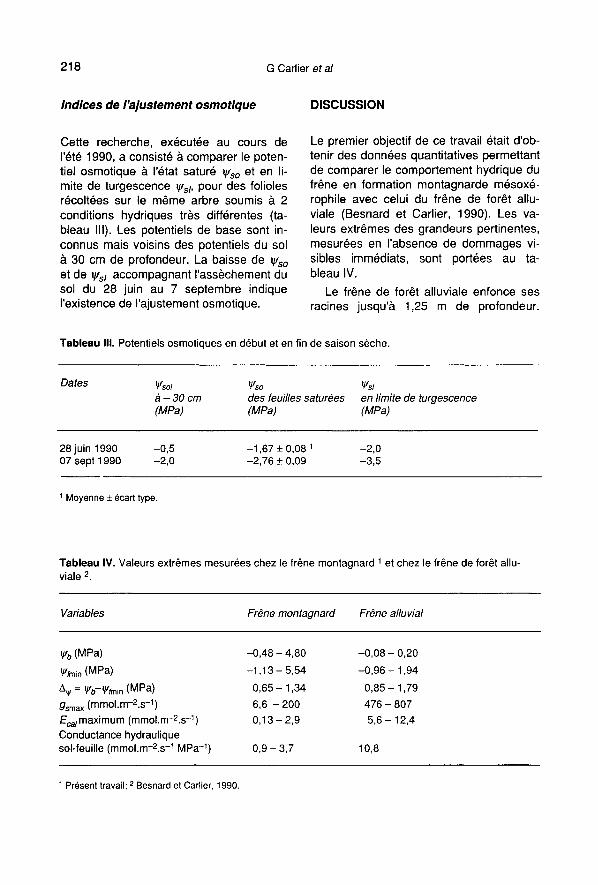
Cette recherche, exécutée au cours del’été 1990, a consisté à comparer le poten-tiel osmotique à l’état saturé ψso et en li-mite de turgescence ψsl, pour des foliolesrécoltées sur le même arbre soumis à 2conditions hydriques très différentes (ta-bleau III). Les potentiels de base sont in-connus mais voisins des potentiels du solà 30 cm de profondeur. La baisse de ψsoet de ψsl accompagnant l’assèchement dusol du 28 juin au 7 septembre indiquel’existence de l’ajustement osmotique.
DISCUSSION
Le premier objectif de ce travail était d’ob-tenir des données quantitatives permettantde comparer le comportement hydrique dufrêne en formation montagnarde mésoxé-rophile avec celui du frêne de forêt allu-viale (Besnard et Carlier, 1990). Les va-leurs extrêmes des grandeurs pertinentes,mesurées en l’absence de dommages vi-sibles immédiats, sont portées au ta-
bleau IV.
Le frêne de forêt alluviale enfonce sesracines jusqu’à 1,25 m de profondeur.

Dans la zone des racines, le potentiel hy-drique du sol ne tombe jamais au-dessousde -0,1 MPa, grâce aux remontées ca-pillaires s’effectuant à partir de la nappephréatique. Le frêne montagnard installésur éboulis de blocs enfonce ses racines
jusqu’à 0,4 m et prélève l’eau du sol au voi-sinage de 0,3 m où le potentiel peut, encas de sécheresse prolongée, s’abaisser à- 5 MPa. En conséquence, le frêne de forêtalluviale n’est jamais soumis à la séche-resse édaphique alors que le frêne monta-gnard l’a été, dans le biotope choisi ici, 3étés de suite (1988, 1989, 1990).
Le frêne de plaine est réputé gros con-sommateur d’eau (Ladefoged, 1963; Aus-senac et Lévy, 1983), réputation méritéeégalement par le frêne de forêt alluviale
(tableau IV). Le frêne montagnard est
beaucoup plus sobre, même en bonnesconditions d’humidité du sol, et restreintnotablement sa transpiration en période desécheresse. Cette différence repose sur
celle des conductances stomatiques maxi-males (tableau IV) et sur la cinétiquediurne de l’ouverture stomatique. Le frênede forêt alluviale garde ses stomates
grand ouverts une bonne partie de la jour-née, sauf en cas de diminution notable del’ensoleillement; le frêne montagnard les
ferme partiellement, au plus tard à midi ethabituellement beaucoup plus tôt (ta-bleau I).
Les jours chauds et secs où sa transpi-ration est intense, le frêne de forêt alluvialesubit une chute abrupte du potentiel hydri-que foliaire sans qu’intervienne aucune ré-gulation stomatique. Cependant, grâce auxbonnes conditions édaphiques et à uneconductance hydraulique sol-feuille remar-quablement élevée, le ψf minimal ne
tombe pas au-dessous de -2 MPa. Lefrêne montagnard fait l’inverse. Sous l’effetdominant de la sécheresse du sol, le ψf de
ses feuilles peut atteindre des valeurs trèsbasses, plus proches de celles d’arbres de
régions arides ou désertiques (Roberts etal, 1981; Nilsen et al, 1984) que de cellesde feuillus de régions tempérées (Hinckleyet al, 1978). En revanche, la chute diurnede ψf est plus modérée que celle du frênede forêt alluviale, grâce à la fermeture par-tielle des stomates; toutefois cette régula-tion n’a qu’une efficacité limitée puisqu’àcompter de son déclenchement, il peut en-core se produire une chute de potentiel at-teignant -0,7 MPa.
Au vu de ces différences de comporte-ment entre frêne de forêt alluviale et frênede montagne, une étude génétique compa-rée serait justifiée.
Un second domaine de discussionconcerne la signification adaptative des
propriétés mises en évidence.Les stomates s’ouvrent d’autant moins
que la sécheresse est plus prononcéecomme le montre la corrélation très signifi-cative entre la conductance maximale et le
potentiel de base, déjà établie pourd’autres arbres (Reich et Hinckley, 1989).En outre, quand le ψb s’abaisse, l’efficacitéde l’éclairement sur l’ouverture stomatiquediminue (tableau III). Davies et Kozlowski(1975) ont montré que la réponse des sto-mates aux changements d’éclairement estinfluencée négativement par la détresse
hydrique chez plusieurs arbres, à des de-grés divers selon l’espèce (Fraxinus ameri-cana est relativement peu sensible).
Ces éléments du comportement du
frêne de montagne contribuent à limiter latranspiration mais ne limitent que très im-parfaitement la diminution de ψf . SelonAussenac et Granier (1978) la chutediurne de potentiel foliaire Δψ est limitée à0,4 MPa quand la transpiration est exclusi-vement cuticulaire; des arbres méditerra-néens ont même un Δψ nul en période es-tivale (Morrow et Mooney, 1974). Cheznotre frêne montagnard, cette situation n’ajamais été observée: même en cas de sé-

cheresse extrême, une ouverture stomati-que significative se produit et Δψ atteint0,74 MPa.
Une autre propriété peu favorable à lastabilisation du ψf réside dans la conduc-tance hydraulique sol-feuille (tableau IV).Non seulement, celle-ci, dans lesmeilleures conditions, est 3 fois plus faiblechez le frêne montagnard que chez le
frêne de forêt alluviale, ce qui, à transpira-tion égale, favorise la chute du ψf, maisencore on observe une diminution pro-gressive de cette conductance quand lasécheresse s’accentue. Selon Reich et
Hinckley (1989), cette diminution de laconductance hydraulique peut être due àl’augmentation de la résistance hydrauli-que du sol et à des mécanismes propres àl’arbres et donc susceptibles de variationsselon l’espèce et le milieu, tels que la dis-parition des fines racines et la cavitation.Ce dernier mécanisme serait prépondé-rant chez la vigne (Schultz et Matthews,1988).
Ainsi, parmi les stratégies adaptativesdéfinies par Ludlow (1989), le frêne mon-tagnard ne pratique pas celle de l’évite-ment puisqu’il ne possède pas de dispositifadéquat pour limiter la chute de son poten-tiel hydrique foliaire. Il survivrait donc aux
épisodes de sécheresse estivale par la
stratégie de tolérance. On peut observerque notre frêne a supporté sans dom-mage, le 27 septembre 1989, des poten-tiels foliaires allant de -4,8 à -5,5 MPapendant que d’autres individus du mêmebiotope subissaient des dommages (cons-tatés plusieurs jours plus tard). Le poten-tiel critique défini par Levitt (1963), corres-pondant à un seuil létal, a donc été
approché.L’un des éléments de la stratégie de to-
lérance est l’ajustement stomatique,«processus par lequel les réponses sto-matiques sont modifiées pour être appro-priées à la nouvelle situation de détresse
hydrique» (Ludlow, 1980). Le frêne de
montagne pratique l’ajustement stomatiquepuisque la relation entre gs et ψf se modi-fie avec l’avancement de la sécheresse (fig6). Un mécanisme possible de l’ajustementstomatique est l’ajustement osmotique(Ludlow, 1980; Hinkley et al, 1980). L’ajus-tement osmotique favorise le maintien dela turgescence. Il est mesuré par l’abaisse-ment du potentiel osmotique dans des si-tuations précises, par exemple à pleine tur-gescence ou à la limite de perte de
turgescence, le second critère étant plussensible (Turner et Jones, 1980). C’estbien ce qui se produit chez le frêne de
montagne (tableau III). L’ajustement osmo-tique a lieu chez l’abricotier (Loveys et al,1987) et aussi, en même temps que l’ajus-tement stomatique, chez Cornus sangui-nea, alors que l’un et l’autre manquentchez d’autres arbres, dont l’olivier (Hinck-ley et al, 1980).
Un dernier point à discuter se rapporteau déterminisme des mouvements stomati-
ques, mais il ne faut pas perdre de vuequ’un travail comme celui-ci, effectué enconditions naturelles incontrôlables, n’est
pas particulièrement approprié à la miseen évidence des facteurs responsables.
L’éclairement intervient de façon pré-pondérante en début et en fin de journée.L’efficacité lumineuse, qui mesure la sensi-bilité des stomates à l’augmentation mati-nale de l’éclairement, est en corrélation si-gnificative avec le potentiel de base
(tableau II) comme l’est aussi, de façontrès significative, la conductance maxi-male. Ces faits peuvent s’interpréter, à lasuite de Gollan et al (1985), comme indi-quant une dépendance du fonctionnementstomatique par rapport aux conditions hy-driques régnant dans le sol au voisinagedes racines. Le signal transmis des ra-
cines aux feuilles pourrait être constitué
par la variation de concentration de l’acide
abscissique dans la sève brute (Wartinger

et al, 1990). Cependant Reich et Hinckley(1989) suggèrent que le signal pourraitêtre constitué par la valeur de la conduc-tance hydraulique sol-feuille. Effective-ment, chez le frêne montagnard, il existedes corrélations très significatives, d’unepart entre la conductance stomatiquemaximale et la conductance hydrauliquesol-feuille (r = 0,833**), d’autre part entrela conductance hydraulique sol-feuille et lepotentiel de base (τ = 0,943**; tableau II,fig 5).
Par ailleurs, la cinétique diurne de laconductance stomatique est dominée parl’existence d’un «décrochement» dans larelation entre conductance et éclairement
(phase c, fig 3). Quel facteur en est res-ponsable? Une littérature abondante se
rapporte à la détermination du potentiel fo-liaire «seuil» au-delà duquel la fermeturedes stomates s’amorce (Hinckley et al,1978) ce qui implique que le potentiel fo-liaire agit instantanément sur les stomates(Ludlow, 1980), hypothèse récusée parGollan et al (1985) et par Wartinger et al(1990). Dans le cas du frêne montagnard,il n’est pas possible d’associer le«décrochement» à une valeur fixe de ψf.Les recherches relatives à l’impact de
l’ABA, apporté aux feuilles par la sève
brute, sur la cinétique diurne des stomatesn’ont pas permis de conclure positivement(Loveys et Düring, 1984; Zhang et Davies,1989). Enfin, l’influence du VPD atmosphé-rique sur les stomates a été démontréedans de nombreux cas (Levy, 1980; Kör-ner et Cochrane, 1985) mais nos donnéesne font pas apparaître d’effet de ce genrechez le frêne de montagne, pas plus quechez le frêne de forêt alluviale (Besnard etCarlier, 1990).
CONCLUSION
Le frêne montagnard mésoxérophile est
moindre gaspilleur d’eau que le frêne de
forêt alluviale, grâce à une ouverture sto-matique moins grande et moins durable, etsensible à l’assèchement du sol. Dans unecertaine mesure, il ménage la réserved’eau limitée présente dans le sol peuabondant des éboulis qu’il occupe. Toute-fois, son maintien dans de tels biotopes,au cours des épisodes de sécheresse esti-vale fréquents dans les Alpes intermé-
diaires, ne repose pas tant sur l’évitementque sur des propriétés de tolérance dontles limites restent à déterminer.
REMERCIEMENTS
Nous remercions M le Maire d’Huez-en-Oisans
qui a facilité la recherche du site, le Dr B Dochequi nous a conseillés dans le choix de celui-ci,et Mme E Sarret qui a autorisé les travaux expé-rimentaux sur la parcelle dont elle est proprié-taire.
RÉFÉRENCES
Aussenac G, Granier A (1978) Quelques résul-tats de cinétique journalière du potentiel desève chez les arbres forestiers. Ann Sci For
35, 19-32
Aussenac G, Lévy G (1983) Influence du dessè-chement du sol sur le comportement hydri-que et la croissance du chêne pédonculé(Quercus pedunculata Ehrl) et du frêne
(Fraxinus excelsior L) cultivés en cases devégétation. Ann Sci For 40, 251-264
Besnard G, Carlier G (1990) Potentiel hydriqueet conductance stomatique des feuilles defrêne (Fraxinus excelsior L) dans une forêt al-luviale du Haut-Rhône français. Ann Sci For47, 353-365
Braun HJ (1977) Growth and water economy ofthe trees Acer pseudoplatanus L and Fraxi-nus excelsior L. Z Pflanzenphysiol 84, 459-462
Bruckler L (1984) Utilisation des micropsychro-mètres pour la mesure du potentiel hydriquedu sol en laboratoire et in situ. Agronomie 4,171-182

Davies WJ, Kozlowski TT (1975) Stomatal res-ponses to changes in light intensity as in-fluenced by plant water stress. For Sci 21,129-133
Devauchelle R, Lévy G (1977) Propriétés sta-tionnelles et croissance du frêne dans l’Estde la France, étude de certaines caractéristi-ques de cette essence. Ann Sci For 34, 231-244
Gollan T, Turner NC, Schulze ED (1985) Theresponse of stomata and leaf gas exchangeto vapor pressure deficits and soil water
content. 3. In the sclerophyllous woody spe-cies Nerium oleander. Œcologia 65, 356-362
Hinckley TM, Lassoie JP, Running SW (1978)Temporal and spatial variations in the waterstatus of forest trees. For Sci 24, Monograph20, 79 p
Hinckley TM, Duhme F, Hinckley AR, Richter H(1980) Water relations of drought hardyshrubs: osmotic potential and stomatal reac-tivity. Plant Cell Environ 3, 131-140
Körner C, Cochrane PM (1985) Stomatal res-ponses and water relations of Eucalyptuspauciflora along an elevational gradient.Œcologia 6, 443-455
Ladefoged K (1963) Transpiration of forest treesin closed stands. Physiol Plant 16, 378-414
Le Goff N, Lévy G (1984) Productivité du frêne(Fraxinus excelsior L) en région Nord-Picardie. B. Étude des relations entre la pro-ductivité et les conditions de milieu. Ann SciFor 41, 135-170
Levitt J (1963) The measurement of drought re-sistance. In: Methodology of plant ecophysio-logy (Eckardt FE, ed) Proc MontpellierSymp, Unesco 407-412
Levy Y (1980) Effect of evaporative demand onwater relations of Citrus limonum. Ann Bot46, 695-700
Loveys BR, Düring H (1984) Diurnal changes inwater relations and abscissic acid in field-
grown Vitis vinifera cultivars. New Phytol 97,37-47
Loveys BR, Robinson SP, Downton WJS (1987)Seasonal and diurnal changes in abscissicacid and water relations of apricot leaves(Prunus armeniaca L). New Phytol 107,15-27
Ludlow M (1980) Significance of stomatal res-ponse to water stress. In: Adaptation of
plants to water and high temperature stress(Turner NC, Kramer PJ, eds) New York 123-138
Ludlow M (1989) Strategies of response to
water stress. In: Structural and functional res-
ponses to environmental stresses. (KreebKH, Richter H et Hinckley TM, eds) SPBAcad Publ bv, La Haye, 269-281
Morrow PA, Mooney HA (1974) Drought adapta-tion in two California evergreen sclerophylls.Œcologia 15, 205-222
Nilsen ET, Sharf MR, Rundel PW (1984) Com-parative water relations of phreatophytes inthe Sonoran desert of California. Ecology 65,767-778
Ozenda P (1985) La végétation de la chaîne al-pine dans l’espace montagnard européen.Masson, Paris, 25
Pautou G (1970) Écologie des formations allu-viales de la Basse Isère. Application à l’étuded’une nappe phréatique et de ses risques depollution. Documents pour la carte de la vé-gétation des Alpes VIII, 73-114
Pautou G, Cadel G, Girel R (1992) Le bassin deBourg d’Oisans, un carrefour phytogéogra-phique des Alpes françaises. Rev Ecol Al-
pine 1, 23-43Reich PB, Hinckley TM (1989) Influence of pre-
dawn water potential and soil-to-leaf hydrau-lic conductance on maximum daily leaf diffu-sive conductance in two oak species. FunctEcol 3, 719-726
Roberts SW, Miller PC, Valamanesh A (1981)Comparative field water relations of four co-occurring chaparral species. Œcologia 48,360-363
Scholander PM, Hammel HT, Bradstreet ED,Hemmingsen EA (1965) Sap pressure in vas-cular plants. Science 148, 339-346
Schultz HR, Matthews MA (1988) Resistance towater transport in shoots of Vitis vinifera L.Relation to growth at low water potential.Plant Physiol 88, 718-724
Sokal RR, Rohlf FJ (1981) Biometry. The prin-ciples and practice of statistics in biologicalresearch. Freeman and Co, 859 p

Turner NC, Jones MM (1980) Turgor mainte-nance by osmotic adjustment: a review andevaluation. In: Adaptation of plants to waterand high temperature stress (Turner NC, Kra-mer PJ, eds) New York, 87-104
Turner NC, Kramer PJ (eds) (1980) Adaptationof plants to water and high temperaturestress. New York, 1-5
Wartinger A, Heilmeier H, Hartung W, SchulzeED (1990) Daily and seasonal courses of leafconductance in the xylem sap of almondtrees (Prunus dulcis [Miller] DA Webb) underdesert conditions. New Phytol 116, 581-587
Zhang J, Davies WJ (1989) Sequential res-
ponse of whole plants water relations to pro-longed soil drying and the involvment of
xylem sap ABA in the regulation of stomatalbehaviour on sunflower plants. New Phytol113, 167-174
ANNEXES
Ecal : intensité de la transpiration (mmol[H2O]m-2.s-1)
gs : conductance stomatique pour la vapeurd’eau (mmol[H2O]m-2.s-1)gsmax: conductance stomatique maximaleQ : éclairement ou «irradiance» (μmol[pho-tons]m-2.s-1)RWC : teneur en eau relative (%)VPD : déficit de pression de vapeur d’eau at-
mosphérique (hPa)
ψb : potentiel hydrique foliaire de base (MPa).ψf: potentiel hydrique foliaire (MPa). ψfmin : po-tentiel hydrique foliaire minimum (MPa). ψsol :potentiel hydrique du sol (MPa). ψs : potentielosmotique (MPa).Δψ = ψb -ψfmin





![MARCHE FRANCAIS - Présentation Excelsior 2012-09 [Lecture seule]](https://static.fdocuments.fr/doc/165x107/586e02621a28ab42788c1114/marche-francais-presentation-excelsior-2012-09-lecture-seule.jpg)












